Группа изобретений относится к биотехнологии, генной и белковой инженерии и предназначена для получения генетически слитого бифункционального гибридного белка, состоящего из одноцепочечного антитела мыши против белка Ε вируса клещевого энцефалита и модифицированного светоизлучающего белка целентеразин-зависимой люциферазы Renilla muelleri.
Вирус клещевого энцефалита, представитель семейства Flaviviridae, является высокопатогенным инфекционным агентом, способным вызывать серьезные поражения нервной системы. Современная эпидемическая ситуация в отношении клещевого энцефалита характеризуется значительным ростом заболеваемости как Российской Федерации, так и в мире [1, 2]. Экстренная профилактика клещевого энцефалита у невакцинированных укушенных пациентов включает введение препарата «Иммуноглобулин человека против клещевого энцефалита», полученного из сыворотки донорской крови. Применение продуктов донорской крови всегда сопровождается биологическим риском. Кроме того, не исключены случаи дефицита препарата «Иммуноглобулин человека против клещевого энцефалита» в период эпидемиологического сезона. Вместе с тем, показано, что в среднем, лишь около 5% клещей являются носителями вируса клещевого энцефалита. Поэтому быстрое, высокочувствительное выявление этого вируса необходимо для экспресс-диагностики для того, чтобы избежать необоснованного применения препарата «Иммуноглобулин человека против клещевого энцефалита» для экстренной профилактики и, как следствие, снижения количества случаев возможных осложнений.
Люцифераза мягкого коралла Renilla muelleri представляет собой сравнительно небольшой, 36 кДа, одноцепочечный полипептид. Фермент катализирует окисление субстрата - целентеразина, молекулярным кислородом с образованием СО2 и молекулы целентерамида в возбужденном состоянии. Переход этого соединения в основное состояние сопровождается выделением света в видимой области спектра (λmах=482 нм). Ген, кодирующий этот белок, был клонирован, получен и изучен рекомбинантный белок дикого типа, а также методом сайт-направленного мутагенеза получен термостабильный вариант этого белка Rm7 [3, 4].
Известны ген и рекомбинантная целентеразин-зависимая люцифераза коралла Renilla reniformis, а также улучшенные производные этого белка. В литературе описаны бифункциональные гибридные белки на основе этой люциферазы, слитой с другими белками, например, зеленым флуоресцентным белком (GFP) [5, 6], рецептором [7], антителом [8] и пр., которые используются для in vivo имиджинга, исследований на основе явления BRET и др.
В доступных источниках информации не обнаружено сведений о рекомбинантных белках, способных связывать белок Ε вируса клещевого энцефалита и обладающих биолюминесцентной активностью.
Конструирование, получение и использование гибридных белков для иммуноанализа вируса клещевого энцефалита с применением люцифераз Renilla reniformis или Renilla muelleri или их аналогов в качестве репортеров в литературе также не описано.
Наиболее ближайшими к заявляемой группе изобретений -прототипом, являются рекомбинантная плазмидная ДНК pGl-Rm7, обеспечивающая синтез гибридного белка Gl-Rm7, штамм бактерий Escherichia coli HB2151- продуцент гибридного белка Gl-Rm7 и гибридный белок, связывающий фактор некроза опухолей и обладающий биолюминесцентной активностью [9].
Недостатками прототипа являются высокая трудоемкость выделения рекомбинантного белка из периплазматического пространства клетки Escherichia coli НВ2151, а также низкий выход целевого гибридного белка, составляющий 150-200 мкг с литра культуры.
Задачей группы изобретений является получение бифункционального гибридного белка, обладающего способностью связывать белок Ε вируса клещевого энцефалита и одновременно биолюминесцентной активностью, как потенциально пригодного высокочувствительного репортера для выявления вируса клещевого энцефалита в биологических образцах методом биолюминесцентного иммуноанализа.
Технический результатом группы изобретений является получение бифункционального гибридного белка, обладающего способностью связывать белок Ε вируса клещевого энцефалита и одновременно биолюминесцентной активностью, а также упрощение процедуры выделения гибридного белка и повышение его выхода.
Указанный результат достигается путем конструирования рекомбинантной плазмидной ДНК pFLAG-sc14D5a-Rm7, содержащей уникальный ген одноцепочечного антитела мыши, направленного против белка Ε вируса клещевого энцефалита, и ген модифицированной люциферазы Renilla muelleri; экспрессией целевого гибридного белка в трансформированных упомянутой плазмидной ДНК клетках Escherichia coli Rosetta-gami с последующими выделением из цитоплазматической фракции и очисткой металл-хелатной хроматографией гибридного белка 14D5a-Rm7, который обладает способностью связывать белок Ε вируса клещевого энцефалита и одновременно биолюминесцентной активностью.
Сущность группы изобретений заключается в следующем:
Генно-инженерными методами получают плазмиду pFLAG-sc14D5a-Rm7, несущую фрагмент ДНК, кодирующий одноцепочечное антитело мыши, направленное против белка Ε вируса клещевого энцефалита, коннекторный пептид (Gly2Ser)4, модифицированную люциферазу Renilla muelleri, содержащую С-концевой пептид НННННН в одной рамке считывания. Клетки Escherichia coli Rosetta-gami, трансформированные сконструированной плазмидой, способны продуцировать в растворенной форме гибридный белок sc14D5a-Rm7, состоящий из одноцепочечного антитела мыши, направленного против белка Ε вируса клещевого энцефалита, коннекторного пептида (Gly2Ser)4 и модифицированной люциферазы Renilla muelleri, содержащей С-концевой пептид НННННН, катализирующей реакцию окисления целентеразина с излучением света в видимой области спектра.
Исходным генетическим материалом для конструирования рекомбинантной плазмиды pFLAG-sc14D5a-Rm7 являются:
а) плазмидный вектор pFLAG-CTS (Sigma-Aldrich, США), обеспечивающий встраивание фрагментов ДНК, кодирующих одноцепочечное антитело sc14D5a, направленное против белка Ε вируса клещевого энцефалита, коннектор (Gly2Ser)4 и модифицированную люциферазу Renilla muelleri, содержащую С-концевой пептид НННННН, и его экспрессию под контролем tac-промотора;
б) клетки мышиной гибридомной линии 14D5 [10] - источник генов, кодирующих вариабельные домены тяжелой и легкой цепей моноклонального антитела 14D5, направленного против белка Ε вируса клещевого энцефалита;
в) ДНК плазмиды pG1-Rm7 [9], кодирующей модифицированную люциферазу Renilla muelleri, содержащую С-концевой пептид НННННН, встроенную по сайтам эндонуклеаз рестрикции NotI и ApaI.
Полученная в результате плазмида pFLAG-sc14D5a-Rm7, характеризуется следующими признаками:
- имеет молекулярную массу 4.5 МДа и размер 6930 п. о.;
- кодирует гибридный белок sc14D5a-Rm7 состоящий из одноцепочечного антитела мыши, направленного против белка Ε вируса клещевого энцефалита, коннекторного пептида (Gly2Ser)4 и модифицированной люциферазы Renilla muelleri, содержащей С-концевой пептид НННННН, катализирующей реакцию окисления целентеразина с излучением света в видимой области спектра;
- состоит из следующих элементов:
а) фрагмент ДНК NdeI-NotI размером 723 п. о., представляющий собой последовательность, кодирующую тяжелую и легкую цепи вариабельного домена моноклонального антитела sc14D5a, объединенные линкером (Gly4Ser)3;
б) фрагмент ДНК NotI - EcoRI размером 1008 п. о., представляющий собой последовательность, кодирующую пептид (Gly2Ser)4 и модифицированную целентеразин-зависимую люциферазу из мягкого коралла Renilla muelleri, содержащую С-концевой пептид НННННН;
- содержит генетические маркеры: amp - ген ампициллин-резистентности (ген β-лактамазы), определяющий устойчивость к ампициллину при трансформации Escherichia coli.
Для получения штамма-продуцента гибридного белка sc14D5a-Rm7 компетентные клетки бактерий Escherichia coli Rosetta-gami2 (Novagen, США), трансформируют сконструированной плазмидой pFLAG-sc14D5a-Rm7.
Полученный таким образом штамм Escherichia coli Rosetta gami2/pFLAG-sc14D5a-Rm7 характеризуется следующими признаками:
Морфологические признаки. Клетки мелкие утолщенной палочковидной формы, грамотрицательные, неспороносные.
Культуральные признаки. Клетки хорошо растут на простых питательных средах. При росте на агаре "Difko" - колонии круглые, гладкие, прижатые, мутные, блестящие серые, край ровный. При росте на жидких средах (на минимальной среде с глюкозой или LB бульоне) образуют интенсивную ровную муть. Клетки растут при температуре 37°C при оптимуме pH от 6.8 до 7.0.
Устойчивость к антибиотику. Клетки проявляют устойчивость к ампициллину (200 мкг/мл), обусловленную наличием плазмиды.
Штамм Escherichia coli Rosetta gami2/pFLAG-sc14D5a-Rm7 обеспечивает индуцируемый изопропилтиогалактозидом (ИПТГ) синтез гибридного белка sc14D5a-Rm7. Клетки данного штамма содержат мутированные гены тиоредоксин редуктазы и глутатион редуктазы в результате чего существенно увеличивается образование дисульфидных связей в белках при экспрессии в цитоплазме. Индикацию экспрессии осуществляют с помощью гель-электрофореза в денатурирующих условиях (SDS-PAGE). Уровень экспрессии определяют с помощью денситометрии полиакриламидного геля, окрашенного Кумaccи-R250 с использованием программного обеспечения Alfalmage, поставляемого с прибором Alphalmager (Alfalnnotech, США). Уровень экспрессии составляет около 0.5 % суммарного клеточного белка.
Весь целевой белок локализован в цитоплазматической фракции. Его выделяют и очищают с помощью металл-хелатной хроматографии. Для получения белка высокой степени очистки проводят дополнительно гель-фильтрацию. Степень очистки определяют сканированием геля на Alphalmager (Alfalnnotech, США). Концентрацию очищенного гибридного белка sc14D5a-Rm7 определяют спектрофотометрически с помощью набора DC Protein Assay (Bio-Rad, США) по протоколу производителя.
Полученный гибридный белок обладает биолюминесцентной активностью, которую определяют с помощью кюветного люминометра (модель БЛМ 8802, СКБ Наука, Россия) в буфере следующего состава 50 мМ Трис-HCl pH 7.0, 25 мМ NaCl, 1 мМ ЭДТА, сразу после внесения целентеразина (10-5 Μ раствор в метаноле). При проведении биолюминесцентного иммуноанализа измерения проводят с помощью планшетного люминометра Mithras LB 940 (Berthold, Германия). Биолюминесценцию инициируют внесением свежеприготовленного раствора целентеразина в растворе 50 мМ Трис-HCl pH 7.0, 25 мМ NaCl, 1 мМ ЭДТА.
Полученный гибридный белок обладает способностью связывать белок Ε вируса клещевого энцефалита, которая была показана с помощью прямого биолюминесцентного твердофазного иммуноанализа. При этом величина Kaff гибридного белка составила 2.65×107 М-1, что близко к значению Kaff исходного моноклонального антитела мыши 14D5 - 8.5×107 М-1 [11].
Таким образом, впервые получена плазмидная ДНК и штамм-продуцент, обеспечивающие продукцию в бактериальных клетках Escherichia coli гибридного белка sc14D5a-Rm7, состоящего из одноцепочечного антитела мыши против белка Ε вируса клещевого энцефалита и способного связывать белок Е, коннекторного пептида (Gly2Ser)4 и модифицированной люциферазы Renilla muelleri, содержащей С-концевой пептид НННННН, обладающей биолюминесцентной активностью, который обеспечивает выявление вируса клещевого энцефалита биолюминесцентным иммуноанализом.
Изобретение иллюстрируется следующими чертежами.
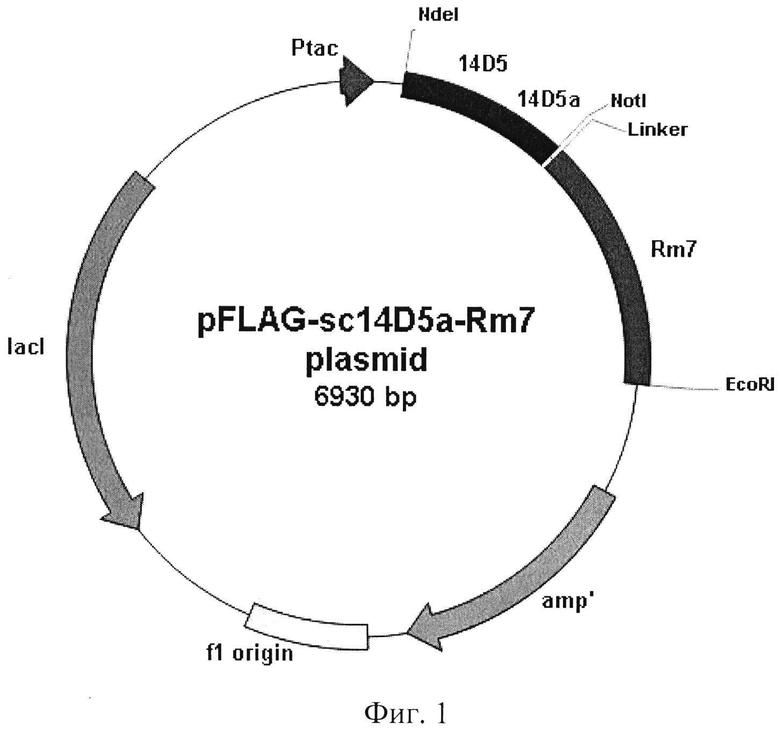
Фиг. 1. Общая схема структурной организации плазмиды pFLAG-sc14D5a-Rm7. Обозначения: sc14D5a-Rm7 - ген, кодирующий одноцепочечное антитело мыши, способное связывать белок Ε вируса клещевого энцефалита, и модифицированную люциферазу Renilla muelleri, содержащую С-концевой пептид НННННН; Ptac - область промотора; amp - ген устойчивости к ампициллину; указаны некоторые сайты рестрикции.
Фиг. 2. Нуклеотидная последовательность гена, кодирующего гибридный белок sc14D5a-Rm7 (одноцепочечное антитело-люцифераза).
Фиг. 3. Аминокислотная последовательность гибридного белка sc14D5a-Rm7.
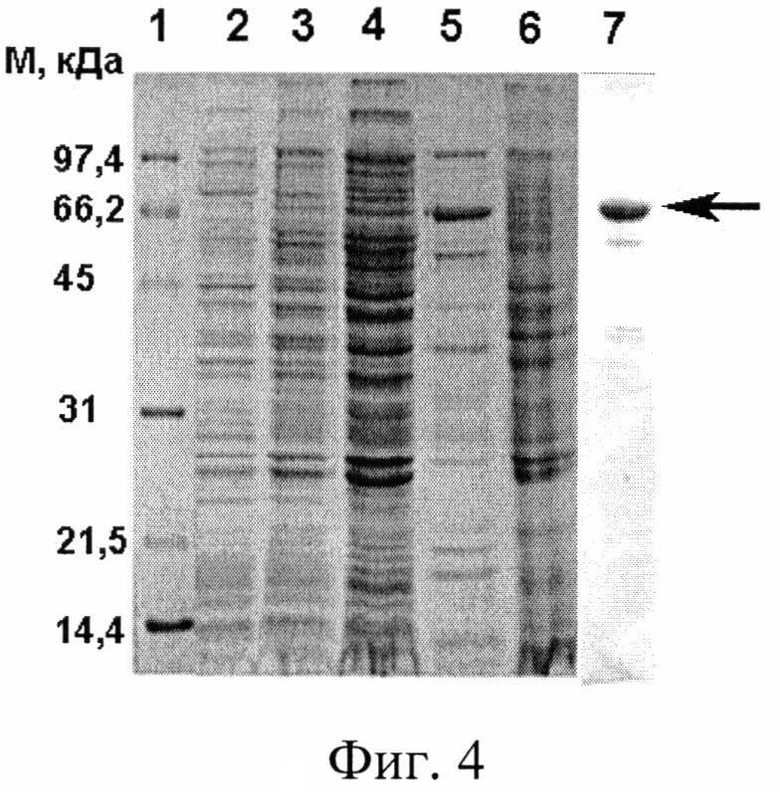
Фиг. 4. Электрофоретическое разделение фракций при выделении гибридного белка sc14D5a-Rm7 в 12,5 %-ном денатурирующем SDS-ΠΑΑΓ, где дорожки: 1 - маркеры молекулярных масс; 2, 3 - лизат клеток Rosetta-gami/ pFLAG-sc14D5a-Rm7 до и после индукции ИПТГ, соответственно; 4 - цитоплазматическая фракция; 5 - препарат гибридного белка после металл-аффинной хроматографии; 6 - фракция телец включения; 7 - препарат гибридного белка после гель-фильтрации. Стрелкой показаны полосы, соответствующие целевому гибридному белку.
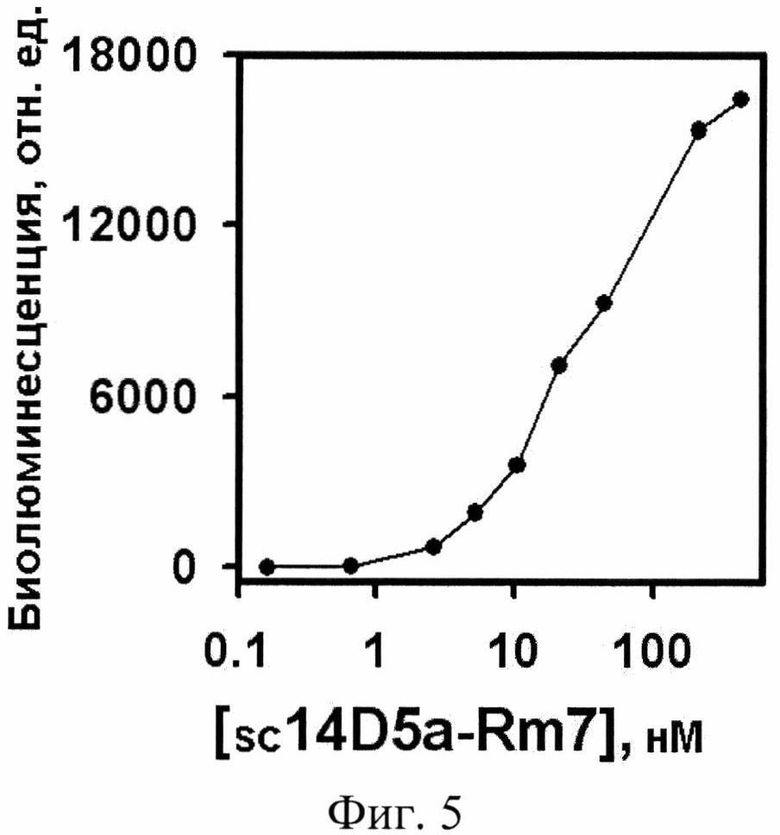
Фиг. 5. Анализ взаимодействия рекомбинантного белка Е, сорбированного на поверхности иммунопланшета, с гибридным белком sc14D5a-Rm7 методом биолюминесцентного иммуноанализа.
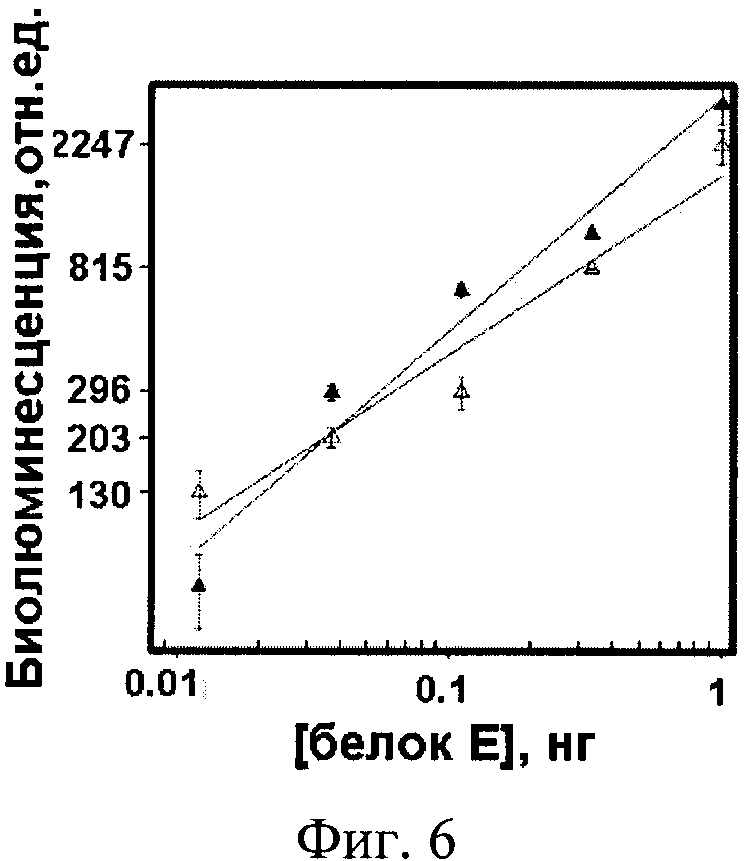
Фиг. 6. Биолюминесцентный твердофазный иммуноанализ сэндвич-типа белка Ε с использованием гибридного белка 14D5a-Rm7 до (Δ) и после (▲) замораживания.
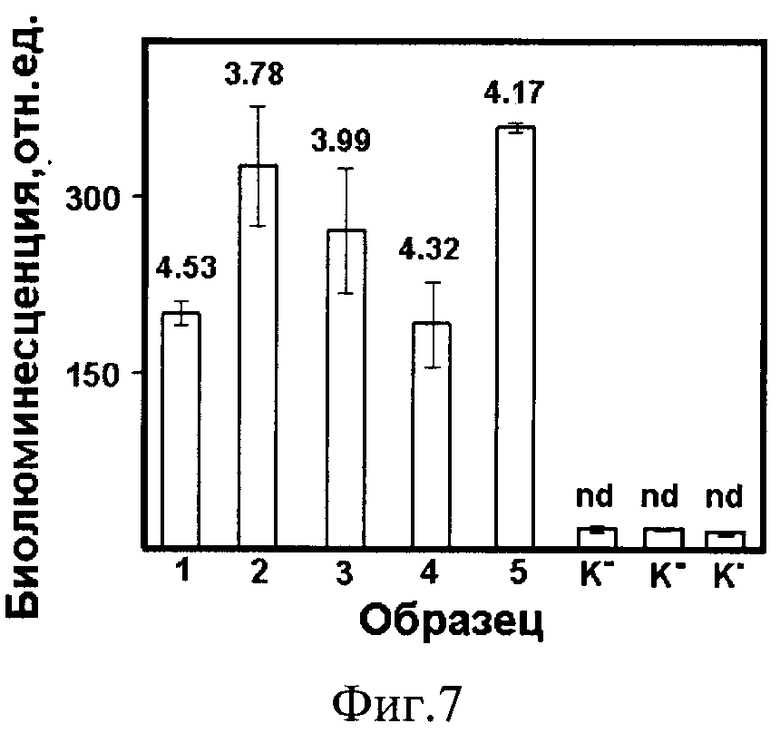
Фиг. 7. Биолюминесцентный иммуноанализ вируса клещевого энцефалита в экстрактах искусственно инфицированных клещей. Числа над столбиками - показатель логарифма (lg) количества копий РНК вируса КЭ, определенный методом РТ-ПЦР. К- - экстракты интактных клещей, nd - РНК не обнаружена.
Для лучшего понимания сущности предлагаемых изобретений, они иллюстрируются следующими примерами осуществления.
Пример 1. Конструирование гена одноцепочечного антитела мыши sc14D5a.
В качестве источника генов тяжелой и легкой цепей МКА 14D5a против вируса клещевого энцефалита используют клетки мышиной гибридомной линии 14D5 [10], продуцирующие моноклональное мышиное антитело 14D5. Из клеток гибридомы выделяют РНК с использованием набора QIAGEN RNeasy Mini Kit (Qiagen, США). Затем проводят реакцию синтеза кДНК с использованием в качестве затравки рандомизированных гексануклеотидов и фермента обратной транскриптазы (Qiagen, США). С полученной кДНК амплифицируют гены вариабельных доменов легкой и тяжелой цепей антитела МКА 14D5 с использованием олигонуклеотидов, специфических к 3′- и 5′-концам гена легкой цепи: 5′-GCAACTGCGGCCCAGCCGGCCATGGCCGARGTGCAGCTGGTGCAGTCTGG-3′ и 5′-TGAGGAGACGGTGACCGTGGTGCCTTGGCCCCA-3′, а также к 3′- и 5′-концам гена тяжелой цепи: 5′-GACATTGAGCTCАСССAGTCTCCA-3′ и 5′-CTCGACTTGCGGCCGCACGTTTGATTTCCASYTTGGTCCC-3′, соответственно. Амплификацию ведут с использованием Taq-ДНК-полимеразы (SibEnzyme, Россия) при следующих условиях: 1 цикл - 94°C, 30 сек, 2 цикл - 56°C, 30 сек, 3 цикл - 72°C, 30 сек.
Отдельно проводят отжиг двух пар олигонуклеотидов, кодирующих гибкий пептидный линкер (Gly4Ser)3:
5′-GTCACCGTCTCCTCAGGTGGCGGTGGCTCGGGCGGTGGT-3′,
5′-GGGTCGGGTGGCGGCGGATCTGACATTGAGCT-3′,
5′-CAATGTCAGATCCGCCGCCACCCGACCCACCACCGCC-3′
5′-CGAGCCACCGCCACCTGAGGAGACG-3′, после чего объединяют выделенные фрагменты генов тяжелой и легкой цепей иммуноглобулина с 10-кратным избытком олигонуклеотидного коннектора и лигируют. Амплифицируют последовательность, кодирующую гены тяжелой и легкой цепей, соединенные последовательностью, кодирующей гибкий пептидный линкер, с использованием в качестве затравки олигонуклеотидов следующего состава 215alRm7Nd5: 5′-CGCAACATATGGCCGAGGTGCAG-3′ и 48pHEN-OLXh3: 5′-ATGGTGATGCTCGAGTGCGGCC-3′, обеспечивающие в амплифицируемом фрагменте наличие сайтов рестрикции для NdeI и NotI.
Пример 2. Конструирование плазмиды pFLAG-sc14D5a-Rm7.
В качестве источника гена одноцепочечного антитела sc14D5a используют плазмидную ДНК pHEN2-sc14D5a. Для синтеза фрагмента ДНК, кодирующего ген одноцепочечного антитела sc14D5a, в реакционную смесь добавляют праймеры 215a1Rm7Nd5: 5′- CGCAACATATGGCCGAGGTGCAG-3′ и 48pHEN-OLXh3: 5′-ATGGTGATGCTCGAGTGCGGCC-3′, обеспечивающие в амплифицируемом фрагменте наличие сайтов рестрикции для NdeI и NotI.
В качестве источника гена модифицированной люциферазы Renilla muelleri, содержащей С-концевой полипептид НННННН, используют плазмиду pG1-Rm7 [9]. Для синтеза фрагмента ДНК, кодирующего ген модифицированной люциферазы Renilla muelleri, содержащей С-концевой полипептид НННННН, в реакционную смесь добавляют праймеры 198GGS4Rm7No5: 5′-СAAACGTGCGGCCGCAGGCGGCTCAGGTGGCTCAGGTGGTTCTG-3′, кодирующий коннектор (Gly2Ser)4 и содержащий сайт рестрикции для NotI, и праймер 137a1R7Ec3: 5′-CCCGGGAATTCTCAGTGGTG-3′, содержащий сайт рестрикции для EcoRI.
Амплификацию генов проводят методом ГЩР с использованием Pfu-ДНК-полимеразы (SibEnzyme, Россия). Все реакции проводят в амплификаторе MJ Mini™ (Bio-Rad, США). Условия проведения ПЦР: предварительная денатурация - 1 минута при 95°С; 20 циклов - 20 секунд при 95°C, 30 секунд при 56°C, 1 минута при 72°C; заключительная стадия - 7 минут при 72°C.
Продукт амплификации, кодирующий ген одноцепочечного антитела sc14D5a, расщепляют ферментами рестрикции NdeI и NotI в реакционной смеси, содержащей 10mМ Трис-HCl, pH 7.6, 10 mM MgCl2, 50 mМ NaCl, 1 mМ DTT, 0.1 мг/мл BSA и по 20 ед. активности соответствующих ферментов. Реакцию ведут 2 часа при 37°C.
Продукт амплификации, кодирующий ген модифицированной люциферазы Renilla muelleri, содержащей С-концевой полипептид НННННН, расщепляют ферментами рестрикции NotI и EcoRI в аналогичных условиях.
Обработку ДНК векторной плазмиды pFLAG-CTS (Sigma-Aldrich, США) ведут рестриктазами NdeI и EcoRI в аналогичных условиях, после чего дефосфорилируют с помощью щелочной фосфатазы CIP (NEB, США). После этого ПЦР-фрагменты и линеаризованный вектор очищают электрофоретически в 1 % агарозном геле с последующим выделением ДНК с помощью набора QIAquick™ Gel Extraction Kit (Quiagen, США) в соответствии с рекомендациями производителя. Лигирование проводят в стандартном буфере. Полученной лигазной смесью трансформируют клетки Escherichia coli XL 1 Blue. С помощью полимеразной цепной реакции отбирают клоны, содержащие вставку нужного размера. Полученную таким образом целевую плазмиду обозначают как pFLAG-sc14D5a-Rm7. Схема плазмидной ДНК pFLAG-sc14D5a-Rm7 представлена на фиг. 1.
Правильность конструирования подтверждают секвенированием. Результат представлен на фиг. 2, 3.
Пример 3. Получение гибридного белка sc14D5a-Rm7.
Клетки Escherichia coli Rosetta-gami2 трансформируют плазмидой pFLAG-sc14D5a-Rm, инкубируют при активном перемешивании в LB среде с добавлением 200 мМ ампициллина при температуре 37°C до оптической плотности ОД600=0,6-0,7. Затем клеточную суспензию охлаждают, экспрессию гибридного белка sc14D5a-Rm7 индуцируют добавлением 0,5 мМ ИПТГ, и клетки культивируют при температуре 37°C еще 20 ч и осаждают центрифугированием. Клетки ресуспендируют в буфере А (5 мМ имидазола, 0,3 Μ NaCl, 20 мМ Tris-HCl pH 7.0), разрушают с использованием ультразвукового дезинтегратора Bandelin Sonopuls (Bandelin, Germany) при охлаждении во льду и центрифугируют.
Супернатант наносят на колонку HisTrap™ HP (GE Healthcare, Швеция), уравновешенную буфером А, и хроматографируют белки градиентом имидазола (50-150 mМ) в том же буфере. Фракции, содержащие целевой белок, объединяют и проводят гель-фильтрацию на колонке Superdex 75 (GE Healthcare, Швеция), уравновешенную буфером 0,5 мМ ЭДТА, 25 мМ NaCl in 20 мМ Tris-HCl pH 7,0. Выход гибридного белка высокой очистки составляет 2-3 мг на 1 л LB среды, что превышает выход прототипного белка в 15-20 раз.
Результаты выделения и очистки гибридного белка представлены на фиг. 4. На фигуре 4 видно (дорожки 2, 3), что после индукции в клетках появляется новый белок, молекулярная масса которого около 65 кДа, близка к расчетной массе гибридного белка sc14D5a-Rm7 (показано стрелкой). Белок находится в цитоплазматической фракции (дорожка 4) и практически отсутствует в тельцах включения (дорожка 6). После очистки цитоплазматической фракции металл-хелатной хроматографией получен препарат, в котором гибридный белок составляет 48,5 % (дорожка 5). Гель-фильтрацией получен белок с чистотой 87 % (дорожка 7). Дорожка 1 - маркеры молекулярных масс.
Пример 4. Взаимодействие гибридного белка с рекомбинантным белком Е, иммобилизованным на поверхности иммунологического планшета.
В лунки непрозрачного иммунологического планшета (Costar, США) вносят по 100 мкл раствора рекомбинантного белка Ε (1 мкг/ мл) в 50 мМ K-Na фосфатном буфере pH 7.0 с 0,15 Μ NaCl (PBS) и инкубируют при 37°C в течение часа. После промывки (PBS, содержащий 0,1 % Tween-20 и 5 мМ ЭДТА) в лунки вносят по 100 мкл 2 % раствора обезжиренного молочного порошка в PBS и инкубируют при 37°C, 1 час. После промывки (PBS, содержащий 0,1 % Tween-20 и 5 мМ ЭДТА) в лунки вносят растворы полученного гибридного белка с концентрациями 418, 209, 44.4, 20.9, 10.45, 5.2, 2.6, 0.65, 0.16 нМ), инкубируют при 23°C 1 час при встряхивании. После промывки (PBS, содержащий 0,1 % Tween-20 и 5 мМ ЭДТА) вносят в лунки по 100 мкл раствора целентеразина (2×10-6М в 50 мМ Трис-HCl pH 7.0, 25 мМ NaCl, 1 мМ ЭДТА) с одновременным измерением биолюминесцентного сигнала связавшегося гибридного белка с помощью планшетного люминометра Mithras LB 940 (Berthold, Германия). Сигнал интегрируют в течение 20 с. Полученный результат приведен на фиг. 5. Полученную зависимость использовали для определения Кафф гибридного белка к белку Е: расчеты проводили нелинейной регрессией экспериментальных данных с использованием математического пакета SigmaPlot 2000, используя зависимость
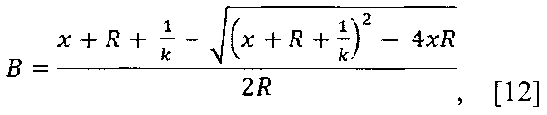
где В=L/Lmax; отношение пика интенсивности сигнала от образца (L) к сигналу от образца с насыщающей концентрацией sc14D5a-Rm7 (Lmax); R - концентрация рекомбинантного белка Ε на поверхности; х- концентрация гибридного белка в образцах, вносимых в лунки; k - константа аффинности. На фиг. 5 видно, что гибридный белок sc14D5a-Rm7 способен связываться с рекомбинантным белком Ε вируса клещевого энцефалита, сорбированным на поверхности лунок иммунологического планшета и демонстрировать биолюминисцентный сигнал при впрыскивании целентразина. Таким образом, оба домена гибридного белка демонстрируют специфические функциональные активности - способность одноцепочечного антитела sc14D5a связывать белок Ε вируса клещевого энцефалита и биолюминисцентную активность люциферазы Renilla muelleri.
Пример 5. Выявление поверхностного белка Ε вируса клещевого энцефалита в биолюминесцентном сэндвич-иммуноанализе с использованием гибридного белка sc14D5a-Rm7 как репортера.
В лунки непрозрачного иммунологического планшета (Costar, США) вносят по 100 мкл раствора мышиных моноклональных антител 14D5 [10] в PBS (5 мкг/мл) и инкубируют при 8°C в течение ночи. После промывки (PBS, содержащий 0,1 % Tween-20 и 5 мМ ЭДТА) в лунки вносят по 100 мкл 2 % раствора обезжиренного молочного порошка в PBS и инкубируют при 37°C, 1 час. После промывки (как описано выше) в лунки вносят растворы нативного белка Ε [13] в PBS, по 100 мкл, с концентрациями от 1 до 0.004 нг/мл) и инкубируют при 37°С, 1 час. Лунки промывают, вносят по 100 мкл раствора гибридного белка 14D5a-Rm7 (1 мкг/мл в PBS), инкубируют при комнатной температуре 1 час, затем промывают. Измерение сигнала гибридного белка связавшегося с мишенью проводят при внесении раствора целентеразина как описано выше. Полученные результаты показаны на фиг. 6, из которой видно, что гибридный белок sc14D5a-Rm7 способен выявлять поверхностный гликопротеин Ε вируса клещевого энцефалита в биолюминисцентном сэндвич-иммуноанализе. Репортерный сигнал зависит от концентрации белка Ε линейно в диапазоне 1-0.015 нг/лунка. Используя данную зависимость как калибровочную кривую, определяют чувствительность биолюминисцентного иммуноанализа, составляющую 0.025 нг белка Е. Полученная величина превышает в десять раз чувствительность коммерческого колориметрического иммуноанализа, базирующегося на использовании в качестве метки пероксидазы.
Пример 6. Применение гибридного белка sc14D5a-Rm7 как репортера в биолюминесцентном иммуноанализе вируса клещевого энцефалита в экстрактах инфицированных клещей.
Экстракты инфицированных клещей Dermacentor marginatus получают на второй день после инфицирования. В качестве контрольных (К-) аналогичные экстракты получают от интактных клещей. Каждого клеща замораживают при -70°C, растирают в гомогенизаторе и суспендируют в 100 мкл PBS. Половину экстракта разводят в 10 раз и используют для иммуноанализа как описано в Примере 5. Вторую половину экстрактов исследуют на присутствие вирусной РНК методом ПЦР в режиме реального времени с использованием набора "Real Best RNA TBEV" (Вектор Бест, Россия) по протоколу производителя. Полученные результаты двух способов определения вируса клещевого энцефалита (ВКЭ) хорошо коррелируют и показаны на фиг. 7, из которой видно, что биолюминисцентные сигналы регистрируются только в экстрактах клещей, инфицированных вирусом клещевого энцефалита, что подтверждается результатами, получаемыми методом ПЦР в режиме реального времени.
Таким образом, впервые получена плазмидная ДНК pFLAG-sc14D5a-Rm7, содержащая в одной рамке считывания ген одноцепочечного мышиного антитела 14D5, способного связывать капсидный белок Ε вируса клещевого энцефалита, и ген модифицированной люциферазы Renilla muelleri; получен штамм бактерий Escherichia coli - продуцент гибридного белка sc14D5a-Rm7; получен рекомбинантный гибридный белок - одноцепочечное антитело-люцифераза (sc14D5a-Rm7), связывающий капсидный белок Ε вируса клещевого энцефалита и обладающий биолюминесцентной активностью, что делает возможным его применение в качестве высокочувствительного репортера для биолюминесцентного иммуноанализа вируса клещевого энцефалита.
ИСТОЧНИКИ ИНФОРМАЦИИ
1. T.G.T. Jaenson, Μ. Hjertqvist, Т. Bergstrom, A. Lundkvist. Why is tick-bome encephalitis increasing? A review of the key factors causing the increasing incidence of human TBE in Sweden. Parasit Vectors. - 2012 - Vol. 5. - P. 184-197.
2. Kunze U; ISW-TBE. Tick-borne encephalitis-a notifiable disease: report of the 15th Annual Meeting of the International Scientific Working Group on Tick-Borne Encephalitis (ISW-TBE) Ticks Tick Borne Dis. - 2013 - Vol. 4(5). - P. 363-365.
3. M.S. Titushin, S.V. Markova, L.A. Frank, N.P. Malikova, G.A. Stepanyuk, J. Lee, E.S. Vysotski. Coelenterazine-binding protein of Renilla muelleri: cDNA cloning, overexpression, and characterization as a substrate of luciferase. // Photochem. Photobiol. Sci. - 2008 - Vol. 7. - P. 189-196.
4. G.A. Stepanyuk, J. Unch, N.P. Malikova, S.V. Markova, J. Lee, E.S. Vysotski. Coelenterazine-v ligated to Ca2+-triggered coelenterazine-binding protein is a stable and efficient substrate of the red-shifted mutant of Renilla muelleri luciferase. Anal. Bioanal. Chem. - 2010 - Vol. 398. - P. 1809-1817.
5. Wang Y., Yu Y.A., Shabahang S., Wang G., Szalay A.A. Renilla luciferase-Aequorea GFP (Ruc-GFP) fusion protein, a novel dual reporter for real-time imaging of gene expression in cell cultures and in live animals. Mol. Genet. Genomics. -2002 - Vol. 268. - P. 160-168.
6. J. Liu, Y. Wang, A.A. Szalay, A. Escher. Visualizing and quantifying protein secretion using a Renilla luciferase-GFP fusion protein. Luminescence. - 2000 - Vol. 15. - P. 45-49.
7. Zeng F.-Y., Mclean A.J., G. Milligan, Lerner M., Chalmers D.T., Behan D.P. Ligand specific up-regulation of a Renilla reniformis luciferase-tagged, structurally unstable muscarinic M3chimeric G protein-coupled receptor. Mol. Pharmacol. - 2003 - Vol. 64. - P. 1474-1484.
8. Venisnik Κ.Μ., Olafsen Т., Loening Α.Μ., Iyer Μ., Gambhir S.S., Wu A.M. Bifunctional antibody-Renilla luciferase fusion protein for in vivo optical detection of tumors. Prot. Eng. Des. Sel. - 2006 - Vol. 19. - P. 453-460.
9. Патент RU 2513686 C1, oп. 20.04.2014.
10. Tsekhanovskaya N.A., Matveev L.E., Rubin S.G., Karavanov A.S., Pressman E.K. Epitope analysis of tick-borne encephalitis (TBE) complex viruses using monoclonal antibodies to envelope glycoprotein of TBE virus (persulcatus subtype). Virus Res. - 1993 - Vol. 30(1) - P. 1-16.
11. Baykov I.K., Matveev A.L., Stronin O.V., Ryzhikov A.B., Matveev L.E., Kasakin M.F., Richter V.A., Tikunova N.V. A protective chimeric antibody to tick-borne encephalitis. Vaccine - 2014 - Vol. 32 - P. 3589-3594.
12. Beatty J.D., Beatty B.G., Vlahos W.G. Measurement of monoclonal antibody affinity by non-competitive enzyme immunoassay. J. Immunol. Meth. - 1987 - Vol. l00 - P. 173-179.
13. Ляпустин B.H., Карганова Г.Г., Соболев С.Г., Гритцун Т.С. Препаративное разделение антигенных структур вируса клещевого энцефалита электрофорезом в жидкой среде. Вопросы вирусологии. - 1988 - Vol. 33(1) - P. 98-102.





Группа изобретений относится к биотехнологии, генной и белковой инженерии. Сконструирована плазмида pFLAG-sc14D5a-Rm7, обеспечивающая в клетках Escherichia coli синтез рекомбинантного белка sc14D5a-Rm7, способного связывать белок Ε вируса клещевого энцефалита и обладающего биолюминесцентной активностью. Создан штамм бактерий Escherichia coli - продуцент гибридного белка sc14D5a-Rm7 в цитоплазматическом растворе. Получен рекомбинантный белок sc14D5a-Rm7, выделенный из клеток Escherichia coli, трансформированных плазмидой pFLAG-sc14D5a-Rm7, имеющий молекулярную массу около 63,9 кДа; состоящий из одноцепочечного антитела человека против белка Ε вируса клещевого энцефалита, коннекторного пептида GGSGGSGGSGGS и рекомбинантной люциферазы Renilla muelleri; имеющий аминокислотную последовательность, кодируемую нуклеотидной последовательностью SEQ ID NO:1, обеспечивает выявление белка Ε вируса клещевого энцефалита в диапазоне от 1 нг до 0,025 нг методом биолюминесцентного твердофазного иммуноанализа. Технический результат: получение бифункционального гибридного белка, обладающего способностью связывать белок Ε вируса клещевого энцефалита и одновременно биолюминесцентной активностью. 3 н.п. ф-лы, 7 ил., 6 пр.
1. Рекомбинантная плазмидная ДНК pFLAG-sc14D5a-Rm7, обеспечивающая синтез гибридного белка sc14D5a-Rm7, имеющая размер 6930 п.о. и молекулярную массу 4.5 Md и содержащая в соответствии с физической и генетической картой плазмиды, приведенной на фиг. 1, уникальные сайты рестрикции NdeI (111), HindIII (171), XhoI (177), EcoRI (182), SmaI (187), XmaI (189), Asp718 (191), KpnI (195), BglII (197), SalI (203), и нуклеотидную последовательность SEQ ID NO:1, кодирующую гибридный белок с аминокислотной последовательностью SEQ ID NO:2, состоящий из одноцепочечного антитела против белка Ε вируса клещевого энцефалита, пептида GGSGGSGGSGGS и модифицированной люциферазы Renilla muelleri.
2. Штамм бактерий Escherichia coli - продуцент гибридного белка sc14D5a-Rm7, полученный трансформацией культуры клеток Escherichia coli Rosetta-gami2, плазмидной ДНК pFLAG-sc14D5a-Rm7, имеющей нуклеотидную последовательность SEQ ID NO:1, представленную на фиг. 2.
3. Гибридный белок sc14D5a-Rm7, связывающий белок Ε вируса клещевого энцефалита и обладающий биолюминесцентной активностью, полученный из клеток Escherichia coli Rosetta-gami2, трансформированных плазмидной ДНК pFLAG-sc14D5a-Rm7, имеющий молекулярную массу 63,9 кДа, состоящий из одноцепочечного антитела против белка Ε вируса клещевого энцефалита, пептида GGSGGSGGSGGS и модифицированной люциферазы Renilla muelleri, имеющий аминокислотную последовательность SEQ ID NO:2, представленную на фиг. 2.
| Штамм гибридных культивируемых клеток животных MUS. мUSсULUS L. - продуцент моноклональных антител к гликопротеину Е /V 3/ вируса клещевого энцефалита | 1989 |
|
SU1666531A1 |
| Зажим для присоединения трех электрических проводников | 1958 |
|
SU130085A1 |
| ПЛАСТМАССОВАЯ ЕМКОСТЬ С НАПРАВЛЯЮЩИМ ПОТОК КОМПОНЕНТОМ | 2016 |
|
RU2716655C2 |

