Изобретение относится к биотехнологии, в частности к генной инженерии животных, конкретно, к бинарным системам ДНК-конструкций. Система состоит из двух искусственных ДНК-конструкций, для первой из которых характерна высокая тканеспецифическая активность транскрипционного промотора РНК-полимеразы II типа, с которого осуществляется экспрессия расположенной ниже последовательности гена, кодирующего цитотоксический продукт. Вторая конструкция содержит ген транспозазы ДНК-транспозона под контролем того же промотора и обеспечивает встраивание пары «промотор-ген» из первой конструкции в геномную ДНК. У человека, активность используемого промотора ограничена герминальными клетками (ГК) и соответствующими раковыми клетками и тканями, например, в клетках семиномы или другого вида рака яичка. Настоящая бинарная система может быть использована для генной терапии герминальных видов рака (например, рака яичка).
Экспрессия генов обеспечивается геномными регуляторными элементами, включающими, в том числе, промоторы, энхансоры и сайленсеры различной структуры. Эти регуляторные элементы содержат уникальный набор более коротких мотивов, связывающих регуляторные белки, например, различные белки-факторы транскрипции. Получающийся уникальный репертуар участков связывания регуляторных белков, по-видимому, в основном и обуславливает транскрипционный контроль генов. Некоторые гены транскрибируются во всех тканях организма, некоторые - в нескольких тканях, а некоторые другие - только в единственной ткани. Регуляторные участки генов, характеризующихся такой тканеспецифической транскрипцией, можно использовать для создания генно-инженерных конструкций с целью специфической экспрессии интересующих последовательностей (РНК или белка) в строго определенной ткани (или в группе тканей) организма.
Такая задача может быть поставлена, например, при создании стратегии генной терапии или при решении исследовательских задач. Генно-терапевтические применения могут быть нацелены на восполнение дефицита какого-либо белкового продукта в организме (например, введение в гематопоэтические стволовые клетки пациентов, страдающих наследственными формами гемофилии, конструкций, содержащих экспрессионную конструкцию с нормальной копией гена фактора свертываемости IX (Mates, L., М.К.Chuah. (2009). "Molecular evolution of a novel hyperactive Sleeping Beauty transposase enables robust stable gene transfer in vertebrates." Nat Genet 41(6); 753-61), a также на борьбу с аутоиммунными и раковыми заболеваниями (de Bruin, Е.С. and J.Р.Medema (2008). "Apoptosis and non-apoptotic deaths in cancer development and treatment response." Cancer Treat Rev 34(8): 737-49).
В качестве средств доставки генно-терапевтических препаратов используют липосомы, вирусные частицы, полиплексы (Sobolev, A.S. (2008). "Modular transporters for subcellular cell-specific targeting of anti-tumor drugs." Bioessays 30(3): 278-87), "голая" ДНК или ДНК-липосахаридные комплексы (Yu, Н. and Е.Wagner (2009). "Bioresponsive polymers for nonviral gene delivery." Curr Opin Mol Ther 11(2): 165-78).
В настоящее время, ведется интенсивная разработка препаратов нового поколения, способных селективно воздействовать на раковые клетки с уменьшенной амплитудой побочных эффектов на нормальные ткани организма. Принцип действия таких препаратов основан на знании природы развития того или иного типа опухолей и происходящих в них биохимических процессов, зачастую - с учетом индивидуальных особенностей пациента.
Например, создаются препараты, специфично узнающие и убивающие клетки, на повышенном уровне экспрессирующие те или иные поверхностные антигены (часто -рецепторы клеточных факторов роста), клетки с инактивированными генами онкосупрессоров, клетки с аберрантно блокированными путями апоптоза (Frenzel, A., F.Grespi. (2009). "Вс12 family proteins in carcinogenesis and the treatment of cancer." Apoptosis 14(4): 584-96).
Этот подход предусматривает введение в клетки организма генно-инженерных конструкций, специфично воздействующих на раковые клетки и приводящие к гибели последних. Широко рассматриваются перспективы специфической активации внутриклеточных онкосупрессорных генов (Garcia-Echeverria, С. and W.R.Sellers (2008). "Drug discovery approaches targeting the PI3K/Akt pathway in cancer." Oncogene 27(41):. 5511-26). Многообещающим подходом представляется доставка в клетку "киллерных" генов (или "генов-убийц") под транскрипционным контролем промоторов, обладающих повышенной активностью в раковых клетках. При этом активность таких промоторов может характеризоваться сильной тканеспецифичностью, то есть быть специфичной для определенных типов тканей (соответственно, и определенных видов рака). Это свойство особенно полезно при лечении метастазов рака одной природы в ткани другого происхождения (например, метастазы рака груди в костную ткань (Rabbani, S.A. and А.Р.Mazar (2007). "Evaluating distant metastases in breast cancer: from biology to outcomes." Cancer Metastasis Rev 26(3-4): 663-74)). Чрезвычайно важна "тонкая настройка" системы ген-промотор, которая должна обеспечить безвредность генно-терапевтического препарата для нормальных тканей организма. Иными словами, должно быть исключено "подтекание" промотора на уровне, обеспечивающем экспрессию гена-убийцы на достаточном для терапевтического эффекта уровне в здоровых клетках. Для этого в ряде случаев используемые промоторные последовательности модифицируют, удаляя или вводя в их состав регуляторные последовательности, как правило - сайты узнавания транскрипционных факторов. Это позволяет модулировать тканеспецифичность экспрессии создаваемых генно-инженерных конструкций.
В то же время, ни одна из вышеперечисленных систем не может обеспечить сильной строго тканеспецифической экспрессии эффекторного гена в герминальных клетках (ГК).
Тканеспецифическая экспрессия генов в ГК обусловлена наличием в этом типе клеток ряда уникальных транскрипционных факторов, узнающих особые нуклеотидные мотивы в составе промоторов регулируемых генов. Специфичность действия большего числа таких транскрипционных факторов пока не установлена, но для промоторов значительного числа генов известна их тканеспецифическая активация именно в ГК млекопитающих (Wu, H., Y.Chen. "Sperm associated antigen 8 (SPAG8), a novel regulator of activator of CREM in testis during spermatogenesis." FEBS Lett 584(13): 2807-15). При этом характеристика регуляторных элементов в составе таких промоторов, как правило, не проведена, а точные границы промоторов не определены. Например, для гена человека MAST2, кодирующего особую форму ассоциированной с микротрубочками серин/треониновой киназы, известна ярко выраженная тканеспецифичность его экспрессии в клетках зародышевого пути, но регуляция экспрессии гена не исследована и последовательность промотора функционально не охарактеризована. Известно также, что в ГК активизируются промоторы некоторых повторяющихся элементов генома, в частности, длинные концевые повторы эндогенных ретровирусов человека семейства HERV-K(HML-2) (Buzdin, A. (2007). "Human-specific endogenous retro viruses." Scientific World Joumal 7: 1848-68). Как и в предыдущем случае, диапазон транскрипционных факторов, ответственных за регуляцию этих элементов в ГК, неизвестен.
Ряд промоторов, преимущественно активных в ГК, был запатентован для получения различных тканеспецифичных генетических конструкций. Так, промотор гена сурвивина может быть использован для преимущественной транскрипции генно-инженерных конструкций в клетках рака различной этиологии.
Однако же, такие промоторы имеют существенные недостатки: они или демонстрируют значительную транскрипционную активность и в других типах клеток, нежели ГК, или/и не могут обеспечить высокую транскрипцию нижележащих генов во вводимых конструкциях.
В качестве киллерных генов наибольшей эффективностью характеризуются те, которые обладают эффектом "bystander", то есть воздействия не только на клетки, получившие генно-инженерную конструкцию, но и на соседние клетки. Например, РНКаза онконаза из лягушки Rana pipens не только убивает продуцирующие ее активную форму клетки, но и обладает способностью проникать в соседние клетки и проявлять цитотоксический эффект и в них (Beck, А.К., Н.I.Pass, (2008). "Ranpimase as a potential antitumor ribonuclease treatment for mesothelioma and other malignancies." Future Oncol 4(3): 341-9). Эффект bystander важен исходя из невысоких показателей эффективности попадания генно-терапевтических агентов внутрь клеточного ядра для клеток из некоторых тканей. Таким образом, даже небольшое количество трансфицированных клеток сможет привести к массированной гибели клеток-мишеней в очаге заболевания (Vigna, Е., G.Pacchiana. (2008). ""Active" cancer immunotherapy by anti-Met antibody gene transfer." Cancer Res 68(22): 9176-83).
Вместе с тем, для тонкой настройки действия киллерных генов полезны двухкомпонентные системы, включающие, например, продукт киллерного гена - фермент и его субстрат. Сам по себе такой киллерный ген безвреден для экспрессирующей его клетки, равно как и его субстрат безвреден для организма. А вот уже энзиматическое превращение субстрата приводит к образованию токсических метаболитов, убивающих клетки. Такая система представляется более прогрессивной, поскольку в ней может осуществляться контроль как введения генно-терапевтического агента, так и его субстрата в определенные ткани или участки организма. К таким системам относятся, например, ген тимидинкиназы вируса простого герпеса - ганцикловир (Song, J., С.Kirn. (2009). "Sleeping Beauty-mediated suicide gene therapy of hepatocellular carcinoma." Biosci Biotechnol Biochem 73(1): 165-8), или рекомбинантная цитозиндезаминаза дрожжей - 5-фторцитозин (Kucerova, L., M.Matuskova. (2008). "Cytosine deaminase expressing human mesenchymal stem cells mediated tumour regression in melanoma bearing mice." J Gene Med 10(10); 1071-82).
Однако же, до настоящего изобретения так и не была разработана система, обеспечивающая высокую экспрессию эффекторного цитотоксического гена исключительно в ГК и исключающая экспрессию такого гена в иных клетках организма.
Прототипом настоящего изобретения является модель доставки генетической информации в геномную ДНК с использованием транспозазы ДНК-транспозона (United States Patent 6,489,458; Hackett, et al. December 3,2002; DNA-based transposon system for the introduction of nucleic acid into DNA of a cell). Эта модель использует свойство транспозазы опознавать специфические последовательности инвертированных повторов и осуществлять перенос ограниченных ими фрагментов ДНК в новую локализацию в составе генетического вектора, или в составе геномной ДНК. Модель предлагается использовать для широкого круга задач, в том числе, для генной терапии различных заболеваний. Вместе с тем, эта модель не содержит каких-либо указаний на промотор, обеспечивающий специфичную экспрессию эффекторного цитотоксического гена в ГК, и в модели отсутствует функциональная спецификация самого доставляемого в новую локализацию гена.
Изобретение решает задачу расширения ассортимента средств для генной терапии различных видов злокачественных опухолей, вызванных ростом герминальных клеток, например, рака яичка.
Поставленная задача решается за счет состава, включающего рекомбинантную генно-инженерную конструкцию, содержащую вышележащую промоторную последовательность гена человека NDUFV1 длиной 3574 пары нуклеотидов с удаленной проксимальной последовательностью длиной 91 п.о. и находящийся под ее контролем эффекторный терапевтический цитотоксический ген, а также включающего рекомбинантную генно-инженерную конструкцию, содержащую вышележащую промоторную последовательность гена человека NDUFV1 длиной 3574 пары нуклеотидов с удаленной проксимальной последовательностью длиной 91 п.о. и находящийся под ее транскрипционным контролем ген транспозазы, при молярном соотношении компонентов 100:1-1:100.
В качестве цитотоксического гена конструкция содержит ген тимидинкиназы вируса простого герпеса, рекомбинантный ген бифункциональной цитозиндезаминазы дрожжей, ген онконазы или ген фотоиндуцируемого высокотоксичного флуоресцентного белка Killer Red.
Поставленная задача решается также за счет гена транспозазы, помещенного под транскрипционный контроль промотора гена человека NDUFV1 длиной 3574 пары нуклеотидов с удаленной проксимальной последовательностью длиной 91 п.о., или же какого-либо иного промотора. Использование такого набора конструкций обеспечивает сильный и высокоспецифичный цитотоксический эффект против клеток герминальных опухолей человека и может использоваться для генной терапии рака, например, различных форм рака яичка.
Промотор гена человека NDUFV1 длиной 3574 пары нуклеотидов с удаленной проксимальной последовательностью длиной 91 п.о. создан при помощи амплификации обширного вышележащего контрольного региона гена NDUFV1 человека и делеции одного из его регуляторных участков. Он обладает выраженной специфичностью экспрессии в герминальных клетках, в том числе, в клетках злокачественных опухолей яичка.
Сам по себе, ген человека NDUFV1 (от английского «NADH dehydrogenase (ubiquinone) flavoprotein 1») является геном домашнего хозяйства и кодирует белок, вовлеченный в осуществление клеточного метаболизма в митохондриях (Dieteren, С.Е., Р.Н.Willems, et al. (2008). "Subunits oimitochondrial complex I exist as part of matrix- and membrane-associated subcomplexes in living cells." J Biol Chem 283(50): 34753-61). Как на уровне РНК, так и на белковом уровне, этот ген экспрессируется во всех тканях человека. Транскрипционная регуляция этого гена осуществляется вышележащей (относительно точки начала транскрипции) последовательностью длиной 3665 пар нуклеотидов, включающей в себя промотор и ряд отдаленных регуляторных элементов.
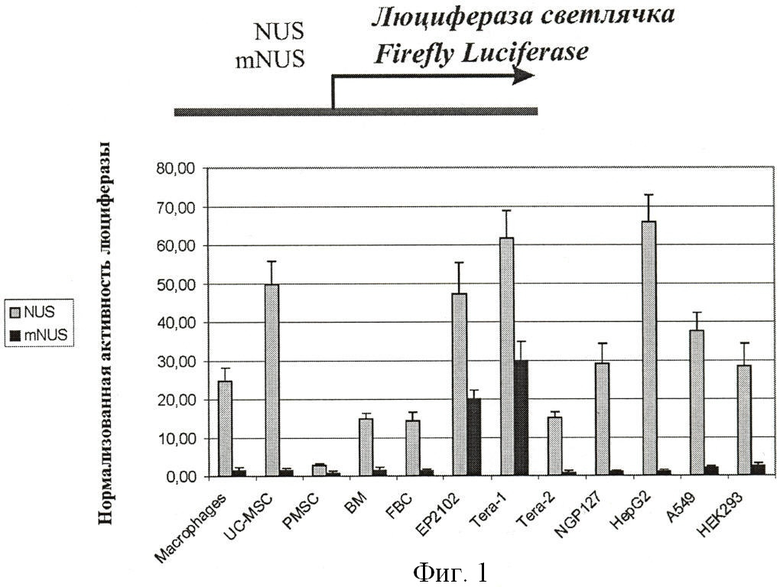
Протестирована промоторная активность вышележащей последовательности NDUFV1 на панели 12 человеческих клеточных культур. Использованы следующие клеточные линии: ЕР2102, Teral (недифференцированный зародышевый рак яичка), Теrа-2 (зародышевый рак яичка, частично дифференцированный в клетки нервной трубки), NGP127 (клетки нейробластомы), HepG2 (гепатокарцинома), А549 (карцинома легкого) и НЕК 293 (эмбриональный рак почки). Кроме того, были использованы первичные человеческие клеточные культуры: макрофаги периферической крови, пуповинные мезенхимальные стволовые клетки (UC-MSC), плацентарные мезенхимальные стволовые клетки (PMSC) и мезенхимальные стволовые клетки костного мозга (BM-MSC), а также трансформированные клетки зародышевого мозга (FBC).
Удаление 91 пары нуклеотидов, располагающихся на самом 3'-конце этой вышележащей последовательности, приводит к резкому падению экспрессии репортерного гена во всех клеточных линиях, кроме линий герминального рака яичка человека Теrа 1 и ЕР-2102 (Фиг.1). Важно, что такая укороченная регуляторная последовательность, являющаяся промотором гена человека NDUFV1 длиной 3574 пары нуклеотидов с удаленной проксимальной последовательностью длиной 91 п.о. (далее в тексте: промотор; первичная нуклеотидная последовательность - SEQ ID NO 1) обеспечивает экспрессию репортерного гена в клеточной линии Теrа 1 на высоком уровне: на уровне, соответствующем стандартному промотору CMV и в 2-2,5 раза превосходящем уровень промотора SV40, тогда как у всех исследованных клеточных культур негерминального происхождения экспрессию детектируют лишь на ничтожно малом уровне (Фиг.2).
Таким образом, полученный промотор можно использовать для сильной тканеспецифической экспрессии генов в клетках герминальных раковых опухолей человека.
В настоящем изобретении промотор осуществляет транскрипционный контроль эффекторного цитотоксического гена. Транскрипцию с промотора осуществляет РНК-полимераза II типа. Поэтому в качестве эффекторного гена, может быть использован любой транскрибируемый РНК-полимеразой II ген, продукт которого обладает выраженным цитотоксическим эффектом.
Например, может быть использован ген высокотоксичного белка онконазы (ранпирназы) из генома лягушки Rana pipens, или ген тимидинкиназы вируса простого герпеса в сочетании с соединением «ганцикловир», или рекомбинантный ген бифункциональной цитозиндезаминазы дрожжей в сочетании с 5-фторцитозином, или ген фотоиндуцируемого высокотоксичного флуоресцентного белка Killer Red. Накопление в клетках продуктов экспрессии эффекторного гена, в зависимости от природы самого используемого терапевтического гена, прямо или косвенно приводит к гибели самих этих клеток, а также, в случае использования эффекта «bystander», и к гибели клеток, расположенных по соседству. В последнем случае, одна раковая клетка, продуцирующая эффекторный белок, убивает также и соседние клетки, что усиливает терапевтический эффект таких конструкций.
Кроме того, генетическая конструкция, содержащая промотор и эффекторный ген, при помощи фермента транспозазы может быть внедрена непосредственно в геном клетки-мишени, что повышает специфичность и амплитуду цитотоксического эффекта. При этом, для введения в клетки берут смесь, включающую вектор, содержащий ген фермента транспозазы, например, транспозазы ДНК-транспозона SleepingBeauty под контролем транскрипционного промотора, и вектор с конструкцией «промотор-эффекторный ген», фланкированной последовательностями инвертированных повторов. Такие инвертированные повторы узнаются транспозазой, которая обеспечивает вырезание конструкции из вводимого вектора и встраивание ее в геномную ДНК клетки (Фиг.3).
Типичным примером работы системы является применение генетической конструкции «промотор - ген тимидинкиназы вируса простого герпеса». Система работает следующим образом: на первом этапе, смесь генетических конструкций доставляется в клетки-мишени в требуемом участке организма. Затем, если конструкции попали в раковые герминальные клетки, происходит экспрессия транспозазы, которую обеспечивает используемый промотор. Транспозаза, в свою очередь, обеспечивает эффективную встройку генетической конструкции в геном клетки. На следующем этапе, в кровь пациенту вводится вещество ганцикловир. Это вещество само по себе безвредно, но продукты его конверсии под действием фермента тимидинкиназы - высокотоксичны как для самой клетки, так и для соседних клеток. Поэтому те клетки, в геноме которых имеется вставка используемой конструкции (то есть раковые герминальные клетки), погибают и убивают соседние клетки, что увеличивает эффективность терапевтического воздействия. Описанный метод терапии безопасен для всех остальных клеток, в которых используемый промотор не активен, и, следовательно, транспозаза и эффекторный ген не нарабатываются.
Помимо транспозазы мобильного элемента SleepingBeauty, может быть также взята транспозаза ДНК-транспозонов PiggyBac, Tol2, Sadhu, и других. В любом случае, генетическую конструкцию «промотор-эффекторный ген» должны фланкировать последовательности соответствующих инвертированных повторов, узнающихся транспозазой используемого типа.
Предложенные в настоящем изобретении рекомбинантные генно-инженерные конструкции можно использовать в качестве генно-терапевтических средств для лечения герминальных опухолей человека, например, большинства видов рака яичка. Ценна высокая специфичность наблюдаемого цитотоксического эффекта предлагаемой системы, поскольку терапевтическая конструкция и соответствующие ей препараты должны селективно убивать раковые клетки заданного типа и не затрагивать другие типы клеток в организме человека.
Изобретение иллюстрируют графические материалы.
Фигура 1. Активность репортерного гена люциферазы светлячка в трансфицированных клеточных линиях. Активность репортерного гена измеряют при помощи метода «двойной люциферазы» и нормируют на активность контрольного вектора pRL-TK (Promega), содержащего промотор тимидинкиназы вируса простого герпеса. По оси абсцисс показана нормализованная активность люциферазы светлячка. Серые столбцы соответствуют трансфекции вектором, содержащим перед геном люциферазы исходный промотор гена NDUFV1 длиной 3665 пары нуклеотидов (NUS); черные столбцы соответствуют трансфекции вектором, содержащим модифицированную регуляторную последовательность гена человека NDUFV1 (mNUS) длиной 3574 пар нуклеотидов. Данные показаны для человеческих клеточных линий следующего происхождения: герминогенные опухоли - недифференцированная (линия Teral) и сильно дифференцированная (Tera2), нейроглиальная опухоль (NGP127), рак печени (HepG2), рак легкого (А549), первичная культура макрофагов, первичная культура плацентарных фибробластов (FBTs), первичная культура фибробластов пуповинной крови (PCF), первичная культура клеток костного мозга (ВМ), рак эмбриональной почки (НЕК293). Для всех линий, кроме НЕК293, трансфекцию проводят в 6 повторностях во время двух независимых экспериментов, для линии НЕК293 - в 4 повторностях во время одного эксперимента.
Фигура 2. Активность репортерного гена люциферазы светлячка в трансфицированных клеточных линиях. По оси абсцисс отложена нормализованная активность люциферазы светлячка. Серые столбцы соответствуют трансфекции вектором, содержащим перед геном люциферазы поздний промотор вируса SV40 (PV); черные столбцы соответствуют трансфекции вектором, содержащим модифицированную регуляторную последовательность гена человека NDUFV1 (mNUS) длиной 3574 пар нуклеотидов. Видно наличие значимой промоторной активности mNUS только в клеточных линиях Теrа-1 и ЕР2102. В этих линиях видна более сильная промоторная активность промотора NDUFV1 по сравнению с промотором вируса SV40.
Фигура 3.
Схема использования системы встройки в геномную ДНК с использованием транспозазы ДНК-транспозона, например, Sleeping Beauty. Транспозаза специфически опознает соответствующие инвертированные повторы и обеспечивает встройку транспозиционной конструкции в геномную ДНК.
Изобретение иллюстрируют примеры.
Пример 1. Создание генно-терапевтической системы с использованием эффекторного гена тимидинкиназы вируса простого герпеса и транспозазы ДНК-транспозона SleepingBeauty
1) Клонирование промотора в вектор, содержащий инвертированные повторы транспозона SleepingBeauty
Промотор гена человека NDUFV1 длиной 3574 пары нуклеотидов с удаленной проксимальной последовательностью длиной 91 п.о. с использованием стандартных методов молекулярного клонирования при использовании эндонуклеаз рестрикции BamHI и BglII, вставляют в вектор рТ2/ВН (Hackett, P.В., S.С.Ekker. (2005). "Sleeping beauty transposon-mediated gene therapy for prolonged expression." Adv Genet 54: 189-232), содержащий последовательности инвертированных повторов транспозона SleepingBeauty, таким образом, чтобы вставка произошла между повторами, на расстоянии не более 500 п.н. от каждого повтора. Наличие вставки проверяют рестрикционным анализом и прямым секвенированием ДНК.
2) Клонирование гена тимидинкиназы вируса простого герпеса под контроль промотора
Ген тимидинкиназы, фланкированный требуемыми рестриктными сайтами, получают при помощи синтеза двуцепочечной ДНК согласно эталонной последовательности (код доступа в GenBank AY426828). Ген тимидинкиназы при помощи эндонуклеаз рестрикции NheI и HaeII клонируют между последовательностями инвертированных повторов SleepingBeauty, под транскрипционный контроль промотора гена человека NDUFV1 длиной 3574 пары нуклеотидов с удаленной проксимальной последовательностью длиной 91 п.о.
3) Клонирование гена транспозазы ДНК-транспозона SleepingBeauty под контроль промотора
Ген транспозазы ДНК-транспозона SleepingBeauty, соответствующий эталонной последовательности в GenBank DD 100289, клонируют в вектор pGL3 basic (Promega) под контроль промотора гена человека NDUFV1 длиной 3574 пары нуклеотидов с удаленной проксимальной последовательностью длиной 91 п.о.
4) Доставка генетических конструкций в клетки
Генетические конструкции в молярном соотношении вектора по п.2) и вектора по п.3) 100:1 доставляют внутрь клеток в клеточных культурах с использованием полиплексных структур (Pathak, A., S.Patnaik. (2009). "Recent trends in non-viral vector-mediated gene delivery." Biotechnol J 4(11): 1559-72). В раковых герминальных клетках, происходит транскрипционная активация гена транспозазы, и конструкция содержащая промотор гена человека NDUFV1 длиной 3574 пары нуклеотидов с удаленной проксимальной последовательностью длиной 91 п.о. и ген тимидинкиназы, стабильно встраивается в геном раковой клетки. Одновременно в раковых клетках нарабатывается и сам фермент тимидинкиназа.
5) Загрузка ганцикловира
Подача ганцикловира (Shashkova, Е.V., L.V.Cherenova. (2005). "Avian adenovirus vector CELO-TK displays anticancer activity in human cancer cells and suppresses established murine melanoma tumors." Cancer Gene Ther 12(7): 617-26) осуществляется непосредственно к клеточным культурам. Количества ганцикловира при введении соответствуют итоговой концентрации этого вещества в среде 250 мкМ. При попадании ганцикловира к клетке, продуцирующей тимидинкиназу вируса простого герпеса, аналогично раковым герминальным клеткам по п.4) настоящего примера, происходит превращение безвредного для клетки ганцикловира в ряд токсических метаболитов, убивающих эту и соседние клетки. Происходит уменьшение количества раковых клеток в опыте на 59-85%, в зависимости от используемой клеточной культуры.
Пример 2. Создание генно-терапевтической системы с использованием эффекторного гена бифункциональной рекомбинантной цитозиндезаминазы дрожжей и транспозазы ДНК-транспозона SleepingBeauty
1) Клонирование промотора в вектор, содержащий инвертированные повторы транспозона SleepingBeauty
Производится аналогично п.1) Примера 1.
2) Клонирование гена бифункциональной рекомбинантной цитозиндизаминазы дрожжей под контроль промотора
Ген бифункциональной рекомбинантной цитозиндизаминазы дрожжей, фланкированный требуемыми рестриктными сайтами, получают при помощи синтеза двуцепочечной ДНК согласно эталонной последовательности по публикации (Graepler, F., М.L.Lemken. (2005). "Bifunctional chimeric SuperCD suicide gene - YCD: YUPRT fusion is highly effective in a rat hepatoma model." World J Gastroenterol 11(44): 6910-9), или из депозитариев известных генетических конструкций. Ген при помощи подходящих рестриктных сайтов клонируют между последовательностями инвертированных повторов SleepingBeauty под транскрипционный контроль промотора гена человека NDUFV1 длиной 3574 пары нуклеотидов с удаленной проксимальной последовательностью длиной 91 п.о.
3) Клонирование гена транспозазы ДНК-транспозона SleepingBeauty под контроль промотора
Производят аналогично п.3) Примера 1.
4) Доставка генетических конструкций в клетки
Производят аналогично п.4) Примера 1, за исключением того, что берут генетические конструкции в молярном соотношении вектора по п.2) и вектора по п.3) 10:1.
5) Загрузка 5-фторцитозина
Подача 5-фторцитозина (Graepler, F., М.L.Lemken. (2005). "Bifunctional chimeric SuperCD suicide gene - YCD: YUPRT fusion is highly effective in a rat hepatoma model." World J Gastroenterol 11(44): 6910-9) осуществляется непосредственно к клеточным культурам, количества 5-фторцитозина при введении соответствуют итоговой концентрации этого вещества в среде 500 мкМ. При попадании 5-фторцитозина к клетке, продуцирующей рекомбинантную цитозиндизаминазу дрожжей, например к раковым герминальным клеткам, происходит превращение безвредного для клетки 5-фторцитозина в ряд токсических метаболитов, убивающих эту и соседние клетки. Происходит уменьшение количества раковых клеток в опыте на 65-90%, в зависимости от используемой клеточной культуры.
Пример 3. Создание генно-терапевтической системы с использованием эффекторного гена бифункциональной рекомбинантной цитозиндизаминазы дрожжей и транспозазы ДНК-транспозона PiggyBac
1) Клонирование промотора в вектор, содержащий инвертированные повторы транспозона PiggyBac
Промотор гена человека NDUFV1 длиной 3574 пары нуклеотидов с удаленной проксимальной последовательностью длиной 91 п.о. с использованием стандартных методов молекулярного клонирования при использовании эндонуклеаз рестрикции KpnI и HindIII, вставляют в вектор рТ2/ВН, содержащий последовательности инвертированных повторов транспозона PiggyBac, таким образом, что вставка происходит между повторами, на расстоянии не более 400 п.н. от каждого повтора. Наличие вставки проверяют рестрикционным анализом и прямым секвенированием ДНК.
2) Клонирование гена бифункциональной рекомбинантной цитозиндизаминазы дрожжей под контроль промотора
При помощи рестриктных сайтов HindIII и EcoRV ген клонируют между последовательностями инвертированных повторов PiggyBac, под транскрипционный контроль промотора гена человека NDUFV1 длиной 3574 пары нуклеотидов с удаленной проксимальной последовательностью длиной 91 п.о.
3) клонирование гена транспозазы ДНК-транспозона PiggyBac под контроль промотора гена человека NDUFV1 длиной 3574 пары нуклеотидов с удаленной проксимальной последовательностью длиной 91 п.о.
Вариант гена транспозазы ДНК-транспозона PiggyBac, соответствующий эталонной последовательности GU937109, клонируют в вектор pGL3(Promega) под контроль промотора гена человека NDUFV1 длиной 3574 пары нуклеотидов с удаленной проксимальной последовательностью длиной 91 п.о., с использованием рестрикционных сайтов MluI и ScaI.
4) Доставка генетических конструкций в клетки
Производят аналогично п.4) Примера 1, за исключением того, что берут генетические конструкции в молярном соотношении вектора по п.2) и вектора по п.3) 1:100.
5) Загрузка 5-фторцитозина
Производится аналогично п.4 примера 2. Наблюдаемый эффект: наблюдается уменьшение количества раковых клеток в опыте на 63-87%, в зависимости от используемой клеточной культуры.
Пример 4. Создание генно-терапевтической системы с использованием эффекторного гена бифункциональной рекомбинантной цитозиндизаминазы дрожжей и транспозазы ДНК-транспозона Тоl2
1) Клонирование промотора в вектор, содержащий инвертированные повторы транспозона Тоl2
Промотор гена человека NDUFVI длиной 3574 пары нуклеотидов с удаленной проксимальной последовательностью длиной 91 п.о., с использованием стандартных методов молекулярного клонирования, при использовании эндонуклеаз рестрикции MluI и NotI, вставляют в вектор рТ2/ВН, содержащий последовательности инвертированных повторов транспозона Тоl2 (Peng, К.С., С.Y.Pan. "Using an improved Тоl2 transposon system to produce transgenic zebrafish with epinecidin-1 which enhanced resistance to bacterial infection." Fish Shellfish Immunol 28(5-6): 905-17), таким образом, что вставка происходит между повторами, на расстоянии не более 350 п.н. от каждого повтора. Наличие вставки проверяют рестрикционным анализом и прямым секвенированием ДНК.
2) Клонирование гена бифункциональной рекомбинантной цитозиндизаминазы дрожжей под контроль промотора гена человека NDUFV1 длиной 3574 пары нуклеотидов с удаленной проксимальной последовательностью длиной 91 п.о.
Ген клонируют между последовательностями инвертированных повторов Тоl2, под транскрипционный контроль промотора с использованием рестрикционных сайтов MluI и ScaI.
3) Клонирование гена транспозазы ДНК-транспозона Тоl2 под контроль промотора гена человека NDUFV1 длиной 3574 пары нуклеотидов с удаленной проксимальной последовательностью длиной 91 п.о.
Вариант гена транспозазы ДНК-транспозона Тоl2, соответствующий последовательности с кодом доступа в GenBank AB032244, клонируют в вектор pTR/t3, под контроль промотора гена человека NDUFV1 длиной 3574 пары нуклеотидов с удаленной проксимальной последовательностью длиной 91 п.о., с использованием рестрикционных сайтов MluI и ScaI.
4) Доставка генетических конструкций в клетки
Производят аналогично п.4) Примера 1.
5) Загрузка 5-фторцитозина
Производят аналогично п.4) Примера 2. Наблюдаемый эффект: наблюдается уменьшение количества раковых клеток в опыте на 58-73%, в зависимости от используемой клеточной культуры.
Пример 5. Создание генно-терапевтической системы с использованием эффекторного гена бифункциональной рекомбинантной цитозиндизаминазы дрожжей и транспозазы ДНК-транспозона Тоl2 под контролем альтернативного тканеспецифичного промотора
1) Клонирование промотора в вектор, содержащий инвертированные повторы транспозона Тоl2
Осуществляют аналогично п.1) Примера 4.
2) Клонирование гена бифункциональной рекомбинантной цитозиндизаминазы дрожжей под контроль промотора
Осуществляют аналогично п.2) Примера 4.
3) Клонирование гена транспозазы ДНК-транспозона Тоl2 под контроль промотора гена MAST2
Ген транспозазы ДНК-транспозона Тоl2 клонируют в вектор pTR/t3, под контроль альтернативного промотора, отличного от промотора гена человека NDUFV1 длиной 3574 пары нуклеотидов с удаленной проксимальной последовательностью длиной 91 п.о. В настоящем примере, в качестве альтернативного промотора используют вышележащую регуляторную последовательность гена MAST2 длиной ~380 п.н., соответствующую координатам (46269185-46269567) по последовательности хромосомы 1 человека, сборка генома от февраля 2009. Этот промотор обеспечивает специфичную экспрессию гена преимущественно в герминальных клетках яичка. Сила этого промотора около двух раз уступает силе промотора SV40. Промотор получают при помощи ПЦР с праймерами, содержащими дополнительно введенные рестриктные сайты HaeI и BglII, необходимые для клонирования промотора в требуемый вектор.
4) Доставка генетических конструкций в клетки
Производят аналогично п.4) Примера 1.
5) Загрузка 5-фторцитозина
Производятся аналогично п.4 примера 2. Наблюдаемый эффект: происходит уменьшение количества раковых клеток в опыте на 34-49%, в зависимости от используемой клеточной культуры.


| название | год | авторы | номер документа |
|---|---|---|---|
| ПРОМОТОР ДЛЯ ТКАНЕСПЕЦИФИЧЕСКОЙ ЭКСПРЕССИИ ГЕНОВ В ГЕРМИНАЛЬНЫХ ТКАНЯХ МЛЕКОПИТАЮЩИХ | 2010 |
|
RU2459870C2 |
| ПОЛИПЕПТИДЫ ТРАНСПОЗАЗЫ И ИХ ПРИМЕНЕНИЕ | 2016 |
|
RU2735700C2 |
| ГЕНЕТИЧЕСКИЕ КОНСТРУКЦИИ ДЛЯ АНТИВИЧ-ТЕРАПИИ | 2010 |
|
RU2426788C1 |
| УНИВЕРСАЛЬНЫЕ РАКОВОСПЕЦИФИЧНЫЕ ПРОМОТОРЫ И ИХ ИСПОЛЬЗОВАНИЕ В ПРОТИВООПУХОЛЕВОЙ ТЕРАПИИ | 2013 |
|
RU2539764C2 |
| УСОВЕРШЕНСТВОВАНИЕ ГЕНЕТИЧЕСКИХ КОНСТРУКЦИЙ ДЛЯ ПОВЫШЕНИЯ ЭФФЕКТИВНОСТИ АНТИВИЧ ТЕРАПИИ | 2013 |
|
RU2533817C1 |
| УЛУЧШЕННЫЕ ЭКСПРЕССИЯ И ПРОЦЕССИНГ ТРАНСГЕНА | 2014 |
|
RU2808756C1 |
| Рекомбинантные плазмидные ДНК lenti sgRNA(MS2)_zeo_Myc, обеспечивающие активацию экспрессии гена MYC в клетках человека, способ получения клеток человека со стабильно повышенной экспрессией гена MYC и моноклональная линия клеток рака молочной железы человека BT549_Myc со стабильно повышенной экспрессией гена MYC | 2022 |
|
RU2812975C1 |
| СПОСОБ ПОЛУЧЕНИЯ БЕЛКА | 2010 |
|
RU2563514C2 |
| ПОЛУЧЕНИЕ ВИРУСНОГО ВЕКТОРА | 2014 |
|
RU2658487C2 |
| В-КЛЕТКИ ДЛЯ ДОСТАВКИ ТЕРАПЕВТИЧЕСКИХ СРЕДСТВ IN VIVO И ИХ ДОЗЫ | 2018 |
|
RU2812477C2 |

Изобретение относится к области биотехнологии, конкретно к бинарным системам ДНК-конструкций, и может быть использовано для генной терапии герминальных видов рака (например, рака яичка). В состав для генной терапии злокачественных опухолей входит рекомбинантная генно-инженерная конструкция, содержащая вышележащую промоторную последовательность гена человека NDUFV1 длиной 3574 пары нуклеотидов с удаленной проксимальной последовательностью длиной 91 п.о. и находящийся под ее контролем эффекторный терапевтический цитотоксический ген, а также рекомбинантная генно-инженерная конструкция, содержащая вышележащую промоторную последовательность гена человека NDUFV1 длиной 3574 пары нуклеотидов с удаленной проксимальной последовательностью длиной 91 п.о. и находящийся под ее транскрипционным контролем ген транспозазы, при молярном соотношении компонентов 100:1-1:100. Изобретение позволяет расширить ассортимент средств для генной терапии различных видов злокачественных опухолей, вызванных ростом герминальных клеток. 3 ил., 5 пр.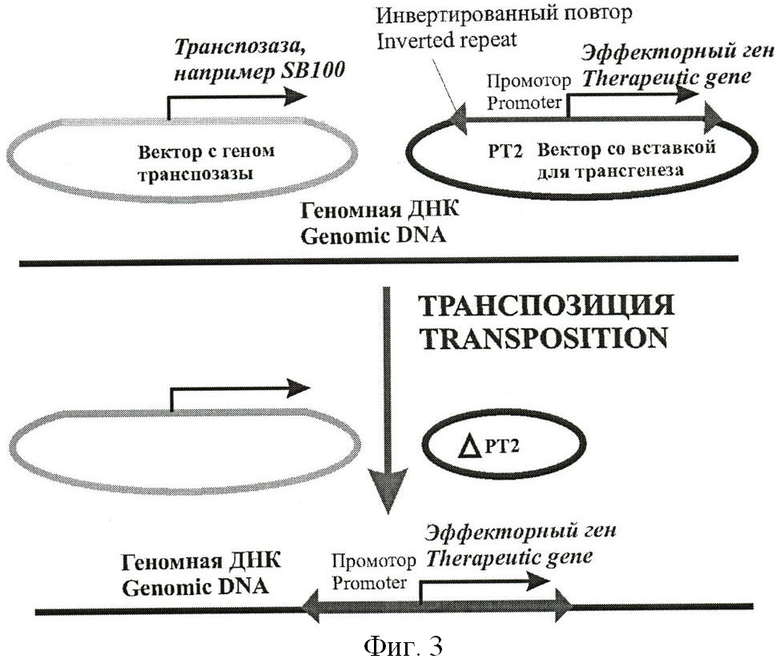
Состав для генной терапии злокачественных опухолей, включающий рекомбинантную генно-инженерную конструкцию, содержащую вышележащую промоторную последовательность гена человека NDLJFV1 длиной 3574 пары нуклеотидов с удаленной проксимальной последовательностью длиной 91 п.о. и находящийся под ее контролем эффекторный терапевтический цитотоксический ген, а также включающий рекомбинантную генно-инженерную конструкцию, содержащую вышележащую промоторную последовательность гена человека NDUFV1 длиной 3574 пары нуклеотидов с удаленной проксимальной последовательностью длиной 91 п.о. и находящийся под ее транскрипционным контролем ген транспозазы, при молярном соотношении компонентов 100:1-1:100.
| US 6489458 B2, 03.12.2002 | |||
| MÄÄTTÄ A.M | |||
| et al | |||
| Transcriptional targeting of virus-mediated gene transfer by the human hexokinase II promoter | |||
| Int | |||
| J | |||
| Mol | |||
| Med., 2006, v.18, n.5, p.901-908 | |||
| МОДИФИЦИРОВАННЫЕ РЕКОМБИНАНТНЫЕ ВАКЦИНИРУЮЩИЕ ВИРУСЫ И ДРУГИЕ МИКРООРГАНИЗМЫ И ИХ ПРИМЕНЕНИЕ | 2004 |
|
RU2376371C2 |

