ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Изобретение относится к применению антисмысловых олигонуклеотидов, направленных против специфичных клеточных рецепторов, отдельно или в комбинации для ингибирования общего воспаления, включая воспаление, связанное с астмой и аллергией, и гиперэозинофилии. Изобретение также относится к применению антисмысловых олигонуклеотидов для ингибирования пролиферации неопластических клеток, напрмер, как при злокачественной опухоли (раке).
УРОВЕНЬ ТЕХНИКИ
Антисмысловые олигонуклеотиды являются новым классом фармацевтических средств. В общем, «антисмысловые» относится к применению небольших синтетических олигонуклеотидов с такими же составляющими, как и составляющие, обнаруживаемые в ДНК или РНК человека. Антисмысловые олигонуклеотиды конструируют в виде последовательности, комплементарной части гена, которая является их мишенью, чтобы они были способны связываться с такой последовательностью и ингибировать экспрессию гена. Экспрессия гена ингибируется в результате гибридизации антисмыслового олигонуклеотида со смысловой мишенью специфичной матричной РНК (мРНК) в соответствии с правилом спаривания оснований Уотсона-Крика, согласно которому аденозин и тимидин (урацил в мРНК) или гуанозин и цитидин взаимодействуют посредством образования водородных связей. Два механизма могут объяснять указанные эффекты, первый представляет собой гибридизацию с нарушенной трансляцией мРНК-мишени, второй представляет собой индукцию РНКазы H или сходных ферментов с разрушением мРНК. Основным преимуществом такой методики является специфичность действия с возможными меньшими побочными эффектами и токсичностью, особенно в случае применения по месту действия (местное лечение). Такая терапевтическая методика потенциально может быть применима по отношению к любому заболеванию, при котором предположительно сверхэкспрессия одного или нескольких генов является причиной наличия или продолжительности заболевания. В результате проведены многочисленные исследования антисмысловых олигонуклеотидов в качестве терапевтических средств для лечения рака и вирусных болезней.
Антисмысловые олигонуклеотиды могут быть использованы для ингибирования экспрессии рецептора интерлейкина (IL-6) и таким образом ингибирования эффектов медиатора острого воспаления интерлейкина-6 на клетках. Проведено несколько исследований для оценки того, можно ли использовать антисмысловые олигонуклеотиды для ингибирования других рецепторов на клетках, которые вовлечены в воспаление, включая без ограничения воспаление, связанное с астмой, и воспаление, связанное атопическими заболеваниями и аллергией, или на раковых клетках.
Астма является заболеванием, которое поражает от 5 до 10% популяции, распространенность которого возросла в два раза в последние 25 лет. Такое увеличение особенно отмечено у новорожденных после вирусной инфекции дыхательных путей (бронхиолит), у детей и в случае профессиональной астмы. Рецидивирующие проблемы дыхания, связанные с астмой, часто стимулируются аллергенами, но точная причина астмы еще не известна. Однако предполагается, что агенты, такие как вирусы, вовлечены в поддержание аномального воспаления, которое выявляют в дыхательных путях пациентов с астмой, и таким образом в устойчивость заболевания.
По этой причине современные рекомендации для терапии астмы первой линии заключаются в применении эффективного противовоспалительного лекарственного средства, такого как средства, содержащие кортикостероиды и антилейкотриены. Хотя такая терапия эффективна у многих пациентов, некоторые пациенты нечувствительны к кортикостероидам. Указанное лекарственное средство также является мощным иммуносупрессором с длительными побочными эффектами, не было показано, что оно является эффективным для профилактики аллергии или астмы. Антилейкотриены оказывают некоторый эффект при аллергии и астме, но не являются такими эффективными, как кортикостероиды.
Несколько воспалительных медиаторов играют роль в появлении и сохранении воспаления в дыхательных путях пациентов с астмой. Некоторые медиаторы привлекают воспалительные клетки в дыхательные пути посредством хемотаксиса эозинофилов (хемокины: RANTES, эотаксины 1, 2, 3, MCP-3, 4, которые главным образом действуют при астматическом воспалении посредством рецептора, называемого CCR3) или посредством активации эндотелиальных клеток (IL-4, -13). Другие медиаторы вызывают примирование и повышенную жизнеспособность воспалительных клеток в дыхательных путях (IL-3, -4, -5, GM-CSF). Указанные медиаторы таким образом включают либо специфичные хемокины для эозинофилов, либо цитокины T-хелперных лимфоцитов с фенотипом типа 2 (Th2: IL-3, -4, -5, -6, -9, -10, -13 и GM-CSF), (John A.E. and Lukacs N.W., 2003 Sarcoidosis Vase Diffuse Lung Dis., 20: 180-189; Blease et al., 2003, Expert. Opin. Emerg. Drugs. 8: 71-81). При астме и общем воспалении дыхательных путей было показано улучшение в случае снижения указанных воспалительных медиаторов в дыхательных путях.
Аллергия представляет собой гиперчувствительность к аллергену, вызывающему нежелательный иммунный ответ. Аллергия является заболеванием, которое чрезвычайно распространено, например атопический ринит и конъюнктивит поражают около 30% популяции. Аллергия характеризуется аномальной продукцией IgE и воспалением в ответ на аллерген. В присутствии IgE и аллергена эффекторные клетки, такие как тучные клетки, дегранулируют и высвобождают медиаторы воспаления, приводящие к рекрутингу таких же воспалительных клеток, которые обнаруживаются при астме. При аллергическом рините (т.е. сенной лихорадке), аллергическом конъюнктивите, полипозе носа, хроническим синусите и экземе, такой как атопический дерматит, обнаруживают такой же избыток медиаторов воспаления, как и избыток в случае астмы. IL-4 и IL-13 необходимы для продукции IgE и индукции клеток с фенотипом Th2 (Barnes P.J., 2003, Cytokine Growth Factor Rev. 14: 511-522; Schuh et al., 2003, Cytokine Growth Factor Rev. 2003, 14: 503-510). Атопические заболевания - общее название аллергических заболеваний, которые развиваются при воздействии аллергенов, особенно у людей с генетической предрасположенностью к тому, что они легко подвергаются сенсибилизации к аллергенам. У людей, имеющих такие факторы предрасположенности, легко развивается аномальный иммунный ответ на пищевые антигены и вдыхаемые вещества. Некоторыми конкретными примерами аллергических заболеваний являются бронхиальная астма, атопический дерматит, крапивница, аллергический ринит, аллергический конъюнктивит и аллергический энтерогастрит.
Неоплазма представляет собой аномальный рост ткани, который является неконтролируемым и прогрессирующим. Злокачественную неоплазму часто характеризуют как рак. Рак является второй лидирующей причиной смертности людей и является общим термином более чем 100 заболеваний, характеризуемых аномальной пролиферацией иммортализованных клеток. Одним из механизмов, который вовлечен в живучесть и увеличение количества таких клеток, является механизм, основанный на высвобождении факторов роста, которые действуют через рецепторы и приводят к клеточной пролиферации. Было показано, что среди таких факторов роста GM-CSF является важным фактором роста для нескольких опухолевых клеток. Рецептор хемокина CCR3 был недавно охарактеризован в злокачественных B-лимфоцитах, полученных от пациентов с хроническим лимфолейкозом (CLL) и с лейкозом ворсистых клеток (HCL), (Trentin et al., 2004, Blood, 104, 502-508). Действительно, обнаружено, что трансактивация рецептора эпидермального фактора роста (EGFR) посредством рецептора хемокина CCR-3 является важным путем, который вызывает активацию MAP-киназ и продукцию цитокинов в эпителиальных клетках бронхов (Adachi et al., 2004, Biochem. Biophys. Res. Commun. 320, 292-396). Ингибирование пролиферации злокачественных клеток блокированием рецепторов для факторов роста и/или для хемокинов может иметь важной значение для терапии некоторых видов рака.
Эозинофилы представляют собой тип белой клетки крови. Они являются гранулированными лейкоцитами с ядром, которое обычно имеет две доли, соединенные тонкой нитью хроматина, и цитоплазмой, содержащей крупные круглые гранулы, которые единообразны по размеру и окрашиваются эозином. Гиперэозинофилия характеризуется повышенным количеством эозинофилов, часто связанным с аллергенами, астмой и инфекциями.
Известны некоторые применения олигонуклеотидов, направленных против конкретных последовательностей нуклеиновых кислот, кодирующих рецепторы, для ингибирования воспалительных реакций. В заявке PCT № WO 99/66037 Renzi описывает антисмысловые олигонуклеотиды, которые применяют для лечения и/или профилактики астмы, аллергии, гиперэозинофилии, генерализованного воспаления и рака. В частности, олигонуклеотиды согласно Renzi направлены против последовательностей нуклеиновых кислот, кодирующих рецептор CCR3, общую субъединицу рецепторов IL-4 и IL-3 или общую субъединицу рецепторов IL-3, IL-5 и GM-CSF. Наряду с прочими в данной публикации описан антисмысловой олигонуклеотид, обозначенный как 107A (5'-GGGTCTGCAGCGGGATGGT-3'), направленный против общей бета (β)-субъединицы рецепторов IL-3, IL-5 и GM-CSF.
Для возможных клинических применений антисмысловые олигонуклеотиды должны обладать стабильностью по отношению к разрушению нуклеазами сыворотки и клеточными нуклеазами, иметь низкое неспецифичное связывание с белками сыворотки и клеток, проявлять повышенную способность узнавать последовательность мРНК-мишени, обладать проницаемостью через клеточные мембраны и активировать клеточные нуклеазы при образовании комплекса с комплементарной мРНК. Документально хорошо подтверждено, что олигонуклеотиды, содержащие природные сахара (D-рибозу и D-2-дезоксирибозу) и фосфодиэфирные (PO) связи, быстро разрушаются нуклеазами сыворотки и внутриклеточными нуклеазами, что ограничивает их применимость в качестве эффективных терапевтических средств. Описаны важные химические модификации олигонуклеотидов для повышения их стабильности и эффективности в качестве терапевтических средств. Основные химические изменения включали модификацию остатка сахара, остатка основания и/или модификацию или замену межнуклеотидной фосфодиэфирной связи. До настоящего времени наиболее широко исследованными аналогами являются фосфоротиоатные (PS) олигодезоксинуклеотиды, в которых один из атомов кислорода, не образующих мостик, в фосфодиэфирном остове заменен атомом серы (Eckstein F., 1985, Ann. Rev. Biochem., 54:367-402). Разработано несколько поколений антисмысловых олигонуклеотидов и использовано для исследований in vitro и in vivo (Goodchild J., 2004, Curr. Opin. Mol. Ther., 2004, 6:120-128; Urban E. and R. Noe CR., 2003, Farmaco. 58:243-258). Недавно Renzi et al. описал применение 2',6'-диаминопурина (DAP) и их аналогов в молекулах нуклеиновых кислот для противовоспалительных композиций (заявка PCT № WO 03/004511 A2). Также в указанной публикации описано получение молекул нуклеиновых кислот, имеющих повышенную физиологическую эффективность in vivo и пониженную токсичность по сравнению с олигонуклеотидами без DAP. Renzi et al. Кроме того описывают, что DAP-замена особенно применима для получения олигонуклеотидов, направленных на лечение легочных/респираторных заболеваний, таких как кистозный фиброз, астма, хронический бронхит, хроническое обструктивное легочное заболевание, эозинофильный бронхит, аллергии, аллергический ринит, фиброз легких, респираторный дистресс-синдром легких, синусит, инфекция респираторным синцитиальным вирусом или другая вирусная инфекция дыхательных путей и рак.
Кроме того, было бы желательно иметь антисмысловые олигонуклеотиды, направленные по меньшей мере против одного специфичного общего рецептора для любого Th2-цитокина или рецепторов для медиаторов, которые привлекают клетки, которые отвечают на Th2-цитокины, чтобы ингибировать воспалительную реакцию, которая имеет место при астме или аллергии, и ингибировать пролиферацию неопластических клеток.
Также было бы крайне желательно иметь антисмысловые олигонуклеотиды, направленные против последовательностей нуклеиновых кислот, кодирующих рецепторы, для того чтобы благодаря ингибированию таких рецепторов указанные олигонуклеотиды можно было бы применять для терапии и/или профилактики астмы, аллергии, гиперэозинофилии, генерализованного воспаления и рака.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к применению антисмысловых олигонуклеотидов, направленных по меньшей мере против одной общей субъединицы клеточного рецептора, такой, например, как общая субъединица бета-рецепторов IL-3, IL-5 и GM-CSF, или рецептора хемокина CCR3, для лечения и/или профилактики по меньшей мере одного из заболеваний: астмы, аллергии, гиперэозинофилии, генерализованного воспаления и рака.
В другом аспекте настоящее изобретение относится к антисмысловым олигонуклеотидам, направленным против последовательности нуклеиновой кислоты, кодирующей общую субъединицу бета-рецепторов IL-3, IL-5 и GM-CSF, с тем чтобы благодаря ингибированию таких рецепторов их можно было применять для лечения и/или профилактики по меньшей мере одного из заболеваний: астмы, аллергии, гиперэозинофилии, генерализованного воспаления и рака.
Настоящее изобретение также относится к антисмысловым олигонуклеотидам, направленным против последовательности нуклеиновой кислоты, кодирующей рецептор CCR3 для хемокинов, с тем чтобы благодаря ингибированию такого рецептора их можно было применять для лечения и/или профилактики по меньшей мере одного из заболеваний: астмы, аллергии, гиперэозинофилии, генерализованного воспаления и рака.
Настоящее изобретение также относится к терапевтически эффективным композициям, содержащим по меньшей мере один антисмысловой олигонуклеотид, направленный против последовательностей нуклеиновой кислоты, кодирующих общую бета-субъединицу рецепторов IL-3, IL-5 и GM-CSF или рецептор CCR3, для лечения и/или профилактики по меньшей мере одного из заболеваний: астмы, аллергии, гиперэозинофилии, генерализованного воспаления и рака.
Настоящее изобретение также относится к терапевтически эффективным композициям, содержащим два антисмысловых олигонуклеотида, каждый из которых направлен против последовательностей нуклеиновой кислоты, кодирующих общую бета-субъединицу рецепторов IL-3, IL-5 и GM-CSF или рецептор CCR3, для улучшенного эффекта при лечении и/или профилактике по меньшей мере одного из заболеваний: астмы, аллергии, гиперэозинофилии, генерализованного воспаления и рака.
Согласно другому аспекту настоящее изобретение относится к способам лечения и/или профилактики по меньшей мере одного из заболеваний: астмы, аллергии, генерализованного воспаления и рака, включающим в себя введение одного или нескольких антисмысловых олигонуклеотидов, направленных против по меньшей мере одной общей субъединицы клеточного рецептора, такой как общая бета-субъединица IL-3, IL-5 и GM-CSF, или рецептора CCR3.
В настоящем изобретении предпринята попытка получить антисмысловые олигонуклеотиды для любого из указанных выше применений, а также химически модифицированные антисмысловые олигонуклеотиды, модифицированные известными способами, которые имеют повышенную стабильность в организме, проявляя при этом повышенную эффективность и пониженную токсичность.
Согласно другому аспекту настоящего изобретения предлагается антисмысловой олигонуклеотид для лечения и/или профилактики по меньшей мере одного из заболеваний: астмы, аллергии, гиперэозинофилии, генерализованного воспаления и рака. Олигонуклеотид направлен против последовательности нуклеиновой кислоты, кодирующей рецептор, выбранный из группы, состоящей из рецептора хемокина CCR3 и общей бета-субъединицы рецепторов IL-3, IL-5 и GM-CSF, и имеет последовательность, выбранную из группы, состоящей из последовательностей SEQ ID NO. 1, SEQ ID NO. 13 и SEQ ID NO. 14.
Согласно другому аспекту изобретения предлагается применение по меньшей мере одного олигонуклеотида для лечения и/или профилактики по меньшей мере одного из заболеваний: астмы, аллергии, гиперэозинофилии, генерализованного воспаления и рака. Предпочтительно применяют олигонуклеотиды, содержащие обе последовательности SEQ ID NO. 13 и SEQ ID NO. 14.
Согласно другому аспекту изобретения предлагается фармацевтическая композиция для лечения и/или профилактики по меньшей мере одного из заболеваний: астмы, аллергии, гиперэозинофилии, генерализованного воспаления и рака, содержащая по меньшей мере один олигонуклеотид вместе с фармацевтически приемлемым носителем. Предпочтительно по меньшей мере один олигонуклеотид содержит обе последовательности SEQ ID NO. 13 и SEQ ID NO. 14.
Согласно другому аспекту изобретения предлагается применение фармацевтической композиции для лечения и/или профилактики по меньшей мере одного из заболеваний: астмы, аллергии, гиперэозинофилии, генерализованного воспаления и рака.
Согласно другому аспекту изобретения предлагается способ лечения и/или профилактики по меньшей мере одного из заболеваний: астмы, аллергии, гиперэозинофилии, генерализованного воспаления и рака, включающий в себя стадию введения эффективного количества (i) по меньшей мере одного олигонуклеотида или (ii) фармацевтической композиции, содержащей по меньшей мере один олигонуклеотид вместе с фармацевтически приемлемым носителем.
Настоящее изобретение также относится к модификациям антисмыслового олигонуклеотида(ов), которые не оказывают значимого неблагоприятного влияния на их способность снижать активность или ингибировать экспрессию белка-мишени, но которые могут усиливать такую способность.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Указанные и другие отличительные признаки предпочтительных вариантов осуществления изобретения станут более понятными в следующем подробном описании, в котором приведены ссылки на прилагаемые чертежи, где:
на фигуре 1A показано выравнивание последовательностей трех клонов, полученных в результате ПЦР-амплификации генов общей бета-цепи рецепторов IL-3, IL-5 и GM-CSF макак-крабоедов (Cynomolgus) с соответствующими ортологами человека, шимпанзе, свиньи, крысы и мыши, окружающими комплементарную последовательность TOP004 человека.
На фигуре 1B показаны рассчитанные аминокислотные последовательности транслируемой области, окружающей комплементарный TOP004 участок, в клонированных последовательностях ДНК общей бета-цепи макак-крабоедов, человека, шимпанзе, свиньи, крысы и мыши.
На фигуре 2A показана пониженная экспрессия мРНК бета-цепи (βc) при различных концентрациях TOP004 в PBMC макак-крабоедов по сравнению с необработанными клетками.
На фигуре 2B показана пониженная экспрессия мРНК CCR3 при различных концентрациях TOP005 в PBMC макак-крабоедов по сравнению с необработанными клетками.
На фигуре 3A показана пониженная экспрессия белка клеточной поверхности бета-цепи (βc) при различных концентрациях TOP004 в PBMC макак-крабоедов по сравнению с необработанными клетками.
На фигуре 3B показана пониженная экспрессия белка клеточной поверхности CCR3 при различных концентрациях TOP005 в PBMC макак-крабоедов по сравнению с необработанными клетками.
На фигуре 4A показана пониженная экспрессия мРНК бета-цепи (βc) при различных концентрациях ASM8 в PBMC макак-крабоедов по сравнению с необработанными клетками.
На фигуре 4B показана пониженная экспрессия мРНК CCR3 при различных концентрациях ASM8 в PBMC макак-крабоедов по сравнению с необработанными клетками.
На фигуре 5 показано влияние олигонуклеотидов на экспрессию мРНК CCR3 в дифференцированных клетках HL60. Показано, что антисмысловой олигонуклеотид A86, направленный против CCR3, снижает экспрессию мРНК CCR3 по сравнению с контролем и смысловыми олигонуклеотидами, не влияя на экспрессию G3PDH.
На фигуре 6 показан анализ мобилизации кальция в обработанных олигонуклеотидом клетках HL-60 cl-15. Снижение мобилизации в ответ на эотаксин в клетках, обработанных A86, сравнивают с клетками в контроле и клетками, обработанными смысловыми олигонуклеотидами.
На фигуре 7 показано влияние олигонуклеотидов на хемотаксический ответ очищенных эозинофилов человека на эотаксин. Относительный хемотаксический ответ обработанных A86 эозинофилов сравнивают с контрольными и обработанными смысловым олигонуклеотидом клетками.
На фигуре 8 показан анализ мобилизации кальция в обработанных олигонуклеотидом эозинофилах в ответ на эотаксин. Мобилизацию кальция сравнивают в эозинофилах, обработанных A86, и в контрольных или обработанных смысловым олигонуклеотидом клетках.
На фигуре 9 показано влияние TOP005 на экспрессию на клеточной поверхности CCR3, представленное в виде процента экспрессии по сравнению с контролями в клетках Eol-I и U937.
На фигуре 10 показано влияние TOP005 на экспрессию мРНК в PBMC человека. Гели, на которых представлена экспрессия G3PDH и CCR3, показаны над гистограммой. Отношение экспрессии мРНК CCR3 к G3PDH, нормализованное по отношению к контролям, представлено снизу.
На фигуре 11A показано модулирование экспрессии мРНК бета-цепи (βc) в обработанных 107A клетках TF-1 с использованием ОТ-ПЦР для выявления экспрессии мРНК бета-цепи (βc) или контрольной мРНК G3PDH.
На фигурах 11B и 11C показано влияние обработки смысловым олигонуклеотидом и 107A на экспрессию бета-цепи на клеточной поверхности клеток TF-1, которое определяли FACS-анализом.
На фигуре 11D показано влияние TOP004 на экспрессию общей бета-цепи на уровне мРНК и белка в клетках U937.
На фигуре 12 показана пролиферация клеток TF-1, обработанных 107A в присутствии GM-CSF, IL-3 или IL-5.
На фигуре 13A показано модулирование жизнеспособности эозинофилом посредством 107A, оцениваемое с использованием анализа на основе исключения при окрашивании трипановым синим.
На фигуре 13B показано модулирование жизнеспособности эозинофилов посредством 107A, которое оценивали в анализе проточной цитометрией, используя протокол с применением аннексин-V-ФИТЦ и йодида пропидия.
На фигуре 14 показан профиль элюирования отдельных продуктов ASM8 (TOP004 и TOP005) при использовании DEAE-анионообменной хроматографии.
На фигуре 15 показан профиль элюирования ASM8 после обработки CH3COOH в течение 3 часов и после того, как его подвергли щелочному лизису перед фракционированием DEAE-анионообменной хроматографией.
На фигуре 16 показан профиль элюирования ASM8 после обработки CH3COOH в течение 6 часов и после того, как его подвергли щелочному лизису перед фракционированием DEAE-анионообменной хроматографией.
На фигурах 17A1, 17A2, 17B1 и 17B2 показана химическая стабильность ASM8 после хранения при разных температурах и последующего элюирования с использованием DEAE-анионообменной хроматографии.
На фигуре 18 показаны кривые плавления TOP004 и TOP005 в 1× PBS.
На фигуре 19 показаны сводные термодинамические данные, основанные на результатах подгонки кривых плавления TOP004 и TOP005 в 1×PBS.
На фигурах 20A и 20B показаны концентрации TOP004 и TOP005 и их метаболитов в плазме обезьян в 1 день после обработки высокой дозой ASM8.
На фигурах 21A и 21B показаны концентрации TOP004 и TOP005 и их метаболитов в плазме обезьян на 14 день после обработки высокой дозой ASM8.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Несколько медиаторов воспаления играют роль в появлении и сохранении воспаления в дыхательных путях пациентов с астмой. Некоторые медиаторы привлекают воспалительные клетки в дыхательные пути посредством хемотаксиса эозинофилов. Многие такие хемокины главным образом действуют при астматическом или аллергическом воспалении через рецептор CCR3. Другие медиаторы вызывают примирование и повышенную жизнеспособность воспалительных клеток в дыхательных путях или коже, такие как IL-3, IL-5 и GM-CSF. Улучшение при астме было показано в случае снижения указанных медиаторов воспаления в дыхательных путях.
Кроме того, рак, характеризуемый аномальной пролиферацией иммортализованных клеток, может быть вызван высвобождением медиаторов воспаления и/или факторов роста, которые действуют через рецепторы и приводят к пролиферации клеток. Из них, как было показано, GM-CSF является важным фактором роста для некоторых опухолевых клеток. Рецептор хемокинов CCR3 характеризовали в злокачественных B-лимфоцитах, извлеченных из организма пациентов с хроническим лимфолейкозом (CLL) и с лейкозом ворсистых клеток (HCL) (Trentin et al., 2004, Blood, 104, 502-508). Действительно, обнаружено, что трансактивация EGFR посредством CCR3 является критическим путем, который вызывает активацию MAP-киназ и продукцию цитокинов в эпителиальных клетках бронхов (Adachi et al., 2004, Biochem. Biophys. Res. Commun. 320, 292-396). Ингибирование пролиферации и метастазов злокачественных клеток при блокировании рецепторов факторов роста или рецептора хемокина CCR3 может иметь важное значение для терапии некоторых видов рака.
В одном варианте изобретения предлагается новый антисмысловой олигонуклеотид, обозначенный как 828 (5'-GTTACTACTTCCACCTGCCTG-3' (SEQ ID NO. 1)) и направленный против рецептора хемокина CCR3. Примеры, описанные в данной публикации, показывают, что 828 является эффективным в снижении или блокировании экспрессии мРНК CCR3 в линиях клеток человека.
В другом варианте изобретения предлагаются новые антисмысловые олигонуклеотиды TOP004 и TOP005, основанные на описанном ранее 107A и указанном выше 828. TOP004 (5'-GGGTCTGCXGCGGGXTGGT-3' (SEQ ID NO. 13), где X означает модификацию DAP остатка аденозина), как в случае 107A является 19-мером, направленным против мРНК общей бета (β)-цепи рецепторов IL-3, IL-5 и GM-CSF. TOP005 (5'-GTTXCTXCTTCCXCCTGC CTG-3' (SEQ ID NO. 14), где X означает модификацию DAP остатка аденозина), как в случае 828 является 21-мером, направленным против мРНК рецептора хемокина CCR3. Композиция, содержащая оба олигонуклеотида TOP004 и TOP005, идентифицирована как часть ASM8.
Как описано в данной публикации, TOP004 и TOP005 обладают активностью в системе примата, отличного от человека, таким образом подтверждая возможность применения макак-крабоедов для оценки безопасности. На фигуре показано секвенирование гена общей бета-цепи макаки-крабоеда. Последовательность бета-цепи макаки-крабоеда, комплементарная TOP004, имела значительную гомологию. Очень высокая степень идентичности между последовательностями бета-цепи обезьян и человека свидетельствует о возможной функциональной активности TOP004 у макак-крабоедов. Эффективность TOP004 и TOP005 в блокировании или снижении экспрессии общей бета-цепи и CCR3 в мононуклеарных клетках периферической крови обезьян показана на фигурах 2, 3, и 4. Результаты показывают, что оба олигонуклеотида TOP004 и TOP005, направленные против генов-мишеней человека, являются эффективными в снижении экспрессии соответствующих им мишеней в мононуклеарных клетках периферической крови (PBMC) обезьян. ASM8, содержащий оба олигонуклеотида TOP004 и TOP005, значимо ингибировал экспрессию общей бета-цепи и рецепторов CCR3 либо в большей степени, либо в такой же степени при более низкой концентрации. Следовательно, TOP004 и TOP005 вместе проявляют синергическое действие в блокировании экспрессии мРНК бета-цепи и CCR3. Кроме того, как показано в таблицах 7 и 8, образцы трахеи, взятые от обезьян, обработанных ASM8, анализировали в отношении уровня экспрессии мРНК. Экспрессию генов-мишеней нормализовали относительно уровней мРНК воспалительных цитокинов (IL-4 и TNF-α). Даже примерно через 24 часа после введения ASM8 относительная экспрессия мРНК βc-субъединицы и CCR3 по отношению к мРНК IL-4 была снижена на 29% и 24%, соответственно, и экспрессия относительно TNF-α была снижена на 30% и 24%, соответственно, у животных, обработанных ASM8.
Как показано на фигурах 5-10, антисмысловые олигонуклеотиды, включая A86 и TOP005, направленные против мРНК CCR3, тестировали в отношении эффективности в клетках человека и линиях клеток. При оценке посредством полуколичественной полимеразной цепной реакции с обратной транскрипцией («ОТ-ПЦР») антисмысловые олигонуклеотиды вызывали ингибирование экспрессии мРНК CCR3. Кроме того, используя FACS-анализ, показано, что экспрессия на клеточной поверхности белка CCR3 также ингибировалась при обработке антисмысловыми олигонуклеотидами. Кроме того, функциональное ингибирование CCR3 было подтверждено ингибированием мобилизации кальция
(Ca++) в очищенных эозинофилах после стимуляции эотаксином. Кроме того, в анализе хемотаксиса олигонуклеотиды ингибировали хемотаксис эозинофилов на 55%.
Как показано на фигурах 11-13, антисмысловые олигонуклеотиды 107A и TOP004 использовали для обработки различных клеток. В клетках TF-1, инкубированных с 107A, наблюдали пониженную экспрессию мРНК бета-цепи. 107A также ингибировал пролиферацию клеток TF-1 в присутствии IL-3, IL-5 или GM-CSF. Кроме того, 107A зависимым от дозы образом снижал противоапоптозное действие EL-5 на эозинофилы. В клетках U937, инкубированных с TOP004, наблюдали пониженную экспрессию общей бета-цепи на уровне мРНК и белка. Таким образом, антисмысловые олигонуклеотиды 107A и TOP004 были высокоэффективными в блокировании экспрессии мРНК и белка бета-цепи и функциональными в блокировании связанных клеточных ответов в культурах клеток человека.
На фигурах 14-17B2 показана стабильность ASM8 при элюировании композиции в разных условиях. ASM8 элюировали с использованием системы фракционирования, основанной на DEAE-анионообменной высокоэффективной жидкостной хроматографии (ВЭЖХ), чтобы оценить целостность ASM8 и продукты его разрушения после хранения при разных температурах. Компоненты ASM8 не подвергались какому-либо заметному разрушению при хранении при -20°C, 4°C, 30°C или 40°C в течение периода времени до 2 месяцев.
На фигурах 18-19 кривые плавления и сводные термодинамические данные, приведенные для ASM8, показывают, что две олигонуклеотидных нити значимо не взаимодействуют в растворе.
Как показано на фигурах 20-21, измеряли концентрации олигонуклеотидных компонентов ASM8 и их первичных метаболитов (n-1) в образцах плазмы обезьян. Образцы собирали во время доклинического испытания токсичности, в котором животных обрабатывали в течение 14 последовательных дней путем ингаляции.
Таким образом, предлагаются антисмысловые олигонуклеотиды, направленные против общей бета-субъединицы рецепторов IL-3, IL-5 и GM-CSF и рецептора CCR3 и против кодирующих их нуклеиновых кислот. Также предлагаются фармацевтические композиции, содержащие олигонуклеотиды и фармацевтически приемлемый носитель. Описаны применения олигонуклеотидов и способы, включающие в себя введение олигонуклеотидов для лечения и/или профилактики по меньшей мере одного из заболеваний: астмы, аллергии, гиперэозинофилии, генерализованного воспаления и рака.
Термины «нуклеиновая кислота» и «молекула нуклеиновой кислоты», которые используют в данном описании взаимозаменяемо, относятся к молекуле, состоящей из нуклеотидов, т.е. рибонуклеотидов, дезоксирибонуклеотидов или и тех, и других. Термин включает мономеры и полимеры рибонуклеотидов и дезоксирибонуклеотидов, при этом рибонуклеотиды и/или дезоксирибонуклеотиды связаны вместе в случае полимеров посредством 5'-3'-связи. Однако связи могут включать любую связь, известную в области синтеза нуклеиновых кислот, включая, например, нуклеиновые кислоты, содержащие 5'-2'-связи. Нуклеотиды, используемые в молекуле нуклеиновой кислоты, могут быть встречающимися в природе нуклеотидами или могут представлять собой синтетически полученные аналоги, которые способны образовывать связи пар оснований с встречающимися в природе основаниями. Примеры не встречающихся в природе оснований, которые способны к образованию связей пар оснований, включают без ограничения аза- и деазапиримидиновые аналоги, аза- и деазапуриновые аналоги и другие гетероциклические аналоги оснований, в которых один или несколько атомов углерода и азота пуринового и пиримидинового циклов были заменены гетероатомами, например атомами кислорода, серы, селена, фосфора и тому подобными.
Термин «остов нуклеиновой кислоты», в используемом в данном описании смысле, относится к структуре химического остатка, связывающего нуклеотиды в молекуле. Остов может включать структуры, образованные из любого и всех средств химического связывания нуклеотидов. Модифицированный остов, в используемом в данном описании смысле, включает модификации химического связывания между нуклеотидами, а также другие модификации, которые могут быть использованы для повышения стабильности и аффинности, такие как модификации структуры сахара. Например, можно использовать альфа-аномер дезоксирибозы, когда основание перевернуто по отношению к природному бета-аномеру. В предпочтительном варианте 2'-OH группа сахара может быть изменена на 2'-O-алкил или 2'-O-алкил-n(O-алкил), который придает резистентность к разрушению, не затрагивая аффинность.
Термин «олигонуклеотид», в используемом в данном описании смысле, относится к молекуле нуклеиновой кислоты, содержащей примерно от 1 до примерно 100 нуклеотидов, более предпочтительно от 1 до 80 нуклеотидов и еще более предпочтительно примерно от 4 до примерно 35 нуклеотидов.
Антисмысловые олигонуклеотидные соединения согласно настоящему изобретению также включают миРНК (малые интерферирующие РНК) и содержащие их RISC (РНК-индуцируемые комплексы репрессии), которые возникают при РНК-и (РНК-интерференции). Методика интерференция РНК (РНК-и), которая была недавно описана, считается новым средством ингибирования экспрессии гена-мишени. Как стало известно несколько лет назад, РНК-и основана на древнем механизме противовирусной защиты у низших эукариот. Она индуцируется двунитевой РНК и ее процессингом до состоящих из 21-23 нуклеотидов малых интерферирующих РНК (миРНК), которые вызывают разрушение гомологичной эндогенной мРНК после гибридизации с мРНК-мишенью в однонитевой форме с помощью комплекса RISC. Работу пути РНК-и еще предстоит более полно выяснить, но он уже служит в качестве наиболее предпочтительного подхода к созданию фенотипов с потерей функции среди широкого множества видов эукариот, таких как нематоды, крылатые насекомые, растения, грибы и млекопитающие.
Антисмысловые олигонуклеотидные соединения согласно настоящему изобретению также включают рибозимы и короткие нуклеотидные последовательности, одно- и двунитевые, РНК или ДНК, которые могут содержать химические модификации, которые описаны выше, способные ингибировать транскрипцию гена и/или трансляцию in vitro и/или in vivo.
Термин «модифицированный олигонуклеотид» и «модифицированная молекула нуклеиновой кислоты» включает антисмысловые олигонуклеотидные соединения, которые были модифицированы без существенного неблагоприятного влияния на их активность, например, посредством инсерции или делеции 1 или нескольких оснований. В частности, добавление или делеция оснований на концах олигонуклеотидов, которые на 100% комплементарны гену, против которого они направлены, в общем могут быть осуществлены без существенной потери ингибирующей активности. Такие модификации могут быть осуществлены для повышения активности или обеспечения повышенной стабильности олигонуклеотида. Кроме того, замена 1 или нескольких оснований в антисмысловых олигонуклеотидных соединениях согласно изобретению также может быть осуществлена без неблагоприятного влияния на активность, например замена пурина другим пурином (аденин, гуанин) и пиримидина пиримидином (цитозин, тимин, урацил). Модифицированный олигонуклеотид и модифицированная молекула нуклеиновой кислоты в используемом в данном описании смысле также включают нуклеиновые кислоты, в том числе олигонуклеотиды, с одной или несколькими химическими модификациями на молекулярном уровне в природных молекулярных структурах всех или любого из оснований нуклеиновых кислот, остатков сахаров, межнуклеозидных фосфатных связей, а также молекулы, имеющие добавленные заместители, такие как диамины, холестерил или другие липофильные группы, или комбинацию модификаций в указанных местах. Межнуклеозидные фосфатные связи могут быть фосфодиэфирными, фосфотриэфирными, фосфорамидатными, силоксановыми, карбонатными, карбоксиметилэфирными, ацетамидатными, карбаматными, тиоэфирными, мостиковыми фосфоранидатными, мостиковыми метиленфосфонатными, фосфоротиоатными, метилфосфонатными, фосфородитиоатными, мостиковыми фосфоротиоатными и/или сульфоновыми межнуклеотидными связями или 3'-3'-, 2'-5'- или 5'-5'-связями и комбинациями таких сходных связей (для получения модифицированных олигонуклеотидов со смешанным остовом). Модификации могут быть внутренними (единичными или повторяющимися) или на конце (концах) молекулы олигонуклеотида могут включать в себя добавления к молекуле межнуклеозидных фосфатных связей, например холестерила, диаминовых соединений с разным количеством атомов углерода между аминогруппами и концевые модификации рибозы, дезоксирибозы и фосфата, которые прилипают или поперечно связываются с противоположными цепями или со связанными ферментами или другими белками. Электрофильные группы, такие как рибоза-диальдегид, могут быть ковалентно связаны с эпсилон-аминогруппой остатка лизила такого белка. Нуклеофильная группа, такая как n-этилмалеимид, связанная с олигомером, может ковалентно связываться с 5'-концом мРНК или с другим электрофильным участком. Термин "модифицированные олигонуклеотиды" также включает олигонуклеотиды, содержащие модификации в остатках сахара, такие как 2'-замещенные рибонуклеотидные или дезоксирибонуклеотидные мономеры, любые из которых связываются вместе посредством 5'-3'-связей. Модифицированные олигонуклеотиды также могут состоять из PNA-модифицированных или модифицированных морфолином остовов, в которых сохраняется целевая специфичность последовательности. Термин "модифицированные олигонуклеотиды" также включает олигонуклеотидные соединения, которые определены в данном описании, в форме, которая существенно не оказывает неблагоприятного влияния на их активность в снижении активности или ингибировании экспрессии белка-мишени, но которая усиливает такую активность.
Модифицированные олигонуклеотиды также включают олигонуклеотиды, которые основаны или сконструированы из остатком арабинонуклеотидов или модифицированных остатков арабинонуклеотидов, включая без ограничения, антисмысловые олигонуклеотидные конструкции, основанные на бета-арабинофуранозе и ее аналогах. Арабинонуклеозиды являются стереоизомерами рибонуклеозидов, отличающимися только по конфигурации в 2'-положении цикла сахара. В заявке PCT № WO 99/67378 Damha et al. (1), которая включена в данное описании в виде ссылки в полном объеме, описаны олигомеры арабинонуклеиновых кислот (ANA) и их аналоги для улучшенного специфичного для последовательности ингибирования экспрессии гена посредством связывания с комплементарной матричной РНК. Dahma et al., кроме того, описывают олигонуклеотиды с модифицированным сахаром, которые образуют дуплекс с последовательностью РНК-мишени, приводя к получению субстрата для РНКазы H. В частности, описаны олигомеры, содержащие бета-D-арабинонуклеотиды и 2'-дезокси-2'-фтор-бета-D-арабинонуклеозиды. В заявке PCT № WO 02/20773 тех же авторов Dahma et al. (2), которая включена в данное описание в виде ссылки в полном объеме, описаны олигонуклеотидные химеры, используемые для ингибирования транскрипции и экспрессии гена зависимым от последовательности образом. В частности, Dahma et al. (2) описывают антисмысловые олигонуклеотиды, сконструированные из арабинонуклеотидов, фланкирующих ряд содержащих дезоксирибозу нуклеотидных остатков различной длины. Антисмысловые олигонуклеотиды, сконструированные таким образом, используют для гибридизации и индукции расщепления комплементарной РНК. В заявке PCT №WO 03/037909 также Dahma et al. (3), которая включена в данное описание в виде ссылки в полном объеме, описаны олигонуклеотиды, имеющие внутренний ациклический линкерный остаток. Антисмысловые олигонуклеотиды, полученные с ациклическим линкером, используют для предотвращения или снижения функции представляющей интерес нуклеиновой кислоты-мишени, такой как РНК. В заявке PCT № WO 03/064441 также Dahma et al. (4), которая включена в данное описание в виде ссылки в полном объеме, описаны олигонуклеотиды, имеющие чередующиеся участки нуклеозидов с модифицированным сахаром и 2'-дезоксинуклеозидов, а также чередующиеся участки нуклеотидов с модифицированным сахаром и 2'-дезоксинуклеотидов. Антисмысловые олигонуклеотиды, имеющие указанные чередующиеся участки, описаны для применения для предотвращения или снижения функции представляющей интерес нуклеиновой кислоты-мишени, такой как РНК.
Термин «в значительной степени резистентные к нуклеазам» относится к нуклеиновым кислотам, которые резистентны к разрушению нуклеазами по сравнению с встречающимися в природе или немодифицированными нуклеиновыми кислотами. Модифицированные нуклеиновые кислоты согласно изобретению по меньшей мере в 1,25 раз более резистентны к разрушению нуклеазами, чем их немодифицированный аналог, более предпочтительно по меньшей мере в 2 раза более резистентны, еще более предпочтительно по меньшей мере в 5 раз более резистентны и наиболее предпочтительно по меньшей мере в 10 раз более резистентны, чем их немодифицированный аналог. Такие в значительной степени резистентные к нуклеазам нуклеиновые кислоты включают без ограничения нуклеиновые кислоты с модифицированными остовами, такими как фосфоротиоаты, метилфосфонаты, этилфосфотриэфиры, 2'-O-метилфосфоротиоаты, 2'-O-метил-п-этоксирибонуклеотиды, 2'-O-алкилы, 2'-O-алкил-n(O-алкил), 3'-O-алкилы, 3'-O-алкил-n(O-алкил), 2'-фторзамещенные, 2'-дезоксиэритропентофуранозилы, 2'-O-метилрибонуклеозиды, метилкарбаматы, метилкабонаты, инвертированные основания (например, инвертированные T) или химерные варианты указанных остовов.
Термины «антисмысловые олигонуклеотиды рецептора CCR3 и общей бета-цепи рецепторов IL-3/IL-5/GM-CSF», в используемом в данном описании смысле, относятся к олигонуклеотидам, мишенями которых соответственно являются последовательности, которые влияют на экспрессию и/или активность рецептора хемокина CCR3 и общей бета-цепи рецепторов IL-3/IL-5/GM-CSF. Такие последовательности включают без ограничения рецептор хемокина CCR3 и общую бета-цепь рецепторов IL-3/IL-5/GM-CSF, кодирующие последовательности ДНК, промоторные последовательности ДНК, энхансерные последовательности ДНК, кодирующие последовательности мРНК и тому подобные.
Как обсуждалось выше, один вариант осуществления настоящего изобретения относится к антисмысловым олигонуклеотидам, мишенями которых являются последовательности, которые влияют на экспрессию и/или активность рецептора хемокина CCR3 и общей бета-цепи рецепторов IL-3/IL-5/GM-CSF. В одном варианте антисмысловой олигонуклеотид может содержать фрагменты или варианты таких последовательностей, как будет понятно специалисту в данной области, которые могут изменять структуру и длину олигонуклеотида, но которые сохраняют или увеличивают активность олигонуклеотида в понижающей регуляции экспрессии гена. В другом варианте настоящее изобретение относится к комбинациям по меньшей мере двух антисмысловых олигонуклеотидов из последовательностей, обозначенных как SEQ ID NO. 1, SEQ ID NO. 13 и SEQ ID NO. 14.
Термины «лечение», «проведение лечения», «терапия» и тому подобные используют в данном описании, чтобы в общем описать получение требуемого фармакологического и/или физиологического эффекта. Эффект может быть профилактическим в смысле полного или частичного предотвращения заболевания или его симптома и/или может быть терапевтическим в смысле частичного или полного излечения заболевания и/или ослабления неблагоприятного эффекта, вызванного заболеванием. «Лечение», в используемом в данном описании смысле, охватывает любое лечение заболеваний у субъекта, которые определены ранее, в частности у человека и включает:
(a) предотвращение возникновения заболевания у субъекта, который может быть предрасположен к заболеванию, но у которого еще не диагностировано наличие заболевания;
(b) подавление заболевания, т.е. задержка его развития; или
(c) ослабление заболевания, т.е. вызывание регрессии заболевания.
Термин «фармацевтически приемлемый» в используемом в данном описании смысле по отношению к носителям, поверхностно-активным веществам и композициям относится к веществам, которые допустимы для применения при лечении пациента, которые не являются токсичными или неприемлемыми по другим причинам для введения любым из путей, описанных в данном описании.
Изобретение в общем направлено на лечение субъектов посредством введения терапевтически эффективных количеств антисмысловых олигонуклеотидных соединений согласно настоящему изобретению, включая миРНК, рибозимы, короткие нуклеотидные последовательности в виде однонитевых или двунитевых молекул, включая РНК и/или ДНК, которые могут быть комплементарны нуклеиновой кислоте-мишени или могут быть необязательно модифицированы, как описано выше, РНК-олигонуклеотид, имеющий по меньшей мере часть указанного РНК-олигонуклеотида, способную гибридизоваться с РНК с образованием дуплекса олигонуклеотид-РНК, или химерный олигонуклеотид, который будет осуществлять понижающую регуляцию или ингибировать экспрессию эндогенного гена in vivo.
Под «терапевтически эффективным» количеством подразумевают нетоксичное, но достаточное количество антисмыслового олигонуклеотидного соединения для получения требуемого терапевтического эффекта. В данном случае такая доза антисмыслового олигонуклеотидного соединения эффективна для облегчения, ослабления или предотвращения симптомов состояния или заболевания, подвергаемого лечению, например заболевания, связанного с аллергией, астмой, воспалительным заболеванием, таким как воспалительное респираторное заболевание.
Термин «аллергия» в используемом в данном описании смысле описывает любой нежелательный иммунный ответ организма на вещество, к которому он стал гиперчувствительным.
Препараты согласно настоящему изобретению предпочтительно вводят непосредственно в место действия и, следовательно, предпочтительно являются препаратами для местного применения, включая без ограничения пероральные, буккальные, внутрилегочные, ректальные, внутриматочные, внутриопухолевые, назальные, интратекальные, ингаляционные, трансдермальные, внутридермальные, внутриполостные, ионтофоретические, глазные, вагинальные, внутрисуставные, ушные, трансмукозальные, ректальные, препараты медленного высвобождения или препараты с кишечнорастворимым покрытием. Не ограничиваясь каким-либо препаратом из вышеперечисленных, препараты согласно настоящему изобретению также могут быть внутричерепными, внутримышечными, подкожными, внутрисосудистыми, интрагландулярными, внутриорганными, внутрилимфатическими, внутрибрюшинными, внутривенными и имплантируемыми. Носители, используемые в препаратах, могут представлять собой, например, твердые и/или жидкие носители.
Можно сослаться на «Remington's Pharmaceutical Sciences», 17th Ed., Mack Publishing Company, Easton, Pa., 1985, в отношении других носителей, которые могут быть подходящими для комбинирования с олигонуклеотидными соединениями согласно настоящему изобретению, чтобы сделать композиции/препараты подходящими для введения при лечении респираторного заболевания.
Необязательно, описанные в данной публикации олигонуклеотиды могут быть приготовлены в виде композиции с использованием множества физиологических молекул носителей. Описанные в данной публикации олигонуклеотиды также могут быть в комплексе с молекулами, которые усиливают их способность проникать в клетки-мишени. Примеры таких молекул включают без ограничения углеводы, полиамины, аминокислоты, пептиды, липиды и молекулы, необходимые для роста клеток. Например, олигонуклеотиды могут быть объединены с липидом, при этом полученная в результате эмульсия олигонуклеотид/липид или суспензия липосом наряду с прочим может эффективно увеличивать время полужизни олигонуклеотида in vivo.
Фармацевтические композиции, предлагаемые в данном изобретении, могут содержать антисмысловые олигонуклеотидные соединения, описанные выше, и одно или несколько фармацевтически приемлемых поверхностно-активных веществ. Подходящие поверхностно-активные вещества или поверхностно-активные компоненты для усиления поглощения антисмысловых олигонуклеотидов согласно изобретению ранее описаны в публикации заявки США №2003/0087845, содержание которой включено в данное описание по отношению к поверхностно-активным веществам. В заявке указано, что подходящие поверхностно-активные вещества «… включают наряду с прочим синтетические и природные, а также полные и укороченные формы поверхностно-активного белка A, поверхностно-активного белка B, поверхностно-активного белка C, поверхностно-активного белка D и поверхностно-активного белка E, динасыщенный фосфатидилхолин (отличный от дипальмитоила), дипальмитоилфосфатидилхолин, фосфатидилхолин, фосфатидилглицерин, фосфатидилинозитол, фосфатидилэтаноламин, фосфатидилсерин; фосфатидную кислоту, убихиноны, лизофосфатидилэтаноламин, лизофосфатидилхолин, пальмитоиллизофосфатидилхолин, дегидроэпиандростерон, долихолы, сульфатидную кислоту, глицерин-3-фосфат, фосфат дигидроацетона, глицерин, глицеро-3-фосфохолин, дигидроацетон, пальмитат, цитидиндифосфат (CDP), диацилглицерин, CDP-холин, холин, холинфосфат; а также природные и искусственные ламеллярные тельца, которые являются природными переносчиками компонентов поверхностно-активного вещества, омега-3-жирные кислоты, полиеновую кислоту, полиеноевую кислоту, лецитин, пальмитиновую кислоту, неионогенные блок-сополимеры этилен- или пропиленоксидов, полиоксипропилен, мономерный и полимерный, полиоксиэтилен, мономерный и полимерный, поли(виниламин) с декстрановыми и/или алканоильными боковыми цепями, Brij 35™, тритон X-100™ и синтетические поверхностно-активные вещества ALEC™, Exosurf™, Survan™ и Atovaquone™. Такие поверхностно-активные вещества можно применять либо отдельно, либо в виде части многокомпонентного поверхностно-активного вещества в препарате, либо в виде ковалентно связанных добавлений на 5'- и/или 3'-концах антисмысловых олигонуклеотидов».
Антисмысловой компонент композиций согласно настоящему изобретению может находиться в фармацевтическом препарате в липидной частице или пузырьке, таких как липосома или микрокристалл. Как описано в патенте США №6025339, липидные частицы могут иметь любую подходящую структуру, например однослойную или многослойную, при условии, что в них содержится антисмысловой олигонуклеотид. Положительно заряженные липиды, такие как N-[1-(2,3-диолеоилокси)пропил]-N,N,N-триметиламмонийэтилсульфат или «DOTAP», являются особенно предпочтительными для таких частиц и пузырьков. Получение таких липидных частиц хорошо известно. Смотри, например, патенты США №4880635, Janoff et al.; 4906477, Kurono et al.; 4911928, Wallach; 4917951, Wallach; 4920016, Allen et al.; 4921757, Wheatley et al.; и т.д.
Композицию согласно изобретению можно вводить любым способом, который доставляет антисмысловое олигонуклеотидное соединение в требуемое место, например, такое как легкие. Антисмысловые соединения, описанные в данной публикации, можно вводить в легкие пациента любым подходящим способом, но предпочтительно вводят путем ингаляции аэрозоля, состоящего из вдыхаемых частиц, которые содержат антисмысловое соединение.
Композиция согласно настоящему изобретению может быть введена в дыхательную систему в виде препарата, содержащего частицы такого размера, которые можно вдыхать, например частицы достаточно небольшого размера, чтобы проходить через нос, рот и гортань при ингаляции и через бронхи и альвеолы легких. В общем, вдыхаемые частицы имеют размер в диапазоне примерно от 0,5 до 10 микрон. Частицы с размером, не позволяющим их вдыхать, которые входят в аэрозоль, имеют тенденцию откладываться в гортани и проглатываться, и, соответственно, количество невдыхаемых частиц в аэрозоле предпочтительно минимизируют. Для назального введения размер частиц в диапазоне 10-500 мкм (микрометров) является предпочтительным для обеспечения удерживания в носовой полости.
Жидкие фармацевтические композиции активного соединения (антисмыслового олигонуклеотидного соединения(ий)) для приготовления аэрозоля могут быть получены посредством объединения антисмыслового соединения с подходящим наполнителем, таким как стерильная апирогенная вода или фосфатно-солевой буфер.
Твердая состоящая из частиц композиция, содержащая антисмысловое соединение, необязательно может содержать диспергирующий агент, который служит для облегчения образования аэрозоля, а также другие терапевтические соединения. Подходящим диспергирующим агентом является лактоза, которая может быть смешана с антисмысловым соединением в любом подходящем соотношении, например в массовом соотношении 1:1.
Антисмысловые композиции могут быть введены в количестве, эффективном против бронхоконстрикции, против аллергии (аллергий) и/или против воспаления, и такое количество зависит от степени заболевания, подвергаемого лечению, состояния пациента, конкретного препарата, пути введения, времени введения субъекту, и т.д. В общем желательны внутриклеточные концентрации олигонуклеотида от 0,05 до 50 мкМ или более конкретно от 0,2 до 5 мкМ. Для введения пациенту-млекопитающему, такому как человек, обычно используют дозу примерно 0,001, 0,01, 0,1 или 1 мг/кг вплоть до примерно 50 или 100 мг/кг или более. Однако также предполагаются и другие дозы. В зависимости от растворимости активного соединения в любом конкретном препарате суточная доза может однократной или может быть разделена на несколько введений дозированных лекарственных форм.
Аэрозоли жидких частиц, содержащие антисмысловое соединение, могут быть получены любым подходящим способом, например с помощью небулайзера. Небулайзеры представляют собой коммерчески доступные устройства, которые преобразуют растворы или суспензии активного ингредиента в терапевтический аэрозольный туман либо посредством ускорения прохождения сжатого газа, обычно воздуха или кислорода, через узкое сопло Вентури, либо посредством перемешивания ультразвуком. Подходящие препараты для применения в неублайзерах содержат активный антисмысловой олигонуклеотидный ингредиент в жидком носителе в количестве до 40% мас./мас., предпочтительно менее 20% мас./мас. препарата. Носителем обычно является вода или разбавленный водный раствор спирта, предпочтительно приготовленный изотоничным жидкостям организма, например добавлением хлорида натрия. Необязательные добавки включают консерванты, если препарат готовят нестерильно, например метилгидроксибензоат, антиоксиданты, противобактериальные средства, корригенты, летучие масла, буферные агенты и эмульгаторы и другие поверхностно-активные вещества для приготовления препаратов.
Аэрозоли твердых частиц, содержащие активное олигонуклеотидное соединение(ия) и фармацевтически приемлемое поверхностно-активное вещество, могут быть получены подобным образом с помощью любого генератора аэрозоля лекарственного средства, состоящего из твердых частиц. Генераторы аэрозоля для введения лекарственных средств в виде твердых частиц субъекту производят частицы, которые являются вдыхаемыми, как объясняется выше, и создают объем аэрозоля, содержащего предварительно определяемую дозу лекарственного средства, со скоростью, подходящей для введения человеку. Активный олигонуклеотидный ингредиент обычно составляет от 0,1 до 100% мас./мас. препарата. Второй тип иллюстративного генератора аэрозоля включает дозирующий ингалятор. Дозирующие ингаляторы представляют собой диспенсеры сжатого аэрозоля, обычно содержащие препарат в виде суспензии или раствора активного ингредиента в сжиженном газе-вытеснителе. Во время применения указанные устройства выпускают препарат через клапан, приспособленный для доставки дозированного объема, обычно от 10 до 150 мкл, чтобы получить тонкодисперсный спрей, содержащий активный ингредиент. Подходящие газы-вытеснители включают некоторые хлорфторуглеродные соединения, например дихлордифторметан, трихлорфторметан, дихлортетрафторэтан или гидрофторалканы и их смеси. Препарат может дополнительно содержать один или несколько сорастворителей, например этанол, эмульгаторы и другие поверхностно-активные вещества для приготовления препаратов, такие как олеиновая кислота или сорбитантриолеат, антиоксиданты и подходящие корригенты.
Аэрозоль, образованный либо из твердых, либо из жидких частиц, может быть получен с помощью генератора аэрозоля со скоростью примерно от 1 до 150 литров в минуту.
В следующем аспекте настоящего изобретения предлагается изделие производства, которое содержит упаковочный материал, внутри которого содержится фармацевтически приемлемая композиция антисмыслового олигонуклеотида, которая является терапевтически эффективной для лечения состояний, связанных с аллергией, астмой, ринитом и воспалительным заболеванием. В одном варианте композиция содержит антисмысловое олигонуклеотидное соединение, которое является эффективным для ингибирования гена рецептора хемокина CCR3 или общей бета-цепи рецепторов для IL-3/IL-5/GM-CSF, при этом указанное олигонуклеотидное соединение является по меньшей мере на 50% комплементарным гену. В другом аспекте композиция содержит по меньшей мере 2 антисмысловых олигонуклеотидных соединения, при этом каждое антисмысловое олигонуклеотидное соединение способно осуществлять понижающую регуляцию гена рецептора хемокина CCR3 и общей бета-цепи рецепторов IL-3/IL-5/GM-CSF, каждое антисмысловое олигонуклеотидное соединение присутствует в концентрации, при которой антисмысловое олигонуклеотидное соединение само по себе практически неэффективно в понижающей регуляции гена, против которого оно направлено, комбинация антисмысловых олигонуклеотидных соединений является эффективной в понижающей регуляции по меньшей мере одного из генов, против которых антисмысловые олигонуклеотиды направлены.
В одном варианте упаковочный материал содержит этикетку, на которой указано, что композицию можно применять для лечения воспалительного респираторного заболевания, и может дополнительно содержаться указание о том, что заболевание представляет собой одно из заболеваний: аллергию, ринит и астму.
В другом варианте упаковочный материал изделия содержит ярлык, на котором указано, что композицию можно применять для лечения воспалительного респираторного заболевания, и может дополнительно содержаться указание, что заболевание представляет собой одно из заболеваний: аллергию, астму, гиперэозинофилию, бронхит, ринит или синусит.
В целях настоящего изобретения упаковочным материалом может быть любой подходящий материал для упаковки содержащей нуклеотиды композиции согласно настоящему изобретению, включая флакон или другую емкость (либо пластиковую, либо стеклянную), картонную коробку, пробирку или другую защитную упаковку. Как будет понятно, упаковка может варьировать в зависимости от природы олигонуклеотидной композиции, например жидкий препарат может быть упакован другим образом, чем аэрозольный препарат.
Настоящее изобретение будет более понятно при обращении к примерам, которые приведены для иллюстрации изобретения, а не для ограничения его объема. Что касается указанных примеров, то использовали следующие способы и материалы.
ПРИМЕРЫ
Материалы и способы
Материалы
Для экспериментов использовали следующие материалы и реагенты: RPMI 1640 (Wisent, № в каталоге 10040 CV); FBS (фетальная сыворотка теленка, Wisent, № в каталоге 80150); пенициллин-стрептомицин (GIBCO, № в каталоге 15140-122); HEPES (Wisent, № в каталоге 26060CI); L-глутамин (Gibco, № в каталоге 25030-081); пируват натрия (Wisent, № в каталоге 25000-Ci); стерильный PBS (GIBCO, № в каталоге 25030-081); сбалансированный солевой раствор Хенкса (HBSS, cellgro, № в каталоге 20021-cv); система синтеза первой нити Superscript для набора ОТ-ПЦР (Invitrogen, № в каталоге 11904-018); dNTP (Invitrogen, № в каталоге 10297-018; олиго-(dT)12-18 (Invitrogen, № в каталоге 11904-018); набор Qiagen RNAeasy Mini Kit (Qiagen, № в каталоге 74106); набор для экстракции из геля Qiagen (Qiagen, № в каталоге 28704); набор для экстракции при ПЦР Qiagen (Qiagen, № в каталоге 28104); β-меркаптоэтанол (Sigma, № в каталоге M-6250); 99% этанол (Commercial alcohols Inc., Brampton, Ontario, Canada); QiaVac 24 Manifold (Qiagen, № в каталоге 19403); одноразовые вакуумные соединительные устройства (Qiagen, № в каталоге 19407); набор ДНКазы I (Fermentas; № в каталоге ENO521); реагент для количественного анализа RiboGreen (Invitrogen-Molecular probes, № в каталоге R-11490); набор для Taq-ПЦР (Qiagen, № в каталоге 201223), набор для окраски Hema-3 (Fisher scientific Co. № в каталоге 122-911, партия №999901); Alamar синий (Biosource № в каталоге DAL1100); пара праймеров CCR3 человека (R&D systems, № в каталоге RDP-209-025); пара праймеров GAPDH человека (R&D systems, № в каталоге RDP-39-025); фикол (Amersham Biosciences; № в каталоге: 17-1440-03); анти-CD16 (набор для очистки эозинофилов, Miltenyi Biotec, Auburn, CA, № в каталоге 130-045-701); rh-эотаксин (Biosource, № в каталоге PMC1434); камера и мембраны для хемотаксиса (NeuroProbe, Nucleopore-Neuroprobe, Cabin John, MD); сывороточный альбумин человека (SIGMA; № в каталоге A9511); антитело против бета-цепи рецепторов IL-3/IL-5/GM-CSF человека (мышиное моноклональное IgG2b; Santa Cruz Biotechnology, № в каталоге sc-457); антитело против IgG2b мыши (моноклональное антитело козы, Alexa Fluor 488, Molecular Probes, № в каталоге A-21141); антитело против CCR3 человека (моноклональное антитело крысы, IgG2a, R&D, № в каталоге MAB155); антитело против IgG крысы (моноклональное антитело козы, Alexa Fluor 633, Molecular Probes, № в каталоге A-21094); rhGM-CSF (R&D systems, № в каталоге 215-GM-005); rhIL-3 (R&D systems, № в каталоге 203-IL-010); rhIL-5 (R&D systems, № в каталоге 205-IL-005), rhIL-2 (R&D systems, № в каталоге 202-IL-010); TOP004-(n-1) (Biosource, олигонуклеотид, содержащий на один нуклеотид меньше на 3'-конце); TOP005-(n-1) (Biosource, олигонуклеотид, содержащий на один нуклеотид меньше на 3'-конце); TOP004-TP (Biosource, Template Probe); TOP004-(n-1-TP-T) (Biosource, зонд матрицы); LP (Biosource, зонд лигирования); TOP005-TP (Biosource); TOP004-(n-1-LP-A, Biosource); покрытые нейтравидином планшеты с высокой связывающей способностью Reacti-Bind (Pierce, № в каталоге 15508); ДНК-лигаза T4 5 ед./мл (Roche, № в каталоге 799 009); анти-DIG-AP-антитело (Roche, № в каталоге 1093274); блокирующий буфер в PBS SuperBlock (Pierce, № в каталоге 37515); субстрат щелочной фосфатазы метилумбеллиферилфосфат (Molecular Probes, № в каталоге M-6491); образцы плазмы обезьян, содержащие ASM8; устройство для промывки планшетов MW96 (Beckman Coulter); 96-луночные планшеты из полипропилена большой емкости, Nunc); микропипетки из Eppendorf Research Brand; черные непрозрачные 96-луночные планшеты (Costar, № в каталоге 3915).
Синтез антисмысловых олигонуклеотидов и идентификация последовательностей
Олигонуклеотиды синтезировали с использованием устройства для сборки генов Gene Assembler-Plus™ (Pharmacia Biotech, Piscataway, NJ, USA), модифицировали фосфоротиоатом и очищали ВЭЖХ. TOP005 использовали в экспериментах, показанных на фигурах 5-7, которые осуществляли с использованием цГМФ-олигонуклеотидов. Антисмысловые последовательности и обозначения описаны в таблице 1.
Таблица 1

Клетки и культура клеток
Использовали следующие линии клеток: TF-1 (линия клеток эритролейкоза человека, ATCC № CRL-2003); EOL-1 (линия клеток острого миелоидного «эозинофильного» лейкоза человека; DSMZ № ACC386) и U937 (линия клеток гистиоцитарной лимфомы человека; ATCC № CRL-1593.2). EOL-land U937 культивировали в RPMI 1640 с 2 мМ L-глутамина; 1,5 г/л бикарбоната натрия; 4,5 г/л глюкозы; 10 мМ Hepes; 1 мМ пирувата натрия; 10% FBS, пенициллином 100 ед./мл, стрептомицином 100 мкг/мл. Такую же среду использовали для культуры TF-1, за исключением того, что добавляли rhGM-CSF в концентрации 2 нг/мл.
Культивирование и дифференцировка клона 15 клеток HL-60
Клон 15 HL-60 подвергали дифференцировке до эозинофилов, как описано в Tiffany et al., 1998, J. Immunol. 160:1385-1392. Коротко, линию клеток промиелоцитов HL-60 поддерживали в среде RPMI 1640 с L-глутамином с добавлением 10% инактивированной нагреванием FBS и 25 мМ N-[2-гидроксиэтил]пиперазин-N'-[2-гидроксипропансульфоновой кислоты] (Sigma Chemical Co., St. Louis, MO), pH 7,6, при 37°C и 5% CO2. Клетки индуцировали к дифференцировке до фенотипа, подобного фенотипу эозинофилов, обрабатывая их 0,5 мкМ масляной кислоты (Sigma Chemical Co., St. Louis, MO) в течение по меньшей мере 5 дней. Анализ FACS использовали для оценки наличия общей бета-цепи рецепторов IL-3/EL-5/GM-CSF после дифференцировки клеток.
Жизнеспособность клеток и обработка антисмысловыми олигонуклеотидами
Жизнеспособность клеток оценивали систематически, используя тест с красителем Alamar синим, следуя способу, предлагаемому производителем. Клетки EOL-1, TF-1, HL-60 или U937 собирали центрифугированием (5 минут, 1500 об/мин, при комнатной температуре), промывали 3×HBSS и ресуспендировали в концентрации 1×106 клеток/мл в среде RPMI без сыворотки. 1×106 клеток инкубировали в трех повторах в течение 5 минут с антисмысловым олигонуклеотидом в точно определенной концентрации (от 0 до 20 мкМ) в стерильной микропробирке. Затем каждую реакционную смесь переносили в 12-луночные планшеты и инкубировали при 37°C, 5% CO2 в течение 5 часов для количественного определения мРНК или в течение 12 часов для анализа белка. Добавляли RPMI/FBS 20% до конечной концентрации 10% FBS и клетки инкубировали при 37°C, 5% CO2 в течение ночи. Клетки собирали центрифугированием (5 минут, 1500 об/мин, при комнатной температуре) и промывали 1×HBSS. Включали контрольные эксперименты, которые состояли из обработок клеток в отсутствие антисмысловых олигонуклеотидов или в присутствии не совпадающих по принципу комплементарности олигонуклеотидов.
Очистка эозинофилов человека
Фракцию гранулоцитов получали центрифугированием цельной крови в градиентах Ficoll-Hypaque (1,077 г/мл при 350 г в течение 30 минут), получая лейкоцитарную пленку. Затем эозинофилы человека очищали посредством негативной селекции с применением покрытых анти-CD 16 иммуномагнитных микрошариков при 4°C, используя магнитную систему сортировки клеток Miltenyi Biotec (Auburn, CA). Чистота популяций эозинофилов, оцениваемая по окрашиванию красителем Гимза, обычно составляла 92%-100%.
Очистка PBMC человека и макак-крабоедов
Свежую кровь макак-крабоедов получали из ITR Laboratories Canada Inc. PBMC выделяли центрифугированием в градиенте плотности Ficoll-Hypaque крови с EDTA K3 от нормальных доноров. PBMC высевали в концентрации 2×106 клеток/мл/лунку в 12-луночные планшеты в среду для культивирования клеток RPMI 1640 с добавлением 10% инактивированной нагреванием FBS, пенициллина 100 ед./мл, стрептомицина 100 мкг/мл. Жизнеспособность клеток оценивали, используя краситель Alamar синий, и она обычно составляла 85%-95%.
Трансфекция PBMC и эозинофилов человека
PBMC человека собирали центрифугированием (5 минут, 1500 об/мин, при комнатной температуре), промывали 3×HBSS и ресуспендировали в концентрации 2×106 клеток/мл в среде RPMI с 5% сыворотки, содержащей 10 мкг/мл ФГА. 2×106 клеток инкубировали в трех повторах в течение 5 минут с антисмысловым олигонуклеотидом в точно определенной концентрации (от 0 до 20 мкМ) в стерильной микропробирке. Затем каждую реакционную смесь переносили в 12-луночные планшеты и инкубировали при 37°C, 5% CO2 в течение ночи для количественного определения мРНК или в течение 48 часов или меньше в указанных случаях для анализа белка. Клетки собирали центрифугированием (5 минут, 1500 об/мин, при комнатной температуре) и промывали 1×HBSS. Включали контрольные эксперименты, которые состояли из обработок клеток в отсутствие антисмысловых олигонуклеотидов или в присутствии не совпадающих по принципу комплементарности олигонуклеотидов.
Очищенные эозинофилы человека собирали центрифугированием (5 минут, 1500 об/мин, при комнатной температуре), промывали 3×HBSS и ресуспендировали в концентрации 2,5×106 клеток/мл в среде RPMI с 10% сыворотки, содержащей 2 нг/мл rhGM-CSF или rhIL-5, в течение ночи. Через сутки клетки дважды промывали HBSS и ресуспендировали в концентрации 2,5×106 клеток/мл в среде RPMI с 5% сыворотки и инкубировали в трех повторах в течение 5 минут с антисмысловым олигонуклеотидом в точно определенной концентрации (от 0 до 20 мкМ) в стерильной микропробирке. Затем каждую реакционную смесь переносили в 12-луночные планшеты и инкубировали при 37°C, 5% CO2 в течение ночи для количественного определения мРНК или в течение 48 часов или меньше в указанных случаях для анализа белка. Клетки собирали центрифугированием (5 минут, 1500 об/мин, при комнатной температуре) и промывали 1×HBSS. Включали контрольные эксперименты, которые состояли из обработок клеток в отсутствие антисмысловых олигонуклеотидов или в присутствии не совпадающих по принципу комплементарности олигонуклеотидов.
Трансфекция PBMC обезьян
PBMC макак-крабоедов собирали центрифугированием (5 минут, 1500 об/мин, при комнатной температуре), промывали 3×HBSS и ресуспендировали в концентрации 2×106 клеток/мл в среде RPMI с 5% сыворотки и 10 мкг/мл ФГА (или 10 нг/мл rhIL-2 в указанных случаях). 2×106 клеток инкубировали в трех повторах в течение 5 минут с антисмысловым олигонуклеотидом в точно определенной концентрации (от 0 до 20 мкМ) в стерильной микропробирке. Затем каждую реакционную смесь переносили в 12-луночные планшеты и инкубировали при 37°C, 5% CO2 в течение ночи для количественного определения мРНК или в течение 48 часов или меньше в указанных случаях для анализа белка. Клетки собирали центрифугированием (5 минут, 1500 об/мин, при комнатной температуре) и промывали 1×HBSS. Включали контрольные эксперименты, которые состояли из обработок клеток в отсутствие антисмысловых олигонуклеотидов или в присутствии не совпадающих по принципу комплементарности олигонуклеотидов.
Анализ проточной цитометрией
Клетки подсчитывали и ресуспендировали в концентрации 1×106 клеток в мл. Клетки центрифугировали при 400g в течение 3 мин при 20-25°C и супернатанты отбрасывали. Затем осадок клеток ресуспендировали в 50 мкл буфера для FACS (1×PBS, pH 7,2-7,4; 0,5% альбумин человека; 2,5% сыворотки человека) и инкубировали при 37°C в течение 30 мин. Не отбрасывая супернатант, первое антитело добавляли непосредственно в пробирку и перемешивали. Инкубировали при 4°C, защищая от света, в течение 1 час (антитело против CCR-3 человека использовали в соотношении 1 мкг на 0,5×106 клеток. Антитело против общей бета-цепи человека использовали в соотношении 2 мкг на 0,5×106 клеток). Промывали 2 мл буфера FACS, центрифугировали при 400g в течение 3 мин и супернатант отбрасывали. Для контролей изотипов осадок клеток ресуспендировали, используя 300 мкл FACSFix (1×PBS, pH 7,2-7,4; 4% параформальдегид), хранили при 4°C защищенным от света.
Для мечения CCR3 и общей бета-цепи ресуспендировали осадок клеток, используя 50 мкл буфера FACS, и добавляли второе антитело (антитело против IgG2a крысы Alexa Fluor 633 использовали в количестве 1 мкг на 0,5×106 клеток. Антитело против IgG2b мыши Alexa Fluor 488 использовали в количестве 2 мкг на 0,5×106 клеток). Инкубировали при 4°C, защищая от света, в течение 1 час. Промывали 2 мл буфера FACS, центрифугировали при 400g в течение 3 мин и супернатант отбрасывали. Меченые клетки фиксировали 300 мкл FACSFix, хранили при 4°C в защищенном от свете виде. Данные анализировали в цитометре FACS Calibur BD biosciences и обрабатывали, используя программу Cell Quest.
Анализ мобилизации кальция
Эозинофилы ресуспендировали в концентрации 1×107 клеток/мл в среде RPMI 1640, содержащей 10% FBS, и нагружали при инкубации с 5 М Fura-2 AM (Molecular Probes, Eugene, OR, USA) при комнатной температуре в течение 30 мин в темноте. Клетки (1×106 клеток/мл) три раза промывали и ресуспендировали в солевом буфере (138 мМ NaCl, 6 мМ KCl, 1 мМ CaCl2, 10 мМ Hepes, 5 мМ глюкоза и 1% БСА, pH 7,4). Затем каждые 2 мл суспензии клеток переносили в кварцевую кювету, которую помещали в люминесцентный спектрофотометр LS50B (Perkin-Elmer, Beaconsfield, UK). Мобилизацию Ca2+клетками измеряли, регистрируя отношение флюоресценции, излучаемой при 510 нм после последовательного возбуждения при 340 и 380 нм в ответ на хемокин.
Анализ хемотаксиса
Хемотаксис in vitro измеряли в 48-луночных камерах (NeuroProbe, Cabin John, MD), используя не содержащие поливинилпирролидона поликарбонатные мембраны с порами размером 5 мкм (Nucleopore-Neuroprobe). Контрольные или обработанные антисмысловым олигонуклеотидом эозинофилы суспендировали в концентрации 1×106 клеток/мл в среде RPMI 1640, содержащей 0,25% БСА. Верхние и нижние лунки содержали 50 мкл и 31 мкл суспензии клеток, соответственно, при этом последняя суспензия с добавлением оптимальной концентрации эотаксина (80 нг/мл). После 1-часовой инкубации при 37°C в 5% CO2 подсчитывали мигрировавшие клетки, присутствующие в нижней лунке. Спонтанную миграцию определяли в отсутствие эотаксина и учитывали в результатах.
Обработка антисмысловыми олигонуклеотидами и исследования токсичности у обезьян
Данный протокол согласован и оценен Комитетом по уходу за животными (ACC) ITR Laboratories Canada Inc. Уход за всеми животными осуществляли согласно принципам, описанным в современном «Руководстве по уходу и использованию экспериментальных животных», которое опубликовано Канадским советом по уходу за животными и в «Руководстве по уходу и использованию лабораторных животных», публикация NIH.
Токсичность ASM8, состоящего из смеси 1:1 двух олигонуклеотидов (TOP004 и TOP005), исследовали для характеристики токсикокинетического профиля составляющих его отдельных олигонуклеотидных компонентов при введении путем ингаляционного воздействия раз в сутки в течение 14 последовательных дней. Также оценивали обратимость любого эффекта ASM8 после 14-дневного периода восстановления. Также оценивали любое состояние системной гиперчувствительности после 14 дней воздействия путем ингаляции к ASM8 (выявляемое при интрадермальной инъекции (ID)).
Контрольный наполнитель представлял собой 0,9% раствор хлорида натрия для инъекции USP, и его использовали непосредственно после получения. Жидкие препараты тестируемого изделия (ASM8) для аэрозолизации готовили смешиванием TOP004 и TOP005 с 0,9% раствором хлорида натрия для инъекции, USP, получая смесь 1:1. Концентрацию целевой дозы в растворе рассчитывали на основе чистого олигонуклеотида. Поэтому при взвешивании и диспергировании компонентов тестируемого изделия применяли поправочный коэффициент для корректировки в отношении чистоты. Поправочные коэффициенты составляли 1,15 для TOP004 и 1,24 для TOP005. Перед началом 14-дневного периода воздействия количества каждого соответствующего олигонуклеотида, необходимые для каждого суточного воздействия, отвешивали, объединяли (в виде порошков) во флаконах для каждого дня воздействия и хранили замороженными при -80°C. В каждый день воздействия соответствующий флакон извлекали из морозильной камеры, где они хранились, содержимое растворяли в наполнителе - физиологическом растворе, фильтровали через стерильный фильтр 0,2 мкм и препарат использовали только для воздействия в этот день.
Количество животных в группе и обработки указаны в таблицах 2 и 3 ниже:
Диапазон возраста: молодые взрослые особи в 1 день обработки
Действующие концентрации ASM8 и уровни доз (4)
(1): На основании оцененной массы тела 2,5 кг.
(2): Животных с группе с использованием контрольного наполнителя подвергали воздействию аэрозоля, созданного из раствора наполнителя в концентрации в аэрозоле, которую считали эквивалентной по массе той концентрации, которую создавали для группы, обрабатываемой высокой дозой.
(3): Целевую дозу и концентрации в аэрозоле рассчитывали на основе абсолютной чистоты тестируемых изделий, которую получали, используя подходящие поправочные коэффициенты в отношении чистоты в процессе приготовления раствора такой дозы.
(4): Достигаемые уровни доз во время периода воздействия оценивали, используя следующую формулу: DL=EC×RMV×T/BW, где
DL=достигаемые уровни доз (мг/кг/сутки)
EC=реальная концентрация, доставляемая животным (мг/л воздуха)
RMV=минутный объем (мл/мин), оцениваемый согласно формуле Bide et al., 2000, J. App.Toxicol., 20, 273-290, а именно: RMV(L)=0,499×W(кг)0,809
T = время, продолжительность ежедневного воздействия (мин)
BW = средняя масса тела (кг) во время периода воздействия.
Считали, что полученное значение достигаемой дозы равно 100% осаждению в дыхательном пути. Прижизненные исследования, включая определение смертности, клинический исследования, определение массы тела, потребления пищи, электрокардиографию, офтальмоскопию, клиническую патологию, определения уровней в плазме, тестирование гиперчувствительности, осуществляли для всех животных.
После завершения периода обработки животных подвергали эвтаназии и проводили тестирование анатомической патологии, вскрытие, взвешивание органов, гистопатологию.
Полуколичественную ОТ-ПЦР использовали для измерения того, имело ли место какое-либо ингибирующее влияние ASM8 на экспрессию мРНК общей бета-цепи и CCR3 в образцах трахеи макак-крабоедов, обработанных высокой дозой, через 24 час после введения ASM8.
HL-ELISA для измерений олигонуклеотидов в плазме обезьян
Образцы крови обезьян (примерно 1 мл каждый) собирали от каждого животного в 1 и 14 день до введения дозы, через 0,5, 1, 3, 6 и 24 часов после введения дозы. Образцы крови центрифугировали при 4°C, чтобы получить плазму, и плазму разделяли и замораживали на сухом льду вплоть до анализа для определения концентраций TOP004 и TOP005 (и ближайших n-1 метаболитов) с использованием количественного анализа ELISA гибридизации/лигирования.
Раствор олигонуклеотида для стандартной кривой для образцов плазмы обезьян готовили серийными разведениями. Обычный рабочий диапазон стандартной кривой составлял от 125 нМ до 0,007629 нМ. Образцы плазмы соответствующим образом разбавляли для измерения в линейной части стандартной кривой, делая более одного разведения для точного измерения.
Каждый стандарт или образец плазмы делили на аликвоты (200 мкл) в 96-луночном планшете из полипропилена, в котором 200 мкл раствора соответствующего зонда матрицы добавляли к 200 мкл образца плазмы и инкубировали при 37°C в течение 60 минут. 150 мкл переносили в планшет, покрытый NeutrAvidin, в двух повторах и инкубировали при 37°C в течение 30 минут. Планшет 4 раза промывали буфером для промывки, используя устройство для промывки планшетов (200 мкл каждый). Добавляли 150 мкл раствора зонда лигирования с последующей инкубацией при комнатной температуре в течение 120 минут. После инкубации образец 2 раза промывали буфером для промывки, используя устройство для промывки планшетов (200 мкл каждый) с последующей промывкой 3 раза ddH2O, используя устройство для промывки (200 мкл). Добавляли 150 мкл анти-DIG-AP в разведении 1:2000 (в Super block, Peirce) с последующей инкубацией при комнатной температуре в течение 30 минут.Образец 4 раза промывали буфером для промывки, используя устройство для промывки (200 мкл). Затем добавляли 150 мкл 10 мкМ реагента MUP с последующей инкубацией при комнатной температуре в течение 60 минут. Регистрировали флуоресценцию при возбуждении при 355 ни/испускании 485 нм.
Растворы, используемые в HL-ELISA
Раствор зонда матрицы (0,05 мкМ зонда матрицы, 60 мМ Na2HPO4 pH 7,4, 0,9 М NaCl, 0,24% твин-20); 10×буфер для лигирования (0,8248 М трис-Cl pH 7,5, 0,0828 М MgCl2, 1,93% DTT); АТФ 100 мМ раствор (приготовленный в воде и доведенный до pH 7±0,5 с использованием NaOH); раствор зонда лигирования (0,067 мкМ олигонуклеотид в 1×буфере для лигирования, 0,025 единиц/мл ДНК-лигазы T4, 0,05 мМ АТФ); буфер для промывки (25 мМ трис-Cl pH 7,2, 0,15 М NaCl, 0,1% твин).
Гомогенизация трахей обезьян и экстракция РНК
Трахеи обезьян гомогенизировали, используя Politron PT 1200 (Brinkmann Instruments), и суммарную РНК экстрагировали, используя набор Qiagen RNAeasy MiniKit (Qiagen, Mississauga ON, Canada), с последующим расщеплением ДНКазой I. Суммарную РНК количественно оценивали, используя флуоресцентный анализ Ribogreen (Invitrogen Corporation, Burlington ON, Canada). кДНК получали из 1-2 мкг РНК, используя синтез первой нити кДНК с применением набора для обратной транскрипции SuperScript™ II RT (Invitrogen Corporation, Burlington ON, Canada).
Экстракция РНК, обратная транскрипция и полимеразная цепная реакция
РНК экстрагировали из осадков клеток согласно протоколу, прилагаемому к набору Qiagen RNAeasy miniKit, используя коллектор QiaVac 24 из Qiagen, и РНК обрабатывали ДНКазой-I согласно способам Fermentas. Количество РНК оценивали, используя реагент RiboGreen согласно протоколу производителя. Или же количество РНК оценивали с использованием спектрофотометра. Получение первой нити кДНК осуществляли с использованием системы для синтеза первой нити Superscript в наборе для ОТ-ПЦР из Invitrogen в общем объеме реакции 20 мкл. Коротко, 1-2,5 мкг РНК сначала денатурировали при 65°C в течение 5 минут с 0,5 мМ каждого из dNTP, 0,5 мкг олиго(dT)12-18 и охлаждали на льду в течение по меньшей мере 1 мин. Смесь инкубировали при 42°C в течение 2 минут и добавляли вторую предварительно приготовленную смесь, содержащую 1×буфер для синтеза первой нити, 10 мМ DTT, 40 единиц RNaseOUT и 40 единиц ОТ Superscript II. Реакционные смеси инкубировали при 42°C в течение 10 минут, при 50°C в течение 1 часа и инактивировали нагреванием при 70°C в течение 15 минут. ПЦР осуществляли с оптимизированным количеством кДНК (100-250 нг в случае CCR3 и 1-10 нг в случае G3PDH) в 1×буфере для ПЦР (10×: трис-HCl, KCl, (NH4)2SO4, 15 мМ MgCl2; pH 8,7) в общем объеме реакции 50 мкл, 0,2 мМ каждого dNTP, 8,5 пмоль каждого праймера для ПЦР и 2,5 единицы ДНК-полимеразы Taq. Смесь нагревали при 94°C в течение 5 минут с последующими 30-35 циклами, каждый из которых состоит из инкубации в течение 1 минуты при 94°C, в течение 45 секунд при 60°C и в течение 45 секунд при 72°C. Дополнительную элонгацию осуществляли при 72°C в течение 10 минут. Продукты ПЦР анализировали посредством электрофореза в 1,5% агарозном геле в присутствии бромида этидия. Количественный анализ продуктов ПЦР осуществляли с использованием компьютерной программы Total Lab (вычитание фона с использованием Rolling Ball; Ultra Lum Inc., модель UC4800). Использовали следующие праймеры для ПЦР: пара праймеров для CCR3 человека (R&D systems, № в каталоге RDP-209-025); пара праймеров для GAPDH человека (R&D systems, № в каталоге RDP-39-025) и праймеры, показанные в таблице 4. Систематически включали контрольные эксперименты, которые заключались в ПЦР на обратно не транскрибированной РНК.
Таблица 4

Химическое разрушение олигонуклеотида
Чтобы индуцировать разрушение TOP004 и TOP005 перед анализом (чтобы удостовериться в разделении продуктов разрушения и интактных молекул), осуществляли следующие обработки:
*Депуринизация: ASM8 ресуспендировали в 30% CH3COOH в конечной концентрации 0,5 мг/мл и инкубировали в течение 3, 4, или 6 часов при комнатной температуре. Реакцию останавливали добавлением 5 объемов воды и смесь помещали при -20°C перед лиофилизацией в Speed-Vac, чтобы удалить уксусную кислоту.
*Расщепление: депуринизированные олигонуклеотиды ресуспендировали в 0,2 М NaOH (0,5 мг/мл), инкубировали при 50°C в течение 1 часа и хранили при -20°C или анализировали ВЭЖХ.
ВЭЖХ-фракционирование TOP004 и TOP005
ASM8 взвешивали и растворяли в PBS в концентрации 0,5 мг/мл (0,25 мг/мл TOP004 и 0,25 мг/мл TOP005). Параметры градиента ВЭЖХ указаны ниже в таблице 5.
Параметры градиента ВЭЖХ
Разделение ВЭЖХ осуществляли с использованием бинарного насоса для ВЭЖХ Waters серии 1500, соединенного с двойным детектором поглощения Waters 2487 λ и оборудованного последовательно соединенным дегазатором, термостатом и ручным инжектором серии 1500, Reodyne 7725i. Смесь олигонуклеотидов фракционировали на анионообменной колонке Waters Protein Pak DEAE 5PW (0,5 см × 75 см), хранили при 60°C и регистрировали УФ-поглощение при 260 нм. Смесь олигонуклеотидов (объем = 25 мкл) наносили на колонку в воде (буфер A:вода (степень очистки MilliQ)) и проводили элюирование, постепенно увеличивая долю буфера B (1 M LiClO4, (профильтрованного через фильтр с диаметром пор 0,22 микрометра)), получая увеличение ионной силы жидкой фазы, которой элюировали олигонуклеотид из твердой фазы (колонки).
В условиях анализа 62,5 мкг либо TOP004, либо TOP005 давали измеряемое изменение >0,15 единиц поглощения (AU) при 260 нм.
Хранение олигонуклеотидов
Аликвоты ASM8 (0,5 мг/мл) в PBS инкубировали при -20°C, 4°C, 30°C и 40°C в течение 2 месяцев. На 4 и 8 неделе получали ВЭЖХ-профиль ASM8. Условием контроля считали ВЭЖХ-профиль ASM8 перед любым периодом хранения (т.е. в нулевой временной точке). Системой ВЭЖХ управляли с помощью компьютерной программы Breeze™ (V 3.30) Waters.
Кривые плавления олигонуклеотидов и сводные таблицы термодинамических данных
TOP004 и TOP005 смешивали в эквимолярных концентрациях в 1×PBS (а также в других буферных системах). Общая концентрация олигонуклеотидов была в диапазоне примерно от 1,2 до 8,7 мМ. Осуществляли стандартные способы УФ-термоденатурации, используя спектрофотометр Beckman DU640 с дополнительным устройством для анализа т.пл. Изменение поглощения регистрировали при 260 нм при изменении температуры на каждый градус от 10 до 90°C. Осуществляли подгонку кривых плавления, используя компьютерную программу MELTWIN™ 3.5, чтобы определить термодинамические параметры. Получали картины скрининга кривых плавления и сводные таблицы термодинамических данных.
Пример 1
Эффективность антисмысловых олигонуклеотидов, направленных к рецептору CCR3
Анализировали несколько антисмысловых олигонуклеотидов, направленных к рецептору хемокина CCR3, в отношении их способности ингибировать экспрессию мРНК рецептора и ингибировать функцию рецептора. Первичный скрининг антисмысловых олигонуклеотидов к CCR3 осуществляли в линиях клеток Eol-1 и U937. Указанные линии экспрессируют мРНК CCR3 в нормальных условиях культивирования клеток, описанных выше. В таблице 6 показаны антисмысловые олигонуклеотиды, направленные против рецептора хемокина CCR3 человека.
Обращаясь к таблице 6, видно, что антисмысловой олигонуклеотид 828, направленный против рецептора CCR3 (828: 5'-GTTACTACTTCCACCTGCCTG-3' SEQ ID NO. 1), эффективен в ингибировании экспрессии мРНК рецептора, как показано в таблице 6.
Олигонуклеотид 828 направлен против гена CCR3 и начинается через 48 оснований после конца экзона 1 и имеет длину 21 основание. Исследования с использованием BLAST осуществляли на 828 и других олигонуклеотидах, не направленных к рецептору CCR3, наиболее близкая гомология, как сообщается, составляет менее 72%. Такая гомология считается незначительной для достижения полной ассоциации двух комплементарных последовательностей. Специфичность 828 оценивали, используя не совпадающий по принципу комплементарности олигонуклеотид (SEQ ID NO. 32). Некомплементарный олигонуклеотид не оказывал влияния на мРНК CCR3 или на ген «домашнего хозяйства» G3PDH, используемый в качестве внутреннего контроля в таких экспериментах. Поэтому антисмысловой олигонуклеотид 828 является специфичным.

Пример 2
Эффективность двух DAP-замещенных олигонуклеотидов в мононуклеарных клетках периферической крови (PBMC) обезьян
Как обсуждается выше, антисмысловые олигонуклеотиды 107A и 828 модифицировали замещением аденозина DAP, чтобы получить антисмысловые олигонуклеотиды TOP004 и TOP005, соответственно. TOP004 (5'-GGGTCTGCXGCGGGXTGGT-3' (SEQ ID NO. 13), где X означает DAP-модификацию остатка аденозина), как в случае 107 A, является 19-мером, направленным к общей бета-цепи рецепторов IL-3, IL-5 и GM-CSF. TOP005 (5'-GTTXCTXCTTCCXCCTGCCTG-3' (SEQ ID NO. 14)), как и в случае 828, является 21-мером, направленным против рецептора хемокина CCR3.
Эффективности TOP004 и TOP005 тестировали отдельно и в комбинации. ASM8 представляет собой композицию, которая содержит частично оба олигонуклеотида TOP004 и TOP005. Исследования эффективности проводили на мононуклеарных клетках периферической крови (PBMC) обезьян, чтобы подтвердить возможность применения этого вида для исследования потенциальных токсических эффектов, возникающих в результате фармакологической активности ASM8.
Для того чтобы ASM8 был эффективным у макак-крабоедов, должна существовать достаточная гомология с их последовательностями-мишенями. Последовательность бета-цепи макак-крабоедов не доступна из опубликованных баз данных, и поэтому клонировали и секвенировали участок, охватывающий область последовательности TOP004. Однако активность TOP004 среди видов приматов может быть оценена непосредственно в соответствующей системе in vitro. В частности, препарат мононуклеарных клеток периферической крови (PBMC) является подходящей системой для тестирования функциональности TOP004, так как общая бета-цепь обнаружена на большей части субпопуляций мононуклеарных лейкоцитов (T- и B-клетки, моноциты и макрофаги).
Информация о последовательности рецептора CCR3 макак-крабоедов доступна только для кодирующей области; информация о последовательности области связывания TOP005 не доступна в опубликованных базах данных. Последовательность-мишень TOP005 начинается через 48 оснований после конца экзона 1 гена человека; указанный интрон охватывает более 20 тысяч пар оснований, что делает его клонирование и секвенирование очень громоздким. Сообщения в литературе показали, что некоторые участки интронных последовательностей консервативны у человека и обезьян (Rahman et al., 2004. Genomics., 8376-84). Эволюционные исследования также показывают, что участки гомологичных интронных последовательностей обнаруживаются в таксоне (человек, кит и тюлень), и что такие участки обнаруживаются более часто вблизи соединений интрон-экзон (Hare M.P. and Palumbi S.R., 2003 Mol. Biol. Evol. 20, 969-978.). Функциональность TOP005 у обезьян тестировали в препарате PBMC, в котором экспрессия рецептора CCR3 обнаружена на подгруппах T- и B-клеток.
Секвенирование общей бета-цепи макак-крабоедов
Анализ генов общей бета-цепи рецепторов IL3/IL-5/GM-CSF человека, шимпанзе, свиньи, мыши и крысы выявил высокую степень сходства последовательностей генов среди позвоночных. Конструировали последовательности праймеров для клонирующей ПЦР. Последовательности праймеров получали из высоко консервативных нуклеотидных последовательностей человека, шимпанзе, свиньи, мыши и крысы, окружающих область олигонуклеотида TOP004 гена общей бета-цепи. В таблице 2 показаны различные праймеры. Указанные праймеры использовали для амплификации специфичных продуктов кДНК PBMC макак-крабоедов. Получали несколько продуктов ПЦР, в зависимости от набора используемых праймеров. Раунд гнездовой ПЦР использовали для оценки специфичности полученных продуктов. Позитивные ампликоны клонировали и секвенировали.
На фигуре 1A показаны последовательности трех клонов (SEQ ID NO. 25, 26 и 27, соответственно), полученных в результате ПЦР-амплификации области TOP004 макак-крабоедов, выравниваемой с последовательностью человека (SEQ ID NO. 28) и соответствующей области в нуклеотидных последовательностях шимпанзе (SEQ ID NO. 33), свиньи (SEQ ID NO. 34), крысы (SEQ ID NO. 35) и мыши (SEQ ID NO. 36). Негомологичные нуклеотиды показаны маленькими буквами, тогда как консервативные области показаны заглавными буквами. Область TOP004 подчеркнута. Последовательность бета-цепи макак-крабоедов, комплементарная области TOP004, проявляла значительную гомологию (18 из 19 оснований идентичны) во всех трех секвенированных клонах. Различие обнаружено в положении 6 (начиная с 5'-конца TOP004), в котором обнаружены и «A» и «G» (при этом «A» является ожидаемым основанием). На фигуре 1B показано выравнивание последовательностей белков, рассчитанных на основе клонированных нуклеотидных последовательностей макак-крабоедов (SEQ ID NO. 37) и известных нуклеотидных последовательностей человека (SEQ ID NO. 38), шимпанзе (вариант AADA01213660) (SEQ ID NO. 39); свиньи (U94688.1) (SEQ ID NO. 40); мыши (NM_007780.1) (SEQ ID NO. 41) и крысы (NM_133555.1) (SEQ ID NO. 42). Различие в нуклеотидах, обнаруженное в положении 6 (начиная с 5'-конца 004, в котором обнаружены и «A» и «G»), соответствует второму основанию кодона глутамина (Q) или лизина (K) в общей бета-цепи доступных последовательностей белков в опубликованном банке данных. Данные, представленные на фигуре 1B, показывают, что высоко эволюционировавшие виды содержат остаток глутамина (стрелка у человека, шимпанзе, свиньи) в области, комплементарной TOP004. Глутамин кодируется 2 кодонами, CAA или CAG. У более низко организованных видов (мышь и крыса) глутамин заменен остатком лизина. Лизин кодируется 2 кодонами, AAA или AAG. В любом случае аденозин во втором положении является консервативным. Как таковой остаток аденозина вероятно является кандидатом для того, чтобы последовательность обезьян была функциональной, так как он имеется у других более высоко организованных позвоночных. Однако полиморфизм бета-цепи GM-CSF не может быть исключен. Freeburn et al. описывают несколько мутаций во внутрицитоплазматической области бета-цепи рецептора, которые могут быть ответственными за чувствительность к лейкозу (Freeburn et al., 1997, Exp.Hematol., 25: 306-311). Данные секвенирования, представленные на фигуре 1A, показывают, что остаток гуанозина может появляться в положении 6, начиная с 5'-конца подчеркнутой последовательности TOP004. В таком случае кодон будет представлять собой CGG, и последовательность белка будет содержать остаток аргинина (R) в данном положении. Наличие основания с основными свойствами (H, K или R) в указанном положении напоминает ситуацию у низших позвоночных и вероятно не относится к приматам.
Несмотря на указанное различие, очень высокая степень идентичности между последовательностями бета-цепи обезьяны и человека свидетельствует о функциональности TOP004 у макак-крабоедов.
Эффективность TOP004 и TOP005 в PBMC макак-крабоедов
TOP004 и TOP005 тестировали отдельно в PBMC макак-крабоедов в отношении их способности избирательно снижать экспрессию бета-цепи и CCR3, соответственно. Очищенные PBMC обезьян инкубировали с разными концентрациями TOP004 и TOP005.
На фигурах 2A и 2B результаты экспериментов, проведенных более чем на 10 образцах крови обезьян, представлены в виде гистограммы A и гистограммы B. Гистограммы показывают пониженную экспрессию мРНК бета-цепи и CCR3 в присутствии TOP004 (A) и TOP005 (B) в PBMC обезьян. Ингибирование было специфичным для TOP004 и TOP005 и не являлось следствием разрушения РНК или потери жизнеспособности клеток, о чем свидетельствует внутренний контроль (продукт длиной 451 п.о., соответствующий мРНК G3PDH, и тест жизнеспособности клеток). TOP004 специфично снижал экспрессию общей β-цепи в первичных PBMC обезьян, которую измеряли с помощью ОТ-ПЦР (фигура 2A). Максимальную эффективность получали в случае концентраций от 10 до 15 мкМ, при этом главным образом наблюдали >50% ингибирование. Ингибирование бета-цепи обезьян олигонуклеотидом TOP004 подтверждалось данными секвенирования (фигура 1A), которые показали очень высокую степень идентичности между последовательностями мРНК бета-цепи человека и обезьян. Подобным образом трансфекция TOP005 в PBMC обезьян снижала экспрессию мРНК CCR3, которую измеряли с помощью ОТ-ПЦР (фигура 2B). Максимальное ингибирование экспрессии мРНК CCR3 олигонуклеотидом TOP005 получали при более низких концентрациях антисмыслового олигонуклеотида (от 0,05 до 2,5 мкМ), чем в случае бета-цепи (от 10 до 15 мкМ).
Ингибирование экспрессии мРНК, измеряемое с помощью ОТ-ПЦР, также подтверждалось на уровне белка при использовании проточной цитометрии (FACS). PBMC обезьян инкубировали в течение 36 часов в среде роста в присутствии различных концентраций либо TOP004, либо TOP005. Количественную оценку проточной цитометрией осуществляли, как описано выше. На фигурах 3A и 3B гистограммы показывают экспрессию на клеточной поверхности бета-цепи и CCR3 в присутствии TOP004 и TOP005, соответственно, в PBMC макак-крабоедов. Графики показывают, что после обработки TOP004 или TOP005 наблюдали, что снижение процента клеток, экспрессирующих бета-цепь и CCR3, более чем на 30% достигается при концентрации 7,5 мкМ и 0,5 мкМ, соответственно.
TOP004 и TOP005 также тестировали в комбинации в соотношении 1:1 (ASM8) в PBMC макак-крабоедов в отношении их способности избирательно снижать экспрессию бета-цепи и CCR3, соответственно. PBMC обезьян инкубировали в течение ночи в присутствии различных концентраций ASM8 перед экспрессией бета-цепи и CCR3, которую оценивали с использованием ОТ-ПЦР. На фигурах 4A и 4B гистограммы, представляющие экспрессию мРНК бета-цепи и CCR3 в присутствии ASM8 в PBMC макак-крабоедов, показаны для более чем пяти (5) образцов крови, полученных от обезьян. Значимое ингибирование бета-цепи наблюдали при концентрациях ASM8 в диапазоне от 2,5 до 5 мкМ (фигура 4A), который ниже, чем диапазон оптимальных концентраций, дающих максимальное ингибирование при воздействии TOP004 отдельно (от 10 до 15 мкМ (фигура 2A)). Полученные результаты показывают, что комбинация антисмысловых олигонуклеотидов TOP004 и TOP005 обеспечивает повышенную эффективность и синергизм в ASM8 при блокировании экспрессии бета-цепи, по сравнению с TOP004 отдельно. Подобным образом трансфекция ASM8 в PBMC обезьян снижала экспрессию мРНК CCR3, которую измеряли, используя ОТ-ПЦР (фигура 4B). Максимальное ингибирование экспрессии мРНК CCR3 при действии TOP005 получали при более низкой концентрации антисмыслового олигонуклеотида (от 0,05 до 5 мкМ), чем в случае бета-цепи (от 2,5 до 5 мкМ). Влияние ASM8 на ингибирование CCR3 не было явно зависимым от концентрации, полученный результат может отражать тот факт, что максимальное ингибирование (плато) достигается при более низкой концентрации в случае CCR3, чем в случае бета-цепи.
В заключение, секвенирование общей бета-цепи макак-крабоедов показало очень высокую степень идентичности (по меньшей мере 18 из 19 оснований, которые охватывают последовательность TOP004). Предположили, что такая высокая степень гомологии с геном человека обеспечит возможность достаточной гибридизации TOP004 с мРНК бета-цепи обезьян, чтобы индуцировать антисмысловую активность и, тем самым, снизить ее экспрессию.
TOP004 трансфицировали в очищенные PBMC макак-крабоедов, чтобы оценить его способность обеспечивать понижающую регуляцию экспрессии β-цепи обезьян. TOP004 эффективно снижал экспрессию мРНК β-цепи, измеряемую с использованием ОТ-ПЦР, на 30-70%.
Подобным образом, TOP005 трансфицировали в PBMC макак-крабоедов и определяли уровень экспрессии CCR3 полуколичественной ОТ-ПЦР. Результаты показывают, что TOP005 осуществляет понижающую регуляцию экспрессии CCR3 макак-крабоедов в диапазоне от 30% до 85%.
Таким же образом трансфекция либо TOP004, либо TOP005 в PMBC обезьян индуцировала специфичное снижение при 0,5 мкМ (>30%) количества клеток, позитивных в отношении бета-цепи или CCR3, которое измеряли проточной цитометрией.
ASM8 также трансфицировали в очищенные PBMC обезьян, чтобы оценить эффективность комбинированной обработки (TOP004 и TOP005) в понижающей регуляции экспрессии мРНК бета-цепи и CCR3 обезьян. В указанных условиях ASM8 значительно снижал экспрессию бета-цепи и CCR3, измеряемую в ОТ-ПЦР, при такой низкой концентрации ASM8, как от 0,1 до 0,5 мкМ. Это также свидетельствует о том, что макаки-крабоеды являются подходящим видом для исследования возможных токсических эффектов вследствие фармакологической активности ASM8.
Пример 3
Влияние антисмысловых олигонуклеотидов, направленных против CCR3, в клетках и линиях клеток человека
Проводили дополнительные эксперименты, чтобы оценить способность A86 и TOP005 ингибировать экспрессию мРНК CCR3 в дифференцированных подобных эозинофилам клетках HL-60 (Lee Tiffany et al, J. Immunol. 1998, 160: 1385-92), клетках U937 и Eol-1, а также в мононуклеарных клетках периферической крови (PBMC). Также исследовали способность A86 и TOP005 ингибировать миграцию эозинофильных клеток и мобилизацию кальция в клетках HL-60 и в очищенных эозинофилах периферической крови человека. A86 является антисмысловым олигонуклеотидом (5' CTG GGC CAT CAG TGC TCT G 3' (SEQ ID NO. 29), который соответствует последовательности 87-105 нуклеотидов кодирующей области (экзон 7) CCR3. Как обсуждалось ранее, TOP005 представляет собой 828, но с замещением трех аденозинов 2,6-диаминопурином (5' GTT XCT XCT TCC XCC TGC CTG 3' (SEQ ID NO. 14)). Последовательность, комплементарная 828, начинается через 48 оснований после конца экзона 1 и имеет длину 21 основание. В качестве контроля для A86 использовали комплементарный смысловой олигонуклеотид (5' CAG AGC ACT GAT GGC CCA G 3' (SEQ ID NO. 30)) и не совпадающий по принципу комплементарности олигонуклеотид (5' CGT GGC ACT CAG TGT CCT G 3' (SEQ ID NO. 31)). В качестве контроля для 828/TOP005 использовали не совпадающий по принципу комплементарности олигонуклеотид (5' CCT TTG ACC TGC CAA TGC TCT 3' (SEQ ID NO. 32)).
Влияние антисмысловых олигонуклеотидов A86 на экспрессию мРНК CCR3 в эозинофилах, полученных из клона 15 клеток HL-60
Известно, что вариант клона 15 клеток HL-60 можно индуцировать обработкой масляной кислотой в дифференцировке в клетки, имеющие множество свойств эозинофилов периферической крови (Lee Tiffany et al., J. Immunol. 1998, 160: 1385-92). Используя такой же протокол дифференцировки, авторы подтвердили экспрессию мРНК CCR3 в дифференцированных клетках HL-60. Затем осуществляли ОТ-ПЦР, чтобы исследовать способности синтетических олигонуклеотидов модулировать экспрессию мРНК, кодирующей рецептор CCR3 в клетках HL-60, дифференцированных в эозинофилы. После обработки клеток 10 мкМ A86 оценивали мРНК CCR3 посредством полуколичественной ПЦР, используя в качестве внутреннего контроля G3PDH. Суммарную РНК выделяли из свежесобранных клеток, как описано выше. На фигуре 5 показано влияние антисмысловых олигонуклеотидов против CCR3 на экспрессию мРНК CCR3 в дифференцированных клетках HL60. В отличие от смысловых олигонуклеотидов и не совпадающих по принципу комплементарности олигонуклеотидов антисмысловые олигонуклеотиды заметно ингибируют экспрессию мРНК CCR3. Экспрессия мРНК CCR3 в клетках, обработанных смысловыми олигонуклеотидами и несовпадающими по принципу комплементарности олигонуклеотидами, значимо не отличалась от экспрессии в необработанных клетках. Кроме того, все олигонуклеотиды в используемой концентрации не влияли на экспрессию мРНК G3PDH. Поэтому антисмысловой олигонуклеотид A86, используемый в данном эксперименте, способен специфично ингибировать экспрессию мРНК CCR3.
Влияние A86 на экспрессию белка CCR3 на клеточной поверхности
Далее исследовали, может ли снижение мРНК CCR3 отражать снижение плотности белка CCR3 на клеточной поверхности. В этой связи осуществляли проточно-цитометрический анализ, чтобы оценить экспрессию рецептора CCR3 на полученных из HL-60 эозинофилах после обработки олигонуклеотидами. После обработки масляной кислотой процент полученных из HL-60 эозинофилов, экспрессирующих рецептор CCR3, составлял 40%. При обработке смысловым и не совпадающим по принципу комплементарности олигонуклеотидами (10 мкМ) процент позитивных клеток немного и не значимо снижался; процент позитивных клеток составлял 35% и 38%, соответственно. Однако плотность рецептора CCR3 на клетках, обработанных A86, снижался значительно (26% позитивных клеток по сравнению с 40% в необработанных клетках). A86 в концентрации 10 мкМ способен снижать экспрессию CCR3 на клеточной поверхности на 65%. Влияние A86 было более значительным при более высоких концентрациях. В частности, использовали 20 и 30 мкМ A86, и результаты показывают, что экспрессия CCR3 на клеточной поверхности снижалась на 75% и 85%, соответственно. A86, антисмысловой олигонуклеотид к CCR3, способен ингибировать экспрессию CCR3 на клеточной поверхности зависимым от дозы образом.
Влияние A86 на индуцированную эотаксином мобилизацию кальция в клетках HL-60
Быстрый временный поток кальция обычно наблюдают в том случае, когда лейкоциты стимулируют хемокинами, к которым они экспрессируют специфичный рецептор. За такой мобилизацией кальция можно следить в режиме реального времени, используя нагруженные Fura-2AM клетки, и мобилизация является подходящей мерой активации рецептора. Хемокин эотаксин является специфичным лигандом рецептора CCR3 и индуцирует быстрый приток кальция и хемотаксис лейкоцитов при связывании с рецептором. На фигуре 6 показано влияние A86 на активацию CCR3. Мобилизация кальция в ответ на эотаксин снижалась в клетках, обработанных A86, по сравнению с контрольным и смысловым олигонуклеотидами. Клетки обрабатывали олигонуклеотидом A86 в концентрации 10 мкМ. Клетки, обработанные смысловым олигонуклеотидом, способны отвечать на эотаксин, как и необработанные клетки. В указанных условиях эотаксин индуцирует увеличение внутриклеточной концентрации Ca++. Однако в клетках, обработанных A86, эотаксин намного меньше индуцировал мобилизацию Ca++. Результаты, представленные в данном описании, показывают, что A86 был эффективным, создавая препятствие для активации рецептора CCR3 в линии клеток HL-60.
Обработка очищенных эозинофилов человека антисмысловыми олигонуклеотидами ингибирует их ответ на эотаксин
На фигуре 7 показано влияние антисмысловых олигонуклеотидов на хемотаксический ответ очищенных эозинофилов человека на эотаксин. Очищенные эозинофилы человека инкубировали в течение ночи с антисмысловыми олигонуклеотидами (квадраты) или смысловыми олигонуклеотидами (кружки) в концентрации 10 мкМ в RPMI 1640 с добавлением 5% FCS и IL-5 (1,5 нг/мл). Контрольные клетки (треугольники) инкубировали в таких же условиях без олигодезоксинуклеотидов (ODNS). Данные получены в одном эксперименте, типичном для трех экспериментов, и представлены в виде среднего количества мигрировавших клеток±стандартное отклонение из определений мигрировавших клеток в 5 полях зрениях при большом увеличении в трех повторах. Миграцию эозинофилов ингибировали антисмысловыми олигонуклеотидами против CCR3 и такое ингибирование было более значительным при увеличении концентрации эотаксина. При концентрации эотаксина 80 нг/мл миграция эозинофилов снижалась на 55,6%.
На фигуре 8 показана мобилизация кальция в эозинофилах, обработанных антисмысловыми олигонуклеотидами. Когда эозинофильные клетки обрабатывали A86 (10 мкМ), мобилизация Ca++, индуцированная эотаксином, также была ингибирована по сравнению с контрольным и смысловым олигонуклеотидами.
Результаты, представленные в данном описании, показывают, что A86 является эффективным средством, препятствующим хемотаксису эозинофилов на эотаксин посредством понижающей регуляции рецептора CCR3.
Эффективность TOP005
Сходные эксперименты осуществляли, используя TOP005. TOP005 был выбран на основании эффективности 828, результатов BLAST-оценки последовательности, показывающей, что 828 не имел гомологии с известными генами, отсутствия гибридизации 828 с TOP004 (эксперименты, осуществляемые с использованием компьютерной программы для ДНК) и его длины (позволяющей отличать его от TOP004 при смешивании вместе и разделении анионообменной ВЭЖХ).
На фигуре 9 показано влияние TOP005 на экспрессию CCR3 на клеточной поверхности. Эффективность TOP005 оценивали на клетках Eol-1 и U937. Экспрессию CCR3 оценивали проточной цитометрией через 36 часов после обработки TOP005. Результаты представлены в виде процента экспрессии по сравнению с контролями в клетках Eol-1 и U937. Гистограммы на фигуре 9 показывают, что TOP005 ингибировал экспрессию белка CCR3 на поверхности клеток U937 и Eol-1.
На фигурах 10A и 10B показано влияние TOP005 на экспрессию мРНК CCR3 в мононуклеарных клетках периферической крови человека (PBMC). PBMC человека либо выделяли свежими, либо культивировали с интерлейкином-2 человека (10 нг/мл в течение 24 часов). Затем их экспонировали с TOP005 и культивировали в течение 18 часов. На фигуре 10A сверху представлены гели, показывающие экспрессию G3PDH и CCR3. Отношение экспрессии мРНК CCR3 к G3PDH, нормализованное по отношению к контролям, представлено снизу. На фигуре 10B гистограмма показывает, что TOP005 является эффективным в снижении экспрессии мРНК CCR3 в PBMC в таких низких дозах, как 1 мкМ.
Следовательно, антисмысловые олигонуклеотиды A86 и TOP005 могут ингибировать экспрессию мРНК CCR3 в клетках Eol-1 (линия эозинофильных клеток человека), клетках HL-60, дифференцированных в эозинофилы, и клетках U937. Ингибирование CCR3 указанными олигонуклеотидами также снижало мобилизацию кальция как в дифференцированных клетках HL-60, так и в эозинофилах человека, а также снижало хемотаксис эозинофилов на эотаксин. Ни соответствующие смысловые олигонуклеотиды, ни некомплементарные олигонуклеотиды не влияли на ответ на эотаксин.
Пример 4
Эффективность TOP004 в снижении экспрессии общей субъединицы β-цепи рецепторов IL-3, IL-5 и GM-CSF и связанных клеточных ответов в линиях клеток человека
Следующие эксперименты осуществляли для того, чтобы тестировать влияние 107A и TOP004 на экспрессию общей субъединицы бета-цепи рецепторов IL-3, IL-5 и GM-CSF.
Модулирование экспрессии мРНК бета-цепи в клетках TF-1 и U937
На фигурах 11A, 11B и 11C показано модулирование экспрессии мРНК бета-цепи в клетках TF-1. Клетки TF-1 обрабатывали антисмысловым олигонуклеотидом 107A в течение 12 часов. Как показано на фигуре 11A, осуществляли ОТ-ПЦР, чтобы выявить экспрессию мРНК бета-цепи и мРНК G3PDH в клетках TF-1. Клетки обрабатывали следующим образом: дорожка 1, необработанный контроль; дорожка 2, смысловой олигонуклеотид (10 мкМ); дорожка 3, 107A (10 мкМ); дорожка 4, некомплементарный олигонуклеотид (10 мкМ). Полуколичественную ОТ-ПЦР в ненасыщающих условиях использовали для оценки экспрессии мРНК бета-цепи и G3PDH (используемой в качестве контроля). Обработка 107A (10 мкМ) почти полностью ингибировала экспрессию бета-цепи в обработанных клетках TF-1 (фигура 11) и U937 (данные не показаны). Ингибирование было специфичным для 107A и не являлось следствием разрушения РНК или потери жизнеспособности клеток, о чем свидетельствует внутренний контроль (продукт длиной 450 п.о., соответствующий мРНК G3PDH) (фигура 11A). В отличие от этого экспрессия мРНК бета-цепи не ингибировалась в необработанных контрольных клетках или в клетках, обработанных смысловым или некомплементарным олигонуклеотидом (фигура 11A). Таким образом, активность 107A было специфичной и эффективной в отношении ингибирования экспрессии мРНК бета-цепи.
На фигурах 11B и 11C показано влияние обработки смысловым олигонуклеотидом и 107A на экспрессию бета-цепи на клеточной поверхности клеток TF-1, которую определяли анализом FACS. На фигуре 11B показаны необработанные контроли (PC) по сравнению с клетками, обработанными смысловым олигонуклеотидом (S-ODN), и NC на фигуре означает негативный контроль. На фигуре 11C показана экспрессия на клеточной поверхности клеток, обработанных 107A (в различных концентрациях 5, 10 и 20 мкМ) в течение 36 часов. Способность антисмысловых олигонуклеотидов ингибировать экспрессию клеточного белка бета-цепи приводила к соответствующей более низкой плотности субъединицы бета-цепи на поверхности клеток, обработанных 107A. Использовали моноклональное антитело (мАт) против белка общей бета-цепи рецепторов GM-CSF/IL-3/IL-5 наряду с FACS-анализом, чтобы измерить экспрессию на клеточной поверхности белка бета-цепи на клетках TF-1. Уровень экспрессии бета-цепи в необработанных клетках TF-1 был очень высоким, и на него не влияла обработка смысловым олигонуклеотидом. Однако возрастающие концентрации 107A (5, 10 и 20 мкМ) значительно снижали уровень экспрессии бета-цепи зависимым от дозы образом (процент клеток, которые были тестированы как позитивные в FACS-анализе, снижался с 69,9% до 27,8%).
На фигуре 11D показано ингибирование экспрессии общей бета-цепи в клетках U937 после обработки TOP004. Клетки U937 инкубировали в присутствии возрастающих концентраций TOP004 (0,01, 0,1, 1 и 10 мкМ) в течение 12 часов в бессывороточной среде перед ОТ-ПЦР и за 48 часов до FACS-анализа. Процент ингибирования мРНК или белка общей бета-цепи определяли, сравнивая полученные значения со значениями для необработанных клеток. Эксперимент осуществляли в трех повторах, и данные представляют собой средние (станд. ошибка). Результаты, представленные на фигуре 11D, показывают, что антисмысловой олигонуклеотид TOP004, который представляет собой последовательность DAP-содержащих остатков, гомологичных антисмысловому олигонуклеотиду 107A, является эффективным в ингибировании общей бета-цепи на уровне мРНК и белка. Кроме того, обнаружено, что небольшие количества TOP004 (например, 1 мкМ) достаточны для «нокаута» мРНК бета-цепи, а также соответствующего белка. Таким образом, полученные данные подтверждают эффективность DAP-химии и ее применение в фармакологической композиции, которая описана выше.
Жизнеспособность клеток и функциональные исследования
На фигуре 12 показана пролиферация клеток TF-1, обработанных антисмысловым олигонуклеотидом 107A, в присутствии GM-CSF, IL-3 или IL-5. Клетки инкубировали с 107A (10 мкМ) в течение 5 часов в бессывороточной среде, содержащей 1 нг/мл GM-CSF или 3 нг/мл IL-3 или 3 нг/мл IL-5. Инкубацию заканчивали через 2 суток и пролиферацию клеток измеряли в анализе с использованием красителя alamar синего (n=3). Результаты выражены в виде среднего поглощения (570-595)±стандартное отклонение.
Для пролиферации клеток TF-1 требуются цитокины GM-CSF, IL-3 или IL-5, и в биологический ответ на указанные цитокины вовлечен путь передачи сигнала посредством бета-цепи. Предположили, что ингибирование экспрессии белка бета-цепи на клеточной поверхности ингибирует пролиферацию клеток TF-1 даже в присутствии таких цитокинов. 107A (10 мкМ) вызывал ингибирование роста клеток TF-1 в присутствии IL-3, IL-5 или GM-CSF. Полученные результаты свидетельствуют, что ингибирование экспрессии белка бета-цепи на клеточной поверхности посредством 107A эффективно ингибировало биологические ответы клеток на все три цитокина.
Эозинофилы экспрессируют рецепторы GM-CSF, IL-3 и IL-5 и играют ключевую роль в воспалении и аллергии. Эозинофилы требуют GM-CSF, IL-3 и особенно IL-5 для своей дифференцировки, активации и жизнеспособности (Oddera et al., 1998, Lung, 176: 237-247; Ohnishi et al., 1993, J. Allergy Clin. Immunol., 92: 607-615). Исследовали способность антисмыслового олигонуклеотида, мишенью которого является мРНК бета-цепи, ингибировать жизнеспособность эозинофилов в ответ на IL-5. На фигурах 13A и 13B показано модулирование жизнеспособности эозинофилов посредством 107A.
Как показано на фигуре 13A, очищенные эозинофилы человека инкубировали с 107A в указанных концентрациях (10, 15 и 20 мкМ) в среде RPMI с добавлением 5% FBS и 1,5 нг/мл IL-5 в течение ночи. Жизнеспособность эозинофилов оценивали, используя анализ исключения на основе окрашивания красителем трипановым синим. Результаты представляют собой средние результаты из трех экспериментов. Обработка 107A в указанных концентрациях значимо снижала жизнеспособность эозинофилов зависимым от дозы образом до 35%±12% (10 мкМ), 43%±2% (15 мкМ) и 54%±7% (20 мкМ) от контрольных уровней (p<0,01). На жизнеспособность эозинофилов существенно не влияла обработка смысловым олигонуклеотидом, а также контролем в концентрации 20 мкМ. Таким образом, 107A, нацеленный на бета-цепь, ингибировал жизнеспособность эозинофилов даже в присутствии культуральной среды, содержащей специфичный цитокин IL-5.
Как показано на фигуре 13B, очищенные эозинофилы человека инкубировали в течение 48 часов в RPMI с добавлением 5% FBS и 2 нг/мл IL-5 в присутствии или отсутствие 107A (15 мкМ). Жизнеспособность эозинофилов оценивали в анализе проточной цитометрией с использованием протокола на основе аннексин-V-ФИТЦ и йодида пропидия, как описано в разделе «Материалы и способы». В том случае, когда эозинофилы обрабатывали 107A, их жизнеспособность снижалась на 64%, 41% вследствие апоптоза. Напротив, в необработанных клетках и клетках, обработанных смысловым олигонуклеотидом, процент мертвых клеток был более низким.
Таким образом, антисмысловой олигонуклеотид 107A специфично ингибирует экспрессию общей бета-цепи в клетках TF-1 и первичных эозинофилах на уровне мРНК и белка, измеряемую в ОТ-ПЦР и FACS. Максимальную эффективность, получаемую на тестированной клеточной системе в используемых экспериментальных условиях, наблюдали при концентрации 20 мкМ. В присутствии 107A пролиферация клеток TF-1 была снижена, независимо от использования IL-3, IL-5 или GM-CSF в качестве трофического фактора. Полученный результат показывает специфичность и эффективность антисмыслового олигонуклеотида 107A в отношении бета-цепи.
Жизнеспособность эозинофилов была ингибирована 107A в присутствии IL-5, и вероятно следствием такого ингибирующего эффекта является апоптоз. Эозинофилы играют ключевую роль в аллергическом воспалении и требуют GM-CSF, IL-3 и IL-5 для своей дифференцировки, активации и жизнеспособности (Adachi et al., 1995, Am. J. Respir. Grit. Care Med. 151:618-623 и Oddera et al., 1998, Lung. 176:237-247). Считается, что в случае астмы накопление и жизнеспособность эозинофилов являются важными факторами, которые вносят вклад в воспаление и повреждение эпителиальной ткани, так как они высвобождают токсичные продукты, включая эозинофильный катионный белок (Walsh et al., 1997, Clin. Exp.Allergy 27: 482-487).
Пример 5
Анализ мРНК генов-мишеней ASM8 в образцах трахеи
Следующие эксперименты проводили для того, чтобы проанализировать образцы трахеи макак-крабоедов в отношении уровней мРНК генов-мишеней, против которых направлен ASM8 (субъединица бета-цепи и CCR3). На 15 день (первый день после последней дозы) образцы трахеи собирали сразу после умерщвления всех животных, включенных в основную фазу испытаний в группах 1 (контроль) и 4 (группа, обработанная высокой дозой; уровень целевой дозы 2,5 мг/кг/сутки), и быстро замораживали в жидком азоте. Замороженные образцы трахеи анализировали в отношении уровней мРНК в ОТ-ПЦР.
Уровни экспрессии генов-мишеней (βC-субъединица и CCR3) определяли в образцах трахеи обезьян, используя подтвержденный, полуколичественный способ ОТ-ПЦР. Специфичные в отношении βC-субъединицы и CCR3 ПЦР-амплификации осуществляли на экстрактах трахеи контрольных и обработанных высокой дозой ASM8 животных (таблица 7).
Результаты анализа образцов в ОТ-ПЦР
Хотя глицеральдегид-3-фосфатдегидрогеназу (G3PDH) обычно используют в качестве внутреннего контроля в анализах реакций ОТ-ПЦР, наблюдали умеренный клеточный инфильтрат легких и трахеи (который обычно наблюдают в случае других антисмысловых олигонуклеотидов в местах отложения). Таким образом, так как клеточный инфильтрат вносил вклад в измеряемые уровни βC-субъединицы и CCR3 (т.е. иммунные клетки экспрессируют βC-субъединицу и CCR3), то считали, что G3PDH не является наиболее подходящим геном для применения в качестве внутреннего контроля в данном случае. Вместо этого экспрессию генов-мишеней нормализовали к уровням мРНК воспалительных цитокинов; т.е. IL-4 и TNF-α. Результаты показывают, что даже примерно через 24 часа после введения ASM8 относительная экспрессия мРНК βC-субъединицы и CCR3 по отношению к мРНК IL-4 снижалась на 29% и 24%, соответственно, и экспрессия относительно TNF-α снижалась на 30% и 24%, соответственно, у обработанных ASM8 животных (таблица 8).
Таблица ингибирования
Таким образом, обработка ASM8 значимо ингибирует экспрессию мРНК βc-субъединицы и CCR3 по сравнению с воспалительными цитокинами IL-4 и TNF-альфа, несмотря на сложность ткани трахеи обезьян и период времени 24 часа, который проходит между введением дозы ASM8 и получением образцов ткани.
Пример 6
Стабильность ASM8 при хранении
Тестирование стабильности проводили для оценки целостности составляющих олигонуклеотидных компонентов ASM8 (TOP004 и TOP005) при разных температурах хранения. Такая информация имеет важное значение для определения оптимальных условий хранения, повторных испытаний и сроков хранения для ASM8.
Капиллярный гель-электрофорез (CGE) и высоко эффективную (высокого давления) жидкостную хроматографию (ВЭЖХ) широко использовали для химического анализа антисмысловых олигонуклеотидов. Так как ASM8 состоит из двух олигонуклеотидов, то система тестирования должна обеспечивать адекватное разделение двух отдельных антисмысловых молекул. Таким образом, будет описан: 1) способ, основанный на анионообменной хроматографии для разделения компонентов ASM8 (TOP004 и TOP005) и продуктов их разрушения, и 2) влияние температуры хранения на стабильность компонентов, составляющих ASM8 (фигуры 14-16).
ASM8 взвешивали и растворяли в PBS в концентрации 0,5 мг/мл (0,25 мг/мл TOP004 и 0,25 мг/мл TOP005). [Коэффициент чистоты для TOP004 составлял 1,15 (т.е. 1,15 г порошка содержит 1 г активной молекулы); коэффициент чистоты для TOP005 составлял 1,24 (т.е. 1,24 г порошка содержит 1 г активной молекулы)].
Чтобы индуцировать разрушение TOP004 и TOP005 перед анализом (чтобы гарантировать возможность разделения продуктов разрушения и интактных молекул), осуществляли следующие обработки:
- Депуринизация: ASM8 ресуспендировали в 30% CH3COOH в конечной концентрации 0,5 мг/мл и инкубировали в течение 3, 4 или 6 часов при комнатной температуре. Реакцию останавливали добавлением 5 объемов воды и смесь помещали при -20°C с последующей лиофилизацией в Speed-Vac, чтобы удалить уксусную кислоту.
- Расщепление: депуринизированные олигонуклеотиды ресуспендировали в 0,2 М NaOH (0,5 мг/мл), инкубировали при 50°C в течение 1 часа и хранили при -20°C или анализировали посредством ВЭЖХ.
Аликвоты ASM8 (0,5 мг/мл) в PBS инкубировали при -20°C, 4°C, 30°C и 40°C в течение 2 месяцев. На 4 и 8 неделе получали ВЭЖХ-профиль ASM8. В качестве контрольного условия определяли ВЭЖХ-профиль ASM8 перед любым периодом хранения (т.е. нулевое время). Системой ВЭЖХ управляли с помощью компьютерной программы Breeze (V 3.30) Waters (фигуры 17A1, 17A2, 17B1 и 17B2).
Разделение ВЭЖХ осуществляли с использованием бинарного насоса для ВЭЖХ Waters серии 1500, соединенного с двойным детектором поглощения Waters 2487 λ и оборудованного последовательно соединенным дегазатором, термостатом и ручным инжектором серии 1500, Reodyne 7725i.
Смесь олигонуклеотидов фракционировали на анионообменной колонке Waters Protein Pak DEAE 5PW (0,5 см × 75 см), хранили при 60°C и регистрировали УФ-поглощение при 260 нм. Смесь олигонуклеотидов (объем = 25 мкл) наносили на колонку в воде (буфер A) и проводили элюирование, постепенно увеличивая долю буфера B (1 M LiClO4), получая увеличение ионной силы жидкой фазы (таблица 9), которой элюировали олигонуклеотид из твердой фазы (колонки).
В условиях анализа 62,5 мкг либо TOP004, либо TOP005 давали измеряемое изменение >0,15 единиц поглощения (AU) при 260 нм.
Градиент ВЭЖХ для разделения ASM8 и продуктов разрушения
Хроматограмма на фигуре 14 показывает профиль элюирования отдельных продуктов ASM8 (TOP004 и TOP005) при анионообменной хроматографии на DEAE. Объем 25 мкл свежеприготовленного ASM8 (0,5 мг/мл) фракционировали на DEAE-анионообменной колонке. В условиях градиента, описанных выше, TOP004 элюировался раньше, чем TOP005; что согласуется с тем, что TOP004 на 2 нуклеотида короче, чем TOP005, и имеет меньше отрицательно заряженных остатков. Олигонуклеотид TOP004 элюировался в точке 81,3 минуты и составлял 48,0% общего материала, поглощающего при 260 нм. TOP005 элюировался в точке 86,8 минуты и составлял 49,3% общего материала, поглощающего при 260 нм.
Чтобы подтвердить адекватное разделение TOP004, TOP005 и продуктов разрушения ASM8, осуществляли двухстадийное химическое разрушение ASM8. Стадию расщепления оставляли без изменения, но период инкубации стадии депуринизации составлял от 3 до 6 часов. Как показано на фигуре 15, ASM8 (0,5 мг/мл) обрабатывали CH3COOH в течение 3 часов и подвергали щелочному лизису (как описано выше) перед фракционированием посредством DEAE-анионообменной хроматографии. Олигонуклеотид TOP004 элюировался в точке 81,5 минуты и составлял 32,4% всего материала, поглощающего при 260 нм. Продукт TOP005 элюировался в точке 86,9 минуты и составлял 28,0% всего материала. Минорные пики представляли продукты разрушения ASM8. Как показано на фигуре 16, ASM8 (0,5 мг/мл) обрабатывали CH3COOH в течение 6 часов и подвергали щелочному лизису, как описано выше, перед фракционированием посредством DEAE-анионообменной хроматографии. При регистрации при 260 нм олигонуклеотид TOP004 элюировался в точке 82,2 минуты, а TOP005 элюировался в точке 86,7 минуты. При увеличении времени депуринизации доля TOP004 снижалась до 20,6%, а TOP005 до 14,5%. Степень разрушения TOP005, по-видимому, немного выше в указанных экспериментальных условиях. Как видно на хроматограммах на фигурах 15 и 16, увеличение времени депуринизации усиливало разрушение ASM8.
Как показано на фигурах 17A1, 17A2, 17B1 и 17B2, оценивали химическую стабильность ASM8 при разных температурах хранения. ASM8 (0,5 мг/мл в PBS) инкубировали при -20°C, 4°C, 30°C или 40°C в течение 4 недель (фигура 17A1 и 17A2) и 8 недель (фигура 17B1 и 17B2) и анализировали посредством DEAE-анионообменной хроматографии. Различные температуры хранения, проверяемые в данном эксперименте, не влияли на профиль элюирования компонентов ASM8. Не наблюдали существенного разрушения ASM8 ни при каких температурах, при которых хранили ASM8 вплоть до 2 месяцев.
Способ разделения, основанный на DEAE-анионообменной ВЭЖХ, для ASM8 описан выше. Вследствие природы данного продукта предпочтительно адекватное разделение компонентов, составляющих ASM8 (олигонуклеотидов TOP004 и TOP005). В описанных условиях градиента время удержания TOP004 было более чем на 5 минут раньше, чем время удержания TOP005 с очень небольшим перекрыванием двух пиков.
Способ также способен выявлять продукты разрушения ASM8. Химическую стабильность ASM8 при разных температурах, влажности и условиях освещения можно оценить способом на основе ВЭЖХ.
Препарат ASM8 в PBS был химически стабильным, и не выявляли значимых продуктов разрушения способом ВЭЖХ после хранения в указанном диапазоне температур в течение периода времени до 2 месяцев.
Пример 7
Термодинамическая оценка ASM8
Следующие эксперименты проводили для того, чтобы убедиться в том, что две нити олигонуклеотидов, TOP004 и TOP005, не взаимодействовали в растворе, используя при этом термодинамические оценки.
TOP004 и TOP005 смешивали в эквимолярных концентрациях в 1×PBS (а также в других буферных системах). Суммарная концентрация олигонуклеотидов была в диапазоне примерно от 1,2 до 8,7 мкМ. Осуществляли стандартные способы УФ-термоденатурации, используя спектрофотометр Beckman DU640 с дополнительным устройством для анализа т.пл. Изменение поглощения регистрировали при 260 нм при изменении температуры на каждый градус от 10 до 90°C. Осуществляли подгонку кривых плавления, используя компьютерную программу MELTWIN™ 3.5, чтобы определить термодинамические параметры. Получали картины скрининга кривых плавления и сводные таблицы термодинамических данных.
На фигуре 18 показаны кривые плавления TOP004 и TOP005 в 1×PBS. Фигура 19 представляет сводные данные термодинамики, основанные на результатах подгонок кривых плавления для TOP004 и TOP005 в 1×PBS. Результаты показывают, что ни одна из комбинаций олигонуклеотидов/условий не давала значимого перехода (прыжка поглощения) в профиле плавления при увеличении температуры. Это свидетельствует о том, что тестированные смеси олигонуклеотидов не образуют значимых взаимодействий вторичной структуры в условиях тестируемых буферов.
Пример 8
Токсичность ASM8 у макак-крабоедов
В данном примере показана токсичность ASM8, состоящего из смеси 1:1 TOP004 и TOP005. Также показан токсикокинетический профиль отдельных составляющих его олигонуклеотидных компонентов при введении путем ингаляции один раз в сутки в течение 14 последовательных дней макакам-крабоедам. Кроме того, 14 суток ингаляционного воздействия ASM8 не вызывало состояния системной гиперчувствительности, выявляемого при интрадермальной инъекции (ID).
Оценка достигаемой дозы
Общие концентрации аэрозоля при воздействии
Осуществляли всестороннюю оценку смертности, клинических признаков, массы тела, потребления пищи, электрокардиографии, офтальмоскопии и клинической патологии. Серийные образцы крови получали в первый и последний день воздействия и в конце периода восстановления, и в конце собирали ткани для определения содержания отдельных олигонуклеотидов. Кроме того, на 25 день животным, выбранным для фазы восстановления, давали интрадермальную инъекцию (ID) ASM8, чтобы оценить возможную системную гиперчувствительность. Всех животных подвергали эвтаназии после 14 дней воздействия (15 день) или после 14-дневного периода восстановления (29 день) и подвергали полной аутопсии со сбором полного набора тканей от каждого животного.
Гистопатологическая оценка состояла в микроскопическом исследовании всех тканей животных в группе, обработанной высокой дозой, и в контрольной группе, и тканей дыхательного пути в группе с более низкой дозой и у животных после восстановления.
Препарат ASM8 легко подвергался аэрозолизации, и при этом получали аэрозоли для воздействия, которые были постоянно стабильными и вдыхаемыми со значениями масс-медианного диметра (MMAD) между группами и геометрического стандартного отклонения (GSD) от 1,7 до 1,8 мкм и от 2,12 до 2,22, соответственно. Получаемые в результате оцениваемые достигаемые дозы были близки к целевым при 0,05, 0,22 и 2,5 мг/кг/сутки для групп 2-4, соответственно, таблица 7-8.
Не наблюдали гибели, и обезьяны хорошо переносили дозы. Не наблюдалось влияния на массу тела, потребление пищи, электрокардиографию, офтальмоскопию или параметры клинической патологии, и в тесте гиперчувствительности не выявили влияния введения ASM8. После вскрытия при измерении массы органов не получили доказательства токсичности. Макроскопические исследования всех органов выявили только слабое обесцвечивание почек у обработанных ASM8 животных. Однако вследствие отсутствия подтверждающих микроскопических изменений, данных клинической патологии или изменений массы органов и того факта, что обесцвечивание не наблюдали после 14 дней восстановления, указанное наблюдение считали неясным с точки зрения биологического и токсикологического значения.
Уровни в плазме TOP004 и TOP005, а также их ближайших (n-1) метаболитов были очень низкими в плазме, при этом в группах, обработанных низкой и средней дозой, ниже предела количественного определения. В случае группы, обработанной высокой дозой (2,5 мг/кг/сутки), концентрации TOP004 и TOP005 обычно были самыми высокими в любой из двух самых ранних временных точек сбора образцов 0,5 часа после введения дозы или во временной точке 1 час. В большинстве временных точек после введения дозы средняя концентрация TOP004 была сходной с концентрацией TOP005, фигура 20A и фигура 20B.
Не происходило накопления какого-либо олигонуклеотидного компонента (или его n-1-метаболитов) в плазме при многократном ежедневном введении в течение 14 дней, как показано на фигуре 21A и фигуре 2B. Не было явных половых различий в концентрациях в плазме. Значительный процент циркулирующего олигонуклеотида присутствовал в виде ближайшего n-1-метаболита как TOP004, так и TOP005, хотя процентное содержание имело тенденцию к немного более низким значениям в случае TOP004. В случае обоих олигонуклеотидов и их n-1-метаболитов клиренс из крови (плазмы) наблюдался на протяжении 24-часового периода сбора. При умерщвлении (через день после последней ингаляции дозы ASM8) заметные количества интактных олигонуклеотидных компонентов ASM8 (TOP004 и TOP005) выявляли в трахее животных, обработанных высокой дозой. В конце 14-дневного периода восстановления (29 день) уровни TOP004 и его n-1-метаболита снижались по сравнению с 15 днем и измерялись на уровне, немного превышающем предел определения анализа. В отличие от этого ни TOP005, ни его метаболит не поддавались количественному определению во временной точке умерщвления после восстановления. Полученные результаты свидетельствуют, что TOP004 обладает более высокой стабильностью в тканях, чем TOP005.
Связанные с обработкой микроскопические изменения не наблюдали ни в одном органе, за исключением дыхательного пути. Все наблюдаемые изменения в дыхательном пути классифицировали по шкале из 4 точек как наименьшие (минимальные). Изменения, которые были отмечены, в случае легких включали: пенистые альвеолярные макрофаги у животных, которым вводили дозу 0,22 или 2,5 мг/кг/сутки, внутриальвеолярное гранулоцитарное воспаление в случае 2,5 мг/кг/сутки, фокальную геморрагию у двух животных и фокальную бронхиолярную метаплазию у одного животного, которому вводили дозу 2,5 мг/кг/сутки; в случае носовой полости: очаги эрозии плоскоклеточного эпителия перегородки носа у 2/6 животных, которым вводили дозу 2,5 мг/кг/сутки, сопровождающиеся острым воспалением и воспалительным экссудатом у одной обезьяны; и в случае бронхиальных лимфатических узлов: пенистые макрофаги у животных, которым вводили дозу 2,5 мг/кг/сутки. Тяжесть изменений, наблюдаемых в легких животных, которым вводили среднюю и высокую дозу, были минимальными и не сопровождались симптомами локального повреждения или клеточной инфильтрации в паренхиму легкого. Воспалительные клетки были редкими и наблюдались только у небольшого количества животных, обработанных высокой дозой ASM8, и распределение изменений соответствовало ингаляции тестируемого материала. Фокальные геморрагии были очень маленькими и вероятно могут быть интерпретированы как случайные. Таким образом, указанные изменения, как правило, согласуются с нормальными легочными механизмами, связанными с фагоцитозом и очищением ингалируемого тестируемого материала. Отмена обработки в течение 14 дней приводила к сохранению присутствия небольшого количества альвеолярных макрофагов без воспаления у одного из двух животных, обработанных высокой дозой. Указанное наблюдение согласуется с постепенной регрессией повреждений и указывает на то, что не происходило прогрессирующего или стойкого изменения паренхимы легкого.
Не обнаружено доказательств влияния обработки тканей носа после 14-дневного периода восстановления. Что касается данных, полученных для бронхиальных лимфатических узлов у животных, обработанных высокой дозой, то пенистые макрофаги в медуллярных синусах согласуются с представлением об очистке тестируемого материала посредством выделения лимфы из легкого. Не получено доказательства повреждения паренхимы, и лимфатические узлы, по-видимому, не находились в реактивном состоянии.
В заключение, ингаляция ASM8 в течение 14 последовательных дней в определенных достигаемых дозах до 2,5 мг/кг/сутки хорошо переносится и не оказывает влияния на массу тела, потребление пищи, электрокардиографию, массу органов, офтальмоскопию или параметры клинической патологии, и при тестировании гиперчувствительности не выявлено влияние введения ASM8. Ряд в основном минимальных гистоморфологических изменений отмечен в легких (0,22 и 2,5 мг/кг/сутки), а также в полости носа и бронхиальных лимфатических узлах (2,5 мг/кг/сутки). Указанные изменения уменьшались по тяжести или отсутствовали после 14 дней восстановления.
Хотя в данной публикации описаны предпочтительные варианты осуществления изобретения, специалистам в данной области будет понятно, что могут быть сделаны изменения, не отходя от сути изобретения или не выходя за рамки объема прилагаемой формулы изобретения.









| название | год | авторы | номер документа |
|---|---|---|---|
| МОЛЕКУЛЫ АНТИТЕЛ ЧЕЛОВЕКА К IL-13 | 2004 |
|
RU2387667C2 |
| АНТИ-GM-CSF АНТИТЕЛА И ИХ ПРИМЕНЕНИЕ | 2006 |
|
RU2447085C2 |
| СПОСОБЫ ДИАГНОСТИКИ И ЛЕЧЕНИЯ, СВЯЗАННЫЕ С ИНГИБИРОВАНИЕМ ТН2 | 2011 |
|
RU2737245C2 |
| СПОСОБЫ ДИАГНОСТИКИ И ЛЕЧЕНИЯ, СВЯЗАННЫЕ С ИНГИБИРОВАНИЕМ TH2 | 2011 |
|
RU2578468C2 |
| СПОСОБ ИНГИБИРОВАНИЯ GMCSFRa У ПАЦИЕНТА | 2012 |
|
RU2639546C2 |
| МАТЕРИАЛЫ И СПОСОБЫ ЛЕЧЕНИЯ РАКА | 2019 |
|
RU2787828C2 |
| АНТИ-IL-17F АНТИТЕЛА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2010 |
|
RU2605318C2 |
| ТЕРАПЕВТИЧЕСКИЕ И ДИАГНОСТИЧЕСКИЕ СПОСОБЫ ДЛЯ ВОСПАЛИТЕЛЬНЫХ ЗАБОЛЕВАНИЙ, ОПОСРЕДОВАННЫХ ТУЧНЫМИ КЛЕТКАМИ | 2019 |
|
RU2795180C2 |
| Новая форма IL33, мутировавшие формы IL33, антитела, анализы и способы их применения | 2016 |
|
RU2736299C2 |
| СПОСОБЫ ДИАГНОСТИКИ И ЛЕЧЕНИЯ, СВЯЗАННЫЕ С ИНГИБИРОВАНИЕМ TH2 | 2020 |
|
RU2832013C2 |

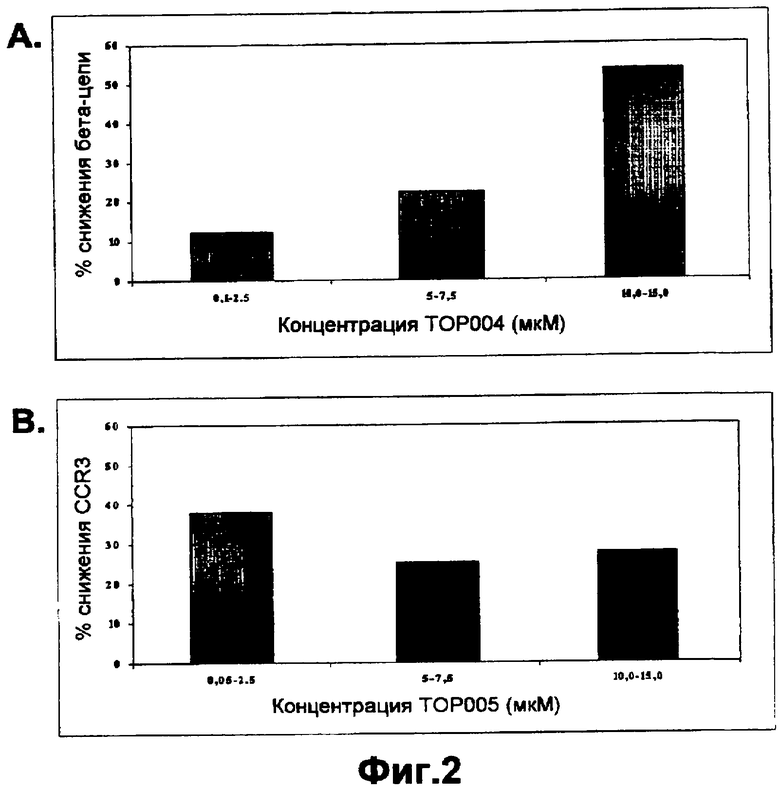
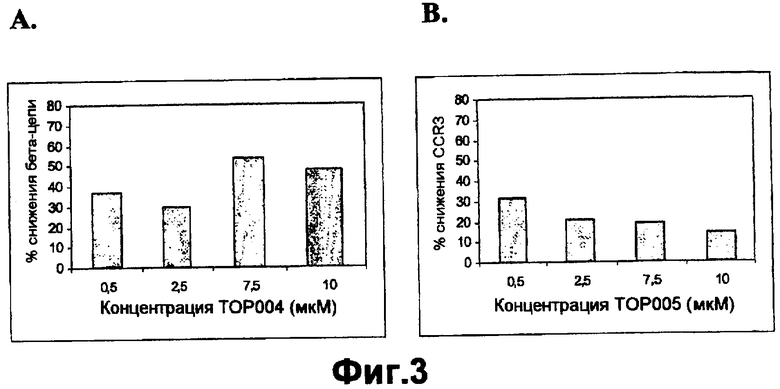
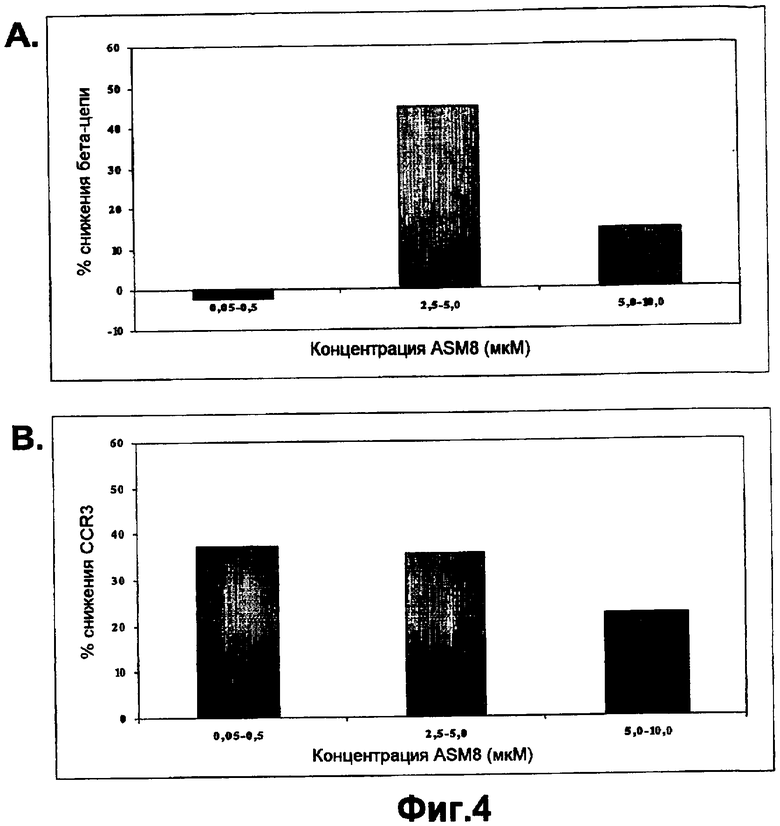



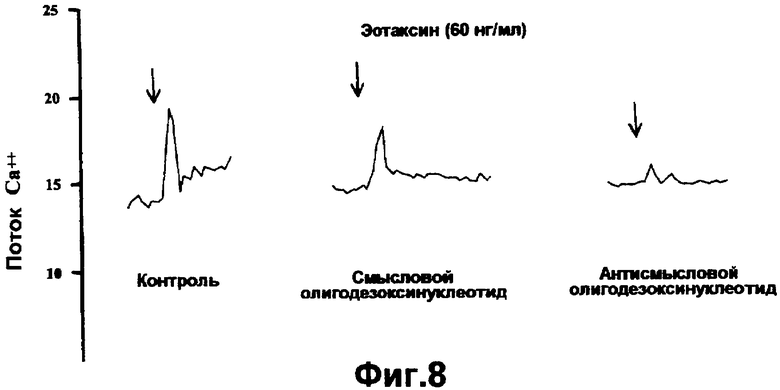
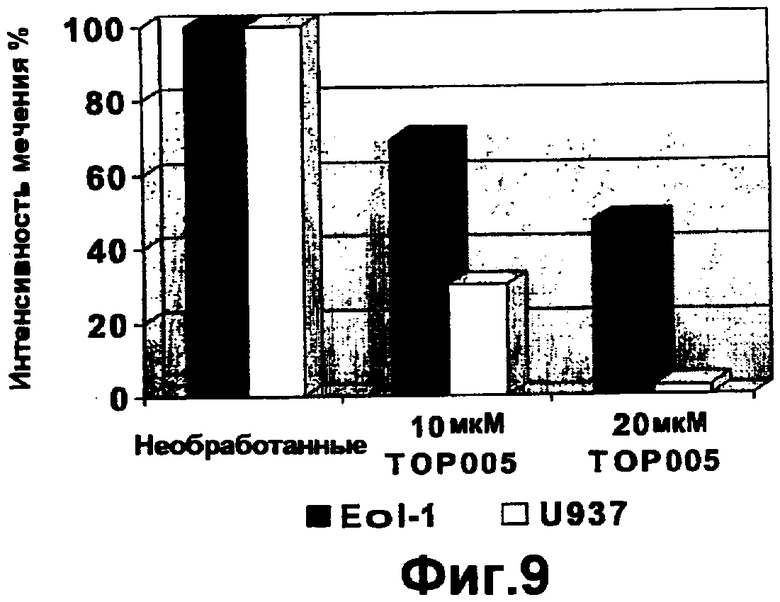


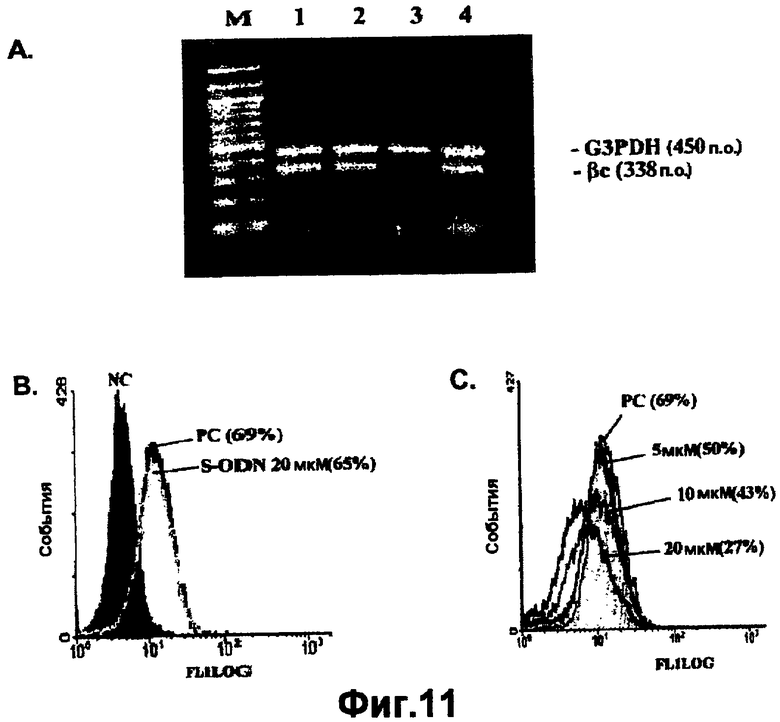
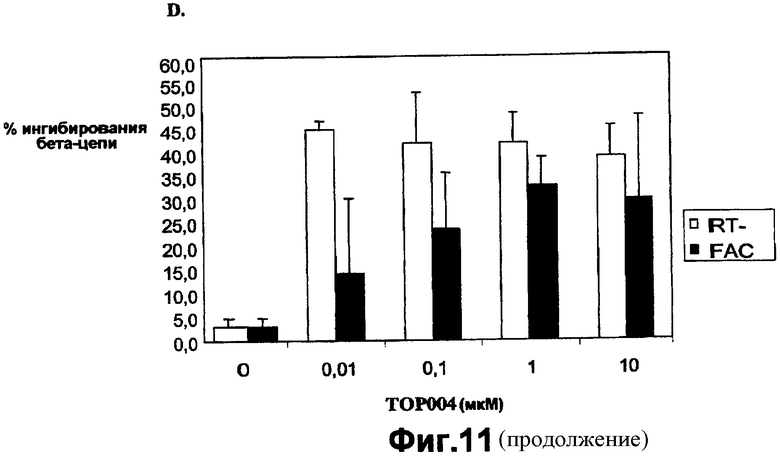



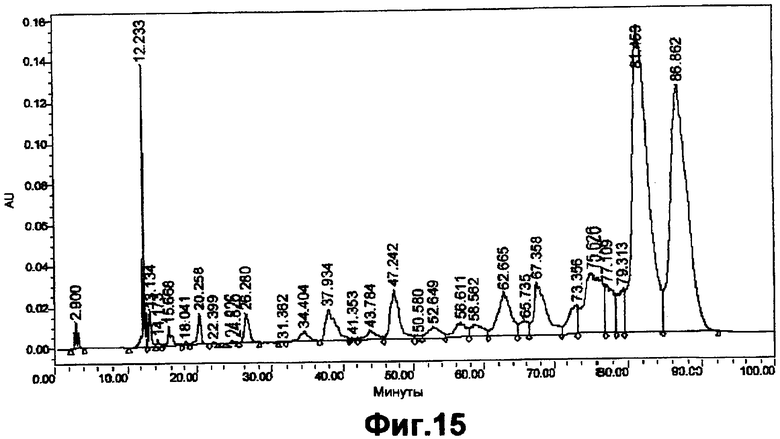
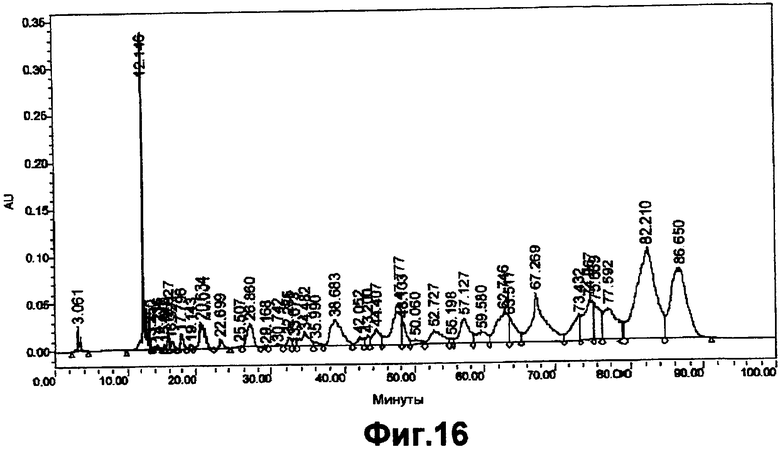

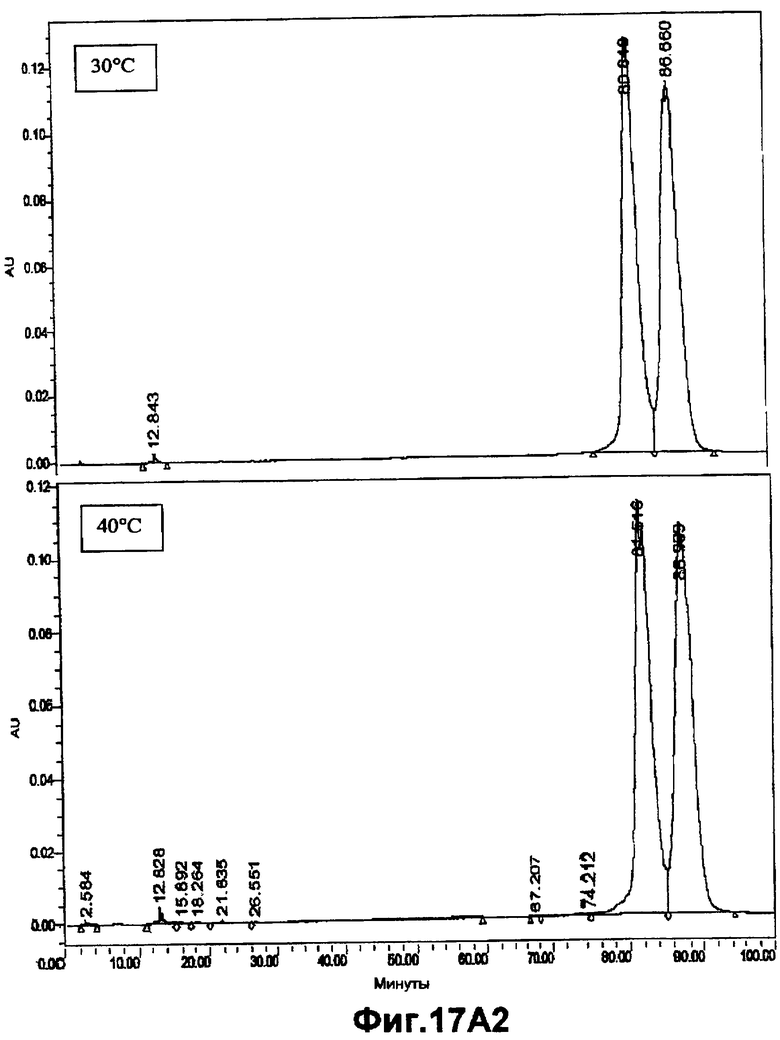



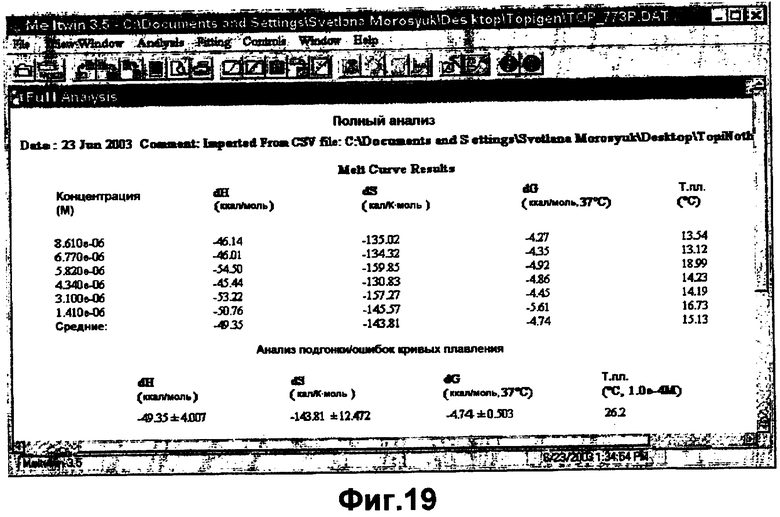


Изобретение относится к области медицины и касается антисмысловых олигонуклеотидов для лечения и/или профилактики, по меньшей мере, одного из заболеваний: астмы, гиперэозинофилии. Сущность изобретения включает олигонуклеотиды, направленные против последовательностей нуклеиновых кислот, кодирующих рецептор, выбранный из группы, состоящей из рецептора CCR3 и общей субъединицы рецепторов IL-3, IL-5 и GM-CSF с последовательностями SEQ ID NO: 1 и SEQ ID NO: 14. Преимущество изобретения заключается в снижении токсичности по сравнению с гормональными препаратами. 8 н. и 31 з.п. ф-лы, 11 табл., 21 ил.
1. Антисмысловой олигонуклеотид, который направлен против последовательности нуклеиновой кислоты, кодирующей рецептор хемокина CCR3, где олигонуклеотид выбран из группы, состоящей из (i) олигонуклеотида с последовательностью, выбранной из группы, состоящей из SEQ ID NO. 1 и SEQ ID NO. 14, и (ii) модифицированного олигонуклеотида с последовательностью, выбранной из группы, состоящей из SEQ ID NO. 1 и SEQ ID NO. 14.
2. Антисмысловой олигонуклеотид по п.1, где олигонуклеотид имеет последовательность, содержащую последовательность SEQ ID NO. 1.
3. Антисмысловой олигонуклеотид по п.1, где олигонуклеотид имеет последовательность, содержащую последовательность SEQ ID NO. 14.
4. Антисмысловой олигонуклеотид по п.1, где антисмысловой олигонуклеотид предназначен для лечения и/или профилактики астмы.
5. Антисмысловой олигонуклеотид по п.1, где антисмысловой олигонуклеотид предназначен для лечения и/или профилактики гиперэозинофилии.
6. Применение, по меньшей мере, одного антисмыслового олигонуклеотида по п.1 для лечения и/или профилактики, по меньшей мере, одного из заболеваний: астмы и гиперэозинофилии.
7. Применение по п.6, где, по меньшей мере, один антисмысловой олигонуклеотид имеет последовательность, содержащую последовательность SEQ ID NO. 1.
8. Применение по п.6, где, по меньшей мере, один антисмысловой олигонуклеотид имеет последовательность, содержащую последовательность SEQ ID NO. 14.
9. Применение по п.6, где, по меньшей мере, один антисмысловой олигонуклеотид представляет собой, по меньшей мере, два антисмысловых олигонуклеотида, имеющих последовательности, содержащие последовательности SEQ ID NO. 13 и SEQ ID NO. 14 соответственно.
10. Применение по п.6, где, по меньшей мере, один олигонуклеотид применяют для лечения и/или профилактики астмы.
11. Применение по п.6, где, по меньшей мере, один олигонуклеотид применяют для лечения и/или профилактики гиперэозинофилии.
12. Фармацевтическая композиция для лечения и/или профилактики, по меньшей мере, одного из заболеваний: астмы и гиперэозинофилии, где композиция содержит, по меньшей мере, один антисмысловой олигонуклеотид по п.1 вместе с фармацевтически приемлемым носителем.
13. Фармацевтическая композиция по п.12, где, по меньшей мере, один антисмысловой олигонуклеотид имеет последовательность, содержащую последовательность SEQ ID NO. 1.
14. Фармацевтическая композиция по п.12, где, по меньшей мере, один антисмысловой олигонуклеотид имеет последовательность, содержащую последовательность SEQ ID NO. 14.
15. Фармацевтическая композиция по п.12, где, по меньшей мере, один антисмысловой олигонуклеотид представляет собой, по меньшей мере, два антисмысловых олигонуклеотида, имеющих последовательности, содержащие последовательности SEQ ID NO. 13 и SEQ ID NO. 14 соответственно.
16. Фармацевтическая композиция по любому из пп.12-15, где фармацевтическая композиция является композицией для местного применения.
17. Фармацевтическая композиция по п.12, где фармацевтическая композиция предназначена для лечения и/или профилактики астмы.
18. Фармацевтическая композиция по п.12, где фармацевтическая композиция предназначена для лечения и/или профилактики гиперэозинофилии.
19. Фармацевтическая композиция по п.12, где, по меньшей мере, один антисмысловой олигонуклеотид представляет собой, по меньшей мере, два олигонуклеотида, имеющих последовательности SEQ ID NO. 1 и SEQ ID NO. 13.
20. Фармацевтическая композиция по п.12, где, по меньшей мере, один антисмысловой олигонуклеотид представляет собой два олигонуклеотида, имеющих последовательности SEQ ID NO. 13 и SEQ ID NO. 14.
21. Фармацевтическая композиция по п.12, где, по меньшей мере, один антисмысловой олигонуклеотид представляет собой, по меньшей мере, два олигонуклеотида, имеющих последовательность SEQ ID NO. 1 и последовательность, имеющую номер доступа NM_000398-1 в Gene Bank.
22. Фармацевтическая композиция по п.12, где, по меньшей мере, один антисмысловой олигонуклеотид представляет собой, по меньшей мере, два олигонуклеотида, имеющих последовательность SEQ ID NO. 14 и последовательность, имеющую номер доступа NM_000398-1 в Gene Bank.
23. Применение фармацевтической композиции по любому из пп.12-22 для лечения и/или профилактики, по меньшей мере, одного из заболеваний: астмы и гиперэозинофилии.
24. Применение по п.23, где фармацевтическую композицию применяют для лечения и/или профилактики астмы.
25. Применение по п.23, где фармацевтическую композицию применяют для лечения и/или профилактики гиперэозинофилии.
26. Способ лечения и/или профилактики, по меньшей мере, одного из заболеваний: астмы и гиперэозинофилии, где способ включает стадию введения эффективного количества, по меньшей мере, одного антисмыслового олигонуклеотида по п.1.
27. Способ по п.26, где, по меньшей мере, один антисмысловой олигонуклеотид имеет последовательность, содержащую последовательность SEQ ID NO. 1.
28. Способ по п.26, где, по меньшей мере, один антисмысловой олигонуклеотид имеет последовательность, содержащую последовательность SEQ ID NO. 14.
29. Способ по п.26, где, по меньшей мере, один олигонуклеотид представляет собой, по меньшей мере, два антисмысловых олигонуклеотида, имеющих последовательности, содержащие последовательности SEQ ID NO. 13 и SEQ ID NO. 14 соответственно.
30. Способ по п.26, где способ является способом лечения и/или профилактики астмы.
31. Способ по п.26, где способ является способом лечения и/или профилактики гиперэозинофилии.
32. Способ лечения и/или профилактики, по меньшей мере, одного из заболеваний: астмы и гиперэозинофилии, где способ включает в себя стадию введения эффективного количества фармацевтической композиции по любому из пп.12-22.
33. Способ по п.32, где способ является способом лечения и/или профилактики астмы.
34. Способ по п.32, где способ является способом лечения и/или профилактики гиперэозинофилии.
35. Применение, по меньшей мере, одного антисмыслового олигонуклеотида по п.1 для получения лекарственного средства для применения при лечении астмы.
36. Применение, по меньшей мере, одного антисмыслового олигонуклеотида по п.1 для получения лекарственного средства для применения при лечении гиперэозинофилии.
37. Применение по любому из пп.35-36, где, по меньшей мере, один олигонуклеотид имеет последовательность, содержащую последовательность SEQ ID NO. 1.
38. Применение по любому из пп.35-36, где, по меньшей мере, один олигонуклеотид имеет последовательность, содержащую последовательность SEQ ID NO. 14.
39. Применение по любому из пп.35-36, где, по меньшей мере, один олигонуклеотид представляет собой, по меньшей мере, два олигонуклеотида, имеющих последовательности, содержащие последовательности SEQ ID NO. 13 и SEQ ID NO. 14 соответственно.
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| Прибор, замыкающий сигнальную цепь при повышении температуры | 1918 |
|
SU99A1 |
| ALLAM M | |||
| et al | |||
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |

