Изобретение относится к биотехнологии и иммунологии и представляет собой способ получения рекомбинатных белков путем синтеза в бактериальных клетках, обладающих антигенной активностью к вирусу гепатита E, и набор белков, полученных этим способом и позволяющих проводить серодиагностику гепатита Е. Гепатит E возникает в результате заражения человека вирусом гепатита Е. Обычным способом заражения вирусом гепатита E является употребление контаминированной фекалиями воды, возможен также трансплацентарный путь заражения ребенка от больной матери. Гепатит E является эндемичным заболеванием для стран с жарким климатом, во время эпидемий эта инфекция характеризуется значительной скоростью распространения и высокой летальностью среди беременных женщин (до 25%) и новорожденных детей, родившихся у заболевших матерей (до 77%). Поэтому для проведения экстренных противоэпидемических, профилактических мероприятий и своевременного начала лечения гепатита E крайне актуальна разработка диагностических тест-систем, позволяющих выявлять инфицированных людей на ранних этапах развития эпидемического процесса.
Многочисленные исследования молекулярно-биологических основ иммунного ответа на вирус гепатита E позволили определить структуру вирусного генома и локализовать основные вирусные антигенные детерминанты. Благодаря определению нуклеотидных последовательностей геномов вирусных изолятов различного географического происхождения и успехам в изучении антигенной структуры вирусных белков доказано существование двух основных серовариантов вируса: серотипа Бирма, к которому относят вирусные изоляты азиатского происхождения, и серотипа Мексика, объединяющего американские изоляты.
Первоначально диагностика гепатита E основывалась на серологическом исключении вирусных гепатитов другой этиологии. Большинство разработаных к настоящему времени методов диагностики гепатита E основано на определении антител к белкам-продуктам второй и третьей открытых рамок считывания генома вируса гепатита E в сыворотках крови (серодиагностика). Для этих целей используются различные подходы, в которых применяется принцип иммуноферментного анализа (ИФА), метода иммунофлюоресценции, иммуноблотинга. Применение этой группы методов базируется на использовании синтетических пептидов-фрагментов вирусных белков или рекомбинантных полипептидов, содержащих фрагменты вирусных антигенов. Одним из наиболее перспективных путей совершенствования диагностических тест-систем является применение в качестве антителосвязывающего субстрата рекомбинантных белков вируса гепатита E, синтезированных в различных экспрессирующих системах. Такие рекомбинантные белки, сохраняющие антигенные детерминанты вируса гепатит E, используются в серодиагностике заболевания, обеспечивая высокие показатели чувствительности и специфичности используемых тестов (патент US 5563032, кл. 435/5, 08.10.96).
Сущность данного технического решения состоит в том, что предложен способ получения рекомбинантных антигенов и набор для определения антител к вирусу гепатита E, содержащий в качестве антигена полипептид/полипептиды, полученные предложенным способом. Рекомбинантные антигены к вирусу гепатита E бирманского сероварианта получены с помощью методов генетической инженерии из штаммов-продуцентов E.coli. трансформированных рекомбинантными ДНК, содержащими фрагменты второй и третьей открытых рамок считывания вирусного генома. В качестве контрольного антигена набор может содержать бета-галактозидазу E. coli. При этом антигены, входящие в набор, иммобилизуют на твердом носителе в смеси или по отдельности в зависимости от решаемой задачи. Твердым носителем могут служить полистироловые или полихлорвиниловые шарики и планшеты для иммунологических реакций, нитроцеллюлоза, найлон, латексы, эритроциты, а также другие твердофазные носители. Повысить чувствительность и специфичность набора позволяет введение в набор дополнительного (по сравнению с прототипом) рекомбинантного антигена, содержащего фрагмент продукта гена ORF2 с 403 по 461 аминокислотные остатки, а также раздельная сорбция различных рекомбинантных антигенов, которая способствует определению спектра антител к индивидуальным вирусным белкам.
Изобретение иллюстрируется примерами.
Пример 1.
Получение фрагмента ДНК НЕ40.
Для получения фрагмента ДНК были использованы олигонуклеотиды d1, d2, r1, r2, приведенные в конце описания.
Олигонуклеотиды смешивают и кинируют с помощью полинуклеотидкиназы фага T4. Для этого в эппендорфовскую пробирку на 1,5 мл вносят по 50 пмолей каждого из олигонуклеотидов, 50 мкл десятикратного буфера для киназы (0,5 М Трис-HCl, pH 7,6, 0,1 М MgCl, 50 mM дитиотреитола, 1 mM спермидина и 1 mM ЭДТА), 250 пмолями (150 мкКИ) [гамма-P] АТФ (удельная активность 3000 Ки/моль), 150 единицами активности полинуклеотидкиназы фага T4 и бидистиллированной воды до объема 500 мкл и инкубируют 30-60 минут при температуре 37oC. Затем реакционную смесь нагревают до температуры 98oC, охлаждают до 16oC и смешивают с ДНК-лигазой. Для этого 100 мкл смеси обработанных киназой олигонуклеотидов смешивают с 12 мкл десятикратного буфера для лигирования (0,66 М Трис-HCl, pH 7,6, 50 mM MgCl, 50 mМ дитиотреитола и 10 mM АТФ), 8 мкл (50 единиц активности) ДНК-лигазы фага T4 и инкубируют при 4oC в течение 5 часов. Затем реакционную смесь нагревают до температуры 65oC в течение 15 минут для инактивации фермента.
10 мкл раствора обработанных киназой и лигазой олигонуклеотидов используют для анализа ДНК методом электрофореза в 12% полиакриламидном геле по обычной методике (Molecular cloning, New York, CSHL, 1982). После радиоавтографии геля виден ряд фрагментов размером больше или равным 92 п.н.
100 мкл раствора полученного фрагмента ДНК гидролизуют рестриктазами BamHI и PstI в буферном растворе для рестрикции (конечная концентрация 20 mМ Трис-HCl, pH 7,5, 50 mМ NaCl, 10 mM MgCl, 1 mМ дитиотреитола) в течение 5 часов при температуре 37oC. Ферменты инактивируют нагреванием при 70oC.
10 мкл раствора фрагментов ДНК, гидролизованных рестрикционными эндонуклезами, используют для анализа ДНК в 12% полиакриламидном геле (после радиоавтографии виден фрагмент размером около 92 п.н.).
ДНК осаждают этиловым спиртом и растворяют в 20 мкл дистиллированной воды. 10 мкл обработанного рестриктазами фрагмента ДНК вносят в эппендорфовскую пробирку, содержащую 7 мкл (0,1 мкг/мл) ДНК векторной плазмиды pUC18, обработанной рестриктазами BamHI и PstI, 2 мкл десятикратного буфера для лигирования, 1 мкл (5 единиц активности) ДНК-лигазы фага T4 и инкубируют при 4oC в течение 5 часов.
Полученной лигазной смесью трансформируют клетки E.coli штамм JM109 по обычной методике (Molecular cloning. New York. CSHL, 1982). Трансформированные клетки высевают на чашки с 1,2% LB-агаром, содержащим 10 мкг/мл ампициллина. Выросшие колонии проверяют на наличие рекомбинантных плазмид. Для этого бактериальные клетки лизируют и выделяют плазмидную ДНК, как описано (Nature, 1981, 145, 1365). Выделенную плазмидную ДНК дорабатывают рестриктазами BamHI и PstI и гидролизат исследуют электрофорезом в 12% полиакриламидном геле. Отбирают штамм, содержащий плазмиду, гидролизат которой имеет на электрофореграмме 1 полосы ДНК размерами 1686 и 92 п.н., что соответствует размерам вектора и синтезированного нами фрагмента ДНК.
Методом Сэнгера (PNAS, 1977, 74, 5463) определяют нуклеотидную последовательность клонированного фрагмента ДНК (см. в конце описания).
Пример 2.
Получение плазмиды pHЕ40.
Фрагмент ДНК HE40 лигируют с ДНК векторной плазмиды рЕL5А, обработанной рестриктазами BamHI и PstI.
Лигазной смесью трансформируют клетки E.coli штамм PLT90. Трансформированные клетки высевают на чашки с 1% LB-агаром. содержащие 50 мкг/мл ампициллина. Выросшие колонии проверяют на наличие рекомбинантных плазмид. Для этого бактериальные клетки лизируют и выделяют плазмидную ДНК. Выделенную плазмидную ДНК обрабатывают рестриктазами BamHI и PstI и гидролизат исследуют электрофорезом в 2% агарозе. Отбирают штамм SHE40. Гидролизат плазмидной ДНК из рекомбинантного штамма SHE40 имеет на электрофореграмме 2 полосы ДНК размерами 6400 и 92 нуклеотида, что соответствует размерам вектора и синтезированного фрагмента ДНК НЕ40.
При определении нуклеотидной последовательности плазмиды pHЕ40 установлено, что синтезированный фрагмент гена ORF2 вируса гепатита E через BamHI сайт присоединен к 3'-концу гена бета-галактозидазы E.coli. Таким образом плазмида pHЕ40 кодирует рекомбинантный белок, содержащий аминокислотные последовательности продукта гена ORF2 вируса гепатита E, слитые с последовательностью β-галактозидазы E.coli.
Пример 3.
Синтез полипептида НЕ40.
Штамм E. coli, содержащий плазмиду pHЕ40, выращивают в 100 мл среды LB при 30oC до плотности 8х10 кл./мл. После этого температуру повышают до 40oC и растят клетки еще в течение 2-х часов. Клетки собирают центрифугированием при 4000 об/мин в течение 15 мин. Клетки лизируют ультразвуком и выделяют белки по описанному методу (EMBO J., 1984, 3, 1429).
Выделенные белки анализируют в 10% ПААГ по стандартному методу Лэммли (Nature, 227, 680-685, 1970). В качестве маркеров молекулярных масс белков используют набор фирмы "Pharmacia", содержащий маркеры молекулярных масс от 20 до 160 кД. В качестве контроля используют полученный аналогичным образом лизат клеток штамма E.coli PLT90, содержащий векторную плазмиду рЕL5А и не содержащий плазмид pHЕ40. Сравнительный анализ белкового состава показывает, что штамм E.coli, содержащий плазмиду pHЕ40, экспрессирует полипептид с молекулярной массой - 122 кД. Ниже представлена аминокислотная последовательность полипептида, определенная на основании нуклеотидной последовательности клонированного фрагмента ДНК, соответствующая фрагменту продукта второй открытой рамки считывания с 633 по 659 аминокислотные осадки:
X - L G L Q G C A F Q S T V A E L Q R L K M K V G K T R E L Q P S L L I D,
где X - аминокислотная последовательность бета-галактозидазы E.coli.
Пример 4.
Получение фрагмента ДНК НЕ60.
Для получения фрагмента ДНК были использованы олигонуклеотиды d1, d2, r1, r2, приведенные в конце описания.
Олигонуклеотиды смешивают и кинируют с помощью полинуклеотидкиназы фага T4. Для этого в эппендорфовскую пробирку на 1,5 мл вносят по 50 пмолей каждого из олигонуклеотидов, 50 мкл десятикратного буфера для киназы (0,5 М Трис-HCl, pH 7,6, 0,1 М MgCl, 50 mM дитиотреитола, 1 mM спермидина и 1 mM ЭДТА), 250 пмолями (150 мкКИ) [гамма-P] АТФ (удельная активность 3000 Ки/моль), 150 единицами активности полинуклеотидкиназы фага T4 и бидистиллированной воды до объема 500 мкл и инкубируют 30-60 минут при температуре 37oC. Затем реакционную смесь нагревают до температуры 98oC, охлаждают до 16oC и смешивают с ДНК-лигазой. Для этого 100 мкл смеси обработанных киназой олигонуклеотидов смешивают с 12 мкл десятикратного буфера для лигирования (0,66 М Трис-HCl, pH 7,6, 50 mM MgCl, 50 mM дитиотреитола и 10 mM АТФ), 8 мкл (50 единиц активности) ДНК-лигазы фага T4 и инкубируют при 4oC в течение 5 часов. Затем реакционную смесь нагревают до температуры 65oC в течение 15 минут для инактивации фермента.
10 мкл раствора обработанных киназой и лигазой олигонуклеотидов используют для анализа ДНК методом электрофореза в 12% полиакриламидном геле по обычной методике (Molecular cloning, New York, CSHL. 1982). После радиоавтографии геля виден ряд фрагментов размером больше или равным 92 п.н.
100 мкл раствора полученного фрагмента ДНК гидролизуют рестриктазами BaniHI и PstI в буферном растворе для рестрикции (конечная концентрация 20 mM Трис-HCl, pH 7,5, 50 mM NaCl, 10 mM MgCl, 1 mM дитиотреитола) в течение 5 часов при температуре 37oC. Ферменты инактивируют нагреванием при 70oC.
10 мкл раствора фрагментов ДНК, гидролизованных рестрикционными эндонуклеазами, используют для анализа ДНК в 12% полиакриламидном геле (после радиоавтографии виден фрагмент размером около 92 п.н.).
ДНК осаждают этиловым спиртом и растворяют в 20 мкл дистиллированной воды. 10 мкл обработанного рестриктазами фрагмента ДНК вносят в эппендорфовскую пробирку, содержащую 7 мкл (0,1 мкг/мл) ДНК векторной плазмиды pUC18, обработанной рестриктазами BamHI и PstI, 2 мкл десятикратного буфера для лигирования, 1 мкл (5 единиц активности) ДНК-лигазы фага T4 и инкубируют при 4oC в течение 5 часов.
Полученной лигазной смесью трансформируют клетки E.coli штамм JM109 по обычной методике (Molecular cloning. New York. CSHL, 1982). Трансформированные клетки высевают на чашки с 1,2% LB-агаром, содержащим 10 мкг/мл ампициллина. Выросшие колонии проверяют на наличие рекомбинантных плазмид. Для этого бактериальные клетки лизируют и выделяют плазмидную ДНК, как описано (Nature, 1981, 145, 1365). Выделенную плазмидную ДНК обрабатывают рестриктазами BamHI и PstI и гидролизат исследуют электрофорезом в 12% полиакриламидном геле. Отбирают штамм, содержащий плазмиду, гидролизат которой имеет на электрофореграмме 1 полосы ДНК размерами 1686 и 107 п.н., что соответствует размерам вектора и синтезированного нами фрагмента ДНК.
Методом Сэнгера (PNAS, 1977. 74, 5463) определяют нуклеотидную последовательность клонированного фрагмента ДНК (см. в конце описания).
Пример 5.
Получение плазмиды pHЕ60.
Фрагмент ДНК HE60 лигируют с ДНК векторной плазмиды pEL5A, обработанной рестриктазами BamHI и PstI.
Лигазной смесью трансформируют клетки E.coli штамм PLT90. Трансформированные клетки высевают на чашки с 1% LB-агаром, содержащие 50 мкг/мл ампициллина. Выросшие колонии проверяют на наличие рекомбинантных плазмид. Для этого бактериальные клетки лизируют и выделяют плазмидную ДНК. Выделенную плазмидную ДНК обрабатывают рестриктазами BamHI и PstI и гидролизат исследуют электрофорезом в 2% агарозе. Отбирают штамм SHE60. Гидролизат плазмидной ДНК из рекомбинантного штамма SHE60 имеет на электрофореграмме 2 полосы ДНК размерами 6400 и 107 нуклеотидов, что соответствует размерам вектора и синтезированного фрагмента ДНК HE60.
При определении нуклеотидной последовательности плазмиды pHE60 установлено, что синтезированный фрагмент гена ORF3 вируса гепатита E через BamHI сайт присоединен к 3'-концу гена бета-галактозидазы E.coli. Таким образом плазмида pHЕ60 кодирует рекомбинантный белок, содержащий аминокислотные последовательности продукта гена ORF3 вируса гепатита E слитые с последовательностью β-галактозидазы E.coli.
Пример 6.
Синтез полипептида НЕ60.
Штамм E. coli, содержащий плазмиду pHЕ60, выращивают в 100 мл среды LB при 30oC до плотности 8х10 кл./мл. После этого температуру повышают до 40oC и растят клетки еще в течение 2-х часов. Клетки собирают центрифугированием при 4000 об/мин в течение 15 мин. Клетки лизируют ультразвуком и выделяют белки по описанному методу (EMBO J., 1984, 3, 1429).
Выделенные белки анализируют в 10% ПААГ по стандартному методу Лэммли (Nature, 227, 680-685, 1970). В качестве маркеров молекулярных масс белков используют набор фирмы "Pharmacia", содержащий маркеры молекулярных масс от 20 до 160 кД. В качестве контроля используют полученный аналогичным образом лизат клеток штамма E.coli PLT90, содержащий векторную плазмиду рЕL5A и не содержащий плазмид pHЕ60. Сравнительный анализ белкового состава показывает, что штамм E.coli, содержащий плазмиду pHЕ60, экспрессирует полипептид с молекулярной массой - 122 кД. Ниже представлена аминокислотная последовательность полипептида, определенная на основании нуклеотидной последовательности клонированного фрагмента ДНК, соответствующая фрагменту продукту третьей открытой рамки считывания с 92 по 123 аминокислотные остатки:
X - S N P P D H S A P L G V T R P S A P P L P H V V D L P Q L G P R R C S Q A C,
где X - аминокислотная последовательность бета-галактозидазы E.coli.
Пример 7.
Получение фрагмента ДНК НЕ70.
Для получения фрагмента ДНК были использованы олигонуклеотиды d1, d2, d3, d4, r1, r2, r3, r4, приведенные в конце описания.
Олигонуклеотиды d1, d2, r1, r2 смешивают и кинируют с помощью полинуклеотидкиназы фага T4. Для этого в эппендорфовскую пробирку на 1,5 мл вносят по 50 пмолей каждого из олигонуклеотидов, 50 мкл десятикратного буфера для киназы (0,5 М Трис-HCl, pH 7,6, 0,1 М MgCl, 50 mM дитиотреитола, 1 mM спермидина и 1 mM ЭДТА), 250 пмолями (150 мкКИ) [гамма-P] АТФ (удельная активность 3000 Ки/моль), 150 единицами активности полинуклеотидкиназы фага T4 и бидистиллированной воды до объема 500 мкл и инкубируют 30-60 минут при температуре 37oC. Затем реакционную смесь нагревают до температуры 98oC. охлаждают до 16oC и смешивают с ДНК-лигазой. Для этого 100 мкл смеси обработанных киназой олигонуклеотидов смешивают с 12 мкл десятикратного буфера для лигирования (0,66 М Трис-HCl, pH 7,6, 50 mM MgCl, 50 mM дитиотреитола и 10 mM АТФ), 8 мкл (50 единиц активности) ДНК-лигазы фага T4 и инкубируют при 4oC в течение 5 часов. Затем реакционную смесь нагревают до температуры 65oC в течение 15 минут для инактивации фермента.
Олигонуклеотиды d3, d4, r3, r4 смешивают и кинируют с помощью полинуклеотидкиназы фага T4. Для этого в эппендорфовскую пробирку на 1,5 мл вносят по 50 пмолей каждого из олигонуклеотидов, 50 мкл десятикратного буфера для киназы (0,5 М Трис-HCl, pH 7,6, 0,1 М MgCl, 50 mM дитиотреитола, 1 mM спермидина и 1 mM ЭДТА), 150 единицами активности полинуклеотидкиназы фага T4 и бидистиллированной воды до объема 500 мкл и инкубируют 30-60 минут при температуре 37oC. Затем реакционную смесь нагревают до температуры 98oC, охлаждают до 16oC и смешивают с ДНК-лигазой. Для этого 100 мкл смеси обработанных киназой олигонуклеотидов смешивают с 12 мкл десятикратного буфера для лигирования (0,66 М Трис-HCl, pH 7,6, 50 mM MgCl, 50 mM дитиотреитола и 10 mM АТФ), 8 мкл (50 единиц активности) ДНК-лигазы фага T4 и инкубируют при 4oC в течение 5 часов. Затем реакционную смесь нагревают до температуры 65oC в течение 15 минут для инактивации фермента.
Полученные фрагменты ДНК смешивают и снова подвергают обработке полинуклеотидкиназой фага T4. Для этого в эппендорфовскую пробирку на 1,5 мл вносят по 50 пмолей каждого из фрагментов ДНК, 50 мкл десятикратного буфера для киназы (0,5 М Трис-HCl, pH 7,6, 0,1 М MgCl, 50 mM дитиотреитола, 1 mM спермидина и 1 mM ЭДТА), 150 единицами активности полинуклеотидкиназы фага T4 и бидистиллированной воды до объема 500 мкл и инкубируют 30-60 минут при температуре 37oC. Затем реакционную смесь нагревают до температуры 98oC, охлаждают до 16oC и смешивают с ДНК-лигазой. Для этого 100 мкл смеси обработанных киназой олигонуклеотидов смешивают с 12 мкл десятикратного буфера для лигирования (0,66 М Трис-HCl, pH 7,6, 50 mM MgCl, 50 mM дитиотреитола и 10 mM АТФ), 8 мкл (50 единиц активности) ДНК-лигазы фага T4 и инкубируют при 4oC в течение 5 часов. Затем реакционную смесь нагревают до температуры 65oC в течение 15 минут для инактивации фермента.
10 мкл раствора обработанных киназой и лигазой олигонуклеотидов используют для анализа ДНК методом электрофореза в 12% полиакриламидном геле по обычной методике (Molecular cloning, New York, CSHL, 1982). После радиоавтографии геля виден ряд фрагментов размером больше или равным 182 п.н.
100 мкл раствора полученного фрагмента ДНК гидролизуют рестриктазами BamHI и PstI в буферном растворе для рестрикции (конечная концентрация 20 mМ Трис-HCl, pH 7,5, 50 mM NaCl, 10mM MgCl, 1 mM дитиотреитола) в течение 5 часов при температуре 37oC. Ферменты инактивируют нагреванием при 70oC.
10 мкл раствора фрагментов ДНК, гидролизованных рестрикционными эндонуклеазами, используют для анализа ДНК в 12% полиакриламидном геле (после радиоавтографии виден фрагмент размером около 92 п.н.).
ДНК осаждают этиловым спиртом и растворяют в 20 мкл дистиллированной воды. 10 мкл обработанного рестриктазами фрагмента ДНК вносят в эппендорфовскую пробирку, содержащую 7 мкл (0,1 мкг/мл) ДНК векторной плазмиды pUC18, обработанной рестриктазами BamHI и PstI, 2 мкл десятикратного буфера для лигирования, 1 мкл (5 единиц активности) ДНК-лигазы. Олигонуклеотиды d3, d4, r3, r4 смешивают и кинируют с помощью полинуклеотидкиназы фага T4. Для этого в эппендорфовскую пробирку на 1,5 мл вносят по 50 пмолей каждого из олигонуклеотидов, 50 мкл десятикратного буфера для киназы (0,5 М Трис-HCl, pH 7,6, 0,1 M MgCl, 50 mM дитиотреитола, 1 mM спермидина и 1 mM ЭДТА), 250 пмолями (150 мкКИ) [гамма-P] АТФ (удельная активность 3000 Ки/моль), 150 единицами активности полинуклеотидкиназы фага Т4 и бидистиллированной воды до объема 500 мкл и инкубируют 30-60 минут при температуре 37oC. Затем реакционную смесь нагревают до температуры 98oC, охлаждают до 16oC и смешивают с ДНК-лигазой. Для этого 100 мкл смеси обработанных киназой олигонуклеотидов смешивают с 12 мкл десятикратного буфера для лигирования (0,66 М Трис-HCl, pH 7,6, 50 mM MgCl, 50 mM дитиотреитола и 10 mM АТФ), 8 мкл (50 единиц активности) ДНК-лигазы фага T4 и инкубируют при 4oC в течение 5 часов. Затем реакционную смесь нагревают до температуры 65oC в течение 15 минут для инактивации фермента. Полученной лигазной смесью трансформируют клетки E.coli штамм JM109 по обычной методике (Molecular cloning, New York. CSHL, 1982). Трансформированные клетки высевают на чашки с 1,2% LB-агаром, содержащим 10 мкг/мл ампициллина. Выросшие колонии проверяют на наличие рекомбинантных плазмид. Для этого бактериальные клетки лизируют и выделяют плазмидную ДНК, как описано (Nature, 1981, 145, 1365). Выделенную плазмидную ДНК обрабатывают рестриктазами BamHI и PstI и гидролизат исследуют электрофорезом в 12% полиакриламидном геле. Отбирают штамм, содержащий плазмиду, гидролизат которой имеет на электорофореграмме две полосы ДНК размерами 1686 и 182 п. н. , что соответствует размерам вектора и синтезированного нами фрагмента ДНК.
Методом Сэнгера (PNAS, 1977, 74, 5463) определяют нуклеотидную последовательность клонированного фрагмента ДНК (см. в конце описания).
Пример 8.
Получение плазмиды pHЕ70.
Фрагмент ДНК НЕ70 лигируют с ДНК векторной плазмиды pEL5A, обработанной рестриктазами BamHI и PstI.
Лигазной смесью трансформируют клетки E.coli штамм PLT90. Трансформированные клетки высевают на чашки с 1% LB-агаром, содержащие 50 мкг/мл ампициллина. Выросшие колонии проверяют на наличие рекомбинантных плазмид. Для этого бактериальные клетки лизируют и выделяют плазмидную ДНК. Выделенную плазмидную ДНК обрабатывают рестриктазами BamHI и PstI и гидролизат исследуют электрофорезом в 2% агарозе. Отбирают штамм SHE70. Гидролизат плазмидной ДНК из рекомбинантного штамма SHE70 имеет на электрофореграмме 2 полосы ДНК размерами 6400 и 182 нуклеотида, что соответствует размерам вектора и синтезированного фрагмента ДНК НЕ70.
При определении нуклеотидной последовательности плазмиды pHЕ70 установлено, что синтезированный фрагмент гена ORF2 вируса гепатита E через BamHI сайт присоединен к 3'-концу гена β-галактозидазы E.coli. Таким образом плазмида pHЕ70 кодирует рекомбинантный белок, содержащий аминокислотные последовательности продукта гена ORF2 вируса гепатита E, слитые с последовательностью β-галактозидазы E.coli.
Пример 9.
Синтез полипептида НЕ70.
Штамм E. coli. содержащий плазмиду pHЕ70, выращивают в 100 мл среды LB при 30oC до плотности 8х10 кл/мл. После этого температуру повышают до 40oC и растят клетки еще в течение 2-х часов. Клетки собирают центрифугированием при 4000 об/мин в течение 15 мин. Клетки лизируют ультразвуком и выделяют белки по описанному методу (EMBO J. 1984, 3, 1429).
Выделенные белки анализируют в 10% ПААГ по стандартному методу Лэммли (Nature, 227, 680-685, 1970). В качестве маркеров молекулярных масс белков используют набор фирмы "Pharmacia", содержащий маркеры молекулярных масс от 20 до 160 кД. В качестве контроля используют полученный аналогичным образом лизат клеток штамма E.coli PLT90, содержащий векторную плазмиду pEL5A и не содержащий плазмид pHЕ70. Сравнительный анализ белкового состава показывает, что штамм E.coli, содержащий плазмиду pHЕ70, экспрессирует полипептид с молекулярной массой - 124 кД. Ниже представлена аминокислотная последовательность полипептида, определенная на основании нуклеотидной последовательности клонированного фрагмента ДНК, соответствующая фрагменту продукта второй открытой рамки считывания с 403 по 461 аминокислотные остатки:
X - A N G E P T V K L Y T S V E N A Q Q D K G I A I P H D I D L G E S R V V I Q D Y D N Q H E Q D R P T P S P A P S R P,
где X - аминокислотная последовательность бета-галактозидазы E.coli.
Пример 10.
Контроль антигенной активности рекомбинантных полипептидов НЕ40, НЕ60, НЕ70.
Контроль антигенной активности полипептидов НЕ40, НЕ60, НЕ70 осуществляют методом иммуноблотинга. Для этого проводят процедуру электрофореза (как описано в примере 3), затем осуществляют перенос электрофоретически разделенных белков на нитроцеллюлозные мембраны ВН85 с помощью аппарата для электроблотинга в режиме 24В, 400 мА в течение 1 часа. Качество переноса проверяют окрашиванием мембран раствором 0,2% Понсо S в 3%-ной ТХУ в течение 5 мин при комнатной температуре. Краску дважды отмывают в течение 30 мин буфером TBS (0,15 М NaCl, 20 mM Трис-HCl, pH 7,4) с 0,05% Твина-20 при комнатной температуре. На следующем этапе проводят блокирование нитроцеллюлозных мембран с иммобилизованными белками раствором 5%-ного обезжиренного сухого молока в 0,1 М Na-фосфатном буфере, pH 7,4, в течение 30 минут при комнатной температуре. После блокирования мембраны в течение 1 ч при 37oC обрабатывают сыворотками, содержащими антитела к вирусу гепатита E, в разведении 1:100. Затем проводят трехкратную отмывку мембран TBS с 0,05%-ным Твином-20 в течение 30 мин при комнатной температуре. Выявление специфических антител осуществляют инкубацией фильтра с антивидовым пероксидазным конъюгатом при 37oC в течение 1 часа. Далее повторяют процедуру трехкратной отмывки. Реакцию проверяют добавлением 100 мкл 30%-ной перекиси водорода и 50 мл раствора диамнобензидина в 50 mM Трис-HCl, pH 7,5. Анализ показывает, что полученные полипептиды связываются с антителами к вирусу гепатита E.
Пример 11.
Набор для массового серологического исследования образцов сыворотки или плазмы крови на присутствие антител к рекомбинантным антигенам, содержащим фрагменты белков - продуктов второй и третьей открытых рамок считывания вируса гепатита Е.
Штаммы бактерий E.coli - продуценты полипептидов:
НЕ70 - продукта второй открытой рамки считывания с аминокислотной последовательностью
A N G E P T V K L Y T S V E N A Q Q D K G I A I P H D I D L G E S R V V I Q D Y D N Q H E Q D R P T P S P A P S R P
HE40 - продукта второй открытой рамки считывания с аминокислотной последовательностью
L G L Q G C A F Q S T V A E L Q R L K M K V G K T R E L Q P S L L I D
НЕ60 - продукта третьей открытой рамки считывания с аминокислотной последовательностью
S N P P D H S A P L G V T R P S A P P L P H V V D L P Q L G P R R C S Q A C
выращивают на среде LB с селективными добавками, клетки собирают центрифугированием, разрушают ультразвуком. Тельца включения, содержащие рекомбинантные полипептиды, промывают с помощью центрифугирования, солюбилизируют. Полипептиды очищают с помощью хроматографических методов, чистоту полученных полипептидов анализируют с помощью ДДС-Na-электрофореза. При изготовлении иммуносорбента для наборов сорбируют смесь полипептидов - продуктов второй открытой рамки считывания (концентрация каждого полипептида - 2,5 мкг/мл), и третьей открытой рамки считывания (5 мкг/мл) растворенных в 0,1 М фосфатно-солевом буфере pH 7,6. Сорбцию ведут при 4oC в течение 18 часов.
Конъюгат пероксидазы хрена с белком A St.aureus или с антивидовыми антителами получают с помощью периодатного окисления с последующей хроматографической очисткой готового конъюгата. Конъюгат, контрольные сыворотки и буферную смесь с гидроперитом (БСГ) для пероксидазной реакции лиофилизируют в ампулах, запаивают в атмосфере инертного газа. В качестве контрольного положительного иммунореагента используют сыворотку крови человека, содержащую антитела к вирусу гепатита E (К+). В качестве контрольного отрицательного иммунореагента (К-) используют сыворотку крови человека, не содержащую антитела к вирусу гепатита Е.
Иммуносорбент представляет собой полистироловые или полихлорвиниловые планшеты для иммунологических реакций с иммобилизованной смесью рекомбинантных антигенов. Иммуносорбент перед использованием трехкратно промывают. Для промывок используют фосфатный буферный раствор pH 7,2 с 0,5% Твином-20 (ФСБ-Т) с помощью автоматического или ручного промывателя, внося не менее 0,2 мл раствора в лунку. По окончании промывки удаляют влагу из планшета.
В 88 из 96 лунок вносят по 90 мкл раствора для разведения сывороток, содержащего 4% раствор клеток E.coli, лизированных ультразвуком, в ФСБ-Т и добавляют по 10 мкл предварительно разведенных (1:10) исследуемых сывороток, перемешивая содержимое лунок пипетированием или с помощью встряхивателя. В незаполненном ряду, например в 1-ом, в две лунки вносят 100 мкл К-, в 4 лунки - по 100 мкл К+ и в 2 лунки по 100 мкл раствора для разведения сывороток (контроль конъюгата). Планшет заклеивают пленкой и выдерживают в течение 60 минут при температуре 37oС.
Содержимое лунок удаляют, планшет 5-кратно промывают и полностью удаляют влагу. Во все лунки планшета вносят по 100 мкл конъюгата. Планшет заклеивают новым листом пленки и выдерживают в течение 30 минут при температуре 37oС.
Содержимое лунок удаляют, планшет 5-кратно промывают и полностью удаляют влагу. В каждую лунку планшета вносят по 100 мкл 0,05% ортофенилендиамина в растворе БСГ. Планшет помещают в защищенное от света место на 15-20 минут при температуре 15-25oС. Реакцию останавливают внесением во все лунки по 50 мкл 2N серной кислоты.
Регистрацию проводят немедленно на спектрофотометре типа "Мультискан", измеряя оптическую плотность (ОП) при длине волны 492 нм. Выведение спектрофотометра на нулевой уровень осуществляют по воздуху или по чистому сухому планшету.
Вычисляют среднее значение ОП (ОПср) для контроля конъюгата, которое должно быть не более 0,15; ОПср К- - не более 0,2; ОПср К+ - в пределах от 0,8 до 1,8. Пороговое значение ОП вычисляют по формуле: ОПср К+, умноженное на расчетный коэффициент, указываемый в паспорте набора. Сыворотку учитывают как положительную, если значение ОП образца выше порогового. Чувствительность препарата не менее 99%, специфичность не менее 98%.
Пример 12.
Набор для повторного (уточняющего и дифференцирующего) серологического исследования образцов сыворотки или плазмы крови на присутствие антител к каждому из соответствующих антигенов вируса гепатита E.
Штаммы бактерий E. coli - продуценты полипептидов продуктов второй и третьей открытых рамок считывания вируса гепатита E, а также продуцент β-галактозидазы выращивают на среде LB с селективными добавками. Клетки собирают центрифугированием, разрушают ультразвуком. Тельца включения, содержащие рекомбинантные полипептиды, промывают с помощью центрифугирования, солюбилизируют. Полипептиды очищают хроматографическими методами, чистоту полученных полипептидов анализируют с помощью ДДС-Na-электрофореза. При изготовлении иммуносорбента для наборов готовят растворы вышеперечисленных полипептидов и "KАг" (контрольный антиген β-галактозидаза), каждый в виде отдельного раствора в 0,1 М фосфатно-солевом буфере, pH 7,6, в концентрации соответственно 5 мкг/мл. Сорбцию ведут при 4oC в течение 18 часов.
Конъюгат пероксидазы хрена с белком A St.aureus или с антивидовыми антителами получают при помощи периодатного окисления с последующей хроматографической очисткой готового конъюгата. Конъюгат, контрольные сыворотки и буферную смесь с гидроперитом (БСГ) для пероксидазной реакции лиофилизируют в ампулах, запаивают в атмосфере инертного газа.
В качестве контрольного положительного иммунореагента используют сыворотку крови человека, содержащую антитела к вирусу гепатита E (K1+). В качестве контрольного отрицательного иммунореагента (К-) используют сыворотку крови человека, не содержащую антител к вирусу гепатита Е.
Иммуносорбент представляет собой полистироловые или полихлорвиниловые планшеты для иммунологических реакций с рекомбинантными антигенами - продуктами второй и третьей открытых рамок считывания генома вируса гепатита E, и контрольным антигеном, иммобилизованными по отдельности. Лунки планшета, в которых иммобилизованы антигены, называют рабочими.
Иммуносорбент перед использованием трехкратно промывают раствором ФСБ-Т, удаляют влагу из планшета. В 88 из 96 лунок вносят по 90 мкл раствора для разведения сывороток, оставляя, как указано в табл. 1, пустыми лунки для К- и К+.
Табл. 1 показывает расположение сорбированных рекомбинантных антигенов и KАг в рядах планшета a, b, c, d соответственно.
Добавляют по 10 мкл предварительно разведенных (1:10) исследуемых сывороток в каждую из 4-х рабочих лунок горизонтального ряда, перемешивая содержимое лунок пипетированием или с помощью встряхивателя. В незаполненные ряды в соответствии с табл. 1 вносят К- и К+ по 100 мкл в каждую из восьми лунок. Планшет заклеивают пленкой и выдерживают в течение 60 минут при температуре 37oC.
Содержимое лунок удаляют с помощью промывателя, затем планшет 5-кратно промывают и полностью удаляют влагу. Во все рабочие лунки планшета вносят по 100 мкл конъюгата. Планшет заклеивают новым листом пленки и выдерживают в течение 30 минут при температуре 37oC.
Содержимое лунок удаляют, планшет 5-кратно промывают и полностью удаляют влагу. В каждую рабочую лунку планшета вносят по 100 мкл 0,05% ортофенилендиамина в растворе БСГ. Планшет помещают в защищенное от света места на 15-20 минут при температуре 15-25oC. Реакцию останавливают внесением во все лунки по 50 мкл 2N серной кислоты.
Регистрацию проводят немедленно на спектрофотометре типа "Мультискан", измеряя оптическую плотность (ОП) при длине волны 492 нм. Выведение спектрофотометра на нулевой уровень осуществляют по воздуху или по чистому сухому планшету.
Вычисляют среднее по 5-ти лункам значение ОП для К-, которое должно быть не более 0,2. Значение ОП К+ по лункам с контрольным антигеном (11Н) должно быть не более 0,2.
Для каждой сыворотки вычисляют отношения ОП по каждому рекомбинантному антигену к ОП по контрольному антигену. Сигнал по антигену учитывают как положительный, если соответствующее отношение превышает коэффициент R, указанный в паспорте. Сигнал по антигену учитывают как отрицательный, если соответствующее отношение меньше коэффициента N, указанного в паспорте. Промежуточный вариант учитывают как сомнительный.
Чувствительность препарата не менее 99%, специфичность не менее 98%. В качестве твердого носителя для нанесения рекомбинантных антигенов могут быть использованы не только планшеты для иммунологических реакций, но и любая другая твердая фаза. Например, полистироловые шарики, нитроцеллюлоза, латекс, эритроциты и др. В этих случаях рекомбинантные антигены выделяют тем же методом, что и в примерах 1 и 2, используют те же либо другие иммунохимические реакции (например, агглютинациии) и результаты учитывают как спектрофотометрически, так и визуально либо при помощи других приборов.
Пример 13.
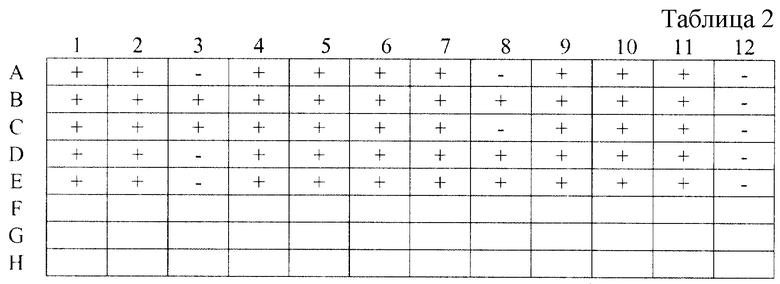
Рекомбинантные белки НЕ40, НЕ60, НЕ70 и β-галактозидазу очищают, как описано в примере 1. Сорбцию антигенов на планшет ведут при 4oC, как описано в примерах 11 и 12. Ряд А планшета сорбируют только полипептидом НЕ40; ряд В сорбируют смесью полипептидов НЕ40, НЕ60 и НЕ70; ряд С смесью полипептидов НЕ40 и НЕ60; ряд D смесью полипептидов НЕ40 и НЕ70; ряд E смесью полипептидов НЕ60 и НЕ70.
В лунки с номерами 1-11 рядов A-E планшета добавляют 11 различных сывороток больных гепатитом E, а в лунки 12 отрицательную сыворотку.
Постановку иммунологических реакций и обработку полученных данных осуществляют как описано в примере 12. Результаты экспериментов представлены в табл. 2. Знаком (+) отмечены лунки, в которых исследуемые сыворотки положительно прореагировали с иммуносорбентом, знаком (-) - лунки с отрицательно прореагировавшими сыворотками.
Как видно из табл. 2, только сорбция всех четырех антигенов (ряд В планшета) приводит к идентификации всех положительных сывороток (чувствительность - 100%). В других вариантах постановки чувствительность тест-системы колеблется от 82 до 90%.
Таким образом, данное изобретение позволяет повысить чувствительность и специфичность набора за счет определения антител к большему числу вирусных белков и спектра антител к индивидуальным вирусным белкам.







Предложен способ получения рекомбинантных полипептидов для использования в наборе для определения антител к вирусу гепатита Е. Набор включает ряд рекомбинантных антигенов, синтезированных в клетках E.coli, трансформированных рекомбинантными ДНК. Рекомбинантные ДНК включают фрагменты второй и третьей открытых рамок считывания генома вируса гепатита Е. В качестве контрольного антигена наборы могут содержать β-галактозидазу E.coli. Набор позволяет повысить чувствительность и специфичность при определении спектра антител к индивидуальным вирусным белкам. 2 с. и 3 з.п. ф-лы, 2 табл.
| RU 93051201 A, 10.01.1997 | |||
| EP 0419182, 27.03.1991. |
