Область, к которой относится настоящее изобретение
Настоящее изобретение относится к моноклональному антителу против человеческого антигена CD20. Настоящее изобретение также относится к химерному моноклональному анти-CD20-антителу и к гуманизованному моноклональному анти-CD20-антителу, продуцируемому посредством рекомбинации генов, а также к терапевтическому средству для лечения В-клеточно-опосредованной опухоли или иммунологического заболевания, содержащему указанные антитела в качестве активного ингредиента.
Предшествующий уровень техники
Известными моноклональными антителами, которые распознают антиген CD20, являются антитела B1, 2B8 (химерное антитело, называемое ритуксимабом), 1F5, 2H7 и т.п. Сначала ритуксимаб, то есть химерное моноклональное анти-CD20-антитело, разработанное Фармацевтической корпорацией IDEC (Pharmaceuticals Corporation, U.S.), было создано как стандартное терапевтическое средство для лечения низкозлокачественной не-ходжкинской лимфомы (НХЛ), но затем было обнаружено, что оно обладает терапевтическим действием против многих В-клеточно-опосредуемых иммунных заболеваний. Так, например, считается, что это средство является эффективным для лечения не только злокачественных опухолей, таких как хронический лимфолейкоз, но и для лечения аутоиммунных заболеваний, при которых продуцируются патогенные аутоантитела, таких как аутоиммунная гемолитическая анемия и идиопатическая тромбоцитопеническая пурпура, а также воспалительных заболеваний, таких как хронический ревматоидный артрит и рассеянный склероз (не-патентные документы 14-17).
CD20 представляет собой молекулу, присутствующую на поверхности В-лимфоцитов, и ее экспрессия наблюдается в нормальных В-клетках периферической крови, селезенки, миндалин и костного мозга и т.п., а также в В-клетках большинства злокачественных опухолей. Эта молекула насчитывает 297 аминокислотных остатков, “прошивает" клеточную мембрану четыре раза; причем С- и N-концы этой молекулы расположены внутри клетки; а ее внеклеточная область представляет собой петлю, не содержащую сахарную цепь и состоящую из 43 аминокислотных остатков, расположенных между третьим и четвертым трансмембранными доменами (не-патентные документы 1 и 9). Предполагается, что молекула CD20 существует в виде тетрамера и, кроме того, образует гетерокомплекс с другими небольшими компонентами (не-патентный документ 18). Поскольку белок CD20 не секретируется за пределы клетки и не расщепляется и, кроме того, он почти не поглощается клеткой при связывании с антителом, то можно предположить, что цитотоксический механизм, опосредуемый антителом против клеток-мишеней, является эффективным (не-патентные документы 1-3).
Молекула CD20 несмотря на свой небольшой размер имеет различные эпитопы, что частично обусловлено характером ее экспрессии как комплекса, находящегося за пределами клетки, и антитела, связывающиеся с этой молекулой, опосредуют различные биологические ответы. Так, например, активности, такие как ингибирование В-клеточных рецепторов, увеличение уровня экспрессии антигенов MHC класса II и адгезивных молекул, активация высвобождения Ca2+ при избыточном перекрестном связывании; ингибирование гомотипической адгезии, не зависимой от функционально связанного антигена 1 лимфоцитов; индуцирование апоптоза и противоположная активность и стимуляция клеточного роста, значительно варьируются (не-патентные документы 4-13). Типичные анти-CD20-антитела, например ритуксимаб, B1, 1F5 и 2H7, также обладают различными свойствами и биологическими функциями, и термин “моноклональное антитело, связывающееся с CD20” не может конкретно определять его биологические свойства.
Молекула, которая составляет внеклеточный домен CD20, является нерастворимой. Хотя молекула CD20, происходящая от клеточного лизата или белка, продуцируемого рекомбинантным геном, может быть солюбилизирована с использованием поверхностно-активного вещества или сильного основания, однако сохранение природной трехмерной структуры в таких условиях обработки представляет определенные трудности. Поэтому CD20-позитивный В-клеточный штамм используется в качестве иммуногена для получения антител. Однако такой иммуноген имеет слабые иммуностимулирующие свойства, а поэтому получение клонов зрелых антителопродуцирующих клеток является нелегкой задачей.
К 2005 году ритуксимаб, то есть химерное антитело мышь/человек, было единственным моноклональным анти-CD20-антителом, разрешенным к применению в качестве терапевтического средства. Поскольку химерные антитела, содержащие гетерологичные молекулы, обладают антигенностью, то обычно они не являются предпочтительными в качестве терапевтических средств. Однако анти-CD20-антитела обладают способностью атаковать и элиминировать все В-клетки, включая нормальные клетки, а поэтому считается, что они, по существу, не обладают антигенностью. Однако были описаны примеры, в которых нейтрализующее антитело индуцируется во время его введения, хотя его уровень составляет лишь несколько процентов, и, по всей вероятности, оно индуцируется в зависимости от дозы и от периода введения дозы. Поэтому желательно получить такое гуманизованное антитело, которое имело бы последовательность, более сходную с последовательностью человеческого антитела, или последовательность человеческого антитела. Другим недостатком химерных антител является их короткое время полужизни в крови, и это время полужизни составляет всего 3 или 4 дня. Клинические исследования, проводимые в США, показали, что эффективный уровень одного ритуксимаба, вводимого для предотвращения рецидивов низкозлокачественной НХЛ, составляет немного ниже 50%, что указывает на то, что у 50% или более пациентов не наблюдается ответ на ритуксимаб либо такой ответ является недостаточным. Уровень ответа у пациентов с умеренно злокачественной НХЛ еще ниже и составляет примерно лишь 30% (не-патентный документ 14). Поэтому необходимо провести исследование по выявлению факторов и причины различных ответов у пациентов и разработать антитело, обладающее более высокой эффективностью.
Не-патентный документ 1: Leukocyte Fact Book 2nd Edition, Academic Press
Не-патентный документ 2: Stashenko P et al., J. Immunol., 1980, 125: 1678-85
Не-патентный документ 3: Anderson KC et al., Blood, 1984, 63: 1424-33
Не-патентный документ 4: Shan D et al., Blood, 1998, 91: 1644-52
Не-патентный документ 5: Flieger D et al., Cell Immunol., 2000, 204: 55-63
Не-патентный документ 6: Mathas S et al., Cancer Res., 2000, 60: 7170-6
Не-патентный документ 7: Cardarelli PM et al., Cancer Immunol. Immunother., 2002, 51: 15-24
Не-патентный документ 8: Pedersen IM et al., Blood, 2002, 99: 1314-9
Не-патентный документ 9: Deans JP et al., Imminol., 2002, 107: 176-82
Не-патентный документ 10: Golay JT et al., J. Immunol., 1992, 149: 300-8
Не-патентный документ 11: Bourger I et al., Eur. J. Immunol., 1993, 23: 768-71
Не-патентный документ 12: White MW et al., J. Immunol., 1991, 146: 846-53
Не-патентный документ 13: Shan D et al., Cancer Immunol. Immnother., 2000, 48: 673-83
Не-патентный документ 14: Coiffier B et al., Blood, 1998, 92: 1927-32
Не-патентный документ 15: Edward JC et al., Rheumatology (Oxford), 2001, 40: 205-11
Не-патентный документ 16: Zaja F et al., Heamatologica, 2002, 87: 189-95
Не-патентный документ 17: Perrotta S et al., Br. J. Haematol., 2002, 116: 465-7
Не-патентный документ 18: Polyak MJ et al., Blood, 2002, 99: 3256-62
Описание сущности изобретения
Целью настоящего изобретения является получение моноклонального антитела, имеющего лучшие биологические функции, чем стандартные терапевтические средства, полученные на основе моноклональных анти-CD20-антител.
Авторами настоящего изобретения с использованием двух или более CD20-антиген-позитивных В-клеточных штаммов были получены мышиные моноклональные анти-CD20-антитела, которые специфически связываются с человеческим антигеном CD20, и методами биотехнологии были сконструированы клетки млекопитающих для экспрессии человеческого антигена CD20 на клеточных мембранах, а также в качестве иммуногена был получен гибрид человеческого белка CD20 с глутатион-S-трансферазой (GST) в произвольной комбинации. Некоторые из этих антител обладают активностью, направленной на ингибирование клеточного роста, включая апоптоз CD20-экспрессирующей клеточной культуры in vitro, в отсутствие эффекторных клеток. Кроме того, независимо от присутствия или отсутствия активностей, стимулирующих ингибирование роста клеток, например апоптоз, на эффективность этих антител, включая другие отобранные мышиные моноклональные анти-CD20-антитела, влияет комплемент- или антителозависимая клеточно-опосредуемая цитотоксичность, вызываемая химеризацией. В результате гуманизации аминокислотных последовательностей антител, которые, как было определено, обладают наиболее желательными биологическими активностями, были получены моноклональные анти-CD20-антитела, которые могут быть использованы в качестве терапевтического средства. И получение таких антител было положено в основу настоящего изобретения.
В настоящем изобретении рассматриваются:
(1) Мышиное моноклональное анти-CD20-антитело, обладающее активностью, направленной на ингибирование роста клеток, включая апоптоз клеток, экспрессирующих человеческий антиген CD20, в культуре клеток, экспрессирующих антиген CD20 в отсутствие эффекторных клеток.
(2) Моноклональное анти-CD20-антитело по п.(1), где аминокислотные последовательности вариабельной области Н-цепи и вариабельной области L-цепи представляют собой последовательности SEQ ID NO: 1 и 7, SEQ ID NO: 2 и 8 или SEQ ID NO: 15 и 17.
(3) Гибридома, продуцирующая моноклональное анти-CD20-антитело по п.(1) или (2).
(4) Химерное моноклональное анти-CD20-антитело, где аминокислотная последовательность вариабельной области моноклонального анти-CD20-антитела по п.(2) и аминокислотная последовательность константной области человеческого иммуноглобулина присоединены друг к другу.
(5) Моноклональное анти-CD20-антитело, гуманизованное с использованием аминокислотной последовательности гипервариабельной области (комплементарность-определяющей области, CDR) вариабельной области моноклонального анти-CD20-антитела по п.(2) и аминокислотной последовательности человеческого иммуноглобулина.
(6) Гуманизованное моноклональное анти-CD20-антитело по п.(5), где комбинация аминокислотных последовательностей вариабельной области Н-цепи и вариабельной области L-цепи представляет собой комбинацию последовательностей SEQ ID NO: 19 и 23, SEQ ID NO: 19 и 24, SEQ ID NO: 19 и 25, SEQ ID NO: 19 и 26, SEQ ID NO: 20 и 23, SEQ ID NO: 20 и 24, SEQ ID NO: 20 и 25, SEQ ID NO: 20 и 26, SEQ ID NO: 21 и 23, SEQ ID NO: 21 и 24, SEQ ID NO: 21 и 25, SEQ ID NO: 21 и 26, SEQ ID NO: 22 и 23, SEQ ID NO: 22 и 24, SEQ ID NO: 22 и 25 или SEQ ID NO: 22 и 26.
(7) Моноклональное анти-CD20-антитело по любому из пп.(4)-(6), которое оказывает цитотоксическое действие на клетки, экспрессирующие антиген CD20, в присутствии человеческого комплемента.
(8) Клетка млекопитающего, в которую была введена нуклеотидная последовательность, кодирующая аминокислотную последовательность моноклонального анти-CD20-антитела по любому из пп.(4)-(7).
(9) Клетка млекопитающего по п.(8), которой является клетка яичника китайского хомячка (СНО).
(10) Мышиное моноклональное анти-CD20-антитело, где комбинация аминокислотных последовательностей вариабельной области Н-цепи и вариабельной области L-цепи представляет собой комбинацию последовательностей SEQ ID NO:3 и 9, SEQ ID NO:4 и 10, SEQ ID NO:5 и 11, SEQ ID NO:6 и 12 или SEQ ID NO:16 и 18.
(11) Гибридома, продуцирующая моноклональное анти-CD20-антитело по п.(10).
(12) Химерное моноклональное анти-CD20-антитело, где аминокислотная последовательность вариабельной области моноклонального анти-CD20-антитела по п.(10) и аминокислотная последовательность константной области человеческого иммуноглобулина присоединены друг к другу.
(13) Моноклональное анти-CD20-антитело, гуманизованное с использованием аминокислотной последовательности CDR вариабельной области моноклонального анти-CD20-антитела по п.(10) и аминокислотной последовательности человеческого иммуноглобулина.
(14) Моноклональное анти-CD20-антитело по п.(12) или (13), которое оказывает цитотоксическое действие на клетки, экспрессирующие антиген CD20, в присутствии человеческого комплемента.
(15) Клетка млекопитающего, в которую была введена нуклеотидная последовательность, кодирующая аминокислотную последовательность моноклонального анти-CD20-антитела по любому из пп.(12)-(14).
(16) Клетка млекопитающего по п.(15), которой является клетка СНО.
(17) Диагностическое средство, содержащее моноклональное анти-CD20-антитело по любому из пп.(2), (4)-(7), (10) и (12)-(14) в качестве активного ингредиента.
(18) Терапевтическое средство, содержащее моноклональное анти-CD20-антитело по любому из пп.(4)-(7) и (12)-(14) в качестве активного ингредиента.
Аминокислотные остатки в аминокислотных последовательностях моноклональных антител, определенных выше, могут быть заменены другими аминокислотными остатками при условии, что такие замены не будут оказывать значительного влияния на вторичные структуры и биологические свойства указанных антител, и такие моноклональные антитела, имеющие модифицированные аминокислотные последовательности, упомянутые выше, также входят в объем настоящего изобретения.
Краткое описание графического материала
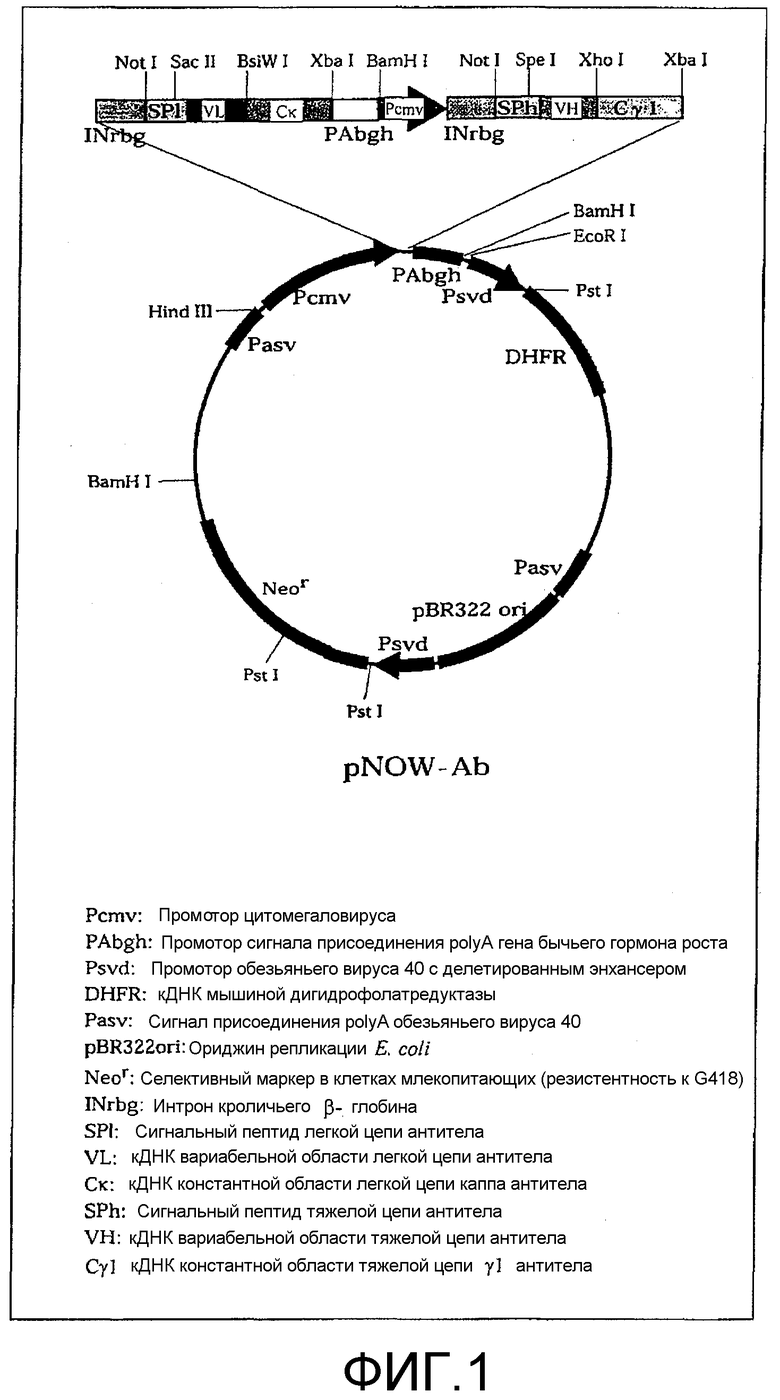
Фиг.1. Структура вектора, экспрессирующего рекомбинантное антитело, pNOW-Ab.
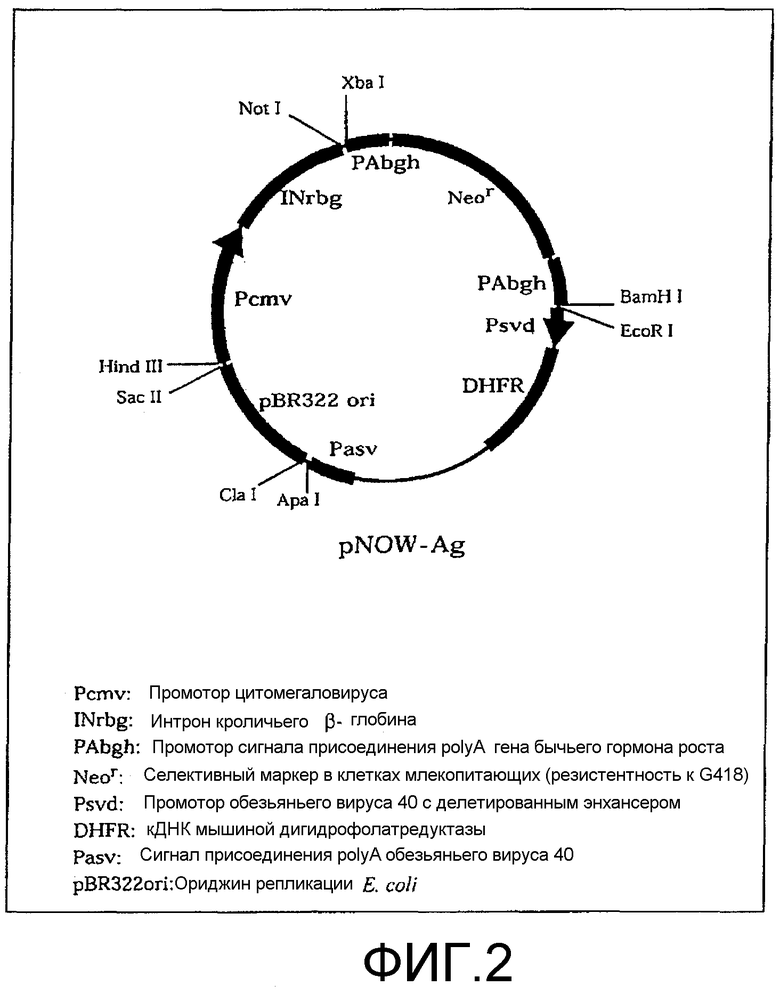
Фиг.2. Структура вектора, экспрессирующего белок, pNOW-Ag.
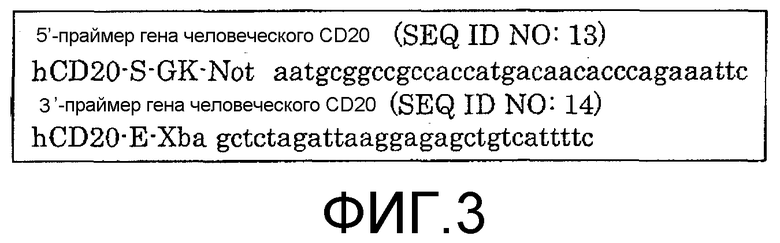
Фиг.3. Последовательности праймеров для клонирования человеческого гена CD20.
Фиг.4. Аминокислотные последовательности вариабельных областей Н-цепи и L-цепи мышиных моноклональных анти-CD20-антител.
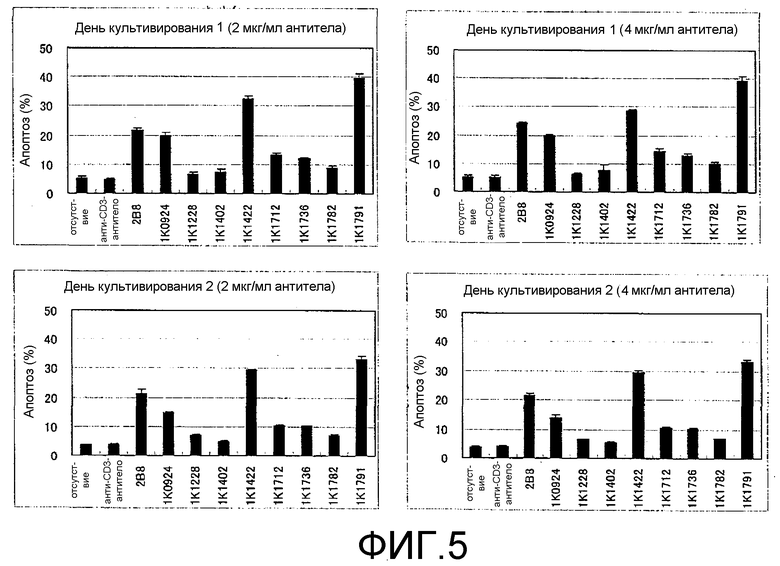
Фиг.5. Результаты теста на апоптоз, проводимого с использованием мышиных моноклональных анти-CD20-антител.
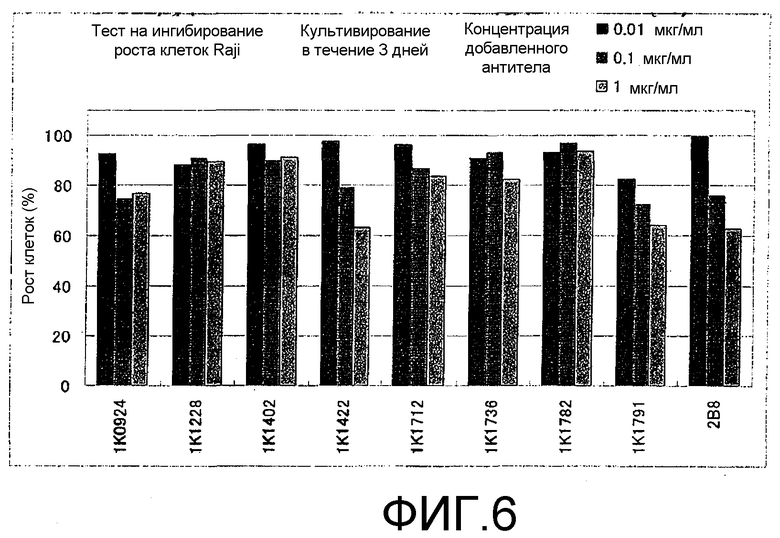
Фиг.6. Результаты теста на ингибирование роста клеток, проводимого с использованием мышиных моноклональных анти-CD20-антител.
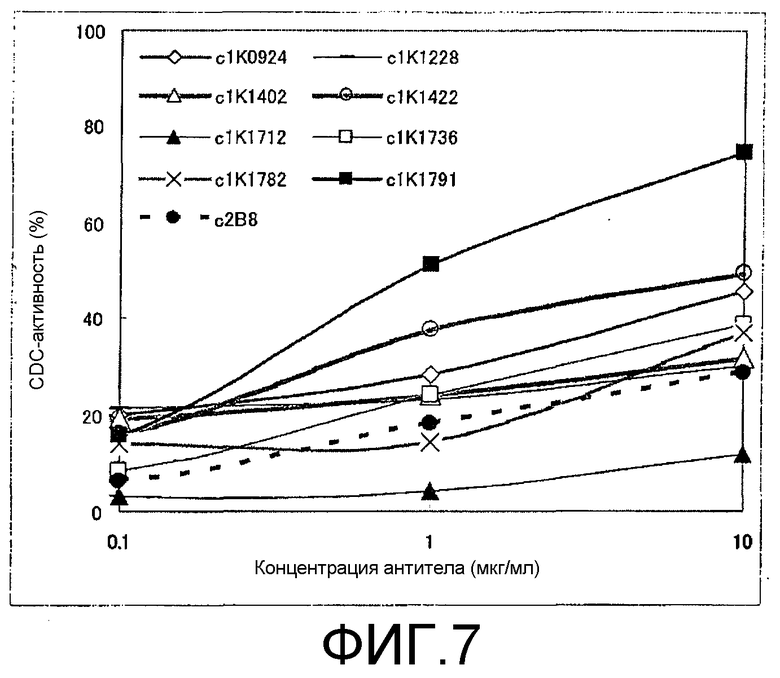
Фиг.7. Результаты теста на комплементзависимую цитотоксичность, проводимого с использованием химерных моноклональных анти-CD20-антител.
Фиг.8А. Аминокислотные последовательности вариабельных областей Н-цепи и L-цепи гуманизованных моноклональных анти-CD20-антител и нуклеотидные последовательности, соответствующие этим последовательностям.
Фиг.8В. Аминокислотные последовательности вариабельных областей Н-цепи и L-цепи гуманизованных моноклональных анти-CD20-антител и нуклеотидные последовательности, соответствующие этим последовательностям.
Фиг.9. Результаты теста на ингибирование роста клеток, проводимого с использованием гуманизованных моноклональных анти-CD20-антител.
Наилучшие варианты осуществления изобретения
Используемый в настоящем изобретении термин “антитело” означает антитело в его общепринятом значении, а также составляющие его H- и L-цепи и их фрагменты.
Настоящее изобретение относится к моноклональному анти-CD20-антителу, которое связывается с человеческим антигеном CD20 на клеточной мембране и обладает желательными биологическими активностями, способствующими индуцированию терапевтического эффекта.
Антителом согласно первому варианту настоящего изобретения является моноклональное антитело, которое специфически связывается с человеческим антигеном CD20 на клеточной мембране и обладает активностями, направленными на ингибирование роста клеток, включая апоптоз клеток, экспрессирующих человеческий антиген CD20 в культуре клеток, экспрессирующих антиген CD20, в отсутствие эффекторных клеток. Таким антителом является, главным образом, мышиное моноклональное анти-CD20-антитело, а также моноклональное анти-CD20-антитело, полученное путем его химеризации или гуманизации. Такие антитела непосредственно обладают активностями, направленными на ингибирование роста клеток, включая апоптоз клеток, экспрессирующих человеческий антиген CD20 в in vitro культуре клеток, экспрессирующих антиген CD20, в отсутствие эффекторных клеток. Такие химерные или гуманизованные моноклональные анти-CD20-антитела обладают комплемент- и/или антителозависимой клеточно-опосредуемой цитотоксичностью.
Уровень связывания с антигеном CD20 на клеточной мембране может быть оценен с помощью ELISA-анализа клеток, в котором CD20-экспрессирующие клетки, такие как клетки SB и Raji, подвергают адгезии к планшету, а затем взаимодействию с тестируемым моноклональным антителом. Однако, поскольку уровни экспрессии антигена CD20 этими клетками являются недостаточными, то и их реактивность является невысокой. Поэтому в настоящем изобретении был разработан способ осуществления клеточного ELISA, в котором клетки СНО, экспрессирующие большое количество CD20 в результате рекомбинации генов (CD20/CHO-клетки), подвергают адгезии к планшету, а затем взаимодействию с тестируемым моноклональным антителом. В предварительном тесте, проводимом в соответствии с настоящим изобретением, было подтверждено, что клеточный ELISA-анализ, осуществляемый с использованием CD20/СНО-клеток, обнаруживает такую же картину, как и клеточный ELISA-анализ, осуществляемый с использованием клеток SB или Raji в тесте на реактивность моноклонального антитела, и обладает высокой специфичностью (см., например, проведение скрининга с помощью клеточного ELISA для CD20/CHO-клеток, таблица 1).
Активности, направленные на прямое ингибирование роста клеток в in vitro культуре клеток, экспрессирующих человеческий антиген CD20, в отсутствие эффекторных клеток, могут быть определены обычным методом (Miyamoto T et al., Avian Dis., Vol. 46(1), 10-16). Кроме того, способность индуцировать апоптоз может быть определена в тесте с применением проточной цитометрии (путем окрашивания аннексином V/иодидом пропидия (PI)).
Примерами антител согласно первому варианту изобретения являются мышиные моноклональные анти-CD20-антитела, имеющие комбинацию аминокислотных последовательностей SEQ ID NO:1 и 7, SEQ ID NO:2 и 8 или SEQ ID NO:15 и 17 в вариабельной области Н-цепи и в вариабельной области L-цепи, а также моноклональные анти-CD20-антитела, полученные путем химеризации или гуманизации этих антител. Такие антитела обладают активностью, направленной на прямое ингибирование роста клеток, включая апоптоз клеток, экспрессирующих человеческий антиген CD20 в in vitro культуре клеток, экспрессирующих антиген CD20, в отсутствие эффекторных клеток. Эти антитела также обладают комплемент- и/или антителозависимой клеточно-опосредуемой цитотоксичностью. Кроме того, настоящее изобретение включает гибридому, продуцирующую мышиное антитело, а также клетки млекопитающих (клетки СНО в примерах), в которые была введена нуклеотидная последовательность, соответствующая любой одной из аминокислотных последовательностей химерных или гуманизованных антител.
Химеризацию осуществляют путем присоединения аминокислотной последовательности вариабельной области Н-цепи мышиного моноклонального антитела к аминокислотной последовательности константной области Н-цепи человеческого иммуноглобулина и аминокислотной последовательности вариабельной области L-цепи к аминокислотной последовательности константной области L-цепи человеческого иммуноглобулина (Ishida T et al., Nippon Rinsho, Vol. 60, №3, 439-444). Гуманизованные антитела конструируют с использованием аминокислотной последовательности CDR вариабельной области мышиного моноклонального антитела и аминокислотной последовательности человеческого иммуноглобулина. Гуманизованные моноклональные анти-CD20-антитела, которые являются предпочтительными для их использования в качестве терапевтических средств, отбирают путем сравнения свойств различных сконструированных антител (Padlan EA, Mol. Immunol., Vol. 28, №4/5, 489-498; Wu TT and Kabat EA, Mol. Immunol., Vol. 29, №9, 1141-1146; Padlan EA et al., FASEB J., Vol. 9, 133-139).
Химерные или гуманизованные моноклональные анти-CD20-антитела также обладают комплементзависимой цитотоксичностью (CDC) и антителозависимой клеточно-опосредуемой цитотоксичностью (ADCC) в присутствии эффекторных клеток. Анализы на CDC и ADCC могут быть проведены общеизвестными методами (Manches O et al., Blood, 2003, 101(3), 949-54; Idusogie EE et al., J. Immunol., 2000, 164, 4178-4184).
Конкретными примерами гуманизованных моноклональных анти-CD20-антител являются антитела, имеющие комбинацию аминокислотных последовательностей SEQ ID NO: 19 и 23, SEQ ID NO: 19 и 24, SEQ ID NO: 19 и 25, SEQ ID NO: 19 и 26, SEQ ID NO: 20 и 23, SEQ ID NO: 20 и 24, SEQ ID NO: 20 и 25, SEQ ID NO: 20 и 26, SEQ ID NO: 21 и 23, SEQ ID NO: 21 и 24, SEQ ID NO: 21 и 25, SEQ ID NO: 21 и 26, SEQ ID NO: 22 и 23, SEQ ID NO: 22 и 24, SEQ ID NO: 22 и 25 или SEQ ID NO: 22 и 26 в вариабельной области Н-цепи и в вариабельной области L-цепи.
Антителом согласно второму варианту изобретения является мышиное моноклональное антитело, которое специфически связывается с человеческим антигеном CD20 на клеточной мембране и не обладает активностью, направленной на ингибирование роста клеток, включая апоптоз, либо оно обладает невысокой активностью. Однако на CDC- или ADCC-активность этих мышиных антител может повлиять их химеризация или гуманизация. Указанные цитотоксические активности также являются очень важными биологическими активностями, а поэтому моноклональное анти-CD20-антитело согласно второму варианту изобретения может также рассматриваться как кандидат на его использование в качестве терапевтического средства.
Примерами антител согласно второму варианту изобретения являются мышиные моноклональные анти-CD20-антитела, имеющие комбинацию аминокислотных последовательностей SEQ ID NO:3 и 9, SEQ ID NO:4 и 10, SEQ ID NO:5 и 11, SEQ ID NO:6 и 12 или SEQ ID NO:16 и 18 вариабельной области Н-цепи и вариабельной области L-цепи, а также моноклональные анти-CD20-антитела, полученные путем химеризации или гуманизации этих антител. Настоящее изобретение также включает гибридому, продуцирующую указанное мышиное антитело, и клетки млекопитающих (клетки СНО в примере), в которые была введена нуклеотидная последовательность, соответствующая любой одной из указанных аминокислотных последовательностей химерных или гуманизованных антител.
Метод определения уровня связывания с антигеном CD20 на клеточной мембране, различные методы анализа на ингибирование роста клеток, апоптоз, ADCC, CDC и т.п. и метод получения химерных или гуманизованных антител аналогичны соответствующим методам, описанным для антитела согласно первому варианту изобретения.
При этом предполагается, что химерное моноклональное анти-CD20-антитело и гуманизованное моноклональное анти-CD20-антитело, рассматриваемое в первом и втором вариантах изобретения, обладают большей эффективностью как терапевтические средства для лечения опосредуемых В-клетками злокачественных опухолей и иммунных заболеваний, в патогенезе которых участвуют В-клетки, а поэтому целью настоящего изобретения является использование этих антител для разработки терапевтического средства, содержащего в качестве активного ингредиента химерное или гуманизованное моноклональное анти-CD20-антитело, а предпочтительно гуманизованное моноклональное анти-CD20-антитело. Примерами рассматривамых заболеваний являются не-ходжкинская лимфома, лимфома Ходжкина, хронический лимфолейкоз, острый лимфолейкоз, хронический ревматоидный артрит, аутоиммунная гемолитическая анемия, идиопатическая тромбоцитопеническая пурпура, системная красная волчанка, синдром, ассоциированный с вырабатыванием антител против фосфолипида, синдром Сьегрена, болезнь Крона, склеродермия, рассеянный склероз, диабет типа 1 и т.п.
Мышиное моноклональное антитело согласно изобретению может быть получено описанным ниже методом.
В качестве иммуногена для сенсибилизации могут быть использованы комбинации клеток SB или Raji, которые представляют собой клеточные штаммы, экспрессирующие человеческий антиген CD20, и клеток СНО, экспрессирующих человеческий антиген CD20. Кроме того, в качестве вспомогательного сенсибилизирующего антигена может быть использован человеческий белок CD20, конъюгированный с GST (GST-CD20).
Моноклональное антитело, продуцируемое гибридомой, может быть получено рядом способов, включая (1) иммунизацию животного, предназначенного для иммунизации (мыши), (2) получение лимфоцитов от указанного иммунизованного животного, (3) получение родительских клеток, (4) слияние лимфоцитов и родительских клеток, (5) скриниг и (6) клонирование (Biochemistry Experiment Method: Monoclonal antibody, written by Ailsa M. Campbell, translated by Toshiaki Osawa, Tokyo Kagaku Dozin Co., Ltd., 1989). Моноклональное анти-CD20-антитело, которое специфически связывается с антигеном CD20 на клеточной поверхности, может быть клонировано посредством реакции тестируемого моноклонального антитела с клеточной ELISA-системой, где клетки CD20/CHO иммобилизованы на планшете. Могут быть также использованы коммерчески доступные экспрессионные векторы. Однако, поскольку необходимо, чтобы антиген CD20 экспрессировался на клетках СНО с высокой плотностью, то может быть использован высокоэкспрессионный вектор млекопитающих, pNOW (Japanese Patent №3582965). Критерием отбора моноклонального антитела может служить его реактивность, сравнимая или превышающая реактивность позитивного контроля.
Химерное или гуманизованное антитело может быть получено в соответствии со стандартной методикой рекомбинации генов. Так, например, для продуцирования антитела в качестве экспрессионного вектора может быть использован вектор pNOW-Ab, который содержит 2 набора сайтов множественного клонирования, расположенных тандемно и предварительно введенных вместе с генами, кодирующими константные области человеческой Н-цепи и L-цепи (фиг.1).
Пример 1
Получение, химеризация и гуманизация моноклональных антител против антигена CD20, а также тест для определения свойств полученных антител будут описаны ниже со ссылками на примеры.
(1) Получение иммуногена для сенсибилизации мышей
Ген, кодирующий человеческий CD20, получали из библиотеки кДНК с использованием 5'-праймера SEQ ID NO:13 и 3'-праймера SEQ ID NO: 14, которые являются специфичными в отношении гена, кодирующего полноразмерную молекулу человеческого CD20 (Multiple Choice cDNA human spleen, Origene Technologies, Inc., 6 Taft Court, Suite 100, Rockville, MD 20850). В частности, были использованы праймеры, представленные на фиг.3. Ген CD20 вводили в вектор pNOW-Ag (фиг.2), используемый в качестве высокоэкспрессионного вектора для экспрессии в клетках млекопитающих, и трансфицировали в клетки CHO, используемые в качестве клеток-хозяев. Рекомбинантные клетки CHO (клетки CD20/CHO), экспрессирующие на своей поверхности молекулы CD20 на высоком уровне, были протестированы с помощью FAСS-анализа. Клетки, которые при их окрашивании ФИТЦ-меченными моноклональными анти-CD20-антителами обнаруживали 5-кратное или большее превышение интенсивности флуоресценции по сравнению с клетками SB, были определены как клетки, имеющие высокий уровень экспрессии. GST-CD20, дополнительный иммуноген, получали путем присоединения GST у N-конца внеклеточного домена CD20, состоящего из 43 аминокислотных остатков, с использованием вектора pGEX-4T2 (G et al. AM, Electrophoresis, Vol. 20(2): 344-348).
(2) Получение иммуногена
Клетки SB или Raji культивировали в среде RPMI 1640, в которую была добавлена 10% FCS. Клетки CD20/CHO культивировали в среде CHO-S-SFM II (GIBCO, Cat. №12052-098), в которую было добавлено 800 мкг/мл G418. Эти культуры центрифугировали (1100 об/мин, 5 минут), а затем в клеточные культуры добавляли среду Дульбекко без PBS (PBS(-)) и суспендировали, после чего суспензию снова центрифугировали. Эту процедуру промывки повторяли еще раз и суспензию, полученную путем добавления к клеткам (количество клеток: 1-3 × 107/мл) физиологического раствора, использовали для иммунизации. Вектор pGEX-4T2, в который был включен GST-CD20, вводили в компетентные клетки E. coli. После культивирования компетентные клетки подвергали лизису и GST-CD20 посредством грубой очистки отделяли от лизированных клеток, а затем солюбилизировали путем добавления 0,1 н. гидроксида натрия.
(3) Иммунизация и слияние клеток
Для иммунизации использовали самок 7-11-недельных мышей Balb/c. Клетки SB, клетки Raji или клетки CD20/CHO повторно вводили два или три раза через различные интервалы времени в несколько дней, а затем клетки отбирали на различные клеточные антигены (клетки SB, клетки Raji или клетки CD20/CHO) и использовали для конечной иммунизации. Число вводимых клеток независимо от их типа составляло 1-3 × 107 на мышь. Затем для части мышей проводили дополнительную иммунизацию с использованием GST-CD20. Через три дня после конечной иммунизации у мышей брали клетки селезенки и суспендировали в среде RPMI, а затем осуществляли слияние этих клеток с мышиными миеломными клетками (NS-1) в присутствии ПЭГ-1500 (Oi, VT et. Herzenberg, 1980, in: Selected Methods in Cellular Immunology; Mishell B et al. (Freeman and Co., San Francisco, CA) p.351).
(4) Разработка метода ELISA-скринига клеток CD20/CHO
Несколько мышиных моноклональных анти-CD20-антител и антител 2B8 подвергали взаимодействию с клетками SB, с клетками Raji, с клетками CD20/CHO и с родительскими CD20-клетками CHO, которые были иммобилизованы на 96-луночных планшетах. Было подтверждено, что в этих клеточных ELISA-тестах наблюдались аналогичные тенденции для концентраций антител, и было обнаружено, что могут быть также проведены относительные сравнения этих антител между собой и с контролем. Из-за высокой плотности антигенов клеточной поверхности, иммобилизованных на планшете, в ELISA-анализе клеток CD20/CHO, наблюдаемый уровень оптической плотности был достаточным для детекции даже в образце с относительно низкой концентрацией тестируемого антитела, в результате чего было установлено, что указанный анализ представляет собой чувствительную систему измерения. Конкретные результаты измерения приводятся в таблице 1.
Сравнение клеток SB, клеток Raji и клеток CD20/CHO в ELISA
(5) Скрининг с помощью клеточного ELISA
Клеточный ELISA осуществляли с использованием 96-луночного планшета, на котором были иммобилизованы клетки CD20/CHO или клетки CHO (родительская клеточная линия CD20), и отбирали лунки, в которых продуцировались антитела, специфически реагирующие с CD20. В качестве позитивного контроля использовали 2B8, а в качестве негативного контроля использовали мышиное моноклональное антитело против человеческого антигена CD3 (BD PharMingen). В частности, для проведения клеточного ELISA использовали клетки CD20/CHO или клетки CHO (родительскую клеточную линию), иммобилизованные на 96-луночном планшете, покрытом поли-L-лизином (Asahi Techno Glass Corporation, Cat. № 11-023-018). В каждую лунку добавляли блокирующий раствор (0,2% желатина и 0,5% раствора BSA в PBS) в объеме 150 мкл и оставляли на 1 час при 37оС. Затем планшет 5 раз промывали 150 мМ NaCl и 0,05% водного раствора твина 20, после чего в каждую лунку добавляли 100 мкл каждого из образцов (разведенного раствора супернатанта культуры) и проводили первичную реакцию при 37°C в течение 1 часа. После промывки в каждую лунку добавляли 100 мкл разведенного раствора меченного антитела [ПХ-меченного кроличьего антитела против мышиных IgG(H+L) (Jackson Lab., Code № 315-035-003) или ПХ-меченного кроличьего антитела против мышиных IgG(Fcγ) (Jackson Lab., Code № 315-035-008)] и проводили вторичную реакцию при 37°C в течение 1 часа. В целях получения реакционных смесей для первичной и вторичной реакции тот же самый раствор использовали в качестве блокирующего раствора. После промывки в каждую лунку добавляли 100 мкл раствора для проявления окраски (OPD), а через 30 минут добавляли 50 мкл 4 н. H2SO4 для прекращения реакции и измеряли оптическую плотность при 492 нм (A492). Затем отбирали лунки, обнаруживающие реактивность, сравнимую с реактивностью антитела 2B8 или значительно превышающую эту реактивность.
(6) Клонирование
Клонирование осуществляли методом лимитирующего разведения. Клетки высевали на 96-луночный планшет и культивировали, а затем для отбора клона, продуцирующего специфическое антитело, проводили клеточный ELISA для клеток CD20/CHO с использованием супернатанта культуры в лунке, содержащей 1 колонию.
(7) Получение очищенного антитела
Клон, продуцирующий специфическое антитело, культивировали в среде RPMI 1640, в которую была добавлена 10% FCS. При достижении клеточной плотности примерно 5 × 105/мл эту среду заменяли бессывороточной средой ASF-104N (Ajinomoto Co. Inc.) и продолжали культивирование. Затем через 2-4 дня культуральную среду центрифугировали и супернатант культуры собирали, а затем очищали на колонке с G-белком. Раствор элюированного моноклонального антитела диализовали против 150 мМ NaCl. Раствор стерилизовали фильтрацией с использованием фильтра с размером пор 0,2 мкм и использовали в качестве тестируемого антитела (мышиного моноклонального антитела против человеческого CD20).
Клоны моноклональных антител, обнаруживающие аффинность связывания, сравнимую с аффинностью связывания позитивного контроля, отбирали с помощью ELISA клеток CD20/CHO. Затем определяли генные последовательности вариабельных областей этих антител, а следовательно, и их аминокислотные последовательности. Последовательности вариабельной области Н-цепи и вариабельной области L-цепи типичных антител представлены в SEQ ID NO: 1 и 7, SEQ ID NO: 2 и 8, SEQ ID NO: 3 и 9, SEQ ID NO: 4 и 10, SEQ ID NO: 5 и 11, SEQ ID NO: 6 и 12, SEQ ID NO: 15 и 17 и SEQ ID NO: 16 и 18 (фиг.4). Кроме того, были исследованы биологические свойства этих клонов.
Тест на биологические свойства (1): Тест на индуцирование апоптоза
Способность тестируемых антител индуцировать апоптоз определяли с помощью проточной цитометрии (путем окрашивания аннексином V/PI). В качестве позитивного контроля использовали 2B8, а в качестве негативного контроля использовали мышиное моноклональное антитело против человеческого CD3 (BD PharMingen). Для этого проводили нижеследующие процедуры.
Был использован набор для апоптоза MEBCYTO (MBL, Cat. № 4700, Lot. 20).
Клетки Raji центрифугировали, а затем суспендировали в свежей среде RPMI 1640 (Sigma, Cat. № R8758, Lot 44K2416), содержащей 10% FCS (инактивированную) (ICN, Cat. № 2916754, Lot 8005C), и в каждую лунку 12-луночного планшета добавляли 1 мл полученной суспензии при плотности 5 × 105 клеток/мл. Для каждого антитела использовали двенадцать лунок и каждое антитело добавляли при конечной концентрации 2 мкг/мл или 4 мкг/мл (3 лунки × 2 различных концентрации × 2 раза, всего 12 лунок).
Через один день и через два дня после начала культивирования культуральную среду, содержащую примерно 2 × 105 клеток, собирали и центрифугировали, а затем клетки один раз промывали PBS. После этого в клетки добавляли 85 мкл связывающего буфера и клетки суспендировали в этом буфере. Затем к полученной суспензии добавляли 10 мкл аннексина V-ФИТЦ и 5 мкл PI, смешивали в достаточной степени и оставляли на 15 минут в светонепроницаемом помещении при комнатной температуре для прохождения реакции. Измерение проводили с помощью проточной цитометрии (FACS Calibur, Becton Dickinson) и результаты анализировали на аппарате CellQuest (Becton Dickinson).
Результаты измерений для 8 видов типичных мышиных моноклональных анти-CD20-антител, позитивного контроля (2B8) и негативного контроля (анти-CD3 антитела) представлены на фиг.5. Вообще говоря, можно утверждать, что апоптоз-индуцирующая активность антитела 2B8 является высокой. Но даже по сравнению с этим антителом, клон, имеющий аминокислотные последовательности вариабельной области Н-цепи и вариабельной области L-цепи, представленные SEQ ID NO: 2 и 8 (1K1791), обладал значительно более высокой апоптоз-индуцирующей активностью. Гибель клеток, явно обусловленная апоптозом, также наблюдалась для клонов, которые имели аминокислотные последовательности вариабельной области Н-цепи и вариабельной области L-цепи, представленные SEQ ID NO: 1 и 7 (1K1422) и SEQ ID NO: 15 и 17 (1K0924).
Тест на биологические свойства (2): Тест на ингибирование роста клеток
Суспензию 5 × 104 клеток Raji/мл получали с использованием среды RPMI 1640, в которую была добавлена 10% FCS, а затем эту суспензию добавляли в 96-луночный планшет в объеме 100 мкл/лунку и проводили культивирование. Через 24 часа добавляли 50 мкл/лунку каждого раствора антитела при концентрации антител 0,01 мкг/мл, 0,1 мкг/мл или 1 мкг/мл и продолжали культивирование. Через 72 часа после добавления антитела добавляли 10 мкл/лунку раствора для проявления окраски (Cell Counting Kit-8; Dojindo Laboratories, Cat. № 343-07623, Lot SG076), клетки культивировали еще 4 часа, а затем измеряли оптическую плотность при 492 нм. Число живых клеток, продуцирующих 8 видов типичных мышиных моноклональных анти-CD20-антител и позитивный контроль (2B8), указано на фиг.6 и выражено в процентах от негативного контроля (100%). Действие, направленное на ингибирование роста клеток, может быть оценено исходя из уровня снижения числа живых клеток по сравнению с негативным контролем. Явное ингибирование роста клеток наблюдалось для 1K0924, 1K1422, 1K1791 и 2B8, используемых в качестве позитивного контроля, а особенно заметным было ингибирование под действием 1K1791. Эта тенденция прослеживалась и в результатах теста на индуцирование апоптоза.
Получение химерных антител
Гены, кодирующие вариабельные области Н-цепи и L-цепи каждого мышиного антитела, встраивали в вектор pNOW-Ab, то есть высокоэкспрессионный вектор для клеток CHO, уже содержащий гены, кодирующие константные области (к) Н-цепи и L-цепи человеческого иммуноглобулина, в виде кластера. Каждый экспрессионный вектор трансфицировали в клетки СНО и для каждого антитела отбирали клоны, обнаруживающие высокую продуктивность.
Тест на способность химерных антител связываться с антигеном CD20
Полученные 8 видов химерных моноклональных анти-CD20-антител были проанализированы на их способность реагировать с человеческим антигеном CD20 с помощью ELISA-клеток CD20/CHO. В качестве позитивного контроля использовали ритуксимаб (c2B8). Результаты теста представлены в таблице 2. Величины, измеренные в клеточном ELISA (A492), представляют собой уровень связывающей активности. Эти антитела обнаруживали аффинность связывания, по существу, сравнимую с аффинностью или превышающую аффинность связывания контрольных антител, за исключением c1K0924 и c1K1422, которые обнаруживали несколько более низкую аффинность по сравнению с контролем.
ELISA-анализ для клеток CD20/CHO, продуцирующих химерные анти-CD20-антитела
Тест на CDC химерных антител
Полученные 8 видов химерных моноклональных анти-CD20-антител оценивали на их CDC-активность. В качестве позитивного контроля использовали ритуксимаб (c2B8). В качестве клеток-мишеней использовали клетки RC-K8 (полученные от Kochi Medical School). Для проведения этого теста приготавливали и использовали среду RHB (базалиная среда: RPMI-1640 с добавками: 0,1% BSA, 20 мM HEPES (pH 7,2), 2 мM глутамин, 100 единиц/мл пенициллина G, 100 мкг/мл стрептомицина). Клетки-мишени промывали средой RHB и ресуспендировали при 106 клеток/мл. В каждую лунку плоскодонного 96-луночного планшета для культивирования тканей (черный) добавляли 50 мкл каждого из растворов тестируемых химерных антител и ритуксимаба в различных концентрациях, 50 мкл 4-кратно разведенного раствора коммерчески доступного человеческого комплемента (Quidel, San Diego, CA, Cat. A113) и 50 мкл клеточной суспензии, содержащей 106 клеток/мл. Концентрация антитела в смеси 150 мкл/лунку составляла 0,1, 1 и 10 мкг/мл. Для стимуляции опосредуемого комплементом лизиса клеток смесь инкубировали при 37°C и 5% CO2 в течение 2 часов. К этой смеси добавляли 50 мкл аламарового синего (неразведенного, изготовленного фирмой AccuMed International, Biosource, Cat. DAL1100) и смесь оставляли на ночь при тех же условиях для прохождения реакции. Затем планшет оставляли на 10 минут при комнатной температуре для охлаждения и измеряли интенсивность флуоресценции при излучении на 590 нм и при возбуждении на 530 нм с использованием флуоресцентного ридера для микротитрационных планшетов. Полученные результаты были представлены как интенсивность флуоресценции (RFU). Уровень CDC-активности вычисляли по следующему уравнению:
% CDC-активности = 100 × {RFU (без добавления антитела) - RFU (с добавлением антитела))/{RFU (без добавления антитела] - RFU (с добавлением тритона X-100)}
Результаты представлены на фиг.7. Все антитела, за исключением c1K1712, обладали CDC-активностями, которые были, по существу, сравнимы с активностью C2B2, используемого в качестве позитивного контроля, или превышали такую активность.
Получение гуманизованных антител
Гуманизованные антитела конструировали на основе CDR вариабельной области мышиного моноклонального анти-CD20-антитела 1K1791. Путем проведения структурного анализа исходя из аминокислотных последовательностей и путем дополнительной модификации такого метода конструирования было получено 4 вида гуманизованных последовательностей для каждой из Н-цепей и L- цепей (Padlan EA, Mol. Immunol., Vol. 28, № 4/5, 489-498; Wu TT and Kabat EA, Mol. Immunol., Vol. 29, № 9, 1141-1146; Padlan EA et al., FASEB J., Vol. 9, 133-139). Были получены антитела со всеми возможными комбинациями четырех типов для каждой из Н- и L-цепей. Эти аминокислотные последовательности представлены в SEQ ID NO: 19 и 23, SEQ ID NO: 19 и 24, SEQ ID NO: 19 и 25, SEQ ID NO: 19 и 26, SEQ ID NO: 20 и 23, SEQ ID NO: 20 и 24, SEQ ID NO: 20 и 25, SEQ ID NO: 20 и 26, SEQ ID NO: 21 и 23, SEQ ID NO: 21 и 24, SEQ ID NO: 21 и 25, SEQ ID NO: 21 и 26, SEQ ID NO: 22 и 23, SEQ ID NO: 22 и 24, SEQ ID NO: 22 и 25 и SEQ ID NO: 22 и 26 (фиг.8A и 8B).
Аминокислотные последовательности 4 видов вариабельных областей Н-цепи и 4 видов вариабельных областей L-цепи были преобразованы в последовательности ДНК (нуклеотидные последовательности) с кодонами, наиболее часто встречающихся в человеческих генных последовательностях, и некоторые из этих нуклеотидов были изменены с учетом возможности их использования в клетках-хозяевах СНО без изменения исходных аминокислотных остатков (Kim CH et al., Gene, 1997, 15; 199 (1-2): 293-301). В частности, нуклеотидными последовательностями, соответствующими аминокислотным последовательностям, являются последовательность SEQ ID NO: 27, соответствующая SEQ ID NO: 19; последовательность SEQ ID NO: 28, соответствующая SEQ ID NO: 20; последовательность SEQ ID NO: 29, соответствующая SEQ ID NO: 21; последовательность SEQ ID NO: 30, соответствующая SEQ ID NO: 22; последовательность SEQ ID NO: 31, соответствующая SEQ ID NO: 23; последовательность SEQ ID NO: 32, соответствующая SEQ ID NO: 24; последовательность SEQ ID NO: 33, соответствующая SEQ ID NO: 25; и последовательность SEQ ID NO: 34, соответствующая SEQ ID NO: 26 (фиг.8A и 8B). В этих нуклеотидных последовательностях нуклеотиды могут быть заменены другими нуклеотидами при условии, что соответствующие аминокислотные последовательности останутся без изменения.
Было синтезировано всего 8 видов нуклеотидных последовательностей вариабельных областей Н-цепи (SEQ ID NO: 27-30) и вариабельных областей L-цепи (SEQ ID NO: 31-34) (Takara Shuzo Co., Ltd.), и эти последовательности встраивали в pNOW-Ab, то есть в экспрессионный вектор для клеток млекопитающих, содержащий сайт множественного клонирования. Экспрессионные векторы, в которые были включены каждый из этих генов гуманизованных антител, трансфицировали в клетки CHO и для каждого антитела отбирали клоны с высокой продуктивностью.
Тесты на биологические свойства гуманизованных антител и на ингибирование ими клеточного роста
Суспензию, содержащую 5 × 104 клеток Raji/мл, получали с использованием среды RPMI 1640, в которую была добавлена 10% FCS, а затем эту суспензию добавляли в 96-луночный планшет в объеме 100 мкл/лунку и проводили культивирование. Через 24 часа добавляли 50 мкл/лунку каждого раствора антитела при концентрации антитела 0,5 мкг/мл и продолжали культивирование. Через 72 часа после добавления антитела добавляли 10 мкл/лунку раствора для проявления окраски (Cell Counting Kit-8; Dojindo Laboratories, Cat. № 343-07623, Lot SG076) и клетки культивировали еще 4 часа, а затем измеряли оптическую плотность при 492 нм. Число живых клеток, продуцирующих 15 (27 клонов) из 16 видов гуманизованных антител, происходящих от 1K1791 и С2В8 (другое название ритуксимаба), и используемых в качестве позитивного контроля, указано на фиг.9 и выражено в процентах от негативного контроля (100%). Действие, направленное на ингибирование роста клеток, может быть оценено исходя из уровня снижения числа живых клеток по сравнению с негативным контролем, и в этом тесте эффект ингибирования клеточного роста наблюдался для всех клонов.
Названия моноклональных антител и номера последовательностей, описанных в настоящей заявке и в прилагаемом графическом материале, систематизированы в нижеследующей таблице.
SEQ ID NO:19 и 25
SEQ ID NO:19 и 26
SEQ ID NO:20 и 23
SEQ ID NO:20 и 24
SEQ ID NO:20 и 25
SEQ ID NO:20 и 26
SEQ ID NO:21 и 23
SEQ ID NO:21 и 24
SEQ ID NO:21 и 25
SEQ ID NO:21 и 26
SEQ ID NO:22 и 23
SEQ ID NO:22 и 24
SEQ ID NO:22 и 25
SEQ ID NO:22 и 26
Гибридомы, продуцирующие эти моноклональные антитела, были названы исходя из названий продуцируемых ими антител и были депонированы согласно Международному договору в Международном депозитарии патентуемых микроорганизмов Национального Института Новых Промышленных Исследований и Технологий (National Institute of Advanced Industrial Science and Technology) (Tsukuba Central 6, 1-1, Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, 305-8566, Japan) 28 марта, 2006, в рамках Будапештского договора под регистрационными номерами FERM BP-10587 (1K1422), FERM BP-10591 (1K1791), FERM BP-10588 (1K1712), FERM BP-10586 (1K1402), FERM BP-10589 (1K1736), FERM BP-10590 (1K1782), FERM BP-10584 (1K0924) и FERM BP-10585 (1K1228).
Промышленное применение
Настоящее изобретение относится к моноклональному антителу, обладающему биологической активностью, подходящей для его использования в качестве терапевтического средства.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ЛЕЧЕНИЯ, АНТИТЕЛА, ГИБРИДОМА | 1993 |
|
RU2139731C1 |
| ПРИМЕНЕНИЕ АНТИ-CD40-АНТИТЕЛ | 2006 |
|
RU2442606C2 |
| ОЧИСТКА АНТИТЕЛ С ПОМОЩЬЮ КАТИОНООБМЕННОЙ ХРОМАТОГРАФИИ | 2008 |
|
RU2498991C2 |
| АНТИТЕЛО ПРОТИВ ILT17 | 2006 |
|
RU2456298C2 |
| АНТИ-СD3-АНТИТЕЛО И МОЛЕКУЛА, СОДЕРЖАЩАЯ ЭТО АНТИТЕЛО | 2017 |
|
RU2790326C2 |
| АНТИТЕЛО ПРОТИВ ILT17 | 2012 |
|
RU2599450C2 |
| АНТИТЕЛА ПРОТИВ IL2 | 2006 |
|
RU2425054C2 |
| ДЕГРАДИРОВАННОЕ АНТИТЕЛО, ЯВЛЯЮЩЕЕСЯ АГОНИСТОМ TPO | 2001 |
|
RU2287534C2 |
| Антитело против PD-1 и его применение | 2017 |
|
RU2739610C1 |
| ЛЕЧЕНИЕ РАКА ПРИ ПОМОЩИ НАПРАВЛЕННЫХ НА МИШЕНЬ АНТИТЕЛ IN VIVO | 2011 |
|
RU2642305C2 |














Изобретение относится к биотехнологии. Описано моноклональное антитело, способное индуцировать специфическую биологическую реакцию посредством его связывания с антигеном CD20 на поверхности клетки. Было клонировано моноклональное антитело, обладающее высокой аффинностью связывания с внеклеточным эпитопом антигена CD20, а также биологическими активностями, включая активность, направленную на ингибирование роста клеток. Указанное моноклональное антитело было подвергнуто химеризации или гуманизации в целях разработки терапевтического средства для лечения заболевания, опосредуемого В-клетками. Изобретение расширяет спектр терапевтических средств, полученных на основе моноклональных анти-СD20-антител. 7 н. и 3 з.п. ф-лы, 9 ил., 3 табл.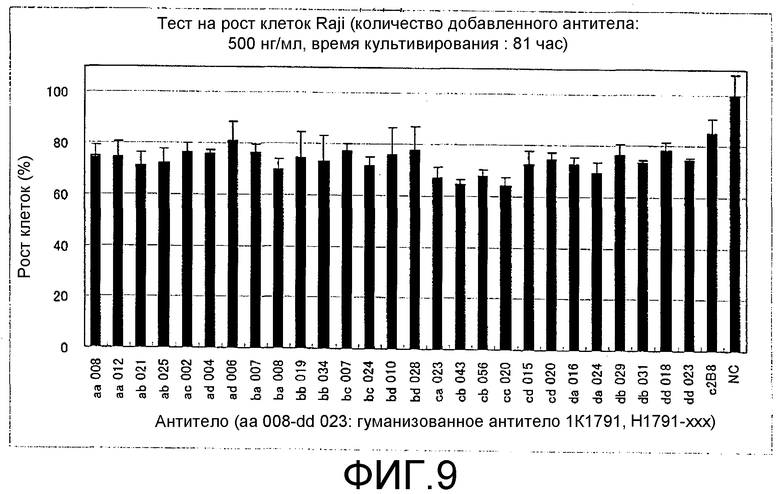
1. Мышиное моноклональное анти-СD20-антитело, обладающее активностью, направленной на ингибирование роста клеток, включая апоптоз клеток, экспрессирующих человеческий антиген CD20, в культуре клеток, экспрессирующих антиген CD20 в отсутствие эффекторных клеток, где аминокислотные последовательности вариабельной области Н-цепи и вариабельной области L-цепи представляют собой последовательности SEQ ID NO:1 и 7 соответственно или SEQ ID NO:2 и 8 соответственно.
2. Гибридома, продуцирующая моноклональное анти-СD20-антитело по п.1, регистрационным номером которой является FERM BP-10587.
3. Гибридома, продуцирующая моноклональное анти-СD20-антитело по п.1, регистрационным номером которой является FERM BP-10591.
4. Химерное моноклональное анти-СD20-антитело, где аминокислотная последовательность вариабельной области Н-цепи, представленная в SEQ ID NO:1, и аминокислотная последовательность вариабельной области L-цепи, представленная в SEQ ID NO:7, слиты с аминокислотными последовательностями константной области Н-цепи и константной области L-цепи человеческого иммуноглобулина соответственно, или аминокислотная последовательность вариабельной области Н-цепи, представленная в SEQ ID NO:2, и аминокислотная последовательность вариабельной области L-цепи, представленная в SEQ ID N0:8, слиты с аминокислотными последовательностями константной области Н-цепи и константной области L-цепи человеческого иммуноглобулина соответственно.
5. Моноклональное анти-СD20-антитело, гуманизованное с использованием аминокислотных последовательностей CDR
вариабельной области Н-цепи, представленной в SEQ ID NO:1, и вариабельной области L-цепи, представленной в SEQ ID NO:7, или
вариабельной области Н-цепи, представленной в SEQ ID NO:2, и вариабельной области L-цепи, представленной в SEQ ID NO:8,
и аминокислотной последовательности человеческого иммуноглобулина.
6. Гуманизованное моноклональное анти-СD20-антитело по п.5, где комбинация аминокислотных последовательностей вариабельной области Н-цепи и вариабельной области L-цепи представляет собой комбинацию последовательностей SEQ ID NO:19 и 23, SEQ ID NO:19 и 24, SEQ ID NO:19 и 25, SEQ ID NO:19 и 26, SEQ ID NO:20 и 23, SEQ ID NO:20 и 24, SEQ ID NO:20 и 25, SEQ ID NO:20 и 26, SEQ ID NO:21 и 23, SEQ ID NO:21 и 24, SEQ ID NO:21 и 25, SEQ ID NO:21 и 26, SEQ ID NO:22 и 23, SEQ ID NO:22 и 24, SEQ ID NO:22 и 25 или SEQ ID NO:22 и 26 соответственно.
7. Моноклональное анти-СD20-антитело по любому из пп.4-6, которое оказывает цитотоксическое действие на клетки, экспрессирующие антиген CD20, в присутствии человеческого комплемента или эффекторных клеток.
8. Клетка млекопитающего, в которую была введена нуклеотидная последовательность, кодирующая аминокислотную последовательность моноклонального анти-СD20-антитела по любому из пп.4-7, благодаря чему продуцируется моноклональное анти-СD20-антитело.
9. Клетка млекопитающего по п.8, которой является клетка СНО.
10. Терапевтическое средство, содержащее эффективное количество моноклонального анти-СD20-антитела по любому из пп.4-7 в качестве активного ингредиента, для лечения неходжкинской лимфомы, лимфомы Ходжкина, хронического лимфолейкоза, острого лимфолейкоза, хронического ревматоидного артрита, аутоиммунной гемолитической анемии, идиопатической тромбоцитопенической пурпуры, системной красной волчанки, синдрома, ассоциированного с вырабатыванием антител против фосфолипида, синдрома Сьегрена, болезни Крона, склеродермии, рассеянного склероза или диабета типа 1.
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |
| Перекатываемый затвор для водоемов | 1922 |
|
SU2001A1 |
| СПОСОБ ЛЕЧЕНИЯ, АНТИТЕЛА, ГИБРИДОМА | 1993 |
|
RU2139731C1 |

