Связанные заявки
Эта заявка относится и основана на заявке на патент США №11/340 661, поданной 27 января 2006 г.
Область техники, к которой относится изобретение
Данная заявка относится к слитым белкам человеческого эритропоэтина.
Уровень техники
Человеческий эритропоэтин (ЕРО), представитель семейства факторов роста системы эритропоэза, синтезируется в основном в почках взрослых и в печени плода в ответ на гипоксию ткани вследствие сниженной доступности кислорода крови [1]. Принципиальная функция ЕРО состоит в том, чтобы действовать непосредственно на определенных прародителей и предшественников эритроцитов (red blood cells, RBC) в костном мозге, чтобы стимулировать синтез гемоглобина и зрелых RBC. Он также регулирует пролиферацию, дифференциацию и созревание RBC. Был получен рекомбинантный ЕРО, имеющий аминокислотную последовательность природного ЕРО, который был разрешен к применению для лечения связанной с нарушением функции почек анемии, рака и других патологических состояний [2]. Результаты недавних исследований [3] показали, что ЕРО, помимо связанных с эритропоэзом свойств, действует также на клетки не из костного мозга, такие как нейроны, что позволяет предположить наличие у ЕРО других возможных физиологических/патологических функций в центральной нервной системе (ЦНС) и других органах/системах. Поскольку рецепторы ЕРО были обнаружены во многих различных органах, ЕРО может иметь многие биологические функции, такие как действие в качестве антиапоптозного агента.
Человеческий ЕРО представляет собой гликопротеин с молекулярным весом 30,4 кДа. Углеводы составляют приблизительно 39% его общей массы. Ген ЕРО расположен в хромосоме 7q11-22 и занимает область длиной 5,4 тысяч пар нуклеотидов (тыс.п.н.) с 5 экзонами и 4 интронами [4]. Предшественник ЕРО состоит из 193 аминокислот. Отщепление лидерной последовательности и последней аминокислоты Arg в ходе пост-трансляционной модификации дает зрелый ЕРО, имеющий 165 аминокислот. Гликозилирование, с тремя N-сайтами в положениях Asn 24, Asn38, Asn83 и одним O-сайтом в положении Ser-126, играет решающую роль в биосинтезе, образовании третичной структуры и биологической активности in vivo ЕРО [5]. ЕРО функционирует, связываясь с рецептором эритропоэтина - гликозилированным и фосфорилированным трансмембранным полипептидом с молекулярным весом 72-78 кДа. Это связывание запускает гомодимеризацию рецепторов, которая приводит к активации нескольких путей передачи сигнала: системы JAK2/STAT5, белка G, кальциевого канала и киназ. Чтобы достигнуть оптимальной активации рецептора, необходимо одновременное связывание двух молекул белка ЕРО с одной молекулой рецептора [6].
Как первый фактор роста системы гематопоэза, разрешенный для лечения людей, рекомбинантный человеческий ЕРО (rHuEPO) использовали для лечения анемии, являющейся результатом хронической почечной недостаточности, рака (первичной анемии, связанной с химиотерапией), аутоиммунных заболеваний, СПИД, хирургических осложнений, трансплантации костного мозга, миелодиспластических синдромов и т.д. Интересно, что недавние исследования показали также, что rHuEPO имеет функции, не связанные с системой крови, и имеет потенциал для использования в качестве нейропротективного лекарства для лечения церебральной ишемии, травм головного мозга, воспалительных заболеваний и нервно-дегенеративных расстройств [7].
В настоящее время в продаже имеются три типа rHuEPO или аналогов rHuEPO, а именно rHuEPO-α, rНuЕРО-β и дарбепоэтин-α([8]. Эти три рекомбинантных белка связываются с одним и тем же рецептором эритропоэтина, но различаются по структуре, степени гликозилирования, сродству в связывании с рецептором и метаболизму in vivo. Co времени начала использования rHuEPO-α в 1980-х гг. клиницисты быстро установили, что существенным недостатком лекарства является необходимость частых инъекций дозы. Средние времена полужизни in vivo rHuEPO-α и rHuEPO-β при внутривенном или подкожном введении составляют соответственно только 8,5 и 17 ч [9, 10]. Поэтому пациентам необходимо производить инъекции ежедневно, дважды в неделю или три раза в неделю, что создает трудности как для пациентов, так и для работников здравоохранения. Поэтому имеется давняя потребность в разработке рекомбинантных аналогов ЕРО, имеющих более долгое время полужизни in vivo и/или повышенную активность в эритропоэзе.
В предшествующей практике делались попытки генетически изменить или химически модифицировать структуру нативного белка ЕРО, чтобы либо замедлить его метаболизм in vivo, либо улучшить его терапевтические свойства. Например, оказывается, что имеется прямая корреляция между количеством в молекуле ЕРО углеводов, содержащих сиаловые кислоты, и его метаболизмом и функциональной активностью in vivo. Поэтому повышение содержания углеводов в молекуле ЕРО приводит к удлинению времени полужизни и повышению активности in vivo [11, 12]. Фирма Amgen сконструировала аналог rHuEPO дарбепоэтин-α так, чтобы включить в rHuEPO в положении 5N две дополнительные углеводные цепи.
Дарбепоэтин-α известен также как «Новый белок, стимулирующий эритропоэз» (Erythropoiesis Stimulating Protein, NESP), и продается под торговой маркой AranespTM. Дарбепоэтин-α отличается от нативного человеческого ЕРО в пяти положениях аминокислот (Ala30Asn, His32Thr, Pro87Val, Trp88Asn, Pro90Thr), что позволяет присоединить два дополнительных N-присоединенных олигосахарида к остаткам аспарагина в положениях 30 и 88. Дарбепоэтин-α связывается с рецептором ЕРО в идентичной нативному ЕРО манере, индуцируя внутриклеточную сигнальную цепь с участием фосфорилирования тирозина киназой JAK-2 и тех же самых внутриклеточных молекул Ras/MAP-k, P13-K и STAT-5. Благодаря повышенному содержанию углеводов, время полужизни дарбепоэтина-α у животных и человека почти втрое длиннее, чем у rHuEPO-α (25,3 ч против 8,5 ч) [9]. Оказывается также, что дарбепоэтин-α (AranespaTM) имеет повышенную биологическую активность по сравнению с природным или рекомбинантным человеческим ЕРО in vivo [13] и был разрешен к применению Американским фармакологическим комитетом (FDA) как лекарство rHuEPO второго поколения. Это лекарство необходимо вводить только один раз в неделю, чтобы получить такое же терапевтическое действие, как и при инъекциях 2-3 раза в неделю rHuEPO [10, 14, 15].
Делались другие попытки удлинить время полужизни ЕРО, сосредоточенные на повышении молекулярного веса белка ЕРО с помощью химической конъюгации с полиэтиленгликолем («ПЭГилирование») и подобных методов. ПЭГилированный ЕРО имеет намного больший молекулярный вес, защищен от выведения из кровотока и поэтому имеет более длительное время полужизни в плазме [16]. Однако ПЭГилирование может изменять структуру белка, что может привести к нежелательным изменениям функции и специфичности части ЕРО молекулы. Имеются также сведения об увеличении молекулярного веса ЕРО другими методами - такими как связывание молекулы ЕРО с белком-носителем (человеческим альбумином), или путем гомодимеризации двух полных молекул ЕРО с помощью пептидных мостиков (длиной 3-17 аминокислот) или химических сшивающих реагентов [17,18, 19, 20]. Хотя все эти способы привели к некоторому успеху в продлении времени полужизни и повышении активности ЕРО, комбинирование молекулы ЕРО с фрагментом Fc человеческого иммуноглобулина (IgG) в слитом белке, как это описано в настоящей заявке, обеспечивает уникальные преимущества.
Человеческий иммуноглобулин IgG состоит из четырех полипептидов (по две идентичных копии легкой цепи и тяжелой цепи), соединенных ковалентно дисульфидными связями. Протеолиз молекулы IgG папаином дает два фрагмента Fab и один фрагмент Fc. Фрагмент Fc состоит из двух полипептидов, соединенных дисульфидными связями. Каждый полипептид от N-конца к С-концу состоит из шарнирной области, домена СН2 и домена СНЗ. Структура фрагмента Fc почти одинакова у всех подтипов человеческого иммуноглобулина. IgG входит в число самых преобладающих белков человеческой крови и составляет от 70 до 75% всех иммуноглобулинов в человеческой сыворотке. Время полужизни IgG в кровотоке наибольшее среди всех пяти типов иммуноглобулина и может достигать 21 дня.
Современная технология биоинженерии была с успехом применена для создания слитых белков, состоящих из фрагментов терапевтических белков, таких как цитокины и растворимые рецепторы, и фрагмента Fc человеческого IgG [21, 22, 23, 24]. Эти слитые белки имеют значительно большее время полужизни in vivo и при этом сохраняют свои биологические и терапевтические свойства. Поэтому содержащие фрагмент Fc слитые белки были с успехом разработаны как биологические лекарственные вещества и разрешены FDA для лечения ревматоидного артрита и хронического бляшечного псориаза [25, 26].
Предыдущими исследованиями было показано, что димеры двух молекул ЕРО, соединенные либо химической сшивкой, либо полипептидом, обладают повышенной активностью in vivo и продленным временем полужизни [17, 19]. Повышенная активность может быть следствием более эффективного связывания димера ЕРО с одним рецептором, а продленное время полужизни in vivo может быть связано с большим размером белка-димера. Однако процесс химической сшивки мало эффективен и его трудно контролировать. Кроме того, пептидный мостик в димере ЕРО может изменять трехмерную структуру молекулы ЕРО, а сам пептид может стимулировать иммуногенные реакции in vivo. Эти недостатки снижают терапевтический потенциал димеров ЕРО, особенно для пациентов с расстройством почек, у которых заместительная терапия ЕРО длится всю жизнь.
Поэтому возникла потребность в аналогах ЕРО, имеющих продленное время полужизни и повышенную эритропоэтическую активность in vivo, но не обладающих усиленными иммуногенными свойствами.
Сущность изобретения
В соответствии с изобретением описан рекомбинантный слитый белок, содержащий эритропоэтиновую пептидную часть из пептида человеческого эритропоэтина, присоединенную к иммуноглобулиновой пептидной части. Слитый белок имеет продленное время полужизни in vivo по сравнению с природным или рекомбинантным нативным человеческим эритопоэтином. В одном из вариантов осуществления изобретения белок имеет время полужизни in vivo, по меньшей мере в 3 раза превышающее время полужизни нативного человеческого эритропоэтина. Слитый белок может также обладать повышенной эритропоэтической биологической активностью по сравнению с нативным человеческим эритропоэтином.
В одном из вариантов осуществления изобретения иммуноглобулиновая пептидная часть является фрагментом Fc, таким как фрагмент lgG1. Фрагмент Fc включает домены СН2 и СН3 и шарнирную область. ЕРО пептидная часть может быть непосредственно присоединена к шарнирной области. Предпочтительно шарнирная область имеет длину по меньшей мере 9 аминокислот. В одном из вариантов осуществления изобретения ЕРО пептидная часть имеет остаток цистеина, ближайший к его С-концу, а шарнирная область включает остаток цистеина, расположенный ближе всего к ЕРО пептидной части. Предпочтительно эти два остатка цистеина разделены между собой по меньшей мере 12 аминокислотами. В одном из вариантов осуществления изобретения ЕРО пептидная часть может представлять собой полную молекулу ЕРО, непосредственно присоединенную к иммуноглобулиновой части (то есть между ЕРО и иммуноглобулиновой частями нет никаких чужеродных пептидных мостиков).
Изобретение относится также к мультимерным белковым конструкциям, содержащим несколько единиц слитого белка согласно настоящему изобретению.
Например, два слитых белка могут быть смонтированы в виде димера, причем шарнирные области белков соединены дисульфидными связями. Димер имеет общую форму молекулы IgG и более стабилен, чем свободные молекулы ЕРО.
Изобретение относится также к нуклеиновой кислоте, кодирующей слитый белок, к соответствующей аминокислотной последовательности и к линиям трансфицированных клеток и способам получения слитого белка. Далее изобретение включает фармацевтические композиции, содержащие слитый белок, и способы использования слитого белка и/или фармацевтических композиций, например, для стимулирования эритропоэза у нуждающихся в лечении индивидуумов.
Краткое описание фигур
Подразумевается, что фигуры, иллюстрирующие различные варианты осуществления изобретения, не должны рассматриваться в ограничивающем смысле.
Фиг.1А - схематическая диаграмма, показывающая общую структуру рекомбинантного человеческого слитого белка EPO-Fc (rHuEPO-Fc) согласно настоящему изобретению.
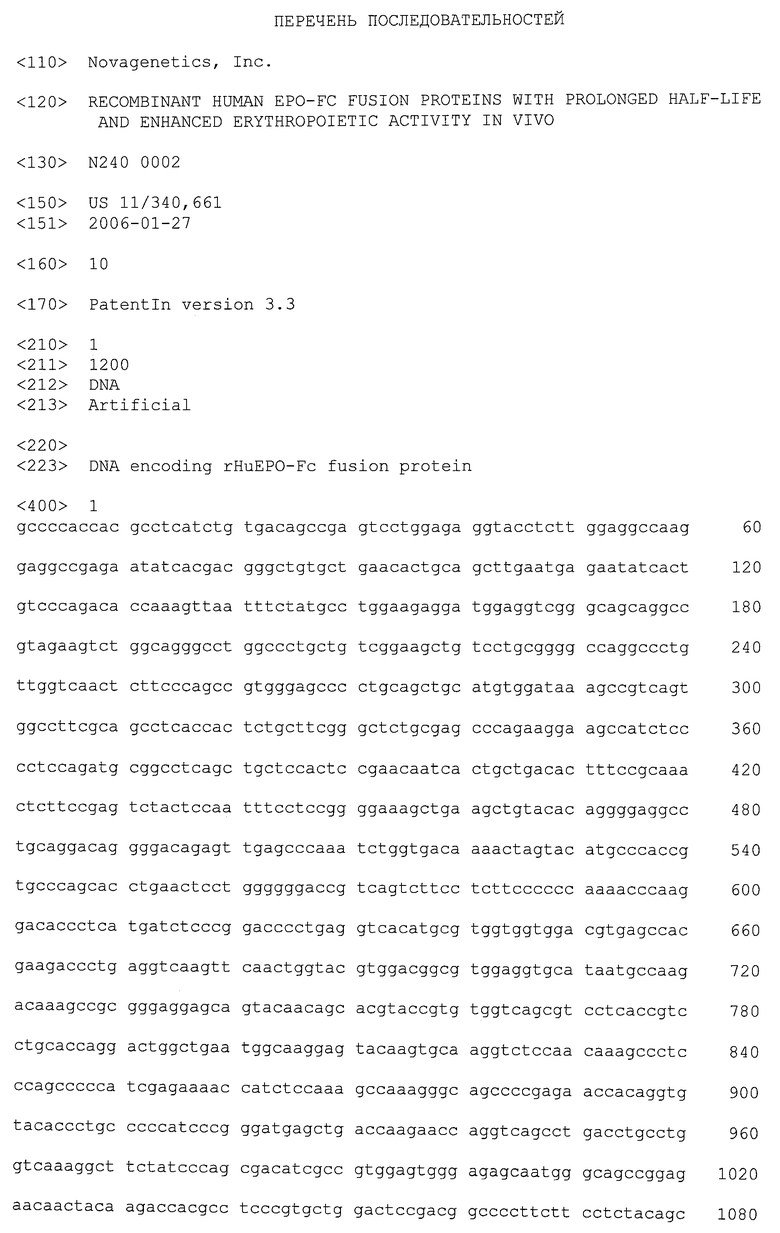
Фиг.1Б - список последовательностей, показывающий нуклеотидную последовательность и выведенную из нее аминокислотную последовательность белка rHuEPO-Fc. Полная длина ДНК равна 1281 п.н. В выведенной белковой последовательности 426 аминокислот (ак) включают 27 ак для сигнального пептида и 399 ак для полного белка rHuEPO-Fc. Полный белок rHuEPO-Fc состоит из домена человеческого ЕРО (166 ак), шарнирной области (16 ак, подчеркнуты) и доменов СН2 и СН3 (217 ак) фрагмента Fc человеческого lgG1. Рассчитанный молекулярный вес полипептида зрелого слитого белка rHuEPO-Fc равен 44,6 кДа, часть которого 18,5 кДа (41,4%) относится к фрагменту ЕРО, а 26,1 кДа (58,6%) - это фрагмент Fc lgG1. Гомодимер образован с помощью двух дисульфидных связей с участием двух остатков цистеина (заключены в рамку) в шарнирной области. В положении 172 зрелого слитого белка (то есть в 6-й аминокислоте шарнирной области) нативный остаток цистеина был замещен глицином (выделен жирным шрифтом).
Фиг.2 - схематическая диаграмма, показывающая структуру и особенности экспрессирующей плазмиды млекопитающих pCD1, использованной для введения последовательности ДНК, кодирующей полипептид слитого белка rHuEPO-Fc, и для трансфекции клеток СНО, экспрессирующих слитый белок rHuEPO-Fc.
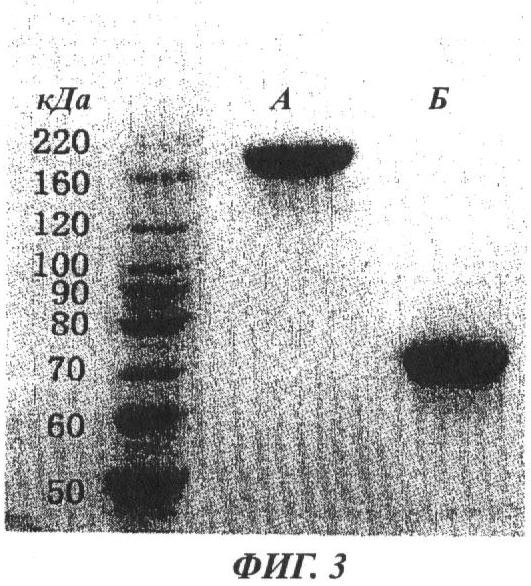
Фиг.3 - изображение зон после электрофореза в полиакриламидном геле с додецилсульфатом натрия (SDS-PAGE), показывающее установленные анализом в SDS-PAGE размеры димерной формы чистого белка rHuEPO-Fc в условиях отсутствия восстановления и мономерной формы чистого белка rHuEPO-Fc при восстановлении. Очищенный белок rHuEPO-Fc из надосадочных жидкостей культивированной линии клеток СНО, экспрессирующих rHuEPO-FC, существует в основном в виде димерной формы и имеет молекулярный вес приблизительно 180 кДа в 8% геле в буфере бис-трис без восстановления. При восстановлении (100 мМ дитиотреитол, DTT), разрушающем дисульфидные связи, димер распадается на 2 идентичные мономерные единицы с молекулярным весом 75 кДа.
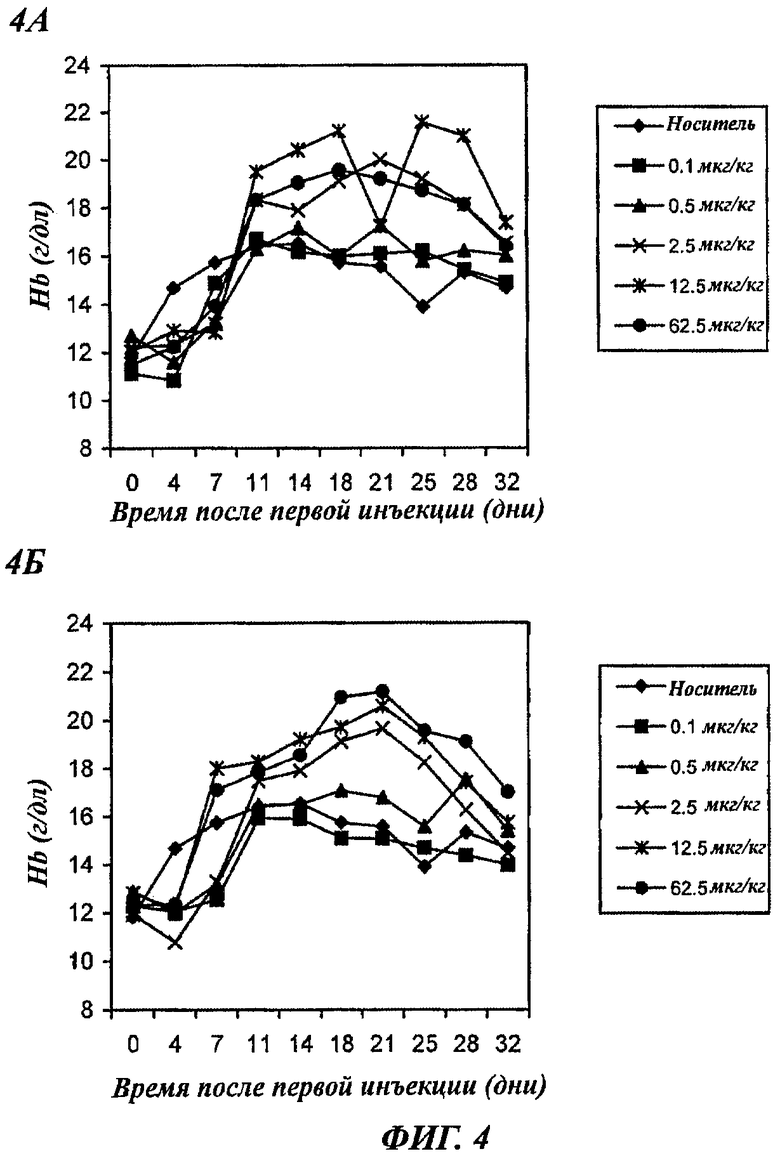
Фигуры 4А и 4Б - графики, показывающие зависящее от дозы повышение уровней содержания гемоглобина (Hb) у нормальных мышей, получавших 3 раза в неделю подкожную (п/к) инъекцию rHuEPO-Fc или rHuEPO. Каждая точка представляет средний уровень Hb в группе (6 мышей). Уровни в день 0 представляют уровни Hb перед воздействием. А: мыши получали rHuEPO-Fc. Б: мыши получали нативный rHuEPO.
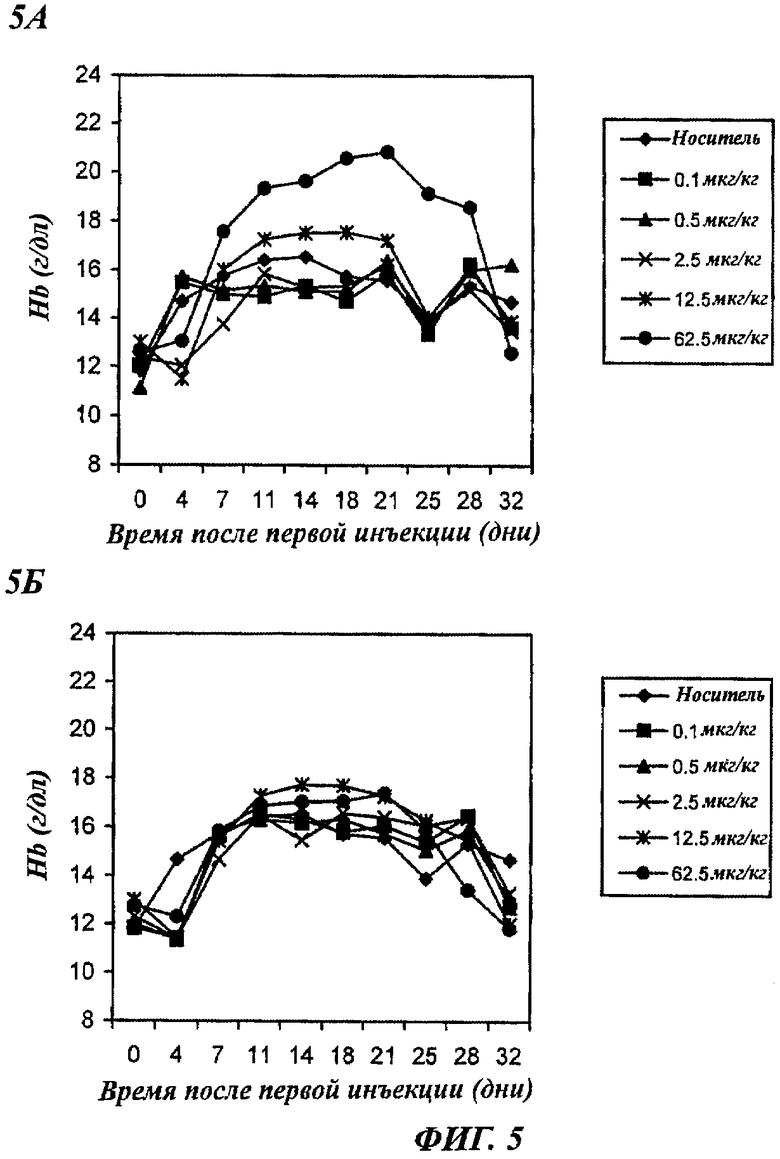
Фигуры 5А и 5Б - графики, показывающие зависящее от дозы повышение уровней содержания гемоглобина (Hb) у нормальных мышей, получавших 1 раз в неделю п/к rHuEPO-Fc или rHuEPO. Каждая точка представляет средний уровень Hb в группе (6 мышей). Уровни в день 0 представляют уровни Hb перед воздействием. А: мыши получали rHuEPO-Fc. Б: мыши получали нативный rHuEPO.
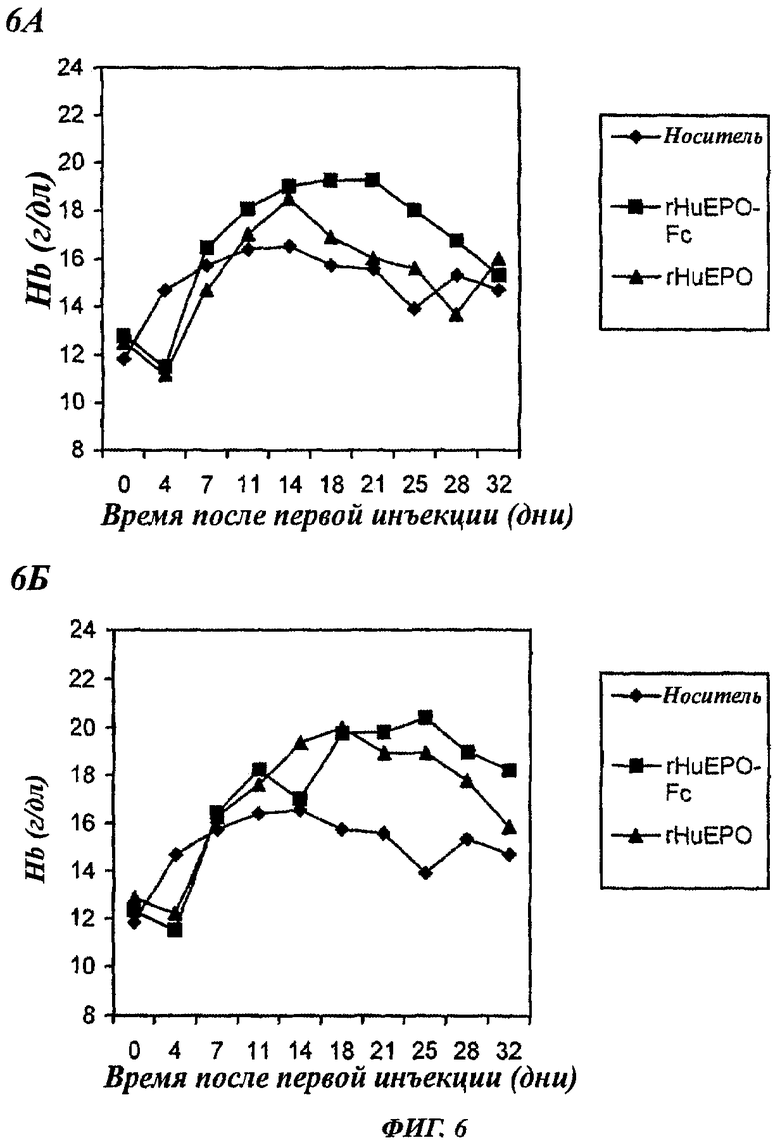
Фигуры 6А и 6Б - графики, показывающие повышение уровней содержания гемоглобина (Hb) у нормальных мышей, получавших внутривенную (в/в) инъекцию 12,5 мкг/кг rHuEPO-Fc или rHuEPO. Каждая точка представляет средний уровень Hb в группе (6 мышей). Уровни в день 0 представляют уровни Hb перед воздействием. А: мыши получали инъекцию 1 раз в неделю. Б: мыши получали инъекцию 3 раза в неделю.
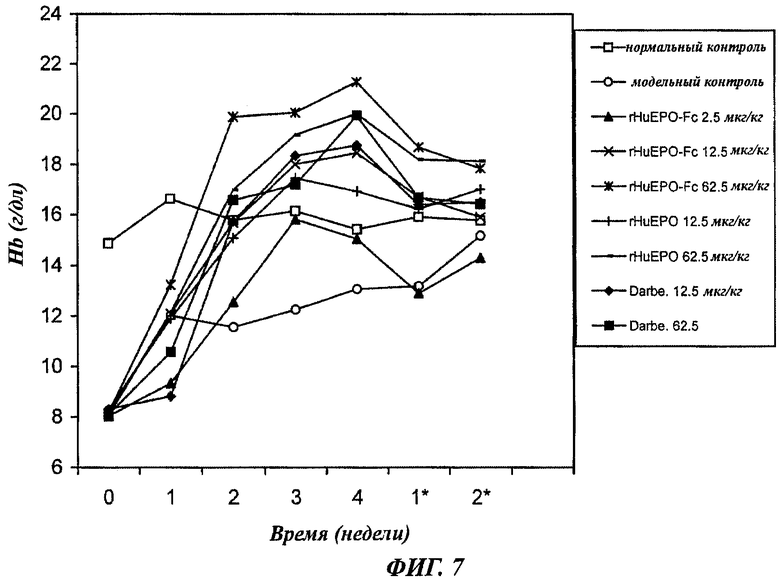
Фиг.7 - график, показывающий зависящее от дозы повышение уровней содержания гемоглобина (Hb) у крыс с нефроэктомией 5/6, получавших п/к 1 раз в неделю rHuEPO-Fc, rHuEPO или дарбопоэтин-α (обозначение Darbe). Каждая точка представляет средний уровень Hb в группе. Нормальный контроль представлен нормальными крысами с инъекцией раствора носителя. Модельный контроль - это крысы с нефроэктомией 5/6, получавшие инъекции раствора носителя. Уровни в неделю 0 представляют уровни Hb до начала воздействия. Звездочкой * отмечены недели после воздействия.
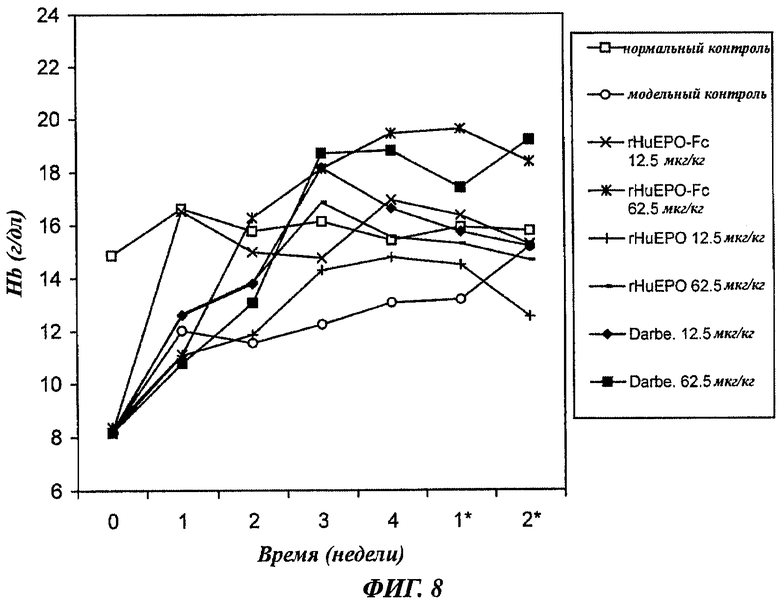
Фиг.8 - график, показывающий зависящее от дозы повышение уровней содержания гемоглобина (Hb) у крыс с нефроэктомией 5/6, получавших п/к 1 раз в 2 недели rHuEPO-Fc, rHuEPO или дарбопоэтин-α (обозначение Darbe). Каждая точка представляет средний уровень Hb в группе. Нормальный контроль представлен нормальными крысами с инъекцией раствора носителя. Модельный контроль - это крысы с нефроэктомией 5/6, получавшие инъекции раствора носителя. Уровни в неделю 0 представляют уровни Hb до начала воздействия. Звездочкой * отмечены недели после воздействия.
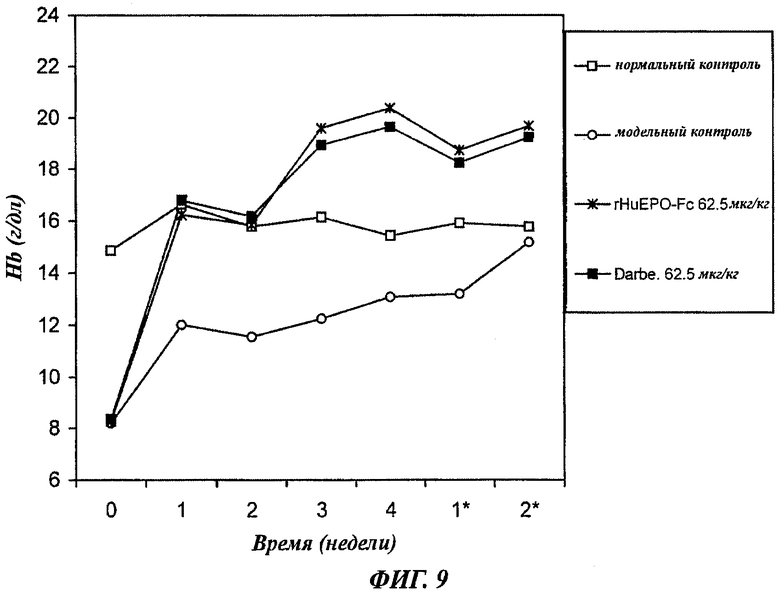
Фиг.9 - график, показывающий зависящее от дозы повышение уровней содержания гемоглобина (Hb) у крыс с нефроэктомией 5/6, получавших в/в 1 раз в 2 недели 62,5 мкг/кг rHuEPO-Fc, rHuEPO или дарбопоэтина-α (обозначение Darbe). Каждая точка представляет средний уровень Hb в группе. Нормальный контроль представлен нормальными крысами с инъекцией раствора носителя. Модельный контроль - это крысы с нефроэктомией 5/6, получавшие инъекции раствора носителя. Уровни в неделю 0 представляют уровни Hb до начала воздействия. Звездочкой * отмечены недели после воздействия.
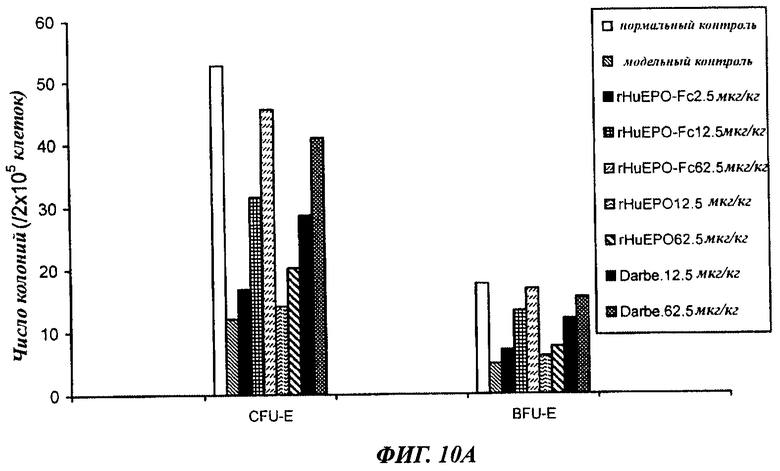
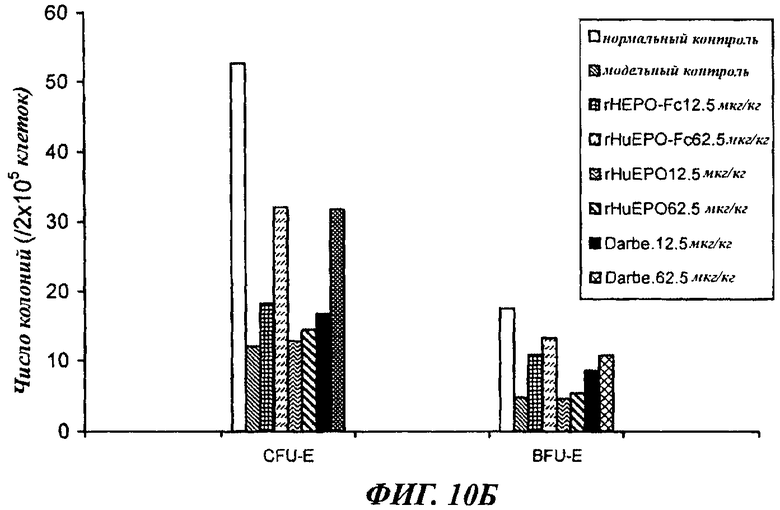
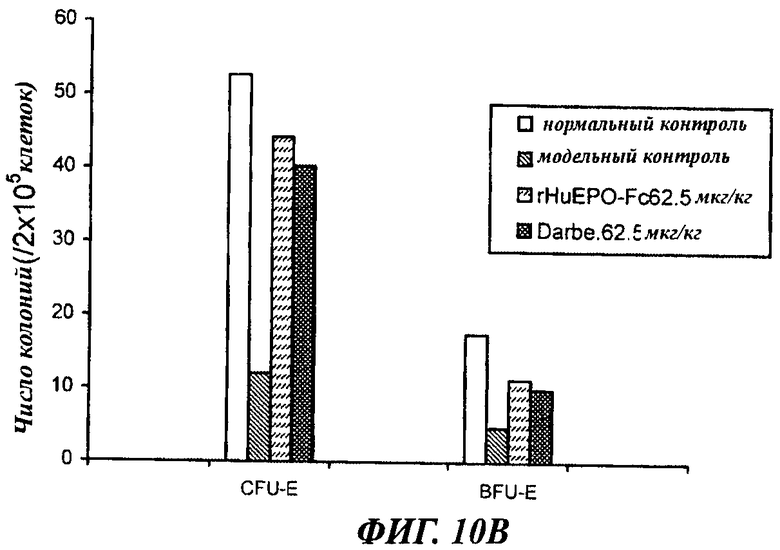
Фигуры с 10А по 10Б показывают сравнение активности rHuEPO-Fc, rHuEPO и дарбопоэтина-α в стимуляции образования колоний CFU-E и BFU-E у крыс с нефроэктомией 5/6, которым вводили разные дозы препаратов в разных режимах. Введение rHuEPO-Fc и дарбопоэтина-α (обозначение Darbe) выявило их одинаковую зависящую от дозы активность в образования колоний CFU-E и BFU-E, а rHuEPO был менее активен. А: п/к введение 1 раз в неделю. Б: п/к введение 1 раз в 2 недели. В: в/в введение 1 раз в 2 недели.
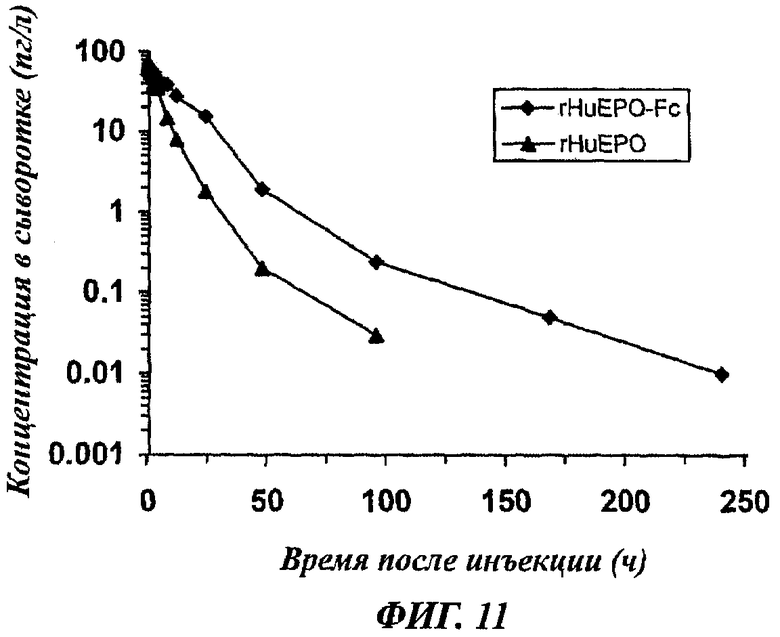
Фиг.11 - график, показывающий уровни содержания в сыворотке rHuEPO-Fc и rHuEPO после внутривенной инъекции обезьянам резус 5 мкг/кг rHuEPO-Fc или rHuEPO (средние уровни у 5 обезьян).
Фиг.12 - список последовательностей, показывающий нуклеотидную последовательность и выведенную аминокислотную последовательность белка rHuEPO-FcC дикого типа. Детали последовательностей такие же, как показаны на фиг.1, за исключением того, что в положении 172 зрелого слитого белка (т.е. в положении аминокислоты 6 шарнирной области) указан остаток цистеина, соответствующий нативному белку дикого типа.
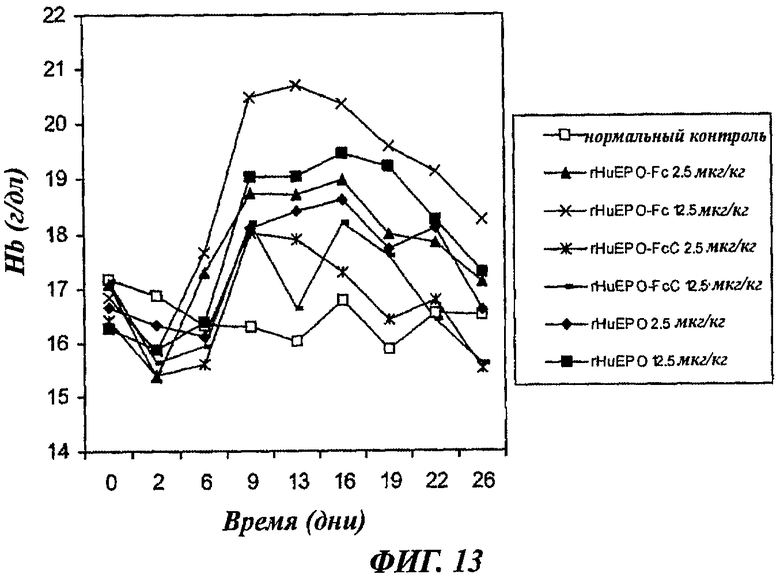
Фиг.13 - график, показывающий зависящее от дозы повышение уровней содержания гемоглобина (Hb) у нормальных мышей, получавших 3 раза в неделю п/к инъекцию rHuEPO-Fc (мутантный слитый белок согласно настоящему изобретению), rHuEPO- FcC (слитый белок дикого типа) и rHuEPO. Каждая точка представляет средний уровень Hb в группе (8 мышей). Нормальный контроль представлен нормальными мышами, которым вводили раствор носителя. День 0 представляет уровни Hb до воздействия.
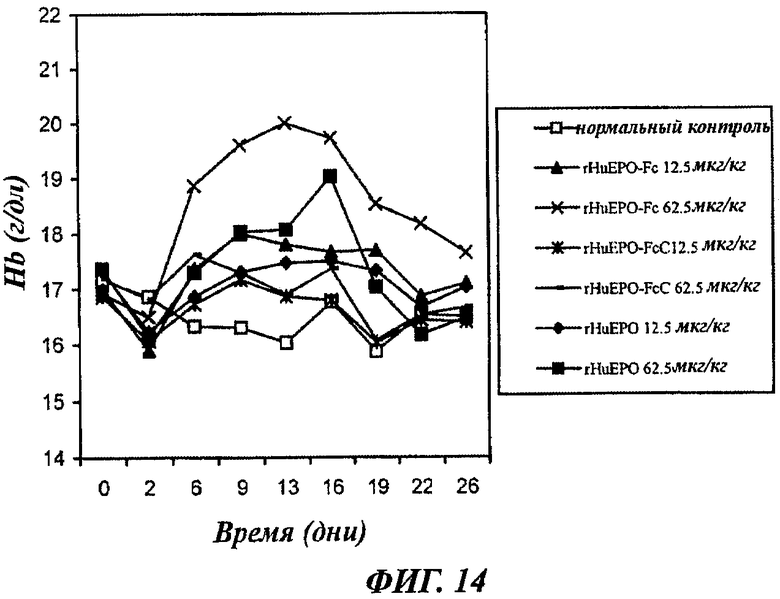
Фиг.14 - график, показывающий зависящее от дозы повышение уровней содержания гемоглобина (Hb) у нормальных мышей, получавших 1 раз в неделю п/к инъекцию rHuEPO-Fc, rHuEPO-FcC и rHuEPO. Каждая точка представляет средний уровень Hb в группе (8 мышей). Нормальный контроль представлен нормальными мышами, которым вводили раствор носителя. День 0 представляет уровни Hb до воздействия.
Детальное описание изобретения
На протяжении последующего описания для лучшего понимания изобретения приводятся конкретные детали. Однако изобретение может быть осуществлено без этих деталей. В других случаях хорошо известные элементы не были показаны или подробно описаны, чтобы избежать нагромождения деталей, которое помешает понять настоящее изобретение. Поэтому описание и фигуры следует рассматривать в иллюстративном, а не в ограничивающем смысле.
Эта заявка относится к новому слитому белку, имеющему эритропоэтические свойства. Слитый белок, обозначенный здесь как rHuEPO-Fc, содержит молекулу человеческого эритропоэтина (ЕРО) с помощью рекомбинантной технологии соединенную с фрагментом Fc иммуноглобулина. Как обсуждается далее, слитый белок может быть в форме димера, состоящего из двух идентичных полипептидных субъединиц. В варианте осуществления изобретения, схематически представленном на фиг.1А, каждая полипептидная субъединица от N-конца к С-концу состоит из полипептидной последовательности молекулы человеческого ЕРО и полипептидной последовательности шарнирной области, домена СН2 и домена СН3 фрагмента Fc человеческого иммуноглобулина lgG1. Две полипептидные субъединицы для образования димерной структуры соединены друг с другом дисульфидными связями между соответственными шарнирными областями. Поэтому димер имеет такую же общую форму, как и молекула IgG, и обладает лучшей стабильностью, чем свободные молекулы ЕРО, что обсуждается далее в примерах.
Как будет очевидно для специалиста в данной области, шарнирная область интактного иммуноглобулина придает белку достаточную гибкость для эффективного связывания антиген-антитело. Подобным же образом в настоящем изобретении шарнирная область включена в конструирование слитого белка rHuEPO-Fc, чтобы сохранить его гибкость, особенно когда слитый белок находится в димерной форме. Как описано далее, это обеспечивает нормальное связывание части ЕРО слитого белка rHuEPO-Fc с рецепторами ЕРО для активации биологических функций ЕРО. Можно с уверенностью считать, что димерная форма слитого белка rHuEPO-FC, предоставляющая две молекулы ЕРО, способна индуцировать оптимальную активацию рецепторов ЕРО (например, способствуя сшивке рецепторов).
Как показано в приведенных далее примерах, слитый белок rHuEPO-Fc был успешно синтезирован с помощью техники рекомбинантной ДНК. В исследованиях на мышах, крысах и приматах было установлено, что слитый белок имеет продленное время полужизни in vivo и повышенную эритропоэтическую активность по сравнению с природным или рекомбинантным нативным человеческим ЕРО. Термины «нативный человеческий эритропоэтин» и «нативный человеческий ЕРО», как они использованы в данной патентной заявке, обозначают ЕРО, имеющий немодифицированную структуру дикого типа. Как понятно специалисту в данной области, нативный человеческий ЕРО может быть природным или полученным по рекомбинантной технологии (например rHuEPOα). Термин «нативный человеческий ЕРО» не включает аналоги rHuEPO, такие как дарбепоэтин-α, где структура ЕРО была существенно модифицирована, как например путем гипергликозилирования.
Последовательность нуклеиновой кислоты для слитого белка rHuEPO-Fc согласно настоящему изобретению представлена как SEQ ID No. 1. Соответствующая выведенная аминокислотная последовательность представлена в SEQ ID No. 2. Длина полного слитого белка rHuEPO-Fc составляет 399 аминокислот. Как показано на фиг.1Б, полный слитый белок rHuEPO-Fc состоит из домена ЕРО (166 аминокислот), шарнирной области (16 аминокислот, подчеркнуты) и доменов СН2 и СН3 (217 аминокислот). Сигнальная и лидерная пептидная последовательность, состоящая из 27 аминокислот, также показана на фиг.1Б. Сигнальный пептид отщепляется в ходе синтеза rHuEPO-Fc. Последовательность нуклеиновой кислоты и аминокислотная последовательность rHuEPO-Fc, включая сигнальный или лидерный пептид, показана соответственно в SEQ ID No. 3 и SEQ ID No. 4.
Как видно лучше всего на фиг.1Б и в последовательности SEQ ID No. 2, домен ЕРО содержит остаток цистеина вблизи его С-конца в положении 161 аминокислотной последовательности. Шарнирная область включает 2 остатка цистеина в положениях 178 и 181 аминокислотной последовательности, которые на фиг.1Б заключены в рамку. Остатки цистеина шарнирной области образуют дисульфидные связи между полипептидными субъединицами гомодимера, как обсуждалось выше. Природная шарнирная область фрагмента человеческого lgG1 также имеет остаток цистеина в положении 6 части шарнирной области (отсчитанном от N-конца). В настоящем изобретении остаток цистеина 6 в шарнирной области был заменен нецистеиновым остатком. В частности, в варианте осуществления изобретения, представленном на фиг.1Б и в последовательности SEQ ID No. 2, аминокислота цистеин была заменена глицином (в положении 172 аминокислотной последовательности rHuEPO-Fc, которое соответствует остатку 6 шарнирной области). Как будет очевидно для специалиста в данной области, цистеин в этом положении может быть также замещен другими нецистеиновыми остатками, чтобы предотвратить образование дисульфидной связи.
В результате аминокислотной замены в положении 172 первый остаток цистеина шарнирной области (в положении 178) отделен 17 аминокислотами от описанного выше остатка цистеина домена ЕРО (в положении 161). Заявители уверены, что минимальное расстояние между остатком цистеина 161 домена ЕРО и первым остатком цистеина шарнирной области должно составлять по меньшей мере 12 аминокислот, чтобы обеспечить успешную сборку и/или связывание с рецептором ЕРО гомодимера rHuEPO-Fc. To есть, если остаток в положении 172 - это остаток цистеина, то возможно образование нежелательной дисульфидной связи, такой как между остатками цистеина 161 и 172. Это может изменить трехмерную структуру молекулы ЕРО, что приведет к исчезновению или снижению биологической активности.
В одном из вариантов осуществления настоящего изобретения домен ЕРО присоединен непосредственно к части, соответствующей фрагменту Fc слитого белка. Исключение чужеродного пептида-мостика сохраняет предпочтительную трехмерную структуру слитого пептида rHuEPO-Fc и сводит к минимуму запуск нежелательной иммуногенной реакции. Шарнирная область фрагмента Fc предпочтительно имеет длину по меньшей мере 9 аминокислот и предпочтительно его длина составляет приблизительно 10-20 аминокислот.
Следующие примеры будут дополнительно иллюстрировать изобретение более детально, хотя следует признать, что изобретение не ограничено конкретными примерами.
Примеры
Пример 1. Конструирование рекомбинантой плазмиды pCdEpo-Fc. кодирующей слитый белок HuEPO-Fc
Молекулу ДНК полной длины, которая кодирует аминокислотную последовательность полипептида rHuEPO-Fc, создавали путем перекрывающей полимеразной цепной реакции (ПЦР) с использованием следующих олигонуклеотидных праймеров (QIAGEN Inc., US):

Последовательности указанных выше праймеров представлены соответственно в последовательностях с SEQ ID No. 5 по SEQ ID No. 8.
Сайты рестрикции EcoR I и Not I ввели соответственно в EF5 и EF3.
Для оптимальной экспрессии белка HuEPO-Fc в клетках млекопитающих в EF5 ввели также последовательность Kozak (GCCACCATGG). EFL5 и EFL3 - это комплементарные последовательности, состоящие из 3'-концевой последовательности ДНК ЕРО (22 п.н.) и 5'-концевой последовательности ДНК для шарнира lgG1 (22 п.н.).
Сначала фрагмент ДНК ЕРО размером 0,6 тыс.нуклеотидов амплифицировали в ПЦР (Platinum Taq DNA Polymerase High Fidelity) с праймерами EF5 и EFL3 из плазмиды р9Е, содержащей кДНК полной длины для человеческого ЕРО, а фрагмент Fc размером 0,7 тыс.нуклеотидов амплифицировали с праймерами EF3 и EFL5 с плазмиды pD, содержащей последовательность кДНК полной длины для человеческого lgG1 (р9Е и pD были получены в лаборатории заявителей). Затем оба фрагмента очищали и смешивали в равных количествах. Используя смесь в качестве матрицы, ДНК rHuEPO-Fc полной длины размером 1,3 тыс.нуклеотидов амплифицировали с праймерами EF5 и EF3. Очищенный фрагмент длиной 1,3 тыс.нуклеотидов расщепляли рестриктазами EcoR I и Not I (New England Biolab Inc. US) и затем клонировали в расщепленный EcoR I/Not I экспрессирующий вектор млекопитающих pCDI (фиг.2). Полученный рекомбинантный вектор был назван pCdEpo-Fc, введенная в него последовательность нуклеиновой кислоты, кодирующая аминокислотную последовательность HuEPO-Fc, была подтверждена секвенированием ДНК.
Пример 2. Получение линии клеток, экспрессирующей rHuEPO-Fc
В качестве клеток-хозяев для экспрессии rHuEPO-Fc использовали клетки яичников китайского хомячка, дефектные по дигидрофолат-редуктазе (dhfr) (CHO/dhfr-, ATCC No. CRL-9096), которые разрешены FDA для продукции биологических веществ.
Клетки CHO/dhfr- трансфицировали рекомбинантным вектором pCdEpo-Fc с использованием липофектамина (Gibco, Cat. No: 18292-037, USA). Надосадочные жидкости из культур отобранных клонов анализировали методом ELISA (Roche, Cat. No: 1-693417, Canada) для определения активности ЕРО. Далее позитивные клоны подвергали скринингу при повышающейся нагрузке метотрексата (МТХ). Одна линия клеток с наиболее высокой экспрессией белка rHuEPO-Fc была отобрана как экспрессирующая rHuEPO-Fc линия клеток СНО и постепенно адаптирована к среде без сыворотки (CD СНО Medium, Gibco, Cat. No: 10743-029, USA). Эта экспрессирующая rHuEPO-Fc линия клеток СНО была использована для продукции белка rHuEPO-Fc.
Пример 3. Очистка белка rHuEPO-Fc
Молекулы белка rHuEPO-Fc, содержащиеся в надосадочных жидкостях, собранных после культивирования экспрессирующих rHuEPO-Fc клеток СНО в среде без сыворотки, были выделены вначале с помощью аффинной хроматографии на белке A (Amersham, Cat. No: 17-0402- 01, Canada). Выделенные белки были дополнительно очищены гель-хроматографией на колонке HiLoad 16/60 Superdex 200pg (Amersham, Cat. No: 17-1069-01, Canada). Чистота белка rHuEPO-Fc была более 98% на основании электрофореза.
Пример 4. Определение размера чистого белка rHuEPO-Fc
Сначала для определения размера чистого белка rHuEPO-Fc был проведен SDS-PAGE. Как показано на фиг.3, при электрофорезе в 8% геле с бис-трис буфером без восстановления, который позволяет измерить общий размер белка с дисульфидными связями, получена одна зона с молекулярным весом около 180 кДа. Это указывает на то, что основная часть молекул белка rHuEPO-Fc продуцируется в димерной форме, как и ожидалось на основании конструирования слитого белка. Если анализ SDS-PAGE проводили в восстанавливающих условиях (100 мМ DTT) для разрушения дисульфидных связей, была идентифицирована только одна линия с молекулярным весом 75 кДа, что согласуется с оценкой молекулярного веса для одиночной полипептидной цепи состава HuEPO-шарнирная область-СН2-СН3.
Точный молекулярный вес чистого слитого белка rHuEPO-Fc с гликозилированием, определенный масс-спектрометрией (MALDI-TOF-MS), составил 111099 Да (111,1 кДа). В этом анализе наблюдался только один пик белка, что указывает на близкую к 100% чистоту белка rHuEPO-Fc. Анализ последовательности белка позволил определить 15 аминокислот N-конца чистого белка rHuEPO-Fc как APPRLICDSRVLERY. Это согласуется с последовательностью первых 15 аминокислот полипептида нативного человеческого ЕРО и подтверждает, что очищенный белок rHuEPO-Fc действительно имеет правильную и полную последовательность молекулы ЕРО, как предсказано по последовательности ДНК, кодирующей аминокислотные последовательности слитого белка rHuEPO-Fc.
Пример 5. Повышенная эритропоэтическая активность rHuEPO-Fc у нормальных мышей
Для подтверждения сохранения эритропоэтической активности белка rHuEPO-Fc и определения его активности в сравнении с rHuEPO и дарбепоэтином-α были проведены опыты in vivo на мышах. В целях сравнения все дозы трех использованных в описанных опытах на животных ЕРО: rHuEPO-Fc согласно настоящему изобретению, rHuEPO (нативный человеческий ЕРО) и дарбепоэтина-α были выражены количествами части только молекулы ЕРО на основании размера молекулы. Для белка rHuEPO-Fc часть ЕРО составляет 41,4% от полного молекулярного веса rHuEPO-Fc, что вычислено на основании веса аминокислот ЕРО и веса всех аминокислот в целой молекуле rHuEPO-Fc (160 ак из общего числа 399 ак). Поэтому количество ЕРО в rHuEPO-Fc было сочтено равным 41,4% от общего количества белка rHuEPO-Fc.
rHuEPO-Fc (концентрация рабочего раствора 0,5 мг/мл, чистота 98,6%) и нативный человеческий rHuEPO (т.е. со структурой природного человеческого ЕРО) (6000 ед./0,5 мл, продукция фирмы Kirin Brewery Co., Japan) разбавляли в растворе носителя (2,5 мг/мл человеческого сывороточного альбумина, 5,8 мг/мл цитрата натрия, 0,06 мг/мл лимонной кислоты и 5,8 мг/мл хлорида натрия, рН 5,5-5,6). Количественную дозу rHuEPO рассчитывали согласно известному для него соотношению активность/количество. Мыши BALB/c (возраст 6-8 недель, вес 18-22 г, равное количество самцов и самок, получены из Experiment Animal Center, AMMS, China) были случайным образом распределены в группы по 6 мышей в каждой группе. Каждая из групп мышей получала одну комбинацию из одной дозы (0,1, 0,5, 2,5, 12,5, 62,5 мкг/кг), одного пути введения (в/в в хвостовую вену или п/к) и одного режима инъекций (3 раза в неделю или 1 раз в неделю). Контрольной группе мышей вводили равный объем раствора носителя. Воздействие длилось в течение 3 недель, а общее время наблюдения составляло 5 недель. Образцы периферической крови (из хвостовой вены) для измерений отбирали перед началом воздействия, на 4-й и на 7-й день каждой недели в течение 5 недель. Количество Hb измеряли показателем абсорбциометрии. Средние стандартные отклонения рассчитывали для данных каждой группы, для сравнения различных групп использовали r-тест.
Введение ЕРО мышам 3 раза в неделю, при условии, что эти ЕРО имеют нормальную эритропоэтическую активность, будет с насыщением индуцировать стимуляцию эритропоэза. Как показано на фиг.4, в обеих группах, подвергавшихся воздействию 3 раза в неделю, наблюдалось существенное повышение уровней даже при дозе 2,5 мкг/кг. Этот опыт показал, что эритропоэтическая активность rHuEPO-Fc in vivo столь же высока, как и у rHuEPO. Повышение уровней Hb в подвергшихся воздействию группах зависело от дозы. Однако насыщение в повышении уровней Hb достигалось у мышей при дозе rHuEPO-Fc 12,5 мкг/кг, тогда как такое же насыщенное повышение уровней Hb при введении rHuEPO достигалось лишь при дозе 62,5 мкг/кг. Индуцируемое rHuEPO-Fc при дозе 2,5 мкг/кг повышение уровней Hb было также выше, чем повышение при индукции rHuEPO при дозе 2,5 мкг/кг. Эти результаты позволяют предположить более сильную стимуляцию эритропоэза при действии rHuEPO-Fc, чем при действии rHuEPO.
Эритропоэтический потенциал rHuEPO-Fc был далее использован для снижения числа п/к инъекций до 1 в неделю. Как показано на фиг.5, в получавших rHuEPO-Fc группах наблюдалось зависящее от дозы повышение уровней Hb при дозах 12,5 или 62,5 мкг/кг. Обе дозы (12,5 и 62,5 мкг/кг) rHuEPO также индуцировали повышение уровней Hb до близкого значения, которое было намного ниже, чем уровни, индуцируемые 62,5 мкг/кг rHuEPO-Fc. Это несомненно указывает на то, что rHuEPO-Fc имеет повышенную эритропоэтическую активность in vivo. Возможно, это является следствием либо продленного времени полужизни rHuEPO-Fc in vivo, либо усиления связывания/активации рецепторов ЕРО димерами молекул ЕРО в белке rHuEPO-Fc, либо объединенными эффектами обоих факторов.
Если такие же дозы (12,5 мкг/кг) rHuEPO-Fc или rHuEPO вводили внутривенно или 3 раза в неделю, или 1 раз в неделю, во всех подвергнутых воздействию группах наблюдалось повышение уровней Hb (фиг.6). Однако в/в введение rHuEPO-Fc 1 раз в неделю индуцировало более значительное и более стойкое повышение уровней Hb, которое длилось дольше после окончания воздействия. Эти данные дополнительно поддерживают мнение о более высокой эритропоэтической активности белка rHuEPO-Fc по сравнению с rHuEPO, имеющим структуру природного белка ЕРО.
Пример 6. Повышенная эритропоэтическая активность rHuEPO-Fc у 5/6 крыс с нефрэктомией
Опыты на нормальных мышах подтвердили повышенную эритропоэтическую активность rHuEPO-Fc in vivo. Чтобы дополнительно проверить эффективность rHuEPO-Fc в стимуляции эритропоэза, были проведены исследования фармакодинамики у крыс с экспериментальной почечной анемией, полученной путем 5/6 нефрэктомии. Эффективность rHuEPO-Fc сравнивали с эффективностью rHuEPO и дарбепоэтина-α (60 мкг/мл, партия №N079, производство фирмы Kirin Brewery Co., Japan).
В данном изобретении для получения модели анемии вследствие нарушения функции почек путем двухэтапной нефрэктомии [27] использовали крыс Wistar (равное число самцов и самок, вес 160-180 г, получены от Vitalriver Experiment Animal Inc., Beijing, China. Licence No. SCXKI 1-00-0008). У крыс под общим наркозом проводили нефрэктомию 5/6 в двух отдельных операциях в стерильных условиях. После резекции 2/3 левой почки крыс оставляли для восстановления в течение 20 дней. Затем аккуратно удаляли правую почку. Для предотвращения инфекции после каждой операции вводили антибиотики. В конечном итоге были удалены 5/6 почечной ткани. У крыс после нефрэктомии постепенно развивались недостаточность функции почек и анемия. Стабильное состояние анемии у крыс достигалось через 50 дней после нефрэктомии, после чего их распределяли случайным образом по группам (9 крыс в группе) для введения препаратов ЕРО. Каждая группа крыс получала одну комбинацию одной дозы (2,5, 12,5, 62,5 мкг/кг), одного пути введения (в/в через хвостовую вену или п/к) и одного режима инъекций (1 раз в неделю или 1 раз в 2 недели). Контрольной группе и модельной группе крыс вводили равный объем раствора носителя. Воздействие продолжали в течение 4 недель, а общее время наблюдения составляло 6 недель.
Все дозы (2,5, 12,5, 62,5 мкг/кг) rHuEPO-Fc, вводимого подкожно 1 раз в неделю, индуцировали зависящее от дозы повышение уровней Hb по сравнению с модельной контрольной группой, где не проводилось введение ЕРО. Дозы 12,5 и 62,5 мкг/кг rHuEPO или дарбепоэтина, вводимые подкожно 1 раз в неделю, также индуцировали повышение уровней Hb. Повышенные уровни Hb в обеих группах, получавших 12,5 или 62,5 мкг/кг rHuEPO-Fc, были значительно выше, чем таковые в группах, получавших 12,5 или 62,5 мкг/кг rHuEPO. Уровни Hb в группе, получавшей 62,5 мкг/кг rHuEPO-Fc, были также несколько выше, чем уровни в группе, получавшей 62,5 мкг/кг дарбепоэтина. После прекращения воздействия снижение уровней Hb в группе, получавшей 62,5 мкг/кг rHuEPO-Fc, было значительно более медленным, и уровни Hb оставались более высокими до конца наблюдения (2 недели после прекращения воздействия), чем уровни в группах нормального контроля и модельного контроля. Это указывало на более сильную и/или продолжительную стимуляцию эритропоэза (данные суммированы на фиг.7).
Для воздействия подкожными инъекциями 1 раз в 2 недели вводили 3 типа ЕРО только в дозах 12,5 или 62,5 мкг/кг (фиг.8). Доза 12,5 мкг/кг rHuEPO слабо повышала уровни Hb в сравнении с модельной контрольной группой, а в группе, получавшей 62,5 мкг/кг rHuEPO, слабая эритропоэтическая реакция не позволила довести уровни Hb до нормального значения в сравнении с нормальной контрольной группой. Воздействия rHuEPO-Fc или дарбепоэтином при дозах 12,5 или 62,5 мкг/кг индуцировало значительное повышение уровней Hb, которые были выше уровней в нормальной контрольной группе. Это служило доказательством эффективной коррекции анемичного статуса как rHuEPO-Fc, так и дарбепоэтином. Между одинаковыми дозами rHuEPO-Fc и дарбепоэтина не наблюдалось существенных различий в смысле эффективности. Высокая доза 62,5 мкг/кг приводила к устойчивому усилению эритропоэза вплоть до окончания наблюдения (2 недели после завершения воздействия). Это подкрепило предположение, что rHuEPO-Fc и дарбепоэтин обладают свойством длительно стимулировать эритропоэз in vivo, что в свою очередь может быть использовано для снижения частоты введений лекарства пациентам в клинике.
В то время как дарбепоэтин был разрешен к клиническому применению с менее частыми инъекциями для лучшего соблюдения пациентами режима лечения и уменьшения нагрузки для медицинского персонала, приведенные здесь экспериментальные данные весомо указывают на то, что раскрытый в настоящем изобретении rHuEPO-Fc имеет по меньшей мере такие же потенциальные преимущества. Как обсуждалось выше, дарбепоэтин, как мутантный аналог молекулы человеческого ЕРО, содержащий дополнительные сахарные компоненты (повышенное гликозилирование), может привносить повышенный риск индукции иммуногенеза in vivo вследствие измененной трехмерной структуры. Только длительное наблюдение пациентов, подвергнутых лечению дарбепоэтином, может дать решающее заключение относительно иммуногенных рисков дарбепоэтина. В противоположность этому, rHuEPO-Fc, не имеющий модификации части с молекулой ЕРО, имеет содержание углеводов, идентичное или очень близкое к содержанию в нативном человеческом ЕРО. Количество сиаловых кислот в предлагаемом заявителями чистом белке rHuEPO-Fc было около 10,0 ммоль на 1 ммоль ЕРО, что согласуется с опубликованными параметрами для rHuEPO. Часть Fc молекулы rHuEPO-Fc не имеет чужеродных аминокислот и/или чужеродного пептидного мостика и представляет основную структуру человеческого lgG1. Теоретически это не должно приводить к иммунологической реакции. rHuEPO-Fc, если он будет разрешен к клиническому применению, может быть более благоприятен для пациентов, чем обычно применяемые rHuEPO и аналоги ЕРО, особенно для пациентов, нуждающихся в длительном введении препаратов.
При внутривенных инъекциях 1 раз в 2 недели и rHuEPO-Fc, и дарбепоэтин (62,5 мкг/кг) были способны индуцировать идентичное повышение уровней Hb у крыс с почечной анемией намного выше нормальных уровней Hb у крыс в нормальном контроле (фиг.9). Это дополнительно продемонстрировало стойкую стимуляцию эритропоэза rHuEPO-Fc, поскольку эффективность дарбепоэтина была доказана клиническими исследованиями.
Данные, полученные в опытах по культивированию клеток костного мозга, отобранных у крыс с нефрэктомией 5/6 после воздействия (п/к или в/в 1 раз в неделю или 1 раз в 2 недели), показали, что rHuEPO-Fc, rHuEPO и дарбепоэтин все стимулировали образование CFU-E и BFU-E. Активности rHuEPO-Fc и дарбепоэтина были сходными и превосходили активность rHuEPO (фиг.10).
Уровни азота мочевины (BUN) и креатинина в крови были сходными в подвергавшихся воздействию группах и в группе модельного контроля. Во всех подвергшихся воздействию группах уровень железа (Fe) в сыворотке был выше, чем уровень в группе модельного контроля. Патологические обследования показали у всех получавших ЕРО крыс повышенное содержание относящихся к эритроцитам (RBC) клеток в костном мозгу и в селезенке.
Пример 7. Фармакокинетические исследования rHuEPO-Fc у обезьян резус
Как обсуждалось выше, заявители сконструировали rHuEPO-Fc таким образом, что часть ЕРО в слитом белке сохраняет функциональные свойства природного ЕРО, такие как свойство стимулировать эритропоэз, а фрагмент Fc человеческого lgG1 обеспечивает стабильное существование слитого белка в кровотоке, что продлевает его время полужизни in vivo. Приведенные выше исследования на животных показали, что эритропоэтическая активность rHuEPO-Fc выше, чем у rHuEPO. Заявители провели также фармакокинетические исследования для сравнения времени полужизни rHuEPO-Fc и rHuEPO in vivo. Для получения данных использовали приматов, поскольку они очень похожи на людей.
Планирование исследований было основано на литературных данных и опытах, проводившихся согласно общим правилам исследования фармакокинетики. Двум группам обезьян резус с 5 обезьянами в каждой группе (вес 3-5 кг, получены из Experiment Animal Center, AMMS, China) вводили в/в инъекцией 5 мкг/кг rHuEPO-Fc или rHuEPO. Образцы крови отбирали перед инъекцией и через 0,017, 0,167, 0,5, 1, 2, 4, 8, 12, 24, 48, 96, 168, 240 ч после инъекции. Сыворотки собирали центрифугированием и определяли уровни содержания в сыворотке rHuEPO-Fc или rHuEPO с помощью наборов для фермент-связанного иммуноферментного анализа (ELISA) на человеческий эритропоэтин (получены от R&D Systems, Minneapolis, MN). Среднее время полужизни (t1/2) rHuEPO-Fc и rHuEPO после в/в инъекции составляло соответственно 35,24±5,15 ч и 8,72±1,69 ч (данные суммированы на фиг.11).
Чтобы определить биологическую доступность rHuEPO-Fc, 5 мкг/кг rHuEPO-Fc вводили инъекцией п/к 5 обезьянам резус. Образцы крови отбирали перед инъекцией и через 1, 2, 5, 8, 10, 12, 15, 24, 48, 72, 96, 168, 240 ч после инъекции. Уровни содержания rHuEPO-Fc в сыворотках определяли с помощью наборов R&D. Рассчитанное значение индекса биологической доступности было равно 35,71±5,37% при п/к инъекции. Это значение идентично опубликованным данным по биологической доступности дарбепоэтина-а (AranespTM) у пациентов с хронической почечной недостаточностью [9, 15].
Эти данные показывают, что rHuEPO-Fc имеет существенно продленное время полужизни у приматов, и время полужизни rHuEPO-Fc in vivo по меньшей мере в 4 раза превышает время полужизни rHuEPO, выпускаемого фирмой Kirin Beer Brewing Co., Japan. Продленное время полужизни in vivo скорее всего способствует повышенной эритропоэтической активности rHuEPO-Fc.
Пример 8. Иммуногенность rHuEPO-Fc у Масаса fascicularis
Как указано выше, внимание к конструированию слитого белка rHuEPO-Fc было вызвано потребностью намеренно устранить или свести к минимуму изменения иммуногенных свойств слитого белка rHuEPO-Fc. Заявители избежали включения/добавления какой-либо чужеродной аминокислоты (каких-либо чужеродных аминокислот) или соединения мостиком пептидных последовательностей в слитом белке. Заявленный слитый белок HuEPO-Fc в варианте осуществления изобретения на фиг.1Б содержит только полипептидные последовательности природного белка ЕРО и фрагмента Fc (шарнирная область, СН2, СН3) человеческого lgG1 и теоретически не должен индуцировать иммуногенную реакцию и продукцию антител к белку rHuEPO-Fc. Как будет признано опытным специалистом в данной области, другие варианты осуществления, имеющие альтернативные структуры, также охватываются настоящим изобретением.
Для выяснения иммуногенности белка rHuEPO-Fc были проведены следующие исследования на приматах. Десяти питающимся крабами макакам (Масаса fascicularis, 5 самцов и 5 самок, возраст около 5 лет, средний вес самцов 4,0±0,3 кг, самок 2,9±0,4 кг, получены из Laboratory Animal Center, AMMS, China), вводили п/к инъекцией 5 мкг/кг очищенного rHuEPO-Fc 3 раза в неделю в течение 4 недель, двум обезьянам вводили равный объем раствора носителя (контроль). Сыворотки собирали 1 раз в неделю в течение 5 недель (начиная через 1 неделю после воздействия) и исследовали на наличие специфических антител к rHuEPO-Fc методом ELISA, используя в качестве покрывающего антигена очищенный rHuEPO-Fc (5 мкг/мл). Кроме того, в течение периода опыта в периферической крови определяли также число RBC и уровни Hb. Полученные данные показывают, что на фоне стимулированного усиления эритропоэза у получавших rHuEPO-Fc макак (среднее число RBC повышалось с 4,74×109/мл до 6,67×109/мл, а средние уровни Hb с 12,2 г/дл до 13,7 г/дл), rHuEPO-Fc не индуцировал заметных количеств специфических антител к слитому белку. Эти результаты показывают, что слитый белок rHuEPO-Fc не вызывает иммуногенности у приматов.
Пример 9. Исследование острой токсичности rHuEPO-Fc у нормальных мышей
Чтобы оценить безопасность слитого белка rHuEPO-Fc, были проведены исследования острой токсичности на животных.
Двум группам мышей BALB/c (n=20, равное число самцов и самок, возраст 5-6 недель, средний вес самок 15,8±0,4 г, самцов 15,9±0,6 г, получены из Chinese Academy of Medicine, China) вводили однократной в/в инъекцией в хвостовую вену избыточное количество rHuEPO-Fc (самцам 13,3 мг/кг, самкам 13,2 мг/кг) или равный объем раствора носителя. Кроме того, в течение 14 дней ежедневно контролировали и регистрировали общие поведение и статус, активность, принятие пищи и испражнение, чтобы установить стойкую реакцию после инъекции. Всех мышей также взвешивали на 7-й день и на 14-й день. На 15-й день после инъекции проводили анатомическое обследование основных органов мышей. Если бы наблюдались какие-либо необычные или подозрительные изменения органов, следовало провести патологическое обследование.
Ни у одной мыши в 2 группах не наблюдалось никакой заметной стойкой реакции после инъекции. В течение периода в 14 дней не наблюдалось заметных изменений поведения, активности, режимов еды и испражнения. Более того, в течение периода наблюдения вес мышей в обеих группах непрерывно увеличивался, и на 7-й и 14-й дни после инъекции не было обнаружено никаких очевидных различий между двумя группами. В тканях мозга, легких, сердца, печени и почек не было обнаружено никаких ненормальных или патологических изменений. Эти результаты показывают, что введение избыточного количества rHuEPO-Fc, намного большего, чем требуется для проявления нормальной эритропоэтической функции, безопасно и не дает очевидных токсических эффектов.
Пример 10. Сравнение слитых белков ЕРО дикого типа и мутантного
Были проведены также исследования для сравнения белков ЕРО дикого типа и мутантной версии. Как было описано выше, один из вариантов осуществления настоящего изобретения включал одиночную аминокислотную мутацию в положении 172 (C172G). Для сравнения был также приготовлен слитый белок дикого типа, имеющий в положении 172 цистеин (фиг.12). Слитый белок дикого типа был приготовлен так, как описано выше в примерах 1-3. Для конструирования рекомбинантной плазмиды использовали следующие олигонуклеотидные праймеры (QIAGEN Inc., US) (изменения для аминокислот в EFL5w и EFL3w по сравнению с праймерами в примере 1 обозначены жирным шрифтом):

Последовательности праймеров EFL5w и EFL3w указаны соответственно в SEQ ID No.9 и SEQ ID No.10.
Для сравнения эритропоэтической активности слитого белка дикого типа (обозначен здесь rHuEPO-FcC) с мутантным слитым белком (т.е. с описанным выше белком rHuEPO-Fc согласно настоящему изобретению) и с рекомбинантным человеческим ЕРО (rHuEPO) были проведены опыты in vivo на мышах. Для возможности сравнения все дозы использованных в этом примере трех белков, а именно rHuEPO-Fc, rHuEPO-FcC и rHuEPO, были выражены количеством только части ЕРО молекулы в молярном выражении. Что касается белков rHuEPO-Fc и rHuEPO-FcC, часть ЕРО составляет 41,4% от общего молекулярного веса, это рассчитано по отношению веса аминокислот в ЕРО к весу всех аминокислот в целых молекулах rHuEPO-Fc и rHuEPO-FcC (т.е. 166 ак из 399 ак).
rHuEPO-Fc (концентрация в рабочем растворе 300 мкг/мл), rHuEPO-FcC (концентрация в рабочем растворе 90 мкг/мл) и rHuEPO со структурой природного человеческого ЕРО (6000 ед./0,5 мл, производство фирмы Kirin Brewery Co., Japan) были разбавлены в растворе носителя (2,5 мг/мл человеческого сывороточного альбумина, 5,8 мг/мл цитрата натрия, 0,06 мг/мл лимонной кислоты и 5,8 мг/мл хлорида натрия, рН 5,5-5,6). Доза rHuEPO в количественном выражении была рассчитана на основании соотношения активность/количество. Мыши BALB/c (возраст 9-10 недель, вес 18-22 г, равное число самок и самцов, получены из Experiment Animal Center, AMMS, China) были разделены случайным образом на группы по 8 мышей в каждой группе. Каждая группа мышей получала одну комбинацию одной дозы (2,5, 12,5, 62,5 мкг/кг), одного пути введения (п/к) и одного режима инъекций (3 раза в неделю или 1 раз в неделю). Контрольной группе мышей вводили равный объем раствора носителя. Воздействие продолжалось 26 дней. Образцы периферической крови для измерений отбирали из хвостовой вены перед воздействием, и на 2-й, 6-й, 9-й, 13-й, 16-й, 19-й, 22-й и 26-й дни воздействия. Уровень Hb измеряли как индекс абсорбциометрии. Средние значения ± стандартные отклонения были рассчитаны по данным для каждой группы, для сравнения различных групп был применен t-тест.
Как показано на фиг.13, введение всех трех белков ЕРО в режиме 3 раза в неделю стимулировало эритропоэз. И в дозе 2,5 мкг/кг, и в дозе 12,5 мкг/кг rHuEPO-Fc индуцировал более значительное повышение уровней Hb, чем rHuEPO. Наивысший подъем уровней Hb был достигнут при дозе 12,5 мкг/кг rHuEPO-Fc. Обе дозы rHuEPO-FcC (2,5 мкг/кг и 12,5 мкг/кг) индуцировали намного более слабый эритропоэз, чем эквивалентные дозы rHuEPO и rHuEPO-Fc, что проявлялось в намного более слабом повышении уровней Hb в группах, получавших rHuEPO-FcC. Действительно, доза 12,5 мкг/кг rHuEPO-FcC индуцировала более слабый подъем уровней Hb, чем 2,5 мкг/кг rHuEPO. Эти результаты позволяют предположить, что HuEPO-FcC имеет более слабую эритропоэтическую активность in vivo, чем rHuEPO, имеющий последовательность природной молекулы ЕРО. Напротив, слитый белок rHuEPO-Fc согласно настоящему изобретению проявляет более сильную эритропоэтическую активность. Введение трех белков ЕРО в режиме 3 раза в неделю позволяет исключить вклад различий во времени полужизни белков.
Эритропоэтическую активность rHuEPO-Fc и rHuEPO-FcC дополнительно оценивали при снижении частоты инъекций до 1 в неделю п/к. Как показано на фиг.14, в получавших rHuEPO-Fc группах при дозах 12,5 мкг/кг или 62,5 мкг/кг обнаружен более значительный подъем уровней Hb, чем в группах, получавших rHuEPO. В противоположность этому, rHuEPO-FcC индуцировал намного более слабый подъем уровней Hb, чем rHuEPO. Например, доза 12,5 мкг/кг rHuEPO индуцировала больший подъем уровней Hb, чем доза 62,5 мкг/кг rHuEPO-FcC в большинстве моментов времени. Это дополнительно указывает на то, что при меньшей частоте введения, когда начинают сказываться различия во времени полужизни, rHuEPO-FcC проявляет намного более слабую эритропоэтическую активность, чем rHuEPO, имеющий молекулярную последовательность природного ЕРО, и чем слитый белок rHuEPO-Fc согласно настоящему изобретению.
В совокупности эти результаты показывают, что rHuEPO-FcC, полученный слиянием природных молекулярных последовательностей человеческого ЕРО и человеческого фрагмента Fc (шарнир, СН2 и СН3), проявляет намного более слабую эритропоэтическую активность in vivo, чем rHuEPO, имеющий молекулярную последовательность природного ЕРО. В частности, эритропоэтическая активность слитого белка rHuEPO-FcC составляет менее 1/5 от активности молекулы природного ЕРО. Это указывает на то, что слияние молекулы ЕРО с природной последовательностью человеческого фрагмента Fc ухудшает функциональные свойства молекулы ЕРО. Одиночное замещение первого цистеина в шарнирной области фрагмента Fc другой аминокислотой дает слитый белок rHuEPO-Fc согласно настоящему изобретению, содержащий последовательность природной молекулы ЕРО и мутантный фрагмент Fc и обладающий намного более высокой эритропоэтической активностью in vivo, чем молекула природного ЕРО. Эти данные позволяют предположить, что первый остаток цистеина в шарнирной области фрагмента Fc дикого типа мешает молекуле ЕРО, вызывая, по-видимому, структурные изменения в молекуле ЕРО, а это в свою очередь ухудшает функциональные свойства молекулы ЕРО в стимуляции эритропоэза.
Как будет очевидно опытным специалистам в данной области из предшествующего описания, в практике осуществления настоящего изобретения возможны многие изменения и модификации без выхода за рамки идеи и охвата настоящего изобретения.









Изобретение относится к области биотехнологии, конкретно к получению эритропоэтина, слитого с Fc-фрагментом человеческого IgG, и может быть использовано в медицине. Рекомбинантным путем получают слитый белок, содержащий природную молекулу человеческого эритропоэтина с остатком цистеина вблизи ее С-конца и фрагмент Fc человеческого IgG, содержащий шарнирную область, причем N-конец указанного Fc-фрагмента присоединен к указанному С-концу указанной молекулы эритропоэтина и при этом указанный Fc-фрагмент является природным, за исключением мутации, состоящей в замещении остатка цистеина в указанной шарнирной области, расположенного ближе всего к указанной эритропоэтиновой молекуле, нецистеиновым остатком, в результате чего первый цистеиновый остаток указанной шарнирной области, расположенный ближе всего к указанному N-концу, отделен по меньшей мере 12 или 17 аминокислотами от указанного остатка цистеина указанной молекулы эритропоэтина. Полученный пептид используют для стимуляции эритропоэза у млекопитающего. Изобретение позволяет получить слитый белок, обладающий активностью эритропоэтина, имеющий продленное время полужизни in vivo по сравнению с нативным человеческим эритропоэтином. 12 н. и 31 з.п. ф-лы, 20 ил.
1. Слитый белок, обладающий активностью эритропоэтина, имеющий продленное время полужизни in vivo по сравнению с природным или рекомбинантным нативным человеческим эритропоэтином, содержащий:
а) природную молекулу человеческого эритропоэтина, содержащую остаток цистеина вблизи ее С-конца; и
б) фрагмент Fc человеческого IgG, содержащий шарнирную область, причем N-конец указанного Fc-фрагмента присоединен к указанному С-концу указанной молекулы эритропоэтина, и при этом указанный Fc-фрагмент является природным, за исключением мутации, состоящей в замещении остатка цистеина в указанной шарнирной области, расположенного ближе всего к указанной эритропоэтиновой молекуле, нецистеиновым остатком, в результате чего первый цистеиновый остаток указанной шарнирной области, расположенный ближе всего к указанному N-концу, отделен по меньшей мере 17 аминокислотами от указанного остатка цистеина указанной молекулы эритропоэтина.
2. Белок, как он определен в п.1, где время полужизни указанного белка по меньшей мере в 3 раза больше времени полужизни указанного нативного человеческого эритропоэтина.
3. Белок, как он определен в п.2, где время полужизни указанного белка по меньшей мере в 4 раза больше времни полужизни указанного нативного человеческого эритропоэтина.
4. Слитый белок по п.2, где указанный слитый белок имеет повышенную эритропоэтическую биологическую активность по сравнению с указанным нативным человеческим эритропоэтином.
5. Слитый белок, как он определен в п.1, где указанный фрагмент Fc - это фрагмент IgG1.
6. Слитый белок, как он определен в п.5, где указанный фрагмент Fc содержит указанную шарнирную область и домены СН2 и СН3.
7. Слитый белок, как он определен в п.1, где указанный белок содержит аминокислотную последовательность, представленную в SEQ ID NO:2.
8. Слитый белок, имеющий продленное время полужизни in vivo по сравнению с природным или рекомбинантным нативным человеческим эритропоэтином, содержащий природную молекулу человеческого эритропоэтина, содержащую остаток цистеина вблизи ее С-конца, и фрагмент Fc человеческого IgG, содержащий шарнирную область, причем N-конец указанного Fc-фрагмента присоединен к указанному С-концу указанной молекулы эритропоэтина, и при этом указанный Fc-фрагмент является природным, за исключением мутации, состоящей в замещении остатка цистеина в указанной шарнирной области, расположенного ближе всего к указанной эритропоэтиновой молекуле, нецистеиновым остатком, в результате чего первый цистеиновый остаток указанной шарнирной области, расположенный ближе всего к указанному N-концу, отделен по меньшей мере 17 аминокислотами от указанного остатка цистеина указанной молекулы эритропоэтина, где указанный белок содержит аминокислотную последовательность, представленную в SEQ ID NO:2, или последовательность, чья степень идентичности с ней составляет по меньшей мере 98%, причем указанный белок имеет остаток нецистеиновой аминокислоты в положении 172.
9. Димерная белковая конструкция, обладающая активностью эритропоэтина, содержащая два слитых белка, как они определены в п.1, связанных дисульфидной связью между двумя шарнирными областями.
10. Димерный белок, содержащий два полипептида, каждый из которых содержит природную молекулу человеческого эритропоэтина, содержащую остаток цистеина вблизи ее С-конца, и фрагмент Fc человеческого IgG, содержащий шарнирную область, причем N-конец указанного Fc-фрагмента присоединен к указанному С-концу указанной молекулы эритропоэтина, и при этом указанный Fc-фрагмент является природным, за исключением мутации, состоящей в замещении остатка цистеина в указанной шарнирной области, расположенного ближе всего к указанной эритропоэтиновой молекуле, нецистеиновым остатком, в результате чего первый цистеиновый остаток указанной шарнирной области, расположенный ближе всего к указанному N-концу, отделен по меньшей мере 17 аминокислотами от указанного остатка цистеина указанной молекулы эритропоэтина, и имеет аминокислотную последовательность, представленную в SEQ ID NO:2, или последовательность, чья степень идентичности с ней составляет по меньшей мере 98%, причем указанный белок имеет остаток нецистеиновой аминокислоты в положении 172.
11. Белок, как он определен в п.10, где указанный димер содержит дисульфидные связи между соответствующими шарнирными доменами указанных полипептидов.
12. Белок, как он определен в п.10, где каждый из указанных полипептидов имеет молекулярную массу около 75 кДа.
13. Белок, как он определен в п.10, где указанный димер имеет молекулярную массу около 180 кДа.
14. Фармацевтическая композиция, обладающая активностью эритропоэтина, содержащая белок по п.1 в эффективном количестве вместе с фармацевтически приемлемым носителем, адъювантом или разбавителем.
15. Последовательность нуклеиновой кислоты, кодирующая полипептид, имеющий степень идентичности по меньшей мере 98% с SEQ ID NO:2, причем указанный полипептид имеет продленное время полужизни in vivo по сравнению с нативным человеческим эритропоэтином и содержит фрагмент молекулы человеческого иммуноглобулина, N-конец которого непосредственно присоединен к С-концу молекулы эритропоэтина, и при этом шарнирная область указанной молекулы иммуноглобулина имеет мутацию, посредством которой ближайший к указанному N-концу остаток цистеина замещен нецистеиновым остатком.
16. Выделенная последовательность нуклеиновой кислоты, имеющая последовательность, представленную в SEQ ID NO:1, или последовательность, имеющую по отношению к ней степень идентичности по меньшей мере 98%, кодирующая полипептид, содержащий фрагмент молекулы человеческого иммуноглобулина, N-конец которого непосредственно присоединен к С-концу молекулы эритропоэтина, и при этом шарнирная область указанной молекулы иммуноглобулина имеет мутацию, посредством которой ближайший к указанному N-концу остаток цистеина замещен нецистеиновым остатком.
17. Вектор экспрессии, содержащий последовательность нуклеиновой кислоты по п.15 или 16.
18. Клетка для продуцирования слитого белка по п.1, трансфицированная вектором экспрессии по п.17.
19. Клетка, как она определена в п.18, где указанная клетка - это клетка CHO.
20. Способ получения белка, как он определен в п.1, включающий культивирование клетки по п.18 и очистку кодированного ею полипептида.
21. Способ стимуляции эритропоэза у млекопитающего, включающий в себя введение указанному млекопитающему белка, как он определен в п.1, или фармацевтической композиции, как она определена в п.14.
22. Способ, как он определен в п.21, где указанное млекопитающее - это примат.
23. Способ, как он определен в п.22, где указанный примат является человеком.
24. Способ, как он определен в п.21, стимуляции эритропоэза у млекопитающего, включающий в себя введение указанному млекопитающему фармацевтической композиции, как она определена в п.14.
25. Способ, как он определен в п.24, где указанное млекопитающее - это примат.
26. Способ, как он определен в п.25, где указанный примат является человеком.
27. Способ, как он определен в п.21, где время полужизни указанного белка в указанном млекопитающем по меньшей мере в 3 раза превышает время полужизни нативного человеческого ЕРО при внутривенном или подкожном введении.
28. Способ, как он определен в п.27, где время полужизни указанного белка в указанном млекопитающем по меньшей мере в 4 раза превышает время полужизни нативного человеческого ЕРО при внутривенном или подкожном введении.
29. Слитый белок, как он определен в п.8, где указанная нецистеиновая аминокислота представляет собой нейтральную аминокислоту.
30. Слитый белок, как он определен в п.29, где указанная нецистеиновая аминокислота представляет собой глицин.
31. Слитый белок, как он определен в п.1, где время полужизни указанного белка при введении млекопитающему по меньшей мере в 3 раза превышает время полужизни нативного человеческого эритропоэтина, введенного указанному млекопитающему таким же способом.
32. Слитый белок, как он определен в п.31, где время полужизни указанного белка при введении млекопитающему по меньшей мере в 4 раза превышает время полужизни нативного человеческого эритропоэтина, введенного указанному млекопитающему таким же способом.
33. Слитый белок, как он определен в п.31, где указанное млекопитающее - это человек.
34. Вектор экспрессии, как он определен в п.17, содержащий последовательность нуклеиновой кислоты, представленную в SEQ ID NO:1.
35. Димерный белок, как он определен в п.10, содержащий два полипептида, каждый из которых имеет аминокислотную последовательность, представленную в SEQ ID NO:2.
36. Слитый белок, имеющий продленное время полужизни in vivo no сравнению с природным или рекомбинантным нативным человеческим эритропоэтином, содержащий:
а) часть из пептида эритропоэтина, содержащего прилегающий к его С-концу остаток цистеина; и
б) фрагмент Fc, содержащий шарнирную область, причем N-конец указанной шарнирной области непосредственно присоединен к указанному С-концу указанной части из пептида эритропоэтина, и при этом указанная шарнирная область имеет мутацию в прилегающем к указанному N-концу положении аминокислоты, состоящую в замещении остатка цистеина нецистеиновым остатком, причем первый цистеиновый остаток указанной шарнирной области, расположенный ближе всего к указанному N-концу, отделен по меньшей мере 12 аминокислотами от указанного остатка цистеина указанной части из пептида эритропоэтина.
37. Слитый белок, как он определен в п.36, где указанная шарнирная область имеет длину по меньшей мере 9 аминокислот.
38. Слитый белок, как он определен в п.37, где указанная шарнирная область представляет собой вариант человеческого фрагмента Fc, имеющий нецистеиновый остаток в положении аминокислоты 6, отмеренном от N-конца указанной шарнирной области.
39. Слитый белок, как он определен в п.38, где указанная шарнирная область имеет аминокислотную последовательность VEPKSGDKTSTCPPCP или последовательность, имеющую по отношению к ней степень идентичности по меньшей мере 90%.
40. Слитый белок, как он определен в п.36, где указанный белок имеет аминокислотную последовательность, указанную в SEQ ID NO:2, или последовательность, имеющую по отношению к ней степень идентичности по меньшей мере 98%.
41. Слитый белок, как он определен в п.36, где указанная часть из пептида эритропоэтина является полной молекулой человеческого эритропоэтина.
42. Слитый белок, как он определен в п.36, где указанный фрагмент Fc - это фрагмент Fc человеческого IgG, содержащий указанную шарнирную область и домены СН2 и СН3.
43. Слитый белок, как он определен в п.42, где указанный фрагмент Fc IgG представляет собой фрагмент IgG1.
| WO 2004004798 A2, 15.01.2004 | |||
| US 2005202538 A1, 15.09.2005 | |||
| WO 2005001025 A2, 06.01.2005 | |||
| RU 2003127387 A, 27.03.2005. |

