Изобретение относится к области биотехнологий, в частности к клеточной и тканевой инженерии, и может быть использовано в медицине и ветеринарии.
Эмбриональные стволовые клетки (ЭСК) млекопитающих, которые получают из клеток внутренней клеточной массы бластоцист, способны пролиферировать вне организма (в культуре in vitro) в течение неограниченного времени, сохраняя при этом свои свойства. Путем модуляции параметров культивирования ЭСК можно добиваться дифференцировки ЭСК in vitro практически во все клеточные типы тканей взрослого организма (за исключением двух внезародышевых клеточных типов - трофоэктодермы и первичной эндодермы). Эти два свойства, обозначенные термином «плюрипотентность», делают ЭСК ценным источником материла для регенеративной медицины (заместительной и восстановительной клеточной терапии). На сегодняшний день получены уже несколько сотен линий ЭСК, стали известны некоторые молекулярные и генетические аспекты механизмов регуляции самообновления и плюрипотентности ЭСК, ведутся широкомасштабные работы по оптимизации методов их дифференцировки в различные клеточные типы. Однако очевидны и серьезные ограничения, заметно отдаляющие перспективу применения ЭСК в клинической практике. Одним из таких ограничений является отсутствие ЭСК во взрослом организме с возникающей вследствие этого проблемой иммунной несовместимости при аллогенной трансплантации дифференцированных производных от уже полученных человеческих ЭС линий. Известно, что недифференцированные ЭСК, хотя и не экспрессируют HLA антигены (и, следовательно, не должны отторгаться при аллогенном переносе), являются высоко туморогенными, т.е. способными инициировать опухолевый процесс при введении в организм реципиента. С другой стороны, полученные из ЭСК дифференцированные клетки (то есть клетки-производные ЭСК), хотя и не являются туморогенными, экспрессируют HLA антигены, что заставляет проводить иммуносупрессию при аллогенной трансплантации таких клеток реципиентам.
Для решения этой проблемы были предложены несколько подходов. Во-первых, создание банков человеческих ЭС клеток, охватывающих все HLA типы, поможет решить эту проблему, но это, по-видимому, будет (если будет) реализовано только в отдаленной перспективе. Во-вторых, терапевтическое клонирование было предложено как альтернативный метод получения аутологичных (пациент-совместимых) ЭСК. При таком подходе ядра соматических клеток пациента подсаживают в безъядерные ооциты. ЭСК затем получают в культуре из развившихся клонированных бластоцист. Это весьма дорогостоящий и малоэффективный метод, который к тому же предполагает уничтожение доимплантационных человеческих эмбрионов, что запрещено в некоторых странах по этическим соображениям. Решением этой проблемы явилась недавно продемонстрированная возможность получения аутологичных (полностью пациент-совместимых) плюрипотнентных клеток. Поэтому получение аутологичных ЭСК или подобных им клеток непосредственно из соматических клеток пациента, не прибегающее к терапевтическому клонированию, представлялось крайне актуальным. Подобная идея доказала свою состоятельность, когда одна группа из Киотского университета путем простого перебора различных комбинаций из 24 генов, экспрессия которых ассоциирована с ЭСК фенотипом, показала возможность индукции плюрипотентного состояния соматических клеток (МЭФ) при помощи форсированной генной экспрессии (Takahashi and Yamanaka, 2006). Как оказалось, экспрессии 4 транскрипционных факторов Oct4, Sox2, cMyc и Klf4, доставленных в фибробласты при помощи ретровирусных векторов, на протяжении нескольких дней было достаточно для получения иПС клеток. Полученные иПС клетки обладали всеми свойствами, присущими ЭС клеткам мыши, т.е. способностью самообновляться и дифференцироваться in vivo и in vitro во все эмбриональные типы клеток, включая половые клетки. За последующие 3 года вышло большое количество работ по данной тематике, описавших альтернативные комбинации транскрипционных факторов, улучшающие эффективность получения иПС клеток у разных видов (включая человека), предложивших различные способы доставки (вирусный, плазмидный, транспозонный, белковый), использовавших различные исходные соматические клеточные типы для получения иПС клеток и т.д. (Ellis et al., 2009; Maherali and Hochedlinger, 2008). Стало совершенно понятно, что клеточная терапия, основанная в первую очередь на иПСК, уже в ближайшие годы сулит здоровью человечества огромные выгоды.
Другой и пока еще нерешенной проблемой, стоящей на пути к использованию и ЭСК, и иПСК в клинической практике, стоит туморогенность этих клеток. Так, известно, что попадание во взрослый организм мыши хотя бы нескольких ЭСК приводит к возникновению тератом (Nishimura, et al., 2003). С другой стороны, существующие способы направленной дифференцировки ЭСК in vitro позволяют получить не «чистые», а весьма гетерогенные популяции клеток - производных ЭСК. В таких гетерогенных популяциях практически всегда присутствуют резидуальные (остаточные) недифференцированные клетки, обладающие высоким туморогенным потенциалом (Brederlau et al., 2006). Описанная выше ситуация в полной мере относится и к иПСК. При этом отдельные недифференцированные клетки (ЭСК и иПСК) способны вызвать в сайте трансплантации развитие тератомы, а клетки-производные ЭСК и иПСК, достигшие невысокого уровня дифференцировки (и сохранившие способность активно пролиферировать), - нетератомные опухоли (например, шванному). Таким образом, присутствие даже небольшого количества недифференцированных ЭСК и иПСК (и даже одной такой клетки) в трансплантируемом пациенту клеточном материале недопустимо ввиду высокого риска возникновения тератом, - а активно пролиферирующих клеток, являющихся их производными - ввиду риска возникновения нетератомных опухолей. Соответственно разработка способов, обеспечивающих полное удаление резидуальных ЭСК и иПСК и их сохранивших пролиферативную активность производных из гетерогенных клеточных суспензий (субстрата трансплантаций) остается одной из самых актуальных на сегодняшний день задач в области клеточных технологий.
Способы получения дифференцированных стволовых клеток, позволяющие селективно очистить гетерогенные клеточные популяции от резидуальных недифференцированных плюрипотентных клеток и пролиферирующих клеток на промежуточных этапах дифференцировки основаны на нескольких принципах, включающих длительную дифференцировку (Brederlau et al., 2006), высокопроизводительный сортинг клеток по известным маркерам (позитивная или негативная селекция) (Pruszak et al., 2007), использование антител к недифференцированным клеткам (US 20030032187) или обработку клеточных популяций факторами, селективно индуцирующими апоптоз в недифференцированных клетках (Choo et al., 2008), включая аналоги церамида (Bieberich et al., 2001). Доступные данные позволяют заключить, что технический уровень достигнутых к настоящему времени результатов не обеспечивает эффективного решения поставленной задачи: а именно полного удаления резидуальных ЭСК и иПСК и их сохранивших пролиферативную активность производных из гетерогенных клеточных суспензий (Анисимов, 2009). Увеличение длительности in vitro дифференцировки стволовых клеток до полного исчезновения в клеточной популяции недифференцированных клеток связано с высокими затратами и возрастанием риска прерывания протокола из-за спонтанного изменения направления дифференцировки, контаминации клеточной системы, и т.д. (Brederiau et al., 2006; Анисимов, 2009). Применение химических факторов, способных селективно индуцировать апоптоз в недифференцированных клетках (синтетический N-олеол-серинол (S18); Bieberich et al., 2003, 2004; US20060014280) обеспечивает лишь частичное снижение риска развития онкологических осложнений трансплантации производных ЭСК, поскольку механизм действия S18 (действительно способного селективно индуцировать апоптоз в недифференцированных клетках) опосредован наличием в клетках-мишенях экспрессии белка prostate apoptosis response-4 (PAR-4). По имеющимся данным, этот белок экспрессирован лишь приблизительно в 10% клеток человека, экспрессирующих маркер плюрипотентности Oct4. Отсюда, специфическая индукция апоптоза в 10% недифференцированных клеток-производных ЭСК человека способна лишь несколько снизить риск «перерастания трансплантата» (неконтролируемой пролиферации прогениторных клеток-производных ЭСК) и развития тератом/нетератомных опухолей в сайте трансплантации, не решив, следовательно, вопрос принципиально.
Помимо проблемы тератогенности ЭСК и иПСК клеток, разработанные методы направленной in vitro дифференцировки этих клеток в клинически востребованные типы клеток являются преимущественно малоэффективными и всегда приводят к получению весьма гетерогенных популяций диффренцированных клеток. Например, технология направленной in vitro дифференцировки ЭСК человека в такие высокоспециализированные клетки, какими являются функциональные дофаминергические нейроны, характеризуется высоким уровнем затрат и вариабельности (что, в частности, обусловлено большой продолжительностью комплекса воздействий, оказываемых на клетки) (Анисимов, 2009). Поэтому проблема увеличения эффективности технологий направленной дифференцировки, а также повышение гомогенности получаемых клеточных типов является крайне актуальной.
Одним из наиболее перспективных подходов к существенному снижению риска возникновения тератом является метод, основанный на генетической сенсибилизации исходной клеточной популяции (ЭСК и иПСК). Данный метод подразумевает введение в геном ЭСК и иПСК и поддержание в стабильном состоянии так называемых «суицидальных генов» или генов-самоубийц: например, гена тимидин киназы (ТК) или мышиного рецептора токсина дифтерии (мДТР) под контролем промоторов, поддерживающих экспрессию этих маркеров исключительно в недифференцированных клетках (ЭСК и иПСК), - например, промоторов генов Oct4 или Nanog. Генетически модифицированные ЭСК и иПСК затем подвергаются дифференцировке in vitro в желаемом направлении, после чего резидуальные ЭСК и иПСК элиминируются добавлением в среду ганцикловира (метаболизируемого ТК) или токсина дифтерии (мДТР).
В качестве прототипа нами выбран способ, описанный в патенте US 6576464. На первом этапе в недифференцированные стволовые клетки вводят молекулу нуклеиновой кислоты, состоящую из 2 элементов. Первый элемент - это последовательность нуклеиновой кислоты, кодирующая продукт, являющийся смертельным для клетки, в которой он экспрессирован, или делающий клетку, в которой он экспрессирован, подверженной действию летального внешнего фактора. Второй элемент - это элемент контроля транскрипции, который обеспечивает экспрессию первого элемента преимущественно в недифференцированных стволовых клетках. Первый элемент содержит гены, кодирующие токсин или белок, индуцирующие клеточную гибель (суицидальные гены), - например фермент (ТК), конвертирующий лекарственный субстрат (широко применяемый противовирусный препарат ганцикловир) в субстрат, летальный для данной клетки. При этом экспрессия кодирующих фермент генов находится под контролем промотора (эндогенного транскрипционного контрольного элемента), специфичного для недифференцированных стволовых клеток (например, промотора генов Oct4 или обратной транскриптазы теломеразы (TERT)). На втором этапе проводят индукцию направленной дифференцировки стволовых клеток: параметры данного этапа зависят от задач протокола (направления дифференцировки клеток). На третьем этапе в культивируемую клеточную популяцию добавляют ганцикловир, который селективно метаболизируется недифференцированными стволовыми клетками (экспрессирующими гены Oct4 или обратной транскриптазы теломеразы (TERT)) и индуцирует в этих клетках клеточную гибель (апоптоз). Это обеспечивает селективную абляцию резидуальных недифференцированных СК из гетерогенной клеточной популяции. По данным авторов изобретения доля оставшихся недифференцированных клеток не превышает 0,02% от общего числа клеток в популяции (US 6576464). Между тем, даже небольшое число недифференцированных клеток, персистирующих в гетерогенной популяции, может могут стать причиной развития тератомы в сайте/сайтах трансплантации клеток. Кроме того, патент-прототип (US 6576464), не предлагает какого-либо способа увеличения эффективности технологий направленной дифференцировки ЭСК.
Целью настоящего изобретения является обеспечение возможности проведения эффективной тканезаместительной терапии с использованием плюрипотентных СК (ЭСК и иПСК) при сведении к нулю риска развития тератом, происходящих из донорских плюрипотентных СК в месте введения или в каком-либо дистальном сайте у реципиента. Достигается это в настоящей заявке путем введении в геном плюрипотентных СК, получаемых из предимплантационных зародышей или соматических клеток разных типов взрослых организмов общепринятыми способами, и поддержании в стабильном состоянии мультикомпонентной генетической конструкции (например, бицистронной кассеты). Такая кассета включает ген-самоубийцу, например ТК или ДТР, под контролем регуляторного элемента, специфического для плюрипотентных СК, такого как энхансер/промотор гена Oct4. Для положительной селекции кассета дополнительно содержит ген устойчивости к антибиотику, например пуромицину, находящемуся под контролем того же регуляторного элемента. После проведения электропорации (при использовании плазмидного вектора) или инфицирования (в случае использования вирусного вектора) клеточная популяция обрабатывается антибиотиком, например пуромицином, для позитивного отбора клонов ЭСК/иПСК, встроивших указанную кассету в свой геном. В настоящей заявке способы введения и поддержания суицидальной кассеты в иПСК и ЭСК не ограничиваюся стабильно интегрирующими в геном плазмидными или вирусными векторами, но также включают, транспозонные, эписомные вектора, а также искусственные хромосомы. Для удаления спонтанно дифференцированных клеток и клеток, в которых по каким-либо причинам произошло затухание экспрессии указанной выше бицистронной кассеты, за 1-2 дня перед введением реципиентам ЭСК/иПСК клоны дополнительно культивируют в присутствии пуромицина. После этого клетки вводят, без какой-либо предварительной дифференцировки в культуре, непосредственно в кровоток, орган, или ткань реципиентов. Через 1-14 дней реципиентам проводится курс терапии ганцикловиром.
Положительный эффект от применения предлагаемого изобретения заключается: 1) в сведении к нулю риска развития у реципиентов тератом после трансплантаций клеточного материала, 2) в повышении клинической эффективности восполнения клеточной массы поврежденного органа или ткани, 3) в сокращении, благодаря отсутствию этапа дифференцировки в культуре, длительности периода ожидания для потенциальных реципиентов.
Изобретение иллюстрируется следующими графическими материалами.
На Фиг.1 изображена кассета, обеспечивающая трансляцию тимидинкиназы (ТК) и маркера резистентности к пуромицину (Puro) с общего бицистронного транскрипта, чья селективная экспрессия в ЭСК и иПСК обеспечивается за счет комбинации энхансера 2А2В из дистальной консервативной области CR4 гена Oct4 (Pou5f1) и минимального промотора гена тимидин киназы (tk). На Фиг.1 использованы следующие сокращения: IRES - внутренний сайт посадки рибосом, BOVGH рА - сайт полиаденилирования из гена гормона роста быка, start - сайт инициации транскрипции.
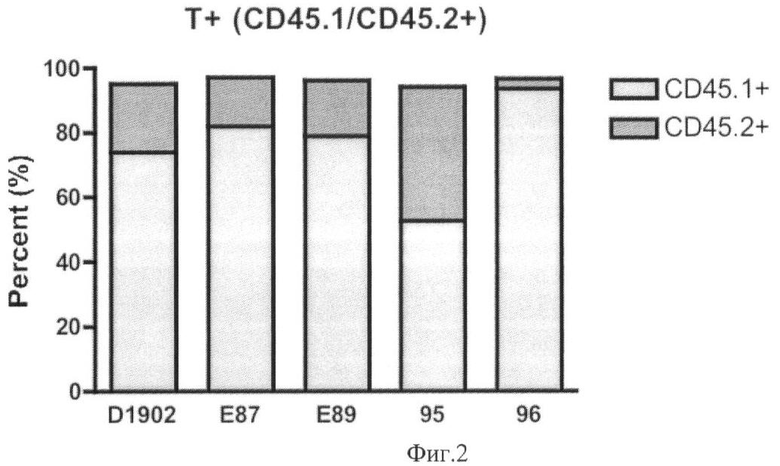
На Фиг.2 показан пример участия ЭСК, несущих в своем геноме указанную на Фиг.1 кассету (клетки были названы ТК-ЭСК) в восстановлении кроветворения у летально облученных мышей. Показан результат анализа с использованием проточной цитофлуориметрии кровяных клеток 5 реципиентов, получивших сразу после облучения инъекцию 106 ТК-ЭСК (CD45.2+), а также прошедших 5-дневный курс инъекции ганцикловира (10 мг/кг веса тела), начиная с 10-го дня после облучения. Использовались специфические антитела к изотипам 1 и 2 антигена CD45 - известного поверхностного маркера В-, Т-лимфоцитов и миелоидных типов клеток. Анализ проводили через 6 недель после трансплантации.
Способ осуществляется следующим образом: полученные общепринятым способом культуры ЭСК или иПСК (Thomson et al., 1998; Reubinoffet al., 2000; Takahashi et al., 2007; Yu et al., 2007; Nakagawa et al., 2008) (в том числе коммерчески доступные линии ЭСК) культивируют согласно существующим рекомендациям, и генетически модифицируют указанной на Фиг.1 кассетой, доставленной в клетки при помощи плазмидного или лентивирусного векторов путем электропорации (плазмидный вектор) или заражения (лентивирусный вектор) и последующего отбора пуромицин-резистентных (1-2 мкг/мл) клонов в течение 2 суток, начатого через 48 ч после заражения или электропорации. Клетки после генетической модификации бицистронной кассетой, обозначаемые в дальнейшем как «ТК-ЭСК», индивидуально отбирают на 8-12-й день после начала селекции; клетки нарабатывают в необходимых для трансплантации количествах (число клеток зависит от организма; например, для мыши - 2-5×106 клеток), а затем вводят реципиентам. По прошествии 1-14 дней проводится 5-7-дневный курс внутримышечных инъекции ганцикловира (не менее 10 мг/кг веса тела).
Работоспособность разработанной бицистронной кассеты иллюстрируется приведенными ниже in vitro тестами.
Тест 1. Оценка эффективности удаления резидуальных ТК-ЭСК при нейрональной дифференцировке in vitro. День -4 ÷ -2 (за 0 принимали день начала дифференцировки): клетки дополнительно культивировали на обработанном 0.1% желатином культуральном пластике в присутствии 1 мкг/мл пуромицина для гарантированного удаления спонтанно дифференцированных ТК-ЭСК и ТК-ЭСК с инактивированной кассетой. Среда содержала 15% сыворотку плодов коровы (Sigma, США), 100 мкМ заменимые аминокислоты (Invitrogen, США), 2 мМ L-глютамин (Invitrogen), 50 мкг/мл пенициллин/стрептомицин (Invitrogen), 100 мкМ (3-меркаптоэтанол (Sigma), 10 нг/мл бактериально экспрессированный и аффинно очищенного LIF. День 0-10: формирование и культивирование эмбриональных тел в той же среде, но на неадгеизивной поверности бактериальных чашек Петри. День 11: перенос эмбриональных тел на чашки, обработанные орнитином и ламинином, и дальнейшее культивирование в среде N2B27 в присутствии 10 нг/мл основного фактора роста фибробластов (bFGF) и 20 нг/мл эпидермального фактора роста (EGF). День 13-20: культивирование прикрепившихся к подложке клеток в присутствии 2 мкМ ганцикловира (Sigma), с дальнейшей окраской на щелочную фосфатазу (маркер ЭСК) или TuJ (маркер нейронов). Таблица 1 (животные 1-12) иллюстрирует типичный результат описанного теста, демонстрирующего отсутствие резидуальных ТК-ЭСК при культивировании дифференцированной клеточной популяции (производной пуромицинрезистентных клеток) в присутствии ганцикловира. Для дальнейшего подтверждения эффективности работоспособности кассеты клеточные суспензии клеток инъецировали под кожу бестимусным мышам линии NUDE. Культивировавшиеся в присутствии ганцикловира клетки тератом не образовывали (Таблица 1, мыши 4-6 и 10-12). Дифференцированные клетки, не прошедшие такую обработку, во всех случаях формировали тератомы (Таблица 1, мыши 1-3 и 7-9), что еще раз подчеркивало проблему остаточных ЭСК в диффренцированных клеточных популяциях.
Тест 2. Оценка эффективности подавления образования тератом in vivo, после трансплантации недифференцированных ТК-ЭСК (резистентных к пуромицину) под кожу мышам линии NUDE. 7-дневный курс внутримышечных инъекций ганцикловира (10 мг/кг веса тела) начинали либо в день инъекции ЭСК, либо спустя несколько дней (до 10 дней), что во всех случаях подавляло возникновение тератом (Таблица 1, мыши 13-15 и 18-20). Наблюдение за этой группой мышей линии NUDE на протяжении 5 месяцев не выявило тератом или каких-либо патологий. С другой стороны, тератомы возникали через 3-4 недели после введения ЭСК у всех мышей, не прошедших курс инъекций ганцикловиром (Таблица 1, мыши 16, 17, 21, 22). Контрольные ЭСК дикого типа образовывали тератомы, независимо от инъекций ганцикловира (Таблица, мыши 23-24), что подчеркивало важность ТК-кодирующей кассеты для обеспечения работоспособности системы.
В таблице приведен анализ трех различных клонов ТК-ЭСК (No. 3, 14 и 15), несущих бицистронную кассету в различных геномных локусах. Несмотря на эту разницу, во всех трех клонах удаление резидуальных ТК-ЭСК было обеспечено с высокой степенью надежности, что указывало на универсальность предложенной в заявке системы сенсибилизации плюрипотнентных СК.
Описанные тесты позволяют убедится в работоспособности предложенной системы in vitro, однако не являются обязательными для проведения заключительного этапа, который заключается в использовании ТК-ЭСК для восстановления функции органа или ткани реципиента in vivo. Достигается это путем прямого введения недифференцированных ТК-ЭСК реципиенту без каких-либо предварительных дифференцировок и абляций резидуальных ТК-ЭСК у реципиента. В прямом использовании недифференцированных ТК-ЭСК и заключается новизна предложенного изобретения, в корне отличающего его от US6576464 и других патентов.
Пример 1. Иллюстрацией предложенного подхода является описанный ниже эксперимент, показывающий восполнения кроветворной фунции костного мозга мышей за счет ТК-ЭСК. Гибридные мыши (129Sv X C57B16, представляющие собой гемизиготы CD45.1/CD45.2) подвергались летальной дозе облучения (2×5,5 Гр с 5-мин интервалом). В тот же день мыши получали внутривенную инъекцию пуромицин-резистентных ТК-ЭСК (2×106 клеток) и, для облегчения визуализации развития тератом, дополнительно подкожную инъекцию (1×106 клеток) различных клонов ТК-ЭСК, представляющих собой CD45.2/CD45.2 гомозиготы. По прошествии 6-10 дней, в течение которых, как ожидалось, основная масса ТК-ЭСК дифференцировалась в клетки крови, проводился 5-7-дневный курс внутримышечных инъекции ганцикловира (10 мг/кг веса тела). Спустя еще 6 недель, (а) регистрировалось наличие тератом в месте введения ТК-ЭСК, (б) проводился анализ клеток крови с использованием методов проточной цитофлуориметрии, имевшего целью обнаружение среди клеток крови потомков донорских ТК-ЭСК (генотип CD45.2/CD45.2) клеток. Во-первых, все облученные реципиенты (n=2), не получившие инъекции ТК-ЭСК, как и ожидалось, погибали на 8-14 день после облучения. Во-вторых, у всех реципиентов (n=5), получивших инъекции ТК-ЭСК клеток, но не прошедших курс ганцикловира (Г-), развились опухоли в месте подкожного введения, и, что обнаруживалось после вскрытия, в других местах (щитовидная железа, костный мозг, легкие, и т.д.). Развитие тератом явилось причиной гибели Г- реципиентов к 6-й неделе после облучения. В то же время, у всех реципиентов (n=7), получивших инъекцию ТК-ЭСК клеток и ганцикловира (Г+), тератомы не развивались. Анализ клеток крови с использованием методов проточной цитофлуориметрии показал значительный (до 40%) вклад ТК-ЭСК как в миелоидные, так и в лимфоидные типы клеток реципиентов как у Г-, так и у Г+ реципиентов, причем у последних - без каких-либо видимых тератомных или иных осложнений (Фиг.2). Таким образом, проведенный эксперимент демонстрирует клиническое приложение изобретения, суть которого заключается в безопасном (безопухолевом) и прямом (не нуждающемся в дорогостоящей, длительной и неэффективной предварительной дифференцировке в культуре), применении недифференцированных плюрипотентных СК для заместительной терапии.
Новизна настоящего изобретения заключается в том, что недифференцированные плюрипотентные СК, несущие описанный выше ген-самоубийцу (например, ТК) и дополнительно, маркер для положительного отбора клеток (например, пуромицин), вводят, без какой-либо предварительной дифференцировки в культуре, в кроветок, орган, или ткань реципиентов, для восполнения клеточной массы или для исправления паракринной, сократительной, нейротрансмиттерной или иной физиологической функции. После определенного инкубационного периода (1-14 дней), в ходе которых введенные плюрипотентные СК колонизируют поврежденный орган, ткань, и самостоятельно дифференцируются в необходимый тип клеток под воздействием микроокружения, реципиентам вводится орально, внутримышечно или внутривенно соответстующий типу гена-самоубийцы препарат (в случае использования ТК применяется известный противовирусный препарат ганцикловир). Экспрессия гена-самоубийцы во всех резидуальных недифференцированных плюрипотентных СК (что обеспечивается предварительной селекцией по резистентности к пуромицину) придает этим клеткам чувствительность к ганцикловиру, что приводит к их апоптотической гибели, тем самым препяствуя возникновению тератом или остановке их роста на ранних стадиях.
Изобретение может найти важное применение при лечении заболеваний, таких как сахарный диабет (путем восполнения пула инсулин-секретирующих бета-клеток поджелудочной железы), печеночной недостаточности (восполнение пула гепатоцитов), последствий инфарктов (восстановление и васкуляризация сердечной мышцы), нейродегенеративных заболеваний и спинномозговых повреждений (восстановление нейрональной ткани), синдрома Дюшена (восстановление скелетномышечной мускулатуры) и т.д.


Изобретение относится к биотехнологии. Плюрипотентные стволовые клетки генетически модифицируют. Затем проводят абляцию резидуальных (остаточных) недифференцированных стволовых клеток противовирусным препаратом. При этом генетическая модификация недифференцированных стволовых клеток проводится бицистронной кассетой, содержащей ген-самоубийцу, под контролем регуляторного элемента, специфического для плюрипотентных стволовых клеток, содержащей и ген устойчивости к антибиотику, также находящийся под контролем указанного регуляторного элемента. Модифицированные клетки отбирают в присутствии соответствующего антибиотика как после модификации, так и непосредственно перед введением реципиенту. Затем отобранные клетки вводят в организм реципиента напрямую, без предварительной дифференцировки in vitro, после чего проводят курс терапии препаратом, активирующим ген-самоубийцу у недифференцированных клеток. Изобретение позволяет клинически эффективно восполнить клеточную массу поврежденного органа или ткани и сократить длительность периода ожидания для потенциальных реципиентов за счет отсутствия этапа дифференцировки стволовых клеток in vitro перед трансплантацией. При этом риск развития у реципиентов тератом после трансплантаций клеточного материала сводится к нулю. 2 ил., 1 табл.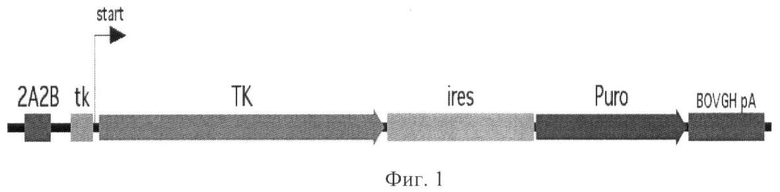
Способ использования стволовых клеток в тканезаместительной терапии, включающий использование плюрипотентных стволовых клеток, генетическую модификацию этих клеток, а затем абляцию резидуальных (остаточных) недифференцированных стволовых клеток противовирусным препаратом, отличающийся тем, что генетическую модификацию недифференцированных стволовых клеток проводят бицистронной кассетой, содержащей ген-самоубийцу, под контролем регуляторного элемента, специфического для плюрипотентных стволовых клеток, содержащей и ген устойчивости к антибиотику, также находящийся под контролем указанного регуляторного элемента, модифицированные клетки отбирают в присутствии соответствующего антибиотика как после модификации, так и непосредственно перед введением реципиенту, а затем отобранные клетки вводят в организм реципиента напрямую, без предварительной дифференцировки in vitro, после чего проводят курс терапии препаратом, активирующим ген-самоубийцу у недифференцированных клеток.
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| СПОСОБ ПОЛУЧЕНИЯ ЭНДОТЕЛИАЛЬНЫХ КЛЕТОК ИЗ ЭМБРИОНАЛЬНЫХ СТВОЛОВЫХ КЛЕТОК ЧЕЛОВЕКА (ВАРИАНТЫ) | 2008 |
|
RU2359030C1 |

