Область, к которой относится изобретение
Настоящее изобретение относится к полиспецифическим антителам и способам получения и применения таких антител.
Предпосылки создания изобретения
Антитела представляют собой специфические иммуноглобулиновые полипептиды, продуцируемые иммунной системой позвоночных в ответ на заражение чужими белками, гликопротеинами, клетками или другими антигенными "чужими" веществами. Важной частью этого процесса является выработка антител, которые специфически связываются с конкретным "чужим" веществом. Специфичность связывания таких полипептидов с конкретным антигеном является очень тонкой (утонченной), и совокупность (множество) специфичностей, способных вырабатываться отдельным позвоночным, является удивительным по своей сложности и вариабельности. Тысячи антигенов способны выявлять ответы (реакции), каждый из которых почти исключительно направлен на конкретный антиген, который выявил этот ответ (реакцию).
Специфическое распознавание антигена является необходимым для функционирования антител при адаптивном иммунном ответе. Комбинаторная ассоциация тяжелой цепи (НС) и легкой цепи (LC) при выработке спектра антител сохраняется у всех позвоночных. Однако в обеих цепях наблюдается асимметрия и разнообразие (вариабельность). Вариабельный домен НС (VH) содержит значительно большее разнообразие последовательностей и вводит детерминанты распознавания антигена чаще, чем вариабельный домен LC (VL). На роль LC в определении специфичности к антигену указывает процесс, называемый "редактированием" рецептора. Постоянная рекомбинация VL генов с целью редактирования В-клеточного рецептора является основным механизмом корректировки предшественников аутореактивных антител, которые, по-видимому, составляют значительную часть начального спектра (~75%). Показано, что изменение легкой цепи гасит (аннулирует) нежелательную специфичность связывания или полиспецифичность.
Специфичность антител и фрагментов антител к конкретному антигену или к конкретным антигенам делает антитела подходящими (хорошими) терапевтическими агентами. Антитела и фрагменты антител можно использовать для нацеливания на конкретные ткани, например на опухоль, и тем самым свести к минимуму потенциальные побочные эффекты неспецифического нацеливания. По этой причине существует постоянная необходимость в идентификации и определении характеристик терапевтических антител, в особенности антител, их фрагментов и производных, применимых для лечения рака и других пролиферативных расстройств.
Сущность изобретения
Настоящее изобретение охватывает выделенное антитело, включающее последовательность гипервариабельной области (HVR) L1, содержащую последовательность NIAKTISGY (SEQ ID NO:1), причем антитело специфически связывает рецептор эпидермального фактора роста 2-го типа (HER2) и сосудистый эндотелиальный фактор роста (VEGF). В одном варианте изобретения антитело дополнительно включает HVR-L2, содержащую последовательность WGSFLY (SEQ ID 2 NO:), и/или HVR-L3, содержащую последовательность HYSSPP (SEQ ID NO:3). В другом варианте изобретения антитело дополнительно включает одну, две или три HVR последовательности, выбранные из (i) HVR-H1, содержащей последовательность NIKDTY (SEQ ID NO:4); (ii) HVR-H2, содержащей последовательность RIYPTNGYTR (SEQ ID NO:5); и (iii) HVR-Н3, содержащей последовательность WGGDGFYAMD (SEQ ID NO:6). В другом варианте изобретения антитело дополнительно включает одну, две или три HVR последовательности, выбранные из (i) HVR-H1, содержащей последовательность NISGTY (SEQ ID NO:7); (ii) HVR-H2, содержащей последовательность RIYPSEGYTR (SEQ ID NO:8); и (iii) HVR-Н3, содержащей последовательность WVGVGFYAMD (SEQ ID NO:9).
В другом аспекте изобретение охватывает выделенное антитело, включающее последовательность HVR-L1, содержащую последовательность X1IX3X4X5X6X7X8X9Y (SEQ ID NO:83), где X1 обозначает любую аминокислоту, за исключением аспарагиновой кислоты, Х3 обозначает любую аминокислоту, за исключением пролина, Х4 обозначает любую аминокислоту, за исключением аргинина, и Х5 обозначает любую аминокислоту, за исключением серина, причем антитело специфически связывает HER2 и VEGF. В одном варианте изобретения антитело, содержащее последовательность X1IX3X4X5X6X7X8X9Y (SEQ ID NO:83), имеет аспарагин в X1, аланин в Х3, лизин в Х4, треонин в Х5, серин в X7 и/или глицин в Х8, или любую их комбинацию. В различных комбинациях этого аспекта изобретения любой из остатков HVR-L1, имеющий, как показано на Фигуре 57, значение F больше чем 1, 5 или 10, представляет собой остаток, который, предпочтительно, сохраняется в виде такого же остатка, находящегося в том же самом положении HVR-L1 в bH1-44 или bH1-81 (SEQ ID NO:1). В других вариантах изобретения любой из остатков HVR-L1, имеющий, как показано в Таблице 14, величину ΔΔG более 1, представляет собой остаток, который, предпочтительно, сохраняется в виде такого же остатка, находящегося в том же самом положении HVR-L1 в bH1-44 или bH1-81 (SEQ ID NO:1). В одном варианте изобретения антитело включает последовательность HVR-H2, содержащую последовательность RX2X3X4X5X6X7X8X9R (SEQ ID NO:84). В одном варианте изобретения антитело дополнительно включает последовательность HVR-L2, содержащую последовательность WGSFLY (SEQ ID NO:2), и/или HVR-L3, содержащую последовательность HYSSPP (SEQ ID NO:3). В другом варианте изобретения антитело дополнительно включает одну, две или три HVR последовательности, выбранные из (i) HVR-H1, содержащей последовательность NIKDTY (SEQ ID NO:4); (ii) HVR-H2, содержащей последовательность RIYPTNGYTR (SEQ ID NO:5); и (iii) HVR-Н3, содержащей последовательность WGGDGFYAMD (SEQ ID NO:6). В другом варианте изобретения антитело дополнительно включает одну, две или три HVR последовательности, выбранные из (i) HVR-H1, содержащей последовательность NISGTY (SEQ ID NO:7); (ii) HVR-H2, содержащей последовательность RIYPSEGYTR (SEQ ID NO:8); и (iii) HVR-Н3, содержащей последовательность WVGVGFYAMD (SEQ ID NO:9).
В другом аспекте изобретение охватывает выделенное антитело, включающее последовательность HVR-H2, содержащую последовательность RX2X3X4X5X6X7X8X9R (SEQ ID NO:85), где Х5 обозначает любую аминокислоту, за исключением треонина, а Х6 обозначает любую аминокислоту, за исключением аспарагина, и при этом антитело специфически связывает HER2 и VEGF. В другом варианте изобретения антитело, содержащее последовательность RX2X3X4X5X6X7X8X9R (SEQ ID NO:84), имеет тирозин в X8. В одном варианте изобретения антитело, содержащее последовательность RX2X3X4X5X6X7X8X9R (SEQ ID NO:84), имеет серин в X5 и/или глутаминовую кислоту в Х6. В другом варианте данного аспекта антитела дополнительно содержат одну, две или три HVR последовательности, выбранные из группы HVR-L1, содержащей последовательность NIAKTISGY (SEQ ID NO:1), HVR-L2, содержащей последовательность WGSFLY (SEQ ID NO:2), и/или HVR-L3, содержащей последовательность HYSSPP (SEQ ID NO:3). В любом из вариантов по данному описанию антитела дополнительно содержат одну или две HVR последовательности, выбранные из (i) HVR-H1, содержащей последовательность NIKDTY (SEQ ID NO:4), и (ii) HVR-H3, содержащей последовательность WGGDGFYAMD (SEQ ID NO:6). В другом варианте изобретения антитела дополнительно содержат одну или две HVR последовательности, выбранные из (i) HVR-H1, содержащей последовательность NISGTY (SEQ ID NO:7), и (ii) HVR-H3, содержащей последовательность WVGVGFYAMD (SEQ ID NO:9).
В различных вариантах этого аспекта изобретения любые из остатков HVR-H2, имеющие, как показано на Фигуре 57, значение F больше чем 1, 5 или 10, представляют собой остатки, которые, предпочтительно, сохраняются в виде такого же остатка, находящегося в том же самом положении HVR-H2 в bH1-44 или bH1-81 (SEQ ID NO:8 и 5, соответственно). В других вариантах изобретения любой из остатков HVR-H2, имеющий, как показано в Таблице 14, величину ΔΔG более 1, представляет собой остаток, который, предпочтительно, сохраняется в виде такого же остатка, находящегося в том же самом положении HVR-H2 в bH1-44 или bH1-81 (SEQ ID NO:8 и 5, соответственно).
В конкретных вариантах изобретения антитело включает HVR-L1 последовательность, содержащую NIAKTISGY (SEQ ID NO:1); HVR-L2 последовательность, содержащую WGSFLY (SEQ ID NO:2); HVR-L3 последовательность, содержащую HYSSPP (SEQ ID NO:3); HVR-H1 последовательность, содержащую NIKDTY (SEQ ID NO:4); HVR-H2 последовательность, содержащую RIYPTNGYTR (SEQ ID NO:5); и HVR-H3 последовательность, содержащую WGGDGFYAMD (SEQ ID NO:6), или включает HVR-L1 последовательность, содержащую NIAKTISGY (SEQ ID NO:1); HVR-L2 последовательность, содержащую WGSFLY (SEQ ID NO:2); HVR-L3 последовательность, содержащую HYSSPP (SEQ ID NO:3); HVR-H1 последовательность, содержащую NISGTY (SEQ ID NO:7); HVR-H2 последовательность, содержащую RIYPSEGYTR (SEQ ID NO:8); и/или HVR-H3 последовательность, содержащую WVGVGFYAMD (SEQ ID NO:9).
В другом конкретном (частном) варианте изобретения выделенное антитело включает области HVR-L1, HVR-L2, HVR-L3, HVR-H1, HVR-H2 и HVR-H3, причем каждая из этих областей, по порядку, содержит последовательность NIAKTISGY (SEQ ID NO:1); WGSFLY (SEQ ID NO:2); HYSSPP (SEQ ID NO:3); NIKDTY (SEQ ID NO:4); RIYPTNGYTR (SEQ ID NO:5) и WGGDGFYAMD (SEQ ID NO:6) и специфически связывает HER2 и VEGF. В другом конкретном варианте изобретения антитело включает области HVR-L1, HVR-L2, HVR-L3, HVR-H1, HVR-Н2 и HVR-H3, причем каждая из этих областей, по порядку, содержит последовательность NIAKTISGY (SEQ ID NO:1); WGSFLY (SEQ ID NO:2); HYSSPP (SEQ ID NO:3); NISGTY (SEQ ID NO:7); RIYPSEGYTR (SEQ ID NO:8) и WVGVGFYAMD (SEQ ID NO:9) и специфически связывает HER2 и VEGF.
В различных вариантах любого из аспектов по данному описанию антитело связывает человеческий и мышиный VEGF с Kd 150 нМ или сильнее, а HER2 с Kd 7 нМ или сильнее. В других вариантах изобретения антитело ингибирует индуцируемую VEGF клеточную пролиферацию и пролиферацию HER2-экспрессирующей клетки по сравнению с контролем. В конкретном варианте изобретения антитело связывает человеческий и мышиный VEGF с Kd 36 нМ или сильнее, а HER2 с Kd 1 нМ или сильнее. В дополнительном варианте изобретения антитело ингибирует связывание VEGF с VEGFR2.
В другом аспекте изобретение включает выделенный фрагмент антитела, который связывает человеческий и мышиный VEGF с Kd 58 нМ или сильнее, а HER2 с Kd 6 нМ или сильнее, и/или ингибирует VEGF-индуцируемую клеточную пролиферацию и пролиферацию HER2-экспрессирующей клетки по сравнению с контролем. В конкретном варианте изобретения фрагмент антитела связывает человеческий и мышиный VEGF с Kd 33 нМ или сильнее, а HER2 с Kd 0,7 нМ или сильнее. В другом конкретном варианте изобретения фрагмент представляет собой Fab фрагмент или одноцепочечный вариабельный фрагмент антитела (scFv).
В любом из вышеописанных аспектов антитело может представлять собой моноклональное антитело. В другом варианте всех вышеуказанных аспектов изобретения антитело может представлять собой IgG антитело. В дополнительных вариантах всех вышеуказанных аспектов изобретения по меньшей мере один участок каркасной последовательности антитела может представлять собой человеческую консенсусную каркасную последовательность.
В другом аспекте изобретение включает в качестве фрагмента антитела фрагмент любого из антител по данному описанию. Один вариант фрагмента антитела представляет собой фрагмент, включающий последовательность HVR-L1, содержащую последовательность NIAKTISGY (SEQ ID NO:1), которая специфически связывает HER2 и VEGF. В другом варианте изобретения фрагмент антитела дополнительно включает одну или две HVR последовательности, выбранные из (i) HVR-L2, содержащей последовательность WGSFLY (SEQ ID NO:2); и (ii) HVR-L3, содержащей последовательность HYSSPP (SEQ ID NO:3). В другом варианте изобретения фрагмент антитела дополнительно включает одну, две или три HVR последовательности, выбранные из (i) HVR-H1, содержащей последовательность NIKDTY (SEQ ID NO:4); (ii) HVR-H2, содержащей последовательность RIYPTNGYTR (SEQ ID NO:5); и (iii) HVR-НЗ, содержащей последовательность WGGDGFYAMD (SEQ ID NO:6). В другом варианте изобретения фрагмент антитела дополнительно включает одну, две или три HVR последовательности, выбранные из (i) HVR-H1, содержащей последовательность NISGTY (SEQ ID NO:7); (ii) HVR-H2, содержащей последовательность RIYPSEGYTR (SEQ ID NO:8); и (iii) HVR-Н3, содержащей последовательность WVGVGFYAMD (SEQ ID NO:9). В конкретных вариантах изобретения фрагмент антитела включает последовательность HVR-L1, содержащую NIAKTISGY (SEQ ID NO:1); последовательность HVR-L2, содержащую WGSFLY (SEQ ID NO:2); и последовательность HVR-L3, содержащую HYSSPP (SEQ ID NO:3); последовательность HVR-H1, содержащую NIKDTY (SEQ ID NO:4); последовательность HVR-H2, содержащую RIYPTNGYTR (SEQ ID NO:5); и последовательность HVR-Н3, содержащую WGGDGFYAMD (SEQ ID NO:6), или включает последовательность HVR-L1, содержащую NIAKTISGY (SEQ ID NO:1); последовательность HVR-L2, содержащую WGSFLY (SEQ ID NO:2); последовательность HVR-L3, содержащую HYSSPP (SEQ ID NO:3); последовательность HVR-H1, содержащую NISGTY (SEQ ID NO:7); последовательность HVR-H2, содержащую RIYPSEGYTR (SEQ ID NO:8); и последовательность HVR-Н3, содержащую WVGVGFYAMD (SEQ ID NO:9). В одном варианте изобретения фрагмент представляет собой Fab фрагмент или одноцепочечный вариабельный фрагмент антитела (scFv). В дополнительных вариантах всех вышеуказанных аспектов изобретения по меньшей мере один участок каркасной последовательности антитела может представлять собой человеческую консенсусную каркасную последовательность.
В других аспектах изобретение включает полинуклеотиды, кодирующие любое антитело или любой фрагмент антитела по данному описанию, а также вектор, содержащий такой полинуклеотид. В конкретных вариантах изобретения кодированное антитело включает HVR-L1 последовательность, содержащую NIAKTISGY (SEQ ID NO:1). Необязательно, или помимо этого, полинуклеотид кодирует антитело, которое также включает последовательность HVR-L2, содержащую WGSFLY (SEQ ID NO:2); и/или последовательность HVR-L3 содержащую HYSSPP (SEQ ID NO:3), или любую их комбинацию. В другом аспекте полинуклеотид может дополнительно кодировать антитело, включающее одну, две или три последовательности из HVR-H1 последовательности, содержащей NIKDTY (SEQ ID NO:4); последовательности HVR-H2, содержащей RIYPTNGYTR (SEQ ID NO:5); и последовательности HVR-Н3, содержащей WGGDGFYAMD (SEQ ID NO:6); или антитело, включающее одну, две или три последовательности из HVR-H1, содержащей NISGTY (SEQ ID NO:7); HVR-H2, содержащей RIYPSEGYTR (SEQ ID NO:8); и/или последовательности HVR-Н3, содержащей WVGVGFYAMD (SEQ ID NO:9).
В дополнительных аспектах изобретения полинуклеотид кодирует последовательность HVR-H1, содержащую NISGTY (SEQ ID NO:7), HVR-H2, содержащую последовательность RIYPSEGYTR (SEQ ID NO:8) или HVR-Н3, содержащую последовательность WVGVGFYAMD (SEQ ID NO:9), или любую их комбинацию.
В других аспектах изобретение включает выделенный полинуклеотид, кодирующий последовательность HVR-L1, содержащую последовательность NIAKTISGY (SEQ ID NO:1), и, необязательно, помимо этого, полинуклеотид кодирует одну, две или три HVR последовательности, выбранные из (i) HVR-H1, содержащей последовательность NIKDTY (SEQ ID NO:4); (ii) HVR-H2, содержащей последовательность RIYPTNGYTR (SEQ ID NO:5); и (iii) HVR-Н3, содержащей последовательность WGGDGFYAMD (SEQ ID NO:6). В дополнительных аспектах изобретение включает выделенный полинуклеотид, кодирующий последовательность HVR-L1, содержащую последовательность NIAKTISGY (SEQ ID NO:1); и (i) последовательность HVR-L2, содержащую последовательность WGSFLY (SEQ ID NO:2) или (ii) последовательность HVR-L3, содержащую последовательность HYSSPP (SEQ ID NO:3), или обе, или, необязательно, полинуклеотид дополнительно кодирует одну, две или три HVR последовательности, выбранные из (i) HVR-H1, содержащей последовательность NIKDTY (SEQ ID NO:4); (ii) HVR-H2, содержащей последовательность RIYPTNGYTR (SEQ ID NO:5); и (iii) HVR-Н3, содержащей последовательность WGGDGFYAMD (SEQ ID NO:6).
В другом аспекте изобретение включает выделенный полинуклеотид, кодирующий последовательность HVR-L1, содержащую последовательность NIAKTISGY (SEQ ID NO:1); HVR-H1, содержащую последовательность NISGTY (SEQ ID NO:7); HVR-H2, содержащую последовательность RIYPSEGYTR (SEQ ID NO:8); и HVR-Н3, содержащую последовательность WVGVGFYAMD (SEQ ID NO:9). Еще в одном аспекте изобретение включает выделенный полинуклеотид, кодирующий последовательность HVR-L1, содержащую последовательность NIAKTISGY (SEQ ID NO:1); последовательность HVR-L2, содержащую последовательность WGSFLY (SEQ ID NO:2); последовательность HVR-L3, содержащую последовательность HYSSPP (SEQ ID NO:3); HVR-H1, содержащую последовательность NISGTY (SEQ ID NO:7); HVR-H2, содержащую последовательность RIYPSEGYTR (SEQ ID NO:8); и HVR-Н3, содержащую последовательность WVGVGFYAMD (SEQ ID NO:9).
В других аспектах изобретение включает выделенный полинуклеотид, кодирующий последовательность HVR-H1, содержащую последовательность NISGTY (SEQ ID NO:7), выделенный полинуклеотид, кодирующий последовательность HVR-Н2, содержащую последовательность RIYPSEGYTR (SEQ ID NO:8), и выделенный полинуклеотид, кодирующий последовательность HVR-Н3, содержащую последовательность WVGVGFYAMD (SEQ ID NO:9). В другом аспекте изобретение включает выделенный полинуклеотид, кодирующий полипептид, включающий последовательность HVR-H1, содержащую последовательность NISGTY (SEQ ID NO:7); последовательность HVR-H2, содержащую последовательность RIYPSEGYTR (SEQ ID NO:8); и последовательность HVR-Н3, содержащую последовательность WVGVGFYAMD (SEQ ID NO:9).
В дополнительном варианте изобретения выделенный полинуклеотид кодирует последовательность HVR-L1, содержащую последовательность X1IX3X4X5X6X7X8X9Y (SEQ ID NO:83), где X1 обозначает любую аминокислоту, аспарагиновой кислоты, Х3 обозначает любую аминокислоту, за исключением пролина, Х4 обозначает любую аминокислоту, за исключением аргинина, и Х5 обозначает любую аминокислоту, за исключением серина. В другом варианте изобретения полинуклеотид кодирует последовательность HVR-L1, содержащую последовательность X1IX3X4X5X6X7X8X9Y (SEQ ID NO:83), где X1 обозначает любую аминокислоту, за исключением Asp, Х3 обозначает любую аминокислоту, за исключением пролина, Х4 обозначает любую аминокислоту, за исключением аргинина, и Х5 обозначает любую аминокислоту, за исключением серина; и последовательность HVR-L2, содержащую последовательность WGSFLY (SEQ ID NO:2), и/или последовательность HVR-L3, содержащую последовательность HYSSPP (SEQ ID NO:3). В дополнительных вариантах данного аспекта изобретения полинуклеотид кодирует антитело, содержащее последовательность X1IX3X4X5X6X7X8X9Y (SEQ ID NO:83), которая имеет аспарагин в X1, аланин в Х3, лизин в Х4, треонин в X5, серин в Х7 и/или глицин в X8, или любую их комбинацию. В различных вариантах этого аспекта изобретения любой из остатков HVR-L1, имеющий, как показано на Фигуре 57, значение F больше чем 1, 5 или 10, представляет собой остаток, который, предпочтительно, сохраняется в виде такого же остатка, находящегося в том же самом положении HVR-L1 в bH1-44 или bH1-81 (SEQ ID NO:1). В других вариантах изобретения любой из остатков HVR-L1, имеющий, как показано в Таблице 14, величину ΔΔG более 1, представляет собой остаток, который, предпочтительно, сохраняется в виде такого же остатка, находящегося в том же самом положении HVR-L1 в bH1-44 или bH1-81 (SEQ ID NO:1).
В дополнительном варианте изобретения выделенный полинуклеотид кодирует последовательность HVR-H2, содержащую последовательность RX2X3X4X5X6X7X8X9R (SEQ ID NO:85), где Х5 обозначает любую аминокислоту, за исключением треонина, а Х6 обозначает любую аминокислоту, за исключением аспарагина. В другом аспекте изобретение включает полинуклеотид, кодирующий последовательность HVR-H1, содержащую последовательность NISGTY (SEQ ID NO:7); последовательность HVR-Н2, содержащую последовательность RX2X3X4X5X6X7X8X9R (SEQ ID NO:85), в которой Х5 обозначает любую аминокислоту, за исключением треонина, а Х6 обозначает любую аминокислоту, за исключением аспарагина; и последовательность HVR-Н3, содержащую последовательность WVGVGFYAMD (SEQ ID NO:9). В дополнительных вариантах изобретения полинуклеотид кодирует последовательность HVR-H2, содержащую последовательность RX2X3X4X5X6X7X8X9R (SEQ ID NO:84), которая имеет серин в Х5, глутаминовую кислоту в Х6 и/или тирозин в Х8, или любую их комбинацию. В различных вариантах этого аспекта изобретения любые из остатков HVR-H2, имеющие, как показано на Фигуре 57, значение F больше чем 1, 5 или 10, представляют собой остатки, которые, предпочтительно, сохраняются в виде такого же остатка, находящегося в том же самом положении HVR-H2 в bH1-44 или bH1-81 (SEQ ID NO:8 и 5, соответственно). В других вариантах изобретения любой из остатков HVR-H2, имеющий, как показано в Таблице 14, величину ΔΔG более 1, представляет собой остаток, который, предпочтительно, сохраняется в виде такого же остатка, находящегося в том же самом положении HVR-H2 в bH1-44 или bH1-81 (SEQ ID NO:8 и 5, соответственно).
В других аспектах изобретение охватывает выделенный полипептид, включающий последовательность HVR-L1, содержащую последовательность NIAKTISGY (SEQ ID NO:1), или выделенный полипептид, включающий последовательность HVR-L1, содержащую последовательность NIAKTISGY (SEQ ID NO:1); последовательность HVR-L2, содержащую последовательность WGSFLY (SEQ ID NO:2); и/или последовательность HVR-L3, содержащую последовательность HYSSPP (SEQ ID NO:3). В другом аспекте изобретение охватывает полипептид, включающий последовательность HVR-L1, содержащую последовательность X1IX3X4X5X6X7X8X9Y (SEQ ID NO:83), в которой X1 обозначает любую аминокислоту, за исключением аспарагиновой кислоты, Х3 обозначает любую аминокислоту, за исключением пролина, Х4 обозначает любую аминокислоту, за исключением аргинина, и Х5 обозначает любую аминокислоту, за исключением серина. В другом варианте данного аспекта изобретения полипептид содержит HVR-L1 последовательность X1IX3X4X5X6X7X8X9Y (SEQ ID NO:83), где X1 обозначает любую аминокислоту, за исключением аспарагиновой кислоты, Х3 обозначает любую аминокислоту, за исключением пролина, Х4 обозначает любую аминокислоту, за исключением аргинина, и X5 обозначает любую аминокислоту, за исключением серина. Необязательно, помимо этого полипептид включает последовательность HVR-L2, содержащую последовательность WGSFLY (SEQ ID NO:2), и/или последовательность HVR-L3, содержащую последовательность HYSSPP (SEQ ID NO:3). В конкретных вариантах любого из вышеуказанных аспектов изобретения, которые включают полипептид, который содержит последовательность X1IX3X4X5X6X7X8X9Y (SEQ ID NO:83), в X1 находится аспарагин, в Х3 находится аланин, в Х4 находится лизин, в Х5 находится треонин, в Х7 находится серин и/или в X8 находится глицин или любая их комбинация. В различных вариантах этого аспекта изобретения любой из остатков HVR-L1, имеющий, как показано на Фигуре 57, значение F больше чем 1, 5 или 10, представляет собой остаток, который, предпочтительно, сохраняется в виде такого же остатка, находящегося в том же самом положении HVR-L1 в bH1-44 или bH1-81 (SEQ ID NO:1). В других вариантах изобретения любой из остатков HVR-L1, имеющий, как показано в Таблице 14, величину ΔΔG более 1, представляет собой остаток, который, предпочтительно, сохраняется в виде такого же остатка, находящегося в том же самом положении HVR-L1 в bH1-44 или bH1-81 (SEQ ID NO:1).
Изобретение также охватывает полипептид, включающий последовательность HVR-H2, содержащую последовательность RX2X3X4X5X6X7X8X9R (SEQ ID NO:85), где X5 обозначает любую аминокислоту, за исключением треонина, а Х6 обозначает любую аминокислоту, за исключением аспарагина. В другом аспекте изобретения полипептид включает последовательность HVR-H2 RX2X3X4X5X6X7X8X9R (SEQ ID NO:85), где Х5 обозначает любую аминокислоту, за исключением треонина, а Х6 обозначает любую аминокислоту, за исключением аспарагина, последовательность HVR-H1, содержащую последовательность NISGTY (SEQ ID NO:7), и последовательность HVR-H3, содержащую последовательность WVGVGFYAMD (SEQ ID NO:9). В различных вариантах вышеуказанных аспектов полипептид, включающий последовательность HVR-H2, содержащую последовательность RX2X3X4X5X6X7X8X9R (SEQ ID NO:84), имеет серин в Х5, глутаминовую кислоту в Х6 и/или тирозин в положении X8, или любую их комбинацию. В различных вариантах данного аспекта изобретения любые из остатков HVR-H2, имеющие, как показано на Фигуре 57, значение F больше чем 1, 5 или 10, представляют собой остатки, которые, предпочтительно, сохраняются в виде такого же остатка, находящегося в том же самом положении HVR-H2 в bH1-44 или bH1-81 (SEQ ID NO:8 и 5, соответственно). В других вариантах изобретения любой из остатков HVR-H2, имеющий, как показано в Таблице 14, величину ΔΔG более 1, представляет собой остаток, который, предпочтительно, сохраняется в виде такого же остатка, находящегося в том же самом положении HVR-H2 в bH1-44 или bH1-81 (SEQ ID NO:8 и 5, соответственно).
Изобретение охватывает также полипептид, содержащий одну, две или три HVR последовательности из последовательности HVR-H1, содержащей последовательность NISGTY (SEQ ID NO:7), последовательности HVR-H2, содержащей последовательность RIYPSEGYTR (SEQ ID NO:8), и/или последовательности HVR-Н3, содержащей последовательность WVGVGFYAMD (SEQ ID NO:9), или любую их комбинацию.
В любом из вышеуказанных аспектов выделенный полипептид может дополнительно содержать одну, две или три HVR последовательности из последовательности HVR-L1, содержащей последовательность NIAKTISGY (SEQ ID NO:1); HVR-H1, содержащей последовательность NIKDTY (SEQ ID NO:4); HVR-H2, содержащей последовательность RIYPTNGYTR (SEQ ID NO:5); и/или HVR-H3, содержащей последовательность WGGDGFYAMD (SEQ ID NO:6), или любую их комбинацию.
В любом из вышеуказанных аспектов выделенный полипептид может дополнительно содержать одну, две или три HVR последовательности из последовательности HVR-L1, содержащей последовательность NIAKTISGY (SEQ ID NO:1); последовательности HVR-L2, содержащей последовательность WGSFLY (SEQ ID NO:2); и/или последовательности HVR-L3, содержащей последовательность HYSSPP (SEQ ID NO:3), или любую их комбинацию.
В любом из вышеуказанных аспектов выделенный полипептид может дополнительно содержать HVR-H1, содержащую последовательность NIKDTY (SEQ ID NO:4); HVR-H2, содержащую последовательность RIYPTNGYTR (SEQ ID NO:5); и/или HVR-Н3, содержащую последовательность WGGDGFYAMD (SEQ ID NO:6), или любую их комбинацию.
В любом из вышеуказанных аспектов выделенный полипептид может дополнительно содержать одну, две или три HVR последовательности, выбранных из HVR-H1, содержащей последовательность NISGTY (SEQ ID NO:7); и последовательности HVR-H2, содержащей последовательность RIYPSEGYTR (SEQ ID NO:8); и/или HVR-Н3, содержащей последовательность WVGVGFYAMD (SEQ ID NO:9), или любую их комбинацию.
В дополнительных аспектах изобретение охватывает выделенный полипептид, включающий последовательность HVR-L1, содержащую последовательность NIAKTISGY (SEQ ID NO:1) и (i) последовательность HVR-L2, содержащую последовательность WGSFLY (SEQ ID NO:2), или (ii) последовательность HVR-L3, содержащую последовательность HYSSPP (SEQ ID NO:3), или и ту, и другую последовательность; и одну, две из трех HVR последовательностей, выбранных из (i) HVR-H1, содержащей последовательность NISGTY (SEQ ID NO:7); (ii) HVR-H2, содержащей последовательность RIYPSEGYTR (SEQ ID NO:8); и/или (iii) HVR-Н3, содержащей последовательность WVGVGFYAMD (SEQ ID NO:9), или любую их комбинацию.
В дополнительных аспектах изобретение охватывает выделенный полипептид, включающий последовательность HVR-L1, содержащую последовательность NIAKTISGY (SEQ ID NO:1) и (i) последовательность HVR-L2, содержащую последовательность WGSFLY (SEQ ID NO:2), или (ii) последовательность HVR-L3, содержащую последовательность HYSSPP (SEQ ID NO:3), или и ту, и другую последовательность; и одну, две из трех HVR последовательностей, выбранных из (i) HVR-H1, содержащей последовательность NIKDTY (SEQ ID NO:4); (ii) HVR-H2, содержащей последовательность RIYPTNGYTR (SEQ ID NO:5); и/или (iii) HVR-Н3, содержащей последовательность WGGDGFYAMD (SEQ ID NO:6), или любую их комбинацию.
В других аспектах изобретение включает выделенный полипептид, включающий последовательность HVR-H1, содержащую последовательность NISGTY (SEQ ID NO:7); выделенный полипептид, включающий последовательность HVR-H2, содержащую последовательность RIYPSEGYTR (SEQ ID NO:8); и выделенный полипептид, включающий последовательность HVR-H3, содержащую последовательность WVGVGFYAMD (SEQ ID NO:9). Еще в одном аспекте изобретение включает выделенный полипептид, включающий последовательность HVR-H1, содержащую последовательность NISGTY (SEQ ID NO:7); последовательность HVR-H2, содержащую последовательность RIYPSEGYTR (SEQ ID NO:8); и последовательность HVR-H3, содержащую последовательность WVGVGFYAMD (SEQ ID NO:9).
В одном варианте изобретение включает вектор, содержащий любой из вышеописанных полинуклеотидов по изобретению. В другом аспекте изобретение включает клетку-хозяина, содержащую любой из векторов по изобретению. В одном варианте изобретения клетка-хозяин является прокариотической клеткой. В другом варианте изобретения клетка-хозяин является эукариотической клеткой, например клеткой млекопитающего.
В другом аспекте изобретение включает способ получения любого из вышеописанных антител или фрагментов антител. Этот способ включает культивирование клетки-хозяина, которая содержит вектор, включающий полинуклеотид, кодирующий антитело, и извлечение антитела. В некоторых вариантах изобретения полинуклеотид кодирует последовательность HVR-L1, содержащую последовательность NIAKTISGY (SEQ ID NO:1), и, необязательно, полинуклеотид дополнительно кодирует HVR-H1, содержащую последовательность NIKDTY (SEQ ID NO:4); HVR-H2, содержащую последовательность RIYPTNGYTR (SEQ ID NO:5); и HVR-H3, содержащую последовательность WGGDGFYAMD (SEQ ID NO:6). В других вариантах изобретения полинуклеотид кодирует последовательность HVR-L1, содержащую последовательность NIAKTISGY (SEQ ID NO:1); последовательность HVR-L2, содержащую последовательность WGSFLY (SEQ ID NO:2); и последовательность HVR-L3, содержащую последовательность HYSSPP (SEQ ID NO:3), и, необязательно, полинуклеотид дополнительно HVR-H1, содержащую последовательность NIKDTY (SEQ ID NO:4); HVR-H2, содержащую последовательность RIYPTNGYTR (SEQ ID NO:5); и HVR-Н3, содержащую последовательность WGGDGFYAMD (SEQ ID NO:6). В другом варианте изобретения полинуклеотид кодирует последовательность HVR-L1, содержащую последовательность NIAKTISGY (SEQ ID NO:1); HVR-H1, содержащую последовательность NISGTY (SEQ ID NO:7); HVR-H2, содержащую последовательность RIYPSEGYTR (SEQ ID NO:8); HVR-Н3, содержащую последовательность WVGVGFYAMD (SEQ ID NO:9). Еще в одном варианте изобретения полинуклеотид кодирует последовательность HVR-L1, содержащую последовательность NIAKTISGY (SEQ ID NO:1); последовательность HVR-L2, содержащую последовательность WGSFLY (SEQ ID NO:2); последовательность HVR-L3, содержащую последовательность HYSSPP (SEQ ID NO:3); HVR-H1, содержащую последовательность NISGTY (SEQ ID NO:7); HVR-H2, содержащую последовательность RIYPSEGYTR (SEQ ID NO:8); и HVR-Н3, содержащую последовательность WVGVGFYAMD (SEQ ID NO:9).
В других вариантах изобретения полинуклеотид кодирует последовательность HVR-L1, содержащую последовательность NISGTY (SEQ ID NO:7), последовательность HVR-H2, содержащую последовательность RIYPSEGYTR (SEQ ID NO:8), или последовательность HVR-Н3, содержащую последовательность WVGVGFYAMD (SEQ ID NO:9). Еще в одном варианте изобретения полинуклеотид кодирует полипептид, включающий последовательность HVR-L1, содержащую последовательность NISGTY (SEQ ID NO:7), последовательность HVR-H2, содержащую последовательность RIYPSEGYTR (SEQ ID NO:8), и последовательность HVR-Н3, содержащую последовательность WVGVGFYAMD (SEQ ID NO:9).
В одном варианте изобретения клетка-хозяин является прокариотической клеткой, а в другом варианте изобретения клетка-хозяин является эукариотической клеткой, такой как клетка млекопитающего.
В другом аспекте изобретение включает способ лечения опухоли у субъекта. Этот способ включает введение субъекту антитела или фрагмента антитела по данному описанию, причем введение проводят в течение такого времени и в таком количестве, которые достаточны для лечения или предупреждения опухоли у субъекта. В одном варианте изобретения опухоль представляет собой колоректальную опухоль, рак молочной железы, рак легкого, почечноклеточную карциному, глиому, глиобластому или рак яичника. В другом варианте изобретения антитело включает последовательность HVR-L1, содержащую последовательность NIAKTISGY (SEQ ID NO:1), и специфически связывает HER2 и VEGF. В одном варианте изобретения антитело дополнительно включает HVR последовательности, выбранные из (i) HVR-L2, содержащей последовательность WGSFLY (SEQ ID NO:2), и/или (ii) HVR-L3, содержащей последовательность HYSSPP (SEQ ID NO:3). В другом варианте изобретения антитело включает одну, две или три HVR последовательности, выбранные из (i) HVR-H1, содержащей последовательность NIKDTY (SEQ ID NO:4); (ii) HVR-H2, содержащей последовательность RIYPTNGYTR (SEQ ID NO:5); и (iii) HVR-Н3, содержащей последовательность WGGDGFYAMD (SEQ ID NO:6). В дополнительном варианте изобретения антитело включает одну, две или три HVR последовательности, выбранные из (i) HVR-H1, содержащей последовательность NISGTY (SEQ ID NO:7); (ii) HVR-H2, содержащей последовательность RIYPSEGYTR (SEQ ID NO:8); и (iii) HVR-Н3, содержащей последовательность WVGVGFYAMD (SEQ ID NO:9). В конкретных вариантах изобретения антитело включает последовательность HVR-L1, содержащую NIAKTISGY (SEQ ID NO:1); последовательность HVR-L2, содержащую WGSFLY (SEQ ID NO:2); последовательность HVR-L3, содержащую HYSSPP (SEQ ID NO:3); последовательность HVR-H1, содержащую NIKDTY (SEQ ID NO:4); последовательность HVR-H2, содержащую RIYPTNGYTR (SEQ ID NO:5); и последовательность HVR-Н3, содержащую WGGDGFYAMD (SEQ ID NO:6), и специфически связывает HER2 и VEGF. В другом варианте изобретения антитело включает последовательность HVR-L1, содержащую NIAKTISGY (SEQ ID NO:1); последовательность HVR-L2, содержащую WGSFLY (SEQ ID NO:2); последовательность HVR-L3, содержащую HYSSPP (SEQ ID NO:3); последовательность HVR-H1, содержащую NISGTY (SEQ ID NO:7); последовательность HVR-H2, содержащую RIYPSEGYTR (SEQ ID NO:8); и последовательность HVR-Н3, содержащую WVGVGFYAMD (SEQ ID NO:9), и специфически связывает HER2 и VEGF.
В одном варианте способ дополнительно включает дополнительную противораковую терапию. В другом варианте изобретения дополнительная противораковая терапия включает введение другого антитела, химиотерапевтического агента, цитотоксического агента, антиангиогенного агента, иммуносупрессора, пролекарства, цитокина, антагониста цитокина, проведение цитотоксической лучевой терапии, введение кортикостероида, противорвотного средства, противораковой вакцины, анальгетика или ингибитора роста.
В другом варианте изобретения дополнительную противораковую терапию проводят до или после введения антитела. В другом варианте изобретения дополнительную противораковую терапию проводят одновременно с введением антитела.
В другом аспекте изобретение включает способ лечения аутоиммунного заболевания у субъекта. Этот способ включает введение субъекту антитела или фрагмента антитела по данному описанию, причем введение проводят в течение такого времени и в таком количестве, которые достаточны для лечения или предупреждения аутоиммунного заболевания у субъекта. В одном варианте изобретения антитело включает последовательность HVR-L1, содержащую последовательность NIAKTISGY (SEQ ID NO:1), и специфически связывает HER2 и VEGF. В одном варианте изобретения антитело включает одну или две HVR последовательности, выбранные из (i) HVR-L2, содержащей последовательность WGSFLY (SEQ ID NO:2); и (ii) HVR-L3, содержащей последовательность HYSSPP (SEQ ID NO:3). В другом варианте изобретения антитело включает одну, две или три HVR последовательности, выбранные из (i) HVR-H1, содержащей последовательность NIKDTY (SEQ ID NO:4); (ii) HVR-H2, содержащей последовательность RIYPTNGYTR (SEQ ID NO:5); и (iii) HVR-Н3, содержащей последовательность WGGDGFYAMD (SEQ ID NO:6). В дополнительном варианте изобретения антитело включает одну, две или три HVR последовательности, выбранные из (i) HVR-H1, содержащей последовательность NISGTY (SEQ ID NO:7); (ii) HVR-H2, содержащей последовательность RIYPSEGYTR (SEQ ID NO:8); и (iii) HVR-Н3, содержащей последовательность WVGVGFYAMD (SEQ ID NO:9). В конкретных вариантах изобретения антитело включает последовательность HVR-L1, содержащую NIAKTISGY (SEQ ID NO:1); последовательность HVR-L2, содержащую WGSFLY (SEQ ID NO:2); последовательность HVR-L3, содержащую HYSSPP (SEQ ID NO:3); последовательность HVR-H1, содержащую NIKDTY (SEQ ID NO:4); последовательность HVR-H2, содержащую RIYPTNGYTR (SEQ ID NO:5); и последовательность HVR-Н3, содержащую WGGDGFYAMD (SEQ ID NO:6), и специфически связывает HER2 и VEGF; или антитело включает последовательность HVR-L1, содержащую NIAKTISGY (SEQ ID NO:1); последовательность HVR-L2, содержащую WGSFLY (SEQ ID NO:2); последовательность HVR-L3, содержащую HYSSPP (SEQ ID NO:3); последовательность HVR-H1, содержащую NISGTY (SEQ ID NO:7); последовательность HVR-H2, содержащую RIYPSEGYTR (SEQ ID NO:8); и последовательность HVR-Н3, содержащую WVGVGFYAMD (SEQ ID NO:9), и специфически связывает HER2 и VEGF.
Еще в одном аспекте изобретение охватывает способ лечения незлокачественного заболевания, включающего патологическую активацию HER2 у субъекта. Этот способ включает введение субъекту антитела или фрагмента антитела по данному описанию, причем введение проводят в течение такого времени и в таком количестве, которые достаточны для лечения или предупреждения незлокачественного заболевания у субъекта. В одном варианте изобретения антитело включает последовательность HVR-L1, содержащую последовательность NIAKTISGY (SEQ ID NO:1), и специфически связывает HER2 и VEGF. В одном варианте изобретения антитело включает одну или две HVR последовательности, выбранные из (i) HVR-L2, содержащей последовательность WGSFLY (SEQ ID NO:2); и (ii) HVR-L3, содержащей последовательность HYSSPP (SEQ ID NO:3). В другом варианте изобретения антитело дополнительно включает одну, две или три HVR последовательности, выбранные из (i) HVR-H1, содержащей последовательность NIKDTY (SEQ ID NO:4); (ii) HVR-H2, содержащей последовательность RIYPTNGYTR (SEQ ID NO:5); и (iii) HVR-Н3, содержащей последовательность WGGDGFYAMD (SEQ ID NO:6). В дополнительном варианте изобретения антитело включает одну, две или три HVR последовательности, выбранные из (i) HVR-H1, содержащей последовательность NISGTY (SEQ ID NO:7); (ii) HVR-H2, содержащей последовательность RIYPSEGYTR (SEQ ID NO:8); и (iii) HVR-Н3, содержащей последовательность WVGVGFYAMD (SEQ ID NO:9). В конкретных вариантах изобретения антитело включает последовательность HVR-L1, содержащую NIAKTISGY (SEQ ID NO:1); последовательность HVR-L2, содержащую WGSFLY (SEQ ID NO:2); последовательность HVR-L3, содержащую HYSSPP (SEQ ID NO:3); последовательность HVR-H1, содержащую NIKDTY (SEQ ID NO:4); последовательность HVR-H2, содержащую RIYPTNGYTR (SEQ ID NO:5); и последовательность HVR-Н3, содержащую WGGDGFYAMD (SEQ ID NO:6), и специфически связывает HER2 и VEGF; или антитело включает последовательность HVR-L1, содержащую NIAKTISGY (SEQ ID NO:1); последовательность HVR-L2, содержащую WGSFLY (SEQ ID NO:2); последовательность HVR-L3, содержащую HYSSPP (SEQ ID NO:3); последовательность HVR-H1, содержащую NISGTY (SEQ ID NO:7); последовательность HVR-H2, содержащую RIYPSEGYTR (SEQ ID NO:8); и последовательность HVR-Н3, содержащую WVGVGFYAMD (SEQ ID NO:9), и специфически связывает HER2 и VEGF.
Дополнительные аспекты изобретения включают применение антител и фрагментов антител по данному описанию для лечения опухоли, аутоиммунного заболевания или незлокачественного заболевания, включающего патологическую активацию HER2 у субъекта, а также применение для получения лекарственного препарата для лечения опухоли, аутоиммунного заболевания или незлокачественного заболевания, включающего патологическую активацию HER2 у субъекта. В одном варианте такого применения антитело включает последовательность HVR-L1, содержащую последовательность NIAKTISGY (SEQ ID NO:1), и специфически связывает HER2 и VEGF. В одном варианте изобретения антитело включает одну или две HVR последовательности, выбранные из (i) HVR-L2, содержащей последовательность WGSFLY (SEQ ID NO:2); и (ii) HVR-L3, содержащей последовательность HYSSPP (SEQ ID NO:3). В другом варианте изобретения антитело дополнительно включает одну, две или три HVR последовательности, выбранные из (i) HVR-H1, содержащей последовательность NIKDTY (SEQ ID NO:4); (ii) HVR-H2, содержащей последовательность RIYPTNGYTR (SEQ ID NO:5); и (iii) HVR-Н3, содержащей последовательность WGGDGFYAMD (SEQ ID NO:6). В дополнительном варианте изобретения антитело включает одну, две или три HVR последовательности, выбранные из (i) HVR-H1, содержащей последовательность NISGTY (SEQ ID NO:7); (ii) HVR-H2, содержащей последовательность RIYPSEGYTR (SEQ ID NO:8); и (iii) HVR-Н3, содержащей последовательность WVGVGFYAMD (SEQ ID NO:9). В конкретных вариантах изобретения антитело включает последовательность HVR-L1, содержащую NIAKTISGY (SEQ ID NO:1); последовательность HVR-L2, содержащую WGSFLY (SEQ ID NO:2); последовательность HVR-L3, содержащую HYSSPP (SEQ ID NO:3); последовательность HVR-H1, содержащую NIKDTY (SEQ ID NO:4); последовательность HVR-H2, содержащую RIYPTNGYTR (SEQ ID NO:5); и последовательность HVR-Н3, содержащую WGGDGFYAMD (SEQ ID NO:6), и специфически связывает HER2 и VEGF; или антитело включает последовательность HVR-L1, содержащую NIAKTISGY (SEQ ID NO:1); последовательность HVR-L2, содержащую WGSFLY (SEQ ID NO:2); последовательность HVR-L3, содержащую HYSSPP (SEQ ID NO:3); последовательность HVR-H1, содержащую NISGTY (SEQ ID NO:7); последовательность HVR-H2, содержащую RIYPSEGYTR (SEQ ID NO:8); и последовательность HVR-Н3, содержащую WVGVGFYAMD (SEQ ID NO:9), и специфически связывает HER2 и VEGF.
В одном варианте способов лечения опухоли, аутоиммунного заболевания или незлокачественного заболевания, включающего патологическую активацию HER2, по данному описанию субъектом является человек.
Также рассматриваются наборы, композиции и изделия, содержащие антитела или фрагменты антител по данному описанию.
Краткое описание Фигур
На Фигуре 1 изображено спланированное разнообразие в различных библиотеках LC (легких цепей).
На Фигуре 2 даны краткие сведения из четырех библиотек легких цепей, используемых для изменения антител против VEGF или антител против Her2 с целью связывания с дополнительной мишенью. Выделенные курсивом NNK и XYZ относятся к наборам кодонов. Ys, Ds, Ts и Ss относятся к программной рандомизации при условии, что тирозин, аспарагиновая кислота, треонин и серин, соответственно, встречаются в течение 50% времени, а любая из 20 аминокислот встречается в течение других 50% времени. D/Ds и T/Ts относятся к программной рандомизации при условии, что D или Т, соответственно, встречаются в течение 75% времени, а любая из 20 аминокислот встречается в течение других 25% времени.
На Фигуре 3 показаны последовательности НС, LC CDR остатков матриц легкой цепи.
На Фигуре 4 показано естественное и спланированное разнообразие CDR легкой цепи. В каждом положении последовательность антитела Герцептин® показана в скобках. "*" обозначает, что инсерция отсутствует в антителе Герцептин®.
На Фигурах 5А и 5В1-5В2 показаны последовательности специфических антигенсвязывающих клонов, выделенных из библиотеки легкой цепи (LC). На Фигуре 5А показаны последовательности LC CDR моноспецифических фаговых клонов, связывающихся с VEGF, DR5 и Fc, а на Фигуре 5В показаны биспецифические Fab, связывающиеся с VEGF/HER2, DR5/HER2 и Fc/HER2. Последовательности каркасной области легкой цепи и тяжелой цепи соответствуют последовательностям антитела Герцептин®, за исключением замены R66G в каркасной области LC.
На Фигуре 6 представлена диаграмма, показывающая специфичность связывания антител, полученных из LC библиотеки. Показаны результаты для антител bH1, bH3, 3-1, bD1, bD2, 4-1 и 4-5. Связанные IgG антитела детектируют спектрофотометрическими методами (оптическая плотность при 450 нм, ось y). В анализе используют следующие белки (слева направо для каждого антитела): человеческий сосудистый эндотелиальный фактор роста A (hVEGF-А (СЭФР-А)), hVEGF-C, hVEGF-D, внеклеточный домен (ECD) hHER2, внеклеточный домен рецептора эпидермального фактора роста (hEGFR), человеческий "рецептор смерти 5" (hDR5), бычий сывороточный альбумин (BSA), казеин, фетальную бычью сыворотку (FBS), лизат клеток WIL2 и лизат клеток NR6.
На Фигуре 7 показаны условия сортинга и обогащение библиотеки С и D.
На Фигуре 8 показаны VEGF-связующие. Остатки 28, 30, 30а, 31, 92, 93 и 93а совершенно различны. Остатки 32, 50, 53, 91 и 94 рестриктированы. Остатки 29, 33 и 51 ограничены (<3).
На Фигуре 9 показаны связующие человеческого VEGF, совместная селекция в планшете и растворе.
На Фигурах 10А и 10В показаны клоны, которые связывают как VEGF, так и HER2.
На Фигуре 11 показаны клоны, которые связывают только VEGF и утрачивают активность связывания с HER2 (HER2-связывающая активность).
На Фигуре 12 показаны клоны, связывающиеся с VEGF.
На Фигурах 13А и 13В показаны клоны, которые блокируют связывание VEGF с VEGFR1-D2 или D1.
На Фигурах 14А и 14В показаны VEGF-связующие и аффинности VEGF-связующих из библиотеки L1/L2/L3-C,D.
На Фигуре 15 показаны клоны, которые могут связывать как hVEGF, так и HER2.
На Фигуре 16 показаны связующие из LC библиотеки, используемые при образовании scFv'2 и визуализуемые на фаге (фаговый дисплей).
На Фигуре 17 показана экспрессия различных клонов в Fab или hIgG форме.
На Фигурах 18А и 18В показаны анализы ELISA (твердофазный ИФА, иммуноферментные анализы) клонов при связывании hIgG формы с hVEGF165.
На Фигуре 19 показаны анализы ELISA клонов при связывании hIgG формы с иммобилизованными белками-мишенями.
На Фигуре 20 показаны конкурентные анализы ELISA клонов в hIgG форме в присутствии Her2 и VEGF или DR5.
На Фигуре 21 показан анализ связывания с VEGF или HER2 методом Biacore.
На Фигуре 22 показано связывание HER2-ECD или hVEGF с IgG или Fab, имеющим легкую цепь, полученную из другого связывающего (связующего) клона.
На Фигурах 23А и 23В показано антитело против VEGF, блокирующее взаимодействие VEGF с VEGFR1 D1-3 и KDR D1-7.
На Фигуре 24 показаны антитела, блокирующие связывание В20-4.1 и VEGF.
На Фигуре 25 показаны антитела, блокирующие связывание антитела Авастин и VEGF.
На Фигуре 26 показаны кристаллические структуры биспецифического bH1 Fab, связанного с HER2 или VEGF.
На Фигуре 27 представлена диаграмма, показывающая, что антитела против VEGF блокируют связывание hVEGF с VEGF рецептором 2 (VEGFR2).
На Фигуре 28 показаны кристаллические структуры биспецифического bH1 Fab, связанного с HER2 или VEGF.
На Фигуре 29 представлен ряд круговых диаграмм, показывающих вклад отдельных CDR в структуру-паратоп для bH1. Размер (площадь) паратопа для VEGF составляет 730Å2, а для HER2 составляет 690Å2. Домены CDR тяжелой цепи показаны серым, а CDR легкой цепи показаны белым.
На Фигуре 30 показана суперпозиция CDR петель VEGF/HER2-связанного bH1 антитела или HER2-связанного антитела Герцептина® в той же ориентации, что и на Фигуре 28.
На Фигуре 31 показаны кристаллические структуры биспецифического bH1 Fab, связанного с HER2 или VEGF. CDR-L1 двух комплексов bH1 показаны в одной и той же ориентации.
На Фигуре 32 показаны энергетически важные сайты связывания bH1 для связывания VEGF и HER2.
На Фигуре 33 показаны кодоны bH1, сканированные методом дробовика (shotgun).
На Фигуре 34 показана конструкция библиотеки.
На Фигуре 35 показан клон антител с мутациями в результате сканирования методом дробовика, скрининг которых осуществляют с помощью связывания с VEGF.
На Фигуре 36 показан клон антител с мутациями в результате сканирования методом дробовика, скрининг которых осуществляют с помощью связывания с HER2.
На Фигурах 37А-37D показаны результаты аланин-сканирующего мутагенеза. На Фигурах 37А и 37В показаны результаты сканирования аланином bH1 для связывания (Фигура 37А) VEGF или связывания (Фигура 37В) HER2 и результаты сканирования гомологом bH1 для связывания (Фигура 37С) VEGF или для связывания (Фигура 37D) HER2.
На Фигуре 38 показаны результаты аланин-сканирующего мутагенеза (сканирования аланином) мутантов bH1 или антитела Герцептина®.
На Фигурах 39А1-39А3 и 39В1-39В3 показан аланин- и гомолог сканирующий мутагенез методом дробовика bH1 Fab для связывания с VEGF и HER2.
На Фигурах 40А-40В показаны энергетически важные сайты связывания bH1 для связывания VEGF и HER2.
На Фигуре 41 показаны последовательности зрелого по VEGF-аффинности клона bH1 и аффинность связывания с VEGF или HER2.
На Фигуре 42 показано ингибирование VEGF-индуцированной пролиферации клеток HUVEC антителами против VEGF.
На Фигуре 43 показано связывание биспецифических антител с HER2, экспрессируемым на NR6 клетках.
На Фигуре 44 показаны результаты экспериментов по конкурентному связыванию для bH1 с VEGF или HER2.
На Фигуре 45 показано, что bH1 и варианты с повышенной аффинностью bH1-44 и bH1-81 IgG ингибируют опосредуемую HER2 и VEGF клеточную пролиферацию in vitro.
На Фигуре 46 показана специфичность связывания биспецифических антител, полученных из LC библиотеки.
На Фигуре 47 показано, что антитела против VEGF блокируют связывание VEGF с VEGFR2 рецепторами. На Фигуре 47А показано связывание человеческого VEGF, а на Фигуре 47В показано связывание мышиного VEGF.
На Фигурах 48А и 48В показано, что VEGF и HER2 конкурируют за связывание с bH1-44 биспецифическим IgG в растворе.
На Фигурах 49А и 49В показано, что биспецифические антитела bH1 и bH1-44 связываются с экспрессирующими HER2 клетками мышиных фибробластов (NR6; Фигура 49В), но не с HER2-негативными NR6 клетками (Фигуре 49А).
На Фигуре 50 показано, что биспецифическое bH1 антитело вызывает специфическую иммунопреципитацию VEGF или HER2, но не других белков, из лизатов мышиных фибробластов (NR6).
На Фигуре 51 показано ингибирование опухоли антителом bH1-44 в ксенотрансплантатах Colo205 и ВТ474М1 у мышей с ослабленным иммунитетом.
На Фигурах 52А, 52В и 53 изображены примеры акцепторных человеческих консенсусных каркасных последовательностей для практического применения настоящего изобретения с нижеприведенными условными обозначениями (идентификаторами) последовательностей:
Консенсусные каркасные последовательности для вариабельной области тяжелой цепи (VH) (Фиг.52А и 52В)
консенсусные последовательности каркасных участков FR1, FR2, FR3 и FR4 человеческой VH подгруппы I без CDR по Kabat (IA: SEQ ID NO:42-45, соответственно)
консенсусные последовательности каркасных участков FR1, FR2, FR3 и FR4 человеческой VH подгруппы I без удлиненных гипервариабельных областей (IB: SEQ ID NOS: 46, 47, 44 и 45, соответственно; IC: SEQ ID NO:46-48 и 45, соответственно; ID: SEQ ID NOS: 42, 47,49 и 45, соответственно)
консенсусные последовательности каркасных участков FR1, FR2, FR3 и FR4 человеческой VH подгруппы II без CDR по Kabat (IIA: SEQ ID NO:50-52 и 45, соответственно)
консенсусные последовательности каркасных участков FR1, FR2, FR3 и FR4 человеческой VH подгруппы II без удлиненных гипервариабельных областей (IIB: SEQ ID NO:53, 54, 52 и 45, соответственно; IIC: SEQ ID NO:53-55 и 45, соответственно; IID: SEQ ID NO:53, 54, 56 и 45, соответственно)
консенсусные последовательности каркасных участков FR1, FR2, FR3 и FR4 человеческой VH подгруппы III без CDR пo Kabat (IIIA: SEQ ID NO:57-59 45, соответственно)
консенсусные последовательности каркасных участков FR1, FR2, FR3 и FR4 человеческой VH подгруппы III без удлиненных гипервариабельных областей (IIIB: SEQ ID NO:60, 61, 59 и 45, соответственно; IIIC: SEQ ID NOS: 60-62 и 45, соответственно; HID: SEQ ID NOS: 60, 61, 63 и 45, соответственно)
акцепторные каркасные участки FR1, FR2, FR3 и FR4 человеческой VH без CDR по Kabat (Акцептор A: SEQ ID NO:64, 58, 65 и 45, соответственно)
акцепторные каркасные участки FR1, FR2, FR3 и FR4 человеческой VH без удлиненных гипервариабельных областей (Акцептор В: SEQ ID NO:60, 61, 65 и 45, соответственно; Акцептор С: SEQ ID NO:60, 61, 66 и 45, соответственно)
каркасные участки FR1, FR2, FR3 и FR4 акцептора 2 человеческой VH без CDR по Kabat (Второй акцептор A: SEQ ID NO:64, 58, 67 и 45, соответственно)
каркасные участки FR1, FR2, FR3 и FR4 акцептора 2 человеческой VH без удлиненных гипервариабельных областей (Второй акцептор В: SEQ ID NO:60, 61, 67 и 45, соответственно; Второй акцептор С: SEQ ID NO:60, 61, 68 и 45, соответственно; Второй акцептор D: SEQ ID NO:60, 61, 69 и 45, соответственно)
Консенсусные каркасные последовательности для вариабельной области легкой цепи (VH) (Фиг.53)
консенсусные последовательности каркасных участков FR1, FR2, FR3 и FR4 человеческой VL каппа подгруппы I (kv1: SEQ ID NO:70-73, соответственно)
консенсусные последовательности каркасных участков FR1, FR2, FR3 и FR4 человеческой VL каппа подгруппы II (kv2: SEQ ID NO:74-76 и 73, соответственно)
консенсусные последовательности каркасных участков FR1, FR2 и FR3 человеческой VL каппа подгруппы III (kv3: SEQ ID NO:77-79 и 73, соответственно)
консенсусные последовательности каркасных участков FR1, FR2 и FR3 человеческой VL каппа подгруппы IV (kv4: SEQ ID NO:80-82 и 73, соответственно)
На Фигуре 54 показаны остатки, которые имеют структурные контакты или энергетически взаимодействуют с HER2, VEGF или и с тем, и с другим. Остатки, которые имеют структурные контакты (>25% "с(о)крыто", экранировано) или энергетическое взаимодействие (ΔΔG>10% общей энергии связывания) с HER2 (светло-серый), VEGF (серый) или с обоими (общий, черный), картированы на поверхности HER2-связанного bH1.
На Фигуре 55 показаны области контакта (интерфейсы, обеспечивающие связывание, интерфейсы связывания) bH1/VEGF и bH1/HER2. Увеличенное изображение интерфейса связывания bH1/VEGF (А) и bH1/HER2 (В) иллюстрирует структурные различия между VEGF и HER2 в областях связывания антител. Изображения поверхностей VEGF (С) и HER2-ECD (D) показаны в одной и той же ориентации относительно bH1 Fab. Остатки, контактирующие с bH1 Fab (расстояние меньше 4.5 Å), выделены. С точки зрения химического состава или топологии очевидного сходства между двумя эпитопами для bH1 не наблюдается.
На Фигуре 56 показано, что антитела bH1 и bH1-44 блокируют связывание человеческого VEGF с VEGFR1. Биотинилированный человеческий VEGF165 инкубируют с увеличивающимися концентрациями IgG (ось x), затем улавливают иммобилизованным человеческим VEGFR1-Fc и детектируют с помощью конъюгата пероксидаза хрена - стрептавидин с добавлением субстрата (нормализованная % 00450, ось y).
На Фигуре 57 показаны результаты сканирования аланином bH1 и bH1-44 мутантов. Аланин-сканирующий мутагенез позволяет идентифицировать функционально важные остатки для связывания VEGF и/или HER2. Значения F представляют собой относительный вклад каждого сканированного остатка в связывание антигена. Величину F определяют для связывания bH1-44 с VEGF и HER2 (черные столбцы) и сравнивают с величиной F для bH1 (бесцветные столбцы). Аминокислоты в скобках представляют собой остатки в bH1-44, которые отличаются от bH1. Эта диаграмма основана на результатах, показанных на Фигуре 56.
На Фигуре 58 показано связывание bH1-44 I29A Y32A bH1-44 и R50A R58A bH1-44 антител с VEGF и HER2. Анализы связывания методом ELISA (твердофазным ИФА, иммуноферментным анализом) показывают способность bH1-44 IgG и двух двойных мутантов связываться с биотинилированным VEGF109 (слева) или HER2-ECD (справа) и конкурировать с иммобилизованным антителом против VEGF или Герцептином, соответственно. У мутанта I29A/Y32A LC наблюдается утрата связывания VEGF, но при этом он сохраняет аффинность к HER2, аналогичную аффинности bH1-44. Мутант R50A/R58A НС утрачивает аффинность к HER2, но сохраняет связывание с VEGF.
На Фигуре 59 показаны результаты калориметрических измерений изменений энтальпии, обусловленных связыванием антигена. На Фигурах 59А-F показаны данные для связывания bH1 с VEGF, связывания bH1 с HER2, связывания bH1-44 с VEGF, связывания bH1-44 с HER2, связывания bH1-44 НС-R50A+R58A с VEGF, связывания bH1-44 LC-I29A+Y32A с HER2, соответственно. На Фигурах показаны отдельные тепловые импульсы (вверху) и величины теплоты реакции (внизу), рассчитанные суммированием (интегрированием) каждого импульса, изображенные в виде графика зависимости от отношения антитела к антигену к концу введения инъекций. Ввиду малой величины изменений энтальпии требуются сравнительно высокие концентрации белка, что препятствует точному определению KD в случае высокой аффинности. На Фигурах 59А-D: Растворы VEGF109 или HER2-ECD в концентрациях 10-20 мкМ титруют с помощью 15 инъекций bH1 или bH1-44 Fab в концентрациях от 100 до 200 мкМ. Фигуры 59Е-F: Растворы VEGF109 или HER2-ECD в концентрациях 10-20 мкМ титруют с помощью 20 инъекций bH1-44 LC-I29A+Y32A Fab или bH1-44 HC-R50A+R58A Fab в концентрациях 150 и 250 мкМ. Титрования номер 1 и 13 на (Фигуре 59Е) исключают из анализа ввиду аппаратурного шума.
На Фигуре 60 показаны термодинамические характеристики bH1 вариантов и антитела Герцептина®. Каждый двойной специфический вариант (bH1, bH1-81 и bH1-44) имеет термодинамические характеристики, отличающиеся предпочтительной энтальпией и энтропией для связывания как VEGF, так и HER2. Варианты HC-R50A+R58A и LC-129A+Y32A, которые утратили аффинность к HER2 или VEGF, соответственно, демонстрируют термодинамические характеристики, аналогичные bH1-44. Термодинамические характеристики взаимодействия bH1-44/HER2 отличаются от термодинамических характеристики взаимодействия Герцептин/HER2.
На Фигуре 61 дано сравнение "горячих точек" (hotspot) антител bH1, bH1-44 и Герцептина® для связывания HER2 на основе результатов аланин-сканирующего мутагенеза. Остатки в "горячих точках" выделены серым цветом, отображенным на структуре Герцептина® или структурах bH1 Fab (bH1, bH1-44). "Горячие точки" (хотспоты, hotspots) определяют как сайты, в которых величина ΔΔG больше или равна 10% от общей свободной энергии связывания (ΔG). Сайты контакта структур (в пределах 4.5 Å от антигенов в структурах) обведены светлыми пунктирными линиями. НС и LC разделены черным пунктиром. Подчеркнутые остатки отличаются в последовательности от Герцептина®.
На Фигуре 62 показаны вычисленные изменения теплоемкости, обусловленные связыванием bH1-44 Fab с VEGF или HER2. ΔСр определяют по тангенсу угла наклона зависимости ДН от температуры между 20°С и 37°С. В этом интервале ΔСр, по видимому, не зависит от Т, если исходить из линейной зависимости между ΔН и Т (R=0.991 для bH1-44/HER2, R=0.9989 для bH1-44/VEGF). Величина ΔСр для связывания Герцептин®/НЕР2 была определена ранее Kelley et al. (Biochemistry, 1992).
На Фигурах 63А-В показана кинетика связывания bH1-44 вариантов, определяемая методом BIAcore. На Фигурах показано наложение типичных графиков зависимости ответа от времени для связывающих взаимодействий между иммобилизованным (A) VEGF109 или (В) HER2-ECD и 0.5 мкМ растворами bH1-44 Fab (А и В: вверху), bH1-44-LC-Y32 (А: второй снизу; В: второй сверху), bH1-44-LC-I29A+Y32A (А: внизу; В: второй снизу) и bH1-44-НС-R50A+R58A (А: второй сверху; В: внизу). Линии обозначают связывание с тем же самым иммобилизованным чипом СМ5, который регенерируется после каждого опыта с Fab. He обнаружено никакого связывания bH1-44-LC-I29A+Y32A с VEGF или bH1-44-HC-R50A+R58A с HER2 при концентрации 0.5 мкМ. Вариант bH1-44-Y32A показывает заметно более слабое связывание с VEGF по сравнению с bH1-44 дикого типа.
На Фигурах 64А-D показано картирование остатков, определяющих специфичность антитела bH1-44 на кристаллической структуре bH1. Остатки, важные для связывания VEGF (LC-I29 и LC-Y32: А и В), и остатки, важные для связывания HER (НС-R50 и НС-R58; С и D), показаны темно-серым цветом в виде полосок (стержней) на кристаллических структурах bH1/VEGF (А и С, разрешение 2.6 Å) или bH1/HER2 (В и D, разрешение 2.9 Å). Остатки I29 и Y32, по-видимому, участвуют во внутрицепных взаимодействиях, которые служат для сохранения конформации петли CDR-L1, необходимой для связывания с VEGF. I29 доступен для растворителя (является гидрофильным) в структуре HER2. Y32 расположен ("упакован") рядом с HER2, но не участвует в продуктивном контакте с антигеном. R50 и R58 расположены ("упакованы") рядом с D560 и Е558 на HER2 и, по-видимому, участвуют во взаимодействиях зарядов (заряд-заряд). R50 and R58 доступны для растворителя (являются гидрофильными) в структуре VEGF в растворе.
На Фигуре 65 показана экспрессия Fabs мутантов Герцептина® (R50A, R58A, и R50A/R58A).
Подробное описание изобретения
Настоящее изобретение включает способы получения полиспецифических антител и фрагментов антител, а также антитела, идентифицированные этими способами, и их применение. Как правило, способы по изобретению включают создание разнообразия (диверсификации) вариабельного домена легкой цепи или вариабельного домена тяжелой цепи антитела с целью получения вариантов, которые могут стабильно экспрессироваться в библиотеке. Затем из этой библиотеки выбирают "диверсифицированные" антитела, способные специфически связывать два эпитопа, и подробно их характеризуют.
Примеры антител, идентифицированных этими способами по изобретению, включают антитела, которые связывают как HER2 (рецептор человеческого эпидермального фактора роста 2), так и VEGF (сосудистый эндотелиальный фактор роста). В частности, результаты, представленные в данном описании, например, в нижеприведенных Примерах, показывают, что мутации в гипервариабельных областях (областях, определяющих комплементарность, CDR) легкой цепи антитела к HER2 сообщает способность к двойственному (двойному) связыванию неродственных белковых антигенов, а также HER2. Одно биспецифическое высокоаффинное HER2/VEGF антитело очень хорошо охарактеризовано (тщательно изучено). Помимо этого продемонстрированы кристаллические структуры этого биспецифического Fab в комплексе с HER2 и VEGF и методом мутагенеза определен энергетический вклад остатков Fab. Сайты связывания для двух антигенов в значительной степени перекрываются; большинство CDR остатков, которые контактируют с HER2, также участвуют в контактах с VEGF. Однако энергетически остатки тяжелой цепи оказывают преобладающее влияние на специфичность к HER2, тогда как на специфичность к VEGF влияние легкой цепи является доминирующим.
Биспецифическое антитело к HER2/VEGF ингибирует in vitro и in vivo клеточную пролиферацию, опосредуемую как HER2, так и VEGF. Эти результаты демонстрируют, что изменение последовательности вариабельного домена легкой цепи антитела может дать антитела с двойной специфичностью и функцией. Например, bH1-44 и bH1-81 можно нацеливать на два механизма прогрессирования опухоли: пролиферацию опухолевых клеток, опосредуемую HER2, и на опухолевый ангиогенез, опосредуемый VEGF. Совместное нацеливание одного антитела на два антигена представляет собой альтернативу комбинированной терапии.
I. Определения
Термин "полиспецифическое антитело" употребляется в самом широком смысле и конкретно охватывает антитело, содержащее вариабельный домен тяжелой цепи (VH) и вариабельный домен легкой цепи (VL), где структурный элемент (сегмент) VHVL имеет полиэпитопную специфичность (т.е. способен связываться с двумя различными эпитопами на одной биологической молекуле или с каждым эпитопом на различных биологических молекулах). Такие полиспецифические антитела включают, но без ограничения, полноразмерные антитела, антитела, имеющие два или более VL и VH доменов, фрагменты антител, такие как Fab, Fv, dsFv, scFv, диатела, биспецифические диатела и триатела, фрагменты антител, связанные ковалентной или нековалентной связью. Выражение "полиэпитопная специфичность" относится к способности специфически связываться с двумя или более различных эпитопов на одной и той же мишени или на различных мишенях. Термин "моноспецифический" относится к способности связывать только один эпитоп. Согласно одному варианту изобретения полиспецифическое антитело представляет собой IgG1 форму и связывается с каждым эпитопом с аффинностью от 5 мкМ до 0.001 пМ, от 3 мкМ до 0.001 пМ, от 1 мкМ до 0.001 пМ, от 0.5 мкМ до 0.001 пМ или от 0.1 мкМ до 0.001 пМ.
Основная четырехцепочечная структурная единица представляет собой гетеротетрамерный гликопротеин, состоящий из двух идентичных легких (L) цепей и двух идентичных тяжелых (Н) цепей (IgM антитело состоит из 5 основных гетеротетрамерных единиц, а также дополнительного полипептида, называемого J цепью, и, следовательно, содержит 10 антигенсвязывающих сайтов, тогда как секретированные IgA антитела могут полимеризоваться с образованием поливалентных сборных структур (совокупностей), содержащих 2-5 основных 4-цепочечных структурных единиц наряду с J цепью). В случае IgG 4-цепочечная структурная единица обычно имеет молекулярную массу 150000 Дальтон. Каждая L цепь связана с Н цепью одной ковалентной дисульфидной связью, тогда как две Н цепи связаны друг с другом одной или более дисульфидными связями в зависимости от изотипа Н цепи. Каждая Н и L цепь имеет также регулярно расположенные внутрицепные дисульфидные мостики. Каждая Н цепь имеет на N-конце вариабельный домен (VH), за которым следуют три константных домена (CH), для каждой из α и γ цепей и четыре CH домена для µ и ε изотипов. Каждая L цепь имеет на N-конце вариабельный домен (VL), за которым следует константный домен (CL) на другом конце. VL выравнивается с VH, a CL выравнивается с первым константным доменом тяжелой цепи (CH1). Полагают, что конкретные аминокислотные остатки образуют область контакта (границу раздела, интерфейс) между вариабельными доменами легкой цепи и тяжелой цепи. Объединение (спаривание) VH и VL дает единый антигенсвязывающий сайт. О структуре и свойствах различных классов антител см., например, Basic and Clinical Immunology. 8th edition, Daniel P. Stites, Abba I. Terr and Tristram G. Parslow (eds.), Appleton & Lange, Norwalk, CT, 1994, page 71 and Chapter 6.
L цепь различных видов млекопитающих можно отнести к одному из двух четко различающихся типов, называемых каппа и лямбда, в зависимости от аминокислотных последовательностей их константных доменов. В зависимости от аминокислотной последовательности константного домена их тяжелых цепей (CH) иммуноглобулины можно отнести к различным классам или изотипам. Существует пять классов иммуноглобулинов: IgA, IgD, IgE, IgG и IgM, имеющих тяжелые цепи, обозначаемые α, δ, γ, ε и µ, соответственно. Классы γ и α дополнительно делятся на подклассы на основании сравнительно незначительных (минорных) различий в CH последовательности и функции, например, в организме человека экспрессируются следующие подклассы: IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2.
Термин "вариабельный" относится к тому факту, что последовательности некоторых сегментов вариабельных доменов антител очень сильно различаются. V домен опосредует связывание антигена и определяет специфичность конкретного антитела к его конкретному антигену. Однако вариабельно неравномерно распределяется на участке вариабельных доменов протяженностью 110 аминокислот. Напротив, V области состоят из сравнительно инвариантных фрагментов, называемых каркасными областями (FR) из 15-30 аминокислот, разделенных более короткими участками чрезвычайной вариабельности, называемых "гипервариабельными областями", каждая из которых имеет протяженность 9-12 аминокислот. Каждый вариабельный домен нативных тяжелых и легких цепей содержит четыре FR, в значительной степени принимающих конфигурацию бета-складок, связанных тремя гипервариабельными областями, которые образуют петли, связывающие, и в некоторых случаях являющиеся частью бета складчатой структуры. Гипервариабельные области удерживаются вместе в тесной близости с помощью FR и с гипервариабельными областями другой цепи вносят вклад в образование антигенсвязывающего сайта антител (см. Kabat et al., Sequences of Proteins of Immunological Interest. 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD. (1991)). Константные домены не принимают непосредственного участия в связывании антитела с антигеном, но проявляют различные эффекторные функции, такие как участие антитела в антителозависимой клеточной цитотоксичности (АЗКЦ, ADCC).
Термин "гипервариабельная область" по данному описанию относится к аминокислотным остаткам антитела, которые отвечают за связывание антигена. Обычно гипервариабельная область содержит аминокислотные остатки из "области, определяющей комплементарность" или "CDR" (например, примерно, в области остатков 24-34 (LI), 50-56 (L2) и 89-97 (L3) в VL, и, примерно, в области остатков 26-35 (H1), 50-65 (H2) и 95-102 (Н3) в VH (в одном варианте изобретения H1 находится, примерно, в области остатков 31-35); Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD. (1991)), и/или такие остатки из "гипервариабельной петли" (например, остатки 26-32 (L1), 50-52 (L2) и 91-96 (L3) в VL и 26-32 (H1), 53-55 (H2) и 96-101 (Н3) в VH; Chothia and Lesk, J. Mol. Biol. 196: 901-917 (1987)).
"Каркасные области" (FR) представляют собой остатки вариабельного домена, отличные от CDR остатков. Обычно каждый вариабельный домен имеет четыре FR, определяемые как FR1, FR2, FR3 и FR4. Если CDR определяются согласно Kabat, FR остатки легкой цепи локализуются, приблизительно, в области остатков 1-23 (LCFR1), 35-49 (LCFR2), 57-88 (LCFR3) и 98-107 (LCFR4), а остатки FR тяжелой цепи локализуются, приблизительно, в области остатков 1-30 (HCFR1), 36-49 (HCFR2), 66-94 (HCFR3) и 103-113 (HCFR4) в тяжелой цепи. Если участки CDR содержат аминокислотные остатки из гипервариабельных петель, FR остатки легкой цепи локализуются, приблизительно, в остатках 1-25 (LCFR1), 33-49 (LCFR2), 53-90 (LCFR3) и 97-107 (LCFR4) в легкой цепи, а FR остатки тяжелой цепи локализуются, примерно, в остатках 1-25 (HCFRI), 33-52 (HCFR2), 56-95 (HCFR3) и 102-113 (HCFR4) в остатках тяжелой цепи. В некоторых примерах, когда CDR содержит аминокислоты как из CDR по Kabat, так и аминокислоты из гипервариабельной петли, FR соответствующим образом корректируются. Например, когда CDRH1 включает аминокислоты Н26-Н35, остатки FR1 тяжелой цепи находятся в положениях 1-25, а остатки FR2 находятся в положениях 36-49.
"Человеческая консенсусная каркасная область" обозначает каркасную область, которая представляет наиболее часто встречающиеся аминокислотные остатки при отборе VL или VH каркасных последовательностей человеческого иммуноглобулина. Обычно отбор VL или VH последовательностей человеческого иммуноглобулина проводят из подгруппы последовательностей вариабельных доменов. Как правило, подгруппа последовательностей представляет собой подгруппу в Kabat. В одном варианте изобретения, для VL подгруппа представляет собой подгруппу каппа I по Kabat. В другом варианте изобретения, для VH подгруппа представляет собой подгруппу каппа III по Kabat.
Термин "моноклональное антитело" по данному описанию относится к антителу из популяции практически гомогенных антител, т.е. индивидуальные антитела, составляющие популяцию, являются практически сходными и связывают один и тот же эпитоп (одни и те же эпитопы), за исключением возможных вариантов, которые могут появляться при продуцировании моноклонального антитела, причем такие варианты обычно присутствуют в минорных количествах. Такое моноклональное антитело, как правило, включает антитело, содержащее вариабельную область, которая связывает мишень, причем антитело получают таким способом, который включает отбор (селекцию) антитела из множества антител. Например, процесс селекции может представлять собой селекцию уникального клона из множества клонов, таких как пул клонов гибридомных клеток, фаговых клонов или клонов рекомбинантной ДНК. Следует понимать, что выбранное антитело можно далее изменять, например, с целью повышения аффинности к мишени, "гуманизации" антитела, повышения его продукции в клеточной культуре, снижения его иммуногенности in vivo, создания полиспецифического антитела и т.д., и что антитело, содержащее измененную вариабельную область, также является моноклональным антителом по данному изобретению. Помимо их специфичности, препараты моноклонального антитела имеют то преимущество, что они обычно не содержат примесей других иммуноглобулинов. Модификатор "моноклональный" указывает на отличительную особенность антитела, как полученного из практически гомогенной популяции антител, но его не следует рассматривать как требующий получения антитела каким-либо конкретным методом. Например, моноклональные антитела для применения по настоящему изобретению можно получать различными методами, включая метод гибридом (например, Kohler et al., Nature, 256: 495 (1975); Harlow et al., Antibodies: A Laboratory Manual, (Cold Spring Harbor Laboratory Press, 2nd ed. 1988); Hammerling et al., in: Monoclonal Antibodies and T-Cell Hybridomas 563-681, (Elsevier, N.Y., 1981), методы рекомбинантной ДНК (см., например, патент США No. 4816567), методы фагового дисплея (см., например, Clackson et al., Nature, 352: 624-628 (1991); Marks et al., J. Mol. Biol., 222: 581-597 (1991); Sidhu et al., J. Mol. Biol. 338(2): 299-310 (2004); Lee et al., J. Mol. Biol. 340(5): 1073-1093 (2004); Fellouse, Proc. Nat. Acad. Sci. USA 101(34): 12467-12472 (2004); и Lee et al. J. Immunol. Methods 284(1-2): 119-132 (2004), и методы получения человеческих и подобных человеческим антител с использованием животных, имеющих фрагменты или целые локусы генов или гены человеческого иммуноглобулина, кодирующие человеческие иммуноглобулиновые последовательности (см., например. Международные патентные заявки WO 98/24893, WO/9634096, WO/9633735 и WO/9110741, Jakobovits et al., Proc. Natl. Acad. Sci. USA, 90: 2551 (1993); Jakobovits et al., Nature, 362: 255-258 (1993); Bruggemann et al., Year in Immuno., 7: 33 (1993); патенты США No. 5545806, 5569825, 5591669 (все принадлежат GenPharm); 5545807; Международная патентная заявка WO 97/17852, патенты США No. 5545807; 5545806; 5569825; 5625126; 5633425; и 5661016, и Marks et al., Bio/Technology, 10: 779-783 (1992); Lonberg et al., Nature, 368: 856-859 (1994); Morrison, Nature, 368: 812-813 (1994); Fishwild et al., Nature Biotechnology, 14: 845-851 (1996); Neuberger, Nature Biotechnology, 14: 826 (1996); и Lonberg and Huszar, Intern. Rev. Immunol., 13: 65-93 (1995).
"Интактное" антитело представляет собой антитело, содержит антигенсвязывающий сайт, а также CL и по меньшей мере константные домены тяжелой цепи CH1, CH2 и CH3. Константные домены могут представлять собой константные домены нативной последовательности (например, константные домены человеческой нативной последовательности) или вариант аминокислотной последовательности. Предпочтительно, интактное антитело имеет одну или более эффекторных функций.
"Фрагменты антитела" содержат участок интактного антитела, предпочтительно, антигенсвязывающую или вариабельную область интактного антитела. Примеры фрагментов антитела включают фрагменты Fab, Fab', F(ab')2 и Fv; диатела; линейные антитела (см. патент США No. 5641870, Пример 2; Zapata et al., Protein Eng. 8(10): 1057-1062 (1995)); молекулы одноцепочечных антител и полиспецифические антитела, образованные из фрагментов антител.
Выражение "линейные антитела" в целом относится к антителам, описанным в Zapata et al., Protein Eng., 8(10): 1057-1062 (1995). Коротко говоря, эти антитела содержат пару тандемных Fd сегментов (VH-CH1-VH-CH1), которые вместе с полипептидами комплементарной легкой цепи образуют пару антигенсвязывающих областей. Линейные антитела могут быть биспецифическими или моноспецифическими.
Расщепление антител папаином дает два идентичных антигенсвязывающих фрагмента, называемых "Fab" фрагментами, и остаточный "Fc" фрагмент, его обозначение отражает способность легко кристаллизоваться. Фрагмент Fab состоит из целой цепи L, а также вариабельного домена цепи Н (VH) и первого константного домена одной тяжелой цепи (CH1). Обработка антитела пепсином дает единственный большой фрагмент F(ab')2, который в грубом приближении соответствует двум связанным дисульфидной связью фрагментам Fab, проявляющий двухвалентную антигенсвязывающую активность и все еще способный к перекрестному связыванию антигена. Фрагменты Fab' отличаются от фрагментов Fab наличием нескольких дополнительных остатков на карбокси-конце CH1 домена, включая один или более цистеиновых остатков из шарнирной области антитела. Fab'-SH обозначает в данном описании Fab', в котором цистеиновый(-ые) остаток (остатки) константной области несет (несут) свободную тиольную группу. Первоначально фрагменты антитела F(ab')2 получали в виде пары фрагментов Fab', между которыми находились шарнирные цистеиновые остатки. Известны также другие примеры связывания фрагментов антитела химической связью.
Фрагмент Fc содержит карбокси-концевые участки обеих цепей Н, связанные дисульфидной связью. Эффекторные функции антител определяются последовательностями в Fc области; эта область представляет собой также участок, узнаваемый Fc рецепторами (FcR), имеющимися в некоторых типах клетках.
"Fv" состоит из димера, состоящего из одного вариабельного домена тяжелой цепи и одного вариабельного домена легкой цепи, прочно связанного нековалентной связью. В результате фолдинга этих двух доменов образуется шесть гипервариабельных петель (по 3 петли из каждой Н и L цепи), которые предоставляют аминокислотные остатки в связывании антигена и сообщают антителу антигенсвязывающую специфичность. Однако даже единичный вариабельный домен (или половина Fv, содержащая только три CDR, специфических к антигену) способен узнавать и связывать антиген, хотя часто с более низкой аффинностью, чем целый сайт связывания.
"Одноцепочечный Fv", также сокращенно обозначаемый как "sFv" или "scFv", представляет собой фрагменты антитела, которые содержат VH и VL домены антитела, связанные в единую полипептидную цепь Предпочтительно, sFv полипептид дополнительно включает полипептидный линкер между VH и VL доменами, который способствует образованию фрагментом sFv структуры, необходимой для связывания антигена. Обзор об sFv см. Pluckthun in The Pharmacology of Monoclonal Antibodies vol. 113, Rosenburg and Moore eds., Springer-Verlag, New York, pp.269-315 (1994); Borrebaeck 1995.
Термин "диатела" относится к малым фрагментам антител, получаемым при составлении sFv фрагментов (см. предыдущий абзац), с короткими линкерами (примерно, 5-10 остатков) между VH и VL доменами так, чтобы осуществлялось не внутрицепное, а межцепное спаривание V доменов, давая в результате бивалентный фрагмент, т.е. фрагмент, имеющий два антигенсвязывающих сайта. Биспецифические диатела представляют собой гетеродимеры двух "кроссоверных" sFv фрагментов, в котором VH и VL домены двух антител находятся в различных полипептидных цепях. Более подробно диатела описаны, например, в Европейском патенте ЕР 404097; Международной патентной заявке WO 93/11161; и в Hollinger et al., Proc. Natl. Acad. Sci. USA, 90: 6444-6448 (1993).
"Гуманизированные" формы антител нечеловеческого происхождения (например, грызуна) представляют собой химерные антитела, которые содержат минимальную последовательность, полученную из нечеловеческого антитела. По большей части гуманизированные антитела представляют собой человеческие иммуноглобулины (реципиентное антитело), в которых остатки из гипервариабельной области реципиента заменены на остатки из гипервариабельной области вида, отличного от человека (донорное антитело), такого как мышь, крыса, кролик или нечеловеческий примат, имеющие заданные специфичность, аффинность и характеристики антитела. В некоторых случаях остатки каркасной области (FR) человеческого иммуноглобулина заменены на соответствующие остатки нечеловеческого иммуноглобулина. Помимо этого гуманизированные антитела могут содержать остатки, которых нет в реципиентном антителе или в донорном антителе. Эти модификации получают для того, чтобы дополнительно уточнить характеристики антитела. Как правило, "гуманизированное" антитело содержит практически все из по меньшей мере одного, и обычно двух, вариабельных доменов, в которых все или практически все гипервариабельные петли соответствуют этим элементам иммуноглобулина нечеловеческого происхождения, а все или практически все области FR представляют собой FR области из человеческой иммуноглобулиновой последовательности. Гуманизированное антитело, необязательно, содержит по меньшей мере часть константной области иммуноглобулина (Fc), как правило, человеческого иммуноглобулина. Более подробные сведения даны в Jones et al., Nature 321: 522-525 (1986); Riechmaim et al., Nature 332: 323-329 (1988); и Presta, Curr. Op. Struct. Biol. 2: 593-596 (1992).
Выражение "набор кодонов" по данному описанию относится к различным последовательностям нуклеотидных триплетов, применяемым для кодирования заданных аминокислотных вариантов. Набор нуклеотидов можно синтезировать, например, твердофазным синтезом, включая последовательности, которые представляют все известные комбинации нуклеотидных триплетов, предоставляемые набором кодонов, и которые будут кодировать заданную группу аминокислот. Стандартной формой обозначения кодонов является IUB код, который известен в уровне техники и представлен в данном описании. Набор кодонов обычно изображается в виде 3 заглавных букв, написанных курсивом, например NNK, NNS, XYZ, DVK и т.п. (например, кодон NNK относится к N=A/T/G/C в положениях 1 и 2 в кодоне, K=G/T в эквимолярном соотношении в положении 3 для кодирования всех 20 природных аминокислот). Таким образом, выражение "неслучайный набор кодонов" по данному описанию относится к набору кодонов, который кодирует выбранные аминокислоты, соответствующие частично, предпочтительно, полностью, критериям для селекции аминокислот по данному описанию. Синтез олигонуклеотидов с "вырожденностью" выбранных нуклеотидов в некоторых положениях хорошо известен в данной области техники, например, метод TRIM (Knappek et al., J. Mol. Biol. 296: 57-86, 1999); Garrard and Henner, Gene 128: 103, 1993). Такие наборы олигонуклеотидов, содержащие определенные наборы кодонов, можно синтезировать в продажных синтезаторах олиго(поли)нуклеотидов (выпускаемых, например, Applied Biosystems, Foster City, CA), или их можно приобретать (например, от компании Life Technologies, Rockville, MD). Следовательно, набор синтезированных олигонуклеотидов, имеющих конкретный набор кодонов, обычно включает множество олигонуклеотидов с различными последовательностями, причем различия определяются набором кодонов в полной последовательности. Олигонуклеотиды, применяемые по изобретению, имеют последовательности, которые способствуют гибридизации с нуклеотидной матрицей вариабельного домена, и также, но необязательно, могут включать сайты расщепления рестриктазами, используемые, например, для клонирования.
Антитело по данному изобретению, "которое связывает" целевой антиген, представляет собой антитело, которое связывает антиген с достаточной аффинностью так, что антитело можно применять в качестве диагностического и/или терапевтического агента при нацеливании на белок или клетку или ткань, экспрессирующую антиген, и в незначительной степени перекрестно реагирует с другими белками. По данным аналитических методов: сортинга флуоресцентно-активированных клеток (FACS), радиоиммунопреципитации (RIA) или ИФА (ELISA), в таких вариантах изобретения степень связывания антитела с белком, не являющимся "мишенью" (с "нецелевым белком"), составляет менее 10% от связывания антитела с конкретным белком-мишенью. По отношению к связыванию антитела с молекулой-мишенью термин "специфическое связывание" или выражения "специфически связывается с" или "специфический к" конкретному полипептиду или эпитопу на конкретном полипептиде-мишени означает связывание, которое заметно (измеримо) отличается от неспецифического взаимодействия (например, в случае bH1-44 или bH1-81 неспецифическое взаимодействие представляет собой связывание с бычьим сывороточным альбумином, казеином, фетальной бычьей сывороткой или нейтравидином). Специфическое связывание можно определять количественно, например, определяя связывание молекулы по сравнению со связыванием контрольной молекулы. Например, специфическое связывание можно определять конкурентной реакцией с другой молекулой, аналогичной мишени, например, с избытком немеченой мишени. В этом случае специфическое связывание указывается, если связывание меченой мишени с зондом конкурентно ингибируется избытком немеченой мишени. В данном описании термин "специфическое связывание" или выражения "специфически связывается с" или "специфический к" конкретному полипептиду или эпитопу на конкретном полипептиде-мишени можно характеризовать на примере молекулы, имеющей Kd к мишени по меньшей мере около 200 нМ, или же по меньшей мере около 150 нМ, или же по меньшей мере около 100 нМ, или же по меньшей мере около 60 нМ, или же по меньшей мере около 50 нМ, или же по меньшей мере около 40 нМ, или же по меньшей мере около 30 нМ, или же по меньшей мере около 20 нМ, или же по меньшей мере около 10 нМ, или же по меньшей мере около 8 нМ, или же по меньшей мере около 6 нМ, или же по меньшей мере около 4 нМ, или же по меньшей мере около 2 нМ, или же по меньшей мере около 1 нМ или выше. В одном варианте изобретения термин "специфическое связывание" относится к связыванию, при котором молекула связывается с конкретным полипептидом или эпитопом на конкретном полипептиде, практически не связываясь с каким-либо другим полипептидом или эпитопом на полипептиде.
"Аффинность связывания" обычно относится к силе совокупных нековалентных взаимодействий между единичным сайтом связывания молекулы (например, антитела) и ее партнером по связыванию (например, антигеном). Если не указано иначе, "аффинность связывания" относится к внутренней (характерной, истинной) аффинности связывания, которая отражает 1:1 взаимодействие между членами пары связывания (например, антителом и антигеном). Аффинность молекулы Х к своему партнеру Y обычно можно представить константной диссоциации (Kd). Желательно, чтобы величина Kd составляла, примерно, 200 нМ, 150 нМ, 100 нМ, 60 нМ, 50 нМ, 40 нМ, 30 нМ, 20 нМ, 10 нМ, 8 нМ, 6 нМ, 4 нМ, 2 нМ, 1 нМ или менее. Аффинность можно измерять обычными методами, известными в уровне техники, включая методы по данному описанию. Низкоаффинные антитела обычно медленно связываются с антигеном и имеют тенденцию легко диссоциировать, тогда как высокоаффинные антитела обычно быстрее связывают антиген и имеют тенденцию дольше оставаться в связанном состоянии. В уровне техники известны различные методы измерения аффинности связывания, любой из этих методов можно использовать для целей настоящего изобретения.
В одном варианте изобретения "Kd" или "величину Kd " по данному изобретению измеряют методами поверхностного плазменного резонанса на приборе BIAcore™-2000 или BIAcore™-3000 (BIAcore, Inc., Piscataway, NJ) при 25°С, используя чипы с иммобилизованным антигеном СМ5 при ~10 относительных единицах (единицах отклика, RU). Коротко говоря, биосенсорные чипы с карбоксиметилдекстраном (СМ5, BIAcore Inc.) активируют гидрохлоридом N-этил-N'-(3-диметиламинопропил)-карбодиимида (EDC) и N-гидроксисукцинимидом (NHS) в соответствии с инструкциями производителя. Антиген разводят 10 мМ раствором ацетата натрия, рН 4.8, до концентрации 5 мкг/мл (~0.2 мкМ), а затем вводят (инжекция) при скорости потока 5 мкл/минута до достижения, примерно, 10 относительных единиц (RU) связанного белка. После введения антигена вводят 1 М раствор этаноламина, чтобы блокировать непрореагировавшие группы. Для кинетических измерений двукратные серийные разведения Fab (например, от 0.78 нМ до 500 нМ) вводят в PBS с 0.05% Tween 20 (PBST) при 25°С при скорости потока, примерно, 25 мкл/мин. Величины скорости ассоциации (kon) и скорости ассоциации (koff) рассчитывают, применяя простую модель Ленгмюра для связывания один-плюс-один (BIAcore Evaluation Software версия 3.2), с помощью одновременного получения сенсограммы ассоциации и диссоциации. Константу равновесной диссоциации (Kd) рассчитывают как отношение koff/kon. См., например, Chen, Y., et al., (1999) J. Mol. Biol. 293: 865-881. Если по данным вышеуказанного метода поверхностного плазмонного резонанса скорость ассоциации превышает 106 М-1 сек-1, тогда ее можно определять методом тушения флуоресценции, который измеряет увеличение или уменьшение интенсивности флуоресцентной эмиссии (возбуждение=295 нм; эмиссия (излучение)=340 нм, полоса 16 нм) при 25°С раствора антитела против антигена (Fab форма) с концентрацией 20 нМ в PBS, рН 7.2, в присутствии увеличивающихся концентраций антигена, измеряемых с помощью спектрометра, такого как спектрофотометр остановленного потока (Aviv Instruments) или спектрофотометр SLM-Aminco (ThermoSpectronie) серии 8000 с кюветой с перемешиванием.
"Скорость ассоциации" ("on-rate") или "kon" по данному изобретению можно также определять тем же самым описанным выше методом поверхностного плазменного резонанса на приборе BIAcore™-2000 или BIAcore™-3000 (BIAcore, Inc., Piscataway, NJ) при 25°С, используя чипы с иммобилизованным антигеном СМ5 при ~10 относительных единицах (единицах отклика, RU). Коротко говоря, биосенсорные чипы с карбоксиметилдекстраном (СМ5, BIAcore Inc.) активируют гидрохлоридом N-этил-N'-(3-диметиламино пропил)-карбодиимида (EDC) и N-гидроксисукцинимидом (NHS) в соответствии с инструкциями производителя. Антиген разводят 10 мМ раствором ацетата натрия, рН 4.8, до концентрации 5 мкг/мл (~0.2 мкМ), а затем вводят (инжекция) при скорости потока 5 мкл/минута до достижения, примерно, 10 относительных единиц (RU) связанного белка. После введения антигена вводят 1 М раствор этаноламина, чтобы блокировать непрореагировавшие группы. Для кинетических измерений двукратные серийные разведения Fab (например, от 0.78 нМ до 500 нМ) вводят в PBS с 0.05% Tween 20 (PBST) при 25°С при скорости потока, примерно, 25 мкл/мин. Величины скорости ассоциации (kon) и скорости ассоциации (koff) рассчитывают, применяя простую модель Ленгмюра для связывания один-плюс-один (BIAcore Evaluation Software версия 3.2), одновременно получая сенсограммы ассоциации и диссоциации. Константу равновесной диссоциации (Kd) рассчитывают как отношение koff/kon. См., например, Chen, Y., et al., (1999) J. Mol. Biol. 293: 865-881. Однако, если по данным вышеуказанного метода поверхностного плазменного резонанса скорость ассоциации превышает 106 М-1 сек-1, тогда ее можно определять методом тушения флуоресценции, который измеряет увеличение или уменьшение интенсивности флуоресцентной эмиссии (возбуждение=295 нм; эмиссия (излучение)=340 нм, полоса 16 нм) при 25°С раствора антитела против антигена (Fab форма) с концентрацией 20 нМ в PBS, рН 7.2, в присутствии увеличивающихся концентраций антигена, измеряемых с помощью спектрометра, такого как спектрофотометр остановленного потока (Aviv Instruments) или спектрофотометр SLM-Aminco (ThermoSpectronic) серии 8000 с кюветой с перемешиванием.
Если специально не указано иначе, выражения "биологически активный", и "биологическая активность", и "биологические характеристики", по отношению к полипептиду по данному изобретению, означают обладание способностью связываться с биологической молекулой.
Выражение "биологическая молекула" относится к нуклеиновой кислоте, белку, углеводу, липиду и их комбинации. В одном варианте изобретения биологическая молекула существует в природе.
Определение "выделенный" ("изолированный"), применяемое для описания различных антител по данному описанию, означает антитело, идентифицированное и выделенное и/или регенерированное из клетки или клеточной культуры, в которой оно экспрессируется. Примеси (загрязняющие компоненты) из природной среды представляют собой материалы, которые, как правило, мешают диагностическому или терапевтическому применению полипептида, и могут включать ферменты, гормоны и другие белковые или небелковые растворенные вещества. В предпочтительных вариантах изобретения антитело очищают (1) до степени, достаточной для получения по меньшей мере 15 остатков N-концевой или внутренней аминокислотной последовательности, при использовании секвенатора с вращающейся стеклянной чашечкой (секвенатора Эдмана), или (2) до гомогенности методом SDS-PAGE в невосстанавливающих или восстанавливающих условиях с применением окрашивания Кумасси бриллиантовым голубым или, предпочтительно, серебром. Выделенное антитело включает антитела in situ внутри рекомбинантных клеток, так как по меньшей мере один компонент природной среды полипептида отсутствует. Обычно, однако, выделенный полипептид получают в результате по меньшей мере одной стадии очистки.
Термин "контрольные последовательности" относится к последовательностям ДНК, необходимым для экспрессии функционально связанной кодирующей последовательности в конкретном организме-хозяине. Контрольные последовательности, которые подходят для прокариот, включают, например, промоторную, необязательно, операторную последовательность и сайт связывания рибосом. Известно, что в эукариотических клетках используются промоторы, сигналы полиаденилирования и энхансеры.
Нуклеиновая кислота является "функционально связанной", если находится в функциональном контакте с последовательностью другой нуклеиновой кислоты. Например, ДНК для препоследовательности или секреторной лидерной последовательности функционально связана с ДНК для полипептида, если она экспрессируется в виде пребелка, который участвует в секреции полипептида; промотор или энхансер функционально связан с кодирующей последовательностью, если он влияет на транскрипцию последовательности; или сайт связывания рибосом функционально связан с кодирующей последовательностью, если он расположен таким образом, который позволяет содействовать трансляции. Как правило, "функционально связанный" означает, что связанные ДНК-последовательности являются прилегающими, непрерывными, а, в случае секреторной лидерной последовательности, прилегающими (смежными, непрерывными) и в фазе считывания. Однако энхансеры не должны быть прилегающими. Связывание выполняют дотированием по подходящим сайтам рестрикции. Если такие сайты отсутствуют, в соответствии с обычной практикой используют синтетические олигонуклеотидные адаптеры или линкеры.
"Идентичность аминокислот в процентах (%)" по отношению к полипептидным последовательностям по данному описанию определяется как процентное содержание аминокислотных остатков в кандидатной последовательности, которые идентичны аминокислотным остаткам в сравниваемом полипептиде, после изменения последовательностей и введения гэпов, при необходимости, для достижения максимальной идентичности последовательностей в процентах, и при этом никакие консервативные замены не рассматриваются как составляющие идентичности последовательностей. Выравнивание с целью процента идентичности аминокислотных последовательностей можно проводить различными способами, известными специалистам в данной области техники, например, используя общедоступные компьютерные программы, такие как BLAST, BLAST-2, ALIGN или Megalign (DNASTAR). Специалисты в данной области техники могут определить соответствующие параметры для количественного определения выравнивания, включая любые алгоритмы, необходимые для достижения максимального выравнивания по всей длине сравниваемых последовательностей. Однако для целей настоящего изобретения величину идентичности аминокислотных последовательностей в % получают, используя компьютерную программу сравнения последовательностей ALIGN-2. Компьютерная программа сравнения последовательностей ALIGN-2 разработана в Genentech, Inc., и исходная программа была подана с документацией для пользователей в Бюро регистрации авторских прав США, Washington D.C., 20559, где она зарегистрирована под No. TXU510087. Программа ALIGN-2 является общедоступной через Genentech, Inc., South San Francisco, California. Программу ALIGN-2 следует компилировать для применения в операционной системе UNIX, предпочтительно, цифровой UNIX V4.0D. Все параметры сравнения последовательностей устанавливаются программой ALIGN-2 и не изменяются.
Если не указано иначе, аминокислотные последовательности по данному описанию представляют собой непрерывные аминокислотные последовательности.
Выражение "структурно несходные" ("несходные по структуре") биологические молекулы по данному изобретению относится к биологическим молекулам, не относящимся к одному и тому же классу (белок, нуклеиновая кислота, липид, углеводы и т.д.), или, например, по отношению к белкам, имеющие идентичность аминокислотных последовательностей менее 60%, идентичность аминокислотных последовательностей менее 50%, идентичность аминокислотных последовательностей менее 40%, идентичность аминокислотных последовательностей менее 30%, идентичность аминокислотных последовательностей менее 20% или идентичность аминокислотных последовательностей менее 10% относительно друг друга.
"Жесткость" ("строгость") условий реакции гибридизации легко определяется рядовым специалистом в данной области техники и обычно является эмпирической, рассчитанной в зависимости от длины зонда, температуры отмывки и концентрации соли. В целом, более длинные зонды требуют более высоких температур для соответствующего отжига, тогда как для более коротких зондов требуются более низкие температуры. Обычно гибридизация зависит от способности денатурированной ДНК вторично отжигаться, когда комплементарные нити находятся в среде с температурой ниже их температур плавления. Чем выше степень заданной гомологии между зондом и гибридизуемой последовательностью, тем выше температура, которую можно применять. Отсюда следует, что более высокие относительные температуры способствуют более жестким условиям реакции, тогда как более низкие температуры менее жестким. Более подробные сведения и объяснение понятия жесткости условий реакций гибридизации даны в Ausubel et al., Current Protocols in Molecular Biology, Wiley Interscience Publishers, (1995).
"Жесткие условия" или "условия высокой жесткости", по определению в данном описании, можно идентифицировать как следующие: (1) применение среды с низкой ионной силой и высокой температуры для отмывки, например, 0.015 М раствор хлорида натрия/0.0015 М раствор цитрата натрия /0.1% раствор додецилсульфата натрия при 50°С; (2) применение в процессе гибридизации денатурирующего агента, такого как формамид, например, 50% (об./об.) формамида с 0.1% альбумина бычьей сыворотки /0.1% фиколла /0.1% поливинилпирролидона/50 мМ натрий-фосфатного буфера с рН 6.5 с 750 мМ хлорида натрия, 75 мМ цитрата натрия при 42°С; или (3) гибридизация в течение ночи в растворе, который содержит 50% формамида, 5 х SSC (0.75 М NaCl, 0.075 М цитрата натрия), 50 мМ фосфата натрия (рН 6.8), 0.1% пирофосфата натрия, 5 х раствора Денхардта, обработанной ультразвуком ДНК из спермы лосося (50 мкг/мл), 0.1% SDS 10% сульфата декстрана при 42°С, с 10-минутной отмывкой при 42°С в 0.2 х SSC (хлорид натрия/цитрат натрия) с последующей 10-минутной отмывкой в условиях высокой жесткости 10, а именно, в 0.1 х SSC содержащем EDTA (ЭДТА) при 55°С.
"Умеренно жесткие условия" можно определить, как описано в Sambrook et al., Molecular Cloning: A Laboratory Manual, New York: Cold Spring Harbor Press, 1989, они включают применение отмывочного раствора и условий гибридизации (например, температура, ионная сила и %SDS), менее жестких, чем описанные выше. Примером умеренно жестких условий является инкубация в течение ночи при 37°С в растворе, содержащем: 20% формамида, 5 х SSC (150 мМ NaCl, 15 мМ тринатрийцитрата), 50 мМ фосфата натрия (рН 7.6), 5 х раствора Денхардта, 10% сульфата декстрана и 20 мг/мл денатурированной ДНК из спермы лосося, с последующей отмывкой фильтров в 1 х SSC при температуре около 37-50°С. Опытный специалист в данной области техники знает, как при необходимости скорректировать температуру, ионную силу и т.д., чтобы они соответствовали таким факторам, как длина зонда и т.п.
"Эффекторные функции" антитела относятся к биологической активности, обусловленной Fc областью (нативной последовательностью Fc области или аминокислотной последовательностью вариантной Fc области) антитела, и изменяются с изменением изотипа антитела. Примеры эффекторных функций антитела включают: C1q связывание и комплементзависимую цитотоксичность; Fc-рецепторное связывание; антителозависимую клеточно-опосредованную (клеточную) цитотоксичность (ADCC); фагоцитоз; даун ("down")-регуляцию рецепторов клеточной поверхности (например В-клеточного рецептора); и активацию В клеток.
Выражение "антителозависимая клеточно-опосредованная (клеточная) цитотоксичность" или "ADCC" относится к форме цитотоксичности, при которой секретированный Ig, связанный с Fc рецепторами (FcR), присутствующими на некоторых цитотоксических клетках (например, естественных киллерных (NK) клетках, нейтрофилах и макрофагах), позволяет этим цитотоксическим эффекторным клеткам специфически связываться с несущей антиген клеткой-мишенью, а затем убивать клетку-мишень с помощью цитотоксинов. Антитела "вооружают" цитотоксические клетки и абсолютно необходимы для такого киллинга. Первичные клетки для опосредования ADCC, NK клетки, экспрессируют только FcγRIII, тогда как моноциты экспрессируют FcγRI, FcγRII и FcγRIII. Экспрессия FcR на гемопоэтических клетках кратко представлена в Таблице 3 на стр.464 обзора Ravetch and Kinet, Annu. Rev. Immunol. 9: 457-92 (1991). Для оценки ADCC активности представляющей интерес (нужной, целевой) молекулы можно провести in vitro ADCC анализ, такой как анализы, описанные в патентах США No. 5500362 или 5821337. Подходящие для таких анализов эффекторные клетки включают мононуклеарные клетки периферической крови (РВМС) и естественные киллерные клетки (NK). Или же, или помимо этого, ADCC активность нужной (целевой) молекулы можно определять in vivo, например, на животной модели, такой как животная модель, раскрываемая в Clynes et al. (Proc. Natl. Acad. Sci. USA) 95: 652-656 (1998).
Термин "Fc рецептор" или "FcR" описывает рецептор, который связывается с областью Fc антитела. Предпочтительный FcR представляет собой человеческий FcR с нативной последовательностью. Более того, предпочтительный FcR представляет собой FcR, который связывает IgG антитело (гамма рецептор) и включает рецепторы FcγRI, FcγRII и FcγRIII подклассов, в том числе аллельные варианты и формы этих рецепторов, образующиеся при альтернативном сплайсинге. FcγRII рецепторы включают FcγRIIA ("активирующий рецептор") и FcγRIIB ("ингибирующий рецептор"), которые имеют сходные аминокислотные последовательности, которые отличаются главным образом в своих цитоплазматических доменах. Активирующий рецептор FcγRIIA содержит иммунорецепторный тирозинсодержащий активационный мотив (ITAM) в своем цитоплазматическом домене. Ингибирующий рецептор FcγRIIB содержит иммунорецепторный тирозинсодержащий ингибиторный мотив (ITIM) в своем цитоплазматическом домене (см. обзор М. in Daeron, Annu. Rev. Immunol. 15: 203-234 (1997)). обзор о FcR в Ravetch and Kinet, Annu. Rev. Immunol. 9: 457-492 (1991); Capel et al., Immunomethods 4: 25-34 (1994); и de Haas et al., J. Lab. Clin. Med. 126: 330-41 (1995). Другие FcR, включая FcR, которые будут определены в будущем, в данном описании охватываются термином "FcR". Этот термин включает также неонатальный рецептор, FcRn, отвечающий за перенос материнских IgG-антител через плаценту (в фетус, плод) (Guyer et al., J. Immunol. 117: 587 (1976) и Kim et al., J. Immunol. 24: 249 (1994)).
"Человеческие эффекторные клетки" обозначают лейкоциты, которые экспрессируют один или более FcR и осуществляют эффекторные функции. Предпочтительно, клетки экспрессируют по меньшей мере FcγRIII и осуществляют ADCC эффекторную функцию. Примеры человеческих лейкоцитов, которые опосредуют ADCC, включают мононуклеарные клетки периферической крови (РВМС), естественные киллерные (NK) клетки, моноциты, цитотоксические Т клетки и нейтрофилы; предпочтительными являются РВМС и NK клетки. Эффекторные клетки можно выделять из нативного источника, например, из крови.
Термин "комплементзависимая цитотоксичность" или "CDC" относится к лизису клетки-мишени в присутствии комплемента. Активация классического пути комплемента инициируется связыванием первого компонента системы комплемента (C1q) с антителами (соответствующего подкласса), которые связываются со "своим" антигеном (партнером по когнатному взаимодействию). Для оценки активации комплемента можно провести CDC анализ, например, описанный в Gazzano-Santoro et al., J. Immunol. Methods 202: 163 (1996).
Термин "терапевтически эффективное количество" относится к количеству антитела или фрагмента антитела для лечения заболевания или расстройства у субъекта. В случае опухоли refers (например, раковой опухоли) терапевтически эффективное количество антитела или фрагмента антитела (например, полиспецифического антитела или фрагмента антитела, которое специфически связывает HER2 и VEGF) может уменьшать число раковых клеток; уменьшать начальный размер опухоли; ингибировать (т.е. до некоторой степени замедлять и, предпочтительно, прекращать) инфильтрацию раковыми клетками периферических органов; ингибировать (т.е. до некоторой степени замедлять и, предпочтительно, прекращать) метастазирование опухоли; ингибировать, до некоторой степени, рост опухоли; и/или облегчать, до некоторой степени, один или более симптомов, обусловленных расстройством. Антитело или фрагмент антитела может, до некоторой степени, предупреждать рост и/или убивать имеющиеся раковые клетки, оно может вызывать цитостатический и/или цитотоксический эффект. При терапии рака эффективность in vivo можно определять, например, оценивая продолжительность жизни, время до прогрессирования заболевания (ТТР), частоту ответа опухоли на лечение (RR), продолжительность ответа и/или качество жизни.
Под выражением "снижает или ингибирует" понимают способность вызывать общее снижение, предпочтительно, на 20% или более, более предпочтительно, на 50% или более и, наиболее предпочтительно, на 75%, 85%, 90%, 95% или более. "Снижает или ингибирует" может относиться к наличию или размеру метастазов, размеру первичной опухоли или к размеру или числу кровеносных сосудов при ангиогенных расстройствах.
Термины "рак" или "раковый" относятся к физиологическому состоянию или описывают физиологическое состояние у млекопитающих, которое, как правило, характеризуется нерегулируемым(/-ой) ростом/пролиферацией клеток. Это определение охватывает доброкачественные и злокачественные раковые заболевания. Примеры раковых заболеваний включают, но без ограничения, карциному, лимфому, бластому, саркому и лейкоз. Более конкретные примеры таких раковых заболеваний включают плоскоклеточный рак, мелкоклеточный рак легкого, немелкоклеточный рак легкого, аденокарциному легкого, плоскоклеточную карциному легкого, рак брюшины, печеночноклеточный рак, рак желудка, включая рак ЖКТ, рак поджелудочной железы, глиобластому, глиому, цервикальный рак, рак яичника, рак печени, рак мочевого пузыря, рак молочной железы, рак толстой кишки, колоректальный рак, карциному эндометрия и матки, карциному слюнных желез, рак почки (почечноклеточную карциному), рак предстательной железы (рак простаты), рак наружных женских половых органов, рак щитовидной железы, печеночную карциному, карциному анального канала, карциному пениса, меланому и различные типы рака головы и шеи.
Под "ранней стадией рака" понимают рак, который не является инвазивным или метастатическим или классифицируется как стадия 0, I или II рака.
Термин "предраковый" относится к состоянию или к росту, которые обычно предшествуют раку или развиваются в раковое заболевание.
Под термином "неметастатический" понимают рак, который является доброкачественным или который остается в первичной области и не проникает в лимфатическую или кровеносную систему или в ткани, отличные от первичной области. Как правило, неметастатический рак обозначает любой рак, который представляет собой стадию 0, I или II рака и, иногда, стадию III рака.
"Незлокачественное заболевание или расстройство, включающее анормальную активацию HER2", представляет собой состояние, исключая рак, при котором анормальная активация HER2 происходит в клетках или в ткани субъекта, имеющего заболевание или расстройство, или предрасположенного к заболеванию или расстройству. Примеры таких заболеваний или расстройств включают аутоиммунное заболевание (например, псориаз), см. определение, данное ниже; эндометриоз; склеродермию; рестеноз; полипы, такие как полипы в толстой кишке, назальные полипы или полипы в желудочно-кишечном тракте; фиброаденому; респираторное заболевание (например, хронический бронхит, астма, включая острую астму и аллергическую астму, муковисцидоз, бронхоэктаз, аллергические и другие формы ринита или синусита, недостаточность α1-антитрипсина, кашель, эмфизема легкого, фиброз легкого или гиперреактивность дыхательных путей, хроническое обструктивное заболевание легких и хроническое обструктивное легочное расстройство); холецистит; нейрофиброматоз; поликистозную болезнь почек; воспалительные заболевания; кожные расстройства, включая псориаз и дерматит; сосудистое заболевание; состояния, включающие анормальную пролиферацию клеток эпителия сосудов; язву желудочно-кишечного тракта; болезнь Менетрие с развитием аденом или синдром потери белков; почечные расстройства; ангиогенные расстройства; глазную болезнь, такую как возрастная дегенерация желтого пятна, синдром предполагаемого глазного гистоплазмоза, ретинальная неоваскуляризация при пролиферативной диабетической ретинопатии, ретинальная васкуляризация или диабетическая ретинопатия; патологии, связанные со скелетом, такие как остеоартрит, рахит и остеопороз; поражение после церебрального ишемического события; фиброзные или отечные заболевания, такие как цирроз печени, легочный фиброз, саркоидоз, тиреоидит, системный синдром гипервязкости крови, болезнь Ослера-Вебера-Рандю, хроническую окклюзионную легочную болезнь, или отек после ожогов, травмы, облучения, удара, гипоксии или ишемии; аллергическую кожную реакцию; диабетическую ретинопатию и диабетическую нефропатию; синдром Гийена-Барре; болезнь трансплантат-против-хозяина или отторжение трансплантата; болезнь Педжета; воспаление костей или суставов; солнечную геродермию (например, вызванную действием УФ-лучей на кожу); доброкачественную гипертрофию простаты; некоторые микробные инфекции, включая микробные патогены, выбранные из аденовируса, гантавирусов, боррелию бургдорфери, Yersinia spp. (иерсинию, возбудителя псевдотуберкулеза) и "коклюшную палочку"; тромбоз, вызванный агрегацией эритроцитов; состояния репродуктивной системы, такие как эндометриоз, синдром гиперстимуляции яичников, преэклампсия, дисфункция маточное кровотечение или менометроррагия; синовит; атерому; острые и хронические нефропатии (включая пролиферативный гломерулонефрит и диабетическую болезнь почек); экзему; образование гипертрофированных рубцов; эндотоксический шок и грибковую инфекцию; семейный аденоматоз (диффузный полипоз); нейродегенеративные заболевания (например, болезнь Альцгеймера, деменция, связанная со СПИДом, болезнь Паркинсона, амиотрофический латеральный склероз, пигментный ретинит, мышечная атрофия спинного мозга и мозжечковая дегенерация); миелодиспластические синдромы; гипопластическую анемию; ишемическое поражение; фиброз легкого, почки или печени; Т-клеточно-опосредованное аллергическое заболевание; детский гипертрофический стеноз привратника; синдром закупорки мочевых путей; псориатический артрит; и тиреоидит Хашимото.
"Аутоиммунное заболевание" по данному описанию означает заболевание или расстройство, обусловленное собственными тканями индивидуума и направленное против собственных тканей индивидуума, или их выделенного компонента или проявления или результирующего состояния. Примеры аутоиммунных заболеваний или расстройств включают, но без ограничения, артрит (ревматоидный артрит, такой как острый артрит, хронический ревматоидный артрит, подагрический артрит, хронический воспалительный артрит, дегенеративный артрит, инфекционный артрит, Лайм-артрит, пролиферативный артрит, псориатический артрит, артрит позвоночника и ювенильный ревматоидный артрит, остеоартрит, артрит хронический проградиентный, деформирующий артрит, полиартрит хронический первичный, реактивный артрит и анкилозирующий спондилоартрит), воспалительные гиперпролиферативные кожные заболевания, псориаз, такой как бляшечный псориаз, каплевидный псориаз, пустулезный псориаз и псориаз ногтей, дерматит, включая контактный дерматит, хронический контактный дерматит, аллергический дерматит, аллергический контактный дерматит, герпесный дерматит и атопический дерматит, гипер-IgM-синдром, крапивницу, такую как хроническая аллергическая крапивница и хроническая идиопатическая крапивница, включая хроническую аутоиммунную крапивницу, полимиозит/дерматомиозит, ювенильный дерматомиозит, токсический эпидермальный некролизис, склеродермию (включая системную склеродермию), склероз, такой как системный склероз, рассеянный склероз (PC, MS), такой как MS с поражением спинного мозга и зрительного нерва, первичный прогрессирующий MS (ППРС, PPMS); и MS на стадии "рецидив-ремиссия" (RRMS), прогрессирующий системный склероз, атеросклероз, артериосклероз и рассеянный склероз с атактическим синдромом, воспалительное заболевание кишечника (IBD) (например, болезнь Крона, аутоиммунные заболевания желудочно-кишечного тракта, колит, такой как язвенный колит, микроскопический колит, коллагенозный колит, полипозный колит, некротизирующий энтероколит и трансмуральный колит, и аутоиммунное воспалительное заболевание кишечника), гангренозную пиодермию, нодозную эритему, первичный склерозирующий холангит, эписклерит), респираторный дистресс-синдром, включая респираторный дистресс-синдром взрослых и острый респираторный дистресс-синдром (ARDS), менингит, воспаление целого увеального тракта или его отдела, ирит, хориоидит, аутоиммунное гематологическое расстройство, ревматоидный спондилит, внезапную потерю слуха, IgE-опосредованные заболевания, такие как анафилаксия и аллергический и атопический ринит, энцефалит, такой как энцефалит Расмуссена и лимбический энцефалит и/или энцефалит головного мозга, увеит, такой как увеит, острый передний увеит, гранулематозный увеит, негранулематозный увит, факогенный увеит, задний увеит или аутоиммунный увеит, гломерулонефрит (GN) с нефротическим синдромом и без нефротического синдрома, такой как хронический или острый гломерулонефрит, например, первичный GN, иммуноопосредованный GN, мембранозный GN (мембранозную нефропатию), идиопатический мембранозный GN или идиопатическую мембранозную нефропатию, мембрано- или мембранозный пролиферативный GN (MPGN), включая Типа I и Типа II и быстро прогрессирующий GN, аллергические состояния, аллергическую реакцию, экзему, включая аллергическую или атопическую экзему, астму, такую как бронхиальная астма и аутоиммунная астма, состояния, включающие инфильтрацию Т клетками и хронические воспалительные реакции, хроническое легочное воспалительное заболевание, аутоиммунный миокардит, недостаточность адгезии лейкоцитов, системную красную волчанку (SLE) или системную эритематозную волчанку (SLE), такую как кожная SLE, подострая кожная красная волчанка, волчаночный синдром новорожденных, волчаночный синдром новорожденных, волчанку (включая нефритную, энцефалитную, педиатрическую, неренальную, экстраренальную, дискоидную алопецию), ювенильный (Типа I) сахарный диабет (IDDM), сахарный диабет взрослых (диабет типа II), идиопатический несахарный диабет (Типа II), аутоиммунный диабет, идиопатический несахарный диабет, иммунные реакции, ассоциированные с острой и аллергией и аллергией замедленного типа, опосредуемой цитокинами и Т-лимфоцитами, туберкулез, саркоидоз, гранулематоз, гранулематоз Вегенера, агранулоцитоз, васкулит (включая васкулит крупных сосудов (включая полимиалгию ревматическую и гигантоклеточный артериит (артериит Такаясу)), васкулит средних сосудов (включая болезнь Кавасаки и нодозный полиартериит), микроскопический полиартериит, васкулит ЦНС, некротизирующий, кожный или аллергический васкулит, системный некротизирующий васкулит и ANCA-ассоциированный васкулит, такой как васкулит или синдром Черга-Страусе (CSS)), временный артериит, апластическую анемию, аутоиммунную апластическую анемию, Кумбс-позитивную анемию или иммунную гемолитическую анемию, включая аутоиммунную гемолитическую анемию (AIHA), злокачественную анемию (пернициозную анемию), болезнь Аддисона, чистую красноклеточную анемию или аплазию (PRCA), недостаточность Фактора VIII, гемофилию А, аутоиммунную нейтропению, панцитопению, лейкопению, заболевания, включающие диапедез лейкоцитов, воспалительные расстройства ЦНС, синдром множественного поражения органов, такой как синдром, вызванный (вторичный) сепсисом, травмой или кровоизлиянием, заболевания, опосредуемые комплексом антиген-антитело, болезнь анти-БМК антител (синдром Гудпасчера), антифосфолипидный синдром (АФС), аллергический неврит, синдром Бехчета, синдром Съегрена (Шегрена), синдром Стивенса-Джонсона, пемфигоид, такой как буллезный пемфигоид, пузырчатку (пемфигус) (включая обыкновенную пузырчатку, листовидную пузырчатку, пемфигоид слизистых оболочек и эритематозную пузырчатку), аутоиммунные полиэндокринопатии, болезнь или синдром Рейтера, иммунокомплексный нефрит, антителоопосредованный нефрит, нейромиелит зрительного нерва, полинейропатии, хроническую нейропатию, такую как IgM полинейропатии или IgM-опосредованная нейропатия, тромбоцитопению (например, такую, которая развивается у больных инфарктом миокарда), включая тромбоцитарную тромбоцитопеническую пурпуру (ТТР) и аутоиммунную или иммунноопосредованную тромбоцитопению, такую как идиопатическая тромбоцитопеническая пурпура (ITP), включая хроническую и острую ITP, аутоиммунное заболевание яичка и яичника, включая аутоиммунный орхит и оофорит, первичный гипертиреоз, гипопаратиреоз, аутоиммунные эндокринные заболевания, включая тиреодит, такой как аутоиммунный тиреодит, болезнь Хашимото, хронический тиреодит (тиреодит Хашимото) или подострый тиреодит, аутоиммунное заболевание щитовидной железы (ЩЖ), идиопатический гипотиреоз, болезнь Грейвза, полигландулярные синдромы, такие как аутоиммунные полигландулярные синдромы (или полигландулярные эндокринные синдромы), паранеопластические синдромы, включая неврологические паранеопластические синдромы, такие как миастенический синдром Ламберта-Итона или синдром Итона-Ламберта, синдром мышечной скованности, энцефаломиелит, такой как аллергический энцефаломиелит и экспериментальный аллергический энцефаломиелит (ЕАЕ), тяжелую псевдопаралитическую миастению, такую, как тяжелая псевдопаралитическая миастения, сопутствующая тимоме, мозжечковую дегенерацию, опсоклонус-миоклонус (OMS), и сенсорную нейропатию, мультифокальную двигательную нейропатию, синдром Шихана, аутоиммунный гепатит, хронический гепатит, волчаночный гепатит, гигантоклеточный гепатит, хронический активный гепатит или аутоиммунный хронический активный гепатит, лимфоидный интерстициальный пневмонит, облитерирующий бронхиолит (не-трансплантат) см. выше NSIP (неспецифическая интерстициальная пневмония), синдром Гийена-Барре, болезнь Берже (IgA нефропатию), идиопатическую IgA нефропатию, линейный IgA дерматоз, первичный билиарный цирроз, пневмоцирроз, синдром аутоиммунной энтеропатии, глютеновую болезнь, глютеновую энтеропатию, рефракторный спру, идиопатический спру, криоглобулинемию, амиотрофический латеральный склероз (БАС, ALS; болезнь Лу Герига), болезнь коронарных артерий, аутоиммунное ушное заболевание, такое как аутоиммунная болезнь внутреннего уха (AIED), аутоиммунная потеря слуха, синдром опсоклонус-миоклонус (OMS), полихондрит, такой как рефракторный или рецидивирующий полихондрит, легочный альвеолярный протеиноз, амилоидоз, склерит, доброкачественный лимфоцитоз, первичный лимфоцитоз, который включает моноклональный В-клеточный лимфоцитоз (например, доброкачественную моноклональную гаммопатию и моноклональную гаммопатию неопределенного значения (MGUS), периферическую нейропатию, паранеопластический синдром, каналопатии, такие как эпилепсия, мигрень, аритмия, мышечные расстройства, глухота, слепота, периодический паралич, и каналопатии ЦНС, аутизм, воспалительную миопатию, фокально-сегментарный гломерулосклероз (FSGS), эндокринную офтальмопатию, увеоретинит, хориоретинит, аутоиммунную болезнь печени, фибромиалгию, множественную недостаточность эндокринных желез, синдром Шмидта, адреналит, атрофию желудка, пресенильную деменцию, демиелинизирующие заболевания, такие как аутоиммунные демиелинизирующие заболевания, диабетическую нефропатию, синдром Дресслера, гнездную алопецию, CREST синдром (кальциноз, феномен Рейно, нарушения моторики пищевода, склеродактилия и телеангиэктазия), мужское и женское аутоиммунное бесплодие, смешанное соединительнотканное заболевание, болезнь Чага, ревматическую лихорадку, привычный выкидыш, легкие фермера, многоформную (мультиформную) эритему, посткардиотомический синдром, синдром Кушинга, легкое птицеводов, аллергический гранулематозный ангиит, доброкачественный лимфоцитарный ангиит, синдром Альпорта, альвеолит, такой как аллергический альвеолит и фиброзирующий альвеолит, интерстициальное заболевание легких, трансфузионную реакцию, проказу, малярию, лейшманиоз, трипаносомиаз, шистосоматоз, аскаридоз, аспергиллоз, синдром Самптера, синдром Каплана, лихорадку денге, эндокардит, эндомиокардиальный фиброз, диффузный интерстициальный легочный фиброз, идиопатический легочный фиброз, цистофиброз, эндофтальмит, стойкую возвышающуюся эритему, эритробластоз плода, эозинофильный фасциит, синдром Шульмана, синдром Фелти, филяриоз, циклит, такой как хронический циклит, гетерохромный циклит, иридоциклит или циклит Фукса, пурпуру Геноха-Шенлейна, инфекцию, вызываемую вирусом иммунодефицита человека (ВИЧ), инфекцию, вызываемую эховирусом, кардиомиопатию, болезнь Альцгеймера, инфекцию, вызываемую парвовирусом, инфекцию, вызываемую вирусом краснухи, поствакцинационные синдромы, конгенитальную инфекцию, вызываемую краснухой, инфекцию, вызываемую вирусом Эпштейна-Барр, вирус свинки, синдром Эванса, аутоиммунную гонадальную недостаточность, хорею Сиденхема, постстрептококковый нефрит, облитерирующий тромбангиит, тиреотоксикоз, сухотку спинного мозга, хориоидит, гигантоклеточную полимиалгию, эндокринную офтальмопатию, хронический аллергический пневмонит, кератоконъюнктивит сухой, эпидемический кератоконъюнктивит, идиопатический нефритный синдром, нефропатию с минимальными изменениями, доброкачественное семейное поражение и ишемию-реперфузию миокарда, ретинальную патологию, связанную с аутоиммунитетом, воспаление суставов, бронхит, хроническое обструктивное заболевание дыхательных путей (легких), силикоз, афты, афтозный стоматит, артериосклеротические расстройства, аспермию, аутоиммунный гемолиз, болезнь Бека (Boeck), криоглобулинемию, контрактуру Дюпюитрена, факоанафилактический эндофтальмит, аллергический энтерит, лепрозную узловатую (нодозную) эритему, идиопатический фациальный паралич, синдром хронической усталости, ревматическую лихорадку, болезнь Хаммана-Рича, сенсоневральную потерю слуха, пароксизмальную гемоглобулинурию, гипогонадизм, илеит региональный, лейкопению, инфекционный мононуклеоз, поперечный миелит, первичную идиопатическую микседему, нефроз, симпатическую офтальмию, гранулематозный орхит, панкреатит, острый полирадикулит, гангренозную пиодермию, тиреоидит де Кервена, приобретенную спинальную атрофию, бесплодие, вызванное антителами против антигенов, вырабатываемых сперматозоидами, незлокачественную тимому, витилиго, заболевания, ассоциированные с ТКИН (тяжелой комбинированной иммунной недостаточностью, SCID) и вирусом Эпштейна-Барр, синдром приобретенного иммунодефицита (СПИД), паразитарные заболевания, такие как лейшманиоз, синдром токсического шока, пищевое отравление, состояния, включающие инфильтрацию Т-клетками, дефицит лейкоцитарной адгезии, иммунные реакции, ассоциированные с острой аллергией и аллергией замедленного типа,, опосредуемой цитокинами и Т-лимфоцитами, заболевания, включающие диапедез лейкоцитов, синдром множественного поражения органов, заболевания, опосредуемые комплексом антиген-антитело, болезнь анти-БМК антител (БМК-базальная мембрана клубочков) (синдром Гудпасчера), аллергический неврит, аутоиммунные полиэндокринопатии, оофорит, первичную микседему, аутоиммунный атрофический гастрит, симпатическую офтальмию, ревматические заболевания, смешанное соединительнотканное заболевание, нефротический синдром, инсулит, полиэндокринную недостаточность, периферическую нейропатию, аутоиммунный полигландулярный синдром типа I, идиопатический гипопаратиреоидизм (AOIH), тотальную алопецию, застойную ("дилатированную") кардиомиопатию, приобретенный буллезный эпидермолиз (ЕВА), гемохроматоз, миокардит, нефротический синдром, первичный склерозирующий холангит, гнойный и негнойный синусит, острый или хронический синусит, синусит - воспаление слизистой оболочки (иногда и костных стенок) придаточных пазух носа: решетчатой (этмоидит), лобной (фронтит), максиллярной (верхнечелюстной) (гайморит), или клиновидной (основной синусит), расстройство, связанное с эозинофилами, такое как эозинофилия, легочная инфильтрационная эозинофилия, синдром эозинофилии-миалгии, синдром Леффлера, хроническую эозинофильную пневмонию, тропическую легочную эозинофилию, бронхолегочный аспергиллез, аспергиллому или гранулемы, содержащие эозинофилы, анафилаксию, серонегативные спондилоартриты, полиэндокринные аутоиммунные болезни, склерозирующий холангит, склерит, эписклерит, хронический слизисто-кожный кандидоз, синдром Брутона, транзиторную гипогаммаглобулинемию детского возраста, синдром Вискотта-Олдрича (СВО), атаксию-телеангиэктазию, аутоиммунные расстройства, ассоциированные с коллагеновой болезнью, ревматизм, неврологическое заболевание, ишемическое/реперфузионное повреждение, реакцию понижения кровяного давления, вегетативно-сосудистую дисфункцию, ангиэктазию, поражение тканей, ишемию миокарда, гипералгезию, церебральную ишемию, и васкуляризацию, сопутствующую заболеванию, аллергическую гиперчувствительность, гломерулонефриты, реперфузионное поражение, реперфузионное поражение миокарда или других тканей, дерматозы с компонентами острого воспаления, острый гнойный менингит или другие воспалительные расстройства центральной нервной системы, воспалительные расстройства глазничной области, синдромы, ассоциированные с трансфузиями гранулоцитов, цитокин-индуцированную токсичность, острое тяжелое воспаление, стойкое хроническое воспаление, пиелит, цирроз легкого, диабетическую ретинопатию, диабетическое поражение крупных артерий, гиперплазию внутренней оболочки артерий, пептидную язву, вальвулит и эндометриоз.
Термин "антиангиогенезный агент" или "ингибитор ангиогенеза" относится к низкомолекулярному веществу, полинуклеотиду, полипептиду, выделенному белку, рекомбинантному белку, антителу или их конъюгатам или слитым белкам, которые ингибируют ангиогенез, васкулогенез или нежелательную сосудистую проницаемость, либо непосредственно, либо опосредованно. Например, антиангиогенезный агент представляет собой антитело или другой антагонист ангиогенного агента по определению выше, например, антитела к VEGF (например, бевацизумаб (AVASTIN7), bH1, bH1-44, bH1-81), антитела к рецепторам VEGF, низкомолекулярные вещества, которые блокируют передачу сигнала от VEGF рецептора (например, PTK787/ZK2284, SU6668, SUTENT/SU11248 (сунитиниба малат), AMG706). Антиангиогенезные агенты включают также нативные ингибиторы ангиогенеза, например, ангиостатин, эндостатин и т.д. См., например, Klagsbrun and D'Amore, Annu. Rev. Physiol., 53: 217-39 (1991); Streit and Detmar, Oncogene, 22: 3172-3179 (2003) (например, в Таблице 3 приводится антиангиогенная терапия при злокачественной меланоме); Ferrara & Alitalo, Nature Medicine 5(12); 1359-1364 (1999); Tonini et al, Oncogene, 22: 6549-6556 (2003) (например, в Таблице 2 приведены антиангиогенные факторы); и Sato Int. J. din. Oncol., 8: 200-206 (2003) (например, в Таблице 1 приводятся антиангиогенные агенты, применяемые в клинических испытаниях). Дисрегуляция ангиогенеза может вызвать множество расстройств, которые можно лечить с помощью композиций и методов по изобретению. Эти расстройства включают состояния как неопухолевого, так и опухолевого генеза.
Термин "цитотоксический агент" по данному описанию относится к веществу, которое ингибирует или предупреждает функцию клетки и/или вызывает деструкцию клетки. Предполагается, что термин включает радиоактивные изотопы (например, At211, I131, I125, Y90, Re186, Re188, Sm153, Bi212, Ra223, P32 и радиоактивные изотопы Lu), химиотерапевтические агенты, например, метотрексат, адриамицин, алкалоиды винка (винкристин, винбластин, этопозид), доксорубицин, мелфалан, митомицин С, хлорамбуцил, даунорубицин или другие интеркалирующие агенты, ферменты и их фрагменты, такие как нуклеолитические ферменты, антибиотики и токсины, такие как низкомолекулярные токсины или ферментативно активные токсины бактериального, грибкового, растительного или животного происхождения, включая их фрагменты и/или варианты, и различные противоопухолевые или противораковые агенты по данному описанию. Другие цитотоксические агенты представлены в данном описании. Агент, уничтожающий опухолевые клетки, вызывает разрушение (деструкцию) этих клеток.
"Химиотерапевтический агент" представляет собой химическое соединение, применимое для лечения рака. Примеры химиотерапевтических агентов включают алкилирующие агенты, такие как тиотепа и CYTOXAN® циклофосфамид; алкилсульфонаты, такие как бусульфан, импросульфан и пипосульфан; азиридины, такие как бензодопа, карбоквон, метуредопа и уредопа; этиленимины и метилмеламины, включая алтретамин, триэтиленмеламин; триэтиленфосфорамид, триэтилентиофосфорамид и триметилолмеламин; ацетогенины (в особенности буллатацин и буллатацинон); дельта-9-тетрагидроканнабинол (дронабинол, MARINOL®); бета-лапахон; лапахол; колхицины; бетулиновая кислота; камптотецин (включая синтетический аналог топотекан (HYCAMTIN®), СРТ-11 (иринотекан, CAMPTOSAR®), ацетилкамптотецин, скополектин и 9-аминокамптотецин); бриостатин; каллистатин; СС-1065 (включая его синтетические аналоги адозелезин, карзелезин и бизелезин); подофиллотоксин; подофилиновая кислота; тенипозид; криптофицины (в частности, криптофицин 1 и криптофицин 8); доластатин; дуокармицин (включая синтетические аналоги KW-2189 и СВ1-ТМ1); элеутеробин; панкратистатин; саркодиктиин; спонгистатин; азотистые иприты, такие как хлорамбуцил, хлорнафазин, хлорфосфамид, эстрамустин, ифосфамид, мехлоретамид, гидрохлорид оксида мехлоретамина, мелфалан, новембицин, фенестерин, преднимустин, трофосфамид, урацил мустард; нитрозомочевины, такие как кармустин, хлорозотоцин, фотемустин, ломустин, нимустин и ранимустин; антибиотики, такие как ендииновые антибиотики (например, калихеамицин, в особенности калихеамицин гамма 1 (см., например, Agnew, Chem Intl. Ed. Engl., 33: 183-186 (1994)); динемицин, включая динемицин А; эсперамицин; а также хромофор неокарзиностатин и родственные хромопротеиновые хромофоры на основе ендииновых антибиотиков), аклациномизины, актиномицин, аутрамицин, азасерин, блеомицины, кактиномицин, карабицин, карцинофилин, хромомицины, дактиномицин, даунорубицин, деторубицин, 6-диазо-5-оксо-L-норлейцин, ADRIAMYCIN® доксорубицин (включая морфолино-доксорубицин, цианоморфолино-доксорубицин, 2-пирролинодоксорубицин и дезоксидоксорубицин), эпирубицин, эзорубицин, идарубицин, марцелломицин, митомицины, такие как митомицин С, микофеноловая кислота, ногаламицин, оливомицины, пепломицин, потфиромицин, пуромицин, квеламицин, родорубицин, стрептонигрин, стрептозопин, туберцидин, убенимекс, зиностатин, зорубицин; антиметаболиты, такие как метотрексат и 5-фторурацил (5-FU); аналоги фолиевой кислоты, такие как деноптерин, метотрексат, птероптерин, триметрексат; аналоги пурина, такие как флударабин, 6-меркаптопурин, тиампирин, тиогуанин; аналоги пиримидина, такие как анцитабин, азацитидин, 6-азауридин, кармофур, цитарабин, дидезоксиуридин, эноцитабин, флоксуридин; андрогенные стероиды, такие как калустерон, дромастанолона пропионат, эпитиостанол, мепитиостан, тестолактон; препараты, блокирующие синтез гормонов коры надпочечников, такие как аминоглютетимид, митотан, трилостан; компенсатор фолиевой кислоты, такой как фолиновая кислота; ацеглатон; альдофосфамида гликозид; аминолевулиновая кислота; енилурацил; амсакрин; бестрабуцил; бисантрен; эдатрексат; дефофамин; демекольцин; диазиквон; элфорнитин; эллиптиния ацетата; эпотилон; этоглюцид; нитрат галлия; гидроксимочевина; лентинан; лонидаинин; майтанзиноиды, такие как майтанзин и ансамитоцины; митогуазон; митоксантрон; мопиданмол; нитраэрин; фенамет; пирарубицин; лозоксантрон; 2-этилгидразид; прокарбазин; PSK® полисахаридный комплекс (JHS Natural Products, Eugene, OR); разоксан; ризоксин; сизофиран; спирогерманий; тенуазоновая кислота; триазиквон; 2,2',2"-трихлортриэтиламин; трихотецены (в особенности Т-2 токсин, верракурин А, роридин А и ангуидин); уретан; виндезин (ELDISINE®, FILDESIN®); дакарбазин; манномустин; митобронитол; митолактол; пипоброман; гацитозин; арабинозид ("Ara-С"); тиотепа; таксоиды, например, TAXOL® паклитаксел (Bristol-Myers Squibb Oncology, Princeton, N.J.), не содержащий кремофора генно-инженерный препарат наночастиц паклитаксела с альбумином ABRAXANE™ (American Pharmaceutical Partners, Schaumberg, Illinois), TAXOTERE® доксетаксел (Rhone-Poulenc Rorer, Antony, France); хлорамбуцил; гемцитабин (GEMZAR®); 6-тиогуанин; меркаптопурин; метотрексат; платиновые аналоги, такие как цисплатин и карбоплатин; винбластин (VELBAN®); платина; этопозид (VP-16); ифосфамид; митоксантрон; винкристин (ONCOVIN®); оксалиплатин; лейковорин (?); винорельбин (NAVELBINE®); новантрон; эдатрексат; дауномицин; аминоптерин; ибандронат; ингибитор топоизомеразы RFS 2000; дифторметилорнитин (DMFO); ретиноиды, такие как ретиноевая кислота; капецитабин (XELODA®); фармацевтически приемлемые соли, кислоты или производные любого из вышеперечисленных агентов; а также комбинации двух или более вышеописанных агентов, такие как CHOP, сокращенное название (аббревиатура) комбинированного терапевтического средства из циклофосфамида, доксорубицина, винкристина и преднизолона, и FOLFOX, аббревиатура схемы лечения оксиплатином (ELOXATIN™) совместно с 5-FU и лейковорином(?).
Также это определение включает антигормональные агенты, которые регулируют, снижают, блокируют или ингибируют действие гормонов, которые могут промотировать рост раковой опухоли, и часто в виде системного или общего лечения. Они сами могут являться гормонами. Примеры включают антиэстрогены и селективные модуляторы рецепторов эстрогена (SERM), в том числе, например, тамоксифен (включая NOLVADEX® тамоксифен), EVISTA® ралоксифен, дролоксифен, 4-гидрокситамоксифен, триоксифен, кеоксифен, LY 117018, онапристон и FARESTON® торемифен; и антипрогестероны; down-регуляторы рецепторов эстрогена (ERD); агенты, которые подавляют или "отключают" яичники, например, агонисты рилизинг-фактора лютеинизирующего гормона (LHRH), такие как LUPRON® и ELIGARD® лейпролида ацетат, гозерелина ацетат, бузерелина ацетат и триптелин; другие антиандрогены, такие как флутамид, нилутамид и бикалутамид; и ингибиторы ароматазы, которые ингибируют фермент ароматазу, регулирующую продуцирование эстрогенов в коре надпочечника, например, такие как 4(5)-имидазолы, аминоглутетимид, MEGASE® магестрола ацетат, AROMASIN® экземестан, форместан, фадрозол, RIVISOR® ворозол, FEMARA® летрозол и ARIMIDEX® анастрозол. Помимо этого, такое определение химиотерапевтических агентов включает бифосфонаты, такие как клодронат (например, BONEFOS® или OSTAC®), DIDROCAL® этидронат, NE-58095, ZOMETA® золедроновая кислота/золедронат, FOSAMAX® алендронат, AREDIA® памидронат, SKELID® тилудронат или ACTONEL® ризедронат; а также (1,3-диоксолановый аналог цитозинового нуклеозида); антисмысловые олигонуклеотиды, в частности, антисмысловые олигонуклеотиды, которые ингибируют экспрессию генов сигнальных путей аберрантной клеточной пролиферации, например, таких как, PKC-альфа, Raf, H-Ras и рецептора эпидермального фактора роста (EGF-R); вакцины, такие как вакцина THERATOPE® и терапевтические вакцины для генной терапии, например, вакцина ALLOVECTIN®, вакцина LEUVECTIN® и вакцина VAXID®; LURTOTECAN® ингибитор топоизомеразы 1; ABARELIX® rmRH; дитозилат лапатиниба (и ErbB-2 и EGFR двойной низкомолекулярный ингибитор тирозинкиназ, также известный как GW572016); и фармацевтически приемлемые соли, кислоты или производные любого из вышеперечисленных агентов.
Термин "ингибитор роста" по данному описанию относится к соединению или композиции, которые ингибируют рост клетки in vitro или in vivo. Так, ингибитор роста может представлять собой такой агент, который заметно снижает процентное содержание клеток в S фазе. Примеры ингибиторов роста включают агенты, которые блокируют прогрессию клеточного цикла (в фазе, отличной от S фазы), такие как агенты, которые индуцируют блокаду G1 и М-фазы. Классические блокаторы М-фазы включают алкалоиды винка (например, винкристин и винбластин), таксаны и ингибиторы топоизомеразы II, такие как доксорубицин, даунорубицин, этопозид и блеомицин. Агенты, которые блокируют фазу G1, также ("перетекают") блокируют S-фазу, например, агенты алкилирования ДНК, такие как тамоксифен, преднизон, дакарбазин, мехлоретамин, цисплатин, метотрексат, 5-фторурацил и ara-С. Более подробные сведения можно найти в The Molecular Basis of Cancer. Mendelsohn and Israel, eds., Chapter 1, entitled "Cell cycle regulation, oncogenes, and antineoplastic drugs" by Murakami et al. (WB Saunders: Philadelphia, 1995), в особенности стр.13. Таксаны (паклитаксел и доцетаксел) представляют собой противораковые лекарства, полученные из тисового дерева. Доцетаксел (TAXOTERE®, Rhone-Poulenc Rorer), полученный из европейского тиса, представляет собой полусинтетический аналог паклитаксела (TAXOL®, Bristol-Myers Squibb). Паклитаксел и доцетаксел инициируют сборку микротрубочек из димеров тубулина и стабилизируют микротрубочки, предупреждая деполимеризацию, что приводит к ингибированию митоза в клетках.
Выражение "противораковая терапия" по данному описанию относится к лечению, которое ослабляет или ингибирует раковое заболевание у субъекта. Примеры противораковой терапии включают цитотоксическую лучевую терапию, а также введение субъекту терапевтически эффективного количества цитотоксического агента, химиотерапевтического агента, ингибитора роста, противораковой вакцины, ингибитора ангиогенеза, пролекарства, цитокина, антагониста цитокина, кортикостероида, иммуносупрессора, противорвотного агента (антиэметика), антитела или фрагмента антитела или анальгетика.as used herein refers to a treatment that reduces or inhibits cancer in a subject.
Термин "пролекарство", применяется в данной заявке, относится к форме предшественника или производного фармацевтически активного вещества, которое является менее цитотоксическим по отношению к опухолевым клеткам, чем исходное лекарство, и способное ферментативно активироваться или превращаться в более активную исходную форму. См., например, Wilman, "Prodrugs in Cancer Chemotherapy" Biochemical Society Transactions, 14, pp.375-382, 615th Meeting Belfast (1986) и Stella et al., "Prodrugs: A Chemical Approach to Targeted Drug Delivery," Directed Drug Delivery, Borchardt et al., (ed.), pp.247-267, Humana Press (1985). Пролекарства включают, но без ограничения, фосфатсодержащие пролекарства, лекарства, модифицированные D-аминокислотами, гликозилированные пролекарства, бета-лактамсодержащие пролекарства, необязательно замещенные феноксиацетамид-содержащие пролекарства или необязательно замещенные фенилацетамид-содержащие пролекарства, 5-фторцитозиновые и другие 5-фторуридиновые пролекарства, которые можно превратить в более активное цитотоксическое свободное лекарство. Примеры цитотоксических лекарств, которые можно дериватизировать в форму пролекарства для применения в данном изобретении, включают, но без ограничения, описанные выше химиотерапевтические агенты.
Термин "цитокин" является родовым термином для белков, высвобождаемых клетками одной популяции, которые действуют на другую клетку как межклеточные медиаторы. Примерами таких цитокинов являются лимфокины, монокины и обычные полипептидные гормоны. Цитокины включают гормон роста, такой как человеческий гормон роста (HGH), N-метионил человеческий гормон роста и бычий гормон роста; паратиреоидный гормон; тироксин; инсулин; проинсулин; релаксин; прорелаксин; гликопротеиновые гормоны, такие как фолликулостимулирующий гормон (FSH), тиреоидстимулирующий гормон (TSH) и лютеинизирующий гормон (LH); эпидермальный фактор роста (EGF); фактор роста печени; фактор роста фибробластов (FGF); пролактин; плацентарный лактоген; фактор некроза опухолей-альфа и -бета; мюллеровский ингибирующий фактор (MIF); мышиный гонадотропин-ассоциированный пептид; ингибин; активин; сосудистый эндотелиальный фактор роста (VEGF); интегрин; тромбопоэтин (ТРО); факторы роста нерва, такие как NGF (ФРН)-альфа; фактор роста тромбоцитов; трансформирующие факторы роста (TGF), такие как TGF-альфа и TGF-бета; инсулиноподобный фактор роста-I и -II; эритропоэтин (ЕРО); остеоиндуктивные факторы; интерфероны, такие как интерферон-альфа, -бета и -гамма; колониестимулирующие факторы (CSF), такие как макрофагальный -CSF (М-CSF); гранулоцитарно-макрофагальный -CSF (GM-CSF); и гранулоцитарный-CSF (G-CSF); интерлейкины (IL), такие как IL-1, IL-1альфа, IL-2, IL-3, IL-4, IL-5, IL-6, IL-7, IL-8, IL-9, IL-10, IL-11, IL-12; факторы некроза опухолей, такие как TNF-альфа или -бета; и другие полипептидные факторы, включая LIF и kit-лиганд (KL). Термин цитокин по данному описанию включает белки из природных источников или из культуры рекомбинантных клеток и биологически активные эквиваленты цитокинов с нативными последовательностями.
Под "антагонистом цитокина" понимают молекулу, которая частично или полностью блокирует, ингибирует или нейтрализует биологическую активность по меньшей мере одного цитокина. Например, антагонисты цитокинов могут ингибировать активность цитокина, ингибируя экспрессию и/или секрецию цитокина, или связываясь с цитокином или рецептором цитокина. Антагонисты цитокинов включают антитела, синтетические или нативные пептиды, иммуноадгезины и низкомолекулярные антагонисты, которые связываются с цитокином или с цитокиновым рецептором. Антагонист цитокина, необязательно, конъюгируется или сливается с цитотоксическим агентом. Примерами TNF антагонистов являются этанерцепт (ENBREL®), инфликсимаб (REMICADE®) и адалимумаб (HUMIRA™).
Термин "иммуносупрессор (иммунодепрессант)" по данному описанию относится к веществам, которые подавляют или маскируют иммунную систему подлежащего лечению субъекта. Эти агенты включают вещества, которые подавляют продуцирование цитокинов, "down"-регулируют или подавляют экспрессию аутоантигенов или маскируют МНС (ГКГ) антигены. Примеры иммуносупрессоров включают 2-амино-6-арил-5-замещенные пиримидины (см. патент США No. 4665077); микофенолята мофетил, такой как CELLCEPT®; азатиоприн (IMURAN®, AZASAN®/6-меркаптопурин; бромокриптин; даназол; дапсон; глутаральдегид (который маскирует МНС антигены, как описано в патенте США No. 4120649); антиидиотипические антитела для МНС антигенов и МНС фрагментов; циклоспорин А; стероиды, такие как кортикостероиды и глюкокортикостероиды, например, преднизон, преднизолон, такой как PEDIAPRED® (преднизолон натрия фосфат) или ORAPRED® (преднизолон натрия фосфат, раствор для перорального приема), метилпреднизолон и дексаметазон; метотрексат (форма для перорального или подкожного введения) (RHEUMATREX®, TREXALL™); гидроксиклорохин/хлорохин; сульфасалазин; лефлюномид; антагонисты цитокинов или рецепторов цитокинов, включая антитела против интерферона -γ, -β или -α, антитела против фактора некроза опухоли -α (инфликсимаб или адалимумаб), анти-TNFα иммуноадгезин (ENBREL®, этанерцепт), антитела против фактора некроза опухоли -β, антитела против интерлейкина -2 и антитела против IL-2 рецептора; антитела против LFA-1, включая антитела против CD11a и против CD18; антитела против L3T4; гетерологичный антилимфоцитарный глобулин; поликлональные или pan-Т антитела, или моноклональные антитела против CD3 или против CD4/CD4a; растворимый пептид, содержащий LFA-3 связывающий домен (Международная патентная заявка WO 1990/08187, опубликованная 26 июля 1990 года); стрептокиназу; TGF-β; стрептодорназу; РНК или ДНК хозяина; FK506; RS-61443; дезоксиспергуалин; рапамицин; Т-клеточный рецептор (Cohen et al., патент США No. 5114721); фрагменты Т-клеточных рецепторов (Offner et al. Science, 251: 430-432 (1991); Международная патентная заявка WO 1990/11294; laneway. Nature, 341: 482 (1989); и Международная патентная заявка WO 1991/01133); антитела к Т-клеточным рецепторам (Европейский патент ЕР 340109), такие как Т10В9; циклофосфамид (CYTOXAN®); дапсон; пеницилламин (CUPRIMINE®); плазмаферез или иммуноглобулин внутривенно (IVIG). Их можно применять самостоятельно или в комбинации друг с другом, в частности, в виде комбинации стероида и другого иммуносупрессора, или в виде таких комбинаций с последующей поддерживающей дозой с нестероидным агентом для уменьшения потребности в стероидах.
Термин "анальгетик (аналгетик)" относится к лекарству, которое ингибирует или подавляет боль у субъекта. Примеры анальгетиков включают нестероидные противовоспалительные препараты (НСПВП, NSAID), включая ибупрофен (MOTRIN®), напроксен (NAPROSYN®), ацетилсалициловую кислоту, индометацин, сулиндак и толметин, включая их соли и производные, а также различные другие лекарственные средства, применяемые для ослабления острой боли, которая может появиться, включая противосудорожные средства (габапентин, фенилоин, карбамазепин) или трициклические антидепрессанты. Конкретные примеры включают ацетаминофен, аспирин, амитриптилин (ELAVIL®), карбамазепин (TEGRETOL®), фенилтоин (DILANTIN®), габапентин (NEURONTIN®), (Е)-N-ванилил-8-метил-6-нонамид (CAPSAICIN®) или блокаторы нейрона.
Термин "кортикостероид" относится к любому из ряда синтетических или природных веществ с общей химической стероидной структурой, которые имитируют или усиливают действие природных кортикостероидов. Примеры синтетических кортикостероидов включают преднизон, преднизолон (включая метилпреднизолон), дексаметазон триамцинолон и бетаметазон.
"Противораковая вакцина" по данному описанию представляет собой композицию, которая стимулирует у субъекта иммунный ответ против рака. Противораковые вакцины обычно состоят из исходного ассоциированного с раком материала или клеток (антиген), которые могут быть аутологичными (собственные материал или клетки субъекта) или аллогенными (от другого субъекта), совместно с другими компонентами (например, адъювантами), чтобы дополнительно стимулировать и усилить иммунный ответ на антиген. Подходящие противораковые вакцины стимулируют иммунную систему субъекта продуцировать антитела к одному или нескольким специфическим антигенам и/или заставляют киллерные Т клетки атаковать раковые клетки, имеющие такие антигены.
Термин "цитотоксическая лучевая терапия" по данному описанию относится к лучевой терапии, которая ингибирует или предупреждает функцию клеток и/или вызывает деструкцию клеток. Лучевая терапия может включать, например, внешнее облучение или терапию меченым агентом, таким как антитело. Предполагается, что термин включает применение радиоактивных изотопов (например. At211, I131, I125, Y90, Re186, Re188, Sm153, Bi212, Ra223, P32 и радиоактивных изотопов Lu).
"Антиэметик" представляет собой соединение, которое ослабляет или предупреждает тошноту у субъекта. Антиэметики включают, например, антагонисты рецепторов нейрокинина-1, антагонисты рецепторов 5НТЗ (такие как ондансетрон, гранисетрон, трописетрон и затисетрон), агонисты рецепторов GABAB, такие как баклофен, кортикостероид, такой как дексаметазон, KENALOG®, ARISTOCORT® или NASALIDE®, антидопаминергические препараты (нейролептики), фенотиазины (например, прохлорперазин, флуфеназин, тиоридазин и мезоридазин), дронабинол, метроклопрамид, домперидон, галоперидол, циклизин, лоразепам, прохлорперазин и левомепромазин.
Термин "субъект" означает позвоночное, предпочтительно, млекопитающее, более предпочтительно, человека. Млекопитающие включают, но без ограничения, сельскохозяйственных животных (таких как крупный рогатый скот), спортивные животные, домашние животные (такие как кошки, собаки и лошади), приматы, мыши и крысы.
Если не указано иначе, продажные реагенты, используемые в Примерах, применяют в соответствии с инструкциями производителя. Исходные клетки, определяемые в нижеприведенных Примерах и по всему описанию с помощью регистрационных номеров в АТСС, находятся в Американской коллекции типовых культур, Manassas, VA. Если не указано иначе, в настоящем изобретении используются стандартные методы рекомбинантной ДНК, такие как методы, представленные выше в данном описании и в следующих руководствах: Sambrook et al., supra; Ausubel et al., Current Protocols in Molecular Biology (Green Publishing Associates and Wiley Interscience, N.Y., 1989); Innis et al., PCR Protocols: A Guide to Methods and Applications (Academic Press, Inc.: N.Y., 1990); Harlow et al., Antibodies: A Laboratory Manual (Cold Spring Harbor Press: Cold Spring Harbor, 1988); Gait, Oligonucleotide Synthesis (IRL Press: Oxford, 1984); Freshnev. Animal Cell Culture. 1987; Coligan et al., Current Protocols in Immunology, 1991.
Следует понимать, что везде в данном описании и в Формуле изобретения слово "содержать" или его варианты, такие как "содержит" или "содержащий", предполагают включение целого числа или группы целых чисел, но не исключение любого другого целого числа или группы целых чисел.
II. Векторы, клетки-хозяева и методы рекомбинантной ДНК
Для получения антитела по изобретению методами рекомбинантной ДНК нуклеиновую кислоту выделяют и включают в реплицируемый вектор для дальнейшего клонирования (амплификации ДНК) или для экспрессии. ДНК, кодирующую антитело, быстро выделяют и секвенируют обычными методами (например, используя олигонуклеотидные зонды, которые могут специфически связываться с генами, кодирующими тяжелые и легкие цепи антитела). Многие векторы являются доступными. Выбор вектора зависит частично от того, какая клетка-хозяин используется. Как правило, предпочтительными клетками-хозяевами являются клетки либо прокариотического, либо эукариотического (обычно млекопитающих) происхождения. Ясно, что для этой цели можно использовать константные области любого изотипа, включая константные области IgG, IgM, IgA, IgD и IgE и что такие константные области можно получать из иммуноглобулинов человека или животного любых видов.
а. Получение антител с применением прокариотических клеток-хозяев:
i. Конструирование векторов
Полинуклеотидные последовательности, кодирующие полипептидные компоненты антитела по изобретению, можно получать методами рекомбинантной ДНК, Заданные полинуклеотидные последовательности можно получать и секвенировать, используя антителопродуцирующие клетки, такие как гибридомные клетки. Или же, полинуклеотиды можно синтезировать, используя синтезатор полинуклеотидов или метод ПЦР. После получения последовательности, кодирующие полипептиды, встраивают в рекомбинантный вектор, способный реплицироваться и экспрессировать гетерологичные полинуклеотиды в прокариотических хозяевах. Для целей настоящего изобретения можно использовать многие векторы, которые имеются и известны в уровне техники. Выбор соответствующего вектора зависит главным образом от размера нуклеиновых кислот, которые нужно включить в вектор, и от конкретной клетки-хозяина, которую нужно трансформировать вектором. Каждый вектор содержит различные компоненты, в зависимости от их функции (амплификация или экспрессия гетерологичного полинуклеотида или и то, и другое) и их совместимости с конкретной клеткой-хозяином, в которой вектор находится. Компоненты вектора обычно включают, но без ограничения, ориджин репликации, селективный маркерный ген, промотор, сайт связывания рибосомы (RBS), сигнальную последовательность, гетерологичный нуклеотид (инсерт, вставка) и последовательность терминации транскрипции.
Как правило, плазмидные векторы, содержащие последовательность репликона и последовательности контроля, полученные из вида, совместимого с клеткой-хозяином, используют применительно к этим хозяевам. Вектор обычно несет сайт репликации, а также маркерные последовательности, способные обеспечивать фенотипическую селекцию в трансформированных клетках. Например, Е.coli обычно трансформируют, используя pBR322, плазмиду, полученную из вида Е.coli. pBR322 содержит гены, кодирующие резистентность к ампициллину (Amp) и тетрациклину (Tet) и тем самым предоставляет простой метод идентификации трансформированных клеток. Плазмида pBR322, ее производные или плазмиды других микроорганизмов или бактериофаг могут также содержать, или могут быть модифицированы таким образом, чтобы содержать промоторы, которые могут использоваться микроорганизмом для экспрессии эндогенных белков. Примеры pBR322 производных, применяемых для экспрессии конкретных антител, подробно описаны Carter et al., в патенте США No. 5648237.
Помимо этого, фаговые векторы, содержащие последовательность репликона и последовательности контроля, совместимые с микроорганизмом-хозяином, можно использовать применительно к этим хозяевам. Например, бактериофаг, такой как λGEM.TM.-11, можно использовать для получения рекомбинантного вектора, который можно использовать для трансформации восприимчивой клетки-хозяина, такой как E.coli LE392.
Экспрессирующий вектор по изобретению может содержать две или более пары промотор-цистрон, кодирующие каждый из компонентов полипептида. Промотор представляет собой нетранслируемую регуляторную последовательность, расположенную выше (5') по отношению к цистрону, которая модулирует его (цистрона) экспрессию. Прокариотические промоторы обычно делятся на два класса, индуцибельные (индуцируемые) и конститутивные. Индуцибельный промотор представляет собой промотор, который инициирует повышенные уровни транскрипции цистрона под своим контролем в ответ на изменения условий в культуре, например, присутствие или отсутствие питательного вещества или изменение температуры.
Известно большое количество промоторов, узнаваемых различными потенциальными клетками-хозяевами. Выбранный промотор можно функционально связывать с участком ДНК (цистрон), кодирующим легкую или тяжелую цепь, удаляя промотор из исходной ДНК расщеплением рестриктазами и встраивая выделенную промоторную последовательность в вектор по изобретению. Как нативную промоторную последовательность, так и многие гетерологичные промоторы можно использовать для непосредственной амплификации и/или экспрессии генов-мишеней. В некоторых вариантах изобретения применяются гетерологичные промоторы, так как обычно они способствуют повышению транскрипции и увеличению выхода экспрессируемого гена-мишени по сравнению с нативным промотором для целевого полипептида.
Промоторы, пригодные для применения в прокариотических хозяевах, включают PhoA промотор, β-галактамазу и лактозную промоторную систему, промоторную систему триптофана (trp) и гибридные промоторы, такие как tac или trc промотор. Однако также применимы другие промоторы, являющиеся функциональными в бактериях (такие как другие известные бактериальные или фаговые промоторы). Их нуклеотидные последовательности опубликованы, что позволяет специалисту лигировать их к цистронам, кодирующим целевые легкую и тяжелую последовательности (Siebenlist et al., (1980) Cell 20: 269), используя линкеры или адаптеры для того, чтобы предоставить любые необходимые сайты рестрикции.
В одном аспекте изобретения каждый цистрон в рекомбинантном векторе содержит компонент системы секреции сигнальной последовательности, который управляет транслокацией экспрессированных пептидов через мембрану. Обычно сигнальная последовательность может представлять собой компонент вектора или она может являться частью ДНК для целевого полипептида, которая встраивается в вектор. Сигнальная последовательность, выбранная для целей настоящего изобретения, должна представлять собой последовательность, которая узнается клеткой-хозяином и процессируется в ней (т.е. расщепляется сигнальной пептидазой). В случае прокариотических клеток-хозяев, которые не узнают и не процессируют сигнальную последовательность, нативную к гетерологичным полипептидам, сигнальная последовательность заменяется прокариотической сигнальной последовательностью, выбранной, например, из группы, состоящей из лидерных последовательностей щелочной фосфатазы, пенициллиназы, Ipp или термостабильного энтеротоксина II (STII), LamB, PhoE, PelB, OmpA и МВР. В одном варианте изобретения сигнальные последовательности, используемые в обоих цистронах экспрессионной системы, представляют собой STII сигнальные последовательности или их варианты.
В другом аспекте изобретения продуцирование иммуноглобулинов по изобретению может происходить в цитоплазме клетки-хозяина и, следовательно, не требует присутствия сигнальных последовательностей, определяющих секрецию, в каждом цистроне. При этом легкая и тяжелая цепи иммуноглобулина экспрессируются, складываются и собираются с образованием функциональных иммуноглобулинов внутри цитоплазмы. Некоторые штаммы-хозяева (например, штаммы Е.coli trxB) обеспечивают состояние цитоплазмы, предпочтительное для образования дисульфидной связи, тем самым обеспечивается соответствующий фолдинг и сборка субъединиц экспрессируемого белка (Proba and Pluckthun, Gene, 159: 203 (1995)).
Прокариотические клетки-хозяева, пригодные для экспрессии антител по изобретению, включают архебактерии (Archaebacteria) и эубактерии (Eubacteria), такие как грамотрицательные или грамположительные организмы. Примеры применимых бактерий включают эшерихии (Escherichia, например, Е.coli), бациллы (Bacilli) (например, В.subtilis), энтеробактерии, род Pseudomonas (например, Р.aeruginosd), Salmonella typhimurium, Serratia marcescans ("чудесная палочка"), клебсиеллы (Klebsiella), протей (Proteus), шигеллы (Shigella), клубеньковую бактерию (микоризу, Rhizobia), витреосциллы (Vitreoscilla) или паракокк (Paracoccus). В одном варианте изобретения используют грамотрицательные клетки. В одном варианте изобретения в качестве хозяев по изобретению используют клетки Е.coli. Примеры штаммов Е.coli включают штамм W3110 (Bachmann, Cellular and Molecular Biology, vol. 2 (Washington, D.C.: American Society for Microbiology, 1987), pp.1190-1219; ATCC Deposit No. 27325) и их производные, включая штамм 33D3, имеющий генотип W3110 ΔfhuA (ΔtonA) ptr3 lac Iq lacL8 ΔompTΔ(nmpc-fepE) degP41 kanR (патент США No. 5639635). Также используются другие штаммы и их производные, такие как Е.coli 294 (ATCC 31446), Е.coli В, Е.coli λ 1776 (ATCC 31537) и Е.coli RV308 (ATCC 31608). Эти примеры являются скорее иллюстративными, нежели ограничивающими. Методы конструирования производных любых вышеприведенных бактерий, имеющих определенные генотипы, известны в уровне техники и описаны, например, в Bass et al., Proteins, 8: 309-314 (1990). Как правило, требуется выбрать подходящие бактерии с учетом реплицируемости репликона в клетках бактерии. Например, Е.coli, рода Serratia или Salmonella целесообразно использовать в качестве хозяина, когда для пополнения репликона применяют общеизвестные плазмиды, такие как pBR322, pBR325, pACYC177 или pKN410. Как правило, клетка-хозяин должна секретировать минимальные количества протеолитических ферментов, и желательно вводить в клеточную культуру минимальные количества ингибиторов протеаз.
ii. Продуцирование антител
Клетки-хозяева трансформируют с помощью вышеописанных экспрессирующих векторов и культивируют в традиционных питательных средах, модифицированных так, как это необходимо дли индукции промоторов, селекции трансформантов или амплификации генов, кодирующих заданные последовательности
Трансформация означает введение ДНК в прокариотического хозяина таким образом, чтобы ДНК могла реплицироваться, либо в виде внехромосомного элемента, либо с помощью компонента хромосом. В зависимости от используемой клетки-хозяина, трансформацию проводят стандартными методами, подходящими для таких клеток. Обработку кальцием с применением хлорида кальция обычно используют для бактериальных клеток, у которых клеточные стенки являются реальным барьером. В другом методе трансформации используют полиэтиленгликоль/ДМСО. Еще в одном методе используют электропорацию.
Прокариотические клетки, используемые для продуцирования полипептидов по изобретению, выращивают в среде, известной в уровне техники и подходящей для культуры выбранных клеток-хозяев. Примеры подходящих сред включают бульон Лурия (LB) плюс необходимые питательные добавки. В некоторых вариантах изобретения среда содержит также агент для селекции, выбранный с учетом конструкции экспрессирующего вектора, с целью селективно обеспечить рост прокариотических клеток, содержащих экспрессирующий вектор. Например, к среде добавляют ампициллин для роста клеток, экспрессирующих ген резистентности к ампициллину.
Любые необходимые добавки, помимо источников углерода, азота и неорганических фосфатов, также можно вводить в соответствующих концентрациях, самостоятельно или в виде смеси с другой добавкой или средой, такой как комплексный источник азота. Необязательно, культуральная среда может содержать один или более восстанавливающих агентов, выбранных из группы, состоящей из глутатиона, цистеина, цистамина, тиогликолята, дитиоэритритола и дитиотреитола.
Прокариотические клетки-хозяева культивируют при соответствующей температуре. Например, для выращивания Е.coli предпочтительные интервалы температур составляют, примерно, от 20°С, примерно, до 39°С, более предпочтительно, примерно, от 25°С, примерно, до 37°С, еще более предпочтительной температурой является температура, примерно, 30°С. рН среды может представлять собой любую величину рН в интервале, примерно, от 5, примерно, до 9, это зависит, главным образом, от организма-хозяина. Для Е.coli рН, предпочтительно, находится в интервале, примерно, от 6.8, примерно, до 7.4, и, более предпочтительно, это значение равно 7.0.
Если в экспрессирующем векторе по изобретению используется индуцибельный промотор, экспрессия белка индуцируется в условиях, подходящих для активации промотора. В одном аспекте изобретения для регуляции (контроля) транскрипции полипептидов используют промоторы PhoA. Соответственно трансформируемые клетки-хозяева культивируют на фосфат-лимитирующей среде. Предпочтительно, фосфат-лимитирующая среда представляет собой среду C.R.A.P (см., например, Simmons et al., J. Immunol. Methods (2002), 263: 133-147). Как известно в уровне техники, в зависимости от используемой векторной конструкции можно применять различные другие индукторы.
В одном варианте изобретения экспрессированные полипептиды по настоящему изобретению секретируются в периплазму и выделяются (извлекаются) из периплазмы клеток-хозяев. Извлечение (выделение) белка обычно включает разрушение микроорганизма такими методами, как осмотический шок, ультразвуковая обработка или лизис. После разрушения клеток клеточный осадок или целые клетки можно удалить центрифугированием или фильтрацией. Белки можно дополнительно очищать, например, аффинной хроматографией на смоле. Или же, белки можно перенести в культуральную среду и выделять в среде. Клетки можно удалить из культуры, а культуральный супернатант отфильтровать и концентрировать с целью дополнительной очистки полученных белков. Экспрессированные полипептиды можно далее выделять и идентифицировать общеизвестными методами, такими как электрофорез в акриламидном геле (PAGE) и Вестерн-блоттинг.
В одном аспекте изобретения продуцирование антител проводят в большом количестве, применяя ферментативный процесс. Для экспрессии рекомбинантных белков существуют различные крупномасштабные ферментативные процессы с периодической подпиткой. В крупномасштабных ферментативных процессах загрузка (рабочий объем) составляет, предпочтительно, по меньшей мере 1000 литров, предпочтительно, примерно, 1000-100000 литров. В этих ферментерах используются роторные мешалки для распределения кислорода и питательных веществ, в особенности глюкозы (предпочтительного источника углерода/энергии). Выражение "ферментация в малом масштабе" обычно относится к ферментации в ферментере с рабочим объемом не более, примерно, 100 литров и может быть в интервале, примерно, от 1 литра, примерно, до 100 литров.
В ферментативном процессе индукцию экспрессии белка обычно инициируют после того, как клетки выращивают в соответствующих условиях до заданной плотности, например, величина оптической плотности OD550 составляет, примерно, 180-220, на этой стадии клетки находятся в ранней стационарной фазе. Как известно в уровне техники и как описано выше, в зависимости от используемой векторной конструкции можно применять различные индукторы. Клетки перед индукцией можно выращивать в течение более короткого времени. Клетки обычно индуцируют в течение, примерно, 12-50 часов, хотя можно индуцировать клетки в течение более или менее продолжительного периода времени.
Для повышения выхода продукта и качества полипептидов по изобретению можно модифицировать различные условия ферментации. Например, для усовершенствования сборки и фолдинга секретированных полипептидов антитела дополнительные векторы, сверхэкспрессирующие белки-шапероны, такие как Dsb белки (DsbA, DsbB, DsbC, DsbD и/или DsbG) или FkpA (пептидил-пролил-цис/транс-изомераза с шаперонной активностью), можно использовать с целью котрансформации прокариотических клеток-хозяев. Было показано, что белки-шапероны содействуют корректным фолдингу и растворимости геторологичных белков, продуцированных в бактериальных клетках-хозяевах. Chen et al., (1999) J. Biol. Chem. 274: 19601-19605; Georgiou et al., патент США No. 6083715; Georgiou et al., патент США No. 6027888; Bothmann and Pluckthun (2000) J. Biol. Chem. 275: 17100-17105; Ramm and Pluckthun, (2000) J. Biol. Chem. 275: 17106-17113; Arie et al., (2001) Mol. Microbiol. 39: 199-210.
Для минимизации протеолиза экспрессированных гетерологичных белков (в особенности белков, чувствительных к протеолизу) в данном изобретении можно использовать некоторые штаммы-хозяева, дефицитные по протеолитическим ферментам. Например, штаммы клеток-хозяев можно модифицировать таким образом, чтобы повлиять на генетическую(-ие) мутацию(-и) в генах, кодирующих известные бактериальные протеазы, такие как Протеаза III, OmpT, DegP, Tsp, Протеаза I, Протеаза Mi, Протеаза V, Протеаза VI и их комбинации. Имеется несколько Е.coli дефицитных по протеазе штаммов, они описаны, например, Joly et al., (1998), Proc. Natl. Acad. Sci. USA 95: 2773-2777; Georgiou et al., патент США No. 5264365; Georgiou et al., патент США No. 5508192; Нага et al., Microbial Drug Resistance, 2: 63-72 (1996).
В одном варианте изобретения в качестве клеток-хозяев в системе экспрессии по изобретению используют штаммы Е.coli, дефицитные по протеолитическим ферментам и трансформированные плазмидами, сверхэкспрессирующими один или более белок-шаперон.
iii. Очистка антител
Можно применять стандартные методы очистки белков, известные в уровне техники. Следующие методы являются примерами подходящих методов очистки: фракционирование на иммуноаффинных или ионообменных колонках; осаждение (преципитация) этанолом, обращенно-фазовая ВЭЖХ, хроматография на силикагеле или на катионообменной смоле, такой как DEAE, хроматофокусирование, SDS-PAGE, осаждение (преципитация) сульфатом аммония и гель-фильтрация, например, на Сефадексе (Sephadex) G-75.
В одном аспекте для иммуноаффинной очистки полноразмерных антител-продуктов по изобретению применяют Белок А, иммобилизованный на твердой фазе. Белок А представляет собой белок клеточной стенки золотистого стафилококка (Staphylococcus aureus) длиной 41 кДа, который с высокой аффинностью связывается с областью Fc антител. Lindmark et al., (1983) J. Immunol. Meth. 62: 1-13. Твердой фазой, на которой иммобилизован Белок А, предпочтительно, является колонка, имеющая поверхность из стекла или оксида кремния, более предпочтительно, колонка со стеклянной насадкой с контролируемыми порами или колонка с насадкой из кремниевой кислоты. В некоторых применениях на колонке иммобилизован реагент, такой как глицерин, с целью предупредить неспецифическую адгезию примесей.
На первой стадии очистки препарат, полученный из культуры клеток, как описано выше, наносят на твердую фазу с иммобилизованным Белком А, чтобы содействовать специфическому связыванию целевого антитела с Белком А. Затем твердую фазу промывают для удаления примесей, неспецифически связанных с твердой фазой. В конце целевое антитело элюируют с твердой фазы.
б. Получение антител с применением эукариотических клеток-хозяев:
Компоненты вектора обычно включают, но без ограничения, один или более следующих элементов: сигнальную последовательность, ориджин репликации, один или более маркерных генов, энхансер, промотор и последовательность терминации транскрипции.
(i) Компонент сигнальной последовательности
Вектор для применения в эукариотической клетке-хозяине содержит также сигнальную последовательность - последовательность сигнального пептида - или другого полипептида, содержащего специфический сайт расщепления на N-конце целевого зрелого белка или полипептида. Предпочтительно выбираемая гетерологичная сигнальная последовательность представляет собой последовательность, которую узнает и процессирует (например, расщепляет с помощью сигнальной пептидазы) клетка-хозяин. Для экспрессии в клетках млекопитающих существуют сигнальные последовательности млекопитающих, а также вирусные секреторные лидерные последовательности, например, gD сигнал герпесвируса.
ДНК для такой области-предшественника лигируют в рамке считывание к ДНК, кодирующей антитело.
(ii) Ориджин репликации
Обычно ориджин репликации не является необходимым для экспрессирующих векторов для работы с клетками млекопитающих. Например, можно обычно использовать ориджин SV40 только потому, что он содержит ранний промотор.
(iii) Ген селекции (Селективный маркерный ген)
Экспрессирующие и клонирующие векторы могут содержать селективный ген, который также называется селективным маркером (селективным маркерным геном). Типичные гены селекции кодируют белки, которые (а) придают резистентность к антибиотикам или другим токсинам, например, к ампициллину, неомицину, метотрексату или тетрациклину, (б) восполняют ауксотрофный дефицит, если это требуется, или (в) поставляет важные питательные вещества, отсутствующие в комплексной среде.
В одном примере в схеме селекции используется лекарство для прекращения роста клетки-хозяина. Такие клетки, которые успешно трансформируются гетерологичным геном, продуцируют белок, сообщающий резистентность к лекарству и, следовательно, выживание в процессе селекции. В примерах такой доминантной селекции используются лекарства неомицин, микофенольная кислота и гигромицин.
Другими примерами подходящих селективных маркеров для работы в клетках млекопитающих являются такие селективные маркеры, которые способствуют идентификации клеток, компетентных, чтобы принять (оттянуть) нуклеиновую кислоту для антитела, например, гены DHFR (дигидрофолатредуктазы), тимидинкиназы, металлотионеина-I и -II, предпочтительно, гены металлотионеина приматов, аденозиндеаминазы, орнитиндекарбоксилазы и т.д.
Например, клетки, трансформированные селективным маркерным геном DHFR, сначала идентифицируют, культивируют все трансформанты в культуральной среде, которая содержит метотрексат (Mtx), конкурентный антагонист DHFR. В том случае, когда используют DHFR дикого типа, подходящей клеткой-хозяином является линия клеток яичника китайского хомячка (СНО), дефицитная по DHFR активности (например, АТСС CRL-9096).
Или же клетки-хозяева (в частности, хозяева дикого типа, содержащие эндогенный DHFR), трансформированные или котрансформированные с использованием ДНК-последовательности, кодирующие антитело, дикого типа белок DHFR и другой селективный маркер, такой как аминогликозид 3'-фосфотрансфераза (АРН), можно отбирать, используя выращивание клеток в среде, содержащей агент селекции для селективного маркера, такой как аминогликозидный антибиотик, например, канамицин, неомицин или G418. См. патент США No. 4965199.
(iv) Промотор
Экспрессирующие и клонирующие векторы обычно содержат промотор, который распознается организмом-хозяином и функционально связывается с нуклеиновой кислотой, кодирующей полипептид для получения антитела. Промоторные последовательности для эукариот известны. Теоретически все гены эукариот имеют AT-богатую область, локализованную, примерно, на 25-30 оснований выше (5', upstream) сайта инициации транскрипции. Другая последовательность, находящаяся на 70-80 оснований выше (5', upstream) начала транскрипции многих генов, представляет собой область CNCAAT, где N может обозначать любой нуклеотид. На 3' конце большинства генов эукариот находится последовательность ААТААА, которая может служить сигналом для присоединения поли А хвоста к 3' концу кодирующей последовательности. Все эти последовательности обычно встраиваются в эукариотические экспрессирующие векторы.
Транскрипция полипептидов для получения антител с применением векторов в клетках хозяев-млекопитающих контролируется, например, промоторами, полученными из геномов вирусов, таких как вирус полиомы, вирус оспы птиц, аденовирус (такой как аденовирус 2), вирус бычьей папилломы, вирус саркомы птиц, цитомегаловирус, ретровирус, вирус гепатита В и обезьяний вирус (Симиан вирус) 40 (SV40), из промоторов гетерологичного млекопитающего, например, промотор гена актина или промотор гена иммуноглобулина, из промоторов генов теплового шока при условии, что такие промоторы совместимы с системами клеток-хозяев.
Ранний и поздний промоторы вируса SV40 обычно получают в виде рестрикционного фрагмента SV40, который также содержит ориджин репликации SV40 вируса. Предранний промотор цитомегаловируса человека обычно получают в виде рестрикционного фрагмента HindIII E. Система для экспрессии ДНК в хозяевах-млекопитающих с применением в качестве вектора вирус бычьей папилломы раскрывается в патенте США No. 4419446. Модификация этой системы описана в патенте США No. 4601978. Или же в качестве промотора можно использовать длинный концевой повтор вируса саркомы Рауса.
(v) Энхансер
Транскрипция ДНК, кодирующей полипептид для получения антитела по данному изобретению в клетках высших эукариот, часто повышается при введении в вектор энхансерной последовательности. В настоящее время известно множество энхансерных последовательностей из генов млекопитающих (глобина, элактазы, альбумина, α-фетопротеина и инсулина). Однако, как правило, используют энхансер эукариотического вируса. Примеры включают SV40 энхансер на "поздней" стороне ориджина репликации (п.о. 100-270), энхансер раннего промотора цитомегаловируса, энхансер вируса полиомы на "поздней" стороне ориджина репликации и энхансеры аденовирусов. См. также Yaniv, Nature 297: 17-18 (1982) об энхансерах для активации эукариотических промоторов. Энхансер может сплайсироваться в вектор в положение 5' или 3' с последовательностью, кодирующей антитело, но, предпочтительно, локализован в сайте 5' от промотора.
(vi) Компонент терминации транскрипции
Экспрессирующие векторы, используемые в эукариотических клетках-хозяевах, обычно содержат также последовательности, необходимые для терминации транскрипции и для стабилизации мРНК. Такие последовательности обычно получают из 5', и иногда из 3', нетранслируемых областей эукариотических или вирусных ДНК или кДНК. Эти области содержат нуклеотидные сегменты, транскрибируемые в виде полиаденилированных фрагментов в нетранслируемом участке мРНК, кодирующей антитело, одним применяющимся компонентом терминации транскрипции является область полиаденилирования бычьего гормона роста. См. Международную патентную заявку WO 94/11026 и раскрываемый в ней экспрессирующий вектор.
(vii) Селекция и трансформация клеток-хозяев
Подходящие клетки-хозяева для клонирования или экспрессии ДНК в векторах по данному описанию включают клетки высших эукариот по данному описанию, в том числе клетки хозяев-позвоночных. Выращивание клеток позвоночных в культуре (тканевой культуре) стало обычной процедурой. Примерами подходящих линий клеток хозяев-млекопитающих являются линия клеток почек обезьяны CV1, трансформированных с помощью SV40 (COS-7, АТСС CRL 1651); линия клеток почки человеческого эмбриона (клетки 293 или 293, субклонированные для выращивания в суспензионной культуре, Graham et al., J. Gen. Virol. 36: 59 (1977)); клетки яичника детеныша китайского хомячка (ВНК, АТСС CCL 10); клетки яичника китайского хомячка/-DHFR (СНО, Urlaub et al., Proc. Natl. Acad. Sci. USA 77: 4216 (1980)); мышиные клетки Сертоли (ТМ4, Mather, Biol. Reprod. 23: 243-251 (1980)); клетки почки обезьяны (CV1 АТСС CCL 70); клетки почки африканской зеленой мартышки (VERO-76, АТСС CRL-1587); клетки цервикальной карциномы человека (HELA, АТСС CCL 2); клетки почки собак (MDCK, АТСС CCL 34); клетки печени крыс Buffalo (BRL 3А, АТСС CRL 1442); клетки легкого человека (W138, АТСС CCL 75); клетки печени человека (Hep G2, НВ 8065); мышиные опухолевые клетки молочной железы (ММТ 060562, АТСС CCL51); клетки TRI (Mather et al., Annals N.Y. Acad. Sci. 383: 44-68 (1982)); клетки MRC 5; клетки FS4; линия клеток гепатомы человека (Hep G2).
Клетки-хозяева трансформируют с использованием вышеописанной экспрессии клонирующих векторов для получения антитела и культивируют в обычных питательных средах, модифицированных соответствующим образом, с целью индукции промоторов, селекции трансформантов или амплификации генов, кодирующих заданные последовательности.
(viii) Культивирование клеток-хозяев
Клетки-хозяева, используемые для продуцирования антитела по данному изобретению, можно культивировать в различных средах. Для культивирования клеток-хозяев пригодны продажные среды, такие как среда Хэма F10 (Sigma), минимальная поддерживающая среда ((MEM), (Sigma), RPMI-1640 (Sigma) и модифицированная по методу Дульбекко среда Игла ((DMEM), Sigma). Помимо этого, в качестве культуральных сред для выращивания клеток-хозяев можно использовать любые среды, описанные в Ham et al., Mem. Enz. 58: 44 (1979), Barnes et al., Anal. Biochem. 102: 255 (1980), патентах США No. 4767704; 4657866; 4927762; 4560655; или 5122469; Международных патентных заявках WO 90/03430; WO 87/00195; или в заменяющем Re. патенте США 30985. Любая из этих сред при необходимости может быть дополнена гормонами и/или другими факторами роста (такими как инсулин, трансферрин или эпидермальный фактор роста), солями (такими как хлорид натрия, кальция, магния и фосфат), буферами (такими как HEPES), нуклеотидами (такими как аденозин и тимидин), антибиотиками (такими как лекарство GENTAMYCIN™ (гентамицин)), микроэлементами (определяемыми как неорганические соединения, присутствующие обычно в конечных концентрациях, выражаемых в микромолях) и глюкозой или эквивалентным источником энергии. Можно также включать любые другие необходимые добавки в соответствующих концентрациях, которые известны специалистам в данной области техники. В культурах применяют условия, такие как температура, рН и т.п., которые ранее использовались для выращивания клеток-хозяев, выбранных для экспрессии, и известны рядовому специалисту в данной области техники.
(ix) Очистка антитела
При использовании методов рекомбинантной ДНК антитело можно продуцировать внутриклеточно или непосредственно секретировать в следу. Если антитело продуцируется внутриклеточно, на первой стадии осадок макрочастиц, либо клеток-хозяев, либо лизированных фрагментов, удаляют, например, центрифугированием или ультрафильтрацией. Если антитело секретируется в среду, супернатанты из таких экспрессирующих систем обычно сначала концентрируют, используя продажный фильтр для концентрации белков, например, установку для ультрафильтрации Amicon или Millipore Pellicon. На любой из предыдущих стадий может быть включен ингибитор протеаз, такой как PMSF, для ингибирования протеолиза, и могут быть включены антибиотики для предупреждения роста случайных примесей.
Композицию антитела, приготовленную с применением этих клеток, можно очищать, используя, например, хроматографию на гидроксилапатите, электрофорез в геле, диализ, причем предпочтительным методом очистки является аффинная хроматография. Возможность применения Белка А в качестве аффинного лиганда зависит от вида и изотипа любого иммуноглобулинового Fc домена, присутствующего в последовательности антитела. Белок A domain that is present in the antibody. Белок А можно применять для очистки антител на основе человеческих γ1, γ2 или γ4 тяжелых цепей (Lindmark et al., J. Immunol. Meth. 62: 1-13 (1983)). Белок G рекомендуется для всех мышиных изотипов и для человеческой γ3 (Guss et al., EMBO J. 5: 15671575 (1986)). Матрицей, с которой связывается аффинный лиганд, очень часто является агароза, но применяются и другие матрицы. Матрицы, устойчивые к механическому воздействию, такие как пористое стекло или сополимер стирола и дивинилбензола с контролируемым размером пор, позволяют повысить скорость потока и уменьшить время процесса по сравнению с агарозой. Если антитело содержит домен СНЗ, для очистки применима смола Bakerbond ABX™ (J. Т. Baker, Phillipsburg, NJ). Также, в зависимости от выделяемого антитела, применяются другие методы очистки белков, такие как фракционирование на ионообменной колонке, преципитация этанолом, обращено-фазовая ВЭЖХ, хроматография на диоксиде кремния, хроматография на гепарин-Сефарозе SEPHAROSE™, хроматография на катионообменной смоле (например, на колонке с аспарагиновой кислотой), хроматофокусирование, SDS-PAGE и преципитация сульфатом аммония.
После любой (любых) стадии(-ий) предварительной очистки можно проводить хроматографию с гидрофобным взаимодействием при низких значениях рН смеси, содержащей целевое антитело и примеси, используя элюирующий буфер с рН, примерно, 2.5-4.5, предпочтительно, с низкими концентрациями соли (например, концентрация соли около 0-0.25 М).
Иммуноконъюгаты
Изобретение включает также иммуноконъюгаты (иначе называемые "конъюгаты антитело-лекарственное вещество" или "ADC"), содержащие любые из анти-Notch 1 NRR антител по данному описанию, конъюгированные с цитотоксическим агентом, таким как химиотерапевтический агент, лекарственное вещество, ингибитор роста, токсин (например, ферментативно активный токсин бактериального, грибкового, растительного или животного происхождения или их фрагменты), или радиоактивный изотоп (т.е. радиоконъюгат).
Применение конъюгатов антитело-лекарственное вещество для локальной доставки цитотоксических или цитостатических агентов, т.е. лекарств для киллинга или ингибирования опухолевых клеток при лечении рака (Sprig's and Epenetos (1999) Anticancer Research 19:605-614; Niculescu-Duvaz and Springer (1997) Adv. Drg. Del. Rev. 26: 151-172; патент США No. 4975278) способствует адресной доставке лекарственной частицы к опухолям и внутриклеточной аккумуляции в них, в том случае, когда системное введение этих неконъюгированных лекарственных агентов может привести к неприемлемым уровням токсичности в отношении нормальных клеток, а также опухолевых клеток, которые хотят ликвидировать (Baldwin et al., (1986) Lancet pp.(Mar. 15, 1986):603-05; Thorpe, (1985) "Antibody Carriers Of Cytotoxic Agents In Cancer Therapy: A Review," in Monoclonal Antibodies '84: Biological And Clinical Applications, A. Pinchera et al. (eds.), pp.475-506). Тем самым пытаются добиться максимальной эффективности при минимальной токсичности. Сообщалось, что в этих методах применимы как поликлональные антитела, так и моноклональные антитела (Rowland et al., (1986) Cancer Immunol. Immunother., 21: 183-87). Лекарства (лекарственные вещества), применяемые в этих методах, включают дауномицин, доксорубицин, метотрексат и виндезин (Rowland et al., (1986) supra). Токсины, используемые в конъюгатах антитело-токсин, включают бактериальные токсины, такие как дифтерийный токсин, токсины растительного происхождения, такие как рицин, низкомолекулярные токсины, такие как гелданамицин (Mandler et al. (2000) Jour. of the Nat. Cancer Inst. 92(19): 1573-1581; Mandler et al., (2000) Bioorganic & Med. Chem. Letters 10: 1025-1028; Mandler et al., (2002) Bioconjugate Chem. 13: 786-791), майтанзиноиды (Европейский патент ЕР 1391213; Liu et al., (1996) Proc. Natl. Acad. Sci. USA 93: 8618-8623) и калихеамицин (Lode et al., (1998) Cancer Res. 58: 2928; Hinman et al., (1993) Cancer Res. 53: 3336-3342). Токсины могут осуществлять свое цитотоксическое или цитостатическое действие по механизмам, включающим связывание тубулина, связывание ДНК или ингибирование топоизомераз. Некоторые цитотоксические лекарства становятся неактивными или менее активными в результате конъюгации с большими антителами или лигандами белковых рецепторов.
Зевалин (ZEVALIN®, ибритумомаб тиуксетан, Biogen/Idec) представляет собой конъюгат антитело-радиоизотоп, состоящий из мышиного IgG1 каппа моноклонального антитела, специфического к антигену CD20, находящемуся на поверхности нормальных и злокачественных В лимфоцитов, и радиоизотопа 111In или 90Y, связанных содержащим тиомочевинную группу линкером-хелатообразующим соединением (Wiseman et al., (2000) Eur. Jour. Nucl. Med. 27(7): 766-77; Wiseman et al., (2002) Blood 99(12): 4336-42; Witzig et al., (2002) J. Clin. Oncol. 20(10); 2453-63; Witzig et al., (2002) J. Clin. Oncol. 20(15); 3262-69), Хотя Зевалин обладает активностью протии В-клеточной неходжкинской лимфомы (NHL), его введение приводит к тяжелой и продолжительной цитопении у большинства пациентов. MYLOTARG™ (милотарг, гемтузумаб озогамицин, Wyeth Pharmaceuticals), конъюгат антитело-лекарственное вещество, состоящий из huCD33 антитела, связанного с калихеамицином, был одобрен в 2000 году для лечения острого миелоидного лейкоза в виде инъекций (Drugs of the Future (2000) 25(7); 686; патенты США No. 4970198; 5079233; 5585089; 5606040; 5693762; 5739116; 5767285; 5773001). Проводится Фаза II испытаний кантузумаба мертансина (Immunogen, Inc.), конъюгата антитело-лекарственное вещество, состоящего из huC242 антитела, связанного дисульфидным линкером SPP с лекарственной частицей, майтанзиноидом DM1, для лечения раковых заболеваний, при которых экспрессируется CanAg, таких как рак толстой кишки, поджелудочной железы, желудка и т.д. Ведутся разработки по применению MLN-2704 (Millennium Pharm., BZL Biologies, Immunogen Inc.), конъюгата антитела с лекарственным веществом, состоящего из моноклонального антитела против простата-специфического мембранного антигена (PSMA), связанного с лекарственной частицей, майтанзоидом DM1, для возможного лечения раковых заболеваний простаты. Проводятся работы по терапевтическому использованию конъюгатов ауристатиновых пептидов ауристатина Е (АЕ) и монометилауристатина (ММАЕ), синтетических аналогов доластатина, с химерными моноклональными антителами cBR96 (специфическим к антигену Lewis Y при карциномах) и сАС10 (специфическим к CD30 при гематологических злокачественных заболеваниях) (Doronina et al., (2003) Nature Biotechnology 21(7): 778-784).
Химиотерапевтические агенты, применимые для получения иммуноконъюгатов, представлены в данном описании (см. выше). Ферментативно активные токсины и их фрагменты, которые можно использовать, включают А цепь дифтерийного токсина, несвязывающие активные фрагменты дифтерийного токсина, цепь А эндотоксина (из синегнойной палочки (Pseudomonas aeruginosa)), А цепь рицина, А цепь абрина, А цепь модекцина, альфа-сарцин, белки Aleurites fordii (тука Форда), белки диантина, белки Phytolacca americana (лаконоса, PAPI, PAPII и PAP-S), ингибитор momordica charantia (момордика харантская), курцин, кротин, ингибитор Saponaria officinalis (мыльнянку лекарственную), гелонин, митогеллин, рестриктоцин, феномицин, эномицин и трикотесены. См., например. Международную патентную заявку WO 93/21232, опубликованную 28 октября 1993 года. Имеются различные радионуклиды для получения радиоконъюгированных антител. Примеры включают 212Bi, 131I, 131In, 90Y и 186Re. Конъюгаты антитела и цитотоксического агента получают, применяя различные бифункциональные белоксвязывающие агенты, такие как N-сукцинимидил-3-(2-пиридилтиол) пропионат (SPDP), иминотиолан (IT), бифункциональные производные имидоэфиров (такие как диметилдиадипимидат (диметиловый эфир диимида адипиновой кислоты) HCl), активные сложные эфиры (такие как дисукцинимидилсуберат), альдегиды (такие как глутаральдегид), бис-азидосоединения (такие как бис-(п-азидобензоил) гександиамин), бис-диазониевые производные (такие как бис-(п-диазонийбензоил)-этилендиамин), диизоцианаты (такие как толуол 2,6-диизоцианат) и бис-активные фторсодержащие соединения (такие как 1,5-дифтор-2,4-динитробензол). Например, иммунотоксин рицин можно получать как описано в статье Vitetta et al., Science, 238: 1098 (1987). Меченная углеродом-14 1-изотиоцианатобензил-3-метилдиэтилен триаминопентауксусная кислота (МХ-DTPA) является типичным хелатирующим агентом для конъюгации радионуклида с антителом. См. Международную патентную заявку WO 94/11026.
Конъюгаты антитела с одним или более низкомолекулярных токсинов, таких как калихеамицин, майтанзиноиды, доластатины, ауростатины, трихотецен и СС1065, и производные этих токсинов, обладающие активностью токсина, также рассматривается в данном описании.
i. Майтанзин и майтанзиноиды
В некоторых вариантах изобретения иммуноконъюгат содержит антитело (полноразмерное или фрагменты) по изобретению, конъюгированное с одной или более молекул майтанзиноида.
Майтанзиноиды представляют собой митотические ингибиторы, действие которых заключается в ингибировании полимеризации тубулина. Впервые майтанзин был выделен из восточно-африканского кустарника Maytenus serrata (патент США No. 3896111). Впоследствии было обнаружено, что некоторые микробы также продуцируют майтанзоиды, такие как сложные эфиры майтанзинола и С-3 майтанзинола (Патент США No. 4151042). Синтетический майтанзинол и его производные и аналоги раскрываются, например, в патентах США No. 4137230; 4248870; 4256746; 4260608; 4265814; 4294757; 4307016; 4308268; 4308269; 4309428; 4313946; 4315929; 4317821; 4322348; 4331598; 4361650; 4364866; 4424219; 4450254; 4362663;и 4371533.
Молекулы майтанзиноидных лекарственных веществ представляют собой привлекательные лекарственные компоненты в конъюгатах антитело-лекарственное вещество, так как они: (i) сравнительно легко получаются ферментативной или химической модификацией, (ii) склонны к дериватизации с использованием функциональных групп, пригодных для конъюгации с антителами через несульфидные линкеры; t (iii) устойчивы в плазме и (iv) эффективны по отношению к различным линиям опухолевых клеток.
Иммуноконъюгаты, содержащие майтанзиноиды, способ их получения и их терапевтическое применение описаны, например, в патентах США No. 5208020, 5416064 и в Европейском патенте ЕР 0425235 В1, раскрытие которых однозначно вводится в данное описание в качестве ссылки. В статье Liu et al., Proc. Natl. Acad. Sci. USA 93: 8618-8623 (1996) описываются иммуноконъюгаты, содержащие майтанзиноид, обозначенный DM1, связанный с моноклональным антителом С242, специфическим к человеческому колоректальному раку. Обнаружено, что этот конъюгат обладает высокой токсичностью к культивированным клеткам рака толстой кишки и проявляет противоопухолевую активность в in vivo анализе роста опухоли. Chari et al., Cancer Research 52: 127-131 (1992) описывают иммуноконъюгаты, в которых майтанзиноид конъюгирован через дисульфидный линкер с мышиным антителом А7, связывающим антиген на линии человеческих клеток рака толстой кишки, или с другим мышиным моноклональным антителом ТА.1, которое связывает онкоген HER-2/neu. Цитотоксичность конъюгата ТА.1-майтанзиноид проверяли in vitro на линии человеческих клеток рака молочной железы SK-BR-3, которая экспрессирует 3×105 HER-2 поверхностных антигенов на клетку. Конъюгат лекарственного вещества достигал степени цитотоксичности, аналогичной степени цитотоксичности свободного майтанзиноидного лекарственного вещества, которую можно повысить, увеличивая число молекул майтанзиноида на молекулу антитела. Конъюгат А7-майтанзиноид показал низкую системную цитотоксичность на мышах.
Конъюгаты антитело-майтанзиноид получают, связывая химической связью молекулу антитела с молекулой майтанзиноида, при этом биологическая активность как антитела, так и майтанзиноида снижается незначительно. См., например, патент США No. 5208020 (раскрытие которого однозначно вводится в данное описание в качестве ссылки). Конъюгаты, содержащие в среднем 3-4 молекулы майтанзиноида, на молекулу антитела, показали эффективное повышение цитотоксичности клеток-мишеней, при этом майтанзиноид не оказывал отрицательного влияния на функцию или растворимость антитела, хотя предполагалось, что даже одна молекула токсина на молекулу антитела повысит цитотоксичность по сравнению со свободным антителом. Майтанзиноиды хорошо известны в уровне техники, и их можно синтезировать известными методами или выделять из природных источников. Приемлемые майтанзиноиды раскрываются, например, в патенте США No. 5208020 и в других патентах и в непатентных публикациях, ссылки на которые даются выше. Предпочтительными майтанзиноидами являются майтанзинол и аналоги майтанзинола, модифицированные в ароматическом кольце и в других положениях молекулы майтанзиноида, такие как различные сложные эфиры майтанзинола.
В уровне техники известно множество связывающих групп для получения конъюгатов антитело-майтанзиноид, включая, например, связывающие группы, раскрываемые в патенте США No. 5208020 или в Европейском патенте ЕР 0425235 B1, Chari et al., Cancer Research 52: 127-131 (1992), и в патентной заявке США No. 10/960602, поданной 8 октября 2004 года, раскрытие которых однозначно вводится в данное описание в качестве ссылки. Конъюгаты антитело-майтанзиноид, содержащие линкерный компонент SMCC, можно получать, как раскрывается в патентной заявке США No. 10/960602, поданной 8 октября 2004 года. Связывающие группы включают дисульфидные группы, простые тиоэфирные группы, лабильные кислотные группы, фотолабильные (неустойчивые на свету) группы, группы, неустойчивые к действию пептидаз или эстераз, раскрываемые в вышеприведенных патентах, причем предпочтительными являются дисульфидные и простые тиоэфирные группы. Дополнительные связывающие группы описываются и приводятся в данном описании.
Конъюгаты антитела и майтанзиноида можно получать с помощью различных бифункциональных агентов, связывающих белки, таких как N-сукцинимидил-3-(2-пиридилтио) пропионат (SPDP), сукцинимидил-4-(N-малеимидометил) циклогексан-1-карбоксилат (SMCC), иминотиолан (IT), бифункциональные производные имидоэфиров (такие как диметилдиадипимидат HCl), активные сложные эфиры (такие как дисукцинимидилсуберат), альдегиды (такие как глутаральдегид), бис-азидосоединения (такие как бис-(п-азидобензоил) гександиамин), бис-диазониевые производные (такие как бис-(п-диазонийбензоил)-этилендиамин), диизоцианаты (такие как толуол 2,6-диизоцианат) и бис-активные фторсодержащие соединения (такие как 1,5-дифтор-2,4-динитробензол). Особенно предпочтительные конденсирующие агенты для введения дисульфидной связи включают N-сукцинимидил-3-(2-пиридилтио) пропионат (SPDP) (Carlsson et al., Biochem. J. 173: 723-737 (1978)) и N-сукцинимидил-4-(2-пиридилтио)пентаноат (SPP).
Линкер может связываться с молекулой майтанзоида в различных положениях, в зависимости от типа связи. Например, сложноэфирную связь можно получать по реакции с гидроксильной группой обычными методами конденсации. Реакция может идти по С-3 положению, содержащему гидроксильную группу, по С-14 положению, модифицированному гидроксиметильной группой, по С-15 положению, модифицированному гидроксильной группой, и по С-20 положению, имеющему гидроксильную группу. В предпочтительном варианте изобретения связь образуется в положении С-3 майтанзинола или майтанзиноидного аналога.
ii. Ауристатины и доластатины
В некоторых вариантах изобретения иммуноконъюгаты содержат антитело по изобретению, конъюгированное с доластатинами или с пептидными аналогами или производными доластатина, ауристатинами (патенты США No. 5635483 и 5780588). Было показано, что доластатины и ауристатины нарушают динамику микротрубочек, гидролиз GTP и деление ядра и клетки (Woyke et al (2001) Antimicrob. Agents and Chemother. 45(12): 3580-3584) и обладают противораковой (патент США No. 5663149) и противогрибковой активностью (Pettit et al., (1998) Antimicrob. Agents Chemother. 42: 2961-2965). Молекулу доластатин или ауристатина можно связывать с антителом по N (амино) концу или по С (карбоксильному) концу пептидной молекулы (Международная патентная заявка WO 02/088172).
Примеры вариантов изобретения с ауристатином включают связанные с N-концом молекулы лекарственного вещества монометилауристатина DE и DF, раскрываемые в патентной заявке США "Monomethylvaline Compounds Capable of Conjugation to Ligands," U.S. Ser. No. 10/983340, поданной 5 ноября 2004 года, раскрытие которой однозначно во всей полноте вводится в данное описание в качестве ссылки.
Как правило, пептидные молекулы лекарственного вещества можно получать образованием пептидной связи между двумя или более аминокислотами и/или пептидными фрагментами. Такие пептидные связи можно получать, например, методом жидкофазного синтеза (см. Е. Schroder and K. Lubke, "The Peptides," volume 1, pp.76-136, 1965, Academic Press), общеизвестными в химии пептидов. Молекулы лекарственного вещества ауристатина/доластатина можно получать методами, описанными в: патентах США No. 5635483 и 5780588; Pettit et al., (1989) J. Am. Chem. Soc. 111: 5463-5465; Pettit et al., (1998) Anti-Cancer Drug Design 13: 243-277; Pettit, G.R., et al., Synthesis, 1996, 719-725; и Pettit et al., (1996) J. Chem. Soc. Perkin Trans. 1 5: 859-863. См. также Doronina (2003) Nat. Biotechnol. 21(7): 778-784; "Monomethylvaline Compounds Capable of Conjugation to Ligands," заявку США US20050238649, опубликованную 27 октября 2005 года, вводимую в данное описание ссылкой во всей полноте (раскрывающую, например, линкеры и методы получения соединений монометилвалина, таких как ММАЕ и MMAF, конъюгированных с линкерами).
iii. Калихеамицин
В других вариантах изобретения иммуноконъюгат содержит антитело по изобретению, конъюгированное с одной или более молекул калихеамицина. Антибиотики семейства калихеамицина способны вызывать разрыва двухнитевой ДНК в субпикомолярных концентрациях. О получении конъюгатов с антибиотиками семейства калихеамицина см. патенты США No. 5712374, 5714586, 5739116, 5767285, 5770701, 5770710, 5773001 и 5877296 (все они принадлежат American Cyanamid Company), Структурные аналоги калихеамицина, которые можно использовать, включают, но без ограничения, γ1 I, α2 I, α3 I, N-ацетил-γ1 I, PSAG и θI 1) (Hinman et al., Cancer Research 53: 3336-3342 (1993), Lode et al., Cancer Research 58: 2925-2928 (1998) и вышеуказанные патенты США, выданные компании American Cyanamid). Другим противораковым лекарством, с которым может конъюгировать антитело, является QFA, представляющее собой антифолат. Как у калихеамицина, так и у QFA, имеются внутриклеточные сайты действия и они с трудом проникают через плазматическую мембрану. Таким образом, клеточное поглощение этих агентов с помощью антитела, опосредующего интернализацию, значительно повышает их цитотоксический эффект.
iv. Другие цитотоксические агенты
Другие противоопухолевые агенты, которые могут конъюгироваться с антителами по изобретению, включают BCNU, стрептозоцин, винкристин и 5-фторурацил, семейство агентов, известных под общим названием комплекс LL-Е33288, описанный в патентах США No. 5053394 и 5770710, а также эсперамицины (патент США No. 5877296).
Ферментативно активные токсины и их фрагменты, которые можно использовать, включают А цепь дифтерийного токсина, несвязывающие активные фрагменты дифтерийного токсина, цепь А эндотоксина (из синегнойной палочки (Pseudomonas aeruginosa)), А цепь рицина, А цепь абрина, А цепь модекцина, альфа-сарцин, белки Aleurites fordii (тука Форда), белки диантина, белки Phytolacca americana (лаконоса) (PAPI, PAPII и PAP-S), ингибитор momordica charantia (момордика харантская), курцин, кротин, ингибитор Saponaria officinalis (мыльнянку лекарственную), гелонин, митогеллин, рестриктоцин, феномицин, эномицин и трикотесены. См., например, Международную патентную заявку WO 93/21232, опубликованную 28 октября 1993 года.
Помимо этого настоящее изобретение рассматривает иммуноконъюгат, образованный антителом и соединением с нуклеолитической активностью (например, рибонуклеазой или ДНК эндонуклеазой, такой как дезоксирибонуклеаза; ДНКаза).
Для селективной деструкции опухоли антитело может содержать атом с высокой радиоактивностью. Для получения радиоактивных конъюгатов антител имеются различные радиоактивные изотопы. Примеры включают At211, I131, I125, Y90, Re186, Re188, Sm153, Bi212, P32, Pb212 и радиоактивные изотопы Lu. Если конъюгат используют для детекции, он может содержать радиоактивный атом для сцинтиграфических исследований, например tc99m или I123, или спиновую метку для визуализации методом ЯМР (ядерного магнитного резонанса, mri (магнитно-резонансного исследования, МРИ)), такую как йод-123, йод-131, индий-111, фтор-19, углерод-13, азот-15, кислород-17, гадолиний, марганец или железо.
Радиоактивная (радио-) или другие метки можно включать в конъюгат известными методами. Например, можно получать методами биосинтеза или химического синтеза аминокислот, используя подходящие аминокислоты-предшественники, содержащие, например, изотоп фтор-19 вместо атома водорода. Метки, такие как Tc99m или I123, Re186, Re188 и In111, можно связывать с помощью остатка цистеина в пептиде. Иттрий-90 можно связывать через лизиновый остаток. Метод введения изотопа с применением йодогена (IODOGEN) (Fraker et al (1978) Biochem. Biophys. Res. Commun. 80: 49-57) можно применять для включения изотопа йода-123. Другие методы подробно описаны в книге "Monoclonal Antibodies in Immunoscintigraphy" (Chatal, CRC Press 1989).
Конъюгаты антитела и цитостатического агента можно получать с помощью различных бифункциональных агентов, связывающих белки (конденсирующих агентов), таких как N-сукцинимидил-3-(2-пиридилтио) пропионат (SPDP), сукцинимидил-4-(N-малеимидометил) циклогексан-1-карбоксилат (SMCC), иминотиолан (IT), бифункциональные производные имидоэфиров (такие как диметилдиадипимидат HCl), активные сложные эфиры (такие как дисукцинимидилсуберат), альдегиды (такие как глутаральдегид), бис-азидосоединения (такие как бис-(п-азидобензоил) гександиамин), бис-диазониевые производные (такие как бис-(п-диазонийбензоил)-этилендиамин), диизоцианаты (такие как толуол 2,6-диизоцианат) и бис-активные фторсодержащие соединения (такие как 1,5-дифтор-2,4-динитробензол). Например, иммунотоксин рицин можно получать, как описано в статье Vitetta et al., Science, 238: 1098 (1987). Меченная углеродом-14 1-изотиоцианатобензил-3-метилдиэтилен триаминопентауксусная кислота (MX-DTPA) является типичным хелатирующим агентом для конъюгации радионуклида с антителом. См. Международную патентную заявку WO 94/11026. Линкер может представлять собой "отщепляемый линкер", способствующий высвобождению цитотоксического лекарственного вещества в клетке. Например, можно использовать линкер, неустойчивый в кислой среде, линкер, чувствительный к действию пептидаз, фотолабильный линкер, линкер, содержащий диметильную или дисульфидную группу (Chari et al., Cancer Research 52: 127-131 (1992); патент США No. 5208020).
Соединения по изобретению однозначно включают, но без ограничения, конъюгаты антитело-лекарственное вещество (ADC), полученные с помощью кросс-линкеров (сшивающих агентов): BMPS, EMCS, GMBS, HBVS, LC-SMCC, MBS, МРВН, SBAP, SIA, SIAB, SMCC, SMPB, SMPH, сульфо-EMCS, сульфо-GMBS, сульфо-KMUS, сульфо-MBS, сульфо SIAB, сульфо-SMCC и сульфо-SMPB, и SVSB (сукцинимидил-(4-винилсульфон)бензоат), который выпускается промышленно (например, Pierce Biotechnology, Inc., Rockford, IL., U.S.A). См. страницы 467-498, 2003-2004 Applications Handbook and Catalog.
v. Получение конъюгатов антитело-лекарственное вещество
В конъюгатах антитела с лекарственным веществом (ADC) по изобретению антитело (Ab)конъюгируется с одной или более молекул лекарственного вещества (D), например, в примерном соотношении от 1 до 20 молекул лекарственного вещества на молекулу антитела, с помощью линкера (L). ADC Формулы I можно получать несколькими способами, применяя химические реакции органических соединений, условия и реагенты для химических реакций, известные специалистам в данной области техники, включая: (1) реакцию нуклеофильной группы антитела с двухвалентным линкером с образованием Ab-L, связанных ковалентной связью, с последующей реакцией с молекулой лекарственного вещества D; и (2) реакцию нуклеофильной группы молекулы лекарственного вещества с дивалентным линкером с образованием D-L, связанных ковалентной связью, с последующей реакцией с нуклеофильной группой антитела. Дополнительные методы получения ADC представлены в данном описании.

Линкер может состоять из одного или более компонентов. Примеры линкерных компонентов включают 6-малеимидокапроил ("МС"), малеимидопропаноил ("МР"), валин-цитруллин ("val-cit"), аланин-фенилаланин ("ala-phe"), п-бензилоксикарбонил ("РАВ"), N-сукцинимидил 4-(2-пиридилтио) пентаноат ("SPP"), N-сукцинимидил 4-(N-малеидометил) циклогексан-1 карбоксилат ("SMCC') и N-сукцинимидил (4-йод-ацетил) аминобензоат ("SIAB"). В уровне техники известны другие линкерные компоненты и некоторые из них представлены в данном описании. См. также патентную заявку США "Monomethylvaline Compounds Capable of Conjugation to Ligands," U.S. Ser. No. 10/983340, поданную 5 ноября 2004 года, содержание которой вводится в данное описание ссылкой во всей полноте.
В некоторых вариантах изобретения линкер может содержать аминокислотные остатки. Типичные аминокислотные линкерные компоненты включают дипептид, трипептид, тетрапептид или пентапептид. Примеры дипептидов включают: валин-цитруллин (vc или val-cit), аланин-фенилаланин (af или ala-phe). Примеры трипептидов включают: глицин-валин-цитруллин (gly-val-cit) и глицин-глицин-глицин (gly-gly-gly). Аминокислотные остатки, которые содержат аминокислотный линкерный компонент, включают натуральные (естественные, природные) аминокислотные остатки, а также минорные аминокислоты и неприродные аналоги аминокислот, такие как цитруллин. Аминокислотные линкерные компоненты можно создать и оптимизировать по селективности к ферментативному расщеплению конкретными ферментами, например, ассоциированной с опухолью протеазой, катепсином В, С и D или протеазой плазмином.
Нуклеофильные группы в антителах включают, но без ограничения: (i) N-концевые аминогруппы, (ii) аминогруппы боковых цепей, например, лизина, (iii) тиольные группы боковых цепей, например, цистеина, и (iv) гидроксильные или аминогруппы сахаров, если антитело является гликозилированным. Амино-, тиольные и гидроксильные группы являются нуклеофильными и способны реагировать с образованием ковалентных связей с электрофильными группами в линкерных частицах и линкерных реагентах, включающих: (i) активные сложноэфирные группы, такие как NHS сложноэфирные группы, HOBt сложноэфирные группы, галогенформиаты и галогенангидриды кислот; (ii) алкил- и бензилгалогениды, такие как галогенацетамиды; (iii) альдегиды, кетоны, карбоксильные и малеимидогруппы. Некоторые антитела содержат восстанавливаемые межцепные дисульфидные группы, например, цистеиновые мостики. Антитела можно сделать способными к конъюгации с линкерными реагентами обработкой их восстановителем, таким как DTT (дитиотреитол). При этом каждый цистеиновый мостик, теоретически, образует две реакционно-способных тиольных нуклеофильных группы. Дополнительные нуклеофильные группы можно ввести в антитела по реакции лизиновых остатков с 2-иминотиоланом (реагентом Траута), при этом аминогруппа превращается в тиольную. Реакционно-способные тиольные группы можно вводить в антитело (или его фрагмент), вводя один, два, три, четыре или более цистеиновых остатка (например, получая мутантные антитела, содержащие один или более не-нативных цистеиновых аминокислотных остатков).
Конъюгаты антитело-лекарственное вещество по изобретению можно также получать модификацией антитела, вводя электрофильные группы, которые могут реагировать с нуклеофильными заместителями в линкерном реагенте или лекарственном веществе. Сахара (углеводные группы) в гликозилированных антителах можно окислять, например, периодатами, с образованием альдегидной или кетогруппы, которые могут реагировать с аминогруппой линкерных реагентов или молекул лекарственного вещества. Полученные иминогруппы шиффова основания могут образовывать стабильную связь или их можно восстанавливать, например, борогидридами, с образованием устойчивых аминных групп. В одном варианте изобретения в результате реакции углеводного фрагмента гликозилированного антитела либо с галактоз-оксидазой, либо с метапериодатом натрия могут образоваться карбонильные (альдегидные и кето-) группы в белке, которые могут реагировать с соответствующими группами в лекарственном веществе (Hermanson, Bioconjugate Techniques). В другом варианте изобретения белки, содержащие N-концевой сериновый или треониновый остатки, могут реагировать с метапериодатом натрия, что приводит к образованию альдегида вместо первой аминокислоты (Geoghegan & Stroh, (1992) Bioconjugate Chem. 3: 138-146; патент США No. 5362852). Такой альдегид может реагировать с лекарственным веществом или с нуклеофильной группой линкера.
Аналогично, нуклеофильные группы в молекуле лекарственного вещества включают, но без ограничения: амино-, тиольную, гидроксильную, гидразидную, оксимную, гидразинную, тиосемикарбазидную, гидразинкарбоксилатную и арилгидразидную группы, способные образовывать ковалентные связи с электрофильными группами в линкерных частицах и линкерных реагентах, включая: (i) активные сложноэфирные группы, такие как NHS сложноэфирные группы, HOBt сложноэфирные группы, галогенформиаты и галогенангидриды кислот; (ii) алкил- и бензилгалогениды, такие как галогенацетамиды; (iii) альдегидные, кето-, карбоксильные и малеимидогруппы.
Или же, слитый белок, содержащий антитело и цитотоксический агент, можно получать, например, методами рекомбинантной ДНК или пептидным синтезом. ДНК может содержать соответствующие области, кодирующие два участка конъюгата, либо прилегающие друг к другу, либо разделенные областью, кодирующей линкерный пептид, который не нарушает заданные свойства конъюгата.
Еще в одном варианте изобретения антитело может конъюгироваться с "рецептором" (таким как стрептавидин) для предварительного нацеливания на опухоль, причем конъюгат антитело-рецептор вводят индивидууму, а затем удаляют несвязанный конъюгат из кровотока, используя агент, содействующий его выведению (клиренсу), а затем вводят "лиганд" (например, авидин), который конъюгирован с цитотоксическим агентом (например, радионуклидом).
Фармацевтические препараты
Терапевтические препараты, содержащие антитело по изобретению, готовят для хранения смешением антитела нужной степени чистоты с необязательными физиологически приемлемыми носителями, эксципиентами или стабилизаторами (Remington: The Science and Practice of Pharmacy 20th edition (2000)) в виде водных растворов, лиофилизированных или других сухих препаратов. Приемлемые носители, эксципиенты или стабилизаторы нетоксичны по отношению к реципиентам в применяемых дозах и в концентрациях и включают буферы, такие как фосфатный, нитратный, гистидиновый и буферные растворы, содержащие другие органические кислоты; антиоксиданты, включающие аскорбиновую кислоту и метионин; консерванты (такие как хлорид октадецилдиметилбензиламмония; хлорид гексаметония; хлорид бензалкония; хлорид бензетония; фенол, бутиловый или бензиловый спирт; алкилпарабены, такие как метил- или пропилпарабен; катехин; резорцин; циклогексанол; 3-пентанол; и м-крезол; низкомолекулярные (содержащие менее 10 аминокислотных остатков) полипептиды; белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстрины; хелатирующие агенты, такие как EDTA; сахара, такие как сахароза, маннит, трегалоза или сорбит; солеобразующие противоионы, такие как катион натрия; комплексы металлов (например, комплексы Zn-белок); и/или неионные поверхностно-активные вещества, такие как Твин (TWEEN™), Плуроникс (PLURONICS™) или полиэтиленгликоль (ПЭГ, PEG).
Если это необходимо по конкретным подлежащим лечению показаниям, препарат по данному описанию может также содержать более одного активного соединения, предпочтительно, с комплементарной активностью, причем эти активные соединения не оказывают отрицательного воздействия друг на друга. Соответственно, такие молекулы присутствуют в комбинации в количествах, эффективных для осуществления предполагаемой задачи.
Можно также приготовить препараты, в которых активные ингредиенты заключены в микрокапсулы, например, методами коацервации или межфазной полимеризации, например, микрокапсулы из гидроксиметилцеллюлозы или желатиновые микрокапсулы или микрокапсулы из полиметилметакрилата, соответственно, препараты в коллоидных системах доставки (например, в виде липосом, альбуминовых микросфер, микроэмульсий, наночастиц и нанокапсул) или в макроэмульсиях. Такие методы раскрываются в руководстве Remington: The Science and Practice of Pharmacy 20th edition (2000).
Препараты для применения in vivo должны быть стерильными. Это легко осуществить фильтрованием через стерильные мембранные фильтры.
Можно приготовить препараты с пролонгированным высвобождением. Подходящие примеры препаратов с пролонгированным высвобождением включают полупроницаемые матрицы из твердых гидрофобных полимеров, содержащие иммуноглобулин по изобретению, причем эти матрицы представляют собой оформленные изделия, например, пленки или микрокапсулы. Примеры матриц с пролонгированным высвобождением включают сложные полиэфиры, гидрогели (например, поли(2-гидроксиэтилметакрилат) или поливиниловый спирт, полилактиды (патент США No. 3773919), сополимеры L-глутаминовой кислоты и γ-этил-L-глутамата, неразлагающийся сополимер этилена с винилацетатом, разлагающиеся (расщепляемые) сополимеры молочной и гликолевой кислот, такие как LUPRON DEPOT™ (инъецируемые микросферы, состоящие из сополимера молочной кислоты с гликолевой кислотой и лейпролидацетата) и поли-D-(-)-3-гидроксимасляная кислота. В то время как полимеры, такие как сополимер этилена с винилацетатом и сополимер молочной кислоты с гликолевой кислотой, способствуют высвобождению молекул в течение более чем 100 дней, некоторые гидрогели высвобождают белки за более короткий период времени. Инкапсулированные иммуноглобулины остаются в организме продолжительное время, они могут денатурироваться или агрегировать под воздействием влаги при 37°С, что приводит к потере биологической активности и возможным изменениям иммуногенности. Можно разработать правильную стратегию стабилизации в зависимости от задействованного механизма. Например, если найдено, что механизм агрегации представляет собой образование внутримолекулярной S-S связи за счет тио-дисульфидного взаимодействия, стабилизацию можно осуществить модификацией сульфгидрильных остатков, лиофилизацией из кислых растворов, контролем содержания влаги, применением соответствующих добавок и разработкой специфических композиций полимерных матриц.
III. Терапевтическое применение
Антитела и фрагменты антител по данному описанию, которые связывают как HER2, так и VEGF, например, bH1-44 или bH1-88 или их фрагменты можно применять для лечения опухолей, включая предраковые, неметастатические и раковые опухоли (например, рак на ранних стадиях), для лечения аутоиммунного заболевания, для лечения нарушения ангиогенеза, для лечения заболевания, включающего анормальную активацию HER2, или для лечения субъекта с повышенным риском развития рака (например, рака молочной железы, колоректального рака, рака легкого, почечноклеточной карциномы, глиомы или рака яичника), нарушения ангиогенеза, аутоиммунного заболевания или заболевания, включающего анормальную активацию HER2.
Термин "рак" охватывает группу пролиферативных расстройств, включая, но без ограничения, предраковые новообразования, доброкачественные опухоли и злокачественные опухоли. Доброкачественные опухоли остаются локализованными в месте образования и не способны проникать, распространяться на дальние расстояния или давать там метастазы. Злокачественные опухоли инвазируют и поражают другие близлежащие ткани. Они могут также приобретать способность отрываться от места их образования и проникать в другие области организма (метастазировать), обычно через кровоток или через лимфатическую систему, в которой локализованы лимфатические узлы. Первичные опухоли классифицируются в зависимости от типа ткани, из которой они возникают; метастатические опухоли делят на классы в зависимости от типа тканей, из которых образованы раковые клетки. С течением времени клетки злокачественной опухоли становятся более анормальными и выглядят менее похожими на нормальные клетки. Это изменение внешнего вида раковых клеток называют стадией опухоли, а раковые клетки описывают как хорошо дифференцированные, умеренно дифференцированные, плохо дифференцированные (слабодифференцированные) или недифференцированные. Хорошо дифференцированные клетки выглядят на вид совершенно нормальными и напоминают нормальные клетки, из которых они произошли. Недифференцированные клетки представляют собой клетки, которые стали настолько анормальными, что более невозможно определить происхождение клеток.
Опухоль может представлять собой солидную опухоль или несолидную опухоль или опухоль мягких тканей. Примеры опухолей мягких тканей включают лейкоз (например, хронический миелогенный лейкоз, острый миелогенный лейкоз, острый лимфобластный лейкоз взрослых, В-клеточный острый лимфобластный лейкоз из зрелых В-клеток, хронический лимфобластный лейкоз, пролимфоцитарный лейкоз или волосатоклеточный лейкоз), или лимфому (например, неходжкинскую лимфому, кожную Т-клеточную лимфому или болезнь Ходжкина). Солидная опухоль включает рак любых тканей организма, отличных от крови, костного мозга или лимфатической системы. Солидные опухоли можно далее разделить на солидные опухоли из эпителиальных клеток и солидные опухоли из неэпителиальных клеток. Примеры солидных опухолей из эпителиальных клеток включают опухоли желудочно-кишечного тракта, толстой кишки, молочной железы, предстательной железы (простаты), легкого, почки, печени, поджелудочной железы, яичника, головы и шеи, полости рта, желудка, двенадцатиперстной кишки, тонкого кишечника, толстого кишечника, ануса, желчного пузыря, губы, носоглотки, кожи, матки, мужских половых органов, органов мочеиспускания и мочевого пузыря. Солидные опухоли неэпителиального происхождения включают саркомы, опухоли головного мозга и опухоли костных тканей.
Эпителиальные раковые опухоли обычно развиваются из доброкачественной опухоли в преинвазивную стадию (например, карциному in situ), в злокачественный рак, который проникает через базальную мембрану и инвазирует субэпителиальную строму.
Полиспецифические антитела, которые связывают как VEGF, так и HER2 (например, bH1-44 или bH1-88 или их фрагмент), желательно применять для лечения рака молочной железы, колоректального рака, рака легкого, почечноклеточной карциномы, глиомы или рака яичника.
В настоящее время точно установлено, что ангиогенез непосредственно связан с патогенезом различных расстройств. Эти расстройства включают солидные опухоли и метастазирование, атеросклероз, ретролентальную фиброплазию, гемангиомы, хроническое воспаление, заболевания с внутриглазной неоваскуляризацией, такие как пролиферативные ретинопатии, например, диабетическая ретинопатия, возрастная дегенерация желтого пятна (AMD), неоваскулярная глаукома, иммунное отторжение трансплантированной роговичной ткани и других тканей, ревматоидный артрит и псориаз. Folkman et al., J. Biol. Chem., 267: 10931-10934 (1992); Klagsbrun et al., Annu. Rev. Physiol. 53: 217-239 (1991); и Gamer A., "Vascular diseases", In: Pathobiology of Ocular Disease. A Dynamic Approach, Gamer A., Klintworth GK, eds., 2nd Edition (Marcel Dekker, NY, 1994), pp. 1625-1710.
Анормальный ангиогенез происходит, когда новые кровеносные сосуды либо растут избыточно, недостаточно или нецелесообразно (например, локализация, время или начало ангиогенеза нежелательно с медицинской точки зрения) в болезненном состоянии, либо таким образом, что это вызывает болезненное состояние. Избыточный, недостаточный или неконтролируемый ангиогенез наблюдается, когда происходит рост новых кровеносных сосудов, который способствует ухудшению болезненного состояния или вызывает болезненное состояние, такое как рак, в особенности васкуляризованные солидные опухоли и метастатические опухоли (включая рак толстой кишки, рак легкого (в особенности мелкоклеточный рак легкого) или рак простаты), заболевания, вызванные глазной неоваскуляризацией, в особенности диабетическая слепота, ретинопатии, главным образом диабетическая ретинопатия или возрастная дегенерация желтого пятна (AMD), диабетический макулярный отек, отек головного мозга (например, связанный с острым инсультом/ закрытым повреждением/травмой головы), синовиальное воспаление, появление паннуса при ревматоидном артрите, оссифицирующий миозит, гипертрофию костной ткани, рефракторный асцит, поликистоз яичника, 3rd spacing of fluid diseases (панкреатит, компартмент синдром, ожоги, кишечное заболевание), фиброму матки, преждевременные роды, неоваскуляризацию радужки (рубеоз), плевральный экссудат при злокачественной опухоли, рестеноз сосудов, гемангиобластому, такую как гемангиома; воспалительные заболевания почек, такие как гломерулонефрит, в особенности мезангиопролиферативный гломерулонефрит, гемолитический уремический синдром, диабетическая нефропатия или гипертонический нефросклероз, различные воспалительные заболевания, такие как артрит, в особенности ревматоидный артрит, воспалительное кишечное заболевание, псориаз, псориатический артрит, псориатические бляшки, саркоидоз, артериосклероз и заболевания после трансплантации, отторжение почечного трансплантата, эндометриоз или хроническая астма и более 70 других состояний. Новые кровеносные сосуды могут снабжать кровью больные ткани, разрушать нормальные ткани и, в случае ракового заболевания, новые сосуды могут способствовать попаданию опухолевых клеток в кровоток и переносу их в другие органы (метастазы опухоли). Недостаточный ангиогенез наблюдается тогда, когда происходит неадекватный рост кровеносных сосудов, который способствует ухудшению болезненного состояния, например, при заболеваниях, таких как коронарный атеросклероз (коронарная артериальная болезнь), "удар" (внезапный мозговой удар) и замедленное заживление ран. Помимо этого язвы, "удары" и инфаркты могут быть результатом отсутствия ангиогенеза, который обычно необходим для естественного заживления. Настоящее изобретение рассматривает лечение пациентов, у которых наблюдаются вышеуказанные заболевания, или пациентов с повышенным риском развития вышеуказанных заболеваний, с применением антитела, которое специфически связывает как VEGF, так и HER2 (например, антитело bH1-81 или bH1-44).
Другими пациентами, кандидатами на получение композиций по изобретению, являются пациенты, у которых наблюдаются следующие заболевания или имеется повышенный риск развития следующих заболеваний: анормальная пролиферация сосудисто-волокнистой ткани, розовые угри, синдром приобретенного иммунодефицита, окклюзия артерий, атопический кератит, бактериальные язвы, болезнь Бехчета, опухоли у новорожденных, закупорка сонных артерий, хориоидальная неоваскуляризация, хроническое воспаление, хроническое отслоение сетчатки, хронический увеит, хронический витреит, переутомление глаз, вызванное контактными линзами, отторжение пересаженной роговицы, неоваскуляризация роговицы, неоваскуляризация пересаженной роговицы, болезнь Крона, перифлебит сетчатки, эпидемический кератоконъюнктивит, грибковые язвы, инфекции простого герпеса, инфекции опоясывающего лишая, синдромы гипервязкости крови, саркома Капоши, лейкоз, липидная дегенерация, болезнь Лайма, маргинальный кератолиз, разъедающая язва роговицы, микобактериальные инфекции, отличные от проказы, близорукость, неоваскуляризация отделов глаза, врожденные ямки в диске зрительного нерва, синдром Ослера-Вебера (Ослера-Вебера-Рандю), остеоартрит, болезнь Педжета, средний увеит (pars planitis), пемфигоид (пузырчатка), фликтенулезный конъюнктивит, полиартериит, осложнения после лечения лазером, протозойные инфекции, эластичная псевдоксантома, птеригий, сухой кератит, радиальная кератотомия, неоваскуляризация сетчатки (ретинальная), ретинопатия недоношенных, ретролентальная фиброплазия, саркоидоз, склерит, серповидно-клеточная анемия, болезнь Шегрена, солидные опухоли, болезнь Старгарта, болезнь Стивенса-Джонсона, "верхний лимбический кератит", сифилис, системная красная волчанка, краевая дегенерация Террьена (Terrien), токсоплазмоз, саркома Юинга, нейробластома, остеосаркома, ретинобластома, рабдомиосаркома, язвенный колит, окклюзия вен, дефицит витамина А, саркоидоз Вегенера, нежелательный ангиогенез, ассоциированный с диабетом, паразитарные заболевания, анормальное заживление ран, гипертрофия после хирургической операции, поражение или травма (например, острый респираторный дистресс-синдром/ARDS), ингибирование (угнетение, замедление) роста волос, ингибирование овуляции и образования желтого тела, ингибирование имплантации и ингибирование развития эмбриона в матке.
Антиангиогенезные терапевтические средства применимы для общего лечения отторжения трансплантата, воспаления легких, первичной легочной гипертензии, нефротического синдрома, преэклампсии и плеврального выпота, заболеваний и расстройств, характеризующихся нежелательной проницаемостью сосудистой стенки, например, отека, ассоциированного с опухолями головного мозга, асцитов, ассоциированных со злокачественными новообразованиями, синдрома Мейгса, воспаления легких, нефротического синдрома, выпота в полость перикарда (такого как ассоциированный с перикардитом), проницаемости, ассоциированной с сердечнососудистыми заболеваниями, такими как состояние после инфаркта миокарда и инсультов и т.п., и сепсиса.
Другие ангиогенез-зависимые заболевания по данному изобретению включают ангиофиброму (отличная от нормы кровь в сосудах, склонных к кровоизлиянию), неоваскулярную глаукому (рост кровеносных сосудов в глазу), артериовенозные мальформации (АВМ, AVM; патологические соединения между артериями и венами), несрастание костей при переломах (незаживающие переломы), атеросклеротические бляшки (уплотнение артерий), пиогенную гранулему (обычное кожное поражение, состоящее из кровеносных сосудов), склеродермию (форму соединительнотканного заболевания), гемангиому (опухоль из кровеносных сосудов), менингиому, гиперплазии щитовидной железы (включая болезнь Грейвса), трахому (основную причину слепоты в странах третьего мира), гемофилическую артропатию, синовит, дерматит, спайки сосудов и гипертрофические рубцы (образование аномальных рубцов).
IV. Дозы и препараты
Композиции антитела (например, bH1-44 или bH1-81) или фрагмента антитела готовят, дозируют и вводят в соответствии с обычной медицинской практикой. Факторы, принимаемые во внимание в этом контексте, включают конкретное расстройство, которое следует лечить, клиническое состояние отдельного субъекта, причину расстройства, область доставки агента, метод введения, схему введения и другие факторы, известные практикующим врачам. При определении "терапевтически эффективного количества" вводимого антитела или фрагмента антитела руководствуются этими соображениями, это "терапевтически эффективно количество" представляет собой минимальное количество, необходимое для предупреждения, ослабления или лечения ракового или аутоиммунного расстройства. Антитело или фрагмент антитела, не обязательно, готовят с одним или более агентами, используемыми в настоящее время для предупреждения или лечения ракового или аутоиммунного расстройства или риска развития ракового или аутоиммунного расстройства. Эффективное количество таких других агентов зависит от количества антитела или фрагмента антитела, присутствующего в препарате, типа расстройства или лечения и других обсуждавшихся выше факторов. Эти другие агенты обычно применяют в тех же дозах и вводят теми же путями, которые применялись ранее, или в количестве, примерно, от 1 до 99% от ранее применявшихся доз. Как правило, облегчение или лечение рака включает уменьшение ослабление одного или более симптомов или медицинских проблем, ассоциированных с раком. Терапевтически эффективное количество лекарства может выполнить одну или комбинацию из следующих задач: уменьшить (по меньшей мере на 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 100% или более) число раковых клеток; уменьшить или ингибировать размер опухоли или опухолевую массу; ингибировать (т.е. понизить (замедлить) до некоторой степени и/или прекращать) инфильтрацию раковыми клетками периферических органов; уменьшить гормональную секрецию в случае аденом; понизить плотность сосудов; ингибировать метастазирование опухоли; уменьшить или ингибировать рост опухоли; и/или до некоторой степени облегчить один или более симптомов, ассоциированных с раком. В некоторых вариантах изобретения антитело или фрагмент антитела используют для предупреждения появления или повторения ракового заболевания или аутоиммунного расстройства у субъекта.
В одном варианте настоящее изобретение можно применять для увеличения продолжительности выживания (выживаемость) пациента, человека, восприимчивого к раковому заболеванию или аутоиммунному расстройству, или пациента, у которого диагностировано раковое заболевание или аутоиммунное расстройство. Продолжительность выживания (выживаемость) определяется как время от первого введения лекарства до момента смерти. Продолжительность жизни (выживаемость) можно также измерять с помощью стратифицированных показателей относительного риска (HR) подопытной группы по сравнению с контрольной группой, которые отражают риск смерти пациента в процессе лечения.
Еще в одном варианте изобретения лечение по настоящему изобретению значительно повышает показатель эффективности (быстродействие) терапии в группе пациентов, восприимчивых к раковому заболеванию, или пациентов, у которых диагностировано раковое заболевание, которых лечат различными противораковыми терапевтическими средствами. Показатель эффективности определяют как процент пролеченных пациентов, которые реагируют (отвечают) на лечение. В одном аспекте изобретения комбинированное лечение по изобретению с применением антитела или фрагмента антитела и хирургической операции, лучевой терапии или одного или более химиотерапевтических агентов значительно повышает показатель отклика (показатель эффективности) в группе пролеченных пациентов по сравнению с группой пациентов, пролеченной с помощью только хирургической операции, лучевой терапии или химиотерапии, причем это повышение имеет Хи-квадрат р-величину менее 0.005.
Другие методы количественного определения терапевтической эффективности при лечении раковых заболеваний описаны в опубликованной патентной заявке США No.20050186208.
Терапевтические препараты готовят стандартными методами, известными в уровне техники, смешением активного ингредиента нужной степени чистоты с необязательными физиологически приемлемыми носителями, эксципиентами или стабилизаторами (Remington's Pharmaceutical Sciences (20th edition), ed. A. Gennaro, 2000, Lippincott, Williams & Wilkins, Philadelphia, PA). Приемлемые носители включают физиологический солевой раствор или буферы, такие как фосфатный, нитратный и буферные растворы, содержащие другие органические кислоты; антиоксиданты, включающие аскорбиновую кислоту; низкомолекулярные (содержащие менее 10 аминокислотных остатков) полипептиды; белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстрины; хелатирующие агенты, такие как EDTA; сахарные спирты, такие как маннит или сорбит; солеобразующие противоионы, такие как катион натрия; и/или неионные поверхностно-активные вещества, такие как Твин (TWEEN™), Плуроникс (PLURONICS™) или полиэтиленгликоль (ПЭГ, PEG).
Необязательно, но предпочтительно, чтобы препарат содержал фармацевтически приемлемую соль, предпочтительно, хлорид натрия, и предпочтительно, в концентрациях, близких физиологическим. Необязательно, препараты по изобретению могут содержать фармацевтически приемлемый консервант. В некоторых вариантах изобретения концентрация консерванта находится в интервале 0.1-2.0%, обычно об./об. Подходящие консерванты включают консерванты, известные в фармацевтике. Предпочтительными консервантами являются бензиловый спирт, фенол, м-крезол, метилпарабен и пропилпарабен. Необязательно, препараты по изобретению могут включать фармацевтически приемлемое поверхностно-активное вещество (ПАВ) в концентрации от 0.005 до 0.02%.
Если это необходимо по конкретным подлежащим лечению показаниям, препарат по данному описанию может также содержать более одного активного соединения, предпочтительно, с комплементарной активностью, причем эти активные соединения не оказывают отрицательного воздействия друг на друга. Соответственно, такие молекулы присутствуют в комбинации в количествах, эффективных для осуществления предполагаемой задачи.
Можно также приготовить препараты, в которых активные ингредиенты заключены в микрокапсулы, например, методами коацервации или межфазной полимеризации, например, микрокапсулы из гидроксиметилцеллюлозы или желатиновые микрокапсулы или микрокапсулы из полиметилметакрилата, соответственно, препараты в коллоидных системах доставки (например, в виде липосом, альбуминовых микросфер, микроэмульсий, наночастиц и нанокапсул) или в виде макроэмульсий. Такие методы раскрываются в руководстве Remington: The Science and Practice of Pharmacy см. выше.
Можно приготовить препараты с пролонгированным высвобождением. Подходящие примеры препаратов с пролонгированным высвобождением включают полупроницаемые матрицы из твердых гидрофобных полимеров, содержащие антитело, причем эти матрицы представляют собой оформленные изделия, например, пленки или микрокапсулы. Примеры матриц с пролонгированным высвобождением включают сложные полиэфиры, гидрогели (например, поли(2-гидроксиэтилметакрилат) или поливиниловый спирт, полилактиды (патент США No. 3773919), сополимеры L-глутаминовой кислоты и γ-этил-L-глутамата, неразлагающийся сополимер этилена с винилацетатом, разлагающиеся (расщепляемые) сополимеры молочной и гликолевой кислот, такие как LUPRON DEPOT™ (инъецируемые микросферы, состоящие из сополимера молочной кислоты с гликолевой кислотой и лейпролидацетата) и поли-D-(-)-3-гидроксимасляная кислота. В то время как полимеры, такие как сополимер этилена с винилацетатом и сополимер молочной кислоты с гликолевой кислотой способствуют высвобождению молекул в течение более чем 100 дней, некоторые гидрогели высвобождают белки за более короткий период времени. Инкапсулированные антитела остаются в организме продолжительное время, они могут денатурироваться или агрегировать под воздействием влаги при 37°С, что приводит к потере биологической активности и возможным изменениям иммуногенности. Можно разработать правильную стратегию стабилизации в зависимости от задействованного механизма. Например, если найдено, что механизм агрегации представляет собой образование внутримолекулярной S-S связи за счет взаимодействия тиогруппа - дисульфидная группа, стабилизацию можно осуществить модификацией сульфгидрильных остатков, лиофилизацией из кислых растворов, контролем содержания влаги, применением соответствующих добавок и разработкой специфических композиций полимерных матриц.
Антитела или фрагменты антител по данному описанию (например, bH1-44 или bH1-81 или их фрагменты) вводят человеку известными методами, такими как внутривенное введение в виде болюса или в виде непрерывного вливания (инфузии) в течение некоторого периода времени, введением внутримышечно, интраперитонеально, интрацереброспинально (в спинной мозг), подкожно, внутрисуставно, подоболочечно, перорально, топически или в виде ингаляций. Локальное введение может быть особенно желательным, если с VEGF и/или HER2 антагонизмом ассоциированы очень сильные побочные эффекты или токсичность. Методы ex vivo включают трансфекцию или трансдукцию клеток, полученных от субъекта, полинуклеотидом, кодирующим антитело или фрагмент антитела. Трансфецированные или трансдуцированные клетки затем возвращают в организм субъекта, клетки могут представлять собой любые клетки из широкого ряда типов, включая, но без ограничения, гемопоэтические клетки, клетки костного мозга, макрофаги, моноциты, дендритные клетки, Т клетки или В клетки), фибробласты, эпителиальные клетки, эндотелиальные клетки, кератиноциты или мышечные клетки.
В одном примере антитело (например, bH1-44 или bH1-81) или фрагмент антитела вводят человеку местно, например, в виде прямой инъекции, если расстройство или расположение опухоли это позволяют, и инъекции можно периодически повторять. Антитело или фрагмент антитела можно также доставлять системно субъекту или непосредственно к опухолевым клеткам, например к опухоли или к ложу опухоли после хирургического удаления опухоли, с целью предупреждения или уменьшения местного рецидива или метастазирования.
V. Готовые изделия и наборы
Другой вариант изобретения включает готовое изделие, содержащее материалы, применимые для лечения аутоиммунных и раковых заболеваний. Изделие содержит контейнер и этикетку или вкладыш в упаковку на контейнере или соединенный с контейнером. Подходящие контейнеры включают, например, флаконы, виалы, шприцы и т.д. Контейнер может быть изготовлен из различных материалов, таких как стекло или пластик. Контейнер содержит композицию, которая эффективна для лечения состояния, и может иметь стерильное входное отверстие (порт доступа) (например, контейнер может представлять собой пакет (мешок) с внутривенным раствором или виалу с пробкой, которую может проткнуть гиподермическая игла для инъекций). По меньшей мере один активный агент в композиции представляет собой полиспецифическое антитело или фрагмент антитела по изобретению. На этикетке или на вкладыше в упаковку указывается, что композиция применяется для лечения конкретного заболевания. Этикетка или вкладыш дополнительно содержат инструкции по введению композиции антитела пациенту. Также рассматриваются изделия и наборы, содержащие средства для комбинированной терапии.
Вкладыш в упаковку содержит инструкции, обычно включаемые в продажные упаковки терапевтических продуктов, которые содержат информацию о показаниях, применении, дозе, введении, противопоказаниях и/или предостережениях, относящихся к применению таких терапевтических продуктов. В других вариантах изобретения на вкладыше в упаковку указывается, что композиция применяется для лечения рака молочной железы, колоректального рака, рака легкого, почечноклеточной карциномы, глиомы или рака яичника.
Помимо этого изделие может дополнительно включать второй контейнер, содержащий фармацевтически приемлемый буфер, такой как бактериостатическая вода для инъекций (BWFI), фосфатно-солевой буферный раствор, раствор Рингера и раствор декстрозы. Помимо этого он может включать другие материалы, необходимые с точки зрения производителя и пользователя, включая буферы, разбавители, фильтры, иглы и шприцы.
Также предоставляют наборы, применимые для различных целей, например, для очистки или иммунопреципитации VEGF или HER2 от клеток. Для выделения и очистки VEGF или HER2 набор может содержать VEGF/HER2 антитело (например, bH1-44 или bH1-81), связанное с гранулами (бусами) (например, сефарозными гранулами (бусами)). Можно предоставить наборы, которые содержат антитела для детекции и количественного определения VEGF или HER2 in vitro, например, методом ИФА (ELISA) или Вестерн-блоттингом. Что касается изделия, набор содержит контейнер и этикетку или вкладыш в упаковку на контейнере или связанный с контейнером. Контейнер содержит композицию, включающую по меньшей мере одно полиспецифическое антитело или фрагмент антитела по изобретению. Могут включаться дополнительные контейнеры, которые содержат, например, разбавители и буферы или контрольные антитела. Этикетка или вкладыш в упаковку может содержать описание композиции, а также инструкции для предполагаемого in vitro или диагностического применения.
Предполагается, что вышеприведенного описания достаточно для того, что специалист в данной области техники смог применить изобретение на практике. Нижеприведенные примеры предлагаются только с целью иллюстрации и никоим образом не претендуют на ограничение объема настоящего изобретения. На самом деле различные модификации изобретения, помимо известных и представленных в данном описании, станут очевидны для специалистов в данной области техники из вышеприведенного описания, и они входят в объем прилагаемой Формулы изобретения.
ПРИМЕРЫ
Пример 1. Дизайн и конструирование библиотеки
Антигенсвязывающий сайт антитела получают ассоциацией вариабельного домена (VH, VL) тяжелой цепи (НС) и легкой цепи (LC), причем каждый из них содержит три CDR петли для распознавания антигена. Во многих случаях один из двух вариабельных доменов, часто VH, определяет антигенную специфичность. Мыши с трансгенным НС, но интактным LC репертуаром вырабатывают титры нейтрализующих антител (Senn et al., Eur. J. Immunol. 33: 950-961, 2003). Мы задались целью изучить, как может возникнуть биспецифичность антитела и действительно ли другое употребление VH и VL доменов может способствовать двойственной антигенсвязывающей специфичности.
Для поиска дизайна диверсификации (создания разнообразия) аминокислотного состава и длины CDR легкой цепи антитела и матрицы библиотеки, позволяющую получить фаг-дисплейную библиотеку функциональных антител, из которой можно выбрать антитела, специфически связывающиеся с белковым антигеном, выбирают полуэмпирический подход. Разнообразие последовательности и длины CDR областей, примерно, 1500 человеческих последовательностей легкой цепи каппа, представленных в базе данных Kabat, служит руководством к процессу создания. Гидрофильные остатки намечают для рандомизации. Создают субпопуляцию (подмножество) рандомизированных положений, чтобы представить аминокислоты, которые являются частью природного (естественного) репертуара в этих сайтах, тогда как остальные сайты рандомизируют таким образом, чтобы они включали все 20 природных аминокислот.
В частности, матрица легкой цепи (вариабельный домен), представленная ниже, модифицирована по данному описанию (подчеркнутые остатки рандомизированы (выбраны произвольно)) (SEQ ID NO:10). DIQMTQSPSSLSASVGDRVTITCRASQD 28 VNTAVAWYQQKPGKAPKLLIYS 50 ASFLY SGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQQH 91 YTTPPTFGQGTKVEIKRTVAAP SVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKD STYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
Создают четыре группы библиотек на основе 3 матриц человеческих Fab и scFv, в которых различные наборы положений были намечены для рандомизации (Фигура 1).
Во всех библиотеках тяжелая цепь остается постоянной, ее последовательность определяется матрицей библиотек. Последовательность матрицы тяжелой цепи (вариабельный домен) представлена ниже (SEQ ID NO:11). EVQLVESGGGLVQPGGSLRLSCAASGFNIKDTYIHWVRQAPGKGLEWVARIYPTNG YTRYADSVKGRFTISADTSKNTAYLQMNSLRAEDTAVYYCSRWGGDGFYAMDYW GQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTS GVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTH
Дизайн библиотеки представлен на Фигуре 1 и Фигуре 2. Все матрицы библиотек содержат стоп-кодон (Sidhu et al., 2004), заключенный в домен CDR L1, предупреждающий присутствие матричной легкой цепи у членов фаг-дисплейной библиотеки антител. Матричные CDR последовательности представлены на Фигуре 3.
В одном примере мы вводим мутации в LC вариабельном домене HER2-специфического антитела, чтобы идентифицировать варианты, которые могут связывать другой (отличный) белковый антиген, при этом сохраняют первоначальную специфичность связывания. Мы выбираем консервативный подход к рандомизации LC CDR доменов, чтобы получить варианты, которые могут стабильно экспрессироваться. Двенадцать гидрофильных (подверженных действию растворителя) LC CDR положений выбирают для рандомизации: пять в CDR1 (28, 29, 30, 31, 32), три CDR2 (50, 51, 53) и четыре в CDR3 (91, 92, 93, 94). Помимо этого, для того чтобы определять дизайн аминокислотного разнообразия в выбранных сайтах, изучают естественное разнообразие этих положений, анализируя около 1500 CDR последовательностей человеческой каппа LC (Johnson and Wu, Nucleic Acids Res. 28:214, 2000; Chothia and Lesk, J. Mol. Biol. 196: 901, 1987) (Фигура 4). Некоторые положения с относительно высоким естественным разнообразием (30, 31, 50, 92, 93) рандомизируют полностью, тогда как другие положения ограничивают не более чем двумя типами аминокислот для имитации природных (натуральных) антител. Изменение длины CDR1 и CDR3 природной LC также отражается в библиотеке (Фигура 4). На Фигуре 4 Х обозначает типы аминокислот, созданные с низкой частотой, как показано. Разнообразие длины конструируют, включая 1-5 остатков между остатками 30 и 31 и между остатками 93 и 94.
Библиотека LC представляет собой полезный репертуар интактных последовательностей (Таблица 1). Приводятся результаты скрининга 95 случайных клонов по окончании четырех циклов селекции. В частности, селекции на новую специфичность связывания осуществляют как описано для иммобилизованных мишеней (VEGF, DR5 и человеческий Fc) (Sidhu et al., J. Mol. Biol. 338:299, 2004). После четырех циклов селекции 95 фаговых клона анализируют методом ИФА (ELISA) на связывание с целевым, HER2, и с нецелевым белком, BSA, чтобы гарантировать специфическое связывание. Для обогащения связывающимися с мишенью клонами, которые сохраняют связывание с HER2, проводят последний цикл селекции на HER2. Позитивные клоны секвенируют. Для идентификации связующих с наивысшей аффинностью конкурентным анализом ИФА (ELISA) (Sidhu et al., J. Mol. Biol. 338: 299, 2004) определяют IC50 для связывания с антигеном. Показано число уникальных клонов, определенное секвенированием последовательностей, и число уникальных клонов, которые сохраняют HER2 связывание (биспецифические клоны). Для этих клонов наблюдается минимальные сопоставимые с фоном сигналы связывания с нерелевантными антигенами, такими как BSA.
Селекция против трех белковых антигенов: человеческого сосудистого эндотелиального фактора роста (hVEGF), рецептора смерти 5 (DR5) и комплементсвязывающего фрагмента IgG (Fc), дает много связывающих клонов (Фигура 5А). Некоторые клоны утрачивают аффинность связывания с HER2, тогда как другие сохраняют HER2-связывание и, следовательно, являются биспецифическими. Секвенирование 131 уникальных вариантов антитела Герцептин® с новой специфичностью связывания позволяет идентифицировать аминокислотные замены и инсерции по сравнению с антителом Герцептин® (Фигура 5В).
Диапазон числа мутаций составляет 3-17. Клоны, которые сохраняют HER2 связывание (биспецифические клоны), в среднем содержат меньше мутаций, чем клоны, утратившие HER2 связывание. Сохраняющая CDR-L3 последовательность антитела Герцептин® является предпочтительной, но недостаточной для того, чтобы сохранить HER2-связывание. Это согласуется с сообщением о том, что CDR-L3 последовательность антитела Герцептин® является наиболее важной LC CDR областью для HER2 связывания (Kelley and O'Connell, Biochemistry 32: 6828, 1993). Характерные VEGF-связывающие клоны экспрессируют в виде белков Fab и IgG (Таблица 2).
Для того чтобы продемонстрировать, что эти антитела специфически связываются со "своими" антигенами и не-специфически не взаимодействуют с другими белками, мы показали, что детектируемое связывание с панелью лизатов клеток млекопитающих и неантигенными белками. Анализ подтвердил моно- и биспецифичность очищенных IgG или Fab (Фигура 6).
Величины аффинности равновесного связывания (KD) полученных из LC библиотеки моноспецифических антител находится в пределах 15-150 нМ. Биспецифические антитела связываются с новыми антигенами (т.е. с VEGF) с аффинностью от высоких величин в нМ до низких величин в мкМ, а с HER2 с аффинностью, выражаемой низкими величинами в нМ (Таблица 2). Из антител, показанных в Таблице 2, антитело bH1 проявляет наивысшую биспецифическую аффинность к двум различным белковым антигенам VEGF (KD=300 нМ) и HER2 (KD=26 нМ).
Материалы
Ферменты и хелперный фаг М13-KO7 получают от New England Biolabs. Клетки Е.coli XL1-Blue получают от фирмы Stratagene. Альбумин бычьей сыворотки (BSA), овальбумин и Твин 20 получают от фирмы Sigma. Нейтравидин, казеин и SuperBlock™ получают от Pierce. Иммобилизованный белок G и конъюгированную с антителом против М13 пероксидазу хрена (HRP) получают от фирмы GE Healthcare (Piscataway, NJ). Иммунопланшеты MaxiSorp получены от фирмы NUNC (Roskilde, Denmark). Субстрат тетраметилбензидина (ТМВ) получают от Kirkegaard and Perry Laboratories (Gaithersburg, MD). Все белковые антигены получены исследовательской группой в компании Genentech, Inc. Варианты вырожденных ДНК представлены с применением IUB кода и, если не указано иначе, представляют собой эквимолярные смеси: N=A/C/G/T, D=A/G/T, V=A/C/G, B=C/G/T, H=A/C/T, K=G/T, M=A/C, R=A/G, S=G/C, W=A/T, Y=C/T.
Например, в некоторых произвольных (рандомизированных) положениях кодон дикого типа заменяют на вырожденный NNK кодон (N=A/T/G/C, K=G/T в эквимолярном соотношении), который кодирует все 20 природных аминокислот. Кодон XYZ относится к кодону с неравными соотношениями нуклеотидов в каждом положении кодонового триплета. Х содержит 38% G, 19% А, 26% Т и 17% С; Y содержит 31% G, 34% А, 17% Т и 18% С; и Z содержит 24% G и 76% С.
Фагмидные векторы для конструирования библиотек
Для конструирования векторов используют стандартные методы молекулярной биологии. Для получения библиотек используют три матрицы. Все матрицы являются производными плазмиды pV0354, используемой в библиотеках тяжелых цепей на основе модифицированных гуманизированных 4D5 (версия 8) (Lee et al., 2004a).
Фагмиду pJB0290 матрицы 2С4 Fab-C конструируют, клонируя вариабельный домен тяжелой цепи 2С4 в вектор pV0354-Fab-С, содержащий промотор щелочной фосфатазы (Lowman et al., 1991) и сигнал секреции stII как для легкой, так и для тяжелой цепи Fab. Его создают таким образом, чтобы он содержал единичный цистеиновый остаток на С-конце вариабельного домена 1 тяжелой цепи, чтобы осуществить дисплей 2С4 Fab с применением фага бивалентного М13, как описано ранее (Lee et al., 2004b). Домены CDR легкой цепи 2С4 вводят в вектор Fab-С с помощью сайт-направленного мутагенеза, используя метод Kunkel et al. (Kunkel et al., 1987). На С-конце легкой цепи добавляют эпитопную метку (gD тэг) (Lasky and Dowbenko, 1984), чтобы способствовать определению уровня дисплея, как описано ранее (Sidhu et al., 2004). Матрицу pV1283 для библиотеки Fab12-G создают, клонируя "высоковизуализируемый" вариабельный домен тяжелой цепи в pV0354-Fab-C, a вариабельный домен легкой цепи модифицируют таким образом, чтобы он содержал CDR-L3 фрагмента Fab-12 (гуманизированное А4.6.1, антитело против VEGF). VH с высокой степенью визуализации выбирают из библиотеки Fab таким образом, чтобы CDR остатки G6 Fab, рандомизированные аланин-сканирующим мутагенезом методом "shotgun" (выстрела из дробового ружья, дробовика) (Liang et al., 2006; Vajdos et al., 2002) с помощью CDR-L3, превращались в Fab-12 (Y91STVPW96; SEQ ID NO:24) пэннингом на иммобилизованном антителе против gD. Дизайн и конструкцию фагмиды pV1384, "бивалентно" выявляющей 4d5 (LC-R66G) scFv на поверхности М13 фаговых частиц, модифицируют при использовании матрицы pS2018, описанной ранее (Sidhu et al., 2004). Фрагмент scFv содержит эпитопную метку gD в линкерной области между легкой цепью и тяжелой цепью. LC каркасный остаток Arg66 мутирует в Gly66, который является преобладающим остатком в этом положении более чем у 95% природных каппа легких цепей. Мутация R66G лишь немного снижает аффинность связывания антитела Герцептин® с HER2 (менее, чем в 2 раза), как описано Kelley and Connell (Biochemistry 32: 6828, 1993). Последовательности CDR матриц библиотек приводятся на Фигуре 3.
Создание библиотек
Фаг-дисплейные библиотеки создают, используя олигонуклеотид-направленный мутагенез, описанный ранее (Sidhu et al., 2004). Векторы матриц для библиотек содержат стоп-кодон (ТАА), заключенный в CDR-L1, который подвергается репарации в процессе мутагенеза с применением вырожденных олигонуклеотидов, которые отжигаются с последовательностями, кодирующими CDR-L1, CDR-L3 (все библиотеки), CDR-L2 (L1/L2/L3-A, -В, -С, +L4-D) и каркасную последовательность 3 легкой цепи (L1/L4 и L1/L2/L3+L4-D). Реакции мутагенеза библиотеки осуществляют по методу Kunkel et al. (Kunkel et al., 1987). Дизайн доменов CDR легкой цепи для библиотек описан на Фигуре 1, на которой представлены вырожденные кодоны в каждом положении для различных библиотек. Три или четыре олигонуклеотида смешивают в определенных соотношениях для каждого CDR домена, чтобы кодировать заданную частоту типов аминокислот в каждом положении, намеченном для рандомизации (Фигура 4). Олигонуклеотиды смешивают в различных соотношениях для тонкой настройки разнообразия, отражающей частоту аминокислот в природных последовательностях легкой цепи каппа в выбранных положениях. Для CDR1 три олигонуклеотида, содержащих кодоны для положений 91-94: CAT NNK RST (SEQ ID NO:25), KMT XYZ XYZ RST (SEQ ID NO:26) или DGG XYZ XYZ RST (SEQ ID NO:27), смешивают в соотношении 1:3:1. XYZ представляет собой вариант NNK с равными пропорциями A/G/T/C для каждого сайта, чтобы уменьшить область охвата алифатических гидрофобных аминокислот (Lee et al., J. Mol. Biol. 340: 1073, 2004). Для CDR2 четыре олигонуклеотида, содержащие кодоны для положений 50-53: NNK GST TCC NNK (SEQ ID NO:28), TGG GST TCC NNK (SEQ ID NO:29), KGG GST TCC TMT (SEQ ID NO:30) или NNK GST TCC TMT (SEQ ID NO:31) смешивают в соотношениях 1:1:2:10. Для CDR3 каждое расстояние охватывается смесью трех олигонуклеотидов, содержащих кодоны для положения 28-33: G70A70C70 RTT NNK NNK TAC STA (SEQ ID NO:32), G70A70C70 RTT NNK NNK DGG STA (SEQ ID NO:33) или G70A70C70 RTT NNK NNK NMT STA (SEQ ID NO:34) в соотношении 1:1:2. G70A70C70 представляет собой "soft" кодон, который предоставляет 70% означенного олигонуклеотида и 10% каждого из трех других, кодируя ~50% Glu и ~50% других аминокислот.
Структурный анализ ряда характерных антител с каппа LC показывает, что CDR1 имеет широкий ряд конформаций, которые, по-видимому, являются результатом изменения длины петли (11-17 остатков между положениями 24 и 34). Поэтому в библиотеку были включены CDR-L1 разной длины (длина 11-16 остатков). Природный CDR-L3 также меняет длину (длина 7-10 остатков между положениями 89-96), что отражается в дизайне библиотеки (длина 8-10; Фигура 4).
На Фигуре 1 дается сравнение природного разнообразия легкой цепи и действительного дизайна библиотеки. Продукты мутагенеза собирают в одну реакционную смесь на библиотеку и подвергают электропорации в клетки Е.Coli SS320, дополненные хелперным фагом KO7 и выращивают в течение ночи при 30°С (Lee et al., J. Mol. Biol. 340:1073, 2004). В реакции электропорации используют ~1011 клеток и ~5-10 мкг ДНК. Библиотечный фаг очищают (Sidhu et al., J. Mol. Biol. 338: 299, 2004). Число трансформантов находится в диапазоне 109-1010. Уровень дисплея интактных Fab или scFv на поверхности фага определяют анализом связывания ИФА (ELISA), в котором 96 произвольно выбранных клонов из каждой библиотеки тестируют на способность связываться с антителом против gD. Уровень дисплея находится в интервале 5-25% (Фигура 2). 25% визуализируемого антитела сохраняет связывание с HER2. Около 150 визуализирующих клонов секвенируют с целью изучения разнообразия данной библиотеки по сравнению с запланированным разнообразием. Часть (~30%) членов библиотеки с визуализированной функциональностью сохраняют последовательность доменов CDR-L2 и/или CDR-L3 антитела Герцептин® вследствие неполного мутагенеза (стоп-кодон матрицы в CDR-1 гарантирует 100% мутацию этого CDR в экспрессированных scFv). Их исключают из секвенирования с целью анализа разнообразия данной библиотеки. В большинстве рандомизированных положений в разнообразии фаг-дисплейной библиотеки визуализирующих клонов не наблюдается значимого отклонения (p>0.05, отношение шансов (коэффициент несогласия)) от запланированного разнообразия. Исключения составляют положение 29 в CDR-L1, т.к. найдено, что Val представлен в этом положении несколько избыточно по сравнению с Ile (р=0.005), и положения 51 и 53 в CDR-L2, где Gly и Ser преобладают по сравнению с Ala и Tyr, соответственно (p<0.01).
Пример 2. Оценка характеристик библиотек
Сортинг и скрининг библиотек
Библиотека считается функциональной, если антитела, специфически связывающиеся с различными белковыми антигенами, можно выделять после 4-5 раундов (циклов) сортинга. Известно, что многие белки-мишени содействуют функциональной иммобилизации для пэннинга библиотек, а специфические антитела получают из проверенных, утвержденных фаг-дисплейных библиотек (Fellouse et al., 2005) (Lee et al., 2004a). Для оценки каждого набора библиотек мы выбираем подмножество этих мишеней для селекции (Фигура 2). Проводят начальный цикл (раунд) селекции библиотек на связывание с антителом против gD или белком L в качестве улавливаемой мишени для того, чтобы исключить клоны, в которых удален ген Fab/scFv, а затем 4-5 циклов селекции антитела. Или же библиотеки подвергают непосредственной селекции путем связывания с мишенью без преселекции с антителом против gD или белком L. Антиген (5 мкг/мл) иммобилизуют на NUNC 96-луночных планшетах MaxiSorp в течение ночи и блокируют в течение 1 часа альтернативными блокирующими агентами (Фигура 7). В первом цикле селекции фаговые растворы с концентрацией 1013 фагов/мл добавляют в покрытые антигеном иммунопланшеты. Концентрацию фага уменьшают в каждом цикле селекции. После инкубации фаговых растворов на иммунопланшетах для связывания с иммобилизованным антигеном планшеты многократно отмывают с помощью PBS и 0.5% Твин 20. Для повышения жесткости время инкубации снижают (4 часа в 1м цикле, 3 часа во 2м, 3 часа в 3м, 2 часа в 4м, 1.75 часа в 5м), а число отмывок увеличивают в каждом цикле селекции (Фигура 7). Связанный фаг элюируют 0.1 М HCl в течение 30 минут и элюент нейтрализуют 1.0 М Tris основанием. Регенерацию фага с покрытой антигеном лунки иммунопланшета рассчитывают и сравнивают с регенерацией с блокированной лунки, не покрытой антигеном, чтобы изучить обогащение фаговых клонов, визуализирующих фрагменты Fab или scFv, которые специфически связаны с целевым антигеном (Фигура 7). Элюированный фаг амплифицируют в клетках Е.coli и используют для дальнейших циклов селекции. Случайные клоны после цикла 4 и 5 отбирают для скрининга и анализируют методом фагового твердофазного ИФА (ELISA), в котором связывание с мишенью и антителом против gD сравнивают со связыванием с нерелевантным белком (BSA) для проверки неспецифического связывания. Клоны, которые связывают антитело против gD и мишень, но не неспецифический белок, полагают специфическими позитивными. Библиотеки L1/L3, L1/L4, L1/L2/L3-A, L1/L2/L3-В_1 and L1/L2/L3-В_2 не дают никаких специфически позитивных клонов, тогда как из библиотек L1/L2/L3-С и L1/L2/L3+L4-D можно выделить специфические антитела к целевым антигенам.
Например, случайные клоны из четвертого цикла анализируют методом фагового твердофазного ИФА (ELISA), в котором связывание индивидуально амплифицированных клонов с мишенью и HER2 сравнивают со связыванием нецелевого белка (BSA), чтобы проверить специфичность связывания. Для обогащения фаговыми клонами, которые сохраняют связывание с HER2, элюированный фаг из третьего и четвертого цикла VEGF или DR5 селекции амплифицируют и проводят с ним следующий цикл селекции на лунках с иммобилизованным HER2. Области VL и VH позитивных клонов амплифицируют с помощью ПЦР и секвенируют.
Процент "хитов" (попаданий) для hFC, hVEGF и hDR5-lf составляет 63, 77 и 85%, соответственно. Области VL позитивных клонов амплифицируют с помощью ПЦР и секвенируют как описано ранее (Sidhu et al., 2004). Секвенирование последовательности ДНК позитивных специфических связующих позволяет определить процентное содержание уникальных клонов 51% (hFC), 55% (hVEGF) и 6.1% (hDR5-lf). Последовательности уникальных hVEGF-связывающих клонов представлены на Фигуре 8.
Комбинированная селекция клонов, связывающих hVEGF, в планшете и в растворе
После четырех циклов сортинга наблюдается высокое разнообразие клонов, связывающих hVEGF. Чтобы идентифицировать клоны, связывающих hVEGF с высокой аффинностью, выбирают методе селекции в растворе после 4х циклов сортинга в планшете. 50 нМ биотинилированного hVEGF инкубируют с фагом, was размноженным из 4го цикла селекции на иммобилизованном антигене. Инкубируют 2 часа при комнатной температуре при встряхивании и hVEGF-связанный фаг улавливают на покрытых стрептавидином и блокированных иммунопланшетах с последующими многократными отмывками. Фаговые клоны элюируют, подвергают скринингу и секвенируют как описано ранее. Последовательности клонов, связывающих hVEGF из последней стадии селекции в растворе, представлены на Фигуре 9.
Выделение биспецифических клонов из библиотек L1/L2/L3-С и L1/L2/L3+L4-D
Матрица для библиотек L1/L2/L3-С и L1/L2/L3+L4-D представляет собой фрагмент scFv, модифицированный с применением hu4D5 антитела, которое связывает Her2 с высокой аффинностью. Картирование на функциональном паратопе hu4D5-5 антитела для Her2 связывания аланин-сканирующим мутагенезом CDR областей показывает, что остатки тяжелой цепи вносят большую часть свободной энергии связывания, тогда как вклад отдельных остатков меньше (Kelley and O'Connell, 1993). Анализ атомной структуры Fab антитела Герцептин® в комплексе с человеческим Her2-ECD показывает, что тяжелая цепь вносит наибольший вклад в структуру интерфейса (области контакта с антигеном) (Cho et al., Nature 421:756, 2003). Мы наблюдали, что некоторые члены функциональных библиотек легких цепей, созданных на основе матрицы антитела Герцептин®, сохраняют способность связываться с Her2. Чтобы выделить из функциональных библиотек L1/L2/L3-С и L1/L2/L3+L4-D биспицифические scFv фрагменты, способные связывать Her2, а также второй антиген, применяют две стратегии. В одном методе позитивные клоны из ранее описанной селекции по целевому антигену подвергают скринингу методом ИФА (ELISA) на клоны, которые сохраняют связывание Her2. Процентное содержание специфических позитивных клонов, способных связывать Her2, меняется в зависимости от специфичности ко второму антигену. Только 1 клон из 61 уникального hFc специфического позитивного клона все еще связывает Her2 (1.6%), 30 из 41 уникального hVEGF-связывающего клона все еще связывают Her2 (73%) и 2 из 5 уникальных hDR5-связующих продолжают связывать Her2 (40%). Помимо метода на основе селекции, применяют метод выделения биспецифических антител отбором Her2 связующих из пула антител, связывающих hVEGF и hDR5. Проводят дополнительный цикл селекции продукта, элюированного после 4 цикла сортинга по целевому антигену, инкубируя 2×1013 фагов/мл в иммунопланшетах MaxiSorp, покрытых Her2 (5 мкг/мл) и блокированных BSA в течение 1 часа. Планшеты отмывают 15 раз с помощью PBS, 0.5% Твин 20 и связанный фаг элюируют как указано выше. Случайные клоны отбирают и анализируют на связывание Her2, антитела против gD и мишени по сравнению с неспецифическим связывание с нерелевантным белком (BSA). Все тестированные 192 клона идентифицируют как специфические позитивные и секвенируют как описано ранее. Секвенирование позволяет выявить 94 уникальных последовательности. Таким образом, этот метод дает возможность получить 94 Her2/hVEGF биспецифических клона из 94 проверенных биспецифических клонов (100%) (Фигура 8). Последовательности всех выделенных уникальных hVEGF/Her2 биспецифических антител из обоих методов выделения приводятся на Фигурах 10А и 10В. Последовательности выделенных клонов, которые полностью утрачивают способность связываться с Her2, показаны на Фигуре 11. Из клонов с двойной специфичностью почти все сохраняют домен CDR-L3 антитела Герцептин®, т.е. весьма правдоподобно, что сохранение домена CDR-L3 важно для сохранения HER2 связывания. В случае hDR5 2 из 7 уникальных Her2-связывающих клонов являются биспецифическими (29%, 12 секвенированных клонов). Один из клонов с двойной специфичностью имеет некоторые гомологичные изменения в CDR-L3.
Высокопроизводительный метод получения характеристик клонов, связывающих hVEGF
Для скрининга на клоны с высокой аффинностью к hVEGF и для изучения профилей VEGFR1-блокады используют высокопроизводительный конкурентный ИФА (ELISA) метод "одного пятна" в 96-луночном формате (Sidhu et al., 2004). Коротко говоря, иммунопланшеты MaxiSorp покрывают 2 мкг/мл hVEGF109 в течение ночи при 4°С и блокируют с помощью 1% (вес./об.) BSA в течение 1 часа. Фагмидные клоны в клетках Е.coli XL1-Blue выращивают в 150 мкл бульона 2YT, дополненного карбенициллином и М13-KO7 хелперным фагом; культуры выращивают при встряхивании в течение ночи при 37°С в 96-луночном формате. Культуральные супернатанты, содержащие фаг, пятикратно разводят в PBST (PBS с 0.05% Твин 20 и 0.5% (вес./об.) BSA) с добавлением или без добавления 100 нМ hVEGF109 для скрининга на аффинность. Для скрининга на блокирование рецептора лунки с иммобилизованным hVEGF инкубируют в присутствии или в отсутствие VEGFR1 Домена 1-3 (D1-3) и VEGFR1 Домена 2 (D2), а затем добавляют супернатант, содержащий фаг, в пятикратном разведении (Liang et al., 2006; Wiesmann et al., 1997). Инкубируют 1 час при комнатной температуре (RT), смеси переносят на покрытые hVEGF109 планшеты и инкубируют в течение 10 минут. Планшеты отмывают РВТ (PBS с 0.05% Твин 20) и инкубируют в течение 30 минут с конъюгатом антитела против М13 с пероксидазой хрена в 5000-кратном разведении в PBST до концентрации 1 нМ. Планшеты отмывают, проявляют ТМВ субстратом, примерно, пять минут, гасят 1.0 М H3PO4 и читают на спектрофотометре при 450 нм. В анализе аффинности методом "одного пятна" в качестве показателя аффинности используют отношение оптической плотности в присутствии hVEGF109 в растворенной фазе к оптической плотности в отсутствие hVEGF109 в растворенной фазе. Низкая величина отношения показывает, что основная часть Fab-фага связывается с hVEGF109 в растворенной фазе на начальной стадии инкубации и, следовательно, фаг становится неспособным захватывать иммобилизованный hVEGF109. Результаты высокопроизводительного анализа аффинности для первого 41 уникального клона представлены на Фигуре 12. Аналогично, для анализа блокирования, низкая величина отношения показывает, что связывание клона с hVEGF109 блокируется взаимодействием hVEGF109 - VEGFR1, это говорит о том, что некоторые клоны имеют перекрывающийся сайт связывания (эпитоп) на VEGF с соответствующими фрагментами VEGF (Фигуры 13А и 13В) и эти клоны, по-видимому, являются визуализирующими блокирование антителами.
Высокопроизводительный метод получения характеристик биспецифических hVEGF/Her2 клонов
Тот же принцип, который описан в предыдущем разделе, применяют для того, чтобы выделить клоны с высокой аффинностью к hVEGF и Her2 для более подробного изучения (Фигура 14А). Для скрининга на клоны с высокой аффинностью к hVEGF и Her2 для изучения профилей VEGFR1-блокады используют высокопроизводительный конкурентный ИФА (ELISA) метод "одного пятна", иммобилизуя на иммунопланшетах MaxiSorp 2 мкг/мл hVEGF109 и Her2-ECD в течение ночи при 4°C с последующим блокированием с помощью 1% (вес./об.) BSA в течение 1 часа. Фаговые клоны, идентифицированные как биспецифические в предыдущем скрининге с помощью ИФА (ELISA) методом "одного пятна", выращивают, как описано ранее и инкубируют в присутствии и в отсутствие 20 нМ Her2-ECD и 50 нМ hVEGF. Инкубируют 1 час при комнатной температуре, растворы наносят на покрытые иммунопланшеты и сигналы связывания регистрируют и анализируют как описано в предыдущем разделе. Клоны с низким соотношением как для hVEGF, так и для Her2 отбирают для более детального изучения. hVEGF-специфические и hVEGF/Her2 биспецифические фаговые клоны, которые дают самые низкие отношения сигналов в конкурентном анализе ИФА (ELISA) методом одного пятна, отбирают для количественного определения аффинности конкурентным ELISA, так же как DR5-связывающие и DR5/Her2 биспецифические фаговые клоны из начального скрининга ELISA методом одного пятна и VEGF-связывающие клоны из комбинированной селекции на планшете и в растворе. Фаговые клоны размножают из единичной колонии, выращивая в 25 мл 2YT культуры, дополненной карбенициллином и KO7 хелперным фагом, в течение ночи при 30°С. Сначала готовят серийные разведения фага, очищенного преципитацией в ПЭГ/NaCl, в PBST, а затем проверяют связывание с антигеном, иммобилизованным на планшете. Разведение, которое дает 50-70% насыщающий сигнал, используют в растворе для анализа связывания, в котором сначала инкубируется фаг с увеличивающимися концентрациями антигена в течение одного-двух часов, а затем переносят на покрытые антигеном планшеты на 10-15 минут для улавливания несвязанного фага. Величину IC50 рассчитывают как концентрацию антигена на стадии связывания в растворе, в котором ингибируется связывание фага с иммобилизованным антигеном на 50% (Lee et al., 2004a). На Фигуре 14В показаны кривые, по которым рассчитывают величину IC50 для анализируемых hVEGF-связывающих клонов из метода сортинга в планшетах. Величины IC50 находятся в интервале от 22 нМ до >1 мкМ (Фигура 14 В). Величины IC50 для hVEGF-связующих, выделенных комбинированной селекцией на планшетах и в растворе, находятся в интервале от 41 нМ до 226 нМ (Фигура 9). Величины IC50 DR5-связывающих клонов находятся в интервале от 20 нМ до >1 мкМ. Величины IC50 для hVEGF/Her2 биспецифических клонов представлены на Фигуре 15.
Пример 3. Получение характеристик антител из библиотеки легкой цепи
Превращение scFvs в Fabs
Чтобы проверить, действительно ли превращение фрагментов scFvs'2, выявляемых на фаге, во фрагменты Fabs влияет на аффинность связывающих клонов из библиотеки, 2 клона (3-7 анти-hVEGF и 4-1 анти-hDR5) выбирают для превращения в Fab и визуализируют на фаге. Область VL фагмидной ДНК для выбранных hVEGF и DR5 scFv фрагментов раразрезают рестриктазами, которые расщепляют ДНК выше (5', upstream) области, кодирующей CDR-LI (EcoRV) и ниже (3', downstream) области, кодирующей CDR-L3 (KpnI). Отщепленный фрагмент ДНК лигируют в аналогично расщепленный (разрезанный) вектор (рАР2009), созданный для фагового дисплея фрагмента Fab hu4D5 слиянием с С-концевым доменом гена-3 для М13 минорного оболочечного белка (Lee et al., 2004b). Полученная бис-цистронная фагмида содержит легкую цепь, слитую с эпитопной (gD) меткой на С-конце, и тяжелую цепь (VH и CH1), слитую с геном для М13 минорного оболочечного белка (p3) по С-концу под контролем промотора щелочной фосфатазы. Первая открытая рамка считывания кодирует полипептид, состоящий из сигнала секреции stil и следующей за ним легкой цепью Fab4D5, где области CDR заменены на CDR 3-7 анти-hVEGF и 4-1 анти-hDR5 scFv'2, с последующей эпитопной gD-меткой. Вторая открытая рамка считывания кодирует слитый полипептид, состоящий из следующих последовательностей: сигнала секреции stil, Fab4D5 тяжелой цепи и амбер (TAG) стоп-кодона, Gly/Ser линкерной последовательности и С-концевого домена g3 белка (сР3). Экспрессия в клетках Е.coli XL-1 Blue, коинфицированных М13-KO7, приводит к получению М13 бактериофага, визуализирующего Fab варианты 3-7 и 4-1 scFv'2. Конкурентный фаговый ИФА (ELISA) используют для оценки аффинности фаг-дисплейных scFvs и Fabs к hVEGF и к hDR5 в виде IC50 значений. Результаты из двух различных форматов хорошо согласуются между собой (данные не показаны).
Для дисплея bH1 Fab на поверхности М13 бактериофага плазмиду рАР2009 модифицируют таким образом, чтобы она кодировала bH1 Fab. Варианты bH1 Fab используют в качестве матриц для библиотеки, содержащих стоп-кодоны (ТАА) либо в трех доменах LC CDR доменах, либо в трех НС CDR доменах для LC и НС библиотеки, соответственно. Отдельные аланин- и гомолог-сканирующие библиотеки тяжелых цепей и легких цепей конструируют, как описано ранее (Vajdos et al., J. Mol. Biol. 320: 415, 2002). Значение вырожденности находится в интервале от 1×105 до 1×108, а размер данных библиотек составляет от 6×109 до 4×1010. Библиотеки создают как описано выше. От двух до трех циклов селекции проводят на иммобилизованных мишенях (VEGF, HER2-ECD, белок L или анти-gD mIgG) (Vajdos et al., J. Mol. Biol. 320: 415, 2002), Мишень-связывающие клоны подвергают скринингу фаговым иммуноферментным анализом ELISA на связывание с мишенью с последующим секвенированием ДНК и выравниванием последовательностей для расчета отношений дикого типа/мутация в каждом положении. Отношения из секвенирования, примерно, 100 уникальных последовательностей VEGF-и HER2-связывающих клонов корректируют на визуализацию и эффект скручивания белка делением с отношениями, рассчитанными для последовательностей более чем 100 анти-gD связывающих клонов, получают значения Fwt/mut. Так как только тяжелая цепь Fab сливается с оболочкой фага, фаговый дисплей gD метки, которая слита с легкой цепью, является показателем корректного фолдинга и корректного соединения легкой цепи и тяжелой цепи. Соответственно, связывание белка L с нелинейным эпитопом на легкой цепи Fab также в результате дает отношения дикого типа/мутация, аналогичные отношениям при селекции по gD метке. Величины Fwt/mut применяют для расчета ΔΔG по формуле ΔΔG=RTln(Ka,wt/Ka,mut)=RTln(Fwt/mut), как описано Vajdos et al. (J. Mol. Biol. 320: 415, 2002).
Экспрессия связующих из библиотеки в виде свободного человеческого Fab HigG
Для точного определения аффинности, специфичности и других свойств антител репрезентативные клоны из группы с каждой специфичностью, проявляющей наивысшую аффинность в конкурентном анализе ELISA, отбирают для экспрессии в виде свободного Fab и hIgG (Фигура 16). Вариабельный домен легкой цепи и тяжелой цепи клонируют в вектор, созданный ранее для экспрессии Fab в Е.coli или транзиторной экспрессии человеческого IgG в клетках млекопитающих (Lee et al., 2004а). Белок Fab получают, выращивая трансформированные клетки 34В8 Е.coli в полной C.R.A.P. среде при 30°С в течение 26 часов как описано ранее (Presta et al., 1997). hIgG экспрессируют транзиторной трансфекцией клеток 293 и hIgG очищают аффинной хроматографией на белке A (Fuh et al., J. Biol. Chem. 273:11197, 1998). 1 л культур E.coli очищают аффинной хроматографией на белке G. Колонки промывают PBS и Fab белок элюируют 100 мМ раствором уксусной кислоты и подвергают диализу против PBS. 4 л культур Е.coli очищают на колонке для аффинной хроматографии с белком А с последующей катионообменной хроматографией, описанной ранее (Muller et al., 1998). Концентрацию белка определяют методом спектрофотометрии. Конечный выход очищенного Fab, выращенного в малом масштабе в культуре во встряхиваемой на качалке колбе, обычно составляет 0.8-15 мг/л. Выход IgG в культуре в мелком масштабе составляет от 6.7 до 60 мг/л, т.е. от среднего до высокого (Фигура 17). Очищенные белки сначала характеризуют методом высокоэффективной гель-хроматографии и методом рассеяния света, чтобы гарантировать, что отсутствует заметная агрегация белка (<5%).
Коротко говоря, экспрессируемые белки Fab и hIgG анализируют методом ELISA на связывание с их соответствующим(-и) антигеном(-ами). Найдено, что все варианты, за исключением одного, связываются со "своим(-и)" антигеном(-ами). Клон 4-6 утрачивает способность связывать hDR5 при превращении в Fab и hIgG. Выбранные клоны антител против VEGF, специфические к укороченной форме hVEGF109, проверяют на связывание с hVEGF165 стандартным методом ELISA (Н3, H4_N, H4_D hIgG) и конкурентным ELISA (bH1, 3-1, 3-6, 3-7 hIgG). G6 hIgG (Fuh et al., 2006) используют в качестве позитивного контроля (Фигуры 18А и 18В). Как и предполагалось, все клоны связывают hVEGF165.
Для изучения степени агрегации белка выбранные клоны анализируют высокоэффективной гель-(проникающей) хроматографией (ГПХ, SEC) с последующим анализом методом рассеяния света (LS) в виде очищенных Fab и IgG. Образцы анализируют в PBS в концентрации 0.5 мг/мл (hIgG) и 1 мг/мл (Fab). Во всех образцах при данной концентрации максимальная наблюдаемая агрегация составляет 5% (Фигура 17), что находится в пределах значений, наблюдавшихся нами ранее для других антител, полученных методом фагового дисплея. Клоны 3-6 и 3-7 не выходят в ожидаемой временной точке, это наводит на мысль, что для "переформатированных" IgG и Fab наблюдается агрегация и/или неспецифическое взаимодействие со смолой (данные не показаны). Эти клоны исключают из набора клонов, которые анализируют далее.
Для того чтобы исключить перекрестную реактивность т неспецифическое связывание, мы изучали связывание выбранного hIgG при высоких концентрациях (100 нМ) с панелью иммобилизованных белков-мишеней, включая все клеточные лизаты, "свои" антигены и гомологи, стандартным анализом ELISA. Помимо антигена, мы иммобилизовали мышиный вариант hVEGF для проверки перекрестно-видовой реактивности клонов антител против hVEGF. В частности, панель белков иммобилизуют на планшетах MaxiSorp и блокируют с помощью 1% BSA в PBS в течение 1 часа. hIgGs (или Fabs) разводят в PBST до концентрации 100 или 500 нМ и переносят на планшеты с иммобилизованными белками. Инкубируют 1 час, планшеты отмывают и инкубируют с конъюгатом HRP - белок А. Сигналы связывания "проявляют" (визуализируют), добавляя ТМВ субстрат, примерно, в течение 5 минут, "тушат" реакцию с помощью 1М H3PO4 и читают на спектрофотометре при А450. Тестированные hIgGs специфически связываются со своим(-и) антигеном(-ами). Наблюдается перекрестная реактивность клонов bH1 и 3-1 по отношению к мышиному VEGF (mVEGF) (Фигура 19).
Чтобы проверить, действительно ли биспецифические антитела bH1, Н3 (анти-hVEGF/Her2) и D1 (анти-hDR5/Her2) могут синхронно (одновременно) связывать свои антигены (в когнатном взаимодействии) или они конкурируют за связывание с антителом, hVEGF и hDR5 иммобилизуют в концентрации 2 мкг/мл. Образцы hIgG с фиксированной концентрацией инкубируют с серийными разведениями Her2-ECD с последующим захватом hIgG на иммобилизованном антигене. Обнаружено, что в каждом случае Her2-ECD связывание является связыванием, конкурентным связыванию с другими антигенами (Фигура 20).
Для точного определения аффинности антител IgG и Fabcs (т.е выделенных из библиотек антител Fab и IgG против hVEGF и против hVEGF/Her2) и для изучения профилей связывания в реальном времени, мы используем метод поверхностного плазмонного резонанса (SPR) на приборе BIAcore™-3000 (BIAcore, Uppsala, Sweden) с иммобилизованными hVEGF, mVEGF, DR5 и Her2-ECD CM5 сенсорные чипы в единицах ответа (относительных единицах (RU)) 40-300 в зависимости от исследуемого аналита. Иммобилизацию проводят как описано ранее (Chen et al., 1999). Для минимизации эффектов авидности аналитов бивалентных IgG в этих случаях нацеливаются на лиганд с более низкой плотностью на сенсорном чипе. Образцы с повышающейся концентрацией в интервале величин, примерно, от в 10 раз более низкой до в 10 раз более высокой, чем расчетная величина KD (на основе экспериментов с конкурентным анализом ELISA), вводят (инжекция) со скоростью 22-30 мкл/минута, и отклики на связывание корректируют, вычитая RU из величины для эталонной проточной ячейки. Помимо этого отклики дважды проверяют по эталону, чтобы нормализовать по дрейфу нуля, вычитая RU из показаний для конъюгированной с лигандом проточной ячейки, в которую введен буфер для образца (PBS с 0.05% Твин 20). В кинетическом анализе фрагментов Fab для расчета kon и koff используют ленгмюровскую модель связывания 1:1. Где это необходимо, (при высоких концентрациях аналита), применяют 1:1 ленгмюровскую модель связывания с ограничением массопереноса. Для IgG аналитов используют модель бивалентного связывания аналита с ограничением или без ограничения массопереноса (BIAcore Evaluation Software 3.2). В случае Н3 hIgG, H4_N Fab и H4_D hIgG соответствие откликов кинетическим моделям связывания является неудовлетворительным. Поэтому анализ равновесного связывания применяют в тех случаях, когда существует зависимость (можно построить кривую зависимости) равновесного отклика от концентрации аналита. Величину КD определяют как ЕС50. Результаты анализа связывания на приборе BIAcore можно найти на Фигуре 21. Найдено, что аффинность hVEGF-связывающих антител 3-1, 3-6 и 3-7 выражается в наномолярных концентрациях. Анализируемые биспецифические антитела (bH1, Н3, H4_N, H4_D) проявляют аффинность к hVEGF от микромолярной до макромолярной. Напротив, аффинность к Her2 находится в диапазоне 8-59 нМ (Fab).
Чтобы определить, может ли легкая цепь антител (связующих) против hVEGF bH1, Н3 и H4_N связывать hVEGF независимо от последовательности ассоциированной тяжелой цепи, вариабельные домены легкой цепи "прививают" на Her2 2С4 Fab, клонируя вариабельные домены легкой цепи в вектор, экспрессирующий 2С4 Fab, pJB0524, тем самым заменяя вариабельный домен легкой цепи 2С4. Фрагменты Fabs экспрессируют как описано ранее. Уровни экспрессии химерных Fab bH1/2С4 и Н3/2С4 не являются заметными. Химерный белок Fab H4_N/2C4 выделяют и тестируют на связывание с hVEGF (начальная специфичность исходного bH1) и Her2 (начальная специфичность исходных bH1, 2С4). Методом стандартного твердофазного ИФА (ELISA) не обнаружено никакого связывания с hVEGF и Her2 (Фигура 22). Результаты показывают, что для связывания антигена необходима тяжелая цепь bH1.
Сравнение анти-hVEGF эпитопов (антител)
Чтобы приблизительно, грубо картировать эпитопы антител против hVEGF на hVEGF, мы изучаем способность этих только что выделенных антител против VEGF конкурировать с другими hVEGF-связывающими антителами и VEGF-рецепторами с известными сайтами связывания (Fuh et al., 2006; Muller et al., 1998; Wiesmann et al., 1997). Анализы проводят в формате конкурентного твердофазного ИФА (ELISA), в котором слитый белок VEGFR1 (Fit) Домена 1-3 и антител против hVEGF Авастина (Avastin®) (IgG), B20-4.1 (IgG), G6 (Fab) и KDR Домена 1-7 Fc иммобилизуют на иммунопланшетах MaxiSorp с концентрацией 2 мкг/мл. Для конкурентного связывания в растворе используют биотинилированный VEGF, уравновешенный с серийными разведениями очищенных IgG белков, и несвязанный биотин-VEGF улавливают на планшетах MaxiSorp с иммобилизованным Fab или IgG и детектируют с помощью конъюгированной со стрептавидином HRP (Lee et al., J. Mol. Biol. 340: 1073, 2004). Антитела, которые блокируют связывание hVEGF с другими hVEGF-связывающими антителами или hVEGF-рецепторами, по-видимому, имеют общие перекрывающиеся эпитопы. Применение высоких концентраций (мкМ) биспецифического связывающего hVEGF/Her2 антитела, bH1, делает возможным полное блокирование связывания hVEGF с его рецепторами, VEGFR1 и VEGFR2, это наводит на мысль, что эпитоп bH1 в достаточной степени перекрывается с VEGFR1 (Фигура 23) и VEGFR2 (Фигура 23). Помимо этого, bH1 блокирует связывание hVEGF с В20-4.1 (Фигура 24). H3, H4_N и H4_D также блокируют связывание hVEGF с обоими рецепторами, это показывает, что эпитопы сходны с эпитопом bH1 (Фигура 23). Профили неполного блокирования, по-видимому, объясняются их относительно низкой аффинностью к hVEGF (Фигура 21). Напротив, 3-1 не блокирует связывание hVEGF с VEGFR1 даже в самой высокой концентрации (0.5 мкМ) (Фигура 23). Помимо этого, мы не смогли детектировать блокирование антитела Авастин® с помощью 3-1 hIgG (Фигура 25). Однако, 3-1 hIgG блокирует связывание hVEGF с VEGFR2 (KDR) (Фигура 23), а также с В20-4.1 (Фигура 24). Эти результаты показывают, что 3-1 имеет уникальный эпитоп по сравнению с эпитопами других антител.
Пример 4. Изучение завимости структура-функция bH1, антитела против hVEGF/Her2
Для того чтобы выяснить природу взаимодействия антитела bH1 с двумя его антигенами, VEGF и HER2, проводят структурные и функциональные исследования (изучение структуры и функции). Антитело Герцептин® и bH1 имеют различия в областях CDR-L1 (V29NTA32 vs. I29PRSISGY32; SEQ ID NO:35 и 36) и CDR-L2 (S50ASF53 vs. W50GSY53; SEQ ID NO:37 и 38). bH1 против VEGF/Her2 выбирают как репрезентативное для характеристики структуры на основании его двойной специфичности и относительно высокой аффинности к VEGF и Her2. С целью изучения функциональных и структурных эпитопов на VEGF и Her2 нами получены кристаллы bH1 Fab в комплексе VEGF109 и внеклеточным доменом антигена hHer2 и проведено решение структуры двух комплексов методом РСА. Помимо этого, мы провели аланин- и гомолог-сканирование мутагенезом методом "shotgun" (выстрела из дробового ружья), используя комбинаторные фаг-дисплейные библиотеки, описанные ранее (Vajdos et al., 2002).
Экспрессия, очистка, кристаллизация и сбор данных
Участок связывания человеческого VEGF с рецептором, состоящий из 8-109, экспрессируют, проводят рефолдинг и очищают, как описано ранее (Christinger et al., 1996).
Остатки 1-624 внеклеточного домена антигена Her2 экспрессируют и очищают, как описано ранее (Franklin et al., 2004; Hudziak and Ullrich, 1991).
Для получения больших количеств bH1 Fab осадок (пеллеты) цельных клеток получают в результате ферментации клеток Е. coli в масштабе 10 л. 220 грамм клеточной пасты размораживают в 1 л PBS, 25 мМ EDTA, 1 мМ PMSF. Смесь гомогенизируют, а затем дважды пропускают через микрофлюидизатор. Затем суспензию центрифугируют при 12k в аликвотах по 250 мл в течение 90 минут. Затем белок наносят на колонку с Белком G (25 мл), уравновешенную PBS при скорости потока 5 мл/минута. Колонку отмывают буфером для уравновешивания, а затем элюируют 0.58% уксусной кислотой. Фракции анализируют методом SDS-PAGE (данные не показаны). Фракции, содержащие bH1 Fab, объединяют, а затем наносят на колонку (50 мл) для катионообменной хроматографии с SP-сефарозой (Pharmacia), уравновешенной 20 мМ MES рН 5.5. Fab элюируют в градиенте хлорида натрия в буфере для уравновешивания. Градиент является линейным относительно 0.5 М NaCl, 20mM MES рН 5.5. Фракции, содержащие Fab, идентифицируют методом SDS-PAGE (данные не показаны) и объединяют. bH1 Fab элюируют при концентрации NaCI около 0.5 М. Концентрацию Fab определяют, измеряя А280. Конечный выход bH1 Fab составляет 67 мг/л при выращивании в ферментере.
Комплексы получают, смешивая очищенный bH1 Fab и VEGF или Her2 ECDв молярном соотношении 2:1, и очищают гель-проникающей хроматографией (SP-200, Pharmacia) в смеси 25 мМ Tris-HCl, рН 7.5 и 0.3 М хлорида натрия для комплекса VEGF-Fab и смесью 25 мМ Tris-HCl, рН 8 и 0.15 М хлорида натрия для комплекса Her2 ECD-Fab. Состав полученных комплексов проверяют методом SDS-PAGE (данные не показаны). Белковый комплекс концентрируют и используют для кристаллизации. Начальные эксперименты с висящей каплей методом диффузии паров при 19°С дают мелкие изоморфные кристаллы в 14 различных состояниях в течение 1 недели в случае комплекса bH1-VEGF. Кристаллы комплекса bH1-Her2 появляются в 4 состояниях в течение недели. В каждом случае для дальнейшей оптимизации отбирают кристаллы в одном состоянии (одного вида).
Для кристаллизации bH1 Fab-VEGF (8-109) равные объемы раствора белкового комплекса (10.6 мг/мл белка, 300 мМ NaCl, 25 мМ Tris-HCl рН 7.5) и буфера для кристаллизации, содержащего 0.15 М D-, L-яблочной кислоты рН 7.0, 20% ПЭГ3350 смешивают и уравновешивают при 19°С. Через 24 часа появляются большие кристаллы, которые относятся к пространственной группе C2221, с размерами ячейки а=100.6, b=198.0, c=77.7. Кристаллические формы содержат в асимметричной ячейке 1 Fab и 1 VEGF мономер. До начала сбора данных проводят криопротекцию кристаллов, осуществляя перенос между каплями, содержащими 5%, 10% и 15% глицерина в искусственном маточном растворе, с последующим быстрым замораживанием в жидком азоте. Данные собирают до 2.6 Å в пучке синхротронного излучения 5.0.1 сверхмощного источника рентгеновского излучения Advanced Light Source (Berkeley).
Кристаллы bH1 Fab-Her2(l-624) получают, смешивая раствор белка (11 мг/мл, 25 мМ Tris рН 8 и 150 мМ хлорида натрия) с буфером для кристаллизации, содержащим 25% вес/об ПЭГ2000, 0.1М MES рН 6.5. Через 12 часов появляются кристаллы, относящиеся к пространственной группе P212121, с размерами ячейки a=62.3, b=115.1, с=208.2. Кристаллы содержат комплекс Her2-Fab в асимметричной ячейке. Перед сбором данных кристаллы быстро замораживают в жидком азоте с 20% этиленгликоля в качестве криопротектора. Данные собирают до 2.9 Å в пучке синхротронного излучения 5.0.1 сверхмощного источника рентгеновского излучения Advanced Light Source (Berkeley).
Обработка данных, определение и уточнение структуры
Данные обрабатывают, используя пакет программ Denzo and Scalepack (Otwinowski, 1997). Структуры комплексов bH1 Fab решены с помощью программы Phaser (L. С. Storoni, 2004; Read, 2001). Комплекс bH1-Fab-VEGF(8-109) решают, используя координаты VEGF из ранее описанного комплекса VEGF-Fab (2FJG) и Fab фрагментов, содержащих либо вариабельные домены VL/VH, либо константные домены CH1/CL антитела Герцептин® комплекса Fab-Her2 (1N8Z). Фрагменты Her2 и вариабельный домен Fab антитела Герцептин® из комплекса Her2-Fab, 1N8Z, используют в качестве моделей для поиска при решении структуры bH1-Her2. Константный домен bH1 Fab нельзя найти, используя константный участок Fab антитела Герцептин® в качестве поисковой модели (1N8Z) и молекулярный докинг необходимо проводить вручную, руководствуясь структурой комплекса антитела Герцептин® Fab-Her2. Построение и уточнение модели осуществляют, используя программы Refmac (Collaborative Computational Project, 1994) и Coot (Emsley and Cowtan, 2004), соответственно. Стереохимические параметры анализируют, используя MolProbity (Lovell et al., Proteins 50:437 (2003)). Структуры уточняют по Rvalue=0.22 и Rfree=0.27 для комплекса Fab-VEGF и Rvalue=0.25 и Rfree=0.31 для комплекса Fab-Her2. Создана модель кристаллической структуры bH1 Fab в комплексе с VEGF, а также Her2-ECD. Расстояние между некоторыми остатками bH1 Fab и антигенами находится в пределах 4.5, 4.0 и 3.5 Å. Два паратопа (область антитела, осуществляющая контакт с антигеном) для двух антигенов на одном и том же антителе заметно перекрываются, а остатки как легкой, так и тяжелой цепи участвуют в связывании с обоими антигенами. bH1 связывает на VEGF эпитоп, аналогичный эпитопу, который связывает на нем антитело Авастин®, а bH1 и антитело Герцептин® связывают на Her2 практически идентичный эпитоп.
Кристаллические структуры bH1 Fab, связанного с внеклеточным доменом (ECD) антигена HER2 (остатки 1-624) и с доменом, связывающим рецептор VEGF (остатки 8-109), определяют с разрешением 2.9 Å и 2.6 Å, соответственно (Фигура 26 и Таблица 3). На Фигуре 26 показана структура кристалла bH1 Fab/HER2, наложенная на комплекс антитело Герцептин®/НЕР2, и структура кристалла комплекса bH1 Fab/VEGF.
В комплексе bH1/HER2 Fab связывается с доменом IV антигена HER2 аналогично антителу Герцептин® (Cho et al., Nature 421: 756, 2003); два комплекса накладываются друг на друга со средним квадратичным отклонением (r.m.s.d.) Ca положений, равным 2.3 Å. В комплексе VEGF bH1 узнает эпитоп, который перекрывается с сайтами связывания VEGF рецепторов VEGFR1 и VEGFR2 и других антител к VEGF (Wiesmann et al., Cell 91: 695, 1997; Muller et al., Proc. Natl. Acad. Sci. USA 94: 7192, 1997). Соответственно, наблюдается блокирование bH1 связывания VEGF со своими рецепторами (Фигура 27). Для данных, показанных на Фигуре 27, биотинилированный человеческий VEGF165 уравновешивают с повышающимися концентрациями IgG (ось x). Несвязанный hVEGF165 улавливают на иммобилизованном слиянии VEGFR2-ECD Fc и детектируют спектрофотометрически (оптическая плотность при 450 нм, ось y).
Как показано на Фигуре 28, сайты связывания для VEGF и HER2 на bH1 в значительной степени перекрываются. Двенадцать из четырнадцати остатков, которые контактируют с HER2, также контактируют с VEGF. Оба сайта связывания включают остатки CDR как из НС, так и из LC. В комплексе HER2 области LC и НС CDR наблюдаются примерно равные области контакта с антигеном (53% и 47%, соответственно), тогда как в комплексе VEGF LC CDR домены составляют почти 70% с(о)крытой ("углубленной") поверхности (Фигура 29). Сайты связывания HER2 на молекуле антитела Герцептин® и на молекуле bH1 аналогичны и различаются только в областях CDR-L1 и -L2, где последовательность антитела Герцептин® не сохраняется в bH1 (Фигура 28). На Фигуре 28 остатки на поверхности Fab bH1 или антитела Герцептин® заштрихованы в соответствии со степенью сокрытия антигеном VEGF или HER2 (темная штриховка и белый шрифт >75% с(о)крыто, штриховка средней интенсивности и белый шрифт 50-75% сокрыто, легкая штриховка и черный шрифт 25-49% сокрыто). Подчеркнуты остатки, различные в bH1 и антителе Герцептин®. Белым пунктиром изображена граница между легкой и тяжелой цепью.
Конформация bH1 Fab в комплексе с HER2 явно аналогична конформации VEGF-связанного Fab (r.m.s.d.=0.7Å, Сα). Домены CDR обеих bH1 Fab структур полностью налагаются друг на друга и на Fv исходного антитела Герцептин® и bH1 Fv (HER2) r.m.s.d.=0.6Å, Fv антитела Герцептин® и bH1 Fv (VEGF) r.m.s.d.=1.2Å. Область CDR-L1 является исключением и значительно отличается в структурах двух комплексов; отклонение составляет 4.6 Å (Сα остатков 27-32). На Фигуре 30 показано, что CDR конформации bH1 Fab, связанного с VEGF, в значительной степени аналогичны конформациям в HER2-связанном bH1 и антителе Герцептин®, за исключением области CDR-L1. На Фигуре 30 представлено наложение CDR петель в виде трубок VEGF-связанного bH1 (темная штриховка), HER2-связанного bH1 (белый) и HER2-связанного антитела Герцептин® (легкая (светлая) штриховка). Петля CDR-L1 имеет значительно отличающуюся конформацию в двух структурах bH1 (r.m.s.d.Cα=4.6 для bH1 остатков 27-32) (Фигура 31). В комплексе HER2 домен CDR-L1 минимально участвует во взаимодействии с антигеном, а гибким является участок петли (остатки 28-30b). Для связывания VEGF вся петля имеет подходящую структуру и вносит 26% от площади поверхности, заглубленной VEGF.
Два остатка CDR-L1, Ile30c и Tyr32, имеют различные конформации и играют различную роль в связывании bH1 с HER2 или VEGF. В комплексе HER2 боковая цепь остатка Ile30c с(о)крыта в гидрофобном ядре, образованном остатками CDR-L1 и CDR-L3. В VEGF комплексе эта боковая цепь образует гидрофобные контакты с VEGF. Cα остатка Tyr32 находится в одинаковом положении в обеих структурах, но боковая цепь остатка повернута на ~130 градусов. В комплексе HER2 остаток Tyr32 располагается (укладывается) напротив рецептора, тогда как в комплексе VEGF боковая цепь, совместно с Ile29, образует гидрофобное ядро и поддерживает конформацию CDR-L1 и CDR-L3. Конформация CDR-L1 дополнительно стабилизируется водородными связями между Tyr32 и LC каркасным остатком Gly72. Структурный анализ подтверждает, что Tyr32 является критически важным для связывания VEGF, так как мутация либо в аланин, либо в фенилаланин не допускается. В отличие от связывания VEGF мутация Tyr32 в аланин (обратная мутация в остаток антитела Герцептин®) является предпочтительной для связывания HER2. Наложение двух комплексов выявляет, что VEGF пришел бы в столкновение с остатком Tyr32 области CDR-L1 в его HER2-связанном состоянии (Фигура 31). На Фигуре 31 боковые цепи остатков Tyr32, Ile30c, Ile29 и Gly72 показаны вертикальными линиями. Остатки с температурными факторами, превышающими средний, выделены более темной штриховкой (остатки 28-30b). Водородная связь между Tyr32 и Gly72 показана пунктиром.
Вышеизложенные результаты показывают, что способность к перегруппировке CDR-L1 необходима для биспецифичности bH1. Показано, что аналогичная конформационная гибкость CDR-L1 важна для распознавания антигенов природными антителами (Jimenez et al., Proc. Natl. Acad. Sci. USA 100: 92, 2003; Mylvaganam et al., J. Mol. Biol. 281: 301, 1998). Изображения на Фигурах 26, 28, 30, 31 и 32 получены с помощью координат кристаллической структуры (PDB коды, 3BDY, ЗВЕ1, 1N8Z) при использовании программы PYMOL (DeLano Scientfic, San Carlos, CA).
Сканирование bH1 методом Shotgun (шот-ган, метод дробовика)
Для изучения антигенсвязывающих сайтов bH1 Fab осуществляют сканирующий комбинаторный мутагенез методом "shotgun" (методом дробовика) с использованием фаг-дисплейных библиотек (Vajdos et al., J. Mol. Biol. 320: 415, 2002; Weiss et al., Proc. Natl. Acad. Sci. USA 97: 8950, 2000). Селекция по связыванию антигенов (hVEGF и Her2-ECD) для выделения функциональных клонов с последующим секвенированием ДНК позволяет рассчитать соотношения дикого типа/мутанта в каждом измененном положении (Vajdos et al., 2002). Затем эти соотношения используются для определения вклада каждой сканированной боковой цепи в связывание с VEGF и Her2. Результаты дают возможность картировать функциональный паратоп для связывания VEGF и Her2.
Дизайн фаг-дисплейных библиотек bH1 методом дробовика (Shotgun)
Гидрофильные (экспонированные с растворителем) остатки в областях CDR сканируют, используя фаг-дисплейные библиотеки, в которых остатки дикого типа варьируют: меняют либо на аланин, либо на остаток дикого типа (Alanine Scan), меняют либо на гомологичный остаток, либо на остаток дикого типа (Homolog Scan). Характер генетического кода требует, чтобы в библиотеку были включены другие замены, помимо остатков Wt/аланин или Wt/гомолог (Фигура 33). Методом аланин- и гомолог-сканирующего мутагенеза конструируют отдельные библиотеки тяжелой и легкой цепи. Библиотеки описаны на Фигуре 34. Вырожденность в диапазоне от 1.3×105 до 1.3×108 и фактический размер библиотек от 6×109 до 4×1010.
Конструирование шотган (Shotgun) - сканирующих фаг-дисплейных библиотек bH1
Как отмечается выше, для содействия визуализации bH1 Fab на поверхности М13 бактериофага, ранее описанную плазмиду АР2009, сконструированную для визуализации на фаге hu4D5Fab, слитого с С-концевым доменом М13 гена-3 минорного оболочечного белка, стандартными методами молекулярной биологии модифицируют таким образом, чтобы она кодировала bH1 Fab. С-конец легкой цепи содержит эпитопную (gD) метку. В качестве матрицы для библиотеки используют варианты "стоп-матрицы" ("Stop template") bH1 Fab (Sidhu et al., 2004). Легкие цепи в аланин- и гомолог-сканирующих библиотеках имеет стоп-кодоны в CDR-LI, CDR-L2 и CDR-L3, а тяжелые цепи в аланин- и гомолог-сканирующих библиотеках имеет стоп-кодоны в каждой области CDR тяжелой цепи. Библиотеки создают описанными ранее методами (Sidhu et al., 2004), используя метод мутагенеза по Kunkel (Kunkel et al., 1987) на соответствующих "стоп-матрицах".
Селекция библиотек
NUNC 96-луночные иммунопланшеты MaxiSorp покрывают мишенью улавливания (захвата) (hVEGF109, Her2-ECD или mIgG против gD) с концентрацией 5 мкг/мл и блокируют с помощью 1% BSA (вес/об) в PBS. Фаг из вышеописанных библиотек выращивают с KO7 хелперным фагом (NEB), как описано ранее (Lee et al., 2004а). Растворы фагов из библиотек с концентрацией 1013 фаговых частиц/мл добавляют в покрытые планшеты и инкубируют 1-2 часа при RT. Планшеты отмывают 8 раз буфером PBST, а затем элюируют 0.1 М HCl в течение 30 минут. Обогащение после каждого цикла селекции определяют, как описано ранее. После 2 циклов целевой селекции 50-1000-кратное обогащение наблюдают для всех библиотек, за исключением LC-Ala и LC-Hom, отсортированных по hVEGF, которые показывают 5-10-кратное обогащение. Некоторое число случайных клонов из каждой библиотеки, в которой наблюдается 50-1000-кратное обогащение, отбирают для секвенирования, описанного ранее (Sidhu et al., 2004). Библиотека LC-Ala подвергается скринингу на hVEGF-связывание методом фагового ИФА (ELISA) (Sidhu et al., 2000). Клоны, для которых наблюдаются hVEGF ELISA сигналы по меньшей мере вдвое превышающие сигналы на контрольных планшетах с иммобилизованным BSA, отбирают для секвенирования. Проводят 1 дополнительный цикл селекции библиотеки LC-Hom на hVEGF с последующим скринингом методом фагового ELISA и секвенированием VEGF-связывающих клонов.
Анализ последовательностей ДНК
"Высококачественные" последовательности из каждой библиотеки из различных вариантов целевой селекции транслируют и выравнивают (Данные не показаны). Число анализируемых последовательностей из каждой библиотеки представлено ниже, в Таблице 4.
Отношения Wt/Mut рассчитывают в каждом варьируемом положении (Фигура 35 и Фигура 36), что позволяет рассчитать приводимые значения Fwt/mut (Фигура 35 и Фигура 36), которые корректируются по визуализации делением отношений из целевой селекции на отношения из "дисплей-селекции", описанных ранее (Vajdos et al., 2002). Значение Fwt/mut выше 1 указывает на то, что в этом положении предпочтителен остаток Wt, и значение Fwt/mut меньше 1 указывает, что в этом положении предпочтительна мутация. Значение Fwt/mut>5 указывает на важную роль в связывании антигена. Важность каждого сканированного остатка CDR иллюстрируется Фигурами 37A-37D. Результаты показывают, что остатки как тяжелой цепи, так и легкой цепи вносят энергетический вклад в связывание обоих антигенов (Her2 и hVEGF). Влияние (воздействие) остатков легкой и тяжелой цепей антитела bH1 на связывание Her2 сравнивают с влиянием на это связывание исходного антитела hu4D5 (Kelley and O'Connell, 1993) (Фигура 38).
На Фигуре 39А и Фигуре 39В показано "шотган" сканирование аланином и гомологом bH1 Fab на VEGF и HER2. Эффекты мутации аланина (m1) или дополнительных мутаций (m2, m3; вследствие ограничений шотган-кодонов для аланина), или на гомологичную аминокислоту (m4) рассчитывают как отношение случаев дикого типа и мутантов (wt/mut) среди клонов, связывающихся с человеческим VEGF (Фигура 39А) или HER2 (Фигура 39В). В тех случаях, когда появляется только остаток дикого типа, отношения показываются как больше чем ">" число дикого типа. Идентичность аминокислотных замен (m1-m4) показана в виде верхнего индекса над значениями F. Когда остатком дикого типа является аланин, он замещается на глицин (m1). Знак "*" указывает на степень (содержание) bH1 остатков, которые с(о)крыты при образовании комплексов с VEGF или HER2 (*25-49% доступной (незащищенной) площади с(о)крыта (экранирована), **50-75% доступной (незащищенной) площади скрыта, ***75% или более % доступной (незащищенной) площади скрыта).
Остатки, которые вносят заметный вклад в энергетические взаимодействия, образуют функциональные паратопы, которые составляют подгруппу структурных сайтов связывания. В отличие от значительного перекрывания между сайтами контакта с антигеном, перекрывание двух функциональных паратопов является ограниченным (Фигуры 32 и 40). В частности, на основании шотган-сканирующего мутагенеза, наносят значения ΔΔG (ось y, ккал/моль) для каждой мутации на аланин (черный столбец) или гомологичную аминокислоту (белый столбец) для связывания VEGF (Фигура 40А) или HER2 (Фигура 40В). Знак "†" обозначает нижний предел, так как в данной позиции не наблюдаются мутации. Знак "*" указывает на размер площади поверхности остатков bH1, с(о)крытых при образовании комплекса VEGF или HER2 (*25-49% с(о)крыто, **50-75%, ***>75%). Связывающее взаимодействие VEGF опосредуется прежде всего LC CDR с Tyr32 области CDR-L1 и His91 области CDR-L3 в качестве "горячих точек" ядра (ΔΔGwt/ala>1.5 ккал/моль). В связывание HER2 основной вклад вносят области НС CDR. На Фигуре 32 показаны кристаллические структуры, в которых остатки bH1 и антитела Герцептин® с(о)крыты на поверхности Fab, в зависимости от их функциональной значимости (важности) (темная штриховка и белый шрифт ΔΔG≥1.5 ккал/моль; штриховка средней интенсивности (нейтральная) и белый шрифт 1≤ΔΔG<1.5 ккал/моль; легкая штриховка и черный шрифт 0.5≤ΔΔG<1 ккал/моль мутации аланина). Черным пунктиром обведена область контакта на Фигуре 28. Белым пунктиром показана граница раздела между легкой и тяжелой цепью.
Для связывания VEGF и связывания HER2 остатки функциональных паратопов распределены вдоль НС и LC, означая синергию двух цепей. Остаток Trp95 из CDR-Н3 является единственной общей "горячей точкой" для обоих взаимодействий (ΔΔGwt/ala>1.5 ккал/моль). Как отмечается выше, связывающее взаимодействие с VEGF опосредуется прежде всего областями LC CDR, тогда как в связывании HER2 доминируют области НС CDR. По сравнению с антителом Герцептин® bH1 с более слабой аффинностью связывания с HER2 (в 300 раз) сохраняет те же самые остатки для связывания с HER2 (Arg50, Trp95 и Tyr100a), тогда как значимость (важность) периферических остатков перераспределяется (Фигура 32). В целом большинство важных боковых цепей, вносящих вклад в связывание hu4D5/Her2, все еще остаются важными для связывания bH1/Her2 (ΔΔG>1.5 ккал/моль). Имеется несколько изменений. В результате вклада наблюдается много перестановок, в остатках легкой цепи содержат - некоторые остатки становятся менее важными, а некоторые более важными. В целом функциональные сайты являются частью структурного интерфейса (области контакта) кристаллической структуры комплексов bH1-VEGF и bH1-Her2.
Вкратце, взаимодействие bH1 с двумя неродственными по структуре большими белками характеризуется участием отдельного набора остатков bH1 в энергетическом взаимодействии с каждым антигеном. Хотя большая часть из двух в значительной степени перекрывающихся сайтов связывания для двух различных антигенов имеет единственную конформацию, гибкость одной CDR петли (L1) способствует аккомодации как HER2, так и VEGF. Механизм напоминает молекулярную приспособляемость (гибкость), наблюдаемую в полиспецифических антителах, связывающих неродственные малые гаптены или пептиды. В более ранних работах описывается полиспецифичность, опосредуемая либо различным позиционированием малых лигандов в пространственно различных областях антитела в одной конформации антитела (Sethi et al., Immunity 24: 429, 2006), либо несколькими уже существующими (предсуществующими) конформациями антигенсвязывающего сайта (James et al., Science 299: 1362, 2003). При рассмотрении гибкости (приспособляемости, адаптации) молекул антитела при распознавании антигена особенно примечательно, как ограниченные LC мутации могут привести к образованию антител, которые связывают два неродственных белковых антигена.
Созревание аффинности bH1
Для того чтобы изучить, можно ли повысить аффинность связывания bH1 с VEGF оптимизацией последовательности легкой цепи до того, как будут получены данные о структуре и функции, создают библиотеку, в которой диверсифицируют (получают большое разнообразие) остатков CDR в положениях с высокой степенью доступности растворителя, если исходить из кристаллической структуры h4D542 Fab (Eigenbrot et al., 2001), который, как полагают, очень похож на bH1 Fab. Нацеленные остатки варьируют: либо они находятся в виде остатков дикого типа, либо в виде нескольких гомологичных остатков (Фигура 34). Библиотеку создают так, как описано в разделе "Создание библиотек, сканируемых методом "дробовика" (Shotgun, шотган)". Метод селекции в растворе используют для селекции VEGF-связующих с повышенной аффинностью. Проводят два цикла селекции в растворе. Жесткость условий повышают в каждом цикле селекции, понижая концентрацию биотинилированного VEGF с 50 нМ в первом цикле до 20 нМ во втором цикле. Секвенируют 138 клонов из последнего цикла селекции. Найдено, что большинство клонов являются уникальными. Метод ИФА (ELISA) с применением высокопроизводительного анализатора с иммобилизованными VEGF (8-109), антителом против gD и Her2-ECD используют для идентификации клонов, которые связываются с VEGF, Her2-ECD и анти-gD mIgG, но не с BSA. Сигналы связывания VEGF-ELISA нормализуют по анти-gD ELISA сигналам для оценки относительной аффинности VEGF-связывающих клонов. Клоны с высокими значениями отношения VEGF/анти-gD отбирают для более подробного изучения. Аффинность выбранных клонов к VEGF и Her2 определяют конкурентным ИФА (ELISA) с применением Fab из фаг-дисплейной библиотеки, как описано ранее. Врианты bH1 проявляют повышенную аффинность связывания с VEGF по сравнению с исходным bH1 клоном. Примечательно, что некоторые клоны имеют немного повышенные значения IC50 по отношению к связыванию Her2, даже несмотря на то что селекция по аффинности к Her2 не проводилась. Это показывает, что возможно осуществлять созревание аффинности клона bH1 к VEGF-связыванию без заметного воздействия на способность связывания Her2. Имеется несколько клонов с повышенной аффинностью к VEGF, которые проявляют пониженную аффинность связывания с Her2 по сравнению с исходным bH1 клоном. Этот результат показывает, что легкая цепь активно способствует способности связывания bH1 с Her2, несмотря на тот факт, что тяжелая цепь является основным "вкладчиком" в энергию связывания, как показывает изучение структуры комплекса bH1-Her2 и "шотган" сканирование аланином. Последовательности и значения IC50 характеризуемых клонов представлены на Фигуре 41. Вывод о том, что большинство последовательностей являются уникальными, наводит на мысль, что последовательность легкой цепи этих вариантов еще не полностью оптимизирована по связыванию VEGF и что возможно дальнейшее повышение аффинности клона bH1 в дополнительных циклах селекции.
Как показано в Таблице 5, можно достигнуть заметного повышения аффинности единственного Fab к двум антигенам, что обычно и делается. Например, КD для человеческого VEGF повышают от 250 (bH1; IgG) до 41 (bH1-81; IgG) или 16 нМ (bH1-44; IgG), a KD для HER2 повышают от 21 (bH1; IgG) до 7 (bH1-81; IgG) или 1 нМ (bH1-44; IgG).
Аффинность повышают, вводя мутации в областях НС и LC CDR антитела bH1. Положения выбирают на основании информации о функциональных паратопах для VEGF и HER2 по данному описанию. Варианты bH1 выделяют в две стадии селекцией и скринингом фаг-дисплейных библиотек по данному описанию. Усовершенствованный клон bH1-81 выделяют селекцией по аффинности описанной библиотеки легкой цепи, полученной шотган-сканированием гомологом. На второй стадии клон с наивысшей аффинностью (bH1-44) выделяют из библиотеки рандомизацией остатков bH1-81. В частности, создают олигонуклеотиды, которые рандомизируют сайты в НС и LC bH1-81 (Таблица 5), кодируя ~50% дикого типа и 50% всех других 19 аминокислот в каждом положении (Gallop et al., Journal of Medicinal Chimistry 37: 1233, 1994).
Величины KD улучшенных по аффинности вариантов bH1 (Таблица 5) определяют для Fab фрагментов и IgG антител. Fab фрагменты и IgG антитела экспрессируют в клетках Е. Coli и клетках 293, соответственно, и очищают как представлено в данном описании. Измерения, сделанные методом поверхностного плазменного резонанса (SPR) на приборе BIAcore 3000, используют для определения величины аффинности связывания фрагментов Fab и IgG антител, как описано Lee et al. (J. Mol. Biol. 340: 1073, 2004). Для изучения аффинности антитела в виде одновалентных Fab фрагментов антигены (hVEGF109, мышиный VEGF102 и HER2 ECD) иммобилизуют с низкой плотностью на BIAcore CM5 чипе. Fab фрагменты в серийных разведениях контактируют с иммобилизованными антигенами и методом SPR количественно определяют отклики связывания. Для расчета kon, koff и KD используют ленгмюровскую модель связывания 1:1. Для определения KD IgG антител IgG захватывается на BIAcore CM5 чипе иммобилизованным антителом против Fc и экспонируется с серийными разведениями hVEGF109, мышиного VEGF102 и HER2-ECD. В случае HER2 с целью определения КD используют простую 1:1 ленгмюровскую модель связывания, тогда как для VEGF требуется двухвалентная модель анализируемого вещества. Все эксперименты проводят при 30°С.
В Таблице 5 рандомизированные положения показаны жирным шрифтом и представлены последовательности CDR (SEQ ID NO:1-9 и 39-41) антитела bH1, bH1-81 и bH1-44 и их аффинности (определенные методом поверхностного плазмонного резонанса).
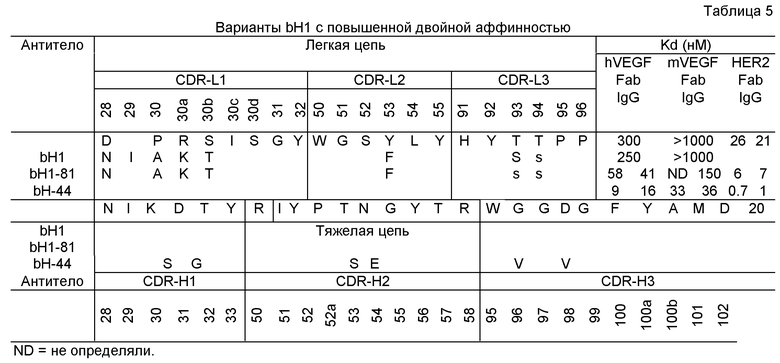
Одновалентную аффинность антител к человеческому VEGF109, мышиному VEGF102 и HER2 ECD определяют на чипе BIAcore. В Таблице 5 показаны характерные константы диссоциации (Kd) для каждого связывающего взаимодействия. Рецептор-связывающий фрагмент VEGF (VEGF109) используют в эксперименте с BIAcore чипом, так как варианты bH1 связывают полноразмерный белок (VEGF165) и VEGF109 с аналогичной аффинностью в конкурентных реакциях в растворе (данные не показаны). Для расчета величины Kd для Fab фрагментов/IgG антител по данному описанию используют различные форматы анализа и расчетные модели. Различные форматы анализа/расчетов (оценки) дают согласующиеся константы диссоциации для отдельных взаимодействий.
Определение IgG активности в клеточных анализах
Для того чтобы определить, могут ли антитела bH1 и 3-1 ингибировать индуцированную hVEGF165 пролиферацию эндотелиальных клеток пупочной вены человека (HUVEC), их тестируют в анализе пролиферации. Выращивают эндотелиальные клетки пупочной вены человека (HUVEC) (Cambrex, East Rutherford, NJ) и анализируют, как описано ранее (Fuh et al., J. Biol. Chem. 273: 11197, 1998). Примерно, 4000 клеток HUVEC засевают в каждую лунку 96-луночного планшета для клеточных культур и инкубируют в смеси сред: модифицированной по методу Дульбекко среды Игла/F-12 среды (1:1), дополненной 1.0% (об/об) фетальной бычьей сыворотки (среда для анализа) в течение 18 часов. Затем в клетки добавляют свежую среду для анализа с фиксированными количествами VEGF (конечная концентрация 0.2 нМ), которую сначала титруют как уровень VEGF, который может стимулировать субмаксимальный уровень синтеза ДНК, а затем к клеткам добавляют антитела против VEGF (например, bH1) с увеличивающимися концентрациями. После инкубации при 37°С в течение 18 часов клетки с добавлением 0.5 мкКи/лунка [3H]Тимидина выдерживают в пульсовом режиме в течение 24 часов и собирают для подсчета на сцинтилляционном счетчике для микропланшетов TopCount. Результаты показывают, что как 3-1, так и bH1 может ингибировать индуцированный VEGF рост клеток HUVEC, предупреждая передачу hVEGF-индуцированного сигнала и последующую пролиферацию. Антитело Авастин® (против VEGF) используют в качестве позитивного контроля, а антитело Гецептин® в качестве негативного контроля (Фигура 42).
Для изучения связывания биспецифических антител против Her2/VEGF с Her2, экспрессируемым на клетках млекопитающих, методом проточной цитометрии изучают связывание антител bH1 и bH3 с фибробластами NR6, сверхэкспрессирующими Her2 (NR6-Her2). Один миллион клеток NR6-Her2 инкубируют со 100 мкг/мл Fab и IgG в течение 1 часа, а затем инкубируют с конъюгированным с Alexa488 мышиным антителом против человеческого IgGв течение 1 часа. В качестве негативного контроля изучают Fab и IgG связывание с неэкспрессирующими клетками NR6. Как показано на Фигуре 43, bH1 и bH3 специфически связываются с Her2 на клетках NR6 в виде Fab и в виде IgG.
На Фигуре 44 показаны результаты экспериментов по конкурентному связыванию bH1 с VEGF или HER2. bH1 ингибирует индуцированную VEGF пролиферацию эндотелиальных клеток пупочной вены человека (HUVEC) с IC50 200 нМ, что согласуется с его аффинностью 300 нМ, и пролиферацию HER2-экспрессирующей линии раковых клеток молочной железы ВТ474 после инкубации в течение 5 дней, хотя с более низкой эффективностью, чем антитело Герцептин®, из-за пониженной аффинности (Фигура 45). Герцептин® IgG антитело и бевацизумаб (анти-VEGF) служат в качестве контроля. Как показано на Фигуре 45, антитела bH1-81 и bH1-44 ингибируют VEGF-индуцированную пролиферацию клеток HUVEC и рост клеток ВТ474 в большей степени, нежели bH1. Повышенная эффективность bH1 вариантов коррелирует с их относительными аффинностями. Вариант с наивысшей аффинностью, bH1-44, ингибирует рост клеток HUVEC и ВТ474 с эффективностью, аналогичной эффективности бевацизумаба или антитела Герцептин, соответственно.
Для проведения этих экспериментов VEGF-стимулированные клетки HUVEC обрабатывают, добавляя увеличивающиеся концентрации человеческого IgG, инкубируют в течение 2 дней и пролиферацию количественно определяют, как описано в Liang et al. (J. Biol. Chem. 281: 951, 2006). Раковые клетки молочной железы ВТ474 культивируют в среде RPMI, дополненной 10% FBS. Для анализов засевают 104 клеток/лунка в лунки 96-луночного планшета и инкубируют в течение ночи (18 часов) при 37°С. К клеткам добавляют увеличивающиеся концентрации человеческого IgG. Клетки инкубируют при 37°С в течение пяти дней, а затем добавляют 10% AlamarBlue (Biosource International, Camarillo, CA) в соответствии с инструкциями производителя. Антителозависимое ингибирование пролиферации клеток, экспрессирующих HER2, определяют, измеряя сигнал флуоресценции через 6 часов.
Пример 6. Анализ специфичности связывания
Определяют специфичность связывания антител, полученных из LC библиотеки. Антитела IgG, связывающиеся с различными иммобилизованными очищенными белками или клеточными лизатами, включая собственные антигены, анализируют твердофазным ИФА (ELISA). Антигены иммобилизуют и инкубируют с hIgG в концентрации 15 мкг/мл в течение часа. Связанные IgG детектируют спектрофотометрически (оптическая плотность при 450 нм; ось y; Фигура 46). Белки, участвующие в анализе, представляют собой (слева направо на Фигуре 46): сосудистый эндотелиальный фактор роста A (VEGF), мышиный сосудистый эндотелиальный фактор роста (мышиный VEGF), сосудистый эндотелиальный фактор роста С, (hVEGF-С), сосудистый эндотелиальный фактор роста D, (hVEGF-D), внеклеточный домен HER2 (HER2 ECD), внеклеточный домен рецептора эпидермального фактора роста (hEGFR), внеклеточный домен ErbB3/HER3 (HER3 ECD), человеческий "рецептор смерти" 5 (hDR5), бычий сывороточный альбумин (БСА, BSA), казеин, фетальную бычью сыворотку (FBS), нейтравидин, 5% молоко, лизат мышиных фибробластов и лизат мышиных фибробластов с добавлением hVEGF-А или HER2 ECD. На Фигуре 46 "усы" представляют стандартную ошибку среднего (SEM) в двух повторах. Антитела bH3, 3-1, bD1, bD2, 4-1 и 4-5 не проверяют на связывание с мышиным VEGF, HER3 ECD, нейтравидином, 5% молоком, клеточным лизатом с добавлением hVEGF-A и клеточным лизатом с добавлением HER2 ECD.
Также определяется способность различных антител (антитело Авастин®, антитело Герцептин®, bH1, bH3, bH4, bH1-81 и bH1-44) блокировать связывание VEGF с VEGF-рецепторами (Фигура 47). Биотинилированный человеческий VEGF 155 (Фигура 47А) или мышиный VEGF164 (Фигура 47В) уравновешивают с увеличивающимися концентрациями IgG (ось x). Несвязанный VEGF захватывают на слитом белке иммобилизованный человеческий VEGFR2-ECD Fc и детектируют на спектрофотометре (оптическая плотность при 450 нм, ось y). Аналогичное ингибирование наблюдают также с VEGFR1. Антитела против VEGF блокируют связывание VEGF с VEGF рецепторами.
Было показано, что антигены VEGF и HER2 конкурируют с bH1-44 биспецифическим IgG антителом в растворе (Фигура 48). Человеческое bH1-44 IgG антитело с концентрацией 0.1 нМ инкубируют с 0.1 нМ раствором биотинилированного человеческого VEGF 155 в присутствии увеличивающихся концентраций HER2 ECD. bH1-44 захватывается (улавливается) иммобилизованным Fc против человеческого иммуноглобулина и bH1-44-связанным биотин-VEGF, детектируемым комплексом стрептавидин-HRP. HER2 ECD, связанный с захваченным bH1-44, детектируют, используя связывание мышиного антитела против HER2 с неперекрывающимся эпитопом на HER2, а затем с конъюгатом HRP-антитела козы против мышиного IgG (Фигура 48А). Конъюгат человеческий bH1-44 IgG с концентрацией 0.2 нМ инкубируют с 0.6 нМ раствором биотинилированного HER2 в присутствии увеличивающихся концентраций человеческого VEGF165. bH1-44 захватывается иммобилизованным антителом против человеческого Fc и связанный с bH1-44-биотин-HER2 обнаруживают конъюгатом стрептавидин-HRP (Фигура 48В).
Специфическое связывание bH1 и bH1-44 с клетками также определяют методом FACS (сортинг флуоресцентно-активированных клеток; Фигура 49). Биспецифические антитела (bH1 и bH1-44) связываются с HER2-экспрессирующими мышиными фибробластами (NR6) (Фигура 49В), но не с HER2-негативными NR6 клетками (Фигура 49А). 0.5-1 миллион клеток инкубируют с 15 мкг/мл hIgG на льду в течение часа. Первичные антитела, связанные с клетками, детектируют, используя вторичное конъюгированное с флуоресцентным красителем РЕ антитело козы против IgG человека. Клетки анализируют на проточном цитометре FACSCalibur. bH1 и bH1-44 не вступают в перекрестную реакцию с крысиным ортологом HER2, поскольку не замечено никакого связывания с мышиными фибробластами, трансфецированными крысиным геном neu (крысиный ортолог HER2).
Для того чтобы более подробно изучить специфичность вариантов антитела bH1, а именно bH1-81 и bH1-44, проводят эксперименты по иммунопреципитации, обнаружено, что варианты антитела bH1 специфически вызывают образование иммунопреципитата VEGF или HER2 из лизатов мышиных фибробластов (NR6), но не других белков (Фигура 50). Клетки NR6 неспецифически биотинилируют, лизируют и клеточные мембранные белки солюбилизируют в присутствии детергентов. Клеточные лизаты, соответствующие по плотности 5-10 миллионов/мл NR6 клеток, NR6 клеток с добавлением 0.1 мкг/мл биотинилированного VEGF165, или HER2 - сверхэкспрессирующих клеток NR6 инкубируют с 15 мкг/мл антитела. Антитело захватывают, используя иммобилизованный на сефарозных гранулах белок А, и связанные белки элюируют. Элюированные белки разделяют методом SDS-PAGE. клеточные лизаты, соответствующие по плотности клеток, примерно, 25-50000 клеток и иммунопреципитат, примерно, 0.12-0.25 миллионов клеток, наносят на гель. Захваченные биотинилированные белки детектируют Вестерн-блоттингом с помощью конъюгата стрептавидин - HRP.
Пример 7. Анализ активности IgG in vivo
Чтобы проверить, транслируется ли двойная активность этих антител in vitro в соответствующую активность in vivo, мы используем мышиные модели опухолевых ксенотрансплантатов, заведомо отвечающие на лечение антителом против VEGF (Colo205, линия клеток колоректального рака) или антителом Герцептин® (ВТ474М1, линия клеток рака молочной железы). В частности, ксенотрансплантаты Colo205 вводят мышам nu/nu, а ксенотрансплантаты ВТ474М1 вводят мышам beige/nude/XID. Все исследования на животных проводят в соответствии с рекомендациями Американской ассоциации по аттестации и аккредитации содержания лабораторных животных и Комиссии по контролю за содержанием и использованием лабораторных животных Genentech.
В частности, ВТ474М1 (получены в собственной лаборатории) и Colo205 (АТСС, Manassas, VA) клетки культивируют в среде RPMI/10% фетальной бычьей сыворотки. Смесь 5×106 клеток ВТ474М1, суспендированных в буферном солевом растворе Хэнкса (HBSS), и матригеля (1:1) инъецируют в жировую ткань молочной железы мышей Harlan beige nude XID mice (Indianapolis, IN), которым подкожно имплантированы гранулы эстрадиола. В случае ксенотрансплантатов Colo205 5×106 клеток Colo205 в HBSS инъецируют подкожно мышам Charles River nu/nu (Hollister, CA). Когда средний размер опухоли достигнет ~200 мм3, мышей произвольно делят на 7 групп по 8 мышей (ВТ474М1) или по 10 мышей (Colo205). Антитела вводят интраперитонеально раз в неделю. Размер опухоли определяют дважды в неделю. Объем рассчитывают как V=0.5ab2 (а обозначает самую большую длину опухоли, a b обозначает длину стороны, перпендикулярной а). Для статистической оценки применяют односторонний анализ, а затем двусторонний t-критерий Стьюдента. Корректировка альфа-уровня за счет множественного сравнения (Bonferroni) не меняет значимости наших выводов. Частичные отклики (PR) определяют как уменьшение объема опухоли на 50-99% по сравнению с V0. Образцы сыворотки берут через 7 дней после первого и третьего введения лекарства. Концентрацию человеческого антитела определяют методом твердофазного ИФА (ELISA). Антитело осла против человеческого IgG Fc иммобилизуют на иммунопланшете. Разведения сыворотки и эталонного антитела инкубируют на планшете в течение 2 часов. Связанное антитело детектируют пероксидазой хрена, конъюгированной с антителом козы против человеческого IgG Fc, а затем раствором ТМВ субстрат/1 М фосфорная кислота. Планшеты считывают при 450/620 нм. Концентрацию образцов определяют, используя четырехпараметрический алгоритм.
Группы, получавшие bH1-44, сравнивают с группами, получавшими антитело против VEGF (B20-4.1) (Liang et al., J. Biol. Chem. 281:951, 2006), антитело Герцептин® или его комбинацию (антитело Герцептин® + анти-VEGF), чтобы подтвердить, что антитело bH1-44 способно ингибировать рост опухоли, опосредованный VEGF и HER2. Во всех группах высокие уровни антитела присутствует в сыворотке из Colo205 ксенотрансплантатов (определено методом ELISA) через 7 дней после начала лечения, что указывает на нормальную фармакокинетику (Таблица 6).
Еженедельно вводимая доза bH1-44 10 мг/кг ингибирует рост опухоли из клеток Colo205 по сравнению с контрольным антителом (р<0.0001, n=10) с эффективностью, аналогичной эффективности антитела против VEGF (10 мг/кг/неделя), тогда как антитело Герцептин® не оказывает никакого влияния на рост опухолевых клеток Colo205 (р=0.12, n=10). Как предполагалось, комбинированная терапия имеет такую же эффективность, как и одно антитело против VEGF. Антитело bH1-44, вводимое в дозе 10 и 20 мг/кг/неделя, вызывает дозозависимую реакцию. На модели ВТ474М1 наблюдают заметное ингибирование роста опухоли в группе мышей, пролеченных антителом bH1-44 (10 мг/кг/неделя, р=0.0005, n=8 и 20 мг/кг/неделя, р=0.0001, n=7). Подобно группам, которым вводят антитело Герцептин® или комбинацию Герцептин®/антитело против VEGF, более чем у половины животных, пролеченных антителом bH1-44, наблюдается регрессия более чем 50% от начального объема (т.е. частичный отклик, Фигура 51). С другой стороны, при введении только одного антитела против VEGF наблюдается незначительное увеличения ингибирующего действия на ВТ474М1 по сравнению с контролем (р=0.06, n=7) и не наблюдается частичный (или парциальный) отклик. Таким образом, показано, что биспецифическое антитело bH1-44 ингибирует два различных (отдельных) механизма роста опухоли in vivo.
Вышеприведенные результаты указывают на возможность (потенциал) вариантов антитела bH1 с повышенной аффинностью (например, bH1-44 и bH1-81) ингибировать два механизма, которые являются важными для роста опухоли in vivo.
Пример 8. Определение характеристик области контакта (интерфейсов) при связывании VEGF-и HER2-с bH1 и bH1-44
Для дальнейшего сравнения структурных характеристик bH1 и bH1-44 идентифицируют области контакта (интерфейсы) при связывании VEGF и HER2 с этими антителами. Структурные контакты, приведенные в Таблице 7, определяют на основании координат кристаллических структур 3BDY (bH1/VEGF) и 3ВЕ1 (bH1/HER2). Область контакта рассчитывают, применяя программу XSAE. Эта программа позволяет определять область контакта (границу раздела) как полярную, гидрофобную и смешанную. В Таблице 7 приводятся остатки bH1 с >25% от общей площади поверхности, с(о)крытой (экранированной) при связывании HER2 или VEGF. В Таблице 7 приводятся также остатки VEGF и HER2 на расстоянии не более 4.5Å от остатков bH1. Площадь поверхности каждого остатка, с(о)крытую при образовании комплекса, рассчитывают, используя программу для моделирования iMOL с учетом координат кристаллических структур 3BDY, 3ВЕ1 и 1N8Z (PDB). Площади полярной и гидрофобной области контакта (интерфейса), приведенные в Таблице 11, показывают площадь полярного интерфейса и половину смешанного. Приведенная площадь гидрофобного интерфейса состоит из гидрофобных областей (площадей) и половины смешанной области.
Изучение кристаллической структуры и сканирование аланином показывают, что bH1 сохраняет тот же самый связывающий эпитоп на HER2, который имеет антитело Герцептин® (Bostrom et al., 2009). Кристаллическая структура Герцептин® Fab в комплексе с HER2 хорошо совмещается со структурой в bH1/HER2 комплексе (r.m.s.d 0.8 Å) (Bostrom et al., 2009; Cho et al., 2003). Кроме того, остатки антитела Герцептин®, которые вносят более 10% общей энергии связывания, на основании сканирования аланином являются консервативными, и многие из них являются также частью связывающих "горячих точек" bH1 и bH1-44 (Bostrom et al., 2009; Kelley and O'Connell, 1993) (Таблица 14, Фигура 62). Интерфейсы между bH1/VEGF и bH1/HER2 экранируют (скрывают) 1506 Å2 и 1579 Å2, соответственно, и являются, главным образом, гидрофобными (60% и 63%, соответственно). Интерфейс связывания Герцептин®/НЕР2 имеет размер и состав, аналогичные размеру и составу интерфейса bH1/HER2 (1524 Å2, 60% гидрофобный. Таблица 11), и также характеризуется высокой комплементарностью конфигурации (Таблица 8) (Bostrom et al., 2009).
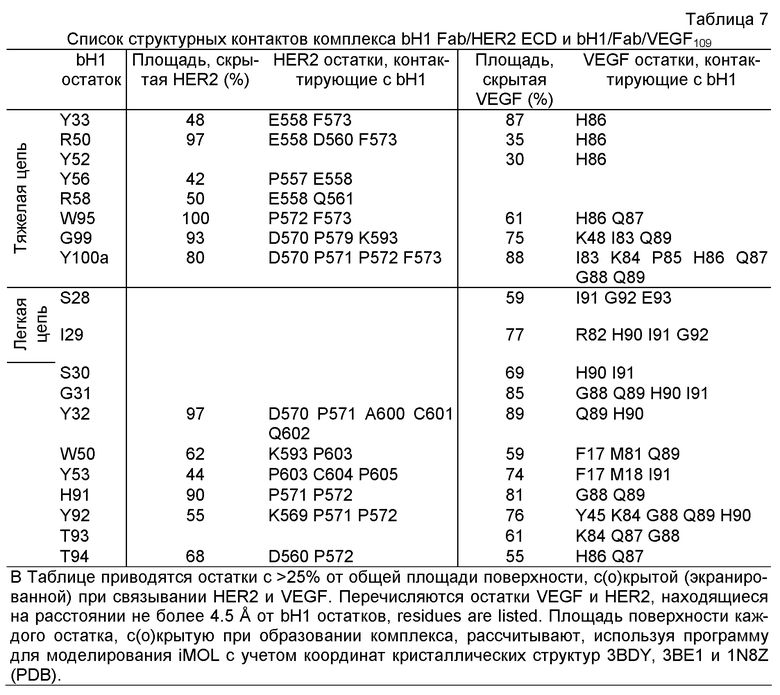
Комплементарность конфигурации (представлена как Sc в Таблице 8) между антителом и антигеном определяют, как описано в (Lawrence et al., 1993). Высокая комплементарность конфигурации в комплексах bH1/VEGF и bH1/HER2, аналогичная комплементарности между антителом Герцептин® и HER2, находится в пределах для описанных комплексов антиген-антитело (Sc ~ 0.64-0.68; Lawrence et al., 1993). Совмещение (наложение) HER2 с bH1 в той конформации, в которой это антитело находится в комплексе с VEGF, или совмещение VEGF с bH1 в той конформации, в которой это антитело находится в комплексе с HER2, выявляет низкую комплементарность конфигурации в том случае, когда антитело помещается рядом с несвязанным антигеном (Sc ~ 0.35; Lawrence et al., 1993). Результаты показывают степень преобразования bH1 в процессе приспособления к двум разным антигенам.
Аффинность bH1 повышают отбором высокоаффинного варианта bH1-44 из фаг-дисплейных библиотек антитела bH1. Аланин-сканирующий мутагенез методом шотган (дробовика) показывает, что bH1-44 сохраняет "горячую точку" для связывания bH1 с антигеном (Таблицы 9А-В, 10 и 14). Аланин-сканирующий мутагенез bH1-44 методом шотган осуществляют описанными выше методами шотган аланин-сканирующего мутагенеза bH1.
В Таблицах 9А-В эффекты мутации на аланин (m1) или дополнительных мутаций (m2, m3; вследствие ограничения шотган кодонов) или на гомологичную аминокислоту (m4) рассчитывают как отношение частоты дикого типа (wt), или wt/mut, для клонов, связывающих VEGF (Таблица 9А) или HER2 (Таблица 9В). Если wt остатком является аланин, он заменяется на глицин (m1). Отношения wt/mut корректируют по эффектам фолдинга/экспрессии белка делением в отношении wt/mut значений из "фаг-дисплейной селекции", получают значения F. "Дисплей-селекцию" осуществляют независимо, отбирая клоны, связывающиеся с белком L, который связывает нелинейный эпитоп на легкой цепи антитела. Коль скоро только тяжелая цепь Fab сливается с оболочечным белком фага (р3), связывание белка L указывает на корректный фолдинг и ассоциацию легкой цепи и тяжелой цепи.
В Таблице 10 приведены остатки антитела bH1 и bH1-44, которые контактируют с VEGF и/или HER2 в кристаллической структуре. Энергетические "горячие точки" (хотспоты, hotspots) для связывания определяются остатками антитела, которые дают в результате величину ΔΔG больше или равную, примерно, 10% от общей связывающей энергии взаимодействия.
Данные в Таблице 11 показывают, что полярность и размер областей контакта аналогичны в комплексах bH1/VEGF, bH1/HER2 и Герцептин®/HER2. Полярность каждой области контакта анализируют, используя программу XSAE. Если не указано иначе, все цифры, приведенные в Таблице 11, представляют собой площадь в Å2.
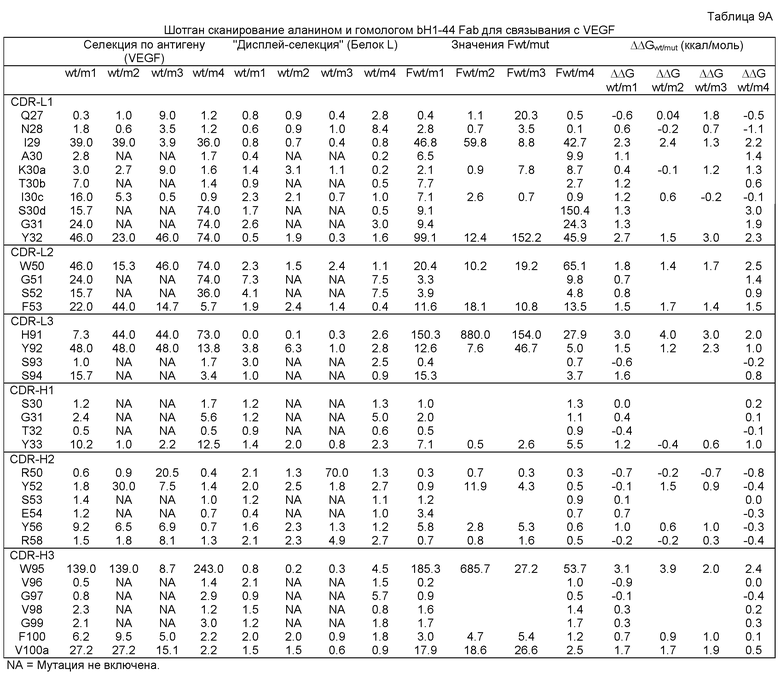
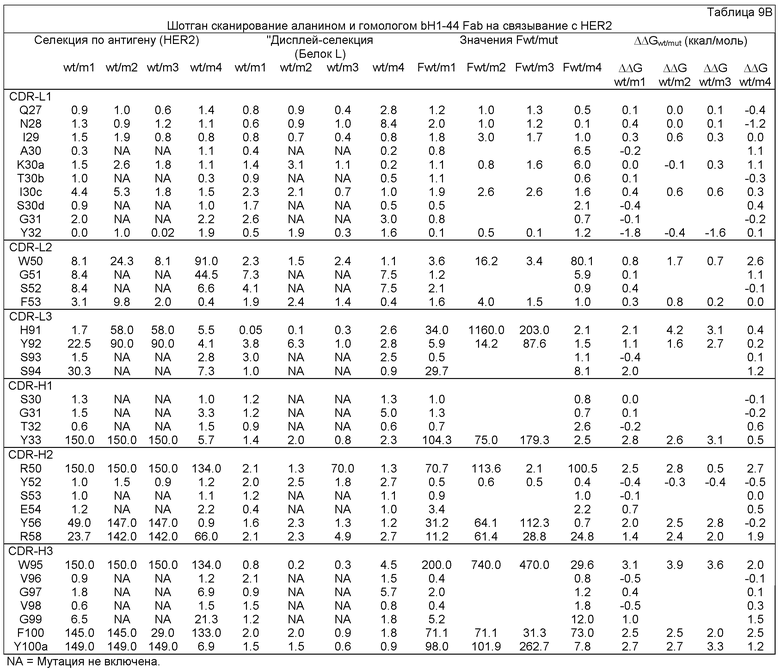
Антитело bH1-44 с двойной специфичностью к HER2/VEGF сохраняет кинетику связывания HER2 антитела Герцептин®.
Для изучения кинетики связывания bH1 и его Fab вариантов с иммобилизованным VEGF или HER2 используют метод поверхностного плазменного резонанса (Таблица 12). SPR анализ проводят на приборе BIAcore 3000. Внеклеточные домены VEGF109 и HER2 иммобилизуют на чипах СМ5 при плотности, которая позволяет достигнуть Rmax в интервале 50-150 RU. Серийные разведения Fab в PBS с 0.05% Твин20 вводят при скорости потока 30 мкл/мин. Значение откликов связывания корректируют, вычитая из величины отклика показания контрольной проточной кюветы и нормализуя по буферным эффектам. Для определения ka (константы скорости ассоциации) и kd (константы скорости диссоциации) используют ленгмюровскую модель 1:1. Значения KD определяют из соотношения ka и kd.
Взаимодействие bH1 Fab/VEGF характеризуется относительно высокой константой скорости ассоциации (kon=3.7×104) и быстрой скоростью диссоциации (koff=0.013), что в результате дает умеренную величину KD 300 нМ. Аффинность при взаимодействии bH1/HER2 (KD=26 нМ, kon=9.6×104, koff=2.4×10-3) в 52 раза ниже, чем при взаимодействии Герцептин®/HER2 (KD=0.5 нМ, kon=7.1×105, koff=3.5×10-4), причем скорость ассоциации медленнее, а скорость диссоциации быстрее. Для вариантов bH1 с повышенной аффинностью, bH1-81 и bH1-44, наблюдается повышение как константы скорости ассоциации, так и константы скорости диссоциации при взаимодействии с VEGF и с HER2. Высокоаффинный клон bHl-44 связывает HER2 с аффинностью, аналогичной аффинности Герцептина® (KD=0.2 нМ, Таблица 12).
В Таблице 12 представлены кинетические профили bH1 вариантов и антитела Герцептин®, определенные методом поверхностного плазменного резонанса на приборе BIAcore при 30°С. В этих экспериментах найдено, что Fab связываются с иммобилизованным VEGF или HER2, а константу скорости ассоциации (ka), константу скорости диссоциации (kd) и константу равновесной диссоциации (KD) определяют, используя ленгмюровскую модель связывания 1:1. Антитело bH1-44 имеет кинетический профиль и аффинность к HER2, аналогичные кинетическому профилю и аффинности антитела Герцептин®. Два двойных мутанта (bH1-44 I29A+Y32A и bH1-44 R50A+R58A), утратившие способность связываться с VEGF или HER2, сохраняют кинетический профиль и аффинность к другому антигену.
Антитела с двойной специфичностью взаимодействуют с HER2 и с VEGF со сходными термодинамическими свойствами.
Изменение энтальпии (ΔН) и энтропии (ΔS) во взаимодействии между bH1 Fab вариантами и двумя антигенами, VEGF (рецептор-связывающим доменом VEGF, VEGF8-109) и внеклеточным доменом (ECD) HER2 также определяют (Фигура 59A-F, Фигура 60, Таблица 13), используя изотермическую титрационную калориметрию (ITC).
Микрокалориметрические измерения взаимодействий между фрагментами Fab и человеческим VEGF109 и внеклеточным доменом антигена HER2 проводят на титрационном калориметре VP-ITC (Microcal Inc.), как описано в (Starovasnik et al., 1999). Белковые растворы подвергают интенсивному диализу против фосфатно-солевого буферного раствора. Диализ антигена и Fabs проводят в одном и том же сосуде, чтобы минимизировать тепловые эффекты смешения из-за разницы буферной композиции. Растворами Fabs в концентрации 100-220 мкМ титруют растворы антигенов (HER2-ECD или VEGF109) с концентрацией 10-22 мкМ. Такая концентрация антигена необходима для точного измерения энтальпии, но препятствует определению КD в тех случаях, когда аффинность связывания высока. Для получения двукратного избытка антитела делают пятнадцать или шестнадцать инъекций. Определяют теплоту реакции, вычитают теплоту разведения Fab и рассчитывают ΔН.
Константы диссоциации (KD), определенные методом поверхностного плазменного резонанса (Таблица 12), используют для расчета свободной энергии связывания (ΔG) по уравнению
ΔG=RT ln (KD).
Изменение энтропии при ассоциации (ΔS) рассчитывают по уравнению
ΔS=(ΔН-ΔG)/Т, где Т обозначает температуру (K).
Для определения ΔСр проводят микрокалориметрические измерения, как описано выше, при различных температурах в диапазоне от 20 до 37°С. ΔСр определяют методом линейной регрессии, строя график зависимости ΔН от температуры (Фигура 62).
Впервые характеризуют взаимодействие антитела с двойной специфичностью, bH1, с любым из двух его антигенов, VEGF и HER2. Связывание bH1 с VEGF и HER2 имеет аналогичные термодинамические характеристики (свойства) (Таблица 13). Оба взаимодействия, изучаемые при 30°С в PBS при рН 7.4, являются экзотермическими (ΔН=-2.4 и -2.4 ккал/моль для VEGF и HER2 соответственно. Таблица 13, Фигура 60) с в высшей степени благоприятствующим вкладе изменения энтропии в энергию связывания (-TΔS=-6.6 и -7.9 ккал/моль для VEGF и HER2 соответственно. Таблица 13, Фигура 60).
В Таблице 13 даны значения ΔG (свободная энергия связывания), ΔS (изменение энтропии) и ΔН (изменение энтальпии) в ккал/моль. Показанные значения аффинности измеряют по меньшей мере в двух независимых экспериментах методом кинетического анализа с помощью BIAcore при 30°С. Значение ΔН измеряют методом ITC, оно является средним результатом, полученным в двух или трех измерениях, с поправкой на стандартное (среднеквадратичное) отклонение. Значения ΔG и ΔS рассчитывают, как описано выше.
Высокоаффинные варианты bH1-81 и bH1-44 имеют термодинамические профили, аналогичные bH1. Их взаимодействие с VEGF и HER2 также характеризуется благоприятной энтальпией и энтропией (Таблица 13, Фигура 60). Для взаимодействия с повышение аффинности ассоциируется со значительно более благоприятным изменением энтальпии (ΔН=-7.1 для bH1-44 по сравнению с -2.4 ккал/моль для bH1 при 30°С) и немного менее позитивным изменением энтропии (-TΔS=-6.6 для bH1-44 по сравнению с -4.7 для bH1 при 30°С, Таблица 13, Фигура 60). Повышенная аффинность к HER2 также ассоциируется с более благоприятным изменением энтальпии (ΔН=-5.3 по сравнению с -2.4 ккал/моль, 30°С, Таблица 13, Фигура 60).
Термодинамика взаимодействия bH1-44 и Герцептина® с HER2 различна.
В отличие от взаимодействия антител с двойной специфичностью взаимодействие HER2/Herceptin® характеризуется большим благоприятным изменением энтальпии (ΔН=-13.6 ккал/моль) без какого-либо заметного изменения энтропии (-TΔS=-0.3 ккал/моль. Фигура 60, Таблица 13) (Kelley et al., 1992). Хотя bH1-44 взаимодействует с HER2 с аффинностью, сходной аффинности Герцептина®, свободная энергия связывания слагается из более значительного энтропийного компонента (-TΔS=-8.1 ккал/моль, 30°С) и меньшего энтальпийного компонента (ΔН=-5.3 ккал/моль, 30°С). Различные термодинамические свойства противоречат многим сходным характеристикам связывания HER2 между Герцептином® и bH1-44, которые включают аффинность, кинетику и множество остатков энергетических "хотспотов" ("горячих точек"). Хотя хотспот - остатки Герцептина®, которые вносят более 10% общей энергии связывания с HER2, аналогичны соответствующим остаткам bH1 и bH1-44, имеются некоторые четкие различия.
В Таблице 14 показаны "горячие точки" (хотспоты) антител bH1, bH1-44 и Герцептина® для связывания HER2, определенные аланин-сканирующим мутагенезом. Мутагенез осуществляют, как описано в Kelley et al., 1993. Цифры в Таблице 14 показывают изменения в свободной энергии связывания (ΔΔGwt-mut) при замене остатка на аланин. Хотспот - остатки в Таблице 14 заштрихованы и определяются тем, что ΔΔG больше или равно 10% от общей свободной энергии связывания (ΔG).
Остатки LC-Thr94, НС-Tyr33, НС-Asp98 сохраняются в последовательности в bH1, но имеют разные функции при связывании HER2 (Таблица 14, Фигура 61). Следовательно, мутации в антигенсвязывающем сайте Герцептина®, которые участвуют в связывании VEGF, по-видимому, вносят некоторые фундаментальные изменения в антигенсвязывающий сайт, которые влияют на взаимодействие с HER2. Антитела с двойной специфичностью приспосабливают введенные мутации, используя другую стратегию распознавания HER2, которая определяет такую же высокую аффинность к HER2, как и у антитела Герцептин®. Интересно отметить, что за исключением LC-Ser94 антитела bH1-44, мутации, которые повышают аффинность к HER2 более чем в 100 раз по сравнению с bH1, не являются участками связывающей "горячей точки", но, по-видимому, оптимизируют имеющиеся взаимодействия.
Большая отрицательная теплоемкость при взаимодействиях с двойной специфичностью
Чтобы глубже понять общую энергетику, управляющую взаимодействиями антител с двойной специфичностью, и как они отличаются от моноспецифического исходного Герцептина®, проводят ряд экспериментов по изучению следующих трех взаимодействий Fab/антиген: bH1-44 с VEGF или HER2 и Герцептина® с HER2. Теплоемкость взаимодействий с двойной специфичностью измеряют, определяя энтальпию связывания (ΔН) при различных температурах в интервале от 20°С до 37°С (ΔТ=17°С, Фигура 62, Таблица 15). Теплоемкость (ΔСр) является функцией ΔН и температуры (Т) и может быть описана уравнением
ΔСр=δ(ΔН)/δТ.
ΔСр определяется по углу наклона кривой зависимости ΔН от температуры линейной регрессией (Фигура 62, Таблица 15). ΔСр антитела bH1-44 определяют до -400 кал/мольK для взаимодействия с VEGF, и -440 кал/мольK для взаимодействия с HER2. Большая отрицательная теплоемкость указывает на важность гидрофобного эффекта, как описано ранее (Kauzmann, 1959), что согласуется с гидрофобной природой структурных поверхностей контакта (интерфейсов) в двух комплексах (Таблица 11). Величина ΔСр для комплекса Герцептин®/HER2, определенная ранее как -370 кал/мольK в аналогичном интервале температур (Kelley et al., 1992), ниже, чем величина ΔСр комплекса bH1-44/HER2, но все еще указывает на важную роль гидрофобного эффекта при связывании HER2.
Изменение общей энтропии (ΔS) свободной энергии связывания представляет собой сумму изменений энтропии из трех источников (Murphy et al., 1994): изменений энтропии, ассоциированных с десольватацией связывающих поверхностей (поверхностей контакта) (ΔSSOLV), изменений энтропии, вызванных потерей степеней свободы вращательного и поступательного движения (ΔSRT), и изменений энтропии вследствие изменений динамических характеристик взаимодействующих молекул, относящихся к конфигурации и конформации (ΔSCONF).
(1) ΔSTOT=ΔSSOLV+ΔSRT+ΔSCONF
Обычно только ΔSSOLV является положительной величиной, тогда как обе величины, ΔSRT и ΔSCONF, являются отрицательными. Важный термин для энтропии, ΔSRT, при ассоциации двух молекул можно оценивать до -8 кал/Kмоль, как описано в (Murphy et al., 1994). Можно предположить, что в ΔSSOLV преобладает гидрофобный эффект вследствие сокрытия площади неполярной поверхности и эту величину можно описать как функцию ΔСр:
(2) ΔSSOLV=ΔCp ln(T/T*), T*=385 K
Таким образом, ΔSCONF можно определить как:
(3) ΔSCONF=ΔSTOT-ΔSRT-ΔSSOLV
Согласно уравнению (3), ΔSSOLV оценивается до 96 кал/моль-1K-1 для bH1-44/VEGF, 105 кал/моль-1K-1 для bH1-44/HER2 и 89 кал/моль-1K-1 для Герцептин®/НЕР2 (Таблица 15). Оно транслируется в ΔSCONF -72 кал/моль-1K-1 для bH1-44/VEGF, -70 кал/моль-1K-1 для bH1-44/HER2 и -80 кал/моль-1K-1 для комплекса Герцептин®/HER2 (Таблица 15).
Чтобы изучить общую стабильность структуры фрагментов Fab с двойной специфичностью по сравнению с исходным Герцептином®, проводят эксперименты по тепловой денатурации с помощью дифференциальной сканирующей калориметрии (DSC). Эксперименты по тепловой денатурации проводят на дифференциальном сканирующем калориметре от Microcal Inc. Фрагменты Fab подвергают диализу против 10 мМ раствора ацетата натрия рН 5, 150 мМ раствора хлорида натрия. Концентрацию растворов доводят до 0.5 мг/ил и нагревают до 95°С со скоростью 1°С/мин. Профили плавления корректируют и нормализуют по базовой (нулевой) линии. Температуры плавления (TM) определяют, используя программное обеспечение, поставляемое производителем. Как и предполагалось, ни один из Fab не визуализирует обратимых профилей обратимой тепловой денатурации (Kelley et al., 1992) (данные не показаны). TM вариантов с двойной специфичностью (77.2°С, 75.6°С, 74.3°С для bH1, bH1-81 и bH1-44, соответственно, Таблица 16) немного ниже, чем TM Герцептина® (82.5°С), но является высокой и находится в диапазоне температур, сообщаемых для других терапевтических антител (Garber and Demarest, 2007).
Кинетика и термодинамика связывания bH1 вариантов с высокой аффинностью только к VEGF или к HER2
Интересно отметить, что антитела с двойной специфичностью получают основную часть своей энергии связывания из совершенно различных областей общего сайта связывания VEGF/HER2. Эти данные показывают, что функция VEGF-или HER2-связывания антител с двойной специфичностью можно селективно нарушать, не оказывая влияния на оставшуюся специфичность связывания. Структурные исследования показывают, что структурные паратопы на bH1 для VEGF и HER2 в значительной степени перекрываются, но шотган аланин-сканирующий мутагенез антител bH1 и bH1-44 показывает, что взаимодействия с VEGF и HER2 опосредуются двумя уникальными группами остатков CDR, степень перекрывания которых мала (Фигуры 54 и 57, Таблицы 9А, 9В и 10). Шотган сканирование аланином антител bH1 и bH1-44 показывает, что некоторые остатки CDR являются исключительно важными для связывания либо VEGF, либо HER2 (Фигуры 54 и 57, Таблицы 9А, 9В и 10), включая LC-Ile29, LC-Tyr32, которые важны для VEGF-связывания, и НС-Arg50, НС-Arg58 для HER2-связывания (Фигуры 54 и 57, Таблицы 9 и 10). Для подтверждения уникальной важности боковых цепей этих остатков в каждом взаимодействии, причем каждый остаток заменяется на аланин в скаффолдах антитела bH1-44 (LC-Ile29, LC-Tyr32, HC-Arg50, HC-Arg58) или антитела Герцептин® (НС-Arg50, HC-Arg58), индивидуально или в комбинации, и мутанты экспрессируются в виде Fabs и IgGs.
Векторы, которые кодируют фрагменты Fabs антител bH1-44 Herceptin®, слитые с N-концом генаIII через тяжелую цепь, используют в качестве матриц для мутагенеза по Кункелю (Kunkel et al., 1987). Создают олигонуклеотиды для введения заданных аланиновых остатков (мутаций) в выбранных положениях. Мутанты Fab, содержащие замены на аланин, экспрессируют в виде фага, а связывание проверяют конкурентным ИФА (ELISA) (Фигура 58). Затем вариабельные домены тяжелой цепи и легкой цепи клонируют в векторы экспрессии Fab и IgG, a Fab и IgG экспрессируют и очищают, как описано в (Bostrom et al., 2009). Методом SDS-PAGE проверяют корректный размер белка (Фигура 65). Анализ методом гель(-проникающей) хроматографии показывает уровни агрегации менее 5%.
Связывание с двумя антигенами изучают конкурентным ИФА (ELISA) и/или BIAcore. Все единичные мутации в аланин в скаффолде bH1-44 в различной степени ослабляют связывание (данные не показаны). Наиболее поразительной единичной мутацией является мутация LC-Y32A, которая в значительной степени нарушает VEGF-связывание, при этом сохраняя аффинность и кинетику связывания с HER2 (Таблица 12, Фигура 58 и Фигура 63). Двойные мутации I29A+Y32A (LC) или R50A+R58A (НС) почти полностью нарушают связывание с VEGF или HER2 соответственно, в то же время сохраняя аффинность и кинетику связывания с другим антигеном (Таблица 12, Фигура 58 и Фигура 63). Мутации в аланин HC-R50A, НС-R58A в скаффолде антитела Герцептин® также до некоторой степени нарушают связывание с HER2, тогда как двойной мутант НС R50A+R58A не проявляет какого-либо детектируемого связывания с HER2 (Таблица 12).
Термодинамические параметры двойных мутантов анализируют подробнее и сравнивают с параметрами bH1-44. Свободная энергия связывания мутантов антитела bH1-44, LC-I29A+Y32A и HC-R50A+R58A, с HER2 или с VEGF, соответственно, слагается из благоприятных вкладов энтальпии и энтропии (ΔН=-7.7 и -TΔS=-3.9 для VEGF, ΔН=-6.4 и -TΔS=-7.6 для HER2, Таблица 13, Фигура 60), что примерно эквивалентно значениям для bH1-44, полученным при 30°С (Таблица 13, Фигура 60). Таким образом, термодинамические и кинетические профили двойных мутантов и bH1-44 одинаковы.
Структурные основания для функций остатков, изменяющих специфичность
Далее анализируют кристаллические структуры bH1 в комплексе с VEGF или HER2 (Bostrom et al., 2009), чтобы выявить специфические взаимодействия связывающих детерминант в каждом комплексе с антигеном (Фигура 64). Итоговый анализ объясняет, как мутации двух определяющих специфичность остатков нарушают способность связываться с одним антигеном, не влияя на аффинность, кинетику и термодинамику связывания с другим антигеном. Домен CDR-L1 антитела bH1 содержит основную часть изменений в последовательности антитела Герцептин® и важен для связывания с VEGF. Конформации CDR-L1 антитела bH1 значительно отличаются в структурах двух комплексов; среднее отклонение составляет 4.6 Å (Сα остатков 27-32). Напротив, полная (абсолютная) конформация bH1 Fab в комплексе с VEGF в имеет заметное сходство с полной конформацией Fab, связанного с HER2 (r.m.s.d.=0.7 Å, для 398 скелетных атомов, Сα). Петля CDR-L1 loop составляет 26% от площади поверхности, экранированной (сокрытой) VEGF, в то время как эта петля расположена на периферии HER2 паратопа и минимально участвует в контакте с HER2.
Наложение двух комплексов показывает, что VEGF приходит в столкновение с Tyr32 и прилегающими остатками CDR-L1 в HER2-связанной конформации. Атом Сα главной цепи остатков Tyr32 находится в одинаковом положении в двух структурах, но его боковая цепь повернута на ~130°. В комплексе с VEGF Tyr32 и Ile29, по-видимому, принимают участие в структуре, способствуя тому, что CDR-L1 принимает конформацию, нужную для связывания VEGF. Мутация Tyr32 либо в Ala, либо в Phe недопустима для связывания VEGF (Bostrom et al., 2009). Хотя боковая цепь Tyr32 направлена к HER2, по-видимому, она не участвует в продуктивных контактах. Ile29 удален от HER2, его боковая цепь является гидрофильной, а мутация Ile29 и Tyr32 в Ala вполне допустима для связывания с HER2.

Также изучена структура остатков с уникальной важностью для связывания HER2 в комплексе bH1/HER2. Боковые цепи остатков Arg50 и Arg58 укладываются напротив кислых остатков на HER2 (Glu558 и Asp560) в структуре bH1-HER2 (Фигура 64). Взаимодействия представляются в высшей степени специфическими к боковым цепям, так как мутации в Lys, а также в Ala, являются деструктивными (Bostrom et al., 2009). Однако, в структуре с VEGF остатки Arg50 and Arg58 являются гидрофильными и находятся далеко от VEGF, и мутации в Ala или Lys вполне допустимы (Bostrom et al., 2009).
Ссылочные материалы:
Baselga, J., L. Norton, J. Albanell, et al., 1998, Cancer Res. V.58, p.2825.
Bostrom, J., S. F. Yu, D. Kan, B. A. Appleton, C. V. Lee, K. Billeci, W. Man, F. Peale, S. Ross, C. Wiesmann, and G. Fuh, 2009, Science, v.323, p.1610-4.
Chen Y., C. Wiesmann, G. Fuh, B. Li, H. W. Christinger, P. McKay, A. M. de Vos, and H. B. Lowman, 1999, J Mol Biol, v.293, p.865-81.
Cho, H. S., K. Mason, K. X. Ramyar, A. M. Stanley, S. B. Gabelli, D. W. Denney, Jr., and D. J. Leahy, 2003, Nature, v.421, p.756-60.
Chothia, C., A. M. Lesk, 1987, J. Mol. Biol., v.196, p.901.
Christinger, H. W., Y. A. Muller, L. T. Berleau, B. A. Keyt, B. C. Cunningham, N. Ferrara, and A. M. de Vos, 1996, Proteins, v.26, p.353-7.
Collaborative Computational Project, N., 1994: Acta Crystallogr. Section D. Biol. Crystallogr., v.50, p.760-763.
Dall'Acqua, W., E. R. Goldman, E. Eisenstein, et al., 1996, Biochemistry, v.35, p.1967.
Emsley, P., and K. Cowtan, 2004, Acta Crystallogr D Biol Crystallogr, v.60, p.2126-32.
Fellouse. F. A., B. Li, D. M. Compaan, A. A. Peden, S. G. Hymowitz, and S. S. Sidhu, 2005, J Mol Biol, v.348, p.1153-62.
Fields, B. A., F. A. Goldbaum, X. Ysern, et al., 1995, Nature, v.374, p.739.
Franklin, M. C., K. D. Carey, F. F. Vajdos, D. J. Leahy, A. M. de Vos, and M. X. Sliwkowski, 2004, Cancer Cell, v.5, p.317-28.
Fuh, G., B. Li, C. Crowley, B. Cunningham, and J. A. Wells, 1998, J Biol Chem, v.273, p.11197-204.
Fuh, G., P. Wu, W. C. Liang, M. Ultsch, C. V. Lee, B. Moffat, and C. Wiesmann, 2006, J Biol Chem, v.281, p.6625-31.
Gallop, M. A., R. W. Barrett, W, J. Dower, et al., 1994, Journal of Medicinal Chemistry, V.37, p.1233.
Garber, E., and S. J. Demarest, 2007, Biochem Biophys Res Commun, v.355, p.751-7.
Hudziak, R. M., and A. Ullrich, 1991, J Biol Chem, v.266, p.24109-15.
James, L. C., Roversi, P., Tawfik, D. S., 2003, Science, v.299, p.1362.
Jimenez, R., G. Salazar, K. K. Baldridge, et al., 2003, Proc. Natl. Acad. Sci. USA, v.100, p.92.
Johnson, G., T. T. Wu, 2000, Nucleic Acids Res., v.28, p.214.
Kauzmann, W., 1959, Adv Protein Chem, v.14, p.1-63.
Kelley, R. F., and M. P. O'Connell, 1993, v.32, p.6828-35.
Kelley, R. F., M. P. O'Connell, P. Carter, L. Presta, C. Eigenbrot, M. Covarrubias, B. Snedecor, J. H. Bourell, and D. Vetterlein, 1992, Biochemistry, v.31, p.5434-41.
Kunkel, T. A., J. D. Roberts, and R. A. Zakour, 1987, Methods Enzymol, v.154, p.367-82.
L. C. Storoni, A. J. M. a. R. J. R., 2004, Acta Cryst., p.432-438.
Lasky, L, A., and D. J. Dowbenko, 1984, DNA, v.3, p.23-9.
Lawrence, M.C., Colman, P.M. et al., 1993, J Mol Biol, v.234, pp.946-950.
Lee, C. V., W. C. Liang, M. S. Dennis, C. Eigenbrot, S. S. Sidhu, and G. Fuh, 2004a, J Mol Biol, v.340, p.1073-93.
Lee, C. V., S. S. Sidhu, and G. Fuh, 2004b, J Immunol Methods, v.284, p.119-32.
Lee, С. V., S. G. Hymowitz, H.J. Wallweber, et al. Liang, W. С., X. Wu, F. V. Peale, C. V. Lee, Y. G. Meng, J. Gutierrez, L. Fu, A. K. Malik, H. P. Gerber, N. Ferrara. and G. Fuh, 2006, J Biol Chem, v.281, p.951-61.
Lovell, S. C., I. W. Davis, W. B. Arendall, et al., 2003, Proteins, v.50, p.437.
Lowman, H. В., S. H. Bass, N. Simpson, and J. A. Wells, 1991, Biochemistry, v.30, p.10832-8.
McCoy, Y. A. J., R. J. Read, and L. C. Storoni, 2004, Acta Cryst, 432.
Muller, Y. A., Y. Chen, H. W. Christinger, B. Li, B. C. Cunningham, H. B. Lowman, and A. M. de Vos, 1998, Structure, v.6, p.1153-67.
Muller, В., H. Li, W. Christinger, et al., 1997, Proc. Natl. Acad. Sci USA, v.94, p.7192.
Murphy, K. P., D. Xie, K. S. Thompson, L. M. Amzel, and E. Freire, 1994, Proteins, v.18, p.63-7.
Mylvaganam, S. E., Y. Paterson, and E. D. Getzoff, 1998, J. Mol. Biol., v.281, p.301.
Nemazee, D., 2006, Nat. Rev. Immunol., v.6, p.728.
Otwinowski, Z., and Minor, W., 1997, Methods Enzymol., v.276, p.307-326.
Pauling, L., 1940, J. Am. Chem. Soc., v.62, p.2643.
Presta, L. G., H. Chen, S. J. O'Connor, V. Chisholm, Y. G. Meng, L. Krummen, M. Winkler, and N. Ferrara, 1997, Cancer Res, v.57, p.4593-9.
Read, R. J., 2001, Acta Cryst., v. D57, p.1373-1382.
Reichert, J. M., Rosenzweig, C. J., Faden, L. В., et al., 2005, Nat. Biotechnol., v.23, p.1703.
Senn, B. M., Lopez-Macias, C., Kalinke, U., et al., 2006, Eur. J. Immunol., v.33, p.950.
Sethi, D. K., Agarwal, A., Manivel, V., et al., 2006, Immunity, v.24, p.429.
Sidhu, S. S., B. Li, Y. Chen, F. A. Fellouse, C. Eigenbrot, and G. Fuh, 2004, J Mol Biol, v.338, p.299-310.
Sidhu, S. S., H. B. Lowman, B. C. Cunningham, and J. A. Wells, 2000, Methods Enzymol, v.328, p.333-63.
Starovasnik, M. A., M. P. O'Connell, W. J. Fairbrother, and R. F. Kelley, 1999, Protein Sci, v.8, p.1423-31.
Vajdos, F. F., C. W. Adams, T. N. Breece, L. G. Presta, A. M. de Vos, and S. S. Sidhu, 2002, J Mol Biol, v.320, p.415-28.
Weiss, G. A., C. K. Watanabe, A. Zhong, et al., 2000, Proc. Natl. Acad. Sci. USA, v.97, p.8950.
Wiesmann, C., G. Fuh, H. W. Christinger, C. Eigenbrot, J. A. Wells, and A. M. de Vos, 1997, Cell, v.91, p.695-704.
Winn, M. D., M. N. Isupov, and G. N. Murshudov, 2001, Acta Crystallogr. D. Biol. Crystallogr., v.57, p.122.
Все патенты, патентные заявки, опубликованные патентные заявки и другие публикации, которые цитируются или приводятся в качестве ссылок в данном описании, вводятся ссылкой в той же степени как если бы конкретно и индивидуально указывалось, что каждый независимый патент, каждая независимая патентная заявка, опубликованная патентная заявка или публикация вводится ссылкой в данное описание.
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛА ПРОТИВ ФАКТОРА РОСТА ЭНДОТЕЛИЯ СОСУДОВ (VEGF) | 2013 |
|
RU2639506C2 |
| АНТИТЕЛА ПРОТИВ ФАКТОРА РОСТА ЭНДОТЕЛИЯ СОСУДОВ (VEGF) | 2008 |
|
RU2514148C2 |
| АНТИТЕЛА ПРОТИВ BV8 И ИХ ПРИМЕНЕНИЕ | 2010 |
|
RU2559542C2 |
| АНТИТЕЛА ПРОТИВ ПОЛИУБИКВИТИНА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2011 |
|
RU2603093C2 |
| СИСТЕМА ЭКСПРЕССИИ И СЕКРЕЦИИ | 2013 |
|
RU2670491C2 |
| АНТИ-ПОЛИУБИКВИТИНОВЫЕ АНТИТЕЛА И СПОСОБЫ ПРИМЕНЕНИЯ | 2012 |
|
RU2630637C2 |
| АНТИТЕЛА ПРОТИВ DLL4 И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2007 |
|
RU2415869C2 |
| ОПТИМИЗИРОВАННЫЕ ВАРИАНТЫ АНТИ-VEGF АНТИТЕЛ | 2016 |
|
RU2763916C2 |
| АНТИТЕЛА ПРОТИВ AXL И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2011 |
|
RU2577986C2 |
| АНТИТЕЛА ПРОТИВ NOTCH2 И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2009 |
|
RU2580029C2 |

























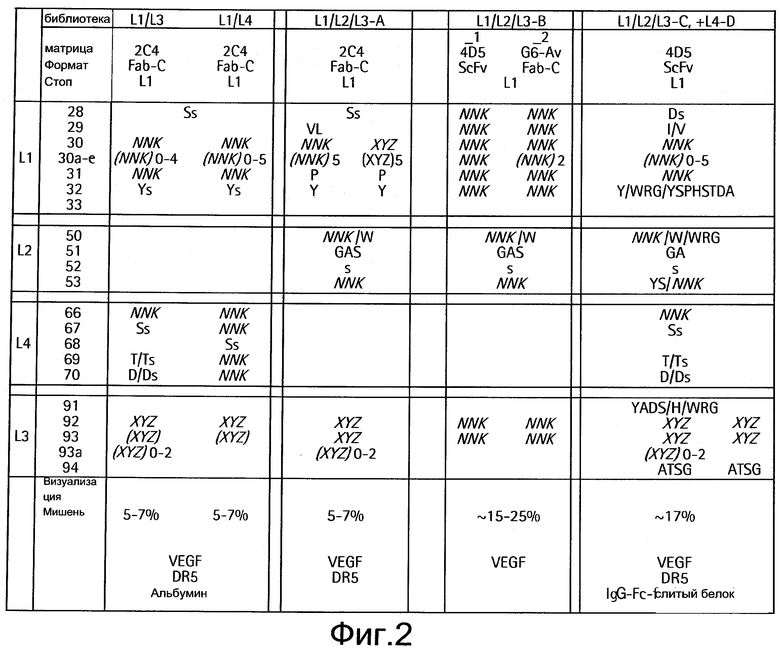





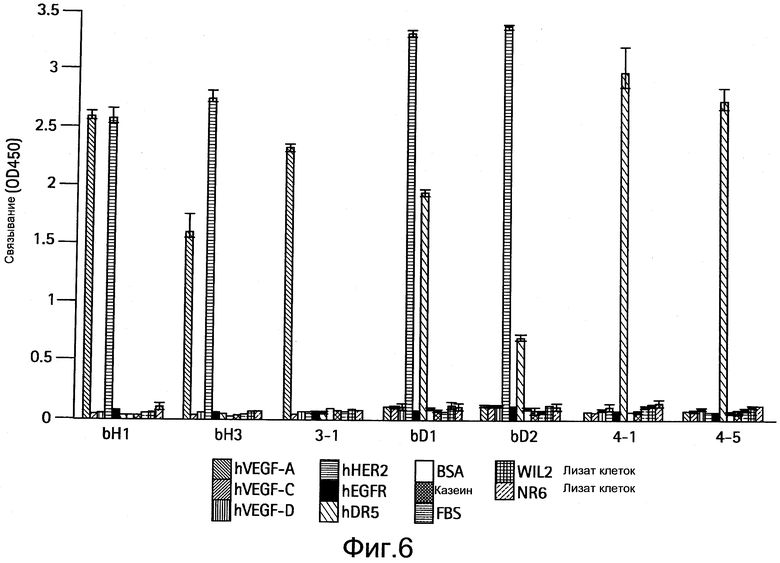
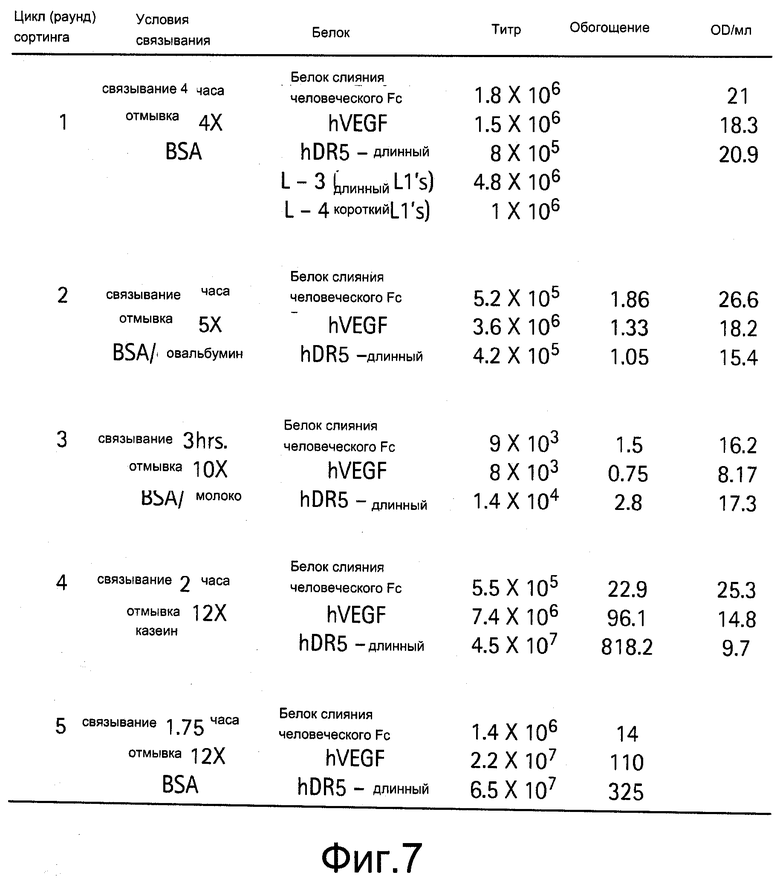

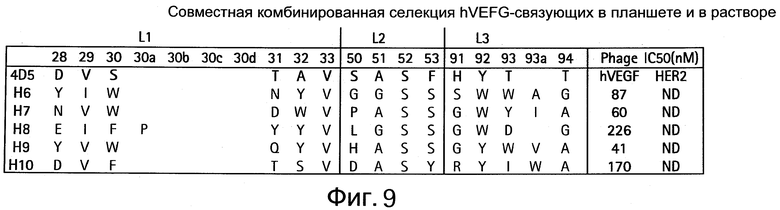
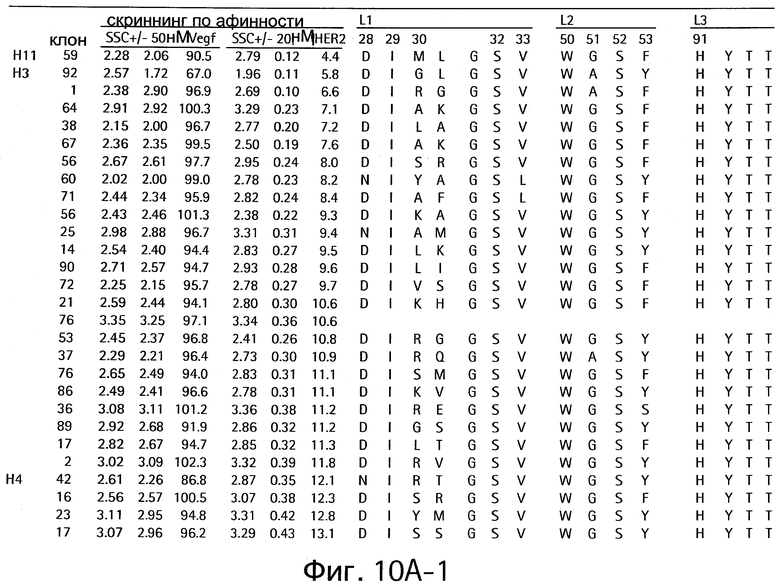
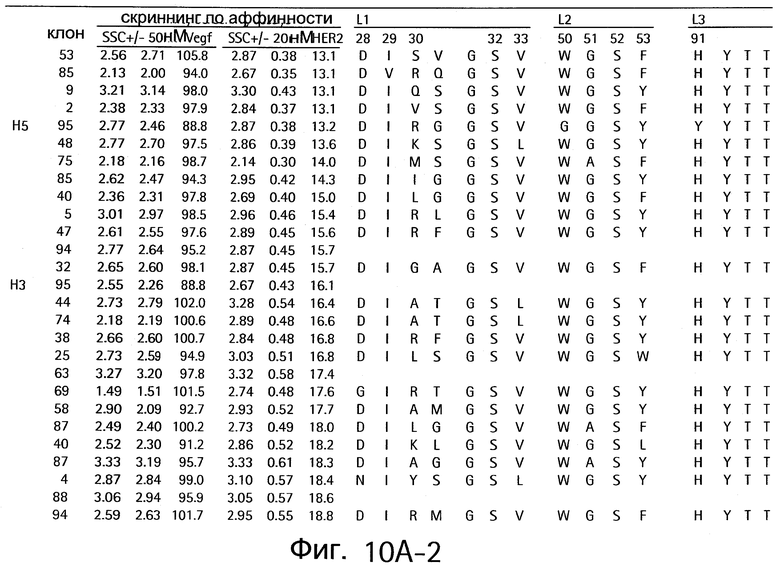
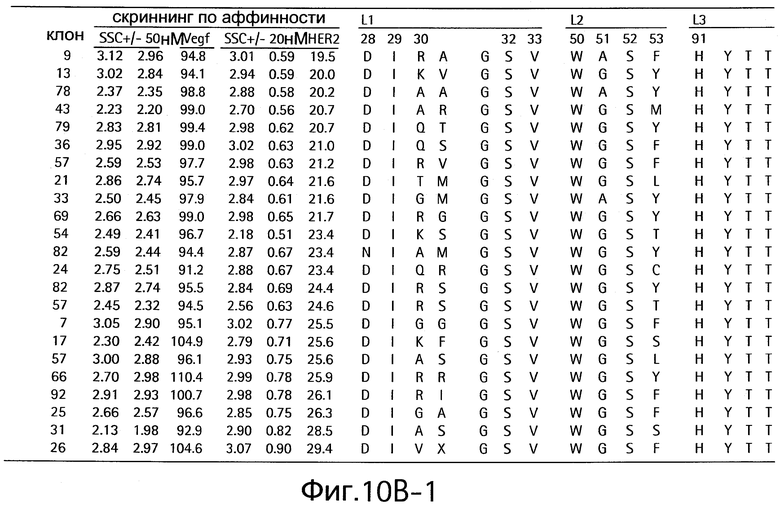
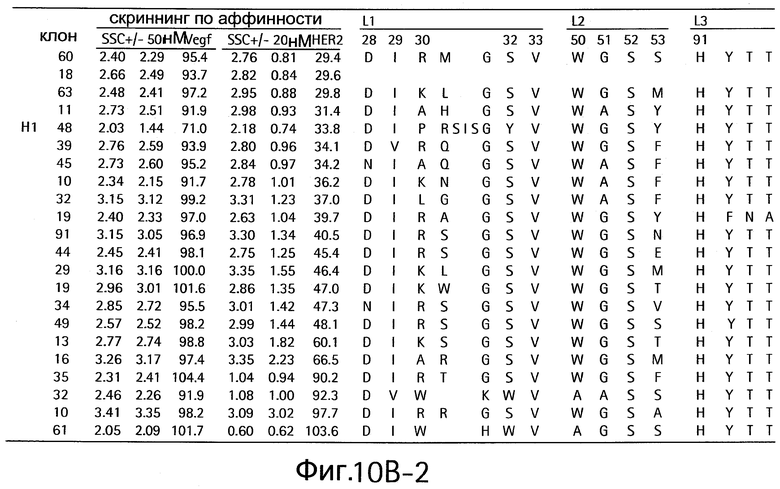
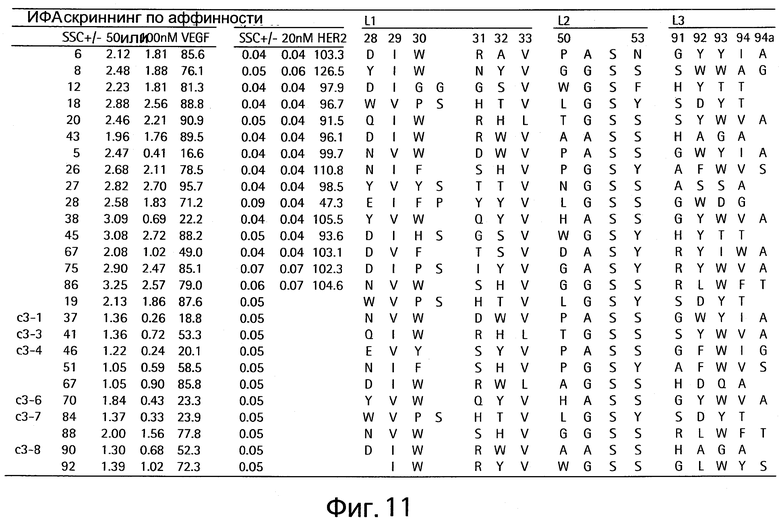


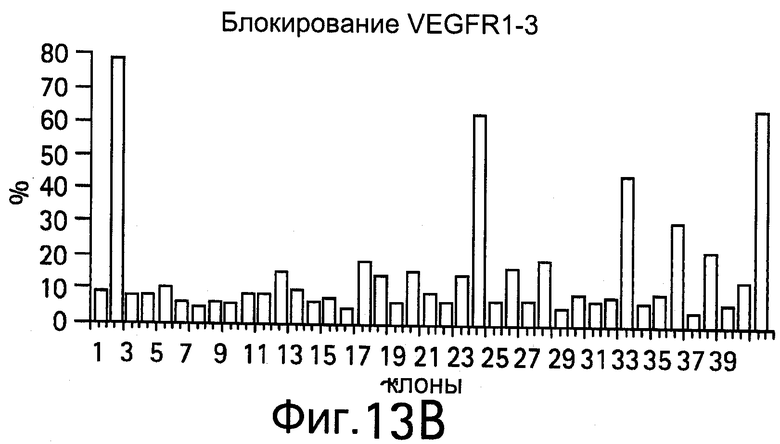
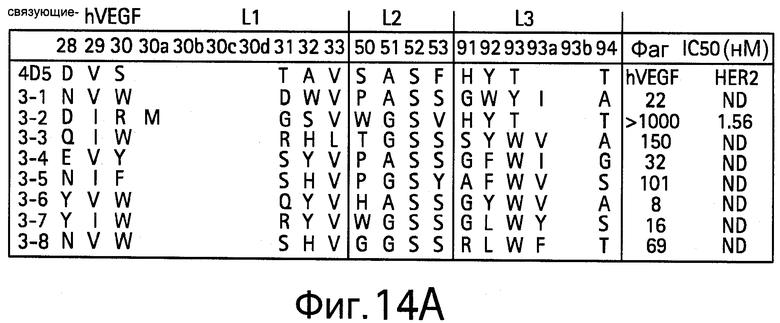



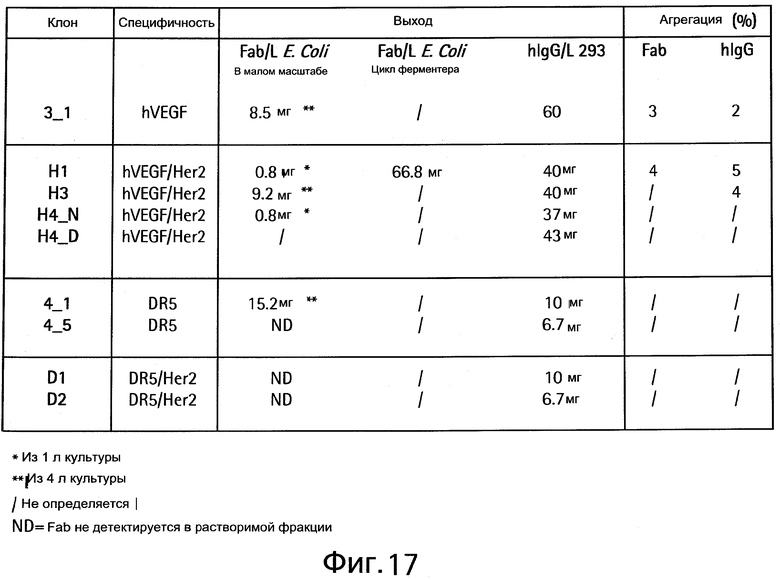

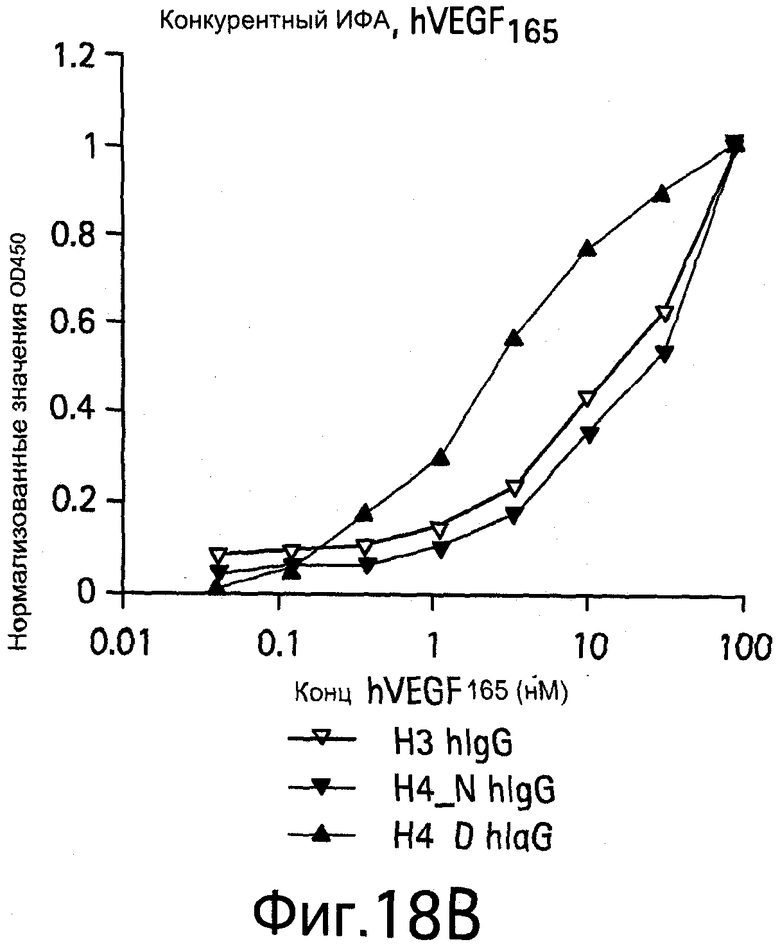
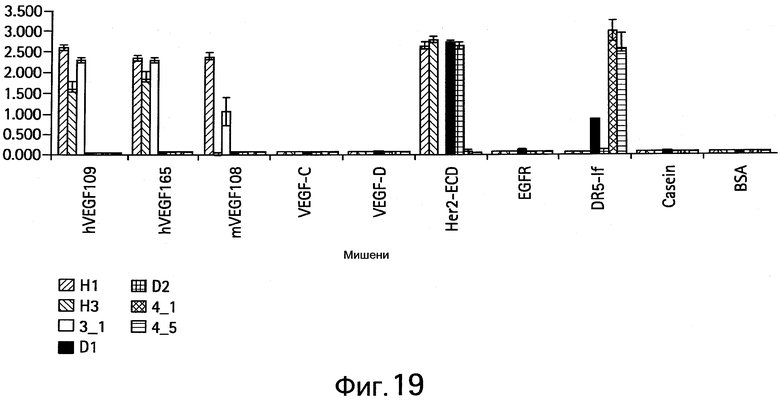
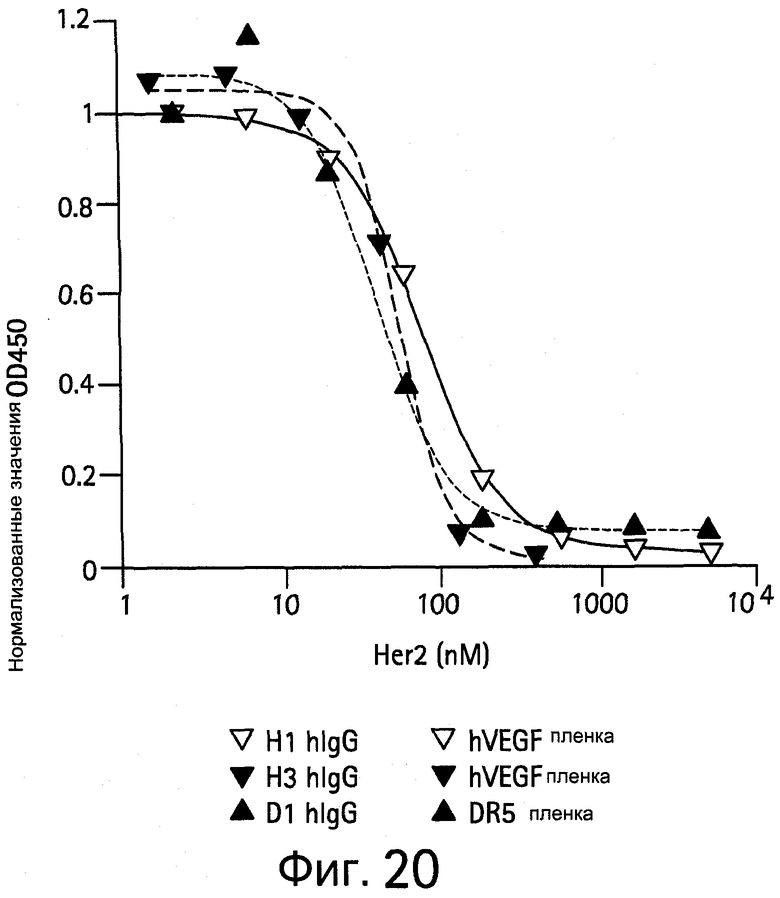
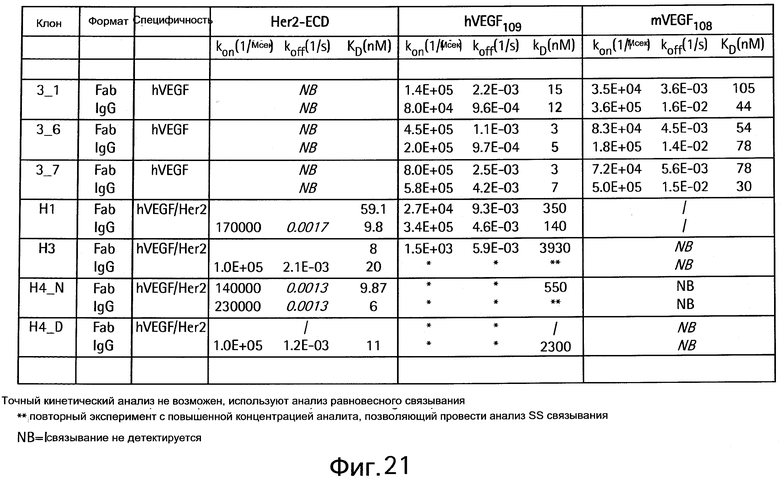
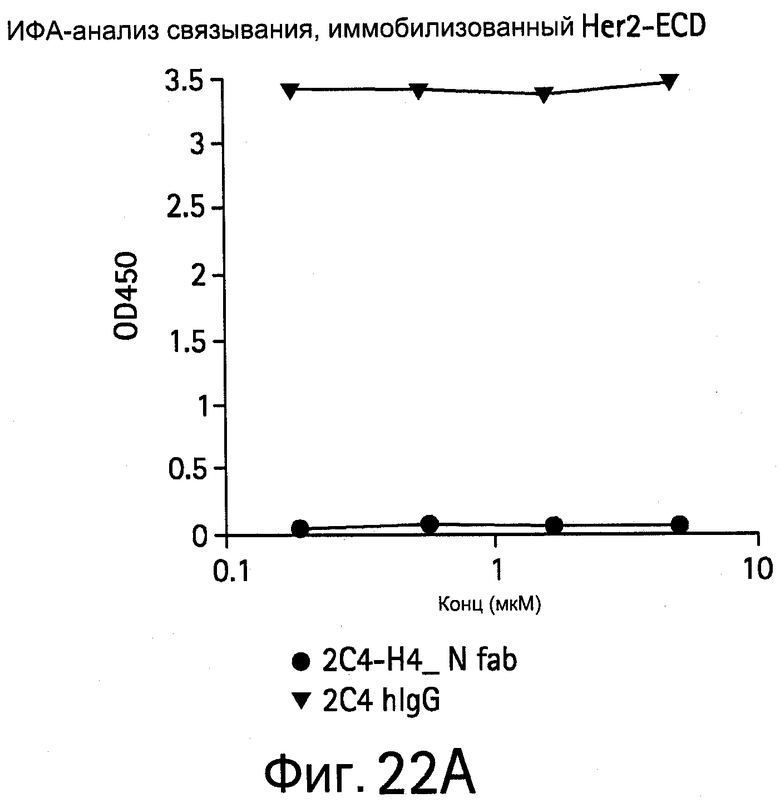

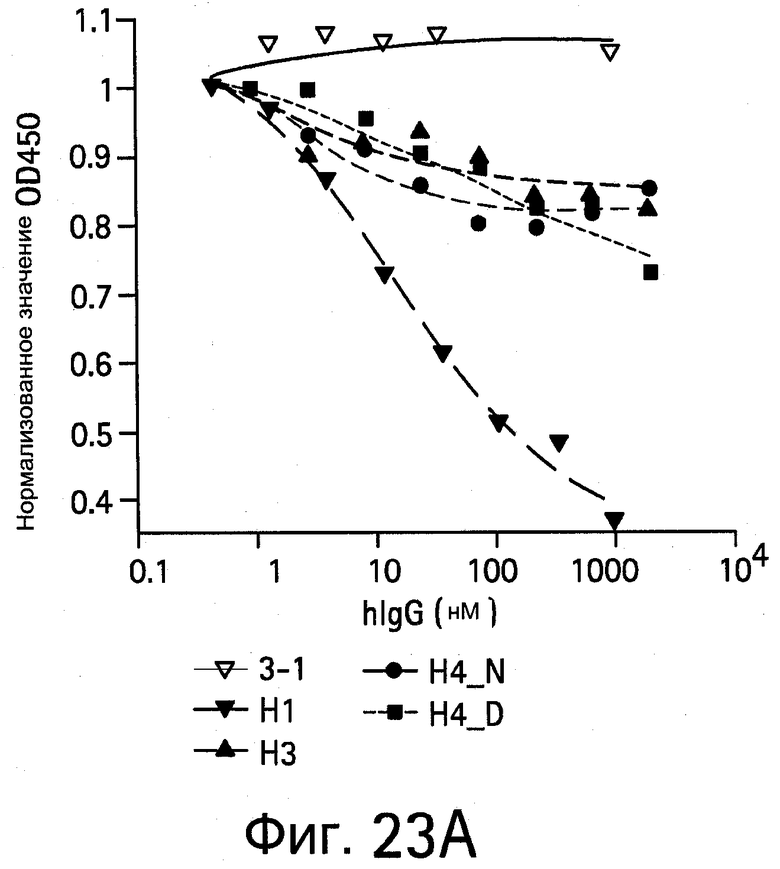

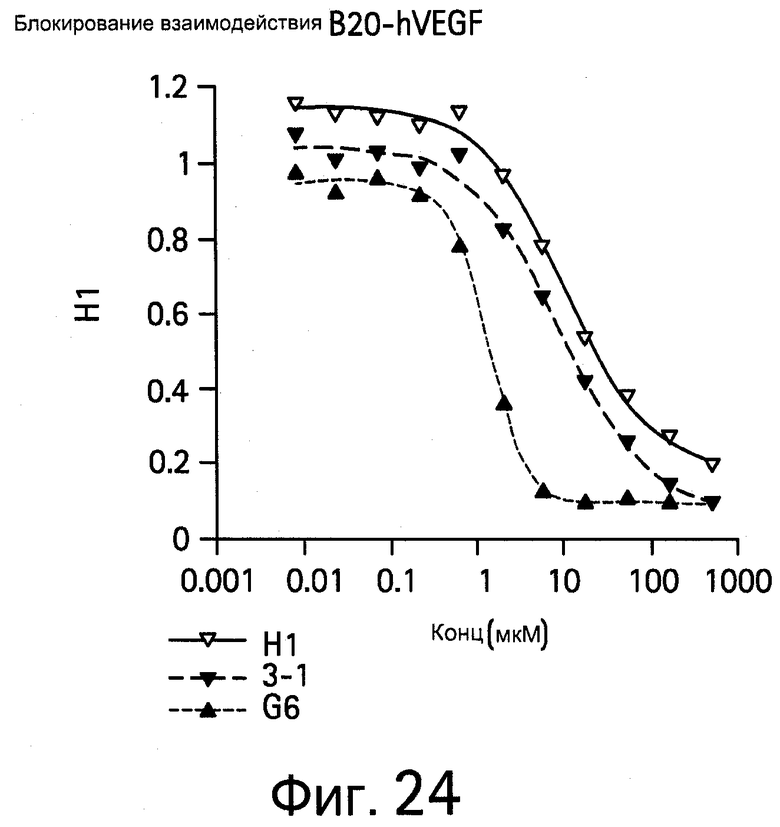

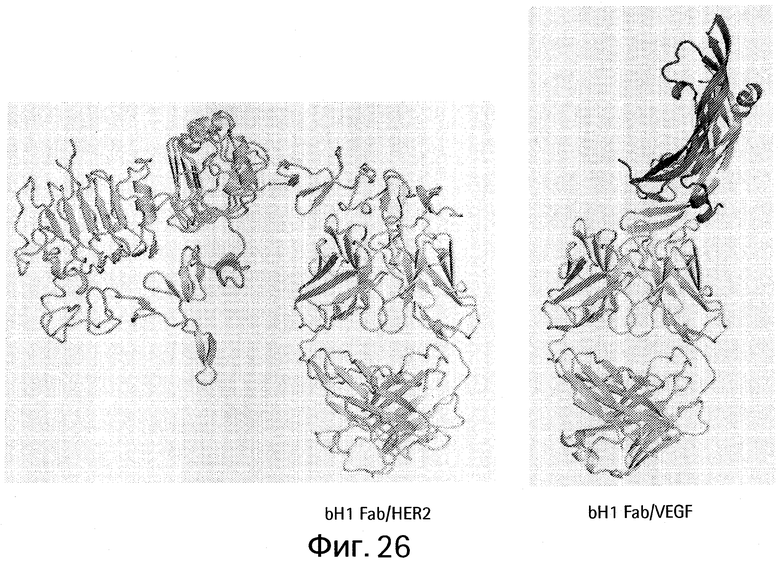
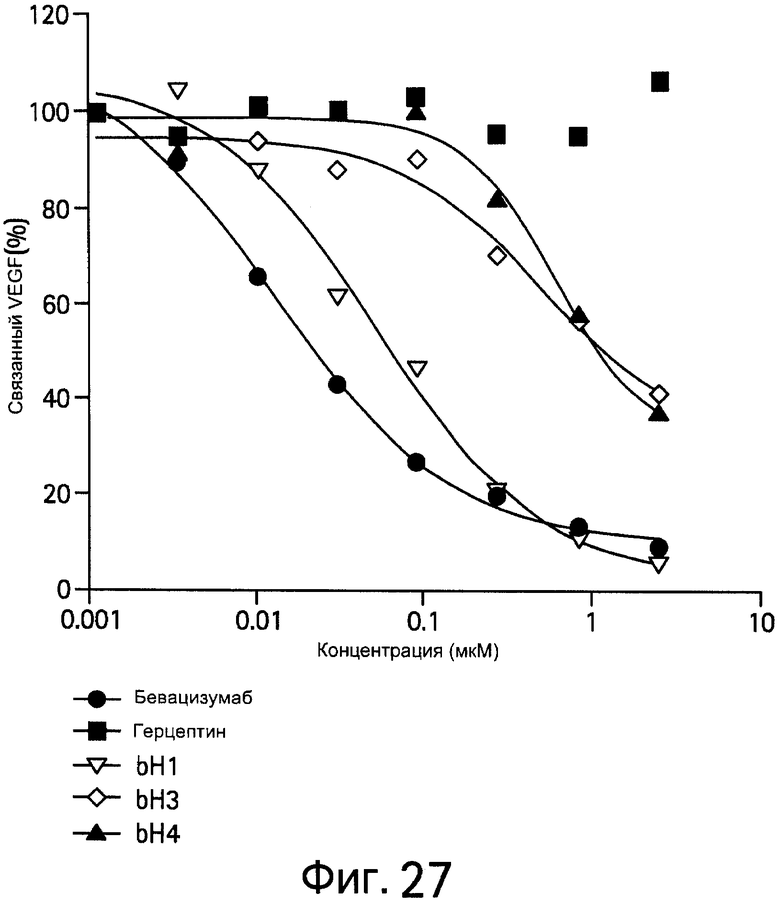
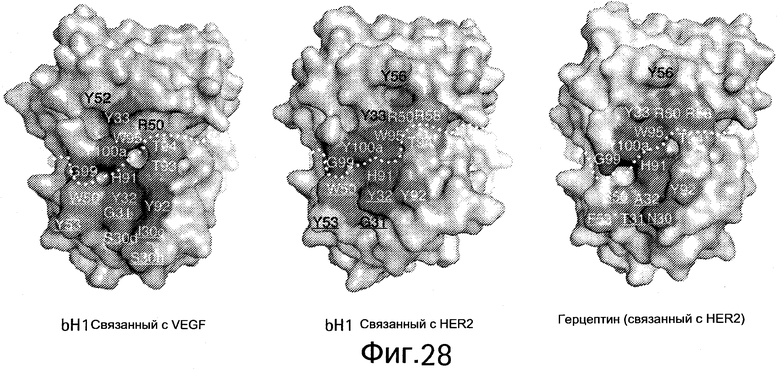
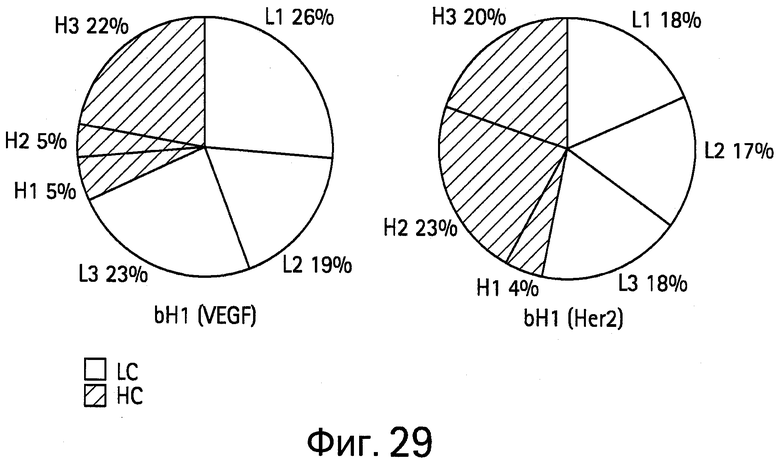
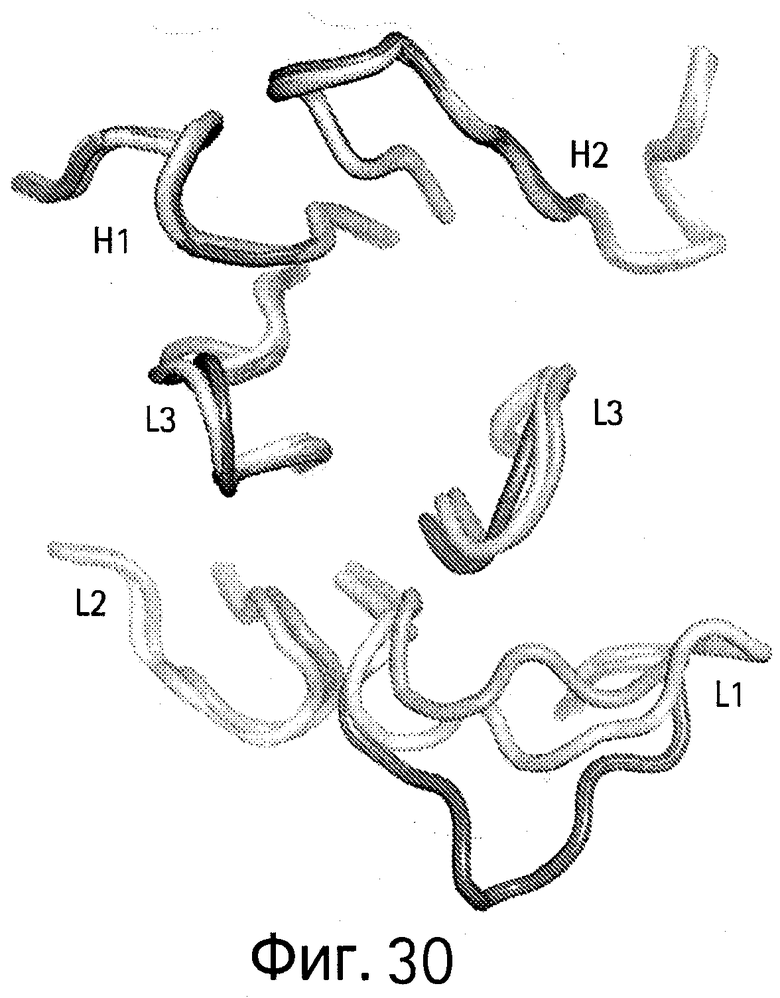


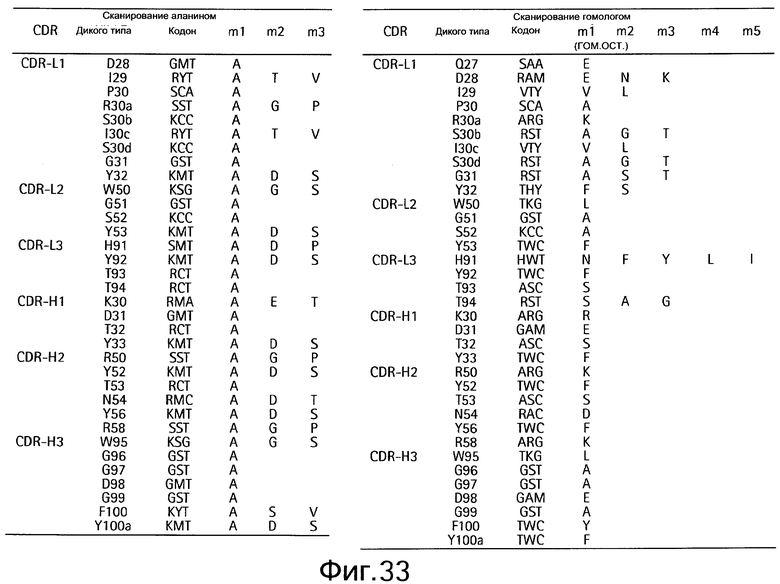

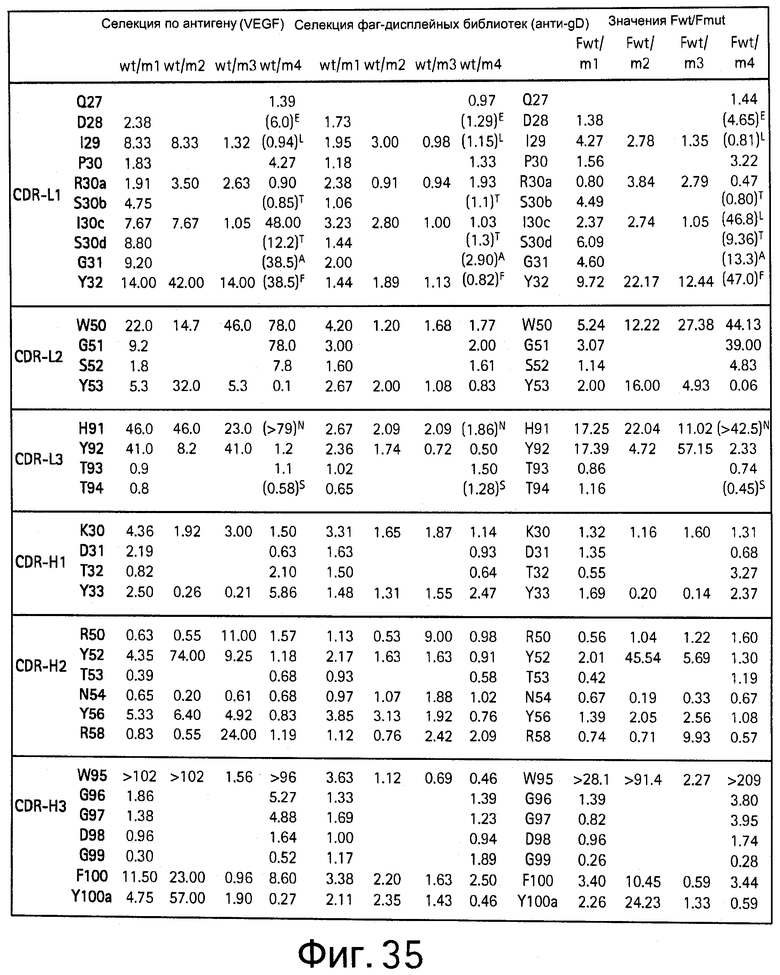


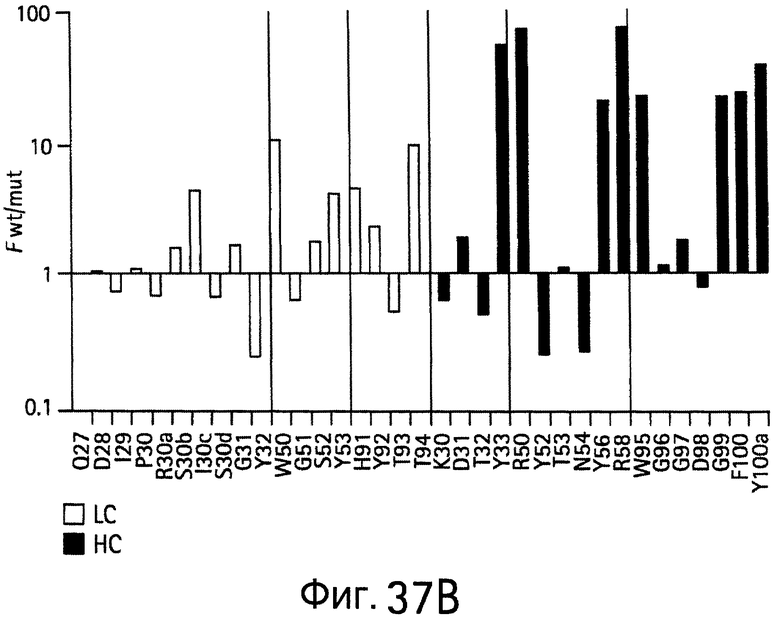
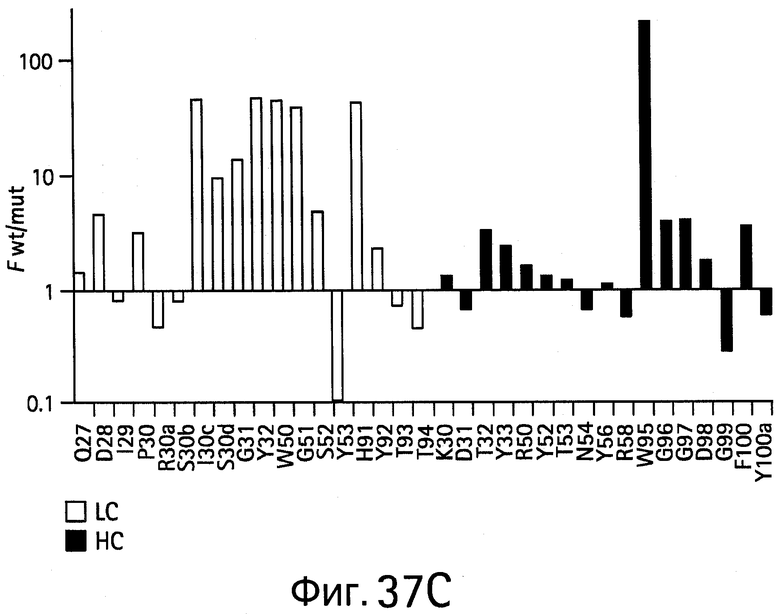
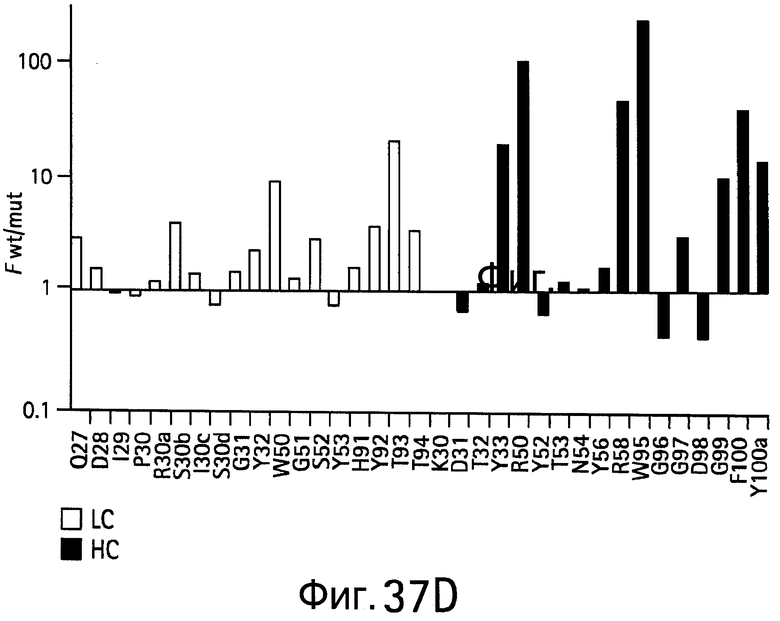

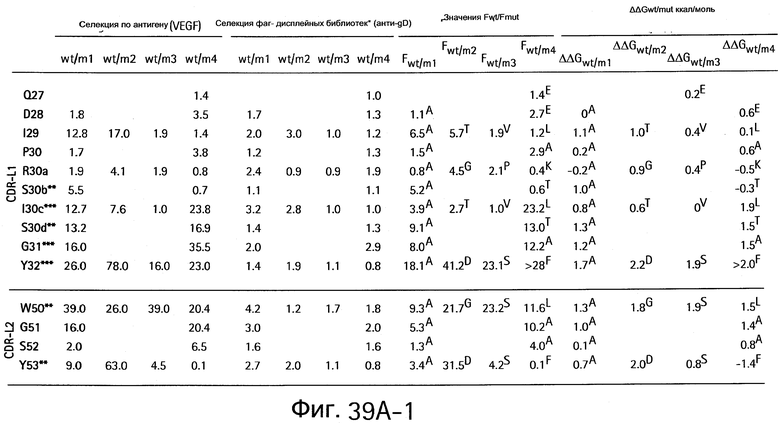
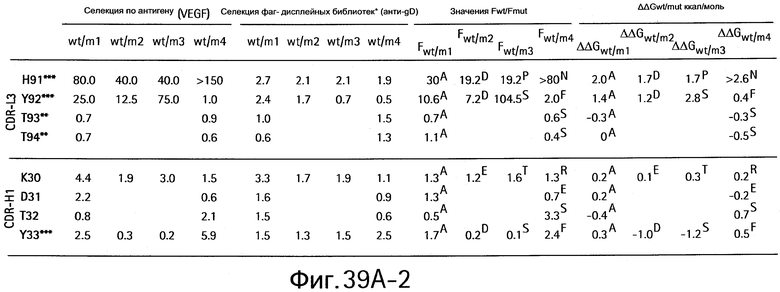
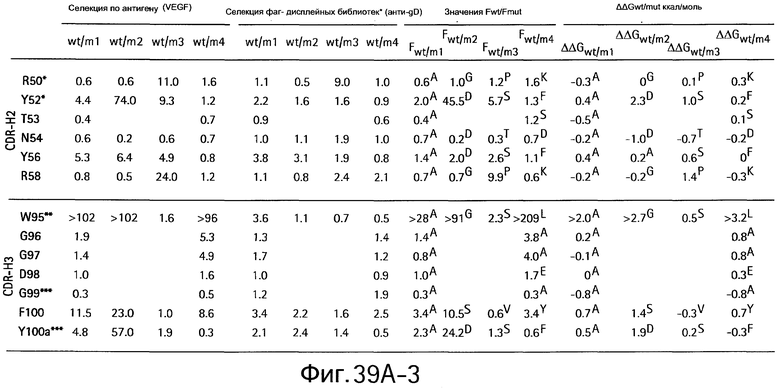
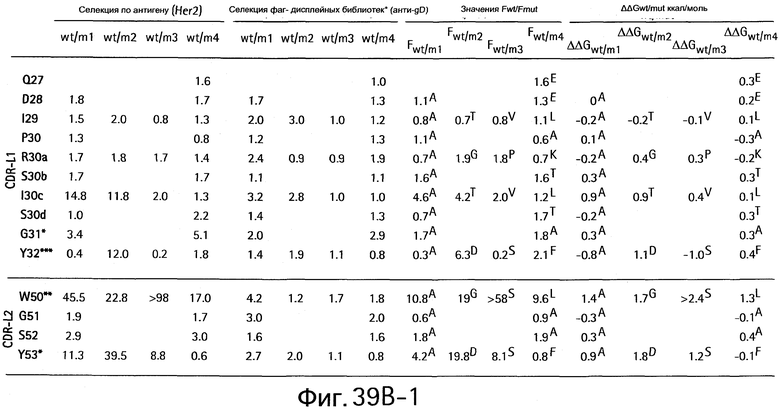
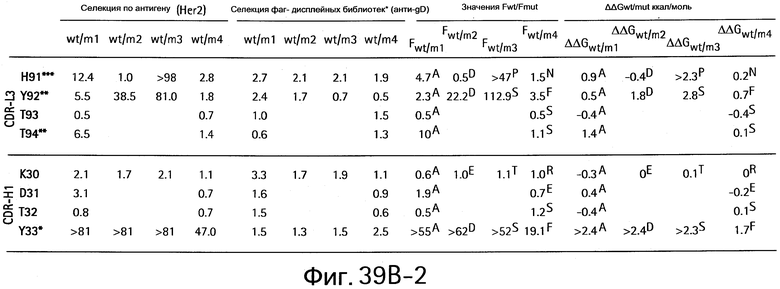

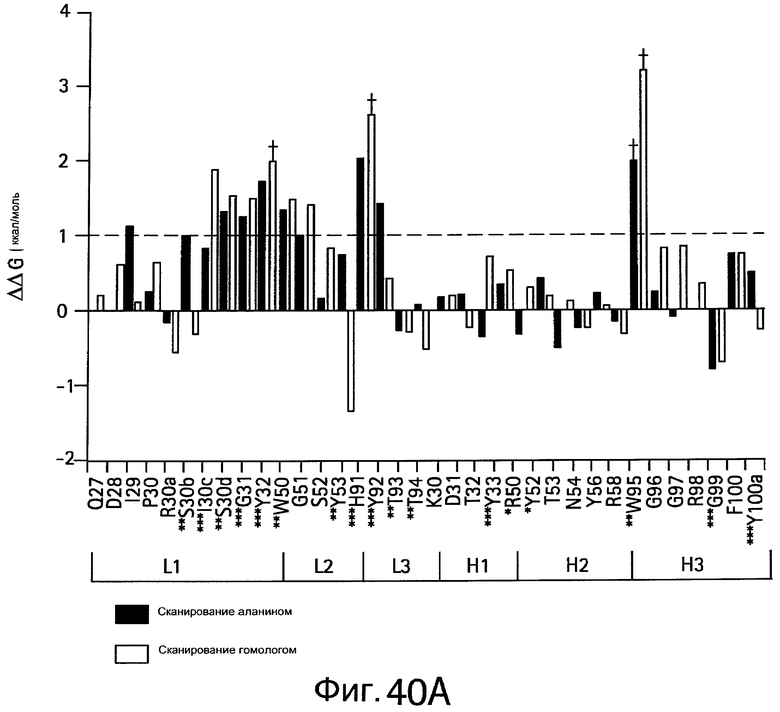

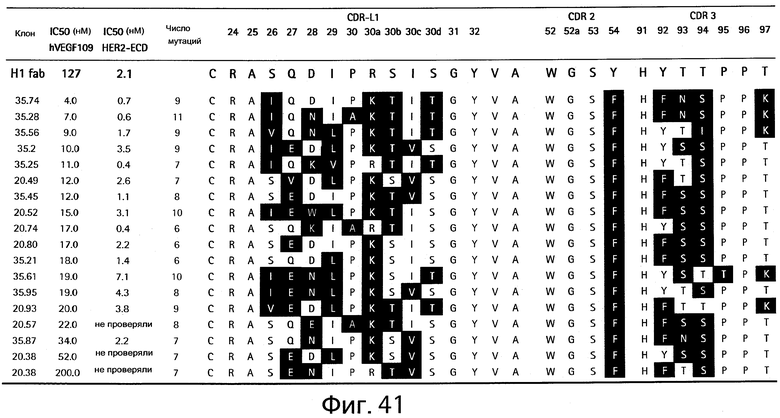
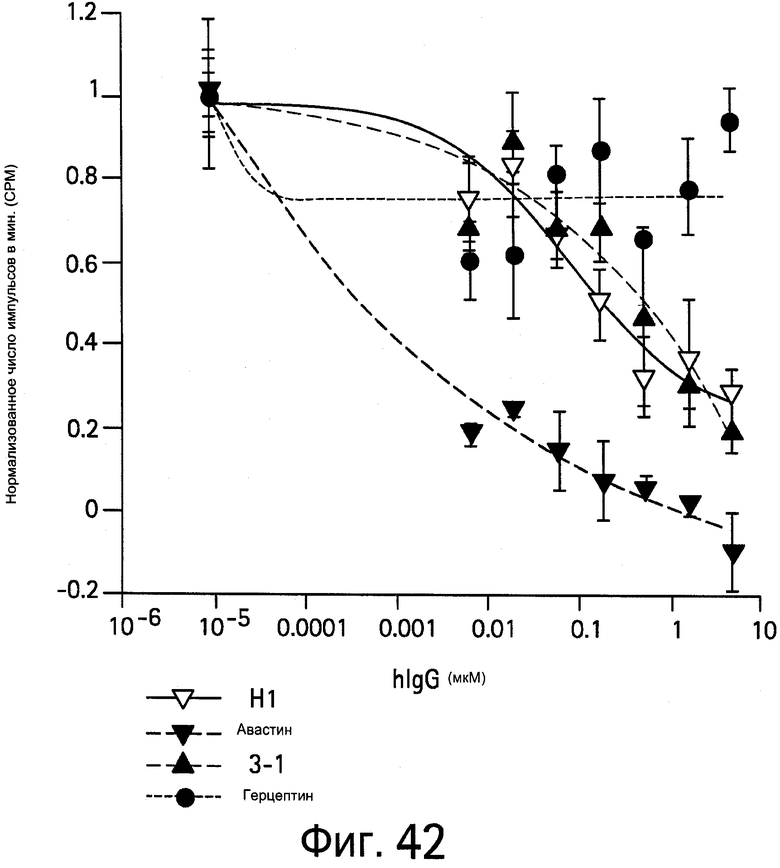
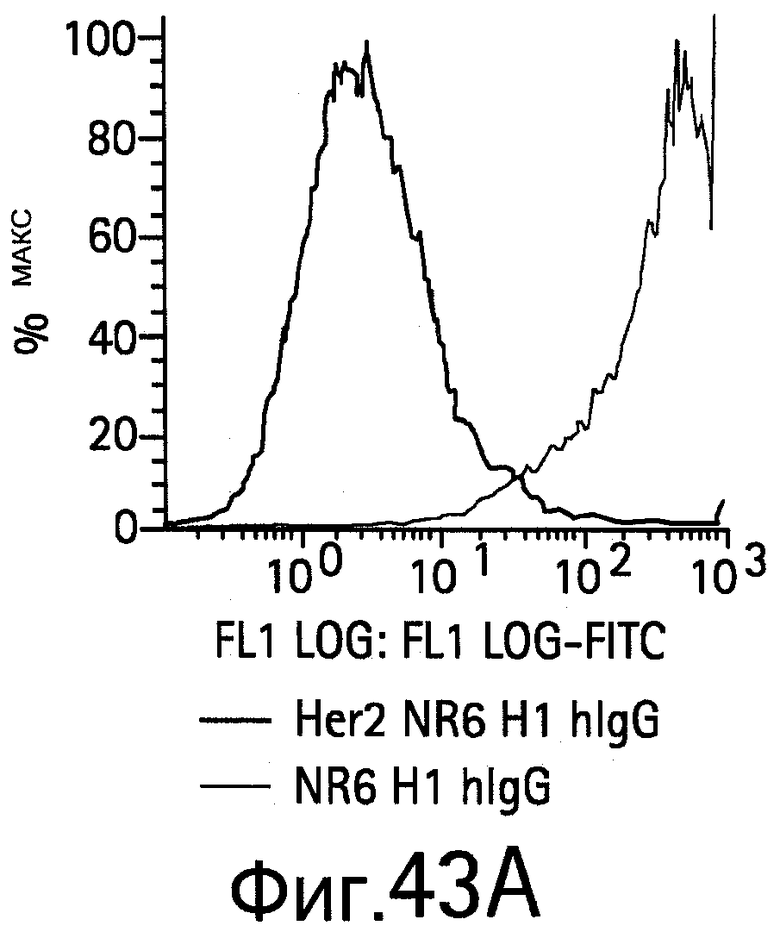
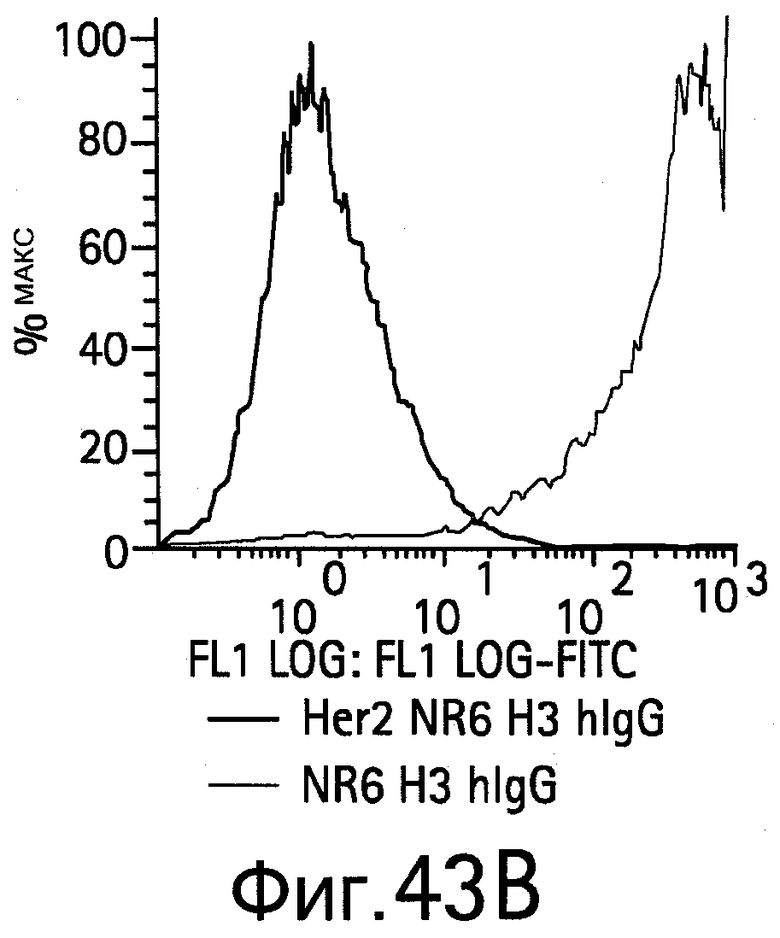
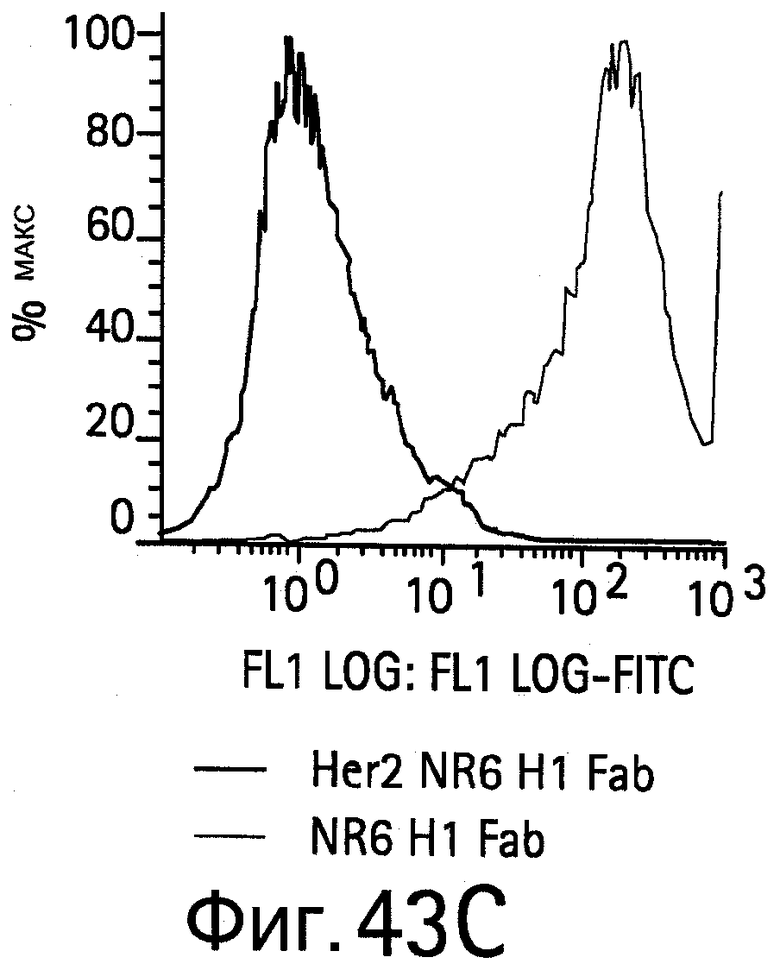
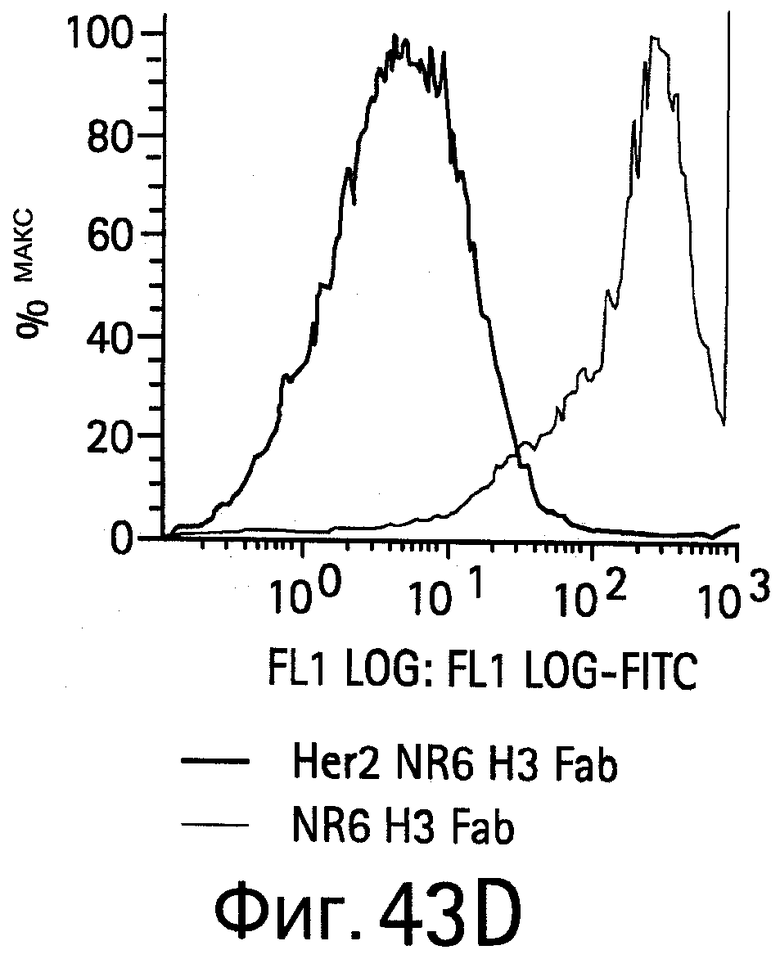

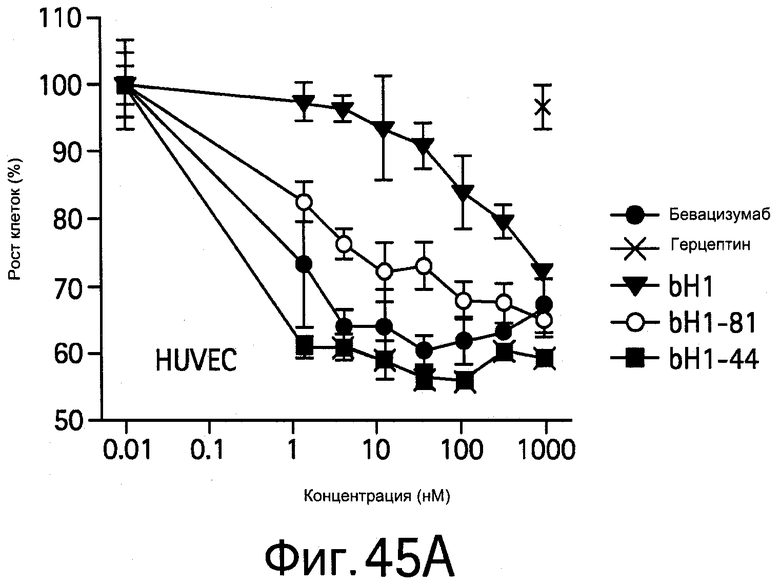
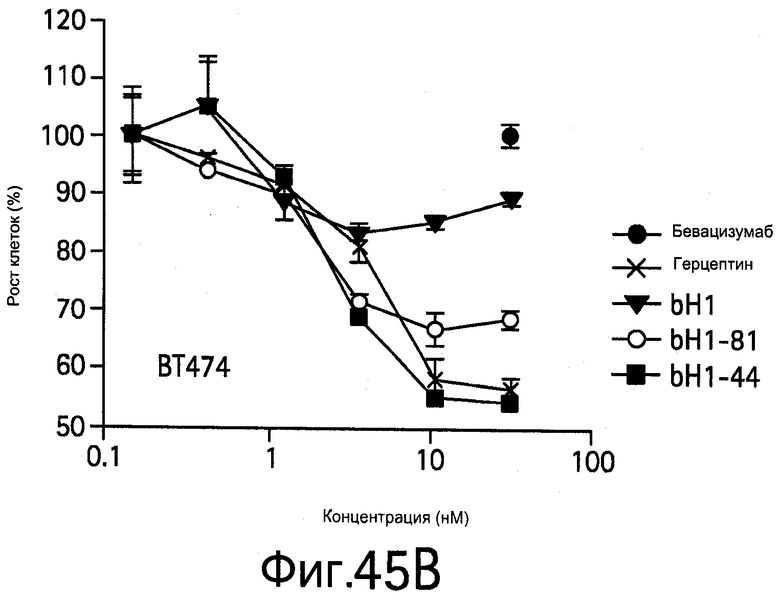

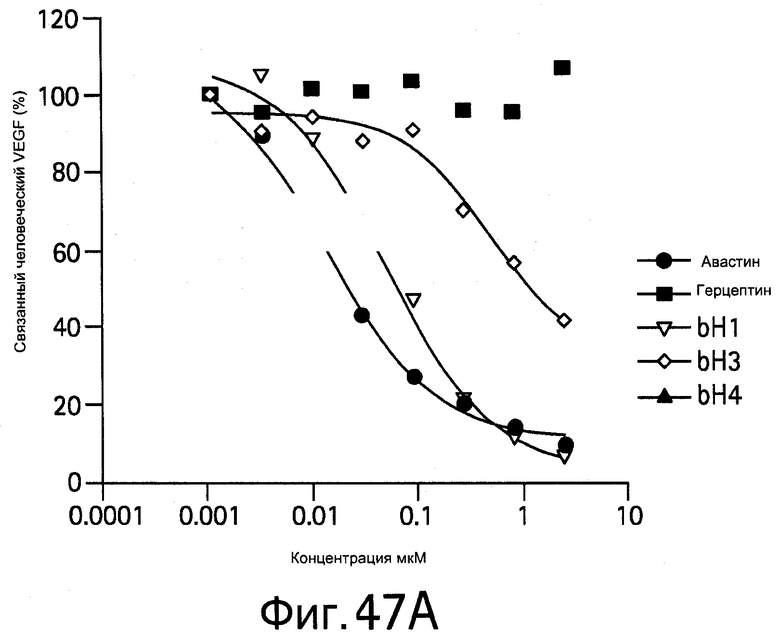



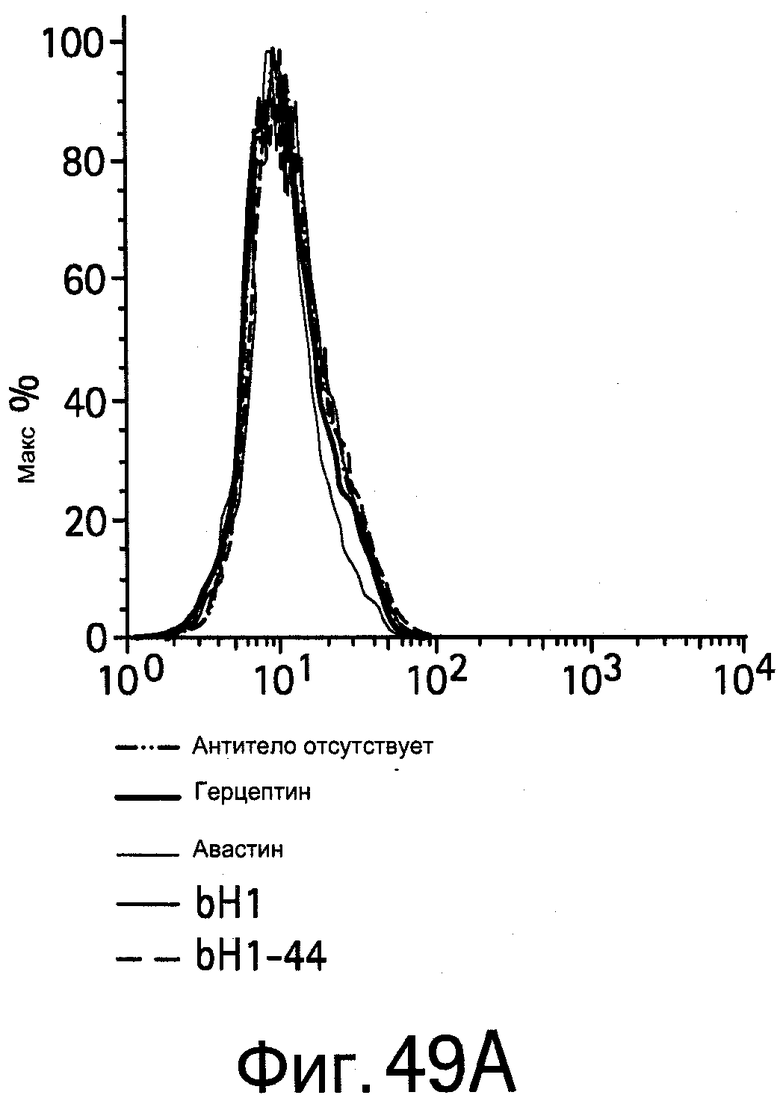

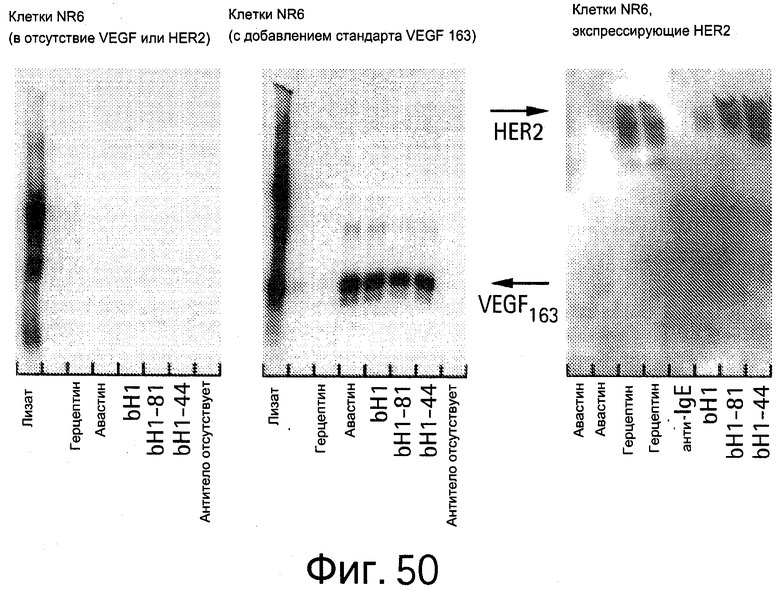
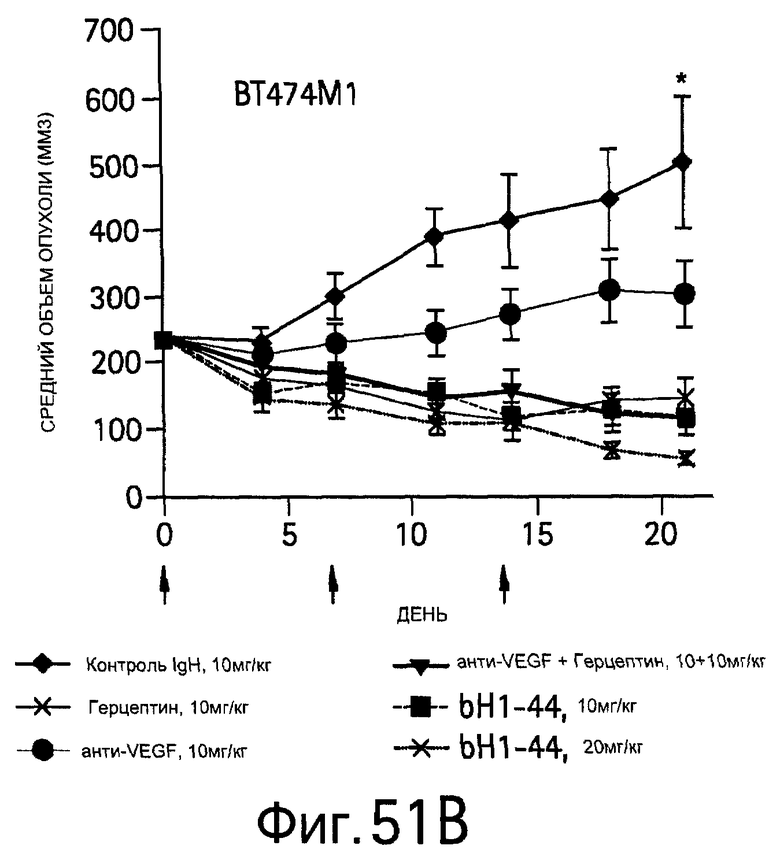
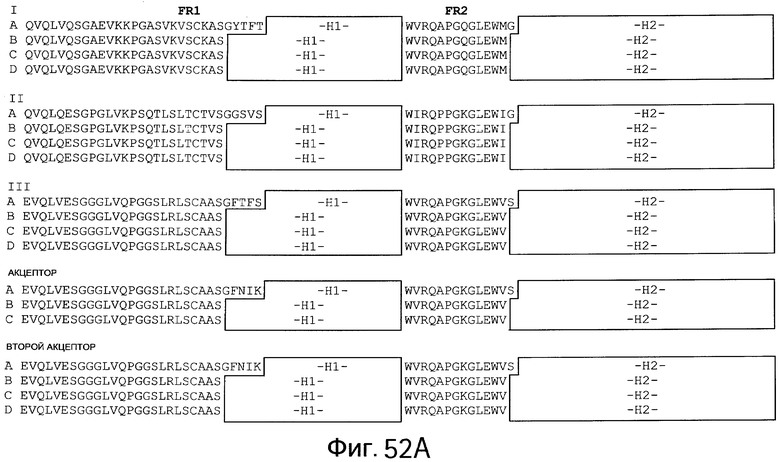
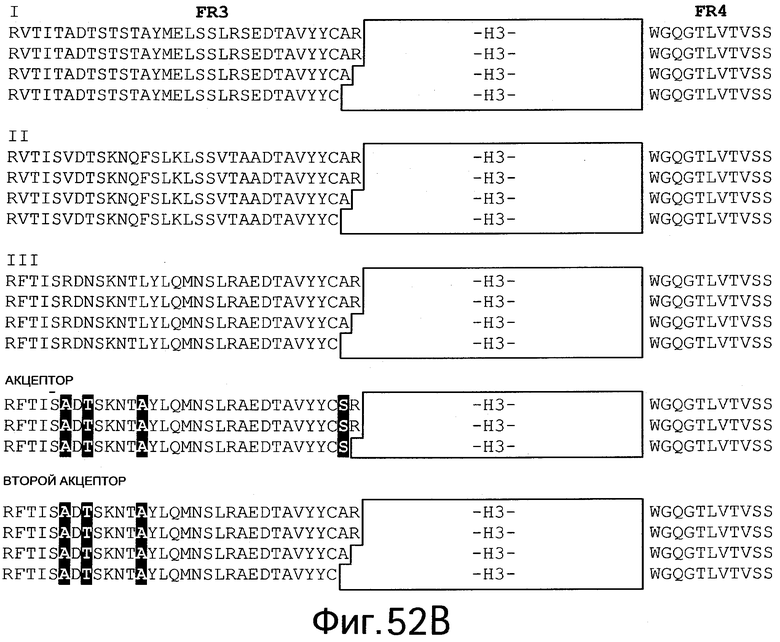

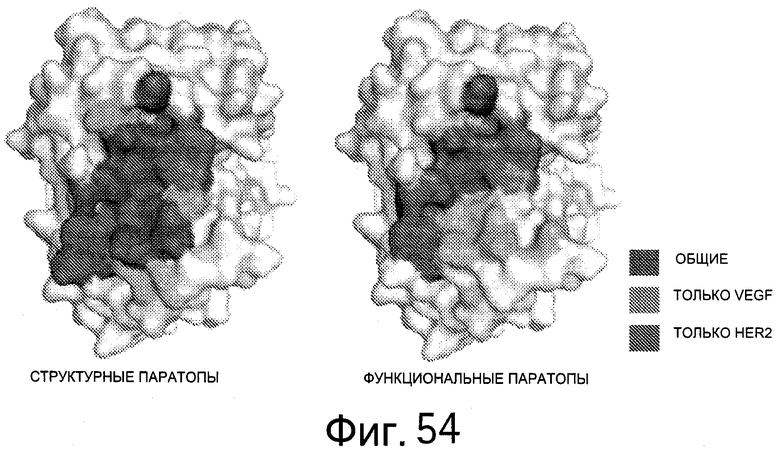
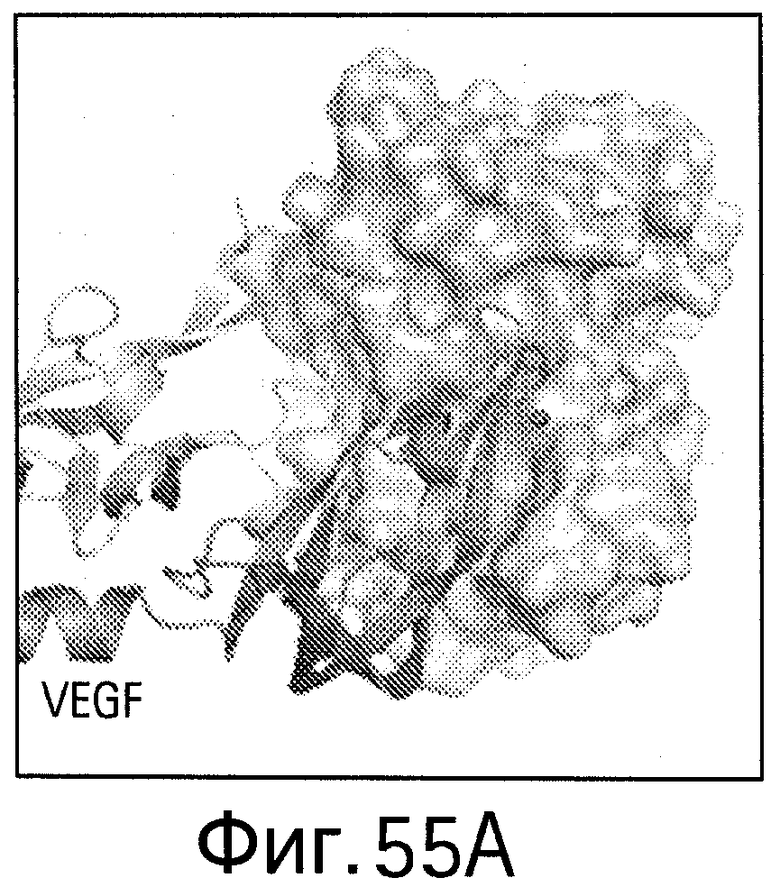
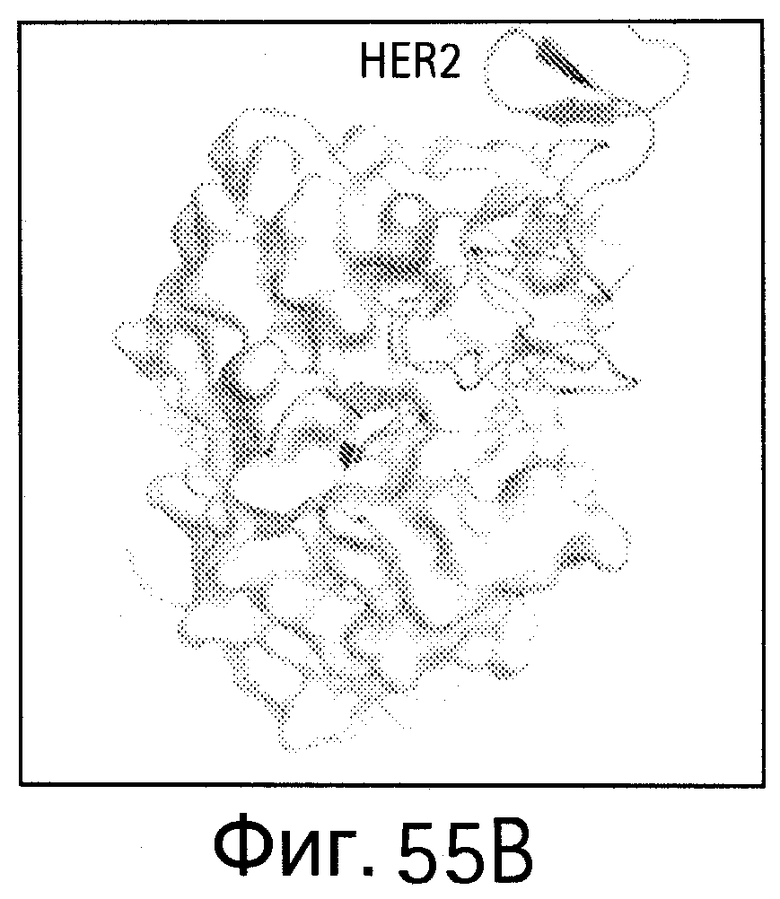
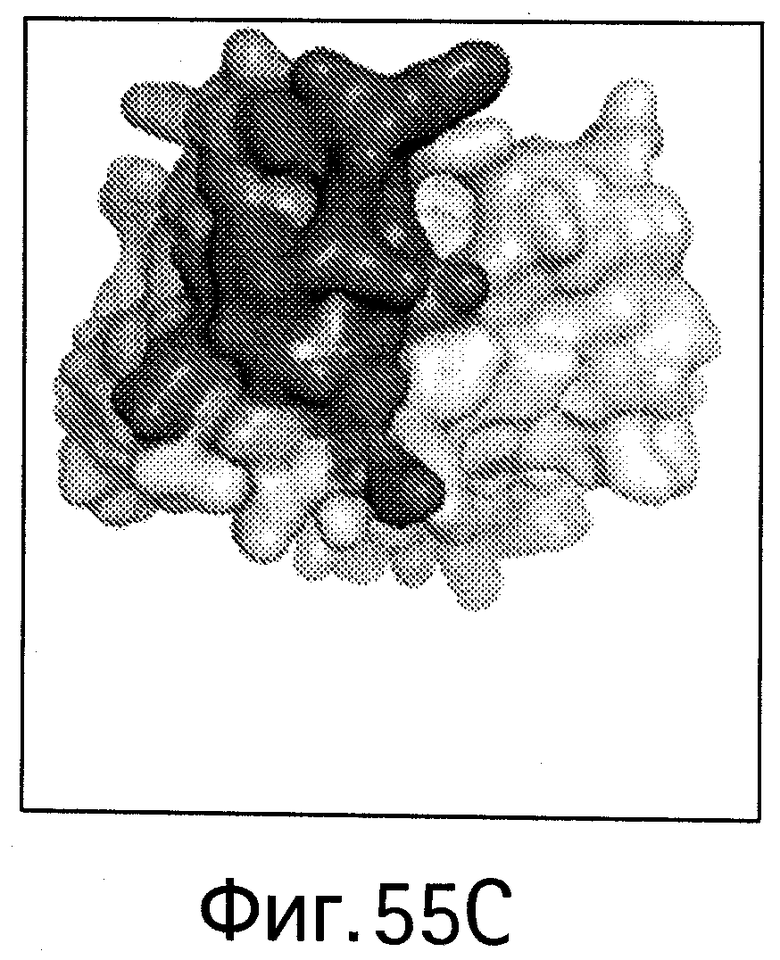
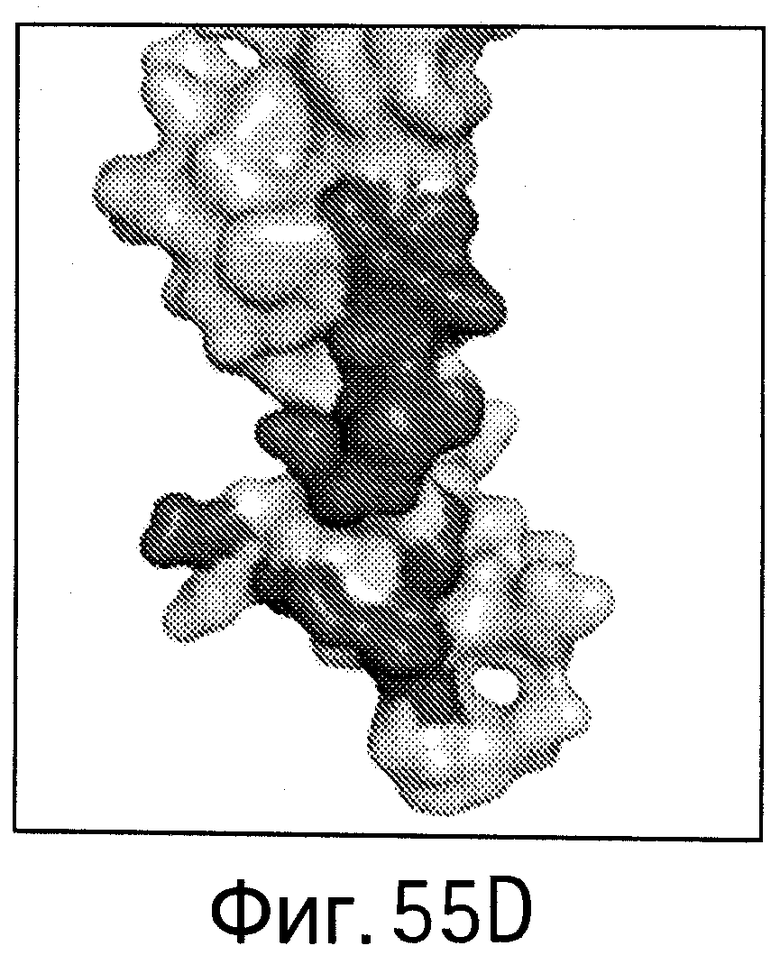
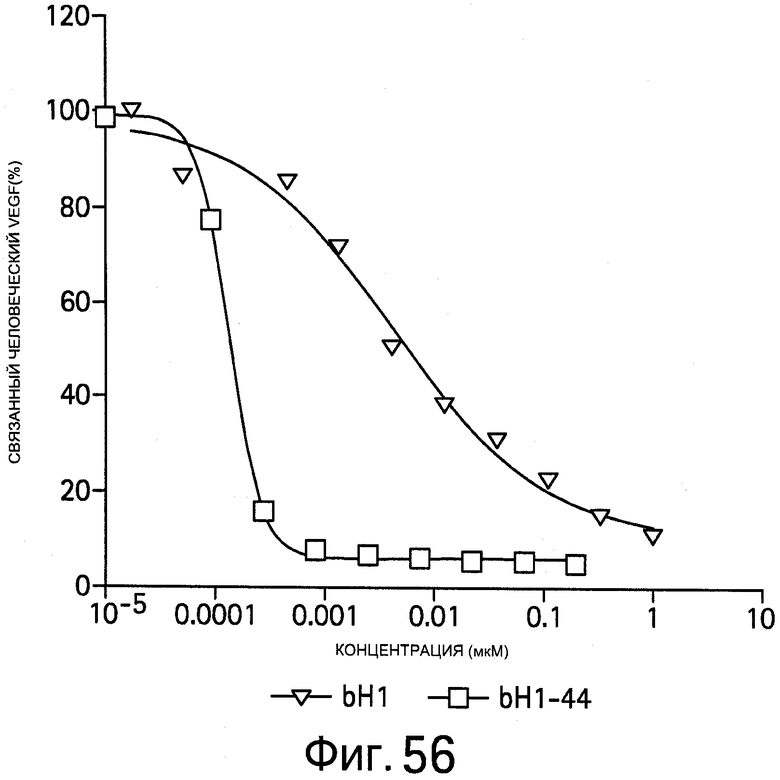
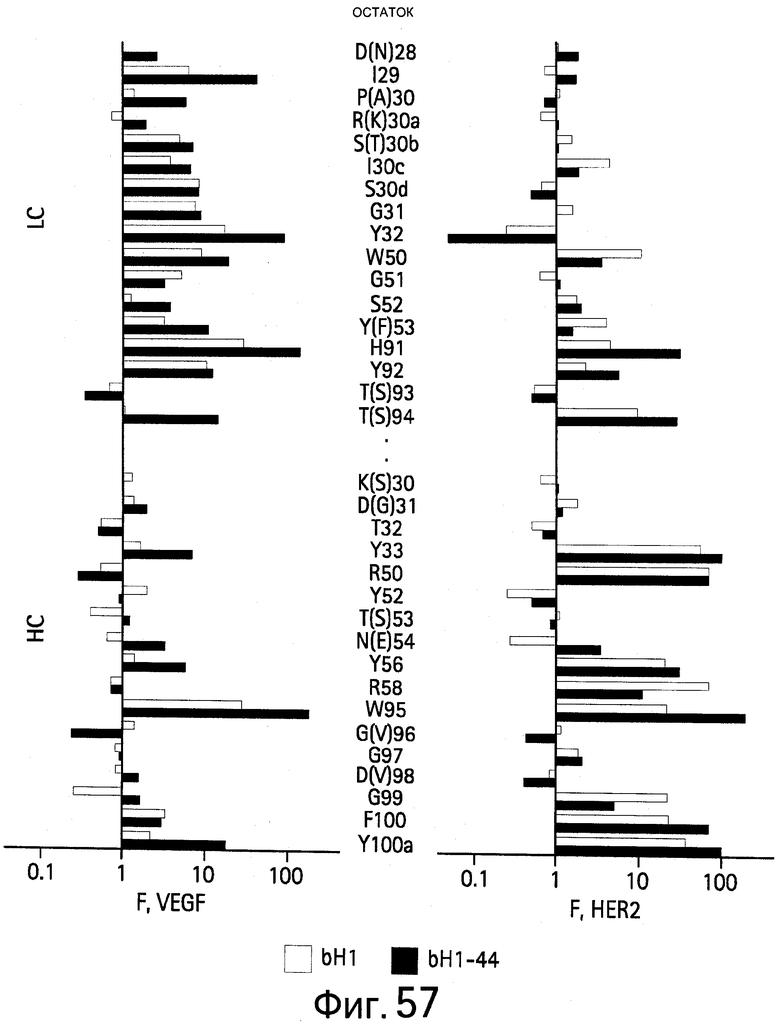

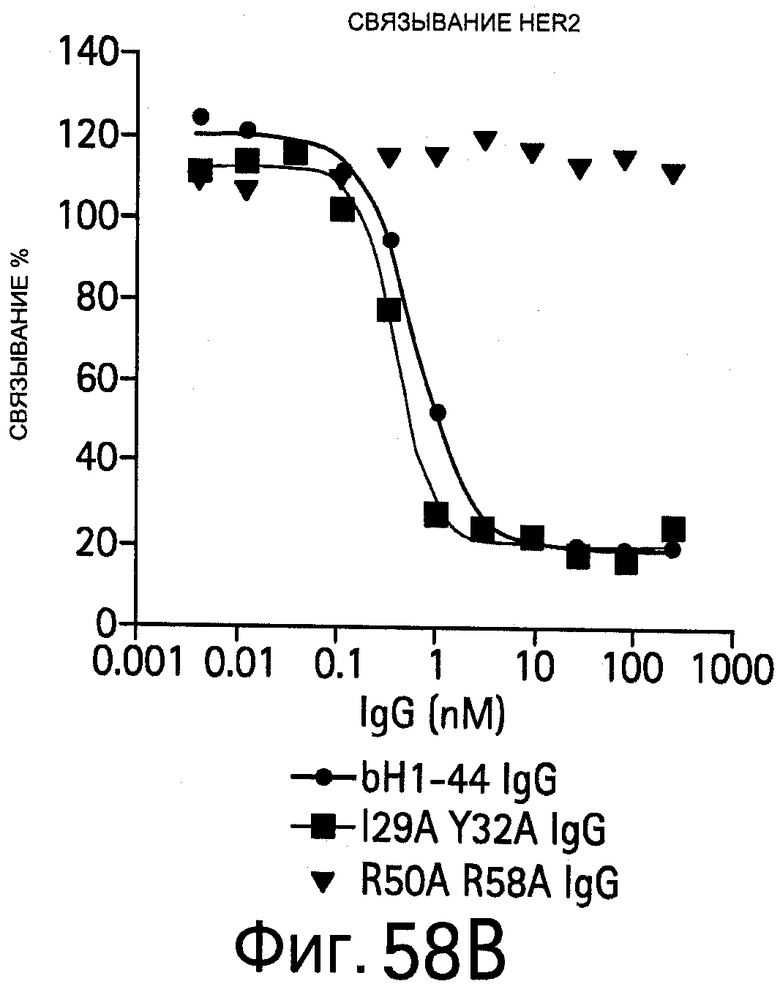
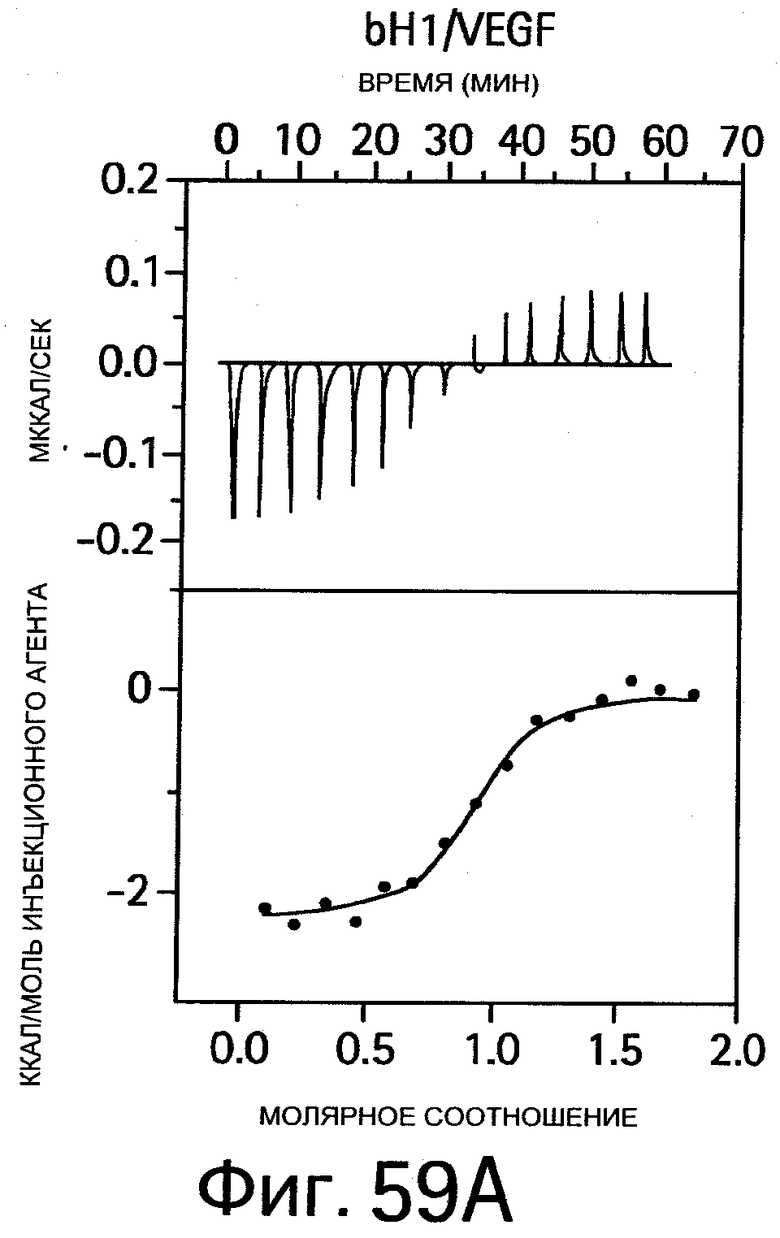
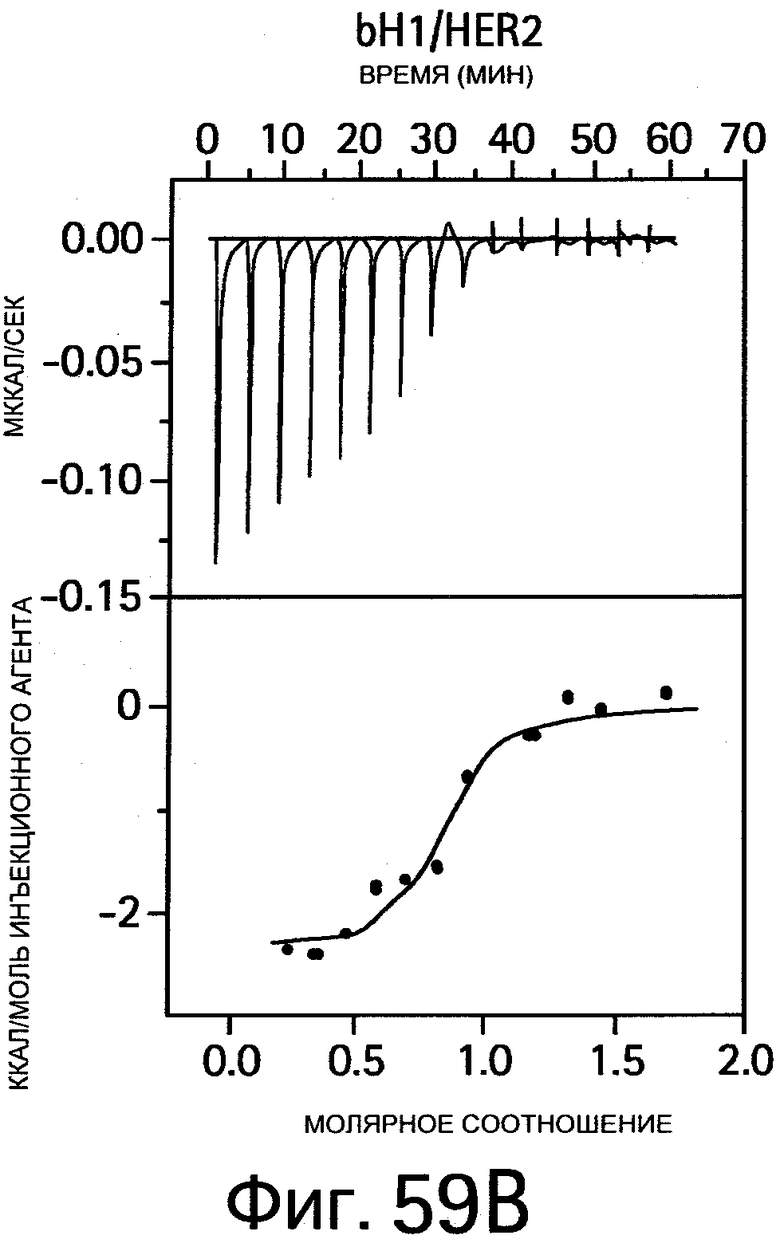

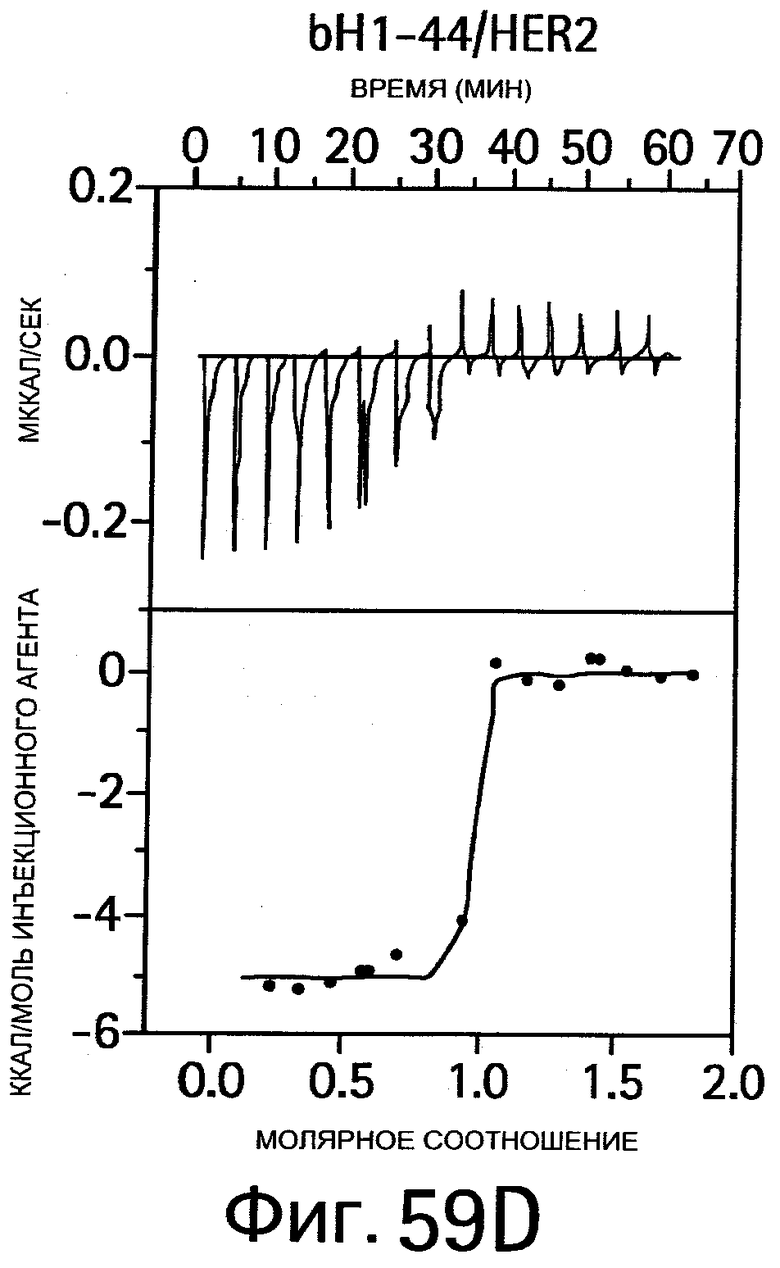
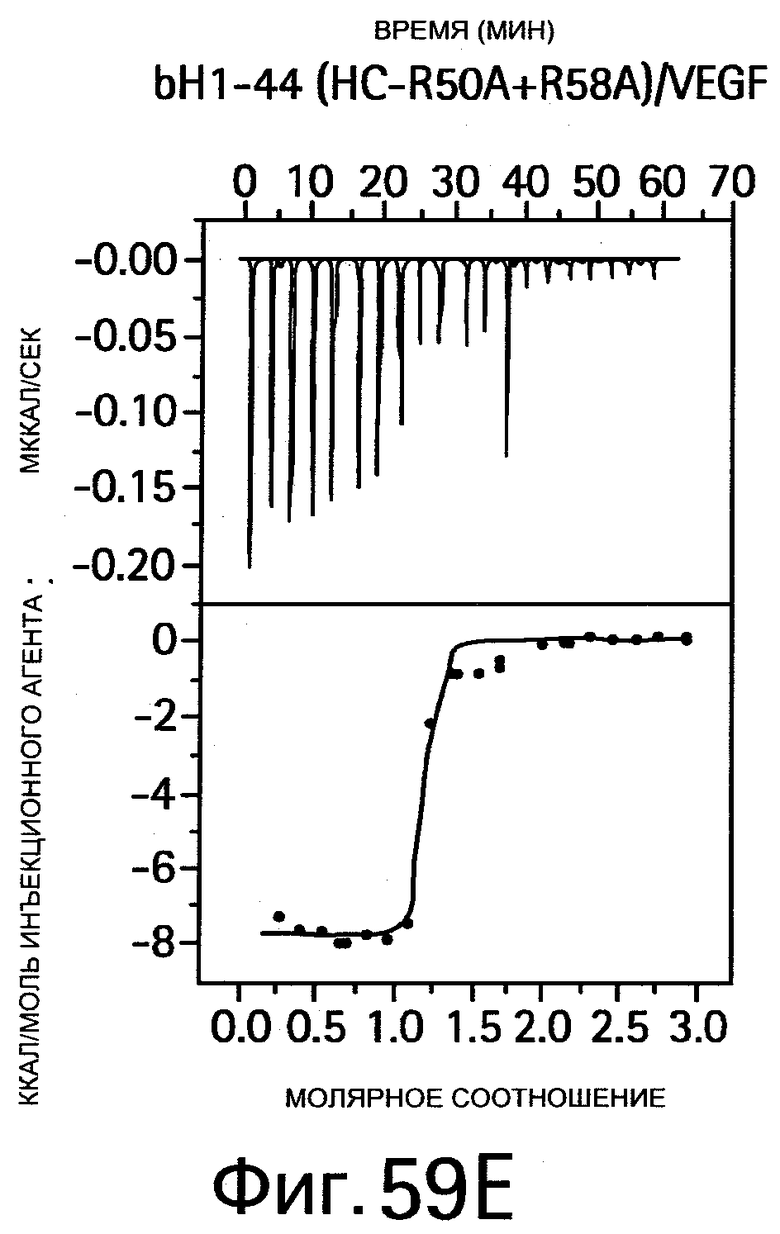
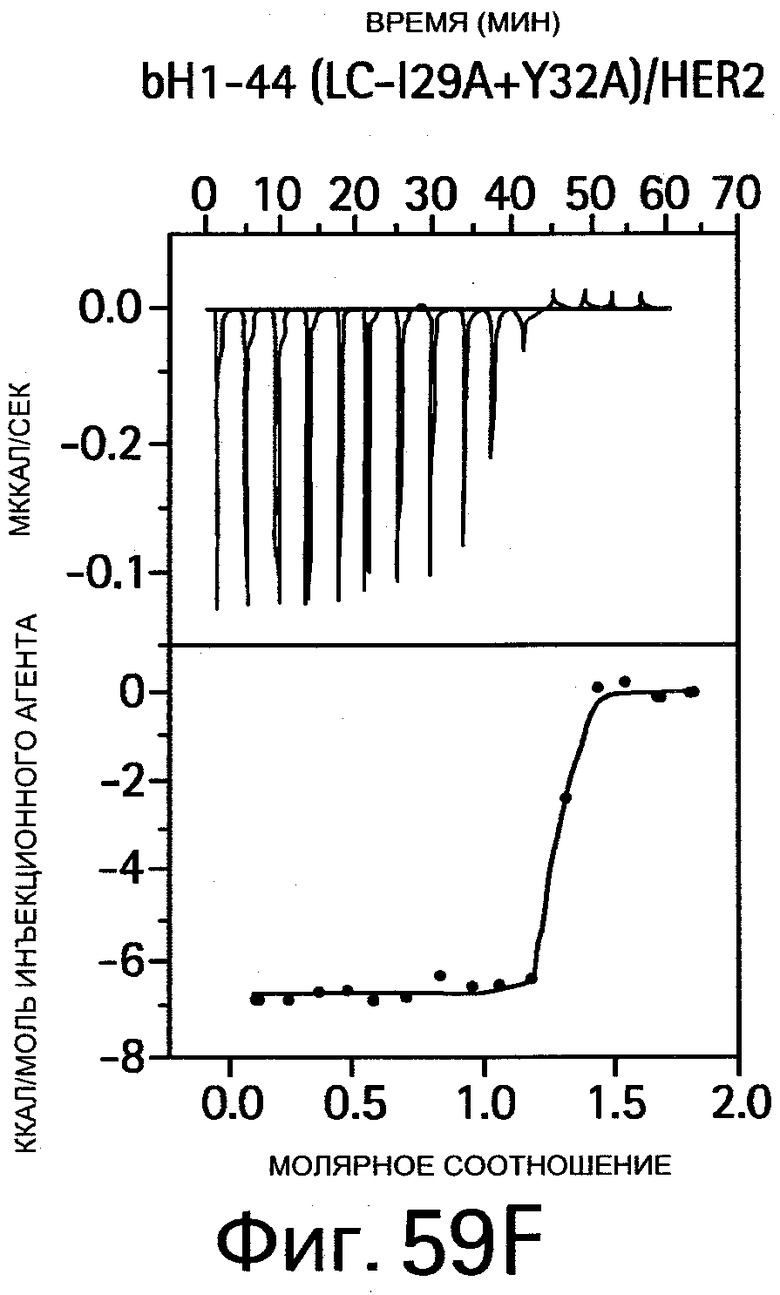



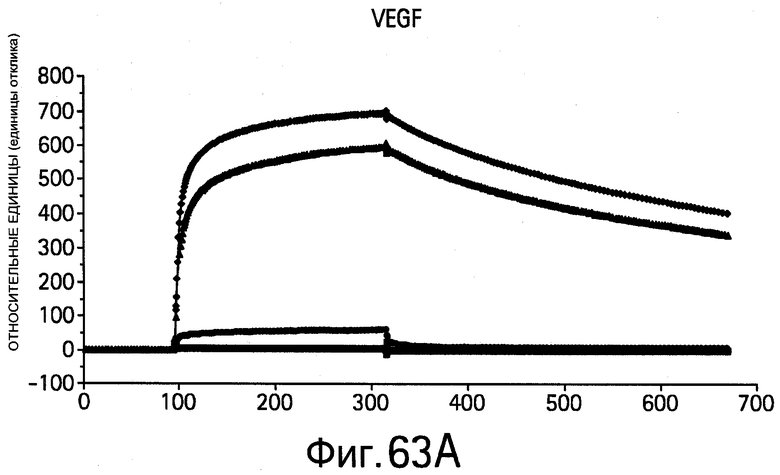
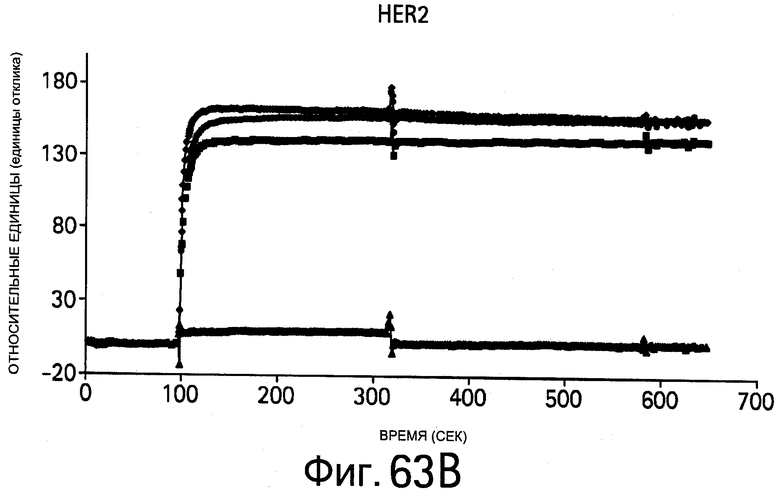
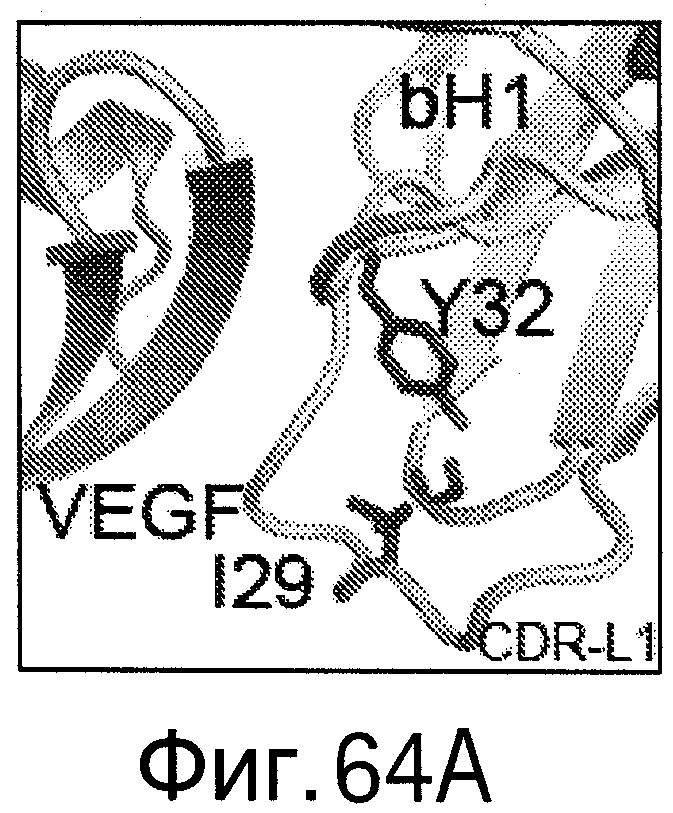
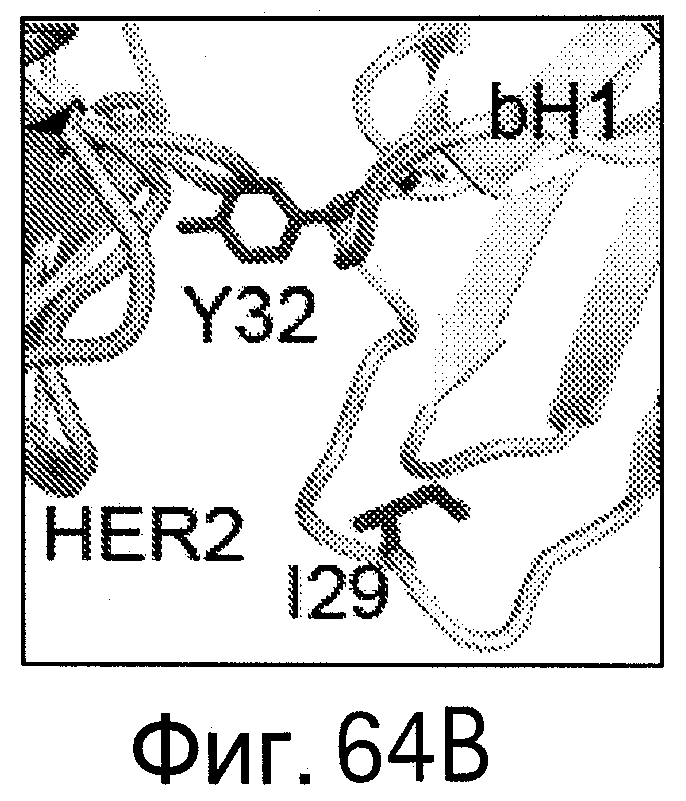
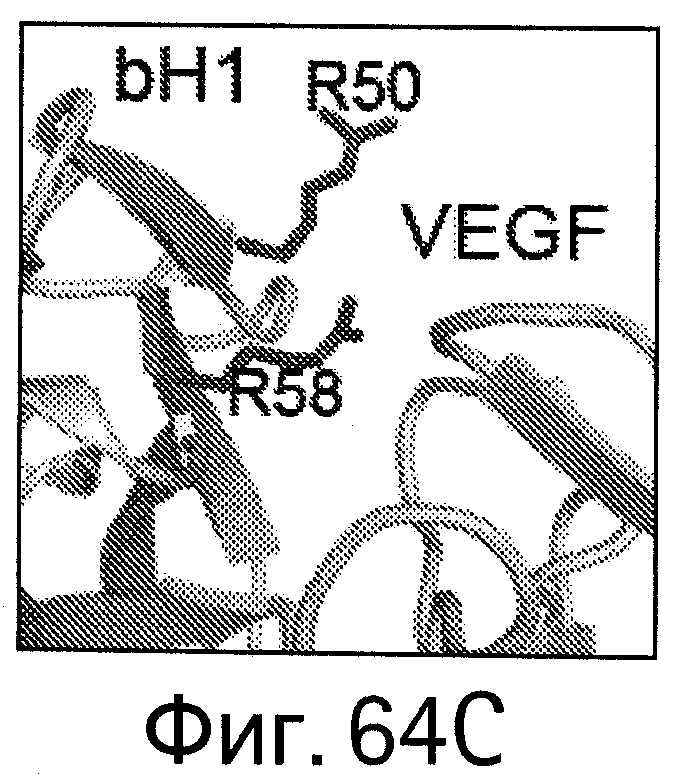
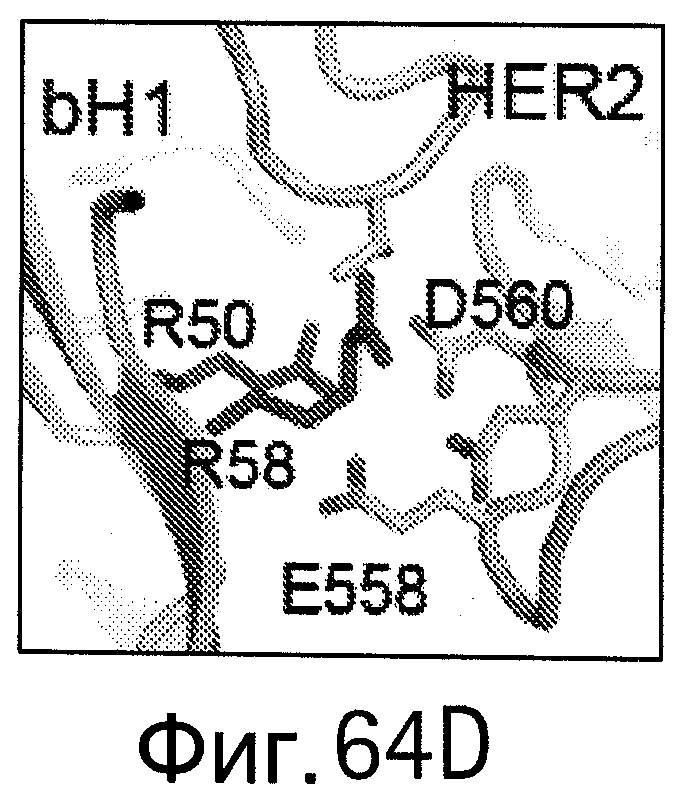

Изобретение относится к области биотехнологии, конкретно к биспецифическим антителам, и может быть использовано в медицине. Конструируют антитело, содержащее одну из следующих групп из шести последовательностей гипервариабельной области (HVR): (a) HVR-L1, включающую последовательность NIAKTISGY; (b) HVR-L2, содержащую последовательность WGSFLY; (c) HVR-L3, содержащую последовательность HYSSPP; (d) HVR-H1, содержащую последовательность NIKDTY; (e) HVR-H2, содержащую последовательность RIYPTNGYTR; и (f) HVR-Н3, содержащую последовательность WGGDGFYAMD; или (a) HVR-L1, включающую последовательность NIAKTISGY; (b) HVR-L2, содержащую последовательность WGSFLY; (c) HVR-L3, содержащую последовательность HYSSPP; (d) HVR-H1, содержащую последовательность NISGTY; (e) HVR-H2, содержащую последовательность RIYPSEGYTR; и (f) HVR-Н3, содержащую последовательность WVGVGFYAMD. Полученное антитело специфически связывает рецептор человеческого эпидермального фактора роста 2 (HER2) и сосудистый эндотелиальный фактор роста (VEGF). Изобретение относится также к выделенному фрагменту Fab указанного антитела, полинуклеотиду, его кодирующему, вектору экспрессии, клетке-хозяину, способу его получения, а также к его применению для лечения HER2-опосредованных заболеваний. Настоящее изобретение позволяет получить биспецифичное антитело, способное связывать одновременно VEGF и HER2 с улучшенной аффинностью. 9 н. и 5 з.п. ф-лы, 65 ил., 16 табл., 8 пр.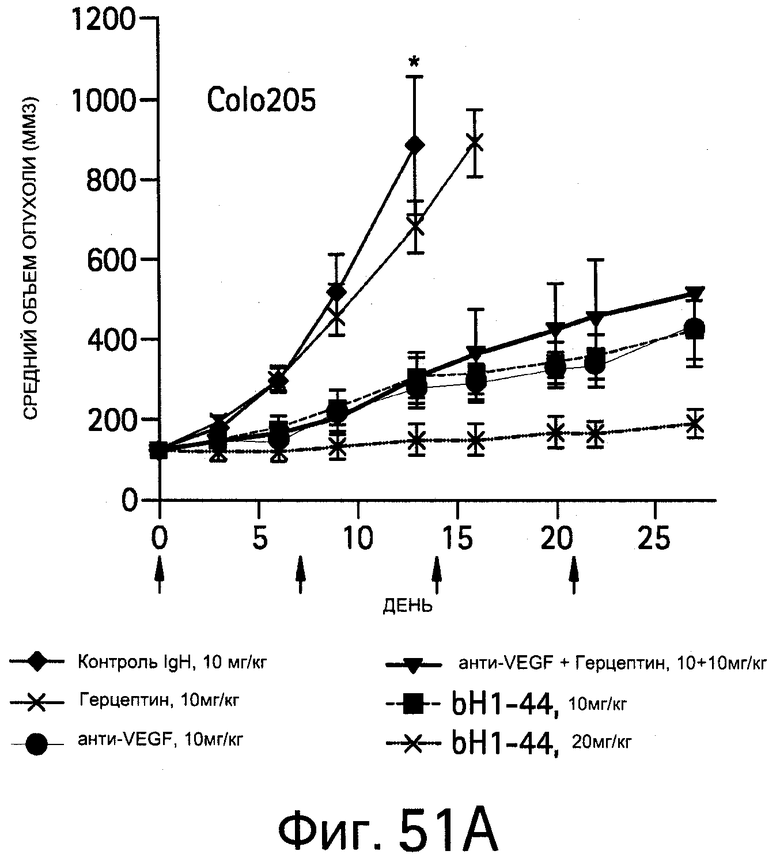
1. Выделенное антитело, специфически связывающее рецептор человеческого эпидермального фактора роста 2 (HER2) и сосудистый эндотелиальный фактор роста (VEGF), содержащее одну из следующих групп из шести последовательностей гипервариабельной области (HVR):
(a) HVR-L1, включающую последовательность NIAKTISGY (SEQ ID NO:1);
(b) HVR-L2, содержащую последовательность WGSFLY (SEQ ID NO:2);
(c) HVR-L3 содержащую последовательность HYSSPP (SEQ ID NO:3)
(d) HVR-H1, содержащую последовательность NIKDTY (SEQ ID NO:4);
(e) HVR-H2, содержащую последовательность RIYPTNGYTR (SEQ ID NO:5); и
(f) HVR-Н3, содержащую последовательность WGGDGFYAMD (SEQ ID NO:6); или
(a) HVR-L1, включающую последовательность NIAKTISGY (SEQ ID NO:1);
(b) HVR-L2, содержащую последовательность WGSFLY (SEQ ID NO:2);
(c) HVR-L3 содержащую последовательность HYSSPP (SEQ ID NO:3);
(d) HVR-H1, содержащую последовательность NISGTY (SEQ ID NO:7);
(e) HVR-H2, содержащую последовательность RIYPSEGYTR (SEQ ID NO:8); и
(f) HVR-Н3, содержащую последовательность WVGVGFYAMD (SEQ ID NO:9).
2. Антитело по п. 1, которое:
(a) связывает человеческий и мышиный VEGF с Kd 150 нМ или сильнее и HER2 с Kd 7 нМ или сильнее;
(b) ингибирует VEGF-индуцированную пролиферацию клеток и пролиферацию HER2-экспрессирующих клеток по сравнению с контролем; или
(c) ингибирует связывание VEGF с VEGF рецептором 2 (VEGFR2).
3. Антитело по п. 1 или 2, которое связывает человеческий и мышиный VEGF с Kd 150 нМ или сильнее и HER2 с Kd 7 нМ или сильнее, и которое ингибирует VEGF-индуцированную пролиферацию клеток и пролиферацию HER2-экспрессирующих клеток по сравнению с контролем, где антитело необязательно связывает человеческий и мышиный VEGF с Kd 36 нМ или сильнее и HER2 с Kd 1 нМ или сильнее.
4. Антитело по любому из пп. 1-3, где указанное антитело является моноклональным антителом, или где указанное антитело представляет собой IgG антитело.
5. Выделенный фрагмент Fab антитела по любому из пп. 1-3, где указанный фрагмент Fab связывает человеческий VEGF с Kd 58 нМ или сильнее и HER2 с Kd 6 нМ или сильнее, и где указанный фрагмент Fab ингибирует VEGF-индуцированную пролиферацию клеток и пролиферацию HER2-экспрессирующих клеток по сравнению с контролем, где указанный фрагмент Fab необязательно связывает человеческий и мышиный VEGF с Kd 33 нМ или сильнее и HER2 с Kd 0.7 нМ или сильнее.
6. Антитело или фрагмент Fab по любому из пп. 1-5, у которого по меньшей мере участок каркасной последовательности представляет собой консенсусную последовательность человеческой каркасной области.
7. Полинуклеотид, кодирующий антитело или фрагмент Fab по любому из пп. 1-6.
8. Вектор экспрессии, содержащий полинуклеотид по п. 7.
9. Клетка-хозяин для экспрессии антитела или фрагмента Fab по любому из пп. 1-6, содержащая вектор по п. 8, где необязательно клетка-хозяин является прокариотической, эукариотической или клеткой млекопитающего.
10. Способ получения антитела или фрагмента Fab по любому из пп. 1-6, который включает культивирование клетки-хозяина, которая содержит вектор, содержащий полинуклеотид по п. 7, и извлечение указанного антитела, где необязательно клетка-хозяин является прокариотической, эукариотической или клеткой млекопитающего.
11. Применение антитела или фрагмента Fab по любому из пп. 1-6 для получения лекарственного средства для лечения:
(a) опухоли, включающей патологическую активацию HER2,
(b) аутоиммунного заболевания, включающего патологическую активацию HER2, или
(c) незлокачественного заболевания у субъекта, включающего патологическую активацию HER2 у субъекта,
где указанное антитело или фрагмент Fab вводят в течение такого времени и в таком количестве, которые достаточны для лечения или предупреждения указанной опухоли, аутоиммунного заболевания, или незлокачественного заболевания у указанного субъекта.
12. Применение по п. 11, где указанная опухоль представляет собой колоректальную опухоль, рак молочной железы, рак легкого, почечноклеточную карциному, глиому, глиобластому или рак яичника.
13. Применение антитела или фрагмента Fab по любому из пп. 1-6 в лечении:
(a) опухоли, включающей патологическую активацию HER2,
(b) аутоиммунного заболевания, включающего патологическую активацию HER2, или
(c) незлокачественного заболевания, включающего патологическую активацию HER2, у субъекта,
причем указанное антитело или фрагмент Fab вводят в течение такого времени и в таком количестве, которые достаточны для лечения или предупреждения указанной опухоли, аутоиммунного заболевания или незлокачественного заболевания у указанного субъекта.
14. Применение антитела или фрагмента Fab по любому из пп. 1-6 в качестве активного ингредиента в композиции для лечения:
(a) опухоли, включающей патологическую активацию HER2,
(b) аутоиммунного заболевания, включающего патологическую активацию HER2, или
(c) незлокачественного заболевания, включающего патологическую активацию HER2, у субъекта,
причем указанное антитело или фрагмент Fab вводят в течение такого времени и в таком количестве, которые достаточны для лечения или предупреждения указанной опухоли, аутоиммунного заболевания или незлокачественного заболевания у указанного субъекта.
| WO 2008027236 А2, 06.03.2008 | |||
| JP 2002355074 A, 10.12.2002 | |||
| РОЙТ А | |||
| и др., Иммунология, Москва, "Мир", 2000, 592 с |

