Настоящее изобретение относится к способу ингибирования или нейтрализации активности альфа цепи рецептора колониестимулирующего фактора гранулоцитов/макрофагов (GM-CSFRα), и, в частности, относится к молекулам анти-GMCSFRα антитела (т.е. антитела к GMCSFRα) для применения по указанному назначению, и способам их получения. Изобретение также относится к лечению воспалительных, респираторных и аутоиммунных заболеваний, опосредуемых GMCSFRα, включая ревматоидные артриты, хроническую обструктивную болезнь легких и рассеянный склероз посредством ингибирования связывания GM-CSF с GM-CSFRα человека.
GM-CSF является провоспалительным цитокином 1 типа, который увеличивает выживаемость, пролиферацию и/или дифференцировку широкого спектра гемопоэтических клеток, включая нейтрофилы, эозинофилы, макрофаги и клетки-предшественники указанных клеток. Рецептор GM-CSF является членом суперсемейства рецепторов гематопоэтинам. Он представляет собой гетеродимер, состоящий из субъединиц альфа и бета. Субъединица альфа обладает высокой специфичностью к GM-CSF, а субъединица бета является общей с другими рецепторами цитокинов, включая рецепторы IL3 и IL5. Данный феномен отражает более широкое распространение субъединицы бета в тканях. Субъединица альфа, GM-CSFRα, экспрессируется главным образом на поверхности миелоидных и клеток, не принадлежащих к гематопоэтическому ряду, таких как нейтрофилы, макрофаги, эозинофилы, отросчатые клетки, клетки эндотелия и клетки дыхательного эпителия. Полноразмерный GM-CSFRα представляет собой мембранный гликопротеин 1 типа длиной 400 аминокислот, который принадлежит к семейству рецепторов цитокинов 1 типа, и состоит из сигнального пептида длиной 22 аминокислоты (положение 1-22), внеклеточного домена длиной 298 аминокислот (положение 23-320), трансмембранного домена, располагающегося от положения 321 до положения 345 и короткого внутриклеточного домена, состоящего из 55 аминокислот. Отщепление сигнального полипептида приводит к образованию зрелой формы GM-CSFRα, состоящей из 378 аминокислот.
Клоны кДНК GM-CSFRα мыши и человека являются доступными, и на уровне белка субъединицы рецепторов имеют 36% идентичность. GM-CSF способен связываться с субъединицей α отдельно с относительно низкой аффинностью (Kd 1-5 нМ), но не способен связываться с отдельными субъединицами β. Однако, присутствие обеих субъединиц (α и β) дает в результате высокоаффинный комплекс лиганд-рецептор (Kd≈100 пМ). Передача сигнала GM-CSF осуществляется путем его первоначального связывания с α-цепью GM-CSFR и последующего перекрестного связывания с большей субъединицей общей β-цепи с возникновением высокоаффинного взаимодействия, которое фосфорилирует путь JAK-STAT. Связывание GM-CSFR с GMCSF рассмотрено в источнике [1]. Данное взаимодействие также обеспечивает передачу сигналов через фосфорилирование тирозина и активации МАР-киназного пути. С точки зрения патологии, было показано, что GM-CSF играет роль в обострении воспалительных, респираторных и аутоиммунных заболеваний. Нейтрализация связывания GM-CSF с GM-CSFRα является, следовательно, терапевтическим подходом к лечению заболеваний и состояний, опосредуемых GM-CSFR.
У Nicola с соавт. [2] описано антитело мыши к GM-CSFRα человека, обозначаемое 2В7-17-А или "2В7", которое, как было показано с помощью нескольких разных биологических тестов, обладает сравнительно высокой аффинностью к GM-CSFRα человека и является сильным ингибитором биологического действия GM-CSF человека.
Антитело 2В7 можно приобрести в Chemicon как продукт МАВ1037, в спецификации которого отмечено, что МАВ1037 является сильным ингибитором биологического действия GM-CSF. 2В7 описано также в WO 94/09149.
Применяя сочетание отборов на фаговых библиотеках интактных (naive) одноцепочечных фрагментов scFv, неспецифичного мутагенеза и соответствующих биохимических и биологических методов анализа (см. Экспериментальную часть ниже) авторы настоящего изобретения идентифицировали молекулы высокоактивных антител, которые связывают GM-CSFRα человека и ингибируют действие GM-CSF человека на его рецептор.
Представленные здесь результаты показывают, что такие антитела связывают с областью или эпитопом GM-CSFRα, которые отличается от эпитопов, которые связывает известное антитело к GM-CSFRα 2B7, и при этом с эффективностью даже превосходящей эффективность антитела 2B7, что продемонстрировано в различных биологических тестах.
Соответственно, настоящее изобретение относится к связывающим агентам, которые связывают GM-CSFRα человека и ингибируют связывание GM-CSF с GM-CSFRα и способны ингибировать и нейтрализовать его активность. Связывающие агенты согласно настоящему изобретению могут быть антагонистами GM-CSFR. Связывающие агенты могут быть конкурентными обратимыми ингибиторами передачи сигнала т GM-CSF через GM-CSFR.
Антитела и другие связывающие агенты согласно настоящему изобретению особенно ценны в связывании и нейтрализации GM-CSFRα и, таким образом, их можно применять в лечении заболеваний, опосредуемых GM-CSFRα, включая воспалительные и аутоиммунные заболевания, как показано в экспериментах, приведенных в данном тексте и в дополнительной технической литературе. Например, мы продемонстрировали в тесте, основанном на клеточной системе, что антитела согласно настоящему изобретению способны ингибировать высвобождение цитокинов (например, IL-6 и TNFα), индуцированное связыванием нативного GM-CSF с его рецептором. Как будет более подробно объяснено ниже, ингибирование активности GM-CSF путем блокирования связывания с GM-CSFRα является терапевтическим подходом к лечению таких заболеваний, как ревматоидный артрит (РА), астма, связанное с курением воспаление дыхательных путей, хроническая обструктивная болезнь легких (ХОБЛ), аллергическая реакция, рассеянный склероз (PC), миелоидная лейкемия и атеросклероз.
Связывающие агенты согласно настоящему в целом связываются с внеклеточным доменом GM-CSFRα. Предпочтительно, чтобы связывающий агент согласно настоящему изобретению связывал, по меньшей мере, один остаток последовательности Tyr-Leu-Asp-Phe-Gln (YLDFQ), SEQ ID NO: 201, в положении от 226 до 230 в зрелого GM-CSFRα человека (SEQ ID NO: 206). Связывающий агент может связывать, по меньшей мере, один остаток в последовательности YLDFQ GM-CSFRα человека, например, он может связывать один, два, три или четыре остатка последовательности YLDFQ. Таким образом, связывающий агент может распознавать один или несколько остатков в пределах этой последовательности и, возможно, может также связывать дополнительные фланкирующие остатки или остатки, находящиеся по соседству в структуре внеклеточного домена GM-CSFRα.
Связывание можно определить любым подходящим способом, например, можно использовать сканирование связывания пептидов, такое как твердофазный иммуноферментный анализ, основанный на системе PEPSCAN, как более подробно описано далее в тексте. В сканировании о связывания пептидов, как, например, в системе, поставляемой PEPSCAN Systems, короткие перекрывающиеся пептиды, полученные из антигена, методично исследуют на предмет связывания со связывающим агентом. Пептиды могут быть ковалентно связаны с поверхностью подложки с образованием матрицы (массива) пептидов. Вкратце, сканирование связывания пептидов (например, "PEPSCAN") включает идентификацию (например, с применением метода твердофазного ИФА) массива пептидов, которые связывает связывающий агент, причем пептиды имеют последовательности аминокислот, соответствующие фрагментам SEQ ID NO: 206 (напр. пептиды, состоящие из примерно 15 последовательных остатков SEQ ID NO: 206) и выравнивание пептидов для определения «отпечатка» (footprint) остатков, связанных связывающим агентом, где отпечаток включает общие для перекрывающихся пептидов остатки. В соответствии с настоящим изобретением отпечаток, полученный методом сканирования связывания пептидов или PEPSCAN, может включать, по меньшей мере, один остаток YLDFQ, соответствующей положениям от 226 до 230 последовательности SEQ ID NO: 206. Отпечаток может включать один, два, три, четыре или все остатки YLDFQ. Связывающий агент согласно настоящему изобретению, может связывать пептидный фрагмент (напр. из 15 остатков) последовательности SEQ ID NO: 206, включающий один или несколько, предпочтительно - все, из остатков YLDFQ, соответствующей положениям от 226 до 230 последовательности SEQ ID NO: 206, например, определенных путем сканирования связывания пептидов или описанным здесь методом PEPSCAN. Таким образом, связывающий агент согласно настоящему изобретению может связывать пептид, имеющий последовательность аминокислот из 15 последовательных остатков последовательности SEQ ID NO: 206, который включает, по меньшей мере, один остаток YLDFQ или, по меньшей мере, частично перекрывается с YLDFQ в положениях с 226 по 230 последовательности SEQ ID NO: 206. Детали метода сканирования пептидов, подходящего для определения связывания, подробно изложены ниже в данном тексте. Для определения связывания остатков антителом и/или для подтверждения результатов метода сканирования пептидов (напр.PEPSCAN), можно применять другие хорошо известные способы, включая сайт-направленный мутагенез, водородно-дейтериевый обмен, масс-спектрометрию, ЯМР, рентгенокристаллографию.
Таким образом, связывающий агент согласно настоящему изобретению предпочтительно нейтрализует GM-CSFRα. Нейтрализация означает уменьшение или ингибирование биологической активности GM-CSFRα, например, уменьшение или ингибирование связывания GM-CSF с GM-CSFRα или передачи сигнала GM-CSFRα, которые измеряют по опосредуемым GM-CSFRα ответам. Уменьшение или ингибирование биологической активности может быть частичным или полным. Степень, в которой антитело нейтрализует GM-CSFRα, называется его эффективностью нейтрализации. Эффективность можно определить или измерить, применяя один или несколько тестов, известных специалистам в данной области и/или в соответствии с описанными или представленными в настоящей заявке тестами. Например, связывающий агент может обладать нейтрализующей активностью в одном или нескольких из следующих тестов:
- Биохимический анализ связывания лигандов
- Тесте на пролиферацию TF-1
- Тесте на изменение формы гранулоцитов человека
- Тесте на изменение формы гранулоцитов примата Cynomolgus (яванский макак), не являющегося человеком
- Тесте на высвобождение TNFα моноцитами
- Тесте на выживаемость гранулоцитов
- Тесте на колониеобразование (ингибировании in vitro GM-CSF связанной дифференциации предшественников эритроцитов)
- Ингибирование in vitro биоактивности GM-CSF, например, у химерных мышей с трансгенным костным мозгом, экспрессирующим GM-CSFR человека
- Тесте на высвобождение цитокинов мононуклеарами периферической крови
Если не указано другое, активность обычно выражают как значение IC50, измеряемое в пМ. В функциональных тестах, IC50 - это концентрация, которая уменьшает биологических ответ на 50% от его максимума. В изучении связывания лигандов, IC50 - это концентрация, которая уменьшает связывание рецептора на 50% от максимального уровня специфичного связывания. IC50 можно рассчитать путем построения графика: % максимального биологического ответа (представленного, например, пролиферацией клеток, которая может быть измерена в тесте на пролиферацию по включению 3H тимидина в импульсах/мин, изменением формы в тесте на изменение формы, высвобождением TNFα в тесте на высвобождение TNFα, выживаемостью в тесте на выживаемость, количеством колоний в тесте на образование колоний или увеличением массы селезенки или уменьшением моноцитов кровяного русла у химерных мышей с трансгенным костным мозгом, экспрессирующим человеческий GM-CSFR в тесте биоактивности) или % специфичного связывания рецептора, к функции от логарифма концентрации связывающего агента, и с использованием такого программного обеспечения, как Prism (GraphPad) для аппроксимации полученных данных сигма-функцией, в результате чего получают IC50.
Значение IC50 может представлять среднее множества измерений. Таким образом, можно получить значение IC50, например, по результатам трех экспериментов, а затем рассчитать среднее значение IC50.
В тесте на пролиферацию TF-1 связывающие агенты согласно настоящему изобретению обычно имеют IC50 менее 1500 пМ, например, IC50 может быть <300, <60, <10 или <1.5 пМ, например примерно 1.0 пМ. Обычно IC50 составляет по меньшей мере 0.5 или 1.0 нМ. IC50 известного антитела 2В7 мыши в этом тесте составляла примерно 1600 пМ. Применяемый здесь тест на пролиферацию TF-1 проводили с конечной концентрацией GM-CSF человека, равной 7 пМ. Таким образом, эффективность нейтрализации IC50 в тесте на пролиферацию TF-1 представляет собой способность связывающего агента ингибировать пролиферацию клеток TF-1, индуцируемую 7 пМ GM-CSF человека. Более подробную информацию можно найти в разделе Материалы и Методы Анализа.
Связывающий агент согласно настоящему изобретению может иметь pA2 более отрицательное, чем -6, -7, -8, -9, -10, -10.5 или -11 в тесте на пролиферацию TF-1. например, рА2 может составлять примерно -10.5 или -11. Расчет и значение значений рА2 подробно обсуждается в экспериментальной части раздела Материалы и Методы Анализа.
IC50 связывающего агента согласно настоящему изобретению в тесте на изменение формы гранулоцитом человека обычно составляет менее 100 пМ, например, менее 50 пМ или менее 30, 25, 20, 15 или 10 пМ. Обычно IC50 составляет по меньшей мере 5, 6 или 7 пМ. Известное антитело 2B7 мыши, напротив, является менее эффективным, поскольку измеренное в этом тесте значение IC50 составляет 477 пМ. Применяемый здесь тест на изменение формы гранулоцитов человека проводили при конечной концентрации GM-CSF человека, равной 7 пМ. Таким образом, эффективность нейтрализации IC50 в тесте на изменение формы гранулоцитов человека представляет способность связывающего агента ингибировать изменение формы гранулоцита человека, индуцируемое 7 пМ GM-CSF человека. Более подробно см. в разделе Материалы и Методы Анализа.
В тесте на изменение формы гранулоцитов о примата Cynomolgus, не являющегося человеком, связывающий агент согласно настоящему изобретению обычно имеет IC50 менее 20 пМ, стандартно - менее 10, 5 или 2.5 пМ. Значение IC50 может составлять по меньшей мере 0.5, 1 или 1.5 пМ. При исследовании в этом тесте известное антитело 2B7 мыши имело значение IC50, равное 26 пМ. Применяемый тест на изменение формы гранулоцитов о примата Cynomolgus, не являющегося человеком, выполняли с конечной концентрацией GM-CSF человека, равной 7 пМ. Таким образом, эффективность нейтрализации IC50 тесте на изменение формы гранулоцитов о примата Cynomolgus, не являющегося человеком, представляет способность связывающего агента ингибировать изменение формы гранулоцитов Cynomolgus, индуцируемое 7 пМ GM-CSF человека. Более подробную информацию можно найти в разделе Материалы и Методы Анализа.
Связывающий агент согласно настоящему изобретению может иметь рА2 более отрицательное, чем -6, -7, -8, -9, -10, -10.5 или -11 в тесте на изменения формы у человека и/или cynomologus. Предпочтительные значения рА2 - примерно -10 или -11.
В тесте на высвобождение TNFα моноцитами связывающие агенты согласно настоящему изобретению в норме имеют IC50 менее 150 пМ, обычно - менее 110 пМ, например, менее 100 пМ. IC50 может составлять по меньшей мере 30 или 40 пМ. Используемый здесь анализ высвобождения моноцита TNFα выполнялся при конечной концентрации человеческого GM-CSF 1 пМ. Таким образом, IC50 нейтрализующая активность в анализе высвобождения моноцита TNFα показывает способность связывающего агента ингибировать высвобождение TNFα из человеческих моноцитов стимулированных 1 пМ человеческого GM-CSF. Более подробную информацию можно найти в разделе Материалы и Методы Анализа.
В тесте на выживаемость гранулоцитов, связывающие агенты согласно настоящему изобретению в норме имеют IC50 менее 1000 пМ, обычно менее 850 пМ. IC50 может быть менее чем 500, 250, 150, 100, 50, 30, 20 или 10 пМ. IC50 может быть по меньшей мере 5 пМ.
Известное антитело 2В7 мыши не активно в этом тесте при концентрации до 83 пМ. Используемый здесь анализ на выживаемость гранулоцитов выполняли с конечной концентрацией GM-CSF человека, равной 7 пМ. Таким образом, эффективность нейтрализации IC50 в тесте на выживаемость гранулоцитов представляет способность связывающего агента ингибировать выживаемость гранулоцитов человека, индуцируемую 7 пМ GM-CSF человека. Более подробную информацию можно найти в разделе Материалы и Методы Анализа.
В тесте на образование колоний связывающие агенты согласно настоящему изобретению могут иметь IC50 менее 5, менее 2.5, менее 1 или менее 0.3 мкг/мл. Предпочтительно, чтобы IC50 была 0.25 мкг/мл или менее, например, менее 0.1 мкг/мл. IC50 может составлять по меньшей мере 0.05 мкг/мл. Известное антитело 2B7 мыши имеет в этом тесте малую активность (либо активность отсутствует) при концентрации до 10 мкг/мл (67 нМ). Используемый здесь тест на образование колоний выполняли с конечной концентрацией GM-CSF человека, равной 10 нМ/мл. Таким образом, эффективность нейтрализации IC50 в тесте на образование колоний представляет способность связывающего агента ингибировать образование колонии, индуцируемое 10 нг/мл GM-CSF человека. Более подробную информацию можно найти в разделе Материалы и Методы Анализа.
Связывающий агент согласно настоящему изобретению может демонстрировать дозозависимую способность ингибировать увеличение массы селезенки и/или ингибировать индуцируемое GM-CSF снижение количества моноцитов в циркулирующей крови у химерных мышей с трансгенным костным мозгом, экспрессирующим GM-CSFR человека, которых обрабатывают GM-CSF человека. IC50 для ингибирования увеличения массы селезенки может быть менее 5, менее 2.5, менее 2, менее 1 или менее 0.75 мг/кг.
Кроме того, кинетику связывания и аффинность связывающих агентов в отношении GM-CSFRα человека можно определить, например, методом поверхностного плазменного резонанса, например, с применением BIAcore. KD связывающих агентов согласно настоящему изобретению обычно менее 5 нМ, в более предпочтительном варианте менее 4, 3, 2 или 1 нМ. Предпочтительно, чтобы KD была менее 0.9, 0.8, 0.7, 0.6, 0.5, 0.4, 0.3, 0.2 или 0.15 нМ.
Связывающие агенты согласно настоящему изобретению обычно связывают, помимо GM-CSFRα человека, GM-CSFRα отличных от человека приматов, например, GM-CSFRα яванского макака (cynomolgous). Поскольку гомология между рецепторами GM-CSF человека и мыши низка (приблизительно 36%), связывающие агенты согласно настоящему изобретению в основном не будут связываться или давать перекрестную реакцию с рецептором мыши.
Связывающий агент для применения в способах согласно настоящему изобретению обычно включает молекулу антитела, например, целого антитела или его фрагмента, как описано более подробно ниже. Предпочтительно, молекула антитела согласно настоящему изобретению представляет собой молекулу антитела человека.
Связывающий агент согласно настоящему изобретению обычно включает VH- и/или VL-домен (т.е. вариабельную область тяжелой цепи или вариабельную область легкой цепи, соответственно) антитела. VH- и VL-домены связывающих агентов также являются частью настоящего изобретения. В пределах каждого из доменов VH и VL располагаются гипервариабельные участки ("CDR") и каркасные области ("FR"), VH- домен включает набор участков HCDR, а VL-домен включает набор участков LCDR. Молекула антитела обычно включает VH-домен, включающий CDR1, CDR2 и CDR3 VH-домена и каркасную область. Кроме того, молекула антитела также может включать VL домен, включающий CDR1, CDR2 и CDR3 VL-домена и каркасную область. Каркасная область VH- или VL-домена включает четыре каркасные области FR1, FR2, FR3 и FR4 между которыми располагаются CDR в виде следующей структуры:
FR1 - CDR1-FR2-CDR2-FR3-CDR3-FR4.
Примеры VH- и VL-доменов антитела, FR-областей и CDR-участков в соответствии с данным изобретением представлены в прилагающемся перечне последовательностей, который является частью настоящей заявки. Все последовательности VH и VL, последовательности CDR, наборы CDR, HCDR и LCDR, раскрытые в настоящей заявке, представляют собой аспекты и варианты реализации настоящего изобретения. Таким образом, VH-домен связывающего агента, соответствующий настоящему изобретению, является аспектом настоящего изобретения. "Набор CDR" включает CDR1, CDR2 и CDR3. Таким образом, под набором HCDR подразумевают HCDR1, HCDR2 и HCDR3, а под набором LCDR подразумевают LCDR1, LCDR2 и LCDR3. Если не указано иное, "набор CDR" включает участки HCDR и LCDR. Типичными связывающими агентами согласно настоящему изобретению являются моноклональные антитела (mAb).
В более подробном описании в Экспериментальной части, авторы настоящего изобретения идентифицировали панель молекул антитела, которые связывают GM-CSFRα. Мы также идентифицировали определенные остатки внутри гипервариабельных участков (CDR) VH- и VL-доменов, которые особенно важны для связывания рецептора и эффективности нейтрализации. Поскольку CDR в первую очередь определяют связывание и специфичность связывающего агента, можно применять один или несколько CDR, имеющих подходящие остатки, определенные в настоящей заявке, и встраивать их в любой подходящий каркас или в остов белка, не являющегося антителом, как описано более подробно в тексте. Например, один или несколько CDR или наборы CDR антитела, можно внедрить (привить) в каркас (например, иммуноглобудина человека) с получением молекулы антитела или различных молекул антитела. Например, молекула антитела может включать CDR, раскрытый в настоящей заявке, и каркасные участки последовательностей сегментов генов зародышевых линий человека. Антитело может быть представлено набором CDR внутри каркаса, который можно подвергнуть эмбрионизации с изменением одного или нескольких остатков внутри каркаса, что обеспечивает соответствие остаткам в эквивалентных положениях в наиболее близких зародышевых линий человека. Таким образом, предпочтительно, чтобы каркасные области антитела были зародышевыми и/или из организма человека.
Мы провели исследование с целью выяснить, какие из остатков потенциально подходящего антитела важны для распознавания антигена, следуя способу, описанному в экспериментальной части, и затем выполнили анализ последовательностей 160 клонов, продемонстрировавших в биологическом тесте активность по меньшей мере в 5 превышающую активность исходного клона антитела. Результаты показали, что следующие позиции вносят вклад в связывание антигена: остатки по Kabat 27A, 27B, 27С, 32, 51, 52, 53, 90, 92 и 96 в VL-домене и остатки по Kabat 17, 34, 54, 57, 95, 97, 99 и 100В в VH-домене. В предпочтительных способах реализации настоящего изобретения один или несколько из этих остатков по Kabat представляют собой остатки по Kabat, присутствующие в данном положении в одном или нескольких клонах антител с номерами 1,2 4-20, последовательности которых раскрыты в представленном перечне последовательностей. В различных вариантах реализации остаток может совпадать с остатком находящимся в данной позиции антитела 3, или отличаться от него.
В ходе исследования мы выявили 4 положения остатков в участках CDR, которые имеют особенно сильное влияние на связывание рецептора: Н97, Н100В, L90 и L92 (нумерация по Kabat). Предпочтительно, чтобы Н97 VH CDR3 представлял собой S. Таким образом, остаток серина, обнаруженный в данном положении во всех 160 клонах, является важным остатком для распознавания антигенов.
Предпочтительно, чтобы VH CDR3 включал один или несколько из следующих остатков:
V, N, А или L в положении остатка по Kabat Н95, более предпочтительно V;
S, F, Н, Р, Т или W в положении остатка по Kabat Н99, более предпочтительно S;
А, Т, Р, S, V или Н положении остатка по Kabat H100B, более предпочтительно А или Т.
Предпочтительно, чтобы остаток по Kabat Н34 в VH CDR1 представлял собой I. Предпочтительно, чтобы VH CDR2 включал Е в положении по Kabat Н54 и/или I в положении по Н57.
Если связывающий агент включает VH-домен антитела, остаток по Kabat Н17 в VH-домене каркасной области предпочтительно является S. Остаток по Kabat Н94 предпочтительно представляет собой I или его консервативную замену (например, L, V, А или М). Обычно Н94 является I.
Предпочтительно, чтобы VL CDR3 включал один или несколько из следующих остатков:
S, Т или М в положении остатка по Kabat L90, более предпочтительно S или Т;
D, Е, Q, S, М или Т в положении остатка по Kabat L92, более предпочтительно D или Е;
А, Р, S, Т, I, L, М или V в положении остатка по Kabat L96, более предпочтительно S, Р, 1 или V, особенно S.
Остаток в положении L95A по Kabat в VL CDR3 предпочтительно представляет собой S.
Предпочтительно, чтобы VL CDR1 включал один или несколько из следующих остатков:
S в положении остатка по Kabat 27A;
N в положении остатка по Kabat 27B;
I в положении остатка по Kabat 27C;
D в положении остатка по Kabat 32.
Предпочтительно, чтобы VL CDR2 включал один или несколько из следующих остатков:
N в положении остатка по Kabat 51;
N в положении остатка по Kabat;
К в положении остатка по Kabat.
В предпочтительном варианте реализации связывающий агент согласно настоящему изобретению включает один или несколько CDR, выбранных из CDR доменов VH и VL, например, VH CDR1, 2 и/или 3 и/или VL CDR 1, 2 и/или 3 любого из антител 1, 2 или с 4 по 20, которые показаны в перечне последовательностей, или из исходного (родительского) антитела 3. В предпочтительном варианте реализации связывающий агент согласно настоящему изобретению включает VH CDR3 любой из следующих молекул антитела: Антитела 1 (SEQ ID NO 5); Антитела 2 (SEQ ID NO 15); Антитела 3 (SEQ ID NO 25); Антитела 4 (SEQ ID NO 35); Антитела 5 (SEQ ID NO 45); Антитела 6 (SEQ ID NO 55); Антитела 7 (SEQ ID NO 65); Антитела 8 (SEQ ID NO 75); Антитела 9 (SEQ ID NO 85); Антитела 10 (SEQ ID NO 95); Антитела 11 (SEQ ID NO 105); Антитела 12 (SEQ ID NO 115); Антитела 13 (SEQ ID NO 125); Антитела 14 (SEQ ID NO 135); Антитела 15 (SEQ ID NO 145); Антитела 16 (SEQ ID NO 155); Антитела 17 (SEQ ID NO 165); Антитела 18 (SEQ ID NO 175); Антитела 19 (SEQ ID NO 185); Антитела 20 (SEQ ID NO 195). Предпочтительно, чтобы связывающий агент дополнительно включал VH CDR1 последовательности SEQ ID NO: 3 или SEQ ID NO: 173 и/или VH CDR2 последовательности SEQ ID NO: 4. Предпочтительно, чтобы связывающий агент, включающий VH CDR3 последовательности SEQ ID NO: 175, включал VH CDR1 последовательности SEQ ID NO: 173, а также, возможно, включал VH CDR1 последовательности SEQ ID NO: 3.
Предпочтительно, чтобы связывающий агент включал набор VH CDR одного из следующих антител: Антитела 1 (Seq ID 3-5); Антитела 2 (SEQ ID 13-15); Антитела 3 (SEQ ID 23-25); Антитела 4 (SEQ ID 33-35); Антитела 5 (SEQ ID 43-45); Антитела 6 (SEQ ID 53-55); Антитела 7 (SEQ ID 63-65); Антитела 8 (SEQ ID 73-75); Антитела 9 (SEQ ID 83-85); Антитела 10 (SEQ ID 93-95); Антитела 11 (SEQ ID 103-105); Антитела 12 (SEQ ID 113-115); Антитела 13 (SEQ ID 123-125); Антитела 14 (SEQ ID 133-135); Антитела 15 (SEQ ID 143-145); Антитела 16 (SEQ ID 153-155); Антитела 17 (SEQ ID 163-165); Антитела 18 (SEQ ID 173-175); Антитела 19 (SEQ ID 183-185); Антитела 20 (SEQ ID 193-195). Дополнительно связывающий агент может включать набор VL CDR одного из следующих антител, причем VL CDR может быть как из того же антитела, что и VH CDR, так и из другого антитела:
Обычно VH домен соединен с VL доменом с получением антиген-связывающего сайта антитела, хотя в некоторых вариантах реализации для связывания антигена можно применять отдельно VH-домен или VL-домен. Разнообразие легкой цепи хорошо известно в данной области и, таким образом, согласно данной заявке, VH и VL домен не обязательно должны быть были из одного клона.
Связывающий агент может включать набор HCDR и/или LCDR любого из 1-20 антител, с одной или несколькими изменениями (десятью или менее заменами), например, с одним, двумя, тремя, четырьмя или пятью заменами в пределах набора HCDR и/или LCDR согласно настоящему изобретению. Предпочтительными являются замены в положениях по Kabat, отличных от 27А, 27B, 27С, 32, 51, 52, 53, 90, 92 и 96 в VL-домене и 34, 54, 57, 95, 97, 99 и 100B в VH-домене.
В случае осуществления замены в этих положениях, предпочтительными являются замены на остатки, указанные в данном описании как предпочтительные для данного положения.
В предпочтительном варианте реализации связывающий агент согласно настоящему изобретению представляет собой выделенную молекулу антитела человека с VH-доменом, включающим набор HCDR в каркасной области зародышевой линии человека, например, из семейства тяжелых цепей VH1 или VH3. В предпочтительном варианте реализации выделенная молекула антитела человека содержит VH-домен, включающий набор HCDR в VH1 DP5 или VH3 DP47 каркасной области зародышевых клеток человека. Таким образом, каркасные области VH-доменов могут включать каркасные области сегмента гена зародышевой линии человека VH1 DP5 или VH3 DP47. Последовательность аминокислот VH FR1 может представлять собой последовательность SEQ ID NO: 251. Последовательность аминокислот VH FR2 может быть последовательность SEQ ID NO: 252, Последовательность аминокислот VH FR3 может представлять собой последовательность SEQ ID NO: 253. Последовательность аминокислот VH FR4 может представлять собой последовательность SEQ ID NO: 254.
Обычно связывающий агент также включает VL домен, включающий набор LCDR, желательно, в каркасной области зародышевой линии человека, например, из семейства легких цепей Vlambda 1 или Vlambda 6.
В предпочтительном варианте реализации выделенная молекула антитела человека включает VL-домен, включающий набор LCDR в каркасной области VLambda 1 DPL8 или VLambda 1 DPL3 или VLambda 6_6a зародышевой линии человека. Таким образом, каркас VL-домена может включать каркасные области сегмента гена зародышевой линии человека VLambda 1 DPL8, VLambda 1 DPL3 или VLambda 6_6a. FR4 VL-домен может включать каркасную область сегмента гена зародышевой линии человека JL2. Последовательность аминокислот VL FR1 может представлять собой SEQ ID NO: 255. Последовательность аминокислот VL FR2 может представлять собой SEQ ID NO: 256. Последовательность аминокислот VL FR3 может представлять собой SEQ ID NO: 257. Последовательность аминокислот VL FR4 может представлять собой SEQ ID NO: 258.
Неэмбрионизированное антитело имеет такие же CDR, что и эмбрионизированное антитело, но другой каркас.
Связывающий агент согласно настоящему изобретению может конкурировать за связывание GM-CSFRα с любым описанным в настоящей заявке связывающим агентом, например, с антителом 3 или с любым из антител 1,2 или 4-20. Таким образом, связывающий агент согласно настоящему изобретению может конкурировать за связывание GM-CSFRα с молекулой антитела, включающей VH-домен любого из антител 1,2 или 4-20. Конкурирование между связывающими агентами легко можно исследовать in vitro, например, путем присоединения к одному из связывающих агентов репортерной молекулы, которую можно детектировать в присутствии одного или нескольких других немеченых связывающих агентов, чтобы обеспечить идентификацию связывающих агентов, которые связывают один эпитоп или перекрывающиеся эпитопы.
Конкурирование можно определять, например, методом твердофазного иммуноферментного анализа (ELISA), в котором, например, внеклеточный домен GM-CSFRα или пептид внеклеточного домена, иммобилизуют на чашке, в которую затем добавляют первый меченый связывающий агент вместе с несколькими другими немечеными связывающими агентами. Присутствие немеченого связывающего агента, который конкурирует с меченым связывающим агентом, определяют по уменьшению сигнала от меченого связывающего агента. Для определения конкурирования между связывающими агентами можно также использовать метод поверхностного плазмонного резонанса.
При исследовании конкурирования можно применять пептидный фрагмент антигена, в частности, пептид, включающий или состоящий по существу из эпитопа или представляющей интерес связывающей области. Можно использовать пептид, имеющий эпитоп или целевую последовательность с одной или несколькими аминокислотами на каждом конце. Связывающие агенты согласно настоящему изобретению могут быть такими, что их связывание с антигеном ингибируется пептидом, имеющим данную последовательность, или включающим ее.
Связывающие агенты, которые связывают пептид могут быть выделены, например, из библиотеки фагового дисплея, путем пэннинга (panning) с пептидом (пептидами).
Настоящее изобретение также обеспечивает применение описанного выше связывающего агента для измерения уровней антигена в тесте на конкурирование, то есть способа измерения уровня антигена в образце с применением связывающего агента, предложенного в настоящем изобретении, в тесте на конкурирование. Это может иметь место в том случае, если нет необходимости в физическом отделении связанного антигена от несвязанного. Одним из подходов к измерению является присоединение репортерной молекулы к связывающему агенту, обеспечивающее изменение физического или оптического параметра при связывании. Репортерная молекула может прямо или косвенно производить обнаруживаемые и, желательно, измеряемые сигналы. Присоединение репортерных молекул может быть прямым или непрямым, например, ковалентным, например, посредством пептидной связи, или нековалентным. Присоединение посредством пептидной связи быть результатом рекомбинантной экспрессии гибридных генов, кодирующих антитело и репортерную молекулу.
Настоящее изобретение также обеспечивает измерение уровней антигена непосредственно путем применения связывающего агента, например, в биосенсорной системе.
Настоящее изобретение обеспечивает способ, включающий вызывания или обеспечения возможности связывания обеспеченного настоящим изобретением связывающего агента с GM-CSFRα. Такое связывание может происходить in vivo, например, после введения связывающего агента или нуклеиновой кислоты, кодирующей связывающий агент, или может происходить in vitro, например, в твердофазном иммуноферментном анализе (ELISA), методе Вестерн-блот (Western blotting), иммуноцитохимии, иммунопреципитации, аффинной хроматографии или в тесте с использованием клеток, например, в тесте с клетками TF-1.
Можно провести количественную оценку связывания связывающего агента с GM-CSFRα. Количественная оценка может относиться к количеству антигена в тестируемом образце, которое может представлять интерес для диагностики или прогнозирования.
Набор, включающий связывающий агент или молекулу антитела в соответствии с аспектом или вариантом реализации согласно настоящему изобретению также является аспектом настоящего изобретения. В наборе согласно настоящему изобретению, связывающий агент или молекула антитела могут быть снабжены меткой, что обеспечивает возможность для определения их активности в исследуемом образце. Компоненты набора обычно стерильны и находятся в герметичных склянках или других контейнерах. Наборы можно применять в диагностических тестах или в других способах, в которых полезны молекулы антител. Набор может включать инструкции по использованию компонентов в способе, например, в способе, согласно настоящему изобретению. В набор согласно настоящему изобретению могут быть включены вспомогательные материалы для облегчения реализации или обеспечения реализации указанного способа.
Химическую активность антител в образце можно определять любыми подходящими способами, например, методом радиоиммуноанализа (РИА). Антиген, меченный радиоактивными метками, смешивают с немеченым антигеном (с тестируемым образцом) и проводят связывание с антителом. Связанный антиген физически отделяют от несвязанных антигенов и радиоактивных антигенов, связанных с определяемым антителом. Чем больше антигена в тестируемом образце, тем меньше радиоактивного антигена будет связываться с антителом. Также можно применять анализ конкурентного связывания с нерадиоактивным антигеном, с использованием антигена или аналога, связанного с репортерной молекулой. Репортерная молекула может представлять собой флуорохром, люминофор или лазерный краситель различным спектральными характеристиками поглощения и испускания. Подходящими флуорохромами являются флуоресцин, родамин, фикоэритрин и Texas Red. Подходящим хромогенным красителем является диаминобензидин. Другими репортерами являются макромолекулярные коллоидные частицы или материал в форме честиц, такой как окрашенные частицы из латекса, магнитные или парамагнитные и биологически или химически активные агенты, которые могут прямо или косвенно обеспечивать обнаруживаемые сигналы, которые можно наблюдать визуально, детектировать при помощи электронных приборов или регистрировать иным способом. Такими молекулами могут быть ферменты, которые, например, катализируют реакции, обеспечивающие появление или изменение окраски или вызывающие изменения в электрических свойствах. Возбуждение таких молекул может приводить к электронному переходу между энергетическими состояниями и в результате давать поглощение или испускание с характеристическим спектром. Молекулы могут включать химические структурные элементы, применяемые в сочетании с биосенсорами. Можно применять такие системы обнаружения как биотин/авидин или биотин/стрептавидин и щелочная фосфатаза.
Сигналы, генерируемые отдельными конъюгатами антитело-репортер, можно использовать для получения количественно измеряемых абсолютных или относительных величин, соответствующих связыванию антитела в образцах (в норме и в тестируемом образце).
Кроме того, настоящей изобретение обеспечивает выделенные нуклеиновые кислоты, которые включают последовательность, кодирующую связывающий агент, VH-домен и/или VL-домен согласно настоящему изобретению. Нуклеиновая кислота может включать ДНК и/или PHK, и может быть полностью или частично синтетической. Термин «последовательность нуклеотидов» в данном описании включает молекулу ДНК с указанной последовательностью и молекулу PHK с указанной последовательностью, в которой тимин (Т) замещен на урацил (U), если контекст не требует иного. В предпочтительном варианте, настоящее изобретение обеспечивает нуклеиновую кислоту, которая кодирует CDR или набор CDR, или VH-домен или VL домен, или антиген-связывающий сайт антитела, или молекулу антитела, например, scFv или lgG1 или lgG4 согласно изобретению, описанному в настоящей заявке. Настоящее изобретение также обеспечивает конструкции в виде плазмид, векторов, кассет транскрипции и экспрессии, которые включают, по меньшей мере, один полинуклеотид, описанный выше.
Другой аспект настоящего изобретения представляет собой клетку-хозяина, трансформированную или содержащую нуклеиновую кислоту согласно настоящему изобретению. Такая клетка-хозяин может существовать как in vitro, так и в культуре клеток. Также клетка-хозяин может быть представлена in vivo. Существование клетки-хозяина in vivo, может обеспечивать внутриклеточную экспрессию связывающих агентов согласно настоящему изобретению, так называемых «интрател» (intrabodies) или внутриклеточных антител, которые можно применять в генной терапии.
В еще одном аспекте предложен способ, включающий введение такой нуклеиновой кислоты в клетку-хозяина. Для введения можно использовать любой доступный способ. Для эукариотических клеток, подходящими способами являются кальций-фосфатная трансфекция, способ с применением DEAE-Dextran, электропорация, липосомная трансфекция и трансдукция с применением ретровирусов или других вирусов, например, вируса коровьей оспы или бакуловирусов для клеток насекомых. Для введения нуклеиновой кислоты в клетку-хозяина, в частности в эукариотическую клетку, можно применять систему, основанную на вирусах или плазмидах. Плазмидную систему можно поддерживать с помощью эписом или можно вводить в клетку-хозяина или в искусственную хромосому. Включение может быть или случайной или целевой интеграцией одной или нескольких копий в единичный или множественный локус. Для бактериальных клеток подходящими способами являются хлоридно- кальциевая трансформация, электропорация и трансфекция с помощью бактериофага.
За введением нуклеиновой кислоты может следовать вызывание или обеспечения возможности экспрессии с данной нуклеиновой кислоты, например, путем культивирования клеток-хозяев в условиях, подходящих для экспрессии данного гена.
В одном варианте реализации нуклеиновую кислоту согласно настоящему изобретению встраивают в геном (например, в хромосому) клетки-хозяина. Интеграцию можно стимулировать путем включения последовательностей, которые стимулируют рекомбинацию с геномом, в соответствии со стандартными способами.
Настоящее изобретение также обеспечивает способ, который включает применение указанной конструкции в системе экспрессии, что обеспечивает экспрессию связывающего агента или указанного полипептида. Таким образом, способы получения связывающего агента, VH-домена и/или VL-домена являются дополнительными аспектами настоящего изобретения. Способ может включать экспрессию указанной нуклеиновой кислоты в условиях для получения указанного связывающего агента, VH-домена и/или VL-домена, и его выделение. Такой способ может включать культивирование клеток-хозяев в условиях для получения указанного связывающего агента или домена антитела.
Способ получения может включать этап выделения и/или очистки продукта. Способ получения может также включать приготовление лекарственной формы из продукта и по меньшей мере одного дополнительного компонента, например, фармацевтически приемлемого наполнителя.
Системы клонирования и экспрессии полипептида в различных клетках-хозяевах хорошо известны. Подходящими клетками-хозяевами являются бактерии, клетки млекопитающих, клетки растений, дрожжи и системы на основе бакуловирусрв, трансгенные растения и животные. Экспрессия антител и фрагментов антител в прокариотических клетках хорошо известна в данной области [3]. Стандартным, предпочтительным бактериальным хозяином является Е. coli.
Специалистам в данной области в качестве варианта получения связывающего агента доступна также экспрессия в культуре эукариотических клеток [4, 5, 6]. К доступным линиям клеток млекопитающих для экспрессии гетерологичных полипептидов относятся клетки яичника китайского хомячка, клетки HeLa, клетки почек детенышей хомяков, клетки миеломы мыши NSO, клетки миеломы крысы YB2/0, клетки почек эмбрионов человека, клетки сетчатки глаза эмбрионов человека и многие другие.
Можно выбрать или сконструировать подходящие векторы, содержащие подходящие регуляторные последовательности, включая промоторные последовательности, терминаторные последовательности, последовательности полиаденилирования, энхансерные последовательности, маркерные гены и другие походящие последовательности. Векторами могут быть плазмиды, например, фагмиды или вирусные плазмиды, например, фагλ [7]. Многие известные способы и протоколы для манипуляций с нуклеиновыми кислотами, например, для получения конструкций нуклеиновых кислот, мутагенеза, секвенирования, введения ДНК в клетки и экспрессии генов, анализа белков подробно описаны у Ausubel с соавт. [8].
Настоящее изобретение обеспечивает способ получения одного или нескольких связывающих агентов, способных связывать антиген, причем указанный способ включает библиотеки связывающих агентов в контакт с указанным антигеном и выделение одного или нескольких связывающих агентов из библиотеки, способных связывать указанный антиген.
Библиотека может быть представлена на частицах или комплексах молекул, например, в виде способных к репликации генетических блоков (replicable genetic packages), таких как дрожжи, бактерии или бактериофаги (например, Т7) или в виде ковалентных, рибосомных или в виде других систем представления in vitro, в которых каждая частица или комплекс молекул содержат нуклеиновую кислоту, кодирующую представляемый на ней вариабельный VH-домен антитела, а также (если он присутствует) представляемый VL-домен. После выделения связывающих агентов, способных связывать антиген и представленных на бактериофаге или частицах другой библиотеки или комплексах молекул, можно получить нуклеиновую кислоту из бактериофага или другой частицы или комплекса молекул, представляющих указанный выделенный связывающий агент. Такую нуклеиновую кислоту можно затем применять в получении связывающего агента или вариабельного домена VH или VL антитела путем экспрессии с нуклеиновой кислоты, имеющей последовательность, взятой из бактериофага или другой частицы или комплекса молекул, представляющих указанный выбранный связывающий агент.
Вариабельный VH-домен антитела, имеющий последовательность аминокислот вариабельного VH-домена антитела вышеуказанного выбранного связывающего агента может быть представлен в изолированной форме, как и связывающий агент, включающий такой VH-домен.
Вариабельный VL-домен антитела, имеющий последовательность аминокислот вариабельного VL-домена антитела указанного выбранного связывающего агента может быть представлен в изолированной форме, как и связывающий агент, включающий такой VL-домен.
Кроме того, можно исследовать способность к связыванию GM-CSFRα, а также способность конкурировать с любым из антител от 1 до 20 (напр. в формате scFv и/или IgG формате, напр. lgG1 или lgG4) за связывание с GM-CSFRα. Также можно исследовать способность нейтрализовать GM-CSFRα.
Варианты VH- и VL-доменов и CDR.согласно настоящему изобретению, включая те, для которых для которых в настоящем описании представлены последовательности аминокислот, могут быть получены путем изменения последовательностей или мутаций и скрининга, и их можно включать в связывающие агенты для GM-CSFRα. Следуя идеям вычислительной химии по применению методов многопараметрического анализа к зависимостям структура/свойство-активность [9], можно получить количественные зависимости активность-свойство для антител с применением хорошо известных математических способов, таких как статистическая регрессия, распознавание и классификация образов [10, 11, 12, 13, 14, 15]. Свойства антител можно определить с помощью теоретических и эмпирических моделей (анализа вероятно контактирующих остатков или расчетных физико-химических свойств) последовательности антитела, функциональной и третичной структур, которые можно рассматривать как отдельно, так и в сочетании.
Антигенсвязывающий сайт антитела, состоящий из VH- и VL-доменов, формируется шестью петлями полипептида: тремя из вариабельного домена (VL) легкой цепи и тремя из вариабельного домена (VH) тяжелой цепи. Анализ с антител с известной атомарной структурой объясняет зависимость между последовательностью и третичной структурой связывающего сайта [16, 17]. Эти зависимости подразумевают, что кроме третьего участка (петли) в VH-доменах, петли связывающего сайта имеют одну из небольшого числа конформаций основной цепи: канонические структуры. Было показано, что каноническая структура, образующаяся в конкретной петле, определяется ее размером и по присутствием определенных остатков в ключевых местах в петле и в каркасных участках [16, 17].
Это исследование зависимости последовательность-структура можно использовать для предсказания тех остатков в антителе, последовательность которого известна, а третичная структура - нет, которые необходимы для поддержания третичной структуры петель его CDR и, следовательно, для поддержания связывания. Такие предсказания могут быть подкреплены их сравнением с результатами экспериментов, проводимых с целью оптимизации базовой структуры. В структурном подходе можно создать модель молекулы антитела [18] путем применения любого антитела, находящегося в свободном доступе или такого доступного на коммерческой основе антитела, как WAM [19]. Затем можно использовать визуализацию белка и пакет программного обеспечения для анализа, такой как Insight II (Accelerys, Inc.) или Deep View [20] для оценки возможных замен в каждом положении CDR. Эту информацию можно затем использовать для получения замен, которые, вероятно, оказывают минимальное или полезное влияние на активность.
Способы, необходимые для осуществления замен внутри последовательностей аминокислот CDR, VH- или VL-доменов антител и связывающих агентов, широкодоступны в данной области. Можно получить различные вариантные последовательности с заменами, относительно которых можно или нельзя предсказать, что они будут оказывать минимальное или полезное влияние на активность, и исследовать их на способность связывать и/или нейтрализовать GM-CSFRα и/или на любое другое желаемое свойство.
Варианты последовательности аминокислот любого из вариабельных доменов VH и VL, последовательности которых раскрыты в настоящей заявке, можно применять в соответствии с настоящим изобретением, как обсуждалось. Конкретные варианты могут включать одно или несколько изменений в последовательности аминокислот (добавление, делецию, замену и/или вставку остатков аминокислот), например, менее примерно 20 изменений, менее примерно 15 изменений, менее примерно 10 изменений или менее 5 изменений, возможно, 5, 4, 3, 2 или 1. Изменения могут быть сделаны в одном или нескольких каркасных участках и/или в одном или нескольких CDR.
Предпочтительно, изменения не приводили к потере функции, то есть желательно, чтобы связывающий агент, включающий измененную таким образом последовательность аминокислот, сохранил способность связывать и/или нейтрализовать GM-CSFRα.
Более предпочтительно, чтобы связывающий агент сохранил ту же величину связывания и/или нейтрализующую способность, что и у связывающего агента, в котором не делали изменений, например, измеренную в описанном здесь тесте. Более желательно, чтобы связывающий агент, включающий измененную таким образом последовательность аминокислот, имел улучшенную способность к связыванию или нейтрализции GM-CSFRα, чем связывающий агент, в котором не делали изменений, например, по результатам измерения данной способности в описанном здесь тесте.
Изменение может включать замену одного или нескольких остатков аминокислот на аминокислоту, не встречающуюся в природе, или на нестандартную форму аминокислоты, модицикацию одного или нескольких остатков аминокислот в с получением неприродной или нестандартной формы или вставку в последовательность одной или нескольких не встречающихся в природе или нестандартных аминокислот. Предпочтительные количества и положения изменений в последовательностях согласно настоящему изобретению описаны в данной заявке. Аминокислоты, встречающиеся в природе, включают 20 "стандартных" L-аминокислот, обозначаемых А, V, L, I, M, P, F, W, S, Т, N, Q, Y, С, К, R, H, D, Е в соответствии с их стандартным однобуквенным кодом. Нестандартные аминокислоты включают любой другой остаток, который может быть включен в остов полипептида или является результатом модификации существующего остатка аминокислоты. Нестандартные аминокислоты могут быть встречающимися или не встречающимися в природе. Известно несколько нестандартных аминокислот, встречающихся в природе, например, 4-гидроксипролин, 5-гидроксилизин, 3-метилгистидин, N-ацетилсерин и т.д. [21]. Те остатки аминокислот, которые модифицированы (дериватизированы) в N-альфа положении могут быть расположены только в N-конце. Обычно в настоящем изобретении аминокислота представляет собой L-аминокислоту, но в некоторых вариантах реализации она может представлять собой D-Аминокислоту. Кроме того, изменения могут включать модификацию L-аминокислоты в D-аминокислоту или замену L-аминокислоты на D-аминокислоту. Также известны метилированные, ацетилированные и/или фосфорилированные формы аминокислот, аминокислоты согласно настоящему изобретению также могут быть подвергнуты таким модификациям.
Последовательности аминокислот в доменах антитела и связывающих агентов согласно настоящему изобретению могут включать искусственные или нестандартные аминокислоты, описанные выше. В некоторых способах реализации нестандартные аминокислоты (например, D-аминокислоты) могут быть введены в последовательность аминокислот в процессе синтеза. В то время как в других способах реализации нестандартные аминокислоты могут быть введены посредством модификации или замены "исходных" стандартных аминокислот после синтеза последовательности аминокислот.
Применение нестандартных аминокислот и/или не встречающихся в природе аминокислот увеличивает структурное и функциональное разнообразие, и может, таким образом, увеличивать возможность достижения желаемой способности нейтрализации и связывания GM-CSFRα у связывающего агента согласно настоящему изобретению. Кроме того, было показано, что профили фармакокинетики после назначения животным у D-аминокислот и их аналогов лучше, чем у стандартных L-аминокислот, в связи с деградацией полипептидов, содержащих, L-аминокислоты in vivo.
Как указано выше, предпочтительно, чтобы представленная здесь последовательность аминокислот CDR входила в вариабельный домен антитела человека или значительной части антитела человека. Последовательности HCDR3, по существу совпадающие с описанными в данном документе, представляют собой предпочтительные варианты реализации настоящего изобретения, и также желательно, чтобы каждая из них входила в качестве HCDR3 в вариабельный домен тяжелой цепи или значительной его значительной части.
Вариабельные домены согласно настоящему изобретению, могут быть получены или произведены из любой зародышевой линии или перестроенного вариабельного домена человека, или могут быть синтетическими, основанными на консенсусных или фактических последовательностях известных вариабельных доменов человека. Последовательность CDR согласно настоящему изобретению (например, CDR3) можно ввести в репертуар вариабельных доменов без CDR (например, без CDR3), с использованием технологии рекомбинантных ДНК.
Например, Marks с соавторами (1992) [22] описывают способы получения репертуаров вариабельных доменов антител, в которых консенсусные праймеры, направленные к 5’-концу области вариабельного домена или расположенные рядом с ним, используют вместе с консенсусными праймерами в третьем каркасном участке генов VH человека, и получают репертуар вариабельных доменов VH без CDR3. Кроме того, Marks с соавт. описали, как эти репертуары можно комбинировать с CDR3 определенного антитела. Используя аналогичные методы, последовательности, полученные из CDR3 согласно настоящему изобретению, можно «перетасовывать» с репертуарами доменов VH или VL без CDR3, а полученные полные домены VH или VL, комбинировать с родственным (когнатным) доменом VL или VH с получением связывающих агентов согласно настоящему изобретению. Затем эти репертуары можно «представить» в подходящей системе-хозяине, такой как система фагового дисплея, описанная WO 92/01047 или любом другом из многочисленных литературных, включая [23], что позволяет осуществлять отбор подходящих связывающих агентов. Репертуар может состоять из более 104 независимых агентов, например, от 106 до 108 или 1010 агентов. Другими подходящими системами-хозяивами являются дрожжевой дисплей, бактериальный дисплей, дисплей на основе Т7, вирусный дисплей, клеточный дисплей, рибосомный дисплей и ковалентный дисплей. Аналогичными перетасовочными или комбинаторными методами являются также методики, раскрытые Stemmer (1994) [24], который описывает методики применительно к гену β-лактамазы, но также отмечает, что данный подход можно применять для наработки антител.
Другой альтернативой является получение новых участков VH или VL, несущих последовательности, являющиеся производными CDR согласно настоящему изобретению, с использованием неспецифического мутагенеза одного или нескольких выбранных генов VH и/или VL, с получением мутаций внутри целого вариабельного домена. Такой способ описан у Gram с соавт. (1992) [25], которые использовали ПЦР с повышенным уровнем ошибок (error-proned). В предпочтительных способах реализации делают одну или две аминокислотные замены внутри набора HCDR или LCDR. Другой способ, который можно использовать - это направленный мутагенез участков CDR генов VH или VL [26, 27].
Другой вариант реализации настоящего изобретения представляет способ получения антигенсвязывающего сайта антитела к антигену GM-CSFRα, причем указанный способ включает получение путем добавления делеции, замены или вставки одной или более аминокислот в приведенной в настоящей заявке последовательности аминокислот VH-домена, VH-домена, который является вариантом последовательности аминокислот указанного NV-домена, возможно, комбинирование полученного VH-домена с одним или несколькими VL-доменами, и исследование данных VH-домена или комбинации VH/VL для идентификации связывающего агента или антигенсвязывающего сайта антитела к антигену GM-CSFRα, которые, возможно, обладают одним или несколькими предпочтительными свойствами, предпочтительно, способностью нейтрализовать активность GM-CSFRα. Указанный VL-домен может иметь последовательность аминокислот, которая по существу совпадает с указанной в настоящей заявке последовательностью.
Можно применять аналогичный способ, в котором один или несколько вариантов последовательностей раскрытого в настоящей заявке VL-домена, комбинируют с одним или несколькими VH-доменами.
Другой вариант настоящего изобретения обеспечивает способ получения связывающего агента к антигену GM-CSFRα, который включает:
(a) представление исходного репертуара нуклеиновых кислот, кодирующих VH-домен, который или включает CDR3, который следует заменить, или в котором отсутствует область, кодирующая CDR3;
(b) комбинирование указанного репертуара с донорной нуклеиновой кислотой, кодирующей последовательность аминокислот, по существу совпадающую с приведенной в настоящей заявке для VH CDR3, при этом указанный донорную нуклеиновую кислоту встраивают в CDR3 область в указанном репертуаре, с получением конечного репертуара нуклеиновых кислот, кодирующих VH-домен;
(c) экспрессию нуклеиновых кислот указанного конечного репертуара;
(d) отбор агента, связывающего GM-CSFRα; и
(e) выделение указанного связывающего агента или кодирующей его нуклеиновой кислоты.
Более того, можно применять аналогичные способы, в которых VL CDR3 согласно настоящему изобретению объединен с репертуаром таких нуклеиновых кислот, кодирующих VL домен, которые или включают заменяемый CDR3 или не включают кодирующую CDR3 область.
Аналогично, один или более, или все три CDR могут быть «привить» в репертуар VH- или VL-доменов, которые затем подвергают скринингу на связывающий агент или связывающие агенты для GM-CSFRα.
В предпочтительном варианте реализации могут быть задействованы один или более HCDR1, HCDR2 и HCDR3, например, набор HCDR антитела 1(SEQ ID NOS: 3-5); Антитела 2 (SEQ ID NOS: 13-15); Антитела 4 (SEQ ID NOS: 33-35); Антитела 5 (SEQ ID NOS: 43-45); Антитела 6 (SEQ ID NOS: 53-55); Антитела 7 (SEQ ID NOS: 63-65); Антитела 8 (SEQ ID NOS: 73-75); Антитела 9 (SEQ ID NOS: 83-85); Антитела 10 (SEQ ID NOS: 93-95); Антитела 11 (SEQ ID NOS: 103-105,); Антитела 12 (SEQ ID NOS: 113-115); Антитела 13 (SEQ ID NOS: 123-125); Антитела 14 (SEQ ID NOS: 133-135); Антитела 15 (SEQ ID NOS: 143-145); Антитела 16 (SEQ ID NOS: 153-155); Антитела 17 (SEQ ID NOS: 163-165); Антитела 18 (SEQ ID NOS: 173-175); Антитела 19 (SEQ ID NOS: 183-185)или Антитела 20 (SEQ ID NOS: 193-195) или, возможно, Антитела 3 (SEQ ID NOS: 23-25), и/или могут быть задействованы один или более LCDR1, LCDR2 и LCDR3, например, набор LCDR Антитела 1 (SEQ ID NOS: 8-10); Антитела 2 (SEQ ID NOS: 18-20); Антитела 4 (SEQ ID NOS: 38-40); Антитела 5 (SEQ ID NOS: 48-50); Антитела 6 (SEQ ID NOS: 58-60); Антитела 7 (SEQ ID NOS: 68-70); Антитела 8 (SEQ ID NOS: 78-80); Антитела 9 (SEQ ID NOS: 88-90); Антитела 10 (SEQ ID NOS: 98-100); Антитела 11 (SEQ ID NOS: 108-110); Антитела 12 (SEQ ID NOS: 118-120); Антитела 13 (SEQ ID NOS: 128-130); Антитела 14 (SEQ ID NOS: 138-140); Антитела 15 (SEQ ID NOS: 148-150); Антитела 16 (SEQ ID NOS: 158-160); Антитела 17 (SEQ ID NOS: 168-170); Антитела 18 (SEQ ID NOS: 178-180); Антитела 19 (SEQ ID NOS: 188-190) или Антитела 20 (SEQ ID NOS: 198-200) или, возможно, Антитела 3 (SEQ ID NOS: 28-30).
Значительная часть вариабельного домена иммуноглобулина будет включать, по меньшей мере, три участка CDR вместе с каркасными областями, распологающимися между ними. Желательно, чтобы эта часть также включала по меньшей мере примерно 50% любой из двух или обеих (первой и четвертой) каркасных областей, причем именно 50% первой каркасной области представляют собой С-конец, а 50% четвертой каркасной области с N-конец. Дополнительные остатки в N-терминальном или С-терминальном конце значительной части вариабельного домена могут представлять собой остатки, которые, как правило, не ассоциированы с областями вариабельного домена природного происхождения. Например, конструирование связывающих агентов согласно настоящему изобретению при помощи технологий рекомбинантных ДНК, привести к введению N- или С-концевых остатков кодируемых линкерами, введенными для облегчения клонирования или других этапов манипуляций. Другие этапы манипуляций включают введение линкеров для присоединения вариабельных доменов согласно настоящему изобретению к дополнительным белковым последовательностям, включая константные области антитела, другие вариабельные домены (например, при получении димеров) или детектируемые/функциональные метки, описанные более подробно в других разделах данного текста.
Хотя в предпочтительном варианте настоящего изобретения связывающие агенты содержат пару VH- и VL-доменов, одиночные связывающие домены на основе последовательностей либо VH-, либо VL-доменов составляют дополнительные варианты настоящего изобретения. Известно, что одиночные домены иммуноглобулина, в частности, VH-домены, способны связывать целевые антигены (см., например, обсуждение dAbs в данном тексте).
В случае любого из одиночных связывающих доменов, эти домены можно применять для скрининга комплементарных доменов, способных к образованию двудоменного связывающего агента, способного связывать GM-CSFRα. Этого можно достичь путем скрининга методами фагового дисплея, с применением так называемого двойного иерархического комбинаторного подхода, описанного в WO 92/01047, в котором отдельную колонию, содержащую клон либо с Н-, либо с L-цепью применяют для инфицирования полной библиотеки клонов, кодирующих другую цепь (L или Н), и, полученный в результате двухцепочечный связывающий агент, подвергают отбору в соответствии с методиками фагового дисплея, как описано указанном источнике и в [22].
Другие варианты реализации настоящего изобретения представляют составы, содержащие связывающие агенты и, по меньшей мере, один дополнительный компонент, напр. состав, включающий связывающий агент и фармацевтически приемлемый наполнитель. Такие составы можно применять в способах ингибирования или нейтрализации GM-CSFRα, включая способы терапевтического лечения тела человека или животного.
Изобретение обеспечивает гетерогенные препараты, включающие молекулы антитела к GM-CSFRα. Такие препараты могут представлять собой, например, смеси антител с полноразмерными тяжелыми цепями и тяжелыми цепями без С-концевого лизина, различной степени гликозилирования, и/или с производными аминокислот, например, с уицлизацией N-концевой глутаминовой кислоты с образованием остатка пироглутаминовой кислоты.
Вариантам настоящего изобретения включают способы лечения, включающие введение связывающего агента согласно настоящему изобретению, фармацевтических композиций, включающих указанный связывающий агент, а также применение указанного связывающего агента в изготовлении медикамента, например, в способе получения медикамента или фармацевтической композиции, включающего приготовление связывающего агента с фармацевтически приемлемым наполнителем.
Анти-GM-CSFRα средства можно вводить орально (например, нанотелам), путем инъекций (например, подкожных, внутривенных, внутриартериальных, внутрисуставных, внутрибрюшинных, внутримышечных), путем ингаляций, внутрипузырным путем (инстилляцией в мочевой пузырь) или местным путем (например, внутриглазным, интраназальным, ректальным путем, через пораженные участки, через кожу). Лечение осуществлять путем пульсовой инфузии, в частности, с уменьшением доз связывающего агента. Путь введения может зависеть от физико-химических характеристикам средства, особенностями заболевания или необходимостью оптимизации эффективности или минимизации побочных эффектов. Предполагается, что анти-GM-CSFRα лечение не будет ограничиваться применением в клинике. Таким образом, также предпочтительны подкожные инъекции, для которых используют устройства без игл.
Композицию можно вводить отдельно или в сочетании с другими способами лечения, либо одновременно, либо последовательно, в зависимости от состояния, которое лечат. Для достижения синергетических эффектов можно применять комбинированное лечение, в частности, комбинацию агента, связывающего GM-CSFRα, с одним или более других лекарств. Связывающий агент в соответствии с настоящим изобретением можно вводить в сочетании или в дополенение к одному или более следующих агентов: НПВП (например, ингибиторы циклооксигеназы, такие как целекоксиб и другие подобные ЦОГ-2 ингибиторы), кортикостероиды (например, преднизон) и модифицирующие заболевание противоревматоидные средства (DMARD), например, Хумира (адалимумаб), метотрексат, Арава, Энбрел, (Этанерцепт), Ремикад (Инфликсимаб), Кинерет (Анакинра), Ритуксан (Ритуксимаб), Оренция (абатацепт), соли золота, антималярийные средства, сульфасалазин, d-пенициламид, циклоспорин А, диклофенак, циклофосфамид и азатиоприн.
В соответствии с настоящим изобретением, представленные композиции можно назначать отдельным лицам. Композиции предпочтительно вводить в "терапевтически эффективном количестве". Терапевтически эффективное количество - это количество, являющееся достаточным для демонстрации пользы для пациента. «Пользой» в данном случае может быть как минимум снижение выраженности как минимум одного симптома. Фактическое вводимое количество, частоту и периодичность введения будут зависеть от природы и тяжести заболевания, которое лечат. Предписание лечения, например, выбор дозы и т.д. находится в сфере ответственности врача общей практики и других врачей, и может зависеть от тяжести симптомов и/или прогрессирования заболевания, которое лечат. Подходящие дозы антитела хорошо известны [28, 29]. Кроме того, можно применять конкретные дозы, указанные в настоящей заявке или в Настольной книге врача (Physician’s Desk Reference) (2003) в зависимости от типа вводимого лекарственного средства. Терапевтически эффективное количество или подходящая доза связывающего агента согласно настоящему изобретению можно определить путем сравнения их активности in vitro и активности in vivo на модельном животном. Известны способы экстраполяции эффективных доз для мыши и других экспериментальных животных на дозы для человека. Точная доза будет зависеть от ряда факторов, включая: для чего будут применять антитело:
для диагностики или лечения, от размера и локализации области лечения, от определенного типа антитела (например, целого антитела, фрагмента или димера) и типа детектируемой метки или другой молекулы, присоединенной к антителу. Обычная доза антитела лежит в диапазоне от 100 мкг до 1 г для общего применения и от 1 мкг до 1 мг для местного применения. Обычно, антитело представляет собой целое антитело, желательно lgG1, lgG2 или более желательно lgG4. Указанная доза представляет собой дозу для однократного введения взрослому пациенту, ее можно пропорционально скорректировать для детей и младенцев, а также скорректировать для антител других форматов пропорционально их молекулярному весу. Лечение (введение) можно повторять ежедневно, дважды в неделю, с недельными или месячными интервалами по выбору лечащего врача. В предпочтительных способах реализации настоящего изобретения лечение является периодическим и период между введениями составляет примерно две недели или более, желательно примерно три недели или примерно один месяц. В других предпочтительных вариантах реализации настоящего изобретения лечение можно осуществлять до и/или после операции, кроме того, более предпочтительно, чтобы лечение было осуществляли или применяли непосредственно в анатомическом участке хирургического вмешательства.
Связывающий агент согласно настоящему изобретению обычно вводят в форме фармацевтической композиции, которая может включать, по меньшей мере, еще один дополнительный компонент. Таким образом, фармацевтические композиции в соответствии с настоящим изобретением и для применения в соответствии с настоящим изобретением могут включать в дополнение к активному ингредиенту фармацевтически приемлемый наполнитель, носитель, буфер, стабилизатор или другие материалы, хорошо известные специалистам в данной области. Такие материалы не должны быть токсичны и не должны снижать эффективность активного ингредиента. Конкретный тип носителя или другого материала будет зависеть от пути введения, который может быть оральным или путем инъекций, например, внутривенных. Фармацевтические композиции для орального введения могут быть в форме таблетки, капсулы, порошка, жидкости или в полутвердой форме. Таблетка может содержать твердый носитель, такой как желатин или адъювант. Жидкие фармацевтические составы в основном содержат жидкий носитель, такой как вода, вазелин, животные или растительные масла, минеральное или синтетическое масло. Можно включать физиологический раствор, раствор декстрозы или другого сахара или гликоли, такие как этиленгликоль, пропиленгликоль или полиэтилен гликоль. Для внутривенной инъекции или инъекции в поврежденный участок активный ингредиент должен иметь форму фармацевтически приемлемого водного раствора, который является апирогенным и имеет подходящие pH, изотоничность и стабильность. Специалисты, компетентные в данной области, могут приготовить подходящие растворы, используя, например, изотонические среды, такие как физиологический раствор для инъекций, раствор Рингера для инъекций, раствор Рингера с лактатом для инъекций. При необходимости можно включать консерванты, стабилизаторы, буферы, антиоксиданты и/или другие добавки. Связывающие агенты согласно настоящему изобретению могут быть приготовлены можно приготовить в жидкой, полутвердой или твердой форме, в зависимости от физико-химических свойств молекулы и пути доставки. Композиции могут включать наполнители или их сочетания, например: сахара, амнокислоты и поверхностно-активные вещества. Жидкие составы могут включать антитела в широком диапазоне концентраций и pH. Твердые составы могут можно получить, например, путем лиофилизациии, распылительной сушки или высушивания по технологии сверхкритического флюида. Составы (лекарственные формы) анти-GM-CSFRα будут зависеть от предполагаемого пути доставки: например, составы для доставки в легкие могут состоять из частиц, физические свойства которых обеспечивают проникновение глубоко в легкие при ингаляции; составы для местного применения могут включать модификаторы вязкости, которые увеличивают время нахождения лекарства в участке действия. В некоторых вариантах реализации связывающий агент можно приготовить с носителем, который будет предохранять агент от быстрого высвобождения, например, в виде состава с контролируемым высвобождением, включая имплантаты, трансдермальные пластыри и микрокапсулированные системы доставки. Можно применять рассасывающиеся, биологически совместимые полимеры, такие как этиленвинилацетат, полиангидириды, полигликолиевая кислота, коллаген, полиортоэфиры и полимолочная кислота. Множество способы приготовления таких составов хорошо известны специалистам в данной области. См. Напр. Робинсон, 1978 [30].
Связывающие агенты согласно настоящему изобретению можно применять в способе лечения или диагностики у человека или животного, например, в способе лечения (которое может включать профилактическое действие) заболевания или расстройства у пациента, который включает назначение пациенту эффективного количества связывающего агента. Состояния, которые можно лечить согласно настоящему изобретению, включают любые состояния, в которых принимает участие GM-CSFRα. В опубликованной технической литературе показана роль GM-CSF в нескольких заболеваниях и состояниях, описанных ниже. Поскольку GM-CSF специфично связывает GM-CSFRα, то патологическому и/или симптоматическому действию может противодействовать путем ингибирования связывания GM-CSF с GM-CSFRα. Таким образом, в дополнение к фармакологическим данным in vivo и in vitro, представленным для описанных в экспериментальной части молекул антитела, опубликованные данные показывают, что связывающие агенты согласно настоящему изобретению можно применять в лечении аутоиммунных и воспалительных состояний, заболеваний и расстройств, например, ревматоидного артрита, астмы, аллергической реакции, рассеянного склероза, миелоидной лейкемии или атеросклероза.
Данные, опубликованные для этих состояний, приведены ниже:
Астма и Аллергические реакции
Бронхиальная астма - это часто встречающееся устойчивое воспалительное заболевание легких, характеризующееся гиперреактивностью дыхательных путей, избыточной продукцией слизи, фиброзом и повышением уровня lgE. Гиперреактивность дыхательных путей (ГДП) представляет собой избыточное сокращение дыхательных путей в ответ на неспецифические стимулы. Считают, что и ГДП, и избыточная продукция слизи вносят свой вклад в снижение проходимости дыхательных путей, что приводит к одышке, характеристическим приступам астмы (обострениям), которые обуславливают смертность, связанную с данным заболеванием (примерно 2000 смертей/год в Великобритании).
Недавние исследования показали, что уровни GM-CSF и рецептора к нему, как в форме белка, так и в форме мРНК, при астме повышены. Более того, уровни экспрессии коррелируют с тяжестью заболевания. Путем измерения определили, что продукция GM-SCF была определена в бронхоальвеолярном лаваже (БАЛ), клетках БАЛ, мокроте, клетках эпителия бронхиол, и стимулированных антигеном мононуклеарах периферической крови, выделенных у пациентов с астмой, повышена относительно продукции у субъектов, не страдающих астмой [31, 32]. Более того, было показано, что уровень экспрессии GM-SCF в дыхательных путях после провокации аллергеном коррелирует со степенью эозинофилии ткани и тяжестью поздних астматических реакций [34]. Более поздние исследования выявили связь повышенной экспрессии GM-SCF с истинной или неатопической астмой, что согласуется с данными об экспрессии в легких [34]. В модели введения и иммунизации овальбумином на мышах, нейтрализации активности GM-SCF поликлональным антителом козы путем антраназального введения перед сенсибилизацией овальбумином предотвращала гиперреактивность дыхательных путей, а также снижала инфильтрацию эозинофилами и секрецию слизи в дыхательных путях [35]. Аналогично, в модели аллергического заболевания на мышах, в которой животным в качестве раздражителя вводили частицы отработанного дизельного топлива, нейтрализация GM-SCF также путем интраназального введения поликлональных антител козы предотвращала гиперреактивность дыхательных путей в ответ на метаколин, снижала количество эозинофилов в БАЛ, и также снижала содержание бокаловидных клеток, продуцирующих слузь в эпителии дыхательных путей [36].
Роль GM-CSF далее исследовали в модели индуцированной толерантности на мышах. У мышей, которым ежедневно вводили повторные дозы овальбумина (путем распыления) без предварительной сенсибилизации, развивалась толерантность к овальбумину, и у них не развивалось эозинофильное воспаление дыхательных путей. Экспрессия GM-CSF в легких, которую вызывали при помощи аденовирусного конструкта, изменяет ответ этих животных и способствует притоку эозинофилов в БАЛ, развитию гистологического фенотипа, характерного для аллергии, и ассоциированной гиперплазии бокаловидных клеток. О развитии типичного ответа Th2 дополнительно свидетельствует увеличение концентрации IL-5 в сыворотке и БАЛ и IL-4 в БАЛ. Дальнейшая работа с этой моделью с применением мышей с нокаутом по ГКГС11 указывает на то, что GM-CSF модулирует взаимодействие между антигенпрезентирующими клетками и Т-клетками, способствуя, таким образом, осуществлению Т-клеточных ответов не овальбумин [37]. Существенно то, что активность GM-CSF в качестве эффективного активатора ответов ТН2 также можно продемонстрировать на мышах, у которых отсутствует IL-13 и/или IL-4, что указывает на то, что нейтрализация активности GM-CSF обеспечивает альтернативный терапевтический подход, не связанный с активностью этих цитокинов.
Аналогичные наблюдения были сделаны в другой модели на мышах, в которой повторяющееся антраназальное введение амброзии приводило к сенсибилизации типа Th2 и умеренному воспалению в ответ на повторное воздействие антигена [38]. Введение анти Gm-CSF антител в комбинации с амброзией снижало продукцию ассоциированных с Th2 цитокинов, предположительно, путем ингибирования эндогенного CG-CSF. Доставка амброзии в микросреду дыхательных путей, обогащенную GM-CSF любо путем многократного совместного введения рекомбинантного GM-CSF, либо путем однократного введения аденовирусного вектора, несущего трансген GM-CSF, наоборот, приводила к значительному усилению эозинофльного воспаления дыхательных путей и специфичного вторичного ответа Th2 на амброзию.
Ревматоидный артрит (РА)
РА представляет собой хроническое воспалительное и разрушающее сустав заболевание, которое поражает приблизительно 1% населения в индустриализированном мире. PA характеризуется гиперплазией и воспалением синовиальной мембраны, воспалением в синовиальной жидкости, и прогрессирующим разрушением окружающей кости и хряща, что обычно приводит к значительной инвалидизации.
Хотя причина РА остается неизвестной, накапливается все больше доказательств роли GM-CSF в прогрессировании РА. Считают, что РА инициируется и направляется опосредованным Т-клетками, антиген-специфичным процессом. Вкратце, считают, что присутствие неидентифицированного антигена в организме восприимчивом хозяина вызывает ответ Т-клеток, который приводит к продукции цитокинов Т-клеток с последующим привлечением воспалительных клеток, включая нейтрофилы, макрофаги и В-клетки.
В суставе, пораженном ревамтоидным артритом продуцируются многие про- и противовоспалительные цитокины. Более того, прогрессирование, возобновление и стихание заболевания опосредованы динамическими изменениями продукции цитокинов в суставе. В частности, считают, что TNF-α и IL-1 играют решающую роль в патогенезе РА, и многие новые разработанные или разрабатываемые методы лечения указанного заболевания рассчитаны на ингибирование активности этих двух провоспалительных цитокинов.
Недавние исследования с применением модели на грызунах позволили сделать предположение о центральной и важной роли GM-CSF в развитии и прогрессировании РА. Введение экзогенного рекомбинантного GM-CSF усиливает патологию в двух различных моделях РА на мышах: коллаген-индуцированный артрит (КИА) [6] и модель односуставного артрита [7]. Вдобавок к этому, было показано, что мыши с нокаутом по GM-CSF (GM-CSF-/-) устойчивы к развитию КИА и что уровни IL-1 и фактора некроза опухоли (TNFα), обнаруживаемые в синовиальной суставной жидкости, были снижены по сравнению с таковыми у мышей дикого типа [8, 9]. Аналогичным образом, индукция моноартрита с помощью внутрисуставной инъекции метилированного бычьего сывороточного альбумина и IL-1 в мышей GM-CSF-/- приводит к снижению тяжести заболевания, по сравнению с мышами дикого типа [10].
Более того, введение анти-GM-CSF MAT мыши значительно снижает выраженность заболевания в моделях КИА и односуставного артрита. В модели КИА, лечение MAT было эффективно для лечения прогрессирования стабильного заболевания, гистопатологии и значительного снижения уровней IL-1 и TNF-α в суставе. Кроме того, лечение MAT перед началом артрита снижало заболевания КИА [11, 10].
В ряде исследований анализировали уровни цитокинов и рецепторов, присутствующих в синовиальной жидкости и биопсийных образцах мембран из тканей человека при артрите. Мононуклеары периферической крови 27 пациентов с РА, включая 13 здоровых волонтеров и 14 пациентов с остеопорозом, оценивали на уровни GM-CSFR с применением GM-CSF, меченного РЕ [12]. В данном исследовании было показано, что у пациентов с РА было обнаружено в два раза больше рецептор-положительных клеток (53%), чем у здоровых контрольных пациентов (20%) и пациентов, проходящих обследование на остеопороз (25%), что позволяет предположить, что моноциты, возможно, опосредуют ответ на локально продуцируемый GM-CSF. Исследование экспрессии генов цитокинов у пациентов с РА [13] методом in situ гибридизации клеток синовиальной жидкости показало повышенные уровни GM-CSF, IL-1, TNF-a и IL-6. Более того, выделенные и культивированные синовиоциты, полученные из фибробластов нормальных волонтеров продемонстрировали повышение уровня белка GM-CSF в ответ на IL-1α IL-1β, TNF-α и TNF-β [14]. Количественный анализ уровней GM-CSF в сыворотке пациентов с РА [15] показал, что уровни белка были выше у пациентов с тяжелым РА (366 пг/мл, n=26) и умеренным РА (376 пг/мл, n=58) по сравнению с контрольной группой (174 пг/мл, n=43), более того, также было показано, что уровень GM-CSF был значительно повышен в синовиальной жидкости пациентов с РА (1300 пг/мл).
Ранее наблюдали, что введение рекомбинантного GM-CSF пациентам, которых лечили от нейтропении, может вызывать обострение РА [16]. Сходные наблюдения были сделаны для пациентов, страдающих синдромом Фелти, после лечения рекомбинантным GM-CSF [17].
Хроническая обструктивная болезнь легких (ХОБЛ)
Хроническую обструктивную болезнь легких (ХОБЛ) определяют как болезненное состояние, описываемое ограничением дыхания, которое не является полностью необратимым. Хроническое ограничение дыхания обычно является прогрессирующим и связано с аномальным воспалительным ответом легких на токсичные частицы или газы. Это ограничение дыхания вызвано одновременным заболеванием малых дыхательных путей (обструктивный бронхиолит) и разрушением паренхимы (эмфизема), причем их относительный вклад различается у разных людей. Конечные характерные симптомы ХОБЛ представляют собой кашель, образование мокроты и диспноэ при физической нагрузке.
ХОБЛ является важной проблемой здравоохранения и четвертой по частоте причине хронической заболеваемости и смертности в США. Указанное заболевание на данный момент лечат лекарственными препаратами, изначально предназначенными для лечения астмы, такими как пероральные или вдыхаемые кортикостероиды, бронходилататоров, включая β-агонисты, или без них. Тем не менее, ни для одного из этих лекарственных препаратов не была показана способность замедлять прогрессирование ХОБЛ [18]. Например, кортикостероиды, которые заметно подавляли эозинофильное воспаление при астме, не проявили какого-либо действия на воспаление, наблюдаемое при ХОБЛ, которое опосредовано преимущественно нейтрофилами [19]. Таким образом, существует необходимость в разработке новых способов лечения ХОБЛ, которые специфично нацелены на воспалительные процессы, лежащие в основе патофизиологии данного заболевания.
GM-CSF, с учетом его роли в функционировании нейтрофилов и макрофагов, может играть важную роль в патогенезе ХОБЛ.
В одном исследовании с применением количественной ПЦР было показано, что в мокроте подходящего по возрасту больного с ХОБЛ, число копий GM-CSF было значительно выше, чем внеобструктивной мокротой курильщика без обструктивных симптомов [20]. Более того, в модели воспаления легких, индуцированного сигаретным дымом, на грызунах, животные, которых лечили путем интраназального введения антитела к GM-CSF за 2 дня, 4 часа и 1 час до воздействия дымом, проявили значительное снижение уровней нейтрофилов, макрофагов и ММР-9 в бронхоальвеолярном лаваже (БЖ) по сравнению с изотипическим контролем антител через 5 дней после воздействия [21]. Эти исследования также поддерживают наши собственные наблюдения при исследовании уровней GM-CSF в вызванной мокроте пациентов с различными степенями тяжести ХОБЛ. В этих исследованиях мы показали, что уровень GM-CSF был повышен в мокроте приблизительно 40% принявших участие в исследовании пациентов с ХОБЛ, безотносительно тяжести заболевания, при этом уровень GM-CSF в некоторых случаях приближался к 500 пг/мл. Оказалось, что уровни GM-CSF не повышены у некурящих и подходящих курящих контрольных пациентов. Эти данные позволяют предположить, что GM-CSF может быть одним из ключевых медиаторов индуцированного дымом воспаления дыхательных путей и ХОБЛ.
Рассеянный склероз (PC)
Было показано, что GM-CSF вовлечен в аутоиммунное заболевание - рассеянный склероз. Путем введения антигена - гликопротеина олигодендроцитов миелина (MOG) - грызунам можно получить модель рассеянного склероза человека, которая демонстрирует множество фенотипов PC, таких как воспаление центральной нервной системы и демиелинизация, которые могут привести к PC-подобному параличу. У мышей, лишенных GM-CSF, MOG был неспособен вызвать фенотип экспериментального аллергического энцефаломиелита [55]. Более того, было показано, что эти мыши давали пониженную пролиферацию Т-клеток в ответ на антиген MOG и пониженную продукцию Th1 цитокинов IL-6 и IFN-γ. Введение нейтрализующих GM-CSF антител одновременно с провокацией антигеном предотвращало начало заболевания в течение 10 дней после лечения с признаками снижения повреждений. Если введение осуществляли после начала заболевания, мыши полностью восстанавливались в течение 20 дней лечения.
Лейкемия
Также было показано, что GM-CSF участвует в миелоидной лейкемии, юношеской хронической миелоидной лейкемии (JCML). Это патологическое состояние представляет собой миелопролиферативное расстройство, которое поражает в первую очередь пациентов младше 4 лет. In vitro клетки-предшественники гранулоцитов-макрофагов периферической крови JCML (CFU-GM) проявили спонтанную пролиферацию при низкой плотности клеток, что ранее не было описано для других миелопролиферативных расстройств. Более того, обеднение этих культур моноцитами приводило к прекращению пролиферации. Впоследствии было показано, что эта спонтанная пролиферация опосредована гиперчувствительностью клеток-предшественников JCML к цитокину, продуцируемому моноцитами, GM-CSF [22, 23, 24, 25, 26, 27]. По-видимому, не чрезмерная продукция или повышенные уровни GM-CSF у пациентов с JCML, a скорее, гиперчувствительность клеток-предшественников JCML вызвана нарушением регуляции GM-CSF индуцированного пути передачи сигнала Ras [28]. Недавние исследования с аналогом GM-CSF (E21R), который антагонизирует действие GM-CSF, как в тестах на связывание, так и в функциональных тестах, показали, что путем ингибирования действия GM-CSF можно значительно уменьшить груз клеток JCML в ксенотрансплантатной модели JCML на мышах с тяжелым комбинированным иммунодефицитом/с генетической предрасположенностью к развитию аутоиммунного диабета 1 типа (SClD/NOD) [29]. Профилактическое системное введение дозы E21R во время трансплантации предотвращало приживление JCML клеток-предшественников в костном мозге, а введение дозы E21R через 4 недели после трансплантации индуцировало ремиссию JCML, со снижением груза клеток. Более того, введение E21R мышам SClD/NOD, которым одновременно вводили нормальные клетки костного мозга человека и клетки костного мозга пациентов с JCML, приводило к уменьшению груза JCML, причем клетки нормального костного мозга не были затронуты.
Атеросклероз
Ишемическая болезнь сердца представляет собой самую распространенную причину смерти во всем мире. За последние годы укрепилось общее представление о том, что воспаление играет значительную роль в патогенезе атеросклероза и что накопление воспалительных клеток происходит вместе с накоплением липидов на стенках артерии.
Когда воспалительные клетки, такие как моноциты и макрофаги, заселяют стенку артерии, они участвуют и поддерживают локальный воспалительный ответ. Эти макрофаги также экспрессируют рецепторы-ловушки для множества липопротеинов и, тем самым, способствуют дифференцировке клеток в «пенистые клетки». Именно смерть этих «пенистых клеток» способствует образованию липидного ядра, классического признака этих поражениц. Поскольку воспаление продолжается внутри атеросклеротических бляшек, такие активированные воспалительные клетки высвобождают фиброгенные медиаторы и факторы роста, которые вызывают пролиферацию гладкомышечных клеток (SMC) и фиброз бляшки. В доплнение к стимуляции фиброза, эти клетки также высвобождают протеолитические ферменты, такие как металлопротеиназы матрикса (ММР), которые способствуют снижению плотности фиброзных бляшек, тем самым делая их подверженным разрывам. Когда эти бляшки разрываются, они высвобождают обломки клеток и факторы свертывания крови, такие как тканевый фактор, в сосуд, стимулируя каскад коагуляции и образование тромбов. Развившийся в результате артериальный тромбоз затем может привести к ишемии миокарда или инфаркту.
Недавно было сделано предположение, что GM-CSF участвует во многих аспектах прогрессирования заболевания при атеросклерозе. Обнаружили, что в атеросклеротических очагах у кроликов, которых кормили холестерином, GM-CSF со-локализован с макрофагами и, в меньшей степени, с эндотелиальными клетками и гладкомышечными клетками [30]. Кроме того, было показано, что экспрессия GM-CSF повышена в в пораженных атеросклерозом сосудах человека в местах скопления макрофагов, в среднем слое клеток гладкой мускулатуры и в клетках эндотелия [31]. Это повышение уровней GM-CSF, отчасти, приписывают непосредственному контакту между моноцитами/макрофагами и эндотелиальными клетками в процессе образования и патогенеза атеросклеротического очага [32]. Другим ключевым элементом при атеротическом поражении являются «пенистые клетки», которые представляют собой макрофаги, которые поглотили окисленные липопротеины низкой плотности (LDL) с участием рецепторов-ловушек на своей поверхности. In vitro это поглощение Ox-LDL может затем стимулировать макрофаги к пролиферации по механизму, зависящему от GM-CSF [33].
Поскольку атеросклероз представляет собой хронический воспалительный процесс, исследовали противовоспалительные агенты, такие как глюкокортикоиды. Дексаметазон, противовоспалительный глюкокортикоид, подавляет развитие атеросклероза в ряде экспериментальных моделей на животных [34, 35, 36, 37]. Его эффективность связывают с ингибированием миграции [38] и пролиферации [39] клеток гладкой мускулатуры, и снижением хемотаксиса моноцитов и лейкоцитов циркулирующей крови [40]. Недавние исследования показали, что ox-LDL может индуцировать высвобождение GM-CSF из перитонеальных макрофагов мыши [41]. Более того, после обработки дексаметазоном, имело место дозозависимое ингибирование высвобождения GM-CSF, что позволяет предположить, что противовоспалительное действие дексаметазона опосредовано ингибированием индуцируемой ox-LDL продукции GM-CSF. Поскольку GM-CSF, по-видимому, играет центральную роль в атеросклерозе, альтернативой глюкокортикоидам может быть ингибирование активности GM-CSF при таком показании.
ТЕРМИНОЛОГИЯ
"И/или" в данной заявке следует понимать как конкретное раскрытие каждого из двух указанных свойств или компонентов вместе с другим или отдельно. Например, "А и/или В" следует понимать, как конкретное раскрытие каждого из: (i) A, (ii) В, и (iii) А и В, как если бы каждый из перечисленных пунктов был указан в данной заявке отдельно.
GM-CSFRα и GM-CSF
GM-CSFRα представляет собой альфа-цепь рецептора колониестимулирующего фактора гранулоцитов-макрофагов. Полноразмерная последовательность GM-CSFRα человека хранится под номером доступа S06945 (gi: 106355) [42] и описана в настоящей заявке как SEQ ID NO: 202. Зрелая форма GM-CSFRα человека, т.е. с отщепленным сигнальным пептидом, описана в настоящей заявке как SEQ ID NO: 206. В данной заявке, если контекст не указывает на иное, GM-CSFRα относится к GM-CSFRα человека или не являющегося человеком примата (например, яванского макака), обычно, человека. GM-CSFRα может быть встречающимся в природе GM-CSFRα или рекомбинантным GM-CSFRα.
Внеклеточный домен α-цепи GM-CSF рецептора человека, состоящий из 298 аминокислот, имеет последовательность аминокислот, обозначенную как SEQ ID NO: 205.
Если контекст не предполагает иное, в данной заявке на GM-CSF относится к GM-CSF человека или не являющегося человеком примата (например, яванского макака), обычно, человека.
GM-CSF обычно связывается с внеклеточным доменом (SEQ ID NO: 205) альфа-цепи зрелого рецептора GM-CSF (SEQ ID NO: 206). Как описано в другом месте данной заявки, это связывание ингибируют связывающие агенты согласно настоящему изобретению.
Встречающиеся в природе сплайсированные варианты GM-CSFRα были идентифицированы: см, например, ссылки [43 и 44]. В известных вплайсированных вариантах указанный внеклеточный домен высоко консервативен. Связывающие агенты согласно настоящему изобретению могут связываться или могут не связываться с одним или более сплайсированных вариантов GM-CSFRα, и могут ингибировать или не ингибировать связывание GM-CSF с одним или более сплайсированных вариантов сплайсинга GM-CSFRα.
Связывающий агент
Этот термин описывает одну из пары молекул, которые связываются друг с другом. Молекулы из связывающейся пары могут быть природными или целиком или частично полученными путем синтеза. Одна из пары молекул имеет на поверхности область, или полость, которая связывается и, таким образом, комплементарна определенной пространственной и полярной структуре другой молекулы из этой пары. Примеры типов связывающихся пар включают пары антиген-антитело, биотин-авидин, гормон-рецептор гормона, рецептор-лиганд, фермент-субстрат. Настоящее изобретение связано с типом реакций антиген-антитело.
Связывающий агент обычно включает молекулу, содержащую антигенсвязывающий сайт (участок). Например, связывающий агент может представлять собой молекулу антитела или белок, не являющийся антителом, который содержит антигенсвязывающий сайт. Антигенсвязывающий сайт можно получить путем расположения гипервариабельных участков в каркасе белка, не являющегося антителом, таких как фибронектин или цитохром В, и т.д. [46, 47, 48], или путем рандомизации или мутирования остатков аминокислот какой-либо петли в каркасе белка, в результати которых белок приобретает способность связываться с желаемой мишенью. Каркасы для конструирования новых сайтов связывания в белках были подробно описаны [48]. Белковые каркасы для миметиков антител раскрыты в WO/0034784, в котором изобретатели описывают белки (миметики антител), которые содержат домен фибронектина III типа, имеющий по меньшей мере одну рандомизированную петлю. Подходящий каркас, к которому можно привить один или более гипервариабельных участков, например, набор гипервариабельных участков человека, можно обеспечить с помощью любого домена-члена суперсемейства генов иммуноглобулинов. Каркасом может быть белок человека или белок, не являющийся белком человека.
Преимущество не являющегося антителом белкового каркаса состоит в том, что он позволяет получить антигенсвязывающий сайт в каркасной молекуле, которая меньше по размеру и/или которую легче получить, чем по меньшей мере некоторые из молекул антител. Малый размер связывающего агента может обеспечить полезные физиологические свойства, такие как способность проникать в клетки, проникать глубоко в ткани или достигать мишеней внутри других структур, или связываться внутри полостей белка целевого антигена.
Применение антигенсвязывающих сайтов в не являющихся антителами белковых каркасах описано в источнике [45]. Типичным примером являются белки, имеющие стабильный остов и одну или более вариабельных петель, в которых последовательность аминокислот петли или петель подвергают направленному или случайному мутагенезу с получением антигенсвязывающего сайта, который связывается с целевым антигеном. Такие белки включают lgG-связывающие домены белка А из S. aureus, трансферрин, тетранектин, фибронектин (например, 10ый домен фибронектина III типа) и липокалины. Другие подходы включают синтетические "Микротела" (Selecore GmbH), на основе на циклотидов - малых белков, имеющих внутримолекулярные дисульфидные связи.
В дополнение к последовательностям антител и/или антигенсвязывающим сайтам, связывающий агент согласно настоящему изобретению может включать другие аминокислоты, например, образующие пептид или полипептид, такой как уложенный домен, или аминокислоты, придающие указанной молекуле другое функциональное свойство в дополнение к способности связывать антиген. Связывающие агенты согласно настоящему изобретению могут нести детектируемую метку, или могут быть конъюгированы с токсином или нацеливающей группой, или ферментом (например, посредством пептидной связи или линкера). Например, связывающий агент может включать каталитический сайт (например, в домене фермента), а также антигенсвязывающий сайт, причем указанный антигенсвязывающий сайт связывается с антигеном и, таким образом, нацеливает каталитический сайт на антиген. Каталитический сайт может ингибировать биологическую функцию антигена, например, путем расщепления.
Хотя, как было упомянуто, гипервариабельные участки могут нести на себе каркасы, такие как фибронектин или цитохром В [46, 47, 48], структурой, несущей гипервариабельный участок или набор гипервариабельных участков согласно настоящему изобретению, будет, как правило, последовательность тяжелой или легкой цепи антитела или существенная ее часть, в которой указанный гипервариабельный участок или набор гипервариабельных участков расположен в положении, соответствующем положению гипервариабельного участка или набора гипервариабельных участков во встречающихся в природе вариабельных доменов VH и VL антител, кодируемых перестроенными генами иммуноглобулинов. Структуры и положения вариабельных доменов иммуноглобулинов можно определить со ссылкой на (Kabat, и др., 1987 [64], и обновления, доступные теперь в сети Интернет (http://immuno.bme.nwu.edu или поиск "Kabat" с использованием любой поисковой системы).
Связывающие агенты согласно настоящему изобретению могут включать константные области антител или их части, предпочтительно константные области антител человека или их части. Например, VL-домен может быть присоединен своим С-концом к константным доменам легкой цепи антитела, включая цепи Cκ или Cλ, человека, предпочтительно цепи Cλ. Аналогично, связывающий агент на основе VH-домена, может быть присоединен своим С-концом ко всей тяжелой цепи иммуноглобулина или к ее части (например, к СН1 домену), происходящей из антитела любого изотипа, например, IgG, IgA, IgE и IgM, и любого из подклассов изотипов, в частности, lgG1, lgG2 и lgG4. Предпочтительным являются lgG1, lgG2 или lgG4. lgG4 является предпочтительным, потому что он не связывает комплемент и не создает эффекторных функций. Любой вариант синтетической или другой константной области, который имеет такие свойства и стабилизирует вариабельные участки, также является предпочтительным для применения в реализации настоящего изобретения.
Связывающие агенты согласно настоящему изобретению можно пометить детектируемой или функциональной меткой. Детектируемые метки включают радиоактивные метки, такие как 131I или 99Tc, которые могут быть присоединены к антителам согласно настоящему изобретению с применением стандартных реагентов, известных в области визуализации антител. Метки включают также ферментативные метки, такие как пероксидаза хрена. Метки дополнительно включают молекулы, такие как биотин, которые можно детектировать путем связывания со специфичной родственной детектируемой молекулой, например, меченым авидином. Таким образом, связывающий агент или молекула антитела согласно настоящему изобретению может иметь форму конъюгата, включающего связывающий агент и метку, возможно, присоединенную при помощи линкера, такого как пептид. Указанный связывающий агент можно конъюгировать, например, с ферментами (например, с пероксидазой, щелочной фосфатазой) или с флюоресцентной меткой, включая, без ограничения, следующие: биотин, флюорохром, зеленый флюоресцентный белок. Дополнительно, метка может включать молекулу токсина, такую как молекула токсина, выбранная из группы, состоящей из токсина синегнойной палочки (РЕ или его цитотоксического фрагмента или мутантного варианта), дифтерийного токсина (его цитотоксического фрагмента или мутантного варианта), токсина ботулизма от А до F, рицина или его цитотоксического фрагмента, абрина или его цитотоксического фрагмента, сапорина или его цитотоксического фрагмента, противовирусного токсина лаконоса или его цитотоксического фрагмента и бриодина 1 или его цитотоксического фрагмента. Если указанный связывающий агент включает молекулу антитела, меченый связывающий агент может быть назван иммуноконъюгатом.
Молекула антитела
Этот термин описывает иммуноглобулин, либо природный, либо частично или целиком полученный синтетическим путем. Указанный термин также охватывает любой полипептид или белок, включающий антигенсвязывающий сайт антитела. Фрагменты антител, которые включают антигенсвязывающий сайт антитела, представляют собой молекулы, такие как Fab, F(ab’)2, Fab’, Fab’-SH, scFv, Fv, dAb, Fd; и диатела (diabodies).
Можно взять моноклональные и другие антитела и использовать методики технологии рекомбинантной ДНК для получения других антител или химерных молекул, которые сохраняют специфичность исходного антитела. Такие методики могут включать введение ДНК, кодирующей вариабельный участок иммуноглобулина или гипервариабельные участки антитела, в константные области, или константные области плюс каркасные участки, другого иммуноглобулина. См., например, ЕР-А-184187, GB 2188638A или ЕР-А-239400, и большой объем следующей ниже литературы. Гибридому или другую клетку, продуцирующую антитело, можно подвергнуть генетической мутации или другим изменениям, которые могут изменять или не изменять связывание полученных антител с мишенью.
Поскольку существует множество способов модификации антител, термин "молекула антитела" следует понимать как охватывающий любой связывающий агент или вещество, имеющее антигенсвязывающий сайт антитела. Таким образом, данный термин охватывает фрагменты и производные антител, включая любой полипептид, содержащий антигенсвязывающий сайт антитела, либо природный, либо полностью или частично синтетический. Соответственно, включены и химерные молекулы, содержащие антигенсвязывающий сайт антитела, или его эквивалент, соединенные (слитые, гибридизованные) с другим полипептидом. Клонирование и экспрессия химерных антител описаны в ЕР-А-0120694 и ЕР-А-0125023, и в большом объеме более поздней литературы.
Дополнительные методики, доступные в области конструирования антител, сделали возможным выделение антител человека и гуманизированных антител. Антитела человека и гуманизированные антитела представляют собой предпочтительные варианты реализации настоящего изобретения, и могут быть получены с помощью любого подходящего способа. Например, можно получить гибридомы человека [49]. Фаговый дисплей, другая устоявшаяся методика получения связывающих агентов, подробно описан во многих публикациях, таких как источник [49] и WO 92/01047 (обсуждается дополнительно ниже). Для выделения антител человека можно применять трансгенных мышей, у которых гены антител мыши инактивированы и функционально заменены на гены антител человека, а другие компоненты иммунной системы мыши оставлены нетронутыми [50]. Гуманизированные антитела можно получать, применяя методики, известные в данной области, например, описанные в WO 91/09967, US 5,585,089, ЕР 592106, US 565,332 и WO 93/17105. Дополнительно, в WO 2004/006955 описаны способы гуманизации антител, основанные на отборе последовательностей каркасных участков вариабельных областей из генов антител человека путем сравнения канонических типов структур гипервариабельных участков для последовательностей гипервариабельных участков вариабельной области антитела, не являющегося антителом человека, с каноническими типами структур гипервариабельных участков для соответствующих гипервариабельных участков из библиотеки последовательностей антител человека, например, сегментами генов антитела эмбрионального типа. Вариабельные области антитела человека, имеющие канонические типы структур гипервариабельных участков, сходные с гипервариабельными участками, не относящимися к человеку, образуют подгруппу последовательностей антител человека, из которых нужно выбрать каркасные последовательности человека. Кандидаты из данной подгруппы могут быть дополнительно ранжированы по схожести аминокислот в последовательностях гипервариабельных участков человека и ГВУ, не являющихся ГВУ человека. В способе, описанном в WO 2004/006955, последовательности человека с наивысшим рангом отбирают для получения каркасных последовательностей для конструирования химерных антител, которые функционально замещают последовательности гипервариабельного участка человека на эквивалентные гипервариабельные участки, не относящиеся к человеку, используя каркасные области человека отобранных кандидатов из подгрупп, в результате получают гуманизированное антитело с высокой аффинностью и низкой иммуногенностью без необходимости сравнения каркасных последовательностей между антителами, не являющимися антителм, и антителами человека. Химерный антитела, полученные согласно данному способу, также описаны.
Синтетические молекулы антител можно получить путем экспрессии генов, полученных с помощью олигонуклеотидов, синтезированных и собранных в подходящих векторах экспрессии [51, 52].
Было показано, что фрагменты целого антитела могут выполнять функцию связывания антигенов. Примерами связывающих фрагментов являются (i) Fab-фрагмент, состоящий из VL-, VH-, CL- и СН1-доменов; (ii) Fd-фрагмент, состоящий из VH- и СН-1 доменов; (iii) Fv-фрагмент, состоящий из VL- и VH-доменов одного антитела; (iv) dAb-фрагмент [53, 54, 55], который состоит из VH- или VL-домена; (v) изолированные гипервариабельные участки; (vi) F(ab’)2-фрагменты, бивалентные фрагменты, включающие два связанных Fab-фрагмента; (vii) одноцепочечные Fv-молекулы (scFv), в котором VH- домен и VL-домены связаны пептидным линкером, который обеспечивает ассоциацию двух указанных доменов с образованием антигенсвязывающего сайта [56, 57]; (viii) биспецифичные одноцепочечные димеры Fv (PCT/US 92/09965) и (ix) "диатела" (diabodies), мультивалентные или мультиспецифичные фрагменты, полученные путем гибридизации генов (WO 94/13804; [58]). Молекулы Fv, scFv или диатела можно стабилизировать путем введения дисульфидных мостиков, связывающих VH- и VL-домены [59]. Также можно получить мини-антитела (minibodies), включающие scFv, присоединенный к СН3-домену [60].
dAb (однодоменное антитело) представляет собой малый мономерный антигенсвязывающий, а именно вариабельный участок тяжелой или легкой цепи антитела [55]. dAb, содержащие VH. встречаются в природе у семейства верблюдовых (например, верблюда, ламы) и могут быть получены путем иммунизации представителя верблюдовых целевым антигеном, выделения антиген-специфичных В-клеток и прямого клонирования генов dAb из отдельных В-клеток. dAb также можно получить в культуре клеток. Их малый размер, хорошая растворимость и термостабильность делает их, в частности, физиологически применимыми и подходящими для селекции и аффинного созревания. Связывающий агент согласно настоящему изобретению может представлять собой dAb, включающее VH- или VL-домен, по существу такой, как описано в данной заявке, или VH- или VL-домен, включающий набор гипервариабельных участков, по существу, как описано в данной заявке. "По существу, как описано" означает, что соответствующий гипервариабельный участок или VH- или VL-домен согласно настоящему изобретению будет либо идентичным, либо в высокой степени сходным с определенными участками, последовательности которых описаны в данной заявке. "В высокой степени сходный" означает, что в гипервариабельном участке и/или VH- или VL-домене может быть сделано от 1 до 5, предпочтительно от 1 до 4, например, от 1 до 3 или 1, или 2, или 3 или 4, изменений аминокислот.
Если необходимо использовать биспецифичные антитела, они могут представлять собой обычные биспецифичные антитела, которые могут быть изготовлены множеством способов [61], например, получены хемически или из гибридных гибридом, или могут быть любыми из фрагментов биспецифичного антитела, упомянутых выше. Примеры биспецифичных антител включают антитела, которые получены с помощью технологии BiTE™, согласно которой можно использовать связывающие домены двух антител с различной специфичностью и напрямую соединять их с помощью коротких пептидов. Это позволяет соединить два антитела в одну короткую полипептидную цепь. Можно сконструировать Диатела и scFv без Fc-участка, используя только вариабельные домены, что может обеспечить снижение эффектов антиидиотипического взаимодействия.
Биспецифичные диатела, в противоположность целым биспецифичным антителам, также могут быть особенно полезны, так как их можно легко сконструировать и экспрессировать в E.coli. Диатела (и многие другие полипептиды, такие как фрагменты антител) с соответствующими специфичностями связывания можно легко отобрать из библиотек в помощью фагового дисплея (WO 94/13804). Если одно плечо диатела нужно сохранить неизменным, например, направленным против GM-CSFRα, можно создать библиотеку, в которой изменено другое плечо и выбрать антитело, связывающееся с соответствующей мишенью. Биспецифичные целые антитела можно получить с помощью методики конструирования «knobs-into-holes» (выпуклости-в-полости) [62].
Антигенсвязывающий сайт
Данный термин описывает часть молекулы, которая связывается и является комплементарной ко всему целевому антигену или его части. В молекуле антитела его называют антигенсвязывающим сайтом, и он включает часть антитела, которая связывается и является комплементарной всему антигену или его части. Если антиген большой, антитело может связываться только с определенной частью антигена, которую называют эпитопом. Антигенсвязывающий сайт антитела может быть образован одним или более вариабельными доменами антитела. В предпочтительном случае, участок антитела, связывающий антиген, включает вариабельную область легкой цепи антитела (VL) и вариабельную область тяжелой цепи антитела (VH).
Нумерация по Kabat
Остатки последовательностей антител в данной заявке обычно обозначены с использованием нумерации по Kabat, описанной у Kabat и др., 1971 [63]. Также см. ссылки [64, 65].
Изолированный (выделенный)
Этот термин относится к состоянию, в котором связывающие агенты согласно настоящему изобретению, или нуклеиновая кислота, кодирующая такие связывающие агенты, будут в целом находиться в соответствии с настоящим изобретением. Изолированные агенты и изолированная нуклеиновая кислота будут свободными или по существу свободными от материала, с которым они связаны в природе, такого как другие полипептиды или нуклеиновые кислоты, с которыми их обнаруживают в естественной среде, или среде, в которой их получают (например, культуре клеток), если такое получение осуществляют по технологии рекомбинантной ДНК, реализуемой in vitro или in vivo. Агенты и нуклеиновая кислота могут входить в состав лекарственной формы вместе с разбавителями или адъювантами, и при этом с практической точки зрения быть изолированными - например, агенты будут, как правило, смешаны с желатином или другими носителями, если их используют для покрытия микротитрационных планшетов для приминения в иммуноанализе, или будут смешаны с фармацевтически приемлемыми носителями или разбавителями, для применения в диагностике или терапии. Связывающие агенты могут быть гликозилированы, либо природным путем, либо с помощью систем гетерологичных эукариотических клеток (например, СНО или NSO (ЕСАСС 85110503) клеток), или они могут быть (например, если получены путем экспрессии в прокариотической клетке) негликозилированными.
КРАТКОЕ ОПИСАНИЕ РИСУНКОВ
Фигура 1. Исследование рА2 двух анти-GM-CSFRα антител в тесте на прорливерацию с клетками TF-1. Пролиферацию клеток TF-1 индуцировали увеличивающимися концентрациями GM-CSF в присутствии возрастающих концентраций двух оптимизированных lgG4, антитела 6 (Фигура 1А) и антитела 1 (Фигура 1B), соответственно. Для данных, показанных на графике 1А и графике 1B, измеряли включение меченного тритием тимидина и рассчитывали ЕС50 GM-CSF при каждой концентрации антитела. Для данных, показанных на графике 1C и графике 1D, затем рассчитывали отношения доз и анализировали с помощью модели регрессии Шилда, в результате чего получали значения рА2.
Фигура 2. Исследование рА2 анти-GM-CSFRα антитела, антитела 6, в тестах на изменение формы гранулоцитов. Гранулоциты человека (графики 2А и 2С) или яванского макака (2B и 2D) обрабатывали возрастающими концентрациями GM-CSF в присутствии возрастающих концентраций lgG4. Изменение формы гранулоцитов измеряли применяя проточную цитометрию и рассчитывали ЕС50 GM-CSF при каждой концентрации антитела (график 2А и график 2B). Затем рассчитывали отношения доз и анализировали с помощью модели регрессии Шилда, в результате чего получали значения рА2 (график 2С и график 2D).
Фигура 3. Эффективность антагонизма двух антител, антител 1 и 6, соответственно, в форме lgG4 в тесте, позволяющем измерить пролиферацию TF-1 клеток, индуцированную 7 пМ GM-CSF человека. Также показаны результаты для положительного контроля lgG4 2B7 и для изотипического контроля lgG4. Результаты представлены в форме среднего со стандартным отклоненим по трезкратному измерению в одном эксперименте.
Фигура 4. Эффективность антагонизма двух антител, антител 1 и 6, соответственно, в форме lgG4 в тесте, позволяющем измерить изменение формы гранулоцитов человека, индуцированное 7 пМ GM-CSF человека. Также показаны результаты для контроля lgG4 2B7 и для изотипического контроля lgG4. Результаты представлены в форме среднего со стандартным отклонением по трехкратному измерению в одном эксперименте.
Фигура 5. Эффективность антагонизма двух антител, антител 1 и 6, соответственно, в форме lgG4 в тесте, позволяющем измерить высвобождение TNFα из моноцитов человека, стимулированное 1 нМ GM-CSF человека. Также показаны результаты для контрольного антитела 2B7 и для изотипического контроля lgG4. Результаты представлены в форме среднего со стандартным отклонением по трехкратному измерению в одном эксперименте.
Фигура 6. Эффективность антагонизма двух антител, антител 1 и 6, соответственно, в форме lgG4 в тесте, выживаемость гранулоцитов человека, индуцированную 7 пМ GM-CSF человека. Также показаны результаты для контрольного антитела 2B7 и для изотипического контроля lgG4. Результаты представлены в форме среднего со стандартным отклонением по трехкратному измерению в одном эксперименте.
Фигура 7. MAT человека со зрелой аффинностью, антитело 1 и антитело 6, в отличе от родительского MAT человека 28G5 (антитело 3) и известного антитела мыши 2B7, ингибировали стимулированную GM-CSF дифференцировку клеток-предшественников гемопоэтического ряда человека. 5×104 размороженных мононуклеарных клеток из пробы для афереза культивировали в полутвердом агаре в присутствии 10 нг/мл GM-CSF и указанной концентрации МАТ. Колонии считали в день 14. График показывает число колоний в зависимости от концентрации MAT в мкг/мл.
Фигура 8. Анализ зависимости доза-ответ для эффективности MAT со зрелой аффинностью в химерных мышах huGM-CSFR Tg. Группам из 5 химерных мышей Tg вводили 500 нг huGM-CSF (или ФБР) подкожно дважды в день в течение 4 дней (Д.1-Д.4), и в Д.0 вводили либо контроль (САТ001), либо тестируемое MAT (Антитело 6) в указанных концентрациях. Вес селезенок оценивали в Д.5.
Фигура 9. Анализ зависимости доза-ответ для эффективности антитела 6 в анализе на высвобождение эндогенных цитокинов мононуклеарами периферической крови человека. 1×106 клеток культивировали в течение 72 часов в присутствии или в отсутствие антитела, и проводили твердофазный иммуноанализ надосадочной жидкости на IL-6 и TNFα. Результаты представлены в виде среднего ингибирования со стандартным отклонением для двухкратного измерения в одном эксперименте.
ЭКСПЕРИМЕНТАЛЬНАЯ ЧАСТЬ
Уровень техники
Фрагменты антител человека можно отобрать in vitro из репертуаров, представленных на поверхности нитевидного бактериофага. Этот способ известен как фаговый дисплей и обеспечивает средство получения фрагментов антител человека. Данный процесс можно использовать для выделения специфичных агентов человека к материалу человека, и его можно может быть модифицировать для получения антител с определенными характеристиками аффинности.
Фрагменты антител, состоящие только из вариабельных доменов тяжелой цепи (VH) и вариабельных доменов легкой цепи (VL), соединенные друг с другом коротким пептидным линкером, содержат всю информацию, определяющую связывание антигена. Такие фрагменты известны как одноцепочечные Fv (scFv). Было показано, что при представлении на поверхности фага scFv и правильно складываются, и связывают антиген. Таким образом были сконструированы обширные репертуары scFv человека, которые обеспечили источник, из которого можно выделять отдельные клоны в качестве кандидатов для разработки лекарственного препарата. Кандидатные scFv затем преобразуют в полные молекулы IgG (обычно, IgG человека) для терапевтических приложений.
Краткое описание
Селекцию (отбор) проводили на библиотеке scFv в формате фагового дисплея, полученной из лимфоцитов селезенки человека, чтобы обогатить по популяции фага, который связывается с GM-CSFRα человека. Мы изолировали (выделяли) антитела scFv, имеющие отобранные свойства и преобразовали эти scFv в lgG4. Применяя ряд тестов, панель антител изолировали, оптимизировали и эмбрионизировали с получением lgG4 с подходящими характеристиками для терапевтического антитела.
19 клонов антитела, последовательности которых показаны как антитела 1, 2 и 4-20 в списке последовательностей, были получены из родительского антитела. Родительское антитело показано как антитело 3 в списке последовательностей, в данной заявке оно также обозначено как 28G5. Указанные 19 клонов отобрали, как демонстрирующие исключительно хорошие свойства в ряде биологических тестов, как описано в экспериментльной части, и им были присвоены номера: антитело 1, 2 и с 4 по 20.
Указанные биотесты должны были отразить воспалительную природу заболеваний, таких как ревматоидный артрит. Например, изменение формы нейтрофилов, необходимое для привлечения их в место действия, высвобождения провоспалительных факторов моноцитами и повышенной выживаемости типов воспалительных клеток в ответ на определенные сигналы. Антитела проявляют высокую эффективность нейтрализации в этих тестах.
Подробные протоколы используемых тест-систем приведены ниже в разделе, названном "Материалы и методы тестирования".
Выделение ведущих антител
Для селекции использовали обширную библиотеку одноцепочечных антител FV (scFv) человека. Она была получена из лимфоцитов селезенки от 20 здоровых доноров и клонирована в фагмидный вектор. ScFv, которые распознавали GM-CSFRα, изолировали из библиотеки в формате фагового дисплея в серии повторных циклов селекции на очищенном GMCSF-Ra, полученном при чрезмерной экспрессии помеченного для очистки, растворимого внеклеточного домена указанного рецептора в клетках НЕК293Т. Этого достигают по существу как описано у Vaughan и др. [68]. Вкратце, после взаимодействия биотинилированного рецептора с фаговой библиотекой, белок со связанным фагом фиксировали на покрытых стрептавидином магнитных гранулах. Несвязавшиеся фаги отмывали. Связавшиеся фаги выделяли, как описано у Vaughan и др., и повторяли процесс селекции. Проводили три раунда селекции при понижении концентрации антигена. Представительную долю scFv из продуктов раундов селекции подвергали секвенированию ДНК.
После этих первоначальных отборов из библиотеки в формате фагового дисплея, идентифицировали панель уникальных scFv в тесте на связывание лиганда, предназначенном для идентификации фагов, экспрессирующих scFv антитела, которые способны ингибировать связывание GM-CSF с очищенным внеклеточным доменом GM-CSFRα. Нейтрализующая эффективноть этих scFv в тесте на связывание лиганда была в диапазоне от 0.65 до 3.3 нМ.
Антитела, которые проявляли активность в биохимическом тесте на связывание лиганда, оценивали на биологическую активность в тесте на пролиферацию клеток TF-1, который позволял измерить эффективность нейтрализации путем тестирования способности указанных антител ингибировать пролиферацию TF-1 клеток, стимулированную GM-CSF. TF-1 представляют собой премиелоидную линию клеток человека, полученную из клеток пациента с эритролейкемией. Эта линия клеток является зависимой от факторов, необходимых для выживаемости и пролиферации, и ее поддерживают по стандартной методике с GM-CSF человека. Ингибирование GM-CSF-зависимой пролиферации определяли путем измерения уменьшения включения меченного тритием тимидина во вновь синтезированную ДНК делящихся клеток. Все scFv имели измеримую эффективность в этом тесте, причем значения IC50 лежали в диапазоне от, приблизительною, 180 до 1200 нМ.
Наиболее эффективные клоны scFv преобразовывали в формат молекулы антител lgG4 человека с константным доменом тяжелой цепи гамма 4 человека и константным доменом легкой цепи лямбда человека. Для наиболее эффективных клонов scFv были сконструированы векторы, что позволило осуществлять экспрессию антител в виде целого антитела lgG4, как описано у Persic и др. [66], с несколькими модификациями. В указанные векторы был включен фрагмент OriP, чтобы облегчить применение в клетках HEK-EBNA 293 и обеспечить возможность эписомной репликации. Вариабельный домен VH клонировали в полилинкер между секреторной лидерной последовательностью и константным доменом гамма 4 человека в вектор экспрессии pEU8.1(+).Вариабельный домен VL клонировали в полилинкер между секреторной лидерной последовательностью и константным доменом лямбда человека в вектор экспрессии pEU4.1(-). Клетки HEK-EBNA 293 котрансфецировали конструкциями, экспрессирующими тяжелую и легкую цепи, и целое антитело очищали от кондиционированной среды, применяя аффинную хроматографию на белке А. Очищенные препараты антител фильтровали через стерильные фильтры и хранили при 4°C в фосфатно-солевом буферном растворе (ФБР) до проведения тестов. Концентрацию белка определяли путем измерения поглощения при 280 нм, используя метод бицинхониновой кислоты (ВСА) (Pierce).
Преобразованные IgG сравнивали с известным антителом мыши 2B7 в тесте на пролиферацию клеток TF-1. Указанные lgG4 сохраняли или приобретали активность в этом тесте, при этом значения IC50 лежали в диапазоне от 6 до приблизительно 1600 нМ.
При воспалительном заболевании, изменение формы нейтрофилов необходимо для их привлечения (рекрутинга) в место действия. Тест на изменение формы гранулоцитов человека был разработан, чтобы имитировать этот биологический ответ, с применением метода сортировки клеток с возбуждением флюоресценции (FACS) для измерения изменения формы гранулоцитов, выделенных из крови после их взаимодействия с GM-CSF. Оценивали способность анти-GM-CSFRα антител в формате lgG4 ингибировать ответ на GM-CSF, выражающийся в изменении формы нейтрофилов, полученные значения IC50 отобранных клонов лежали в диапазоне от приблизительно 15 до 350 нМ. Типичное антитело 28G5 нейтрализовало GM-CSF-R яванского макака в тесте на изменение формы гранулоцитов яванского макака с IC50, приблизительно равной 5 нМ. Известное антитело мыши 2B7 также было способно нейтрализовать биологический ответ, возникающий в результате связывания GM-CSF с рецептором яванского макака.
Аффинность указанных антител к рецептору затем измеряли, применяя BIAcore, при этом расчетные значения KD лежали в диапазоне от 32 до 377 нМ.
Оптимизация
В попытке улучшить эффективность 28G5 провели программу оптимизации. Получали библиотеки антител, в которых проводили неспецифический мутагенез гипервариабельных участков CDR3 VH или VL. Каждый CDR3 был рандомизирован в двух блоках из 6 аминокислот, чтобы покрыть весь гипервариабельный участок, что давало библиотеки Н1 (N-концевой блок из 6 аминокислот VH CDR3), Н2 (С-концевой блок из 6 аминокислот в VH CDR3), L1 (N-концевой блок из 6 аминокислот в VL CDR3) и L2 (С-концевой блок из 6 аминокислот в VL CDR3). Полученные библиотеки подвергали повторным циклам селекции на связывание с GM-CSFRa человека. Клоны, выделенные в этом селекционном процессе, затем применяли для конструирования комбинированной фаговой библиотеки, которая содержала scFv как с мутированными CDR3 тяжелой цепи, так и с мутированными CDR3 легкой цепи. Эти библиотеки также подвергали той же процедуре селекции.
На каждой стадии процесса оптимизации, scFv, которые были способны ингибировать связывание 28G5 lgG4 с GM-CSF рецептором, идентифицировали методом конкурентного анализа связывания эпитопа с 28G5 и указанным рецептором, и затем оценивали в тесте на пролиферацию клеток TF-1, как описано ниже.
После случайного мутагенеза последовательностей CDR3 тяжелой цепи 28G5, была идентифицирована панель scFv с эффективностью нейтрализации, которую можно было измерить в тесте с TF-1. Большинство улучшений эффективности было получено при рандомизации а 3’-конаца VH CDR3.
После случайного мутагенеза последовательностей CDR3 легкой цепи 28G5, была идентифицирована панель scFv с эффективностью нейтрализации, которую можно было измерить в тесте с TF-1. Большинство улучшений мощности было получено при рандомизации 3’-конца VL CDR3.
После комбинирования библиотек случайного мутагенеза CDR3 тяжелой и легкой цепи, была идентифицирована панель scFv, эффективность которых в тесте на пролиферацию клеток TF-1, была лучше, чем у родительского scFv 28G5. Выделили ScFv с эффективностью, улчшенной в >60000 раз по сравнению с родительским 28G5. Все комбинации библиотек давали улучшенные scFv, т.е. H1/L1, H1/L2, H2/L1, H2/L2. Данный результат представляет особый интерес, поскольку ранее не удавалось изолировать улучшенные scFv из 1 библиотеки L.
Панель из 19 scFv, идентифицированных в процессе оптимизации 28G5, преобразовали в другой и экспрессировали в формате lgG4 описанными выше способами. Указанная панель состояла из клонов антител 1, 2 и с 4 по 20. Некоторые из наиболее эффективных клонов в этой панели были получены из комбинированных библиотек мутированных Н и L CDR3. Оценивали активность антител lgG4 в этой панели оценивали в тесте на пролиферацию клеток TF-1 и сравнивали с известным антителом мыши 2B7. Все оптимизированные lgG4 были в этом тесте более эффективными, чем 2B7. В данном случае 2B7 имело расчетную IC50 приблизительно равную 1.6 нМ, тогда как клоны имели расчетные значения IC50 в диапазоне от приблизительно 1 пМ до приблизительно 1100 пМ. Результаты представлены в Таблице 1 ниже и суммированы, следующим образом:
IC50<1500 пМ антитела 1, 2 и с 4 по 20
IC50<300 пМ антитела 1, 2, 4-12 и 14-20
IC50<60 пМ антитела 1, 2, 4-6, 8-11, 14 и 16-20
IC50<10 пМ антитела 1, 5, 6, 11 и 20.
На Фигуре 3 показана эффективность антагонистической активности двух примеров антител согласно настоящему изобретению, антитела 1 и антитела 6, в сравнении с известным антителом 2B7 в тесте на пролиферацию клеток TF-1.
Для оценки параметров кинетики взаимодействия некоторых из ведущих оптимизированных lgG4 с рекомбинантным меченным для очистки внеклеточным доменом рецептора GM-CSF применяли систему BIAcore 2000 (Pharmacia Biosensor). Аффинность антител была значительно улучшена, расчетные значения KD составляли от 0.127 нМ до приблизительно 5 нМ. Результаты показаны в Таблице 2. Улучшения были получены как для скоростей ассоциации, так и для скоростей диссоциации. Корреляция между аффинностью lgG4 к растворимому внеклеточному домену GM-CSFR а и поведением в тесте с TF-1 была очень хорошей, с коэффициентом корреляции Пирсона, равным 0.85 (р<0.0001). Для сравнения отдельно рассчитали KD 2B7, и было показано, что она равна приблизительно 7 нМ.
Антитела lgG4, идентифицированные в процессе оптимизации 28G5, оценивали в тесте на изменение формы гранулоцитов человека и сравнивали с известным антителом мыши 2B7. Все антитела, которые оценивали в этом тесте (антитела 1, 2, 5, 6, 9-11, 16 и 20) были очень эффективными, с IC50 в диапазоне от 7.8 до 90 пМ. Из них антитела 1, 2, 5, 6, 9, 16 и 20 имели IC50 меньше, чем 50 пМ, а антитела 1, 2, 6, 16 и 20 имели IC50 меньше, чем 25 пМ. Наши антитела были более мощными, чем 2B7, которое имело IC50, равную 477 пМ. Результаты показаны в Таблице 3. На Фигуре 4 показана эффективность антагонизма двух примеров антител согласно настоящему изобретению, антитела 1 и антитела 6, в сравнении с известным антителом 2B7 в тесте на изменение формы гранулоцитов человека.
Антитела lgG4, идентифицированные в процессе оптимизации 28G5, оценивали в тесте на изменение формы гранулоцитов яванского макака. Все антитела были способны нейтрализовать активность GM-CSF в отношении рецептора яванского макака, также как и рецептора человека, и все антитела были более эффективными, чем 2B7. 2B7 имело IC50, равную 26 пМ, тогда как типичные антитела (антитело 6, антитело 1 и антитело 2) из указанный панели имели значения IC50, равные 1.73, 2.03 и 3.2 пМ, соответственно.
Оценивали эффективность нейтрализации панели lgG4, идентифицированных в процессе оптимизации 28G5 в тесте на высвобождение TNFα моноцитами. Этот тест позволяет исследовать способность ингибировать высвобождение провоспалительного фактора TNFα из моноцитов человека при обработке моноцитов GM-CSF. Тестировали антитела 1, 2, 5, 6, 9 и 10, и все они были активны в данном тесте и были способны полностью нейтрализовать цействие GM-CSF на его рецептор (IC50 в диапазоне от приблизительно 43 до 139), тогда как 2B7 при концентрации 333 нМ могло обеспечить лишь 50% ингибирование индуцированного GM-CSF высвобождения TNFα, что указывает на то, что данное антитело является лишь частичным ингибитором в этом тесте. На Фигуре 5 показана эффективность антагонизма двух типичных антител согласно настоящему изобретению в сравнении с известным антителом 2B7 в тесте на высвобождение TNFα моноцитами. Результаты показаны в Таблице 4, их можно суммировать следующим образом:
<150 пМ антитела с номерами 1, 2, 5, 6, 9 и 10
<110 пМ антитела с номерами 1, 2, 5, 6 и 9
<100 пМ антитела с номерами 1, 5, 6 и 9
Признаком воспалительного заболевания является повышение выживаемости воспалительных типов клеток в ответ на определенные сигналы. Гранулоциты способны выживать в течение большего времени в присутствии GM-CSF, и поэтому способность антител lgG4, выделенных в процессе оптимизации 28G5, ингибировать этот ответ оценивали в тесте на выживаемость гранулоцитов. Все анти-GM-CSFRα lgG4 из ведущей оптимизации были активны в этом тесте, и характерные значения эффективности нейтрализации (IC50) лежали в диапазоне от 7.0 до 843.7 пМ. Это отличалось от известного антитела мыши 2B7, которое было полностью не активно вплоть до концентрации 83 нМ. На Фигуре 6 показана эффективность антагонизма двух типичных антител согласно настоящему изобретению, антитела 1 и антитела 6, в сравнении с известным антителом 2B7 в тесте на выживаемость гранулоцитов.
Эти результаты, показанные на Фигурах 3-6, указывают на то, что наши антитела имеют свойства, в значительной степени отличающиеся от свойств известного антитела мыши 2B7. Например, типичные антитела согласно настоящему изобретению ингибировали выживаемость гранулоцитов и пролиферацию TF-1, стимулируемую 7 пМ GM-CSF, в тесте на выживаемость гранулоцитов и в тесте на пролиферацию клеток TF-1, соответственно, тогда как 2B7 не ингибировало выживаемость гранулоцитов, но ингибировало пролиферацию TF-1 (хотя и в меньшей степени, чем наши антитела). Эти результаты указывают на то, что связывающие агенты согласно настоящему изобретению имеют более высокую аффинность и улучшенную способность ингибировать ряд биологических эффектов, опосредуемых GM-CSF-R, по сравнению с известными анти-GM-CSFRα антителами.
Полученную последовательность аминокислот 28G5 и его производных выравнивали с известными последовательностями зародышевой линии человека в базе данных VBASE, и определяли наиболее близкие зародышевые последовательности по сходству последовательностей. Наиболее близкие зародышевые последовательности для VH-домена 28G5 и его производных были идентифицированы как VH1 DP5. 28G5 VH имело 14 изменений в каркасных участках по сравнению с зародышевым VH 1-24 (DP5). Наиболее близкой зародышевой последовательностью для VL домена была VLambda1 VL 1-е (DPL8), в которой было лишь 5 изменений по сравнению с зародышевой последовательностью в каркасных участках. Каркасные участки 28G5 и его производных эмбрионизировали путем сайтнаправленного мутагенеза, чтобы они идентично совпадали с нативными антителами человека. Все, кроме одной аминокислоты, удалось эмбрионизировать лишь с небольшим изменением эффективности антитела. Аминокислота изолейцин в положении 94 тяжелой цепи (используется нумерация по Kabat, Kabat и др. 1971) не может быть изменена на зародышевый треонин без полной потери активности. Это единственное отличие от зародышевой последовательности было, соответсвенно, сохранено в каркасном участке антитела.
Полный анализ рА2Двух анти-GM-CSFRα антител, антитела 6 и антитела 1, проводили в тесте на пролиферацию клеток TF-1. Результаты подтвердили, что эти антитела являются высоко эффективными антагонистами в этих системах: расчетные значения рА2 составили -11.3±0.2 и -11.0±0.2, соответственно (Фигура 1).
Полный анализ рА2одного из анти-GM-CSFRα антител, антитела 6, проводили в тесте на изменение формы гранулоцитов человека и яванского макака. Результаты подтвердили, что это антитело является высоко эффективным антагонистом в этих системах: расчетные значения рА2 составили -10.58 и -10.78 в тестах с гранулацитами человека и яванского макака, соответственно (Фигура 2).
GM-CSF запускает дифференцировку клеток-предшественников гематопоэтического ряда в колонии гранулоцитов и макрофагов в тестах на полутвердом агаре. Соответственно, антитело 6 и антитело 1 со зрелой аффинностью, родительское MAT антитело 3 (28G5) и отрицательный контроль (САТ001) тестировали на их способность антагонизировать эту специфичную активность GM-CSF с применением клеток-предшественников, полученных из периферической крови, в тесте на образование колоний. Результаты, представленные на Фигуре 7, показывают, что оба представителей MAT со зрелой аффинностью были эффективными ингибиторами опосредуемого GM-CSF человека образования колоний гематопоэтических клеток in vitro.
Приблизительные значения IC50 были равны 0.08 мкг/мл (антитело 6) и 0.25 мкг/мл (антитело 1) для MAT со зрелой аффинностью. Интересно, что известное антитело мыши 2B7 вплоть до концентрации 66 нМ, если и имело какую-то ингибиторную активность в этом тесте, то она была очень низкой.
В контрольных экспериментах указанные MAT со зрелой аффинностью, как и ожидалось, не влияли на образование колоний, опосредуемое комбинацией SCF+IL-3+G-CSF, и в отсутствие цитокинов образование колоний было незначительным (<4 колонии/культуру).
Для in vivo анализа huGM-CSFRα-специфичной антагонистической активности MAT можно применять трансплантацию костного мозга от трансгенных (Tg) мышей, экспрессирующих и α и β цепи GM-CSFR человека, мышам дикого типа, в результате которой получают химерных животных у которых экспрессия трансгенного huGM-CSFR ограничена гематопоэтическими клетками, произошедшими из костного мозга, и таким образом она наиболее близка к профилю экспрессии эндогенного рецептора. В таких Tg химерных мышах введение huGM-CSF приводит к увеличению веса селезенки и маргинализации моноцитов циркулирующей крови. Антитело 6 со зрелой аффинностью и отрицательный контроль MAT, CAT001, оценивали на их способность антагонизировать эти опосредуемые GM-CSF in vivo ответы. Для анализа зависимости доза-ответ, 6 группам по 5 Tg химерных мышей дважды в день в течение 4 дней (день 1-4) вводили подкожно 500 нг huGM-CSF, а семи контрольным группам по пять животных вводили только ФБР. Четырем из 6 групп животных, которым вводили huGM-CSF, в день вводили тестируемое MAT (антитело 6) в концентрации 16 мг/кг, 5.3 мг/кг, 1.78 мг/кг или 0.59 мг/кг 0, а пятой группе животных, которым вводили huGM-CSF, в день обводили контрольное CAT001 в концентрации 16 мг/кг. Результаты, представленные на Фигуре 8, показывают, что по сравнению с контролем ФБР, введение huGM-CSF индуцировало значительное увеличение веса селезенки и снижение моноцитов циркулирующей крови. Как и ожидалось, введение 16 мг/кг контрольного CAT001 не влияло ни на увеличение веса селезенки, ни на снижение числа моноцитов крови. Напротив, наблюдался четкий эффект доза-ответ после введения тестируемого MAT антитела 6, поскольку 16 мг/кг этого антитела устраняло увеличение веса селезенки и, хотя все еще выраженный, указанный эффект сильно снижался при 0.59 мг/кг MAT. IC50, по-видимому, была где-то между 0.59 мг/кг и 1.78 мг/кг. Сходный результат наблюдали дляиндуцируемого GM-CSF уменьшения числа циркулирующих моноцитов - введение тестируемого MAT антитела 6 при 16 мг/кг прекращало снижение, a MAT при 0.59 мг/кг оказывало лишь небольшое влияние на этот ответ. Эти результаты показывают, что анти-GM-CSFRα антитело является антагонистом GM-CSFRα человека in vivo.
Чтобы дополнительно исследовать противовоспалительные свойства этих анти-GM-CSFRα антител, антитело 6 оценивали в тесте на высвобождение цитокинов мононуклеарами периферической крови. В этом тесте TNFα и IL-6 могут эндогенно высвобождаться в зависимости от донора. В этом тесте также имеет место эндогенная продукция GM-CSF, а не его экзогенное добавление, и таким образом, результаты, наблюдаемые в этом анализе, представляют ингибирование биологических эффектов связывания нативного эндогенного GM-CSF с его рецептором.
После введения антитела 6 имело место дозозависимое ингибирование обоих цитокинов, как показано на фигуре 9. Эти результаты указывают на то, что данные антитела могут ингибировать активность нативного GM-CSF и что посредством ингибирования передачи сигнала GM-CSF можно ингибировать ключевые провоспалительные цитокины, такие как IL-6 и TNFα, каждый из которых вовлечен в ряд воспалительных заболеваний, таких как ревматоидный артрит.
Более того, на основании этого результата с антителом 6, можно ожидать, что каждое из антител с 1 по 20 также будет проявлять ингибирование в этом тесте, посколько все антитела с 1 по 20, как считают, связывают один и тот же участок GM-CSFRα.
Картирование остатков, важных для распознавания антигена, и анализ последовательности
Мы определяли вариабельность остатков в положениях в последовательности scFv эмбрионизированного антитела 6, чтобы выявить положения, которые обычно консервативны, для связывания лиганда и какие положения вариабельные в антителе, сохраняющем активность связывания лиганда.
Положениями, участвующими в связывании антигена, оказались остатки 27А, 27B, 27С, 32, 51, 52, 53, 90, 92 и 96 (нумерация по Kabat) в VL-домене и остатки 17, 34, 54, 57, 95, 97, 99 и 100B (нумерация по Kabat) в VH-домене.
Были определены семь положений, которые оказались важными для связывания антигена: Н95, Н97, Н99, H100B, L90, L92 и L96. Затем мы анализировали остатки в этих положениях в последовательностях 160 вариантов, изолированных в процессе оптимизации антитела 28G5, каждый из которых проявил по меньшей мере минимум пятикратное улучшение эффективности в тесте на пролиферацию клеток TF-1.
Таблице 5 ниже собраны различные аминокислоты (из 20 возможных), которые наблюдали в каждом из этих положений, и в L95A. Когда положения строго консервативны в отношении аминокислот, присутствующих в 28G5 и/или антителе 6, это является хорошим свидетельством того, что данные аминокислоты являются ключевыми для связывания антигена. Например, остатки в следующих положениях являются строго консервативными: Н97, H100B, L90, L92.
Метод
Последовательность ДНК, кодирующая эмбрионизированное scFv антитело 6 со зрелой аффинностью, была переведена в формат рибосомного дисплея, по существу как описано в источнике [67]. Проводили ПЦР с повыщенным уровнем ошибок на последовательности антитела 6, применяя условия с высокой частотой мутаций (7.2 мутации на 1000 п.о.) по протоколу производителя (BD Bioscience), в результате библиотеку вариантных последовательностей 574D04, содержащих рандомизированные точечные мутации. Эту библиотеку экспрессировали на рибосомах и инкубировали с меченным для очистки GM-CSFRα, чтобы обеспечить связывание. Варианты, способные связывать меченный GM-CSFRα, фиксировали и удаляли, применяя парамагнитные гранулы, покрытые белком G (Dynal). Несвязавшиеся варианты, оставшиеся в популяции, добавляли к пулу из четырех биотинилированных анти-идиотипических антител, которые были получены ранее из обширной библиотеки антител человека в формате фагового дисплея, описанной в [68], и, как было известно, связывались с scFv антителом 6. Варианты, которые связались с биотинилированными анти-идиотипическими антителами, фиксировали на стрептавидиновых гранулах, а несвязавшиеся варианты отмывали. Этот процесс повторяли в двух дополнительных раундах селекции из рибосомного дисплея, следуя основной методике из [67].
Представительную долю вариантов из продуктов селекции клонировали в фагмидный вектор и экспрессировали варианты scFv на фаге для тестирования методом твердофазного ИФА, применяя такой же способ, какой описан у Edwards ВМ и др. (2003) Journal of Molecular Biology, том 334:103. Варианты, которые не демонстрировали связывания с меченным для очистки GM-CSFRa, тестировали на связывание с пулом из четырех анти-идиотипических антител, которые применяли для селекции. Варианты, которые в тесте на связывание с анти-идиотипическими антителами продемонстрировали связывание, разное или большее, чем у scFv антитело 6, подвергали секвенированию и анализу последовательности, чтобы выявить положения, в которых с высокой частотой происходили мутации.
Средний уровень мутаций в популяции вариантов был примерно равен 3.05 аминокислот на цепь VH или VL, из 486 последовательностей для VH-цепей и 451 последовательностей для VL-цепей. Их анализировали на «горячие» точки мутирования путем построения графика частоты мутаций по отношению к их положению вдоль scFv. Анализ был сосредоточен на тех клонах, которые имели по меньшей мере одну мутацию в гипервариабельном участке на VH и VL и менее 4 мутации на VH и VL. Из этой панели из 123 последовательностей VH и 148 последовательностей VL, определили «горячие» точки: точки, имеющие частоту мутаций, равную 5% или более.
Путем отрицательной селекции из рибосомного дисплея были выделены семь положений внутри VHCDR3 и VLCDR3 антитела 6 как предполагаемые положения, важные для связывания антигена. Затем выполняли анализ на 160 вариантах последовательности, изолированных в процессе оптимизации антитела 28G5, в котором целые последовательности VHCDR3 и VLCDR3 рандомизировали и отбирали по наиболее высокой аффинности. Все последовательности (включая антитело 6) представляют собой варианты 28G5, которые проявили как минимум пятикратное улучшение эффективности в тесте на пролиферацию клеток TF-1.
Определение линейного эпитопа
Мы провели скрининг антитела 6 и известного антитела 2B7 против 2442 пептидов, каждый из которых представлял собой короткий участок последовательности аминокислот из внеклеточной части GM-CSFR-α, применяя способ PEPSCAN. Усредняли сигналы связывания для каждого антитела против всех пептидов с получением среднего фонового сигнала, и для каждого пептида рассчитывали отношение сигнал/фон. Для обоих антител, 6 и 2B7, отношение сигнал/фон, равное четырем или выше, считали специфичным, положительным сигналом. Последовательности пептидов, дающие специфичный, положительный сигнал, анализировали на консервативные связывающие мотивы, и обнаружили, что антитело 6 предпочтительно связывалось с мотивом YLDFQ, соответствующим остаткам с 226 по 230 зрелого GM-CSFRα человека, а 2B7 антитело предпочтительно связывалось с мотивом DVRI, соответствующим остаткам с 278 по 281 зрелого GM-CSFRα человека. Нумерация последовательности аминокислот для зрелого рецептора приведена в SEQ ID NO: 206.
Способ PEPSCAN (сканирование связывания пептидов)
Синтезировали перекрывающиеся, преимущественно состоящие из 15 мономеров синтетические пептиды, имеющие последовательности, полученные из GMCSF, и подвергали скринингу с помощью мини-PEPSCAN карт формата кредитной карты (455-луночный планшет с 3 мкл лунками), как описано ранее [69]. Связывание антитела с каждым пептидом тестировали в основанном в твердофазном иммуноферментном анализе (ELISA) на основе PEPSCAN. 455-луночные полипропиленовые карты формата кредитной карты, содержащие ковалентно связанные пептиды, инкубировали с образцом (например, 10 мкг/мл антитела или сыворотки, разбавленных 1/1000 в растворе ФБР, который содержит 5% лошадиной сыворотки (в объемном отношении, об./об.) и 5% овальбумина (вес/объем, в./об.)) и 1% Tween 80 или, в случае легкого блокирования, в растворе ФБР с 4% лошадиной сывороткой (об./об.) и 1% Tween 80 (4°C, в течение ночи). После промывки указанные пептиды инкубировали с меченными пероксидазой антителами к антителам (разведение 1/1000, например, меченными пероксидазой антителами кролика к антителам мыши, Dako) (1 ч., 25°C), и затем, после промывки, добавляли субстрат пероксидазы 2,2’-азиноди-(3-этилбензтиазолин-6-сульфонат) (ABTS) и 2 мкл/мл 3% H2O2. Через 1 час измеряли развитие цвета. Развитие цвета ELISA оценивали количественно с помощью ПЗС-камеры и системы обработки изображений. Установка состоит из ПЗС-камеры и 55 мм объектива (видео кмера Sony CCD XC-77RR, объектив Nikon micro-nikkor 55 мм f/2.8), адаптера камеры (Sony Camara adaptor DC-77RR) и пакета программного обеспечения для обработки изображений Optimas, версия 6.5 (Media Cybernetics, Сильвер Спринг, Мэриленд 20910, США). Optimas запускали на компьютере pentium.
Материалы и методы анализа
Биохимический тест на связывание лиганда
Очищенные препараты scFv получали, как описано в Примере 3 в WO 01/66754 [70]. Концентрации белка очищенных препаратов scFv определяли методом ВСА [71]. 96-луночные микротитрационные планшеты Fluoronunc™ покрывали в течение ночи при 4°C 50 мкл/лунку антител к IgG4 человека, разбавленных до 2.5 мкг/мл в ФБР. Планшеты промывали 3 раза, используя 300 мкл/лунку ФБР/О. 1% Tween-20, затем блокировали в течение 1 часа при комнатной температуре с помощью 300 мкл/лунку 3% бычьего сывороточного альбумина (БСА) в ФБР. Планшеты снова промывали 3 раза с помощью 300 мкл/лунку ФБР/0.1% Tween-20 и затем добавляли в каждую лунку 50 мкл GM-CSFRα человека, разбавленного до 62.5 нг/мл в 1% БСА/ФБР, и планшеты инкубировали в течение 1 часа при комнатной температуре. После 3-кратной промывки, как описано выше, в каждую лунку добавляли 25 мкл материала образца, после чего добавляли 25 мкл биотинилированного GM-CSF, разбавленного до 2 нМ в 1% БСА/ФБР. Чтобы определить общее связывание, использовали только буфер в качестве материала образца. Чтобы определить неспецифичное связывание в качестве материала образца применяли немеченый GM-CSF, разбавленный до 100 нМ в 1% БСА. Планшеты инкубировали в течение 1 часа при комнатной температуре перед 3-кратной промывкой, как описано выше. Добавляли в каждую лунку планшета 50 мкл стрептавидина, меченного европием (PerkinElmer), разбавленного до 100 нг/мл в буфере для теста DELFIA™ и инкубировали в течение 30-60 минут при комнатной температуре, а затем промывали 7 раз буфером для промывки DELFIA™. Добавляли в планшеты 50 мкл/лунку усиливающего раствора DELFIA™ и образцы считывали при 615 нм на считывающем устройстве для планшетов.
Тест на пролиферацию клеток TF-1
Клетки TF-1, приобретенные в R&D Systems и поддерживаемые по стандартной методике в RPMI 1640 с 10% ФБР, 1 мМ пирувата натрия и 4 нг/мл GM-CSF, истощали путем 3-кратной промывки в среде для анализа (RPMI 1640, 5% ФБР, 1 мМ пируват натрия), ресуспендировали в среде для анализа и инкубировали в течение 7-24 часов при 37°C в 5% CO2. Затем клетки ресуспендировали при 1×105/мл в среде для анализа и добавляли по 100 мкл в каждую лунку 96-луночного планшета с плоским дном для выращивания культуры. Тестируемые образцы получали путем стерильной фильтрации исходного образца с последующим разведением в среде для теста. Затем в каждую лунку с клетками добавляли 50 мкл тестируемого материала и инкубировали в течение 45-60 минут при 37°C в 5% CO2. Затем в каждую лунку добавляли 50 мкл GM-CSF, разбавленного до значения ЕС80 в среде для анализа (или 0.4 нг/мл для некоторых GM-CSF), и планшеты инкубировали в течение 16 часов при 37°C в 5% CO2 в увлажнительной камере. Это представляло конечную концентрацию 7 пМ GM-CSF. Для измерения пролиферации клеток, 20 мкл 3H-тимидина, разбавленного до 5.0 мкКи/мл в среде для анализа, добавляли в каждую лунку планшета, и планшеты инкубировали в течение 4 часов ±30 минут при 37°C в 5% CO2. Клетки собирали на 96-луночные планшеты GF/C Unifilter™, применяя планшетный коллектор, и промывали. После добавления 50 мкл MicroScint 20™ в каждую лунку фильтровального планшета, планшеты запечатывали и считывали на регистрирующем устройстве для планшетов TopCount.
Тест на изменение формы гранулоцитов человека
Лейкоцитарные пленки человека (сгустки крови человека из службы переливания крови) смешивали с равным объемом 3% декстрана Т-500 в 0.9% NaCl. Затем смесь инкубировали в вертикальном положении до формирования границы раздела. Верхний слой собирали и наносили на градиент плотности Histopaque 1.077, который потом центрифугировали при 400 g в течение 40 минут и позволяли остановиться без торможения. Верхние слои этого градиента удаляли, оставляя осадок гранулоцитов. Все оставшиеся эритроциты в осадке лизировали путем ресуспендирования клеток в 20 мл ледяной воды в течение 30 секунд, после чего сразу же добавляли ледяной 1.8% хлорид натрия. Затем клетки переосаждали при 1200 об/мин и ресуспендировали в среде для анализа (RPMI1640, 10% ФБР, 100 мкл/мл пенициллина, 100 мкг/мл стрептомицина, 25 мМ HEPES) при 1×106/мл. Затем в каждую лунку 96-луночного планшета с плоским дном добавляли 100 мкл клеток для выращивания культуры. Тестируемые образцы получали путем стерильной фильтрации исходных образцов и соответствующего разбавления в среде для анализа.
Для выделения ведущих элементов, к клеткам затем добавляли 50 мкл тестируемого образца, и планшеты инкубировали в течение 45-60 минут при 37°C в 5% СО2. Это эквивалентно конечной концентрации 7 пМ GM-CSF. После этого следовали добавление 50 мкл GM-CSF, разбавленного до 0.4 нг/мл в среде для анализа, в каждую лунку и 4-часовая инкубация при 37°C в 5% CO2 в увлажнительной камере.
Для оптимизации ведущих элементов, фильтрованные lgG4, разбавленные в среде для анализа, смешивали с равным объемом GM-CSF при 0.4 нг/мл в среде для анализа. Это эквивалентно конечной концентрации 7 пМ GM-CSF. 100 мкл смеси антитело/GM-CSF затем добавляли в каждую лунку. После этого следовала 3-часовая инкубация при 37°C в 5% CO2 в увлажнительной камере.
Холодный формальдегид добавляли до конечной концентрации 1.25% и клетки фиксировали в течение ночи при 4°C. Методом проточной цитометрии анализировали 2000-5000 событий на лунку. Затем получали среднее геометрическое значение прямого светорассеяния (FSC) для каждого образца, применяя CellQuest. Клетки отделяли по пороговым значениям, чтобы исключить посторонние популяции (например, мертвые клетки/обломки) при рассчете среднего геометрического значения.
Тест на изменение формы гранулоцитов яванского макака
Антитела оценивали в тесте, позволяющем измерить изменение формы гранулоцитов яванского макака после стимуляции GM-CSF. Гранулоциты очищали от остальных элементов крови яванского макака и проводили анализ по существу как описано для анализа на изменение формы гранулоцитов человека.
Значения аффинности связывания, полученные с помощью биосенсорного анализа
Для оценки параметров кинетики взаимодействия между scFv и lgG4 и рекомбинантными рецепторами применяли BIAcore 2000 System (Pharmacia Biosensor). В биосенсорном анализе оптические эффекты поверхностного плазменного резонанса используют для изучения изменений в поверхностной концентрации в результате взаимодействия молекулы аналита с молекулой лиганда, которая ковалентно присоединена к декстрановой матрице. Как правило, частицы аналита в свободном растворе проходят через связанный лиганд, и любое связывание детектируют как возрастание локального сигнала ППР. После этого следует период промывки, во время которого наблюдают диссоциацию частиц аналита как понижение сигнала ППР, после которого любые остатки аналита десорбируют с лиганда и повторяют процедуру при нескольких различных концентрациях аналита. Во время эксперимента обычно применяют серию контролей, чтобы удостовериться, что ни абсолютная связывающая способность или кинетический профиль связанного лиганда не изменяются значительно. Фирменный буферный солевой раствор HEPES (HBS-EP) обычно используют в качестве основного разбавителя образцов аналита и растворителя фазы диссоциации. Экспериментальные данные фиксировали в резонансных единицах (непосредственно соответствующих ППР сигналу) по отношению к времени. Резонансные единицы прямо пропорциональны размеру и количеству связавшегося аналита. Затем можно использовать пакет программного обеспечения BIAevaluation, чтобы определить константы скорости фазы диссоциации (единицы скорости диссоциации с-1) и фазы ассоциации (единицы скорости ассоциации М-1 с-1). Эти значения затем позволяют рассчитать аффинные константы ассоциации и диссоциации.
Аффинность lgG4 оценивали, применяя один тест, в котором происходит нековалентный захват lgG4 поверхностью аминного белка А. Затем через lgG4 последовательно пропускали серию разведении от 100 до 6.25 нМ рекомбинантного меченного для очистки внеклеточного домена рецептора GM-CSF. Рассчитывали молярность рецептора, используя концентрацию (Bradford) и предсказанную массу не зрелого полипептида после посттрансляционной модификации (39.7 кДа). Каждый из двух отдельных наборов данных анализировали одинаковым образом. Откорректированные по эталонному элементу данные аппроксимировали кривой по модели Ленгмюра 1:1 для одновременного глобального расчета скоростей ассоциации и диссоциации, со значением Rmax, приравненным к глобальному. Оценивали уровень lgG4, захваченного во время каждого цикла, чтобы удостовериться, что захваченное количество оставалось стабильным на протяжении всего эксперимента. Дополнительно оценивали скорость диссоциации lgG4, чтобы определить, требуется ли корректировка отклонения базового уровня. Тем не менее, оказалось, что оба взаимодействия белка А достаточно воспроизводимы и стабильны. Достоверность данных была ограничена рассчитанными хи2 и Т-значения (параметр значение/отклонение), которые должны быть <2 и >100 соответственно.
Получение меченного для очистки внеклеточного домена GM-CSFRα:
Вектор экспрессии pEFBOS [72], включающий последовательность, кодирующую внеклеточный домен рецептора α GM-CSF человека (SEQ ID NO: 205, представляющая аминокислоты с 1 по 298 зрелого GM-CSF R) с сигнальной последовательностью IL-3 мыши, и включающая N-концевую метку для очистки, применяли для получения рекомбинантного меченного по N-концу полипептида внеклеточного домена (ВКД) рецептора GM-CSF. Меченный полипептид ВКД экспрессировали в клетках СНО, применяя вектор pEFBOS, используя стандартные процедуры. Этот полипептид также может быть назван очищенным внеклеточным доменом GM-CSFRα, или растворимым внеклеточным доменом GM-CSFRα.
Можно применять любую подходящую метку для очистки, например, пептид Flag (DYKDDDE - SEQ ID NO: 204), Fc, биотин или гистидиновую метку. Очистку можно проводить с помощью любой подходящей методики, например, Flag-меченный полипептид ВКД (SEQ ID NO: 203) может быть очищен на колонке М2для аффинной хроматографии и элюирован пептидом FLAG.
Тест на высвобождение TNFα моноцитами
Очистка моноцитов (Monocyte Isolation Kit - Miltenyi Biotec - 130-053-301):
Лейкоцитарные пленки человека (сгустки крови человека из службы переливания крови) наносили на градиент плотности Histopaque 1.077 (Sigma, Cat No. 1077-1) и клетки центрифугировали при 400×g в течение 40 минут. Центрифуга останавливалась без применения тормоза. МНПК затем собирали с поверхности раздела фаз. Клетки промывали в ФБР и осаждали при 300×g в течение 10 минут, а затем лизировали оставшиеся эритроциты путем ресуспендирования в 20 мл ледяной воды в течение 15 с, после чего немедленно добавляли ледяной 1.8% NaCl. Клетки затем осаждали при 1200 об/мин в течение 5 минут и ресуспендировали в 600 мкл буфера MACS (ФБР, 2 мМ EDTA). 200 мкл неблокирующего реагента, поставляемого в наборе, добавляли к клеткам и перемешивали перед добавлением 200 мкл коктейля гаптен-антитело (также поставляемого в наборе) и перемешиванием. Клетки затем инкубировали при 4°C в течение 15 минут, после чего дважды промывали в 50 мл MACS буфера. Осадок клеток ресуспендировали в 600 мкл буфера MACS, а затем добавляли 200 мкл Fc-блокирующего реагента и перемешивали, после чего добавляли 200 мкл анти-гаптеновые микрогранул в MACS и перемешивали. Клетки инкубировали в течение 45 минут при 4°C перед промывкой в 50 мл буфера MACS" и ресуспендированием в 500 мкл буфера MACS. Приготовили одну колонку (Miltenyi Biotec 130-042-401) путем пропускания через нее 3 мл буфера MACS, а затем на колонку нанесли суспензию клеток. Собирали элюат в виде обогащенной моноцитами фракции. Колонку промывали 2 раза/3 мл буфера MACS и собирали элюат. Чистоту моноцитов проверяли путем окрашивания анти-CD14-PE, применяя стандартные способы проточной цитометрии. Клетки последний раз ресуспендировали при 4×106/мл в среде для анализа (RPMI 1640, 10% ФБС, 100 ед/мл пенициллина, 100 мкг/мл стрептомицина).
Стимуляция моноцитов;
50 мкл клеток добавляли в каждую лунку Costar 96-луночного планшета с плоским дном для выращивания культуры. Во все лунки добавляли 25 мкл 150 мкг/мл rhIFNγ (R&D systyems). Фильтрованные lgG4, разбавленные в среде для анализа, смешивали с равным объемом GM-CSF при 56 нг/мл (4 нМ) в среде для анализа. Это эквивалентно конечной концентрации 1 нМ GM-CSF. Затем в каждую лунку добавляли 75 мкл смеси антитело/GM-CSF. Контрольными были лунки, содержащие только GM-CSF или без GM-CSF и без антитела. Затем планшеты инкубировали в течение 18 часов при 37°C с 5% CO2 в увлажнительной камере. Затем собирали супернатант, чтобы проверить на уровни TNF-α методом ELISA.
Твердофазный ИФА на TNFα (R&D Systems ELISA Development System DY210):
Планшеты Fluoronunc Immunosorb ELISA покрывали в течение ночи при комнатной температуре 100 мкл захватывающего антитела при 4 мкг/мл в ФБР. Планшеты затем промывали три раза ФБР/0.1% Tween и блокировали 300 мкл/лунку 3% Marvel на ФБР в течение 1 часа при комнатной температуре. Планшеты промывали 3 раза в ФБР/0.1% Tween. 100 мкл супернатантов из планшетов для анализа переносили на планшет ELISA и добавляли в контрольные лунки титрованный TNF-α разбавленный в среде для анализа. Планшеты инкубировали при комнатной температуре в течение 2 часов, а затем промывали 4-5 раз в ФБР/0.1% Tween. 100 мкл детектирующего антитела, разбавленного до 300 нг/мл в 1% Marvel/ФБР, добавляли в каждую лунку указанного планшета и планшеты инкубировали в течение дополнительных 2 часов при комнатной температуре, а затем промывали 4-5 раз в ФБР/0.1% Tween. Стрептавидин-Европий (PerkinElmer 1244-360) разбавляли 1:1000 в буфере для анализа DELFIA (PerkinElmer 4002-0010) и добавляли 100 мкл/лунку перед инкубированием в течение 45 минут при комнатной температуре. Затем планшеты промывали 7 раз в буфере для промывки DELFIA, после чего добавляли 100 мкл/лунку усиливающего раствора (PerkinElmer 4001-0010) и считывали сигнал при 615 нм на считывающем устройстве для планшетов.
Тест на выживаемость гранулоцитов
Клетки выделяли из лейкоцитарных пленок человека, как описано для анализа на активацию нейтрофилов (теста на изменение формы), промывали, в среде для анализа (RPMI-1640 Glutamax, 10% ФБС, 100 Ед/мл пенициллина, 100 мкг/мл стрептомицина) и ресуспендировали при 1×106/мл в среде для анализа. В каждую лунку 96-луночного планшета Costar с плоским дном для выращивания культуры добавляли 100 мкл клеток. Фильтрованные исходные растворы антител разбавляли в среде для анализа и смешивали с равным объемом GM-CSF при 0.4 нг/мл. Это эквивалентно конечной концентрации 7 пМ GM-CSF. Контрольные лунки содержали только среду или только GM-CSF. Затем в каждую лунку планшета добавляли 100 мкл смеси тестируемого образца/GM-CSF и клетки инкубировали в течение 68 часов при 37°C /5% CO2 в увлажнительной камере. 20 мкл красителя AlamarBlue добавляли в каждую лунку и инкубировали планшеты в течение дополнительных 24 часов при 37°C/5% CO2 в увлажнительной камере. Планшеты затем считывали при 560 нм и 590 нм на считывающем устройстве для планшетов.
Анализ рА2 анти-GM-CSFRα антител в тесте на пролиферацию клеток TF-1 и в тестах на изменение формы гранулоцитов человека и яванского макака
Основным фармакологическим инструментом для количественного анализа аффинности конкурентного антагониста является анализ Шилда. Используя этот подход, можно определить независимые от системы средства оценки антагонистической аффинности в функциональном тесте. Указанный способ основан на концепции, что концентрация антагониста и его аффинность определяют антагонизм ответу агониста. Поскольку, антагонизм можно оценить количественно и концентрация антагониста известна, можно определить аффинность антагониста. Этот антагонизм оценивали количественно путем измерения соотношения эквиактивных концентраций агонистов, измеренных в присутствии и в отсутствие антагониста (т.н. отношение доз (DR)).
Отношения доз можно рассчитать по отношению ЕС50 агониста (обычно, GM-CSF) в отсутствие связывающего агента к ЕС50 агониста в присутствии одной концентрации связывающего агента. Отношения доз, выраженные в виде log(DR-1), можно затем использовать в линейной регрессии по логарифму концентрации связывающего агента (log[связывающего агента]), чтобы получить регрессионную модель Шилда. Таким образом, каждой концентрации связывающего агента будет соответствовать значение DR; эти значения наносят на график в виде регрессии log(DR-1) по отношению к log [связывающего агента]. Если антагонизм является конкурентным, будет иметь место линейная зависимость между log (DR-1) и log [связывающего агента] согласно уравнению Шилда, при этом уравнение имеет следующую форму:
Log(DR-1)=log[A]-logKA
При этих условиях, нулевое значение по ординате даст пересечение оси х, при котором log[a]=logKA. Таким образом, концентрация связывающего агента, которая дает log(DR-1)=0 будет равной logKA, равновесной константе диссоциации комплекса связывающий агент - рецептор. Это является независимым от системы количественным анализом аффинности связывающего агента, который должен быть правильным для каждой клеточной системы, содержащей рецептор.
Поскольку значения KA получены по графику в логарифмическом масштабе, они распределены по логарифмически нормальному закону. Отрицательной логарифм конкретной концентрации называют эмпирически рА2, концентрацией антагониста, которая вызывает двухкратный сдвиг кривой ответа на дозу агониста. Эффективность антагонистической активности можно оценить количественно путем расчета рА2 по одной концентрации антагониста, которая дает одно значение для соотношения доз из уравнения, в котором:
рА2=log (DR-1)-log[a]
[а]= молярная концентрация антагониста, которая делает необходимым удвоение концентрации агониста для достижения исходного субмаксимального ответа.
DR= отношение доз, определенное путем измерения отношения эквиактивных концентраций агониста, измеренных в присутствии и в отсутствие антагониста.
рА2 можно рассчитать по результатам теста доза-ответ.
Ингибирование in vitro GM-CSF-опосредованной дифференцировки клеток-предшественников клеток крови в тесте на образование колоний
Мононуклеары периферической крови, обогащенные клетками-предшественниками гематопоэтического ряда, были получены от доноров, которые прошли мобилизацию клеток-предшественников и аферез, как часть стандартного клинического лечения. Образцы анонимизировали и клетки перед применением не замораживали. 5×104 мононуклеаров культивировали в полутвердом агаре [73] в присутствии GM-CSF человека в конечной концентрации, равной 10 нг/мл. Тестируемые MAT человека со зрелой аффинностью, и известное антитело мыши 2B7, добавляли в культуры на агаре в конечной концентрации 10, 5, 1, 0.5, 0.1 или 0.05 мкг/мл. Родительское MAT человека 28G5 и изотипически сходный отрицательный контроль MAT человека, САТ001, тестировали в одной концентрации, равной 10 мкг/мл. Для контроля MAT также тестировали на способность блокировать образование колоний, стимулируемое комбинацией SCF, IL-3 и G-CSF (Croker и др., 2004) и на вклад в образование колоний в отсутствие цитокинов. Образование колоний (агрегаты из >40 клеток) оценивали через 14 дней инкубации при 37°C с 10% CO2 в атмосфере. Колонии фиксировали глютаровым альдегидом и подсчитывали, используя препаративный микроскоп при увеличении 35Х.
Ингибирование биологической активности GM-CSF in vivo в мышах, с трансгенным GM-CSFRαβ человека
Получили трансгенные (Tg) мыши, экспрессирующие как α-, так и β-цепи GM-CSFR человека под контролем промотора главного комплекса гистосовместимости класса I, и исследовали in vivo ответы клеток селезенки и крови на введение huGM-CSF [74]. Для in vivo анализа huGM-CSFRa-специфичной антагонистической активности MAT, можно применять трансплантацию костного мозга от Tg мышей мышам дикого типа с получением химерных животных, у которых экспрессия трансгенного huGM-CSFRαβ ограничена гематопоэтическими клетками, происходящими из костного мозга, и, таким образом, имеет профиль, наиболее близкий профилю экспрессии эндогенного рецептора. В этих Tg химерных мышах huGM-CSFRαβ введение huGM-CSF приводит к увеличению веса селезенки и маргинализации моноцитов циркулирующей крови.
Получение Tg химерных мышей:
У Tg мышей-доноров удаляли бедренные кости и большеберцовые кости и вымывали костный мозг с помощью стерильного ФБР с 3% фетальной бычьей сыворотки (ФБС). Сгустки костного мозга затем пропускали через иглу 23G для шприца, в результате чего получали гомогенную суспензию клеток, затем клетки промывали один раз холодным ФБР+3% ФБС и пропускали через сито из нержавеющей стали. Эритроциты затем удаляли путем лизирования в буфере с 0.168 М хлорида аммония, после чего клетки промывали еще два раза фосфатно-солевым буферным раствором (ФБР)+3% ФБС, а затем снова пропускали через сито из нержавеющей стали. Чтобы дополнительно удалить мертвые клетки и обломки клеток, суспензию центрифугировали через градиент ФБС. Жизнеспособные клетки собирали из осадка, промывали один раз ФБР и ресуспендировали в ФБР при 2.5×107/мл. Мышей-реципиентов C57/BL6 в возрасте от 5 до 8 недель подвергали летальному облучению 2 дозами по 550 Rad, с интервалом 3 часа. Мышам-реципиентам вводили внутривенно (i.v) 0.2 мл суспензии клеток (т.е. 5×106 клеток/мышь) и затем поселяли в закрытых боксах с 0.02 М неомицином в питьевой воде на 3 недели. Восстановление оценивали через 6 недель с помощью анализа периферической крови методом FACS с применением MAT, специфичных к α- и β- цепям huGMCSFR.
Лечение GM-CSF и последующий анализ Tg химерных мышей:
Tg химерным мышам в течение 4 дней дважды в день подкожно (s.c) вводили 500 нг huGM-CSF. Для анализа антагонистической активности антител группам из 5 мышей вводили выбранные дозы MAT (см. ниже) интраперитонеальным (i.p) путем за 1 день до начала лечения GM-CSF. В день 5, 0.2 мл крови брали на анализ популяций циркулирующих лейкоцитов, в частности, моноцитов крови, применяя ADVIA™ Hematology System (Bayer Diagnostics). Затем животных умерщвляли и удаляли селезенки для измерения веса.
Ингибирование TNFα и IL-6 человека, эндогенно экспрессируемых мононуклеарами периферической крови человека.
Лейкоцитарные пленки человека (кровяные сгустки человека из службы переливания крови) наносили на градиент плотности Histopaque 1.077 (Sigma, номер в каталоге 1077-1) и клетки центрифугировали при 400×g в течение 40 минут. Центрифуга останавливалась без применения тормоза. МНПК затем собирали с поверхности раздела. Клетки промывали в ФБР и осаждали при 300×g в течение 10 минут перед, а затем лизировали оставшиеся эритроциты путем ресуспендирования в 20 мл ледяной воды в течение 15 с, после чего немедленно добавляли ледяной 1.6% NaCl. Затем клетки осаждали при 1200 об/мин в течение 5 минут и ресуспендировали в 10 мл 10% ФБС/RPMI и 1% пенициллина/стрептомицина. Затем клетки разбавляли до 5×106/мл. 110 мкл клеток распределяли по лункам (5.5×105/лунку) и клеткам позволяли оседать в течение 1 часа при 37°C, 5% CO2. Следующие реагенты добавляли в качестве отдельных контролей конечной концентрации: РНА (5 мкг/мл), LPS (25 мкг/мл), GM-CSF (10 нг/мл) и изотипического контроля (50 мкг/мл). Антитело 6 добавляли до начальной конечной концентрации 50 мкг/мл, с последующим пятью разведениями. Затем планшеты инкубировали в течение 72 часов при 37°C, е% CO2. Супернатанты собирали через 72 часа и уровни рассчитывали TNFa и IL-6, применяя следующие наборы от R&D для твердофазного ИФА (hTNF-a R&D Duoset ELISA development system DY210 и hlL-6 R&D Duoset ELISA development system DY206). Твердофазный ИФА выполняли, следуя рекомендациям производителя.
Результаты для антител 9 и 11 были разными в двух одинаковых экспериментах.
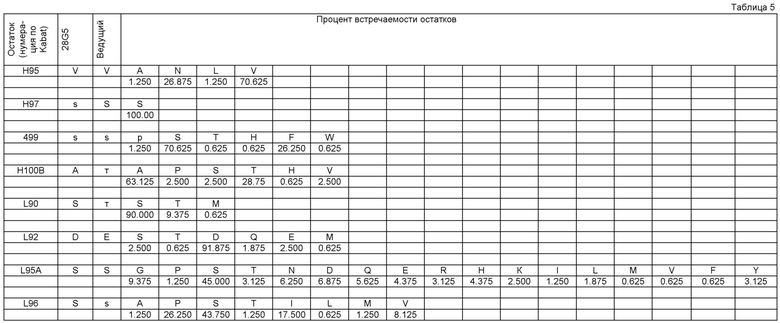
Ключ к списку последовательностей
В списке последовательностей, приведены последовательности нуклеиновых кислот и аминокислот ("PRT") для 20 клонов антител, включая родительский клон и 19 клонов из панели оптимизации. Антитела пронумерованы от АТ1 до АТ20. Родительский клон представляет собой антитело 3, представленное последовательностями SEQ ID NO: 21-30 и SEQ ID NO:211-212.
Следующий список устанавливает номера SEQ ID NO, в которых показаны последовательности указанных молекул:
(нт= последовательность нуклеотидов; ак= последовательность аминокислот
Последовательности нуклеотидов VL-домена антител с 1 по 20 не включают кодон дед, показанный на 3’-конце в последовательностях SEQ ID NO: 6,16, 26, 36, 46, 56,66,76,86,96,106,116,126, 136, 146, 156, 166, 176,186 и 196. Соответственно, последовательности аминокислот VL-домена не включают С-концевой остаток Ala (остаток 113) в последовательностях SEQ ID NO: 7, 17, 27, 37, 47, 57, 67, 77, 87, 97, 107, 117, 127, 137, 147, 157, 167, 177, 187 и 197, соответственно. Остаток А1а113 и соответствующий кодон дед не экспрессировались в антителах с 1 по 20. Сравнение написанных последовательностей с сегментами зародышевых генов, в частности, JL2, указывает на то, что остаток Ala и соответствующий кодон дед не являются частью VL-домена.
Остаток Gly в положении 112 присутствовал в экспрессируемых последовательностях scFv и IgG. Однако, этот остаток не присутствовал в последовательностях сегмента j зародышевой линии человека, которые образуют каркасный участок 4 VL-домена, например, JL2. Остаток Gly не рассматривают как часть VL домена.
Чтобы экспрессировать легкую цепь IgG, обеспечили последовательность нуклеотидов, кодирующую легкую цепь антитела, включающую первый экзон, кодирующий VL-домен, второй экзон, кодирующий CL-домен, и интрон, разделяющий первый экзон и второй экзон. В обычных обстоятельствах, интрон вырезается при сплайсинге механизмом процессинга мРНК клетки, соединяющим 3’-конец первого экзона с 5’-концом второго экзона. Таким образом, когда ДНК, имеющую указанную последовательность нуклеотидов, экспрессировали в PHK, первый и второй экзоны соединялись в результате сплайсинга. Трансляция сплайсированной PHK дает полипептид, включающий VL- и CL-домены. После сплайсинга, Gly в положении 112 кодируется последним основанием (д) последовательности каркасного участка 4 VL-домена и первыми двумя основаниями (gt) CL-домена.
Последовательности VL-доменов антител с 1 по 20 приведены в SEQ ID NO: с 186 по 246, как указано выше. Последовательности нуклеотидов VL-домена оканчиваются последним кодоном cta, и последним остатком аминокислоты Leu в соответствующих последовательностях аминокислот VL-домена.
Последовательности неэмбрионизированных VH- и VL-доменов антитела 6 показаны в SEQ ID NO: 247-250, в дополнение к последовательностям эмбрионизированных VH- и VL-доменов, показанных в SEQ ID NO: 51, 52, 56, 57, 216и217.
Ссылки
Все документы, упомянутые где-либо в данном описании, включены в данную заявку посредством ссылки.
1 Haman et al., Journal of Biological Chemistry 274 (48): 34155-34163 1999
2 Nicola NA; Wycherley A; Boyd AW; Layton JE; Gary D; Metcalf D Blood, 82(6) p!724-31 (1993)
3 Pluckthun, A. Bio/Technology 9: 545-551 (1991)
4 Chadd HE and Charaow SM (2001) Current Opinion in Biotechnology 12: 188-194
5 Andersen DC and Krummen L (2002) Current Opinion in Biotechnology 13: 117
6 Larrick JW and Thomas DW (2001) Current Opinion in Biotechnology 12: 411-418
7 Sambrook and Russell, Molecular Cloning: a Laboratory Manual: 3rd edition, 2001, Cold Spring Harbor Laboratory Press
8 Ausubel et al. eds., Short Protocols in Molecular Biology: A Compendium of Methods from Current Protocols in Molecular Biology, John Wiley & Sons, 4th edition 1999
9 Wold, et al. Multivariate data analysis in chemistry. Chemometrics -Mathematics and Statistics in Chemistry (Ed.: B. Kowalski), D. Reidel Publishing Company, Dordrecht, Holland, 1984 (ISBN 90-277-1846-6)
10 Norman et al. Applied Regression Analysis. Wiley- Intel-science; 3rd edition (April 1998) ISBN: 0471170828
11 Kaiidel, Abraham & Backer, Eric. Computer-Assisted Reasoning in Cluster Analysis. Prentice Hall PTR, (May 11, 1995), ISBN: 0133418847
12 Krzanowski, Wojtek. Principles of Multivariate Analysis: A User’s Perspective (Oxford Statistical Science Series, No 22 (Paper)). Oxford University Press; (December 2000), ISBN: 0198507089
13 Witten, lan H. & Frank, Elbe. Data Mining: Practical Machine Learning Tools and Techniques with Java Implementations. Morgan Kaufmann; (October 11, 1999), ISBN: 1558605525
14 Denisori David G. T. (Editor), Christopher C. Holmes, Bani K.
15 Ghose, Amp K. & Viswanadhan, Vellarkad N.. Combinatorial Library Design arid Evaluation Principles, Software, Tools, and Applications in Drug Discovery. ISBN: 0-8247-0487-8
16 Chothia С.efc al. Journal Molecular Biology (1992) 227, 799-817
17 AI-Lazikani, et al. Journal Molecular Biology (1997) 273 (4), 927-948
18 Chothia, et al. Science, 223,755-758 (1986)
19 Whitelegg, N.R.u. and Rees, A.R (2000). Prot. Eng., 12,815-824
20 Guex, N. arid Peitsch, M.C. Electrophoresis (1997) 18, 2714- 2723
21 Voet & Voet, Biochemistry, 2nd Edition., (Wiley) 1995.
22 Marks et al Bio/Technology, 1992, 10^779-783
23 Kay, В.К., Winter, J., and McCafferty, J. (1996) Phage Display of Peptides and Proteins: A Laboratory Manual, San Diego: Academic Press.
24 Stemmer, Nature, 1994, 370:389-391
25 Gram et al., 1992, Proc. Natl. Acad. Sci„ USA, 8_9-. 3576-35BO
26 Barbas et al., 1994, Proc. Natl. Acad. Sci., USA, 9I-.3B09- 3813
27 Sch.ier et al., 1996, J. Mol. Biol. 263:551-567
28 Ledermann J.A. et al. (1991) Int. J. Cancer 47: 659-664
29 Bagshawe K.D. et al. (1991) Antibody, Immunoconjugates and Radiopharmaceuticals 4: 915-922
30 Robinson, J. R. ed., Sustained and Controlled Release Drug Delivery Systems, Marcel Dekker, Inc., New York, 1978
31 Ritz, S. A., M. J. Cundall, et al. (2002). Am J Respir Cell Mol Biol 27(4); 428-35.
32 Ritz, S. A., M. R. Stampfli, et al. (2002). Trends Immunol 23(8): 396-402.
33 WOOI. LEY, K.L. et al. (1995). Am J Respir Cr.it Care Med, 151, 1915-24.
34 Kotsimbos, А. Т., M. Humbert, et al. (1997). J Allergy Clin Immunol 99(5): 666-72.
35 Yamashita N. et al., Cell. Immunol 2002 oct; 219(2); 92-97
36 Ohta К., et al. J Allergy Clin Immunol. 1999 Nov; 104(5); 1024-30
37 Stampfli, M. R.r R. E. Wiley, et al. (1998). J Clin Invest 102 (9): 1704-14.
38 Gates, E. С., В. U. Gajewska, et al. (2003). J Allergy Clin Immunol 111(5): 1076-86.
39 CAMPBELL, I.К., et al. (1997) Annal -Res Dis, 56, 364-368.
40 BISCHOF, R.J, D, ZAFIEOPOULOS, J.A. HAMILTON AND I.K. CAMPBELL (2000) Clin Exp Immunol, 119, 361-367.
41 Campbell, I.К., M.J. Rich, et al. (1998). J Immunol 161 (7); 3639-44.
42 Hamilton, J. A. (2002). Trends Immunol 23(8): 403-8.
43 YANG. Y.H. AND J.A. HAMILTON (2001) Arthritis Rheiimatol, 44, 111-119
44 Cook, A.D., E.L. Braine, et al. (2001). Arthritis Res 3(5); 293-8.
45 Field, M. and L. Clinton (1993). The Lancet 342: 1244.
46 Firestein GS, Alvaro-Gracia JM, Maki R. 1990 J. Immunol may 1; 144 (9): 3347-53
47 Leizer T, et al. 1990 Blood 1990 Nov 15; 76(10): 1989-96
48 Fiehii С, et al., Z Rheumatol 1992 May-Jun; 51 (3): 1221-126
49 de Vries, E.G., et al. (1991) Lancet, 338, 517-518.
50 Hazenberg BP, et al. Blood 1989 Dec; 74(8): 2769-70
51 Barnes and Hansel (2004). Lancet, 364: 985-96.
52 Barnes, P.J. (2000). Chest, 117: 103-143
53 McManus, T. E., et al (2005) American Thoracic Society Meeting, May 2005.
54 Vlahos, R., et al. (2005). "GM-CSF is a key pathogenic mediator in experimental COPD." European Respiratory Society Meeting, Sept 2005
55 McQualter JL, et al. (2001) J Exp Med. 194: 873-82
56 Bagby GC Jr, et al. 1988 J Clin Invest. 4: 1430-6.
57 Estrov Z, et al. (1986) Cancer Res. 46: 6456-61.
58 Barak Y, et al. 1981 Am J Hematol. 10: 269-75.
59 Gualtieri RJ, et al. 1988 Exp Hematol. 16:613-9.
60 SudaT.etal. 1982 Leuk Res. 6: 43-53.
61 Gualtieri RJ, et al. 19B9 Blood. 74: 2360-7
62 Largaespada DA, et al. (1996) Nat. Genet. 12: 137-43
63 Iversen, P. 0., et al. (1997). Blood 90(12): 4910-7.
64 Wang et al., 1994 Exp mol Pathol
65 Plenz et al., Art. Thrombosis and Vascular biology 1997
66 Takashi etal., 1996 Circulation 93, 1185-1193
67 Biwa T et al., JBC 1998 273: 28305-28313
68 Makheja et al., Atherosclerosis 1989, 76 155-661
69 Naito et al., 1992 J Nutri sci Vitamin 38: 255-264
70 Hayashi etal., Atherosclerosis 1991 91: 107-116
71 Villa et al 1994 J Clin Invest 93: 1243
72 Van Put DJ et al., 1995 Eur J Pharmacol 294: 753-761
73 Voisard et al., 1994 IntJ Cardiol 43: 257-267
74 Yamada et al., 1993: Artery 20: 253-267
75 Sakai et al., 1999 Arterioscler Thromb vase biol 19: 1726-1733
76 Gearing, et al. EMBO J. 8 (12): 3667-3676 (1989)
77 Crosier, К. et al. PNAS 88:7744-7740 1991
78 Raines, M. et al. PNAS 88:8203-8207 1991 Wess, L. In: BioCentury, The Bernstein Report on BioBusiness, 12(42), A1-A7, 2004
79 Wesa, L In: BioCentury, The Bernstein Report on BioBusiness, 12 (42), A1-A7, 2004
80 Haan & Maggos (2004) BioCentury, 12(5): A1-A6
81 Koideetal. (1998) Journal of Molecular Biology, 284: 1141-1151.
82 Nygren et al. (1997) Current Opinion in Structural Biology, 7: 463-469
83 Kontermann, R & Dubel, S, Antibody Engineering, Springer-Verlag New York, LLC; 2001, ISBN: 3540413545
84 Mendez, M. et al. (1997) Nature Genet, 15(2): 146-156
85 Knappik et al. J. Mol. Biol. (2000) 296, 57-86
86 Krebs et al. Journal of Immunological Methods 254 2001 67-84
87 Ward, E.S. et al., Nature 341, 544-546 (1989)
88 McCafferty et al (1990) Nature, 348, 552-554
89 Holt et al (2003) Trends in Biotechnology 21, 484-490
90 Bird et al, Science, 242, 423-426, 1988;
91 Huston et al, PNAS USA, 85, 5879-5883, 1988
92 Holliger, P. et al, Proc. Natl. Acad. Sci. USA 90 6444-6448, 1993
93 Reiter, Y. et al, Nature Biotech, 14, 1239-1245, 1996
94 Hu, S. et al, Cancer Res., 56, 3055-3061, 1996.
95 Holliger, P. and Winter G. Current Opinion Biotechnol 4, 446-449 1993 9G Ridgeway, J. В. В. etal, Protein Eng., 9, 616-621, 1996
97 Ann NYAcadSci. 1971 Dec 31; 190:382-93.
98 Rabat, E.A. et al, Sequences of Proteins of Immunological Interest. 4th Edition. US Department of Health and Human Services. 1987
99 Kabat, E.A. et al. (1991) Sequences of Proteins of Immunological Interest, 5th Edition. US Department of Health and Human Services, Public Service, NIH, Washington.
100 Persic etal 1997 Gene 187; 9-18
101 Hanes J et al (2000) Methods in Enzymology, Vol 328:24.
102 Vaughan TJ etal (1996) Nature Biotechnology Vol 14:309
103 Slootstia-LJW; Puijk-WC; Idgtvoet-GJ; Langeveld-JP; Meloen-RH (1996). Mol-Divers. 1: 87-96
104 WO01/66754 Cambridge Antibody Technology Limited; Vaughan; Wilton; Smith; Main
105 P.К. Smith, et al., Anal. Biochem. 150 (1985), pp.76-85
106 S. Mizushima and S. Nagata Nucleic Acids Research, Vol.18; No 17 1990 pp 5322
107 Clin Haematol. 1979 Juri; 8 (2): 263-85. Metcalf D.
108 Nishijima, I., T. Nakahata, et al. (1997). Blood 90(3): 1031-8.
| название | год | авторы | номер документа |
|---|---|---|---|
| СВЯЗУЮЩИЙ ЭЛЕМЕНТ ДЛЯ РЕЦЕПТОРА GM-CSF | 2007 |
|
RU2495050C2 |
| Лечение остеоартрита | 2009 |
|
RU2712273C2 |
| АНТИ-GM-CSF АНТИТЕЛА И ИХ ПРИМЕНЕНИЕ | 2006 |
|
RU2447085C2 |
| АНТИТЕЛА К ЧЕЛОВЕЧЕСКОМУ NKG2D И ИХ ПРИМЕНЕНИЯ | 2008 |
|
RU2563343C2 |
| ВЫДЕЛЕННОЕ АНТИТЕЛО ЧЕЛОВЕКА ИЛИ ЕГО АНТИГЕН-СВЯЗЫВАЮЩИЙ ФРАГМЕНТ (ВАРИАНТЫ), СПОСОБ ЛЕЧЕНИЯ ЗАБОЛЕВАНИЙ, ОПОСРЕДОВАННЫХ hTNFα С ИХ ПОСРЕДСТВОМ (ВАРИАНТЫ), ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, РЕКОМБИНАНТНОЕ АНТИТЕЛО ЧЕЛОВЕКА ИЛИ ЕГО АНТИГЕН-СВЯЗЫВАЮЩИЙ ФРАГМЕНТ, ВЫДЕЛЕННАЯ НУКЛЕИНОВАЯ КИСЛОТА (ВАРИАНТЫ) | 2005 |
|
RU2458704C9 |
| МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА, КОТОРЫЕ СВЯЗЫВАЮТСЯ С hGM-CSF, И СОДЕРЖАЩИЕ ИХ КОМПОЗИЦИИ МЕДИЦИНСКОГО НАЗНАЧЕНИЯ | 2008 |
|
RU2517596C2 |
| ANTI-CXCR4 АНТИТЕЛА И КОНЪЮГАТЫ АНТИТЕЛО-ЛЕКАРСТВО | 2014 |
|
RU2675516C2 |
| Молекулы, связывающиеся с 4-1ВВ | 2011 |
|
RU2710717C2 |
| Лечение боли | 2009 |
|
RU2630969C2 |
| ВЫДЕЛЕННОЕ АНТИТЕЛО ПРОТИВ IP-10, ИММУНОКОНЪЮГАТ И БИСПЕЦИФИЧЕСКАЯ МОЛЕКУЛА НА ЕГО ОСНОВЕ, ИХ КОМПОЗИЦИИ, СПОСОБ ЛЕЧЕНИЯ (ВАРИАНТЫ), КОДИРУЮЩАЯ МОЛЕКУЛА НУКЛЕИНОВОЙ КИСЛОТЫ, СООТВЕТСТВУЮЩИЙ ЭКСПРЕССИОННЫЙ ВЕКТОР, КЛЕТКА-ХОЗЯИН И ГИБРИДОМА | 2004 |
|
RU2486199C2 |
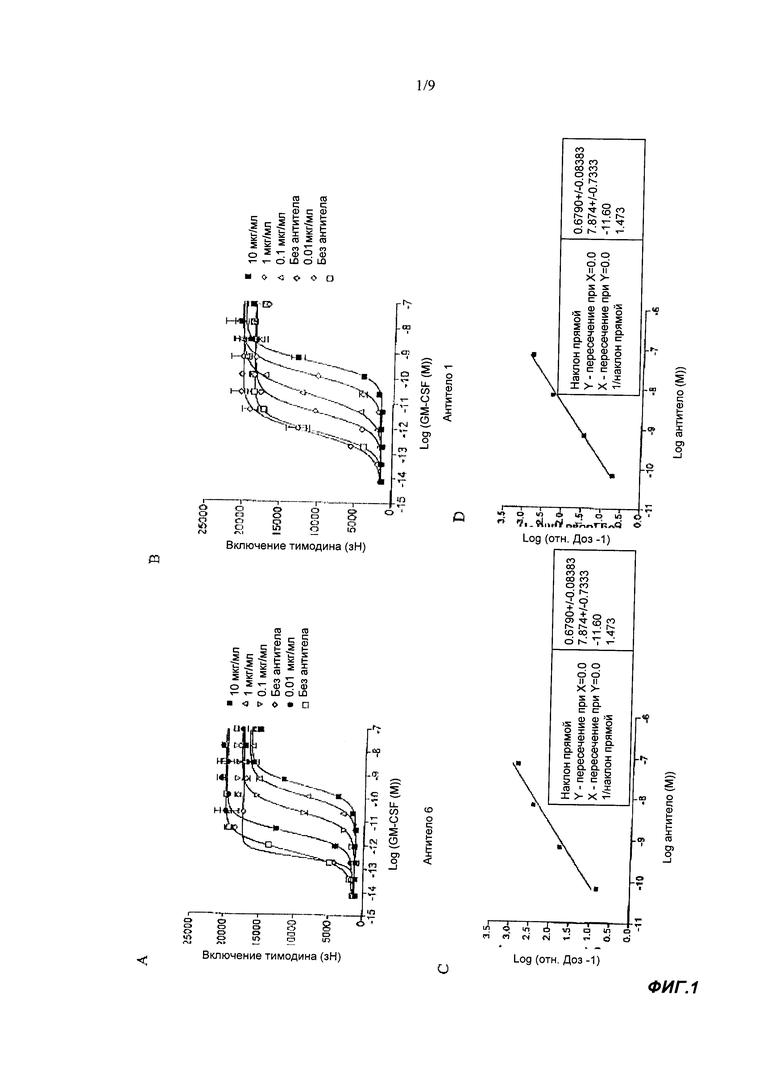
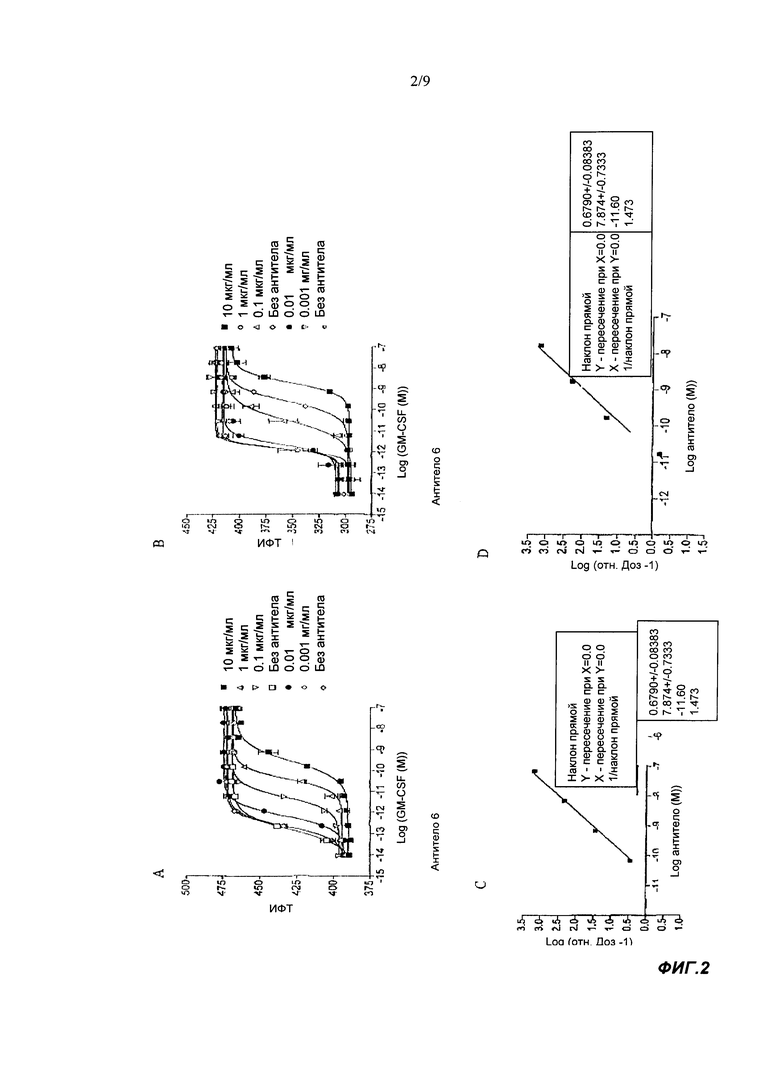
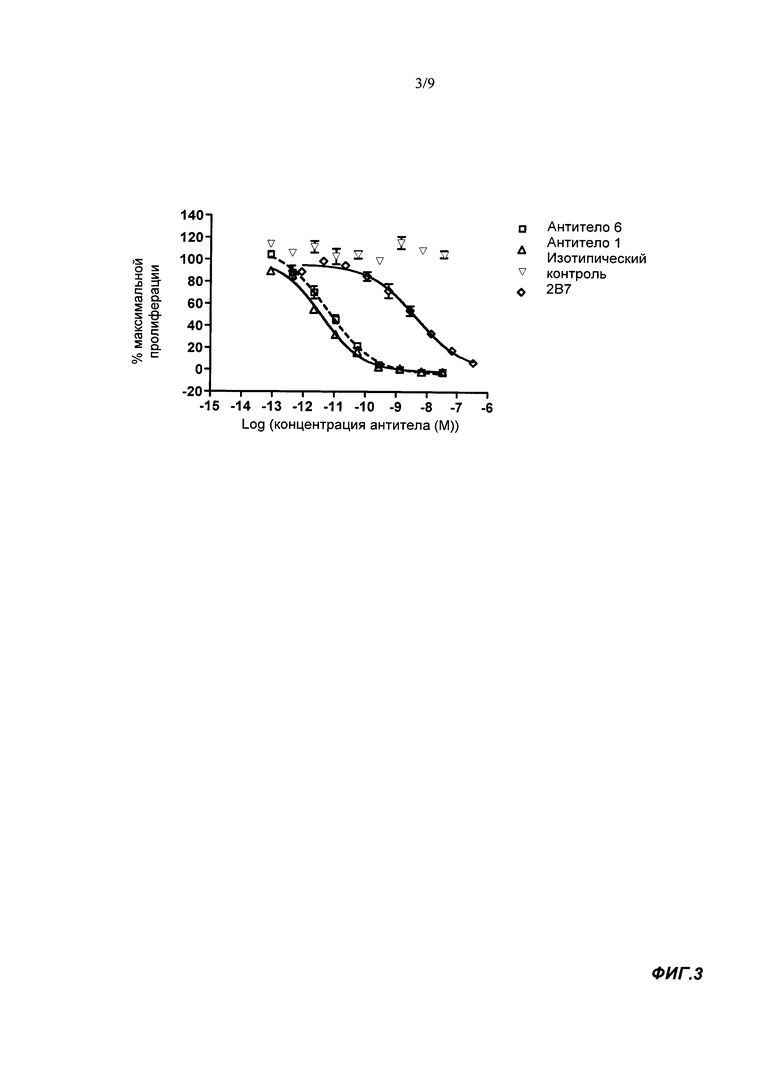
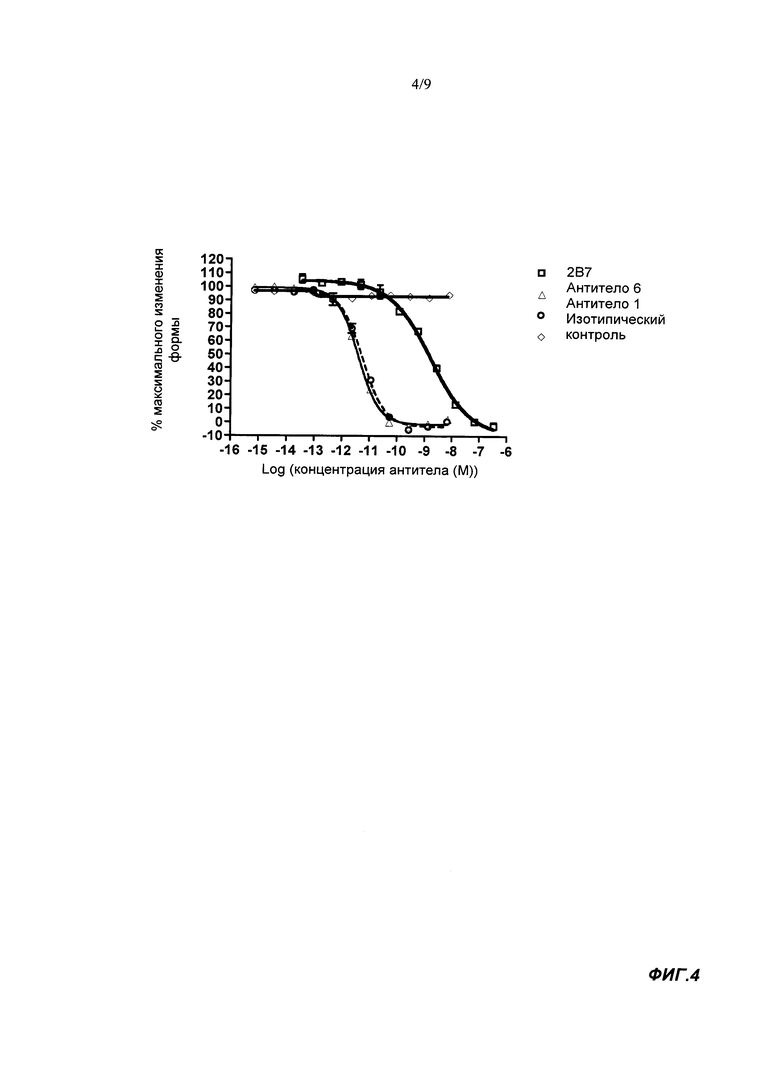
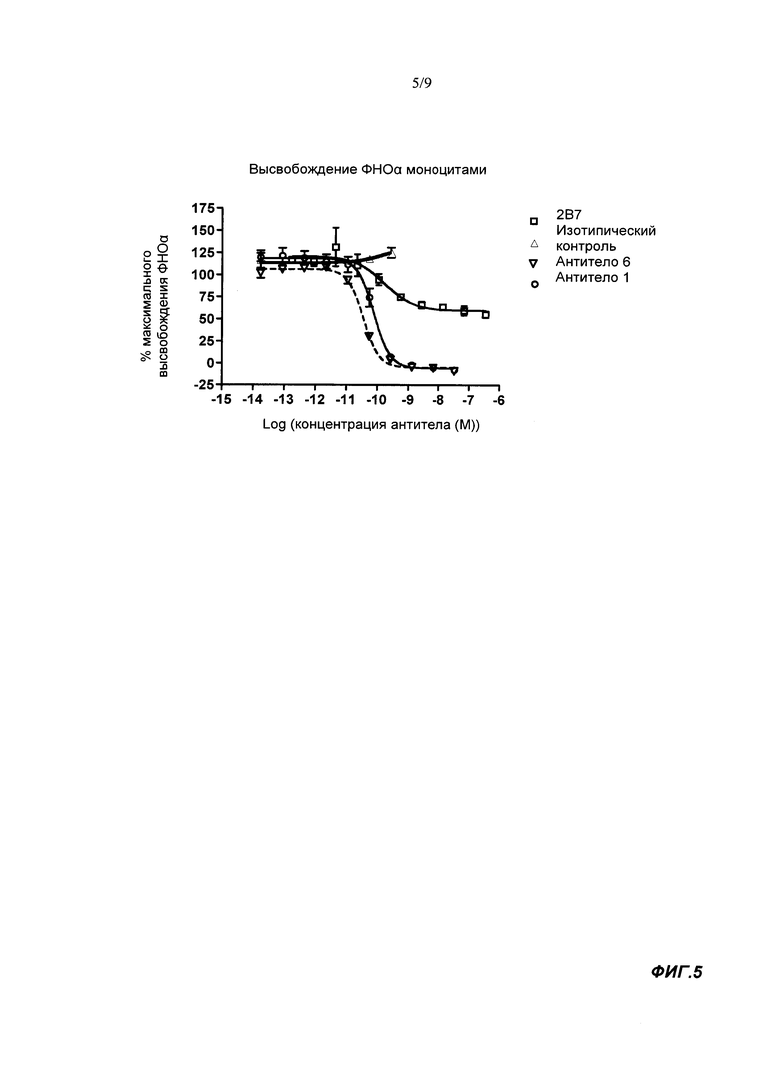
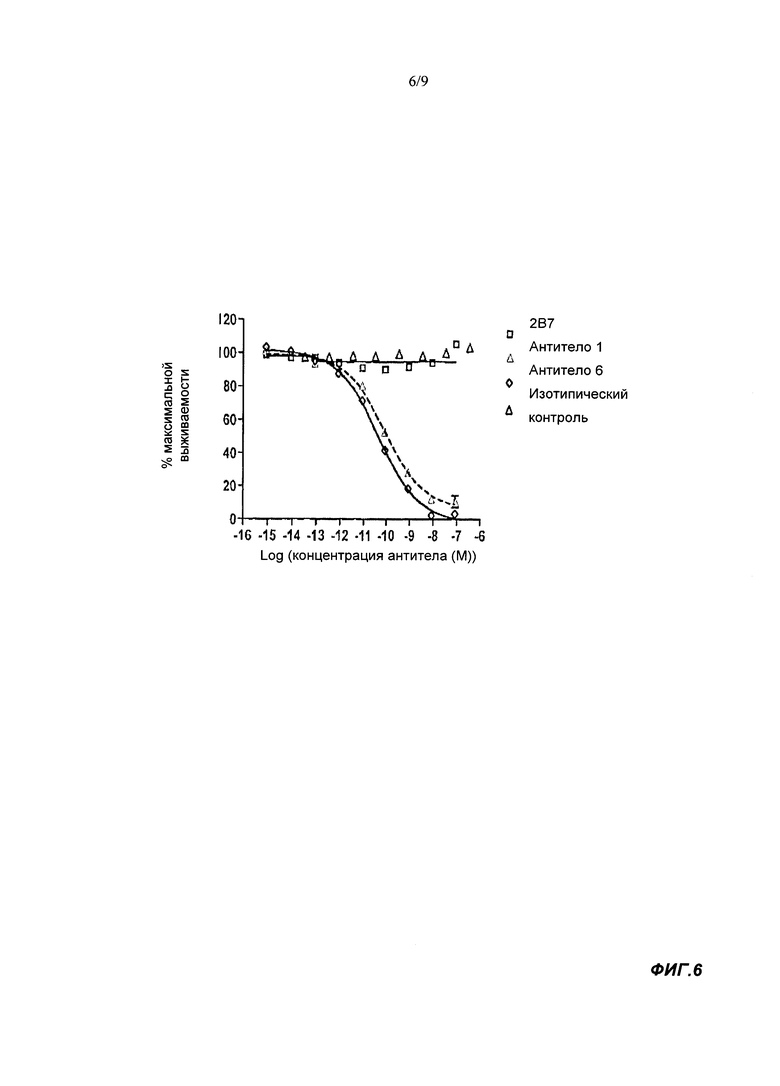
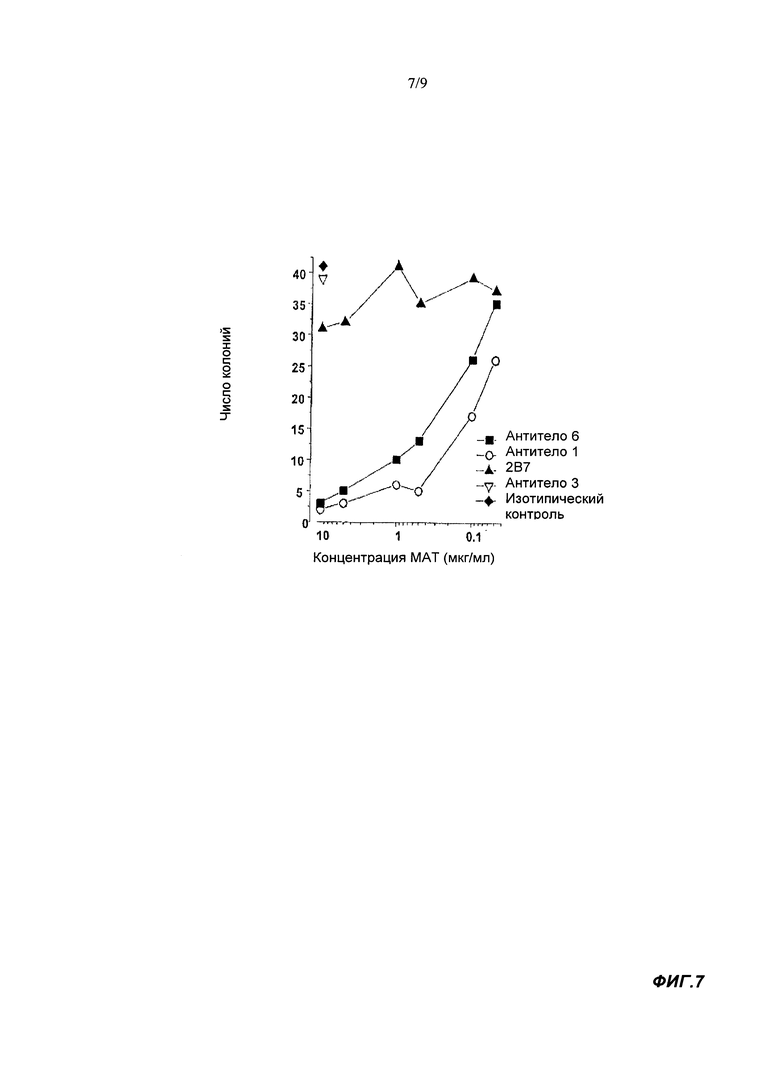
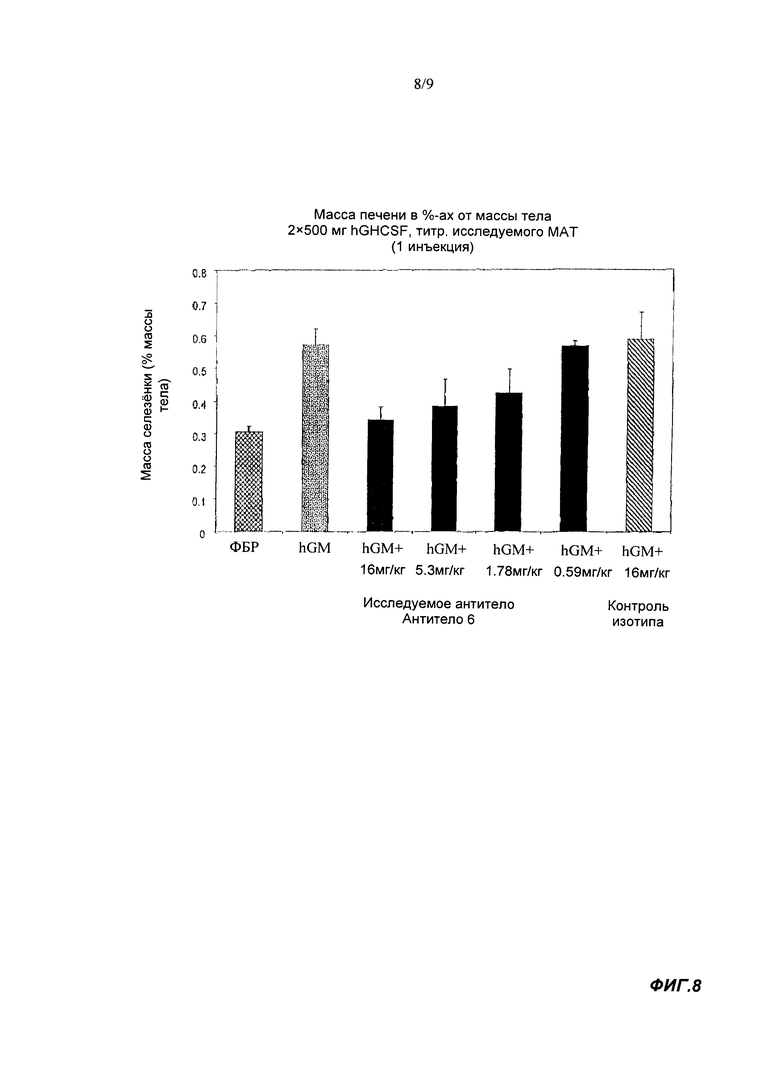
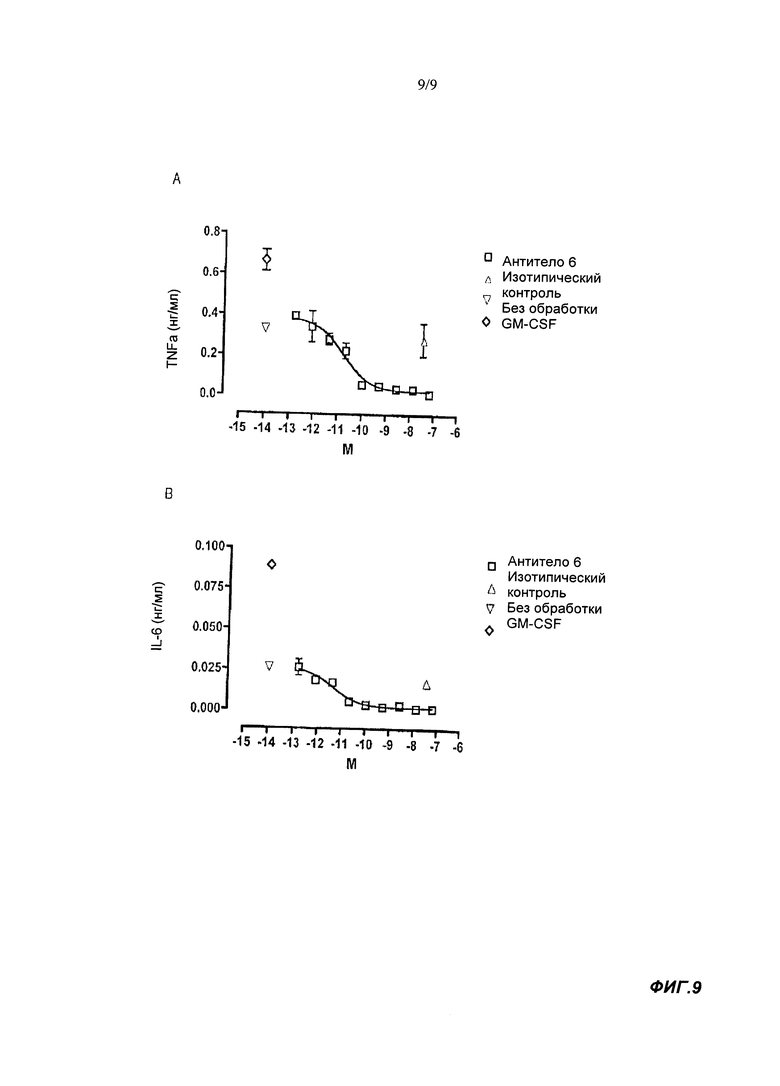
Изобретение относится к иммунологии. Предложены способ ингибирования или нейтрализации активности альфа цепи рецептора колониестимулирующего фактора гранулоцитов/макрофагов (GM-CSFRα) у пациента, включающие введение пациенту композиции, содержащей эффективное количество антитела к GM-CSFRα человека. Кроме того, рассмотрены способы лечения ревматоидного артрита, астмы, хронической обструктивной болезни легких или миелоидной лейкемии у пациента. Данное изобретение может найти применение в терапии других заболеваний, ассоциированных с GM-CSFRα. 4 н. и 8 з.п. ф-лы, 9 ил., 5 табл.
1. Способ ингибирования или нейтрализации активности альфа-цепи рецептора колониестимулирующего фактора гранулоцитов/макрофагов (GM-CSFRα) у пациента, включающий введение указанному пациенту композиции, содержащей эффективное количество антитела к GM-CSFRα человека, при этом указанное антитело:
ингибирует связывание GM-CSF с GM-CSFRα человека; и
имеет значение KD меньше чем 5 нМ при связывании GM-CSFRα человека при определении с помощью поверхностного плазмонного резонанса,
и при этом указанное антитело содержит:
набор гипервариабельных участков (CDR) VH-домена: VH CDR1, включающий последовательность SEQ ID NO: 53, VH CDR2, включающий последовательность SEQ ID NO: 54 и VHCDR3, включающий последовательность SEQ ID NO: 55, с каркасными участками VH-домена; и
набор CDR VL-домена: VL CDR1, включающий последовательность SEQ ID NO: 58, VL CDR2, включающий последовательность SEQ ID NO: 59 и VL CDR3, включающий последовательность SEQ ID NO: 60, с каркасными участками VL-домена.
2. Способ лечения ревматоидного артрита, астмы, хронической обструктивной болезни легких или миелоидной лейкемии у пациента, включающий введение указанному пациенту композиции, содержащей эффективное количество молекулы антитела, при этом указанная молекула антитела:
ингибирует связывание GM-CSF с GM-CSFRα человека;
имеет значение KD меньше чем 5 нМ при связывании GM-CSFRα человека при определении с помощью поверхностного плазмонного резонанса,
и при этом указанное антитело содержит:
набор гипервариабельных участков (CDR) VH-домена: VH CDR1, включающий последовательность SEQ ID NO: 53, VH CDR2, включающий последовательность SEQ ID NO: 54 и VHCDR3, включающий последовательность SEQ ID NO: 55, с каркасными участками VH-домена; и
набор CDR VL-домена: VL CDR1, включающий последовательность SEQ ID NO: 58, VL CDR2, включающий последовательность SEQ ID NO: 59 и VL CDR3, включающий последовательность SEQ ID NO: 60 с каркасными участками VL-домена.
3. Способ ингибирования или нейтрализации активности альфа цепи рецептора колониестимулирующего фактора гранулоцитов/макрофагов (GM-CSFRα) у пациента, включающий введение указанному пациенту композиции, содержащей эффективное количество молекулы антитела, при этом указанная молекула антитела содержит VH-домен и VL-домен, причем VH-домен содержит гипервариабельные участки (CDR) тяжелой цепи VH CDR1, VH CDR2 и VH CDR3 и в VL домен содержит гипервариабельные участки легкой цепи VL CDR1, VL CDR2 и VL CDR3, при этом указанные участки CDR имеют следующие аминокислотные последовательности:
VH CDR1 - SEQ ID NO: 53;
VH CDR2 - SEQ ID NO: 54;
VH CDR3 - SEQ ID NO: 55;
VL CDR1 - SEQ ID NO: 58;
VL CDR2 - SEQ ID NO: 59; и
VL CDR3 - SEQ ID NO: 60.
4. Способ лечения ревматоидного артрита, астмы, хронической обструктивной болезни легких или миелоидной лейкемии у пациента, включающий введение указанному пациенту композиции, содержащей эффективное количество молекулы антитела, при этом указанная молекула антитела содержит VH-домен и VL-домен, причем VH-домен содержит гипервариабельные участки (CDR) тяжелой цепи VH CDR1, VH CDR2 и VH CDR3 и VL-домен содержит гипервариабельные участки легкой цепи VL CDR1, VL CDR2 и VL CDR3, при этом указанные участки CDR имеют следующие аминокислотные последовательности:
VH CDR1 - SEQ ID NO: 53;
VH CDR2 - SEQ ID NO: 54;
VH CDR3 - SEQ ID NO: 55;
VL CDR1 - SEQ ID NO: 58;
VL CDR2 - SEQ ID NO: 59; и
VL CDR3 - SEQ ID NO: 60.
5. Способ по п.3 или 4, отличающийся тем, что указанная молекула антитела представляет собой молекулу гуманизированного антитела или антитела человека.
6. Способ по п.3 или 4, отличающийся тем, что указанная молекула антитела содержит VH-домен антитела с последовательностью аминокислот SEQ ID NO: 52.
7. Способ по п.3 или 4, отличающийся тем, что указанная молекула антитела содержит VL-домен антитела с последовательностью аминокислот SEQ ID NO: 218.
8. Способ по п.3 или 4, отличающийся тем, что указанная молекула антитела представляет собой lgG4.
9. Способ по п.8, отличающийся тем, что указанная молекула антитела представляет собой молекулу антитела lgG4 человека, содержащую VH-домен, который имеет последовательность SEQ ID NO: 52 и VL-домен, который имеет последовательность SEQ ID NO: 218.
10. Способ по любому из пп.1-3, согласно которому указанную композицию вводят в комбинации или в дополнение к одному или более из следующего: НПВП, кортикостероиды и модифицирующие заболевание противоревматоидные средства (DMARD).
11. Способ по п.10, согласно которому указанную композицию вводят в комбинации с одним или более из следующего: ЦОГ-2 ингибиторы, преднизон, Хумира (адалимумаб), метотрексат, Арава, Энбрел, (Этанерцепт), Ремикад (Инфликсимаб), Кинерет (Анакинра), Ритуксан (Ритуксимаб), Оренция (абатацепт), соли золота, антималярийные средства, сульфасалазин, d-пенициламид, циклоспорин А, диклофенак, циклофосфамид и азатиоприн.
12. Способ по п.11, согласно которому указанную композицию вводят в комбинации с метотрексатом.
| NICOLA N.A | |||
| et al., "Neutralizing and nonneutralizing monoclonal antibodies to the human granulocyte-macrophage colony-stimulating factor receptor alpha-chain." Blood, 1993, 82(6): 1724-1731 | |||
| Экономайзер | 0 |
|
SU94A1 |
| Экономайзер | 0 |
|
SU94A1 |
| Бесколесный шариковый ход для железнодорожных вагонов | 1917 |
|
SU97A1 |
| WOODCOCK J.M | |||
| et al., "The Human Granulocyte-Macrophage Colony-Stimulating Factor (GM-CSF) Receptor Exists as a Preformed Receptor Complex That Can be Activated by GM-CSL, Interleukin-3, or Interleukin-5", Blood, 1997, 90(8): 3005-3017 | |||
| ЛАХТИНА О.Е | |||
| и др., "Определение гранулоцитарно-макрофагального колониестимулирующего фактора человека с помощью моноклональных антител." Биоорганическая Химия, 1999, 25(9):673-678. | |||

