Настоящее изобретение относится к способу введения молекул пептидной нуклеиновой кислоты (ПНК), конъюгированных с положительно заряженными пептидами, в клетки, предпочтительно в цитозоль и/или ядро клеток, с использованием фотосенсибилизирующего агента и облучения клеток светом длиной волны, эффективной для активации фотосенсибилизирующего агента, и к использованию данного способа для оценки или изменения активности гена, например, антисмысловые или антигенные технологии, и для таких последующих применений, как в высокопроизводительной системе скрининга влияния продуктов понижающей регуляции генов.
ПНК представляют собой синтетические аналоги ДНК, в которых нормальная фосфодиэфирная связь, находящаяся в основной цепи ДНК, заменена на связь 2-аминоэтил-глицин. Нуклеотидные основания связаны с незаряженными повторяющимися единицами основной цепи посредством метилкарбонильных линкеров.
В результате такого связывания ПНК незаряжены. Они также химически стабильны и устойчивы к гидролитическому расщеплению и связываются с комплементарными цепями нуклеиновых кислот (ДНК или РНК) с более высокой аффинностью, чем природные нуклеиновые кислоты.
Хотя гибридизация ПНК к комплементарным ДНК и РНК сопровождается образованием водородной связи по Уотсону-Крику, можно получить как параллельные, так и антипараллельные двойные спирали. Кроме того, их гибридные комплексы проявляют превосходную термостабильность и показывают уникальные свойства ионной силы. Ввиду данных преимуществ и того факта, что ПНК устойчивы к действию нуклеаз и протеаз, ПНК использовали in vitro в антисмысловых (препятствующих трансляции мРНК) или антигенных (препятствующих репликации или транскрипции гена) применениях. Двойные спирали ПНК-РНК не являются субстратами для РНазы H и поэтому могут вызывать антисмысловые эффекты, основанные на стерическом блокировании либо трансляции РНК, либо процессинга. Тройные спирали являются результатом связывания ПНК и ДНК, которые могут препятствовать репликации или транскрипции, вызывая антигенные эффекты. Не наблюдали никаких признаков какой-либо общей токсичности ПНК.
Таким образом, связываясь молекулами-мишенями нуклеиновых кислот, ПНК обладают существенным влиянием на процессы репликации, транскрипции и трансляции. ПНК, используемая в антигенных или антисмысловых методиках, как показали, препятствует активностям ДНК- и РНК-полимераз, обратной транскриптазы, теломеразы и рибосомы.
Для таких воздействий, которые будут успешно опосредованы, необходимо, чтобы молекулы ПНК проникли в клетки и, для большинства методик, в ядро, которое содержит некоторые РНК и все ДНК, кроме митохондриальной ДНК. Клеточный и ядерный захват, кроме того, очень медленен и самопроизвольно не происходит. Улучшение клеточного и ядерного захвата ПНК поэтому представляет собой основную проблему, которую необходимо решить прежде, чем может появиться любая реальная перспектива развития ее в качестве терапевтического лекарственного средства или лечения или для ее широкого применения.
Один подход для доставки ПНК в клетку заключается в использовании микроинъекции (рассмотренный у Ray and Norden, (2000), FASEB J.14, 1041-1060). Микроинъекция, кроме того, является трудоемкой и требующей больших затрат времени. Кроме того, инъекция должна быть индивидуально проведена в каждую клетку, и это, следовательно, больше всего подходит для небольшого числа клеток и не подходит для множества методик in vivo. Повреждение клеток также является проблемой.
Доставку также осуществляли электропорацией (Shammas et al., (1999), Oncogene 18, 6191-6200), которая также обладает недостатками, например, она не подходит для использования in vivo.
Также проверяли такие способы, разрушающие мембраны, как временная проницаемость с использованием стрептолизина О (Faruqi et al. (1997), P.N.A.S. USA 95, 1398-1403), проницаемость мембраны клетки при помощи лизолектина (Boffa et al. (1996), J. Biol. Chem. 271, 13228-13233) или такие детергенты, как Tween (Norton et al. (1996), Nat. Biotech 14, 625-620). Данные способы также не подходят для использования in vivo и могут вызвать повреждение клеток.
Даже если можно с усилием внедрить молекулы ПНК в клетку, ядерный захват, возможно, не произойдет. ПНК с усилием внедряли для поглощения клетками при высоких концентрациях in vitro, однако для осуществления этого была необходима очень высокая концентрация (от 10 до 20 мкМ) (Folini et al. (2003), Cancer Research 63, 3490-3494). Может поэтому было замечено, что необходимы улучшенные способы введения ПНК в клетки.
Клеточная доставка ПНК in vitro, как также показали, происходит при введении с катионным липидом в виде комплекса. В данной определенной методике молекулы ПНК, связанные с функциональным пептидом, были гибридизированы с перекрывающимися олигонуклеотидами, и комплекс смешивали с катионным липидом. Тогда катионный комплекс липид-ДНК-ПНК интернализировался, неся ПНК в качестве пассивного груза (Hamilton et al. (1999), Chem. Biol. 6, 343-351).
В ПНК отсутствуют полианионные заряды, необходимые для конденсации и комплексообразования с катионными липосомами посредством электростатических взаимодействий. Гибриды ПНК-ДНК, кроме того, обладают распределенным отрицательным зарядом, который обеспечивается ДНК. Конденсированные частицы могут образоваться при взаимодействии гибридов ПНК-ДНК с катионными липидами, и такие липоплексы быстро встраиваются в клетки млекопитающих в культуре (Borgatti et al. (2003), Oncol. Res. 13(5), 279-287; Borgatti et al. (2002), Biochem. Pharmacol. 64(4), 609-616; Nastruzzi et al. (2000), J. Control Release 68(2), 237-249). ПНК можно также переносить в клетки посредством ковалентного связывания с липидами (Muratovska et al. (2001), Nucleic Acids Res. 29(9), 1852-1863; Ljungstrom et al. (1999), Bioconjug. Chem. 10(6), 965-972; Filipovska et al. (2004), FEBS Lett. 556 (1-3), 180-186).
Предпринимались также попытки получить конструкции пептид-ПНК, которые могут поглощаться клетками более эффективно. Branden и Smith (2002, Methods in Enzymology 346, 106-124) использовали так называемую систему Bioplex, при помощи которой ПНК использовали для связывания функционального пептида с ДНК для того, чтобы увеличить доставку ДНК. Можно также добавлять полиэтиленимин (PEI), чтобы улучшить конденсацию нуклеиновой кислоты.
Такая система предназначена для доставки нуклеиновой кислоты в клетку, и она использует преимущество ПНК в качестве средства для связывания ДНК, которую необходимо доставить к пептидам, которые разработаны для улучшения доставки ДНК.
Определенные пептиды, как известно, опосредуют доставку молекул через клеточную мембрану. Также пытались осуществить связывание ПНК с такими клеточными пептидами-транспортерами или проникающими внутрь клетки пептидами, чтобы попытаться улучшить способность ПНК проникать в клетку. Было разработано множество различных пептидов-транспортеров с целью транспортировки ПНК в клетку.
ПНК, разработанную в качестве средства анти-теломеразной активности, конъюгировали с пептидом для интернализации ВИЧ-tat (SEQ ID NO: 1 RKKRRQRRR) и с проникающим внутрь клетки пептидом Antennapedia (SEQ ID NO: 2 RQIKIWFQNRRMKWKK), и она, как было показано, обладала небольшим эффектом в качестве антисмысловой молекулы, уменьшающей теломеразную активность. Данные эксперименты, кроме того, показали только небольшое уменьшение теломеразной активности; конъюгированная с tat ПНК уменьшала теломеразную активность только до 73% от контрольного уровня через 48 часов, и конъюгированная с Antennapedia ПНК осуществляла 50% ингибирование только при очень высоких концентрациях >30 мкМ (Folini et al., 2003, см. выше).
Также были описаны пептиды, которые способны опосредовать транспорт ПНК в ядро. Для только что синтезируемых ядерных белков, как показали, необходимо наличие определенной аминокислотной последовательности для того, чтобы достичь ядра и пройти через ядерную мембрану. Такие сигналы ядерной локализации, в тех случаях, когда присутствуют в белках, не присутствующих эндогенно в ядре, могут также направлять такие белки в ядро.
ПНК также конъюгировали с сигналом ядерной локализации (NLS) (SEQ ID NO: 3 PKKKRKV) в попытке направить ПНК в ядро клетки. Такой NLS, как было показано, опосредует перенос большого T-антигена SV40 через ядерную мембрану. Когда вводили 10 мкМ ПНК-NLS в клетки, его присутствие в ядре было показано через 24 часа. Такой эффект, как было показано, не зависит от последовательности ПНК, но сильно зависит от последовательности NLS; рандомизированная последовательность NLS (SEQ ID NO: 4 KKVKPKR), конъюгированная с ПНК, демонстрировала только минимальные количества ПНК в ядре (Cutrona et al., (2000), Nature Biotechnology 18, 300-303). Данные результаты сравнивали в функциональном исследовании, где было показано, что ПНК-NLS (дикий тип) (где ПНК представляла собой антиген к myc) ингибировала рост клеток, тогда как ПНК, конъюгированная с рандомизированной последовательностью NLS, оказывала воздействия, заметно более подобные воздействиям контрольной ПНК.
Branden et al. (1999, Nature Biotechnology 17, 784-787) сходным образом показали, что хотя конъюгация ПНК с пептидами могла увеличивать ядерный транспорт ПНК зависимым от последовательности NLS образом, никакой ядерной локализации не наблюдали при инверсии последовательности NLS.
Дополнительные исследования также наводили на мысль, что для получения ПНК, которая будет успешно транспортироваться в ядро, необходимо конъюгировать молекулу ПНК как с клеточным мембранным пептидом-транспортером, так и с NLS (Braun et al. (2002), J. MoI. Biol. 318, 237-243). Клеточный мембранный пептид-транспортер, как полагают, импортирует ПНК, и NLS, как полагают, затем далее передает ПНК в ядро. В таких экспериментах NLS, как показали, существенен для ядерного транспорта, поскольку конструкции, содержащие проникающий внутрь клетки пептид с одной только пептидной последовательностью лизин-лизин, оставались в цитозоле.
Интерпретация упомянутых выше результатов осложняется тем фактом, что Richard et al. (J. Biol. Chem., (2003), 278(1), 585-590) продемонстрировали, что фиксация клеток, даже при мягких условиях фиксации, может вызывать артефакты в таких экспериментах, причем ядерное окрашивание замечали при отсутствии ПНК в ядре даже при мягких контролируемых условиях.
Таким образом, хотя показано, что при определенных условиях ПНК или ПНК, конъюгированная с проникающими внутрь клетки пептидами, может войти в клетку, или, как недавно показано, в эндосомы (Richard et al., 2003, выше), в большинстве случаев для биологического эффекта, опосредуемого ПНК, необходимо переместить ПНК в ядро.
Можно заметить, что остается необходимость в таком надежном и воспроизводимом способе введения молекул ПНК, что осуществляется захват в клетку, например цитозоль, предпочтительно в ядро, без необходимости использования высоких концентраций ПНК.
Авторы изобретения с удивлением обнаружили, что молекулы ПНК, которые конъюгированы с положительно заряженными пептидами, подвергаются эндоцитозу, и при высвобождении из эндосом с использованием методики фотохимической интернализации (PCI) данные молекулы транспортируются в ядро.
Таким образом, по первому аспекту изобретение обеспечивает способ введения молекулы ПНК в цитозоль, предпочтительно в ядро клетки, предусматривающий контактирование указанной клетки с молекулой ПНК и фотосенсибилизирующим агентом и облучение клеток светом длины волны, эффективной для активации фотосенсибилизирующего агента, в котором указанная молекула ПНК конъюгирована с положительно заряженным пептидом.
PCI представляет собой методику, которая использует фотосенсибилизирующий агент в комбинации со стадией облучения для активации такого агента и осуществляет интернализацию молекул, совместно вводимых в клетку. Такая методика позволяет молекулам, которые захватываются клеткой в такие органеллы, как эндосомы, высвобождаться из таких органелл в цитоплазму после облучения. Основной способ фотохимической интернализации (PCI) описан в WO 96/07432 и WO 00/54802, которые включены сюда в качестве ссылки. Как изложено выше, интернализуемая молекула (которая для использования по настоящему изобретению представляет собой конъюгат ПНК-пептид) и фотосенсибилизирующий агент приводятся в контакт с клеткой. Фотосенсибилизирующий агент и интернализуемая молекула захватываются клеточным мембраносвязанным субкомпартментом внутри клетки. При воздействии на клетку светом соответствующей длины волны активизируется фотосенсибилизирующий агент, который непосредственно или опосредованно вызывает образование токсичных продуктов, которые разрушают внутриклеточные мембраны компартментов. Это позволяет интернализуемой молекуле высвободиться в цитозоль.
Такие способы используют фотохимический эффект в качестве механизма для введения молекул, иным образом не проникающих через мембрану, в цитозоль клетки способом, который не приводит к общему разрушению клетки или гибели клетки, если методология соответственно приспособлена для того, чтобы избежать чрезмерного образования токсичных продуктов, например, уменьшением времени облучения или дозы фотосенсибилизатора.
Особенно удивительно то, что когда для высвобождения ПНК в клетку используется способ PCI, ни определенной проникающей внутрь клетки последовательности, ни последовательности NLS не требуется для входа ПНК в клетку и для ее последующего перемещения в ядро. Все, что необходимо, чтобы ПНК была связана с пептидом, который имеет, по меньшей мере, единичный суммарный положительный заряд.
Таким образом, не желая быть связанными с теорией, оказывается, что, при использовании PCI, наличие положительно заряженного пептида может способствовать захвату молекулы ПНК в клетку, в такие клеточные компартменты, как эндосомальные компартменты, и дополнительно, после высвобождения или интернализации молекулы ПНК в цитозоль, заряженный пептид затем дополнительно опосредует направленное перемещение молекулы ПНК в ядро. Как следствие этого, необходима только минимальная модификация молекулы ПНК для направленного перемещения ее к необходимому месту локализации, и не требуется конъюгации длинных аминокислотных последовательностей или множественных аминокислотных последовательностей с молекулой ПНК.
Также удивительно, что только один пептид необходим для выполнения обеих из этих функций, то есть осуществление направленного захвата молекулы ПНК в клетку, и также облегчение переноса в ядро, как только молекула ПНК-пептид высвобождается или интернализуется в цитозоль.
Сигналы ядерной локализации были изучены довольно подробно, и было показано, что определенные аминокислотные консенсусные последовательности необходимы для того, чтобы осуществлялся эффективный направленный ядерный перенос. В частности, метаболический путь импортина идентифицировали в качестве способа, при помощи которого молекулы могут проходить в ядро. Такие "классические" NLS с высоким содержанием аргинина/лизина, как последовательность большого антигена SV40T, взаимодействует с белками-импортинами α+β. Комплекс перемещается через центральный канал комплекса ядерной поры и диссоциирует в ядре. Стадии ассоциации и диссоциации являются энергозависимыми механизмами (рассмотренными у Cartier et al. (2002), Gene Therapy 9, 157-167). Существуют, как полагают, другие метаболические пути для ядерного импорта, хотя они не так хорошо охарактеризованы.
Поэтому удивительно, что при использовании способа PCI по изобретению не требуется не только классической последовательности NLS, но и более того, любая последовательность с суммарным положительным зарядом один или более способна опосредовать ядерную локализацию. Это демонстрируется тем фактом, что последовательность SEQ ID NO: 5 GHHHHHG, функционирующая также как SEQ ID NO: 3 PKKKRKV, и дополнительно, что трипептид только с единичным положительным зарядом (SEQ ID NO: 6 AKL) обладали способностью направлять ПНК сначала в эндосому и впоследствии в ядро (смотрите примеры).
Далее, удивительное наблюдение состоит в том, что последовательности, которые были первоначально идентифицированы по их способности направлять белки в такие клеточные органеллы, как пероксисомы и митохондрии, при конъюгации с молекулами ПНК, также способны направлять молекулы ПНК сначала в эндосому и впоследствии в ядро (смотрите примеры).
Точная роль PCI в способе по изобретению не известна, но она, определенно, является основной в осуществлении способа, поскольку без PCI молекулы ПНК, несущие положительно заряженный пептид, не проникают в цитозоль или ядро в какой-либо существенной степени.
Эффект также, по-видимому, не зависит от общей длины конъюгируемого пептида, причем положительно заряженные пептиды длиной 3 аминокислоты одинаково функционируют так же, как и пептиды длиной 29 аминокислот. Эффект также не зависит от соотношения заряда к длине положительно заряженного пептида и конкретных заряженных аминокислот, которые включены в состав последовательности.
Используемый здесь термин «ПНК» относится к молекуле пептидной нуклеиновой кислоты, которая действует в качестве аналога ДНК и основана на псевдопептидном скелете, к которому присоединены нуклеотидные основания. ПНК может находиться в свободной линейной форме или может быть в виде двойной спирали или самозамыкающемся виде, например, bis-ПНК.
Также рассматриваются производные стандартной формы ПНК, например, в которых один или несколько псевдопептидных мономеров, составляющих полимер, можно модифицировать или получить их производное, например, чтобы получить измененные свойства, например, используя лизин или другие аминокислотные аналоги. Точно так же, одно или несколько из используемых оснований можно модифицировать, если необходимо, например, используя варианты, не встречающиеся в природе. Таким образом ПНК включает производные стандартной формы, при условии, что такие производные сохраняют соответствующие функциональные свойства, то есть способны к образованию зависимого от последовательности комплекса с ДНК и/или РНК. Другими словами, производное ПНК соответствует в отношении заряда и структуры, учитывая комплементарность, последовательности ДНК или РНК.
Молекула ПНК может представлять собой любую последовательность или составлять любую длину. Предпочтительно молекула ПНК содержит в длину менее 25, например менее 20, оснований. Предпочтительно молекула ПНК содержит в длину более 6 оснований. Например, можно использовать молекулы от 6 до 20 оснований. Длина олигомера ПНК из 12-17 единиц оптимальна. Длина последовательности, прежде всего, определяется необходимой спецификой используемого способа. Методики ДНК, для которых необходимо наличие более 25 оснований, можно обычно выполнять с гораздо более короткими образцами ПНК. Длинные олигомеры ПНК, в зависимости от последовательности, склонны к агрегации и их трудно очистить и охарактеризовать. Кроме того, чем короче последовательность, тем более она специфична. Следовательно, влияние ошибки больше, чем для короткой последовательности. Кроме того, олигомеры ПНК в 20 единиц использовали без какой-либо проблемы, связанной с агрегацией.
Такие молекулы, их химические свойства и способы синтеза известны в данной области (Ray and Norden, 2000, выше), и их можно получить любыми подходящими способами.
Молекула ПНК может представлять собой антисмысловую молекулу ПНК или молекулу ПНК, комплементарную гену (антигенная молекула), которая может образовывать характерную тройную структуру. Молекула ПНК также может представлять собой зонд, то есть ее можно связывать с последовательностью-мишенью нуклеиновой кислоты, и она соответственно может нести метку.
Способ по изобретению осуществляет перенос конъюгата ПНК-пептид в цитозоль, предпочтительно в ядро. Кроме того, в полной мере будет понятно, что захват всех до одной молекул, контактирующих с клеткой, не достижим. Существенный и улучшенный захват относительно предшествующих уровней, при которых не используется никакой PCI, кроме того, достижим. Предпочтительно, способы по изобретению позволяют осуществлять захват молекул ПНК на таком достаточном уровне, что их влияние на репликацию, транскрипцию или трансляцию очевидно по экспрессированным продуктам таких клеток. Соответствующую концентрацию конъюгатов ПНК-пептид, которые будут контактировать с клеткой, можно подбирать, чтобы достичь этой цели, например, достичь снижения экспрессии гена-мишени более чем на 10%, например, более чем на 20, 30, 40 или 50% снижения после инкубации с клетками в течение, например, 24, 48, 72 или 96 часов, например, от 24 до 48 часов (смотрите, например, фиг.9). Уровень снижения количества белка зависит от времени полужизни белка, то есть ранее существовавший белок будет удаляться в соответствии с его временем полужизни. Таким образом, снижение экспрессии более чем на 10, 20, 30, 40 или 50% осуществляется относительно экспрессии за тот же самый момент времени без ПНК так, что время полужизни принимается во внимание.
Термин «клетка» используется здесь для того, чтобы охватить все эукариотические клетки (включая клетки насекомых и грибковые клетки). Типичные примеры «клеток» таким образом включают все типы клеток животных млекопитающих и немлекопитающих, клетки растений, клетки насекомых, грибковые клетки и простейшие. Кроме того, предпочтительно клетки являются клетками млекопитающих, например кошек, собак, лошадей, ослов, овец, свиней, коз, коров, мышей, крыс, кроликов, морских свинок, но наиболее предпочтительно людей.
Используемый здесь термин «контактирование» относится к приведению клеток и фотосенсибилизирующего агента и/или конъюгата ПНК-пептид в физический контакт друг с другом при условиях, подходящих для интернализации в клетки, например, предпочтительно при 37°C в подходящей питательной среде.
Фотосенсибилизирующий агент представляет собой агент, который активируется светом соответствующей длины волны и интенсивности, чтобы образовались активированные продукты. Обычно такой агент может представлять собой агент, который локализуется во внутриклеточных компартментах, особенно эндосомах или лизосомах. Ряд таких фотосенсибилизирующих агентов известен в данной области и описан в литературе, включающей WO 96/07432, который включен сюда в качестве ссылки. В этом отношении можно упоминуть ди- и тетрасульфонированный фталоцианин алюминия (например, AlPcS2a), сульфонированные тетрафенилпорфины (TPPSn), нильский голубой, хлориновые производные e6, уропорфирин I, филоэритрин, гематопорфирин и метиленовый синий, которые, как было показано, локализуются в эндосомах и лизосомах клеток в культуре. Это происходит в большинстве случаев вследствие захвата фотосенсибилизатора путем эндоцитоза. Таким образом, фотосенсибилизирующий агент предпочтительно представляет собой агент, который захватывается во внутренние компартменты лизосом или эндосом. Дополнительно соответствующие фотосенсибилизаторы для использования по изобретению описаны в WO 03/020309, который также включен сюда в качестве ссылки, а именно, сульфонированные мезо-тетрафенилхлорины, предпочтительно TPCS2a.
Кроме того, другие фотосенсибилизирующие агенты, которые локализуются в других внутриклеточных компартментах, например эндоплазматическом ретикулуме или аппарате Гольджи, также можно использовать. Также предположительно, что могут существовать механизмы, при действии которых воздействия фотохимической обработки приходятся на другие компоненты клетки (то есть компоненты, отличные от ограниченных мембраной компартментов). Таким образом, например, одна возможность может состоять в том, что фотохимическая обработка разрушает молекулы, важные для внутриклеточного транспорта или слияния везикул. Такие молекулы необязательно могут располагаться в ограниченных мембраной компартментах, но фотохимическое повреждение таких молекул может, кроме того, привести к фотохимической интернализации молекул-переносчиков, например, посредством механизма, при котором фотохимические воздействия на такие молекулы приводят к сниженному транспорту интернализуемой молекулы (то есть молекулы ПНК) в такие разрушающие пузырьки, как лизосомы, так, чтобы интернализуемая молекула могла высвобождаться в цитозоль до деградации. Примеры молекул, необязательно расположенных в ограниченных мембраной компартментах, представляют собой несколько такие молекул транспортной системы из микротрубочек, как динеин и компоненты динактина; и, например, rab5, rab7, фактор, чувствительный к N-этилмалеимиду (NSF), растворимый белок присоединения NSF (SNAP) и так далее.
Классы подходящих фотосенсибилизирующих агентов, которые можно упомянуть, таким образом, включают порфирины, фталоцианины, пурпурины, хлорины, нафталоцианины бензопорфиринов, катионные красители, тетрациклины и лизомотропные слабые основания или их производные (Berg et al., J. Photochemistry and Photobiology, 1997, 65, 403-409). Другие подходящие фотосенсибилизирующие агенты включают тексафирины, феофорбиды, порфицены, бактериохлорины, кетохлорины, производные гематопорфирина и их производные, эндогенные фотосенсибилизаторы, индуцируемые 5-аминолевулиновой кислотой и их производные, димеры или другие конъюгаты фотосенсибилизаторов.
Предпочтительные фотосенсибилизирующие агенты включают TPPS4, TPPS2a, AlPcS2a, TPCS2a и другие амфифильные фотосенсибилизаторы. Другие подходящие фотосенсибилизирующие агенты включают соединение 5-аминолевулиновой кислоты или сложные эфиры 5-аминолевулиновой кислоты или ее фармацевтически приемлемые соли.
Термин «облучение» клетки для активации фотосенсибилизирующего агента относится к применению света непосредственно или опосредованно, как описано далее. Таким образом, клетки можно освещать при помощи источника света, например, непосредственно (например, единичные клетки in vitro) или косвенно, например, in vivo, когда клетки находятся под поверхностью кожи или находятся в виде слоя клеток, не все из которых освещаются непосредственно, то есть без экранирования других клеток.
Термин «пептид», как определено здесь, включает любую молекулу, содержащую любое количество аминокислот, то есть одну или несколько аминокислот. Предпочтительно, кроме того, пептид представляет собой полимер последовательных аминокислот.
Предпочтительно положительно заряженный пептид состоит из от 3 (или 4, 5 или 6) до 30 аминокислот в длину, более предпочтительно от 3 (или 4, 5 или 6) до 25, от 3 (или 4, 5 или 6) до 20 или от 3 (или 4, 5 или 6) до 15 аминокислот в длину. В наиболее предпочтительном осуществлении пептид состоит из менее 10 аминокислот в длину, например 3, 4, 5 или 6.
Пептиды можно получить любыми удобными способами, например непосредственным химическим синтезом или посредством рекомбинантных способов, посредством экспрессии молекулы нуклеиновой кислоты соответствующей последовательности в клетке.
Положительно заряженная молекула способна к перемещению молекулы ПНК, с которой она конъюгирована, в клетку и затем в цитозоль и также, предпочтительно, в ядро.
Упоминаемая здесь фраза «положительно заряженный» обозначает, что общий, или суммарный, заряд пептида составляет +1 или более при физиологическом значении pH, то есть при pH 7,2. Аминокислоту считают +1, если преобладающий тип аминокислот при физиологическом значении pH положительно заряжен, в тех случаях, когда находится в составе пептида. Каждая такая аминокислота в пептиде вносит дополнительный положительный заряд при вычислении окончательного заряда пептида. Пептид может содержать один или несколько отрицательно заряженных аминокислотных остатов, а также нейтральные остатки, при условии, что суммарный заряд пептида (вычисленный сложением вместе заряда, вносимого каждой аминокислотой) является положительным. Молекула ПНК незаряжена и, таким образом, не вносит вклад в общий заряд молекулы. Кроме того, необходимо понимать, что такой заряд представляет собой заряд части пептида, которая является важной и которую оценивают при определении наличия положительно заряженного пептида.
Заряд пептида, следовательно, зависит от его аминокислотного состава. Некоторые аминокислоты заряжены при нормальном физиологическом значении pH. Положительно заряженные аминокислоты представляют собой лизин (K), аргинин (R) и гистидин (H) и, как полагают, составляют +1 по описанной выше шкале. Аспарагиновая кислота (D) и глутаминовая кислота (E) несут отрицательный заряд при большинстве физиологических значениях pH и рассматриваются как -1 по упомянутой выше шкале. Другие встречающиеся в природе аминокислоты, как полагают, не несут никакого заряда. Может присутствовать любое количество положительно заряженных или отрицательно заряженных аминокислот, при условии, что общий заряд пептида представляет собой +1 или более.
Используемые в пептидах аминокислоты для применения по изобретению необязательно должны представлять собой встречающиеся в природе аминокислоты. Одну из большего количества аминокислот в пептиде можно заменить на не встречающуюся в природе, например, дериватизированную аминокислоту. Такие аминокислоты подобным образом оценивают на основании их вклада в заряд пептида. Таким образом, подобно встречающимся в природе аминокислотам, если преобладающий тип аминокислот положительно заряжен при физиологическом значении pH, то несущественно, происходит ли или нет такой заряд из дериватизированной группы (например, введенной аминогруппы) или группы, также присутствующей во встречающейся в природе аминокислоте, при условии, что общий заряд составляет +1 или более.
Пептид может присутствовать в качестве участка гибридной молекулы, например, связанной с такой небелковой молекулой, как органический полимер, который можно, например, использовать в качестве сшивающей группы. Пептид может также присоединяться к отдельному компоненту, который может быть белковым по природе, но который эффективно не зависит от пептида, например, незаряжен или находится в отдельной структурной конфигурации. В таких случаях пептид представлял бы собой экспонированный, предпочтительно периферийный участок, и определяли бы заряд такого участка в качестве соответственного пептида.
Положительно заряженный пептид можно конъюгировать либо с N-концом, либо с C-концом молекулы ПНК, и может присоединяться с или без такой сшивающей группы, как 8-амино-3,6-диоксаноктановая кислота, 2-аминоэтокси-2-этоксиуксусная кислота (AEEA) или дисульфидные линкеры. Предпочтительно, кроме того, пептид конъюгируют непосредственно ковалентным связыванием. Особенно предпочтительно, в конъюгате не присутствует никаких других компонентов, кроме ПНК и пептида.
Предыдущие исследования показали, что только классические сигналы ядерной локализации транспортируют конъюгированные молекулы в ядро. Однако, как упоминалось выше, как это ни удивительно, показано, что способность к ядерной локализации пептида зависит только от заряда, а не от последовательности, когда осуществляют способ интернализации, используя PCI. Пептиды с суммарным зарядом +5 показали самую высокую степень поглощения, и это, как обнаружили, не зависело от последовательности, обеспечивающей такой заряд. Заряд пептида составляет >1, предпочтительно от +1 до +10, например, от +2 до +8, такой как от +3 до +6, например, +4 или +5.
Предпочтительно, пептиды для присоединения к ПНК богаты остатками K, R и/или H. Особенно предпочтительно используют ряды последовательных заряженных остатков. Предпочтительно, другие остатки, используемые в пептиде, нейтральны. Таким образом, например, пептид может обладать или содержать последовательность: Xn-(Y)m-Xo, в которой X представляет собой нейтральные остатки и Y представляет собой положительно заряженный остаток, который может быть одним и тем же или другим в каждом положении, в котором они появляются, и n, m и o представляют собой целые числа ≥1, например, в диапазоне от 1 до 10, и n и o предпочтительно представляют собой 1 или 2, и m предпочтительно представляет собой от 2 до 5. Особенно предпочтительно Y представляет собой один и тот же остаток в каждом положении и представляет собой K, R или H.
Особенно предпочтительные пептиды представляют собой SEQ ID NO: 7 MSVLTPLLLRGLTGSARRLPVPRAKIHSL, SEQ ID NO: 6 AKL и SEQ ID NO: 5 GHHHHHG. SEQ ID NO: 7 MSVLTPLLLRGLTGSARRLPVPRAKIHSL и SEQ ID NO: 6 AKL представляют собой последовательности для направленного переноса в митохондрии и пероксисомы соответственно, и все же они оказались способны к направленному переносу в ядро с использованием описанного здесь способа PCI. Такая неожиданная находка иллюстрирует зависимость от заряда, но не от последовательности пептидов, которые применимы по изобретению.
Положительно заряженный пептид предпочтительно не представляет собой такой NLS, как SEQ ID NO: 3 PKKKRKV или рандомизированный NLS SEQ ID NO: 4 KKVKPKR или обратный NLS SEQ ID NO: 8 VKRKKKP, или такой классический проникающий внутрь клетки пептид, как Tat ВИЧ SEQ ID NO:1 RKKRRQRRR или проникающий внутрь клетки пептид Antennapedia SEQ ID NO: 2 RQIKIWFQNRRMKWKK. Это можно оценить, например, определяя степень ядерного переноса или проникновения в клетку без PCI. Пептиды, способные к значительному ядерному переносу или проникновению в клетки, в таких обстоятельствах считали бы NLS или проникающими внутрь клетки пептидами. Пептид также предпочтительно не представляет собой полилизин. Дополнительно, ПНК или пептид могут содержать такие дополнительные модификации, как флуоресцентные метки на концах.
Конъюгаты ПНК-пептид, как описано здесь, составляют дополнительные аспекты изобретения.
Упоминаемый здесь термин «конъюгация» относится к связыванию вместе пептида и молекулы ПНК для образования единой структурной единицы при физиологических условиях. ПНК и пептид предпочтительно связаны ковалентной связью.
Молекулу ПНК и пептид можно синтезировать или очистить отдельно и затем объединить, например, используя такую молекулу-спейсер, как Fmoc-NC603H11-OH (Branden et al., 1999, см. выше), или их можно химически синтезировать в качестве единой молекулы, например, используя стратегию (Btoc). В данном способе мономеры ПНК синтезируются в олигомеры длиной 20 оснований, используя протоколы стандартного пептидного синтеза. Для мономеров ПНК используют защиту флуоренилметоксикарбонилом (Fmoc) N-концевой аминогруппы мономера и бензгидрилоксикарбонил (Bhoc) для защиты A, C и G экзоциклических аминогрупп. Группа Bhoc, связанная с XAL-меткой для синтеза, позволяет осуществить быстрое удаление защиты и отщепление олигомера ПНК от смолы. Типичные выходы связанного продукта составляют >95%. Синтез заканчивается отщеплением олигомера TFMSA от смолы. Олигомер очищают при помощи ВЭЖХ с обращенной фазой (Viirre et al. (2003), J. Org. Chem. 68(4), 1630-1632; Neuner et al. (2002), Bioconjug. Chem. 13 (3), 676-678).
Таким образом, положительно заряженный пептид, по-видимому, ответственен и за поглощение ПНК в клетку, и, как только она высвобождается из внутриклеточного компартмента, за ее ядерный захват.
Более одного типа молекулы ПНК, то есть молекулы ПНК различных последовательностей, можно вводить или вносить одновременно. Точно так же можно одновременно вводить или вносить молекулы ПНК, несущие более одного типа положительно заряженного пептида.
Необязательно, один или другой, или оба из фотосенсибилизирующего агента и конъюгированной молекулы ПНК, которые необходимо ввести в клетки, можно присоединить или связать, или конъюгировать с одной или несколькими молекулами носителя или с молекулами для направленного переноса, которые могут оказывать воздействие на обеспечение или увеличение захвата фотосенсибилизирующего агента или конъюгированной молекулы ПНК, или могут оказывать воздействие на направленный перенос или доставку данных объектов к определенному типу клеток, ткани или внутриклеточному компратменту. В случае конъюгированной молекулы ПНК направленный перенос в ядро можно достичь уже компонентом пептида, конъюгированного по изобретению.
Примеры систем носителей включают полилизин или другие поликатионы, сульфат декстрана, различные катионные липиды, липосомы, воссозданные LDL-частицы или стерически устойчивые липосомы. Такие системы носителя могут, в общем случае, улучшить фармакокинетику и увеличить клеточный захват конъюгированной молекулы ПНК и/или фотосенсибилизирующего агента и могут также направлять молекулу ПНК и/или фотосенсибилизирующий агент во внутриклеточные компартменты, которые особенно благоприятны для осуществления фотохимической интернализации, но они, в общем случае, не обладают способностью направлять молекулу ПНК и/или фотосенсибилизирующий агент к определенным клеткам (например, раковым клеткам) или ткани. Кроме того, чтобы достичь такого определенного или селективного направленного переноса, молекулу носителя, молекулу ПНК и/или фотосенсибилизатор можно связать или конъюгировать с определенными молекулами для направленного переноса, которые обеспечат определенный клеточный захват молекулы ПНК в необходимые клетки или ткани. Такие молекулы для направленного переноса могут также направлять молекулу во внутриклеточные компартменты, которые особенно благоприятны для осуществления фотохимической интернализации.
Можно применять множество различных молекул для направленного переноса, например, как описано у Curiel (1999), Ann. New York Acad. Sci. 886, 158-171; Bilbao et al., (1998), в Gene Therapy of Cancer (Walden et al., eds., Plenum Press, New York); Peng and Russell (1999), Curr. Opin. Biotechnol. 10, 454-457; Wickham (2000), Gene Ther. 7, 110-114.
Молекулу носителя и/или молекулу для направленного переноса можно ассоциировать, связать или конъюгировать с молекулой ПНК, фотосенсибилизирующим агентом или с обоими и можно использовать тот же самый или другой носитель или молекулы для направленного переноса. Как упомянуто выше, можно одновременно использовать более одного носителя и/или молекулы для направленного переноса.
Предпочтительный носитель для использования по настоящему изобретению включает такие поликатионы, как полилизин (например, поли-L-лизин или поли-D-лизин), полиэтиленимин или дендримеры (например, такие катионные дендримеры, как SuperFect®); такие катионные липиды, как DOTAP или липофектин, и пептиды.
Способ по изобретению можно осуществить, как описано ниже. В способе по изобретению интернализуемая молекула и фотосенсибилизирующее соединение применяют одновременно или последовательно в клетках, после чего фотосенсибилизирующее соединение и молекула подвергаются эндоцитозу или другими способами перемещаются в эндосомы, лизосомы или другие внутриклеточные ограниченные мембраной компартменты.
Конъюгат ПНК-пептид и фотосенсибилизирующее соединение можно применять к клеткам вместе или последовательно. Они могут поглощаться клеткой в одни и те же или различные внутриклеточные компартменты (например, они могут перемещаться совместно). Конъюгат ПНК-пептид затем высвобождается при воздействии на клетки светом подходящих длин волн для активации фотосенсибилизирующего соединения, который в свою очередь приводит к разрушению внутриклеточных мембран компартментов и последующему высвобождению молекулы, которая может располагаться в том же самом компартменте, что и фотосенсибилизирующий агент, в цитозоль. Таким образом, в таких способах заключительная стадия воздействия на клетки светом приводит к высвобождению интересующей молекулы из того же самого внутриклеточного компартмента, что и фотосенсибилизирующий агент, и она оказывается в цитозоле.
Совсем недавно в WO 02/44396 (который включен сюда в качестве ссылки) описан способ, в котором порядок стадий можно изменять так, что, например, фотосенсибилизирующий агент взаимодействует с клетками и активизируется облучением до того, как молекула, которая будет интернализоваться таким образом доставляться в клетку, взаимодействует с клетками. Данный адоптированный способ использует преимущество такого факта, что для интернализуемой молекулы нет необходимости присутствовать в том же самом клеточном субкомпартменте, что и фотосенсибилизирующий агент.
Таким образом, в предпочтительном осуществлении указанный фотосенсибилизирующий агент и указанную молекулу ПНК применяют на клетке вместе или последовательно. Как следствие, они могут поглощаться клеткой в один и тот же внутриклеточный компартмент, и затем можно осуществить указанное облучение.
В альтернативном осуществлении указанный способ можно осуществить посредством контактирования указанной клетки с фотосенсибилизирующим агентом, контактирования указанной клетки с молекулой ПНК, которую необходимо ввести, и облучения указанной клетки светом длины волны, эффективной для активации фотосенсибилизирующего агента, где указанное облучение осуществляют до клеточного захвата указанной молекулы ПНК во внутриклеточный компартмент, содержащий указанный фотосенсибилизирующий агент, предпочтительно до клеточного захвата указанной молекулы в любой внутриклеточный компартмент.
Указанное облучение можно осуществить после клеточного захвата молекулы во внутриклеточный компартмент, локализованы ли или нет указанная молекула ПНК и фотосенсибилизирующий агент в одних и тех же внутриклеточных компартментах во время воздействия светом. В одном предпочтительном осуществлении облучение, кроме того, осуществляют до клеточного захвата интернализуемой молекулы.
Используемый здесь термин «интернализация» обозначает цитозольную доставку молекул. В настоящем случае «интернализация» также включает стадию высвобождения молекул из внутриклеточных/мембраносвязанных компартментов в цитозоль клетки.
Используемые здесь термины «клеточный захват» или «перемещение» относятся к одной из стадий интернализации, на которой молекулы, внешние относительно мембраны клетки, поглощаются клеткой так, что их обнаруживают внутри расположенной наружу мембраны клетки, например, при эндоцитозе или других соответствующих механизмах захвата, например, во внутриклеточном ограниченном мембраной компартменте или связанных с ним, например, эндоплазматическим ретикулумом, аппаратом Гольджи, лизосомами, эндосомами и так далее.
Стадию контактирования клеток с фотосенсибилизирующим агентом и с конъюгатом ПНК-пептид можно осуществить любым подходящим или необходимым способом. Таким образом, если стадию взаимодействия необходимо провести in vitro, клетки можно подходящим образом поддерживать в такой водной среде, как, например, соответствующая среда для культивирования клеток, и в соответствующий момент времени фотосенсибилизирующий агент и/или конъюгат ПНК-пептид можно просто добавить к среде при соответствующих условиях, например, при соответствующей концентрации и в течение соответствующего периода времени.
Фотосенсибилизирующий агент приводят в контакт с клетками при соответствующей концентрации и в течение соответствующего периода времени, который легко может определить специалист в данной области, использующий обычные способы, и будет зависеть от таких факторов, как определенный используемый фотосенсибилизирующий агент и тип клетки-мишени и расположение. Концентрация фотосенсибилизирующего агент должна быть такой, что при поглощении в клетку, например, в один или несколько из ее внутриклеточных компартментов или связанный с одним или несколькими из ее внутриклеточных компартментов и при активации облучением, одна или несколько клеточных структур разрушаются, например, один или несколько внутриклеточных компартментов лизируются или разрушаются. Например, фотосенсибилизирующие агенты, используемые здесь в примерах, можно использовать при концентрации, например, от 10 до 50 мкг/мл. Для использования in vitro диапазон может быть гораздо шире, например, 0,05-500 мкг/мл. Для лечения человека in vivo фотосенсибилизирующий агент можно использовать в диапазоне 0,05-20 мг/кг массы тела при системном введении или 0,1-20% в растворителе для местного введения. Для меньших животных диапазон концентрации может отличаться и может подбираться соответственно.
Время инкубации клеток с фотосенсибилизирующим агентом (то есть время «контактирования») можно изменять от нескольких минут до нескольких часов, например, даже до 48 часов или более, например, от 12 до 20 часов. Время инкубации должно быть таким, чтобы фотосенсибилизирующий агент захватился соответствующими клетками, например, во внутриклеточные компартменты в указанных клетках.
Инкубация клеток с фотосенсибилизирующим агентом может необязательно сопровождаться периодом инкубации с не содержащей фотосенсибилизатор средой до того, как клетки подвергнут воздействию света или как добавят молекулу ПНК, например, в течение от 10 минут до 8 часов, особенно от 1 до 4 часов.
Молекулу ПНК приводят в контакт с клетками при соответствующей концентрации и в течение соответствующего периода времени.
Определение подходящих доз молекул ПНК для использования в способах по настоящему изобретению является обычной практикой для специалиста в данной области. Для применений in vitro примерная доза молекул ПНК составляла бы приблизительно 0,1-500 мкг ПНК в мл и для применений in vivo приблизительно 10-6-1 г ПНК на инъекцию для человека. Например, конъюгат ПНК-пептид можно вводить при уровнях менее 50 мкМ, например, менее 30 мкМ, особенно предпочтительно, менее 10 мкМ, например, от 0,1 до l мкМ, или от 5 до 30 мкМ, где обозначенная концентрация отражает уровни при контактировании с клеткой.
Как упоминалось выше, было обнаружено, что контактирование может начаться даже спустя несколько часов после того, как добавляли фотосенсибилизирующий агент и происходило облучение.
Подходящую концентрацию можно определить в зависимости от эффективности захвата интересующей молекулы ПНК в интересующие клетки и конечной концентрации, которой необходимо достичь в клетках. Таким образом, «время трансфекции» или «время клеточного захвата», то есть время, в течение которого молекулы контактируют с клетками, может составлять от нескольких минут до нескольких часов, например, можно использовать время трансфекции от 10 минут до 24 часов, например, от 30 минут до 10 часов или, например, от 30 минут до 2 часов или 6 часов. Можно также использовать более длительное время инкубации, например, от 24 до 96 часов или более, например, 5-10 дней.
Увеличенное время трансфекции обычно приводит к повышенному захвату интересующей молекулы. Кроме того, более короткое время инкубации, например, от 30 минут до 1 часа, по-видимому, приводит к улучшенной специфичности захвата молекулы. Таким образом, при выборе времени трансфекции для любого способа соответствующий баланс необходимо соблюдать между получением достаточной степени захвата молекулы, сохраняя достаточную специфичность обработки PCI.
In vivo соответствующий способ и время инкубации, за которое молекула ПНК и фотосенсибилизирующие агенты приходят в контакт с клетками-мишенями, будут зависеть от таких факторов, как способ введения и типы молекулы ПНК и фотосенсибилизирующих агентов. Например, если молекула ПНК вводится в опухоль, ткань или орган, который необходимо лечить, то клетки около места инъекции проконтактируют с молекулой ПНК и, следовательно, будут иметь тенденцию захватить молекулу ПНК более быстро, чем клетки, расположенные на бóльших расстояниях от места инъекции, которые, вероятно, проконтактируют с молекулой ПНК за более длительный период времени и при более низких концентрациях.
Дополнительно, для молекулы ПНК, вводимой внутривенной инъекцией, может понадобиться некоторое время для достижения клетки-мишени, и это может потребовать более длительное пост-введение, например, несколько дней, для того, чтобы достаточное или оптимальное количество молекулы ПНК накопилось в клетке-мишени или ткани. Те же самые рассуждения, конечно, относятся ко времени ведения, необходимому для захвата фотосенсибилизирующего агента в клетки. Время введения, необходимое для отдельных клеток in vivo, таким образом, вероятно, изменится в зависимости от этих и других параметров.
Тем не менее, хотя ситуация in vivo более сложная, чем in vitro, основная концепция настоящего изобретения все же та же самая, то есть время, за которое молекулы приходят в контакт с клетками-мишенями, должно быть таким, что прежде, чем произойдет облучение, соответствующее количество фотосенсибилизирующего агента поглощается клетками-мишенями и также: (i) до или во время облучения молекула ПНК либо была поглощена, либо будет поглощена после достаточного контактирования с клеткой-мишенью в тот же самый или другой внутриклеточный компартмент; или (ii) после облучения молекула ПНК контактирует с клетками в течение периода времени, достаточного для осуществления ее захвата в клетки. В том случае, если молекула ПНК поглощается во внутриклеточные компартменты, подвергающиеся воздействию при активации фотосенсибилизирующего агента (например, компартменты, в которых присутствует агент), молекула ПНК может поглощаться до или после облучения.
Стадия облучения светом для активации фотосенсибилизирующего агента может происходить согласно способам и процедурам, хорошо известным в данной области. Например, длина волны и интенсивность света может быть выбрана в соответствии с используемым фотосенсибилизирующим агентом. В данной области известны подходящие источники света.
Время, в течение которого клетки подвергаются воздействию света, в способах по настоящему изобретению, можно изменять. Эффективность интернализации молекулы ПНК в цитозоль увеличивается при увеличенном воздействии света до максимума, после которого возрастает повреждение клетки и, следовательно, гибель клетки.
Предпочтительный период времени для стадии облучения зависит от таких факторов, как мишень, фотосенсибилизатор, количество фотосенсибилизатора, накопленного в клетках-мишенях или тканях, и перекрывание между спектром поглощения фотосенсибилизатора и спектром испускания источника света. В общем случае период времени для стадии облучения составляет порядка от минут до нескольких часов, например, предпочтительно, до 60 минут, например, от 0,5 или 1 до 30 минут, например, от 0,5 до 3 минут или от 1 до 5 минут, или от 1 до 10 минут, например, от 3 до 7 минут и, предпочтительно, приблизительно 3 минуты, например, от 2,5 до 3,5 минут.
Соответствующие дозы света может выбрать специалист, и они снова будут зависеть от фотосенсибилизатора и количества фотосенсибилизатора, накопленного в клетках-мишенях или тканях. Например, дозы света, обычно используемые для фотодинамического лечения раковых образований фотосенсибилизатором фотофрином и 5-аминолевулиновой кислотой предшественника протопорфирина, находятся в диапазоне 50-150 Дж/см2 при диапазоне плотности энергии менее 200 мВт/см2, чтобы избежать перегревания. Дозы света обычно ниже, если используют фотосенсибилизатор с более высокими коэффициентами экстинкции в красной области видимого спектра. Кроме того, для лечения нераковых тканей с меньшим количеством накопленного фотосенсибилизатора общее количество необходимого света может быть существенно выше, чем для лечения раковых образований. Более того, если необходимо поддержать жизнеспособность клетки, нужно избегать образования чрезмерных уровней токсичных продуктов и можно соответственно подбирать подходящие параметры.
Способы по изобретению могут неизбежно вызывать гибель некоторых клеток при фотохимической обработке, то есть посредством образования токсичных продуктов при активации фотосенсибилизирующего агента. В зависимости от предполагаемого использования такая гибель клетки может не иметь последствий и может, на самом деле, оказаться выгодной для некоторых применений (например, лечение рака). Предпочтительно, кроме того, гибели клетки избегают. Способы по изобретению можно изменять так, что фракция или соотношение выживающих клеток регулируют, выбирая дозу света относительно концентрации фоточувствительного агента. Опять же такие способы известны в данной области.
В применениях, в которых требуются жизнеспособные клетки, по существу все клетки не погибают, или по существу не погибает большинство клеток (например, по меньшей мере, 50%, более предпочтительно, по меньшей мере, 60, 70, 80 или 90% клеток).
Независимо от степени гибели клеток, вызванной активацией фотосенсибилизатора, для воздействия ПНК на клетки важно, чтобы дозу света регулировали так, что некоторые из отдельных клеток, в которых проявляется эффект PCI, не погибали при одной только фотохимической обработке (хотя они могут впоследствии погибнуть от молекул, введенных в клетки, если такие молекулы обладают цитотоксическим действием).
Цитотоксических эффектов можно достичь при использовании, например, генотерапии, при которой антисмысловая молекула ПНК интернализуется в ядро опухолевых клеток способом по изобретению, например, для понижающей регуляции гена.
Способы по изобретению можно использовать in vitro или in vivo, например, либо для обработки in situ, либо для обработки ex vivo, сопровождающейся введением обработанных клеток в организм для различных целей, включающих: (i) ингибирование экспрессии определенных генных продуктов, связывая мРНК или промежуточные продукты сплайсинга; (ii) влияние на транскрипцию определенных генов, непосредственно влияя на ген (например, ингибируя факторы транскрипции); (iii) использование в качестве зондов для in situ гибридизации; (iv) использование при исследованиях-скрининге, и (v) для осуществления сайт-направленного мутагенеза или репарации дефектных генов в клетке-мишени.
Таким образом, в настоящем изобретении предлагается способ ингибирования транскрипции или экспрессии гена-мишени введением молекулы ПНК в клетку, содержащую указанный ген-мишень способом, описанным выше, где указанная молекула ПНК специфически связывается с указанным геном-мишенью или продуктом его репликации или транскрипции. Таким образом, например, указанную ПНК можно связывать с ДНК и/или РНК.
Термин «специфическое связывание» обозначает зависимое от последовательности связывание ПНК с молекулой-мишенью, которая может представлять собой РНК или ДНК. Термин «ген-мишень» означает ген или его фрагмент, с которым способна связываться ПНК и который является мишенью исследования.
В настоящем изобретении также предлагается способ идентификации или анализа уровня гена-мишени или продукта его репликации или транскрипции, где указанный способ предусматривает введение молекулы ПНК в клетку, содержащую указанный ген-мишень или продукт его репликации или транскрипции, способом, описанным выше, где указанная молекула ПНК специфически связывается с указанным геном-мишенью или продуктом его репликации или транскрипции, и анализ уровней связанной ПНК, чтобы определять наличие или уровень указанного гена-мишени или продукта его репликации или транскрипции. Удобным образом для данного способа молекула ПНК может нести молекулу с репортерной группой, которую можно идентифицировать при анализе, например, радиоактивную метку или средства генерации сигнала. Анализ может быть качественным и/или количественным.
В настоящем изобретении также предлагается способ проведения сайт-направленного мутагенеза или репарации гена-мишени, предпочтительно, дефектного гена в клетке, где указанный способ предусматривает введение молекулы ПНК и молекулы олигонуклеотида, содержащей необходимую последовательность, в клетку, содержащую указанный ген-мишень, способом, описанным выше, где указанная молекула ПНК специфически связывается с указанным геном-мишенью, чтобы сформировать крепление ПНК. Такая деформация нормальной нуклеиновой кислоты, формирующая тройную спираль, происходит на участке-мишени и обеспечивает восстановление или рекомбинацию на данном определенном участке. Донорный нуклеотид, который можно связать с ПНК или просто ввести совместно с ПНК, содержит необходимую нуклеотидную последовательность. ПНК, следовательно, действует в качестве промотора репарации/рекомбинации.
Данные способы можно использовать для исследовательских, например, диагностических целей, или для изменения профиля экспрессии клеток, например, для получения желаемого продукта для выделения, или в терапевтических целях.
Способы по изобретению можно таким образом использовать в диагностических целях, где наличие определенного гена или продукта его репликации или транскрипции информативно ввиду наличия, стадии или прогноза развития заболевания, состояния или нарушения. Таким образом, в настоящем изобретении дополнительно предлагается способ диагностики заболеваний, состояний или нарушений, предусматривающих введение молекулы ПНК в клетку (которое можно осуществлять in vitro, in vivo или ex vivo) способом, описанным выше, где указанная молекула ПНК специфически связывается с геном-мишенью или продуктом его репликации или транскрипции, который свидетельствует о наличии указанного заболевания, состояния или нарушения, и анализ уровня связанной ПНК, чтобы определить наличие, стадию или прогноз развития указанного заболевания, состояния или нарушения.
Способы по изобретению можно также использовать при лечении любого заболевания, при которых благоприятна понижающая регуляция, репарация или мутация одного или нескольких генов. Например, для генов, которые высоко экспрессируются при раке, можно осуществить понижающую регуляцию введением соответствующей ПНК-молекулы.
ПНК, ингибирующую экспрессию мутантного, вызывающего заболевание гена, можно было бы также вводить в комбинации с заменяющим геном (то есть, при генотерапии, которая включает терапевтическую передачу генов или модификацию существующих генов в клетках пациента), например, при лечении кистозного фиброза, рака, сердечно-сосудистых заболеваний, вирусной инфекции и диабета. Другие заболевания, при лечении которых имела бы пользу понижающая регуляция одного или нескольких генов, включают лейкемию и панкреатическую карциному (Cogoi et al. (2003) Nucleosides Nucleotides Nucleic Acids 22(5-8), 1615-1618), амиотрофический латеральный склероз (AMS) (Turner et al. (2003), Neurochem. 87(3), 752-763), болезнь Хантингтона (Lee et al. (2002) J. Nucl. Med. 43(7), 948-956) и болезнь Альцгеймера (McMahon et al. (2002) J. MoI. Neurosci. 19(1-2), 71-76).
Как описано выше, ПНК можно также использовать для изменения существующего гена, таким образом, можно использовать для репарации дефектного гена, например, для лечения заболевания, которое причинно связано с экспрессией данного дефектного гена или недостаточной экспрессией нормальной формы данного гена (Rogers et al. (2002), PNAS U.S.A. 99(26), 16695-16700; Faruqi et al. (1998), PNAS U.S.A. (5(4), 1398-1403).
Таким образом, в дальнейшем аспекте изобретения предлагается композиция, содержащая молекулу ПНК и также необязательно отдельно фотосенсибилизирующий агент, как описано здесь, где указанная молекула ПНК конъюгирована с положительно заряженным пептидом. В дальнейшем аспекте изобретения предлагается указанная композиция для использования в терапии.
Альтернативно описано, что в настоящем изобретении предлагается использование молекулы ПНК, как описано здесь, в препаратах лекарственного средства для лечения или профилактики заболевания, нарушения или инфекции путем изменения экспрессии одного или нескольких генов-мишеней у указанного пациента. Предпочтительно указанное лекарственное средство используют для генотерапии, то есть для лечения заболевания или нарушения, для которого типична аномальная экспрессия гена. Указанное изменение может включать понижающую регуляцию указанной экспрессии или повышающую регуляцию модифицированного вида указанного гена.
Согласно различным осуществлениям, изложенным выше, указанный фотосенсибилизирующий агент и указанная молекула ПНК контактируют с клетками или тканями пациента одновременно или последовательно, и указанные клетки облучают светом длины волны, эффективной для активации фотосенсибилизирующего агента, и облучение осуществляют до, во время или после клеточного захвата указанной молекулы ПНК во внутриклеточный компартмент, содержащий указанный фотосенсибилизирующий агент, предпочтительно, до клеточного захвата указанной молекулы во внутриклеточный компартмент. Таким образом, в альтернативном аспекте изобретения предлагается способ лечения или профилактики развития заболевания, нарушения или инфекции у пациента, предусматривающий введение молекулы ПНК в одну или несколько клеток in vitro, in vivo или ex vivo согласно способам, описанным выше, и где необходимо (то есть, когда перенос осуществляется in vitro или ex vivo) введение указанных клеток указанному пациенту, где указанная молекула ПНК конъюгирована с положительно заряженным пептидом.
Как определено здесь, термин «лечение» относится к сокращению, облегчению или устранению одного или нескольких симптомов заболевания, нарушения или инфекции, которое необходимо вылечить, относительно симптомов до лечения.
Термин «профилактика» относится к задержке или предотвращению возникновения симптомов заболевания, нарушения или инфекции.
Композиции по настоящему изобретению могут также содержать клетку, содержащую молекулу ПНК, которая интернализовалась в цитозоль или ядро указанной клетки способом по изобретению, где указанная молекула ПНК конъюгирована с положительно заряженным пептидом. Изобретение дополнительно относится к таким композициям для использования в терапии, особенно при лечении рака или генотерапии.
Таким образом, в еще одном дополнительном аспекте изобретения предлагается клетка или популяция клеток, содержащих молекулу ПНК, которая интернализовалась в цитозоль или ядро указанной клетки, в котором клетку можно получить способом по настоящему изобретению, где указанная молекула ПНК конъюгирована с положительно заряженным пептидом.
В еще одном дополнительном аспекте изобретения предлагается использование такой клетки или популяции клеток для приготовления композиции или лекарственного средства для использования при терапии, как описано выше, предпочтительно терапии рака или генотерапии, где указанная молекула ПНК конъюгирована с положительно заряженным пептидом.
В изобретении дополнительно предлагается способ лечения пациента, предусматривающий введение указанному пациенту клеток или композиций по настоящему изобретению, то есть способ, предусматривающий стадии введения молекулы в клетку, как описано выше, и введение таким образом полученной указанной клетки указанному пациенту. Предпочтительно указанные способы используют для лечения рака или в генотерапии.
Можно использовать любой способ введения in vivo, обычный или стандартный в данной области, например, инъекцию, вливание, местное введение и к внутренним, и к внешним поверхностям организма и так далее. Для применения in vivo изобретение можно использовать в отношении любой ткани, которая содержит клетки, в которых локализованы фотосенсибилизирующий агент и молекула ПНК, включая локализацию в жидких средах организма так же, как и в твердых тканях. Все ткани можно обрабатывать при условии, что фотосенсибилизатор захватывается клетками-мишенями, и свет может доставляться должным образом.
Таким образом, композиции по изобретению можно приготавливать любым удобным образом в соответствии с методиками и способами, известными в фармацевтической области, например, используя один или несколько фармацевтически приемлемых носителей или наполнителей. Упоминаемый здесь термин «фармацевтически приемлемый» относится к ингредиентам, которые совместимы с другими ингредиентами композиций так же, как и физиологически приемлемы для реципиента. Природу композиции и носителей или веществ-наполнителей, дозировки и так далее можно выбрать обычным образом в соответствии с выбором и необходимым способом введения, целью лечения и так далее. Дозировки аналогично можно определять обычным образом, и они могут зависеть от природы молекулы, цели лечения, возраста пациента, способа введения и так далее. Применительно к фотосенсибилизирующему агенту потенциальную возможность/способность разрушать мембраны при облучении следует также принять во внимание.
Способы, описанные выше, можно альтернативно использовать для создания средства для скрининга для высокопроизводительных способов скрининга, особенно для анализа эффектов подавления определенных генов. ПНК, направленную на один или несколько конкретных генов, можно получить и использовать ее по способу изобретения, как описано выше. ПНК можно таким образом использовать для снижения экспрессии гена в популяции клеток. Полученную в результате клеточную популяцию можно затем использовать в качестве средства для скрининга, чтобы идентифицировать эффекты подавления генов в прямом направлении по цепи последовательности при помощи стандартных способов. Таким образом можно идентифицировать, например, гены, на которые также влияет подавление гена-мишени. Необязательно к таким идентифицированным генам можно направить ПНК на дальнейших стадиях скрининга, чтобы, например, определить молекулы, вовлеченные в определенное событие передачи сигнала.
Таким образом, изобретение также относится к способу скрининга клеток с измененным паттерном экспрессии генов, предусматривающему: a) анализ экспрессии гена-мишени или одного или нескольких дополнительных генов клетки или популяции клеток, которые получали введением молекулы ПНК согласно способу по изобретению, где указанная ПНК специфически связывается с указанным геном-мишенью или продуктом его репликации или транскрипции и изменяет экспрессию указанного одного или нескольких дополнительных генов; и b) сравнение экспрессии указанной мишени и/или одного или нескольких дополнительных генов с экспрессией указанных генов в клетках сравнения, предпочтительно клетках дикого типа.
Паттерн экспрессии можно определять, используя любую подходящую методику, известную в данной области, например, используя микрочипы, несущие зонды, которые связываются с молекулами мРНК (или кДНК) и которые можно использовать для оценки количества каждого транскрипта. Клетки сравнения (контрольные клетки) обозначают любые клетки, с которыми сравнивается экспрессия. Предпочтительно, такие клетки представляют собой контрольные клетки, к которым не вводили ПНК. Особенно предпочтительно, указанные клетки представляют собой контрольные клетки дикого типа, например клетки, которые не подвергались такой генетической манипуляции, как при использовании ПНК.
Предшествующие попытки уменьшить экспрессию генов при помощи нормального и химически модифицированных антисмысловых олигонуклеотидов были ограничены проблемами нуклеазной деградации антисмысловых олигонуклеотидов, возникновения неспецифичных эффектов и/или недостаточной аффиностью к мишени. При использовании способа по изобретению для введения ПНК можно преодолеть такие проблемы.
Таким образом, в дальнейшем аспекте изобретение обеспечивает способ изменения паттерна экспрессии генов в клетке (например, популяции клеток), чтобы подготовить клетку (или популяцию клеток) для использования в качестве средства для скрининга (например, для высокопроизводительного скрининга), предусматривающий контактирование молекулы ПНК, способной ингибировать или уменьшать экспрессию гена, и фотосенсибилизирующего агента с клеткой (например, популяцией клеток) и освещение клетки (например, популяции клеток) светом длины волны, эффективной для активации фотосенсибслизирующего агента, где указанная молекула ПНК конъюгирована с положительно заряженным пептидом. Изобретение дополнительно относится к таким клеткам и способу скрининга таких клеток, где определенные свойства таких клеток, например, уровни эксперссии мРНК таких клеток, исследованы, например, на микрочипах.
Выражение «измененный паттерн экспрессии гена» означает, что как следствие наличия указанной молекулы ПНК в ядре клетки оказывается влияние на транскрипцию или трансляцию гена, к которому она направлена.
Как следствие такого изменения в экспрессии гена можно влиять на экспрессию других генов. Таким образом, влияя на нормальную экспрессию изучаемого гена, можно определить изменения в паттерне экспрессии других генов. Идентификация таких генов и влияние, которое оказывает на них экспрессия изучаемого гена, позволяет исследователю делать выводы о функциях гена, например, об их функции в прямом направлении по последовательности. Гены, на которые влияет изменение нормальной экспрессии изучаемого гена, можно подвергнуть повышающей регуляции или понижающей регуляции, но общее изменение в паттерне экспрессии указывает на роль гена в нормальной функции клетки и последствий нарушения его регуляции.
Используя стандартные способы, хорошо известные в данной области, можно изучать эффект пониженной регуляции или исчезновения экспрессии интересующего гена. Это можно, например, осуществлять посредством поиска таких функциональных изменений в клетках (или популяции клеток), как изменения клеточной адгезии, выделение белка или морфологические изменения. Альтернативно, профиль экспрессии генов можно непосредственно изучать, анализируя паттерны мРНК и/или экспрессии белка, снова используя стандартные способы, которые хорошо известны в данной области.
Ингибируя или уменьшая экспрессию гена, следует понимать, что экспрессия интересующего гена уменьшается при сравнении с клеткой, которую не подвергали воздействию по способу, то есть клеткой дикого типа или нормальной клеткой. Изменение уровня экспрессии гена можно определять стандартными способами, известными в данной области.
Возможно такое полное ингибирование экспрессии, что не существует никакой обнаруживаемой экспрессии гена, то есть не обнаруживаются мРНК или белок, или возможно частичное ингибирование экспрессии, то есть уменьшение, посредством чего количество экспрессии гена ниже, чем в клетке дикого типа или нормальной клетке. Это можно оценить и регулировать посредством сравнения эффекта ПНК с определенной последовательностью с эффектом ПНК с рандомизированной последовательностью, то есть с тем же самым составом нуклеотидов, но в разном порядке следования. Предпочтительно для данной применимой методики снижение экспрессии до менее 80% от контрольных уровней, например, <50%, предпочтительно <20, 10 или 5% от контрольных уровней. Используемая клетка (клетки) предпочтительно представляет собой популяцию клеток, отдельные клетки которой генетически идентичны. Клетки могут представлять собой любые клетки, как обсуждалось выше.
До развития данной новой методики доставки ПНК невозможно было использовать ПНК для такой системы. Способность использовать ПНК в данной системе имеет несколько преимуществ. Такие известные способы введения молекул в клетку, как использование агентов трансфекции, часто привносят отклонения в клеточные исследования, используемые в крупномасштабных системах скрининга так, что с трудом можно установить, какие эффекты вызваны нарушением экспрессии гена, а какие вызваны самой методикой трансфекции. Опосредованная PCI доставка обладает немногими такими эффектами, и это также возможно при использовании соответствующих контролей.
Некоторые из других веществ, используемых для доставки молекул в клетку, могут также вызывать неспецифические эффекты на исследование при скрининге. Например, короткая препятствующая РНК (siPHK), которую используют для методик подавления гена, как сообщали, влияла на экспрессию генов интерферонов (Sledz et al. (2003), Natl. Cell Biol. 5(9), 834-839). Стабильность ПНК высока, и поскольку такие воздействия, которыми она обладает, на экспрессию гена продолжительны даже после однократного введения.
Эффективность ПНК не зависит от систем определенных ферментов, поскольку ее ингибирующее действие зависит от химических взаимодействий с нуклеотидными молекулами. Также степень ингибрования постоянна в различных типах клеток. Дело обстоит не так для siPHK, например, которая зависит от определенных ферментов.
Было обнаружено, что, как это не удивительно, методика PCI не имеет ожидаемых проблем появления неспецифичных воздействий на экспрессию гена.
Клетку или популяцию клеток, полученную согласно способам изобретения, можно использовать для создания библиотеки, которая составляет дополнительный аспект изобретения.
Теперь изобретение будет описано более подробно при помощи следующих ниже неограничивающих примеров со ссылками на следующие чертежи, на которых:
На фиг.1 показано влияние заряда на интернализацию ПНК, используя анализ при помощи проточной цитометрии FITC-ПНК захвата в клетки OHS, HeLa и FEMXIII. Клетки инкубировали с различными FITC-ПНК в концентрации 1000 нМ в течение 24 часов при 37°C и анализировали при помощи проточной цитометрии, как описано в экспериментальных протоколах. «-1» ПНК383, «0» ПНК385, «+1» ПНК384, «+5» ПНК381. Результаты показаны в качестве среднего значения интенсивности флуоресценции относительно суммарного заряда различных молекул ПНК. Столбцы показывают три отдельных эксперимента, с 6 параллелями каждый. Планки погрешностей показывают стандартное отклонение от среднего значения.
На фиг.2 показана релокализация FITC-ПНК-NLS из внутриклеточных везикул в ядро при использовании обработки PCI, используя ПНК 200 в клетках OHS: (A) до и после обработки PCI (3 часа), (B) до обработки PCI: (i) при фазовоконтрастной микроскопии, (ii) при помощи окрашивания FITC-ПНК, (iii) при окрашивании LysoTracker, (iv) при окрашивании Hoechst и (v) показывающее комбинированное окрашивание; и (C) после обработки PCI при окрашивании, как для (B).
На фиг.3 показана ядерная локализация в различных типах клеток после PCI и с использованием различных молекул ПНК, используя флуоресцентную микроскопию. Клетки инкубировали с различными FITC-ПНК в концентрации 1000 нМ в течение 24 часов и анализировали при помощи флуоресцентной микроскопии, как описано в экспериментальных протоколах. (A) OHS-ПНК-NLS (ПНК381), (B) OHS-ПНК-MITO (ПНК382), (C) OHS-ПНК-GHHHHHG (ПНК457), (D) HeLa-ПНК-NLS (ПНК381), (E) FEMXIII-ПНК-NLS (ПНК381), полученные в результате, слева направо, фазовоконтрастного изображения, окрашивания FITC-ПНК, Hoechst, LysoTracker.
На фиг.4 показано, что доставка ПНК в ядро не зависит от локализации используемого флуорофора. Клетки OHS инкубировали с ПНК с FITC в присутствии C- или N-конца (1000 нМ) в течение 24 часов и анализировали при помощи флуоресцентной микроскопии, как описано в экспериментальных протоколах. Результаты, слева направо, показывают FITC, связанный либо с C-, либо с N-концом.
На фиг.5 показан захват ПНК 200 в ядро различных клеток после PCI: (A) до PCI, (B) после PCI, определяя краситель FITC-ПНК; клетки FEMX1, FEMX5, HeLa, OHS, SW620, HCT116, WiDr, 293 и SaOs, соответственно.
На фиг.6 показано, что захват ПНК зависит от температуры; клетки OHS подвергали воздействию 1000 нМ ПНК 200 при: (A) 4°C в течение 5 часов, и (B) 37°C в течение 5 часов. Результаты показаны слева направо, фазовоконтрастное изображение, FITC/ПНК, комбинированное изображение; увеличение составляет 10× на верхних изображениях и 32× на нижних изображениях.
На фиг.7 показано, что доставка ПНК в ядро не зависит от типа используемого флуорофора. Клетки инкубировали с ПНК455 (1000 нМ), конъюгированной с родамином, в течение 24 часов и анализировали при помощи флуоресцентной микроскопии, как описано в экспериментальных протоколах. Результаты, показанные слева направо, представляют фазовоконтрастное изображение, изображение с родамином и комбинированное изображение.
На фиг.8 показано влияние по-разному заряженных молекул ПНК на ядерный импорт после PCI. Клетки OHS инкубировали с ПНК (1000 нМ), как описано в экспериментальных протоколах. (A) 383, (B) 385, (C) 456, (D) 384, (E) 381, (F) 455. Результаты, показанные слева направо, представляют фазовоконтрастное изображение, FITC изображение, комбинированное изображение.
На фиг.9 показано ингибирование экспрессии S100A4 в клетках OHS различными ПНК (1000 нМ), как оценено при помощи вестерн-блоттинга: (A) зависимое от дозы ингибирование ПНК200, (B) зависимое от времени ингибирование различными ПНК. Результаты представлены в качестве процентного соотношения к контрольным клеткам, и столбы обозначают среднее значение для 3 отдельных экспериментов. Планки погрешностей показывают стандартное отклонение от среднего значения. Типичные примеры вестерн-блотта для экспериментов показаны на изображениях от (C) до (E); (C) и (D) контрольная загрузка (α-тубулин), (E) ингибирование через 96 часов представляет слева направо контроль, рандомизированную ПНК (ПНК201), ПНК200, ПНК381, (F) зависимое от дозы ингибирование с ПНК200 через 96 часов представляет слева направо контроль, 100 нМ, 500 нМ, 1000 нМ, 2000 нМ.
На фиг.10 показаны результаты MTS после обработки PCI клеток OHS. Результаты показывают, что одни только ПНК нетоксичны.
На фиг.11 показаны результаты анализа вестерн-блотт, показывающие, что никакого влияния на уровни белков в клетках OHS не наблюдается при направленном переносе ПНК к кодирующему региону S100A4 (ПНК452): (A) верхние полоски представляют собой α-тубулин в качестве контрольной загрузки, нижние полоски представляют собой S100A4, на дорожке 1 находится контроль без фотосенсибилизатора и без обработки светом, на дорожке 2 находится образец с фотосенсибилизатором, но без обработки светом, на дорожке 3 находится образец без фотосенсибилизатора, но с обработкой светом, на дорожке 4 находится образец с фотосенсибилизатором и обработкой светом; (B) верхние полоски представляют собой α-тубулин в качестве контрольной загрузки, нижние полоски представляют собой S100A4, дорожка 1 - контроль, дорожка 2 - рандомизированная ПНК (ПНК202), дорожка 3 - (ПНК452) 1000 нМ, дорожка 4 - (ПНК452) 2000 нМ.
На фиг.12 показана относительная экспрессия мРНК S100A4 в обработанных ПНК/PCI клетках OHS по сравнению с контролем. A) Полную РНК выделяли из клеток OHS после обработки ПНК/PCI при помощи ПНК-AUG, ПНК-5'UTR и ПНК-рандомизированная. Обработанные PCI клетки OHS выделяли в качестве контроля в дополнение к рандомизированной ПНК. Все образцы подвергали действию обратной транскриптазы и растворы кДНК подвергали анализу ПЦР в реальном времени, используя SYBRGreen I в качестве реагента для детекции. CT-значения, полученные для различных образцов, показывали небольшие отличия в экспрессии гена. B) Анализ кривой плавления, показывающий только один интересующий продукт.
На фиг.13 показаны уровни белка TYR через 72 часа, используя вестерн-иммуноблоттинг. Дорожки нагружали, как показано ниже: 1. Контроль (без ПНК), 2. Рандомизированная ПНК TYR (1 мМ), 3. Рандомизированная ПНК TYR (10 мМ), 4. ПНК TYR UTR (1 мМ), 5. ПНК TYR UTR (10 мМ), 6. ПНК TYR AUG (1 мМ), 7. ПНК TYR AUG (10 мМ). Альфа-тубулин показан в качестве контрольной загрузки.
ПРИМЕРЫ
Экспериментальные протоколы
Клеточная линия и условия культивирования
Клеточные линии человека HeLa (аденокарцинома шейки матки), WiDr (карцинома прямой кишки) и 293 (эмбрионная почка) получали из Американской коллекции типовых культур (Manassas, VA, USA). Человеческие OHS (остеосаркома) и FEMXIII (меланома) собирали в Norwegian Radium Hospital (Fodstad et al, (1986), Int. J. Cancer 38(1), 33-40; Fodstad et al., (1988), Cancer Res. 48(15), 4382-8). Все клеточные линии культивировали в среде RPMI-1640 (Bio Whittaker, Verviers, Belgium), за исключением клеточной линии 293, которую культивировали в среде DMEM (Bio Whittaker, Verviers, Belgium). Обе среды использовали без антибиотиков, но с добавлением 10% сыворотки зародыша теленка (FCS; PAA Laboratories, Linz, Austria) и 2 мМ L-глутамина (Bio Whittaker, Verviers, Belgium). Клеточные культуры растили и инкубировали при 37°C в увлажненной атмосфере, содержащей 5% CO2. Все клеточные линии тестировали и обнаруживали, что они были отрицательны в отношении инфицирования Mycoplasma.
Конструирование ПНК
ПНК, специфичные для гена S100A4, включающие рандомизированные ПНК, получали из Oswell DNA Service (Southampton, UK). Модификации выполняли на одном или обоих концах (смотрите таблицу 1). Мишени к 5′UTR (номер доступа в GeneBank NM_002961, 2-15), стартовому региону AUG (63-82) и кодирующему региону во втором экзоне (98-118) выбирали, основываясь на предыдущих экспериментах по ингибированию ПНК (Doyle et al., (2001), Biochem. 40, 53-64; Mologni et al., (1999), Biochem. Biophys. Res. Comm. 264, 537-543) и подавления гена S100A4 рибозимами (Hovig et al., (2001), Antisense Nucleic Acid Drug Dev. Apr 11(2), 67-75). Последовательности выравнивали по базе данных генома человека с использованием поиска в BLAST для того, чтобы исключить последовательности со значительной гомологией к другим генам. Маточные растворы (1 мМ) приготавливали растворением ПНК в 10% трифторуксусной кислоте и перед использованием нагревали до 50°C для того, чтобы гарантировать, что ПНК растворялась полностью. Перед использованием ПНК дополнительно разводили до рабочих растворов (10 мкМ) в стерильной воде и хранили при -20°C.
Конструирование siPHK и отжиг
Основываясь на правилах, предложенных Elbashir et al. (Elbashir et al, (2001), Genes Dev. 15, 188-200), две мишени выбирали против кодирующего региона гена S100A4. Первая мишень представляла собой мишень против последовательности AA(N)19 (номер доступа в GeneBank NM_002961, 343-361) и вторая мишень представляла собой последовательность AA(N19)TT (264-282). В дополнение, получали рандомизированную контрольную siPHK и флуоресцентно-меченную siPHK. Все siPHK заказывали в Eurogentec (Seraing, Belgium). Мечение проводили по обоим нитям с использованием FITC на 5'-конце антисмысловой нити и родамина на 3'-конце смысловой нити. Содержание GC двойной спирали сохраняли в диапазоне 40-70% и все siPHK синтезировали с dTdT липкими концами на их 3'-концах для оптимальной стабильности двойной спирали siPHK. Две целевые последовательности также выравнивали по базе данных генома человека с использованием поиска в BLAST с целью исключения последовательностей со значительной гомологией к другим генам. Высушенные олигонуклеотиды siPHK ресуспендировали в 100 мкМ обработанной DEPC воде и хранили при -20°C. Отжиг siPHK выполняли путем раздельного получения аликвот и разведения каждого олига РНК до концентрации 50 мкМ. Затем 30 мкл каждого раствора олига РНК и 15 мкл 5× буфера для отжига смешивали в конечной концентрации 50 мМ Tris, pH 7,5, 100 мМ NaCl в обработанной DEPC воде. Затем раствор инкубировали в течение 2 минут на водяной бане при 95°C, с последующим постепенным охлаждением в течение 45 минут на рабочем столе. Успешный отжиг подтверждали неденатурирующим электрофорезом в полиакриламидном геле.
Трансфекция siPHK и электропорация ПНК
Клетки OHS культивировали, как описано выше, и культивировали в течение 24 часов в 6-луночных планшетах до 30-60% слияния перед трансфекцией. Транфекцию проводили в несодержащей сыворотку среде OPTI-MEM I (Invitrogen Corp, Paisley, UK) с различными концентрациями siPHK, используя реактив липофектин из Life Technologies Inc. (Gaithersburg, MD, USA), реактив липофектамин из Invitrogen (Catlsbad, CA, USA), (N-(1-(2,3-диолеоксилокси)пропил)-N,N,N,-триметиламмония-метил-сульфат (DOTAP) из Boehringer Mannheim (Mannheim, Germany), FuGene из Roche Diagnostics (Mannheim, Germany), средство трансфекции липидов siPORT из Ambion (Austin, TX, USA) и гидробромид поли-L-лизина (МВ 15000-30000) из Sigma (St. Louis, MO, USA), согласно инструкциям производителя. При электропорации культивируемые клетки OHS собирали и ресуспендировали в свежей среде. Приблизительно 4×106 клеток смешивали с ПНК (1-10 мкМ) в 300 мкл среды и инкубировали на льду в течение 10 минут. Клетки электропорировали в кювете размером 0,4 см с установленными параметрами 950 мкФ/250 В (ECM399, BTX, A Division Of Genetronics, CA). После электропорации клетки инкубировали на льду в течение 30 минут, разводили в колбах T25 и инкубировали при 37°C с 5% CO2 в течение 24 часов и анализировали посредством флуоресцентной микроскопии.
PCI технология и обработка
Сенсибилизатор дисульфонированный тетрафенилпорфин (TPPS2a) закупали у Porphyrin Products (Logan, UT, USA). TPPS2a сначала растворяли в 0,1 M NaOH и впоследствии разводили в забуфференном фосфатом изотоническом растворе (PBS), pH 7,5, до концентрации 5 мг/мл и конечной концентрации 0,002 M NaOH. Фотосенсибилизатор защищали от света и хранили при -20°C до использования. При облучении клетки, обработанные TPPS2a, подвергали воздействию голубого света с использованием LumiSource прототипа (PCI Biotech AS, Oslo, Norway), содержащего ряд из четырех флуоресцентных трубок (Osram 18W/67) с наивысшей интенсивностью потока приблизительно 420 нм.
Перед использованием клетки культивировали в течение 24 часов в 6-луночных планшетах при 37°C под 5% CO2. Затем клетки инкубировали с различными ПНК и сенсибилизатором TPPS2a (1 мкг/мл) в течение 18 часов. После захвата клетки промывали 3 раза свежей средой и инкубировали в среде без сенсибилизатора в течение 4 часов. В заключение, клетки подвергали воздействию голубого света в течение 30 секунд и повторно инкубировали в течение 24, 48 и 96 часов. Во время эксперимента клетки защищали от света алюминиевой фольгой.
Флуоресцентная микроскопия
Клетки анализировали с использованием инвертированного микроскопа Zeiss, Axiovert 200, оборудованного фильтрами для FITC (450-490 нм фильтр возбуждения BP, 510 нм светоделитель FT и 515-565 нм фильтр излучения LP), радомина (546/12 нм фильтр возбуждения BP, 580 нм светоделитель FT и 590 нм фильтр излучения LP) и DAPI (365/12 нм фильтр возбуждения BP, 395 нм светоделитель FT и 397 нм фильтр излучения LP). Изображения получали с использованием Carl Zeiss AxioCam HR, Версия 5.05.10, и программного обеспечения AxioVision 3.1.2.1. Специфичные для органелл маркеры использовали для подтверждения внутриклеточной локализации ПНК. Локализацию лизосом определяли с использованием флуоресцентного микроскопа и LysoTracker Red DND-99 (Molecular Probes, Eugene OR). Ядерную локализацию определяли с использованием Hoechst H33342 (Molecular Probes, Eugene OR).
Анализ проточной цитометрии
Клетки гидролизовали трипсином, центрифугировали, ресуспендировали в 400 мкл среды для культивирования и фильтровали через нейлоновый фильтр с ячейками в 50 мкм перед тем, как анализировать на проточном цитометре FACS-Calibur (Becton Dickinson). Для каждого образца собирали 10000 случаев. Меченный FITC ПНК измеряли с фильтром 510-530 нм после возбуждения аргоновым лазером (15 мВ, 488 нм). Мертвые клетки отделяли от единичных жизнеспособных клеток путем селекции прямого рассеивания против бокового рассеивания. Данные анализировали с использованием программного обеспечения CELLQuest (Becton Dickinson).
Вестерн-блоттинг
Лизаты белков готовили в 50 мМ Tris-HCl (pH 7,5), содержащем 150 мМ NaCl и 0,1% NP-40 с пепстатином в концентрации 2 г/мл, апротинина (Sigma Chemical компания, St Louis, MO) и леупептина (Roche Diagnostics, Mannheim, Germany). Общий лизат белков (30 мкг) из каждого образца, разделяли электрофорезом в 12% SDS-полиакриламидном геле и переносили на мембраны Immobilon-P (Millipore, Bedford, MA) согласно инструкции производителя. В качестве контроля загрузки и переноса мембраны окрашивали 0,1% амидочерным. Мембраны затем инкубировали в 20 мМ Tris-HCl (pH 7,5), содержащим 0,5 M NaCl и 0,25% Tween 20 (TBST), с 10% сухим молоком (блокирующий раствор) перед инкубацией с кроличьими поликлональными антителами против S100A4 (разведенными 1:300, DAKO, Glostrup, Denmark) и мышиными моноклональными антителами против альфа-тубулина (разведенными 1:250, Amersham Life Science, Buckinghamshire, England) в TBST, содержащем 5% сухое молоко. После промывки иммунореактивные белки визуализировали с использованием вторичных антител, конъюгированных с пероксидазой хрена (разведенных 1:5000 DAKO, Glostrup, Denmark), и системы усиленной хемолюминесценции (Amersham Pharmacia Biotech, Buckinghamshire, England). Уровни белка S100A4 представляли как процентные соотношения от контрольных образцов, и альфа-тубулин использовали в качестве контрольной загрузки.
Измерение клеточной жизнеспособности с использованием MTS анализа
100 мкл клеток помещали в 96-луночные планшеты и растили в течение 24 часов при 37°C. Также охватывали отрицательный контроль, не содержащий клеток. 20 мкл реагента MTS (тетразолевая соль) добавляли по 100 мкл к каждой лунке и инкубировали в течение от 2 до 4 часов при 37°С в темноте. Затем считывали поглощение при 490 нм.
ПЦР с обратной транскриптазой в реальном времени
Клетки культивировали и обрабатывали, как описано выше. После фотохимической обработки клетки инкубировали с различными ПНК в течение 96 часов и затем собирали для выделения РНК. Общую клеточную РНК выделяли с использованием набора GenElute Mammalian Total RNA Miniprep (Sigma-Aldrich, Steinheim, GER) согласно инструкциям производителя. Для синтеза кДНК смесь праймеров, содержащих 50 пмоль олиго-dT, 3 мкг общей РНК и dH2O до 12 мкл, готовили для каждого образца. Смесь денатурировали в течение 5 минут при 65°С, затем быстро охлаждали на льду и смешивали с 18 мкл реакционной смеси до конечной концентрации 1× буфера First Strand buffer (Invitrogen), 10 мМ DTT, 0,3 мМ dNTP, 6,5 нг/мкл дрожжевой тРНК и 200 Ед (6,6 Ед/мкл) фермента Superscript II. Синтез кДНК проводили при 42°С в течение 50 минут, с последующей инактивацией в течение 15 минут при 72°C.
Для ПЦР анализа готовили 3 десятикратных разведения из каждого образца кДНК и приводили трижды все реакции, получая всего 9 пробирок для ПЦР на образец кДНК. ПЦР проводили в общем объеме 25 мкл, используя конечную концентрацию 3 мМ MgCl2, 200 мкМ dNTP, 1× буфер для ПЦР, 0,5 Ед платинового Taq, 2 мкл кДНК и 300 нМ праймеров, специфичных для гена S100A4 (прямой праймер 5'-AAGTTCAAGCTCAACAAGTCAGAAC-3' (SEQ ID NO: 9) и обратный праймер 5'-CATCTGTCCTTTTCCCCAAGA-3' (SEQ ID NO: 10)). Кроме того, все реакции метили с 1 нм флуоресцеином, как требуется для iCycler. Результаты ПЦР в реальном времени получали, используя конечное разведение 1:100000 SYBR Green I (Molecular Probes, Eugene OR) в качестве детектирующего агента. Циклы амплификации представляли собой следующие ниже: 5 минут - начальная денатурация при 95°C, с последующими 40 циклами в течение 15 секунд при 95°C/30 секунд при 60°C для амплификации продукта. Определение продуктов ПЦР в реальном времени осуществляли с использованием оптических 96-луночных планшетов и детектирующей системы iCycler iQ, произведенной Bio-Rad Laboratories, CA. Для каждого образца процедуру повторяли трижды. Для определения ложных продуктов амплификации или димеров праймеров (которые в равной степени помечены введением SYBRGreen и могут, таким образом, влиять на считывание флуоресценции) кривую плавления, то есть потерю флуоресценции при денатурации, включали в конец протокола ПЦР-амплификации. Профили плавления для каждого образца сравнивали с профилями, полученными для стандартных образцов.
Чипы кДНК
Микрочипы, используемые в данной заявке, производились в условиях лаборатории, с использованием роботизированного принтера Micro Grid II (Bio Robotics, Cambridge, UK). Данные чипы 15k кДНК человека печатали на слайдах, покрытых аминосиланом (CMT GAPS Corning Life Sciences, Corning, NY). Для подробного описания содержания чипа авторы изобретения ссылаются на: http://www.med.uio.no/dnr/microarray/index.html.
Очистка и мечение РНК
Общую РНК из культивируемых клеток, обработанных с PCI и различными ПНК, выделяли, как описано выше. Для анализа возможных эффектов в прямом направлении по последовательности в качестве следствия подавления гена S100A4, каждый отдельный чип гибридизовали с кДНК из клеток, обработанных с активной ПНК (ПНК382 или 453) и рандомизированной или контрольной ПНК (ПНК452 (контроль) или 454 (рандомизированная)). кДНК получали из 50 мкг общей РНК из каждой из данных клеточных культур и дифференциально метили Cy3- или Cy5-dCTP (Amersham Pharmacia Biotech AB) во время обратной транскрипции. Реакционная смесь содержала заякоренные 20-мерные праймеры олиго-dT (4 мкг), 40 Ед РНКзина (Promega, Madison, WI), 1 стандартный буфер, 0,01 M DTT, 0,5 мМ dATP, dCTP, dGTP и 0,2 мМ dTTP. Смесь инкубировали при 65°C на водяной бане в течение 5 минут. Затем пробирки переносили в нагревательный блок при температуре 42°С и добавляли 4 мкл (4 нмоль) одного из двух флуорофоров в соответствующие пробирки в дополнение к 400 Ед Superscript II (Invitrogen, Groningen, The Netherlands). Через 60 минут реакцию инактивировали 5 мкл 0,5 M ЕДТА (pH 8,0). Для гидролиза остаточной РНК добавляли 10 мкл 1 M NaOH и инкубировали пробирку при 65°С в течение 60 минут. Добавляли 25 мкл 1 M Tris-HCl (pH 7,5) для нейтрализации смеси. Меченную Cy3- и Cy5-кДНК разводили 0,5 × TE-буффером (pH 7,5) перед удалением несвязанного красителя и концентрированием образцов с использованием колонок Microcon YM (Ambion, Millipore Corporation, Bedford, MA).
Предварительная гибридизация слайдов
Слайды перекрестно сшивали UV при 150 кДж в течение 60 секунд. Непосредственно перед использованием слайды предварительно гибридизовали, чтобы инактивировать реакционные группы на поверхности слайдов и затем отмывали несвязанную ДНК. Маленький держатель слайда, заполненный раствором для предварительной гибридизации, предварительно нагревали при 50°С в течение 30 минут. Гибридизационный раствор содержал 1% (мас./об.) фракции V бычьего сывороточного альбумина (BSA) (Sigma-Aldrich), 3,5 × SSC и 0,1% SDS. Слайды инкубировали в предварительно нагретом растворе при 50°С в течение 25 минут, непосредственно после инкубации; слайды переносили на чистую подставку для слайдов и дважды споласкивали со встряхиванием в ультрачистой воде при комнатной температуре. Для денатурации ДНК до вида однонитевой цепи слайды встряхивали в только что вскипяченной воде в течение 2 минут и затем быстро погружали в пропан-2-ол и встряхивали в течение 30 секунд. Слайды высушивали центрифугированием.
Гибридизация и сканирование
Гибридизационная смесь в количестве 45 мкл состояла из 15 мкл каждого из меченых зондов, 16 мкг поли A (Amersham Pharmacia Biotech AB), 4 мкг дрожжевой т-РНК, 1,25 × раствора Денхарта, 5 мкг BSA, 3,5 × SSC (pH 7,5) и 0,3% SDS. Конечную смесь нагревали в течение 2 минут при 100°С и центрифугировали в течение 10 минут при 13 К перед нанесением на микрочип под LifterSlip (Erie Scientific Company, Portsmouth, NH). Затем слайд помещали в гибридизационную камеру ArrayIT (Telechem, Sunnyvale, CA) и инкубировали в течение ночи на водяной бане при 65°С. Перед сканированием покровное стекло удаляли в растворе 0,5 × SSC и 0,1% SDS. Затем слайд дважды отмывали в том же самом растворе в течение 5 минут при комнатной температуре, с последующей отмывкой 2 раза по 5 минут в промывочном растворе 0,06 × SSC. Слайд в заключение высушивали центрифугированием. Сканирование проводили с использованием сканера ScanARRAY 4000 (Packard Biosciences, Biochip Technologies LLC, Meriden, CT) и получали данные из изображений с использованием программного обеспечения GenePix Pro 4.0 (Axon Instruments Inc., Union City, CA). Данные хранили, анализировали и обрабатывали с использованием BASE (Lao H et al. BioArray Software Environment: A Platform for Comprehensive Management and Analysis of Microarray Data Genome Biology 3(8): software 0003.1-0003.6 (2002).), и вычисляли корректированную по фону интенсивность для каждого пятна путем вычитания среднего значения пикселей в локальном участке фона из среднего значения пикселей в пятне.
Пример 1: Клеточный захват молекул ПНК
Первая серия экспериментов относится к вопросу клеточного захвата, то есть происходит ли проникновение ПНК, связанных с короткими пептидами с различными суммарными зарядами, через клеточную мембрану в различных раковых клеточных линиях человека или нет. Для детекции соединений внутри клетки ПНК метили FITC либо по N-, либо по C-концу. В таблице 1 подробно представлены ПНК, использованные в эксперименте, включая их целевые последовательности, химические модификации и заряд.
Клеточный захват измеряли методом проточной цитометрии. ПНК, связанная с пептидом с суммарным отрицательным зарядом (ПНК383), не проникала в клетки OHS (фиг.1, суммарный заряд -1). Поскольку захват отрицательно заряженных молекул практически отсутствовал даже через 24 часа, авторы изобретения исследовали возможность транфекции в клетки посредством электропорации. В данном случае захват также являлся очень слабым. В отличие от отрицательно заряженной ПНК нейтральная ПНК без заряда и безо всякого связанного пептида (ПНК456) и ПНК, связанная с пептидом с нейтральным суммарным зарядом (ПНК385), обе интернализовались в низкой степени (фиг.1, суммарный заряд 0 для ПНК385, ПНК456 не показан).
В дальнейшем авторы изобретения исследовали положительно заряженные ПНК. Захват явным образом наблюдали, когда ПНК связывали с пептидом с суммарным зарядом +1 (ПНК384) (фиг.1, суммарный заряд +1). Однако, когда авторы изобретения увеличивали суммарный заряд до +5 (ПНК381), авторы изобретения наблюдали почти 5-кратное увеличение клеточного захвата, по сравнению с ПНК с зарядом +1 (фиг.1, суммарный заряд +5).
Результаты описанных выше экспериментов, проведенных на клетках OHS, FEMXII и Hela, суммированы в таблице 2.
Для идентификации локализации молекул ПНК использовали окрашивание. Флуоресценцию меток, присоединенных к молекуле ПНК, использовали для идентификации ПНК внутри клетки. Окрашивание Hoechst и окрашивание LysoTracker использовали для идентификации ядерных и лизосомальных компартментов соответственно. На фиг.2A показано, что после PCI молекулы ПНК распределяются внутри клетки. На фиг.2B и C показано, что молекулы ПНК распределяются в ядрах после PCI (то есть, их распределение совпадает с окрашиванием Hoechst).
Для того чтобы исследовать, зависит ли клеточный захват от конформации пептида NLS или только от заряда, авторы изобретения связывали ПНК с 29-аминокислотным сигналом митохондриального импорта с суммарным зарядом +5 (ПНК382) (фиг.3B). В качестве второго контроля к ПНК381 (фиг.3A) авторы изобретения заменили первоначальную аминокислотную последовательность NLS PKKKRKV (SEQ ID NO: 3) на альтернативную последовательность GHHHHHG (+5) (SEQ ID NO:5, ПНК457) (фиг.3C). Относительные уровни клеточного захвата для трех различных конструкций ПНК представляли те же самые уровни, которые обнаруживали с использованием микроскопии. Во всех случаях ПНК локализовалась в ядрах после PCI. Это также происходило, когда осуществляли процедуру на клетках HeLa и FEMXIII (фиг.3D и E).
Для того чтобы исследовать любые возможные различия в клеточном захвате, которые могли бы относится к ориентации NLS, авторы изобретения связывали пептиды и с N- (ПНК453), и с C-концами (ПНК381). Никаких различий в уровнях захвата не наблюдали (фиг.4). Авторы изобретения также тестировали различные ПНК и их клеточный захват в различных клеточных линиях (HeLa, WiDr, 293, OHS, FEMX5, SW620, HCT116, SaOs), но не наблюдали значительных изменений (фиг.5).
Данные авторов изобретения показывают, что захват химерных ПНК сильно зависит от суммарного заряда молекулы пептида и не зависит от конформации аминокислот. Здесь авторы изобретения показали, что захват молекул ПНК увеличивается, когда положительный суммарный заряд на конъюгированном пептиде становится больше.
Пример 2: Механизм и локализация захвата ПНК
Для того чтобы оценить процесс захвата для модифицированных ПНК, авторы изобретения, во-первых, исследовали их захват при различных температурах. Результаты авторов изобретения не показали захвата клеткой при 4°C, хотя захват наблюдали при 37°C (фиг.6). Более того, данные авторов изобретения по флуоресцентной микроскопии показывали зерноподобные флуоресцирующие пятна в отдельных областях в непосредственной близости от ядерной оболочки (фиг.6B). Наконец, авторы изобретения наблюдали хорошее перекрытие между внутриклеточной локализацией конструкций ПНК и маркера для эндосом/лизосом (LysoTracker Red DND) (данные не показаны).
Результаты авторов изобретения показывают, что вовлечен эндоцитоз. Это может быть вызвано интернализацией через окаймленные пузырьки. В экспериментах авторов изобретения захват ПНК авторов изобретения блокировался при 4°С.
Результаты авторов изобретения подтверждаются открытиями Kuismanen и Saraste (Kuismanen E et al. (1989) Methods. Cell. Biol. 32, 257-274), которые показали, что эндоцитоз можно блокировать при низких температурах. Температурная зависимость, кроме того, подтверждалась перекрывающейся локализацией ПНК и LysoTracker.
Эндоцитоз можно разделить на несколько основных типов: обусловленный рецептором зависимый от кларитина, независимый от кларитина и фагоцитоз. Однако для выяснения определенного типа эндоцитоза необходимо проводить дальнейшие исследования. Предположение заключается в том, что молекулы ПНК авторов изобретения захватываются посредством независимого от кларитина эндоцитоза, поскольку нет доказательств обусловленного рецептором зависимого от кларитина эндоцитоза. ПНК связывали с различными пептидными сигналами с одинаковым зарядом; результаты были одинаковые, указывая на то, что захват не зависит от определенного рецептора. Заключение состоит в том, что положительно заряженная молекула ПНК, наиболее вероятно, имеет тесную связь с клеточной мембраной, чем отрицательно заряженная, которая в свою очередь будет увеличивать эндоцитозный захват.
Пример 3: Эффект обработки PCI
Данные по микроскопии авторов изобретения показали, что ПНК с нейтральным/положительным суммарным зарядом локализуются в эндосомах/лизосомах. Данные по микроскопии авторов изобретения ясно показали, что конструкции ПНК перемещаются из эндосом/лизосом в ядро после обработки PCI (фиг.2 и 3). Для того чтобы подтвердить релокализацию ПНК, авторы изобретения также использовали окрашивание ядра Hoechst, как описано выше, смотрите фиг.2 и 3.
Пример 4: Ядерный импорт молекул ПНК
Для ядерного направленного переноса ПНК необходимо преодолеть основные барьеры: самые важные представляют собой мембрану клетки, внутриклеточные мембраны и ядерный конверт. После высвобождения ПНК из эндосом/лизосом, авторы изобретения хотели исследовать способность к локализации пептида NLS и являлась ли ориентация пептида NLS важной для ядерной локализации. Чтобы разрешить данные вопросы, авторы изобретения связывали пептид NLS с ПНК как с N-, так и с C-концом. Данные по микроскопии авторов изобретения показали, что ПНК, связанные с пептидом NLS либо через N-, либо через C-конец, перемещаются в ядро (фиг.4). Усиление сигнала флюоресценции наблюдали, увеличивая время воздействия и концентрацию конструкции FITC/ПНК. Для того чтобы проанализировать любую возможную несовместимость между различными типами флуорофоров, авторы изобретения меняли флуорофоры ПНК от FITC до родамина (Rho). Это, кроме того, не дало никакого видимого изменения в локализации (фиг.7). Авторы изобретения также связывали FITC либо с N-, либо с C-концами ПНК, снова не было изменения в локализации или эффективности (фиг.4).
Далее авторы изобретения исследовали, являлась ли способность к ядерной локализации PKKKRKV (SEQ ID NO: 3) просто следствием изменения заряда в заряде ПНК или следствием определенной аминокислотной последовательности. Для того чтобы проконтролировать способность к ядерной локализации пептида NLS, авторы изобретения тестировали ПНК с альтернативным пептидом, обладающим тем же самым суммарным зарядом (+5), но с замещенными аминокислотами (ПНК457, фиг.3C). Также авторы изобретения исследовали ядерный импорт нейтральных ПНК (ПНК456 и 385, фиг.8C и B соответственно), и также с ПНК, связанными с пептидными сигналами для импорта, направленными к митохондриям и пероксисомам (ПНК382, ПНК384, фиг.3B и 8D соответственно). Как это ни удивительно, данные авторов изобретения демонстрировали, что все нейтральные и положительно заряженные исследуемые ПНК перемещались в ядро после обработки PCI.
В заключение, результаты авторов изобретения демонстрировали, что ПНК с нейтральным/положительным суммарным зарядом не только самопроизвольно перемещаются из среды в лизосомы на высоком уровне, но также и перемещаются из цитозоля в ядро после фотохимической обработки. Эффективность между нейтральными и положительно заряженными ПНК изменяется, вероятно, в качестве непосредственного следствия изменений клеточного захвата, а не ядерного захвата.
Пример 5: Ингибирование экспрессии S100A4 при помощи ПНК/PCI
Для того чтобы оценить функциональную возможность ПНК в качестве ингибитора, авторы изобретения синтезировали ПНК для направленного переноса к трем различным целевым участкам гена S100A4. Авторы изобретения выбрали химерную гомопуриновую ПНК из 14 н.п. для направленного переноса к концу 5'-UTR (ПНК381) и две ПНК со смешанными основаниями из 20 н.п. к стартовому кодону (ПНК200) и кодирующей области во втором экзоне (ПНК452). Авторы изобретения хотели исследовать, можно ли осуществить понижающую регуляцию S100A4 зависимым от дозы образом. Авторы изобретения поэтому воздействовали на клетки OHS различными концентрациями (100-2000 нм) ПНК200 в течении 96 часов и проверяли на жизнеспособность и присутствие белка S100A4 вестерн-блоттингом (фиг.9A и F). Данные авторов изобретения ясно демонстрируют зависимое от дозы ингибирование активности S100A4, со снижением передачи сигналов, начинающимся с концентрации 100 нМ при использовании ПНК200. Подходящие типичные примеры контролей показаны на фиг.9D. Более того, результаты авторов изобретения показывали, что ПНК200 при концентрации 1000 нМ вызывала максимальное ингибирование экспрессии S100A4. На основании данных MTS, которые измеряют митохондриальную целостность в качестве меры жизнеспособности клетки (смотрите экспериментальные протоколы для подробного описания), не существует никакой заметной токсичности при использовании концентраций ПНК менее 2000 нМ (фиг.10, линии 5 и 6). Таким образом, концентрацию в 1000 нМ ПНК выбрали для всех последующих экспериментов.
Чтобы оценить, происходила ли понижающая регуляция уровней белка Sl00A4 зависимым от времени образом, авторы изобретения инкубировали клетки с ПНК в течение 24, 48 и 96 часов (фиг.9B). Через 24 часа уровень белка S100A4 уменьшался на 45% (ПНК200) и 35% (ПНК381). При более длительном времени воздействия (48 часов) экспрессия снижалась до 25% (ПНК200) и 35% (ПНК381), по сравнению с контрольным уровнем, соответственно. Наконец, экспрессия S100A4 снижалась до 10% (ПНК200) и 20% (ПНК381), по сравнению с контролем в клетках, инкубированных с ПНК в течение 96 часов после PCI (фиг.9B, C и E). Данные авторов изобретения указывали на то, что ПНК, направленные и к стартовому участку AUG (ПНК200), и к концу 5'-UTR (ПНК381), ингибируют экспрессию S100A4, и что ПНК, направленная к стартовому участку AUG, представляла собой наиболее эффективный ингибитор. Напротив, авторы изобретения не обнаруживали никакого ингибирования экспрессии S100A4 при помощи ПНК, направленной ко второму экзону (ПНК452), как определяли вестерн-блоттингом (фиг.11).
Также исследовали относительную экспрессию мРНК S100A4. Полную РНК выделяли из клеток OHS после обработки ПНК/PCI при помощи ПНК-AUG, ПНК-5'-UTR и ПНК-рандомизированной. Обработанные PCI клетки OHS выбирали в качестве контроля в дополнение к рандомизированной ПНК. Все образцы подвергали действию обратной транскриптазы и растворы кДНК подвергали анализу ПЦР в реальном времени, используя SYBRGreen I в качестве реагента для детекции. CT-значения, полученные для одинаковых растворов, показывали небольшие отличия в экспрессии гена. Результаты показаны в таблице 3.
Пример 6: Ингибирование экспрессии S100A4 при помощи siPHK
Авторы изобретения сравнивали способность ПНК ингибировать экспрессию S100A4 со способностью siPHK делать то же самое. Для анализа эффективности трансфекции siPHK и распределения, авторы изобретения метили одну из четырех siPHK родамином и FITC. Авторы изобретения затем тестировали различные реагенты трансфекции и концентрации. Данные микроскопии авторов изобретения не показывали никакого захвата при использовании ни FuGene, Lipofectamin, siPORT, ни Lipofectin (данные не показаны). Однако захват показывали как с использованием DOTAP, так и поли-L-лизина, причем поли-L-лизина в качестве наиболее эффективного агента. Авторы изобретения поэтому использовали поли-L-лизин во всех последующих экспериментах.
В экспериментах авторов изобретения siPHK конструировали согласно Elbashir et al. ((2001), Genes Dev. 15, 188-200). В дополнение к siPHK, сконструированной к выбранному гену-мишени, конструировали контрольную siPHK, получая рандомизированную siPHK, поиск BLAST осуществляли в GenBank, чтобы удалить ложную гибридизацию. Клетки OHS инкубировали с siPHK в течение различных периодов времени и с различными концентрациями, с последующими измерениями уровня белка S100A4, проводимыми при помощи вестерн-блотта. Кроме того, авторы изобретения не наблюдали никакой понижающей регуляции экспрессии S100A4 через 24, 48 и 96 часов, используя 20, 50, 100 нМ siPHK, направленной к двум регионам гена S100A4 (данные не показаны).
ПНК может действовать, блокируя транскрипционные процессы посредством их способности образовывать устойчивые структуры тройных спиралей, комплекс, встроенный в нить или со смещенной нитью с ДНК. Такие комплексы могут создать структурные помехи для блокирования устойчивой функцию РНК-полимеразы и могут таким образом оказаться способными к действию в качестве антигенных средств. На уровне трансляции эффект антисмысловой ПНК основан на стерическом блокировании либо процессинга РНК, транспорта в цитоплазму, либо трансляции. Неспособность ПНК активировать РНазу H устраняет вероятность непреднамеренной деградации нецелевых мРНК. Дополнительно, отсутсвие отрицательно заряженной основной цепи предотвращает связывание ПНК со многими белками внутри и снаружи клеток, что обычно происходит со связанными отрицательно заряженными макромолекулами. Ингибирующий эффект ПНК381 находится в согласии с Doyle et al. (2001, см. выше), который продемонстрировал, что ПНК, направленные к концу 5'-UTR, представляли собой эффективные ингибиторы мРНК люциферазы. Более того, эксперименты по трансляции, проведенные на бесклеточных экстрактах, показали, что ПНК блокировала трансляцию зависимым от дозы образом, когда была направлена близко к стартовому кодону AUG РНК (Knudsen & Nielsen (1996), Nucleic Acids Res. 24, 494-500). Никакого эффекта не наблюдали, когда ПНК направляли к последовательностям в кодирующей области. Эти результаты подтверждают результаты авторов изобретения с ПНК200 и ПНК452, направленными к стартовому участку AUG и второму экзону S100A4, соответственно (фиг.9 и 11). Паттерны экспрессии S100A4 в клетках, подвергавшихся воздействию рандомизированной ПНК201/202, и контрольных клетках без ПНК, но с сенсибилизатором, фактически идентичны необработанным клеткам. Дополнительно авторы изобретения проверяли эксперссию S100A4 в клетках OHS с или без сенсибилизатора. Опять же никаких различий в уровнях белка не смогли наблюдать (фиг.11А).
Пример 7: Анализ ПЦР с обратной транскриптазой в реальном времени
Для того чтобы исследовать основной механизм подавления гена, авторы изобретения измеряли относительные уровни мРНК S100A4 при помощи ОТ-ПЦР в реальном времени до и после обработки ПНК/PCI. Цель заключалась в том, чтобы исследовать, оказывали ли молекулы ПНК по изобретению воздействие на уровне транскрипции, или на неком другом уровне процесса синтеза белка. Как можно заметить по фигуре амплификации, не существовало никаких отличающих различий между величинами CT, полученными из обработанных ПНК/PCI образцов и необработанных контролей через 96 часов после обработки (фиг.12). Рандомизированную ПНК201 использовали в качестве внутренней контрольной ПНК для ПНК200 и ПНК381, соответственно.
Ранее Demidov et al. ((1995) Proc. Natl. Acad. Sci. U.S.A., 92, 2637-2641) исследовали кинетику и механизм связывания ПНК с двойной спиралью ДНК. Результаты показывали, что образование тройного комплекса встраивания зависит от гомопиримидиновых ПНК, связывающихся с гомопуриновой целевой ДНК. Второй комплекс, называемый двойным комплексом встраивания, можно также получить, но с гомопуриновыми ПНК. Обычный триплекс, по-видимому, образован только с богатыми цитозином гомопиримидиновыми ПНК. Такие результаты подразумевают, что единственная молекула ПНК, которую использовали авторы изобретения, которая способна к направленному переносу к двойной спирали ДНК, представляла собой гомопуриновую ПНК381, направленную к концу 5'-UTR. Кроме того, даже при том, что гомопуриновую ПНК (ПНК381) по изобретению конструировали для того, чтобы связать ее и с ДНК, и с РНК, данные ОТ-ПЦР в реальном времени указывают на то, что она действует на уровне трансляции. Другие ПНК со смешанными основаниями по составу в соответствии с теорией не способны к образованию тройного или двойного комплекса встраивания, который необходим для блокирования транскрипционных процессов. Это согласуется с результатами ОТ-ПЦР в реальном времени авторов изобретения и поддерживает теорию, что ПНК со смешанными основаниями (ПНК200) неспособна блокировать транскрипционные процессы.
Пример 8: Анализ микрочипов
Для того, чтобы исследовать возможные эффекты ингибирования S100A4 на транскрипцию гена в экспериментах с микрочипами кДНК, авторы изобретения сравнивали обработанные ПНК клетки с клетками, подвергающимися той же самой обработке в то же самое время, но рандомизированной ПНК. Исследовали молекулы ПНК, направленные к двум целевым последовательностям генов. Все эксперименты проводили в двух повторах. Только небольшое количество генов сообразно показывали относительное изменение экспрессии больше, чем в два раза. Используя иерархическое объединение в кластеры, определяли кластер, который показывал паттерн стойкой повышающей регуляции при обработке ПНК, которая вызывала наибольшее снижение уровней S100A4, и меньшей повышающей регуляции с менее эффективной ПНК. Данный кластер содержал девять названных генов (GAS2, UBE4B, FREQ, SHC1, PON3, CTSD, WNT3A, SCD и RAB6A). Они представляют собой гены, вовлеченные в процессы, включающие в себя ответ на стресс, апоптоз и связывание кальция. Проверяли уровни понижающей регуляции транскриптов, используя ПЦР в реальном времени. Для того чтобы продемонстрировать, что наблюдаемые изменения являлись результатом воздействия ПНК определенной последовательности на ген-мишень, ПЦР в реальном времени, проведенная на каждой стадии процесса отдельно, подтверждала данное заключение (данные не показаны).
В качестве первой иллюстрации способности к использованию ПНК/PCI/LS для систематического подавления генов, авторы изобретения исследовали глобальные изменения уровня экспрессии мРНК, используя микрочипы кДНК. Поскольку влияние на экспрессию генов добавления ПНК и/или фотохимической обработки в настоящий момент не известны, авторы изобретения проводили эксперименты с микрочипами, используя клетки, обработанные с рандомизированной ПНК в качестве канала сравнения. Это было сделано для того, чтобы минимизировать потенциальное мешающее влияние режима обработки. Поскольку незначительное изменение в обработке, времени воздействия и так далее все еще может иметь место, авторы изобретения проводили ПЦР в реальном времени на каждой стадии процесса обработки, чтобы исключить то, что наблюдаемые изменения экспрессии являлись следствием обработки, а не определенного воздействия на S100A4. В частности, PCI могла бы привести к транскрипционным изменениям, связанным с процессами, включающими в себя апоптоз (Ferreira S. D. et al, 2004, Lasers Med Sci 18(4): 207-12). Таким образом, в качестве общего средства ПНК/PCI/LS, как думали бы, не так хорошо подходит для контролирования генного подавления генов, связанных с такими процессами. Кроме того, стратегия обладает множеством привлекательных аспектов в легкости конструирования целевой последовательности, стабильности ПНК, высокопроизводительного синтеза и введения, и регулируемая во времени доставка делает данную систему хорошим выбором для систематического подавления клеточных линий in vitro. Генное подавление siPHK, как также продемонстрировали, представляет собой очень жизнеспособную стратегию, но с некоторыми проблемами, связанными с конструированием целевой последовательности и стабильностью (Amarzguioui M et al., 2004: Biochem. Biophys. Res. Commun. 316(4): 1050-8).
Способность S100A4 модулировать экспрессию генов вообще неизвестна, но поскольку он представляет собой белок, который, как предполагают, вовлечен в реконструирование цитоскелета, и как таковой является белком клеточной структуры, а не регулятором транскрипции, ожидаются относительно незначительные эффекты. Соответственно, наблюдаемые изменения уровня транскрипции относительно незначительны и в отношении амплитуды, и в отношении количества затронутых генов. Среди генов, для которых можно наблюдать стойкое изменение, фреквенин является особенно интересным, будучи связывающим кальций белком с четырьмя связывающими доменами (поликлональное антитело доступно на www.abcam.com).
Пример 9: Генное подавление гена тирозиназы (TYR) в клеточной линии меланомы (FEMX V), используя ПНК/PCI
Авторы изобретения конструировали ПНК к различным генам, вовлеченным в биосинтез меланина; включающим в себя тирозиназу (TYR), с тирозиназой связанный белок 1 (TRP-I) и фактор транскрипции микрофтальмии (MITF). Цель авторов изобретения заключается в том, чтобы подавить все три гена при использовании метода ПНК/PCI. TYR, TRP-I и MITF связаны друг с другом, что делает их интересными в качестве модельной системы. Так как эти гены связаны друг с другом, будет очень интересно исследовать возможное влияние на уровни белков TYR/TRP-1, когда авторы изобретения выключат MITF.
Клеточная линия и условия культивирования
Клетки FEMX V (меланомы) собирали в Norwegian Radium Hospital. Клетки культивировали в среде RPMI-1640 (Bio Whittaker, Verviers, Belgium). Среды использовали без антибиотиков, но с добавлением 10% сыворотки зародыша теленка (FCS; PAA Laboratories, Linz, Austria) и 2 мМ L-глутамина (Bio Whittaker, Verviers, Belgium). Клетки растили и инкубировали при 37°C в увлажненной атмосфере, содержащей 5% CO2. Клетки тестировали и обнаруживали, что они отрицательны в отношении инфицирования Mycoplasma.
Конструирование ПНК
Специфичные для гена тирозиназы (TYR) ПНК, включающие рандомизированные ПНК, получали из Oswell DNA Service (Southampton, UK). Модификации проводили на обоих концах (FAM и последовательностью NLS). Мишени к стартовому кодону AUG выбирали, основываясь на предыдущих исследованиях по ингибированию ПНК. Последовательности выравнивали по базе данных генома человека при поиске в BLAST для того, чтобы исключить последовательности со значительной гомологией к другим генам.
Использовали следующие ниже последовательности ПНК:

Каждую молекулу ПНК метили FAM на N-конце (5'-конец) и пептидом NLS на C-конце (3'-конец), то есть FAM-L-L-ПНК-L-L-PKKKRKV, где L представляет собой линкер (2-аминоэтокси-2-этоксиуксусная кислота (AEEA)).
Маточные растворы (1 мМ) готовили растворением ПНК в 0,1% трифторуксусной кислоте и перед использованием нагревали до 50°C для того, чтобы гарантировать, что ПНК растворялась полностью. Перед использованием ПНК дополнительно разводили до рабочих растворов (100 мкМ) в стерильной воде и хранили при -20°C.
Молекулы ПНК вводили в клетки FEMX V, используя способ PCI, как описано в примере 6, и уровни белков определяли при помощи вестерн-блоттинга.
Результаты, как показано на фиг.13, демонстрируют, что TYR можно подавить способом ПНК/PCI, однако дальнейшая оптимизация приведет к более сильному эффекту генного подавления. В частности, на дорожке номер 7 показана понижающая регуляция белка TYR после инкубации с 10 мМ ПНК для направленного переноса к области стартового кодона.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ВВЕДЕНИЯ siRNA В КЛЕТКИ ФОТОХИМИЧЕСКОЙ ИНТЕРНАЛИЗАЦИЕЙ | 2007 |
|
RU2510826C2 |
| ПОЛИКОНЪЮГАТЫ ДЛЯ ВВЕДЕНИЯ IN VIVO ПОЛИНУКЛЕОТИДОВ | 2007 |
|
RU2430740C2 |
| ФОТОСЕНСИБИЛИЗИРУЮЩИЕ КОМПОЗИЦИИ | 2010 |
|
RU2574019C2 |
| МОЛЕКУЛЯРНЫЕ КОНЪЮГАТЫ С ПОЛИКАТИОННЫМ УЧАСТКОМ И ЛИГАНДОМ ДЛЯ ДОСТАВКИ В КЛЕТКУ И ЯДРО КЛЕТКИ ДНК И РНК | 2009 |
|
RU2537262C2 |
| ФОТОХИМИЧЕСКАЯ ИНТЕРНАЛИЗАЦИЯ ДЛЯ ДОСТАВКИ МОЛЕКУЛ В ЦИТОЗОЛЬ | 2001 |
|
RU2333246C2 |
| Способ получения плазмидной ДНК в составе полимерных наносфер для доставки | 2022 |
|
RU2817116C1 |
| УСОВЕРШЕНСТВОВАННЫЕ ЛИПОСОМЫ И ИХ ПРИМЕНЕНИЕ | 2008 |
|
RU2482837C2 |
| ОЛИГОРИБОНУКЛЕОТИДЫ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ ДЛЯ ЛЕЧЕНИЯ ОСТРОЙ ПОЧЕЧНОЙ НЕДОСТАТОЧНОСТИ И ДРУГИХ ЗАБОЛЕВАНИЙ | 2005 |
|
RU2434942C2 |
| ОЛИГОРИБОНУКЛЕОТИДЫ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ ДЛЯ ЛЕЧЕНИЯ ОСТРОЙ ПОЧЕЧНОЙ НЕДОСТАТОЧНОСТИ | 2005 |
|
RU2584609C2 |
| ОЛИГОПЕПТИДНЫЕ СОЕДИНЕНИЯ И ИХ ПРИМЕНЕНИЯ | 2009 |
|
RU2549675C2 |








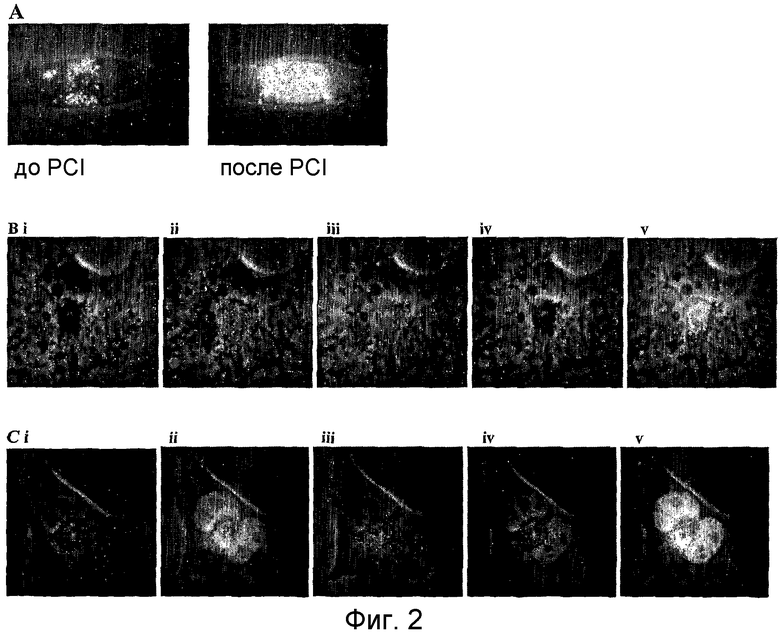






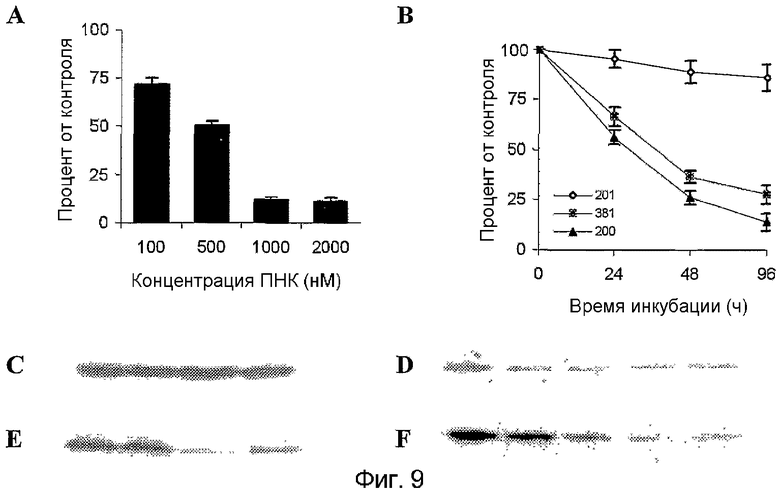
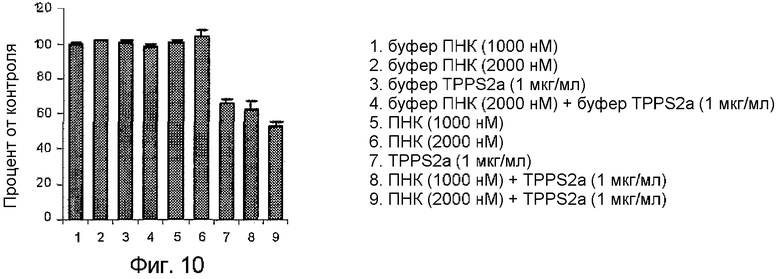


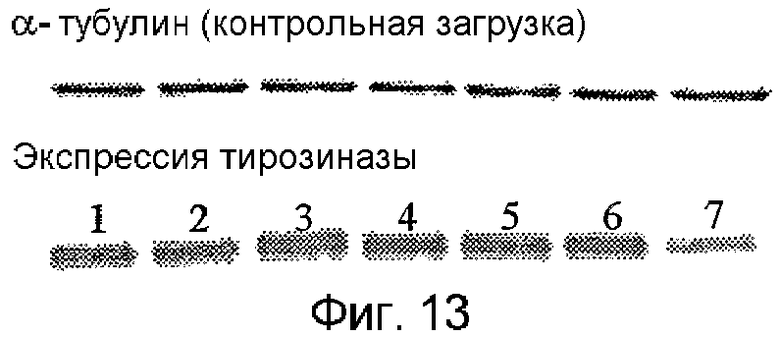
Изобретение относится к биотехнологии, к способу введения молекулы пептидной нуклеиновой кислоты (ПНК) в цитозоль или в ядро клетки. Способ предусматривает контактирование указанной клетки с молекулой ПНК и фотосенсибилизирующим агентом и облучение клетки светом длиной волны, эффективной для активации фотосенсибилизирующего агента, где указанная молекула ПНК конъюгирована с положительно заряженным пептидом. Также описаны композиции, содержащие такие конъюгированные молекулы ПНК, клетки, полученные при использовании способа, и применения способа. Описанный способ обеспечивает эффективный захват клеткой молекул ПНК, конъюгированных с положительно заряженным пептидом. 15 н. и 22 з.п. ф-лы, 13 ил.
1. Способ введения молекул ПНК в цитозоль клетки млекопитающего, предусматривающий контактирование указанной клетки с молекулами ПНК и фотосенсибилизирующим агентом и облучение клетки светом длиной волны, эффективной для активации фотосенсибилизирующего агента, где указанные молекулы ПНК конъюгированы с положительно заряженным пептидом.
2. Способ введения молекул ПНК в ядро клетки млекопитающего, предусматривающий контактирование указанной клетки с молекулами ПНК и фотосенсибилизирующим агентом и облучение клетки светом длиной волны, эффективной для активации фотосенсибилизирующего агента, где указанные молекулы ПНК конъюгированы с положительно заряженным пептидом.
3. Способ по п.1 или 2, в котором указанные молекулы ПНК составляют в длину менее 25 оснований.
4. Способ по п.1 или 2, в котором указанные молекулы ПНК представляют собой антисмысловые молекулы, комплементарные гену молекулы или зонды.
5. Способ по п.1 или 2, в котором введение указанных молекул ПНК происходит при такой концентрации, которая вызывает снижение экспрессии гена-мишени более чем на 10% после инкубации с клетками в течение 24 ч.
6. Способ по п.1 или 2, в котором указанный фотосенсибилизирующий агент выбран из TPPS4, TPPS2a, AlPcS2a, ТРС52а5-аминолевулиновой кислоты и сложных эфиров 5-аминолевулиновой кислоты.
7. Способ по п.1 или 2, в котором указанный положительно заряженный пептид представляет собой полимер последовательных аминокислот.
8. Способ по п.1 или 2, в котором указанный положительно заряженный пептид составляет в длину от 3 до 30 аминокислот.
9. Способ по п.1 или 2, в котором указанный положительно заряженный пептид конъюгирован непосредственно с молекулой ПНК посредством ковалентного связывания.
10. Способ по п.1 или 2, в котором указанный положительно заряженный пептид имеет заряд от +1 до +10, предпочтительно от +3 до +6.
11. Способ по п.1 или 2, в котором указанный положительно заряженный пептид содержит последовательность Xn-(Y)m-Xo, где Х представляет собой нейтральный остаток и Y представляет собой положительно заряженный остаток, который может быть одинаковым или разным в каждом положении, и n, m и о представляют собой целые числа ≥1.
12. Способ по п.11, в котором Y одинаков в каждом положении и представляет собой К, R или Н.
13. Способ по п.1 или 2, в котором указанный положительно заряженный пептид представляет собой SEQ ID NO: 7 MSVLTPLLLRGLTGSARRLPVPRAKIHSL, SEQ ID NO: 6 AKL или SEQ ID NO: 5 GHHHHHG.
14. Способ по п.1 или 2, в котором более одного типа молекул ПНК вводят одновременно, где каждый тип имеет различную последовательность.
15. Способ по п.1 или 2, в котором один или оба из фотосенсибилизирующего агента и молекулы ПНК присоединены, или связаны, или конъюгированы с одной или несколькими молекулами носителя или молекулами для направленного переноса.
16. Способ по п.15, в котором указанная молекула носитель или молекула для направленного переноса представляет собой поликатион, катионный липид, липофектин или пептид.
17. Способ по п.1 или 2, в котором указанный фотосенсибилизирующий агент и указанные молекулы ПНК направляют в клетку совместно или последовательно.
18. Способ по п.1 или 2, в котором указанный способ осуществляют посредством контактирования указанной клетки с фотосенсибилизирующим агентом, контактирования указанной клетки с молекулами ПНК, которые необходимо ввести, и облучения указанной клетки светом длиной волны, эффективной для активации фотосенсибилизирующего агента, где указанное облучение осуществляют до клеточного захвата указанных молекул ПНК во внутриклеточный компартмент, содержащий указанный фотосенсибилизирующий агент, предпочтительно, до клеточного захвата указанных молекул в любой внутриклеточный компартмент.
19. Способ по п.1 или 2, в котором облучение осуществляют вплоть до 60 мин.
20. Способ по п.1 или 2, осуществляемый in vitro или ex vivo.
21. Способ ингибирования транскрипции или экспрессии гена-мишени посредством введения молекулы ПНК в клетку, содержащую указанный ген-мишень, способом по любому из пп.1-20, в котором указанная молекула ПНК специфически связывается с указанным геном-мишенью или продуктом его репликации или транскрипции.
22. Способ определения или оценки уровня гена-мишени или продукта его репликации или транскрипции в клетке, где указанный способ предусматривает введение молекулы ПНК в клетку, содержащую указанный ген-мишень или продукт его репликации или транскрипции, способом по любому из пп.1-20, в котором указанная молекула ПНК специфически связывается с указанным геном-мишенью или продуктом его репликации или транскрипции, и оценку уровней связанной ПНК, чтобы определить наличие или уровень указанного гена-мишени или продукта его репликации или транскрипции.
23. Способ осуществления репарации гена-мишени, предпочтительно дефектного гена в клетке, где указанный способ предусматривает введение молекулы ПНК способом по любому из пп.1-20 и введение молекулы донорного олигонуклеотида, содержащей необходимую последовательность, в клетку, содержащую указанный ген-мишень, в которой указанная молекула ПНК специфически связывается с указанным геном-мишенью, чтобы сформировать крепление ПНК.
24. Способ in vitro диагностики заболевания, состояния или нарушения, предусматривающий введение молекулы ПНК в клетку согласно способу по любому из пп.1-20, в котором указанная молекула ПНК специфически связывается с геном-мишенью или продуктом его репликации или транскрипции, который свидетельствует о наличии указанного заболевания, состояния или нарушения, и оценку уровня связанной ПНК, чтобы определить наличие, стадию или прогноз развития указанного заболевания, состояния или нарушения.
25. Способ лечения заболевания, при котором благоприятна понижающая регуляция, репарация или мутация одного или нескольких генов, предусматривающий введение молекулы ПНК в клетку способом по любому из пп.1-20, в котором указанное заболевание предпочтительно представляет собой злокачественную опухоль, кистозный фиброз, сердечно-сосудистое заболевание, вирусную инфекцию, диабет, амиотрофический латеральный склероз, болезнь Хантингтона или болезнь Альцгеймера.
26. Клетка или популяция клеток, содержащая молекулу ПНК, которая была интернализована в цитозоль или ядро указанной клетки способом по любому из пп.1-20, где указанная молекула ПНК конъюгирована с положительно заряженным пептидом и, где клетка предназначена для применения при лечении или профилактике заболевания, нарушения или инфекции у пациента.
27. Композиция, содержащая молекулу ПНК и фотосенсибилизирующий агент, для применения при лечении или профилактике заболевания, нарушения или инфекции, для которых типична аномальная экспрессия гена или при которых благоприятна понижающая регуляция, репарация или мутация одного или нескольких генов, где композиция предназначена для использования в способе, который содержит введение молекулы ПНК в цитозоль или ядро клетки, предусматривающем контактирование указанной клетки с молекулой ПНК и фотосенсибилизирующим агентом и облучение клетки светом длиной волны, эффективной для активации фотосенсибилизирующего агента, и в которой указанная молекула ПНК конъюгирована с положительно заряженным пептидом, где предпочтительно указанная молекула ПНК составляет в длину менее 25 оснований, и/или указанная молекула ПНК представляет собой антисмысловую молекулу, комплементарную гену молекулу или зонд, и/или указанный положительно заряженный пептид представляет собой полимер последовательных аминокислот, и/или указанный положительно заряженный пептид составляет в длину от 3 до 30 аминокислот, и/или указанный положительно заряженный пептид конъюгирован непосредственно с молекулой ПНК посредством ковалентного связывания, и/или указанный положительно заряженный пептид имеет заряд от +1 до +10, предпочтительно от +3 до +6, и/или указанный положительно заряженный пептид содержит последовательность Xn-(Y)m-Xo, где Х представляет собой нейтральный остаток и Y представляет собой положительно заряженный остаток, который может быть одинаковым или разным в каждом положении, и n, m и о представляют собой целые числа ≥1, и/или Y одинаков в каждом положении и представляет собой К, R или Н, и/или указанный положительно заряженный пептид представляет собой SEQ ID NO: 7 MSVLTPLLLRGLTGSARRLPVPRAKIHSL, SEQ ID NO: 6 AKL или SEQ ID NO: 5 GHHHHHG.
28. Композиция, содержащая клетку или популяцию клеток по п.26, для применения при лечении или профилактике заболевания, нарушения или инфекции, для которых типична аномальная экспрессия гена или при которых благоприятна понижающая регуляция, репарация или мутация одного или нескольких генов.
29. Композиция по п.27 или 28 для применения при терапии злокачественной опухоли или генотерапии.
30. Применение молекулы ПНК при приготовлении лекарственного средства для лечения или профилактики заболевания, нарушения или инфекции у пациента посредством изменения экспрессии одного или нескольких генов-мишеней у указанного пациента, где лекарственное средство предназначено для использования в способе, который содержит введение молекулы ПНК в цитозоль или ядро клетки, предусматривающем контактирование указанной клетки с молекулой ПНК и фотосенсибилизирующим агентом и облучение клетки светом длиной волны, эффективной для активации фотосенсибилизирующего агента, где указанная молекула ПНК конъюгирована с положительно заряженным пептидом и, где предпочтительно указанная молекула ПНК составляет в длину менее 25 оснований, и/или указанная молекула ПНК представляет собой антисмысловую молекулу, комплементарную гену молекулу или зонд, и/или указанный положительно заряженный пептид представляет собой полимер последовательных аминокислот, и/или указанный положительно заряженный пептид составляет в длину от 3 до 30 аминокислот, и/или указанный положительно заряженный пептид конъюгирован непосредственно с молекулой ПНК посредством ковалентного связывания, и/или указанный положительно заряженный пептид имеет заряд от +1 до +10, предпочтительно от +3 до +6 и/или указанный положительно заряженный пептид содержит последовательность Xn-(Y)m-Xo, где Х представляет собой нейтральный остаток и Y представляет собой положительно заряженный остаток, который может быть одинаковым или разным в каждом положении, и n, m и о представляют собой целые числа ≥1, и/или Y одинаков в каждом положении и представляет собой К, R или Н, и/или указанный положительно заряженный пептид представляет собой SEQ ID NO: 7 MSVLTPLLLRGLTGSARRLPVPRAKIHSL, SEQ ID NO: 6 AKL или SEQ ID NO: 5 GHHHHHG.
31. Применение клетки или популяции клеток по п.26 при производстве композиции или лекарственного средства для лечения или профилактики заболевания, нарушения или инфекции посредством изменения экспрессии одного или нескольких генов-мишеней у указанного пациента.
32. Применение по п.30 или 31, в котором указанное лекарственное средство представляет собой средство для генотерапии или терапии злокачественной опухоли.
33. Способ лечения или профилактики заболевания, нарушения или инфекции у пациента, предусматривающий введение молекулы ПНК в одну или несколько клеток in vitro, in vivo или ex vivo согласно способу по любому из пп.1-20, и в котором необходимо введение указанных клеток указанному пациенту.
34. Способ по п.33, в котором указанный способ используют для лечения злокачественной опухоли или используют при генотерапии.
35. Применение клетки или популяции клетки по п.26 в качестве средства для скрининга.
36. Способ in vitro скрининга клеток с измененными паттернами экспрессии гена, предусматривающий:
а) анализ экспрессии гена-мишени, или одного, или нескольких дополнительных генов клетки, определенной в п.26, где указанная ПНК специфически связывается с указанным геном-мишенью или продуктом его репликации или транскрипции и изменяет экспрессию указанного одного или нескольких дополнительных генов; и
b) сравнение экспрессии указанного гена-мишени и/или одного или нескольких дополнительных генов с экспрессией указанных генов в контрольных клетках, предпочтительно клетках дикого типа.
37. Способ по п.36, в котором экспрессия указанного гена-мишени снижается менее чем на 80% от контрольных (дикого типа) уровней.
| ИНТЕГРАЛЬНЫЙ БЛОК ТРАНСФОРМАТОРОВ | 0 |
|
SU244396A1 |
| RU 2002109476, 13.10.2000 | |||
| Folini M et al | |||
| Photochemical Intemalization of a Peptide Nucleic Acid Targeting the Catalytic Subunit of Human Telomerase | |||
| Способ приготовления сернистого красителя защитного цвета | 1915 |
|
SU63A1 |

