Настоящее изобретение относится к области биотехнологии и фармацевтической промышленности, в частности к активной иммунизации, направленно воздействующей на молекулы, определяющие развитие кровеносных сосудов.
Процесс образования новых кровеносных сосудов из ранее существовавших называется ангиогенезом. Данное явление в широких пределах регулируется равновесием про- и антиангиогенных факторов. Болезни, обусловленные индукцией проангиогенных факторов и образованием новых кровеносных сосудов в аномальной форме, включают в себя: (а) рак (как первичные опухоли, так и их метастазы), (b) острые и хронические воспалительные заболевания, такие как астма, респираторный дистресс-синдром, эндометриоз, атеросклероз и отек тканей, (с) инфекционные заболевания, такие как гепатит и саркома Капоши, (d) аутоиммунные заболевания, такие как диабет, псориаз, ревматоидный артрит, тиреоидит, и (е) другие заболевания и нарушения, такие как диабетическая ретинопатия и ретинопатия новорожденных, отторжение трансплантированного органа, дегенерация желтого пятна, неоваскулярная глаукома, гемангиома и ангиофиброма (Carmelliet P. y Jain R.K. Nature 407:249, 2000; Kuwano M. et al., Intern. Med. 40:565, 2001). Потенциально привлекательным методом лечения многих из указанных заболеваний является ингибирование активности проангиогенных факторов, стимулирующих аномальное образование кровеносных сосудов, путем нейтрализации указанных факторов или их рецепторов либо устранения источников продуцирования таких факторов.
Эндотелиальные факторы роста сосудов образуют семейство молекул, которые индуцируют специфическое и прямое образование новых сосудов (Leung Science 246:1306, 1989; Klagsburn M., Annual Rev. Physiol. 33:217, 1991). Данное семейство включает фактор проницаемости сосудов, известный также как эндотелиальный фактор роста сосудов VPF/VEGF (в настоящее время получивший название VEGF-A), фактор роста плаценты PlGF, тромбоцитарные факторы роста PDGF-A и PDGF-B и четыре другие новые молекулы, структурно и функционально связанные с VEGF-A, получившие название VEGF-B/VRF, VEGF-C/VRP, VEGD-D/FlGF и VEGF-E. (Olofsson B. et al., PNAS USA 13:2576, 1996; Joukov V. et al., EMBO J. 15:290, 1996; Yamada Y. et al., Genomics 42:483, 1997; Ogawa S. et al., J. Biol. Chem. 273:31273, 1998).
VEGF-A является гомодимерным гликопротеином, образуемым двумя субъединицами длиной 23 кДа (Ferrara N. et al., Biochem. Biophys. Res. Comun. 165:198, 1989), из которого в результате дифференциального сплайсинга РНК можно выделить пять мономерных изоформ. Указанные изоформы включают две изоформы, которые остаются связанными с клеточной мембраной (VEGF 189 и VEGF 206), и три растворимые изоформы (VEGF 121, VEGF 145 и VEGF 165). VEGF 165 является изоформой, наиболее распространенной в тканях млекопитающих за исключением легкого и сердца, где преобладает изоформа VEGF 189 (Neufeld G. et al., Canc. Met. Rev. 15:153, 1995), и плаценты, где преобладает экспрессия VEGF 121 (Shibuya M.A. et al., Adv. Canc. Res. 67:281, 1995).
VEGF-A является наиболее изученным белком данного семейства и его изменение описано при исследовании большого числа заболеваний. Сверхэкспрессия указанного фактора характерна для опухолей разного происхождения и локализации и их метастазов (Grunstein J. et al., Cancer Res. 59:1592, 1999), хронических воспалительных заболеваний, таких как неспецифический язвенный колит и болезнь Крона (Kanazawa S. et al., Am. J. Gastroenterol. 96:822, 2001), псориаз (Detmar M. et al., J. Exp. Med. 180:1141, 1994), респираторный дистресс-синдром (Thickett D.R. et al., Am. J. Respir. Crit. Care Med. 164:1601, 2001), атеросклероз (Celletti F.L. et al., Nat. Med, 7:425, 2001; Couffinhal T. et al., Am. J. Pathol. 150: 1653, 1997), эндометриоз (McLaren J. Hum. Reprod. Update 6:45, 2000), астма (Hoshino M. et al., J. Allergy Clin. Immunol. 107:295, 2001), ревматоидный артрит и остеоартрит (Pufe T. et al., J. Rheumatol 28:1482, 2001), тиреоидит (Nagura S. et al., Hum Pathol. 32:10, 2001), диабетическая ретинопатия и ретинопатия новорожденных (Murata T. et al., Lab. Invest. 74:819, 1996; Reynolds J.D., Paediatr Drugs 3:263, 2001), дегенерация желтого пятна и глаукома (Wells J.A. et al., Br. J. Ophthalmol. 80:363, 1996; Tripathi R.C. et al., Ophthalmology 105:232, 1998), отек тканей (Kaner R.J. et al., Am. J. Respir. Cell Mol. Biol. 22:640, 2000; Ferrara N. Endocrinol. Rev. 13:18, 1992), ожирение (Tonello C. et al., FEBS Lett. 442: 167, 1999), гемангиома (Wizigmann S. y Plate K.H. Histol. Histopathol., 11:1049, 1996), а также для синовиальной жидкости субъектов, страдающих артритом (Bottomley M.J. et al., Clin. Exp. Immunol 119:182, 2000), и отторжения трансплантата (Vasir B. et al., Transplantation 71:924, 2001). В случае опухолей клетки экспрессирующие три основные изоформы VEGF-A: 121, 165 и 189 растут быстрее in vivo; причем на последних стадиях развития большинства опухолей экспрессия указанных факторов ограничивается изоформой VEGF 165 и при ее отсутствии комбинацией изоформ 121 и 189, которая, не являясь аддитивной, свидетельствует о кооперации, усиливающей развитие сосудистой сети опухоли (Grunstein J. Mol. Cell Biol. 20:7282, 2000).
Фактор PlGF, описанный в 1991 г., не способен индуцировать пролиферацию эндотелия в гомодимерной форме (Maglione D. et al., Proc. Natl. Acad. Sci. USA 88:9267, 1991, DiSalvo J. et al., J. Biol. Chem. 270: 7717, 1995). В результате увеличения концентрации PlGF и, как следствие этого, усиления сигнала, передаваемого VEGFR1, повышается реакция эндотелиальных клеток на VEGF в процессе изменения ангиогенного фенотипа, обусловленного определенными патологиями (Carmeliet P. et al., Nat. Med. 7:575, 2001). Экспрессия PlGF вызывает васкуляризацию менингиомы и глиомы человека (Nomura M. et al., J. Neurooncol. 40:123, 1998). Данная молекула образует гетеродимеры с VEGF 165, обладающие проангиогенной активностью, и сверхэкспрессия указанных факторов, обнаруженная в кондиционированных средах некоторых линий опухолевых клеток (Cao Y. et al., J. Biol. Chem. 271:3154, 1996), вызывает развитие ревматоидного артрита и первичного артрита (Bottomley M.J. et al., Clin. Exp. Immunol. 119:182, 2000).
Сверхэкспрессия остальных членов семейства VEGF, хотя и является менее изученной, также связана с рядом патологий. VEGF-B имеет отношение к опухолям молочной железы, яичника и почки, меланомам и фибросаркомам (Sowter H.M. et al., Lab. Invest. 77:607, 1997; Salven P. Am. J. Pathol. 153:103, 1998, Gunningham S.P. et al., Cancer Res. 61:3206, 2001). Дифференцированная экспрессия изоформы VEGF-B 167 in vitro была обнаружена в опухолевых клетках разного происхождения (Li X. et al., Growth Factors 19:49, 2001). VEGF-C и VEGF-D участвуют в регуляции образования лимфатических сосудов (Joukov V. et al., EMBO J. 15: 290, 1996), и сверхэкспрессия VEGF-C связана с отеком тканей, опухолями молочной железы, легкого, головы и шеи, пищевода и желудка, лимфомой, раком предстательной железы и метастатическими узлами (Kajita T. et al., Br. J. Cancer 85:255, 2001; Kitadi Y. et al., Int. J. Cancer 93:662, 2001; Hashimoto I. et al., Br. J. Cancer 85:93, 2001; Kinoshita J. et al., Breast Cancer Res. Treat 66:159, 2001; Ueda M. et al., Gynecol. Oncol. 82:162, 2001; Salven P. Am. J. Pathol. 153:103, 1998; O-Charoenrat P. et al., Cancer 92:556, 2001). Сверхэкспрессия VEGF-D в опухолевых клетках вызывает увеличение in vivo сосудистой сети лимфов в опухолях и образование метастазов в лимфатических узлах (Stacker S.A. et al., Nat. Med. 7:186, 2001; Marconcini L. et al., Proc. Natl. Acad. Sci. USA 96:9671, 1999).
Изменения функции эндотелиальных клеток под воздействием молекул семейства VEGF опосредованы связыванием указанных молекул с рецепторами клеток класса тирозинкиназы 3, которые включают: VEGFR1 (Flt1), VEGFR2 (KDR/Flk1) и VEGFR3 (Flt4) (Kaipainen A. J. Exp. Med. 178:2077, 1993). Установлено, что N-концевой домен 2 является ответственным за связывание с лигандами, стимуляцию фосфорилирования цитоплазматического домена и передачу сигнала (Davis-Smyth T. et al., EMBO 15:4919, 1996).
Лиганды, идентифицированные для VEGFR1, включают в порядке убывания сродства VEGF-A, PlGF и VEGF-B (Shibuya M. Int. J. Biochem. Cell Biol. 33:409, 2001). В эндотелиальных клетках данный рецептор улавливает циркулирующий фактор VEGF (Gille H. et al., EMBO J. 19: 4064, 2000). Связывание VEGF-A с VEGFR1, экспрессированным в клетках гемопоэтической линии, значительно активирует фактор транскрипции NFκB в клетках-предшественниках дендритных клеток и В- и Т-лимфоцитах. Указанное последним взаимодействие определяет in vivo возникновение неблагоприятного иммунологического баланса, в результате которого сокращается созревание дендритных клеток и фракции Т-лимфоцитов, причем указанное явление характерно для субъектов с иммуносупрессией и, в частности, больных раком (Dikov M.M. et al., Canc. Res. 61:2015, 2001; Gabrilovich D. et al., Blood 92:4150, 1998). Сверхэкспрессия VEGFR1 связана с псориазом, раком эндометрия и злокачественной гепатомой (Detmar M. et al., J. Exp. Med. 180:1141, 1994; Ng IO Am. J. Clin. Patol. 116:838, 2001; Yokoyama Y. et al., Gynecol. Oncol. 77: 413, 2000).
Рецептор VEGFR2 (KDR/Flk1) опосредует биологическое действие VEGF-A, а также связывает VEGF-C и VEGF-D. Данный рецептор дифференцированно экспрессирован в активированном эндотелии и в некоторых линиях клеток опухолевого происхождения, где он создает аутокринные пути с секретированным фактором VEGF. Независимо от участия в вышеуказанных патологиях, которые обусловлены сверхэкспрессией его лигандов, сверхэкспрессия VEGFR2 имеет непосредственное отношение к развитию рака эндометрия (Giatromanolaki A. et al., Cancer, 92:2569, 2001), злокачественной мезотелиомы (Strizzi L. et al., J. Pathol. 193:468, 2001), астроцитомы (Carroll R.S. et al., Cancer 86:1335, 1999), первичного рака молочной железы (Kranz A et al., Int. J. Cancer 84:293, 1999), рака желудка кишечного типа (Takahashi Y. et al., Clin. Cancer Res. 2:1679, 1996), мультиформной глиобластомы, анапластической олигодендроглиомы и некротической эпендимомы (Chan A.S. et al., Am. J. Surg. Pathol. 22:816, 1998). Сверхэкспрессия KDR также связана с аутосомным заболеванием VHL и гемангиобластомой (Wizigmann-Voos S. et al., Cancer Res. 55:1358, 1995), развитием диабетической ретинопатии (Ishibashi T. Jpn. J. Ophthalmol. 44:324, 2000) и в сочетании со сверхэкспрессией Flt-1 имеет отношение к аллергическим реакциям замедленного типа (Brown L.F. et al., J. Immunol. 154:2801, 1995).
Лимфангиогенез, опосредованный VEGF-C и VEGF-D, возникает в результате связывания указанных факторов с рецептором FLT4 или VEGFR3, экспрессированным в эндотелии лимфатических сосудов. В некоторых случаях, даже при отсутствии сверхэкспрессии указанных лигандов, сверхэкспрессия рецептора имеет непосредственное отношение к неблагоприятному прогнозу в отношении группы патологий, включающих диабетическую ретинопатию (Smith G. Br. J. Ophthalmol. 1999 Apr; 83(4):486-94), хронические воспаления и язвы (Paavonen K. et al., Am. J. Pathol. 156:1499, 2000), образование метастазов в лимфатических узлах и развитие рака молочной железы (Gunningham S.P., Clin. Cancer Res. 6: 4278, 2000; Valtola R. et al., Am. J. Pathol. 154:1381, 1999), опухоли носоглотки и плоскоклеточный рак (Saaristo A. et al., Am. J. Pathol, 157:7, 2000; Moriyama M. et al., Oral Oncol, 33:369, 1997). Кроме того, сверхэкспрессия VEGFR3 является четко выраженным показателем саркомы Капоши, гемангиоэндотелиомы Дабска и кожного лимфангиоматоза (Folpe A.L. et al., Mod. Pathol. 13:180, 2000; Lymboussaki A. et al., Am. J. Pathol. 153:395, 1998).
В настоящее время идентифицированы два рецептора для VEGF, получившие название NRP1 и NRP2. Указанные рецепторы относятся к семейству нейрофилинов (NRP) и действуют в качестве корецепторов для некоторых специфических изоформ белков семейства VEGF: VEGF-A145, VEGF-A165, VEGF-B167 и PlGF1, усиливая их митогенную способность. Экспрессия NRP1 является показателем развития рака предстательной железы, имеет отношение к увеличению ангиогенеза в меланомах и апоптозу в случае рака молочной железы (Latil A. et al., Int. J. Cancer 89:167, 2000; Lacal P.M. J. Invest. Dermatol. 115:1000, 2000; Bachelder R.E. Cancer Res. 61:5736, 2001). Скоординированная сверхэкспрессия NRP1, KDR и VEGF-A165 связана с пролиферацией сосудов фиброзной ткани в случае диабетической ретинопатии и ревматоидного артрита (Ishida S. et al., Invest. Ophthalmol. Vis. Sci. 41:1649, 2000; Ikeda M. et al., J. Pathol. 191:426, 2000). Рецептор NRP2 сверхэкспрессирован в остеосаркомах, где он стимулирует ангиогенез и рост опухоли (Handa A. et al., Int. J. Oncol. 17:291, 2000).
В основе большинства методов лечения, направленных на ингибирование ангиогенеза, особенно в случае рака, лежит блокирование молекул семейства VEGF и их рецепторов, причем для достижения указанной цели в клинических испытаниях используют: (1) моноклональные антитела, блокирующие VEGF или рецептор KDR, (2) ингибиторы металлопротеиназы, такие как неовастат и приномастат, (3) ингибиторы VEGF, такие как талидомид, сурамин, тропонин I, IFN-α и неовастат, (4) блокаторы рецепторов VEGF, такие как SU5416, FTK787 и SU6668, (5) индукторы апоптоза эндотелия опухоли, такие как эндостатин и СА4-Р, и (6) рибозимы, уменьшающие экспрессию VEGF или рецепторов VEGF (ангиозим). Благодаря высокой гомологии, существующей между VEGF человека, его рецепторами KDR и Flt-1 и гомологами мыши (соответственно ˜90%, 81% и 89%), обычно используют разные животные модели для оценки предклинической эффективности антиангиогенных соединений, воздействующих на указанную систему (Hicklin D.J. et al., DDT 6: 517, 2001).
Пассивное введение антител к VEGF или VEGFR успешно исследовано на разных стадиях клинических испытаний с участием людей (Hicklin D.J. et al., DDT 6:517, 2001). Гуманизированное моноклональное антитело А.4.6.1 против VEGF (Genentech, San Francisco, United States) проходит стадию III клинического испытания по лечению опухолей ободочной кишки, молочной железы, почки и легкого (Kim K.J. et al., Nature 362:841, 1993; Boersig C. R&D Directions, Oct. 7:44, 2001). В частности, было создано моноклональное антитело к рецептору KDR (IMC-1C11, ImClone), которое узнает N-концевой внеклеточный домен данного рецептора и ингибирует пролиферацию и миграцию лейкозных клеток человека, повышая выживаемость ксенотрансплантированных мышей. В настоящее время воздействие данного антитела исследуют у субъектов с метастазами рака ободочной кишки (Dias S. et al., J. Clin. Invest. 106:511, 2000). В вышеуказанных испытаниях было продемонстрировано отсутствие сопутствующих вредных эффектов при введении рассмотренных моноклональных антител.
Новым методом лечения, ранее не применявшимся для блокирования неоангиогенеза, является специфическая активная иммунотерапия (SAI). При лечении рака методом SAI используют антигены, такие как пептиды, белки или ДНК, в сочетании с приемлемыми адъювантами. Методы SAI направлены на стимуляцию иммунной реакции гуморального типа (активация В-лимфоцитов) и клеточного типа (активация Т-клеток-хелперов, цитотоксических лимфоцитов и естественных клеток-киллеров) в дендритных клетках, которые являются профессиональными антигенпредставляющими клетками в МСН I и МНС II (Bystryn J.C., Medscape Hematology-Oncology 4:1, 2001; Parker K.C. et al., J. Immunol 152:163, 1994; Nestle F.O. et al., Nature Medicine 7:761, 2001; Timmerman J.M., Annual Review Medicine 50:507, 1999).
Специфическая активная иммунотерапия является быстроразвивающейся областью экспериментальных и клинических исследований с интересными возможностями применения в онкологии, где было выполнено более 60 клинических испытаний по методу SAI, причем указанное число в настоящее время превышает количество клинических испытаний с применением моноклональных антител. В случае рака антигены, используемые в качестве иммуногенов для SAI, отбирают с учетом их физиологической пригодности, сложности замены в процессе фенотипического дрейфа опухоли (Bodey B. et al., Anticancer Research 20:2665, 2000) и специфической связи с ростом и развитием опухолевых тканей.
Кроме того, при лечении рака методом SAI предпочтительно идентифицируют антигены, экспрессированные в опухолях разных типов, что может увеличить число показаний к применению одного вакцинного препарата. Примерами таких антигенов являются карциноэмбриональный антиген (СЕА), HER2-neu, теломераза человека и ганглиозиды (Greener M., Mol. Med. Today 6:257, 2000; Rice J. et al., J. Immunol. 167:1558, 2001; Carr A. et al., Melanoma Res. 11:219, 2001; Murray J.L. et al., Semin. Oncol. 27:71, 2000).
В случае опухолей человека VEGF сверхэкспрессирован в опухолевом компартменте (Ferrara N., Curr. Top. Microbiol. Immunol. 237:1, 1999), при этом высокие уровни VEGF и его рецепторов обнаружены в сосудистой сети опухоли (Brekken R.A., J. Control Release 74:173, 2001). Стромальные клетки также продуцируют VEGF под воздействием трансформированных клеток, в результате чего даже после удаления опухолевых клеток у субъектов сохраняются высокие уровни VEGF. Наличие VEGF и его рецепторов имеет практическое значение для прогнозирования и определения стадии развития опухолей предстательной железы, шейки матки и молочной железы (George D.J. et al., Clin. Cancer Res. 7:1932, 2001; Dobbs S.P. et al., Br. J. Cancer 76:1410, 1997; Callagy G. et al., Appl. Immunohistochem. Mol. Morphol. 8:104, 2000). С другой стороны, VEGF входит также в группу растворимых факторов, которые вместе с другими цитокинами, такими как IL-10, TNF-α и TGF-β (Ohm J.E. y Carbone D.P., Immunol. Res. 23:263, 2001), могут участвовать в иммуносупрессии, характерной для больных раком (Staveley K. et al., Proc. Natl. Acad. Sci. USA 95:1178, 1998; Lee K.H. et al., J. Immunol. 161:4183, 1998). Иммуносупрессивное действие, по-видимому, обусловлено связыванием с рецептором Flt1 (Gabrilovich D. et al., Blood 92:4150, 1998).
В настоящем изобретении описано применение SAI для лечения экспериментальных опухолей, вызванных при помощи молекул семейства VEGF и их рецепторов. Достигнутое противоопухолевое действие может быть обусловлено по меньшей мере четырьмя разными механизмами и их возможными комбинациями: (а) прямое разрушение раковых и стромальных клеток, продуцирующих VEGF, цитотоксическими лимфоцитами, (b) лизис эндотелиальных клеток сосудов опухоли вследствие улавливания или нейтрализации циркулирующего VEGF при помощи антител, (с) прямое разрушение эндотелиальных клеток, экспрессирующих рецепторы VEGF, цитотоксическими лимфоцитами или комплементсвязывающими антителами, (d) активация локальной иммунной реакции вследствие улавливания или нейтрализации циркулирующего VEGF и последующего устранения его иммуносупрессивного действия.
Указанные методы лечения можно использовать для предотвращения образования метастазов или снижения вероятности их появления, для уменьшения или устранения первичных опухолей в результате терапии первого или второго порядка в сочетании с другими противоопухолевыми средствами или без использования таких средств.
Активная иммунизация, направленная на подавление семейства VEGF и их рецепторов, может быть также эффективной при осуществлении монотерапии или комбинированной терапии острых и хронических воспалительных заболеваний (астма, респираторный дистресс-синдром, эндометриоз, атеросклероз, отек тканей), инфекционных заболеваний (гепатит, саркома Капоши), аутоиммунных заболеваний (диабет, псориаз, ревматоидный артрит, тиреоидит, синовит), диабетической ретинопатии и ретинопатии новорожденных, отторжения трансплантированного органа, дегенерации желтого пятна, неоваскулярной глаукомы, гемангиомы, ангиофибромы и других заболеваний.
Подробное описание изобретения
В соответствии с настоящим изобретением введение in vivo олигонуклеотидных последовательностей, кодирующих белки семейства VEGF, их рецепторы, корецепторы или фрагменты, и их полипептидных вариантов индуцирует клеточную и гуморальную иммунную реакцию с достижением антиангиогенного и противоопухолевого действия.
Иммуногены полипептидного происхождения, представляющие интерес для настоящего изобретения, и их фрагменты можно выделить из природных источников или получить путем синтеза или методом рекомбинантной ДНК. Указанные полипептиды можно также получить путем слияния с белками, обладающими известной адъювантной активностью, такими как р64K (R. Silva et al., US 5286484 y EP 0474313), или путем ковалентного связывания с предварительно полученными белками. Другим приемлемым способом является получение природного полипептида, его мутированных или модифицированных вариантов и их фрагментов в виде части петель, существующих или отсутствующих в бактериальных белках, таких как ОМР1, которые являются частью иммуностимулирующих препаратов, в данном конкретном случае VSSP (R. Perez et al., US 5788985 y 6149921). Кроме того, можно получить полипептидный иммуноген, экспрессированный на поверхности вирусной частицы (HbsAg, VP2 парвовируса и т.д.) и связанный со специфическими пептидами, которые направленно воздействуют на клетки или органы, индуцирующие иммунную реакцию (CTLA4, Fc-сегмент Ig и т.д.), или с белками, способными увеличивать биологическое распределение подобно VP22.
Основные природные источники белков, представляющих интерес для настоящего изобретения, экспрессированы, главным образом, в плаценте, активированных эндотелиальных клетках и опухолевых клетках. мРНК указанных клеток или тканей используют для получения комплементарной ДНК (кДНК) известными методами. Экстрагированную РНК используют в качестве матрицы для амплификации при помощи полимеразной реакции синтеза цепи (PCR) кДНК, соответствующей выбранному антигену. В каждом случае используемые затравки соответствуют характеристикам вектора, в который предполагается ввести данную кДНК, и ранее известным последовательностям представляющего интерес белка. Альтернативно и предпочтительно, в случае рецепторов, амплифицированных PCR, которые являются самыми крупными антигенами, используемыми в настоящем изобретении, кодирующие области амплифицируют в двух или более перекрывающихся фрагментах. Указанные фрагменты включают общий сайт лигирования, используемый для сборки интактной ДНК, начиная с ее фрагментов.
Альтернативой клонирования представляющих интерес антигенов является выбор из коммерчески доступных библиотек ДНК, полученных из эндотелия человека или опухолей того же происхождения. Иногда может быть желательно мутировать некоторые антигены, являющиеся объектом настоящего изобретения, чтобы избежать индукции ангиогенеза, вызванного вакцинацией, особенно в случае семейства VEGF. Указанные мутации предпочтительно создают на сайтах связывания рецепторов, ранее описанных в научной литературе. Для достижения указанной цели создают затравки, которые охватывают оба конца требуемой молекулы, и продукты PCR используют в качестве матрицы для получения мутированной молекулы. Данные мутированные варианты не обладают биологической активностью, но воспроизводят иммуногенные свойства выбранного антигена.
Молекулы кДНК, полученные описанными выше способами, вводят в соответствующий вектор, представляющий собой вирус, плазмиду, бактериальную искусственную хромосому или тому подобное. Данный вектор несет элементы, необходимые для соответствующей экспрессии гена в клетках-мишенях, а также остальные элементы, необходимые для продуцирования вектора в клеточной системе хозяина в зависимости от его природы. Молекулы ДНК согласно изобретению могут содержать один или несколько представляющих интерес генов, образуемых одной или несколькими нуклеиновыми кислотами (кДНК, гДНК, синтетическая или полусинтетическая ДНК и тому подобное), которые в результате транскрипции или трансляции (в зависимости от необходимости) в клетках-мишенях образуют продукты, обладающие терапевтической/вакцинной ценностью.
Ген лечебного вакцинного продукта согласно изобретению обычно контролируется промотором транскрипции, который функционирует в клетке-мишени или организме (млекопитающих), а также 3'-концевой областью, которая формирует сигналы, необходимые для терминации и полиаденилирования мРНК в представляющем интерес продукте, экспрессирующем данный ген. Промотор может быть естественным промотором гена или гетерологичным промотором, который является транскрипционно-активным в клетке-мишени. Промотор может быть получен из эукариотических или вирусных клеток. Можно использовать любой эукариотический промотор или выделенную последовательность, которая стимулирует или подавляет транскрипцию гена, оказывая специфическое или неспецифическое, индуцируемое или неиндуцируемое, сильное или слабое действие. Кроме того, промоторную область можно модифицировать путем введения активаторных или индукторных последовательностей, обеспечивающих тканеспецифическую или предпочтительную экспрессию рассматриваемого гена.
Кроме того, представляющий интерес ген может содержать сигнальную последовательность, необходимую для субклеточной локализации, благодаря которой можно модифицировать локализацию или секрецию синтезированного гена в клетке, где он экспрессирован, или в любом другом месте. Указанный ген может также содержать последовательность, кодирующую область специфического связывания с лигандом, характерным для иммунной ткани, которая непосредственно связана с сайтом возникновения реакции, обеспечивая достижение терапевтического/вакцинного действия.
Кроме того, представляющему интерес гену может предшествовать кодирующая последовательность, необходимая для репликации мРНК, в результате которой происходит амплификация мРНК в клетке-мишени, усиливающая экспрессию указанного гена и вместе с тем терапевтическое/вакцинное действие согласно изобретению. Рассматриваемый механизм репликации может принадлежать ориджину альфа-вируса (Schlesinger S., Expert Opin. Biol. Ther. 1:177, 2001), в частности, он может быть выделен из вирусов Sindbis, или Semliki, или подобных. В данном конкретном случае транскрипцию представляющего интерес гена контролирует субгеномный промотор, который обеспечивает амплификацию мРНК в клетках-мишенях после интернализации молекул согласно изобретению. Кроме того, вектор на основе ДНК может содержать последовательности, делающие возможной репликацию молекул, являющихся объектом настоящего изобретения, в клетках млекопитающих. Это позволяет увеличить уровни экспрессии и/или терапевтическое/вакцинное действие (Collings A., Vaccine 18:4601, 1999).
Вектор на основе ДНК может быть очищен стандартными методами, применяемыми для очистки плазмидной ДНК. Указанные методы включают очистку в градиенте плотности хлорида цезия или в присутствии бромида этидия, альтернативно применение ионообменных колонок или любого другого ионообменника или отделение молекул ДНК (Ferreira G.N. et al., Trends Biotechnol. 18:380, 2000).
Настоящее изобретение относится к применению векторов на основе плазмидной ДНК, предпочтительно относящихся к семейству РАЕС компактных векторов для ДНК-иммунизации и генотерапии человека (Herrera et al., Biochem. Biophys. Res. Commu. 279:548, 2000). Данное семейство включает векторы рАЕС-К6 (номер доступа AJ278712), рАЕС-М7 (номер доступа AJ278713), рАЕС-Δ2 (номер доступа AJ278714), рАЕС-SPE (номер доступа AJ278715) и рАЕС-SPT (номер доступа AJ278716). Указанные векторы содержат только основные элементы, необходимые для экспрессии представляющего интерес продукта в клетках млекопитающего, включая клетки человека, и единицу репликации Escherichia coli. Единица транскрипции образуется предранним промотором цитомегаловируса человека (CMV), универсальным сайтом мультиклонирования для вставки представляющего интерес продукта и последовательностями для терминации транскрипции и полиаденилирования, выделенными из вируса 40 обезьяны (SV40). В единице репликации вектор содержит ген устойчивости к канамицину (Tn903) и ориджин репликации pUC19 (ColE1) для гарантии получения большого числа копий и селекции бактерий, несущих представляющую интерес плазмиду.
Настоящее изобретение далее относится к применению векторов на основе плазмидной ДНК, предпочтительно относящихся к семейству РМАЕ компактных векторов для ДНК-иммунизации человека. Указанные векторы содержат такие же функциональные элементы в бактериях, что и векторы серии РАЕС, а также предранний промотор CMV и сайт мультиклонирования. Кроме того, указанные векторы имеют синтетический интрон и синтетическую последовательность для терминации транскрипции и полиаденилирования, выделенные из β-глобина кролика. Известно, что при помощи последовательностей, аналогичных вышеуказанной последовательности, можно достичь более высоких уровней экспрессии клонированного гена (Norman J.A. et al., Vaccine 15:801, 1997). Векторы указанной серии далее включают последовательные повторы иммуностимулирующих последовательностей (CpG-фрагменты), которые стимулируют природную иммунную систему у мышей и человека с последующей активацией гуморальной и клеточной реакций против представляющей интерес молекулы (Krieg A.M., Vaccine 19:618, 2001).
Иммунизация рекомбинантным вирусом (аденовирус, аденоассоциированный вирус, вирус коровьей оспы, поксвирус цыплят, поксвирус канареек и подобные) вызывает сильную цитотоксическую реакцию клеток в хозяине. Для введения представляющей интерес последовательности в рекомбинантный вирус используют векторы, содержащие интегрирующие последовательности и промоторы, специально предназначенные для каждого типа вирусов. Данный способ также входит в объем настоящего изобретения, причем предпочтительно используют поксвирус цыплят и вектор pFP67xgpt. Вектор pFP67xgpt используют для клонирования генов под контролем сильного раннего/позднего синтетического промотора между открытыми рамками считывания 6 и 7 фрагмента BamHI длиной 11,2 т.п.н. поксвируса цыплят FP9. Указанная плазмида также содержит Ecogpt, контролируемый промотором вируса коровьей оспы р7.5К, который используют для идентификации рекомбинантного вируса.
Другой вариант осуществления настоящего изобретения относится к иммунизации белками семейства VEGF и их рецепторов и/или корецепторов. Молекулы кДНК, полученные вышеописанным способом, клонируют в векторах для экспрессии в вирусах, дрожжах, фаге, растениях или клетках высших организмов для получения белковых вариантов антигенов после того, как их последовательность была проверена традиционными методами автоматического секвенирования. Было описано несколько экспрессирующих векторов, предназначенных для получения рекомбинантных белков. Указанные векторы содержат по меньшей мере одну последовательность, которая контролирует экспрессию, функционально связанную с последовательностью ДНК или экспрессируемого фрагмента. Примеры последовательностей, пригодных для контроля экспрессии, включают системы lac, trp, tac и trc, промоторные области и главный оператор лямбда-фага, контролирующую область поверхностного белка fd, гликолитические промоторы дрожжей (например, 3-фосфоглицераткиназа), промоторы кислой фосфатазы дрожжей (например, Pho5), дрожжевые промоторы спаривающего альфа-фактора и промоторы, выделенные из полиомы, аденовируса, ретровируса, вируса обезьян (например, ранний/поздний промоторы SV40) и другие известные последовательности, которые регулируют экспрессию генов в прокариотических и эукариотических клетках, вирусах и их комбинациях.
Хозяева, используемые для репликации указанных векторов и получения рекомбинантных белков, являющихся объектом настоящего изобретения, включают прокариотические и эукариотические клетки. Прокариотические клетки включают E. coli (DHI, MRCI, HB101, W3110, SG-936, X1776, X2282, DH5a), Pseudomonas, Bacillus subtilis, Streptomices и другие. Эукариотические клетки включают дрожжи и грибы, клетки насекомых, животные клетки (например, COS-7 и СНО), клетки человека, растительные клетки, тканевые культуры и тому подобное. После экспрессии в выбранной системе и соответствующей среде полипептиды или пептиды могут быть выделены известными способами.
Применение адъювантов
Даже если установлено, что вакцинация депротеинизированной ДНК или белками является эффективной в некоторых животных моделях, неизвестно, каким образом будут реагировать на способ лечения согласно изобретению субъекты, имеющие опухоли или аутоиммунные заболевания. Для стимуляции иммунной реакции ДНК-содержащие или белковые вакцины могут быть объединены с ранее описанными иммунопотенциаторами, такими как минеральные соли (например, гидроксид алюминия, фосфат алюминия, фосфат кальция), иммуностимуляторами, такими как цитокины (например, IL-2, IL-12, GM-CSF, IFN-α, IFN-γ, IL-18), молекулами (например, CD40, CD154, инвариантная цепь МНС типа I, LFA3), сапонинами (например, QS21), производными MDP, олигонуклеотидами CpG, LPS, MPL и полифосфазенами, липидными частицами, такими как эмульсии (например, адъювант Фрейнда, SAF, MF59), липосомами, виросомами, искомами, кохелаторами, адъювантами в виде микрочастиц, такими как микрочастицы PLG, полоксамеры вирусного типа (например, HBcAg, HCcAg, HBsAg) и бактериального типа (то есть VSSP, OPC) и слизистыми адъювантами, такими как термолабильный энтеротоксин (LT), холерный энтеротоксин и мутантные токсины (например, LTK63 и LTR72), микрочастицы и полимеризованные липосомы. В случае вакцинации ДНК представляющий интерес антиген может быть экспрессирован вместе с некоторыми вышеуказанными молекулами иммунопотенциаторов в бицистронном векторе.
Эксперименты, подробно рассмотренные в примерах, показывают, что ДНК может быть нековалентно связана с некоторыми указанными частицами и что применение таких смесей уменьшает оптимальную концентрацию, необходимую для достижения противоопухолевой реакции, которая аналогична реакциям, возникающим при более высоких дозах депротеинизированной ДНК.
Введение млекопитающему
Вакцинные препараты согласно изобретению вводят в лечебных целях млекопитающему, предпочтительно человеку, в фармацевтически приемлемой дозе нижеследующими способами: через слизистую оболочку, подкожно, внутримышечно, внутрибрюшинно, в лимфу, местно и путем ингаляции. Указанные препараты могут быть введены в интерстициальное пространство ткани, включая мышцы, кожу, головной мозг, легкое, печень, костный мозг, селезенку, тимус, сердце, лимфатические узлы, кровь, кость, хрящ, поджелудочную железу, почку, мочевой пузырь, желудок, кишечник, яички, яичник, матку, прямую кишку, глаз, железы и соединительную ткань. Экспрессия векторов, предназначенных для переноса олигонуклеотидов, предпочтительно направлена в соматические дифференцированные клетки, хотя может быть направлена в недифференцированные или менее дифференцированные клетки, такие как фибробласты кожи и плюрипотентные клетки крови.
Иммуноген в требуемых дозах можно вводить в фармацевтически приемлемых носителях, не обладающих токсичностью или терапевтическим действием. Примеры таких носителей включают иониты, оксид алюминия, эстеараты алюминия, лецитин, сывороточные белки, такие как альбумин, буферные растворы, такие как фосфаты, глицин, сорбиновая кислота, сорбат калия, смеси неполного глицерида с насыщенными жирными кислотами растительного происхождения, воду, соли или электролиты, такие как сульфат протамина, динатрийгидрофосфат, хлорид натрия, соли цинка, коллоидный диоксид кремния, трисиликат магния, поливинилпирролидон, вещества на основе целлюлозы и полиэтиленгликоль. В настоящем изобретении в качестве носителей вакцинных препаратов предпочтительно используют фосфатные буферы.
Используемые белки и пептиды могут быть ковалентно или нековалентно конъюгированы с молекулами, известными в качестве носителей, действующих подобно адъювантам. Такими молекулами являются KLH, p64K, OPC (Musacchio A. et al., Vaccine 19; 3692, 2001) и VSSP. Комбинация депротеинизированной ДНК, вирусных векторов и белковых иммуногенов является альтернативой, которая также входит в объем настоящего изобретения. Применение плазмидной ДНК позволяет создать препараты, содержащие в вакцине одну или несколько представляющих интерес молекул. Таким образом, молекулы согласно изобретению можно вводить в соответствии со схемой вакцинации, используя комбинацию векторов разных типов (вариант повторной стимуляции индукции при помощи ДНК, белков, вирусных векторов).
Векторы на основе ДНК можно вводить непосредственно субъекту или указанными векторами можно модифицировать клетки-хозяева in vivo или ex vivo. Последний способ введения может быть объединен с введением при помощи сайтспецифической рекомбинации или иммунизации методом соматического трансгенеза, который направляет экспрессию вектора в определенные клетки. Кроме того, в качестве носителей для переноса векторов на основе ДНК in vivo могут быть использованы бактериальные хозяева.
Молекулы, несущие гены согласно изобретению, можно использовать в форме депротеинизированной ДНК или в комбинации с разными векторами: химическими/биохимическими/биологическими, природными/синтетическими или рекомбинантными. Указанные молекулы могут быть связаны или объединены с катионными пептидами, уплотняющими молекулами (например, PEG, PEI), ядерными локализующими пептидами (NLP) и т.д. Данные молекулы можно также вводить вместе с катионами, способными образовывать преципитаты ДНК в составе липосомных препаратов, в которые указанные молекулы добавляют до слияния с мембраной, и в синтетических векторах липидного типа или образованных катионными полимерами (например, DOGS или DOTMA). Для введения векторов на основе ДНК можно также использовать химерные белки, способные уплотнять ДНК и опосредовать перенос образованного комплекса, и селективный эндоцитоз, опосредованный определенными клетками. Молекулы ДНК, несущие терапевтические/вакцинные гены согласно изобретению, можно использовать для переноса генов в клетки при помощи физических методов переноса, таких как бомбардировка частицами, электропорация (in vitro, in vivo или ex vivo), или непосредственно in vivo путем местного применения, ингаляции тонкоизмельченного вещества и т.д. Живые векторы включают в себя аденовирусы или таких же хозяев, у которых были получены молекулы согласно изобретению.
Приемлемые дозы полипептидов и/или олигонуклеотидов могут быть определены с учетом разных параметров, в частности, в зависимости от гена или белка, вводимого в качестве иммуногена, способа введения, предназначенного для лечения заболевания, периода лечения и в случае использования олигонуклеотидов от вектора, применяемого для иммунизации. Изменение схемы применения или способа введения по сравнению с описанными в нижеследующих примерах не предполагает выхода за пределы объема настоящего изобретения, позволяя оптимизировать схему иммунизации для достижения лучшей реакции.
Терапевтическое применение
Настоящее изобретение имеет преимущество по сравнению с пассивной иммунотерапией, которая проходит последние стадии клинических испытаний с использованием таких же молекул в качестве мишеней. По сравнению с пассивным переносом иммунитета в результате введения моноклональных антител (например, антител против VEGF) иммунизация, производимая при помощи белка или олигонуклеотида, индуцирует эндогенное продуцирование антител, а также пролиферацию и размножение специфических цитотоксических CD8+-лимфоцитов.
Преимуществом настоящего изобретения по сравнению с методами лечения, направленными на блокирование системы VEGF-VEGFR, является то, что указанные методы только уменьшают уровни циркулирующего VEGF или блокируют KDR. Предложенный метод, помимо достижения вышеуказанного действия, также разрушает источник VEGF (то есть опухолевые клетки и связанную строму) и/или клетки, экспрессирующие их рецепторы (эндотелий опухоли и некоторые опухолевые клетки). В предшествующих работах, выполненных в данной области, описана только гуморальная реакция в качестве главного компонента наблюдаемого действия. Не ограничивая объем настоящего изобретения определенным механизмом, примеры показывают, что, помимо специфической гуморальной реакции, вакцинные композиции способны вызывать реакцию CD8+-клеток, которая взаимодействует с гуморальной реакцией; и в случае опухолей комбинация обеих реакций способна оказывать противоопухолевое действие, наблюдаемое в примере 9.
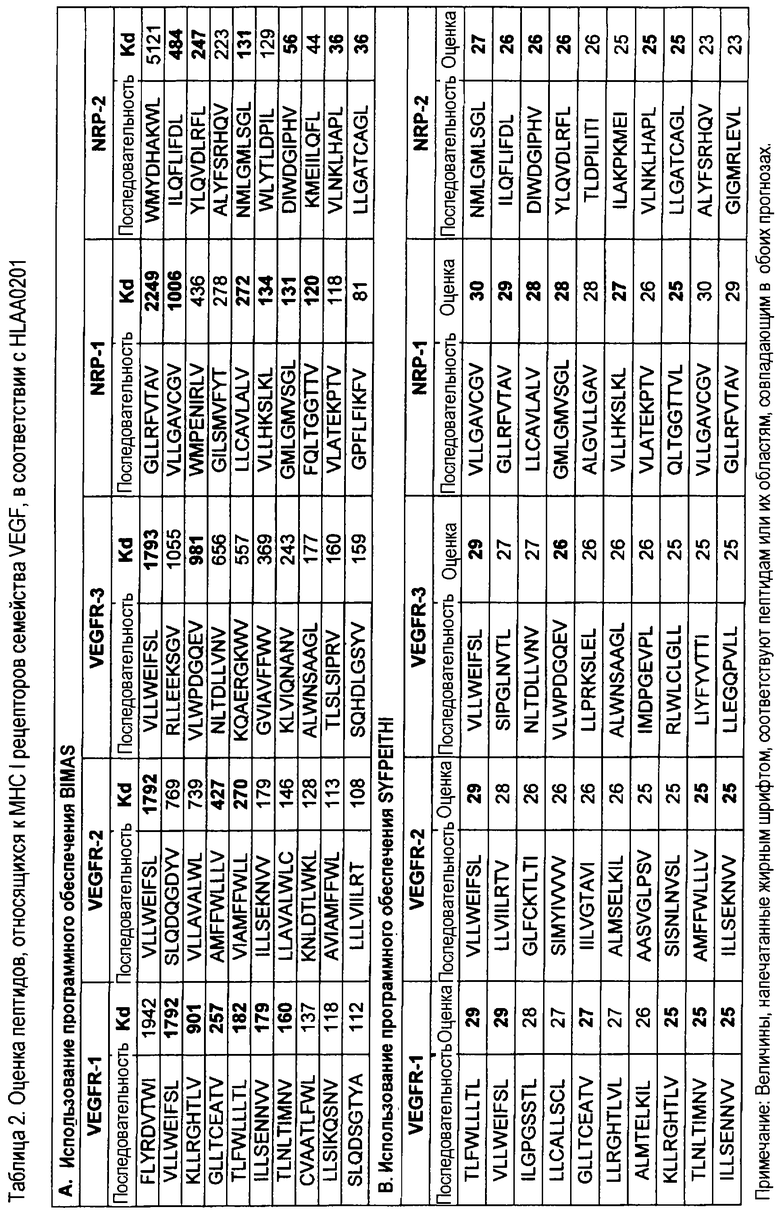
Вполне возможно, что реакция цитотоксических клеток опосредована узнаванием некоторых пептидов, приведенных в таблицах 1 и 2. В указанных таблицах представлены некоторые пептидные сегменты, которые могут иметь отношение к реакции клеток, направленной на выбранные мишени в семействе VEGF, их рецепторах и корецепторах. Данная информация была получена в результате выполнения компьютерных анализов общедоступных баз данных, находящихся в NIH и институте Хейделберга (http://bimas.dcrt.nih.gov/molbio/hla_bind и www.bmi-heidelberg.com/scripts/MHCServer.dll/home.htm), при помощи программного обеспечения BIMAS и SYFPHEITI. Отмеченные пептиды и другие последовательности, выделенные из представляющих интерес антигенов, можно использовать для активной иммунотерапии вышеуказанных заболеваний в виде монотерапии или комбинированной терапии отдельно или в составе молекул, обладающих свойствами адъювантов. Указанные пептиды можно также использовать для получения вакцины в виде олигонуклеотидных вариантов.
Способы ингибирования ангиогенеза и обусловленных им патологий включают введение млекопитающему любыми способами эффективного количества ДНК или белка некоторых молекул, описанных в данном изобретении, с использованием некоторых ранее описанных иммунопотенциаторов или адъювантов. Данное млекопитающее является предпочтительно человеком.
Необратимое и нерегулируемое развитие кровеносных сосудов характерно для целого ряда заболеваний. Система, включающая семейство факторов VEGF, их рецепторы и корецепторы, сверхэкспрессирована во многих патологиях, как было описано выше. Способы лечения согласно изобретению позволяют получить эффективные результаты при лечении таких заболеваний, как (а) рак (первичные опухоли и их метастазы), (b) острые и хронические воспалительные заболевания, такие как астма, респираторный дистресс-синдром, эндометриоз, атеросклероз и отек тканей, (с) инфекционные заболевания, такие как гепатит и саркома Капоши, (d) аутоиммунные заболевания, такие как диабет, псориаз, ревматоидный артрит, тиреоидит, и (е) другие заболевания и нарушения, такие как диабетическая ретинопатия и ретинопатия новорожденных, отторжение трансплантированного органа, дегенерация желтого пятна, неоваскулярная глаукома, гемангиома и ангиофиброма.
В случае рака вакцинация иммуногенами согласно изобретению позволяет получить эффективные результаты при лечении карцином, сарком и васкуляризованных опухолей. Некоторые примеры опухолей, которые можно лечить предложенными способами, включают эпидермоидные опухоли, плоскоклеточные опухоли, поражающие голову и шею, опухоли прямой и ободочной кишки, предстательной железы, молочной железы, легкого (включая мелкие и крупные клетки), поджелудочной железы, щитовидной железы, яичника и печени. Указанные способы также являются эффективными при лечении других типов опухолей, таких как саркома Капоши, неоплазия центральной нервной системы (нейробластома, капиллярная гемангиома, менингиома и метастазы в головном мозге), меланомы, карциномы почек и желудочно-кишечного тракта, рабдомиосаркома, глиобластома и лейомиосаркома.
В частности, применение VEGF-A и/или рецепторов VEGFR-1 и VEGFR-2 в качестве иммуногена может быть эффективным при лечении опухолей разного происхождения и локализации и их метастазов, гемангиомы, эндометриоза, отека тканей, хронических воспалительных заболеваний, таких как неспецифический язвенный колит и болезнь Крона, атеросклероза, ревматоидного артрита и остеоартрита, артрита, псориаза, респираторного дистресс-синдрома, астмы, тиреоидита, диабетической ретинопатии и ретинопатии новорожденных, дегенерации желтого пятна и глаукомы, аутосомного заболевания VHL, ожирения и отторжения некоторых трансплантированных органов. С другой стороны, реакция против PlGF полезна в случае ревматоидного артрита и для лечения первичного артрита.
Применение VEGF-B в качестве иммуногена является полезным в случае опухолей молочной железы, яичника и почек, а также меланомы и фибросаркомы. Применение VEGF-C и его рецептора VEGFR-3 позволяет получить эффективные результаты при лечении отека тканей, диабетической ретинопатии, хронического воспаления, язв, опухолей молочной железы, легкого, головы и шеи, пищевода, желудка, лимфоидной ткани, предстательной железы, метастатических узелков, саркомы Капоши, гемангиоэндотелиомы типа Дабска и кожного лимфангиоматоза. Иммунизацию VEGF-D можно использовать специально для лечения метастазов лимфатических узлов.
Применение корецепторов NRP1 и NRP2 для иммунизации млекопитающих является полезным для лечения, в частности, пролиферации сосудов фиброзной ткани в случае рака предстательной железы, меланомы, остеосаркомы, метастазов рака молочной железы, диабетической ретинопатии и ревматоидного артрита.
Исследования на основе пассивной иммунотерапии, осуществляемой путем введения антител, показали, что комбинация антител против VEGF-A и KDR является более эффективной в моделях сингенных опухолей. Таким образом, использование двух или большего числа иммуногенов согласно изобретению позволяет получить особенно эффективные результаты при ингибировании ангиогенеза и роста опухоли. Указанные иммуногены можно вводить отдельно или попарно, используя бицистронные векторы, описанными способами. Кроме того, вакцинные композиции согласно изобретению можно использовать вместе с лекарственными или химиотерапевтическими средствами, предназначенными для лечения конкретных заболеваний, при одновременном или последовательном введении.
Представленные ниже результаты показывают, что антиангиогенные и противоопухолевые реакции опосредованы совокупностью гуморальных и клеточных реакций. В частности, VEGF и его рецептор участвуют в процессе созревания дендритных клеток и воздействуют на клетки-предшественники В- и Т-лимфоцитов. В примере 10 показано, что предложенный способ лечения, помимо снижения уровней VEGF в сыворотке, также способствует нормализации соотношения В- и Т-лимфоцитов и зрелых дендритных клеток. Указанный эффект способствует презентации опухолевых антигенов в MHC I, что улучшает качество и интенсивность иммунной противоопухолевой реакции, направленной не только на иммуноген, но и на другие опухолеассоциированные, опухолеспецифические и сверхэкспрессированные антигены в опухоли.

Примеры
Пример 1
Клонирование и временная экспрессия антигенов
VEGF человека, его изоформы и функциональные мутанты
Изоформы VEGF клонировали при помощи полимеразной реакции синтеза цепи (PCR), используя в качестве матрицы кДНК, полученную из ранее выделенной мРНК линии клеток CaSki (АТСС CRL 1550), в соответствии с инструкциями изготовителя (Perkin-Elmer) и затравки SEQ ID NO:1 и SEQ ID NO:2. Полосы, соответствующие продуктам амплификации изоформ VEGF 121, 165 и 189, вырезали из 2% агарозных гелей. Полосы гидролизовали эндонуклеазами BamHI и EcoRI, кДНК из изоформ VEGF очищали и независимо клонировали в векторе РАЕСΔ2 (патентованный вектор CIGB). Полученные плазмиды секвенировали, при этом не были обнаружены мутации аминокислотных последовательностей, о которых сообщалось EMBL (www.embl-heidelberg.de) для клонированных изоформ. кДНК, соответствующую изоформам VEGF, затем клонировали Kpnl/EcoRV в векторе рМАЕ5Δ5, который наряду с прочими характеристиками отличается от рАЕСΔ2 наличием 5'-концевых иммуностимулирующих сайтов CpG.
кДНК из варианта VEGF с отсутствием связывания с рецептором KDR (VEGFKDR(-)) была получена прямым мутагенезом предварительно клонированной изоформы VEGF121 в соответствии с описанием, приведенным Siemeister G. и др. (Siemeister G. et al., J. Biol. Chem. 273: 11115, 1998).
Мутированный вариант был получен при помощи PCR с использованием нижеследующих затравок:
(А) амплификация 5'-концевого фрагмента (315 п.н.) при помощи затравок с последовательностями SEQ ID NO:3 и SEQ ID NO:4;
(В) амплификация 3'-концевого фрагмента (93 п.н.) при помощи затравок с последовательностями SEQ ID NO:5 и SEQ ID NO:6.
Амплифицированные таким образом фрагменты очищали вышеописанным способом и использовали в эквимолярных концентрациях в качестве матрицы для слияния при помощи PCR с использованием затравок, соответствующих последовательностям SEQ ID NO:7 и SEQ ID NO:8. Полученную кДНК, содержащую мутацию, гидролизовали BamHI/EcoRI, очищали и клонировали в векторе рАЕСΔ2. Введенные мутации проверяли секвенированием, и ДНК, соответствующую VEGFKDR(-), субклонировали Kpnl/EcoRV в вектор рМАЕ5Δ5, в результате чего был получен вектор рМАЕ5Δ5 VEGFKDR(-).
Плазмиды, используемые как для трансфекции, так и для вакцинации животных, очищали в условиях без эндотоксина в соответствии с описанием, приведенным Whalen R. и др. (Whalen R.G. y Davis H.L., Clin. Immunol. Immunopathol. 75:1, 1995). ДНК очищали, используя системы QIAGEN без эндотоксина, в соответствии с инструкциями изготовителя, после чего ДНК подвергали второй преципитации. Наконец, ДНК растворяли в физиологическом растворе с фосфатным буфером (PBS) без эндотоксина (Sigma, USA) до конечной концентрации, равной 4 мг/мл.
1.2. Рецептор VEGF человека (KDR/Flk1)
кДНК, кодирующие внеклеточный домен рецептора KDR фактора VEGF (KDR1-3), трансмембранные и внутриклеточные домены данного рецептора (KDR TC), получали при помощи RT-PCR с использованием мРНК линии эндотелиальных клеток HUVEC (Clonetic, USA) и обрабатывали VEGF человека (Sigma) и гепарином (Sigma).
В случае внеклеточных доменов 1-3 используемые затравки соответствуют последовательностям SEQ ID NO:9 и SEQ ID NO:10. Амплифицированный фрагмент (943 п.н.) гидролизовали эндонуклеазами BamHI и EcoRI, после чего кДНК, кодирующую домены 1-3 KDR, очищали и клонировали в векторе рАЕСΔ2. Положительные клоны, выявленные рестрикционным анализом, проверяли секвенированием соответствующей ДНК. кДНК, соответствующую KDR 1-3, субклонировали Kpnl/EcoRV в уже описанном векторе рМАЕ5Δ5 (рМАЕ5Δ5 KDR1-3).
Трансмембранные и цитозольные области рецептора клонировали двухстадийным способом. Для вставки первого сегмента использовали затравки, соответствующие SEQ ID NO:11 и SEQ ID NO:12. Сегмент длиной 747 п.н. гидролизовали XbaI/BglII, продукт клонировали в векторе рМАЕ5, который предварительно гидролизовали такими же ферментами, в результате чего была получена плазмида PMAE5 KDR 747. Указанную плазмиду гидролизовали BglII/NotI, чтобы вставить остальной карбоксиконцевой фрагмент длиной 1091 п.н., который был амплифицирован при помощи затравок, соответствующих последовательностям SEQ ID NO:13 и SEQ ID NO:14. Положительные клоны, выявленные рестрикционным анализом, проверяли путем секвенирования ДНК, после чего им было присвоено название рМАЕ5 KDR C.
1.2.1. Клонирование трансмембранных и цитозольных областей KDR в вирусном векторе
Трансмембранные и цитозольные области рецептора VEGF (KDR) клонировали в поксвирусе цыплят, используя затравки, соответствующие последовательностям SEQ ID NO:15 и SEQ ID NO:16. Фрагмент длиной 953 п.н. гидролизовали ферментами StuI/SmaI, продукт клонировали в векторе pFP67xgpt, который предварительно гидролизовали такими же ферментами. В тот же вектор, гидролизованный SmaI/BamHI, вставляли остальной фрагмент длиной 919 п.н., который амплифицировали из исходной кДНК при помощи затравок, соответствующих последовательностям SEQ ID NO:17 и SEQ ID NO:18. Положительные клоны, выявленные рестрикционным анализом, проверяли путем секвенирования ДНК, после чего им было присвоено название pFP67xgpt KDR C.
Поксвирус цыплят (FWPV) реплицировали в фибробластах куриного зародыша (CEF) в среде DMEM, содержащей 2% фетальной телячьей сыворотки (FBS). Клон pFP67xgpt KDR C трансфицировали, используя липофектин (Gibco BRL, Grand Island, USA), в CEF, которые предварительно инфицировали ослабленным штаммом FP9. Через 24 часа добавляли свежую среду и клетки культивировали еще 3-4 дня. Затем клетки трижды замораживали и оттаивали. Рекомбинантные вирусы, экспрессирующие ген, кодирующий фермент Ecogpt, очищали в избирательной среде, используя микофенольную кислоту (25 мкг/мл), ксантин (250 мкг/мл) и гипоксантин (15 мкг/мл) (МХН). Правильное включение гена в рекомбинантные вирусы проверяли при помощи PCR. Рекомбинантные вирусы получили название FPKDRgpt, нерекомбинантные вирусы использовали в качестве отрицательного контрольного штамма FP.
Пример 2
Экспрессия антигенов in vivo
Чтобы подтвердить потенциал конструкций, созданных для экспрессии белков in vivo, указанные конструкции инъецировали мышам С57BL6 в четырехглавую мышцу бедра (3 мыши в группе).
1. рМАЕ5Δ5-VEGF121 (10 и 50 мкг/мышь) в PBS, рН 7,2
2. рМАЕ5Δ5-VEGF165 (10 и 50 мкг/мышь) в PBS, рН 7,2
3. рМАЕ5Δ5-VEGF189 (10 и 50 мкг/мышь) в PBS, рН 7,2
4. рМАЕ5Δ5-VEGFKDR(-) (10 и 50 мкг/мышь) в PBS, рН 7,2
5. рМАЕ5Δ5-KDR 1-3 (10 и 50 мкг/мышь) в PBS, рН 7,2
6. рМАЕ5 KDR C (10 и 50 мкг/мышь) в PBS, рН 7,2
7. FPKDRgpt (2,5*107 КОЕ) в PBS, рН 7,2
8. PBS, рН 7,2 (отрицательная контрольная группа)
Через 48 часов после инъекции животных умерщвляли и полностью удаляли мышцы, в которые была сделана инъекция. Часть мышечной ткани гомогенизировали в присутствии ингибиторов протеазы и неионных детергентов. Наличие VEGF в белковых экстрактах анализировали методами дот-блоттинга и вестерн-блоттинга, используя поликлональное антитело, распознающее все изоформы VEGF человека (sc-152G), в соответствии с описанными способами. РНК экстрагировали из остальной мышечной ткани, используя TRI-реагент (Sigma). В общей сложности 20 мкг РНК, полученной в каждом эксперименте, очищали электрофорезом в 1% агарозном геле, содержащем формальдегид. РНК переносили на нейлоновый фильтр (HYBOND) и гибридизировали с кДНК изоформы VEGF 121, меченной АТР32, которая распознает все изоформы VEGF, или с кДНК рецептора KDR, меченной аналогичным образом. В обоих случаях фильтры повторно гибридизировали кДНК, соответствующей структурному гену: глицеральдегид-3-фосфатдегидрогеназа (GAPDH). Во всех анализированных конструкциях идентифицировали полосы, соответствующие VEGF человека, и клонированные фрагменты рецептора KDR.
Пример 3
Эксперименты по созданию иммунитета in vivo путем вакцинации плазмидой, содержащей фрагменты гена рецептора KDR фактора VEGF
Группы, состоящие из 10 мышей С57BL/6, вакцинировали или не вакцинировали в соответствии с нижеследующими вариантами:
1. рМАЕ5Δ5-KDR 1-3 (1, 10, 50 и 100 мкг/мышь) в PBS, рН 7,2
2. рМАЕ5 KDR C (1, 10, 50 и 100 мкг/мышь) в PBS, рН 7,2
3. FPKDRgpt (2,5*107 КОЕ)
4. PBS, рН 7,2 (отрицательная контрольная группа)
5. FP (2,5*107 КОЕ) (отрицательная контрольная группа 3)
Во всех экспериментах мышей иммунизировали путем внутримышечной инъекции (im) общим объемом 50 мкл в заднюю левую лапу. Всех животных повторно иммунизировали через 15 дней в соответствии с первоначальной схемой иммунизации. Контрольное заражение опухолью производили через тридцать дней после последней иммунизации путем подкожной (sc) инъекции 104 клеток меланомы В16-F10 (АТСС, CRL-6475) всем животным в правую часть брюшной области. Рост опухоли контролировали, производя три измерения в неделю до самой смерти животных.
У мышей, иммунизированных плазмидой рМАЕ5Δ5-KDR 1-3, уменьшение размера опухоли наблюдалось при введении доз, равных 50 и 100 мкг ДНК/мышь, при этом опухоль была значительно меньше, чем у животных в отрицательной контрольной группе (таблица 3). Результаты анализа выживаемости, выполненного на 33-й день, свидетельствовали о значительном увеличении (по сравнению с отрицательной контрольной группой) данного параметра у животных, иммунизированных указанной ДНК в дозах, равных 50 и 100 мкг/мышь, по сравнению с неиммунизированными мышами (группа PBS рН 7,2). В случае применения рМАЕ5Δ5-KDR C (таблица 3) значительное уменьшение объема опухоли наблюдалось при всех четырех использованных дозах, причем выживаемость увеличивалась при введении доз от 100 до 10 мкг/животное. При использовании вирусных векторов наблюдалось уменьшение объема опухоли и увеличение выживаемости в условиях, аналогичных применению конструкции FPKDRgpt (таблица 3), по сравнению с соответствующей отрицательной контрольной группой (группа мышей, иммунизированных вектором без вставки FPgpt).
Объем опухоли и выживаемость мышей, иммунизированных фрагментами гена рецептора (KDR) VEGF
Пример 4
Эксперименты по созданию иммунитета in vivo путем вакцинации плазмидами, содержащими изоформы VEGF и мутированный вариант
Группы, состоящие из 10 мышей С57BL/6, вакцинировали или не вакцинировали в соответствии с нижеследующими вариантами:
1. рАЕСΔ2-VEGF121 (1, 10, 50 и 100 мкг/мышь) в PBS, рН 7,2
2. рМАЕ5Δ5-VEGF121 (1, 10, 50 и 100 мкг/мышь) в PBS, рН 7,2
3. рМАЕ5Δ5-VEGF165 (1, 10, 50 и 100 мкг/мышь) в PBS, рН 7,2
4. рМАЕ5Δ5-VEGF189 (1, 10, 50 и 100 мкг/мышь) в PBS, рН 7,2
5. рМАЕ5Δ5-VEGFKDR(-) (1, 10, 50 и 100 мкг/мышь) в PBS, рН 7,2
6. PBS, рН 7,2 (отрицательная контрольная группа)
Во всех экспериментах мышей иммунизировали путем внутримышечной инъекции (im) общим объемом 50 мкл в заднюю левую лапу. Всех животных повторно иммунизировали через 15 дней в соответствии с первоначальной схемой иммунизации. Контрольное заражение опухолью производили через тридцать дней после последней иммунизации путем подкожной инъекции 104 клеток меланомы В16-F10 (АТСС, CRL-6475) всем животным в правую часть брюшной области. Рост опухоли контролировали, производя три измерения в неделю до самой смерти животных.
В тех случаях, когда мышей иммунизировали депротеинизированной ДНК в векторе серии рАЕС при введении дозы 100 мкг/животное, наблюдалось замедление роста опухоли по сравнению с отрицательной контрольной группой (таблица 4). При использовании вариантов, включающих вектор серии рМАЕ5Δ5 с 5'-концевыми сайтами CpG независимо от изоформы VEGF, размер опухоли значительно уменьшался в группах мышей, иммунизированных ДНК в дозах 10, 50 или 100 мкг, по сравнению с отрицательной контрольной группой. В случае использования мутированного варианта рМАЕ5Δ5 VEGFKDR(-) значительное уменьшение размера опухоли было достигнуто при введении доз, аналогичных используемым при иммунизации рМАЕ5Δ5-VEGF121.
Результаты анализа выживаемости, выполненного на 43-й день, свидетельствовали о значительном увеличении (по сравнению с отрицательной контрольной группой) данного показателя у животных, иммунизированных вариантами рМАЕ5Δ5-VEGF121, рМАЕ5Δ5-VEGF165, рМАЕ5Δ5-VEGF189 и рМАЕ5Δ5-VEGFKDR(-), при введении доз, равных 50 и 100 мкг/животное (таблица 4).
Объем опухоли и выживаемость мышей, иммунизированных разными вариантами депротеинизированной ДНК, содержащей разные изоформы гена VEGF и мутированный вариант
Пример 5
Эксперименты по созданию иммунитета in vivo путем иммунизации рМАЕ5Δ5-VEGF121 и рМАЕ5Δ5-KDR 1-3 в модели индуцированного коллагеном артрита
Группы, состоящие из 20 мышей, вакцинировали или не вакцинировали в соответствии с нижеследующими вариантами:
1. рМАЕ5Δ5-VEGF121 (50 мкг ДНК/мышь) в PBS, рН 7,2
2. рМАЕ5Δ5-KDR 1-3 (50 мкг ДНК/мышь) в PBS, рН 7,2
3. PBS, рН 7,2 (отрицательная контрольная группа)
Во всех экспериментах мышей иммунизировали путем внутримышечной инъекции (im) общим объемом 50 мкл в заднюю левую лапу. Всех животных повторно иммунизировали через 15 дней в соответствии с первоначальной схемой иммунизации.
На 5-й день вызывали аутоиммунный артрит путем иммунизации куриным коллагеном типа II (Sigma) в соответствии с моделью, описанной Campbell и др. (Campbell I.K. et al., Eur. J. Immunol. 30:1568, 2000). Указанную иммунизацию повторяли на 26-й день. У каждой мыши ежедневно оценивали состояние всех четырех конечностей на основании показателя поражения артритом, для чего делали 0-3 пункции каждой конечности для выявления признаков эритемы (1), воспаления (2) или неподвижности (3) при максимальной оценке, равной 12. У мышей появлялись клинические симптомы развития артрита на 23-й день после иммунизации, при этом наибольшая заболеваемость была выявлена через 50 дней. В таблице 5 приведены данные заболеваемости артритом у животных в разных экспериментальных группах. На 40-й и 55-й день было отмечено значительное снижение заболеваемости артритом в вакцинированных группах (1 и 2) по сравнению с контрольной группой.
Заболеваемость артритом в определенные дни (40 и 55)
Пример 6
Антиангиогенное действие вакцинации in vivo
Группы, состоящие из 15 мышей, вакцинировали или не вакцинировали в соответствии с нижеследующими вариантами:
1. рМАЕ5Δ5-VEGF121 (50 мкг ДНК/мышь) в PBS, рН 7,2
2. рМАЕ5Δ5-KDR 1-3 (50 мкг ДНК/мышь) в PBS, рН 7,2
3. рМАЕ5 KDR C (50 мкг/мышь) в PBS, рН 7,2
4. PBS, рН 7,2 (отрицательная контрольная группа)
Во всех экспериментах мышей С57ВL/6 иммунизировали путем внутримышечной инъекции (im) общим объемом 50 мкл в заднюю левую лапу. Всех животных повторно иммунизировали через 15 дней в соответствии с первоначальной схемой иммунизации. Через тридцать дней после последней иммунизации у животных оценивали степень развития кровеносных сосудов in vivo, используя матригель аналогично описанию, приведенному Coughlin M.C. и др. (Coughlin M.C. et al., J. Clin. Invest. 101:1441, 1998). Предварительно вакцинированных животных распределяли в группы по 5 животных в каждой и в середину брюшной области подкожно инъецировали 500 мкл матригеля (Becton Dickinson and Co., Franklin Lakes, New Jersey, USA), содержащего:
1. VEGF 50 нг/мл, гепарин 50 ед/мл
2. 105 клеток меланомы В16-F10
3. Физиологический раствор с фосфатным буфером (PBS)
Через шесть дней после вакцинации животных умерщвляли и извлекали пробку матригеля. Содержание гемоглобина в пробках анализировали в соответствии с инструкциями изготовителя (набор реагентов Драбкина; Sigma Diagnostics Co., St. Louis, Missouri, USA). Вакцинация плазмидами, кодирующими VEGF или его рецептор KDR, значительно подавляет (р<0,001) развитие кровеносных сосудов, вызванное VEGF, а также индуцированное более сложными системами, такими как опухолевые клетки.
Пример 7
Получение иммуногена путем нековалентного связывания рМАЕ5Δ5-VEGF121 с разными адъювантами
Разные иммуностимулирующие средства были использованы в смеси с конструкцией рМАЕ5Δ5-VEGF121 в соответствии с нижеописанным способом. Белок Opc, выделенный из наружной мембраны Neisseria meningitidis, очищали методом Musacchio и др. (Musacchio et al., Vaccine, 67:751, 1997). 50 мкг/мл рМАЕ5Δ5-VEGF121 добавляли к 10 мкг/мл белка Орс при осторожном встряхивании смеси с кислотным показателем рН. Полученный комплекс диализовали в течение ночи в PBS без эндотоксина с рН 7,2 (Sigma). Уровень ассоциации белок Орс-плазмидная ДНК (Орс-рМАЕ5Δ5-VEGF121) проверяли путем визуализации ДНК с использованием 1% агарозного геля. Более 50% плазмидной ДНК было ассоциировано с белком Орс.
В комбинации с представляющей интерес плазмидной ДНК использовали очень мелкие частицы (VSSP) комплекса белков наружной мембраны (ОМРС) Neisseria meningitides, предоставленные Центром молекулярной иммунологии (R. Perez et al., заявки на патент США № 5788985 и 6149921). VSSP (1 мг) инкубировали с 5 мг рМАЕ5Δ5-VEGF121 в течение ночи при осторожном перемешивании. Полученное вещество продолжительное время диализовали в PBS без эндотоксина, рН 7,2 (Sigma). Уровень ассоциации VSSP-плазмидная ДНК (VSSP-pMAE5Δ5-VEGF121) проверяли путем визуализации ДНК с использованием 1% агарозного геля. Более 50% плазмидной ДНК было ассоциировано с частицами VSSP.
Ядерные антигены гепатита С и гепатита В (HCcAg и HBcAg) в виде частиц были получены в соответствии с описанием, приведенным в научной литературе (Lorenzo L.J. et al., Biochem. Biophys. Res. Commun. 281:962, 2001). 1 мг антигенов смешивали с 5 мг плазмиды и инкубировали в течение ночи. Уровни ассоциации HCcAg или HBcAg-плазмидная ДНК (соответственно HCcAg-pMAE5Δ5-VEGF121 и HBcAg-pMAE5Δ5-VEGF121) проверяли путем визуализации ДНК с использованием 1% агарозного геля. Более 50% ДНК было ассоциировано с частицами антигена в каждом эксперименте.
Пример 8
Эксперименты по созданию иммунитета in vivo при помощи конструкции рМАЕ5Δ5-VEGF121 и адъювантов, вызывающих иммунную реакцию
Группы, состоящие из 10 мышей С57BL/6, вакцинировали или не вакцинировали в соответствии с нижеследующими вариантами:
1. рМАЕ5Δ5-VEGF121 (1, 10 и 50 мкг ДНК/мышь) в PBS, рН 7,2
2. Орс-рМАЕ5Δ5-VEGF121 (1, 10 и 50 мкг ДНК/мышь)
3. VSSP-рМАЕ5Δ5-VEGF121 (1, 10 и 50 мкг ДНК/мышь)
4. HBcAg-рМАЕ5Δ5-VEGF121 (1, 10 и 50 мкг ДНК/мышь)
5. HCcAg-рМАЕ5Δ5-VEGF121 (1, 10 и 50 мкг ДНК/мышь)
6. PBS, рН 7,2 (отрицательная контрольная группа 1)
7. Орс (отрицательная контрольная группа 2)
8. VSSP (отрицательная контрольная группа 3)
9. HBcAg (отрицательная контрольная группа 4)
10. HCcAg (отрицательная контрольная группа 5)
Иммунизацию, контрольное заражение опухолью и измерение объема опухоли выполняли аналогично предыдущему примеру. Варианты вакцины в дозах, равных 10 мкг ДНК/мышь или выше, замедляли рост опухоли по сравнению с соответствующими отрицательными контрольными группами (таблица 6). Значительно более высокая выживаемость по сравнению с аналогичным показателем в соответствующей контрольной группе наблюдалась у животных, иммунизированных геном VEGF, ассоциированным или не ассоциированным с Орс, VSSP, HCcAg и HBcAg, которые использовали в качестве иммунопотенциаторных носителей. Все варианты с носителем обеспечивали значительно более высокую выживаемость по сравнению с соответствующей контрольной группой при дозах от 10 мкг/мышь и выше, в то время как вариант, представляющий собой депротеинизированную ДНК с вектором рМАЕ5Δ5-VEGF121, вызывал результаты, существенно отличные от отрицательной контрольной группы, при дозе, равной 50 мкг/мышь (таблица 6).
Объем опухоли и выживаемость у мышей, иммунизированных разными иммуностимулирующими средствами
(5 мкг/мышь/доза)
(5 мкг/мышь/доза)
Пример 9
Эксперимент по созданию иммунитета in vivo при помощи VEGF в форме белка
Группы, состоящие из 10 мышей С57BL/6, вакцинировали или не вакцинировали в соответствии с нижеследующими вариантами:
VEGF165 (20 мкг/мышь) с полным или неполным адъювантом Фрейнда;
полный и неполный адъювант Фрейнда (отрицательная контрольная группа).
Антиген VEGF165 был получен из коммерческого источника (Sigma) с чистотой более 97%. Мышей иммунизировали при помощи внутрибрюшинной инъекции, используя полный адъювант Фрейнда (Sigma), и повторно иммунизировали через 15 и 30 дней тем же способом, но используя неполный адъювант Фрейнда. Контрольное заражение опухолью и измерения объема опухоли производили аналогично предыдущему примеру.
В группе, иммунизированной VEGF, наблюдалось значительное уменьшение объема опухоли и увеличение выживаемости по сравнению с контрольной неиммунизированной группой. Достигнутый эффект был аналогичен обнаруженному в предыдущих примерах с использованием ДНК VEGF.
Пример 10
Эксперименты по созданию иммунитета in vivo у мышей C57BL/6 с тяжелым комбинированным иммунодефицитом (SCID)
Мышей C57BL/6 иммунизировали 50 мкг дозами рМАЕ5Δ5-VEGF121 ДНК/мышь способами, описанными в примере 5, или не иммунизировали. Мышей умерщвляли через 45 дней после первой иммунизации. CD8+, CD4+ и В-лимфоциты указанных мышей отделяли при помощи магнитных шариков (Dynabeads, USA) в соответствии с инструкциями изготовителя.
Мышей C57BL/6 в возрасте шести недель с SCID в группах, состоящих из 10 животных, подвергали иммуновосстановительной терапии, включающей введение нижеследующих комбинаций ранее удаленных лимфоцитов.
Группа 1: CD8+ Т-лимфоциты и CD4+ Т-лимфоциты, полученные у мышей, иммунизированных ДНК рМАЕ5Δ5-VEGF121. В-лимфоциты не восстанавливали.
Группа 2: В-лимфоциты и CD4+ Т-лимфоциты иммунизированных мышей и CD8+ Т-лимфоциты неиммунизированных мышей.
Группа 3: В-лимфоциты, CD8+ Т-лимфоциты и CD4+ Т-лимфоциты иммунизированных мышей, использованные в данном эксперименте в качестве положительного контрольного средства.
Группа 4: В-лимфоциты, CD8+ Т-лимфоциты и CD4+ Т-лимфоциты неиммунизированных мышей, использованные в данном эксперименте в качестве отрицательного контрольного средства.
Мышей с SCID, подвергнутых иммуновосстановительной терапии, заражали при помощи подкожной инъекции 104 клеток меланомы В16-F10. Рост опухоли контролировали, производя три измерения в неделю до самой смерти животных. Уровни антитела против VEGF анализировали лабораторным методом ELISA. 96-Луночные планшеты инкубировали в течение ночи с 0,5 мкг/мл раствора VEGF165 (Sigma). Лунки блокировали раствором PBS с 1% BSA (BDH, UK) и затем инкубировали с несколькими разведениями животной сыворотки. Культуру промывали PBS с 0,05% твина и добавляли полученное коммерческим путем поликлональное антитело против IgG мыши (Sigma, A0168). Сигнал амплифицировали в присутствии коммерческого субстрата ортофенилендиамина (OPD, Sigma).
В таблице 7 приведены показатели объема опухоли (24-й день) и выживаемости (40-й день) мышей из разных групп, зараженных опухолью. Начиная с 15-го дня после иммуновосстановительной терапии, у животных в группах 1-3 наблюдалось уменьшение размера опухоли по сравнению с группой 4, которой вводили лимфоциты неиммунизированных мышей. Таким образом, воздействие, возбуждающее иммунную систему у иммунизированных мышей, в результате которого уменьшается размер опухоли, связано с гуморальными и клеточными реакциями, причем указанная последней реакция относится к цитотоксическому типу (CTL) вследствие отсутствия антител против VEGF в группе 1. Тем не менее, в созданных экспериментальных условиях выживаемость повысилась только в группе 3 (В- и Т-лимфоциты иммунизированных мышей) по сравнению с остальными группами (таблица 7). У животных с отсутствием реакций В- или Т-лимфоцитов типа CTL (соответственно группы 1 и 2), которые были подвергнуты неполной иммуновосстановительной терапии, выживаемость не отличалась от аналогичного показателя в отрицательной контрольной группе. Полученные результаты показывают, что комбинация гуморальных и клеточных реакций (группа 4) оказывает синергичное действие, которое способно вызывать эффективную реакцию, позволяющую увеличить выживаемость мышей, зараженных опухолью.
Объем опухоли и выживаемость у мышей с SCID, подвергнутых восстановительной иммунотерапии с использованием лимфоцитов мышей, иммунизированных рМАЕ5Δ5-VEGF121
лимфоциты
лимфоциты
Пример 11
Иммунологическое восстановление в результате иммунной реакции, определяемое по уменьшению циркулирующего VEGF
Группам, состоящим из 15 самок мышей С57BL/6, внутримышечно инъецировали нижеследующие варианты:
1. рМАЕ5Δ5-VEGF121 (50 мкг/мышь) в PBS, рН 7,2
2. PBS, рН 7,2
Во всех экспериментах мышей иммунизировали путем внутримышечной инъекции (im) общим объемом 50 мкл в заднюю левую лапу. Всех животных повторно иммунизировали через 15 дней в соответствии с первоначальной схемой иммунизации. Через тридцать дней после последней иммунизации 5 произвольно отобранных животных из каждой группы умерщвляли для анализа иммунологического состояния иммунизированных и контрольных животных, а также токсического воздействия вакцинации на органы и ткани при помощи макроскопических и гистологических анализов.
Остальным животным во всех группах подкожно инъецировали 104 клеток меланомы В16-F10 в правую часть брюшной области. Через 15 и 30 дней после инъекции опухолевых клеток умерщвляли по 5 мышей из каждой группы и производили вышеописанные исследования.
У исследованных животных не было обнаружено токсическое воздействие на макроскопическом уровне, и гистопатологический анализ не выявил повреждения каких-либо органов при исследовании через 30 дней после последней иммунизации. Иммунологический анализ включал: (1) определение уровней VEGF мышей в сыворотке; (2) содержание Т- и В-лимфоцитов, а также степень зрелости дендритных клеток в селезенке и бронхиальных вспомогательных и паховых лимфатических узлах.
Анализ уровней VEGF (набор R&D для определения VEGF у мышей) в сыворотке неиммунизированных мышей показал, что с увеличением времени развития опухоли уровни VEGF в сыворотке увеличивались в соответствии с увеличением размера опухоли с течением времени. В группах мышей, иммунизированных против VEGF человека, было обнаружено значительное снижение (р<0,001 по методу ANOVA, пост-критерий Бонферрони) уровней VEGF у мышей, которое сохранялось через 30 дней после контрольного заражения опухолью.
Состояние иммунной системы животных, умерщвленных в каждый период времени, анализировали, исследуя соотношения популяций клеток в лимфатических узлах и селезенке способом, описанным Gabrilovich и др. (Gabrilovich D. et al., Blood 92:4150, 1998). Для выполнения указанных исследований использовали коммерческие моноклональные антитела, распознающие молекулы CD3, CD19, CD11c и CD86 (B7-2) (Pharmingen), меченные флуоресцеинизотиоцианатом (FITC) и фикоэритрином (РЕ), которые позволяли визуализировать популяции клеток при помощи проточного цитометра (FACS). Полученные результаты приведены в таблице 8.
Краткий обзор результатов анализа FACS популяций клеток на основании поверхностных маркеров
Анализы популяций лимфоидных клеток и созревания дендритных клеток у животных через 30 дней после иммунизации показывают, что вакцинация ДНК VEGF не вызывает какого-либо изменения состояния иммунной системы у животного. Тем не менее, через 30 дней после имплантации опухоли у невакцинированных животных наблюдается уменьшение соотношения Т-лимфоциты/В-лимфоциты (CD3/CD19) как в лимфатических узлах, так и в селезенке по сравнению с соотношением, имевшим место до заражения опухолью. Кроме того, в селезенке наблюдается значительное уменьшение числа лимфоидных клеток. У указанных животных также было обнаружено сокращение числа зрелых дендритных клеток в лимфатических узлах и селезенке. В группе мышей, вакцинированных ДНК VEGF, наблюдалось значительное восстановление всех показателей, которое можно коррелировать с уменьшением уровней VEGF в сыворотке, обнаруженным у животных в данной группе.
| название | год | авторы | номер документа |
|---|---|---|---|
| РЕКОМБИНАНТНЫЕ АНТИТЕЛА ПРОТИВ ФАКТОРА РОСТА ЭНДОТЕЛИЯ СОСУДОВ (VEGF), ПОЛУЧЕННЫЕ ПОСРЕДСТВОМ МУТАГЕНЕЗА ВАРИАБЕЛЬНОЙ ОБЛАСТИ | 2011 |
|
RU2557309C2 |
| ДНК-ВАКЦИНА ПРОТИВ ПРОЛИФЕРИРУЮЩИХ ЭНДОТЕЛИАЛЬНЫХ КЛЕТОК И СПОСОБЫ ЕЕ ПРИМЕНЕНИЯ | 2003 |
|
RU2318019C2 |
| КОМПОЗИЦИЯ ДНК ДЛЯ ВЫЗОВА ИММУННОГО ОТВЕТА ПРОТИВ ОПУХОЛЕАССОЦИИРОВАННЫХ МАКРОФАГОВ | 2007 |
|
RU2459631C2 |
| ДНК-ВАКЦИНЫ ПРОТИВ ОПУХОЛЕВОГО РОСТА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2004 |
|
RU2343195C2 |
| РЕКОМБИНАНТНЫЙ БЕЛОК, НЕСУЩИЙ ЭПИТОПЫ ВИРУСА ПАПИЛЛОМЫ ЧЕЛОВЕКА, ВСТРОЕННЫЕ В БЕЛОК АДЕНИЛАТЦИКЛАЗУ ИЛИ ЕГО ФРАГМЕНТ, ЕГО ТЕРАПЕВТИЧЕСКОЕ ПРИМЕНЕНИЕ | 2005 |
|
RU2441022C2 |
| ВАКЦИННЫЙ СОСТАВ, ПОТЕНЦИРОВАННЫЙ КОМБИНАЦИЕЙ ДНК И АНТИГЕНА | 2002 |
|
RU2294212C2 |
| НАНОАНТИТЕЛО V9, СВЯЗЫВАЮЩЕЕ VEGF, И СПОСОБ ЕГО ПОЛУЧЕНИЯ, КОДИРУЮЩАЯ V9 НУКЛЕОТИДНАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ И СОДЕРЖАЩИЙ ЕЕ ВЕКТОР, СПОСОБ ИНГИБИРОВАНИЯ ПРОЛИФЕРАЦИИ ЭНДОТЕЛИАЛЬНЫХ КЛЕТОК | 2008 |
|
RU2395521C1 |
| АГЕНТЫ И СПОСОБЫ, ОСНОВАННЫЕ НА ПРИМЕНЕНИИ ДОМЕНА EDA ФИБРОНЕКТИНА | 2006 |
|
RU2430738C2 |
| КОМБИНАЦИИ РЕГУЛИРУЮЩИХ РОСТ ФАКТОРОВ И ГОРМОНОВ ДЛЯ ЛЕЧЕНИЯ НЕОПЛАЗИИ | 2003 |
|
RU2321422C2 |
| ПЕПТИДНЫЙ АНТАГОНИСТ ИНТЕРЛЕЙКИНА-15 | 2005 |
|
RU2396276C2 |
Изобретение относится к области медицины и касается иммунотерапии против ангиогенеза. Сущность изобретения включает иммуногенные композиции для лечения расстройств, ассоциированных с усилением ангиогенеза, содержащие олигонуклеотиды, кодирующие полипептиды VEGFR2, вводимые в виде части плазмидных или вирусных векторов, а также полипептиды VEGFR2, олигонуклеотиды, кодирующие аутологичные VEGF с нарушенной способностью к активации рецептора, полипептиды VEGF и их комбинации. Иммуногенные композиции могут применяться для лечения злокачественных новообразований и их метастазов, при доброкачественных новообразованиях и при хронических воспалительных и аутоиммунных болезнях. Преимущество изобретения заключается в индукции заявленными композициями гуморального и клеточного иммунитета. 9 н. и 10 з.п. ф-лы, 8 табл.
| WO 00/53219A, 14.09.2000 | |||
| US 6316603 A, 13.11.2001 | |||
| US 6361946 A, 26.03.2002 | |||
| WEI YQ et al., Immunogene therapy of tumors with vaccine based on Xenopus homologous vascular endothelial growth factor as a model antigen, Proc | |||
| Natl | |||
| Acad | |||
| Sci | |||
| USA, 2001, Sep 25, 98(20), pp.11545-50. |

