Предпосылки изобретения
Родственные заявки
По настоящей заявке испрашивается приоритет предварительной патентной заявки США №60/552678, поданной 11 марта 2004, на приоритет которой претендует настоящая заявка в соответствии с § 119 35 U.S.C. и содержание которой приведено здесь в качестве ссылки.
Область изобретения
Изобретение относится к способу получения полипептида, гетерологичного для E. coli. Более конкретно изобретение относится к применению органофосфата для улучшения выхода таких полипептидов.
Описание родственной области техники
Экспрессия гетерологичных белков в Escherichia coli, которой способствует хорошо изученная молекулярная биология микроорганизма и относительная простота генетических манипуляций с ним, является очень производительной и в лабораторных условиях, и на производстве. Как правило, для регуляции экспрессии гетерологичного белка применяют индуцибельный промотор (например, промотор щелочной фосфатазы, промотор tac, арабинозный промотор и т.д.). Потребность в индукционном событии предоставляет исследователю возможность контролировать синхронизацией экспрессии белка-мишени. Эта возможность особенно важна для тех гетерологичных белков, высокая концентрация которых не допускается у хозяина. Посредством достижения высокой плотности клеток перед индукцией экспрессии можно максимально увеличить объемный выход желательного белка.
Клетки перестают расти, когда микроорганизм лишают требуемого питательного вещества. Лимитирующим компонентом может являться углерод, азот, фосфат, кислород или любой из требуемых клетке элементов. В таких условиях клетки выходят из фазы роста. Способом облегчить ответы культуры на стресс, вызванный ограничением питательного вещества, является предоставление питания отсутствующим компонентом. Общепринятые компоненты питания, вводимые в процессы подпитываемой ферментации, включают в себя глюкозу, аминокислоты, кислород и т.д.
В случае клеточного фосфора (P), необходимость источника фосфата неудивительна, с учетом того, что P представляет собой наиболее широко распространенный элемент в клетке кроме углерода, кислорода, азота и водорода. Slanier, Adelberg and Ingraham, The Microbial World. 4th ed. (Prentice Hall, NJ 1976), p. 1357. Фосфор является необходимым компонентом многочисленных макромолекул, таких как нуклеиновые кислоты, липополисахариды и мембранные липиды. Кроме того, его роль в макроэргических фосфоангидридных связях делает его особенно важным в энергетическом обмене. E. coli способна утилизировать в качестве первичного источника P неорганический фосфат (Pi), органофосфат или фосфонат. Поглощения Pi из окружающей среды можно достичь посредством двух систем транспортеров, систем Pit и Pst. Что касается органофосфатов, то большинство из них не являются транспортабельными, и сначала их нужно гидролизовать ферментативно в периплазме, прежде чем высвобожденный Pi может доставлять транспортная система (системы) Pi. Только немногие органофосфаты являются транспортабельными, и глицерол-3-фосфат (G3P) является одним из таких примеров. G3P и глицерофосфат-1-фосфат (G1P) известны как альфа-глицерофосфаты. В ответ на ограничение Pi и ограничение углерода E. coli способна поглощать доступный интактный G3P из внешней среды во внутриклеточный компартмент, где G3P подвергают метаболизму для получения необходимого фосфата или углерода. Wanner, "Phosphorus Assimulation and Control of the Phosphate Regulon", in Escherichia coli and Salmonella Cellular and Molecular Biology, Neidhardt, ed. (второе издание), American Society for Microbiology Press (1996), pp. 1357-1365.
Дополнительными ссылками на G3P являются Silhavy et al., J. Bacteriol., 126: 951-958 (1976) о периплазматическом белке, связанном с транспортной системой sn-глицерол-3-фосфата в E. coli; Argast et al., J. Bacteriol., 136: 1070-1083 (1978) о второй транспортной системе sn-глицерол-3-фосфата в E. coli; Elvin et al., J. Bacteriol., 161: 1054-1058 (1985) об обмене Pi, опосредованном glpT-зависимой системой транспорта G3P; Rao et al., J. Bacteriol., 175: 74-79 (1993) об эффекте мутаций glpT и glpD на экспрессию гена phoA в E. coli; и Elashvili et al., Appl. Environ. Microbiol., 64: 2601-2608 (1998) о генах phnE и glpT, усиливающих утилизацию органофосфатов в E. coli K-12. Кроме того, в статье Vergeles et al. Eur. J. Biochem., 233: 442-447 (1995) описана высокая эффективность глицерол-2-фосфата (G2P), известного иначе как бета-глицерофосфат, и G3P как нуклеотидил-акцепторов в реакциях эстерификации фосфодиэстеразой в яде змеи.
Современное представление о двух транспортных системах для поглощения экзогенного G3P в E. coli, транспортных систем Ugp и GlpT, подробно суммировано в книге Escherichia coli and Salmonella, Cellular and Molecular Biology edited by Neidhardt et. al. (второе издание), выше, pp. 1364 в отношении ссылок 13 и 81. Оперон Ugp относится к регулону pho. Его индуцируют ограничением фосфата и положительно регулируют белком phoB. Система Ugp представляет собой периплазматическую зависимую от связывания белка многокомпонентную транспортную систему с ugpB, кодирующим периплазматический связывающий белок, ugpA и ugpC, кодирующими интегральные белки мембранных каналов, и ugpC, кодирующим АТФазу. GlpT является частью системы glp, опосредующей поглощение и метаболизм глицерина, G3P и глицерол-фосфорил фосфодиэфиров (Lin et al., Annu. Rev. Microbiol., 30: 535-578 (1976); Chapter 20; pg 307-342 Dissimilatory Pathways for sugars, polyols and carboxylates. Escherichia coli and Salmonella, Cellular and Molecular Biology, второе издание). Такая транспортная система представляет собой анионообменник, для которого известно, что он опосредует отток Pi из цитоплазмы посредством обмена с внешним G3P. В штамме дикого типа, выращиваемом на G3P, в то время как немного Pi высвобождают клетки, поглощающие G3P посредством системы Ugp, возможно высвобождение Pi в периплазму при поглощении G3P через систему GlpT. При высвобождении репрессивного количества Pi в результате опосредованного glpT-пермеазой оттока активность регулона pho, включая систему Ugp, выключается. В определенных условиях GlpT представляет собой единственный путь выхода Pi из клетки посредством обмена с внешним G3P. Elvin et al., J. Bacteriol., 161: 1054-1058 (1985); Rosenberg, ”Phosphate transport in prokaryotes," p. 205-248. In B. P. Rosen and S. Silver (ed.), Ion Transport in Prokaryotes (Academic Press, Inc., New York, 1987).
Когда сравнивают производительность систем Ugp и GlpT по транспорту G3P, максимальные скорости двух систем одинаковы. Кажущаяся аффинность для G3P выше у системы Ugp, чем у системы GlpT. Вероятно, обе системы способны поставлять достаточно G3P для роста клетки, если он доступен в среде для роста. Однако G3P, транспортируемый исключительно через систему Ugp, может служить единственным источником фосфата, но не углерода, тогда как GlpT-транспортированный G3P может служить единственным источником обоих (Schweizer et al., J. Bacteriol., 150: 1154-1163(1982)). Картированы два гена ugp, кодирующие pho-регулон-зависимую систему транспорта G3P (Schweizer et al., J. Bacteriol., 150: 1164-1171 (1982)), характеризована область ugp, содержащая данные гены (Schweizer et al., Mol. and Gen. Genetics, 197: 161-168 (1984)), и изучена регуляция оперона ugp (Schweizer et al., J. Bacteriol., 163: 392-394 (1985); Kasahara et al., J. Bacteriol., 173: 549-558 (1991); Su et al., Molecular & General Genetics, 230: 28-32 (1991); Brzoska et al., "ugp-dependent transport system for sn-glycerol 3-phosphate of Escherichia coli" p. 170-177 in A. Torriani-Gorini, F.G. Rothman, S. Silver, A. Wright, and E. Yagil (ed.), Phosphate Metabolism and Cellular Regulation in Microorganisms (American Society for Microbiology, Washington, D.C., 1987); Brzoska et al., J. Bacteriol., 176: 15-20 (1994); and Xavier et al., J. Bacteriol., 177: 699-704 (1995)).
В штаммах дикого типа существует стабильный внутриклеточный пул G3P, поддерживаемый при приблизительно 200 мкМ. Внутри G3P можно синтезировать ферментативным превращением глицерина в G3P глицеролкиназой (кодируемой glpK) при росте на глицерине как единственном источнике углерода, или восстановлением гликолитического промежуточного соединения, дигидроксиацетон-фосфата, G3P-синтазой, продуктом гена gpsA, при росте на источниках углерода, отличных от глицерина. Так как G3P представляет собой важное промежуточное соединение, формирующее каркас всех фосфолипидных молекул, внутренние глицерол-фосфаты можно получить также разложением фосфолипидов и триацилглицерина. Как метаболит внутренний G3P можно направить в путь биосинтеза фосфолипидов или окислить G3P-дегидрогеназой для формирования дигидроксиацетон-фосфата и ввести в гликолитический путь.
В ситуациях, где промотор AP служит для регуляции экспрессии гетерологичного белка в E. coli, поскольку индукция происходит только после истощения Pi в среде, клетки с индуцированной активностью промотора AP обычно голодают по фосфату, и их жизнеспособность снижена. Они могут быть вынуждены извлекать фосфат, необходимый для клеточных функций. Возможные последствия такого извлечения фосфата могут включать в себя оборачиваемость рибосом, низкую клеточную энергетику и увеличение экспрессии протеаз и протеолиза (St. John and Goldberg, J. Bacteriol., 143: 1223-1233 (1980)), потенциально приводя к менее здоровым клеткам со сниженной способностью накапливать белок.
Улучшение метаболического состояния E. coli, вероятно, может увеличить способность клеток синтезировать белки. Если подавать фосфат медленно, клетки могут только почувствовать низкую концентрацию Pi в периплазме, таким образом индуцируя регулон pho без внутриклеточного голодания по атому P (см. U.S. Pat. No 5304472). Существует необходимость предоставления дополнительных способов для продукции гетерологичных полипептидов в E. coli.
Сущность изобретения
Настоящее изобретение относится к способу для улучшения экспрессии гетерологичных полипептидов в E. coli. Показано, что подпитка транспортируемым органофосфатом, таким как альфа-глицерофосфат, различных хозяев - E. coli, включая хозяев с и без гена glpT дикого типа и хозяев с и без гена phoA дикого типа, таких как, например, E. coli (ugp+ ΔglpT phoA-), улучшает экспрессию гетерологичного белка в масштабе и встряхиваемой колбы, и 10-л-ферментера, и ожидают проводить подобное в большем масштабе, таком как 10000 л. Наблюдали улучшение выхода продукта для экспрессии гетерологичных белков в многочисленных модельных системах, применяющих множество промоторов, включая индуцибельные промоторы, такие как промотор tac, T7 или AP. Дополнительным преимуществом является то, что продукт можно получить раньше в фазе активного роста, т.е. за более короткое время, чем другим способом. В конкретных вариантах осуществления больше продукта можно получить раньше в фазе активного роста для значительного улучшения производительности.
Таким образом, настоящее изобретение соответствует формуле изобретения. В одном из аспектов настоящее изобретение относится к способу получения гетерологичного для E. coli полипептида, предусматривающему (a) культивирование клеток E. coli, содержащих кодирующую полипептид нуклеиновую кислоту, в культуральной среде при подпитке культуральной среды транспортабельным органофосфатом, так что нуклеиновая кислота является экспрессированной, и (b) выделение полипептида из клеток. В предпочтительном варианте осуществления органофосфат представляет собой глицерофосфат, более предпочтительно альфа-глицерофосфат и/или бета-глицерофосфат, и еще более предпочтительно смесь глицерол-2-фосфата и глицерол-3-фосфата или один глицерол-3-фосфат. В другом предпочтительном аспекте культивирование проводят во встряхиваемой колбе или ферментере, предпочтительно в ферментере. В еще одном предпочтительном варианте осуществления полипептид выделяют из цитоплазмы, периплазмы или культуральной среды клеток. Также предпочтительно, чтобы экспрессия нуклеиновой кислоты являлась регулируемой индуцибельным промотором, таким как промотор щелочной фосфатазы, промотор tac, или промотор T7, и предпочтительно экспрессию нуклеиновой кислоты начинают в фазе активного роста на стадии культивирования. В одном из вариантов осуществления E. coli представляет собой дикий тип. В другом варианте осуществления E. coli является дефектной по хромосомальному glpT и хромосомальному phoA, но предпочтительно не является дефектной по хромосомальному ugp. Предпочтительно неорганический фосфат также присутствует на стадии культивирования.
Без ограничения какой-либо одной теорией считают, что при данном способе клетки подпитывают транспортабельными фосфорорганическими соединениями, так что pstS системы Pho не будет воспринимать доставку фосфата, но будет все еще обеспечивать фосфат после расщепления в цитоплазме, и далее что подача транспортабельного органофосфата, такого как G3P, потенциально обогащает клетки утилизируемым промежуточным соединением, которое можно легко направлять в важные метаболические пути.
Краткое описание чертежей
На фиг.1 показана экспрессия секретируемого фрагмента антитела ламы в хозяине - E. coli BL21 с применением промотора tac в культуре во встряхиваемой колбе, с применением или воды, или 200 мМ G3P в качестве дополнения к низкофосфатной (CRAP) или высокофосфатной (THCD) среде.
На фиг.2 показана экспрессия цитоплазматического Apo2L в хозяине - E. coli HMS174 с применением промотора T7 в культуре во встряхиваемой колбе, с применением или воды, или 200 мМ G3P в качестве дополнения к среде CRAP.
На фиг.3 показан эффект подпитки G3P во время ферментации на накопление со временем секретируемого IGF-I. Применяли хозяина - E. coli дикого типа, промотор AP, и непрерывную подпитку глюкозой.
На фиг.4 показан эффект мутации glpT и подпитки G3P во время ферментации на накопление со временем секретируемого IGF-I. Применяли хозяина - E. coli ΔglpT, промотор AP и различную скорость подачи G3P.
На фиг.5 показана схема плазмиды pAPApo2-P2RU.
На фиг.6 показана нуклеотидная последовательность кДНК лиганда Apo-2 человека (SEQ ID NO: 1) и производная от нее аминокислотная последовательность (SEQ ID NO:2). «N» в положении нуклеотида 447 (в SEQ ID NO: 1) применяют, чтобы указывать, что нуклеотидное основание может представлять собой «T» или «G».
На фиг.7 показан эффект подпитки G3P на специфическое накопление Apo2L в хозяине - E. coli ΔglpT (43F6), при трех различных скоростях подачи и контроле без подпитки G3P.
На фиг.8 показано преимущество подпитки глицерофосфатом над неорганическим фосфатом для специфического общего накопления Apo2L для хозяина дикого типа по glpT (43E7), где плотность клеток увеличена до превышающей 200 OD550.
На фиг.9 показан эффект на специфическое общее накопление Apo2L замены неорганического фосфата на глицерофосфат в хозяине - E. coli дикого типа по glpT (43E7) и хозяине - E. coli ΔglpT (43F6).
На фиг.10 показан эффект на специфическое общее накопление Apo2L замены альфа-глицерофосфата на смесь 50:50 альфа- и бета-глицерофосфата в качестве подпитки по сравнению с контролем без подпитки в хозяине - E. coli ΔglpT (61G1).
Подробное описание предпочтительных вариантов осуществления
Определения
Как применяют здесь, «полипептид» относится в общем смысле к пептидам и белкам, обладающим более чем десятью аминокислотами. «Гетерологичные» полипептиды представляют собой полипептиды, чужеродные для применяемой клетки-хозяина, такие как человеческий белок, производимый E. coli. В то время как полипептид может являться прокариотическим или эукариотическим, предпочтительно он является эукариотическим, более предпочтительно относящимся к млекопитающим, и наиболее предпочтительно человеческим.
Примеры полипептидов млекопитающих включают в себя такие молекулы, как, например, реннин; гормон роста, включая гормон роста человека или бычий гормон роста; фактор, высвобождающий гормон роста; паратиреоидный гормон; тиреостимулирующий гормон; липопротеины; 1-антитрипсин; A-цепь инсулина; B-цепь инсулина; проинсулин; тромбопоэтин; фолликулостимулирующий гормон; кальцитонин; лютеинизирующий гормон; глюкагон; факторы свертывания крови, такие как фактор VIIIC, фактор IX, тканевой фактор и фактор фон Виллебранда; противосвертывающие факторы, такие как белок C; атриальный натрийуретический фактор; легочный сурфактант; активатор плазминогена, такой как урокиназа или человеческий урокиназный или тканевой активатор плазминогена (t-PA); бомбезин; тромбин; гемопоэтический фактор роста; фактор некроза опухолей-альфа и -бета; антитела против домена(доменов) ErbB2, такие как 2C4 (WO 01/00245; гибридома ATCC HB-12697), связывающие область во внеклеточном домене ErbB2 (например, один или несколько остатков в области от приблизительно 22 остатка до приблизительно 584 остатка ErbB2, включительно); энкефалиназа; мюллеров ингибирующий фактор; A-цепь релаксина; B-цепь релаксина; прорелаксин; гонадотропин-ассоциированный пептид мыши; бактериальный белок, такой как бета-лактамаза; ДНКаза; ингибин; активин; фактор роста эндотелия сосудов (VEGF); рецепторы для гормонов и факторов роста; интегрин; белок A или D; ревматоидные факторы; нейротрофический фактор, такой как мозговой нейротрофический фактор (BDNF), нейротрофин-3, -4, -5, или -6 (NT-3, NT-4, NT-5, или NT-6), или фактор роста нервов, такой как NGF; кардиотрофины (факторы гипертрофии сердца), такие как кардиотрофин-1 (CT-1); тромбоцитарный фактор роста (PDGF); фактор роста фибробластов, такой как aFGF и bFGF; эпидермальный фактор роста (EGF); трансформирующий фактор роста (TGF), такой как TGF-альфа и TGF-бета, включая TGF-1, TGF-2, TGF-3, TGF-4 или TGF-5; инсулиноподобный фактор роста-I и -II (IGF-I и IGF-II); des(1-3)-IGF-I (мозговой IGF-I); белки, связывающие инсулиноподобный фактор роста; CD-белки, такие как CD-3, CD-4, CD-8 и CD-19; эритропоэтин; остеоиндуктивные факторы; иммунотоксины; морфогенетический белок кости (BMP); интерферон, такой как интерферон-альфа, -бета и -гамма; сывороточный альбумин, такой как человеческий сывороточный альбумин (HSA) или бычий сывороточный альбумин (BSA); колониестимулирующие факторы (CSF), например, M-CSF, GM-CSF и G-CSF; интерлейкины (IL), например, IL-1 - IL-10; анти-HER-2-антитело; лиганд Apo2 (Apo2L); супероксиддисмутаза; Т-клеточные рецепторы; поверхностно-мембранные белки; стимулятор гомолиза; вирусные антигены, такие как, например, часть оболочки AIDS; транспортные белки; рецепторы хоминга; адресины; регуляторные белки; антитела; и фрагменты любых из перечисленных выше полипептидов.
Предпочтительные интересующие полипептиды включают в себя такие полипептиды, как HSA, BSA, анти-IgE, анти-CD20, анти-IgG, t-PA, gp120, анти-CD11a, анти-CD18, 2C4, анти-VEGF, VEGF, TGF-бета, активин, ингибин, анти-HER-2, ДНКаза, IGF-I, IGF-II, мозговой IGF-I, гормон роста, цепи релаксина, фактор, высвобождающий гормон роста, цепи инсулина или проинсулин, антитела и фрагменты антител, NGF, NT-3, BDNF, Apo2L и урокиназа. Наиболее предпочтительно полипептид представляет собой IGF-I или Apo2L.
Термины «лиганд Apo2», «Apo2L», и «TRAIL» применяют здесь попеременно в отношении последовательности полипептида, содержащей аминокислотные остатки 114-281, включительно, остатки 95-281, включительно, остатки 92-281, включительно, остатки 91-281, включительно, остатки 41-281, включительно, остатки 15-281, включительно, или остатки 1-281, включительно, аминокислотной последовательности, показанной на фиг.6 (SEQ ID NO:2), так же как биологически активные фрагменты, и делеционные, инсерционные и замещенные варианты вышеуказанных последовательностей. В одном из вариантов осуществления последовательность полипептида содержит остатки 114-281 из фиг.6 (SEQ ID NO:2). Не обязательно, последовательность полипептида содержит остатки 92-281 или остатки 91-281 из фиг.6 (SEQ ID NO:2). Полипептиды Apo2L может кодировать природная нуклеотидная последовательность, показанная на фиг.6 (SEQ ID NO:1). Не обязательно, кодон, кодирующий остаток Pro119 (фиг.6; SEQ ID NO:1), может представлять собой «CCT» или «CCG». В другом предпочтительном варианте осуществления фрагменты или варианты являются биологически активными и обладают по меньшей мере приблизительно 80% идентичностью аминокислотной последовательности, более предпочтительно по меньшей мере приблизительно 90% идентичностью последовательности, и даже более предпочтительно, по меньшей мере 95%, 96%, 97%, 98%, или 99% идентичностью последовательности с любой из вышеуказанных последовательностей. Определение охватывает замещенные варианты лиганда Apo2, в которых по меньшей мере одну из его природных аминокислот замещают остатком аланина. Определение охватывает также лиганд Apo2 с природной последовательностью, выделенный из природного источника лиганда Apo2, или полученный рекомбинантными или синтетическими способами. Лиганд Apo2 по изобретению включает в себя полипептиды, на которые ссылаются как на лиганд Apo2 или TRAIL, описанные в WO 97/01633, WO 97/25428 и WO 01/00832. Термины «лиганд Apo2» и «Apo2L» применяют для ссылки в общем смысле на формы лиганда Apo2, включающие в себя мономерную, димерную или тримерную формы полипептида. Для всей нумерации аминокислотных остатков, на которые ссылаются в последовательности Apo2L, применяют нумерацию в соответствии с фиг.6 (SEQ ID NO:2), если конкретно не указано иначе. Например, «D203» или «Asp203» относится к остатку аспарагиновой кислоты в положении 203 в последовательности, приведенной на фиг.6 (SEQ ID NO:2).
Термин «внеклеточный домен лиганда Apo-2» или «ECD лиганда Apo2» относится к форме лиганда Apo2, по существу свободной от трансмембранного и цитоплазматического доменов. Обычно ECD обладает менее чем 1% данных трансмембранного и цитоплазматического доменов, и предпочтительно обладает менее чем 0,5% данных доменов. «Биологически активный» или «биологическая активность» в отношении Apo2L относится к (a) обладанию способностью индуцировать или стимулировать апоптоз по меньшей мере в одном типе клеток опухоли или инфицированных вирусом клеток млекопитающих in vivo или ex vivo; (b) способности вызывать продукцию антитела (т.е. иммуногенности), (c) способности связывать и/или стимулировать рецептор для Apo2L; или (d) сохранению активности нативного или природного полипептида Apo2L.
«Контрольные последовательности» экспрессии относится к последовательностям ДНК, необходимых для экспрессии функционально связанной кодирующей последовательности в конкретном организме-хозяине. Контрольные последовательности, подходящие для прокариот, включают в себя промотор, необязательно, последовательность оператора и участок связывания рибосомы.
Нуклеиновая кислота «функционально связана», когда ее помещают в функциональную взаимосвязь с другой последовательностью нуклеиновой кислоты. Например, ДНК для препоследовательности или секреторного лидера функционально связана с ДНК для полипептида, если ее экспрессируют как пребелок, участвующий в секреции полипептида; промотор функционально связан с кодирующей последовательностью, если он действует на транскрипцию последовательности; или участок связывания рибосомы функционально связан с кодирующей последовательностью, если он расположен так, чтобы содействовать трансляции. Как правило, «функционально связанный» означает, что связываемые последовательности ДНК являются смежными и, в случае секреторного лидера, смежными и в фазе считывания. Связывание осуществляют лигированием в подходящие участки рестрикции. Если таких участков не существует, можно применять синтетические олигонуклеотидные адаптеры или линкеры в соответствии с общепринятой практикой.
Как применяют здесь, выражения «клетка», «линия клеток» и «культура клеток» применяют попеременно, и все данные обозначения включают в себя потомство. Так, слова «трансформанты» и «трансформированные клетки» включают в себя указанную первичную клетку и производные от нее культуры, независимо от числа переносов. Понятно также, что потомство может не являться точно идентичным по содержанию ДНК, из-за намеренных или случайных мутаций. Сюда включают мутантное потомство, обладающее той же функцией или биологической активностью, по какой отбирали первоначальные трансформированные клетки. Где подразумевают особые обозначения, это станет ясно из контекста.
Термин «органофосфат», как применяют здесь, относится к фосфатному соединению, содержащему один или несколько атомов углерода, которое может содержать также атомы галогенида. Данное фосфатное соединение должно являться таким, чтобы им можно было подпитывать культуру клеток, и она могла его утилизировать. Данные соединения часто применяют как пестициды. «Транспортабельные» органофосфаты можно транспортировать из внешнего окружения клетки в клетку без пре-гидролиза каким-либо образом. Если штамм E. coli не растет хорошо на органофосфате, утилизацию такого органофосфата можно улучшить сверхэкспрессией в E. coli продукта гена phnE. Данный ген после трансформации сообщает штамму E. coli фенотип непосредственной утилизации органофосфата. Смотри Elashvili et al., выше. Примеры подходящих органофосфатов включают в себя алкил-галофосфаты, такие как диизопропилфторфосфат, алкилфосфаты, такие как диизопропилфосфат и 3,4-дигидроксибутил-1-фосфат, так же как сахар- или алканол-содержащие фосфаты, такие как гексоза-6-фосфат и глицерол-3-фосфат. Предпочтительными являются глюкоза-1-фосфат, гексоза-6-фосфат и глицерофосфаты, такие как глюкоза-1-глицерофосфат, фруктоза-6-глицерофосфат, альфа-глицерофосфаты, такие как глицерол-1-фосфат и глицерол-3-фосфат, и бета-глицерофосфат (глицерол-2-фосфат), более предпочтительными являются глицерофосфаты, еще более предпочтительными являются альфа- и/или бета-глицерофосфаты, и еще более предпочтительными являются глицерол-2-фосфат и/или глицерол-3-фосфат, и самыми предпочтительными для применения здесь являются смесь глицерол-2- и глицерол-3-фосфата или глицерол-3-фосфат. Как применяют здесь, термин «G3P», не присутствующий в смеси, или «один G3P» относится к композиции, содержащей по меньшей мере приблизительно 80% глицерол-3-фосфата; она может содержать вплоть до приблизительно 20% примесей, таких как G2P. Смесь G3P и G2P содержит менее чем приблизительно 80% G3P.
Неорганический фосфат представляет собой фосфатное соединение, не содержащее атомов углерода, с фосфатом, обычно связанным с щелочным или щелочноземельным металлом, такое как фосфат калия, кальция, магния или натрия.
«Фаза активного роста» относится к фазе стадии культивирования, где клетки являются активно растущими и не строго ограниченными в питании, как находящиеся в стационарной фазе клетки.
Способы осуществления изобретения
Настоящее изобретение относится к способу получения гетерологичных для E. coli полипептидов. По данному способу клетки E. coli, содержащие кодирующую полипептид нуклеиновую кислоту, культивируют в культуральной среде при подпитке культуральной среды транспортабельным органофосфатом, так что нуклеиновая кислота является экспрессированной. Затем полипептид выделяют из клеток. Выделять можно из цитоплазмы, периплазмы или культуральной среды клеток. Культивирование можно проводить в любом подходящем сосуде, предпочтительно во встряхиваемой колбе или ферментере, более предпочтительно в ферментере.
Можно применять параметры культивирования и проводить производство полипептида общепринятым способом, таким как способы, описанные ниже.
A. Выбор нуклеиновой кислоты и ее модификации
Нуклеиновая кислота, кодирующая интересующий полипептид, представляет собой подходящую РНК, кДНК или геномную ДНК из любого источника при условии, что она кодирует интересующий полипептид(ы). Хорошо известны способы выбора подходящей нуклеиновой кислоты для экспрессии гетерологичных полипептидов (включая их варианты) в E. coli.
Если производят моноклональные антитела, ДНК, кодирующую моноклональные антитела, легко выделить и секвенировать с применением общепринятых способов (например, с применением олигонуклеотидных зондов, способных специфически связываться с генами, кодирующими тяжелые и легкие цепи мышиных антител). Предпочтительным источником данной ДНК являются клетки гибридомы. После выделения ДНК можно помещать в экспрессирующие векторы, которыми затем здесь трансформируют бактериальные клетки-хозяева для получения синтеза моноклональных антител в рекомбинантных клетках-хозяевах. Обзорные статьи по рекомбинантной экспрессии в бактериях ДНК, кодирующей антитело, включают в себя Skerra et al., Curr. Opinion in Immunol., 5: 256-262 (1993) и Plückthun, Immunol. Revs., 130: 151-188(1992).
Способы гуманизации не относящихся к человеку антител описаны в данной области. Предпочтительно, гуманизированное антитело обладает одним или несколькими аминокислотными остатками, введенными в него из источника, не относящегося к человеку. На данные не относящиеся к человеку аминокислотные остатки часто ссылаются как на «импортируемые» остатки, которые обычно взяты из «импортируемого» вариабельного домена. Гуманизацию по существу можно проводить по способу Winter и соавторов (Jones et al., Nature, 321: 522-525 (1986); Riechmann et al., Nature, 332: 323-327 (1988); Verhoeyen et al., Science, 239: 1534-1536 (1988)) посредством замещения последовательностей гипервариабельной области на соответствующие последовательности человеческого антитела. Соответственно, такие «гуманизированные» антитела представляют собой химерные антитела (U.S. Pat. No 4816567), где существенно меньше, чем интактный человеческий вариабельный домен замещен соответствующей последовательностью из не относящихся к человеку видов. На практике гуманизированные антитела обычно представляют собой человеческие антитела, в которых некоторые остатки гипервариабельной области и, возможно, некоторые остатки FR замещают остатками из аналогичных участков антител грызунов.
Выбор человеческих вариабельных доменов, легких и тяжелых, для применения в получении гуманизированных антител является очень важным для уменьшения антигенности. По так называемому способу «наилучшего соответствия» последовательность вариабельного домена антитела грызунов отбирают по сравнению с полной библиотекой известных человеческих последовательностей вариабельных доменов. Затем человеческую последовательность, наиболее близкую к последовательности грызуна, принимают в качестве человеческой каркасной области (FR) для гуманизированного антитела (Sims et al., J. Immunol., 151: 2296 (1993); Chothia et al., J. Mol. Biol., 196: 901 (1987)). По другому способу применяют конкретную каркасную область, выведенную из консенсусной последовательности всех человеческих антител конкретной подгруппы легких или тяжелых цепей. Один и тот же каркас можно применять для нескольких различных гуманизированных антител (Carter et al., Proc. Natl. Acad. Sci. USA, 89: 4285 (1992); Presta et al., J. Immunol., 151: 2623 (1993)).
Кроме того, важно, чтобы антитела являлись гуманизированными с сохранением высокой аффинности к антигену и других удобных биологических свойств. Для достижения данной цели по предпочтительному способу гуманизированные антитела получают способом анализа исходных последовательностей и различных умозрительных гуманизированных продуктов с применением трехмерных моделей исходных и гуманизированных последовательностей. Трехмерные модели иммуноглобулинов общедоступны и знакомы специалистам в данной области. Доступны компьютерные программы, иллюстрирующие и демонстрирующие возможные трехмерные конформационные структуры выбранных иммуноглобулиновых последовательностей-кандидатов. Проверка данных демонстраций позволяет анализ возможной роли остатков в функционировании иммуноглобулиновых последовательностей-кандидатов, т.е. анализ остатков, влияющих на способность иммуноглобулина-кандидата связывать его антиген. Таким образом можно выбирать и комбинировать остатки FR из последовательностей реципиента и импортируемых, так чтобы достигать желаемых характеристик антитела, таких как увеличенная аффинность для антигена(антигенов). Вообще, остатки гипервариабельной области непосредственно и наиболее существенно вовлечены во влияние на связывание антигена.
Рассматривают различные формы гуманизированного антитела или аффинно зрелого антитела. Например, гуманизированное антитело или аффинно зрелое антитело может представлять собой фрагмент антитела, такой как Fab, не обязательно, конъюгированный с одним или несколькими направляющими средствами (средством) для получения иммуноконъюгата. Альтернативно, гуманизированное антитело или аффинно зрелое антитело может представлять собой интактное антитело, такое как интактное антитело IgG1.
Fab'-SH-фрагменты можно выделить непосредственно из E. coli и химически объединить для получения F(ab')2-фрагментов (Carter et al., Bio/Technology, 10: 163-167 (1992)). По другому способу F(ab')2-фрагменты можно выделить непосредственно из культуры рекомбинантных клеток-хозяев. Другие способы получения фрагментов антитела очевидны практикующему специалисту в данной области. В других вариантах осуществления выбранное антитело представляет собой одноцепочечный Fv-фрагмент (scFv) (WO 93/16185; U.S. Pat. No 5571894 и 5587458). Фрагмент антитела может также представлять собой «линейное антитело», например, как описано в U.S. Pat. No 5641870. Данные линейные фрагменты антитела могут являться моноспецифическими или биспецифическими.
Биспецифические антитела представляют собой антитела, обладающие связывающими специфичностями по меньшей мере для двух различных эпитопов. Примерные биспецифические антитела могут связывать два различных эпитопа одного и того же белка. Биспецифические антитела можно получить как полноразмерные антитела или фрагменты антитела (например, биспецифические антитела F(ab')2). Они могут являться слитыми из различных цепей антитела или могут являться одноцепочечными. Одна тяжелая цепь может являться достаточной самостоятельно.
По одному способу получения биспецифических антител, биспецифический иммуноадгезин получают введением в клетку-хозяина последовательностей ДНК, кодирующих первый слитный белок, состоящий из первого связывающего домена, слитого с последовательностью константного домена тяжелой цепи иммуноглобулина с отсутствующим участком связывания легкой цепи; второй слитный белок, состоящий из второго связывающего домена, слитого с последовательностью константного домена тяжелой цепи иммуноглобулина, сохраняющего участок связывания легкой цепи; и легкую цепь иммуноглобулина соответственно. Затем клетку-хозяина культивируют, так чтобы экспрессировать последовательности ДНК для получения смеси из (i) гетеротримера, содержащего первый слитный белок, ковалентно связанный с парой второй слитный белок - легкая цепь иммуноглобулина; (ii) гетеротетрамера, содержащего две ковалентно связанные пары второй слитный белок - легкая цепь иммуноглобулина; и (iii) гомодимера, содержащего две ковалентно связанные молекулы первого слитного белка. Смесь продуктов удаляют из культуры клеток и гетеротример отделяют от других продуктов. Данный способ описан в WO 94/04690. Дополнительные подробности получения биспецифических антител смотри, например, в Suresh et al., Methods in Enzymology, 121: 210 (1986).
Согласно другому подходу, описанному в U.S. Pat. No 5731168, поверхность раздела между парой молекул антитела можно сконструировать, чтобы максимизировать процент гетеродимеров, выделяемых из культуры рекомбинантных клеток. Предпочтительная поверхность раздела содержит по меньшей мере CH3-домен константного домена антитела. По данному способу одну или несколько боковых цепей небольших аминокислот из поверхности раздела первой молекулы антитела заменяют более крупными боковыми цепями (например, тирозина или триптофана). Компенсаторные «впадины» идентичного или похожего размера с большой боковой цепью(цепями) создают на поверхности раздела второй молекулы антитела заменой больших боковых цепей аминокислот на меньшие (например, аланина или треонина). Это предоставляет механизм для увеличения выхода гетеродимера над другими нежелательными конечными продуктами, такими как гомодимеры.
Биспецифические антитела включают в себя перекрестно сшитые или «гетероконъюгированные» антитела. Например, одно из антител в гетероконъюгате может быть соединено с авидином, другое с биотином. Данные антитела, например, предлагают для нацеливания клеток иммунной системы на нежелательные клетки (U.S. Pat. No 4676980), и для лечения инфекции HIV (WO 91/00360, WO 92/200373 и EP 03089). Гетероконъюгированные антитела можно получить с применением любых подходящих способов перекрестного сшивания. Подходящие средства для перекрестного сшивания хорошо известны в данной области и описаны, например, в U.S. Pat. No 4676980, вместе с рядом способов перекрестного сшивания.
Способы получения биспецифических антител из фрагментов антитела также описаны в литературе. Например, биспецифические антитела можно получить с применением химической связи. Brennan et al., Science, 229: 81 (1985) описали способ, где интактные антитела протеолитически расщепляют для получения F(ab')2-фрагментов. Данные фрагменты восстанавливают в присутствии арсенита натрия, образующего комплексы с дитиолами, для стабилизации смежных дитиолов и предупреждения формирования межмолекулярного дисульфида. Затем полученные Fab'-фрагменты переводили в производные тионитробензоата (TNB). Одно из Fab'-TNB производных затем переводили обратно в Fab'-тиол восстановлением меркаптоэтиламином и смешивали с эквимолярным количеством другого Fab'-TNB производного для формирования биспецифического антитела. Полученные биспецифические антитела можно применять в качестве средств для селективной иммобилизации ферментов.
Кроме того, Fab'-SH-фрагменты можно выделять непосредственно из E. coli и химически соединять для формирования биспецифических антител (Shalaby et al., J. Exp. Med., 175: 217-225 (1992)).
Описаны также различные способы для получения и выделения биспецифических фрагментов антител непосредственно из культуры рекомбинантных клеток. Например, биспецифические антитела получены с применением лейциновых молний (Kostelny et al., J. Immunol., 148: 1547-1553 (1992)). Пептиды с лейциновыми молниями из белков Fos и Jun присоединяют к Fab'-частям двух различных антител посредством слияния генов. Гомодимеры антитела восстанавливают в шарнирной области для получения мономеров и затем вновь окисляют для формирования гетеродимеров антитела. Данный способ можно применять также для получения гомодимеров антитела. Способ «диател», описанный Hollinger et al., Proc. Natl. Acad. Sci. USA, 90: 6444-6448 (1993), предоставляет альтернативный механизм для получения биспецифических фрагментов антител. Фрагменты, содержащие вариабельный домен тяжелой цепи (VH), соединяют с вариабельным доменом легкой цепи (VL) посредством линкера, слишком короткого, чтобы позволить спаривание между двумя доменами одной и той же цепи. Следовательно, домены VH и VL одного фрагмента вынуждены спариваться с комплементарными доменами VL и VH другого фрагмента, таким образом формируя два антигенсвязывающих участка. Опубликована также другая стратегия для получения биспецифических фрагментов антител с применением димеров одноцепочечного Fv (sFv) (Gruber et al., J. Immunol., 152: 5368 (1994)).
Рассматривают антитела с более, чем двумя валентностями. Например, можно получить триспецифические антитела (Tutt et al., J. Immunol., 147: 60 (1991)).
Молекулы нуклеиновых кислот, кодирующие варианты полипептидов, получают множеством способов, известных в данной области. Данные способы включают в себя в качестве неограничивающих примеров выделение из природного источника (в случае встречающихся в природе вариантов аминокислотной последовательности) или получение посредством олигонуклеотид-опосредованного (или сайт-специфического) мутагенеза, PCR-мутагенеза, или кассетного мутагенеза полученной ранее вариантной или не-вариантной версии полипептида.
Может являться желательным модифицировать антитело по изобретению по отношению к эффекторной функции, например, так, чтобы усиливать связывание рецептора Fc. Этого можно достичь введением одной или нескольких аминокислотных замен в Fc-область антитела. Альтернативно или дополнительно, в Fc-область можно вводить остаток(остатки) цистеина, таким образом, позволяя формирование в данной области межцепочечной дисульфидной связи.
Для увеличения полужизни антитела в сыворотке можно вводить эпитоп связывания рецептора выживания в антитело (особенно во фрагмент антитела), как описано, например, в U.S. Pat. 5739277. Как применяют здесь, термин «эпитоп связывания рецептора выживания» относится к эпитопу Fc-области молекулы IgG (например, IgG1, IgG2, IgG3 или IgG4), ответственному за увеличение полужизни молекулы IgG в сыворотке in vivo.
Здесь рассматривают другие модификации антитела. Например, к антителу можно присоединить ряд небелковых полимеров, например полиэтиленгликоль, полипропиленгликоль, полигидроксиалкены или сополимеры полиэтиленгликоля и полипропиленгликоля.
B. Вставка нуклеиновой кислоты в реплицируемый вектор
Гетерологичную нуклеиновую кислоту (например, кДНК или геномную ДНК) соответствующим образом вставляют в реплицируемый вектор для экспрессии в E. coli под контролем подходящего промотора. Для данной цели доступны многие векторы, и выбор подходящего вектора в основном зависит от размера нуклеиновой кислоты, подлежащей вставке в вектор, и конкретной клетки-хозяина, подлежащей трансформации вектором. Каждый вектор содержит различные компоненты в зависимости от конкретной клетки-хозяина, с которой он совместим. В зависимости от конкретного типа хозяина компоненты вектора, как правило, включают в себя в качестве неограничивающих примеров один или несколько из следующих: сигнальную последовательность, точку начала репликации, один или несколько маркерных генов, промотор и последовательность терминации транскрипции.
Как правило, в связи с хозяевами - E. coli применяют плазмидные векторы, содержащие репликон и контрольные последовательности, происходящие из видов, совместимых с клеткой-хозяином. Обычно вектор несет участок репликации, так же как маркерные последовательности, способные предоставлять отбор по фенотипу трансформированных клеток. Например, E. coli обычно трансформируют с применением pBR322, плазмиды, происходящей из видов E. coli (см., например, Bolivar et al., Gene, 2: 95 (1977)). pBR322 содержит гены для устойчивости к ампициллину и тетрациклину и таким образом предоставляет простые способы для идентификации трансформированных клеток. Плазмида pBR322, или другая бактериальная плазмида или фаг, также обычно содержит, или модифицирована, чтобы содержать промоторы, которые хозяин - E. coli может применять для экспрессии генов селективных маркеров.
(i) Компонент сигнальной последовательности
ДНК, кодирующую интересующий полипептид, можно экспрессировать не только непосредственно, но также в качестве слитного белка с другим полипептидом, предпочтительно сигнальной последовательностью или другим полипептидом, обладающим специфическим участком расщепления на N-конце зрелого полипептида. Как правило, сигнальная последовательность может являться компонентом вектора или может являться частью кодирующей полипептид ДНК, вставляемой в вектор. Выбранная гетерологичная сигнальная последовательность должна быть узнаваемой и процессируемой (т.е. расщепляемой сигнальной пептидазой) в клетке-хозяине.
Для прокариотических клеток-хозяев, которые не узнают и не процессируют природную или эукариотическую сигнальную последовательность полипептида, сигнальную последовательность заменяют на прокариотическую сигнальную последовательность, например, выбранную из группы, состоящей из лидеров щелочной фосфатазы, пенициллиназы, lpp, или термостабильного энтеротоксина II.
(ii) Компонент точки начала репликации
Экспрессирующие векторы содержат последовательность нуклеиновой кислоты, позволяющую реплицикацию вектора в одной или нескольких выбранных клетках-хозяевах. Такие последовательности хорошо известны для множества бактерий. Точка начала репликации из плазмиды pBR322 подходит для большинства грамотрицательных бактерий, таких как E. coli.
(iii) Компонент селективного гена
Экспрессирующие векторы, как правило, содержат селективный ген, называемый также селективным маркером. Данный ген кодирует белок, необходимый для выживания или роста трансформированной клетки-хозяина на селективной культуральной среде. Клетка-хозяин, не трансформированная вектором, содержащим селективный ген, не будет выживать в культуральной среде. Данный селективный маркер является отдельным от генетических маркеров, как применяют и определяют по настоящему изобретению. Типичные селективные гены кодируют белки, которые (a) придают устойчивость к антибиотикам или другим токсинам, например ампициллину, неомицину, метотрексату или тетрациклину, (b) восполняют ауксотрофные недостаточности, отличные от вызванных присутствием генетического маркера(маркеров), или (c) снабжают критическими питательными веществами, недоступными из составной среды, например ген, кодирующий рацемазу D-аланина для Bacilli.
В одном из примеров схемы отбора применяют вещество для ареста роста клетки-хозяина. В данном случае, клетки, успешно трансформированные интересующей нуклеиновой кислотой, продуцируют полипептид, сообщающий устойчивость к веществу, и таким образом выживают в режиме селекции. В примерах такой доминантной селекции применяют вещества неомицин (Southern et al, J. Molec. Appl. Genet., 1: 327 (1982)), микофеноловую кислоту (Mulligan et al., Science, 209: 1422 (1980)) или гигромицин (Sugden et al., Mol. Cell. Biol., 5:410-413 (1985)). В трех данных выше примерах применяют бактериальные гены под эукариотическим контролем для передачи устойчивости к подходящему веществу G418 или неомицину (генетицину), xgpt (микофеноловой кислоте), или гигромицину, соответственно.
(iv) Компонент промотора
Экспрессирующий вектор для получения интересующего полипептида содержит подходящий промотор, узнаваемый E. coli, и функционально связанный с нуклеиновой кислотой, кодирующей интересующий полипептид. Промоторы, пригодные для применения в хозяевах - E. coli, включают в себя промоторные системы бета-лактамазы и лактозы (Chang et al., Nature, 275: 615 (1978); Goeddel et al., Nature, 281: 544 (1979)), промоторную систему арабинозы (Guzman et al., J. Bacteriol., 174: 7716-7728 (1992)), щелочной фосфатазы, промотор T7, промоторную систему триптофана (trp) (Goeddel, Nucleic Acids Res., 8: 4057 (1980) и EP 36776) и гибридные промоторы, такие как промотор tac (deBoer et al., Proc. Natl. Acad. Sci. USA, 80: 21-25 (1983)). Однако пригодны другие известные бактериальные промоторы. Их нуклеотидные последовательности опубликованы, таким образом позволяя специалисту в данной области функционально лигировать их с ДНК, кодирующей интересующий полипептид (Siebenlist et al., Cell, 20: 269 (1980)) с применением линкеров или адаптеров для добавления любых требуемых участков рестрикции.
Предпочтительно, промотор, применяемый здесь, представляет собой индуцибельный промотор, т.е. активируемый индуцирующим средством или состоянием (таким как истощение периплазматического фосфата). Предпочтительные здесь такие индуцибельные промоторы представляют собой промотор щелочной фосфатазы, промотор tac или промотор T7.
Промоторы для применения в бактериальных системах также, как правило, содержат последовательность Шайна-Дальгарно (S.D.), функционально связанную с ДНК, кодирующей интересующий полипептид. Промотор можно удалить из ДНК из бактериального источника посредством расщепления рестрикционным ферментом и вставить в вектор, содержащий желаемую ДНК.
(v) Конструирование и анализ векторов
Конструирование подходящих векторов, содержащих один или несколько перечисленных выше компонентов, предусматривает общепринятые способы лигирования. Выделенные плазмиды или фрагменты ДНК расщепляют, достраивают и лигируют вновь в форме, желательной для получения требуемых плазмид.
Для анализа для подтверждения правильных последовательностей сконструированных плазмид смеси для лигирования применяют для трансформации E. coli K12 штамма 294 (ATCC 31446) или других штаммов, и получившиеся трансформанты отбирают по устойчивости к ампициллину или тетрациклину, где подходит. Плазмиды из трансформантов выделяют, анализируют расщеплением рестрикционными эндонуклеазами и/или секвенируют по способу Sanger et al., Proc. Natl. Acad. Sci. USA, 74: 5463-5467 (1977) или Messing et al., Nucleic Acids Res., 9: 309 (1981), или по способу Maxam et al., Methods in Enzymology, 65: 499 (1980).
C. Выбор и трансформация клеток-хозяев
Хозяева - E. coli, пригодные в качестве родительских хозяев для экспрессирующих плазмид здесь, включают в себя E. coli W3110 (ATCC 27325), E. coli 294 (ATCC 31446), E coli B и E. coli X1776 (ATCC 31537). Данные примеры являются скорее иллюстративными, чем лимитирующими. Мутантные клетки любого из вышеупомянутых штаммов можно применять также в качестве исходных хозяев, в которые затем дополнительно вводят мутации, чтобы они содержали по меньшей мере минимальный генотип, требуемый здесь. E. coli штамма W3110 является предпочтительным родительским штаммом, поскольку он является общепринятым штаммом-хозяином для ферментаций рекомбинантного продукта ДНК. Примеры исходных хозяев - E. coli для применения в качестве родительских хозяев вместе с их генотипами включены в таблицу ниже:


Также пригодными являются промежуточные продукты получения штамма 36F8, т.е. 27B4 (U.S. Pat. No 5304472) и 35E7 (изолят спонтанной теплостойкой колонии, растущий лучше, чем 27B4). Дополнительным подходящим штаммом является штамм E. coli с мутантной периплазматической протеазой(протеазами), описанный в U.S. Pat. No 4946783, изданном 7 августа 1990.
В одном из вариантов осуществления применяемая клетка-хозяин E. coli представляет собой дикий тип по отношению к гену glpT или в связи с ним, такой как 43E7, или является дефектной по гену glpT, такой как 43F6 или 61G1. В другом варианте осуществления, применяемая клетка-хозяин E. coli представляет собой дикий тип по отношению к гену phoA или в связи с ним. В предпочтительном варианте осуществления E. coli является дефектной по хромосомальному phoA. В другом предпочтительном варианте осуществления E. coli является дефектной по хромосомальному glpT и по хромосомальному phoA. В более предпочтительном варианте осуществления E. coli является дефектной по хромосомальному glpT и по хромосомальному phoA, но не по хромосомальному ugp. Наиболее предпочтительными из данных мутантных хозяев - E. coli являются 43F6 или 61G1, генотипы которых даны в таблице выше. Как применяют здесь, «дикий тип по отношению к glpT» относится к хозяевам - E. coli, представляющим собой glpT+ или glpT-компетентные клетки, т.е. не являющимся дефектными по хромосомальному glpT. Подобным образом, как применяют здесь, «дикий тип по отношению к phoA» относится к хозяевам - E. coli, представляющим собой phoA+ или phoA-компетентные клетки, т.е. не являющимся дефектными по хромосомальному phoA.
Штаммы по настоящему изобретению можно получать хромосомной интеграцией родительского штамма или другими способами, включая изложенные в примерах ниже.
Нуклеиновую кислоту, кодирующую полипептид, вставляют в клетки-хозяева. Предпочтительно, это осуществляют трансформацией клеток-хозяев описанными выше экспрессирующими векторами и культивированием в общепринятой питательной среде, модифицированной подходящим образом для индукции различных промоторов.
Трансформация означает введение ДНК в организм, так что ДНК является реплицируемой, или в качестве внехромосомного элемента, или посредством интеграции в хромосому. В зависимости от применяемой клетки-хозяина трансформацию осуществляют с применением обычных способов, подходящих для данных клеток. Обработку кальцием с применением хлорида кальция, как описано в разделе 1.82 Sambrook et al., Molecular Cloning: A Laboratory Manual (New York: Cold Spring Harbor Laboratory Press, 1989), обычно применяют для прокариотических клеток или других клеток, содержащих подходящие барьеры клеточной стенки. По другому способу трансформации применяют полиэтиленгликоль/DMSO, как описано в Chung and Miller, Nucleic Acids Res., 16: 3580 (1988). Еще одним способом является применение техники, называемой электропорацией.
D. Культивирование клеток-хозяев
Клетки E. coli, применяемые для получения интересующего полипептида, культивируют в подходящей среде, как описано в основном в Sambrook et al. выше. Условия культивирования, такие как температура, pH, и подобные, являются такими, какие применяли предварительно для выбранной для экспрессии клетки-хозяина, и очевидны обычному специалисту в данной области.
Клетки культивируют при подпитке культуральной среды транспортабельным органофосфатом, таким как глицерофосфат, например, альфа-глицерофосфат и/или бета-глицерофосфат, и особенно глицерол-2-фосфат и/или глицерол-3-фосфат. Культивирование можно проводить во встряхиваемой колбе или ферментере, предпочтительно в ферментере. Полипептид предпочтительно выделяют из цитоплазмы, периплазмы или культуральной среды клеток.
По способу по настоящему изобретению экспрессию нуклеиновой кислоты можно начинать в любой фазе стадии культивирования. Однако предпочтительно экспрессию нуклеиновой кислоты начинают, когда плотность клеток все еще увеличивается. Этого можно достичь индукцией промотора подходящим индуктором или индуцирующим условием до прекращения роста клеток.
Скорость подачи органофосфата в культуральную среду, применяемая для максимальной продукции полипептида, зависит от многих факторов, включая тип органофосфата, концентрацию органофосфата, тип продуцируемого полипептида, тип промотора, применяемый штамм клетки-хозяина и плотность клеток в культуре. Если полипептид представляет собой IGF-I, и органофосфат представляет собой глицерол-3-фосфат, предназначенный для продления продолжительности продукции, при описанных условиях культивирования и с применением 10-л-процесса, скорость подачи органофосфата предпочтительно составляет приблизительно от 1 до 7 ммоль/час на приблизительно 8-10 литров (см. фиг.4), более предпочтительно приблизительно от 1 до 6 ммоль/час, и еще более предпочтительно приблизительно от 2 до 6 ммоль/час, еще более предпочтительно приблизительно от 2 до 5 ммоль/час, и наиболее предпочтительно приблизительно от 3 до 4 ммоль/час. Оптимальная скорость подачи зависит от способа, плотности клеток, интенсивности дыхания, и т.д.
Также в предпочтительном варианте осуществления, где полипептид представляет собой Apo2L и органофосфат представляет собой глицерол-3-фосфат, предназначенный для сдвига экспрессии продукта для совпадения с фазой активного роста, с применением 10-л-процесса, скорость подачи органофосфата составляет приблизительно от 4 до 17 ммоль/час на приблизительно 8-10 литров (см. фиг.7), более предпочтительно приблизительно от 6 до 16 ммоль/час, еще более предпочтительно приблизительно от 8 до 15 ммоль/час, и наиболее предпочтительно приблизительно от 10 до 14 ммоль/час. Оптимальную скорость подачи органофосфата нужно определять для отдельного процесса, применяемого для экспрессии специфического гетерологичного белка.
Можно вводить также любые другие необходимые ингредиенты среды помимо источников углерода, азота и неорганического фосфата, в подходящих концентрациях, вводимые самостоятельно или в смеси с другим компонентом среды, таким как комплексный источник азота. Предпочтительно неорганический фосфат также присутствует в культуральной среде в начале стадии культивирования. Если присутствует такой неорганический фосфат, предпочтительно фосфат натрия и/или калия, соотношение неорганического фосфата к органофосфату зависит от таких факторов, как тип экспрессируемого полипептида и применяемый органофосфат. Соотношение может представляет собой любую пропорцию, как легко определяют специалисты в данной области, обычно в диапазоне приблизительно от 1:10 (одна часть Pi на 10 частей органофосфата) до 1:0,25. Для лиганда Apo2 оно предпочтительно лежит в диапазоне приблизительно от 1:4 до 1:0,25, и более предпочтительно приблизительно от 1:3 до 1:0,5, и еще более предпочтительно приблизительно от 1:3 до 1:1, и еще более предпочтительно приблизительно от 1:2 до 1:1, и наиболее предпочтительно приблизительно 1:1. Данные соотношения позволяют более раннюю индукцию экспрессии белка, и в некоторых случаях позволяют раньше производить больше продукта. pH среды может представлять собой любой pH от приблизительно 5-9, в основном, в зависимости от организма-хозяина.
Если промотор представляет собой индуцибельный промотор, чтобы произошла индукция, клетки обычно культивируют до достижения конкретной оптической плотности, например, A550 приблизительно 200 для процесса с высокой плотностью клеток, и в данной точке инициируют индукцию (например, добавлением индуктора, истощением компонента среды, и т.д.), для индукции экспрессии гена, кодирующего интересующий полипептид.
Где применяют промотор щелочной фосфатазы, клетки E. coli, применяемые для продукции интересующего полипептида по настоящему изобретению, культивируют в подходящей среде, в которой можно индуцировать промотор щелочной фосфатазы, как в основном, описано, например, в Sambrook et al., выше. Сначала среда может содержать неорганический фосфат для роста бактерий в количестве, достаточно большом, чтобы поддерживать значительный рост клеток и избежать индукции синтеза заданного гетерологичного полипептида под контролем промотора. По мере роста клеток и утилизации фосфата, у них уменьшается уровень неорганического фосфата в среде, таким образом, вызывая индукцию синтеза полипептида при истощении неорганического фосфата. Добавлением, например, подпитки, составленной из смеси G2P и G3P, или подпитки G3P, дальнейший рост до более высокой плотности клеток, такой как вплоть до 200 OD550 или выше, происходит в отсутствие неорганического фосфата или при уровнях голодания по неорганическому фосфату в периплазме и поддерживающей культуру среде, приводя к увеличению или расширению накопления продукта.
E. Детекция экспрессии
Экспрессию гена в образце можно измерять непосредственно, например, общепринятым нозерн-блоттингом для количественной оценки транскрипции мРНК (Thomas, Proc. Natl. Acad. Sci. USA, 77: 5201-5205 (1980)), дот-блоттингом (анализом РНК) или гибридизацией in situ с применением подходящим образом меченного зонда, основанного на последовательностях, кодирующих полипептид. Можно применять различные метки, наиболее распространены радиоизотопы, особенно 32P. Однако можно применять другие способы, такие как применение модифицированных биотином нуклеотидов для введения в полинуклеотид. Затем биотин служит участком связывания авидина или антител, которые можно метить широким множеством меток, таких как радиоизотопы, люминофоры, ферменты и подобные. Альтернативно для детекции белка можно применять анализы в гелях.
Для секреции продукта экспрессированного гена клетку-хозяин культивируют в условиях, подходящих для секреции продукта гена. Данные условия включают в себя, например, условия температуры, питательного вещества и плотности клеток, позволяющие секрецию из клетки. Кроме того, данные условия являются такими, при которых клетка может осуществлять основные клеточные функции транскрипции, трансляции и переноса белков от одного клеточного компартмента к другому, как известно специалистам в данной области.
F. Очистка полипептидов
Следующие способы, по отдельности или в сочетании, являются примерами подходящих способов очистки с конкретным применяемым способом(способами), зависящим от типа полипептида: фракционирование на иммуноаффинных или ионообменных колонках; осаждение этанолом; обращеннофазовая HPLC; хроматография с гидрофобным взаимодействием; хроматография на силикагеле; хроматография на ионообменной смоле, такой как S-SEPHAROSE™ и DEAE; хроматофокусирование; SDS-PAGE; осаждение сульфатом аммония; и гель-фильтрация с применением, например, наполнителя SEPHADEX™ G-75. Моноклональные антитела можно соответствующим образом отделять от культуральной среды посредством общепринятых способов очистки антитела, таких как, например, на носителе белок A-SEPHAROSE™, хроматографией на гидроксиапатите, электрофорезом в геле, диализом или аффинной хроматографией.
Изобретение можно более полно понять в отношении следующих примеров. Однако их не следует истолковывать как ограничивающие объем и сущность изобретения. Все цитаты из литературы и патентов приведены здесь в качестве ссылки.
Пример 1
Подпитка культуры G3P во встряхиваемой колбе для продукции фрагмента (тяжелой цепи) антитела ламы и Apo2L
Предпосылки
Включение 200 мМ G3P (конечная концентрация) в низкофосфатную (CRAP) или в высокофосфатную культуральную среду (THCD) сравнивали с соответствующей контрольной добавкой (вода) для экспрессии гетерологичного белка в культуре во встряхиваемой колбе. В первой части данного примера заданный гетерологичный белок представляет собой 13кДа анти-HCG ламы - монотело верблюдовых. Ранее показано, что верблюдовые обладают антителами 2 видов, классической молекулой IgG, состоящей из двух тяжелых плюс двух легких цепей, и молекулой IgG из тяжелой цепи с отсутствующей легкой цепью, на которую ссылаются как на монотело. Монотело верблюдовых экспрессировали посредством BL2l, штамма E. coli B, с применением промотора tac в низкофосфатной (CRAP)- или высокофосфатной (THCD)-богатой среде. Сигнальная последовательность связывающего белка malE, предшествующая последовательности, кодирующей фрагмент антитела, направляет секрецию экспрессированного белка в периплазму хозяина. Во второй части данного примера промотор T7 применяли для регуляции экспрессии лиганда Apo2 в HMS174, штамме E. coli K12, в G3P-дополненной и не дополненной среде CRAP. Продукцию гетерологичного белка в обоих экспериментах индуцировали добавлением IPTG при достижении желаемой плотности клеток.
Материалы и методы:
Конструкция плазмиды pCB36624_86.RIG:
pCB36624_86.RIG конструировали модификацией вектора pL1602 (Sidhu et al, J. Mol. Biol., 296:487-495 (2000)). Вектор pS1602, обладающий последовательностью промотора pTac и сигнальной последовательностью секреции malE, содержал последовательность человеческого гормона роста, слитую с C-концевым доменом гена-3 минорного белка оболочки (p3) фага mu. Последовательность, кодирующую hGH, удаляли, и полученная последовательность вектора служила каркасом вектора для вставки синтетического фрагмента ДНК, кодирующего анти-HCG-антитело ламы (Spinelli et al., Nat. Struct. Biol. 3(9): 752-757 (1996)). Полученная фагмида (pCB36624) кодировала гибридный продукт под контролем IPTG-индуцибельного промотора Ptac (Amman and Brosius, Gene, 40: 183-190 (1985)). Экспрессированный полипептид содержал сигнальный пептид связывающего мальтозу белка с последующей кодирующей областью анти-HCG, с последующей меткой - эпитопом FLAG, с последующим Gly/Ser-богатым линкерным пептидом, содержащим супрессируемый стоп-кодон, с последующим P3C (C-концевым доменом белка оболочки фага).
Библиотеки фаговых дисплеев конструировали с применением способа Sidhu et al., J. Mol. Biol., 296: 487-495 (2000) с надлежащим образом сконструированными фагмидами - «остановочными матрицами». Для библиотеки NNS17 производное pCB36624, содержащее стоп-кодоны TAA на месте кодонов 93, 94, 100 и 101, применяли в качестве матрицы для способа мутагенеза по Kunkel (Kunkel et al., Methods Enzymol., 154: 367-382 (1987)) с мутагенным олигонуклеотидом NNS17, сконструированным, чтобы одновременно исправлять стоп-кодоны и вводить 17 вырожденных кодонов NNK между кодонами, кодирующими Gly95 и Trp 103.
NNS17 : GCC GTC TAT ACT TGT GGT GCT GGT NNS NNS NNS NNS NNS NNS NNS NNS NNS NNS NNS NNS NNS NNS NNS NNS NNS TGG GGT CAG GGT (SEQ ID NO:3)
Подобно всем монотелам, анти-HCG ламы является членом семейства Vh3, и, таким образом, его узнает белок A. Связывающее взаимодействие с белком A применяли как заменитель CDR3-опосредованной стабильности. Полученные фаговые библиотеки сортировали посредством нескольких циклов против белка A как показателя стабильности каркаса и экспрессии. Сортированные библиотеки анализировали на селективное смещение в распределении аминокислот в библиотеке NNS. Каркас RIG, названный по последовательности в положениях 96, 97 и 98, оказался наиболее преобладающим клоном на основании секвенированных остатков. Последовательность CDR3 длиной 17 аминокислот для каркаса RIG определили являющейся RIGRSVFNLRRESWVTW (SEQ ID NO:4). Фагмиду с каркасом RIG переименовали в pCB36624_86.RIG с последовательностью ДНК:

Конструкция плазмиды pet19b.nohis
С применением общепринятых способов молекулярной биологии кодоны 114-281 Apo2L амплифицировали посредством полимеразной цепной реакции из полноразмерного клона Apo2L, выделенного из человеческой плацентарной кДНК. Дополнительные нуклеотиды, содержащие участки рестрикции для облегчения клонирования, добавляли к 5'- и 3'-последовательностям соответственно. 5' олигонуклеотидный праймер обладал последовательностью:
5' GCTTGCTACATATGGTGAGAGAAAGAGGTCCTCAGAGA 3' (SEQ ID NO:6),
содержащей подчеркнутый участок рестрикции Nde I. 3' олигонуклеотидный праймер обладал последовательностью: 5' CTTGAATAGGATCCCTATTAGCCAACTAAAAAGGCCCCAAAAAAACTGGC 3' (SEQ ID NO:7),
содержащей подчеркнутый участок рестрикции BamH I. Полученный фрагмент субклонировали с применением участков рестрикции Nde I и BamH I в модифицированный бакуловирусный экспрессирующий вектор pVL1392 (Pharmingen) в рамке и ниже последовательности, содержащей метку His 10 и сайт расщепления энтеропептидазой (Pitti et al., J. Biol. Chem., 271:12687-12690 (1997)). PVL1392-Apo2L расщепляли Nde I и BamH I, и полученный фрагмент Nde I-BamH I субклонировали в pET-l9b (Novagen), также расщепленный Nde I и BamH I. Полученную плазмиду назвали petl9b.nohis.
Бактериальные штаммы:
Компетентные клетки BL2l (Stratagene) и HMS174 (Merck) трансформировали pCB36624_86.RIG и pet19b.nohis соответственно с применением общепринятых способов. Трансформанты отбирали после роста на LB-чашке, содержащей 50 мкг/мл карбенициллина (LB+ карбенициллин CARB50™), очищали штрихами и выращивали в среде LB с 50 мкг/мл карбенициллина CARB50™ в инкубаторе при 30°C. pCB36624_86.RIG сообщала устойчивость к карбенициллину полученному хозяину BL21/pCB36624_86.RIG, а pet19b.nohis - HMS174/pet19b.nohis, позволяя трансформированным хозяевам расти в присутствии антибиотика.
Среда для ферментации:
И низкофосфатную (CRAP) культуральную среду, и высокофосфатную (THCD) культуральную среду применяли для оценки продукции фрагмента антитела ламы и лиганда Apo2. Состав среды (с количествами каждого компонента, применяемыми на литр исходной среды) перечислен ниже:
до pH 7,3
Для получения 200 мМ G3P-дополненной среды 5 мл 1 М DL-альфа-глицерофосфата (G3P) (Sigma Chem. Co.) добавляли к 20 мл среды с низким PO4 с 50 мкг/мл карбенициллина (среда с низким PO4 + карбенициллин CARB50™) или среды с высоким PO4 с 50 мкг/мл карбенициллина (среда с высоким PO4 + карбенициллин CARB50™) перед инокуляцией. Для недополненной (контрольной) среды вместо G3P применяли 5 мл воды.
Ферментация во встряхиваемой колбе:
Ферментацию во встряхиваемой колбе проводили в 125-мл колбе с отражателем, содержащей 25 мл контрольной или G3P-дополненной среды. Ночную культуру BL21/pCB36624_86.RIG или HMS174/pet19b.nohis, выращенную на LB+карбенициллин CARB50™, снова разводили приблизительно 1:100 для инокуляции в контрольную или G3P-дополненную среду. Культуры инкубировали при 30°C во встряхивателе при 250 об/мин, и экспрессию продукта индуцировали добавлением 1 мМ IPTG, когда плотность клеток достигала приблизительно 50-60% потенциального роста клеток, поддерживаемого средой. Получали осадки клеток из 1 мл жидкой культуры, взятые непосредственно перед добавлением индуктора и приблизительно через 24 часа после инокуляции и хранили при -20°C.
Накопление фрагмента антитела ламы, анализируемое посредством PAGE и денситометрии:
Замороженный (-20°C) осадок клеток, полученный из 1 мл образца культуры, оттаивали и ресуспендировали в достаточном количестве l0 мМ Трис, pH 7,6 + 1 мМ ЭДТА, pH 8,0 (TE) для доведения суспензии клеток до концентрации 1 OD/25 мкл. 25 мкл суспензии клеток в TE смешивали с 25 мкл 2X буфера для нанесения образцов, содержащего бета-меркаптоэтанол. Смесь нагревали при >95°C в течение 5 мин перед нанесением 10 мкл (эквивалент 0,2 OD) на лунку в предварительно залитый 10% Бис-Трис гель NU-PAGE™ (Novex). Электрофорез проводили в буфере MES (2-(N-морфолино)этансульфоновая кислота в деионизированной воде, доведенная до подходящего pH, например, 1 Н NaOH). После разделения гель окрашивали COOMASSIE BLUE R250™, и затем удаляли окраску. Интенсивность полосы 13-кДа фрагмента антитела определяли с применением программного обеспечения для обработки изображений Kodak DIGITAL SCIENCE ID™ после сканирования влажного геля системой для обработки изображений Kodak.
Накопление лиганда Apo2, анализируемое посредством обращеннофазовой HPLC:
Замороженный (-20°C) осадок клеток, полученный из 1 мл образца культуры, оттаивали и ресуспендировали в достаточном количестве буфера TE для доведения суспензии клеток до концентрации 1 OD/25 мкл. 20 мкл суспензии клеток смешивали с 480 мкл 6 М гуанидина-HCl, pH 9,0 + 100 мМ дитиотреитола (DTT), и инкубировали при комнатной температуре в течение часа перед центрифугированием при 13000 об/мин в течение 15 мин для получения вновь супернатанта/экстракта. Экстракт фильтровали через центрифугируемый фильтр MILLIPORE™, перед тем как 20 мкл наносили на колонку HPLC (наполнитель PerSeptive Biosystems POROS® R1/10) для обращеннофазовой хроматографии. Разделение HPLC проводили при 80°C с подвижными фазами, протекающими при 1,0 мл/мин, и применяли градиент от 28% до 35% ацетонитрила с 0,1% TFA в течение 20 минут для отделения Apo2L от загрязняющих белков. Пики детектировали при длине волны 280 нм. Количество мономера, присутствующего в образцах, вычисляли с применением среднего фактора отклика (мЕА/мкг), выведенного из площади под пиком, соответствующим 5-20 мкг очищенных стандартов, анализируемых таким же способом.
Результаты:
На фиг.1 показано, что антитело экспрессировано на более высоком уровне и в среде с высоким PO4 (THCD), и в среде с низким PO4 (CRAP), дополненной 200 мМ G3P, по сравнению с контролем.
На фиг.2 показано, что белок Apo2L экспрессирован на более высоком уровне в среде с низким PO4 (CRAP), дополненной 200 мМ G3P, по сравнению с контролем.
Пример 2
Подпитка G3P в 10-л ферментере культуры хозяина дикого типа или (ΔglpTphoA-ugp+) для продукции IGF-I, регулируемой промотором щелочной фосфатазы
Материалы и методы
Плазмида pBKIGF-2B для экспрессии IGF-I:
Плазмиду pBKIGF-2, применяемую для экспрессии IGF-I здесь, конструировали, как подробно описано в U.S. Pat. No 5342763. Данную плазмиду конструировали из основного каркаса pBR322. Транскрипционные и трансляционные последовательности, необходимые для экспрессии гена IGF-I в E.coli, предоставляли посредством промотора щелочной фосфатазы и последовательностей Шайна-Дальгарно trp. Терминатор транскрипции лямбда tO располагали рядом с терминирующим кодоном IGF-I. Секрецию белка из цитоплазмы направляли сигнальной последовательностью lamB. Большинство rhIGF-I обнаружили в периплазматическом пространстве клетки. Плазмида pBKIGF-2B сообщала устойчивость к тетрациклину трансформированному хозяину.
Бактериальные штаммы и условия роста:
Хозяева, применяемые для ферментации IGF-I, являются производными E. coli W3110 (Bachmann, Cellular and Molecular Biology, vol. 2 (Washington, D.C.: American Society for Microbiology, 1987), pp. 1190-1219). Эксперименты, относящиеся к хозяину с glpT дикого типа, проводили со штаммом 43E7 (E. coli W3110fhuA(tonA)Δ(argF-lac)ptr3 degP41 ΔompTΔ(nmpc-fepE)ilvG+ phoA), а эксперименты, относящиеся к хозяину с мутацией ΔglpT, проводили со штаммом 43F6 (E. coli W3110fhuA(tonA) Δ(argF-lac)ptr3 degP41 ΔompTΔ(nmpc-fepE)ilvG+ phoA ΔglpT). Компетентные клетки 43E7 или 43F6 трансформировали pBKIGF-2B с применением общепринятых способов. Трансформанты отбирали после роста на LB-чашке, содержащей 20 мкг/мл тетрациклина (LB + тетрациклин TET20™), очищали штрихами и выращивали в среде LB с 20 мкг/мл тетрациклина TET20™ во встряхивателе/инкубаторе при 37°C перед тестированием в ферментере. pBKIGF-2B сообщает устойчивость к тетрациклину продуцируемому хозяину и позволяет трансформированному хозяину расти в присутствии антибиотика.
Процесс 10-л ферментации:
Состав среды для ферментации и протокол прогона, применяемый для экспрессии IGF-I, являлись до некоторой степени сходными с применяемыми для процесса IGF-I, описанного в U.S. Pat. No 5342763. Кратко, инокулируемую культуру во встряхиваемой колбе 43E7/pBKIGF-2 или 43F6/pBKIGF-2 применяли для инокуляции в богатую среду для продукции. Состав среды (с количествами каждого компонента, применяемыми на литр исходной среды) перечислен ниже:
*Порцию глюкозы, дрожжевого экстракта, метионина и NZ амина AS первоначально добавляли в среду, остальное подавали в ходе ферментации.
10-литровая ферментация представляла собой подпитываемый процесс со следующими параметрами ферментации:
В экспериментах, предусматривающих подпитку глицерол-3-фосфатом (G3P), подходящее количество 1М маточного раствора G3P добавляют в комплексную подпитку азотом и последующее соотношение дополнительная подпитка-подпитка увеличивают для доставки желательного количества комплексного азота плюс G3P в культуру.
Влияние мутации ΔglpT с подпиткой G3P или без нее оценивали по разнице в накоплении IGF-I. Общее количество IGF-I в образце, растворенном в 6M гуанидине+100 мМ DTT, измеряли способом обращеннофазовой HPLC, как описано в U.S. Pat. No 6559122.
Результаты:
На фиг.3 показано, что для хозяина дикого типа (43E7) с промотором AP и непрерывной подпиткой глюкозой количество секретированного IGF-I было заметно выше, когда среду подпитывали G3P, чем когда G3P не добавляли.
На фиг.4 показано, что с хозяином ΔglpT (43F6) и промотором AP количество секретируемого IGF-I было заметно выше, когда культуру подпитывали G3P при 1,18 или 3,28 ммоль/час, на приблизительно 8 литров, чем когда G3P не добавляли, но не было выше, когда культуру подпитывали G3P при 8,22 ммоль/час, на приблизительно 8 литров. Оптимальную скорость подачи специалист в данной области может легко определить на основе продукта, типа органофосфата, и т.д. При условиях описанного процесса ферментации, культивирования в 10-литровом ферментере для производства IGF-I, существует оптимальная скорость подачи, приблизительно на 8-10 литров, в предпочтительном диапазоне приблизительно 1-7 ммоль/час, более предпочтительно приблизительно 1-6 ммоль/час, еще более предпочтительно приблизительно 2-6 ммоль/час, еще более предпочтительно приблизительно 2-5 ммоль/час, и наиболее предпочтительно приблизительно 3-4 ммоль/час. Данный диапазон скорости подачи не только увеличивает количество продукта по сравнению с контролем, но также расширяет продолжительность продукции относительно контроля.
Пример 3
Подпитка глицерол-3-фосфатом в 10-л процессе для улучшения накопления лиганда Apo2
Предпосылки для лиганда Apo2:
Апоптоз-индуцирующий лиганд 2 (Apo2L) (Pitti et al., J. Biol. Chem., 271: 12687-12690 (1996)), известный также как лиганд, индуцирующий апоптоз с участием фактора некроза опухолей (TRAIL) (Wiley et al., Immunity, 3: 673-682 (1995)), представляет собой мембранный белок типа II и член TNF-семейства лигандов. Apo2L/TRAIL запускает апоптоз в широком множестве опухолевых клеток, но не в большинстве нормальных клеток, посредством связывания с родственными ему рецепторами смерти (WO 99/00423; Ashkenazi, FASEB J., 13: (7) A1336 (April 23, 1999); Ashkenazi, Nature Reviews - Cancer, 2: 420-430 (2002)). Растворимый фрагмент внеклеточного домена лиганда Apo2, соответствующий аминокислотным остаткам 114-281 (отсюда в обозначенном Apo2L/TRAIL), в настоящее время подлежит изучению для потенциальных клинических исследований, и его успешно экспрессировали в E. coli.
Общее описание процесса ферментации:
Экспрессирующий вектор кодирует промотор щелочной фосфатазы (AP) для регуляции продукции приблизительно 19,5-кДа полипептида. Возникающие экспрессируемые полипептиды, при высвобождении из рибосом, сворачиваются в мономеры в цитоплазме и затем ассоциируют, становясь биологически активным гомотримером. В ходе ферментации параметры процесса устанавливают так, что клеточные активности проводят на пике скоростей поглощения кислорода приблизительно 3,0 ммоль/л-мин. После сбора культуры, удерживаемый в цитоплазме гетерологичный белок высвобождают посредством механического разрушения клеток в лизат клеток, откуда его можно выделить.
Материалы и методы:
Конструкция плазмиды pAPApo2-P2RU:
pAPApo2-P2RU описана в WO 01/00832, опубликованном 4 января 2001. Кратко, данная плазмида, конструкция которой показана на фиг.5, кодирует коэкспрессирующиеся Apo-2L (аминокислотные остатки 114-281) и тРНК для редких кодонов, кодируемые pro2 и argU, где коэкспрессию регулирует промотор щелочной фосфатазы. Основанную на pBR322 плазмиду (Sutcliffe, Cold Spring Harbor Symp. Quant. Biol., 43:77-90 (1978)), pAPApo2-P2RU, применяли для продукции Apo-2L в E. coli. Транскрипционные и трансляционные последовательности, необходимые для экспрессии Apo-2L, предоставлены промотором щелочной фосфатазы и последовательностью Шайна-Дальгарно trp, как описано для плазмиды phGH1 (Chang et al., Gene, 55:189-196 (1987)). Кодирующая последовательность для Apo-2L (от 114-281) локализована ниже последовательностей промотора и Шайна-Дальгарно, и ей предшествует инициирующий метионин. Кодирующая последовательность содержит нуклеотиды (показанные на фиг.6), кодирующие остатки 114-281 Apo-2L (фиг.6 - SEQ ID NO:1 и 2 соответственно для нуклеотидной и аминокислотной последовательностей), за исключением того, что кодон, кодирующий остаток Pro 119, заменен на «CCG» вместо «CCT» для удаления потенциальной вторичной структуры. Последовательность, кодирующая терминатор транскрипции лямбда t0 (Scholtissek et al., Nucleic Acids Res., 15: 3185 (1987)), следует за кодирующей последовательностью Apo-2L.
Кроме того, данная плазмида также содержит последовательности для экспрессии тРНК pro2 (Komine et al., J. Mol. Biol., 212:579-598 (1990)) и argU/dnaY (Garcia et al., Cell, 45:453-459 (1986)). Данные гены клонировали посредством ПЦР из E. coli W3110 и помещали ниже последовательности терминатора транскрипции лямбда t0. Данная плазмида сообщает продуцирующему хозяину устойчивость и к тетрациклину, и к ампициллину.
Бактериальные штаммы и условия роста:
Штамм 43E7 (E. coli W3110fhuA(tonA) phoA Δ(argF-lac)ptr3 degP ompT ilvG+) применяли в качестве продуцирующего хозяина дикого типа для сравнения с 43F6, glpT-мутантным хозяином, по экспрессии лиганда Apo2 и тРНК для редких кодонов. Компетентные клетки 43E7 или 43F6 получали и трансформировали pAPApo2-P2RU с применением общепринятых способов. Трансформанты отбирали после роста на LB-чашках, содержащих 20 мкг/мл тетрациклина (LB+Tet20), очищали штрихами и выращивали в среде LB с 20 мкг/мл тетрациклина во встряхивателе/инкубаторе при 30°C перед хранением в DMSO при -80°C.
Процесс ферментации для продукции Apo2L:
Инокулят во встряхиваемой колбе получали посредством засева инокуляции стерильной среды LB, содержащей 4-6 мМ фосфата натрия, маточной культурой из свежеразмороженной ампулы. Для осуществления селективного давления для обеспечения сохранения плазмиды в среду добавляли подходящие антибиотики. Культуры в колбах инкубировали со встряхиванием при приблизительно 30°C (28°C-32°C) в течение 14-18 часов. Затем данную культуру применяли для инокуляции ферментационного сосуда для продукции. Инокулируемый объем составлял между 0,1% и 10% исходного объема среды.
Продукцию Apo2L проводили в среде для продукции, данной в таблице 1, для достижения конечного объема культуры приблизительно 10 литров. Процесс ферментации проводили при приблизительно 30°C (28-32°C), и pH контролировали при приблизительно 7,0 (6,5 - 7,5). Скорость аэрации и скорость перемешивания устанавливали для обеспечения адекватного переноса кислорода к культуре. Непосредственно перед истощением загруженного фосфата (приблизительно при 75-85 OD) начинали подпитку DL-альфа-глицерофосфатом (поставщик в спецификации указывает чистоту продукта 80-90%, с бета-глицерофосфатом, указанным в качестве основной примеси) и подпитывали при желаемой скорости подачи. На всем протяжении процесса ферментации культуру клеток подпитывали глюкозой в качестве первичного источника углерода на основе компьютерного алгоритма для обеспечения аэробных условий.
На протяжении процесса ферментации проводили две дополнительные загрузки по приблизительно 50-150 мкМ (конечная концентрация) ZnSO4, одну непосредственно перед индукцией экспрессии продукта, другую приблизительно в середине периода продукции, для улучшения сборки гомотримера. В данном примере добавления проводили при оптической плотности приблизительно 80-120 OD550 и приблизительно через 28 часов после инокуляции.
Ферментацию проводили приблизительно 34-45 часов перед сбором.
Состав среды для продукции в экспрессирующей системе
с промотором AP
Оценка накопления растворимого продукта в ходе процесса ферментации посредством способа ионообменной хроматографии HPLC:
Образцы культуры отбирали на протяжении времени процесса ферментации. Клетки из 1 миллилитра образцов культуры, разведенных до плотности клеток 20 OD550, собирали центрифугированием, и полученные осадки клеток до анализа хранили при -20°C. Осадки клеток оттаивали, ресуспендировали в 0,5 мл буфера для экстракции (50 мМ HEPES, pH 8,0, 50 мМ ЭДТА и 0,2 мг/мл лизоцима яичного белка курицы) и механически разрушали для высвобождения продукта из цитоплазмы. Твердые вещества удаляли из лизатов клеток центрифугированием перед нанесением осветленных лизатов на колонку для HPLC (наполнитель DIONEX PROPAC™ IEX) для количественной оценки тримера. Способом анализа HPLC продукт отделяли от примесных белков E. coli посредством применения 5%-22% градиента 1М NaCl в 25-мМ фосфатном (pH 7,5) буфере в течение 25 минут при скорости потока 0,5 мл/мин.
Оценка общей экспрессии мономерного Apo2L в ходе процесса ферментации посредством обращеннофазовой хроматографии HPLC:
Для количественной оценки общей продукции мономера применяли свежую культуру или предварительно замороженные и затем оттаявшие образцы. 20 мкл образца смешивали с 480 мкл 6 М гуанидина-HCl, pH 9,0 с 100 мМ DTT и инкубировали при комнатной температуре в течение часа перед центрифугированием при 13000 об/мин в течение 15 мин для повторного получения экстракта. Экстракт фильтровали через центрифугируемый фильтр MILLIPORE™, перед тем как 20 мкл наносили на колонку HPLC (наполнитель PerSeptive Biosystems POROS® R1/10) для обращеннофазовой хроматографии. Разделение HPLC проводили при 80°C с подвижными фазами, протекающими при 1,0 мл/мин, и применяли градиент от 28% до 35% ацетонитрила с 0,1% TFA в течение 20 минут для отделения Apo2L от загрязняющих белков. Пики детектировали при длине волны 280 нм. Количество мономера, присутствующего в образцах, вычисляли с применением среднего фактора отклика (мЕА/мкг), выведенного из площади под пиком, соответствующим 5-20 мкг очищенных стандартов, анализируемых таким же способом.
Результаты:
На фиг.7 показаны улучшенные титры специфического продукта (выраженные как специфические титры в мкг/OD-мл на графике) с оптимальной скоростью подачи G3P к хозяину ΔglpT (43F6). Все прогоны с подпиткой G3P проходили лучше, чем прогоны без подпитки. В данном примере при увеличении скорости подачи от 6 до 12 ммоль/час для приблизительно 8-литровой культуры, титр специфического продукта улучшался, но при увеличении скорости выше от 12 ммоль/час до 18 ммоль/час специфический титр был ниже. Оптимальную скорость подачи G3P специалист в данной области может легко определить на основании продукта, типа органофосфата, и т.д. При данных конкретных условиях, культивировании в 10-литровом ферментере клеток для производства данного конкретного продукта, Apo2L, предпочтительная скорость подачи G3P, приблизительно на 8-10 литров, предпочтительно лежит в диапазоне приблизительно от 4 до 17 ммоль/час, более предпочтительно приблизительно от 6 до 16 ммоль/час, еще более предпочтительно приблизительно от 8 до 15 ммоль/час, и наиболее предпочтительно приблизительно от 10 до 14 ммоль/час.
На фиг.8 показан улучшенный титр специфического продукта (выраженный как специфическое общее накопление в мкг/OD-мл на графике) при подпитке G3P по сравнению с подпиткой неорганическим фосфатом для хозяина дикого типа glpT (43E7). В то время как подпитка глицерофосфатом увеличивает специфическое общее накопление Apo2L, подпитка неорганическим фосфатом отрицательно влияет на специфическое общее накопление по сравнению с контролем без подпитки. Подобных тенденций ожидают при применении более низкой подпитки глицерофосфатом, чем применяли. Результаты здесь предназначены, чтобы показать, и показывают, что высокий уровень экспрессии можно получить посредством подпитки глицерофосфатом хозяина дикого типа glpT. Кроме того, в данном конкретном эксперименте, так же как в случае подпитки неорганическим фосфатом, плотность клеток в культуре увеличивалась до более 200 OD550 при подпитке глицерофосфатом, но не в ситуации без подпитки.
Пример 4
Управляемая промотором AP экспрессия продукта Apo2L в фазе активного роста
Применяли такие же конструкции плазмид, продуцирующий штамм-хозяин, состав среды, процесс ферментации и способы анализа продукта, как описаны в примере 3, за исключением загрузки фосфата и добавления G3P. Порцию неорганического фосфата, обычно включаемую в солевую загрузку в контрольном процессе, заменяли эквивалентным числом молей G3P, добавляемых немедленно после инокуляции, или за несколько часов до истощения загруженного неорганического фосфата. В данных примерах ожидали, что добавленный G3P станет источником фосфата для значительной части роста клеток после добавления.
Процесс ферментации для продукции Apo2L в фазе активного роста:
Протокол подготовки инокулята являлся таким же, как описано в примере 3. Продукцию Apo2L проводили в среде для продукции, данной в таблице 1, за исключением того, что или 75%, или 50% фосфатных солей удаляли из начальной загрузки и заменяли эквивалентным количеством молей G3P, добавленным обратно в качестве дополнительной загрузки после инокуляции. Ферментацию проводили при приблизительно 30°C (28-32°C), и pH контролировали при приблизительно 7,0 (6,5-7,5), как при общепринятом протоколе. Скорость аэрации и скорость перемешивания составляли, как описано в примере 3. В случае, где 50% неорганического фосфата заменяли G3P, неорганический фосфат загружали перед стерилизацией среды, тогда как замену глицерол-3-фосфата проводили приблизительно за 1-2 часа перед ожидаемым окончанием загруженного фосфата (при приблизительно 30-40 OD550). В случае, где 75% неорганического фосфата заменяли G3P, и неорганический фосфат, и G3P добавляли немедленно после инокуляции ферментера. На всем протяжении процесса ферментации культуру клеток подпитывали глюкозой в качестве первичного источника углерода на основе компьютерного алгоритма для обеспечения аэробных условий. Добавления Zn проводили в ходе процесса ферментации, как описано в более раннем разделе. Ферментацию проводили приблизительно 34-45 часов.
Результаты:
На фиг.9 показана индукция экспрессии гетерологичного белка, происходящая значительно раньше в фазе активного роста, где 50%- 75% загруженного PO4 заменяют добавлением G3P и для хозяев дикого типа, и для glpT-мутантных хозяев, сдвигая кривую общего специфического накопления влево от дублированных контрольных случаев, проводимых для хозяина дикого типа без замены G3P. Это показывает преимущество настоящего изобретения, в котором продукт можно получить раньше в ходе процесса ферментации.
В то время как для всех соотношений Pi к G3P, тестированных здесь, получили их преимущества независимо от типа хозяина, в таблице 2 показано, что при применении соотношения Pi к G3P 1:1 или 1:3 для glpT-мутантного хозяина 43F6 получают наивысший уровень объемной производительности Apo2L (в среднем приблизительно 0,34 против в среднем приблизительно 0,24 мг/мл-час для контрольного хозяина). Кроме того, с применением любого соотношения и хозяина дикого типа или мутантного пика специфического накопления (в мкг/OD-мл) достигают раньше (от 22 до 26 часов по сравнению с от 28 до 30 часов). Это показывает, что в конкретных предпочтительных вариантах осуществления по изобретению можно достичь таких же, если не больших, количеств мономерного Apo2L за приблизительно на 10% - 25% меньшее время ферментации, чем иным способом, для значительного улучшения производительности процесса.
Эффект замены первоначальной загрузки неорганического фосфата добавлением глицерофосфата в течение первых 30 часов ферментации
(г/л)
Пример 5
Управляемая промотором AP экспрессия продукта Apo2L с применением 50/50 смеси альфа- и бета-глицерофосфата
Следовали способу, подобному описанному в примере 3, за исключением более дешевой стадии применения для загрузки приблизительно 50:50 смеси альфа- и бета-глицерофосфата вместо G3P с применением штамма 61G1 (glpT-мутантный хозяин).
Результаты:
На фиг.10 показано, что похожее улучшение выхода по сравнению с контролем без подпитки получали с применением смеси или вещества G3P более высокой степени очистки. Применение смеси альфа/бета будет уменьшать стоимость сырья без подвергания риску результатов продукции.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБЫ ПОЛУЧЕНИЯ ДВУЦЕПОЧЕЧНЫХ БЕЛКОВ В БАКТЕРИЯХ | 2015 |
|
RU2705274C2 |
| СПОСОБ ОЧИСТКИ ГЕТЕРОЛОГИЧНОГО ПОЛИПЕПТИДА | 2004 |
|
RU2337968C2 |
| СПОСОБЫ ПОЛУЧЕНИЯ ДВУЦЕПОЧЕЧНЫХ БЕЛКОВ В БАКТЕРИЯХ | 2015 |
|
RU2828630C2 |
| СПОСОБЫ ПОЛУЧЕНИЯ ДВУЦЕПОЧЕЧНЫХ БЕЛКОВ В БАКТЕРИЯХ | 2015 |
|
RU2739500C2 |
| СПОСОБЫ И КОМПОЗИЦИЯ ДЛЯ СЕКРЕЦИИ ГЕТЕРОЛОГИЧНЫХ ПОЛИПЕПТИДОВ | 2010 |
|
RU2585488C2 |
| СПОСОБ ПОЛУЧЕНИЯ ГЕТЕРОЛОГИЧНЫХ БЕЛКОВ | 2008 |
|
RU2487168C2 |
| СПОСОБЫ ПРИМЕНЕНИЯ АГОНИСТОВ АРО-2L-РЕЦЕПТОРОВ И АКТИВАТОРОВ NK-КЛЕТОК | 2005 |
|
RU2395294C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ СЕКРЕЦИИ ГЕТЕРОЛОГИЧНЫХ ПОЛИПЕПТИДОВ | 2015 |
|
RU2748026C2 |
| СПОСОБ ОЧИСТКИ АРО-2 LIGAND/TRAIL ПОСРЕДСТВОМ КРИСТАЛЛИЗАЦИИ НА ХОЛОДЕ | 2006 |
|
RU2435781C2 |
| НОВАЯ СИСТЕМА СЕЛЕКЦИИ | 2006 |
|
RU2415935C2 |





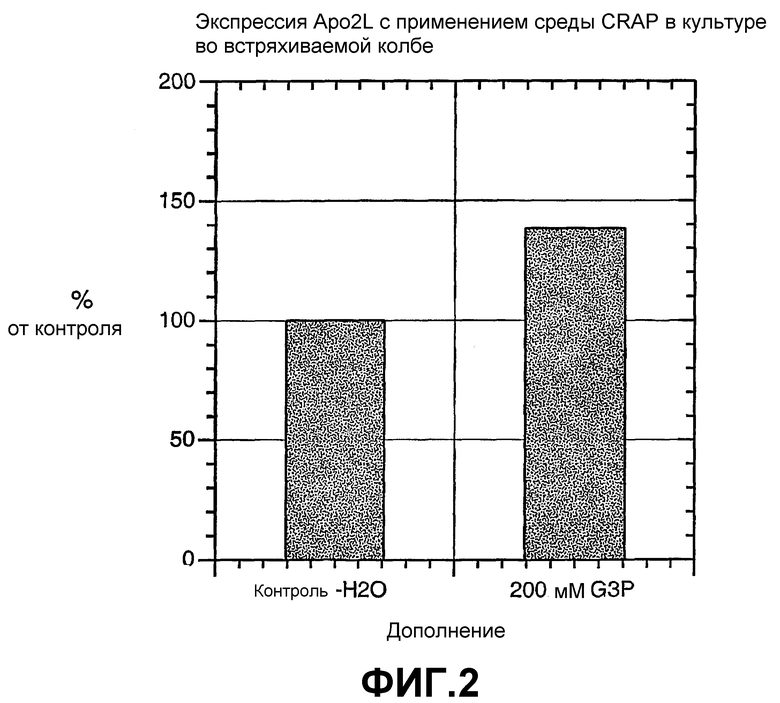
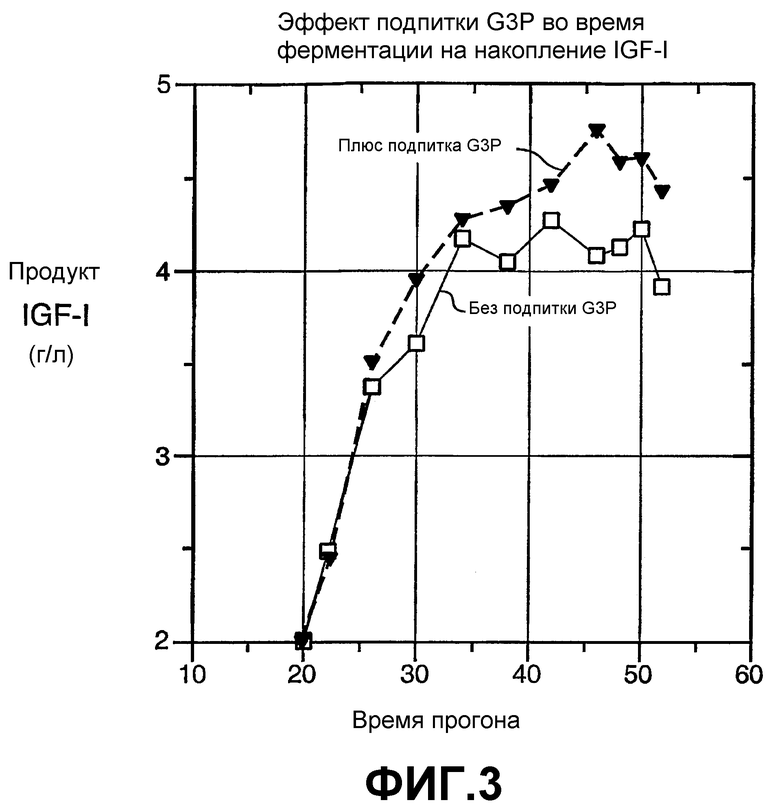
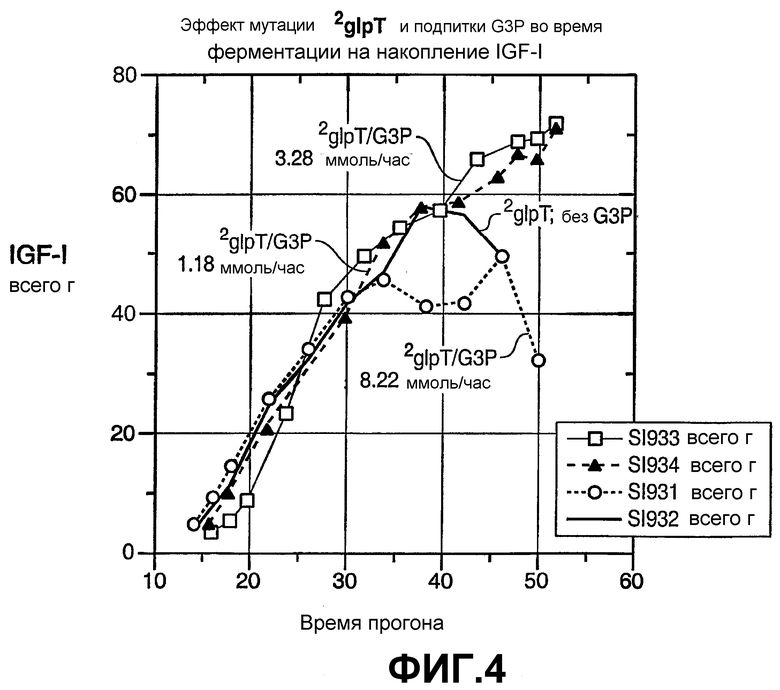
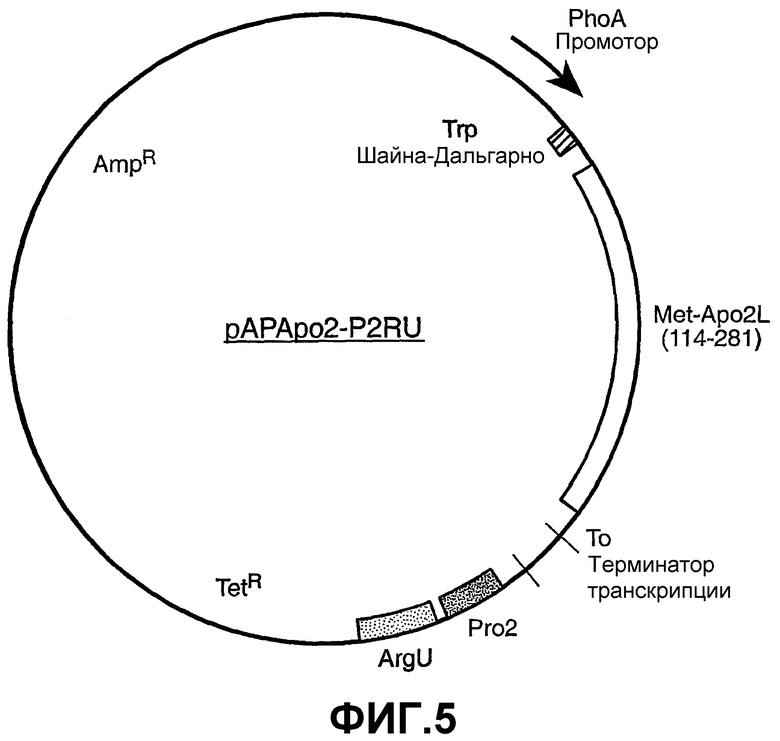

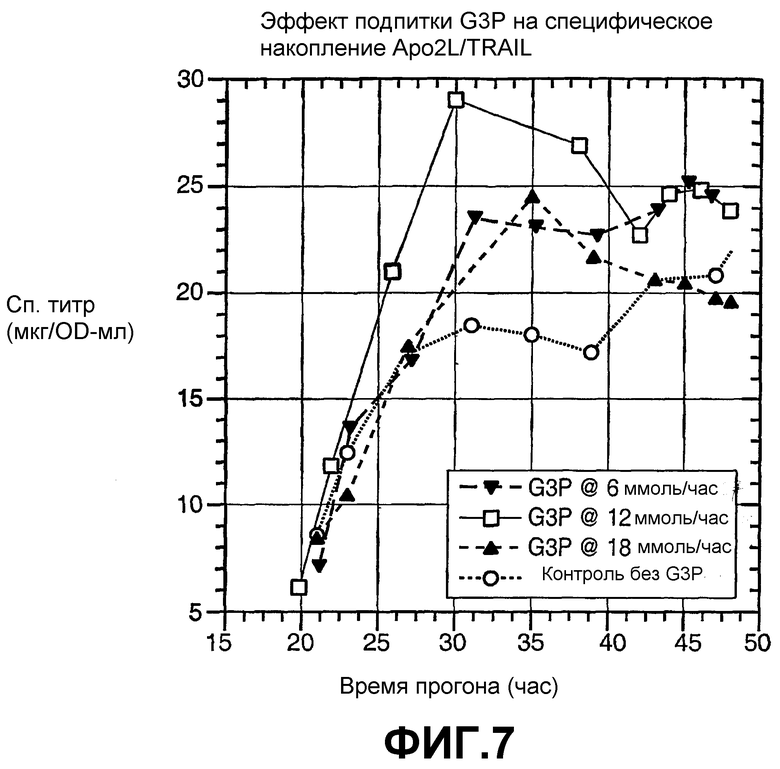
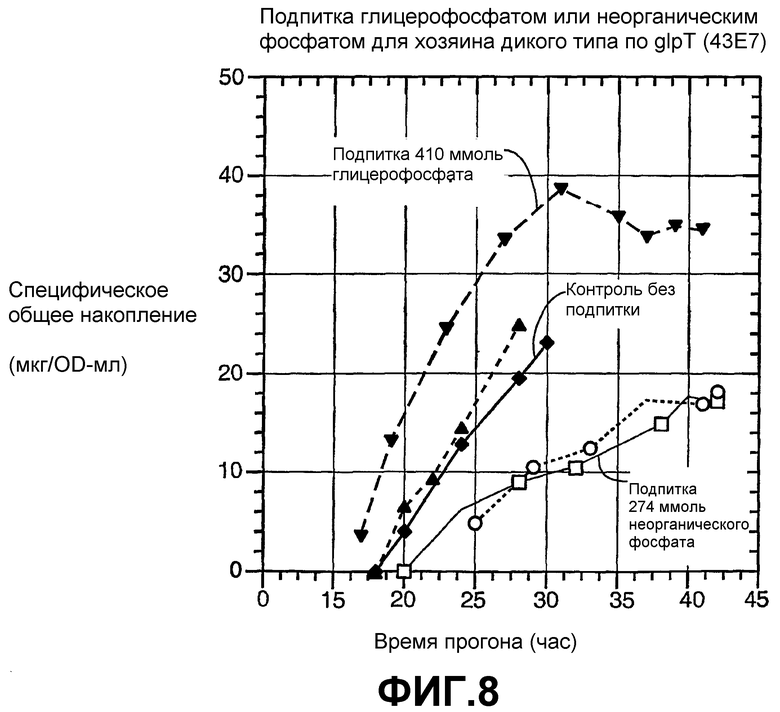
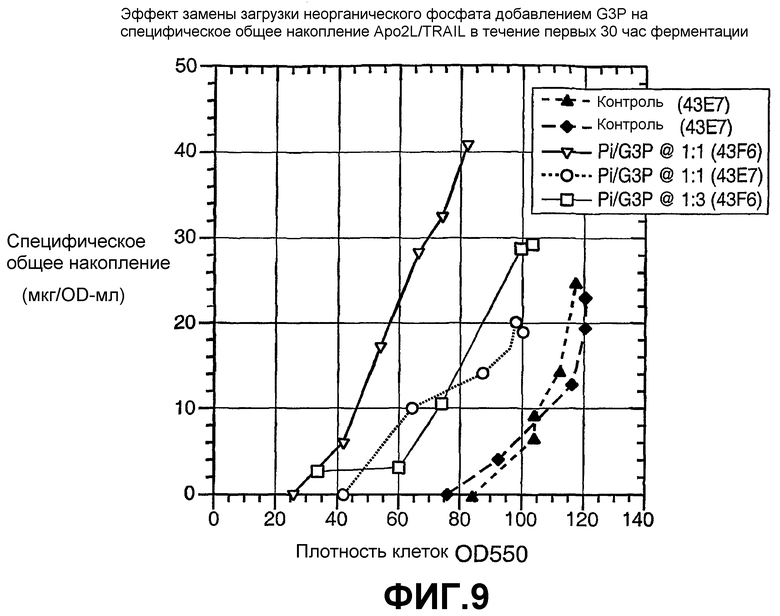
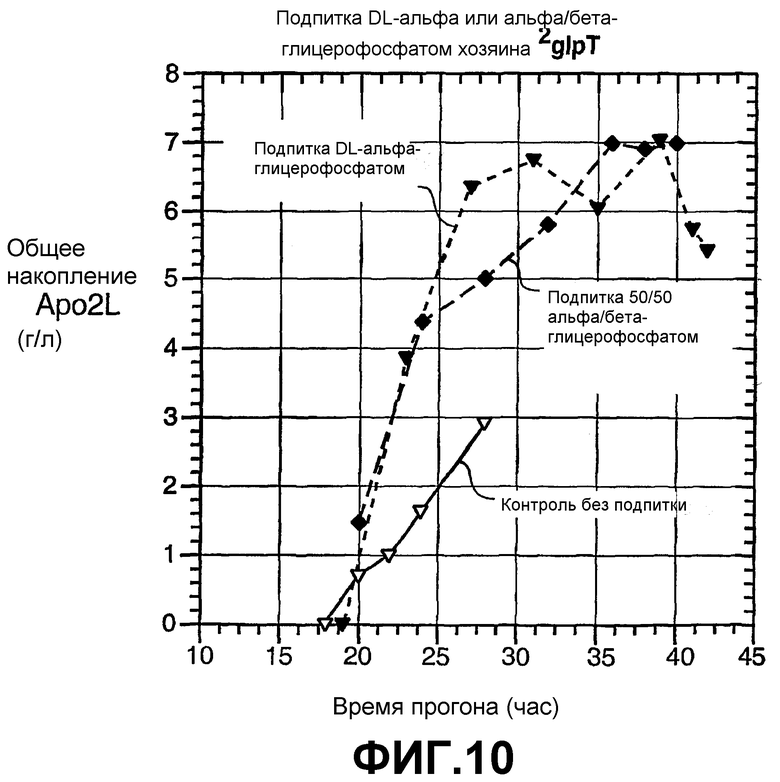
Изобретение относится к области биотехнологии и касается способа получения полипептида, гетерологичного для E.coli. Представленный способ включает: (а) культивирование клеток Е.coli, содержащих нуклеиновую кислоту, кодирующую полипептид, в культуральной среде, подпитываемой транспортабельным органофосфатом, выбранным из группы, сосотоящей из альфа-глицерофосфата, бета-глицерофосфата, глицерол-3-фосфата, и смеси глицерол-2-фосфата и глицерол-3-фосфата, так, что экспрессируется нуклеиновая кислота, и (b) выделение полипептида из клеток, причем во время культуральной стадии в культуральную среду добавляют неорганический фосфат. Представленное изобретение позволяет улучшить экспрессию гетерологичного белка и повысить выход конечного продукта. 28 з.п. ф-лы, 10 ил., 5 табл.
1. Способ получения полипептида, гетерологичного для Е. coli, включающий:
(a) культивирование клеток Е. coli, содержащих нуклеиновую кислоту, кодирующую полипептид, в культуральной среде, подпитывая культуральную среду транспортабельным органофосфатом, который выбран из группы, состоящей из альфа-глицерофосфата, бета-глицерофосфата, глицерол-3-фосфата, и смеси глицерол-2-фосфата и глицерол-3-фосфата, так что экспрессируется нуклеиновая кислота, и
(b) выделение полипептида из клеток,
причем во время культуральной стадии в культуральную среду добавляют неорганический фосфат.
2. Способ по п.1, где глицерофосфат представляет собой альфа-глицерофосфат, или бета-глицерофосфат, или их смесь.
3. Способ по п.2, где глицерофосфат представляет собой смесь глицерол-2-фосфата и глицерол-3-фосфата или представляет собой глицерол-3-фосфат.
4. Способ по любому из пп.1-3, где культивирование проводят во встряхиваемой колбе или ферментере.
5. Способ по любому из пп.1-3, где полипептид выделяют из цитоплазмы, периплазмы или культуральной среды клеток.
6. Способ по любому из пп.1-3, где экспрессию нуклеиновой кислоты регулируют индуцибельным промотором.
7. Способ по п.6, где индуцибельный промотор представляет собой промотор щелочной фосфатазы.
8. Способ по п.6, где индуцибельный промотор представляет собой промотор tac.
9. Способ по п.6, где индуцибельный промотор представляет собой промотор Т7.
10. Способ по п.6, где экспрессию нуклеиновой кислоты начинают в фазе активного роста на стадии культивирования.
11. Способ по п.1, где Е. coli является дефектной по хромосомальному phoA.
12. Способ по п.1, где Е. coli представляет собой дикий тип по отношению к хромосомальному glpT.
13. Способ по п.1, где Е. coli является дефектной по хромосомальному glpT.
14. Способ по п.1 или 13, где Е. coli является дефектной по хромосомальным phoA и glpT.
15. Способ по п.14, где Е. coli не является дефектной по хромосомальному ugp.
16. Способ по п.1, где полипептид представляет собой эукариотический полипептид.
17. Способ по п.1, где полипептид представляет собой полипептид млекопитающих.
18. Способ по п.1, где полипептид представляет собой инсулиноподобный фактор роста-1.
19. Способ по п.18, где скорость подачи органофосфата составляет приблизительно от 1 до 7 ммоль/ч на 8-10 л, и культивирование проводят в 10-литровом ферментере.
20. Способ по п.19, где скорость подачи органофосфата составляет от 2 до 6 ммоль/ч на 8-10 л.
21. Способ по п.20, где скорость подачи органофосфата составляет от 3 до 4 ммоль/ч на 8-10 л.
22. Способ по п.1, где полипептид представляет собой Apo2L.
23. Способ по п.22, где скорость подачи органофосфата составляет от 4 до 17 ммоль/ч на 8-10 л, и культивирование проводят в 10-литровом ферментере.
24. Способ по п.23, где скорость подачи составляет от 6 до 16 ммоль/ч на 8-10 л.
25. Способ по п.24, где скорость подачи составляет от 8 до 15 ммоль/ч на 8-10 л.
26. Способ по п.25, где скорость подачи составляет от 10 до 14 ммоль/ч на 8-10 л.
27. Способ по п.1, где соотношение неорганического фосфата к органофосфату находится в диапазоне от 1:10 до 1:0,25.
28. Способ по п.27, где полипептид представляет собой Apo2L, и соотношение составляет от 1:3 до 1:0,5.
29. Способ по п.28, где соотношение составляет 1:1.
| KIM DM et al., Regeneration of adenosine triphosphate from glycolytic intermediates for cell-free protein synthesis, Biotechnol Bioeng., 2001 Aug 20, v.74(4), p.309-316 | |||
| СПОСОБ ИЗГОТОВЛЕНИЯ БЕСКОНЕЧНЫХ АБРАЗИВНЫХ ШЛИФОВАЛЬНБГХ ЛЕНТ | 0 |
|
SU240497A1 |
| ПИТАТЕЛЬНАЯ СРЕДА ДЛЯ ВЫДЕЛЕНИЯ ВОЗБУДИТЕЛЯ ГЕМОРРАГИЧЕСКОГО КОЛИТА С ГЕМОЛИТИКО-УРЕМИЧЕСКИМ СИНДРОМОМ, ВЫЗЫВАЕМОГО КИШЕЧНОЙ ПАЛОЧКОЙ 0157:Н7 (СОРБИТОЛ E.COLI 0157:Н7АГАР) | 1997 |
|
RU2139343C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pSS5, КОДИРУЮЩАЯ СИНТЕЗ РЕКОМБИНАНТНОГО ЧЕЛОВЕЧЕСКОГО АЛЬФА-2b ИНТЕРФЕРОНА, ШТАММ ESCHERICHIA COLI SS5 - ПРОДУЦЕНТ РЕКОМБИНАНТНОГО ЧЕЛОВЕЧЕСКОГО АЛЬФА-2b ИНТЕРФЕРОНА И СПОСОБ ПОЛУЧЕНИЯ ИНТЕРФЕРОНА АЛЬФА-2b | 1999 |
|
RU2165455C1 |

