Область техники изобретения
Данное изобретение относится к способу получения представляющего интерес белка, включающему введение вектора экспрессии белка, который включает генный фрагмент, содержащий ДНК, кодирующую представляющий интерес белок, и ген селектируемого маркера, а также транспозонные последовательности на обоих концах генного фрагмента, в суспензионную клетку млекопитающего; интегрирование генного фрагмента, вставленного между парой транспозонных последовательностей, в хромосому клетки млекопитающего для получения клетки млекопитающего, способной экспрессировать представляющий интерес белок; и суспензионное культивирование клетки млекопитающего; при этом суспензионная клетка млекопитающего способна экспрессировать представляющий интерес белок.
Уровень техники изобретения
Получение экзогенных белков методами рекомбинантной ДНК используется в различных отраслях промышленности, таких как фармацевтическая и пищевая промышленность. В большинстве случаев получение рекомбинантных белков осуществляют путем введения вектора экспрессии, содержащего нуклеотидную последовательность, кодирующую представляющий интерес белок, в хозяина, такого как Escherichia coli, дрожжи, клетка насекомого, растительная клетка и животная клетка, отбора трансформанта, в котором вектор экспрессии интегрирован в хромосому, с последующим культивированием клеточной линии в соответствующих условиях культивирования.
Однако для создания хозяина, способного эффективно продуцировать экзогенный белок, необходимо выбрать клетку-хозяина, обладающую хорошей производительностью для каждого представляющего интерес белка, вследствие этого необходимы дальнейшие технические инновации в области методов производства экзогенных белков в отдельных хозяевах.
В бактериальных системах, таких как Escherichia coli, и дрожжевых системах, отличающихся от животных клеток, во многих случаях трудно добиться пост-трансляционных модификаций, таких как модификация сахарных цепей, в результате чего возникает проблема с получением белка, обладающего активностью.
Поскольку в системе клеток насекомых полученный белок подвергается пост-трансляционной модификации, такой как фосфорилирование и добавление сахарных цепей, такая система имеет то преимущество, что может экспрессироваться белок, обладающий присущей ему физиологической активностью. Однако, поскольку структура сахарной цепи секретируемого белка отличается от таковой в клетках млекопитающих, антигенность и тому подобное становится проблемой, если белок используют в фармацевтических целях.
Кроме того, поскольку при введении экзогенного гена в системе клеток насекомых используют рекомбинантный вирус, существует проблема его инактивации, и сдерживание вируса необходимо с точки зрения безопасности.
В системе животных клеток пост-трансляционные модификации, такие как фосфорилирование, добавление сахарных цепей и сворачивание, можно осуществлять с белками, происходящими из высших животных, включая человека, более простым способом, чем при их продукции в живом организме. Такие точные пост-трансляционные изменения необходимы для воссоздания в этом рекомбинантном белке физиологической активности, изначально присущей белку, и система продукции белка, в которой используют клетку млекопитающего в качестве хозяина, обычно применяется для фармацевтической продукции и тому подобного, где необходима такая физиологическая активность.
Однако система экспрессии белков, в которой используют в качестве хозяина клетку млекопитающего, как правило, отличается низкой производительностью, и во многих случаях существует проблема стабильности введенных генов. Повышение уровня продукции белка при использовании культуры клеток млекопитающих в качестве хозяев не только имеет большую важность для производства лекарственных средств, диагностических реагентов и тому подобного, но также вносит значительный вклад в их исследование и разработку. Таким образом, необходимо в срочном порядке разработать систему экспрессии генов, которая легко позволяет получать клеточную линию с высокой продуктивностью при использовании культуры клеток млекопитающего, в частности, клеток яичника китайского хомячка (клетки CHO), в качестве хозяина.
Транспозон представляет собой мобильный генетический элемент, который может перемещаться из одного локуса в другой локус на хромосоме. Транспозон является мощным инструментом исследования в области молекулярной биологии и генетики и используется для таких целей, как мутагенез, захват генов и получение трансгенных организмов, у насекомых или нематод (например, Drosophila melanogaster или Caenorhabditis elegans) и растений. Однако разработка такой методики для позвоночных животных, включая клетки млекопитающих, отстает.
Тем не менее, в последнее время сообщали о транспозонах, обладающих активностью также у позвоночных животных, и, как показано, некоторые из них обладают активностью в клетках млекопитающих, например, в клетках мыши и человека. Типичные примеры включают транспозоны Tol1 (патентная ссылка 1) и Tol2 (непатентная ссылка 1), клонированные из оризии (рыба семейства карпозубых), транспозон «спящая красавица», восстановленный из неавтономного транспозона, существующего в геноме рыбы Onchorhynchus (непатентная ссылка 2), искусственный транспозон «лягушачий принц» (непатентная ссылка 3) из лягушки и транспозон piggyBac (непатентная ссылка 4) из насекомых.
Эти ДНК-транспозоны использовали для мутагенеза, захвата генов, получения трансгенных организмов, экспрессии белков устойчивости к лекарственным средствам, и тому подобного, в качестве инструмента переноса генов для привнесения нового фенотипа в геном клетки млекопитающего (непатентные ссылки 5-12).
В случае насекомых изучен метод, в котором экзогенный ген вводят в хромосому шелкопряда при помощи транспозона piggyBac из чешуекрылого насекомого для экспрессии белка, кодируемого указанным экзогенным геном, и описан способ продукции белка с использованием вышеуказанного метода (патентная ссылка 2).
Однако, поскольку уровень экспрессии экспрессируемого представляющего интерес белка недостаточен и он продуцируется во всем организме шелкопряда, возникает экономическая проблема из-за необходимости передовой технологии очистки для извлечения экспрессированного экзогенного белка в высокоочищенном виде из жидкости организма, содержащей большое количество посторонних белков.
Кроме того, известен пример, когда белок, связанный с устойчивостью к G418, экспрессировали в клетке млекопитающего при помощи транспозона Tol2 из оризии (непатентная ссылка 12).
Список литературы
Патентная литература
Патентная литература 1 WO2008/072540
Патентная литература 2 Японская опубликованная нерассмотренная патентная заявка № 2001-532188
Непатентная литература
Непатентная литература 1 Nature, 383, 30 (1996)
Непатентная литература 2 Cell, 91, 501-510 (1997)
Непатентная литература 3 Nucleic Acids Res, 31, 6873-6881 (2003)
Непатентная литература 4 Insect Mol. Biol., 5, 141-151 (1996)
Непатентная литература 5 Genetics, 166, 895-899 (2004)
Непатентная литература 6 PLoS Genet, 2, e169 (2006)
Непатентная литература 7 Proc. Natl Acad. Sci. USA 95, 10769-10773 (1998)
Непатентная литература 8 Proc. Natl. Acad Sci. USA 98, 6759-6764 (2001)
Непатентная литература 9 Nature 436, 221-226 (2005)
Непатентная литература 10 Nucleic Acids Res., 31, 6873-6881 (2003)
Непатентная литература 11 Nucleic Acids Res., 35, e87 (2007)
Непатентная литература 12 Proc. Natl. Acad. Sci. USA, 103, 15008-15013 (2006).
Сущность изобретения
Техническая проблема
Для получения и анализа представляющего интерес белка необходимо выбрать клеточную линию, которая стабильно и на высоком уровне экспрессирует представляющий интерес белок, используя культуру клеток млекопитающего, однако получение и культивирование клетки, продуцирующей представляющий интерес белок, потребует значительных затрат труда и времени.
Кроме того, хотя известно, что представляющий интерес белок экспрессируется в клетке млекопитающего при помощи транспозонной последовательности, получение клетки, которая способна на высоком уровне экспрессировать представляющий интерес белок и, следовательно, может быть использована в качестве системы продукции белка при помощи транспозонной последовательности; способ получения клетки млекопитающего, которая способна на высоком уровне продуцировать представляющий интерес белок при помощи транспозонной последовательности; а также способ получения белка при помощи такой клетки остаются неизвестными.
Как описано выше, существует потребность в экспрессии представляющего интерес белка в больших количествах путем создания системы продукции белка, способной эффективно и за короткое время продуцировать на высоком уровне представляющий интерес белок с использованием культуры клеток млекопитающего. Таким образом, целью изобретения является получение клетки, способной на высоком уровне экспрессировать представляющий интерес белок, которую можно эффективно переводить в культуру, а также способ получения представляющего интерес белка с помощью такой клетки.
Способ решения проблем
Для решения вышеуказанных проблем авторы настоящего изобретения провели интенсивные исследования и в результате обнаружили, что клетку млекопитающего, способную на высоком уровне экспрессировать представляющий интерес белок, можно эффективно получать путем введения вектора экспрессии белка, включающего генный фрагмент, содержащий ДНК, кодирующую представляющий интерес белок, и ген селектируемого маркера, а также транспозонные последовательности на обоих концах генного фрагмента, в суспензионную клетку млекопитающего; и интегрирования генного фрагмента, вставленного между парой (двумя) транспозонных последовательностей, в хромосому клетки млекопитающего. Кроме того, было установлено, что представляющий интерес белок можно эффективно продуцировать при помощи такой клетки, и, таким образом, было совершено изобретение.
Подробное описание изобретения
В частности, изобретение состоит в следующем:
1. Способ получения представляющего интерес белка, включающий введение вектора экспрессии белка, который включает генный фрагмент, содержащий ДНК, кодирующую представляющий интерес белок, и ген селектируемого маркера, а также транспозонные последовательности на обоих концах генного фрагмента, в суспензионную клетку млекопитающего; интегрирование генного фрагмента, вставленного между парой транспозонных последовательностей, в хромосому клетки млекопитающего для получения клетки млекопитающего, способной экспрессировать представляющий интерес белок; и суспензионное культивирование клетки млекопитающего;
2. Способ получения представляющего интерес белка, включающий следующие этапы от (A) до (C):
(A) этап одновременного введения следующих векторов экспрессии (a) и (b) в суспензионную клетку млекопитающего:
(a) вектор экспрессии, включающий генный фрагмент, содержащий ДНК, кодирующую представляющий интерес белок, и транспозонные последовательности на обоих концах генного фрагмента,
(b) вектор экспрессии, содержащий ДНК, кодирующую транспозазу, которая распознает транспозонные последовательности и обладает активностью переноса генного фрагмента, вставленного между парой транспозонных последовательностей, в хромосому,
(B) этап временной экспрессии транспозазы с вектора экспрессии, введенного на этапе (A), для интегрирования генного фрагмента, вставленного между парой транспозонных последовательностей, в хромосому клетки млекопитающего для получения суспензионной клетки млекопитающего, способной экспрессировать представляющий интерес белок, и
(C) этап суспензионного культивирования суспензионной клетки млекопитающего, способной экспрессировать представляющий интерес белок, полученной на этапе (B), для получения представляющего интерес белка;
3. Способ получения суспензионной клетки млекопитающего, способной экспрессировать представляющий интерес белок, включающий введение вектора экспрессии белка, который включает генный фрагмент, содержащий ДНК, кодирующую представляющий интерес белок, и ген селектируемого маркера, а также транспозонные последовательности на обоих концах генного фрагмента, в суспензионную клетку млекопитающего; и интегрирование генного фрагмента, вставленного между парой транспозонных последовательностей, в хромосому клетки млекопитающего;
4. Способ, описанный в любом из вышеуказанных пунктов 1-3, в котором суспензионная клетка млекопитающего представляет собой клетку, способную выживать и пролиферировать в бессывороточной среде;
5. Способ, описанный в любом из вышеуказанных пунктов 1-4, в котором суспензионная клетка млекопитающего представляет собой по меньшей мере одну клетку, выбранную из суспензионной клетки CHO, адаптированной к суспензионному культивированию, клетки PER.C6, клетки крысиной миеломы YB2/3HL.P2.G11.16Ag.20 (или также называемой YB2/0) и суспензионной клетки мышиной миеломы NS0, адаптированной к суспензионному культивированию;
6. Способ, описанный в вышеуказанном пункте 5, в котором клетка CHO представляет собой по меньшей мере одну клетку, выбранную из CHO-K1, CHO-K1SV, DUKXB11, CHO/DG44, Pro-3 и CHO-S;
7. Способ, описанный в любом из вышеуказанных пунктов 1-6, в котором ген селектируемого маркера представляет собой ген устойчивости к циклогексимиду;
8. Способ, описанный в вышеуказанном пункте 7, в котором ген устойчивости к циклогексимиду представляет собой ген, кодирующий мутант человеческого рибосомного белка L36a;
9. Способ, описанный в вышеуказанном пункте 8, в котором мутант представляет собой мутант, в котором пролин в положении 54 человеческого рибосомного белка L36a заменен на другую аминокислоту;
10. Способ, описанный в вышеуказанном пункте 9, в котором другая аминокислота представляет собой глутамин;
11. Способ, описанный в любом из вышеуказанных пунктов 1-10, в котором пара транспозонных последовательностей представляет собой нуклеотидные последовательности, происходящие из пары транспозонов ДНК-типа, функционирующих в клетке млекопитающего;
12. Способ, описанный в вышеуказанном пункте 11, в котором нуклеотидные последовательности, происходящие из пары транспозонов ДНК-типа, представляют собой нуклеотидные последовательности, происходящие из пары транспозонов Tol1, или нуклеотидные последовательности, происходящие из пары транспозонов Tol2;
13. Способ, описанный в вышеуказанном пункте 12, в котором нуклеотидные последовательности, происходящие из пары транспозонов Tol2, представляют собой нуклеотидную последовательность, содержащую нуклеотидную последовательность, приведенную в SEQ ID NO: 2, и нуклеотидную последовательность, приведенную в SEQ ID NO: 3;
14. Способ, описанный в вышеуказанном пункте 12, в котором нуклеотидные последовательности, происходящие из пары транспозонов Tol1, представляют собой нуклеотидную последовательность, приведенную в SEQ ID NO: 14, и нуклеотидную последовательность, приведенную в SEQ ID NO: 15;
15. Суспензионная клетка млекопитающего, способная продуцировать представляющий интерес белок, в которую введен вектор экспрессии белка, включающий генный фрагмент, содержащий ДНК, кодирующую представляющий интерес белок, и ген селектируемого маркера, а также транспозонные последовательности на обоих концах генного фрагмента, для интегрирования генного фрагмента, вставленного между парой транспозонных последовательностей, в хромосому;
16. Суспензионная клетка млекопитающего, способная продуцировать представляющий интерес белок, в которую введен вектор экспрессии (a), включающий генный фрагмент, содержащий ДНК, кодирующую представляющий интерес белок, и ген селектируемого маркера, а также транспозонные последовательности на обоих концах генного фрагмента, и вектор экспрессии (b), содержащий ДНК, кодирующую транспозазу (трансферазу), которая распознает транспозонные последовательности и обладает активностью переноса генного фрагмента, вставленного между парой транспозонных последовательностей, в хромосому для интегрирования генного фрагмента, вставленного между парой транспозонных последовательностей, в хромосому;
17. Клетка, описанная в вышеуказанном пункте 15 или 16, способная выживать и пролиферировать в бессывороточной среде;
18. Клетка, описанная в любом из вышеуказанных пунктов 15-17, представляющая собой по меньшей мере одну суспензионную клетку млекопитающего, выбранную из суспензионной клетки CHO, при этом клетка CHO адаптирована к суспензионному культивированию, клетки PER.C6, клетки крысиной миеломы YB2/3HL.P2.G11.16Ag.20 (или также называемой YB2/0) и суспензионной клетки мышиной миеломы NS0, адаптированной к суспензионному культивированию;
19. Клетка, описанная в вышеуказанном пункте 18, где клетка СНО представляет собой по меньшей мере одну клетку, выбранную из CHO-K1, CHO-K1SV, DUKXB11, CHO/DG44, Pro-3 и CHO-S;
20. Клетка, описанная в любом из вышеуказанных пунктов 15-19, в которой ген селектируемого маркера представляет собой ген устойчивости к циклогексимиду;
21. Клетка, описанная в вышеуказанном пункте 20, в которой ген устойчивости к циклогексимиду представляет собой ген, кодирующий мутант человеческого рибосомного белка L36a;
22. Клетка, описанная в вышеуказанном пункте 21, в которой мутант представляет собой мутант, в котором пролин в положении 54 человеческого рибосомного белка L36a заменен на другую аминокислоту;
23. Клетка, описанная в вышеуказанном пункте 22, в которой другая аминокислота представляет собой глутамин;
24. Клетка, описанная в любом из вышеуказанных пунктов 15-23, в которой пара транспозонных последовательностей представляет собой нуклеотидные последовательности, происходящие из пары транспозонов ДНК-типа, функционирующих в клетке млекопитающего;
25. Клетка, описанная в вышеуказанном пункте 24, в которой нуклеотидные последовательности, происходящие из пары транспозонов ДНК-типа, представляют собой нуклеотидные последовательности, происходящие из пары транспозонов Tol1, или нуклеотидные последовательности, происходящие из пары транспозонов Tol2;
26. Клетка, описанная в вышеуказанном пункте 25, в которой нуклеотидные последовательности, происходящие из пары транспозонов Tol2, представляют собой нуклеотидную последовательность, приведенную в SEQ ID NO: 2, и нуклеотидную последовательность, приведенную в SEQ ID NO: 3;
27. Клетка, описанная в вышеуказанном пункте 25, в которой нуклеотидные последовательности, происходящие из пары транспозонов Tol1, представляют собой нуклеотидную последовательность, приведенную в SEQ ID NO: 14, и нуклеотидную последовательность, приведенную в SEQ ID NO: 15;
28. Вектор экспрессии белка, включающий генный фрагмент, содержащий ДНК, кодирующую представляющий интерес белок, и ген селектируемого маркера, а также транспозонные последовательности на обоих концах генного фрагмента;
29. Вектор экспрессии белка, описанный в вышеуказанном пункте 28, в котором пара транспозонных последовательностей представляет собой нуклеотидные последовательности, происходящие из пары транспозонов Tol1, или нуклеотидные последовательности, происходящие из пары транспозонов Tol2;
30. Вектор экспрессии белка, описанный в вышеуказанном пункте 29, в котором нуклеотидные последовательности, происходящие из пары транспозонов Tol2, представляют собой нуклеотидную последовательность, приведенную в SEQ ID NO: 2, и нуклеотидную последовательность, приведенную в SEQ ID NO: 3; и
31. Вектор экспрессии белка, описанный в вышеуказанном пункте 29, в котором нуклеотидные последовательности, происходящие из пары транспозонов Tol1, представляют собой нуклеотидную последовательность, приведенную в SEQ ID NO: 14, и нуклеотидную последовательность, приведенную в SEQ ID NO: 15.
Полезные эффекты изобретения
В соответствии со способом получения белка по изобретению можно эффективно продуцировать представляющий интерес белок, используя клетку млекопитающего. Кроме того, клетку по изобретению можно использовать в качестве продуцирующей белок клетки для производства рекомбинантного белка с высокой эффективностью.
Краткое описание чертежей
На фигуре 1 представлено схематическое изображение транспозонного вектора для экспрессии антитела против белка M2 вируса гриппа человека. Tol2-L представляет собой левый конец транспозона Tol2 (SEQ ID NO: 2), Tol2-R представляет собой правый конец транспозона Tol2 (SEQ ID NO: 3), CMV представляет собой промотор CMV, поли-A представляет собой сайт полиаденилирования, Hc представляет собой кДНК H-цепи антитела человека, Lc представляет собой кДНК L-цепи антитела человека, и CHX-r представляет собой ген устойчивости к циклогексимиду.
На фигуре 2 представлено схематическое изображение вектора экспрессии антитела против белка M2 вируса гриппа человека. CMV представляет собой промотор CMV, поли-A представляет собой сайт полиаденилирования, Hc представляет собой кДНК H-цепи антитела человека, Lc представляет собой кДНК L-цепи антитела человека, и CHX-r представляет собой ген устойчивости к циклогексимиду.
На фигуре 3 представлено схематическое изображение вектора экспрессии транспозазы Tol2. CAGGS представляет собой промотор CAGGS, поли-A представляет собой сайт полиаденилирования, и кДНК TPазы представляет собой кДНК транспозазы Tol2.
На фигуре 4A представлен результат изучения уровня экспрессии антитела против белка M2 вируса гриппа человека в суспензионной клетке CHO-K1 при использовании транспозонного вектора Tol2 для экспрессии антитела против белка M2 вируса гриппа человека. Ось ординат показывает уровень продукции антитела (мкг/мл), а ось абсцисс показывает число трансгенных клонов суспензионной клетки CHO-K1.
На фигуре 4B представлен результат изучения уровня экспрессии антитела против белка M2 вируса гриппа человека в прикрепляющейся клетке CHO-K1 при использовании транспозонного вектора Tol2 для экспрессии антитела против белка M2 вируса гриппа человека. Ось ординат показывает уровень продукции антитела (мкг/мл), а ось абсцисс показывает число трансгенных клонов прикрепляющейся клетки CHO-K1.
На фигуре 5 представлено схематическое изображение транспозонного вектора Tol1 для экспрессии антитела против белка M2 вируса гриппа человека. Tol1-L представляет собой левый конец транспозона Tol1 (SEQ ID NO: 14), Tol1-R представляет собой правый конец транспозона Tol1 (SEQ ID NO: 15), CMV представляет собой промотор CMV, поли-A представляет собой сайт полиаденилирования, Hc представляет собой кДНК H-цепи антитела человека, Lc представляет собой кДНК L-цепи антитела человека, и CHX-r представляет собой ген устойчивости к циклогексимиду.
На фигуре 6 представлено схематическое изображение вектора экспрессии транспозазы Tol1. CAGGS представляет собой промотор CAGGS, поли-A представляет собой сайт полиаденилирования, и кДНК TPазы представляет собой кДНК транспозазы Tol1.
На фигуре 7 представлен результат изучения уровня экспрессии антитела против белка M2 вируса гриппа человека в суспензионной клетке CHO-K1 при использовании транспозонного вектора Tol1 для экспрессии антитела против белка M2 вируса гриппа человека. Ось ординат показывает уровень продукции антитела (мкг/мл), а ось абсцисс показывает число трансгенных клонов суспензионной клетки CHO-K1.
Данное изобретение относится к способу получения представляющего интерес белка, включающему введение вектора экспрессии белка, который включает генный фрагмент, содержащий ДНК, кодирующую представляющий интерес белок, и ген селектируемого маркера, а также транспозонные последовательности на обоих концах генного фрагмента, в суспензионную клетку млекопитающего; интегрирование генного фрагмента, вставленного между парой (двумя) транспозонных последовательностей, в хромосому клетки млекопитающего для получения клетки млекопитающего, способной экспрессировать указанный представляющий интерес белок; и суспензионное культивирование клетки млекопитающего.
Примеры способа получения представляющего интерес белка по настоящему изобретению включают способ, включающий следующие этапы от (A) до (C):
(A) этап одновременного введения следующих векторов экспрессии (a) и (b) в суспензионную клетку млекопитающего:
(a) вектор экспрессии, включающий генный фрагмент, содержащий ДНК, кодирующую представляющий интерес белок, и транспозонные последовательности на обоих концах генного фрагмента,
(b) вектор экспрессии, содержащий ДНК, кодирующую транспозазу, которая распознает транспозонные последовательности и обладает активностью переноса генного фрагмента, вставленного между парой транспозонных последовательностей, в хромосому,
(B) этап временной экспрессии транспозазы с вектора экспрессии, введенного на этапе (A), для интегрирования генного фрагмента, вставленного между парой транспозонных последовательностей, в хромосому клетки млекопитающего для получения суспензионной клетки млекопитающего, способной экспрессировать представляющий интерес белок, и
(C) этап суспензионного культивирования суспензионной клетки млекопитающего, способной экспрессировать представляющий интерес белок, полученной на этапе (B), для получения представляющего интерес белка.
Кроме того, настоящее изобретение относится к суспензионной клетке млекопитающего, способной продуцировать представляющий интерес белок, в которую введен вектор экспрессии белка, включающий генный фрагмент, содержащий ДНК, кодирующую представляющий интерес белок, и ген селектируемого маркера, а также транспозонные последовательности на обоих концах генного фрагмента, для интегрирования генного фрагмента, вставленного между парой транспозонных последовательностей, в хромосому.
Кроме того, настоящее изобретение относится к суспензионной клетке млекопитающего, способной продуцировать представляющий интерес белок, в которую введен вектор экспрессии (a), включающий генный фрагмент, содержащий ДНК, кодирующую представляющий интерес белок, и ген селектируемого маркера, а также транспозонные последовательности на обоих концах генного фрагмента, и вектор экспрессии (b), содержащий ДНК, кодирующую транспозазу (трансферазу), которая распознает транспозонные последовательности и обладает активностью переноса генного фрагмента, вставленного между парой транспозонных последовательностей, в хромосому для интегрирования генного фрагмента, вставленного между парой транспозонных последовательностей, в хромосому.
Термин «транспозон» в настоящем описании относится к мобильному генетическому элементу и означает генетический фрагмент, который перемещается на хромосоме или из одной хромосомы в другую хромосому (транспозиция), сохраняя при этом определенную структуру.
Транспозон включает генетический фрагмент из повторяющихся транспозонных последовательностей (также называемых последовательностью инвертированного повтора (IR-последовательностью) или последовательностью концевого инвертированного повтора (TIR-последовательностью)), расположенных в прямом или обратном направлении на обоих концах генетического фрагмента, и нуклеотидную последовательность, кодирующую транспозазу, которая распознает транспозонную последовательность, для переноса гена, находящегося между транспозонными последовательностями.
Транспозаза, транслируемая с транспозона, может переносить ДНК путем распознавания транспозонных последовательностей на обоих концах транспозона, вырезания фрагмента ДНК, вставленного между парой транспозонных последовательностей, и встраивания фрагмента в сайт переноса.
Термин «транспозонная последовательность» в данном описании означает нуклеотидную последовательность транспозона, распознаваемую транспозазой, и имеет то же значение, что и IR-последовательность или TIR-последовательность. ДНК, содержащая нуклеотидную последовательность, может содержать несовершенный повторяющийся фрагмент при условии, что он может быть перенесен (встроен в другой участок генома) благодаря активности транспозазы, и включает транспозонную последовательность, специфическую для транспозазы.
Что касается транспозонной последовательности, используемой по изобретению, предпочтительной является нуклеотидная последовательность, происходящая из транспозона ДНК-типа, и более предпочтительной является нуклеотидная последовательность, происходящая из пары природных или искусственных транспозонов ДНК-типа, которые могут распознаваться транспозазой и переноситься в клетках млекопитающих.
Примеры нуклеотидной последовательности, происходящей из транспозона ДНК-типа, включают нуклеотидные последовательности, происходящие из транспозона Tol1 и транспозона Tol2 рыбы оризии, транспозона «спящая красавица», восстановленного из неавтономного транспозона, существующего в геноме рыбы Onchorhynchus, искусственного транспозона «лягушачий принц» из лягушки и транспозона PiggyBac из насекомого.
В частности, среди них предпочтительными являются нуклеотидные последовательности, происходящие из транспозона Tol2 рыбы оризии, содержащие нуклеотидную последовательность, приведенную в SEQ ID NO: 6, и транспозона Tol2 рыбы оризии, содержащие нуклеотидную последовательность, приведенную в SEQ ID NO: 13.
Примеры нуклеотидной последовательности, происходящей из пары транспозонов Tol2, включают нуклеотидную последовательность в положениях 1-2229 и нуклеотидную последовательность в положениях 4148-4682 в нуклеотидной последовательности транспозона Tol2, приведенной в SEQ ID NO: 6 в списке последовательностей.
В качестве нуклеотидной последовательности, происходящей из пары транспозонов Tol2, более предпочтительными являются нуклеотидная последовательность в положениях 1-200 (SEQ ID NO: 2) (далее в данном документе называемая «последовательность Tol2-L») и нуклеотидная последовательность в положениях 2285-2788 (SEQ ID NO: 3) (далее в данном документе называемая «последовательность Tol2-R») в нуклеотидной последовательности транспозона Tol2, приведенной в SEQ ID NO: 1 в списке последовательностей.
Примеры нуклеотидной последовательности, происходящей из пары транспозонов Tol1, включают нуклеотидную последовательность, содержащую нуклеотидную последовательность в положениях 1-157 и нуклеотидную последовательность в положениях 1748-1855 в нуклеотидной последовательности транспозона Tol1, приведенной в SEQ ID NO: 13 в списке последовательностей.
В качестве нуклеотидной последовательности, происходящей из пары транспозонов Tol1, более предпочтительными являются нуклеотидная последовательность в положениях 1-200 (SEQ ID NO: 14) (далее в данном документе называемая «последовательность Tol1-L») и нуклеотидная последовательность в положениях 1351-1855 (SEQ ID NO: 15) (далее в данном документе называемая «последовательность Tol1-R») в нуклеотидной последовательности транспозона Tol2, приведенной в SEQ ID NO: 1 в списке последовательностей.
Примеры транспозонной последовательности для использования по изобретению включают транспозонные последовательности, в случае которых реакции переноса контролируются путем использования частичной последовательности из транспозонной последовательности, происходящей из вышеуказанного транспозона, посредством регулирования длины нуклеотидной последовательности и посредством модификации нуклеотидной последовательности за счет добавления, делеции или замены.
Что касается контроля реакции переноса транспозона, реакция переноса может быть ускорена или замедлена путем ускорения или замедления распознавания транспозонной последовательности транспозазой, соответственно.
Термин «транспозаза» в настоящем описании означает фермент, который распознает нуклеотидные последовательности, имеющие транспозонные последовательности, и переносит ДНК, находящуюся между нуклеотидными последовательностями, в хромосому или из хромосомы в другую хромосому.
Примеры транспозазы включают Tol1 и Tol2 из рыбы оризии, транспозон «спящая красавица», восстановленный из неавтономного транспозона, существующего в геноме рыбы Onchorhynchus, искусственный транспозон «лягушачий принц» из лягушки и транспозон PiggyBac из насекомых.
Что касается транспозазы, можно использовать природный фермент, и можно использовать любую транспозазу, в которой часть аминокислот заменены, делетированы, вставлены и/или добавлены, при условии сохранения такой же способности к переносу, что и у транпозазы. Контролируя ферментативную активность транспозазы, можно контролировать реакцию переноса ДНК, находящейся между транспозонными последовательностями.
Для анализа того, обладает ли фермент активностью переноса, подобной активности транспозазы, ее можно измерять при помощи 2-компонентной аналитической системы, раскрытой в японской опубликованной нерассмотренной патентной заявке № 235575/2003.
Для иллюстрации, может ли неавтономный элемент Tol2 быть перенесен и вставлен в хромосому клетки млекопитающего вследствие активности транспозазы, можно анализировать, используя отдельно плазмиду, содержащую транспозон Tol2, лишенный транспозазы Tol2 (производный от Tol2 неавтономный транспозон), и плазмиду, содержащую транспозазу Tol2.
В настоящем описании термин «неавтономный транспозон» означает транспозон, который утратил транспозазу, находящуюся внутри транспозона, и вследствие этого не способен автономно осуществлять свой перенос. Неавтономный транспозон может переносить ДНК, вставленную между транспозонными последовательностями неавтономного транспозона, в хромосому клетки-хозяина в случае одновременного присутствия в клетке белка транспозазы, мРНК, кодирующей белок транспозазу, или ДНК, кодирующей белок транспозазу.
Ген транспозазы означает ген, кодирующий транспозазу. В целях повышения эффективности его экспрессии в клетке млекопитающего последовательность, которая регулирует пространство между консенсусной последовательностью Козака (Kozak M., Nucleic Acids Res., 12, 857-872 (1984)) или последовательностью связывания рибосомы, последовательностью Шайна-Дальгарно, и инициирующим кодоном до соответствующего расстояния (например, от 6 до 18 оснований), можно присоединять выше по ходу транскрипции от кодона инициации трансляции ATG гена.
В соответствии со способом по изобретению, для того чтобы интегрировать генный фрагмент, содержащий ДНК, кодирующую представляющий интерес белок, и ген селектируемого маркера в векторе экспрессии, в хромосому клетки-хозяина, вектор экспрессии, включающий генный фрагмент, содержащий ДНК, кодирующую представляющий интерес белок, и ген селектируемого маркера, а также транспозонные последовательности на обоих концах генного фрагмента, вводят в клетку-хозяина, и транспозаза получает возможность проявлять активность с вовлечением транспозонных последовательностей, находящихся в векторе экспрессии, введенном в клетку.
Чтобы дать возможность транспозазе проявлять активность с вовлечением транспозонных последовательностей, находящихся в векторе экспрессии, введенном в клетку, транспозазу можно вводить в клетку инъекцией, либо вектор экспрессии, содержащий ДНК, кодирующую транспозазу, можно вводить в клетку-хозяина совместно с вектором экспрессии, содержащим ДНК, кодирующую представляющий интерес белок, и ген селектируемого маркера. Кроме того, транспозаза может экспрессироваться в клетке в результате введения РНК, кодирующей ген транспозазы, в клетку-хозяина.
Вектор экспрессии не имеет конкретных ограничений. Можно использовать любой вектор экспрессии, по желанию выбирая из векторов экспрессии, известных специалистам в данной области, в зависимости от клетки-хозяина, в которую вводят вектор экспрессии, содержащий ген транспозазы, от применения и тому подобного.
Для того чтобы с помощью способа по изобретению получать белок, составленный из двух или более полипептидов, ДНК можно интегрировать в хромосому клетки путем интегрирования ДНК, кодирующей два или более полипептидов, в один и тот же или в разные векторы экспрессии, а затем вводить векторы экспрессии в клетку-хозяина.
Транспозазу можно встраивать в вектор экспрессии для экспрессии совместно с представляющим интерес белком, либо можно встраивать в вектор, отличный от вектора экспрессии. Можно обеспечивать временное проявление активности, либо постоянное проявление активности транспозазы, однако предпочтительно, чтобы транспозаза действовала временно, чтобы подготовить клетку к стабильной продукции.
В число примеров способов обеспечения временной активности транспозазы входит способ, включающий получение вектора экспрессии, который содержит ДНК, кодирующую транспозазу, и вектора экспрессии, содержащего ДНК, кодирующую представляющий интерес белок, с последующим введением обеих плазмид экспрессии одновременно в клетку-хозяина.
В настоящем описании термин «вектор экспрессии» означает вектор экспрессии, используемый для введения в клетку млекопитающего с целью экспрессии представляющего интерес белка. Вектор экспрессии, используемый по изобретению, обладает структурой, в которой по меньшей мере пара транспозонных последовательностей присутствует на обоих концах экспрессионной кассеты.
В настоящем описании термин «экспрессионная кассета» означает нуклеотидную последовательность, содержащую контролирующую экспрессию гена область, необходимую для экспрессии представляющего интерес белка, и последовательность, кодирующую представляющий интерес белок. Примеры областей, контролирующих экспрессию гена, включают энхансер, промотор и терминатор. Экспрессионная кассета может содержать ген селектируемого маркера.
Можно использовать любой промотор при условии, что он способен функционировать в животной клетке. Примеры включают промотор IE (предраннего) гена цитомегаловируса (CMV), ранний промотор SV40, промотор ретровируса, промотор металлотионеина, промотор белка теплового шока, промотор SRα, мышиный вирус лейкемии Молони, энхансер и тому подобное. Кроме того, вместе с промотором можно использовать энхансер IE гена человеческого CMV.
«Ген селектируемого маркера» означает произвольный маркерный ген, который можно использовать, чтобы отличить клетку, в которую введен плазмидный вектор, от клетки, не содержащей вектор.
Примеры генов селектируемого маркера включают ген устойчивости к лекарственному средству (ген устойчивости к неомицину, ген DHFR, ген устойчивости к пуромицину, ген устойчивости к бластицидину, ген устойчивости к гигромицину и ген устойчивости к циклогексимиду (японская опубликованная нерассмотренная патентная заявка № 262879/2002)), флуоресцентные и биолюминесцентные маркерные гены (такие как зеленый флуоресцентный белок GFP) и тому подобное.
Предпочтительным селектируемым маркером по изобретению является ген устойчивости к лекарственному средству, и особенно предпочтительным селектируемым маркером является ген устойчивости к циклогексимиду. Кроме того, путем генной модификации гена селектируемого маркера можно также изменять проявление устойчивости к лекарственному средству и люминесцентное проявление белка селектируемого маркера.
Циклогексимид (иногда в данном документе называемый CHX) представляет собой ингибитор белкового синтеза, и, как примеры использования гена устойчивости к циклогексимиду в качестве гена селектируемого маркера, известны случаи с дрожжами (Kondo K. J. Bacteriol., 177, 24, 7171-7177 (1995)) и животными клетками (японская опубликованная нерассмотренная патентная заявка № 262879/2002).
В случае животных клеток было установлено, что устойчивость к циклогексимиду обеспечивается трансформантом, который экспрессирует белок, кодируемый нуклеотидной последовательностью, приведенной в SEQ ID NO: 7 списка последовательностей, в котором пролин в положении 54 субъединицы L36a человеческого рибосомного белка, кодируемой нуклеотидной последовательностью, приведенной в SEQ ID NO: 5 списка последовательностей, заменен на глутамин.
Способ введения вышеуказанного вектора экспрессии белка, содержащего транспозонную последовательность, плазмидного вектора, экспрессирующего транспозазу, и РНК не имеет конкретных ограничений. Примеры включают кальций-фосфатную трансфекцию, электропорацию, липосомный метод, метод генной пушки, липофекцию и тому подобное.
Примеры способа прямого введения транспозазы в форме белка включают введение в клетку с помощью микроинъекции или эндоцитоза. Перенос гена можно осуществлять методом, описанным в книге Shin Idenshi Kogaku Handbook (New Genetic Engineering Handbook) под редакцией Masami Muramatsu и Tadashi Yamamoto, опубликованной Yodo-sha, ISBN 9784897063737.
Клетка-хозяин может являться любой клеткой млекопитающего при условии, что она может быть субкультивирована и может стабильно экспрессировать представляющий интерес белок. Примеры клеток-хозяев включают клетку PER.C6, клетку человеческого лейкоза, клетку Намальвы, COS-клетку обезьяны, клетку крысиной миеломы YB2/3HL.P2.G11.16Ag.20 (также называемую YB2/0), клетку мышиной миеломы NS0, клетку мышиной миеломы SP2/0-Agl4, клетку BHK сирийского хомячка, HBT5637 (японская нерассмотренная публикация патентной заявки № 1998-000299), клетку CHO яичника китайского хомячка (Journal of Experimental Medicine, 108, 945 (1958); Proc. Natl. Acad. Sci. USA., 601275 (1968); Genetics, 55, 513 (1968); Chromosoma, 41, 129 (1973); Methods in Cell Science, 18, 115 (1996); Radiation Research, 148, 260 (1997); Proc. Natl. Acad. Sci. USA., 77, 4216 (1980); Proc. Natl. Acad. Sci., 60, 1275 (1968); Cell, 6, 121 (1975); Molecular Cell Genetics, приложение I, II (стр. 883-900)), CHO/DG44, CHO-K1 (ATCC CCL-61), DUKXB11 (ATCC CCL-9096), Pro-5 (ATCC CCL-1781), CHO-S (Life Technologies, каталожный № 11619), Pro-3 и подштамм клетки CHO.
Кроме того, вышеуказанную клетку-хозяина можно также использовать в способе получения белка по изобретению, модифицируя ее таким образом, чтобы она подходила для продукции белка, путем модификации хромосомной ДНК, введения экзогенного гена и тому подобного.
Кроме того, с целью контроля структуры сахарной цепи, связанной с представляющим интерес белком, который предстоит продуцировать, в качестве клетки-хозяина можно также использовать клетку Lec 13, которая приобрела устойчивость к лектину [Somatic Cell and Molecular Genetics, 12, 55 (1986)], и клетку CHO, у которой делетирован ген α1,6-фукозилтрансферазы (WO 2005/35586, WO 2002/31140).
Представляющий интерес белок может являться любым белком при условии, что он может быть экспрессирован с помощью способа по изобретению. В частности, примеры включают человеческие сывороточные протеины, пептидные гормоны, факторы роста, цитокины, факторы свертывания крови, белки системы фибринолиза, антитела, а также частичные фрагменты различных белков и тому подобное.
Предпочтительные примеры представляющего интерес белка включают моноклональное антитело, такое как химерное антитело, гуманизированное антитело и антитело человека; Fc-слитый белок и альбумин-связанный белок; а также их фрагменты.
Эффекторную активность моноклонального антитела, полученного способом по настоящему изобретению, можно контролировать различными методами. Например, известными методами являются метод контроля количества фукозы (далее в данном описании также называемой «сердцевинной фукозой»), которая связана с N-ацетилглюкозамином (GlcNAc) посредством α-1,6-связи в восстановительном конце сложного типа N-связанной сахарной цепи, связанной с остатком аспарагина (Asn) в положении 297 Fc-области антитела (WO 2005/035586, WO 2002/31140 и WO 00/61739), метод контроля эффекторной активности моноклонального антитела путем модификации аминокислотной группы(групп) Fc-области антитела, и тому подобные. Эффекторную активность моноклонального антитела, полученного способом по настоящему изобретению, можно контролировать, используя любой из этих методов.
«Эффекторная активность» означает антитело-зависимую активность, которая индуцируется через Fc-область антитела. В качестве антитело-зависимой активности известны антитело-зависимая клеточно-опосредованная цитотоксичность (активность ADCC), комплемент-зависимая цитотоксичность (активность CDC), антитело-зависимый фагоцитоз (активность ADP) фагоцитарными клетками, такими как макрофаги или дендритные клетки, и тому подобное.
Кроме того, путем контролирования содержания сердцевинной фукозы сложного типа N-связанной сахарной цепи Fc-области моноклонального антитела можно увеличивать или уменьшать эффекторную активность антитела.
В качестве способа снижения содержания фукозы, которая связана со сложного типа N-связанной сахарной цепью, связанной с Fc-областью антитела, можно получать антитело, с которым не связана фукоза, путем экспрессии антитела с использованием клетки CHO, лишенной гена, кодирующего α1,6-фукозилтрансферазу. Антитело, с которым не связана фукоза, обладает высокой активностью ADCC.
С другой стороны, в качестве способа повышения содержания фукозы, которая связана со сложного типа N-связанной сахарной цепью, связанной с Fc-областью антитела, можно получать антитело, с которым связана фукоза, путем экспрессии антитела с использованием клетки-хозяина, в которую введен ген, кодирующий α1,6-фукозилтрансферазу. Антитело, с которым связана фукоза, обладает пониженной активностью ADCC по сравнению с антителом, не связанным с фукозой.
Кроме того, путем модификации аминокислотного остатка(ов) в Fc-области антитела активность ADCC или активность CDC можно увеличивать или уменьшать. Например, активность CDC антитела можно увеличивать путем использования аминокислотной последовательности Fc-области, описанной в US 2007/0148165.
Кроме того, активность ADCC или активность CDC антитела можно увеличивать или уменьшать путем модификации аминокислоты, как описано в патентах США №№ 6737056 или 7297775 или 7317091.
В настоящем изобретении термин «суспензионная клетка млекопитающего» означает клетку, которая не прикрепляется к культуральной клеточной «якорной» подложке, используемой для облегчения прикрепления клеток в культуре, такой как микрогранулы, культуральный контейнер для культуры тканей (также называемый контейнером для тканевой культуры или прикрепляющейся культуры, и так далее) и тому подобное, и способна выживать и расти, будучи суспендированной в культуральной жидкости.
Если клетка не прикрепляется к культуральной клеточной «якорной» подложке, она способна выживать и расти в состоянии одиночной клетки в культуральной жидкости, либо выживать и расти в состоянии клеточной массы, образованной в результате скопления двух или более клеток.
Кроме того, в качестве суспензионной клетки млекопитающего, используемой по настоящему изобретению, предпочтительной является клетка, которая способна выживать и расти в бессывороточной среде, не содержащей эмбриональную телячью сыворотку (далее в данном документе называемую FCS) и тому подобное, будучи суспендированной в культуральной жидкости без прикрепления к культуральной клеточной «якорной» подложке, и более предпочтительной является клетка млекопитающего, которая способна выживать и расти, будучи суспендированной в безбелковой среде, которая не содержит белка.
В качестве культурального контейнера для тканевой культуры можно использовать любой культуральный контейнер, такой как колба, чашка Петри и тому подобное, при условии, что на него нанесено покрытие для прикрепляющейся культуры. В частности, например, является ли клетка суспензионной клеткой млекопитающего можно убедиться путем использования коммерчески доступной колбы для тканевой культуры (производитель Greiner), колбы для прикрепляющейся культуры (производитель Sumitomo Bakelite) и тому подобного.
В качестве суспензионной клетки млекопитающего, используемой по настоящему изобретению, можно использовать либо клетку, полученную путем дополнительной адаптации клетки, изначально имеющей суспензионные свойства, к суспензионному культивированию, либо суспензионную клетку млекопитающего, полученную путем адаптации прикрепляющейся клетки млекопитающего к условиям суспензионного культивирования.
Примеры клетки, изначально имеющей суспензионные свойства, включают клетку PER.C6, клетку крысиной миеломы YB2/3HL.P2.G11.16Ag.20 (или также называемую YB2/0), клетку CHO-S (производитель Invitrogen) и тому подобное.
Вышеуказанную «суспензионную клетку млекопитающего, полученную путем адаптации прикрепляющейся клетки млекопитающего к условиям суспензионного культивирования» можно получать способом, описанным в Mol. Biotechnol., 2000, 15(3), 249-57, или способом, описанным далее, и можно получать путем создания клетки, демонстрирующей способность пролиферировать и способность выживать, аналогичные таковым до процесса адаптации к условиям суспензионного культивирования, либо превосходящие таковые до процесса адаптации к условиям суспензионного культивирования (J. Biotechnol., 2007, 130(3), 282-90).
Определение «аналогичные таковым до процесса адаптации к условиям суспензионного культивирования» означает, что коэффициент выживания, скорость пролиферации (время удвоения) и тому подобное у клетки, адаптированной к суспензионному культивированию, практически такие же, что и у клетки до адаптации к суспензионному культивированию.
Примеры способа адаптации прикрепляющейся клетки млекопитающего к условиям суспензионного культивирования по настоящему изобретению включают следующий способ. Содержание сыворотки в содержащей сыворотку среде уменьшают до 1/10, и субкультивирование повторяют при относительно высокой концентрации клеток. Когда клетка млекопитающего становится способной выживать и пролиферировать, содержание сыворотки вновь уменьшают и субкультивирование повторяют. С помощью такого способа можно получать суспензионную клетку млекопитающего, способную выживать и пролиферировать в бессывороточных условиях.
Кроме того, суспензионную клетку млекопитающего можно также получать способом, включающим культивирование с добавлением соответствующего неионного сурфактанта, такого как Pluronic-F68 или тому подобное, в культуральную жидкость.
Примеры прикрепляющейся клетки млекопитающего, которая приобретает суспензионные свойства путем адаптации к условиям суспензионного культивирования, включают клетку мышиной миеломы NS0, клетку CHO и тому подобное.
В настоящем изобретении считается, что клетка обладает свойством суспензионной клетки млекопитающего, если при суспензионном культивировании клеток при плотности 2×105 клеток/мл концентрация клеток после культивирования в течение 3 или 4 дней предпочтительно составляет 5×105 клеток/мл или более, более предпочтительно 8×105 клеток/мл или более, особенно предпочтительно 1×106 клеток/мл или более, наиболее предпочтительно 1,5×106 клеток/мл или более.
Кроме того, время удвоения суспензионной клетки млекопитающего по настоящему изобретению предпочтительно составляет 48 часов или менее, более предпочтительно 24 часа или менее, особенно предпочтительно 18 часов или менее, наиболее предпочтительно 11 часов или менее.
Примеры среды для суспензионного культивирования включают коммерчески доступные среды, такие как среда CD-CHO (производитель Invitrogen), среда EX-CELL 325-PF (производитель SAFC Biosciences), среда SFM4CHO (производитель HyClone) и тому подобное. Кроме того, ее также можно получать, смешивая сахариды, аминокислоты и аналогичные кислоты, необходимые для культивирования клеток млекопитающих.
Суспензионную клетку млекопитающего можно культивировать, используя культуральный контейнер, который можно использовать для суспензионного культивирования в условиях культивирования, соответствующих суспензионному культивированию. Примеры культуральных контейнеров включают 96-луночный планшет для клеточных культур (производитель Corning), T-колбу (производитель Becton Dickinson), колбу Эрленмейера (производитель Corning) и тому подобное.
Что касается условий культивирования, например, клетку можно статически культивировать в атмосфере 5% CO2 при температуре культивирования 37°C. Можно также использовать оборудование для встряхивания культуры, такое как культивационное оборудование исключительно для суспензионного культивирования, Wave Bioreactor (производитель GE Healthcare Bioscience).
Что касается условий суспензионного культивирования суспензионной клетки млекопитающего с использованием оборудования Wave Bioreactor, то клетку можно культивировать методом, описанным на сайте GE Healthcare Bioscience http://www.gelifesсienсes.co.jp/tech-support/manual/pdf/cellcult/wave-03-16.pdf.
В дополнение к культивированию при встряхивании, можно также использовать культивирование с помощью оборудования для ротационного перемешивания, такого как биореактор. Культивирование в биореакторе можно проводить методом, описанным в Cytotechnology, (2006) 52: 199-207, и тому подобными методами.
В настоящем изобретении, если используют клеточную линию, отличную от суспензионных клеток млекопитающего, то можно использовать любую клеточную линию при условии, что она является линией клеток млекопитающего, адаптированной к суспензионному культивированию вышеуказанным способом, и представляет собой клеточную линию, которую можно использовать в способе получения белка по настоящему изобретению.
Очистку представляющего интерес белка, полученного с помощью суспензионной клетки млекопитающего, осуществляют путем отделения представляющего интерес белка от примесей, отличных от представляющего интерес белка, в культуральной жидкости или клеточном гомогенате, которые содержат представляющий интерес белок. Примеры способов разделения включают центрифугирование, диализ, осаждение сульфатом аммония, колоночную хроматографию, фильтрование и тому подобное. Разделение можно проводить на основании различий физико-химических свойств представляющего интерес белка и примесей, а также на основании различий их сродства к колоночному носителю.
Очистку представляющего интерес белка можно проводить, например, методом, описанным в Protein Experimentation Note (первый том) - Extraction, Separation and Expression of Recombinant Protein (перевод руководства, написанного по-японски) (под редакцией Masato Okada и Kaori Miyazaki, опубликовано Yodo-sha, ISBN 9784897069180).
Полное содержание всех литературных источников, таких как научные документы, патенты, патентные заявки, цитированных в данном документе, включено в данное описание посредством ссылок в той же степени, что и тех, которые описаны иллюстративно, соответственно.
Настоящее изобретение описано выше путем демонстрации предпочтительных вариантов его осуществления ради облегчения понимания. Здесь и далее настоящее изобретение дополнительно описано, в частности, на основе примеров, однако вышеуказанные объяснения и следующие далее примеры приведены лишь с целью иллюстрации и не предназначены для ограничения изобретения. Соответственно, объем изобретения не ограничен вариантами осуществления и примерами, которые конкретно описаны в данном документе, но ограничен лишь формулой изобретения.
Различные экспериментальные методы, связанные с генетической рекомбинацией, описанные далее в данном документе, такие как клонирование и тому подобное, осуществляли в соответствии с методами генетической инженерии, описанными в руководстве Molecular Cloning, 2е издание под редакцией J. Sambrook, E. F. Frisch и T. Maniatis, руководстве Current Protocols in Molecular Biology под редакцией Frederick M. Ausubel et al., опубликованном Current Protocols, и тому подобными.
Примеры
[Пример 1]
Получение транспозонного вектора для экспрессии антитела против белка M2 вируса гриппа человека
В качестве плазмидного вектора для экспрессии белка использовали плазмиду, содержащую генную экспрессионную кассету для клеток млекопитающего, содержащую произвольный ген человеческого антитела и маркерный ген устойчивости к лекарственному средству, вставленные между парой транспозонных последовательностей Tol2.
Каждую ДНК используемых генов химически или искусственно синтезировали на основании известной нуклеотидной последовательности, либо получали путем создания праймеров для обеих его концевых последовательностей с последующим проведением ПЦР, используя соответствующий источник ДНК в качестве матрицы. Для последующего проведения манипуляций с генами к концу праймера добавляли сайт рестрикции для фермента рестрикции.
В нуклеотидной последовательности неавтономного транспозона Tol2, описанного в японской опубликованной нерассмотренной патентной заявке № 235575/2003 (SEQ ID NO: 1), нуклеотидную последовательность в положениях 1-200 (последовательность Tol2-L) (SEQ ID NO: 2) и нуклеотидную последовательность в положениях 2285-2788 (последовательность Tol2-R) (SEQ ID NO: 3) использовали в качестве транспозонных последовательностей.
Каждый из фрагментов синтетической ДНК, содержащих пару транспозонных последовательностей (производитель TAKARA BIO INC.), получали следующим методом. Получали фрагмент ДНК, содержащий нуклеотидную последовательность, в которой последовательность распознавания фермента рестрикции NruI была присоединена как к 5'-концу, так и к 3'-концу последовательности Tol2-R. Затем получали фрагмент ДНК, содержащий нуклеотидную последовательность, в которой последовательность распознавания фермента рестрикции FseI была присоединена к 5'-концу последовательности Tol2-L, а фермента рестрикции AscI была присоединена к его 3'-концу.
Затем полученные таким образом фрагменты ДНК, содержащие последовательность Tol2-R и последовательность Tol2-L, встраивали в вектор экспрессии N5LG1-M2-Z3 (WO 2006/061723), содержащий нуклеотидную последовательность, кодирующую аминокислотную последовательность антитела Z3G1 против белка M2 вируса гриппа человека.
Вектор N5LG1-M2-Z3 (WO 2006/061723), в который нуклеотидная последовательность (SEQ ID NO: 8), кодирующая H-цепь антитела Z3G1 против белка M2 вируса гриппа человека (№ депозита в ATCC PTA-5968: депонировано 13 марта 2004 г., Американская коллекция типовых культур, Manassas, VA, USA), и нуклеотидная последовательность (SEQ ID NO: 10 и SEQ ID NO: 11), кодирующая L-цепь (SEQ ID NO: 9) того же антитела, были встроены под контролем энхансера/промотора CMV, использовали в качестве генной экспрессионной кассеты антитела.
Фрагмент ДНК, содержащий последовательность Tol2-R, встраивали в сайт рестрикции фермента NruI вектора N5LG1-M2-Z3 на 5'-концевой стороне генного фрагмента, содержащего генную экспрессионную кассету антитела и ген маркера устойчивости. Затем фрагмент ДНК, содержащий последовательность Tol2-L, встраивали в сайты рестрикции ферментов FseI и AscI на 3'-концевой стороне.
Кроме того, сконструировали транспозонный вектор для экспрессии антитела против белка M2 вируса гриппа человека (фигура 1) путем встраивания экспрессионной кассеты гена устойчивости к циклогексимиду, связанной с нуклеотидной последовательностью (SEQ ID NO: 5), кодирующей ген устойчивости к циклогексимиду (ген, в котором пролин в положении 54 человеческого рибосомного белка L36a заменен на глутамин), в сайт распознавания FseI вектора N5LG1-M2-Z3, связанного с транспозонной последовательностью Tol2, под контролем энхансера/промотора CMV.
С другой стороны, вектор, не содержащий транспозонных последовательностей, назвали вектором экспрессии антитела против белка M2 вируса гриппа человека и использовали в качестве контрольного вектора (фигура 2).
[Пример 2]
Получение вектора экспрессии транспозазы
Транспозазу экспрессировали при помощи вектора экспрессии, независимого от вектора экспрессии представляющего интерес антитела. То есть, ген, кодирующий транспозазу Tol2 из рыбы оризии (SEQ ID NO: 4), встраивали ниже по ходу транскрипции от промотора CAGGS вектора pCAGGS (Gene, 108, 193-200, 1991) и использовали в качестве вектора экспрессии (фигура 3).
[Пример 3]
(1) Получение суспензионной клетки CHO
Прикрепляющуюся клетку CHO, которую культивировали в среде α-MEM (производитель Invitrogen), содержащей 10% сыворотку (FCS), открепляли от подложки и извлекали путем обработки трипсином и культивировали со встряхиванием в инкубаторе при 37°C в атмосфере 5% CO2, используя свежую среду α-MEM, содержащую 10% FCS. Через несколько дней рост этих клеток подтверждали, после чего культивирование со встряхиванием продолжали, высеяв их в среду α-MEM, содержащую 5% FCS, в концентрации 2×105 клеток/мл.
Через несколько дней после этого аналогичным образом проводили инокуляцию, используя среду α-MEM, содержащую 5% FCS. В конечном итоге получали клетку, адаптированную к суспензионному культивированию, путем повторяющегося субкультивирования и встряхивания культуры с использованием бессывороточной среды α-MEM, подтверждая, что клетки имеют такую же способность к росту, что и при культивировании их в присутствии сыворотки.
(2) Получение клетки CHO, продуцирующей антитело
В качестве векторов экспрессии использовали транспозонный вектор для экспрессии антитела против белка M2 вируса гриппа человека, полученный в примере 1 и примере 2 (далее называемый транспозонный вектор), и вектор экспрессии транспозазы Tol2 pCAGGS-T2TP (фигура 3, Kawakami K. & Noda T., Genetics, 166, 895-899 (2004)). Кроме того, в качестве контроля использовали вектор экспрессии антитела против белка M2 вируса гриппа человека, не имеющий транспозонных последовательностей.
Путем введения вышеуказанных векторов экспрессии в адаптированную к суспензионному культивированию клетку CHO-K1 (каталожный № CCL-61 в американской коллекции типовых культур) или клетку HEK293 (клетку FreeStyle 293F, производитель Invitrogen) сравнивали частоту получения клонов, устойчивых к циклогексимиду.
Каждый вид клеток (4x106 клеток) суспендировали в 400 мкл PBS, и транспозонный вектор для экспрессии антитела против белка M2 вируса гриппа человека (10 мкг) и вектор экспрессии транспозазы Tol2 (25 мкг) совместно трансфицировали напрямую в форме кольцевой ДНК методом электропорации. В связи с этим, для того чтобы транспозаза Tol2 экспрессировалась временно, вектор экспрессии транспозазы Tol2 вводили напрямую в форме кольцевой ДНК с целью предотвращения интеграции в хромосому хозяина.
Кроме того, в качестве контроля вектор экспрессии антитела против белка M2 вируса гриппа человека (10 мкг) линеаризировали с помощью фермента рестрикции, а затем вводили в каждый вид клеток стандартным методом переноса генов с помощью электропорации.
Электропорацию проводили, используя кювету с шириной щели 4 мм (производитель Bio-Rad), в электропораторе (Gene Pulser Xcell System (производитель Bio-Rad)) в следующих условиях: напряжение 300 В, электростатическая емкость 500 мкФ и комнатная температура.
После транфекции методом электропорации каждый вид клеток высевали в три 96-луночных планшета и культивировали в CO2-инкубаторе в течение 3 дней, используя среду EX-CELL 325-PF, производимую SAFC Biosciences, для клетки CHO и среду FreeStyle-293 (производимую Invitrogen) для клетки HEK293.
Затем, со дня смены среды на 4-й день после трансфекции добавляли 3 мкг/мл циклогексимида к среде так, что клетки культивировались в присутствии циклогексимида, с последующим культивированием в течение 3 недель с еженедельной сменой среды.
После культивирования в течение 3 недель подсчитывали число лунок, в которых были обнаружены устойчивые к циклогексимиду колонии. Результаты приведены в таблице 1 и таблице 2.
Сравнение числа устойчивых к циклогексимиду клеток
(клетки CHO)
Сравнение числа устойчивых к циклогексимиду клеток
(клетки HEK293)
Как показано в таблице 1, каждый из транспозонного вектора экспрессии антитела против белка M2 вируса гриппа человека или вектора экспрессии антитела против белка M2 вируса гриппа человека вводили в суспензионную клетку CHO-K1. В результате, устойчивые к циклогексимиду трансформанты не были получены из клетки, в которую ввели вектор экспрессии антитела против белка M2 вируса гриппа человека, как и в случае с другими клеточными линиями, однако устойчивые к циклогексимиду трансформанты были получены с высокой частотой из клетки, в которую ввели транспозонный вектор для экспрессии антитела против белка M2 вируса гриппа человека.
С другой стороны, как показано в таблице 2, устойчивые к циклогексимиду трансформанты не были получены в случае, когда транспозонный вектор для экспрессии антитела против белка M2 вируса гриппа человека и вектор экспрессии антитела против белка M2 вируса гриппа человека были введены в клетку HEK293.
Исходя из этих результатов, было установлено, что намеченный кодирующий белок ген и ген устойчивости к циклогексимиду, вставленные между парой транспозонных последовательностей, эффективно встраиваются в хромосому клетки-хозяина, а именно, в суспензионную клетку млекопитающего.
(3) Изучение продукции антитела суспензионной клеткой CHO и прикрепляющейся клеткой CHO
Для изучения эффективности продукции антитела суспензионной клеткой CHO или прикрепляющейся клеткой CHO исследовали количество антител, продуцируемых соответствующими клеточными линиями. В качестве суспензионной клетки CHO использовали суспензионную клетку CHO-K1, адаптированную к суспензионному культивированию. Кроме того, в качестве прикрепляющейся клетки CHO использовали прикрепляющуюся клетку CHO-K1 до адаптации к суспензионному культивированию.
Транспозонный вектор экспрессии антитела против белка M2 вируса гриппа человека (10 мкг) и вектор экспрессии транспозазы Tol2 (25 мкг) вводили методом электропорации, соответственно, в суспензионную клетку CHO-K1 и прикрепляющуюся клетку CHO-K1. Затем суспензионную клетку CHO-K1 и прикрепляющуюся клетку CHO-K1 высевали в три 96-луночных планшета для каждого вида клеток.
Среду для суспензионных клеток (EX-CELL 325-PF, производитель SAFC Biosciences) использовали для суспензионной клетки CHO-K1, а среду α-MEM, содержащую 10% сыворотку, использовали для прикрепляющейся клетки CHO-K1. Каждый вид клеток культивировали в CO2-инкубаторе в течение 3 дней. Со дня смены среды на 4-й день после трансфекции добавляли 3 мкг/мл циклогексимида к среде так, что клетки культивировались в присутствии циклогексимида, с последующим культивированием в течение 3 недель. В этом случае среду меняли каждую неделю.
В случае суспензионной клетки CHO-K1, 1x106 клеток высевали в 6-луночный планшет и культивировали со встряхиванием в CO2-инкубаторе в течение 3 дней, количество белка антитела против белка M2 вируса гриппа человека измеряли методом ВЭЖХ в культуральном супернатанте.
В случае прикрепляющейся клетки CHO-K1, среду меняли, когда клетки достигали конфлюэнтности в 6-луночном планшете (2×106 клеток), и через 3 дня после статического культивирования количество белка антитела измеряли методом ВЭЖХ в культуральном супернатанте.
Концентрацию антитела в культуральном супернатанте определяли методом, описанным в Yeast Res., 7 (2007), 1307-1316. Результаты представлены на фигуре 4A и фигуре 4B.
Как показано на фигуре 4A, было получено большое количество клеток, демонстрирующих заметно повышенный уровень экспрессии антитела при использовании клетки CHO-K1, адаптированной к суспензионному культивированию. С другой стороны, как показано на фигуре 4B, было получено количество клеток, демонстрирующих уровень экспрессии лишь на грани предела обнаружения ВЭЖХ (5 мкг/мл) или менее, когда использовали прикрепляющуюся клетку CHO-K1.
Исходя из этих результатов, было установлено, что при экспрессии представляющего интерес белка с использованием транспозонного вектора представляющий интерес белок можно экспрессировать на высоком уровне, если использовать суспензионную клетку млекопитающего.
Кроме того, из результатов примеров 1-3 следует, что способ по изобретению можно использовать как новый способ получения представляющего интерес белка, эффективно получая продуцирующую клетку, которая способна экспрессировать на высоком уровне экзогенный ген, с использованием суспензионной клетки млекопитающего, адаптированной к суспензионному культивированию.
[Пример 4]
Получение транспозонного вектора Tol1 для экспрессии антитела против белка M2 вируса гриппа человека
Таким же образом, как и в примере 1, в качестве плазмидного вектора экспрессии белка использовали плазмиду, содержащую генную экспрессионную кассету для клеток млекопитающих, содержащую произвольный ген человеческого антитела и маркерный ген устойчивости к лекарственному средству, вставленные между парой транспозонных последовательностей Tol1.
Каждую ДНК используемых генов химически искусственно синтезировали на основании известной информации о последовательности, либо получали путем создания праймеров для обеих его концевых последовательностей и проведения ПЦР с использованием соответствующего источника ДНК в качестве матрицы. Для последующего проведения манипуляций с генами к концу праймера добавляли сайт, расщепляемый ферментом рестрикции.
В нуклеотидной последовательности неавтономного транспозона Tol1, приведенной в SEQ ID NO: 13 списка последовательностей (WO 2008/072540), нуклеотидную последовательность в положениях 1-200 (последовательность Toll-L) (SEQ ID NO: 14) и нуклеотидную последовательность в положениях 1351-1855 (последовательность Tol1-R) (SEQ ID NO: 15) использовали в качестве транспозонных последовательностей.
Каждый из фрагментов синтетической ДНК, содержащих пару траспозонных последовательностей, получали следующим методом. Получали фрагмент ДНК, содержащий нуклеотидную последовательность, в которой последовательность распознавания фермента рестрикции NruI была присоединена как к 5'-концу, так и к 3'-концу последовательности Tol1-R. Затем получали фрагмент ДНК, содержащий нуклеотидную последовательность, в которой последовательность распознавания фермента рестрикции FseI была присоединена к 5'-концу последовательности Tol1-L, а фермента рестрикции AscI была присоединена к его 3'-концу.
Затем полученные таким образом фрагменты ДНК, содержащие последовательность Tol1-R и последовательность Tol1-L, встраивали в вектор экспрессии N5LG1-M2-Z3. Фрагмент ДНК, содержащий последовательность Tol1-R, встраивали в сайт рестрикции фермента NruI вектора N5LG1-M2-Z3, существующий на 5'-концевой стороне генного фрагмента, содержащего генную экспрессионную кассету антитела и ген маркера устойчивости, а фрагмент ДНК, содержащий последовательность Tol1-L, встраивали в сайты рестрикции ферментов FseI и AscI, существующие на 3'-концевой стороне.
Кроме того, сконструировали транспозонный вектор Tol1 для экспрессии антитела против белка M2 вируса гриппа человека (фигура 5) путем встраивания экспрессионной кассеты гена устойчивости к циклогексимиду, связанной с геном устойчивости к циклогексимиду (ген, в котором пролин в положении 54 человеческого рибосомного белка L36a заменен на глутамин), в сайт распознавания FseI вектора N5LG1-M2-Z3, связанного с транспозонной последовательностью Tol1, под контролем энхансера/промотора CMV.
[Пример 5]
Получение вектора экспрессии транспозазы Tol1
Транспозазу экспрессировали при помощи вектора экспрессии, независимого от вектора экспрессии представляющего интерес антитела. То есть, экспрессионную кассету гена транспозазы Tol1, связанную с фрагментом ДНК, кодирующим транспозазу Tol1 из рыбы оризии, содержащую нуклеотидную последовательность, приведенную в SEQ ID NO: 16 списка последовательностей, встраивали в pBluescriptII SK (+) (производитель Stratagene) под контролем энхансера/промотора CMV и использовали в качестве вектора экспрессии pTol1азы (фигура 6).
[Пример 6]
(1) Получение клетки CHO, продуцирующей антитело
В качестве векторов экспрессии использовали транспозонный вектор Tol1 для экспрессии антитела против белка M2 вируса гриппа человека (далее в данном документе называемый транспозонный вектор Tol1) и вектор экспрессии транспозазы Tol1 pTol1азы из примера 4 и примера 5. Кроме того, в качестве клетки использовали клетку CHO-K1, полученную путем адаптации к суспензионному культивированию таким же образом, как в примере 3(1).
Вышеуказанные векторы экспрессии вводили в клетку CHO-K1, адаптированную к суспензионному культивированию, и измеряли частоту появления клонов, устойчивых к циклогексимиду. Клетки CHO-K1, адаптированные к суспензионному культивированию (4x106 клеток), суспендировали в 400 мкл PBS, и транспозонный вектор Tol1 для экспрессии антитела против белка M2 вируса гриппа человека (10 мкг) и вектор экспрессии транспозазы Tol1 (50 мкг) совместно трансфицировали напрямую в форме кольцевой ДНК методом электропорации. Чтобы получить временную экспрессию транспозазы Tol1, вектор экспрессии транспозазы Tol1 вводили напрямую в форме кольцевой ДНК с целью предотвращения интеграции в хромосому хозяина.
Электропорацию проводили, используя кювету с шириной щели 4 мм (производитель Bio-Rad), в электропораторе (Gene Pulser Xcell System (производитель Bio-Rad)) в следующих условиях: напряжение 300 В, электростатическая емкость 500 мкФ и комнатная температура.
После транфекции методом электропорации каждый вид клеток высевали в два 96-луночных планшета и культивировали в CO2-инкубаторе в течение 3 дней, используя среду EX-CELL 325-PF (производитель SAFC Biosciences) для клетки CHO. Затем, со дня смены среды на 4-й день после трансфекции добавляли 3 мкг/мл циклогексимида к среде так, что клетки культивировались в присутствии циклогексимида, с последующим культивированием в течение 3 недель с еженедельной сменой среды.
После культивирования в течение 3 недель подсчитывали число лунок, в которых были обнаружены устойчивые к циклогексимиду колонии. Результаты приведены в таблице 3. Каждый из тестов 1-3 в таблице 3 демонстрирует результат троекратного проведения переноса генов.
Как показано в таблице 3, если транспозонный вектор Tol1 для экспрессии антитела против белка M2 вируса гриппа человека вводили в суспензионную клетку CHO-K1, устойчивые к циклогексимиду трансформанты появлялись с высокой частотой, как в примере 3, в котором вводили транспозонный вектор Tol2 для экспрессии антитела против белка M2 вируса гриппа человека.
Исходя из этих результатов, было установлено, что ген антитела и ген устойчивости к циклогексимиду, вставленные между парой транспозонных последовательностей, эффективно трансдуцируют в хромосому клетки-хозяина, а именно, в суспензионную клетку млекопитающего, и в случае использования транспозона Tol1.
(2) Изучение продукции антитела суспензионной клеткой CHO-K1
Эффективность продукции антитела суспензионной клеткой CHO-K1 изучали, используя суспензионную клетку CHO-K1. Транспозонный вектор для экспрессии антитела против белка M2 вируса гриппа человека (10 мкг) и вектор экспрессии транспозазы Tol1 (50 мкг) вводили методом электропорации в суспензионную клетку CHO-K1, адаптированную к суспензионному культивированию.
Затем клетки высевали в соответствующие два 96-луночных планшета и культивировали в течение 3 дней в CO2-инкубаторе, используя среду для суспензионного культивирования EX-CELL 325-PF. Со дня смены среды на 4-й день после электропорации клетки культивировали в течение 3 недель в присутствии 3 мкг/мл циклогексимида. В этом случае среду меняли каждую неделю.
В случае суспензионной клетки CHO-K1, 1×106 клеток высевали в 6-луночный планшет и культивировали со встряхиванием в CO2-инкубаторе в течение 3 дней, количество белка антитела против белка M2 вируса гриппа человека измеряли методом ВЭЖХ в культуральном супернатанте.
Концентрацию антитела в культуральном супернатанте определяли методом, описанным в Yeast Res., 7 (2007), 1307-1316. Результаты представлены на фигуре 7.
Как показано на фигуре 7, было получено большое количество клеток, демонстрирующих заметно повышенный уровень экспрессии антитела, и в случае использования транспозона Tol1. На основании этих результатов было установлено, что, как и в случае использования нуклеотидной последовательности из транспозона Tol2, суспензионную клетку млекопитающего, способную на высоком уровне экспрессировать представляющий интерес белок, можно также получать, если в качестве транспозонной последовательности использовать нуклеотидную последовательность из транспозона Tol1.
Хотя изобретение описано подробно и со ссылкой на конкретные варианты его осуществления, специалисту в данной области будет очевидно, что в изобретение можно вносить различные изменения и модификации без отхода от его сущности и объема. Данная заявка основана на японской заявке № 2009-140626, поданной 11 июня 2009 г., и предварительной патентной заявке США № 61/186138, поданной 11 июня 2009 г., полное содержание которых включено в данное описание посредством ссылок, и все литературные источники, цитированные в данном описании, включены в него посредством ссылок.
Промышленная применимость
С помощью способа получения белка по настоящему изобретению можно эффективно получать представляющий интерес белок, используя суспензионную клетку млекопитающего. Клетку по настоящему изобретению можно использовать в качестве продуцирующей белок клетки для получения рекомбинантного белка.
Список последовательностей
SEQ ID NO: 1 - Описание искусственной последовательности: нуклеотидная последовательность неавтономного транспозона Tol2
SEQ ID NO: 2 - Описание искусственной последовательности: последовательность Tol2-L
SEQ ID NO: 3 - Описание искусственной последовательности: последовательность Tol2-R
SEQ ID NO: 7 - Описание искусственной последовательности: нуклеотидная последовательность гена устойчивости к циклогексимиду
SEQ ID NO: 8 - Описание искусственной последовательности: аминокислотная последовательность белка, кодируемого геном устойчивости к циклогексимиду
SEQ ID NO: 9 - Описание искусственной последовательности: нуклеотидная последовательность, кодирующая H-цепь антитела M2Z3
SEQ ID NO: 9 - Описание искусственной последовательности: нуклеотидная последовательность, кодирующая H-цепь антитела M2Z3
SEQ ID NO: 10 - Описание искусственной последовательности: нуклеотидная последовательность, кодирующая H-цепь антитела M2Z3
SEQ ID NO: 11 - Описание искусственной последовательности: нуклеотидная последовательность, кодирующая L-цепь антитела M2Z3
SEQ ID NO: 12 - Описание искусственной последовательности: аминокислотная последовательность, кодирующая L-цепь антитела M2Z3
SEQ ID NO: 13 - Описание искусственной последовательности: нуклеотидная последовательность неавтономного транспозона Tol1
SEQ ID NO: 14 - Описание искусственной последовательности: последовательность Tol1-L
SEQ ID NO: 15 - Описание искусственной последовательности: последовательность Tol1-R.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ БЕЛКА | 2011 |
|
RU2598255C2 |
| УЛУЧШЕННЫЕ ЭКСПРЕССИЯ И ПРОЦЕССИНГ ТРАНСГЕНА | 2014 |
|
RU2808756C1 |
| Интегральный плазмидный вектор pVEAL2-B7, обеспечивающий экспрессию и секрецию рекомбинантного белка В7 вируса натуральной оспы (ВНО) в клетках млекопитающих, рекомбинантный штамм клеточной линии СНО-К1-В7 и рекомбинантный белок В7 ВНО, продуцируемый штаммом клеточной линии СНО-К1-В7 и используемый для получения иммунобиологических препаратов | 2023 |
|
RU2817380C1 |
| ЭЛЕМЕНТЫ РЕКОМБИНАНТНОГО ВЕКТОРА ЭКСПРЕССИИ (REVES) ДЛЯ УСИЛЕНИЯ ЭКСПРЕССИИ РЕКОМБИНАНТНЫХ БЕЛКОВ В КЛЕТКАХ-ХОЗЯЕВАХ | 2008 |
|
RU2518340C2 |
| ИНДУЦИРУЕМЫЕ КАСПАЗЫ И СПОСОБЫ ИСПОЛЬЗОВАНИЯ | 2017 |
|
RU2757058C2 |
| СПОСОБЫ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ РЕКОМБИНАНТНОГО АЛЬФА-1-АНТИТРИПСИНА (AAT) И КОМПОЗИЦИЙ НА ЕГО ОСНОВЕ | 2020 |
|
RU2817416C2 |
| ВЕКТОРЫ ДЛЯ ПРОИЗВОДСТВА БЕЛКОВ | 2019 |
|
RU2820218C2 |
| ЛИНИЯ КЛЕТОК, СОДЕРЖАЩАЯ НОВЫЙ МАРКЕР ОТБОРА, И ЕЕ ПРИМЕНЕНИЕ ДЛЯ ПРОДУЦИРОВАНИЯ БЕЛКОВ | 2020 |
|
RU2820830C2 |
| ПОЛУЧЕНИЕ ВИРУСНОГО ВЕКТОРА | 2014 |
|
RU2658487C2 |
| Интегративный плазмидный вектор pVEAL3-RBDdel, обеспечивающий синтез и секрецию рекомбинантного белка рецепторсвязывающего домена RBDdelta коронавируса SARS-CoV-2 в клетках млекопитающих, рекомбинантный штамм клеточной линии CHO-K1- RBDdelta и рекомбинантный белок RBDdelta SARS-CoV-2, продуцируемый штаммом клеточной линии | 2023 |
|
RU2816175C1 |




























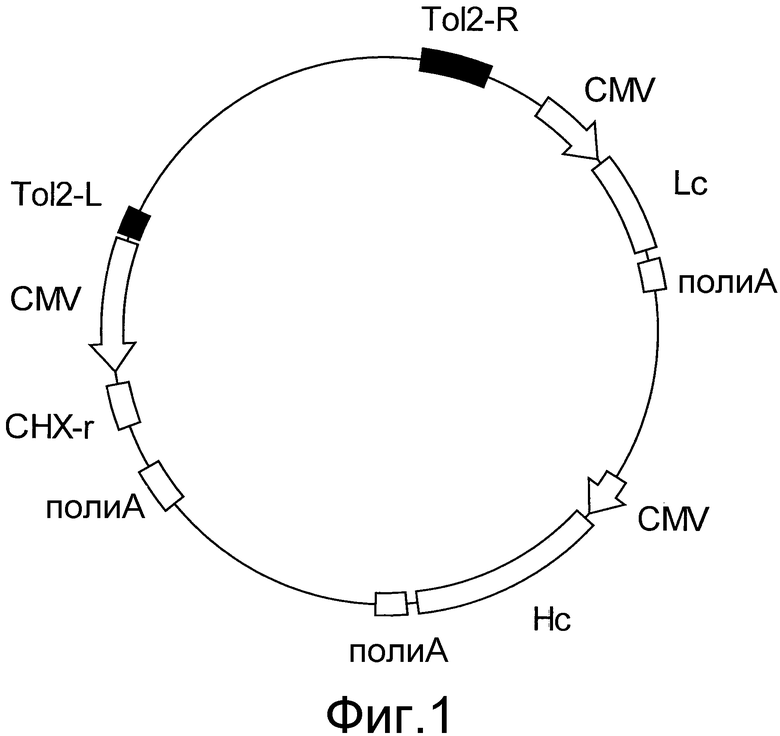

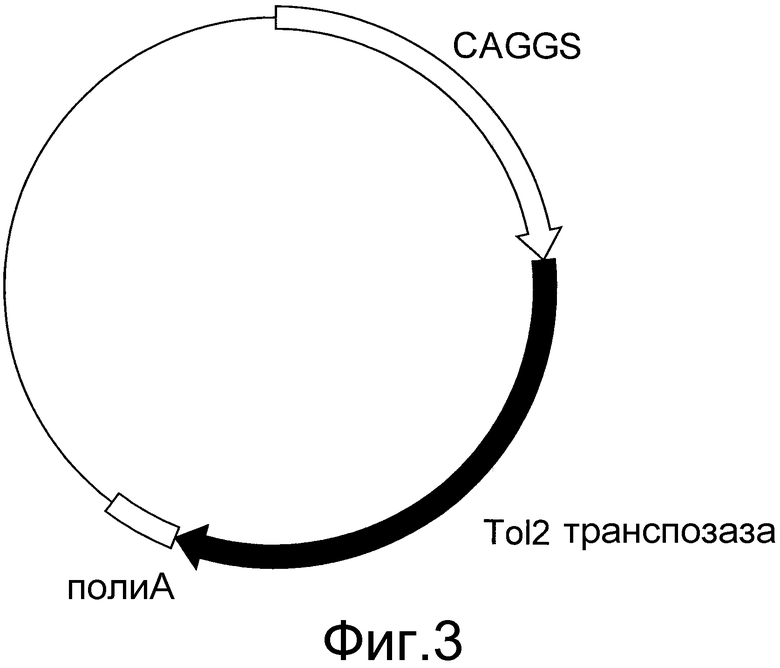
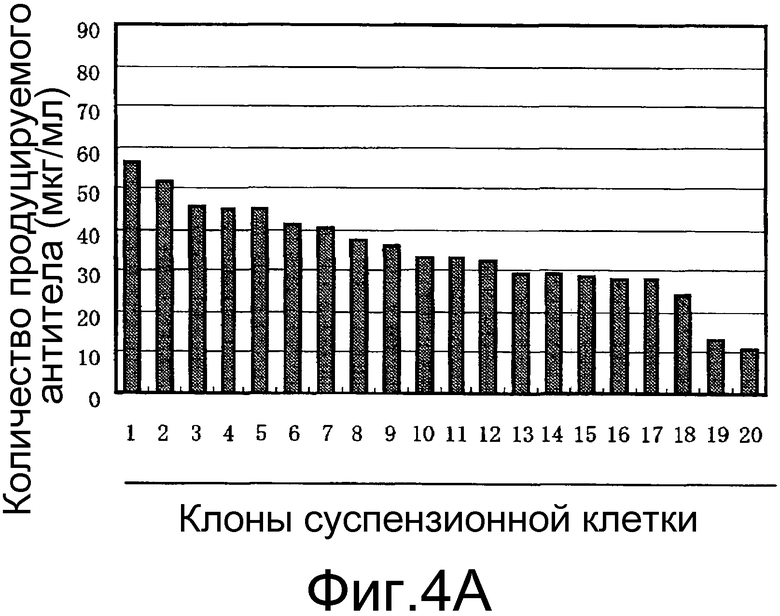
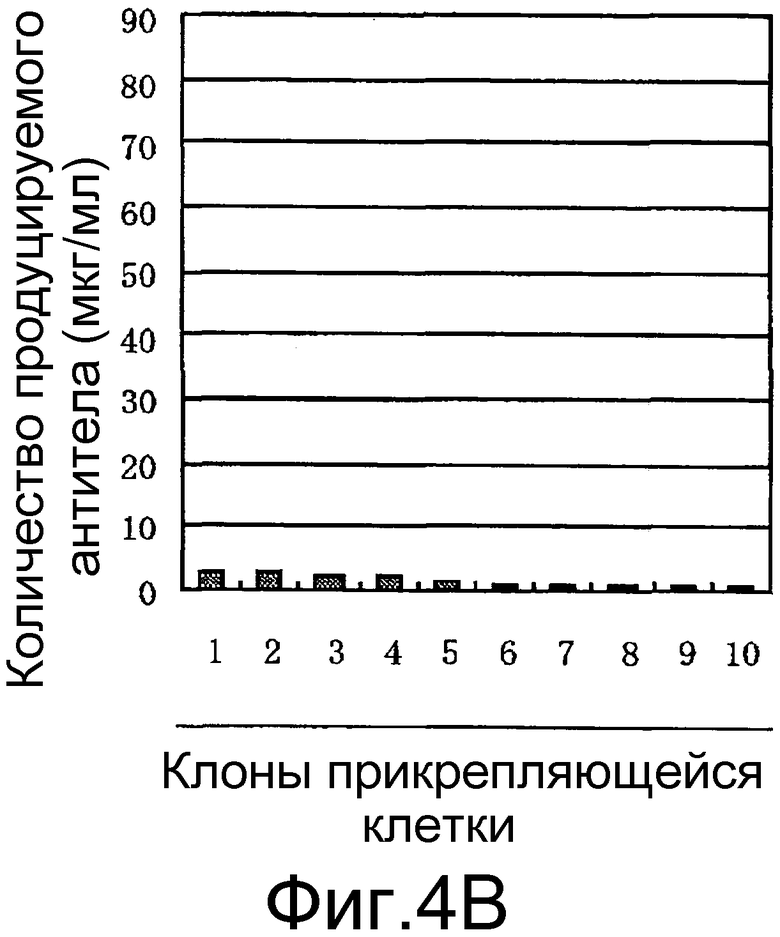
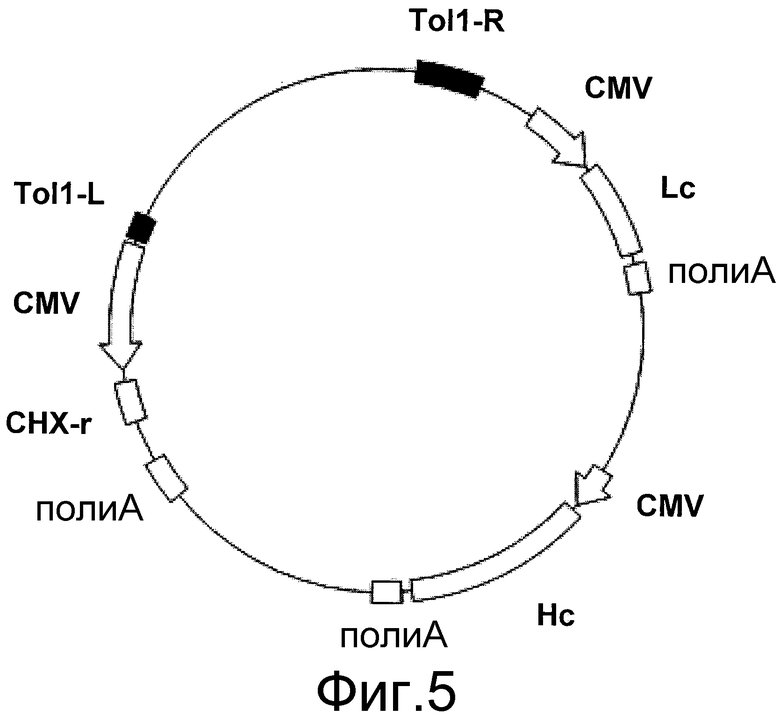
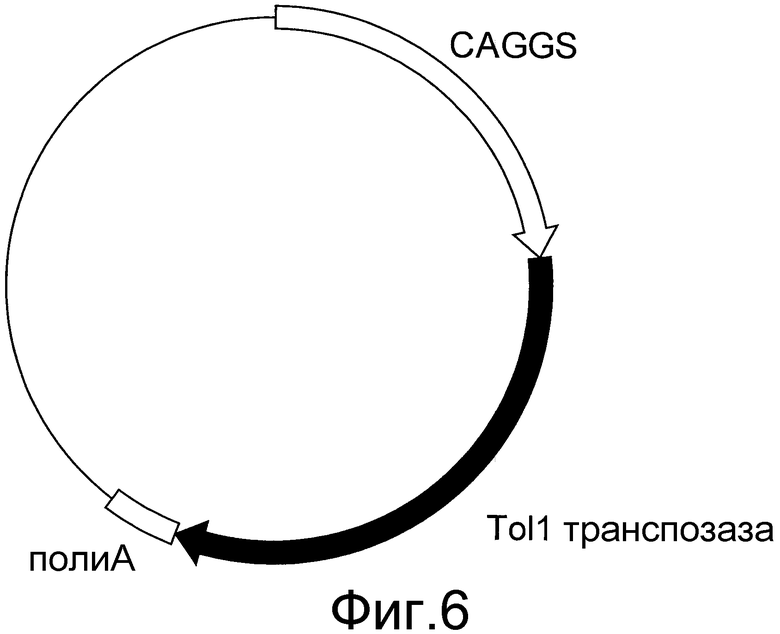
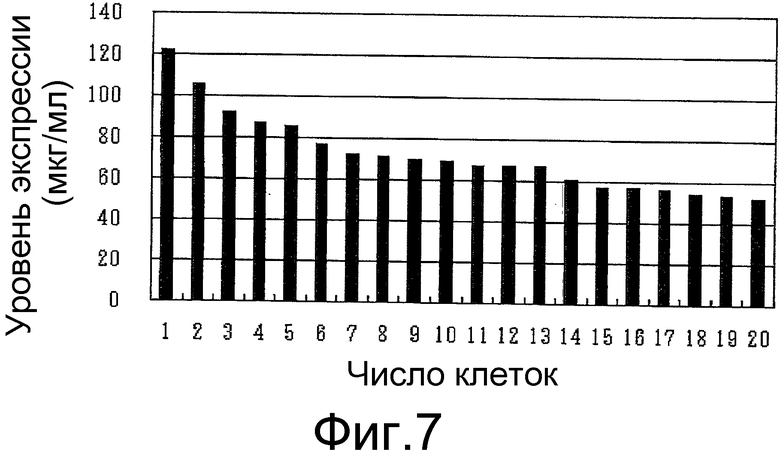
Изобретение относится к области молекулярной биологии и генетической инженерии. Предложен способ получения представляющего интерес белка, включающему введение вектора экспрессии белка, который включает генный фрагмент, содержащий ДНК, кодирующую представляющий интерес белок, и ген селектируемого маркера, а также транспозонные Tol1 или Tol2 последовательности на обоих концах генного фрагмента, в суспензионную клетку млекопитающего СНО, адаптированную к суспензионному культивированию, или клетку PER.C6, клетки крысиной миеломы YB2/3HL.Р2.G11.16Ag.20 (или также называемой YB2/0), или клетку мышиной миеломы NS0, адаптированную к суспензионному культивированию; интегрирование генного фрагмента, вставленного между парой транспозонных последовательностей, в хромосому клетки млекопитающего для получения клетки млекопитающего, способной экспрессировать представляющий интерес белок; и суспензионное культивирование клетки млекопитающего; при этом суспензионная клетка млекопитающего способна экспрессировать представляющий интерес белок, а также предложены способ получения клетки млекопитающего, соответствующая рекомбинантная клетка и применение вектора экспрессии. 6 н. и 23 з.п. ф-лы, 8 ил., 3 табл., 6 пр.
1. Способ получения представляющего интерес белка, включающий
введение вектора экспрессии белка, который содержит генный фрагмент, содержащий ДНК, кодирующую представляющий интерес белок, и ген селектируемого маркера, а также транспозонные последовательности на обоих концах генного фрагмента, в клетку млекопитающего, способную выживать и пролиферировать в бессывороточной среде;
интегрирование генного фрагмента, вставленного между парой транспозонных последовательностей, в хромосому клетки млекопитающего для получения клетки млекопитающего, способной экспрессировать представляющий интерес белок; и
суспензионное культивирование клетки млекопитающего,
где клетка млекопитающего, способная выживать и пролиферировать в бессывороточной среде, представляет собой по меньшей мере клетку, выбранную из клетки СНО, адаптированной к суспензионному культивированию, клетки PER.C6, клетки крысиной миеломы YB2/3HL.Р2.G11.16Ag.20 (или также называемой YB2/0) и клетки мышиной миеломы NS0, адаптированной к суспензионному культивированию,
где транспозонные последовательности представляют собой нуклеотидные последовательности, происходящие из пары транспозонов Tol1, или нуклеотидные последовательности, происходящие из пары транспозонов Tol2.
2. Способ получения представляющего интерес белка, включающий следующие стадии (А)-(С):
(A) стадию одновременного введения следующих векторов экспрессии (а) и (b) в клетку млекопитающего, способную выживать и пролиферировать в бессывороточной среде:
(a) вектор экспрессии, который содержит генный фрагмент, содержащий ДНК, кодирующую представляющий интерес белок, ДНК, кодирующую ген селектируемого маркера, и транспозонные последовательности на обоих концах генного фрагмента,
(b) вектор экспрессии, который содержит ДНК, кодирующую транспозазу, которая распознает транспозонные последовательности и обладает активностью переноса генного фрагмента, вставленного между парой транспозонных последовательностей, в хромосому,
(B) стадию временной экспрессии транспозазы с вектора экспрессии, введенного на этапе (А), для интегрирования генного фрагмента, вставленного между парой транспозонных последовательностей, в хромосому клетки млекопитающего с получением клетки млекопитающего, способной выживать и пролиферировать в бессывороточной среде и способной экспрессировать представляющий интерес белок, и
(C) стадию суспензионного культивирования клетки млекопитающего, способной выживать и пролиферировать в бессывороточной среде и способной экспрессировать представляющий интерес белок, полученной на стадии (В), с получением представляющего интерес белка,
где клетка млекопитающего, способная выживать и пролиферировать в бессывороточной среде, представляет собой по меньшей мере клетку, выбранную из клетки СНО, адаптированной к суспензионному культивированию, клетки PER.C6, клетки крысиной миеломы YB2/3HL.Р2.G11.16Ag.20 (или также называемой YB2/0) и клетки мышиной миеломы NS0, адаптированной к суспензионному культивированию,
где транспозонные последовательности представляют собой нуклеотидные последовательности, происходящие из пары транспозонов Tol1, или нуклеотидные последовательности, происходящие из пары транспозонов Тоl2.
3. Способ получения клетки млекопитающего, способной выживать и пролиферировать в бессывороточной среде и способной экспрессировать представляющий интерес белок, включающий
введение вектора экспрессии белка, который включает генный фрагмент, содержащий ДНК, кодирующую представляющий интерес белок, и ген селектируемого маркера, а также транспозонные последовательности на обоих концах генного фрагмента, в клетку млекопитающего, способную выживать и пролиферировать в бессывороточной среде; и
интегрирование генного фрагмента, вставленного между парой транспозонных последовательностей, в хромосому клетки млекопитающего,
где клетка млекопитающего, способная выживать и пролиферировать в бессывороточной среде, представляет собой по меньшей мере клетку, выбранную из клетки СНО, адаптированной к суспензионному культивированию, клетки PER.C6, клетки крысиной миеломы YB2/3HL.Р2.G11.16Ag.20 (или также называемой YB2/0) и клетки мышиной миеломы NS0, адаптированной к суспензионному культивированию,
где транспозонные последовательности представляют собой нуклеотидные последовательности, происходящие из пары транспозонов Tol1, или нуклеотидные последовательности, происходящие из пары транспозонов Tol2.
4. Способ по любому из пп. 1-3, в котором клетка СНО представляет собой по меньшей мере одну клетку, выбранную из CHO-К1, CHO-K1SV, DUKXB11, CHO/DG44, Pro-3 и CHO-S.
5. Способ по любому из пп. 1-3, в котором ген селектируемого маркера представляет собой ген устойчивости к циклогексимиду.
6. Способ по п. 5, в котором ген устойчивости к циклогексимиду представляет собой ген, кодирующий мутант человеческого рибосомного белка L36a.
7. Способ по п. 6, в котором мутант представляет собой мутант, в котором пролин в положении 54 человеческого рибосомного белка L36a заменен на глутамин.
8. Способ по любому из пп. 1-3, в котором пара транспозонных последовательностей представляет собой нуклеотидные последовательности, происходящие из пары транспозонов ДНК-типа, функционирующих в клетке млекопитающего.
9. Способ по п. 8, в котором нуклеотидные последовательности, происходящие из пары транспозонов Tol2, представляют собой нуклеотидную последовательность, содержащую нуклеотидную последовательность SEQ ID NO: 2 и нуклеотидную последовательность SEQ ID NO: 3.
10. Способ по п. 8, в котором нуклеотидные последовательности, происходящие из пары транспозонов Tol1, представляют собой нуклеотидную последовательность SEQ ID NO: 14 и нуклеотидную последовательность SEQ ID NO: 15.
11. Рекомбинантная клетка млекопитающего для получения представляющего интерес белка, где указанная клетка способна выживать и пролиферировать в бессывороточной среде и способна продуцировать представляющий интерес белок, в которую введен вектор экспрессии белка, который включает генный фрагмент, содержащий ДНК, кодирующую представляющий интерес белок, и ген селектируемого маркера, а также транспозонные последовательности на обоих концах генного фрагмента, для интегрирования генного фрагмента, вставленного между парой транспозонных последовательностей, в хромосому,
где клетка млекопитающего, способная выживать и пролиферировать в бессывороточной среде, представляет собой по меньшей мере клетку, выбранную из клетки СНО, адаптированной к суспензионному культивированию, клетки PER.C6, клетки крысиной миеломы YB2/3HL.Р2.G11.16Ag.20 (или также называемой YB2/0) и клетки мышиной миеломы NS0, адаптированной к суспензионному культивированию,
где транспозонные последовательности представляют собой нуклеотидные последовательности, происходящие из пары транспозонов Tol1, или нуклеотидные последовательности, происходящие из пары транспозонов Tol2.
12. Клетка по п. 11, где клетка СНО представляет собой по меньшей мере одну клетку, выбранную из СНО-К1, CHO-K1SV, DUKXB11, CHO/DG44, Pro-3 и CHO-S.
13. Клетка по п. 11, в которой ген селектируемого маркера представляет собой ген устойчивости к циклогексимиду.
14. Клетка по п. 13, в которой ген устойчивости к
циклогексимиду представляет собой ген, кодирующий мутант человеческого рибосомного белка L36a.
15. Клетка по п. 14, в которой мутант представляет собой мутант, в котором пролин в положении 54 человеческого рибосомного белка L36a заменен на глутамин.
16. Клетка по п. 11, в которой пара транспозонных последовательностей представляет собой нуклеотидные последовательности, происходящие из пары транспозонов ДНК-типа, функционирующих в клетке млекопитающего.
17. Клетка по п. 16, в которой нуклеотидные последовательности, происходящие из пары транспозонов Tol2, представляют собой нуклеотидную последовательность SEQ ID NO: 2 и нуклеотидную последовательность SEQ ID NO: 3.
18. Клетка по п. 16, в которой нуклеотидные последовательности, происходящие из пары транспозонов Tol1, представляют собой нуклеотидную последовательность SEQ ID NO: 14 и нуклеотидную последовательность SEQ ID NO: 15.
19. Рекомбинантная клетка млекопитающего для получения представляющего интерес белка, где указанная клетка способна выживать и пролиферировать в бессывороточной среде и способна продуцировать представляющий интерес белок, в которую введен вектор экспрессии (а), включающий генный фрагмент, содержащий ДНК, кодирующую представляющий интерес белок, и ген селектируемого маркера, а также транспозонные последовательности на обоих концах генного фрагмента, и вектор экспрессии (b), содержащий ДНК, кодирующую транспозазу (трансферазу), которая распознает транспозонные последовательности и обладает
активностью переноса генного фрагмента, вставленного между парой транспозонных последовательностей, в хромосому для
интегрирования генного фрагмента, вставленного между парой транспозонных последовательностей, в хромосому,
где клетка млекопитающего, способная выживать и пролиферировать в бессывороточной среде, представляет собой по меньшей мере клетку, выбранную из клетки СНО, адаптированной к суспензионному культивированию, клетки PER.C6, клетки крысиной миеломы YB2/3HL.Р2.G11.16Ag.20 (или также называемой YB2/0) и клетки мышиной миеломы NS0, адаптированной к суспензионному культивированию,
где транспозонные последовательности представляют собой нуклеотидные последовательности, происходящие из пары транспозонов Tol1, или нуклеотидные последовательности, происходящие из пары транспозонов Tol2.
20. Клетка по п. 19, где клетка СНО представляет собой по меньшей мере одну клетку, выбранную из СНО-К1, CHO-K1SV, DUKXB11, CHO/DG44, Pro-3 и CHO-S.
21. Клетка по п. 19, в которой ген селектируемого маркера представляет собой ген устойчивости к циклогексимиду.
22. Клетка по п. 21, в которой ген устойчивости к циклогексимиду представляет собой ген, кодирующий мутант человеческого рибосомного белка L36a.
23. Клетка по п. 22, в которой мутант представляет собой мутант, в котором пролин в положении 54 человеческого рибосомного белка L36a заменен на глутамин.
24. Клетка п. 19, в которой пара транспозонных последовательностей представляет собой нуклеотидные последовательности, происходящие из пары транспозонов ДНК-типа, функционирующих в клетке млекопитающего.
25. Клетка по п. 24, в которой нуклеотидные последовательности, происходящие из пары транспозонов Tol2, представляют собой нуклеотидную последовательность SEQ ID NO: 2 и нуклеотидную последовательность SEQ ID NO: 3.
26. Клетка по п. 24, в которой нуклеотидные последовательности, происходящие из пары транспозонов Tol1, представляют собой нуклеотидную последовательность SEQ ID NO: 14 и нуклеотидную последовательность SEQ ID NO: 15.
27. Применение вектора экспрессии белка, включающего генный фрагмент, содержащий ДНК, кодирующую представляющий интерес белок, и ген селектируемого маркера, а также транспозонные последовательности на обоих концах генного фрагмента,
где транспозонные последовательности представляют собой нуклеотидные последовательности, происходящие из пары транспозонов Tol1, или нуклеотидные последовательности, происходящие из пары транспозонов Tol2, для интегрирования генного фрагмента, вставленного между транспозонными последовательностями, в хромосому суспензионной СНО клетки, способной выживать и пролиферировать в бессывороточной среде.
28. Применение вектора экспрессии белка по п. 27, в котором нуклеотидные последовательности, происходящие из пары транспозонов Tol2, представляют собой нуклеотидную последовательность SEQ ID NO: 2 и нуклеотидную последовательность SEQ ID NO: 3.
29. Применение вектора экспрессии белка по п. 27, в котором нуклеотидные последовательности, происходящие из пары транспозонов Tol1, представляют собой нуклеотидную последовательность SEQ ID NO: 14 и нуклеотидную последовательность SEQ ID NO: 15.
| KAWAKAMI K., To12:a versatile gene transfer vector in vertebrates, Genome Biology, 2007, Vol | |||
| Топка с несколькими решетками для твердого топлива | 1918 |
|
SU8A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| ANDREAS LACKNER, A bicistronic baculovirus vector for transient and stable protein expression in mammalian cells, Analytical Biochemistry, 2008, pages 146-148 | |||
| EP 243153 A, 22.04.1987 | |||
| WO 1991009313 A, 27.06.1991 | |||
| CN | |||

