Область техники
Настоящее изобретение относится к способу, который используется в ферментирующей промышленности. Более конкретно, настоящее изобретение относится к способу для эффективного производства L-треонина посредством ферментации при помощи бактерии Escherichia. L-Треонин является незаменимой аминокислотой и может быть использован в качестве ингредиента в питательных смесях медицинского назначения. Далее, она имеет различные применения в питательных подкормках для животных, а также среди реагентов в фармацевтической и химической отраслях.
Уровень техники
L-Аминокислоты, такие как L-треонин и L-изолейцин, получают в промышленных масштабах посредством ферментации, используя бактерии, производящие аминокислоты, например коринеформные бактерии или Escherichia, которые могут производить L-аминокислоты. Помимо таких бактерий, производящих аминокислоты, также используют природные бактериальные штаммы или их искусственные мутанты. С целью повышения производительности используют рекомбинантные штаммы бактерий, в которых ферменты, участвующие в биосинтезе L-аминокислот, повышены в силу рекомбинации генов или подобных способов.
В частности, известны мутантные штаммы бактерий Escherichia, которые продуцируют L-треонин, такие как 6-диметиламинопурин-устойчивые (выложенный Патент Японии (Kokai) No 5-304969) и боррелидин-устойчивый штамм (Международная патентная публикация WO 98/04715). Известны способы получения L-треонина с помощью бактерий Escherichia, особенно с помощью штамма, в котором треониновый оперон амплифицирован при помощи плазмиды (US 5175107) или у которого ген фосфоенолпируваткарбоксилазы и ген аспартазы амплифицированы при помощи плазмиды (заявка на патент США No. 2002/0110876).
В качестве генов, кодирующих ферменты, принимающих участие в биосинтезе L-треонина у Escherichia coli, известны следующие гены: ген аспартокиназы III (lysC), ген аспартатсемиальдегиддегидрогеназы (asd), ген аспартокиназы I-гомосериндегидрогеназы (thrA), ген гомосеринкиназы (thrB) и ген треонинсинтазы (thrC). Гены thrA, thrB и thrC (thrABC) составляют треониновый оперон. Треониновый оперон образует структуру аттенюатора, и экспрессия его генов ингибируется в присутствии изолейцина и треонина в культуральной среде. Известно, что если удалить из треонинового оперона лидерную последовательность участка аттенюатора, то выход ферментативного производства улучшается (Патент США No. 5538873, Biotechnology Letters, Vol.24, No. 21, November 2002, и Международная патентная публикация WO 05/049808).
На сегодняшний день к уже разработанным способам получения L-треонина относятся использование периодической культуры, выращиваемой в ферментере, в котором изначально содержатся все питательные вещества, а также культуры с подпиткой, выращиваемой в ферментере, который содержит базовый набор питательных веществ и непрерывно пополняется одним или более дополнительными питательными компонентами (Патент СШA No. 5538873 и Европейский патент No 593792), и способ регулируемой концентрации сахаридов, которую поддерживают на определенном уровне или ниже (Международные патентные публикации WO 05/014840 и WO 05/014843). Помимо этого, разработан еще один способ получения L-треонина, согласно которому культуральная среда пополняется питательными компонентами таким образом, что фосфорная кислота и источники углерода являются сдерживающими факторами роста (Патент СШA No 5763230).
Сера является незаменимым компонентом для роста бактерий и обычно добавляется в среду для производства L-треонина посредством ферментации в виде сульфата аммония. Однако, что касается производства L-треонина путем ферментации, способы регуляции концентрации серы в ферментационной среде, а также влияние снижения концентрации серы еще неизвестны.
Описание изобретения
Целью настоящего изобретения является разработка способа эффективного получения L-треонина с помощью бактерий рода Escherichia, которые способны продуцировать L-треонин.
Авторы данного изобретения провели всесторонние исследования в целях достижения указанной цели. В итоге было установлено, что концентрация серы в среде может повлиять на результаты ферментации и что выход L-треонина в процессе ферментации может быть улучшен путем регуляции концентрации серы в ферментационном растворе на определенном уровне или ниже, и это изложено в настоящем изобретении.
Целью данного изобретения является разработка способа для получения L-треонина, который включает культивирование микроорганизма, принадлежащего к роду Escherichia, который способен продуцировать L-треонин в ферментационной среде, содержащей источник углерода, источник азота и источник серы, а также выделение L-треонина из среды, при этом концентрация серы в среде регулируется на определенном уровне или ниже.
Следующей целью настоящего изобретения является разработка изложенного выше способа, при этом концентрация серы в ферментационой среде регулируется таким образом, что она составляет 0,35 г/л или ниже.
Следующей целью настоящего изобретения является изложенный выше способ, при этом микроорганизмом является Escherichia coli.
Следующей целью настоящего изобретения является изложенный выше способ, при этом фермент биосинтеза L-треонина в микроорганизме изменен таким образом, что он не является мишенью ингибирования L-треонином по механизму обратной связи.
Следующей целью настоящего изобретения является изложенный выше способ, в котором фермент биосинтеза L-треонина выбран из группы, в которую входят аспартокиназа, гомосеринкиназа, треонинсинтаза и их комбинации.
Следующей целью настоящего изобретения является изложенный выше способ, при этом источник серы выбран из группы, в которую входят сульфаты, тиосульфаты, сульфиты, цистеин, цистин, глутатион и их комбинации.
Следующей целью настоящего изобретения является изложенный выше способ, в котором способ культивирования выбран из группы, в которую входят способ периодического культивирования, способ подпитываемого культивирования и способ непрерывного культивирования.
Следующей целью настоящего изобретения является изложенный выше способ, в котором способом культивирования является способ подпитываемого культивирования или способ непрерывного культивирования, при этом подпитывающую среду, содержащую источник серы, добавляют к ферментирующей культуре.
Следующей целью настоящего изобретения является изложенный выше способ, в котором указанная подпитывающая среда содержит также источник углерода и питательный компонент, ускоряющий рост, при этом указанную подпитывающую среду добавляют к ферментационой среде непрерывно или периодически так, чтобы концентрация источника углерода в культуральной среде поддерживалась на уровне 30 г/л или ниже после окончания периода логарифмического роста микроорганизма.
Следующей целью настоящего изобретения является разработка способа получения кормовых добавок для животных на основе ферментации, включая
А) культивирование микроорганизма из рода Escherichia, который способен к продуцированию L-треонина в ферментационной среде, которая включает источник углерода, источник азота и источник серы,
Б) проведение ферментации, при этом концентрация серы в среде регулируется таким образом, что она поддерживается на определенном уровне или ниже,
В) высушивание необработанного ферментационного бульона до состояния, когда содержание воды составляет 10% по массе или менее.
Краткое описание чертежа
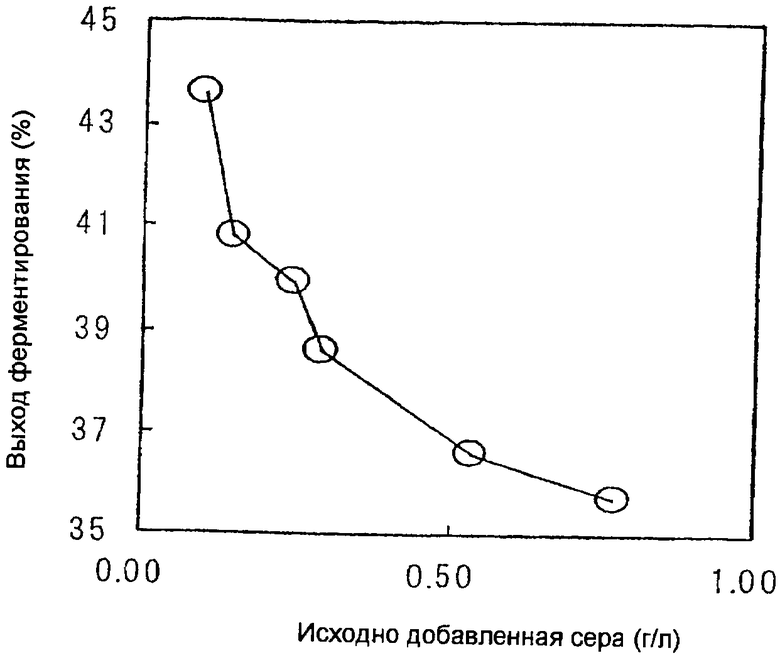
На чертеже изображена взаимосвязь между количеством исходно добавленной к среде серы и выходом L-треонина.
Подробное описание предпочтительного варианта осуществления
1. Способ настоящего изобретения
Способом настоящего изобретения является способ получения L-треонина методом культивирования микроорганизма, принадлежащего к роду Escherichia, который обладает способностью к продуцированию L-треонина в среде для ферментации, в которой содержится источник углерода, источник азота и источник серы, причем L-треонин продуцируется в среде, где концентрация серы регулируется таким образом, что она поддерживается на определенном уровне или ниже. В настоящем изобретении термин «концентрация серы» обозначает концентрацию источника серы, имея в виду концентрацию атомов серы.
Среда, используемая в настоящем изобретении, может быть любой жидкой средой, которая содержит источник углерода, источник азота и источник серы в виде питательных компонентов, и среда не особо ограничена, за исключением того, что в ней регулируется концентрация серы, которая поддерживается на определенном уровне или ниже. Источником серы может быть любое вещество, содержащее серу. Желательно использовать соли серной кислоты, такие как сульфаты, тиосульфаты и сульфиты, а также серосодержащие аминокислоты, такие так цистеин, цистин и глутатион. Среди данных особенно предпочтителен сульфат аммония. Перечень таких солей не особенно ограничен, также можно использовать соли аммония, соли кальция, соли натрия, соли калия, соли магния, соли марганца и соли железа. Более того, в среде может содержаться и один тип, и два типа, и даже больше типов таких веществ. Под обозначением концентрации «на определенном уровне или ниже» может быть любая концентрация, при которой выход L-треонина улучшен по сравнению с условиями высокой концентрации серы или в стандартной ферментационой среде. В особенности, концентрация серы в ферментационной среде предпочтительна 0,35 г/л или ниже, более предпочтительна 0,25 г/л или ниже и особенно предпочтительна 0,10 г/л или ниже. Одной из особенностей настоящего изобретения является то, что концентрация серы в ферментационой среде подлежит регуляции. Для способа, описанного в данном изобретении, могут быть использованы периодическая культура, культура с подпиткой и/или постоянная культура, и концентрацию серы в среде можно регулировать для удержания на определенном уровне или ниже в исходной среде, или ограничена на определенном уровне или ниже путем контролирования концентрации серы в подпитывающей среде, или можно использовать эти методы в сочетании. Для исходной среды и подпитывающей среды может быть использован один и тот же источник серы или же в подпитывающей среде может быть использован другой источник серы, нежели в исходной среде.
В настоящем изобретении среда с подпиткой обозначает метод культивирования, при котором среда добавляется непрерывно или периодически в сосуд во время культивирования, не удаляя среду из сосуда до окончания культивирования. Непрерывное культивирование означает метод непрерывного культивирования или периодического добавления среды в сосуд в течение культивирования и забор среды из сосуда (обычно в объеме, равном объему подпитывающей среды). Термин «исходная среда» означает среду, которая используется для периодической культуры до добавления подпитывающей среды при использовании подпитываемой культуры или постоянной культуры, а термин «подпитывающая среда» означает среду, которая вносится в ферментер, если производится культивирование с подпиткой или непрерывное культивирование. Подпитывающая среда может содержать все или только часть необходимых для роста микроорганизма компонентов. В настоящем изобретении термин «ферментационная среда» означает среду в ферментере, и L-треонин выделяется из данной ферментационной среды. Далее, в данном изобретении термин «ферментер» означает сосуд, в котором проводится ферментирование L-треонина, и он не ограничен по форме. Могут быть использованы бак или колба для ферментирования. Более того, у ферментера нет ограничения по объему до тех пор, пока можно получить и выделить L-треонин.
Несмотря на то, что концентрация серы предпочтительно ограничена на определенном уровне или ниже в течение всего процесса культивирования, ограничение по концентрации серы может иметь место только для части процесса. Например, когда способ данного изобретения включает стадию, когда клетки пролиферируют (фаза роста), и стадию, когда продуцируется L-треонин (L-треонин продуцирующая фаза), достаточно, чтобы концентрация серы была ограничена определенным уровнем или ниже в течение фазы продукции L-треонина. В фазе роста (во время пролиферации клеток) сера может присутствовать в среде в более высокой, чем определенная, концентрации, или же концентрация серы может быть в пределах определенного уровня или ниже. Более того, в фазу продукции L-треонина содержание серы не обязано находится в пределах вышеупомянутых значений в течение всего периода данной фазы, и содержание серы может быть в пределах вышеупомянутого уровня или выше его на раннем этапе данной фазы, и может быть сокращено со временем культивирования. И далее, серу можно периодически добавлять, кода ее уровень снижается. Термин «фаза роста», использующийся в настоящем изобретении, означает период в течение 3 часов, предпочтительно 6 часов, особенно предпочтительно 10 часов от начала культивирования, в течение которого источник углерода главным образом расходуется на рост бактериальных клеток, в логарифмической фазе роста. Термин “фаза продуцирования L-треонина”, использующийся в настоящем изобретении, означает период в течение 3 часов, предпочтительно 6 часов, особенно предпочтительно 10 часов, от начала культивирования, в течение которого источник углерода главным образом расходуется на продуцирование L-треонина.
Достаточно, чтобы в ферментационной среде содержалось минимальное количество серы, которое необходимо для роста микроорганизма; однако количество серы может временно подходить к концу. Фраза «подходить к концу» означает, что концентрация серы понижена в сравнении с предшествующими временными точками культивирования и даже может становится «0». Термин «временно» означает, что, например, в определенное время содержание серы может подойти к концу, и этот период времени соответствует примерно 20%, примерно 40% или в крайней степени примерно 60% от всего периода ферментации. Хотя концентрация серы временно может достигать 0, в течение этого времени содержание серы в ферментационной среде составляет 1 мкг/л или более, 10 мкг/л или более, или 100 мкг/л или более. Таким образом, даже если концентрация серы временно становится 0, выражение «культивирование микроорганизма, принадлежащего роду Escherichia, в ферментационной среде, содержащей источник углерода, источник азота и источник серы», подходит для описания культивирования в среде, содержащей серу в любое время. Концентрация серы в среде для культивирования можно измерять методом ионной хроматографии и методом горячего флакона.
Более того, согласно настоящему изобретению, концентрация серы в подпитывающей среде может быть доведена таким образом, чтобы она была в пределах предустановленной или ниже, если используется метод культивирования с подпиткой. Например, если концентрация серы ограничена при культивировании с подпиткой, предпочтительно следили за тем, чтобы концентрация серы в ферментационной среде была 0,35 г/л или менее, желательно 0,25 г/л или менее и более желательно 0,10 г/л или менее.
В настоящем изобретении примером источников углерода могут служить глюкоза, глицерин, фруктоза, сахароза, мальтоза, манноза, галактоза, гидролизированный крахмал и меласса. Особенно предпочтительны глюкоза и сахароза. В дополнение, могут быть использованы по одиночке или в сочетании с другими источниками углерода органические кислоты, такие как уксусная кислота и лимонная кислота, и спирты, такие как этанол. Более того, необработанным источником углерода может быть тростниково-сахарная меласса, меласса сахарной свеклы и цитрусовая меласса, а также гидролизаты природных сырых материалов, таких как целлюлоза, крахмал, зерновые культуры, крупы и тапиока. Помимо этого, в качестве источника углерода можно использовать растворенную в среде двуокись углерода. Данные источники углерода можно использовать для приготовления исходной среды и/или подпитывающей среды. В среде могут содержаться один или более видов этих источников углерода. Более того, для приготовления исходной и подпитывающей сред можно использовать один и тот же источник углерода или для приготовления подпитывающей среды можно использовать иной источник углерода, нежели при приготовлении исходной среды. К примеру, глюкозу можно использовать для приготовления исходной среды, а сахарозу - для приготовления подпитывающей среды.
В настоящем изобретении примером источников азота могут служить аммиак, соли аммония, такие как сульфат аммония, карбонат аммония, хлорид аммония, фосфат аммония, ацетат аммония, а также мочевина, нитраты и так далее. Газообразный аммиак или водный раствор аммиака, которые применяются для доведения рН, также могут быть использованы в качестве источника азота. Более того, пептон, дрожжевой экстракт, мясной экстракт, солодовый экстракт, жидкий кукурузный экстракт, гидролизат соевых бобов и тому подобные также могут быть использованы. В среде могут содержаться один или более данных видов источников азота. Данные источники азота также могут быть использованы при приготовлении исходной среды и/или подпитывающей среды. Более того, для приготовления исходной и подпитывающей сред можно использовать один и тот же источник азота или для приготовления подпитывающей среды можно использовать иной источник азота, нежели при приготовлении исходной среды.
Более того, в настоящем изобретении среда предпочтительно содержит в добавление к источнику углерода, источнику азота и серы еще и источник фосфорной кислоты. В качестве источника фосфорной кислоты могут быть использованы дигидрофосфат калия, гидрофосфат калия и фосфатные полимеры, такие как пирофосфорная кислота.
Кроме того, в настоящем изобретении среда может содержать факторы роста (питательные компоненты, которые обладают эффектом стимуляции роста) в дополнение к источнику углерода, источнику азота и серы. В качестве факторов роста могут быть использованы микроэлементы, аминокислоты, витамины, жирные кислоты, нуклеиновые кислоты, такие как пептон, казаминовая кислота, дрожжевой экстракт, гидролизат белков соевых бобов.
Примерами микроэлементов служат железо, марганец, магний, кальций, калий, натрий и так далее. Примерами витаминов служат витамин В1, витамин В2, витамин В6, никотиновая кислота, никотинамид, витамин В12, и так далее. Эти факторы роста могут содержаться в исходной среде или в подпитывающей среде.
При этом, если используется ауксотрофный мутант, который нуждается в аминокислоте или подобном веществе для своего роста, предпочтительно добавлять в среду требуемый питательный компонент. В особенности, в силу того, что путь биосинтеза L-треонина проактивирован, а способность к разложению L-треонина ослаблена у тех L-треонинсинтезирующих бактерий, которые могут быть использованы в рамках настоящего изобретения, как указано выше, предпочтительно добавлять L-лизин, L-гомосерин, L-изолейцин, L-метионин или их комбинации.
Состав исходной и подпитывающей сред может быть одинаковым или различаться. Более того, концентрация серы в исходной среде и в подпитывающей среде может быть одинаковая или различная. Кроме того, если подпитывающая среда добавляется на нескольких этапах, состав подпитывающей среды, добавляемой на каждом из этапов, может быть одинаковым или различным.
Культивирование предпочтительно проводить в условиях аэрации, предпочтительно проводить ферментирование при температуре от 20 до 45°С, особенно предпочтительно от 33 до 42°С. Концентрация кислорода предпочтительно настроена на уровне от 5 до 50% и более предпочтительно на уровне примерно 10%. Кроме того, аэрацию предпочтительно проводить в условиях рН от 5 до 9. Если в процессе культивирования рН снижается, то можно для нейтрализации добавить, к примеру, карбонат кальция или основания, такие как газообразный аммиак или водный раствор аммиака. Если культивирование производится при данных условиях, предпочтительно в течение примерно от 10 до 120 часов, в среде накапливается значительное количество L-треонина. Концентрация накопившегося L-треонина не ограничена в силу того, что L-треонин можно удалить и выделить из среды, и она составляет 50 г/л или выше, желательно 75 г/л или выше и более желательно 100 г/л или выше.
В настоящем изобретении культивирование микроорганизма можно проводить в рассеянной культуре и/или в основной культуре с целью добиться накопления L-треонина в концентрации выше определенного уровня. Рассеянное культивирование можно проводить путем встряхивания, используя колбу или же что-то подобное, или культивирование в условиях одного производственного цикла. Основную культуру можно вести как культуру с подпиткой или постоянную культуру. Как вариант, рассеиваемую культуру и основную культуру можно вести как периодическую культуру.
При данных методах культивирования, если концентрация L-треонина достигает определенного уровеня, можно удалить часть ферментационного бульона и добавить свежей среды для повторения культивирования. В качестве свежей среды предпочтительно использовать среду, содержащую источник углерода и питательные компоненты, способные стимулировать рост (факторы, стимулирующие рост), и предпочтительно, если она содержит серу в определенной концентрации или ниже. Выражение «определенная концентрация или ниже» означает, что добавляемая среда приготовлена таким образом, что концентрация серы в ферментационном бульоне составляет 0,35 г/л или менее, желательно 0,25 г/л или менее и более желательно 0,10 г/л или менее. В качестве источника углерода предпочтение отдается глюкозе, сахарозе и фруктозе. В качестве факторов роста предпочтение отдается источнику азота, фосфорной кислоте, аминокислотам и так далее. В качестве источника азота можно использовать аммиак, соли аммония, такие как сульфат аммония, карбонат аммония, хлорид аммония, фосфат аммония, ацетат аммония и мочевина, нитраты и так далее. Более того, в качестве источника фосфорной кислоты можно использовать дигидрофосфат калия и гидрофосфат калия. В качестве аминокислот, в случае применения штамма ауксотрофного мутанта, предпочтительно снабжать необходимой аминокислотой.
Если в рамках данного изобретения используется способ периодического культивирования или непрерывного культивирования, добавление подпитывающей среды можно временно приостановить, таким образом снабжение сахаридами или питательными компонентами временно приостановится. Внесение подпитывающей среды предпочтительно приостановить самое большее на 30%, желательно на 20% и особенно желательно на 10% от периода подпитки. Под «периодом подпитки» понимается период от начала первого внесения подпитывающей среды до конца последнего добавления подпитывающей среды. Приостановление добавления подпитывающей среды может быть меньше 30% и наиболее предпочтительно на менее чем 10%. Если подпитывающая среда добавляется периодически, то подпитывающую среду можно в первый раз добавить через определенное время, а повторно и все последующие разы можно производить добавки так, чтобы они начались после того, как компьютер зафиксирует повышение рН или концентрации растворенного кислорода. Данные опознания обычно происходят при истощении источника углерода в ферментационной среде во время добавления/приостановления перед этапом определенного добавления, и таким образом, концентрация субстрата в емкости для культивирования следует автоматически и в нужной мере поддерживать на низком уровне (Патент СШA No 5912113).
Подпитывающая среда, которая используется для культивирования с подпиткой, предпочтительно содержит источник углерода и питательный компонент с эффектом стимуляции роста (фактор, стимулирующий рост), и может содержать серу, причем концентрация серы в ферментационной среде поддерживается на определенном уровне или ниже. Выражение «определенная концентрация или ниже», использующееся здесь, означает, что добавляемая среда готовится таким образом, что концентрация серы в ферментационной среде поддерживается на уровне 0,35 г/л или ниже, желательно 0,25 г/л или ниже и более желательно 0,10 г/л или ниже. Несмотря на то, что концентрация серы в подпитывающей среде может быть в или вне пределов вышеуказанного промежутка концентраций, но предпочтительно в пределах вышеуказанного промежутка концентраций.
В качестве источника углерода предпочтение отдается глюкозе, сахарозе и фруктозе. В качестве факторов роста предпочтение отдается источнику азота, фосфорной кислоте, аминокислотам и так далее. В качестве источника азота можно использовать аммиак, соли аммония, такие как сульфат аммония, карбонат аммония, хлорид аммония, фосфат аммония, ацетат аммония и мочевина, нитраты и так далее. Более того, в качестве источника фосфорной кислоты можно использовать дигидрофосфат калия и гидрофосфат калия. В качестве аминокислот, в случае применения штамма ауксотрофного мутанта, предпочтительно снабжать необходимой аминокислотой. Помимо этого, подпитывающая среда может быть одинаковая или же быть композицией из двух или более типов сред. Если используются два или более типов подпитывающих сред, то среду можно смешивать и добавлять, используя одну емкость или используя две или более емкостей.
Более того, если применяется культивирование с подпиткой, то добавление осуществляется предпочтительно таким образом, что количество сахаридов в конечной ферментационной среде или общей среде, применяемой для культивирования с подпиткой, не превышает 30 г/л, предпочтительно 20 г/л и более предпочтительно 10 г/л. В особенности, концентрацию сахаридов следует контролировать таким образом, чтобы она находилась в пределах вышеуказанных концентраций после завершения фазы логарифмического роста микроорганизма. Частоту подпитывания источником углерода можно контролировать способом, описанным в Патенте США No. 5912113. Помимо этого, подпитывание сахаридами и фосфорной кислотой осуществляется таким образом, что концентрации сахаридов и фосфорной кислоты являются лимитирующими факторами роста бактериальных клеток. Содержание фосфорной кислоты в подпитывающей среде является таким, что отношение фосфор/углерод (Ф/У) составляет 2 или меньше, предпочтительно 1,5 или меньше и более предпочтительно 1 или меньше (Патент СШA No. 5763230).
Если в рамках настоящего изобретения использовать метод непрерывного культивирования, забор и добавление среды можно осуществлять одновременно или часть среды может быть отобрана и после этот среда добавляется. Более того, методика культивирования может быть основана на способе непрерывного культивирования, при этом при заборе культуральной среды, содержащей L-треонин и бактериальные клетки, осуществляется рециркуляция только бактериальных клеток в ферментер (Патент Франции No. 2669935). В качестве способа непрерывного или периодического добавления питательных компонентов используется такой же способ, как и при культивировании с подпиткой.
При периодическом заборе среды с культурой часть L-треонина может быть выделена, если концентрация L-треонина достигает определенного уровня, и для продолжения культивирования добавляется свежая среда. Более того, что касается количества свежей среды, которую требуется добавить, то конечный общий объем среды после добавления свежей среды равен объему среды для культивирования до момента экстракции. Термин «равен», использующийся здесь, означает приблизительно от 93 до 107% от объема культуральной среды до экстракции.
Если же культуральная среда непрерывно отводится, забор предпочтительно начать в то же время или после, что и добавление питательной среды. К примеру, время начала составляет максимально 5 часов, предпочтительно 3 часа, более предпочтительно 1 час после начала добавления. Более того, количество среды, которое забирается, предпочтительно равно количеству среды, которое добавляется.
Способ непрерывного культивирования с многократным использованием бактериальных клеток является способом, при котором периодически или непрерывно забирается среда из ферментационной среды, когда концентрация аминокислоты достигает определенного уровня, причем забирается только L-треонин, и после рециркулирующей фильтрации остатки, включая бактериальные клетки, попадают в ферментер, и это можно воспроизвести, ссылаясь, например, на Патент Франции No. 2669935.
Анализ на L-треонин и другие аминокислоты можно провести с помощью анионообменной хроматографии и последующим анализом продукта обработки нингидрином, как описано у Spackman и соавт. (Analytical Chemistry 30:1190-1206(1958)), или с помощью ВЭЖХ с обратной фазой, как описано у Lindroth и соавт. (Analytical Chemistry 51:1167-1174).
2. Способ производства пищевой добавки для животных, основанный на ферментировании бульона, проведенном по способу данного изобретения
Изготовление пищевой добавки для животных согласно данному изобретению можно осуществить, придерживаясь следующей методики выделения.
Для удаления биомассы или уменьшения биомассы можно использовать такие способы отделения L-треонина, как центрифугирование, фильтрация, удаление хлопьев осадка или их комбинации.
Бульон, полученный согласно данному изобретению, можно уплотнить или сконцентрировать при помощи известных методик, таких как роторное эвапорирование, тонкослойное эвапорирование, обратный осмос или нанофильтрация (FR 8613346В, US 4997754, EP410005В, JP1073646В).
Сконцентрированный бульон далее подвергается воздействию таких способов, как замораживание-высушивание, спрэй-высушивание, спрэй-гранулирование, или другим обработкам для придания предпочтительно свободносыпучести, высокозернистости порошку для использования в качестве пищевой добавки для животных. Этот свободносыпучий, мелкозернистый порошок может быть переработан в крупнозернистый, высокосыпучий, устойчивый и в высокой степени свободный от пыли продукт, применяя подходящие процедуры прессования или гранулирования. В конечном итоге, более 90% воды удаляется таким способом, и в пищевой добавке остается лишь менее 10% воды, предпочтительно менее 5% от массы.
Содержание белка в пищевой добавке может быть менее 10%, предпочтительно менее 5% по весу, и концентрация L-треонина может быть более 50%, предпочтительно более 85% и более предпочтительно более 95% (US 5431933, JP 121463GB, US 4956471, US 4777051, US 4946654, US 5840358, US 6238714, US 2005/0025878).
Этап отделения, описанный выше, необязательно должен быть выполнен, но может быть совмещен технически подходящим способом.
3. Бактерии Escherichia, которые могут быть использованы в рамках настоящего изобретения
Бактерии Escherichia, которые могут быть использованы в рамках настоящего изобретения, это бактерии Escherichia, которые способны продуцировть L-треонин, и термин «способны продуцировать L-треонин», использованный в настоящем изобретении, означает способность, если бактерии культивируются в среде, продуцировать свободный L-треонин в среду, то есть за пределы клетки, в таком количестве, чтобы L-треонин можно было выделить из среды. Предпочтительно, чтобы в рамках настоящего изобретения использовали такой штамм бактерии Escherichia, который производит большее количество L-треонина в сравнении со штаммами дикого типа или родственными штаммами. В особенности предпочтительно, если бактерия Escherichia продуцирует L-треонин в количестве 30 г/л или более, более предпочтительно 50 г/л или более, особенно предпочтительно 75 г/л или выше, при использовании обычных методов культивирования, в которых не контролируется концентрация серы.
Список исходных штаммов бактерий Escherichia, которые можно использовать для получения бактерий Escherichia с целью использовать их в рамках настоящего изобретения, особенно не ограничен, и конкретные примеры таковых упоминаются в работе Neidhardt и соавт. (Neidhardt, F.C. и соавт., Escherichia coli and Salmonella Typhimurium, American Society for microbiology, Washington D.C., 1029, таблица 1). Среди указанных, например, предпочтительно использовать Escherichia coli. Конкретными примерами Escherichia coli являются Escherichia coli W3110 (ATCC 27325) и Escherichia coli MG1655 (ATCC 47076), которые (оба) происходят от одного прототипа - штамма К12 дикого типа, и так далее.
Что касается возможности приобретения указанных штаммов, то их можно получить, например, в Американском фонде типовых культур (адрес: P.O. box 1549 Manassas, VA 20108, United States of America). Каждый бактериальный штамм имеет соответствующий регистрационный номер, и любой штамм можно заказать, указав его регистрационный номер. Регистрационные номера, соответствующие бактериальным штаммам, представлены в каталоге Американского фонда типовых культур.
3-1. Привнесение способности продуцировать L-треонин
Здесь и далее будет описан метод внедрения способности продуцировать L-треонин в бактерии Escherichia.
Для получения способности продуцировать L-треонин могут быть использованы такие общепринято адаптированные для разведения бактерий Escherichia или коринеформных бактерий методы, как получение ауксотрофного мутанта, штамма, устойчивого к аналогу, или штамма с мутацией на уровне регуляции метаболизма, каждый из которых обладает способностью продуцировать L-треонин в той же степени, как создание рекомбинантного штамма с усиленной активностью системы биосинтеза L-треонина. Для примера, мутант или рекомбинантный штамм могут быть изменены таким образом, чтобы фермент биосинтеза L-треонина не являлся мишенью ингибирования по механизму обратной связи, или рекомбинантный штамм может быть модифицирован с целью усиления экспрессии гена, кодирующего фермент биосинтеза L-треонина. При разведении бактерий, продуцирующих L-треонин такими способами, можно придать им свойства, такие как ауксотрофность, устойчивость к аналогу и мутацию в системе регуляции метаболизма, отдельно или в сочетании. При увеличении активности фермента биосинтеза L-треонина может быть увеличен один или более типов активностей данного фермента. Более того, придание свойств, упомянутых выше, и усиление активности фермента, как упомянуто выше, могут быть применены в сочетании.
Ниже следует описание варианта способа по внедрению способности к продуцированию L-треонина в бактерию Escherichia или усилению способности к продуцированию L-треонина посредством усиления активности фермента биосинтеза L-треонина. Для того чтобы усилить ферментативную активность можно, например, внести мутацию в ген, кодирующий фермент, или увеличить число копий гена, и в результате внутриклеточная активность фермента возрастет. Такого можно достичь посредством применения методик генной рекомбинации.
В число генов, кодирующих ферменты биосинтеза L-треонина, входят ген аспартокиназы III (lysC), ген аспартатсемиальдегиддегидрогеназы (asd), ген аспартокиназы I в составе thr оперона (thrA, последовательность нуклеотидов с 337 по 2799 в последовательности SEQ ID NO:1), ген гомосеринкиназы (thrB, последовательность нуклеотидов с 2801 по 3733 в последовательности SEQ ID NO:1) и ген треонинсинтазы (thrC, последовательность нуклеотидов с 3734 по 5020 в последовательности SEQ ID NO:1). В скобках указаны аббревиатуры названий генов. Два или более этих генов могут быть введены в бактерию. Гены биосинтеза L-треонина можно ввести в бактерию Escherichia, в которой подавлены механизмы разрушения L-треонина. Примерами бактерий Escherichia, у которых подавлены механизмы разрушения треонина, являются штамм TDH6, который дефицитен по активности треониндегидрогеназы ЕР1149911А, и так далее.
Энзиматическая активность ферментов биосинтеза L-треонина подавляется L-треонином как конечным продуктом. В силу этого особое значение имеет создание L-треонинпродуцирующей бактерии с модифицированными генами биосинтеза L-треонина таким образом, чтобы ферменты не являлись мишенью ингибирования L-треонином по механизму обратной связи. Вышеупомянутые гены thrA, thrB и thrC составляют треониновый оперон, и треониновый оперон имеет структуру аттенюатора. Экспрессия треонинового оперона подавляется в присутствие изолейцина и треонина в культуральной среде и это осуществляется по механизму аттенюации. Вышеупомянутые модификации можно получить посредством удаления лидерной последовательности в участки аттенюатора (SEQ ID NO:6) или путем удаления аттенюатора (SEQ ID NO:1) (Международная патентная публикация WO 02/26993; Biotechnology Letters, Vol.24, No. 21, November 2002; Международные патентные публикации WO 05/049808, WO 98/04715, WO 03/097839). Особенно желательно модифицировать треониновый оперон таким образом, чтобы последовательность нуклеотидов со 188 по 310 была удалена из последовательности SEQ ID NO:1. В качестве альтернативы желательно модифицировать треониновый оперон таким образом, чтобы последовательность нуклеотидов с 148 по 320 была удалена из последовательности SEQ ID NO:1.
В участке, предшествующем треониновому оперону, находится его нативный промотор, который можно заменить другим не нативным промотором (согласно WO 98/04715). В качестве альтернативы можно сделать такой треониновый оперон, в котором экспрессия генов, вовлеченных в биосинтез треонина, регулируется репрессором и промотором фага лямбда (согласно Европейскому патенту No. 0593792). Помимо этого, можно получить модифицированную бактерию Escherichia с недействующей системой ингибирования L-треонином по механизму обратной связи путем выведения штамма, устойчивого к α-амино-β-гидроксивалериановой кислоте (АГВ) (согласно WO 03/097839).
Более того, предпочтительно внести несколько копий треонинового оперона с модификацией, избавляющей его от подавляющего воздействия L-треонина по механизму обратной связи. В качестве альтернативы оперон можно поместить вместе с сильным промотором и в результате получить повышенную экспрессию. Получить несколько копий L-треонинового оперона можно путем амплификации, используя плазмиду и/или перенеся его на хромосому посредством транспозона, Mu-фага или подобным способом (US 5175107).
Помимо этого, предпочтительно использовать ген аспартокиназы III (lysC), модифицированный таким образом, что он не является мишенью подавления L-треонином по механизму обратной связи. Такой ген lysC можно получить способом, изложенным в Патенте США No. 5932453.
Также предпочтительно наряду с ферментами биосинтеза L-треонина усилить экспрессию генов, относящихся к гликолитическому пути, циклу ТКК или дыхательной цепи, генам, регулирующим экспрессию гена, генов, отвечающих за потребление сахаридов. Примерами генов, влияющих на продукцию L-треонина, служат ген трансгидрогеназы (pntAB) (Европейский Патент No. 733712), ген фосфоенолпируваткарбоксилазы (pepC) (Международная патентная публикация WO 95/06114, US2005-0136518), ген фосфоенолпируватсинтазы (pps) (Европейский патент No. 877090) и ген пируваткарбоксилазы коринеформных бактерий или бактерий Bacillus (Международная патентная публикация WO 99/18228; Публикация Европейского патента No. 1092776).
Помимо этого, предпочтительно иметь повышенную экспрессию гена, который придает устойчивость к L-треонину, или гена, придающего устойчивость к L-гомосерину, или их обоих в комбинации у хозяина. Примером гена, придающего устойчивость, является ген rhtA (Res. Microbiol., 2003 Mar., 154(2): 123-35), ген rhtB (Публикация Европейского патента No 0994190), ген rhtC (Публикация Европейского патента No. 1013765), ген yfiK и ген yeaS (Публикация Европейского Патента No 1016710). Боле того, способ наделения хозяина устойчивостью к L-треонину описан в Публикации Европейского патента No. 0994190 и Международной патентной публикации WO 90/04636.
Активность ферментов, кодируемых вышеуказанными генами, можно повысить посредством увеличения экспрессии генов, например, при увеличении числа копий генов в клетке с помощью методик рекомбинации генов. Например, фрагмент ДНК, содержащий ген-мишень, можно лигировать в функционирующий в данном микроорганизме-хозяине вектор, предпочтительно многокопийный вектор, и микроорганизм-хозяин может быть трансформирован данным ДНК-вектором.
Если в качестве гена-мишени выбран ген Escherichia coli, то этот ген можно получить посредством изготовления праймеров, со ссылкой на известную последовательность гена в Escherichia coli MG1655 или W3110 в GenBank, и проведения полимеразной цепной реакции (ПЦР) с использованием в качестве матрицы хромосомной ДНК Escherichia coli (согласно White, T.J. и соавт., Trends Genet. 5, 185 (1989)). В качестве гена-мишени можно выбрать ген другого микроорганизма и с помощью ПЦР поднять его с хромосомной ДНК или библиотеки хромосомных ДНК микроорганизмов с помощью праймеров, полученных на основе известной информации о генах микроорганизмов или последовательности из GenBank, или путем гибридизации с использованием олигонуклеотида, приготовленного на основе вышеперечисленных последовательностей в качестве пробы. Хромосомная ДНК может быть получена из организма-донора, например, по способу Saito и Miura (согласно H. Saito и K.Miura, Biochem. Biophys. Acta, 72, 619 (1963); Text for Bioengineering Experiments, Edited by the Society for Bioscience and Bioengineering, Japan, pp.97-98, Baifukan, 1992) и так далее.
Более того, поскольку нуклеотидные последовательности генов, кодирующих ферменты-мишени, могут различаться в зависимости от вида бактерий Escherichia или штамма бактерии, число генов-мишеней, использующихся в данном изобретении, не ограничивается лишь известными генами или последовательностями генов, зарегистрированными в GenBank, и могут включать мутантные гены и искусственно модифицированные гены. Такой мутантный или искусственно модифицированный ген может кодировать белок, в последовательности которого есть замены, делеции, инсерции, добавки или что-либо подобное, одной или нескольких аминокислотных остатков в одном или более мест, в такой мере, что способность закодированного белка-мишени продуцировать L-треонин может быть только улучшена путем увеличения числа копий гена. Термин «несколько», использованный здесь, означает число, которое различно и зависит от положения аминокислотного остатка в трехмерной структуре белка и типа аминокислотного остатка. Однако в особенности это число от 1 до 20, предпочтительно от 1 до 10, более предпочтительно от 1 до 5.
Предпочтительно, если вышеупомянутые замены являются консервативными заменами, то есть нейтральными мутациями, не приводящими ни к каким функциональным изменениям. Консервативная мутация это такая мутация, в случае которой замена осуществляется между Phe, Trp и Tyr, если замена осуществляется в положении ароматической аминокислоты; между Leu, Ile и Val, если замена осуществляется в положении гидрофобной аминокислоты; между Gln и Asn, если замена осуществляется в положении полярной аминокислоты; между Lys, Arg и His, если замена осуществляется в положении основной аминокислоты; между Asp и Glu, если замена осуществляется в положении кислой аминокислоты; между Ser и Thr, если замена осуществляется в положении аминокислоты с гидроксильной группой. Более конкретно, примерами консервативных замен служат замена Ser или Thr на Ala, замена Gln, His или Lys на Arg, замена Glu, Gln, Lys, His или Asp на Asn, замена Asn, Glu или Gln на Asp, замена Ser или Ala на Cys, замена Asn, Glu, Lys, His, Asp или Arg на Gln, замена Gly, Asn, Gln, Lys или Asp на Glu, замена Pro на Gly, замена Asn, Lys, Gln, Arg или Tyr на His, замена Leu, Met, Val или Phe на Ile, замена Ile, Met, Val или Phe на Leu, замена Asn, Glu, Gln, His или Arg на Lys, замена Ile, Leu, Val или Phe на Met, замена Trp, Tyr, Met, Ile или Leu на Phe, замена Thr или Ala на Ser, замена Ser или Ala на Thr, замена Phe или Tyr на Trp, замена His, Phe или Trp на Tyr и замена Met, Ile или Leu на Val.
Более того, можно использовать гомологичный ген, если он функционально такой же, как и ген-мишень. В особенности, гомологичный ген должен кодировать белок, который гомологичен последовательности известного белка на 80% или более, предпочтительно на 90% или более, более предпочтительно на 95% или более, в особенности предпочтительно на 97% или более. Более того, поскольку вырожденность гена зависит от хозяина, которому передают данный ген, то можно использовать ген с заменами в кодонах, которые бы легко использовался хозяином. Подобно этому, поскольку способность гена продуцировать L-треонин может быть улучшена путем его амплификации, то можно укоротить или удлинить ген с N- или с С-конца. Размер удлинения или укорочения, например, составляет 50 или менее, предпочтительно 20 или менее, более предпочтительно 10 или менее, особенно предпочтительно 5 или менее, аминокислотных остатков. Более определенно, аминокислотная последовательность белка может быть укорочена на 5-50 аминокислотных остатков с N-конца и/или с С-конца.
Гомологичный гену-мишени ген можно получить путем изменения нуклеотидной последовательности, например, путем сайт-специфического мутагенеза, при этом в закодированном белке содержатся замены, делеции, инсерции или добавки аминокислотных остатков в определенном положении. Более того, такой ген также можно получить с помощью известных мутагенных методик, которые описаны ниже. Примерами таких мутагенизирующих протоколов являются способ с обработкой нуклеотидной последовательности гена-мишени гидроксиламином или подобным соединением in vitro, способ с обработкой микроорганизма, несущего ген, к примеру бактерию Escherichia coli, УФ-излучением или типичным мутагенизирующим соединением, таким как N-метил-N'-нитро-N-нитрозогуанидин (НТГ) или этилметансульфонат (ЭМС), и способ внесения мутации посредством склонного к ошибке ПЦР. Помимо этого, вышеперечисленные замены, делеции, инсерции, добавки, инверсии или подобные им, аминокислотных остатков включают мутации, происходящие в естественных условиях (мутанты или варианты), такие как мутации, основанные на индивидуальных различиях и/или видовых различиях микроорганизмов, несущих ген-мишень. С целью выяснения того, улучшают ли белки, кодируемые этими генами, способность продуцировать L-треонин, при условии, что уровень их экспрессии повышен, можно, например, ввести эти гены в штамм Escherichia coli дикого типа или штамм, способный продуцировать L-треонин, и проверить, приводит ли это к улучшению способности продуцировать L-треонин.
В роли гена-мишени также может выступать ДНК, которая гибридизуется с нуклеотидной последовательностью-мишенью, или комплементарная ей последовательность, или проба, приготовленная из данной последовательности в строгих условиях, и кодирующая белок, который улучшает способность бактерии Escherichia coli продуцировать L-треонин посредством усиления его экспрессии. Под «строгими условиями», на которые здесь ссылаются, понимаются такие условия, при которых образуется так называемый специфический гибрид, а неспецифические гибриды не образуются. Четкое описание смысла этого термина с использованием цифр представляется достаточно трудным. Однако, например, строгие условия включают условия, при которых ДНК имеют высокую гомологию, к примеру ДНК, имеющие гомологию 50% или более, предпочтительно 60% или более, более предпочтительно 70% или более, более предпочтительно 80% или более, более предпочтительно 90% или более, и еще более предпочтительно 95% или более, и наиболее предпочтительно 97% или более, гибридизуются друг с другом, а ДНК с гомологией меньше, чем указано выше, не гибридизуются друг с другом. В качестве альтернативы строгие условия подразумевают отмывку один раз, предпочтительно от 2 до 3 раз, в условиях для гибридизации при концентрации соли, типичной для отмывки при Саузерн гибридизации, т.е. 1хSSC, 0,1% SDS при 60°С, предпочтительно 0,1% SSC, 0,1% SDS при 68°С.
Далее, рекомбинантная ДНК готовится путем лигирования гена-мишени, амплифицированного ПЦР, в ДНК вектор, который может функционировать в микроорганизме-хозяине. К подобным векторам относятся вектора, которые автономно размножаются в клетке микроорганизма-хозяина. Примерами таких автономно размножающихся в Escherichia coli векторов служат pUC19, pUC18, pHSG299, pHSG399, pHSG398, pACYC184 (pHSG и pACYC имеются в наличие у Takara bio Inc.), RSF1010, pBR322, pMW219 (pMW имеется в наличие у Nippon Gene) и так далее.
Рекомбинантная ДНК, полученная, как описано выше, может быть включена в микроорганизм посредством известных методов трансформации. Примерами служат способы обработки клеток реципиента хлоридом кальция для повышения проницаемости ДНК, как показано для Escherichia coli K-12 (Mandel, M. и Higa, A., J. Mol. Biol., 53, 159 (1970)), метод приготовления компетентных клеток из клеток, находящихся в фазе роста, и последующего включения в них ДНК, как показано для Bacillus subtilis (Duncan, C.H., Wilson, G.A. и Young, F.E., Gene, 1, 153, (1977)), и так далее. В дополнение к этому также можно воспользоваться методом перевода ДНК клетки-реципиента с протопласта или сферопласта, которые могут охотно захватывать рекомбинантную ДНК после добавления рекомбинантной ДНК к ДНК в клетках-реципиента, что известно и применимо в случае Bacillus subtilis, актиномицетов и дрожжей (Chang, S. и Cohen, S.N., Molec. Gen. Genet., 168, 111, 1979; Bibb, M.J., Ward, J.M. и Hopwood, O.A., Nature, 274, 398, 1978; Hinnen, A., Hicks, J.B. и Fink, G.R., Proc. Natl. Sci. USA, 75, 1929 (1978)).
Достичь увеличения числа копий гена можно путем введения многих копий гена в хромосомную ДНК микроорганизма. Для того чтобы ввести множество копий, можно провести гомологичную рекомбинацию, используя в качестве последовательностей-мишеней последовательности, представленные в хромосомной ДНК в множестве копий. В качестве последовательностей, которые представлены в хромосомной ДНК в множестве копий, можно использовать ДНК повторы и инвертированные повторы, расположенные на концах мобильных элементов. Ген-мишень также можно поместить в последовательность ненужного гена на хромосоме посредством гомологичной рекомбинации или поместить в ненужный район тандем из генов-мишеней. В качестве альтернативы, как описывается в выложенном патенте Японии No 2-109985, можно также ген-мишень встроить в транспозон и позволить ему перемещаться с включением множества копий гена в хромосомную ДНК (см. Патент СШA No. 5595889). Для проверки того, встроился ли ген-мишень в хромосомную ДНК, можно сделать гибридизацию по Саузерну, используя в качестве пробы часть гена-мишени.
Более того, помимо увеличения числа копий гена экспрессия гена-мишени может быть усилена посредством замены последовательности, которая регулирует экспрессию, такой как промотор гена-мишени на хромосомной ДНК или в плазмиде на более сильный промотор путем амплификации регуляторной последовательности, которая усиливает экспрессию, или удаления/подавления регуляторной последовательности, которая ослабляет экспрессию, как описано в Международной патентной публикации WO 00/198935. К примеру, lac-промотор, trp-промотор, trс-промотор, Pr-промотор, происходящие из фага лямбда, и т.п., как известно, являются сильными промоторами. Более того, промотор гена-мишени может быть усилен посредством нуклеотидных замен в область промотора. Способ оценки силы промотора, как и для описания известных сильных промоторов, приведен у Goldstein и соавт. (Procaryotic promoters in biotechnology, Biotechnol. Annu. Rev., 1995, 1, 105-128). Помимо этого, известно, что замена спейсера между участком связывания рибосомы (RBS) и старт-кодоном, в частности замена нескольких нуклеотидов в последовательности непосредственно выше старт-кодона, имеет большое влияние на эффективность трансляции мРНК, и эти нуклеотиды можно модифицировать. Участки, ответственные за регуляцию экспрессии, такие как промотор, можно определить с помощью вектора для нахождения промотора или программного обеспечения для анализа генов, такого как GENETYX. Экспрессия гена-мишени усиливается в результате замены или модификации таких промоторов. Участок, ответственный за регуляцию экспрессии, может быть заменен при помощи, например, температурочувствительной плазмиды.
В результате повышения активности при помощи указанных методик, ферментативная активность возрастает как минимум на 10%, 25%, 50%, 100%, 200%, 400%, 600% или 1000% по сравнению со штаммом дикого типа или немодифицированным штаммом. Помимо этого, способы повышения ферментативной активности могут быть использованы как в сочетании друг с другом, так и с понижением ферментативной активности. Примерами бактерий Escherichia, которые служат в качестве примера, являются штаммы дикого типа, Escherichia coli W3110 (ATCC 27325), Escherichia coli MG1655 (ATCC 47076) и так далее, которые происходят от прототипного штамма дикого типа K-12.
Помимо этого, в настоящем изобретении у бактерий, у которых активность фермента, катализирующего реакцию, ответвляющуюся от пути биосинтеза L-треонина и продукции вещества, отличного от L-треонина, может быть уменьшена или отсутствовать. В число примеров таких ферментов входят треониндегидрогеназа, треониндеаминаза и треониндегидратаза. Штаммы с пониженной или отсутствием активности указанных ферментов приведены в Международной патентной публикации WO 95/23864, WO 96/17930, WO 03/080843, WO 04/087895 и так далее.
Помимо этого, в настоящем изобретении у бактерий активность фермента, вовлеченного в путь гликолиза, цикл ТКК или дыхательную цепь, и который негативно влияет на продукцию L-треонина, фермент, регулирующий экспрессию гена, или фермент из системы биосинтеза стороннего продукта может быть уменьшен или сделан неполноценным, в добавление к системе ферментов деградации L-треонина. Примером гена, кодирующего такой фермент, служит ген, кодирующий σ (сигма) фактор РНК полимеразы (rpoS, Международная патентная публикация WO 01/05939 и WO 03/074719), ген фосфоенолпируваткарбоксилазы (pckA, Международная патентная публикация WO 02/29080), ген изомеразы фосфоглюкозы (pgi, Molecular and General Genetics, 217(1): 126-31 (1989)), ген пируватоксидазы (poxB, Международная патентная публикация WO 02/29080) и так далее.
Для того чтобы избавиться от активности фермента, как описано выше, в ген вышеупомянутого фермента на хромосоме с помощью обычных мутагенизирующих методик можно вставить мутацию, которая бы сокращала или элиминировала внутриклеточную активность данного фермента. Этого можно добиться, например, с помощью удаления кодирующего фермент гена на хромосоме посредством генной рекомбинации, модификации последовательности, контролирующей экспрессию гена, такой как промотор и последовательности Шайн-Дельгарно (SD), и так далее. Более того, этого также можно достичь внесением одной или более аминокислотных замен (миссенс мутации), вставкой стоп-кодона (нонсенс мутация), внесением мутации со сдвигом рамки считывания, которая добавляет или удаляет один или два нуклеотида, или удалить часть или целый участок гена (Journal of Biological Chemistry, 272: 8611-8617 (1997)) в области, кодирующей фермент, на хромосоме. Помимо этого, ферментативную активность также можно понизить или свести к нулю посредством сборки гена, кодирующего мутантный фермент, у которого отсутствует кодирующая область, и заменить этим геном нормальный ген на хромосоме путем гомологичной рекомбинации или подобным образом, или путем вставки транспозона или IS фактора в ген. Более того, это так же может быть получено путем подавления экспрессии с помощью антисмысловых РНК (Proceedings of the National Academy of Sciences, USA, 95 5511-5515 (1998); Journal of Biological Chemistry, 266, 20833-20839 (1991)).
Для внесения мутации, сокращающей или избавляющей от активности фермента, как упоминалось ранее, посредством генной рекомбинации, например, используется следующий способ. Мутированный вариант гена может заменить ген-мишень на хромосоме посредством модификации части последовательности гена-мишени для того, чтобы приготовить мутантный ген таким образом, что не образуется нормально функционирующего фермента, и трансформировать бактерию Escherichia ДНК, содержащей такой ген, который приведет к рекомбинации между мутантным геном и геном на хромосоме. Подобный сайт-специфичный мутагенез, вовлекающий внесение замены при помощи гомологичной рекомбинации, уже известен, и примерами таковых служат способ с использованием линейной ДНК, способ с использованием плазмиды, содержащей температурочувствительный сайт начала репликации (Патент США No 6303383; Открытый Патент Японии No 05-007491), и так далее. Более того, вышеупомянутый сайт-специфичный мутагенез посредством замены гена при помощи гомологичной рекомбинации также может быть проведен, используя плазмиду без способности к репликации в хозяине.
В результате снижения активности или экспрессии посредством такого метода, активность фермента сокращается до 75%, 50%, 25% или 10% в сравнении со штаммом дикого типа или немодифицированным штаммом, или его активность полностью исчезает. Помимо этого, два или более таких способов по снижению активности фермента могут быть применены в комбинации или могут быть использованы наряду с повышением ферментативной активности, описанной выше. Примерами бактерий Escherichia, выступающих в роли ссылок, являются Escherichia coli W3110 (ATCC 27325), Escherichia coli MG1655 (ATCC 47076) и так далее, которые происходят от прототипного штамма дикого типа К-12.
В качестве L-треонинпродуцирующей бактерии, используемой в рамках настоящего изобретения, может использоваться штамм, модифицированный таким образом, что он способен усваивать в качестве источника углерода сахарозу. Например, усваивающий сахарозу штамм можно получить из штамма E.coli H155 путем Р1 трансдукции, используя в качестве показателя способность расти на сахарозе как единственном источнике углерода. Для получения L-треонинпродуцирующей бактерии, усваивающей сахарозу в качестве источника углерода, можно обратиться к Международной патентной публикации WO 90/04636.
Помимо этого, в добавление к вышеупомянутым манипуляциям с генами, примерами способа получения L-треонинпродуцирующей бактерии являются способ воздействия на Escherichia coli УФ-излучением или обычными мутагенными агентами, такими как N-метил-N'-нитро-N-нитрозогуанидин (НТГ) или азотная кислота, с целью получить штамм, устойчивый к L-аминокислоте или аналогу L-аминокислоты, или ауксотрофный по L-аминокислоте штамм, и селекция штамма с улучшенной способностью к продукции L-треонина. Примерами таковых являются штамм, устойчивый к боррелидину (Патент США No 5939307), мутантный штамм, устойчивый к диаминосукциниловой кислоте (Международная патентная публикация WO 00/09661), мутантный штамм, устойчивый к уксусной кислоте (Патент США No. 5919670), мутантный штамм, устойчивый к фторпирувату (Международная патентная публикация WO 00/09661), мутантный штамм, устойчивый к α-метилсерину (Международная патентная публикация WO 00/09661), мутантный штамм, устойчивый к треонину (Международная патентная публикация WO90/04636), и мутантный штамм, устойчивый к AHV (Genetika, 16: 206 (1978)).
3-2. Примеры L-треонинпродуцирующих бактерий, которые могут быть использованы в рамках настоящего изобретения
Примеры бактерий Escherichia, наделенных способностью продуцировать L-треонин, которые могут быть использованы в рамках настоящего изобретения, будут описаны ниже. Однако бактерии, которые могут быть использованы в рамках данного изобретения, не ограничиваются этими примерами при условии, что бактерии обладают способностью продуцировать L-треонин.
В качестве примера бактерии, которая может быть использована в рамках данного изобретения, является штамм Escherichia coli VKPM B-3996 (Патент СШA No. 5175107). Этот штамм VKPM B-3996 хранится во Всероссийской Национальной Коллекции Промышленных Микроорганизмов (ВКПМ) (Россия, Москва, Дорожный проезд, 1) от 19 ноября 1987 с регистрационным номером VKPM В-3996. Штамм VKPM В-3996 несет плазмиду pVIC40 (Международная патентная публикация WO 90/04636), полученную при вставлении генов биосинтеза треонина (треониновый оперон thrABC) в pAYC32, которая является плазмидой у широкого спектра хозяев и несет маркер устойчивости к стрептомицину (Сhistorerdov, A.Y., Tsygankov, Y.D., Plasmid, 1986, 16, 161-167). В pVIC40 ингибитор аспартокиназы I-гомосериндегидрогеназы, закодированной как thrA в треониновом опероне, по механизму обратной связи посредством L-треонина удален.
Примером L-треонинпродуцирующей бактерии, которая может быть использована в рамках настоящего изобретения, служит штамм Escherichia coli VKPM B-5318 (Европейский патент No. 0593792). Данный штамм VKPM B-5318 был занесен во Всероссийскую Национальную Коллекцию Промышленных Микроорганизмов (ВКПМ) (Россия, Москва, Дорожный проезд, 1) 3 мая 1990 г. с регистрационным номером VKPM B-5318. Этот штамм VKPM B-5318 не нуждается в изолейцине, имеет термочувствительный С1 репрессор, PR-промотор и треониновый оперон. Это значит, что гены, вовлеченные в биосинтез треонина, у которых участок аттенюатора и область естественной регуляции транскрипции удалены и помещены позади гена с N-концевого участка белка Cro фага лямбда. Помимо этого штамм содержит ДНК рекомбинантную плазмиду, которая работает таким образом, что экспрессия генов, вовлеченных в биосинтез треонина, регулируется репрессором и промотором фага лямбда.
В качестве примера предпочтительной для использования бактерии, продуцирующей L-треонин, является Escherichia coli 427T23 (Патент США No. 5631157). Штамм 427Т23 имеет гомосериндегидрогеназу, которая не является мишенью ингибирования L-треонином по механизму обратной связи. Также у данного штамма подавлена активность треониндеаминазы, и он может утилизировать сахарозу в качестве источника углерода. Штамм 427Т23 занесен в Американскую коллекцию типовых культур с регистрационным номером ATCC 98082.
Штамм Escherichia coli kat-13 (Патент СШA No 5175107) также предпочтителен для использования в качестве бактерии для продуцирования L-треонина. Штамм kat-13 устойчив к боррелидину, имеет гомосериндегидрогеназу, не являющуюся мишенью для ингибирования L-треонином по механизму обратной связи, подавленную активность треониндеаминазы и может утилизировать сахарозу в качестве источника углерода.
Штамм Escherichia coli TDH-6 (патент Японии в открытом доступе No. 2001-346578), трансформированный генами, кодирующими ферменты биосинтеза L-треонина, также предпочтителен для использования в качестве бактерии для продуцирования L-треонина. Штамм TDH-6 дефицитен по активности треониндегидроназы и получен путем удаления pVIC40, которая несет гены, кодирующие ферменты биосинтеза треонина, из штамма B-3996 (США 5631157) и занесен во Всероссийскую Национальную Коллекцию Промышленных Микроорганизмов (ВКПМ) (Россия, Москва, Дорожный проезд, 1) с регистрационным номером VKPM B-3420.
Помимо этого, другими примерами L-треонинпродуцирующих бактерий, которые могут быть использованы в рамках настоящего изобретения, являются следующие бактериальные штаммы:
Escherichia coli MG-442 (CMIMB-1628, Патент СШA No 4278765)
Escherichia coli VL334/pYN7 (Патент СШA No 4278765)
Escherichia coli H-4225 (FERM BP-1236, Патент СШA No 5017483)
Escherichia coli Н-7256 (FERM BP-2137)
Escherichia coli DSM9807 (KCCM-10168)
Примеры
Настоящее изобретение будет объяснено более подробно со ссылкой на следующие примеры. Однако настоящее изобретение не ограничено данными примерами.
Пример 1. Влияние регуляции изначально добавленной серы при культивировании с подпиткой
Во-первых, оценивали влияние ограничения по сере в начальной среде основной культуры до определенной концентрации или менее в течение продуцирования L-треонина при культивировании с подпиткой.
Штамм VKPM В-5318 культивировали на чашке с агаром в LB среде (10 г/л триптона, 5 г/л дрожжевого экстракта, 5 г/л NaCl, 15 г/л агара), содержащей 20 мг/л стрептомицина сульфата при 37°С в течение 24 часов, и 1/10 часть клеток со всей чашки соскребали с одной чашки и инокулировали в 50 мл среды LB (10 г/л триптона, 5 г/л дрожжевого экстракта, 5 г/л NaCl), содержащей 20 мг/л стрептомицина сульфата, в закрытой колбе для получения посевной культуры при 40°С и 144 об/мин в течение 6 часов.
После завершения рассеивания культуры посев культуральной среды в объеме, эквивалентном 16% объема основной культуральной среды, вносили в 1-литровую колбу ферментера, заправленного 300 мл основной культуральной среды, и культивирование происходило при 40°С и рН 7,0. Состав основной культуральной среды написан ниже.
Состав основной культуральной среды:
Аммония сульфат добавляли в таком количестве, чтобы изначально содержание серы составляло от 0,78 до 0,10 г/л.
В процессе культивирования рН доводили до 7,0 посредством добавления газообразного аммиака. После утилизации сахаридов в среде, при культивировании с подпиткой, добавляли водный раствор сахарозы 600 г/л.
После 42 часов культивирования концентрацию L-треонина определяли по ВЭЖХ.
В результате выход L-треонина при ферментации возрастал при уменьшении серы и значительно возрастал, в особенности, при концентрации серы 0,29 г/л или ниже, как приведено в таблице 1.
Пример 2. Влияние регуляции серы в подпитывающей среде в культуре с подпиткой
Во-первых, таким же способом, как и в примере 1, штамм VKPM B-5318 культивировали в чашках на агаре со средой LB (10 г/л триптона, 5 г/л дрожжевого экстракта, 5 г/л NaCl, 15 г/л агара), содержащей 20 мг/л стрептомицина сульфата при 37°С в течение 24 часов, и 1/10 часть бактериальных клеток со всей чашки соскребали с одной чашки и инокулировали в 50 мл среды LB (10 г/л триптона, 5 г/л дрожжевого экстракта, 5 г/л NaCl), содержащей 20 мг/л стрептомицина сульфата, в закрытой колбе для получения посевной культуры при 40°С и 144 об/мин в течение 6 часов.
После завершения рассеивания культуры посев культуральной среды в объеме, эквивалентном 16% объема основной культуральной среды, инокулировали в 1-литровую колбу ферментера, заправленного 300 мл основной культуральной среды, и культивирование происходило при 40°С и рН 7,0. Состав основной культуральной среды был таким же, как и в примере 1.
В процессе культивирования рН доводили до 7,0 посредством добавления газообразного аммиака. После утилизации сахаридов в среде ферментирование проводили с добавлением подпитывающей среды, содержащей сахарозу, и поддерживали концентрацию серы в ферментационном растворе на определенном уровне или ниже. Результаты показаны в таблице 2. Концентрация серы в подпитывающей среде контролировалась на уровне от 0 до 1,22 г/л и культивирование контролировалось таким образом, что в ферментационной среде содержалось от 0,144 до 0,548 г/л серы на протяжении всего основного культивирования.
В результате было подтверждено, что выход L-треонина улучшается также и при культивировании с подпиткой путем регуляции концентрации серы в ферментационной среде на уровне 0,35 г/л или ниже.
Согласно изобретению выход ферментации и продукции L-треонина можно улучшить в способе продуцирования L-треонина путем ферментации с помощью микроорганизма, относящегося к роду Escherichia, который обладает способностью продуцировать L-треонин.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ | 2009 |
|
RU2518677C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ МЕТОДОМ ФЕРМЕНТАЦИИ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИЙ, ОБЛАДАЮЩИХ ПОВЫШЕННОЙ ЭКСПРЕССИЕЙ ГЕНОВ УТИЛИЗАЦИИ КСИЛОЗЫ | 2005 |
|
RU2283346C1 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ | 2008 |
|
RU2515044C2 |
| МУТАНТНАЯ АДЕНИЛАТЦИКЛАЗА, ДНК, КОДИРУЮЩАЯ ЕЕ, БАКТЕРИЯ СЕМЕЙСТВА ENTEROBACTERIACEAE, СОДЕРЖАЩАЯ УКАЗАННУЮ ДНК, И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ | 2010 |
|
RU2471868C2 |
| БАКТЕРИЯ, ПРИНАДЛЕЖАЩАЯ К РОДУ ESCHERICHIA - ПРОДУЦЕНТ L-ТРЕОНИНА И СПОСОБ ПОЛУЧЕНИЯ L-ТРЕОНИНА | 2004 |
|
RU2288264C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ ESCHERICHIA | 2005 |
|
RU2311454C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-треонина С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ Escherichia | 2006 |
|
RU2351646C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИЙ, ПРИНАДЛЕЖАЩИХ К РОДУ ESCHERICHIA | 2005 |
|
RU2304615C2 |
| СПОСОБ ПОЛУЧЕНИЯ ДИПЕПТИДА (ВАРИАНТЫ) | 2005 |
|
RU2316596C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-ТРЕОНИНА С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ Escherichia, В КОТОРОЙ ИНАКТИВИРОВАН ГЕН yahN | 2007 |
|
RU2392322C2 |













Изобретение относится к биотехнологии и представляет собой способ получения L-треонина, который включает в себя культивирование микроорганизма, принадлежащего к роду Escherichia, который способен продуцировать L-треонин, в ферментирующей среде, содержащей источник углерода, источник азота и источник серы, и выделение L-треонина из среды, при этом концентрация серы в среде регулируется таким образом, чтобы ее концентрация составляла 0,35 г/л или ниже. Изобретение позволяет получать L-треонин с высокой степенью эффективности. 2 н. и 17 з.п. ф-лы, 1 ил., 2 табл.
1. Способ получения L-треонина, который включает в себя
а) культивирование микроорганизма, принадлежащего к роду Escherichia, который способен продуцировать L-треонин, в ферментирующей среде, содержащей источник углерода, источник азота и источник серы,
б) выделение L-треонина из среды, при этом концентрация серы в среде регулируется таким образом, чтобы ее концентрация составляла 0,35 г/л или ниже.
2. Способ по п.1, где концентрация серы в ферментирующей среде регулируется таким образом, что она составляет 0,305 г/л или ниже.
3. Способ по п.2, где микроорганизмом является Escherichia coli.
4. Способ по п.3, где фермент биосинтеза L-треонина в микроорганизме изменен таким образом, что он не является мишенью для ингибирования L-треонином по механизму обратной связи.
5. Способ по п.4, где фермент биосинтеза L-треонина выбран из группы, состоящей из аспартокиназы, гомосеринкиназы, треонинсинтазы и их комбинаций.
6. Способ по п.5, где источник серы выбран из группы, состоящей из сульфатов, тиосульфатов, сульфитов, цистеина, цистина, глутатиона и их комбинаций.
7. Способ по любому из пп.1-6, где способ культивирования выбран из группы, состоящей из способа периодического культивирования, способа подпитываемого культивирования и способа непрерывного культивирования.
8. Способ по п.7, где способом культивирования является способ подпитываемого культивирования или способ непрерывного культивирования, при этом подпитывающая среда, содержащая источник серы, добавляется в ферментер.
9. Способ по п.8, где указанная подпитывающая среда дополнительно содержит источник углерода и питательный компонент, вызывающий эффект ускорения роста, при этом указанную подпитывающую среду добавляют к ферментирующей среде непрерывно или периодически, так, чтобы концентрация источника углерода в культуральной среде поддерживалась на уровне 30 г/л или ниже после окончания периода логарифмического роста микроорганизма.
10. Способ получения кормовых добавок для животных на основе ферментационного бульона, включающий в себя
A) культивирование микроорганизма из рода Escherichia, который способен к продуцированию L-треонина, в ферментирующей среде, которая содержит источник углерода, источник азота и источник серы, Б) проведение ферментации, при этом концентрация серы в среде регулируется таким образом, чтобы она поддерживалась на уровне 0,35 г/л или ниже,
B) высушивание необработанного ферментационного бульона до состояния, когда содержание воды составляет 10% по массе или менее.
11. Способ по п.10, где концентрация серы в ферментирующей среде регулируется таким образом, что она составляет 0,305 г/л или ниже.
12. Способ по п.11, где микроорганизмом является Escherichia coli.
13. Способ по п.12, где фермент биосинтеза L-треонина в микроорганизме изменен таким образом, что он не является мишенью для ингибирования L-треонином по механизму обратной связи.
14. Способ по п.13, где фермент биосинтеза L-треонина выбран из группы, состоящей из аспартокиназы, гомосеринкиназы, треонинсинтазы и их комбинаций.
15. Способ по п.14, где источник серы выбран из группы, состоящей из сульфатов, тиосульфатов, сульфитов, цистеина, цистина, глутатиона и их комбинаций.
16. Способ по п.15, где способ культивирования выбран из группы, состоящей из способа периодического культивирования, способа подпитываемого культивирования и способа непрерывного культивирования.
17. Способ по п.16, где способом культивирования является способ подпитываемого культивирования или способ непрерывного культивирования, при этом подпитывающая среда, содержащая источник серы, добавляется в ферментер.
18. Способ по п.16, где указанная подпитывающая среда дополнительно содержит источник углерода и питательный компонент, вызывающий эффект ускорения роста, при этом указанную подпитывающую среду добавляют к ферментирующей среде непрерывно или периодически, так, чтобы концентрация источника углерода в культуральной среде поддерживалась на уровне 30 г/л или ниже после окончания периода логарифмического роста микроорганизма.
19. Способ по любому из пп.10-18, где в состав указанной пищевой добавки входит более чем 95% L-треонина.
| WO 2005049808 A1, 02.06.2005 | |||
| EP 0001085087 A2, 21.03.2001 | |||
| JP 2002051787 A, 19.02.2002 | |||
| ФРАГМЕНТ ДНК, КОДИРУЮЩИЙ АСПАРТОКИНАЗУ III, СПОСОБ ПОЛУЧЕНИЯ L-ТРЕОНИНА | 1993 |
|
RU2113484C1 |
| US 20050025878 A1, 03.02.2005. | |||

