ПРИМЕНЕНИЕ
Настоящее изобретение относится к способу получения не являющегося аденовирусом вируса-мишени или белков-мишеней с использованием сильной экспрессирующей клеточной линии, обладающей стабильно интегрированным в ее геном геном, кодирующим специфичный гетерологичный регуляторный белок.
Известный уровень техники
Биофармацевтические продукты из эукариотических клеток являются составной частью современной медицины. Однако повышенная продуктивность и повышенная безопасность представляют собой важные параметры, которые все еще требуют существенной оптимизации. Наиболее значимым узким местом в попытках оптимизации является сам клеточный субстрат. Безопасный трансген, который может быть введен в клеточные линии, уже выпущенный для применения в биофармацевтических процессах для повышения выхода продуктов из таких обработанных клеток, является предельно дорогим.
Аденовирусы являются безоболочечными (голыми) вирусами с двухцепочечной ДНК, которые инфицируют широкий спектр животных. Среди наиболее хорошо охарактеризованных членов можно назвать аденовирус серотипа 5 (Ad5). Этот вирус является частой причиной общих симптомов охлаждения, и инфицирование часто происходит в детстве.
Аденовирусы поддаются генетическим манипуляциям, и не способные к репликации аденовирусы, включая Ad5 (номер поступления в GenBank для последовательности: AC_000008), используются в качестве векторов для генной терапии и для терапевтической вакцинации. Для получения не способных к репликации вирусов большие области геномной ДНК замещают невирусными последовательностями. Потеря вирусных функций обеспечивается транс-действием линий клеток-хозяев, которые стабильно трансфицированы генами, которые удалены в векторе. Одна из ранних систем, которые были разработаны, состоит из аденовирусных векторов с делецией регуляторной области E1, созданной в клеточных линиях, обеспечивающих соответствующими белками E1.
Наиболее обычной клеточной линией для этой цели является клеточная линия 293. Эта линия была создана в 1977 г. путем трансфекции фрагментированной аденовирусной геномной ДНК в первичные клетки человека (Graham et al., J. Gen. Virol. 36, 59-74 (1977)), как раз перед тем как было обнаружено, что аденовирусы могут служить в качестве векторов для генной терапии. Полученная клеточная линия, как показано впоследствии, содержала нуклеотиды с 1 по 4344 геномной ДНК (Louis et al., Virology 233, 423-429 (1997)), которая включает область E1; эта характеристика показала, что область E1 может быть использована для иммортализации и трансформации первичных клеток.
Сразу за областью E1 следует ген для pIX («белка 9»; нуклеотиды с 3609 до 4031 в геномной ДНК). Промотор для pIX встроен в компонент E1B области E1. Механизм, называемый окклюзией промотора, позволяет экспрессироваться pIX в раннее отставленное время в цикле инфицирования вирусом при начале репликации вирусной ДНК с увеличивающимся числом копий вирусной ДНК (Fessler and Young, J. Virol. 72, 4049-4056 (1998)). Хотя ген присутствует в пределах 4344 нуклеотидов, интегрированных в клетки 293, экспрессия pIX не может быть определена даже чувствительными методами с радиоактивной меткой (Spector et al., J. Virol. 36, 860-871 (1980)).
Как описано выше, аденовирусные векторы с недостаточной репликацией созданы с помощью делеции области E1 из вирусного генома. Продукты E1 существенны для репликации вирусов, и функция области E1, следовательно, должна состоять в обеспечении транс-действия через стабильные трансгены E1 в клетке, упаковывающей аденовирус (такой как клеточная линия 293). Благодаря проксимальному положению в отношении области E1 ген pIX вызывал частую рекомбинацию между аденовирусными векторами с делецией и E1 в геноме хозяина. Это событие рекомбинации создавало компетентный в отношении репликации аденовирус (RCA), существенное загрязнение при получении векторов. Для подавления этого события рекомбинации pIX удаляли из аденовирусного генома. В ходе этих экспериментов было установлено, что pIX стабилизирует аденовирусный капсид в отношении термического и стерического стресса путем усиления взаимодействия главных строительных блоков, гексонов (Colby and Shenk, J. Virol. 39, 977-980 (1981); Ghosh-Choudhury et al., EMBO J. 6, 1733-1739 (1987)). Морфогенез в отсутствие pIX дает чувствительные к температуре вирусы, которые не могут доставлять молекулы геномной ДНК более крупные, чем 105% от 35938 пар оснований у дикого типа. Для того чтобы все еще придать нормальную упаковывающую способность и термическую стабильность, белок pIX стабильно вводят в клеточные линии, предназначаемые в качестве упаковывающих клеток для аденовирусных векторов (Krougliak and Graham, Hum. Gene Ther. 6, 1575-1586 (1995); патент WO 99/57296 и Imler et al., Gene Therapy 3, 75-84 (1996)).
Также при выяснении того, что pIX отделывает поверхность вирионов, были созданы слитые белки pIX с целью расширения диапазона хозяев аденовирусных векторов или для прослеживания морфогенеза и внутриклеточного перемещения вирусных частиц (суммировано Parks, Mol. Ther. 11, 19-25 (2005)).
Несмотря на то, что он является чисто структурным белком, pIX как регуляторный белок вовлечен также в репликацию аденовирусов. PIX, как предполагается, функционирует в качестве трансактиватора транскрипции для усиления экспрессии E1A, функции, возможно, даже оказывающей влияние в качестве вирокина путем поступления белка PIX из капсида на стадии инфицирования (суммировано Parks, Mol. Ther. 11, 19-25 (2005)). PIX совместно с ранним белком E4 Orf3 так же, как описано, взаимодействует с субъядерными включениями, называемыми тельцами PML (Puvion-Dutilleul et al., Exp. Cell. Res. 218, 9-16 (1995); Leppard and Everett, J. Gen. Virol. 80, 997-1008 (1999)). Они представляют собой динамические агрегаты размером от 250 нм до 500 нм и, как предполагается, участвуют в регуляции клеточной дифференцировки (Wang et al., Science 279, 1547-1551 (1998)), контроле апоптоза (Quignon et al., Nature Gen. 20, 259-265 (1998)) и в ответ на вирусную инфекцию (Moller and Schmitz, Arch Immunol. Ther. Exp. (Warsz) 51, 295-300 (2003)).
Из-за его плейотропных эффектов заявители ввели белок PIX в клеточные линии для изучения того, будет ли PIX обладать свойствами усиления клеточной пролиферации или продукции для биофармацевтических продуктов, которые не относятся к аденовирусу или аденовирусным векторам. Существует повсеместная потребность в факторах, которые модулируют эти свойства в устоявшихся клеточных линиях.
Например, аттенуированные (ослабленные) вирусы являются многообещающими кандидатами для вакцин: при инокуляции они имитируют природную инфекцию, но предоставляют больше времени для установления желаемого защитного иммунного ответа путем вакцинации. При растущем количестве иммунодефицитных больных (например, из-за ВИЧ-инфекции) желательны крайне ослабленные штаммы. Поскольку ослабленные штаммы все еще продолжают инфицировать (обычно в легкой форме), высоко ослабленные штаммы блокируются на клеточном уровне даже в отсутствие функционирующей иммунной системы. Часто используемым методом для установления и поддержания аттенуации являются пассажи вирусов на различных тканях хозяина. Например, вирусы кори и свинки, предназначенные для вакцинации человека, пассируют в первичных клетках либо яиц с эмбрионами цыплят, либо происходящих из них культур. Новая генерация вакцин основана на сильно ослабленных поксвирусах, продукция которых также зависит от первичных клеток цыплят. Клеточная линия, которая может заменить первичные клетки цыплят и в то же время которая даже менее эффективно защищает себя против вирусной инфекции из-за вторичных манипуляций, таких как введение трансгена PIX, следовательно, дает крайне желаемый субстрат.
Для других целей может быть предпочтительна клеточная линия млекопитающих (а не птиц). Такие предпочтительные клеточные линии уже существуют и прошли проверку органов здравоохранения в отношении безопасности и рисков, установленных для производных биофармацевтических продуктов. Здесь также крайне желательна вторичная манипуляция (такая как введение трансгена PIX) для повышения спектра доступных применений или эффективности продукции без компрометации характеристик безопасности.
Краткое изложение существа изобретения
При получении клеточных линий, которые стабильно трансфицированы аденовирусным геном pIX (или его химерным гибридным аналогом), заявители неожиданно обнаружили, что pIX проявляет фенотипический эффект в клетках птиц и человека. Для клеток птиц это особенно удивительно, потому что клетки птиц не могут быть инфицированы аденовирусами человека и авиаденовирусы (такие как CELO или аденовирус типа 8 домашней птицы) не кодируют гомолог PIX (Ojkic and Nagy, J. Gen. Virol. 81, 1833-1837 (2000)). Более того, заявители обнаружили, что стабильное присутствие PIX повышает восприимчивость клетки к индукции аналогом двухцепочечной РНК, вероятно, через толл-подобный рецептор 3. В этом контексте, вероятно, заявители также неожиданно обнаружили, что присутствие белка PIX повышает выходы высоко ослабленного поксвируса в клетках-хозяевах птиц. Так как клетки, инфицированные поксвирусом, меньше страдали от индукции аналогом двухцепочечной РНК, заявители могли найти взаимодействие между белком PIX и антиинтерфероновыми генами поксвируса, которое не было описано ранее. Заявители также выявили неожиданное увеличение выхода белкового продукта (не только вируса), секретируемого стабильно трансфицированной клеточной линией.
Таким образом, настоящее изобретение относится к
(1) способу получения неаденовирусного вируса-мишени или одного или более белков-мишеней, включающему
(a) культивирование экспрессирующей клетки, получаемой с помощью инфицирования или трансфекции клетки-хозяина, обладающей стабильно интегрированным в ее геном геном, кодирующим PIX аденовируса или его функциональный вариант в качестве гетерологичного регуляторного белка, и стабильно экспрессирующей указанный регуляторный белок или его функциональный вариант, с указанным вирусом-мишенью или с вектором, несущим последовательности нуклеиновой кислоты, кодирующие указанный вирус-мишень, или с вектором, несущим последовательности нуклеиновой кислоты, кодирующие указанные один или более белки-мишени (где не аденовирусный вирус-мишень не содержит указанный регуляторный белок, и белок(и)-мишень(и) отличаются от указанного регуляторного белка или его функционального варианта), и
(b) выделение вируса-мишени или белка(ов)-мишени(ей);
(2) предпочтительному варианту осуществления (1), указанного выше, где указанный белок PIX или его функциональный вариант кооперируется с неродственными вирусными факторами, модулирует субклеточное распределение факторов, отличных от указанного белка, модулирует транскрипцию и/или рост клеток и повышает продуктивность клетки в отношении продукции вируса, не содержащего указанный регуляторный белок, и в отношении продукции белка, отличающегося от указанного регуляторного белка или его функционального варианта;
(3) предпочтительному варианту осуществления (1) или (2), указанных выше, где pIX аденовируса предлагается как слитый белок с другим белком, который модулирует или увеличивает активность или субклеточное распределение pIX аденовируса, например, гибридизованный с рецептором альфа-ретиноевой кислоты, предпочтительно, представляет собой слитый белок, имеющий последовательность, представленную в SEQ ID NO:4, или гибридизованный с GFP и содержащий NLS, предпочтительно, представляет собой слитый белок, имеющий последовательность, представленную в SEQ ID NO:23;
(4) предпочтительному варианту осуществления с (1) по (3), указанных выше, где хозяин и экспрессирующая клетка представляют собой клетку позвоночных, включая клетки млекопитающих и клетки птиц, предпочтительно клетка млекопитающих представляет собой клетку, происходящую из мозга человека, и клетка птиц представляет собой клетку, происходящую из сетчатки уток, или клетку, происходящую из сомитов утки;
(5) предпочтительному варианту осуществления с (1) по (4), указанных выше, где хозяин и экспрессирующая клетка происходят из мозга человека, предпочтительно из мозга плода человека, и наиболее предпочтительно представляют собой клетку NC5T11, и указанная клетка несет последовательность нуклеиновой кислоты, кодирующую PIX аденовируса или его функциональный вариант в качестве гетерологичного регуляторного белка, предпочтительно указанный гетерологичный регуляторный белок кодируется нуклеиновой кислотой, представленной SEQ ID NO:1, SEQ ID NO:3 или SEQ ID NO:22;
(6) предпочтительному варианту осуществления по (5), указанному выше, где клетка-хозяин представляет собой клетку NC5T11#34 и экспрессирующая клетка происходит от клетки NC5T11#34, причем указанная клетка NC5T11#34 депонирована с DSMZ под номером поступления DSM ACC2744;
(7) предпочтительному варианту осуществления с (1) по (4), указанных выше, где хозяин и экспрессирующая клетка представляют собой клетку птиц, предпочтительно происходящую из сетчатки утки или сомита утки, и указанная клетка несет последовательность нуклеиновой кислоты, кодирующую PIX аденовируса или его функциональный вариант в качестве гетерологичного регуляторного белка, предпочтительно указанный гетерологичный регуляторный белок кодируется нуклеиновой кислотой, представленной SEQ ID NO:1, SEQ ID NO:3 или SEQ ID NO:22;
(8) предпочтительному варианту осуществления по (7), указанному выше, где клетка-хозяин представляет собой клетку CR.PIX (17a11b) и экспрессирующая клетка происходит от клетки CR.PIX (17a11b), причем указанная клетка CR.PIX (17a11b) депонирована с DSMZ под номером поступления DSM ACC2749;
(9) экспрессирующей клетке для продукции вируса-мишени или одного или более белков-мишеней, как определено в с (1) по (8) выше, предпочтительно экспрессия является такой, как определено в с (5) по (8) выше;
(10) способу получения экспрессирующей клетки, как определено в (9) выше, который включает инфицирование или трансфекцию линии клеток-хозяев, как определено в с (1) по (8) выше, вирусом или вектором, несущим последовательности нуклеиновой кислоты, кодирующие указанный вирус, или вектором, несущим последовательности нуклеиновой кислоты, кодирующие указанный один или более белков-мишеней;
(11) клетке-хозяину, как определено в с (5) по (8) выше;
(12) способу получения клетки-хозяина по (11) выше, который включает трансфекцию подходящей исходной клетки вектором, несущим указанный регуляторный белок или его функциональный вариант;
(13) применению экспрессирующей клетки, как определено в (7) выше, для получения вируса-мишени или одного или более белков-мишеней;
(14) слитому белку, включающему по меньшей мере один первый домен, включающий регуляторный белок, и по меньшей мере один второй домен, включающий белок или пептид, действующий в качестве модулятора транскрипции и/или в качестве сигнала для субклеточной направленности, как определено в (2) или (3) выше; и
(15) нуклеотидной последовательности, кодирующей слитый белок, как определено в (14) выше.
Краткое описание фигур
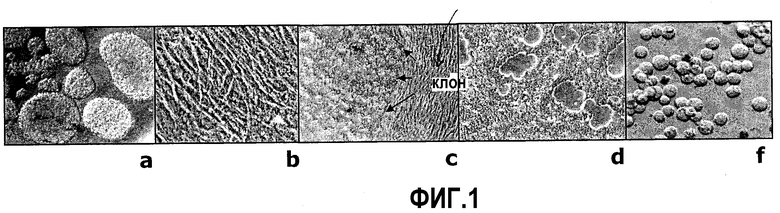
Фиг.1: история клеточной линии NC5T11. A - Нейросферы первичных клеток, выращенных в бессывороточных условиях. B - Монослой первичных клеток, выращенных в DMEM/F12 с 5% FCS. C - Первичный фокус через две недели после трансфекции p79. Стрелки указывают границу между первичными и иммортализованными клетками. D - Гомогенная адгезивная культура иммортализованных клеток в DMEM/F12 с 5% FCS. E - Суспензионная культура NC5T11 во встряхиваемых пробирках в EXcell VPRO.
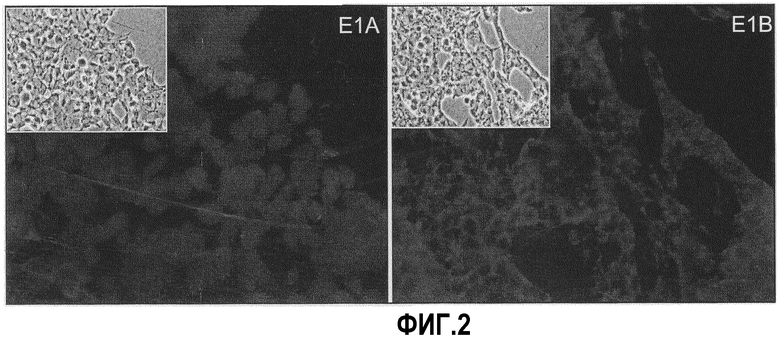
Фиг.2: Иммунофлуоресцентное определение E1A и E1B в NC5T11. После фиксации в метаноле клетки обрабатывали крысиными антителами, направленными против E1A и E1B 55k соответственно с последующей обработкой техасским красным, конъюгированным с антикрысиными антителами. Как показано на фиг.2, все клетки в образце демонстрируют ядерное окрашивание, типичное E1A и цитоплазматическое окрашивание для E1B. После фиксации метанолом клетки обрабатывали крысиными антителами, направленными против E1A и E1B 55k соответственно с последующей обработкой техасским красным, конъюгированным с антикрысиными антителами. Как здесь показано, все клетки в образце демонстрируют ядерное окрашивание, типичное E1A и цитоплазматическое окрашивание для E1B.
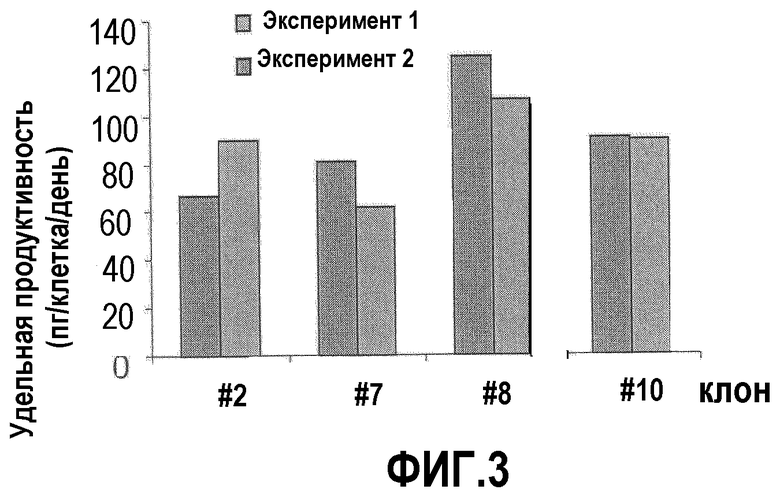
Фиг.3: Конкретная продуктивность отобранных клонов aat. Клеточные клоны NC5T11, трансфицированные C55 и отобранные с помощью пуромицина, высевали по 7×104 клеток в 12-луночные планшеты, число клеток и титр aat определяли на 1, 2 и 3 день после высевания и рассчитывали максимальную продуктивность конкретных клеток в день.
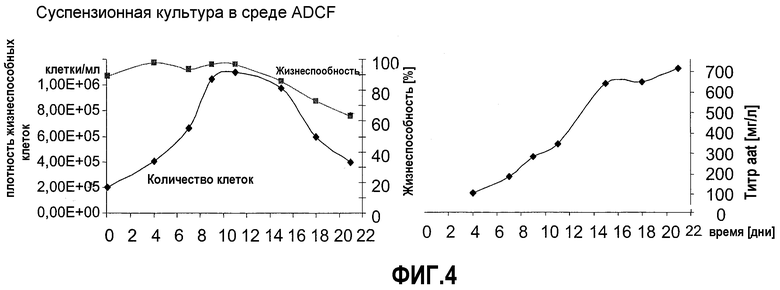
Фиг.4: Экспрессионный анализ NC5T11puro#8 во встряхиваемой суспензионной культуре. Клетки высевали в 12 мл EXCELL VPRO при 6×104 клеток/мл в 50-мл полипропиленовых пробирках для встряхивания (TPP, Switzerland) и подвергали вращению с радиусом 1 см и скоростью 160 об/мин. Образцы 200 мкл отбирали на 4, 7, 9, 11, 15, 18 и 21 дни. Плотность и жизнеспособность клеток определяли после окрашивания трипановым-синим с применением гемоцитометра.
Фиг.5: Определение кДНК pIX-RARA в пуле клонов с помощью ПЦР. Пул клонов, несущих ген pIX-RARA, создавали из NC5Tllpuro#8 с помощью трансфекции F67 и отбора с помощью гигромицина. РНК экстрагировали с применением набора для экстракции РНК (Machery Nagel, Germany) и кДНК синтезировали с применением обратной транскриптазы AMV (Invitrogen). кДНК для одного фрагмента pIX или целого гена pIXRARA амплифицировали с праймером 1 и 1 и 164 соответственно.
Фиг.6: Определение последовательностей pIX как части слитого белка в устойчивых к гигромицину субклонах NC5T11puro#8 и NC5T11 (#34, 35, 36, 37, 38) с помощью ПЦР. ДНК выделяли из клеточных клонов, выращенных в 6 лунках с помощью лизиса в ДДС-Na, с последующей экстракцией фенолом и преципитацией. ДНК амплифицировали с помощью праймеров 1 и 2 в течение 28 циклов.
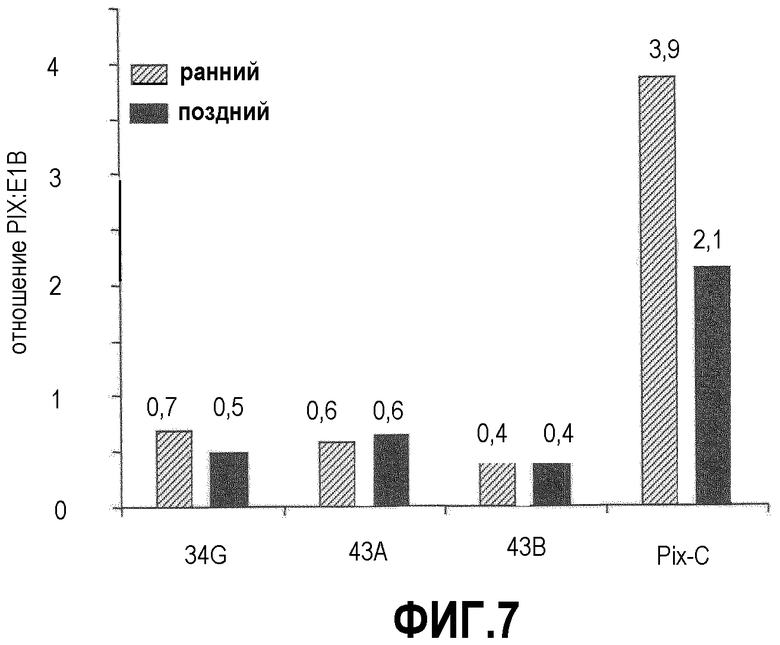
Фиг.7: Стабильность генов pIXRARA и pIX в стабильно трансфицированных NC5T11 определяли относительно E1B: E1A+E1B и pIXRARA вводили в отдельных трансфекциях, и гены E1 поддерживались в отсутствие селекции в течение >2 лет и, следовательно, рассматривались как стабильно интегрированные. Геномную ДНК выделяли из выбранных клеточных клонов в 2 временные точки: непосредственно сразу после прекращения селекции hyg (ранняя); через 2 месяца в отсутствие процедуры селекции (поздняя). Уровни ДНК PIX и E1B определяли с помощью ПЦР в реальном времени.
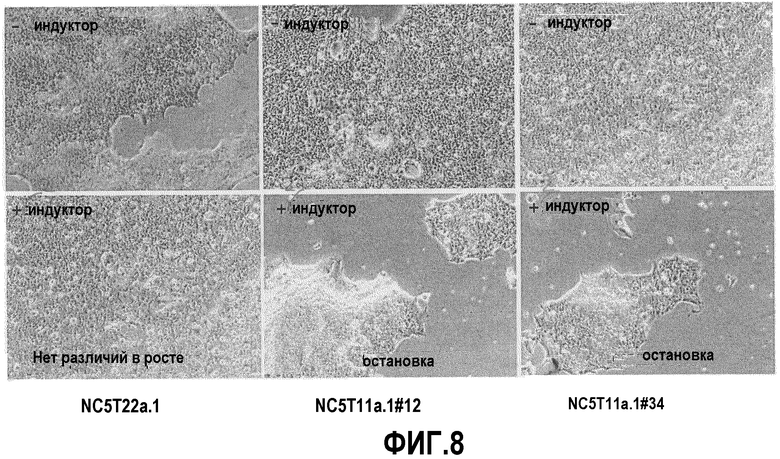
Фиг.8: Задержка роста, индуцированная обработкой RA в рекомбинантных pIXRARA клонах NC5T11 и NC5T11puro#8. Клетки высевали при 2×105 в 6-луночные планшеты в присутствии или в отсутствие 6 мкг/мл ретиноевой кислоты. Фотографии, демонстрирующие задержку роста в обработанных RA клонах #12 и #34, но не в NC5T11, были получены через 4 дня после высевания с использованием фазово-контрастного изображения при увеличении х4.
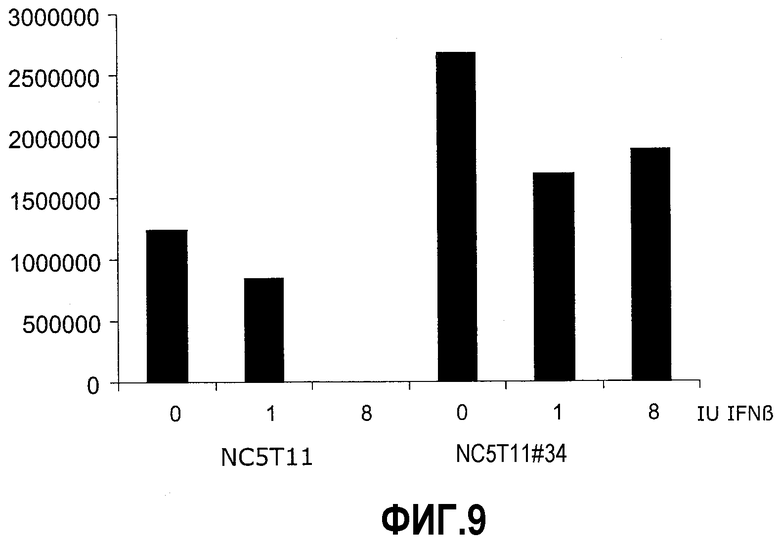
Фиг.9: PIXRARA предотвращает ингибирование репликации вируса интерфероном. Клетки NC5T11 и NC5T11#34 инфицировали EMCV, чувствительным к интерферону вирусом, при MOI 0,004 в клетках, обработанных интерфероном-бета. Лизаты клеток подвергали титрованию по образованию бляшек на клетках A549.
Фиг.10: Экспрессионный анализ клеточной линии NC5T11puro#8, экспрессирующей альфа-1-антитрипсин с вектора C55, и ее субклонов #10, #12, #14, несущих вектор F67 (pEFpIX-RARA) в дополнение к C55. Клетки высевали в EXCELL VPRO (JRH Biosciences) при 6×104 клеток/мл в 50 мл полипропиленовых пробирках для встряхивания (TPP, Switzerland) в суммарном объеме 12 мл и подвергали вращению с радиусом 1 см и скоростью 160 об/мин.
Фиг.11: Экспрессия PIX в клетках CR сетчатки утки. Левая панель: реакция ПЦР против гена PIX в геномной ДНК клеток pIX. Нанесение на гель слева направо: маркер 1 т.п.н. (Invitrogen); ПЦР на геномной ДНК из CRpIX; нематричный контроль; позитивный контроль с плазмидой, применяемой при трансфекции CRpIX. Правая панель: вестерн-блоттинг для определения белка pIX в клетках CRpIX. Дорожка 1: клетки 293; дорожка 2: белок pIX в клетках CRpIX.
Фиг.12: Стабильное поддержание трансгена PIX в клетках сетчатки уток. Верхняя левая панель: MCX и DXS представляют собой независимые аликвоты клона CRpIX, культивируемые в течение >3 месяцев параллельно и без селекции. После трех месяцев ПЦР TagMan применяли для подсчета количества копий E1B и PIX трансгена в MCX и DXS. Так как трансген E1B поддерживается независимо от PIX, отношение двух генов является показателем поддержания трансгена PIX. Отношение не меняется между MCX и DXS. Более того, отношение мРНК PIX к мРНК E1B также не изменялось, и мРНК PIX была в избытке по отношению к мРНК E1B, что указывает на экспрессию PIX. Правая панель: MCX и родительские (PIX-негативные) CR. Клетки HS культивировали с или без давления селекции гигромицином в течение 2 недель (показано в виде черной полосы на оси X). В различные временные точки геномную ДНК выделяли (показано как “A”, “B” и “C”) и подвергали количественному определению с TaqMan (нижняя левая). В то время как родительские клетки уничтожались гигромицином, клетки MCX выживали. Отношение трансгенов оставалось постоянным, снова указывая на стабильное поддержание трансгена PIX также в присутствии давления селективного отбора. Время удвоения для родительских клеток составляет приблизительно 32 час, но только 41 час для клеток MCX.
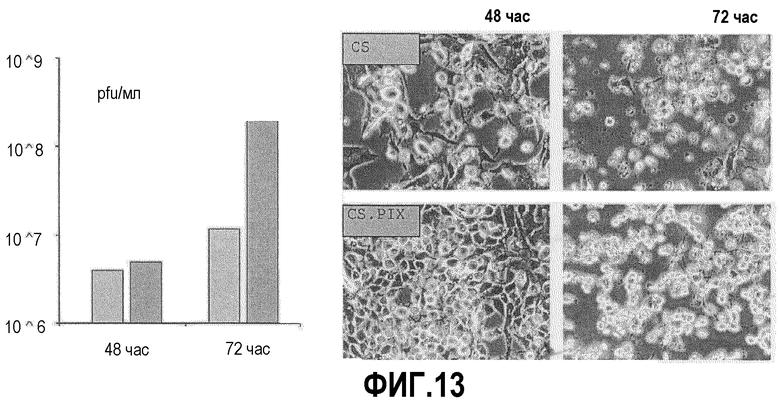
Фиг.13: Эффект PIX на репликацию MVA в клетках CS. Клетки CS и CSpIX инфицировали MVA и тестировали на репликацию MVA через 48 час и 72 час после инфицирования. Выход MVA был значительно выше в PIX-позитивных клетках CS. Также, что очевидно в фазово-контрастном микроскопе, цитопатический эффект появлялся с задержкой у клеток CSpIX на 48 час. Однако обе культуры являются восприимчивыми к вирусу в сходной степени, так как полный лизис очевиден через 72 час после инфицирования.
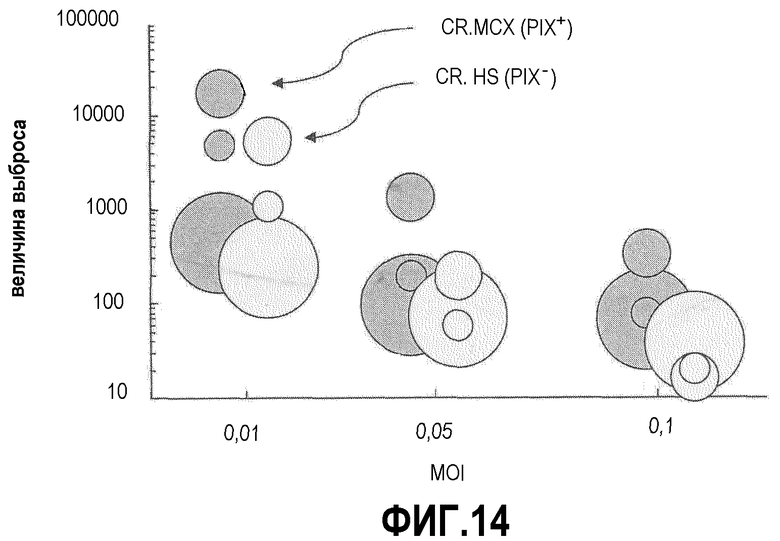
Фиг.14: Эффект PIX на репликацию MVA в клетках CR. Клетки CR и CRpIX в суспензии инфицировали различными сочетаниями и при различной плотности высевания клеток. Выход MVA оценивали через 48 час после инфицирования, и он показан на оси Y. Размер пузырьков отражает плотность высевания клеток. Закрашенные пузырьки показывают величины для клеток CRpIX, белые пузырьки показывают величины для родительских клеток. Во всех конфигурациях PIX придает отчетливое увеличение скоростей амплификации MVA.
Фиг.15: Субклеточное распределение PIX-GFP в клетках уток. Меченый GFP PIX появляется в нескольких цитоплазматических ярких пятнах и диффузном цитоплазматическом окрашивании, практически исключая ядра. Другие клоны проявляют более сильное диффузное цитоплазматическое окрашивание и более интенсивную аккумуляцию непосредственно вокруг ядра.
Фиг.16: Субклеточное распределение вариантов PIX-GFP в клетках CHO. Для исследования индуцируемых изменений распределения PIX в клетках были созданы гибридные варианты, включая вставку сайта ядерной локализации (NLS) и гибридизацию с ретиноидным рецептором альфа, клеточным белком, который уже содержит NLS. Представлены клетки CHO, временно трансфицированные экспрессионными плазмидами для вариантов PIX-GFP.
Фиг.17: Эффект индукции интерферона в присутствии PIX. Клетки CSpIX и CRpIX обрабатывали поли I:поли C, известным индуктором ответа интерферона типа I, и сравнивали с родительскими клетками. Клетки CSpIX реагировали с более высокой восприимчивостью к поли I:поли C, чем клетки CS. Клетки CRpIX и CR реагировали сравнимо и в меньшей степени, чем клетки CS.
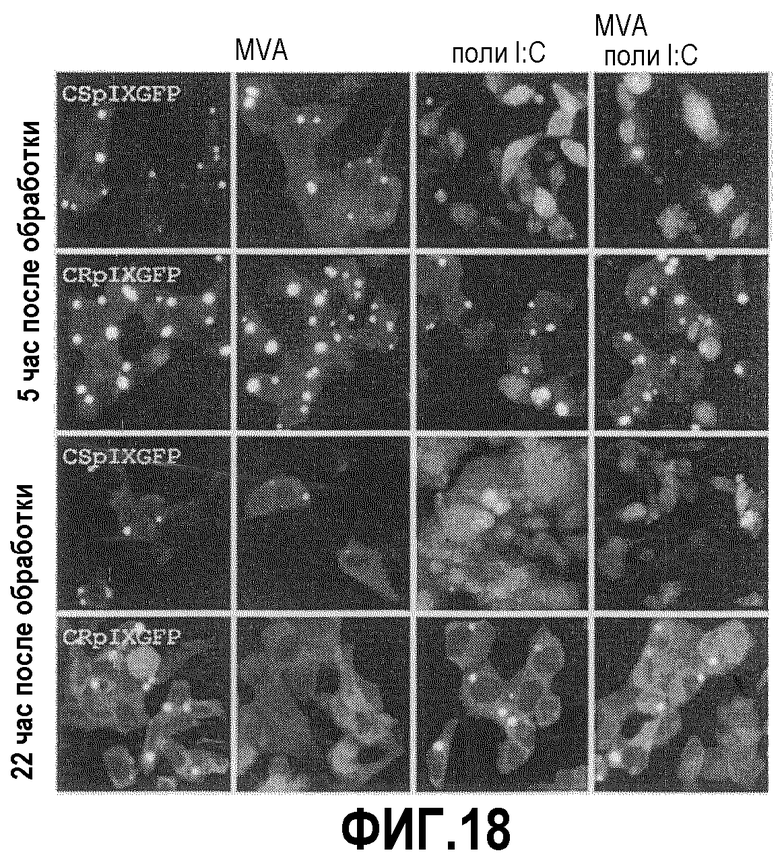
Фиг.18: Эффект инфицирования MVA и индукции интерферона в присутствии PIX-GFP. Клетки CSp9GFP и CRp9GFP обрабатывали поли I:поли C и инфицировали MVA при M.O.I. 0,1, как указано. Вновь происходящие от CS клетки реагировали на индукцию интерферона более сильно. Неожиданно оказалось, что количество ярких телец PIX, очевидно, снижается при индукции или инфицировании в обеих клеточных линиях, в то время как общая интенсивность сигнала PIX-GFP растет. Более того, эффект индукции поли I:поли C улучшается при параллельном инфицировании MVA, на что указывает большее количество клеток CS, все еще прикрепленных через 22 час после обработки.
Подробное описание изобретения
В способе согласно варианту осуществления (1) изобретения используют экспрессирующую клетку, обладающую интегрированным в ее геном геном, кодирующим гетерологичный регуляторный белок, называемый pIX или его функциональный вариант. Указанный регуляторный белок обладает следующими свойствами:
1. Он модулирует транскрипцию и, особенно, если связан с соответствующими модуляторами или регуляторами, также влияет на клеточный рост.
2. Он увеличивает продуктивность клеточной линии в отношении продукции вируса, не содержащего указанный регуляторный белок (т.е. регуляторный белок не замещает белок, удаленный из вируса), и/или в отношении продукции белка, отличающегося от указанного регуляторного белка или его функционального варианта.
В соответствии с изобретением гетерологичный регуляторный белок представляет собой белок pIX аденовируса серотипа 5 (например, имеющий последовательность а.к., представленную SEQ ID NO:2), его мутанты (включая мутанты с добавкой, заменой и/или делецией), его варианты (например, варианты, полученные из аденовируса другого серотипа) и тому подобное.
«Функциональный вариант гетерологичного регуляторного белка» в соответствии с изобретением включает гомологи из аденовируса серотипа, отличного от серотипа 5, все типы мутации (добавку, замену и/или делецию) конкретного(ых) аминокислотного(ых) остатка(ов) соответствующего регуляторного белка дикого типа, модификации путем слияния, которые дополнительно активируют белковую или пептидную последовательности и тому подобное. Особенно предпочтительными являются слитые белки, а именно слитые белки, включающие по меньшей мере один первый домен, включающий регуляторный белок, как определено здесь выше, и по меньшей мере один второй домен, включающий белок или пептид, действующий в качестве модулятора транскрипции. В предпочтительном варианте осуществления изобретения указанный модулятор транскрипции представляет собой транскрипционный фактор, включая рецептор альфа ретиноевой кислоты, который может присутствовать в полной или неполной (т.е. усеченной) форме. Модулятор может также представлять собой переносящий пептид, который включает последовательности NLF, например, как представлено в SEQ ID NO:21. В соответствии с изобретением первый и второй домен(ы) либо прямо, либо через пептидный линкер ковалентно соединены друг с другом. Подходящие пептидные линкеры включают гибкие и гидрофильные структуры, такие как поли gly-ser.
Термины «клетка» и «клеточная линия», используемые в последующем подробном описании, относятся к экспрессирующим клеткам/экспрессирующим клеточным линиям и к клеткам-хозяевам/клеточным линиям-хозяевам.
В соответствии с изобретением предпочтительно, чтобы гетерологичный регуляторный белок или его функциональный вариант экспрессировался в клетке в количестве по меньшей мере 1 пг/мкг клеточного белка, предпочтительно в количестве по меньшей мере 10 пг/мкг клеточного белка, так, чтобы его экспрессия могла быть определена с помощью вестерн-блоттинга.
В клетке по изобретению гетерологичный регуляторный белок или его функциональный вариант находится предпочтительно под контролем стабильного гомологичного или гетерологичного промотора. Подходящими промоторами являются конститутивный клеточный промотор или его варианты, такие как промотор фактора 2 элонгации при транслокации человека. Конкретный вариант промотора фактора 2 элонгации при транслокации человека, применяемый в изобретении, имеет последовательность, представленную в SEQ ID NO:12. Представленная последовательность представляет собой «короткий» вариант промотора, который локализован на хромосоме человека 19:3935325-3936638 (собрание по геному человека, май 2004) и дает стабильный средний уровень экспрессии. Для более сильной экспрессии может быть использован «более длинный» вариант промотора, локализованный на хром. 19: 3935349-3938957 (собрание по геному человека, май 2004).
В соответствии с изобретением клетка представляет собой клетку позвоночных, включая клетки млекопитающих, клетки птиц и тому подобное. Подходящими клетками млекопитающих являются клетки человека и клетки грызунов, включая мышь, крысу, хомячка и т.п. Особенно предпочтительными клетками млекопитающих в соответствии с изобретением являются клетки, происходящие из мозга человека, особенно из мозга плодов человека, клетка NSO или Sp2/0 мыши, клетка BHk или CHO. Подходящими клетками птиц являются клетки утки, птенцов перепелки и гуся. Особенно предпочтительными клетками птиц в соответствии с изобретением являются клетки, происходящие из сетчатки утки или клетка сомита.
С другой стороны, клетка по изобретению может происходить от первичной клетки или от ранее иммортализованной клетки. Более того, клетка может нести дополнительные иммортализующие (вирусные) гены, включая белок E1 аденовируса, такой как белок E1 мастаденовируса группы C типа 5 и тому подобное. Особенно предпочтительным в соответствии с настоящим изобретением является то, что клетка дополнительно несет ген аденовируса E1A и/или E1B, представленный в SEQ ID NO:5.
Клетка, используемая в способе согласно варианту осуществления (1) изобретения, может дополнительно нести функциональные последовательности, например, последовательности, требуемые для ее применения в качестве экспрессирующей клетки, такие как последовательности маркера селекции, последовательности сайтов сплайсинга донора/акцептора и/или последовательности, узнаваемые рекомбиназой, обеспечивающие интеграцию последовательности нуклеиновой кислоты-мишени, подлежащей экспрессии в клетке, и т.д.
«Инфицирование», «трансфекция» и «трансформирование», выполненные способами осуществлений (1), (10) и (12) изобретения, могут осуществляться в соответствии со стандартными процедурами, известными специалисту в данной области техники. Указанные способы могут дополнительно охватывать подходящие стадии селекции, выделения и экспансии, если это требуется.
В способе согласно варианту осуществления (1) вирус-мишень включает формы дикого типа, вирус с мутацией или делецией, адаптированный к холоду, или ослабленный вирус, штаммы вакцин, вирусные векторы, несущие гетерологичный(ые) ген(ы), вирусные векторы, такие как лентивирус, поксвирус, вирус, ассоциированный с аденовирусом (aav), вирус герпеса, флавивирус и тому подобное.
Далее в способе согласно варианту осуществления (1) один или более белков-мишеней включают антитела, рекомбинантные белки, такие как эритропоэтин, альфа-1-антитрипсин, факторы свертывания VIII и IX и интерфероны, вирусные антигены, такие как HA и NA гриппа и M, HBV-S, белок G герпеса и белок G бешенства, пептидные гормоны и тому подобное. Хотя способ позволяет продуцировать клетки, способные к одновременной экспрессии более одного белка-мишени, особенно предпочтительно, чтобы они кодировали только один белок-мишень.
Культивирование и выделение в способе согласно варианту осуществления (1) изобретения может быть осуществлено в соответствии со стандартными процедурами, легко доступными специалисту в данной области техники. Указанный способ может дополнительно включать стандартные стадии очистки, а также последующие стадии модификации вируса-мишени или белка(ов)-мишени(ей).
Что касается предпочтительных вариантов экспрессии клеточных линий-хозяев согласно вариантам осуществления (9) и (11) соответственно, а также способов продукции для указанных клеточных линий согласно вариантам осуществления (10) и (12), дается ссылка на подробное обсуждение, предложенное здесь выше в связи с вариантом осуществления (1).
В соответствии с вариантами осуществления (14) и (15) в изобретении предлагается слитый белок, включающий по меньшей мере один первый домен, включающий регуляторный белок, и по меньшей мере один второй домен, включающий белок или пептид, действующий в качестве модулятора транскрипции, как определено здесь выше, и нуклеотидная последовательность, кодирующая указанный слитый белок, соответственно. Изобретение также относится к применению указанного слитого белка (например, к диагностическому и фармацевтическому применению и т.д.) и нуклеотидной последовательности, кодирующей его в осуществлениях (14) и (15) соответственно, например, во всех типах векторных конструктов, клеточных линиях, тканевой культуре, трансгенных животных и т.д.
Клеточная линия NC5T11#34 была депонирована DSMZ-Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH, Mascheroder Weg Ib, 38124 Braunschweig, Germany, 4 ноября 2005 г. под номером поступления DSM ACC2744. Клеточная линия CR.PIX (17a11b) была депонирована DSMZ-Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH, Mascheroder Weg Ib, 38124 Braunschweig, Germany, 24 ноября 2005 г. под номером поступления DSM ACC2749.
Изобретение будет объяснено более подробно с помощью последующих примеров, которые, однако, не истолковываются как ограничивающие изобретение.
Примеры
Пример 1
Формирование клеточной линии ТС5Е11
Клеточную линию формировали из смеси клеток эмбрионального мозга путем иммортализации генами E1A и B аденовируса 5 с помощью невирусной трансфекции.
Образец ткани отбирали из перивентрикулярной зоны мозга плода после индуцированного аборта. Его вырезали ножницами и гомогенизировали отсасыванием пипеткой для культуры ткани в нейрональной культуральной среде на основе DEMEM/F12, содержащей 20 нг/мл ФРФч (Invitrogen, Carlsbad, CA 92008, США), 20 нг/мл ЭФРч (Invitrogen), добавку 1x N2 (Invitrogen) и 8 мкг/мл гепарина (Sigma Aldrich, St. Louis, США). Клетки осаждали при 200 г в течение 3 мин. Жизнеспособность клеток оценивали с помощью трипанового синего и йодида пропидия с использованием проточного цитометра (BD Biosciences, Jose, CA 95131, США). Жизнеспособность составляла 75%. 0,5×106 клеток высевали во флаконы T25. Клетки инкубировали при 37°С и 5% CO2. К 5 дню культивирования клетки образовывали нейросферы. Нейросферы показаны на фиг.1A. На 8 день клетки переносили на DMEM/F12 с добавкой 5% FCS для обеспечения прикрепления и стимуляции пролиферации. Клетки образовывали однородный монослой, как показано на фиг.1B, и их пассировали 1:5 раз в неделю. Эта первичная нейрональная клеточная культура была названа NC5.
После 2 недель в среде с сывороткой клетки трансфицировали вектором p79 в субконфлюентных 6-луночных планшетах с использованием Effectene (Quiagen, 40724 Hilden, Германия) в качестве реагента трансфекции в соответствии с инструкциями производителя. Плазмида p79 содержит следующие элементы: в качестве основы плазмиды служит pBluescript (Stratagene, США), в которой маркер устойчивости к ампициллину был заменен геном устойчивости к канамицину под контролем бактериального промотора, который обеспечивает рост и селекцию в E.coli. Вектор несет фрагмент аденовируса типа 5 дикого типа (SEQ ID NO:5), содержащий открытые рамки считывания для E1A (сплайсинговые варианты 13S и 12S, включающие или не включающие домен CR3 соответственно) и E1B 55k и 19k, а также последовательности выше E1A. Гену E1A предшествует промотор фосфоглицераткиназы (мыши). После последовательностей аденовируса расположен сигнал полиаденилирования из гена тимидинкиназы Herpes simplex, служащий в качестве замены сигнала полиаденилирования E1B. Элементы были получены из соответствующих организмов или из донорных плазмид с помощью ПЦР и клонирования с использованием традиционных способов рекомбинантной ДНК и подтверждены секвенированием. Через два дня после трансфекции клетки трипсинизировали и переносили на 10-см чашку. Спустя две недели образовывались очаги небольших клеток с высоким соотношением ядро/цитоплазма и четко различимыми границами (фиг.1C) при использовании трансфицированных, но не ложнообработанных клеток. Из 11-й трансфекции было выделено восемь независимых очагов с использованием трипсина и цилиндров для клонирования (Corning, США), которые высевали в лунки 24-луночного планшета и наращивали в 12-луночном планшете, 6-луночном планшете и флаконах T25. Все выделенные очаги содержали два типа клеток: мелкие клетки с четкими границами и крупные фибробластоподобные клетки.
Через три недели после трансфекции стали видимыми 15 дополнительных клонов при трансфекции T11, некоторые из которых могли возникнуть из оставшихся клеток уже выделенных первичных клонов.
Клоны T11a.1 и T11a.6 проявили наиболее быстрый рост, и они были сохранены замораживанием в DMEM/F12, 10% ДМСО, 25% сыворотке через 8 недель после трансфекции. В это время все клоны все еще содержали фракцию клеток с увеличенной цитоплазмой, напоминающих исходный фенотип. Однако при отношении разделения 1:5 для T11a.1 и T11a.6 они переросли и элиминировались приблизительно через 3 месяца после трансфекции. Были проведены эксперименты с иммунофлуоресценцией для выявления связи между измененной морфологией и экспрессией E1A и E1B. После фиксации метанолом клетки обрабатывали антителами крысы против E1A и E1B 55k соответственно и затем техасским красным, конъюгированным с антикрысиными антителами. Как показано на фиг.2, все клетки образца проявляют ядерное окрашивание, типичное для E1A, и цитоплазматическое окрашивание для E1B.
Через три месяца после трансфекции клоны T11a.1 и T11a.6 были перенесены в бессывороточную среду ProPER (Cambrex, Бельгия) высеванием 1,6×106 клеток после обработки трипсином. Клетки собирали центрифугированием и среду меняли центрифугированием раз в неделю. Клеточная популяция выжила, но даже через шесть месяцев после трансфекции клетки все еще проявляли низкую жизнеспособность и росли с большим временем удвоения, составляющим 60-80 час.
Когда клетки были вновь перенесены в DMEM/F12 с 5% FCS после 3 месяцев в среде ProPER, оба клона дали гомогенные и высокожизнеспособные культуры с временем удвоения 40 час. Для дальнейших экспериментов был выбран T11a.1 как наиболее здоровый клеточный клон и был назван NC5T11 (фиг.1D).
Пример 2
Формирование клона #8 NC5T11puro
Продукция белка линией клеток зависит от эффективности способов трансфекции и селекции. Пригодность обычно применяемых маркеров селекции G418, пуромицина, гигромицина, бластицидина, MTX, гистидинола для NC5T11 была неизвестна. На первой стадии оценки тестировали различные имеющиеся в продаже реагенты трансфекции Lipofectamine (Invitrogene), Fugene (Roche, Германия), Polyfect (Qiagen) и Effectene в соответствии с протоколами производителей. Для определения эффективности временной трансфекции использовали плазмиду pEGFP-N1 (Clonetech, США), экспрессирующую gfp из промотора CMV человека. Наибольшая эффективность была получена с Effectene (Qiagen), достигающая 20-50%, в зависимости от плотности клеток при использовании прикрепленных клеток, растущих в DMEM/F12 10% FCS.
Для тестирования генов маркеров селекции были сконструированы экспрессионные векторы, содержащие ген альфа-1-антитрипсина (aat) человека под контролем промотора CMV человека с последующим сигналом PolyA гормона роста быка и соответствующий маркер селекции, контролируемый слабым промотором. Конкретнее, для тестирования селекции пуромицином использовали вектор C55, который содержит ген устойчивости к пуромицину, управляемый промотором PGK человека, сопровождаемый ранним сигналом полиаденилирования SV40.
После трансфекции клетки высевали в DMEM/F12 5% FCS, содержащую 0,75 мкг/мл пуромицина, и подвергали селекции в течение 3 недель при еженедельной смене среды. Был образован пул клона, и экспрессия была подтверждена с помощью иммунофлуоресценции с использованием поликлонального антитела козы против альфа-1-антитрипсина (Innogenetics, США) и затем меченного биотином вторичного антитела кролика против козы и конъюгата стрептавидин-техасский красный.
Для выделения индивидуальных клеточных клонов 10000 клеток из пула клона высевали в 15-см чашку. Через 4 недели клоны выделяли с помощью цилиндров для клонов (Corning) и клоны #1, 2, 7, 8, 9, 10 анализировали на предмет экспрессии aat. Конкретнее, клетки высевали в количестве 7×104 клеток в 12-луночные планшеты и определяли количество клеток и экспрессию aat. Вычисленные показатели удельной продуктивности клеток показаны на фиг.3. Клон NC5T11puro#8 показал удельную продуктивность более 100 пг/клетку·день и был выбран для дальнейшей работы.
Пример 3
Изменение NC5T11 и NC5T11puro#8 для среды продуцирования и анализ экспрессии во встряхиваемой порционной культуре
Скорость роста и NC5T11, и NC5T11puro#8 в среде ProPER (Cambrex) была приблизительно в два раза ниже, чем в содержащей сыворотку среде. Поэтому в качестве альтернативы была апробирована Xcell VPRO (JRH Biosciences). Адаптацию проводили прямым высеванием 4,5×106 клеток в 5 мл EXCELL VPRO во флаконе T25. Через 2 недели клеточный рост инициировали повторно, и установилось недельное отношение разделения 1:3. В качестве следующей стадии оба клона адаптировали к росту при наличии напряжения сдвига. Клетки высевали в 12 мл EXCELL VPRO при 6×104 клеток/мл в 50-мл полипропиленовых встряхиваемых пробирках (TPP, Швейцария) и подвергали вращению с радиусом 1 см и скоростью 160 об/мин. После повторных пассажей проводили анализ партий для определения максимальной плотности клеток и аккумуляции продукта во время экспоненциальной и стационарной фаз роста. Были сделаны следующие наблюдения: клон NC5T11puro проявил высокую жизнеспособность, которая оставалась выше 70% до 18 дня. Максимальная плотность клеток не превышала 1×106. Накопление продукта продолжалось во время стационарной фазы (фиг.5). Было сделано заключение, что для достижения высокой плотности клеток, сравнимой с получаемой с клетками CHO, необходимо создание специфических сред или дальнейшее улучшение клеток. Это, вероятно, должно вести к существенному увеличению титров продукта.
Пример 4
Конструирование слитого белка pIX-RARA и интегрирующих векторов и для pIX-RARA, и для pIX
Последовательности для pIX аденовируса и рецептора альфа-ретиноевой кислоты (RARA) были получены с помощью ПЦР из аденовируса типа 5 и геномной ДНК человека соответственно с использованием праймеров
1. AACCAGCGCTACCATGAGCACCAACTCGT (SEQ ID NO:6)
2. AATGGTGGCAACCGCATTGGGAGGGGAGG (SEQ ID NO:7) для PIX и
3. CCAATGCGGTTGCCACCATTGAGACCCAGA (SEQ ID NO:8)
4. AAGGAGCGCTGGCGAGGGCTGTGTCCAT (SEQ ID NO:9) для RARA.
Два фрагмента амплифицировали раздельно. Перекрытие между праймерами 2 и 3 обеспечивало соединение двух фрагментов посредством ПЦР с использованием праймеров 3 и 4. Гибридный ген клонировали в вектор pEFpromhyg, что дало pEFpIXRARA. Этот вектор содержит промотор фактора элонгации 2 человека, расположенный на chrl9:3935325-3936638 (сборка генома человека май 2004; SEQ ID NO:12). Он содержит расположенные выше элементы и первый интрон гена EF2. Природный стартовый кодон EF2, расположенный в 5' направлении от первого интрона, был удален мутагенезом. В векторе pEFpromhyg последовательность EF2 сопровождается уникальным сайтом рестрикции (AfeI), используемым для вставки pIX-RARA. Внутренний сайт связывания с рибосомой после сайта AF1 обеспечивает экспрессию гена устойчивости к гигромицину в качестве второй открытой рамки считывания в объединенной с pIX-RARA РНК. Такая конфигурация поддерживает экспрессию pIX-RARA во всех устойчивых к гигромицину клетках.
Ген pIX дикого типа амплифицировали с помощью праймеров AACCAGCGCTACCATGAGCACCAACTCGT (SEQ ID NO:10) и ACCGAGCGCTTGTTTTAAACCGCATTGG (SEQ ID NO:11) и интегрировали в сайт Afe pEFpromhyg вместо pIX-RARA с получением pEFpIX.
Пример 5
Модификация клеточных линий NC5T11 и NC5Tllpuro#8 геном pIX и геном слитого белка pIX-рецептор ретиноевой кислоты
Были получены клеточные клоны NC5T11 и NC5T11puro#8 для трансфекции в 6-луночных планшетах высеванием 1,3×106 клеток/лунку в DMEM/F12, 5% FCS. Плазмиду F67 (pEFpIXRARA) и плазмиду F76 (pEFpIX) выравнивали ферментом рестрикции Asp700 (Roche) и трансфицировали после осаждения этанолом с помощью Effectene (Qiagen): ДНК смешивали с 16 мкл усилителя и 200 мкл буфера EC. Через 2 мин при комнатной температуре добавляли 18 мкл Effectene и давали образоваться липосоме в течение 10 мин при комнатной температуре. Смесь трансфекции добавляли к 1 мл культуры клеток, высеянных до 80% конфлюентности в 6-луночные планшеты накануне. Высокая эффективность трансфекции была подтверждена параллельной трансфекцией pEGFP-Nl (Clontech, Palo Alto, CA 94303-4230, США). Через день после трансфекции среду заменяли DMEM/F12 с 10% FCS и добавляли 75 или 100 мкг/мл (NC5T11) и 50 мкг/мл (NC5Tllpuro#8) гигромицина. Селекционную среду меняли дважды в неделю. Через 18 дней в каждой лунке становились видимыми 15-20 четких клонов. Пулы клонов обрабатывали трипсином и селекцию продолжали еще в течение двух недель. Пулы клонов тестировали на экспрессию PIXRARA с помощью ОТ-ПЦР. В большинстве пулов клонов был выявлен сильный сигнал с праймерами 1 и 2 и более слабый сигнал с праймерами 1 и 4. Для выделения индивидуальных клеточных клонов 10000 и 1000 клеток пулов клонов высевали на 15-см чашки. Через 4 недели клоны выделяли с помощью цилиндров для клонирования (Corning). Для клонов, возникших в результате трансфекции F76, использовали другую стратегию. Пулы клонов обрабатывали трипсином, переносили в 50-мл встряхиваемую пробирку в EXCELL VPRO, содержащую 75 мкг/мг гигромицина, и через 2 недели 2000 жизнеспособных клеток суспензионной культуры смешивали с 12 мл матрикса клонирования (Genetix, UK), высевали в шестилуночный планшет (Greiner) (2 мл/лунку) и подвергали автоматизированному отбору колоний (clonepixFL, Genetics, UK). Клонам, происходящим от NC5Tllpuro#8, были присвоены номера #10-21. Клонам, происходящим от NC5Tll, были присвоены номера #34-45. Клоны, трансфицированные pEF2PIX, были названы NC5T11PIXA-NC5T11PIXE. Клоны тестировали на наличие соответствующего вектора с помощью ПЦР. Первоначальный тест с использованием праймеров 1 и 2 представлен на фиг.6. Все клоны, давшие положительный сигнал, были перенесены для выращивания в ExCell VPRO. Повторную адаптацию проводили прямым высеванием 4,5×106 клеток в 5 мл EXCELL VPRO во флаконе T25. Через 6 недель стационарную культуру клеток переносили в 50-мл полипропиленовые встряхиваемые пробирки (TPP, Швейцария) при плотности высевания 6×104 клеток/мл в EXCELL VPRO при общем объеме 12 мл и подвергали вращению с радиусом 1 см и при скорости 160 об/мин. По наибольшей жизнеспособности в условиях напряжения сдвига были отобраны клоны #10, #12, #14 (NC5Tllpuro#8pIXRARA) и #34, #43 (NC5T11pIXRARA) и NC5T11PIXB, NC5T11PIXC. Клоны #34, 43 и NC5T11PIXC повторно клонировали и анализировали на стабильность интегрированной ДНК. Стабильность оценивали определением соотношения генов PIX и E1B, поскольку E1A+E1B и pIXRARA вводили при раздельных трансфекциях, и гены E1 вводили и сохраняли в отсутствие селекции в течение >2 лет и поэтому рассматриваются как стабильно интегрированные. Геномную ДНК выделяли из отобранных клеточных клонов в два момента времени: сразу после прекращения селекции hyg (ранний момент); через 2 месяца в отсутствие давления отбора (поздний момент). Уровни ДНК PIX и E1B определяли с помощью ПЦР в реальном времени (ABI 7000, CYBR Green) при 2 концентрациях ДНК. Примеры этой оценки показаны на фиг.7. Вектор pEF2PIXRARA стабильно сохраняется в субклонах #34 и #43 при приблизительно 0,4-0,6 копиях PIXRARA на E1B. Клон NC5T11PIXC исходно содержит 4 копии pIX на E1B. Это число снижается в два раза в ходе эксперимента.
Пример 6
Зависимый от ретиноевой кислоты рост NC5T11pIX-RARA
Ретиноевая кислота (RA) является индуктором клеточной дифференцировки. Дифференцировка обычно ассоциирована со сниженным ростом клеток. Ответ на RA зависит от экспрессии ретиноевого рецептора альфа, транскрипционного фактора, активируемого RA. RA оказывает сильное влияние на PML (промиелоцитарный лейкоз), обращая трансформированный фенотип путем остановки роста.
Как показано на фиг.8, обработка RA в концентрации 6 мкг/мл не оказывала никакого влияния на рост ни NC5T11, ни экспрессирующего альфа-1-антитрипсин клона NC5Tllpuro#8. Однако рост клонов #14 (NC5Tllpuro#8pIXRARA) и #34 (NC5T11pIXRARA) прекращался обработкой 6 мкг/мл в течение 5 дней в прикрепленной культуре в DMEM/F12 с 5% FCS.
Пример 7
pIXRARA предотвращает ингибирование репликации вируса интерфероном
Клетки NC5 T11 и NC5T11#34 высевали в 6-луночные планшеты в количестве 8,0×105 клеток/лунку в DMEM/F12 5% FCS. Через 12 час в лунки добавляли интерферон бета (rebif 44 of Serono) в концентрации 1 и 8 МЕ/мл. Через 30 мин обработанные интерфероном клетки и контрольные клетки NC5 T11 и NC5T11#34 инфицировали вирусом энцефаломиокардита (EMCV) с кратностью инфекции (MOI) 0,004. Через 24 час культуры собирали и замораживали при -80ºС для разрушения клеток. Эту суспензию оттаивали и осветляли центрифугированием при 800×g в течение 10 мин. Лизаты подвергали титрованию на вирус анализом образования бляшек на клетках A549: Вкратце, клетки A549 высевали в 24-луночные планшеты и выращивали до достижения конфлюентности, затем инкубировали с осветленными лизатами, разбавленными от 10 до 2×108-кратно, покрывали 0,2% слабо желатинизирующейся агарозой типа VII в RPMI 10% FCS и инкубировали при 37°С в течение 24 час. Агарозный слой отсасывали, клетки фиксировали 2% глутаральдегидом в PBS в течение 20 мин при 20°С, промывали водой и инкубировали в течение 30 мин при комнатной температуре с 1% раствором Kristallviolett в 50% этаноле для подсчета вирусных бляшек. Результаты, показанные на фиг.9, демонстрируют, что репликация EMCV весьма неэффективна в клетках NC5T11 и что она очень чувствительна к обработке интерфероном перед инфекцией. При 8 МЕ/мл не удается извлечь жизнеспособный вирус из культуры. Напротив, репликация вируса в NC5T11#34 лишь слабо зависит от обработки интерфероном. Более того, даже в присутствии экзогенного ИФН для NC5T11#34 наблюдаются более высокие титры. Показанные феномены моделируют типичные процессы вакцинирования, поскольку многие штаммы вакцин индуцируют ответ ИФН в продуцирующих его клетках с интактным путем ИФН, и инфекция при низкой кратности инфекции (MOI) предпочтительно ведет к процессам продукции, которые вызывают секреции ИФН, что ведет к блокированию репликации вируса в клетках, которые не были исходно инфицированы. Следовательно, экспрессия PIX может обеспечивать высокий титр продукции вируса при исходной низкой MOI.
Пример 8
Стимуляция pIX-RARA продукции белка
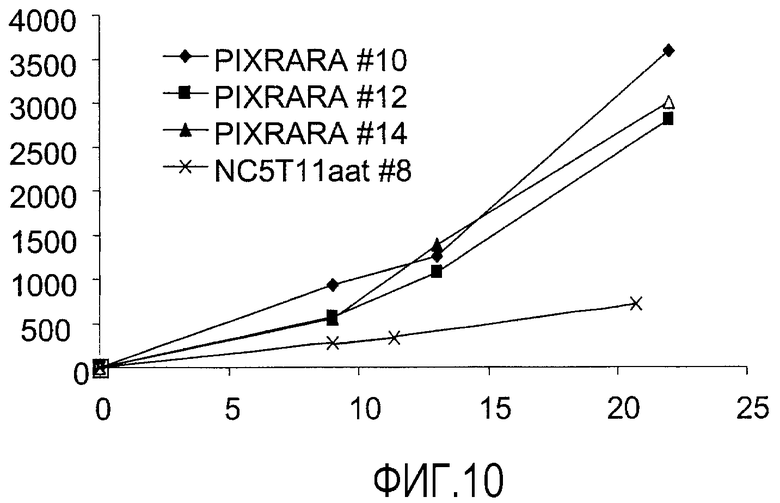
NC5T11puro#8pIXRARA, содержащие вектор F67, а также альфа-1-антитрипсин, сравнивали с исходным клоном NC5T11puro#8 по способности продуцировать альфа-1-антитрипсин в тесте порционной встряхиваемой культуры. Клетки высевали в EXCELL VPRO (JRH Biosciences) при 6×104 клеток/мл в 50-мл полипропиленовых встряхиваемых пробирках (TPP, Швейцария) при суммарном объеме 12 мл и подвергали вращению с радиусом 1 см и скоростью 160 об/мин. Культивирование продолжали до 22 дня. Кроме суспендированных клеток образовывалось кольцо кластеров клеток, прикрепленных к стенке пробирки. Клетки этих кластеров сохраняли жизнеспособность более 70% на протяжении процесса. Поскольку диссоциация этих кластеров под действием трипсина или акутазы (PAA) не происходила, кинетику роста и максимальную плотность клеток не определяли. Образцы отбирали на 9, 13 и 22 дни и определяли титры с помощью ELISA альфа-1-антитрипсина. Результаты показаны на фиг.10.
Пример 9
Получение клеток сетчатки утки, экспрессирующих pIX
Клеточная линия CR, берущая начало от первичных клеток сетчатки утки и иммортализованная в соответствии с правилами определенного риска, описана в другом месте (патентная заявка WO 05042728, трансфицированные плазмидой 60E клетки сетчатки). Клетки культивировали в среде DMEM:F12 (Invitrogen, Carlsbad, CA 92008, США) с добавкой 5% сыворотки плодов телят (Biochrom AG, 12213 Berlin, Германия) при 39°С и 7,5% CO2. Для пассирования клетки в течение короткого периода времени обрабатывали TrypLE Express (Invitrogen). Плазмиду 76F pEFPIXl выравнивали ферментами рестрикции Ssp I и Xmn I (оба от New England Biolabs, Beverly, MA 01915-5599, США) и очищали до 400 нг/мкл аффинной хроматографией (набор для экстракции геля от Qiagen, 40724 Hilden, Германия). 5 мкл (2 мкг) очищенной ДНК трансфицировали в клетки CR с помощью реагентов Effectene (Qiagen): ДНК смешивали с 16 мкл усилителя и 200 мкл буфера EC. Через 2 мин при комнатной температуре добавляли 18 мкл Effectene и давали образоваться липосоме в течение 10 мин при комнатной температуре. Смесь трансфекции добавляли к 1 мл культуры клеток, высеянных до 80% конфлюентности в 6-луночные планшеты накануне. Высокая эффективность трансфекции была подтверждена параллельной трансфекцией pEGFP-Nl (Clontech, Palo Alto, CA 94303-4230, США).
Трансфицированные pIX культуры наращивали во флаконах T75 через три дня после трансфекции и селекцию инициировали 25 мкг/мл гигромицина B (Invitrogen). Среду меняли раз в неделю с увеличением гигромицина B до 50 мкг/мл. Через три недели выжило в общей сложности четыре больших узла, которые были повторно высеяны во флакон T25: клетки отделяли TrypLE и собирали при 100×g в течение 10 мин, ресуспендировали в свежей среде и помещали во флакон T25.
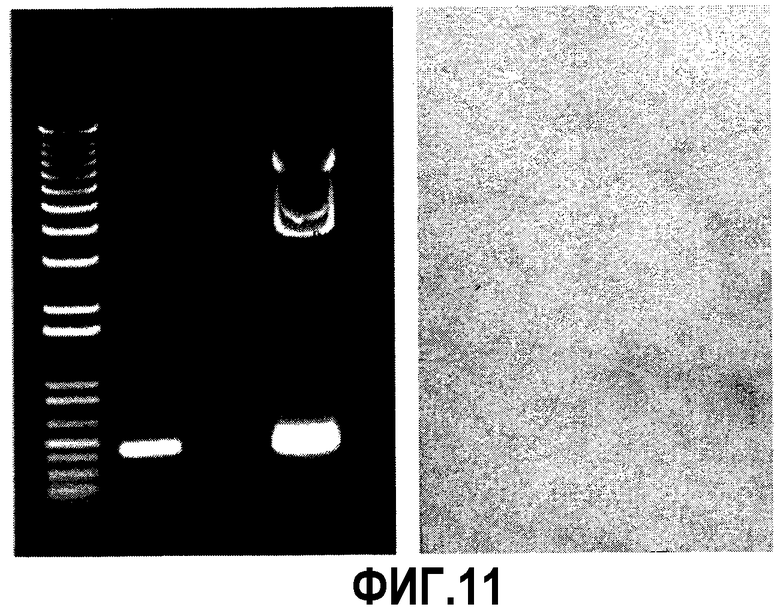
Через четыре недели после трансфекции клетки из здоровой субконфлюентной культуры во флаконе T25 отделяли TrypLE в 5 мл DMEM:F12, 5% FCS. 2 мл полученного смешивали со средой #63032-1000M (JRH Biosciences, KS 66215, США), средой, не содержащей компонентов животного происхождения, предназначенной для поддержания суспензионных культур. Добавляли гигромицин B до 50 мкг/мл. Выделяли геномную ДНК из 2 мл культуры и проводили ПЦР для трансгена pIX с праймерами V293 и V294. Отрицательный контроль обеспечивался параллельной реакцией без ДНК, а положительный контроль - параллельной реакцией с плазмидой 76F. Ожидаемый продукт ПЦР на фиг.11 (левая часть) демонстрирует стабильную вставку трансгена pIX в клетки CR, называемые теперь “CRpIX”.
Экспрессия белка PIX была подтверждена иммуноблоттингом: 6×106 клеток были разрушены кипячением в (20 мМ трис pH 7,4, 300 мМ NaCl, 1% Na-дезоксихолат, 1% тритон® X-100, 0,1% ДДС-Na), и белок разделяли гель-электрофорезом и затем переносили на нейлоновую мембрану. PIX выявляли первичным антителом против pIX (любезно предоставленным д-ром W. Seidel, Ernst-Moritz-Arndt-Universitat Greifswald, Германия) и затем вводили во взаимодействие со вторичным антителом, направленным против первого антитела и меченным щелочной фосфатазой.
Сигнал ожидаемого размера на фиг.11 (правая часть) подтверждает экспрессию pIX в клетках CRpIX. Сигнал отсутствовал в отрицательном контроле из клеток 293, полученном в параллельной реакции.
Клетки культивировали в среде #63032-1000M для двух дополнительных пассажей, затем переносили в среду #14561-1000M (также от JRH Biosciences) для двух дополнительных пассажей. Во все среды JRH добавляли 1 × Glutamax I (Invitrogen) и 100 мкг/мл гигромицин B.
При втором пассаже в среде #14561-1000M небольшую часть (приблизительно 2% из 5 мл суспензионной культуры) тщательно ресуспендировали в среде DMEM:F12, FCS и помещали в чашки Петри диаметром 15 см. Через 6 дней одиннадцать очагов переносили в отдельные лунки 12-луночного планшета и поддерживали в среде DMEM:F12, FCS. Для перенесения клонов среду отсасывали и диски клонирования (Sigma, MO, USA), пропитанные TrypLE, на короткое время прямо наносили на клоны, после чего переносили в лунки со средой. Отдельные клоны размножали для последующих экспериментов и определения ростовых свойств.
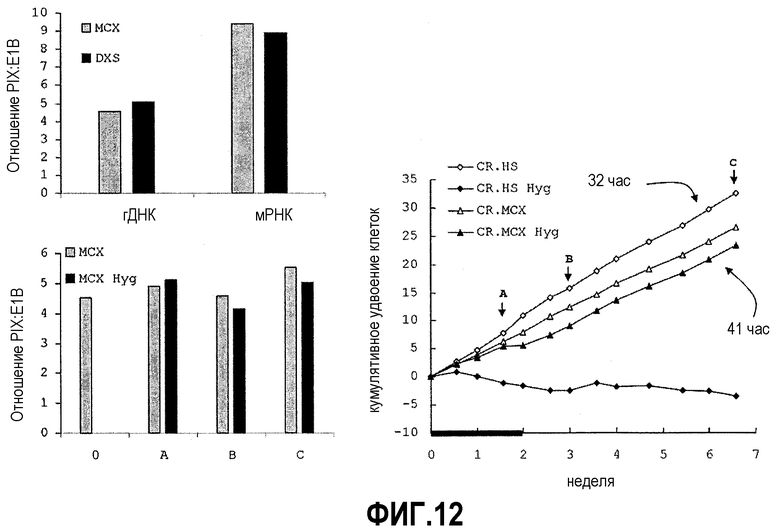
Для анализа сохранения PIX в стабильно трансфицированных клетках геномную ДНК выделяли из 1×106 клеток с помощью набора для ДНК крови QIAamp (Qiagen) и количество молекул E1B и PIX определяли в 25 нг и 50 нг геномной ДНК в реакции ABI 7000 TaqMan с использованием химической детекции SYBR Green (ABI). Для количественного определения использовали праймеры gTggTTgCTTCATgCTAgTg (SEQ ID NO:13) и TCTTCAgCAggTgACAgTTg (SEQ ID NO:14) для E1B, ACCTACgAgACCgTgTCTg (SEQ ID NO:15) и gAgCCgTCAACTTgTCATC (SEQ ID NO:16) для PIX. Поскольку E1B вводили независимо и он должен был сохраняться для поддержания жизнеспособности клеток, этот ген служит внутренним маркером для стандартизации количества копий PIX и эффективности экспрессии гена.
На фиг.12 показано стабильное сохранение трансгена PIX в суспензионных клетках CR даже в отсутствие давления отбора. На нем также показано, что PIX-позитивные клетки страдают сниженной скоростью роста. Заявители постоянно наблюдали пониженную пролиферацию всех их клеток (птичьих и человеческих клеток) после стабильной трансфекции экспрессирующей PIX плазмидой, что демонстрирует влияние PIX на биохимию клетки-хозяина. Более того, заявители наблюдали тенденцию PIX-позитивных, прикрепленных клеток CR к уклонению от роста до конфлюентных слоев (повышенное контактное ингибирование).
Пример 10
Создание сомитных клеток утки, экспрессирующих pIX
Клеточная линия CS берет начало от эмбриональных сомитов, также описанных в патентной заявке WO 05042728. Создание PIX-позитивных клеток CS осуществляли параллельно с описанной выше процедурой для линии CR. В отличие от CRpIX суспензионные культуры для CSpIX не были сформированы. Особенностью клеток CS является полная зависимость от прикрепления. Клетка, которая не пролиферирует в суспензии, может рассматриваться как менее злокачественная, поскольку ее потенциал к метастазированию серьезно подорван. Белок PIX не влияет на данное свойство клеток CS. Поэтому, несмотря на плейотропность, данный белок, по-видимому, не оказывает влияния на фенотип трансформации. Это наблюдение поддерживает предположение заявителей о том, что PIX можно безопасно применять в биофармацевтических процессах.
Пример 11
Влияние PIX на репликацию MVA в клетках CS
Клетки CS, стабильно трансфицированные для экспрессии PIX, и исходные клетки CS в качестве репера высевали в 6-луночный планшет в количестве 2×106 клеток/лунку и на следующий день инфицировали MVA при m.o.i. 0,1. Через 48 и 72 час после инфекции были получены снимки для документирования различий до CPE. Выход вируса в супернатанте через 48 час после инфекции и полный выход (супернатант вместе с лизированным осадком клеток) через 72 час после инфекции определяли с помощью микрофокусного анализа. Результаты показаны на фиг.13: наличие PIX в клетках CS, очевидно, задерживает наступление CPE через 48 час после инфекции по сравнению с исходным репером. Через 72 час после инфекции обе культуры полностью лизированы, что позволяет предполагать, что различия в степени CPE не обусловлены наличием смеси устойчивых к MVA и чувствительных к MVA клеток в популяции CS.PIX.
Насколько известно заявителям, PIX-позитивные клетки не тестировали когда-либо вирусами, отличными от родственного аденовируса.
Пример 12
Влияние PIX на репликацию MVA в клетках CR
Положительное действие белка PIX измеряли количественно также в клетках CR. В отличие от клеток CS клетки CR были адаптированы для суспензии. Суспензионные культуры являются предпочтительными для многих промышленных применений. Поэтому заявители определили влияние PIX в такой системе, как противоположной прикрепленной культуре.
Для клеток CR поддерживающий эффект PIX не является столь же выраженным, как для клеток CS, но заявители постоянно наблюдали более высокие титры MVA. Выход MVA оптимизировали в серии экспериментов, и было обнаружено, что он является наибольшим через 48 час после инфекции при средней плотности клеток. На фиг.14 показано сравнение суспензионного клона CR.HS (отобранного из нескольких клонов в качестве клеточной линии с наибольшим выходом MVA) и PIX(+) линии клеток CR.MCX. На оси абсцисс показана m.o.i. (0,01, 0,05 или 0,1 соответственно), на оси ординат показаны выброс и размер пузырьков, указывающих на плотность клеток (8, 2 или 0,8×106 клеток/мл соответственно). Выброс представляет собой отношение выхода вируса (или выход) к введенному вирусу (прививка зависит от количества клеток и m.o.i.) и таким образом служит мерой эффективности амплификации. При всех сочетаниях линии клеток PIX(+) превосходили линию PIX(-) по производительности.
Пример 13
Исключение PIX из ядра
Гибридный ген PIX-GFP был создан для визуализации распределения белка PIX в живых клетках и преодоления слабой связывающей активности имеющегося антитела: плазмиду 76F pEFPIXl обрабатывали ферментами рестрикции Acc I и Dra I и концы делали тупыми с помощью полимеразы Кленова (все от New England Biolabs, Beverly, MA 01915-5599, США). Из сложной смеси полос желаемый фрагмент 447 п.н., содержащий кодирующую PIX последовательность, выделяли экстракцией агарозного геля (набор для экстракции геля от Qiagen, 40724 Hilden, Германия). Фермент рестрикции Dra I узнает последовательность "ttTAAa", разрезаемую после последнего остатка тимидина, где центральный триплет “TAA” является стоп-кодоном открытой рамки считывания PIX. Таким образом, фрагмент 447 п.н. лишен стоп-кодона. Слияние этого фрагмента с геном EGFP в плазмиде pEGFP-Nl, раскрытой Sma I (также New England Biolabs), создает непрерывный гибридный ген PIX, сопровождаемого EGFP. Полученная плазмида была названа p9GFP, а экспрессируемый слитый белок в данном тексте будет именоваться PIX-GFP.
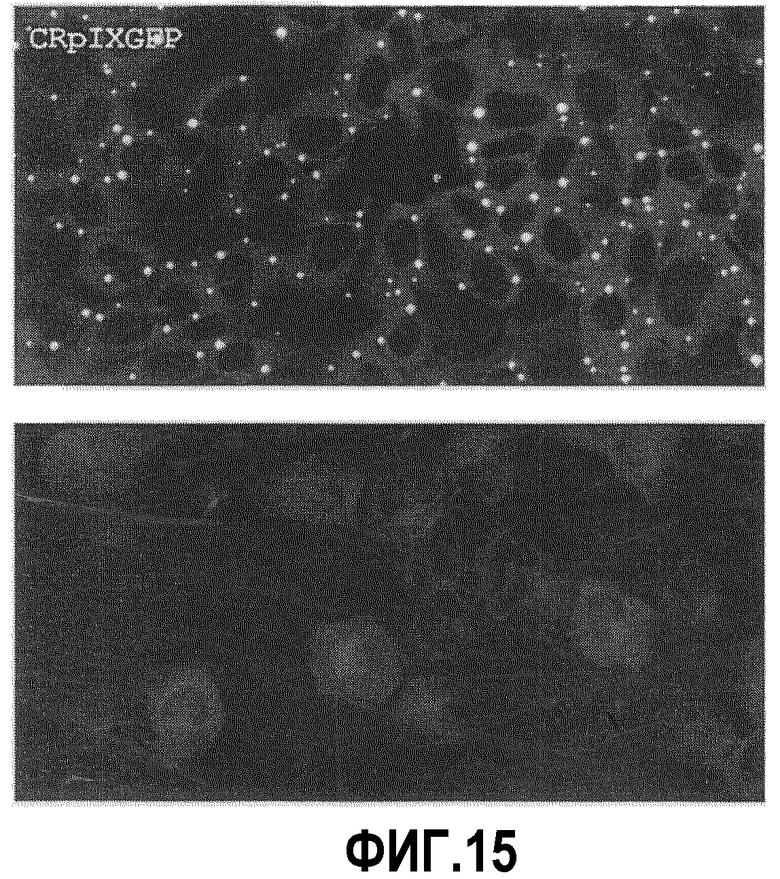
Клетки CS и CR трансфицировали p9GFP и подвергали селекции с помощью 300 мкг/мл генетицина (Invitrogen) на стабильную экспрессию PIX-GFP. В пределах двух недель наблюдали две различающиеся по экспрессии PIX-GFP популяции: одна популяция обычно содержала от 2 до 5 ярких точек PIX-GFP в цитоплазме, а другая популяция имела более однородную экспрессию PIX-GFP в цитоплазме и более диффузную аккумуляцию сигнала GFP в приближенной к ядру области. В обоих случаях клеточные ядра выглядели в виде темной области, что позволяет предполагать, что слитый белок не входит или входит очень слабо в ядро. Типичные примеры клонов обоих типов в CRpIXGFP показаны на фиг.15.
В соответствии с этим результатом заявители обнаружили последовательность экспорта из ядра в открытой рамке считывания PIX при применении поискового алгоритма NetNES (Cour et al., Protein Eng. Des. Sel. 17, 527-536 (2004)) к первичной последовательности PIX. Идентифицированная этой программой NES представляет собой 313-GCACAATTGGATTCTTTGACCCGGGAACTT-342 (SEQ ID NO:17) (транслированная: AQLDSLTREL; SEQ ID NO:18). По сведениям заявителей такой сигнал в белке PIX был описан впервые. И наоборот, последовательность ядерной локализации (NLS) для PIX не была описана в литературе, и заявители не могли выявить такой сигнал (например, при использовании алгоритма, предлагаемого Колумбийским университетом на сайте http://cubic.bioc.columbia.edu/cgi/var/nair/resonline.pl).
Таким образом, цитоплазматическая локализация PIX-GFP в клетках утки находится в соответствии с первичной последовательностью PIX.
Пример 14
Варианты гибрида PIX-GFP
Неожиданное исключение PIX GFP из ядра исследовали с помощью дополнительных гибридных конструкций PIX. Экспрессионная плазмида PIX-GFP-NLS была получена на основе описанного выше конструкта p9GFP путем вставки синтетических олигонуклеотидов i185 и i186. Эти олигонуклеотиды денатурировали при 80°С в течение 5 мин и отжигали в 10 мМ MgCl2 постепенным охлаждением до комнатной температуры с получением:
5'-GATGTACAAAGATCCGAAGAAGAACCGCAAAGGTTAACGCGGCCGCAC-3' (SEQ ID NO:19)
3'-CTACATGTTTCTAGGCTTCTTCTTGGCGTTTCCAATTGCGCCGGCGTG-5' (SEQ ID NO:20).
Этот двухцепочечный олигонуклеотид гидролизовали BsrG I и Not I и вставляли в те же сайты p9GFP. Транслированная вставка добавляет последовательность NLS (PKKNRK; SEQ ID NO:21) к слитому белку PIX-GFP, что напоминает NLS обезьяньего вируса 40. Новый сайт Hpa I, вводимый этой вставкой, служит в качестве диагностического маркера для подтверждения успешного клонирования. Полученная плазмида названа p9 GFP NLS, экспрессируемый белок PIX-GFP-NLS и открытая рамка считывания, кодирующая белок PIX-GFP-NLS, показаны в SEQ ID NO:23 и 22 соответственно.
Меченный GFP слитый белок PIX-RARA получали посредством амплификации ПЦР фрагмента, содержащего PIX-RARA, с праймерами EBR 44A (5'-GGATCCTTCCTCCTCGGGCGGGTGT-3'; SEQ ID NO:24), V293 (5'-AACCAGCGCTACCATGAGCACCAACTCGT 3'; SEQ ID NO:25) и плазмиды #67F pEF PIX RARA NEO в качестве матрицы. Ампликон из 1926 п.н. обрабатывали полинуклеотидкиназой и вставляли в pEGFP-C2 (Clontech), выровненную EcoR I и затупленную ферментом Кленова (все ферменты от New England Biolabs). Полученная плазмида названа p9 GFP RARA, а экспрессируемый белок PIX GFP RARA.
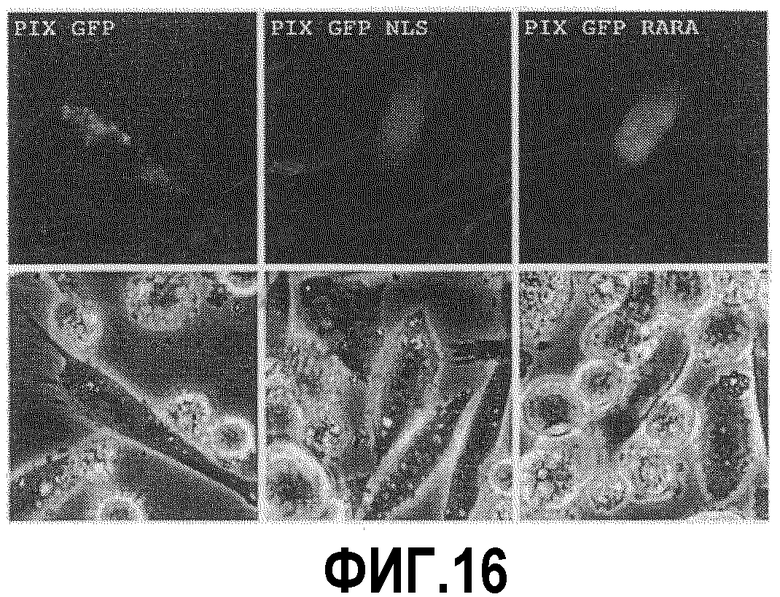
На фиг.16 показано внутриклеточное распределение различных меченных GFP белков во временно трансфицированных клетках CHO.
Пример 15
PIX и интерферон
Многие вирусы индуцируют врожденный клеточный иммунный ответ через TLR-3 (toll-подобный рецептор 3). TLR-3 узнает двухцепочечную РНК, отличительный признак репликации вируса. Одной из функций TLR-3 является активация NFκB и интерферона типа I (Alexopoulou et al., Nature 413, 732-738 (2001)). Интерферон опосредует противовирусное состояние клетки-хозяина. Был исследован эффект индукции интерферона искусственной двухцепочечной РНК, поли I:поли C (полиинозиновая-полицитидиловая кислота, Sigma). Удивительно, что клетки CR и CRpIX проявили очень низкую чувствительность к поли I:поли C, в то время как клетки CS четко отвечали на индуктор. Неожиданно PIX-позитивные клетки CS отвечали быстрее и при более низких концентрациях, чем исходные клетки CS (фиг.17). По сведениям заявителей, связь между PIX и интерфероном не была описана в современной литературе.
Поддерживающий эффект PIX в отношении MVA в клетках CS был сильнее, чем в клетках CR, и клетки CS отвечали на суррогат двухцепочечной РНК лучше, чем клетки CR. Известно, что MVA индуцирует интерферон и что этот вирус также снабжен белками, которые смягчают ответ интерферона. Поэтому определяли действие индуктора поли I:поли C на клетки CRp9GFP и CSp9GFP. Фиг.18 позволяет предполагать, что инфицированные MVA клетки страдают от поли I:поли C в меньшей степени, чем неинфицированный контроль. Это наблюдение находится в соответствии с тем фактом, что MVA и индуцирует клеточный врожденный иммунный ответ, и препятствует ему и связано с неожиданным наблюдением заявителей того, что PIX из аденовируса человека в клетках птиц действует совместно с противовирусными белками крайне ослабленного поксвируса с повышением выхода последнего вируса.
Перечень последовательностей - свободный текст

















Настоящее изобретение относится к области биотехнологии и генной инженерии. Способ получения отличного от аденовируса вируса-мишени или одного или более белков-мишеней предусматривает культивирование экспрессирующей клетки позвоночного и выделение целевого продукта. При этом клетка представляет собой клетку позвоночного, содержащую стабильно интегрированный в ее геном ген, кодирующий PIX аденовируса или его функциональный вариант, в качестве гетерологичного регуляторного белка. Клетка стабильно экспрессирует указанный регуляторный белок или его функциональный вариант вирусом-мишенью, не являющимся аденовирусом, или вектором, несущим последовательности нуклеиновой кислоты, кодирующие указанный вирус, или вектором, несущим последовательности нуклеиновой кислоты, кодирующие указанные один или более белков-мишеней. Предложенный способ позволяет повысить выход целевого продукта при его осуществлении. 7 н. и 17 з.п. ф-лы, 18 ил., 15 пр.
1. Способ получения отличного от аденовируса вируса-мишени, включающий
(a) культивирование экспрессирующей клетки позвоночного, получаемой инфицированием или трансфекцией клетки-хозяина, представляющей собой клетку позвоночного, содержащей стабильно интегрированный в ее геном ген, кодирующий PIX аденовируса или его функциональный вариант, в качестве гетерологичного регуляторного белка, и стабильно экспрессирующей указанный регуляторный белок или его функциональный вариант, вирусом-мишенью, не являющимся аденовирусом, или вектором, несущим последовательности нуклеиновой кислоты, кодирующие указанный вирус, и
(b) выделение вируса-мишени, не являющегося аденовирусом.
2. Способ по п.1, где
(i) гетерологичный регуляторный белок модулирует транскрипцию и/или клеточный рост и повышает продуктивность клетки в отношении продукции вируса, не содержащего указанный регуляторный белок, и продукции белка, отличного от указанного регуляторного белка или его функционального варианта; или
(ii) функциональный вариант гетерологичного регуляторного белка представляет собой слитый белок, причем предпочтительно указанный слитый белок включает по меньшей мере один первый домен, включающий аденовирусный регуляторный белок PIX, как определено выше, и по меньшей мере один второй домен, который модулирует или расширяет активность или внутриклеточное распределение аденовирусного PIX; или
(iii) указанный в (ii) по меньшей мере один второй домен включает белок или пептид, действующий в качестве модулятора транскрипции, предпочтительно указанный модулятор транскрипции представляет собой транскрипционный фактор, включая рецептор альфа-ретиноевой кислоты, представляет собой маркерный белок, указанный маркерный белок предпочтительно представляет собой флуоресцентный маркер, включая GFP, DsRed и их варианты, или представляет собой фермент, включая LacZ, или представляет собой транзитный пептид, включая NLS; или
(iv) указанные первый и второй домены в (ii) и (iii) прямо или посредством пептидного линкера ковалентно соединены друг с другом; или
(v) гетерологичный регуляторный белок или его функциональный вариант экспрессируется в клетке-хозяине и экспрессирующей клетке в количестве по меньшей мере 1 пг/мкг клеточного белка, предпочтительно в количестве по меньшей мере 10 пг/мкг клеточного белка; или
(vi) гетерологичный регуляторный белок или его функциональный вариант находится под контролем стабильного гомологичного или гетерологичного промотора, предпочтительно указанный промотор является конститутивным клеточным промотором, наиболее предпочтительно промотором фактора 2 человека элонгации при транслокации.
3. Способ по п.2, где гетерологичный регуляторный белок
(i) представляет собой PIX аденовируса, предпочтительно представляет собой белок, имеющий последовательность, показанную в SEQ ID NO:2; или
(ii) представляет собой слитый белок PIX аденовируса и рецептора альфа-ретиноевой кислоты, предпочтительно представляет собой слитый белок, имеющий последовательность, показанную в SEQ ID NO:4; или
(iii) представляет собой слитый белок PIX аденовируса и GFP и содержащий последовательность NLS, предпочтительно представляет собой слитый белок, имеющий последовательность, показанную в SEQ ID NO:23; или
(iv) представляет собой слитый белок PIX аденовируса и GFP или PIX и выделенной последовательности NLS.
4. Способ по п.1, где
(i) клетка-хозяин и экспрессирующая клетка представляют собой клетку млекопитающего или клетку птицы, где клетка млекопитающего предпочтительно является клеткой человека или клеткой грызуна, включая клетку мыши, крысы или хомячка, и где клетка птицы предпочтительно является клеткой утки, курицы, перепела или гуся; и/или
(ii) клетка-хозяин и экспрессирующая клетка происходят от первичной клетки или предварительно иммортализованной клетки; и/или
(iii) клетка-хозяин и экспрессирующая клетка несут дополнительно иммортализующие (вирусные) гены, включая белок Е1 аденовируса, предпочтительно мастаденовируса группы С типа 5, наиболее предпочтительно клетка несет ген Е1А и/или Е1В аденовируса, показанный в SEQ ID NO:5; и/или
(iv) клетка дополнительно несет функциональные последовательности, такие как последовательности маркера селекции, донор/акцепторные сайты сплайсинга и/или последовательности, узнаваемые рекомбиназой, обеспечивающие интеграцию последовательности нуклеиновой кислоты-мишени, подлежащей экспрессии в клетке.
5. Способ по п.1, где вирус-мишень выбран из дикого типа, мутантного вируса или вируса с делецией, адаптированного к холоду, или ослабленного вируса, штаммов вакцины, вирусных векторов, несущих гетерологичный(гетерологичные) ген(ы), вирусных векторов, таких как лентивирус, поксвирус, включая вирус коровьей оспы, такой как модифицированная вакцина Анкара, вирус птичьей оспы или вирус канареечной оспы и аденоассоциированный вирус (aav).
6. Способ по п.1, где клетка-хозяин и экспрессирующая клетка происходят из мозга человека и наиболее предпочтительно представляют собой клетку NC5T11, и указанная клетка несет последовательность нуклеиновой кислоты, кодирующую PIX аденовируса или его функциональный вариант в качестве гетерологичного регуляторного белка, предпочтительно указанный гетерологичный регуляторный белок кодируется нуклеиновой кислотой, показанной в SEQ ID NO:1, SEQ ID NO:3 или SEQ ID NO:22.
7. Способ по п.6, где клетка-хозяин представляет собой клетку NC5T11#34, и экспрессирующая клетка происходит от клетки NC5T11#34, причем указанная клетка NC5T11#34 депонирована DSMZ под регистрационным номером DSM АСС2744.
8. Способ по п.1, где клетка-хозяин и экспрессирующая клетка являются клеткой птицы, происходящей из сетчатки утки или сомита утки, и указанная клетка несет последовательность нуклеиновой кислоты, кодирующую PIX аденовируса или его функциональный вариант в качестве гетерологичного регуляторного белка, предпочтительно указанный гетерологичный регуляторный белок кодируется нуклеиновой кислотой, показанной в SEQ ID NO:1, SEQ ID NO:3 или SEQ ID NO:22.
9. Способ по п.8, где клетка-хозяин представляет собой клетку CR.PIX (17a11b), а экспрессирующая клетка происходит от клетки CR.PIX (17a11b), при этом указанная клетка CR.PIX (17a11b) депонирована DSMZ под регистрационным номером DSM АСС2749.
10. Способ получения одного или более белков-мишеней, включающий
(а) культивирование экспрессирующей клетки позвоночного, получаемой инфицированием или трансфекцией клетки-хозяина, представляющей собой клетку позвоночного, содержащей стабильно интегрированный в ее геном ген, кодирующий PIX аденовируса или его функциональный вариант, в качестве гетерологичного регуляторного белка, и стабильно экспрессирующей указанный регуляторный белок или его функциональный вариант, вектором, несущим последовательности нуклеиновой кислоты, кодирующие указанные один или более белков-мишеней, и
(b) выделение одного или более белка(белков)-мишени(мишеней).
11. Способ по п.10, где
(i) гетерологичный регуляторный белок модулирует транскрипцию и/или клеточный рост и повышает продуктивность клетки в отношении продукции вируса, не содержащего указанный регуляторный белок, и продукции белка, отличного от указанного регуляторного белка или его функционального варианта; или
(ii) функциональный вариант гетерологичного регуляторного белка представляет собой слитый белок, причем предпочтительно указанный слитый белок включает по меньшей мере один первый домен, включающий аденовирусный регуляторный белок PIX, как определено выше, и по меньшей мере один второй домен, который модулирует или расширяет активность или внутриклеточное распределение аденовирусного РIХ; или
(iii) указанный в (ii) по меньшей мере один второй домен включает белок или пептид, действующий в качестве модулятора транскрипции, предпочтительно указанный модулятор транскрипции представляет собой транскрипционный фактор, включая рецептор альфа-ретиноевой кислоты, представляет собой маркерный белок, указанный маркерный белок предпочтительно представляет собой флуоресцентный маркер, включая GFP, DsRed и их варианты, или представляет собой фермент, включая LacZ, или представляет собой транзитный пептид, включая NLS; или
(iv) указанные первый и второй домены в (ii) и (iii) прямо или посредством пептидного линкера ковалентно соединены друг с другом; или
(v) гетерологичный регуляторный белок или его функциональный вариант экспрессируется в клетке-хозяине и экспрессирующей клетке в количестве по меньшей мере 1 пг/мкг клеточного белка, предпочтительно в количестве по меньшей мере 10 пг/мкг клеточного белка; или
(vi) гетерологичный регуляторный белок или его функциональный вариант находится под контролем стабильного гомологичного или гетерологичного промотора, предпочтительно указанный промотор является конститутивным клеточным промотором, наиболее предпочтительно промотором фактора 2 человека элонгации при транслокации.
12. Способ по п.11, где гетерологичный регуляторный белок
(i) представляет собой PIX аденовируса, предпочтительно представляет собой белок, имеющий последовательность, показанную в SEQ ID NO:2; или
(ii) представляет собой слитый белок PIX аденовируса и рецептора альфа-ретиноевой кислоты, предпочтительно представляет собой слитый белок, имеющий последовательность, показанную в SEQ ID NO:4; или
(iii) представляет собой слитый белок PIX аденовируса и GFP и содержащий последовательность NLS, предпочтительно представляет собой слитый белок, имеющий последовательность, показанную в SEQ ID NO:23; или
(iv) представляет собой слитый белок PIX аденовируса и GFP или PIX и выделенной последовательности NLS.
13. Способ по п.10, где
(i) клетка-хозяин и экспрессирующая клетка представляют собой клетку млекопитающего или клетку птицы, где клетка млекопитающего предпочтительно является клеткой человека или клеткой грызуна, включая клетку мыши, крысы или хомячка, и где клетка птицы предпочтительно является клеткой утки, курицы, перепела или гуся; и/или
(ii) клетка-хозяин и экспрессирующая клетка происходят от первичной клетки или предварительно иммортализованной клетки; и/или
(iii) клетка-хозяин и экспрессирующая клетка несут дополнительно иммортализующие (вирусные) гены, включая белок Е1 аденовируса, предпочтительно мастаденовируса группы С типа 5, наиболее предпочтительно клетка несет ген Е1А и/или Е1В аденовируса, показанный в SEQ ID NO:5; и/или
(iv) клетка дополнительно несет функциональные последовательности, такие как последовательности маркера селекции, донор/акцепторные сайты сплайсинга и/или последовательности, узнаваемые рекомбиназой, обеспечивающие интеграцию последовательности нуклеиновой кислоты-мишени, подлежащей экспрессии в клетке.
14. Способ по п.10, где один или более белков-мишеней выбраны из антител, рекомбинантных белков, таких как эритропоэтин, альфа-1-антитрипсин, факторы свертывания крови VIII и IX и интерфероны, вирусных антигенов, таких как НА и NA гриппа, HBV-S, белок G герпеса и белок G бешенства, и пептидных гормонов.
15. Способ по п.10, где клетка-хозяин и экспрессирующая клетка происходят из мозга человека и наиболее предпочтительно представляют собой клетку NC5T11 и указанная клетка несет последовательность нуклеиновой кислоты, кодирующую PIX аденовируса или его функциональный вариант в качестве гетерологичного регуляторного белка, предпочтительно указанный гетерологичный регуляторный белок кодируется нуклеиновой кислотой, показанной в SEQ ID NO:1, SEQ ID NO:3 или SEQ ID NO:22.
16. Способ по п.15, где клетка-хозяин представляет собой клетку NC5T11#34 и экспрессирующая клетка происходит от клетки NC5T11#34, причем указанная клетка NC5T11#34 депонирована DSMZ под регистрационным номером DSM АСС2744.
17. Способ по п.10, где клетка-хозяин и экспрессирующая клетка являются клеткой птицы, происходящей из сетчатки утки или сомита утки, и указанная клетка несет последовательность нуклеиновой кислоты, кодирующую PIX аденовируса или его функциональный вариант в качестве гетерологичного регуляторного белка, предпочтительно указанный гетерологичный регуляторный белок кодируется нуклеиновой кислотой, показанной в SEQ ID NO:1, SEQ ID NO:3 или SEQ ID NO:22.
18. Способ по п.17, где клетка-хозяин представляет собой клетку CR.PIX (17a11b), а экспрессирующая клетка происходит от клетки CR.PIX (17a11b), при этом указанная клетка CR.PIX (17a11b) депонирована DSMZ под регистрационным номером DSM АСС2749.
19. Экспрессирующая клетка позвоночного для продукции вируса-мишени, не являющегося аденовирусом, или одного или более белков-мишеней, как определено в любом из пп.1-18, которая стабильно экспрессирует PIX аденовируса или его функциональный вариант в качестве гетерологичного регуляторного белка и которая инфицирована или трансфицирована вирусом-мишенью, не являющимся аденовирусом, или вектором, несущим последовательности нуклеиновой кислоты, кодирующие указанный вирус, или вектором, несущим последовательности нуклеиновой кислоты, кодирующие указанные один или более белков-мишеней.
20. Экспрессирующая клетка позвоночного по п.19, которая представляет собой клетку, определенную в пп.6-9 или 15-18.
21. Способ получения экспрессирующей клетки позвоночного по п.19 или 20, который включает инфицирование или трансфекцию линии клетки-хозяина, представляющей собой клетку позвоночного, определенную в любом из пп.1-18, вирусом-мишенью, не являющимся аденовирусом, или вектором, несущим последовательности нуклеиновой кислоты, кодирующие указанный вирус, или вектором, несущим последовательности нуклеиновой кислоты, кодирующие указанные один или более белков-мишеней.
22. Клетка-хозяин, представляющая собой клетку позвоночного, такая как определено в пп.6-9 или 15-18, которая стабильно экспрессирует PIX аденовируса или его функциональный вариант в качестве гетерологичного регуляторного белка.
23. Способ получения клетки-хозяина, представляющей собой клетку позвоночного, по п.22, который включает трансфекцию подходящей исходной клетки позвоночного вектором, несущим последовательности нуклеиновой кислоты, кодирующие PIX аденовируса, или его функциональный вариант в качестве гетерологичного регуляторного белка.
24. Применение экспрессирующей клетки позвоночного по п.19 или 20 для получения вируса-мишени, не являющегося аденовирусом, или одного или более белков-мишеней.
| KROUGLIAK V | |||
| еt al | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| IMLER J.-L | |||
| et al | |||
| ЭЛЕКТРОМЕХАНИЧЕСКИЙ СНЕГООЧИСТИТЕЛЬ ДЛЯ ЖЕЛЕЗНЫХ ДОРОГ | 1922 |
|
SU549A1 |

