ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННУЮ ЗАЯВКУ
По настоящей заявке испрашивается приоритет согласно предварительной патентной заявке США с серийным № 60/758439, поданной 12 января 2006 г., полное содержание которой включено в данное описание посредством ссылки, включая любые фигуры, таблицы или чертежи.
ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к получению не-трансгенного растения, устойчивого или толерантного к гербициду фосфонометилглициновой группы, например глифосату. Настоящее изобретение также относится к использованию рекомбинагенного олигонуклеооснования для получения желаемой мутации в хромосомных или эписомных последовательностях растения в гене, кодирующем 5-енолпирувилшикимат-3-фосфатсинтазу (EPSPS). Мутантный белок, который по существу сохраняет каталитическую активность белка дикого типа, делает возможной повышенную устойчивость или толерантность растения к гербициду фосфонометилглициновой группы и по существу позволяет растению, его органам, тканям или клеткам нормально расти или развиваться в сравнении с растением дикого типа независимо от присутствия или отсутствия гербицида. Настоящее изобретение также относится к клетке E. coli, имеющей мутантный ген EPSPS, клетке не-трансгенного растения, в которой ген EPSPS мутирован, регенерированному из нее не-трансгенному растению, а также растению, полученному в результате скрещивания, с использованием в качестве одного из родителей регенерированного не-трансгенного растения, имеющего мутантный ген EPSPS. Настоящий мутантный белок EPSPS изменен по аминокислотным позициям 159, 178, 182, 193, 244, 273 и/или 454 в белке EPSPS Arabidopsis (NM 130093) или по аналогичному аминокислотному остатку в паралоге EPSPS.
УРОВЕНЬ ТЕХНИКИ ИЗОБРЕТЕНИЯ
Фосфонометилглициновые гербициды
Толерантные к гербициду растения могут снизить потребность в обработке почвы для борьбы с сорняками, что приводит к эффективному уменьшению почвенной эрозии. Один из гербицидов, который является предметом интенсивного изучения с этой точки зрения, представляет собой N-фосфонометилглицин, обычно называемый глифосат. Глифосат ингибирует каскад реакций шикимовой кислоты, который приводит к биосинтезу ароматических соединений, включая аминокислоты, гормоны и витамины. В частности, глифосат контролирует превращение фосфоенолпировиноградной кислоты (PEP) и 3-фосфошикимовой кислоты в 5-енолпирувил-3-фосфошикимовую кислоту, ингибируя фермент 5-енолпирувилшикимат-3-фосфатсинтазу (далее называемую EPSP-синтаза или EPSPS). Для целей настоящего изобретения термин «глифосат» включает любую эффективную в качестве гербицида форму N-фосфонометилглицина (включая любую его соль), другие формы, которые приводят к образованию аниона глифосата в растениях, а также любые другие гербициды фосфонометилглициновой группы.
Толерантность растений к глифосату можно повысить путем введения в геном растения мутантного гена EPSPS с измененной последовательностью, кодирующей аминокислоты EPSPS. Примеры некоторых мутаций в гене EPSPS для индуцирования толерантности к глифосату описаны в следующих патентах: патент США № 5310667; патент США № 5866775; патент США № 5312910; патент США №5145783. Предложенные там мутанты обычно имеют более высокую Ki для гликосата, чем фермент EPSPS дикого типа, что придает толерантный к глифосату фенотип, однако данные варианты также характеризуются высокой Km для PEP, что делает фермент кинетически менее эффективным (Kishore et al., 1998, Ann. Rev. Biochem. 57:627-663; Schulz et al., 1984, Arch. Microbiol. 137:121-123; Sost et al., 1984, FEBS Lett. 173:238-241; Kishore et al., 1986, Fed. Proc. 45: 1506; Sost and Amrhein, 1990, Arch. Biochem. Biophys. 282: 433-436). Многие мутации гена EPSPS выбирают таким образом, чтобы получить фермент EPSPS, который устойчив к гербицидам, но, к сожалению, фермент EPSPS, произведенный мутантным геном EPSPS, имеет существенно более низкую ферментативную активность, чем EPSPS дикого типа. Например, кажущаяся Km для PEP и кажущаяся Ki для глифосата для EPSPS дикого типа из E. coli составляют 10 мкМ и 0,5 мкМ соответственно, тогда как для толерантной к глифосату культуры, имеющей одиночную аминокислотную замену глицина аланином в позиции 96, эти значения составляют 220 мкМ и 4,0 мМ соответственно. При помощи мутагенеза был получен ряд генов EPSPS, толерантных к глифосату. Опять-таки, толерантная к глифосату EPSPS имела более низкую каталитическую эффективность (Vmax/Km), о чем свидетельствовало увеличение Km для PEP и небольшое снижение Vmax у растительного фермента дикого типа (Kishore et al., 1998, Ann. Rev. Biochem. 57:627-663).
Поскольку кинетические константы вариантных ферментов снижены в отношении PEP, предположили, что высокие, 40-80-кратные, уровни сверхпродукции вариантного фермента потребуются для поддержания нормальной каталитической активности в растениях в присутствии глифосата (Kishore et al., 1988, Ann. Rev. Biochem. 57:627-663). Было показано, что толерантные к глифосату растения можно получать, встраивая в геном растения способность производить более высокий уровень EPS-синтазы в хлоропластах клетки (Shah et al., 1986, Science 233, 478-481), где указанный фермент предпочтительно является толерантным к глифосату (Kishore et al., 1988, Ann. Rev. Biochem. 57:627-663).
Введение экзогенных мутантных генов EPSPS в растение задокументировано. Например, согласно патенту США № 4545060, с целью повышения устойчивости растения к глифосату в растительный геном введен ген, кодирующий вариант EPSPS, имеющий по меньшей мере одну мутацию, которая делает фермент более устойчивым к его конкурентному ингибитору, то есть глифосату. Однако с такими трансгенными растениями, содержащими мутантные гены EPSPS, связано много сложностей и проблем. Много таких мутаций приводит к низкой экспрессии продукта мутантного гена EPSPS или к продукту гена EPSPS со значительно более низкой ферментативной активностью по сравнению с диким типом. Низкая экспрессия или низкая ферментативная активность мутантного фермента приводит к аномально низким уровням роста и развития растения.
Несмотря на то, что была доказана пригодность подобных вариантов EPSP-синтаз для получения трансгенных растений, толерантных к глифосату, гораздо большие преимущества дало бы получение продукта вариантного гена EPSPS, который в высокой степени толерантен к глифосату, однако все еще является кинетически эффективным, так чтобы повышенную толерантность можно было получать при уровне экспрессии, как у дикого типа.
Рекомбинагенные олигонуклеооснования
Рекомбинагенные олигонуклеооснования и их использование для осуществления генетических изменений в эукариотических клетках описаны в патенте США № 5565350, выданном Kmiec (Kmiec I). Kmiec I сообщают о способе введения специфических генетических изменений в целевой ген. В Kmiec I раскрыты, в числе прочего, рекомбинагенные олигонуклеооснования, имеющие две нити, в которых первая нить содержит два сегмента из, по меньшей мере, 8 РНК-подобных нуклеотидов, которые разделены третьим сегментом из примерно 4-50 ДНК-подобных нуклеотидов, называемым «промежуточным ДНК-сегментом». Нуклеотиды первой нити образуют пары с ДНК-подобными нуклеотидами второй нити. Первая и вторая нити дополнительно связаны сегментом из однонитевых нуклеотидов так, что первая и вторая нити являются частями одной олигонуклеотидной цепи. В Kmiec I дополнительно сообщается о способе введения специфических генетических изменений в целевой ген. В соответствии с Kmiec I, последовательности РНК-сегментов выбирают так, чтобы они были гомологичными, то есть идентичными последовательности первого и второго фрагмента целевого гена. Последовательность промежуточного ДНК-сегмента является гомологичной последовательности целевого гена между первым и вторым фрагментом, за исключением области несовпадения, называемой «гетерологичной областью». Гетерологичная область может вызывать вставку или делецию, либо может содержать одно или несколько оснований, которые не совпадают с последовательностью целевого гена так, что это вызывает замену. В соответствии с Kmiec I, последовательность целевого гена изменяется под контролем гетерологичной области, так что целевой ген становится гомологичным последовательности рекомбинагенного олигонуклеооснования. В Kmiec I конкретно указывается, что нуклеотиды, содержащие рибозу и 2'-O-метилрибозу, то есть 2'-метоксирибозу, можно использовать в рекомбинагенных олигонуклеооснованиях и что природные содержащие дезоксирибозу нуклеотиды можно использовать в качестве ДНК-подобных нуклеотидов.
В патенте США № 5731181, выданном Kmiec (Kmiec II), в частности, раскрыто использование рекомбинагенных олигонуклеооснований для создания генетических изменений в растительных клетках и раскрыты дополнительные примеры аналогов и производных РНК-подобных и ДНК-подобных нуклеотидов, которые можно использовать для создания генетических изменений в конкретных целевых генах. Другие патенты, в которых обсуждается использование рекомбинагенных олигонуклеооснований, включают: патенты США №№ 5756325; 5871984; 5760012; 5888983; 5795972; 5780296; 5945339; 6004804 и 6010907, а также в международном патенте № PCT/US00/23457; и в международных патентных публикациях №№ WO 98/49350; WO 99/07865; WO 99/58723; WO 99/58702 и WO 99/40789. Рекомбинагенные олигонуклеооснования включают смешанные двунитевые олигонуклеотиды, молекулы, не содержащие нуклеотидов, указанные в Kmiec II, и другие молекулы, указанные в приведенных выше патентах и патентных публикациях.
В патенте США 6870075 (патент '075) раскрыт способ получения не-трансгенных устойчивых или толерантных к гербициду растений с применением рекомбинагенных олигонуклеооснований по методам, описанным в Kmiec I и Kmiec II. Мутанты EPSPS, раскрытые в патенте '075, содержат изменения, произведенные на следующих аминокислотных позициях белка EPSPS: Leu173, Gly177, Thr178, Ala179, Met180, Arg181, Pro182, Ser98, Ser255 и Leu198 в белке EPSPS Arabidopsis или по аналогичному аминокислотному остатку в паралоге EPSPS.
В опубликованной патентной заявке США 20030084473 также раскрыто применение рекомбинагенных олигонуклеооснований для получения не-трансгенных устойчивых к гербициду растений, где белок EPSPS изменен по аминокислотным позициям 126, 177, 207, 438, 479, 480 и/или 505 в белке EPSPS Arabidopsis или по аналогичному аминокислотному остатку в паралоге EPSPS.
Настоящее изобретение относится к дополнительным аминокислотным мутациям, которые можно осуществить в любом гене EPSPS из любых видов для получения генного продукта, который обладает устойчивостью к глифосату.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Вкратце, в соответствии с настоящим изобретением получают не-трансгенное растение или растительную клетку, имеющее одну или несколько мутаций в гене EPSPS. Полученное растение обладает повышенной устойчивостью или толерантностью к члену фосфонометилглициновой группы, такому как глифосат, и проявляет по существу нормальный рост или развитие растения, его органов, тканей или клеток в сравнении с соответствующим растением или клеткой дикого типа. Мутантный ген продуцирует генный продукт, имеющий замену в одной или нескольких из аминокислотных позиций 160, 179, 183, 194, 244, 273 и/или 454 в белке EPSPS Arabidopsis (AF360224) или по аналогичному аминокислотному остатку в паралоге EPSPS. Предпочтительно, мутантное растение является устойчивым к глифосату и обладает по существу такой же каталитической активностью в сравнении с белком EPSPS дикого типа.
Кроме того, настоящее изобретение охватывает мутантный ген EPSPS из E. coli и мутантные клетки E. coli, которые продуцируют генный продукт, имеющий замену в одной или нескольких из аминокислотных позиций 82, 97, 101, 114, 164, 193 и 374. Мутантный ген EPSPS E. coli можно использовать для тестирования in vitro мутантного генного продукта. Если будут выявлены активные мутанты E. coli, тогда соответствующие мутации можно осуществлять в гене EPSPS желаемой сельскохозяйственной культуры для придания культуре устойчивости к гербициду.
Настоящее изобретение также относится к способу получения не-трансгенного растения, имеющего мутантный ген EPSPS, который по существу сохраняет каталитическую активность белка дикого типа независимо от присутствия или отсутствия гербицида фосфонометилглициновой группы. Способ включает введение в растительную клетку рекомбинагенного олигонуклеооснования с направленной мутацией в гене EPSPS, который продуцирует генный продукт, имеющий одну или несколько из вышеуказанных аминокислотных замен. Способ дополнительно включает идентификацию клетки, семени или растения, имеющего мутантный ген EPSPS, и методы культивирования и регенерации для получения растения, которое производит семена, далее «фертильное растение», а также производство семян и дополнительных растений из такого фертильного растения, включая растения-отпрыски (потомство), которые содержат мутантный ген EPSPS.
Кроме того, изобретение относится к способу избирательной борьбы с сорняками в поле. В поле находятся растения с описанными изменениями гена EPSPS и сорняки. Способ включает обработку поля фосфонометилглициновым гербицидом, к которому указанные растения стали устойчивыми, и сорняки погибают. Предпочтительным гербицидом является глифосат.
Изобретение также относится к новым мутациям в гене EPSPS и полученному в результате генному продукту, который придает устойчивость или толерантность к члену фосфонометилглициновой группы, например глифосату, растению и, где мутантная EPSPS обладает по существу такой же ферментативной активностью в сравнении с EPSPS дикого типа. Кроме того, настоящее изобретение относится к продукту мутантного гена EPSPS E. coli (белку), который используют для проведения скрининга мутантов EPSPS с целью использования в качестве мутаций устойчивости к гербициду в растениях.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На фиг.1 представлена белковая последовательность продукта гена EPSPS (AroA ген) в E. Coli, где позиции мутантных аминокислот обведены рамкой. Замещенная аминокислота на этих позициях представлена под последовательностью.
На фиг.2 представлена белковая последовательность AtEPSPS кДНК - At2g45300, транслированная с образца Genbank NM_130093 (Arabodopsis).
На фиг.3 представлена белковая последовательность AtEPSPS кДНК - At1g48860, транслированная с образца Genbank AF360224T (Arabodopsis).
На фиг.4 представлена белковая последовательность BnEPSPS кДНК - BN-2 2-23 (Canola).
На фиг.5 представлена белковая последовательность BnEPSPS кДНК - 2-28 от трансляции гДНК X51475 (Canola).
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Определения
Данное изобретение следует понимать в соответствии со следующими определениями.
Олигонуклеооснование представляет собой полимер нуклеиновых оснований, где полимер способен гибридизоваться по принципу спаривания оснований Уотсона-Крика с ДНК, обладающей комплементарной последовательностью.
Нуклеиновые основания включают основания, которые представляют собой пурин, пиримидин, либо их производные или аналоги. Нуклеиновые основания включают пептидные нуклеиновые основания, субъединицы нуклеиновых кислот пептидов, и морфолиновые нуклеиновые основания, а также нуклеозиды и нуклеотиды. Нуклеозиды представляют собой нуклеиновые основания, которые содержат пентозофуранозильный фрагмент, например, необязательно замещенный рибозид или 2'-дезоксирибозид. Нуклеозиды могут быть соединены одним или несколькими связывающими фрагментами, которые могут содержать или не содержать фосфор. Нуклеозиды, которые соединены посредством незамещенных фосфодиэфирных связей, называются нуклеотидами.
Цепь олигонуклеооснований имеет по одному 5'- и 3'-концу, которые представляют собой крайние нуклеиновые основания полимера. Отдельная цепь олигонуклеооснований может содержать нуклеиновые основания всех типов. Соединение олигонуклеооснований представляет собой соединение, включающее одну или несколько цепей олигонуклеооснований, которые являются комплементарными и гибридизуются по принципу спаривания оснований Уотсона-Крика. Нуклеиновые основания могут быть либо дезоксирибо-типа, либо рибо-типа. Нуклеиновые основания рибо-типа представляют собой нуклеиновые основания, содержащие пентозофуранозил, где углерод в 2' положении представляет собой метилен, замещенный гидроксилом, алкилокси или галогеном. Нуклеиновые основания дезоксирибо-типа представляют собой нуклеиновые основания, отличные от нуклеиновых оснований рибо-типа, и включают все нуклеиновые основания, которые не содержат пентозофуранозильный фрагмент.
Нить олигонуклеооснований главным образом включает как цепи олигонуклеооснований, так и сегменты или участки цепей олигонуклеооснований. Нить олигонуклеооснований имеет 3'-конец и 5'-конец. Если нить олигонуклеооснований и цепь одинаковы по протяженности, то 3'- и 5'-концы нити являются также 3'- и 5'-концами цепи.
Согласно настоящему изобретению, по существу нормальный рост растения, органа растения, ткани растения или клетки растения определяют как скорость роста или скорость деления клеток растения, органа растения, ткани растения или клетки растения, которая составляет по меньшей мере 35%, по меньшей мере 50%, по меньшей мере 60%, или по меньшей мере 75% от скорости роста или скорости деления клеток в соответствующем растении, органе растения, ткани растения или клетке растения, экспрессирующего белок EPSPS дикого типа.
Согласно настоящему изобретению, по существу нормальное развитие растения, органа растения, ткани растения или клетки растения определяют как наличие одного или нескольких событий развития в растении, органе растения, ткани растения или клетке растения, которые являются по существу такими же, как те, что имеют место в соответствующем растении, органе растения, ткани растения или клетке растения, экспрессирующих белок EPSPS дикого типа.
Согласно настоящему изобретению, органы растения включают, но не ограничиваются ими, листья, стебли, корни, вегетативные почки, цветочные почки, меристемы, зародыши, семядоли, эндосперм, чашелистики, лепестки, пестики, плодолистики, тычинки, пыльники, микроспоры, пыльцу, пыльцевые трубки, семяпочки, завязи и плоды, либо взятые от них секции, срезы или диски. Ткани растения включают, но не ограничиваются ими, ткани каллюса, покровные ткани, проводящие ткани, запасающие ткани, меристематические ткани, ткани листьев, ткани побегов, ткани корня, ткани галлов, ткани опухолей растений и репродуктивные ткани. Клетки растения включают, но не ограничиваются ими, выделенные клетки с клеточными стенками, их агрегаты различного размера и протопласты.
Растения являются по существу «толерантными» к глифосату, если при обработке им кривая доза/ответ, полученная для них, смещена вправо при сравнении с таковой, полученной для аналогичным образом обработанных растений нетолерантного типа. В таких кривых доза/ответ по оси Х отмеряют значения «дозы» и по оси Y отмеряют «процент гибели», «гербицидный эффект» и так далее. Для толерантных растений потребуется больше гербицида, чем для растений нетолерантного типа, чтобы достичь определенного гербицидного эффекта. Растения, которые являются по существу «устойчивыми» к глифосату, имеют мало, если вообще имеют, некротических, литических, хлоротических или других повреждений при обработке глифосатом в тех концентрациях и режимах, которые обычно используют в агрохимической практике для уничтожения сорняков в поле. Растения, которые являются устойчивыми к гербициду, также являются толерантными к гербициду. Термины «устойчивый» и «толерантный» следует толковать в контексте настоящей заявки как «толерантный и/или устойчивый».
Термин «гомолог EPSPS» или любая его вариация означает ген EPSPS или продукт гена EPSPS, обнаруженный в других видах растений, который выполняет ту же или в основном ту же биологическую функцию, что и гены EPSPS, раскрытые в данном описании, и где последовательности нуклеиновых кислот или полипептидные последовательности (продукта гена EPSPS), как утверждают, являются «идентичными» или, по меньшей мере, на 50% сходными (также называемыми «с процентной идентичностью» или «в основном идентичными»), как описано ниже. Два полинуклеотида или полипептида являются идентичными, если последовательность нуклеотидов или аминокислотных остатков, соответственно, в двух последовательностях является той же самой при выравнивании для максимального совпадения, как описано ниже. Термины «идентичный» или «процентная идентичность» в контексте двух или более последовательностей нуклеиновых кислот или полипептидов относятся к двум или более последовательностям или подпоследовательностям, которые являются одинаковыми или имеют определенный процент аминокислотных остатков или нуклеотидов, которые являются одинаковыми, при сравнении и выравнивании для максимального совпадения в окне сравнения, что определяют, используя один из следующих алгоритмов сравнения последовательностей, либо при выравнивании вручную и визуальном контроле. Для полипептидов, в которых последовательности различаются в консервативных заменах, процент идентичности последовательностей можно увеличивать, чтобы внести поправку на консервативную природу замены. Способы проведения такой корректировки хорошо известны специалистам в данной области. Обычно сюда относится оценивание консервативной замены скорее как частичного, нежели полного несовпадения, что приводит к повышению процента идентичности последовательностей. Таким образом, например, если идентичной аминокислоте присваивают 1 балл и неконсервативной замене присваивают ноль баллов, то консервативной замене присваивают балл между нулем и 1. Оценку консервативных замен рассчитывают, например, по алгоритму Meyers & Miller, Computer Applic. Biol. Sci. 4:11-17 (1988), что, например, используется в программе PC/GENE (Intelligenetics, Mountain View, Calif., USA).
Словосочетания «по существу идентичные» или «процентная идентичность» в контексте двух нуклеиновых кислот или полипептидов относится к последовательностям или подпоследовательностям, которые имеют по меньшей мере 50%, преимущественно 60%, предпочтительно 70%, более предпочтительно 80% и наиболее предпочтительно 90-95% идентичность нуклеотидов или аминокислотных остатков при выравнивании для максимального совпадения в окне сравнения, что определяют, используя один из следующих алгоритмов сравнения последовательностей, либо при выравнивании вручную и визуальном контроле. Данное определение также относится к комплементу тестируемой последовательности, который имеет существенную комплементарность последовательности или подпоследовательности, когда тестируемая последовательность имеет существенную идентичность с последовательностью сравнения.
Специалист в данной области признает, что два полипептида могут также быть «по существу идентичными», если два полипептида являются иммунологически схожими. Таким образом, белковые структуры в целом могут быть схожими, в то время как первичные структуры двух полипептидов имеют существенные различия. Вследствие этого способ определения, являются ли два полипептида по существу идентичными, включает определение связывания моноклональных или поликлональных антител с каждым полипептидом. Два полипептида являются по существу идентичными, если антитела, специфичные для первого полипептида, связывают второй полипептид с афинностью, составляющей по меньшей мере одну треть от афинности для первого полипептида. При сравнении последовательностей обычно одна последовательность играет роль последовательности сравнения, с которой сравнивают тестируемые последовательности. При использовании алгоритма сравнения последовательностей тестируемые последовательности и последовательность сравнения вносят в компьютер, задают координаты подпоследовательностей, если необходимо, и задают программные параметры алгоритма для последовательностей. Затем алгоритм сравнения последовательностей вычисляет процент идентичности последовательностей для тестируемой последовательности(тей) относительно последовательности сравнения на основании заданных программных параметров.
Оптимальное выравнивание последовательностей для сравнения можно выполнять, например, с помощью алгоритма локальной гомологии Smith & Waterman, 0.4dv. Appl. Math. 2:482 (I 98 I), с помощью алгоритма выравнивания гомологии Needleman & Wunsch, J. Mol. Biol. 48:443 (1970), методом поиска сходства Pearson & Lipman, Proc. Nat'I. Acad. Sci. USA 5 85:2444 (1988), с помощью компьютеризированной реализации данных алгоритмов (GAP, BESTFIT, FASTA и TFASTA в пакете программ Wisconsin Genetics, Genetics Computer Group, 575 Science Dr., Madison, Wis.), с помощью программ для выравниваний, таких как VECTOR NTI Version №6 компании InforMax, Inc. MD, США, с помощью методик, описанных в ClustalW., Thompson, J. D., Higgins, D. G. and Gibson, T. J. (1994) CLUSTALW: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Research, 22:4673-4680, либо с помощью визуального контроля (смотри в основном Protocols in Molecular Biology, F. M. Ausubel et al., eds., Current Protocols, a joint venture between Greene Publishing Associates, Inc. and John Wiley & Sons, Inc., (1995 Supplement) (Ausubel)).
Примерами алгоритмов, подходящих для определения процентной идентичности последовательностей и сходства последовательностей, являются алгоритмы BLAST и BLAST 2.0, которые описаны в Altschul et al. (1990) J. Mol. Biol. 215:403-410 и Altschul et al. (1977) Nucleic Acids Res. 25:3389-3402, соответственно. Программное обеспечение для выполнения анализа BLAST общедоступно через Национальный центр информации по биотехнологии (National Center for Biotechnology Information) (http://www.ncbi.nlm.nih.gov/). Данный алгоритм включает в первую очередь выявление пар с высоким баллом в последовательности (HSPs) путем выявления коротких слов длины W в запрашиваемой последовательности, которые либо соответствуют, либо удовлетворяют некоторому положительному пороговому значению T при выравнивании со словом той же длины в последовательности из базы данных. T называют пороговым значением соседнего слова (Altschul et al, выше). Эти первоначальные совпадения соседнего слова играют роль затравок для начала поисков с целью обнаружения более длинных HSPs, содержащих их. Совпадения слов затем продлевают в обоих направлениях вдоль каждой последовательности до тех пор, пока суммарный балл выравнивания может увеличиваться. Суммарные баллы вычисляют, используя для нуклеотидных последовательностей параметры M (призовой балл за пару совпадающих остатков, всегда>0) и N (штрафной балл за несовпадающие остатки; всегда<0). Для аминокислотных последовательностей, матрицу подсчета баллов используют для вычисления суммарного значения. Совпадения слов прекращают продлевать в каждом направлении, если: суммарный балл выравнивания снижается на величину X от максимальной достигнутой им величины; суммарный балл опускается до нуля или ниже, из-за накопления одного или более выравниваний остатков с отрицательными баллами или достигается конец любой последовательности. Параметры W, T и X алгоритма BLAST определяют чувствительность и скорость выравнивания. В программе BLASTN (для нуклеотидных последовательностей) используются в качестве параметров по умолчанию длина слова (W), равная 11, ожидание (E), равное 10, M=5, N=-4 и сравнение обеих нитей. Для аминокислотных последовательностей в программе BLASTP используются в качестве параметров по умолчанию длина слова (W), равная 3, ожидание (E), равное 10, и матрица подсчета баллов BLOSUM62 (смотри Henikoff& Henikoff, Proc. Natl. Acad. Sci. USA 89:10915 (1989)). В дополнение к вычислению процентной идентичности последовательностей алгоритм BLAST также выполняет статистический анализ сходства между двумя последовательностями (смотри, например, Karlin & Altschul, Proc. Nat'l. Acad. Sci. USA 90:5873-5787 (1993)). Один критерий сходства, предоставляемый алгоритмом BLAST, представляет собой вероятность наименьшей суммы (P(N)), который является показателем вероятности, с которой совпадение между двумя нуклеотидными или аминокислотными последовательностями произошло бы случайно. Например, нуклеиновую кислоту считают схожей с последовательностью сравнения, если вероятность наименьшей суммы при сравнении тестируемой нуклеиновой кислоты с нуклеиновой кислотой сравнения составляет меньше чем примерно 0,1, более предпочтительно меньше чем примерно 0,01 и наиболее предпочтительно меньше чем примерно 0,001.
Применяя на практике настоящее изобретение, получали не-трансгенное растение или растительную клетку, имеющие одну или несколько мутаций в гене EPSPS. Полученное растение обладает повышенной устойчивостью или толерантностью к члену фосфонометилглициновой группы, такому как глифосат, и демонстрирует по существу нормальный рост или развитие растения, его органов, тканей или клеток в сравнении с соответствующим растением или клеткой дикого типа. Мутантный ген продуцирует генный продукт, имеющий замену в одной или нескольких аминокислотных позициях 160, 179, 183, 194, 244, 273 и 454 продукта AF 360244 гена EPSPS Arabidopsis или в аналогичной аминокислотной позиции в гомологе EPSPS. Предпочтительно мутантное растение является устойчивым к глифосату и обладает по существу такой же каталитической активностью в сравнении с белком EPSPS дикого типа.
Чтобы выявить мутантные гены EPSPS, которые будут продуцировать генный продукт, который придаст устойчивость к глифосату, для экономии времени и ресурсов можно проводить скрининг в бактериальной системе in vitro. Для оценки способности мутантных генов EPSPS придавать устойчивый к глифосату фенотип можно строить кривые роста бактериальных колоний, экспрессирующих мутантные гены-кандидаты EPSPS. Например, в патенте США 6870075 описывают тест на устойчивость к глифосату у Salmonella с использованием мутантных генов EPSPS Arabidopsis, трансформированных в штамм LacZ-Salmonella typhi. В другом варианте осуществления настоящего изобретения можно использовать ген EPSPS E. coli, также называемый AroA ген, чтобы оценить мутанты EPSPS на устойчивость к глифосату. Тесты с построением кривых роста и ферментативные тесты для измерения значений Ki и Km мутантов-кандидатов проводят по хорошо известным методикам тестирования. По выявлении активного устойчивого к глифосату мутанта гена EPSPS E. coli проводят мутацию аналогичной аминокислоты в гене EPSPS растения при помощи рекомбинагенных нуклеооснований, как описано в данном документе, чтобы получить устойчивое к глифосату растение.
Предпочтительные аминокислотные замены в продукте гена EPSPS E. coli (AroA) включают следующие:
Leu82Ser
Thr97Ile или Ala
Pro101Ala или Thr или Leu или Cys или GIy
Val114Ala
Asp164Ala
Asn193Ala и
X374Leu
где аминокислота слева от числа, записанного подстрочным индексом, представляет собой природную аминокислоту, а аминокислота справа от числа, записанного подстрочным индексом, представляет собой мутантную аминокислоту. Буквой «X» обозначена аминокислотная позиция 374 потому, что в продукте гена EPSPS E. coli природная аминокислота представляет собой Leu. Однако было установлено, что у многих видов растений аминокислота в позиции 374 не является Leu, и когда в этой позиции происходит замена на Leu, растение проявляет устойчивость к глифосату и сохраняет достаточную ферментативную активность, чтобы поддерживать нормальный рост растения.
В соответствии с настоящим изобретением производят замены в соответствующих аминокислотных позициях у растительных видов, чтобы получить не-трансгенное устойчивое к гербициду растение. Ниже приведен список некоторых предпочтительных сельскохозяйственных культур, в котором перечислены аминокислотные позиции в гене EPSPS, в которых следует произвести замены. Предпочтительные аминокислотные замены перечислены справа от номера аминокислотной позиции.
Для кукурузы следующие аминокислотные замены являются предпочтительными:
Leu84Ser
Thr102Ile или Ala
Pro106Ala или Thr или Leu или Cys или Gly
Val114Ala
Asp164Ala
Asn193Ala и
X117Leu
Для хлопчатника следующие аминокислотные замены являются предпочтительными:
Leu82Ser
Thr97Ile или Ala
Pro101Ala или Thr или Leu или Cys или Gly
Val114Ala
Asp164Ala
Asn193Ala и
X184Leu
Для риса следующие аминокислотные замены являются предпочтительными:
Leu150Ser
Thr169Ile или Ala
Pro173Ala или Thr или Leu или Cys или Gly
Val114Ala
Asp164Ala
Asn193Ala и
X184Leu
Для Brassica napus (2-28 от трансляции гДНК X51475) следующие аминокислотные замены являются предпочтительными:
Leu155Ser
Thr174Ile или Ala
Pro178Ala или Thr или Leu или Cys или Gly
Val114Ala
Asp164Ala
Asn193Ala и
X189Leu
Для Arabidopsis thaliana (AF360224) следующие аминокислотные замены являются предпочтительными:
Leu160Ser
Thr179Ile или Ala
Pro183Ala или Thr или Leu или Cys или Gly
Val114Ala
Asp164Ala
Asn193Ala и
X194Leu
Для Petunia hybrida следующие аминокислотные замены являются предпочтительными:
Leu155Ser
Thr174Ile или Ala
Pro178Ala или Thr или Leu или Cys или Gly
Val114Ala
Asp164Ala
Asn193Ala и
X189Leu
Как следует принять во внимание, E. coli не является растением, однако ее рассматривают в настоящем изобретении, поскольку ген E. coli можно подвергать мутации в системе культуры бактериальных клеток, а затем продукт мутантного гена E. coli (фермент) можно тестировать на ферментативную активность (Ki и Km), что укажет на устойчивость к глифосату, и функцию, как необходимый ферментативный продукт, который является незаменимым для растений. По выявлении мутанта мутированной E. coli проводят мутацию в растительной клетке, применяя рекомбинагенные олигонуклеооснования, описанные в данном документе, чтобы получить не-трансгенное устойчивое к гербициду растение. По этой причине белки мутантной E. coli и мутантной Area рассматривают как часть настоящего изобретения.
В следующей таблице перечислены предпочтительные позиции для аминокислотных замен по номеру аминокислоты, для разных видов. При осуществлении аминокислотных замен в одной или нескольких из данных позиций будут получены устойчивые к глифосату растения:
Как можно увидеть из приведенной выше таблицы и фигур 1-5, существуют некоторые незначительные вариации среди генов EPSPS между видами и внутри видов. Этого следовало ожидать. Данные незначительные вариации следует принимать в расчет при получении мутантов в соответствии с настоящим изобретением. Аминокислоты на аналогичных позициях в различающихся генах подвергают мутациям для получения устойчивых к глифосату растений. Например, мутация в позиции 179 (T>A) в Arabidopsis AF360224 будет эквивалентна мутации T>A в позиции 178 в Arabidopsis NM 130093. Другой пример касается позиции L82 в гене EPSPS у E. coli. У большинства растений в аналогичной позиции находится L, однако у Arabidopsis находится аналогичная F в позиции 159 или 160 в зависимости от гена Arabidopsis, как указано в приведенной выше таблице.
Кроме того, некоторые растения имеют больше одного гена EPSPS. В таком случае один или несколько генов подвергают мутации в соответствии с настоящим изобретением для получения устойчивого к глифосату мутанта. Если уровни экспрессии различных генов EPSPS известны, и они различаются, тогда предпочтительно подвергать мутации гены EPSPS с более высоким уровнем экспрессии. В предпочтительном варианте осуществления все гены EPSPS в сельскохозяйственной культуре подвергают мутации для получения глифосатного фенотипа. Например, известно, что у канолы имеется четыре гена EPSPS. Два гена представлены на фиг.4 и 5. При сравнении заметна небольшая разница между этими двумя генами.
Растение, подвергнутое мутации в соответствии с настоящим изобретением, может относиться к любому виду двудольных, однодольных или голосемянных растений, включая любой вид древесных растений, который представляет собой дерево или кустарник, любые травянистые виды или любые виды, которые производят съедобные плоды, семена или овощи, либо любые виды, которые производят яркие или ароматные цветы. Например, растение можно выбирать из видов растений группы, состоящей из канолы, подсолнечника, табака, сахарной свеклы, сладкого картофеля, ямса, хлопчатника, кукурузы, пшеницы, ячменя, риса, сорго, помидора, манго, персика, яблони, груши, клубники, банана, дыни, картофеля, моркови, салата, лука, видов сои, сахарного тростника, гороха, арахиса, конских бобов, тополя, винограда, цитрусовых, люцерны, ржи, овса, газонных и кормовых трав, льна, масличных культур, огурца, ипомеи, бальзамина, перца, баклажана, календулы, лотоса, капусты, маргаритки, гвоздики, тюльпана, ириса, лилии и производящих орехи растений, поскольку о них конкретно не упоминалось.
Рекомбинагенные олигонуклеооснования можно вводить в растительную клетку при помощи любого способа, обычно применяемого в данной области, включая, но не ограничиваясь ими, микроносители (биолистическая доставка), микроволокна (усики), электропорацию, прямое поглощение ДНК и микроинъекции.
Иллюстративные примеры рекомбинагенных олигонуклеооснований обсуждаются ниже.
Изобретение можно осуществлять на практике с помощью рекомбинагенных олигонуклеооснований, имеющих конформации и химический состав, описанные в патентах Kmiec I и Kmiec II, которые включены в данное описание посредством ссылок. В Kmiec I сообщают о способе внесения специфических генетических изменений в целевой ген. Рекомбинагенные олигонуклеооснования в Kmiec I и/или Kmiec II содержат две комплементарные нити, одна из которых содержит, по меньшей мере, один сегмент нуклеотидов РНК-типа («РНК-сегмент»), которые спарены с нуклеотидами ДНК-типа другой нити.
В Kmiec II описано, что нуклеотиды можно заменять на не-нуклеотиды, содержащие пуриновые и пиримидиновые основания. В патентах США №№ 5756325; 5871984; 5760012; 5888983; 5795972; 5780296; 5945339; 6004804 и 6010907, а также в международном патенте № PCT/USOO/23457 и в международных патентных публикациях №№ WO 98/49350; WO 99/07865; WO 99/58723; WO 99/58702; WO 99/40789; США 6870075 и публикации патентной заявки США 20030084473, полное содержание которых включено в данное описание посредством ссылок, описывают дополнительные рекомбинагенные молекулы, которые можно использовать по настоящему изобретению. Термин «рекомбинагенные олигонуклеооснования» используют в данном документе для обозначения молекул, которые можно использовать в методах по настоящему изобретению и включает смешанные двунитевые олигонуклеотиды, молекулы, не содержащие нуклеотидов, указанные в Kmiec II; однонитевые олигодезоксинуклеотиды и другие рекомбинагенные молекулы, указанные в вышеперечисленных патентах и патентных публикациях.
В одном варианте осуществления рекомбинагенное олигонуклеооснование представляет собой смешанный двунитевой олигонуклеотид, в котором нуклеотиды РНК-типа смешанного двунитевого олигонуклеотида делают устойчивыми к РНКазе, заменяя 2'-гидроксил на фтористую, хлористую или бромистую функциональную группу, либо помещая заместитель на 2'-O. Подходящие заместители включают заместители, указанные в Kmiec II. Альтернативные заместители включают заместители, указанные в патенте США № 5334711 (Sproat) и заместители, указанные в патентных публикациях EP 629 387 и EP 679 657 (обобщенно, заявки Martin), которые включены в данное описание посредством ссылок. Как используют в данном документе, 2'-фтористое, хлористое или бромистое производное рибонуклеотида или рибонуклеотид, в котором 2'-OH замещен на заместитель, описанный в заявках Martin или Sproat, называется «2'-замещенный рибонуклеотид». Как используют в данном документе, термин «нуклеотид РНК-типа» означает 2'-гидроксильный или 2'-замещенный нуклеотид, который соединен с другими нуклеотидами смешанного двунитевого олигонуклеотида (MDON) незамещенной фосфодиэфирной связью или любой из неприродных связей, указанных в Kmiec I или Kmiec II. Как используют в данном документе, термин «нуклеотид дезоксирибо-типа» означает нуклеотид, где имеется 2'-H, который можно соединять с другими нуклеотидами MDON незамещенной фосфодиэфирной связью или любой из неприродных связей, указанных в Kmiec I или Kmiec II.
В одном варианте осуществления настоящего изобретения рекомбинагенное олигонуклеооснование представляет собой смешанный двунитевой олигонуклеотид, который соединен исключительно незамещенными фосфодиэфирными связями. В альтернативных вариантах осуществления соединение образовано посредством замещенных фосфодиэфиров, производных фосфодиэфира и связями не на основе фосфора, как указано в Kmiec II. В еще одном варианте осуществления каждый нуклеотид РНК-типа в смешанном двунитевом олигонуклеотиде представляет собой 2'-замещенный нуклеотид. Особенно предпочтительными вариантами осуществления 2'-замещенных рибонуклеотидов являются 2'-фтор-, 2'-метокси-, 2'-пропилокси-, 2'-аллилокси-, 2'-гидроксилэтилокси-, 2'-метоксиэтилокси-, 2'-фторпропилокси- и 2'-трифторпропилоксизамещенные рибонуклеотиды. Более предпочтительными вариантами осуществления 2'-замещенных рибонуклеотидов являются 2'-фтор-, 2'-метокси-, 2'-метоксиэтилокси- и 2'-аллилоксизамещенные нуклеотиды. В другом варианте осуществления смешанный двунитевой олигонуклеотид соединен незамещенными фосфодиэфирными связями.
Хотя смешанный двунитевой олигонуклеотид, имеющий только один тип 2'-замещенного нуклеотида РНК-типа, более удобно синтезировать, способы данного изобретения можно осуществлять на практике, используя смешанные двунитевые олигонуклеотиды, имеющие два и более видов нуклеотидов РНК-типа. Функция РНК-сегмента может не нарушаться прерыванием, вызванным вставкой дезоксинуклеотида между двумя тринуклеотидами РНК-типа, соответственно, термин РНК-сегмент охватывает такой «прерванный РНК-сегмент». Непрерывный РНК-сегмент называют сплошным РНК-сегментом. В альтернативном варианте осуществления РНК-сегмент может содержать чередующиеся РНКаза-устойчивые нуклеотиды и нуклеотиды с незамещенным 2'-OH. Смешанные двунитевые олигонуклеотиды предпочтительно содержат меньше 100 нуклеотидов, и более предпочтительно меньше 85 нуклеотидов, однако больше 50 нуклеотидов. Первая и вторая нити спарены по принципу Уотсона-Крика. В одном варианте осуществления нити смешанного двунитевого олигонуклеотида ковалентно связаны линкером, таким как однонитевой гекса-, пента- или тетрануклеотид так, что первая и вторая нити являются сегментами одной олигонуклеотидной цепи, имеющей один 3'- и один 5'-конец. 3'- и 5'-концы можно защитить добавлением «шпилечного кэпа», при помощи которого 3'- и 5'-концевые нуклеотиды образуют пары по принципу Уотсона-Крика с соседними нуклеотидами. Второй шпилечный кэп можно дополнительно поместить на соединение между первой и второй нитями в отдалении от 3'- и 5'-концов так, чтобы стабилизировать спаривание по принципу Уотсона-Крика между первой и второй нитями.
Первая и вторая нити содержат две области, которые гомологичны с двумя фрагментами целевого гена EPSPS, то есть, имеют такую же последовательность, что и целевой ген. Гомологичная область содержит нуклеотиды РНК-сегмента и может содержать один или более нуклеотидов ДНК-типа соединяющего ДНК-сегмента, а также может содержать нуклеотиды ДНК-типа, которые не находятся внутри промежуточного ДНК-сегмента. Две области гомологии разделены, и каждая из них граничит с областью, имеющей последовательность, которая отличается от последовательности целевого гена, называемой «гетерологичной областью». Гетерологичная область может содержать один, два или три несовпадающих нуклеотида. Несовпадающие нуклеотиды могут примыкать друг к другу или, наоборот, могут быть разделены одним или двумя нуклеотидами, которые гомологичны с целевым геном. Альтернативно, гетерологичная область может также содержать вставку из одного, двух, трех или из пяти или менее нуклеотидов. Альтернативно, последовательность смешанного двунитевого олигонуклеотида может отличаться от последовательности целевого гена только делецией одного, двух, трех или пяти или менее нуклеотидов из смешанного двунитевого олигонуклеотида. В таком случае длиной и положением гетерологичной области считается длина делеции, хотя ни один нуклеотид смешанного двунитевого олигонуклеотида не находится внутри гетерологичной области. Расстояние между фрагментами целевого гена, которые являются комплементарными двум гомологичным областям, идентично длине гетерологичной области, если предполагают замену или замены. Если гетерологичная область содержит вставку, гомологичные области тем самым разнесены в смешанном двунитевом олигонуклеотиде на большее расстояние, чем их комплементарные гомологичные фрагменты разнесены в гене, применимо и обратное утверждение, если гетерологичная область кодирует делецию.
Каждый из РНК-сегментов смешанных двунитевых олигонуклеотидов является частью гомологичной области, то есть области, которая идентична по последовательности фрагменту целевого гена, каковые сегменты вместе предпочтительно содержат, по меньшей мере, 13 нуклеотидов РНК-типа и предпочтительно от 16 до 25 нуклеотидов РНК-типа или еще более предпочтительно 18-22 нуклеотидов РНК-типа, или наиболее предпочтительно 20 нуклеотидов. В одном варианте осуществления РНК-сегменты гомологичных областей разделены и граничат с, то есть, «связаны посредством» промежуточного ДНК-сегмента. В одном варианте осуществления каждый нуклеотид гетерологичной области является нуклеотидом промежуточного ДНК-сегмента. Промежуточный ДНК-сегмент, который содержит гетерологичную область смешанного двунитевого олигонуклеотида называется «сегментом-мутатором».
Изменения, которые следует внести в целевой ген EPSPS, кодируются гетерологичной областью. Изменения, которые следует внести в ген EPSPS, могут быть изменениями в одном или нескольких основаниях последовательности гена EPSPS, которые меняют природную аминокислоту в данной позиции на желаемую аминокислоту.
В другом варианте осуществления настоящего изобретения рекомбинагенное олигонуклеооснование представляет собой однонитевой олигодезоксинуклеотидный мутационный вектор, или SSOMV, который описан в международной патентной заявке PCT/US00/23457, полное содержание которой включено в данное описание посредством ссылки. Последовательность SSOMV основана на тех же принципах, что и мутационные векторы, описанные в патентах США №№ 5756325; 5871984; 5760012; 5888983; 5795972; 5780296; 5945339; 6004804 и 6010907, а также в международных публикациях №№ WO 98/49350; WO 99/07865; WO 99/58723; WO 99/58702; WO 99/40789; США 6870075 и публикации патентной заявки США 20030084473. Последовательность SSOMV содержит две области, которые гомологичны целевой последовательности, разделенные областью, которая содержит желаемое генетическое изменение, называемой областью-мутатором. Область-мутатор может иметь последовательность той же длины, что и последовательность, которая разделяет гомологичные области в целевой последовательности, однако отличную от нее последовательность. Такая область-мутатор вызовет замену.
Нуклеотиды SSOMV представляют собой дезоксирибонуклеотиды, которые соединены немодифицированными фосфодиэфирными связями за исключением того, что 3'-концевая и/или 5'-концевая межнуклеотидная связь или, наоборот, две 3'-концевые и/или 5'-концевые межнуклеотидные связи могут представлять собой фосфоротиоат или фосфоамидат. Как используют в данном документе, межнуклеотидная связь представляет собой связь между нуклеотидами в SSOMV и не включает связь между 3'-концевым нуклеотидом или 5'-концевым нуклеотидом и блокирующим заместителем, смотри выше. В определенном варианте осуществления длина SSOMV составляет от 21 до 55 дезоксинуклеотидов и длины гомологичных областей составляют, соответственно, общую длину в, по меньшей мере, 20 дезоксинуклеотидов и по меньшей мере каждая из двух гомологичных областей должна иметь длину по меньшей мере 8 дезоксинуклеотидов.
SSOMV можно конструировать так, чтобы он был комплементарен либо кодирующей, либо некодирующей нити целевого гена. Если желаемая мутация представляет собой замену одного основания, предпочтительно, чтобы оба нуклеотида мутатора были пиримидинами. В тех случаях, когда это соответствует достижению желаемого функционального результата, предпочтительно, чтобы как нуклеотид мутатора, так и целевой нуклеотид в комплементарной нити были пиримидинами. Особенно предпочтительными являются SSOMV, которые кодируют мутации трансверсии, то есть, C или T нуклеотид мутатора неправильно сочетается, соответственно с C или T нуклеотидом в комплементарной нити.
Кроме олигодезоксинуклеотида SSOMV может содержать 5'-блокирующий заместитель, который присоединен к 5'-концевым атомам углерода через линкер. Химический состав линкера не важен, в отличие от его длины, которая предпочтительно должна составлять по меньшей мере 6 атомов, и линкер должен быть гибким. Можно использовать множество нетоксичных заместителей, таких как биотин, холестерин или другие стероиды, либо неинтеркалирующий катионный флуоресцентный краситель. Особенно предпочтительными в качестве реагентов для получения SSOMV являются реагенты, продаваемые под названиями Cy3™ и Cy5™ компанией Glen Research, Sterling VA, которые представляют собой блокированные фосфороамидиты, которые при встраивании в олигонуклеотид образуют красители 3,3,3',3'-тетраметил-N,N'-изопропилзамещенный индомонокарбоцианин и индодикарбоцианин, соответственно. Cy3 является наиболее предпочтительным. Если индокарбоцианин является N-оксиалкилзамещенным, его можно легко соединять с 5'-концом олигодезоксинуклеотида при помощи фосфодиэфира с 5'-концевым фосфатом. Химический состав красителя-линкера между красителем и олигодезоксинуклеотидом не важен и выбирается по принципу удобства синтеза. Если коммерчески доступный фосфорамидит Cy3 используют в соответствии с указаниями, полученная 5'-модификация состоит из блокирующего заместителя и линкера, которые вместе являются N-гидроксипропил-, N'-фосфатидилпропил-3,3,3',3'-тетраметилиндомонокарбоциацином.
В предпочтительном варианте осуществления индокарбоцианиновый краситель является тетразамещенным в 3- и 3'-положениях индольных колец. Без ограничения в части теории данные замещения препятствуют тому, чтобы краситель был интеркалирующим красителем. Природа заместителей в данных положениях не важна. SSOMV может дополнительно иметь 3'-блокирующий заместитель. Опять-таки химический состав 3'-блокирующего заместителя не важен.
В другом предпочтительном варианте осуществления рекомбинагенный олигонуклеотид представляет собой однонитевой олигодезоксинуклеотид, имеющий 3'-концевой нуклеотид, 5'-концевой нуклеотид, содержащий по меньшей мере 25 дезоксинуклеотидов и не более 65 дезоксинуклеотидов и обладающий последовательностью, включающей по меньшей мере две области, каждая из которых состоит по меньшей мере из 8 дезоксинуклеотидов, каждая из которых является, соответственно, идентичной по меньшей мере двум областям целевого хромосомного гена, каковые области вместе составляют по меньшей мере 24 нуклеотида в длину, и каковые области разделены по меньшей мере одним нуклеотидом в последовательности целевого хромосомного гена или в последовательности олигодезоксинуклеотида, или в обеих так, что последовательность олигодезоксинуклеотида не идентична последовательности целевого хромосомного гена. Смотри патент США 6271360, который включен в данное описание посредством ссылки.
Микроносители и микроволокна
Применение металлических микроносителей (микросфер) для введения больших фрагментов ДНК в растительные клетки, имеющие целлюлозные клеточные стенки, путем пробивания частицами хорошо известно специалистам в соответствующей области (далее биолистическая доставка). В патентах США №№ 4945050; 5100792 и 5204253 описывают общую методику выбора микроносителей и устройств для их выталкивания. В патентах США №№ 5484956 и 5489520 описывают получение фертильной трансгенной кукурузы при помощи бомбардировки микрочастицами ткани каллюса кукурузы. Биолистические методики также применяют при трансформации незрелых зародышей кукурузы.
Конкретные условия применения микроносителей в методах настоящего изобретения описаны в международной публикации WO 99/07865. По иллюстративной методике охлажденные до температуры льда микроносители (60 мг/мл), смешанный двунитевой олигонуклеотид (60 мг/мл), 2,5 M CaCl2 и 0,1 M спермидин добавляют в указанном порядке; смесь осторожно перемешивают, например, на вихревой мешалке, в течение 10 минут и оставляют стоять при комнатной температуре в течение 10 минут, после чего микроносители разводят в 5 объемах этанола, центрифугируют и ресуспендируют в 100% этаноле. Хорошие результаты можно получить при концентрации в отвечающем условиям растворе, равной 8-10 мкг/мкл микроносителей, 14-17 мкг/мл смешанного двунитевого олигонуклеотида, 1,1-1,4 M CaCl2 и 18-22 мМ спермидин. Оптимальные результаты наблюдали при следующих условиях: 8 мкг/мкл микроносителей, 16,5 мкг/мл смешанного двунитевого олигонуклеотида, 1,3 M CaCl2 и 21 мМ спермидин.
Рекомбинагенные олигонуклеооснования можно также вводить в растительные клетки для применения на практике настоящего изобретения, используя микроволокна для проникновения через клеточную стенку и клеточную мембрану. В патенте США № 5302523, выданном Coffee et al., описывают использование 30-кратно 0,5 мкм и 10-кратно 0,3 мкм карбидокремниевых волокон для облегчения трансформации суспензионных культур кукурузы сорта мексиканская сладкая черная. Любой механический метод, который можно использовать для введения ДНК с целью трансформации растительной клетки при использовании микроволокон, можно применять для доставки рекомбинагенных олигонуклеооснований с целью получения настоящих мутантов EPSPS. Способ, описанный Coffee et al. в патенте США № 5302523, можно использовать с материалами в виде регенерируемых растительных клеток для введения настоящих рекомбинагенных олигонуклеооснований, чтобы провести мутацию гена EPSPS, в результате чего можно восстановить целое мутантное растение, которое имеет устойчивый к глифосату фенотип.
Иллюстративная методика доставки рекомбинагенного олигонуклеооснования при помощи микроволокон заключается в следующем: стерильные микроволокна (2 мкг) суспендируют в 150 мкл растительной культуральной среды, содержащей примерно 10 мкг смешанного двунитевого олигонуклеотида. Суспензионной культуре дают осесть и равные объемы клеточного осадка и стерильной суспензии смеси волокно/нуклеотид перемешивают на вихревой мешалке в течение 10 минут и высевают на чашки. Селективную среду добавляют сразу же или с отсрочкой вплоть до примерно 120 часов в зависимости от того, что нужно для конкретного признака.
Электропорация
В альтернативном варианте осуществления рекомбинагенные олигонуклеооснования можно доставлять в растительную клетку посредством электропорации протопласта, полученного из части растения, в соответствии с методикой, хорошо известной рядовому специалисту в данной области. Смотри, например, Gallois et al., 1996, in Methods in Molecular Biology 55:89-107, Humana Press, Totowa, N.J.; Kipp et al., 1999, in Methods in Molecular Biology 133:213-221, Humana Press, Totowa, N.J.
Рекомбинагенные олигонуклеооснования можно также вводить в микроспоры посредством электропорации. После высвобождения тетрады микроспора является одноядерной и тонкостенной. Она начинает увеличиваться и образует проростковую пору до того, как образуется экзина. На этой стадии микроспора потенциально более подвержена трансформации экзогенной ДНК, чем другие растительные клетки. Кроме того, развитие микроспоры можно изменять in vitro для получения либо гаплоидных зародышей, либо эмбриогенного каллюса, который можно регенерировать в растения (Coumans et al., Plant Cell Rep. 7:618-621, 1989; Datta et al., Plant Sci. 67:83-88, 1990; Maheshwari et al., Am. J Bot. 69:865-879, 1982; Schaeffer, Adv. In Cell Culture 7:161-182, 1989; Swanson et al., Plant Cell Rep. 6:94-97, 1987). Таким образом, трансформированные микроспоры можно регенерировать непосредственно в гаплоидные растения или дигаплоидные фертильные растения при удвоении хромосом стандартными методами. Смотри также совместно рассматриваемую заявку США серийный № 09/680858, озаглавленную Compositions and Methods for Plant Genetic Modification, включенную в данное описание посредством ссылки.
Электропорацию микроспор можно проводить с любыми видами растений, для которых возможно получить микроспоровую культуру, включая, но не ограничиваясь ими, растения из семейства злаковых, бобовых, крестоцветных, пасленовых, тыквенных, розоцветных, злаковых, лилейных, рутовых, виноградовых, включая такие виды как кукуруза (Zea mays), пшеница (Triticum aestivum), рис (Oryza sativa), овес, ячмень, канола (Brassica napus, Brassica rapa, Brassica oleracea и Brassica juncea), хлопчатник (Gossypium hirsuitum L.), различные виды бобовых (например, соевые бобы [Glycine max], горох [Pisum sativum] и так далее), виноград [Vitis vinifera] и множество других важных сельскохозяйственных культур. Микроспоровый эмбриогенез, как из пыльника, так и из микроспоровой культуры, описан для более чем 170 видов, принадлежащих к 68 родам и 28 семействам двудольных и однодольных (Raghavan, Embryogenesis in Agniosperms: A Developmental and Experimental Study, Cambridge University Press, Cambridge, England, 1986; Rhagavan, Cell Differentiation 21:213-226, 1987; Raemakers et al., Euphytica 81:93-107, 1995). Для подробного обсуждения выделения, культивирования микроспор и регенерации двойных гаплоидных растений из полученных из микроспор зародышей [MDE] Brassica napus L., смотри Nehlin, The Use of Rapeseed (Brassica napus L.) Microspores as a Tool for Biotechnological Applications, doctoral thesis, Swedish University of Agricultural Sciences, Uppsala, Sweden, 1999; также Nehlin et al., Plant Sci. 111:219-227, 1995, и Nehlin et al., Plant Sci. 111:219-227, 1995). Удвоение хромосом из культуры микроспор или пыльника является хорошо разработанной методикой получения двойных гаплоидных гомозиготных растительных линий у различных сельскохозяйственных культур (Heberle-Bors et al., In vitro pollen cultures: Progress and perspectives. In: Pollen Biotechnology. Gene expression and allergen characterization, vol. 85-109, ed. Mohapatra, S. S., and Knox, R. B., Chapman and Hall, New York, 1996).
Способы электропорации микроспор описаны в Jardinaud et al., Plant Sci. 93:177-184, 1993, и Fennell and Hauptman, Plant Cell Reports 11:567-570, 1992. Способы электропорации MDON в растительные протопласты можно также приспособить для использования при электропорации микроспор.
Усики и микроинъекция
В еще одном альтернативном варианте осуществления рекомбинагенное олигонуклеооснование можно доставлять в растительную клетку при помощи усиков или микроинъекции в растительную клетку. Так называемая методика усиков выполняется в основном, как описано в Frame et al., 1994, Plant J. 6:941-948. Рекомбинагенное олигонуклеооснование добавляют к усикам и используют для трансформации растительных клеток. Рекомбинагенное олигонуклеооснование можно совместно инкубировать с плазмидами, содержащими последовательности, кодирующие белки, способные образовывать рекомбиназные комплексы в растительных клетках так, что катализируется рекомбинация между олигонуклеотидом и целевой последовательностью в гене EPSPS.
Селекция устойчивых к глифосату растений
Растения или растительные клетки можно тестировать на устойчивость или толерантность к фосфонометилглициновому гербициду при помощи общеизвестных в данной области методов, например, выращивая растение или растительную клетку в присутствии фосфонометилглицинового гербицида и измеряя скорость роста в сравнении со скоростью роста контрольных растений в отсутствие гербицида. В случае глифосата в селективной среде используют концентрации примерно от 0,01 до 20 мМ.
Следующие примеры иллюстрируют осуществление на практике настоящего изобретения, однако их не следует истолковывать как ограничивающие его объем.
Пример 1: P178A мутанты в Brassica napus (канола)
Получали следующий генопласт (рекомбинагенное олигонуклеооснование) для внесения изменения P178A в зародышевую плазму Brassica napus (канола):
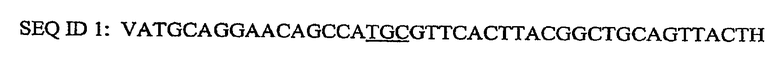
где V представляет собой флуоресцентный краситель (V=Cy3) и H представляет собой обратный нуклеотид или обратное основание (H=3'DMTdCCPG). Подчеркнутые нуклеооснования представляют собой гетерологичную область (кодон), где имеет место мутация в геноме канолы, то есть A. Генопласт получают в соответствии с хорошо известной методикой и генопласт предпочтительно доставляют в растительную клетку канолы посредством бомбардировки микрочастицами, то есть биолистики. Регенерировавшие растения канолы, которые содержат P178A мутант, являются устойчивыми к глифосату, применяемому в коммерческих нормах.
Пример 2: P173A мутанты в Oryza sativa (рис)
Получали следующий генопласт (рекомбинагенное олигонуклеооснование) для внесения изменения P173A в зародышевую плазму Oryza sativa (рис):
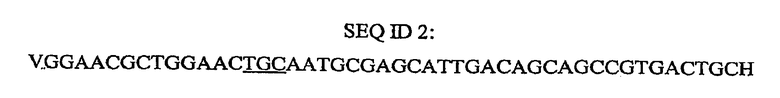
где V представляет собой флуоресцентный краситель (V=Cy3) и H представляет собой обратный нуклеотид или обратное основание (H=3'DMTdCCPG). Подчеркнутые нуклеооснования представляют собой гетерологичную область (кодон), где имеет место мутация в геноме риса, то есть A. Генопласт получают в соответствии с хорошо известной методикой и генопласт предпочтительно доставляют в растительную клетку риса посредством бомбардировки микрочастицами, то есть биолистики. Регенерировавшие растения риса, которые содержат P173A мутант, являются устойчивыми к глифосату, применяемому в коммерческих нормах.
Пример 3: Мутанты E Coli и Arabidposis
В следующей далее таблице перечислены мутации EPSPS в E. coli (Area) и Arabidopsis NM 130093, которые приводят к устойчивому к глифосату фенотипа. Конкретное изменение кодона указано в правой колонке.

* Не истинная гомологичная аминокислота в E. coli. Наиболее близкой гомологичной аминокислотой в E. coli является N111. Также обратите внимание, что в природной E. coli на позиции 82 находится L, а аналогичная аминокислота в Arabodposis на позиции 159 представляет собой F.
В следующем перечне (a-g) настоящие мутации представлены более подробно. Все ссылки на «Arabidopsis» относятся к гену NM 130093 Arabidopsis. Последовательности представляют собой генные последовательности природного гена EPSPS (вверху) и мутантного гена EPSPS (внизу). Мутантный кодон выделен жирным шрифтом и подчеркнут, при этом измененный нуклеотид представлен строчной буквой.







Настоящее изобретение относится к биотехнологии. Описан способ получения не-трансгенного растения, устойчивого или толерантного к гербициду фосфонометилглициновой группы, включающий: введение в растительные клетки рекомбинагенного олигонуклеооснования с направленной мутацией в гене EPSPS для получения растительных клеток с мутантным геном EPSPS, экспрессирующим белок EPSPS, который является мутантным по одной или нескольким аминокислотным позициям, где указанные позиции выбраны из группы, состоящей из Leu160, Thr179, Pro183, Val114, Asp164, Asn193 и X194 в белке EPSPS Arabidopsis (AF360224), или по аналогичному аминокислотному остатку в паралоге EPSPS, где Х не является Leu; отбор растительной клетки, проявляющей повышенную толерантность к глифосату по сравнению с соответствующей растительной клеткой дикого типа; и регенерирование не-трансгенного устойчивого или толерантного к гербициду растения, имеющего мутантный ген EPSPS, из указанной выбранной растительной клетки. Представлено растение, полученное данным способом. Изобретение расширяет арсенал средств для получения растений, устойчивых к гербициду фосфонометилглициновой группы. 6 н. и 27 з.п. ф-лы, 5 ил.
1. Способ получения не-трансгенного растения, устойчивого или толерантного к гербициду фосфонометилглициновой группы, включающий:
введение в растительные клетки рекомбинагенного олигонуклеооснования с направленной мутацией в гене EPSPS для получения растительных клеток с мутантным геном EPSPS, экспрессирующим белок EPSPS, который является мутантным по одной или нескольким аминокислотным позициям, где указанные позиции выбраны из группы, состоящей из Leu160, Thr179, Pro183, Val114, Asp164, Asn193 и X194 в белке EPSPS Arabidopsis (AF360224), или по аналогичному аминокислотному остатку в паралоге EPSPS, где Х не является Leu;
отбор растительной клетки, проявляющей повышенную толерантность к глифосату по сравнению с соответствующей растительной клеткой дикого типа; и
регенерирование не-трансгенного устойчивого или толерантного к гербициду растения, имеющего мутантный ген EPSPS, из указанной выбранной растительной клетки.
2. Способ получения не-трансгенного растения, устойчивого или толерантного к гербициду фосфонометилглициновой группы, включающий:
введение в растительные клетки рекомбинагенного олигонуклеооснования с направленной мутацией в гене EPSPS для получения растительных клеток с мутантным геном EPSPS, экспрессирующим мутантный белок EPSPS, который является мутантным по одной или нескольким аминокислотным позициям, где указанные позиции выбраны из группы, состоящей из Leu160, Thr179, Pro183, Val114, Asp164, Asn193 и X194 в белке EPSPS Arabidopsis (AF360224), или по аналогичному аминокислотному остатку в паралоге EPSPS, где Х не является Leu;
идентификацию растительной клетки, имеющей мутантный белок EPSPS, который проявляет, по существу, такую же каталитическую активность, что и белок EPSPS дикого типа, и который проявляет данную активность даже в присутствии глифосата; и
(с) регенерирование не-трансгенного устойчивого или толерантного к гербициду растения, имеющего мутантный ген EPSPS, из указанной растительной клетки.
3. Способ по п.1 или 2, в котором рекомбинагенное олигонуклеооснование представляет собой смешанный двунитевой нуклеотид или SSMOV.
4. Способ по п.3, в котором смешанный двунитевой нуклеотид содержит первую гомологичную область, которая имеет последовательность, идентичную последовательности из, по меньшей мере, 6 пар оснований первого фрагмента целевого гена EPSPS, и вторую гомологичную область, которая имеет последовательность, идентичную последовательности из, по меньшей мере, 6 пар оснований второго фрагмента целевого гена EPSPS, а также промежуточную область, которая содержит, по меньшей мере, одно нуклеооснование, гетерологичное целевому гену EPSPS, где указанная промежуточная область соединяет первую и вторую гомологичную область.
5. Способ по п.1 или 2, в котором рекомбинагенное олигонуклеооснование вводят посредством электропорации.
6. Способ по п.1, в котором аминокислотные позиции выбраны из группы, состоящей из Leu84, Thr102, Pro106, Val117, Asp164, Asn193 и Х374 в паралоге Zea mays.
7. Способ по п.1, в котором аминокислотные позиции выбраны из группы, состоящей из Leu155, Thr174, Pro178, Val114, Asp164, Asn193 и X189 в паралоге Brassica sp. (X51475.1).
8. Способ по п.1, в котором аминокислотные позиции выбраны из группы, состоящей из Leu155, Thr174, Pro178, Val114, Asp164, Asn193 и X189 в паралоге Petunia hybrida.
9. Способ по п.1 или 2, в котором растительные клетки выбраны из группы, состоящей из кукурузы, пшеницы, риса, ячменя, соевых бобов, хлопчатника, сахарной свеклы, масличных культур, канолы, льна, подсолнечника, картофеля, табака, томата, люцерны, тополя, сосны, эвкалипта, яблони, салата, гороха, чечевицы, винограда, газонных трав и Brassica sp.
10. Способ по п.2, в котором аминокислотные позиции выбраны из группы, состоящей из Leu84, Thr102, Pro106, Val117, Asp164, Asn193 и Х374 в паралоге Zea mays.
11. Способ по п.2, в котором аминокислотные позиции выбраны из группы, состоящей из Leu155, Thr174, Pro178, Val114, Asp164, Asn193 и X189 в паралоге Brassica sp.
12. Способ по п.2, в котором аминокислотные позиции выбраны из группы, состоящей из Leu155, Thr174, Pro178, Val114, Asp164, Asn193 и X189 в паралоге Petunia hybrida.
13. Способ получения не-трансгенного растения, устойчивого или толерантного к гербициду фосфонометилглициновой группы, включающий:
введение в растительные клетки рекомбинагенного олигонуклеооснования с направленной мутацией в гене EPSPS для получения растительных клеток с мутантным геном EPSPS, экспрессирующим белок EPSPS, который является мутантным по аминокислотным позициям Thr179 и Pro183 в белке EPSPS Arabidopsis (AF360224) или по аналогичному аминокислотному остатку в паралоге EPSPS, где Thr179 заменен на Ile и Pro183 заменен на Thr или Ala;
отбор растительной клетки, проявляющей повышенную толерантность к глифосату по сравнению с соответствующей растительной клеткой дикого типа; и
регенерирование не-трансгенного устойчивого или толерантного к гербициду растения, имеющего мутантный ген EPSPS, из указанной отобранной растительной клетки.
14. Способ по п.13, в котором аминокислотные позиции представляют собой Thr102 и Pro106 в паралоге Zea mays.
15. Способ по п.13, в котором аминокислотные позиции представляют собой Thr174 и Pro178 в паралоге Brassica sp.
16. Способ по п.13, в котором аминокислотные позиции представляют собой Thr174 и Pro178 в паралоге Petunia hybrida.
17. Устойчивое к гербициду фосфонометилглициновой группы растение, которое экспрессирует продукт мутантного гена EPSPS, где ген EPSPS мутирован по позиции, чтобы изменить одну или несколько аминокислотных позиций в продукте гена, где указанные аминокислотные позиции выбраны из группы, состоящей из Leu160, Thr179, Pro183, Val114, Asp164, Asn193 и X194 в белке EPSPS Arabidopsis (AF360224), или по аналогичной аминокислотной позиции в гомологе EPSPS, где Х не является Leu.
18. Растение по п.17, где растение представляет собой Zea mays и аминокислотные позиции выбраны из группы, состоящей из Leu84, Thr102, Pro106, Val117, Asp164, Asn193 и Х374.
19. Растение по п.17, где растение представляет собой Brassica napus и аминокислотные позиции выбраны из группы, состоящей из Leu155, Thr174, Pro178, Val114, Asp164, Asn193 и X189.
20. Растение по п.17, где растение представляет собой Petunia hybrida и аминокислотные позиции выбраны из группы, состоящей из Leu155, Thr174, Pro178, Val114, Asp164, Asn193 и X189.
21. Растение по п.17, где растение выбрано из группы, состоящей из кукурузы, пшеницы, риса, ячменя, соевых бобов, хлопчатника, сахарной свеклы, масличных культур, канолы, льна, подсолнечника, картофеля, табака, томата, люцерны, тополя, сосны, эвкалипта, яблони, салата, гороха, чечевицы, винограда и газонных трав.
22. Растение по любому из пп.17-21, в котором мутация гена приводит к одной или нескольким из следующих аминокислотных замен в продукте гена EPSPS по сравнению с последовательностью дикого типа: Leu160Ser, Thr179Ile или Ala, Pro183Ala или Thr или Leu или Cys или Gly, Val114Ala, Asp164Ala, Asn193Ala и X194Leu или по аналогичному аминокислотному остатку в паралоге EPSPS, где аминокислота слева от числа, записанного подстрочным индексом, представляет собой природную аминокислоту, а аминокислота справа от числа, записанного подстрочным индексом, представляет собой мутантную аминокислоту и Х представляет собой любую аминокислоту, за исключением Leu.
23. Способ по п.1, в котором мутация гена приводит к одной или нескольким из следующих аминокислотных замен в продукте гена EPSPS по сравнению с последовательностью дикого типа: Leu160Ser, Thr179Ile или Ala, Pro183Ala или Thr или Leu или Cys или Gly, Val114Ala, Asp164Ala, Asn193Ala и X374Leu или по аналогичному аминокислотному остатку в паралоге EPSPS, где аминокислота слева от числа, записанного подстрочным индексом, представляет собой природную аминокислоту, а аминокислота справа от числа, записанного подстрочным индексом, представляет собой мутантную аминокислоту и Х представляет собой любую аминокислоту за исключением Leu.
24. Способ по п.23, где две аминокислоты являются мутантными.
25. Способ по п.24, где аминокислоты на следующих позициях являются мутантными: (a) Thr179Ile и Pro183Thr или (b) Thr179Ile и Pro183Ala.
26. Способ по п.2, в котором мутация гена приводит к одной или нескольким из следующих аминокислотных замен в продукте гена EPSPS по сравнению с последовательностью дикого типа: Leu160Ser, Thr179Ile или Ala, Pro183Ala или Thr или Leu или Cys или Gly, Val114Ala, Asp164Ala, Asn193Ala и X374Leu, где аминокислота слева от числа, записанного подстрочным индексом, представляет собой природную аминокислоту, а аминокислота справа от числа, записанного подстрочным индексом, представляет собой мутантную аминокислоту и Х представляет собой любую аминокислоту за исключением Leu.
27. Способ по п.26, где две аминокислоты являются мутантными.
28. Способ по п.27, где аминокислоты на следующих позициях являются мутантными: (a) Thr179Ile и Pro183Thr или (b) Thr179Ile и Pro183Ala.
29. Способ получения не-трансгенного растения, устойчивого или толерантного к гербициду фосфонометилглициновой группы, включающий:
введение в растительные клетки рекомбинагенного олигонуклеооснования с направленной мутацией в гене EPSPS для получения растительных клеток с мутантным геном EPSPS, экспрессирующим белок EPSPS, который является мутантным по одной или нескольким аминокислотным позициям, где указанные позиции выбраны из группы, состоящей из Leu160, Thr179, Pro183, Val114, Asp164, Asn193 и X194 в белке EPSPS Arabidopsis (AF360224), или по аналогичному аминокислотному остатку в паралоге EPSPS, где Х не является Leu;
отбор растительной клетки, проявляющей повышенную толерантность к глифосату по сравнению с соответствующей растительной клеткой дикого типа; и
регенерирование не-трансгенного устойчивого или толерантного к гербициду растения, имеющего мутантный ген EPSPS, из указанной выбранной растительной клетки, где указанное растение выбрано из группы, состоящей из кукурузы, пшеницы, сахарной свеклы, масличных культур, канолы, картофеля и Brassica sp.
30. Способ по п.29, в котором мутация гена приводит к одной или нескольким из следующих аминокислотных замен в продукте гена EPSPS по сравнению с последовательностью дикого типа: Leu160Ser, Thr179Ile или Ala, Pro183Ala или Thr или Leu или Cys или Gly, Val114Ala, Asp164Ala, Asn193Ala и X374Leu или по аналогичному аминокислотному остатку в паралоге EPSPS, где аминокислота слева от числа, записанного подстрочным индексом, представляет собой природную аминокислоту, а аминокислота справа от числа, записанного подстрочным индексом, представляет собой мутантную аминокислоту и Х представляет собой любую аминокислоту за исключением Leu.
31. Способ получения не-трансгенного растения, устойчивого или толерантного к гербициду фосфонометилглициновой группы, включающий:
введение в растительные клетки рекомбинагенного олигонуклеооснования с направленной мутацией в гене EPSPS для получения растительных клеток с мутантным геном EPSPS, экспрессирующим белок EPSPS, который является мутантным по аминокислотным позициям Thr179 и Pro183 в белке EPSPS Arabidopsis (AF360224) или по аналогичному аминокислотному остатку в паралоге EPSPS, где Thr179 заменен на Ile и Pro183 заменен на Thr или Ala;
отбор растительной клетки, проявляющей повышенную толерантность к глифосату по сравнению с соответствующей растительной клеткой дикого типа; и
регенерирование не-трансгенного устойчивого или толерантного к гербициду растения, имеющего мутантный ген EPSPS, из указанной отобранной растительной клетки;
где указанное растение выбрано из группы, состоящей из кукурузы, пшеницы, сахарной свеклы, масличных культур, канолы, картофеля и Brassica sp.
32. Устойчивое к гербициду фосфонометилглициновой группы растение, которое экспрессирует продукт мутантного гена EPSPS, где ген EPSPS мутирован по позиции, чтобы изменить одну или несколько аминокислотных позиций в продукте гена, где указанные аминокислотные позиции выбраны из группы, состоящей из Leu160, Thr179, Pro183, Val114, Asp164, Asn193 и X194 в белке EPSPS Arabidopsis (AF360224), или по аналогичной аминокислотной позиции в гомологе EPSPS, где Х не является Leu;
где указанное растение выбрано из группы, состоящей из кукурузы, пшеницы, сахарной свеклы, масличных культур, канолы, картофеля и Brassica sp., и где указанный гербицид представляет собой глифосат.
33. Растение по п.32, в котором мутация гена приводит к одной или нескольким из следующих аминокислотных замен в продукте гена EPSPS по сравнению с последовательностью дикого типа: Leu160Ser, Thr179Ile или Ala, Pro183Ala или Thr или Leu или Cys или Gly, Val114Ala, Asp164Ala, Asn193Ala и X194Leu или по аналогичному аминокислотному остатку в паралоге EPSPS, где аминокислота слева от числа, записанного подстрочным индексом, представляет собой природную аминокислоту, а аминокислота справа от числа, записанного подстрочным индексом, представляет собой мутантную аминокислоту и Х представляет собой любую аминокислоту за исключением Leu.
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| Бесколесный шариковый ход для железнодорожных вагонов | 1917 |
|
SU97A1 |
| ВЫДЕЛЕННЫЙ ПОЛИНУКЛЕОТИД, СПОСОБНЫЙ ПРИДАВАТЬ РАСТЕНИЮ УСТОЙЧИВОСТЬ ИЛИ ТОЛЕРАНТНОСТЬ К ГЛИФОСАТНОМУ ГЕРБИЦИДУ, ВЕКТОР, СПОСОБ ПОЛУЧЕНИЯ РАСТЕНИЙ, ТОЛЕРАНТНЫХ ИЛИ УСТОЙЧИВЫХ К ГЛИФОСАТНОМУ ГЕРБИЦИДУ, СПОСОБ РЕГЕНЕРАЦИИ ТРАНСФОРМИРОВАННОГО РАСТЕНИЯ И СПОСОБ СЕЛЕКТИВНОЙ БОРЬБЫ С СОРНЯКАМИ | 2000 |
|
RU2235778C2 |

