Изобретение относится к областям генной и белковой инженерии, молекулярной биологии и биотехнологии, в частности изобретение направлено на средства оценки межмолекулярных взаимодействий биомолекул, основанные на комплементации белковых фрагментов репортера.
Данный подход использует свойства некоторых белков восстанавливать свою активность при сборке его нативной структуры из определенных независимых фрагментов во время стерического сближения (комплементация белковых фрагментов). Такие комплементирующие фрагменты белка-репортера, по отдельности генетически слитые в единой рамке считывания или присоединенные иным образом к исследуемым биомолекулам, позволяют детектировать взаимодействие этих биомолекул в составе гибридного белка по восстановлению активности белка-репортера. Стерическое сближение исследуемых биомолекул во время взаимодействия в составе гибридных белков ведет соответственно к реассоциации комплементирующих частей репортера с формированием его функционально активной структуры (фиг.1). По регистрируемой активности репортера можно судить о взаимодействии двух исследуемых биомолекул, каждая из которых соединена с отдельным фрагментом репортера.
Известны различные варианты зеленых флуоресцентных белков (ЗФБ) и их мутантов, способные давать комплементирующие фрагменты [US 7176287, C07K 19/00, Feb. 2007], [US 7166424, C12Q 1/25, Jan. 2007 (прототип)], которые формируют при стерическом сближении функционально активный ЗФБ-репортер. Полученные комплементирующие фрагменты ЗФБ были использованы для анализа межмолекулярных взаимодействий.
Основными недостатками таких репортеров являются довольно высокий фон, присущий флуоресцентным репортерам, маленький линейный диапазон измерений, что делает затруднительным количественную оценку взаимодействий, и фотобличинг (фотовыцветание).
Основной задачей, на решение которой направлены заявляемые изобретения, является расширение арсенала средств оценки межмолекулярных взаимодействий в молекулярной биологии и биотехнологии.
Единым техническим результатом, достигаемым при осуществлении заявленной группы изобретений, является получение измеряемой биолюминесцентной активности при стерическом сближении заявляемых комплементирующих фрагментов молекулы фотопротеина класса Hydrozoa с формированием активного репортера, биолюминесцентная активность которого используется для анализа белок-белковых взаимодействий.
Указанный технический результат достигается сочетанием комплементирующих фрагментов молекулы фотопротеина из Hydrozoa, полученной путем деления на две части молекулы фотопротеина в области наружной петли между 4-й и 5-й α-спиралями глобулы белка, комплементирующие фрагменты которой при ассоциации генерирует детектируемый биолюминесцентный сигнал.
Технический результат достигается также тем, что применение композиции комплементирующих фрагментов молекулы фотопротеина из Hydrozoa для анализа межмолекулярных взаимодействий включает генно-инженерное соединение комплементирующих фрагментов с исследуемыми полипептидами, с образованием первого и второго гибридных белков анализируемой пары, при этом взаимодействие исследуемых полипептидов приводит к стерическому сближению и сборке комплементирующих фрагментов в функционально активный фотопротеин, при этом данное взаимодействие регистрируют по восстановлению биолюминесцентной активности фотопротеина. И тем, что первый и второй гибридные белки анализируемой пары получены генно-инженерным слиянием фотопротеинового фрагмента с исследуемым полипептидом через гибкий гидрофильный мостик, при этом наибольшая эффективность конструкции достигается при использовании последовательности из 14 аминокислотных остатков (Seq.9).
Настоящее изобретение предлагает в качестве репортеров фрагментированные специальным образом фотопротеины класса Hydrozoa. Биолюминесцентные свойства фотопротеинов Hydrozoa позволяют широко использовать их в качестве репортерных белков в различных биоаналитических приложениях. Фотопротеин Hydrozoa представляет собой устойчивый фермент-субстратный комплекс из белковой части (апофотопротеин), связанных молекул кислорода и субстрата целентеразина. Биолюминесцентная реакция запускается ионами Са2+ и представляет собой реакцию внутримолекулярного окисления целентеразина с испусканием кванта света в голубой области. Большой квантовый выход реакции, простота ее запуска, легкость и неинвазивность регистрации квантов, отсутствие токсичности для живой клетки, широкий динамический диапазон, сопоставимый с радиоизотопной меткой, обусловливают использование фотопротеинов в качестве чувствительного биолюминесцентного репортера, в том числе в живых клетках.
Основным преимуществом предлагаемых репортеров для анализа на основе комплементации белковых фрагментов по сравнению с выбранным прототипом являются преимущества биолюминесцентного репортера перед флуоресцентным - высокая чувствительность, сопоставимая с радиоизотопной меткой, широкий линейный диапазон измерений, и практически полное отсутствие фона в случае фотопротеинов, поскольку биолюминесцентная реакция запускается ионами Са2+.
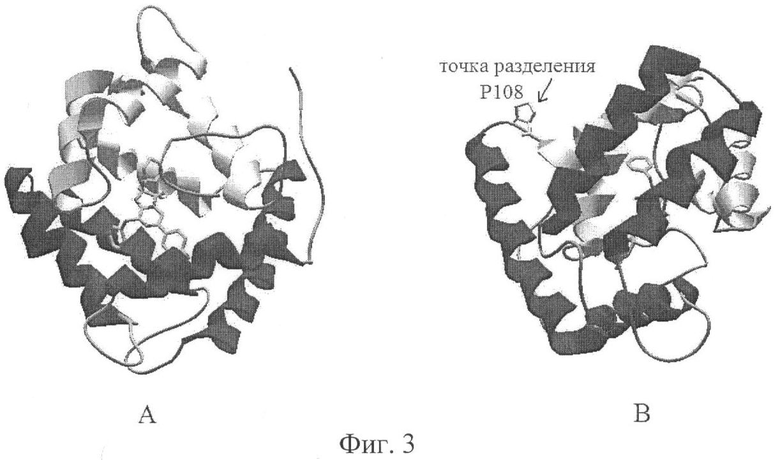
Установление трехмерных структур для ряда фотопротеинов класса Hydrozoa: акворин (PDB код 1EJ3) и клитин (PDB код) из медуз Aequorea и Clytia, обелин из гидроида Obelia longissima (PDB код 1QV0) и Obelia geniculata (PDB код 1JF0) выявило их существенное структурное подобие, несмотря на невысокую гомологию первичных последовательностей (см. выравнивание последовательностей на фиг.2). Пространственная структура фотопротеина представляет собой компактную глобулярную структуру, формируемую двумя структурными доменами из четырех α-спиралей в виде чашек, соединенных краями (фиг.3). Результатом изобретения является получение комплементирующих фрагментов фотопротеина, способных восстанавляивать активность энзима из двух синтезируемых по отдельности последовательностей после реассоциации, путем разделения последовательности фотопротеина на 2 фрагмента в области наружной петли между 4-й и 5-й α-спиралями глобулы белка (фиг.2 и 3). Две таких половинки фотопротеина формируют функциональную структуру при стерическом сближении, способную после инкубации с субстратом целентеразином образовывать стабильный фермент-субстратный компелкс, что сопровождается восстановлением биолюминесцентной активности. Место разделения последовательностей указано стрелочкой и косой чертой на фиг.2. Последовательности комплементирующих фрагментов фотопротеинов описаны в Seq №1-№8, где последовательности Seq. №1 и Seq. №2 соответствуют комплементирующим первому и второму фрагментам обелина из Obelia longissima, последовательности Seq №2 и Seq №3 соответствуют комплементирующим первому и второму фрагментам обелина из Obelia geniculata, последовательности Seq. №5 и Seq №6 соответствуют комплементирующим первому и второму фрагментам акворина, последовательности Seq №7 и Seq №8 соответствуют комплементирующим первому и второму фрагментам клитина.
Подобные пары комплементирующих фрагментов могут быть получены данным способом из других гомологичных фотопротеинов, таким образом изобретение обеспечивает способ конструирования фрагментов биолюминесцентных фотопротеинов из Hydrozoa для анализа на основе комплементации белковых фрагментов.
Изобретение обеспечивает также схему конструирования гибридных белков (фьюжинов) между комплементирующими фрагментами и исследуемыми полипептидами (пример 2). Каждый комплементирующий фрагмент фотопротеина генно-инженерно слит через гибкий гидрофильный мостик-линкер, обеспечивающий стерическую свободу соединяемых компартментов, с отдельным анализируемым полипептидом, образуя первый и второй гибридные белки анализируемой пары. Взаимодействие исследуемых полипептидов приводит к стерическому сближению связанных с ними фрагментов фотопротеина, которые реассоциируют с восстановлением детектируемой биолюминесценции.
Также обеспечивается гибкий гидрофильный мостик-линкер из 14 аминокислотных остатков (Seq 9), соединяющий исследуемый полипептид и один из комплементирующих фрагментов фотопротеина. Присутствие гидрофильного линкера такой длины обеспечивает возможность беспрепятственной сборки активного фотопротеина из его комплементирующих фрагментов вне зависимости от размеров присоединенных молекулярных доменов. В отличие от прототипа, где для присоединения комплементирующих фрагментов зеленого флуресцентного белка используется линкер (GGGGS)2 [Sec.19, US 7166424, C12Q 1/25, Jan. 2007], структура линкера по изобретению является более длинной и протеазоустойчивой, что повышает чувствительность анализа, особенно в живых клетках.
Результатом изобретения также является способ использования полученных комплементирующих фрагментов фотопротеинов для обнаружения молекулярных взаимодействий, где реассоциация комплементирующих вышеупомянутых фрагментов запускается молекулярными доменами, слитыми с каждым фрагментом, при этом реассоциация фрагментов идет независимо от других молекулярных процессов и регистрируется посредством восстановления биолюминесцентной активности фотопротеина. Использование в данном способе комплементирующих фрагментов нового репортера (фотопротеины) в отличие от прототипа, где используются комплементирующие фрагменты зеленого флуоресцентного белка и его аналоги, внедряет в анализ на основе комплементации белковых фрагментов все преимущества биолюминесцентного анализа.
Таким образом, перечисленные выше отличительные от прототипа признаки позволяют сделать вывод о соответствии заявляемых технических решений критерию «новизна».
Признаки, отличающие заявляемые технические решения от прототипа, не выявлены в других технических решениях и, следовательно, обеспечивают заявляемым решениям соответствие критерию «изобретательский уровень».
В настоящей заявке на выдачу патента соблюдено требование единства изобретения, поскольку заявлены композиция и ее применение. Заявленные изобретения решают одну и ту же задачу - расширение арсенала средств оценки межмолекулярных взаимодействий в молекулярной биологии и биотехнологии за счет одного и того же технического результата при осуществлении изобретений - получение специфических комплементирующих фрагментов из фотопротеинов класса Hydrozoa, которые способны восстанавливать биолюминесцентную активность при стерическом сближении и их использование для анализа белок-белковых взаимодействий.
Сущность изобретения поясняется с помощью графических материалов.
На фиг.1 представлена стратегия анализа на основе комплементации белковых фрагментов биолюминесцентного репортера. Реконструкция функциональной молекулы фотопротеина (круглая внизу) происходит при стерическом сближении его комплементирующих фрагментов. При этом реассоциации фрагментов репортера запускается сближением при взаимодействии исследуемых полипептидов А и В (вверху).
На фиг.2 представлен элаймент (выравнивание) аминокислотных последовательностей некоторых фотопротеинов класса Hydrozoa: обелина из гидроидов Obelia longissima (PDB код 1QVO) и Obelia geniculata, акворина (PDB код 1EJ3) и клитина из медуз Aequorea и Clytia. В последовательности затемнены α-спиральные участки структуры по указанным PDB-структурам. Точками обозначены одинаковые аминокислотные остатки в одинаковых положениях. Стрелкой и косыми чертами обозначено место разделения последовательностей на комплементирующие фрагменты в области наружной петли между 4-й и 5-й α-спиралями глобулы белка.
На фиг.3 представлена пространственная структура Са2+-регулируемого фотопротеина обелина из Obelia longissima (PDB 1QV1) с различных сторон (А и Б).
Глобулярная структура молекулы формируется двумя доменами из α-спиралей, которые могут быть представлены в форме двух чашек (показаны темным и светлым), соединенных краями друг с другом. Показана точка разделения молекулы обелина (Р108/Т109) на 2 комплементирующих фрагмента, которые формируют нативную структуру активного фермента при стерическом сближении.
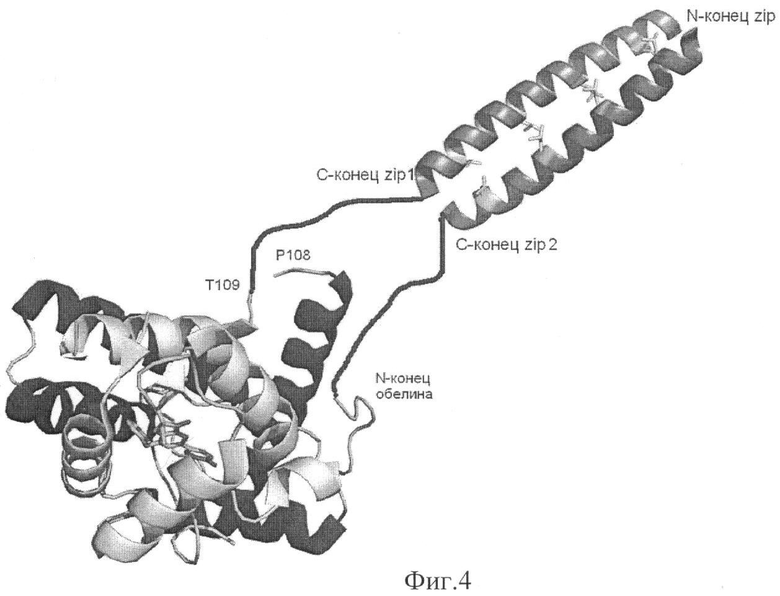
На фиг.4 представлена структурная схема взаимодействия комплементирующих фрагментов обелина, направляемого лейциновым зиппером (пропорции структур приблизительные). Между взаимодействующими доменами гибкий мостик из 14 аминокислотных остатков.
Описание представленных аминокислотных последовательностей
Последовательность №1 представляет первый (с 1-й по 108-й аминокислотный остаток) из пары комплементирующих фрагментов обелина из Obelia longissima.
Последовательность №2 представляет второй (со 109-й по 195-й аминокислотный остаток) из пары комплементирующих фрагментов обелина из Obelia longissima.
Последовательность №3 представляет первый из пары комплементирующих фрагментов обелина из Obelia geniculata.
Последовательность №4 представляет второй из пары комплементирующих фрагментов обелина из Obelia geniculata.
Последовательность №5 представляет первый из пары комплементирующих фрагментов акворина из Aequorea victoria.
Последовательность №6 представляет второй из пары комплементирующих фрагментов акворина из Aequorea victoria.
Последовательность №7 представляет первый из пары комплементирующих фрагментов клитина из Clytia gregaria.
Последовательность №8 представляет второй из пары комплементирующих фрагментов клитина из Clytia gregaria.
Последовательность №9 представляет гибкий гидрофильный мостик-линкер из 14 аминикислотных остатков для присоединения исследуемых полипептидов к комплементирующим фрагментам.
Изобретение иллюстрируется следующими примерами.
Пример 1. Получение первого и второго комплементирующих фрагментов фотопротеина.
Фотопротеин для получения первого и второго комплементирующих фрагментов разделяется на две части в области наружной петли между 4-й и 5-й α-спиралями глобулы белка (фиг.2 и 3). Такая точка (фиг.2) определена по имеющимся пространственным структурам 4-х фотопротеинов и для других гомологичных фотопротеинов, в случае отсутствия установленной трехмерной структуры, также может быть установлена по элайменту (выравниванию) последовательностей.
ДНК-последовательности, кодирующие комплементирующие фрагменты фотопротеина, могут быть получены либо сборкой гена из синтетических олигонуклеотидов, либо синтезом полимеразной цепной реакцией (ПЦР) с праймерами, комплементарными соответствующим концам каждого фрагмента, при использовании природной кДНК-матрицы гена фотопротеина. Природная матрица кДНК может быть либо выделена из природного источника (ткани организма), либо это клонированный кДНК ген фотопротеина.
Получение указанных первого и второго комплементирующих фрагментов фотопротеина показано на примере обелина из Obelia longissima. ДНК-последовательности, кодирующие оба фрагмента обелина получены ПЦР на матрице полноразмерного клонированного кДНК гена обелина с использованием праймеров, комплементарных концам каждого фрагмента. Одновременно ПЦР-праймерами были введены Nde I и Xho I сайты, в обелиновые фрагменты OL1 и OL2, а также стоп-сайт ТАА в OL1 для последующего клонирования в экспрессионную кассету.
Первый фрагмент OL1:
прямой праймер: 5'-GGCTTcatatgTCTTCAAAATACGCAG-3',
обратный: 5'-AAGctcgagAGGTTCGTTTCTTGCCC-3'.
Второй фрагмент OL2:
прямой праймер: 5'-GGCTTcatatgGGCACTCTCATTCGTGAATGG-3',
обратный: 5'-AAGctcgagTTAAGGTTCGTTTCTTGCCC-3', где маленькими буквами обозначены введенные рестриктные сайты.
Для прямой бактериальной экспрессии, после очистки и рестрикции соответствующими рестриктазами, синтезированные ПЦР ДНК-последовательности фрагментов были клонированы в Nde I и Xho I сайты экспрессионного вектора pET22b + (Novagen).
Первый фрагмент OL1, подобно полноразмерному обелину, синтезировался в больших количествах (~70% от всех клеточных белков) в виде нерастворимых телец включения, откуда может легко выделен в больших количествах промыванием телец включения и хроматографией. Второй фрагмент синтезировался в сравнительно небольших количествах в растворимой форме, поэтому для его получения в чистом виде был введен отщепляемый полигистидин His 10 для аффинной очистки. Соответствующая конструкция получена путем переклонирования по сайтам Nde I и Xho I в pET19b + (Novagen). Полученные чистые комплементирующие фрагменты могут быть использованы для конъюгации in vitro с исследуемыми биомолекулами для исследования межмолекулярных взаимодейстсвий в различных вариантах анализа.
Пример 2. Введение гибкого гидрофильного линкера между комплементирующим фрагментом и анализируемым полипептидом и оптимизация его структуры.
Для демонстрации свойств полученных комплементирующих фрагментов в исследованиях белок-белковых взаимодействий были получены парные гибридные белки путем слияния соответствующих ДНК-последовательностей в единой рамке считывания. Каждый из гибридных белков включал один исследуемый полипептид, соединенный с одним из комплементирующих фрагментов репортера через специальную последовательность, структурно представляющую собой гибкий гидрофильный линкер.
Наличие гибкого гидрофильного линкера между структурными компартментами гибридного белка в данном случае, состоящего из анализируемого полипептида и одним из пары комплементирующих фрагментов репортера, является необходимым условием правильного сворачивания гибридного белка (фолдинга) и его функционирования, Такой линкер также обеспечивает необходимые степени свободы (пространственную свободу) для соединенных молекулярных компартментов при их независимом взаимодействии с другими молекулами. Общеизвестно, что максимальную гибкость (степени свободы) обеспечивают полиглициновые последовательности, максимальную гибкость плюс гидрофильность обеспечивают полисериновые последовательности, или их сочетания.
Например, для соединения исследуемого полипептида (например, С-концом Zip-фрагмента) и одного из комплементирующих фрагментов фотопротеина (в примере - фрагменты обелина) были использованы следующие варианты последовательности гибких гидрофильных мостик-линкеров для получения гибридных белков:
короткий GSGSGH, длиной 6 аминокислотных остатков
средний GSTPATTPGTTPGH, длиной 14 аминокислотных остатков,
длинный GSGSGGSGSGGSGSGGSGSGGSGSGSGH, длиной 28 аминокислотных остатков.
Для создания экспрессионных конструкций, последовательности, кодирующий гибкие гидрофильные линкеры были собраны из синтетических олигонуклеотидов. В 5'-конец каждого линкера введен сайт BamHI для соединения с последовательностью, кодирующей исследуемый полипептид (например, с С-концом Zip-фрагмента), в 3'-конец введен сайт NdeI для соединения с одним из комплементирующих фрагментов репортера (например, с N-концом OL1 обелина).
Анализ эффективности восстановления биолюминесцентной активности на примере комплементационных фрагментов обелина при принудительном стерическом сближении, направляемом взаимодействующими фрагментами гибридных белков, проведенный на грубых лизатах бактериальных клеток E.coli, экспрессирующих данные белки, показал, что применение короткого варианта гидрофильного мостика GSGSGH, так же как и самого длинного в 28 аминокислотных остатка, характеризовалось более низким восстановлением биолюминесцентного сигнала - приблизительно на 50% меньше по сравнению с другим линкером GSTPATTPGTTPGH. Дальнейший анализ, проведенный на частично очищенных препаратах гибридных белков, показал, что если низкое восстановление биолюминесцентного сигнала для гибридных белков с коротким линкером связано скорее всего с недостаточной длиной линкера, то для длинного в 28 аминокислотных остатков четко связано с частичной протеолитической деградацией гибридного белка, содержащего такой линкер, а также, видимо, с его общей нестабильностью. Кроме того, малый уровень экспрессии гибридных белков, соединенных самым коротким и самым длинным линкерами, также свидетельствовал об их неустойчивости к действию клеточных протеаз.
Таким образом, наиболее функциональным гибким гидрофильным мостиком для конструирования гибридных белков, состоящих из исследуемых полипептидов и комплементирующих фрагментов фотопротеинов, является вариант линкерной последовательности из 14 аминикислотных остатков: GSTPATTPGTTPGH (Seq. 9), спроектированный с учетом устойчивости к действию протеаз. Такой мостик обеспечивает сборку комплементирующих фрагментов в функционально активный фотопротеин, стерически независимо от структур и размеров присоединенных исследуемых полипептидов. Кроме того, данный линкер устойчив к действию клеточных протеаз, о чем свидетельствует высокий уровень экспрессии гибридных белков в стабильном растворимом виде, полученный в бактериальных клетках.
Пример 3. Конструирование гибридных белков из комплементирующих фрагментов фотопротеина и исследуемых полипептидов для анализа межмолекулярных взаимодействий при использовании лейцинового зиппера в качестве сближающего агента.
Для демонстрации свойств полученных комплементирующих фрагментов обелина в исследованиях белок-белковых взаимодействий была использована последовательность лейцинового зиппера дрожжевого белка GCN4/YEL009C в 31 аминокислотный остаток (далее Zip):
MGRMKQLEDKVEELLSKNYHLENEVARLKKLVGER.
Небольшой простой структурный мотив «лейциновый зиппер» (фиг.4) обеспечивает белок-белковое связывание в ряде онкогенов, факторах транскрипции, ДНК-связывающих белках и других белках, регулирующих генную экспрессию. С помощью лейцинового зиппера белки формируют гомо- или гетеродимеры. В основном взаимодействие происходит за счет гидрофобного слипания лейциновых остатков, находящихся в каждом седьмом положении каждой из α-спиралей, существуют и другие варианты взаимодействий. В данном примере лейциновый зиппер белка GCN4 (Zip или Z) использован как димеризационный фрагмент.
ДНК, кодирующая Zip-фрагмент, синтезирована в ходе двух раундов гнездовой ПНР при использовании хромосомной ДНК обычных пекарских дрожжей. Синтез проведен сначала праймерами, комплементарными началу и концу гена белка GCN4, затем во втором раунде, праймерами, комплементарными началу и концу Zip-фрагмента. В синтезируемую ДНК Zip-фрагмента для последующей сборки гибридного белка Zip-OL1 и Zip-OL2 были введены праймерами перекрывающиеся сайты KpnI/NcoI в 5'-конец (для большей универсальности) и BamHI в 3'-конец (схема на фиг.4).
Далее всю экспрессионную конструкцию Zip-линкер плюс комплементирующий фрагмент собирали и клонировали либо в NcoI-XhoI сайты экспрессионного вектора pET19b (Novagen) для бактериальной экспрессии, либо в KpnI-XhoI сайты pcDNA3.1(+) (Invitrogen) для эукариотической экспрессии. Такая последовательность фрагментов (Zip, слитый в рамке с N-концами обелиновых фрагментов) в гибридном белке была выбрана, поскольку показано, что генноинженерное слияние дополнительных последовательностей с N-концом обелина не влияет существенным образом на его биолюминесцентную активность. И наоборот, для родственного обелину фотопротеина акворина показано, что модификации С-конца могут привести к полной инактивации белка.
Пример 4. Определение межмолекулярных взаимодействий in vitro no восстановлению биолюминесценной активности фотопротеина при использовании лейцинового зиппера в качестве сближающего агента.
Для детекции межмолекулярных взаимодействий in vitro по восстановлению биолюминесценной активности комплементирующих фрагментов обелина, направляемой димеризацией Zip-фрагментов (далее Z) при взаимодействии полученных гибридных белков Z-OL1 и Z-OL2, была проведена очистка гибридных белков, экспрессированных в клетках E.coli. Индукция синтеза Z-OL1 и Z-OL2 в E.coli с полученных в примере 2 конструкций приводит к появлению рекомбинантных гибридных белков Z-OL1 и Z-OL2 в виде доминантных полос на электрофорезном геле, при этом основная часть белка Z-OL1 и приблизительно половина Z-OL2 находятся в клетках в нерастворимом виде (тельца включения). Очистку полученных гибридных белков Z-OL1 и Z-OL2 проводили по одинаковой схеме путем выделения и интенсивной последовательной промывки телец включения 0.9% NaCI в 20 mM Tris-HCl (pH 7,0), затем 3 раза 1% Тритон Х-100 в 20 mM Tris-HCl. Далее белки экстрагировали 6 М мочевиной в 20 mM Tris-HCl (pH 7,0) в течение 3 часов на холоде. Полученную концентрацию и чистоту гибридных белков определяли по электрофорезу в сравнении с известным стандартом обелина. Препараты были практически гомогенны.
Восстановление активности репортера из слитых с Zip-последовательностями фрагментов обелина оценивали также при эквимолярном смешивании очищенных гибридных белков Z-OL1 и Z-OL2. Рефолдинг и зарядку фотопротеинового комплекса осуществляли разведением гибридных белков из 6 М мочевины в 30 раз зарядочным буфером O В (20 mM Tris-HCl pH 7.0, 1 mM EDTA) + 10 mM DTT + целентеразина ~10-5 M + 100 мМ KCl в течение 2-х часов. 100 мМ KCl в данном случае приблизительно имитировал ионную силу клеточного содержимого. Биолюминесцентный сигнал измеряли сразу после впрыска 200 мкл 0,1 М раствора CaCl2 в 500 мкл измерительного буфера 0,1 М Tris-HCl pH 8,8, содержащего аликвоту измеряемого образца. Для примера представляем результаты одного из экспериментов в таблице 1.
Восстановление активности обелинового репортера из данных фрагментов Z-OL1+Z-OL2 in vitro в модельном эксперименте с использованием леейцинового зиппера в качестве сближающего агента составило ~2-4% от активности полноразмерного обелина, что является довольно высоким биолюминесцентным сигналом. Незначительное восстановление биолюминесцентной активности, наблюдаемое при смешивании очищенных OL1 и OL2 рекомбинантных белков (комплементирующих фрагментов обелина без димеризующих Zip-фрагментов) без сближающего агента, можно практически не учитывать вследствие его очень малой величины.
Кинетика биолюминесцентного сигнала от восстановленной молекулы практически не изменилась.
Аналогичные результаты были получены для всех указанных в Seq. №1-8 фрагментов.
Пример 5. Определение межмолекулярных взаимодействий по восстановлению биолюминесценной активности обелина in vivo
Способность к восстановлению биолюминесценной активности при ассоциации комплементирующих фрагментов обелина in vivo, направляемой димеризацией Zip-фрагментов (далее Z), продемонстрирована при взаимодействии полученных гибридных белков Z-OL1 и Z-OL2 в клетках НЕК293. Полученные генно-инженерные конструкции из примера 2 для гибридных белков Z-OL1 и Z-OL2 были перенесены под эукариотический промотор в экспрессионный вектор pcDNA3.1(+) (Invitrogene) no сайтам KpnI-XhoI.
За 24 ч до трансфекции клетки НЕК293 были посеяны в 98-луночный планшет с плоским дном, затем котрансфецированы 0.2 мкг плазмидной ДНК в смеси, содержащей обе конструкции для детекции межмолекулярных взаимодействий (димеризация Zip-Zip). В качестве трансфецирующего агента использован FreeStyle MAX (Invitrogene) согласно инструкции производителя. Через 24 ч экспрессии среда удалена, клетки промыты PBS, разрушены замораживанием-оттаиванием и залиты 100 мкл зарядочного буфера O В (20 mM Tris-HCl, рН 7.0, 1 mМ EDTA) + 10 mМ DTT + целентеразина ~10-5 М + 100 мМ KCl. После инкубации в течение 2 часов аликвоты взяты для анализа на биолюминесцентную активность.
Восстановление биолюминесцентной активности комплементирующих фрагментов обелина in vivo, направляемых димеризацией Zip-фрагментов, составило приблизительно 10% от активности обелина дикого типа в аналогичной конструкции и условиях.
Пример 6. Использование композиции комплементирующих фрагментов фотопротеина обелина для оценки межмолекулярных взаимодействий на примере рапамицин-связывающего домена (FRB) киназы mTORC с цитозольным белком FKBP12
Для демонстрации свойств полученной композиции комплементирующих фрагментов фотопротеина мы также использовали хорошо охарактеризованную взаимодействующую систему белков: рапамицин-опосредованную ассоциацию рапамицин-связывающего домена (FRB) киназной субъединицы внутриклеточных мультимолекулярных сигнальных комплексов mTORC (регулируют клеточный рост и выживание) с цитозольным белком FKBP12 (FK506 binding protein 12). В клетке рапамицин (или сиролимус, антибиотик) связывается с 12-kDa цитозольным белком FKBP12, после чего комплекс рапамицин-РКВР12 связывается с серин-треониновой киназой mTOR и блокирует ее сигнальный путь, регулирующий рост, пролиферацию и подвижность клетки.
Фрагмент, кодирующий рапамицин-связывающий домен (FRB) человеческого mTOR (остатки 2024-2113) и FKBP1A ген человека, кодирующий 12 kDa белок FKBP12, были амплифицированы из тотальной кДНК клеток кожи человека специфическими праймерами, содержащими NcoI и BamHI сайты для клонирования. Далее амплифицированные фрагменты были клонированы вместо вырезанного по сайтам NcoI-BamHI Zip-фрагмента в соответствующих векторах pET19b-Z-OL1 и pET19b-Z-OL2, полученных в примере 3. В результате были получены конструкции, направляющие синтез гибридных белков FRB-OL1 и FKBP12-OL2, где структурные домены взаимодействующих полипептидов были связаны с комплементирующими фрагментами репортера через гибкий гидрофильный мостик (Seq.9).
Гибридные белки были очищены и проанализированы как описано в примере 4 на предмет детекции по биолюминесцентной активности взаимодействия между комплексом рапамицин-FRB и FKBP12. Результаты представлены в таблице 2.
Как можно видеть, биолюминесцентный сигнал достоверно регистрируется при ожидаемом взаимодействии белковых доменов FRB и FKBP12 только под действием рапамицина. Как и ожидалось, не наблюдалось никакого биолюминесцентного сигнала в невзаимодействующих парах гибридных белков FRB-OL1 плюс Z-OL2 (из примера 3) и Z-OL1 плюс FKBP12-OL2. Более высокий биолюминесцентный сигнал, полученный в случае взаимодействия FRB-OL1 плюс FKBP12-OL2 под действием рапамицина, в отличие от взаимодействия пары Z-OL1/Z-OL2 (пример 4) в аналогичных условиях, видимо, обусловлен взаимодействием двух разных белков FRB и FKBP12, которые образуют только гетеродимеры. Взаимодействующая пара Z-OL1/Z-OL2 приводит и к образованию гомодимеров Z-OL1/Z-OL1 и Z-OL2/Z-OL2, что должно приводить к снижению биолюминесцентного сигнала вследствие образования меньшего количества восстановленных молекул репортера.
Пример 7. Использование композиции комплементирующих фрагментов фотопротеина обелина для детекции in vivo межмолекулярных взаимодействий на примере Y2R рецептора нейропептида Y с бета-аррестином-2
Для демонстрации свойств полученной композиции комплементирующих фрагментов молекулы фотопротеина мы также использовали другую пару взаимодействующих белков: взаимодействие постсинаптического рецептора нейропептида Y второго типа (NPY2R) с бета-аррестином-2 (ARR2) человека при активации рецептора нейропептидом Y (NPY, агонист). Рецептор нейропептида Y второго типа (Y2R) вовлечен в контроль ряда поведенческих реакций, включая циркадные ритмы, аппетит, тревогу. Все 5 известных видов NPY2 рецепторов могут быть терепевтическими мишенями для ожирения и других заболеваний. Аррестины являются фундаментальными регуляторами рецепторов, подобных PY2R, при связывании перенаправляют сигнал с NPY2-рецептора на альтернативный G-белок независимый сигнальный путь.
Фрагменты кДНК, кодирующие цитозольный белок бета-аррестин-2 человека (NM_004313.3, 409 а/к, ~46 kDa) и рецептор нейропептида Y второго типа (NPY2R) человека (ВС075053, 381 а/к, ~43 kDa) были амплифицированы специфическими праймерами, содержащими Kpn I и Nde I сайты для клонирования. Далее амплифицированные фрагменты были клонированы вместо вырезанного по сайтам KpnI - NdeI фрагмента Zip + гибкий линкер в векторах pcDNA3.1-ZOL1 и pcDNA3.1-ZOL2 для эукариотической экспрессии, полученных в примере 5. В результате были получены конструкции, направляющие синтез гибридных белков NPY2R-OL1 и ARR2-OL2, где структурные домены взаимодействующих полипептидов были напрямую связаны с комплементирующими фрагментами репортера.
За 24 ч до трансфекции клетки НЕК293 были посеяны в 98-луночный планшет с плоским дном, затем котрансфецированы 0.2 мкг плазмидной ДНК в смеси, содержащей обе конструкции для детекции межмолекулярных взаимодействий. В качестве трансфецирующего агента использован Lipofectamine2000 ((Invitrogene) согласно инструкции производителя. Для активации взаимодействия через 24 ч экспрессии в часть лунок с клетками, котрансфецированными обеими конструкциями, добавлен 1 мкм нейропептида Y. Через час инкубации среда удалена, клетки собраны, разрушены замораживанием-оттаиванием и залиты 100 мкл зарядочного буфера OB (20 mM Tris-HCl рН 7.0, 1 mM EDTA) + 10 mM DTT + целентеразина ~10-5 М + 100 мМ KCl. После инкубации в течение 2-х часов аликвоты взяты для анализа на биолюминесцентную активность. Результаты представлены в таблице 3.
Как можно видеть, биолюминесцентный сигнал достоверно регистрируется при ожидаемом взаимодействии белковых доменов NPY2R и ARR2 в рекомбинантных фьюжинах только под действием нейропептида Y. Как и ожидалось, не наблюдалось никакого биолюминесцентного сигнала в клетках, экспрессирующих по отдельности полученные рекомбинантные белки-фьюжины NPY2R-OL1 или ARR2-OL2. И очень малый биолюминесцентный сигнал наблюдался без стимуляции взаимодействия нейропетидом Y. То есть, ожидаемое взаимодействие NPY2R + ARR2 специфично подтверждается по биолюминесцентному сигналу от трансфецированных клеток.
Таким образом, полученные по изобретению композиции комплементирующих фрагментов молекул фотопротеинов из Hydrozoa являются новыми репортерными молекулами для оценки биомолекулярных взаимодействий, что расширяет арсенал средств оценки межмолекулярных взаимодействий в биохимии, молекулярной биологии и биотехнологии, а также увеличивает чувствительность и диапазон измерений в анализах на основе комплементации белковых фрагментов.



Изобретение относится к биотехнологии. Описана композиция комплементирующих фрагментов молекулы фотопротеина из Hydrozoa для анализа межмолекулярных взаимодействий, полученная путем деления на две части молекулы фотопротеина в области наружной петли между 4-й и 5-й α-спиралями глобулы белка, комплементирующие фрагменты которой при ассоциации генерирует детектируемый биолюминесцентный сигнал. Предложено применение описанной композиции для анализа межмолекулярных взаимодействий, включающее генно-инженерное соединение комплементирующих фрагментов с исследуемыми полипептидами, с образованием первого и второго гибридных белков анализируемой пары, которое обеспечивает сборку комплементирующих фрагментов в функционально активный фотопротеин при взаимодействии исследуемых полипептидов, при этом данное взаимодействие регистрируют по восстановлению биолюминесцентной активности фотопротеина. Изобретение расширяет арсенал средств для анализа белок-белковых взаимодействий. 2 н. и 1 з.п. ф-лы, 4 ил., 3 табл.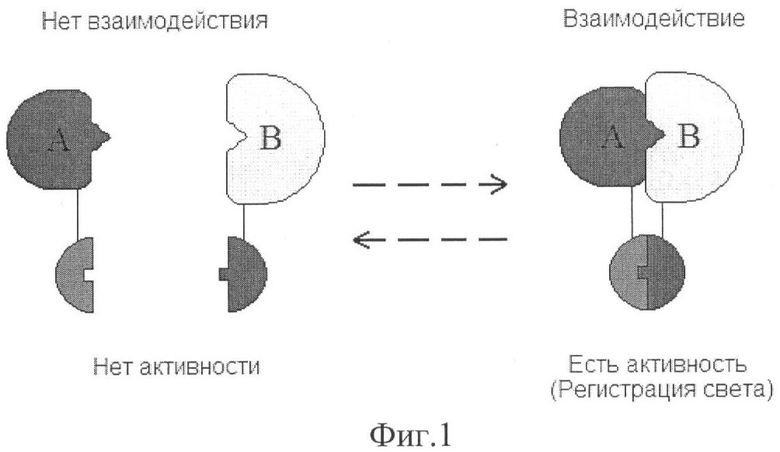
1. Композиция комплементирующих фрагментов молекулы фотопротеина из Hydrozoa для анализа межмолекулярных взаимодействий, полученная путем деления на две части молекулы фотопротеина в области наружной петли между 4-й и 5-й α-спиралями глобулы белка, комплементирующие фрагменты которой при ассоциации генерирует детектируемый биолюминесцентный сигнал.
2. Применение композиции комплементирующих фрагментов молекулы фотопротеина из Hydrozoa по п.1 для анализа межмолекулярных взаимодействий, включающее генноинженерное соединение комплементирующих фрагментов с исследуемыми полипептидами, с образованием первого и второго гибридных белков анализируемой пары, которое обеспечивает сборку комплементирующих фрагментов в функционально активный фотопротеин при взаимодействии исследуемых полипептидов, при этом данное взаимодействие регистрируют по восстановлению биолюминесцентной активности фотопротеина.
3. Применение по п.2, отличающееся тем, что первый и второй гибридные белки анализируемой пары получают генетическим слиянием фотопротеинового фрагмента с исследуемым полипептидом через гибкий гидрофильный мостик из 14 аминокислотных остатков (Seq.9).
| Способ получения нафтеновых кислот | 1960 |
|
SU137515A1 |
| Способ восстановления хромовой кислоты, в частности для получения хромовых квасцов | 1921 |
|
SU7A1 |
| Способ восстановления хромовой кислоты, в частности для получения хромовых квасцов | 1921 |
|
SU7A1 |
| Jennifer C Lewis, Sylvia Daunert "Photoproteins as luminescent labels in binding assays" Fresenius J Anal Chem (2000) 366: 760-768 | |||
| RU 98118374 A, 20.07.2000. | |||

