ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к слитым белкам и укороченным белкам, позволяющим воздействовать на пути передачи сигнала, распространяющегося после активации иммунных клеток, и модулировать их.
УРОВЕНЬ ТЕХНИКИ
Адоптивная иммунотерапия с использованием аутологичных T-клеток включает выделение T-клеток из пациента с последующей их стимуляцией, модификацией и/или экспансией ex vivo для получения популяции T-клеток, демонстрирующих противоопухолевую специфичность. После повторной инфузии пациенту эти клетки способны распознавать экспрессируемые опухолью антигены и опосредовать отторжение опухоли.
В ряде исследований в различных условиях уже показано, что этот подход обладает потенциалом стать мощным, эффективным лечением злокачественных новообразований с длительным сроком действия. Например, EBV-регулируемые опухоли, такие как лимфопролиферативное заболевание после трансплантации солидных органов, можно эффективно лечить с помощью подвергнутых экспансии ex vivo EBV-специфических T-клеток.
Аналогичная терапия невирусных злокачественных новообразований включает инфильтрирующие опухоль лимфоциты (TIL), выделенные из резецированных фрагментов опухоли, а затем подвергнутые стимуляции и экспансии с использованием аутологичных образцов опухоли. Подвергнутые экспансии культуры T-клеток, демонстрирующие реактивную способность в отношении опухоли, затем можно повторно инфузировать пациенту.
Вместо селекции и оптимизации специфичности T-клеток с использованием повторяющегося воздействия антигенов, желаемую противоопухолевую специфичность можно придавать T-клеткам посредством генной модификации и встраивания опухолеспецифического T-клеточного рецептора (TCR) или химерного антигенного рецептора (CAR). Эти клетки подвергают экспансии ex vivo для получения достаточных количеств клеток для достижения значимых клинических ответов у пациента.
Однако подходы, подробно описанные выше, имеют ограничения. Например, подвергнутые адоптивному переносу T-клетки могут демонстрировать ограниченное персистирование и экспансию in vivo по причине недостаточной передачи сигнала, отсутствия ИЛ-2 или дифференцировки. В качестве дополнительного примера, подвергнутые адоптивному переносу T-клетки могут подвергаться воздействию ингибиторных стимулов внутри опухолевого микроокружения. Например, они могут становиться истощенными, подвергаться индуцируемой активацией гибели клеток, являющейся результатом гиперактивации, или могут вызывать специфические для мишени, внеопухолевые эффекты.
Другим многообещающим подходом для активации терапевтического противоопухолевого иммунитета является блокада иммунных контрольных точек. Иммунные контрольные точки относятся к различным ингибиторным путям иммунной системы, важным для поддержания аутотолерантности и модуляции длительности и величины физиологических иммунных ответов в периферических тканях.
Известно, что опухоли используют конкретные пути иммунных контрольных точек в качестве основного механизма иммунной резистентности, в частности, против T-клеток, специфических для опухолевых антигенов. Многие из иммунных контрольных точек активируются лиганд-рецепторными взаимодействиями, что означает, что их можно блокировать с помощью антител или модулировать с помощью рекомбинантных форм лигандов или рецепторов. Антитела против антигена 4, ассоциированного с цитотоксическими T-лимфоцитами (CTLA4), являлись первыми из этого класса иммунотерапевтических средств, получивших одобрение Управления по санитарному надзору за качеством пищевых продуктов и медикаментов (FDA). Недавно разработаны блокаторы дополнительных белков иммунных контрольных точек, таких как белок программируемой гибели клеток 1 (PD1), и показано, что они усиливают противоопухолевый иммунитет.
Одной из проблем, связанных с использованием ингибиторов иммунных контрольных точек, является то, что существует множество ингибиторных путей, запускаемых множеством лиганд-рецепторных взаимодействий. При использовании антитела или рекомбинантной формы лиганда/рецептора будут лишь блокировать один такой ингибиторный путь, что оставляет возможность того, что опухоль может компенсировать блокаду специфических иммунных контрольных точек с использованием других молекул.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Авторы настоящего изобретения разработали систему для модуляции и/или воздействия на пути передачи сигнала в иммунных клетках, таких как T-клетки и естественные киллеры (NK).
Внутриклеточные пути передачи сигнала инициируются и контролируются обратимой посттрансляционной модификацией белков. Авторы настоящего изобретения определили, что активирующие и ингибиторные пути передачи сигнала в T-клетках можно модулировать и/или подвергать воздействию с помощью слитых белков или укороченных белков, содержащих домены SH2 из белков прямой передачи T-клеточного сигнала. Другими словами, активирующие и ингибиторные пути передачи сигнала в T-клетках можно модулировать и/или подвергать воздействию с помощью слитых белков или укороченных белков, содержащих домены SH2 из белков, способных связываться с фосфорилированными иммунорецепторными тирозиновыми активирующими мотивами (ITAM) или фосфорилированным иммунорецепторным тирозиновым ингибирующим мотивом (ITIM).
Таким образом, в первом аспекте настоящее изобретение относится к клетке, содержащей химерный антигенный рецептор (CAR) и белок, модифицирующий передачу сигнала, выбранный из одного из следующих:
(i) укороченного белка, содержащего домен SH2 из белка, связывающегося с фосфорилированным иммунорецепторным тирозиновым активирующим мотивом (ITAM), но в котором отсутствует киназный домен;
(ii) укороченного белка, содержащего домен SH2 из белка, связывающегося с фосфорилированным иммунорецепторным тирозиновым ингибирующим мотивом (ITIM), но в котором отсутствует фосфатазный домен;
(iii) слитого белка, содержащего (a) домен SH2 из белка, связывающегося с фосфорилированным иммунорецепторным тирозиновым активирующим мотивом (ITAM), или из белка, связывающегося с фосфорилированным иммунорецепторным тирозиновым ингибирующим мотивом (ITIM); и (ii) гетерологичный домен.
Белок, модифицирующий передачу сигнала, может являться укороченным белком, содержащим домен SH2 ZAP70, но в котором отсутствует киназный домен ZAP70.
Белок, модифицирующий передачу сигнала, может являться укороченным белком, содержащим SH2 PTPN6, но в котором отсутствует фосфатазный домен PTPN6.
Белок, модифицирующий передачу сигнала, может являться укороченным белком, содержащим домен SH2 SHP-2, но в котором отсутствует фосфатазный домен SHP-2.
Белок, модифицирующий передачу сигнала, может являться слитым белком, содержащим (i) домен SH2 из белка, связывающегося с фосфорилированным иммунорецепторным тирозиновым активирующим мотивом (ITAM); и (ii) фосфатазный домен.
Слитый белок, например, может содержать домен SH2 ZAP70, фосфатазный домен PTPN6 или SHP-2.
Белок, модифицирующий передачу сигнала, может являться слитым белком, содержащим (i) домен SH2 из белка, связывающегося с фосфорилированным иммунорецепторным тирозиновым ингибирующим мотивом (ITIM); и (ii) киназный домен.
Слитый белок может содержать домен SH2 из PTPN6 или SHP-2.
Слитый белок может содержать киназный домен Zap70.
Слитый белок может содержать киназный домен AKT или JAK.
Белок, модифицирующий передачу сигнала может являться слитым белком, содержащим (i) домен SH2 из белка, связывающегося с фосфорилированным иммунорецепторным тирозиновым активирующим мотивом (ITAM), или из белка, связывающегося с фосфорилированным иммунорецепторным тирозиновым ингибирующим мотивом (ITIM); и (ii) гетерологичный сигнальный домен.
Слитый белок может содержать домен SH2 из ZAP70, PTPN6 или SHP-2.
Гетерологичный сигнальный домен можно получать из сигнальной молекулы, как правило, не активированной рецептором, содержащим ITAM или ITIM.
Гетерологичный сигнальный домен может являться костимуляторным доменом. В связи с этим, слитый белок может содержать костимуляторный домен CD28, OX40 или 41BB.
Гетерологичный сигнальный домен может являться ингибиторным доменом. В связи с этим, ингибиторный домен может являться эндодоменом CD148 или CD45 или содержать его. Альтернативно, гетерологичный сигнальный домен является эндодоменом ICOS, CD27, BTLA, CD30, GITR или HVEM или содержит его.
Белок, модифицирующий передачу сигнала, может являться слитым белком, содержащим (i) домен SH2 из белка, связывающегося с фосфорилированным иммунорецепторным тирозиновым активирующим мотивом (ITAM); и (ii) ITAM-содержащий домен.
Слитый белок может содержать домен SH2 ZAP70.
ITAM-содержащий домен может являться эндодоменом CD3 дзета или содержать его.
Белок, модифицирующий передачу сигнала, может являться слитым белком, содержащим (i) домен SH2 из белка, связывающегося с фосфорилированным иммунорецепторным тирозиновым ингибирующим мотивом (ITIM); и (ii) ITIM-содержащий домен.
Слитый белок может содержать домен SH2 из PTPN6 или SHP-2.
ITIM-содержащий домен может являться эндодоменом из PD1, PDCD1, BTLA4, LILRB1, LAIR1, CTLA4, KIR2DL1, KIR2DL4, KIR2DL5, KIR3DL1 или KIR3DL3 или содержать его.
Белок, модифицирующий передачу сигнала, может являться слитым белком, содержащим (i) домен SH2 из белка, связывающегося с фосфорилированным иммунорецепторным тирозиновым активирующим мотивом (ITAM), или из белка, связывающегося с фосфорилированным иммунорецепторным тирозиновым ингибирующим мотивом (ITIM); и (ii) протеазный домен.
Слитый белок может содержать домен SH2 из ZAP70, PTPN6 или SHP-2.
Протеазный домен может являться протеазой вируса гравировки табака (TeV) или содержать ее.
Клетка также может содержать мембраносвязанный фактор транскрипции, имеющий участок расщепления протеазой. Расщепление по участку расщепления протеазой может высвобождать фактор транскрипции, что приводит к повышенной экспрессии гена-мишени.
Ген-мишень кодирует цитокин, например, цитокин, выбранный из следующей группы: ИЛ-2, ИЛ-7, ИЛ-15 и ИЛ-12.
В этом варианте осуществления химерный антигенный рецептор (CAR) может являться целевым CAR, содержащим внутриклеточный участок расщепления протеазой.
Целевой CAR может содержать активирующий или костимуляторный эндодомен, и расщепление по участку расщепления протеазой приводит к удалению эндодомена из целевого CAR.
Альтернативно, целевой CAR может содержать ингибиторный эндодомен, и расщепление по участку расщепления протеазой приводит к удалению ингибиторного эндодомена из целевого CAR. Ингибиторный эндодомен может содержать эндодомен CD148 или CD45.
Клетка по настоящему изобретению может содержать два CAR: активирующий CAR, содержащий ITAM-содержащий эндодомен; и целевой CAR, как определено выше.
Альтернативно, клетка по настоящему изобретению может содержать два CAR: ингибиторный CAR, содержащий ITIM-содержащий эндодомен; и целевой CAR, как определено выше.
Во втором аспекте настоящее изобретение относится к конструкции нуклеиновой кислоты, содержащей:
первую последовательность нуклеиновой кислоты, кодирующую химерный антигенный рецептор; и
вторую последовательность нуклеиновой кислоты, кодирующую укороченный белок или слитый белок, как определено в связи с первым аспектом изобретения.
Конструкция нуклеиновой кислоты также может содержать третью последовательность нуклеиновой кислоты, кодирующую мембраносвязанный фактор транскрипции, как определено выше.
Конструкция нуклеиновой кислоты также может содержать третью последовательность нуклеиновой кислоты, кодирующую целевой CAR как определено выше.
Конструкция нуклеиновой кислоты также может содержать четвертую последовательность нуклеиновой кислоты, кодирующую активирующий CAR или ингибиторный CAR, как определено выше.
В третьем аспекте вектор, содержащий конструкцию нуклеиновой кислоты по второму аспекту изобретения или первую и вторую и, необязательно, третью и/или четвертую последовательности нуклеиновой кислоты, как определено выше.
Настоящее изобретение также относится к набору векторов, содержащему первую и вторую и, необязательно, третью и/или четвертую, последовательности нуклеиновой кислоты, как определено выше.
Вектор или набор векторов могут являться ретровирусными или лентивирусными векторами.
В четвертом аспекте настоящее изобретение относится к фармацевтической композиции, содержащей множество клеток по первому аспекту изобретения.
В пятом аспекте настоящее изобретение относится к фармацевтической композиции по четвертому аспекту изобретения для применения в лечении и/или профилактике заболевания.
В шестом аспекте настоящее изобретение относится к способу лечения и/или профилактики заболевания, включающему стадию введения индивидууму фармацевтической композиции по четвертому аспекту изобретения.
Способ может включать следующие стадии:
(i) выделение образца, содержащего клетки, из индивидуума;
(ii) трансдукция или трансфекция клеток с использованием конструкции нуклеиновой кислоты по второму аспекту изобретения, вектора или набора векторов по третьему аспекту изобретения; и
(iii) введение индивидууму клетки по (ii).
В седьмом аспекте настоящее изобретение относится к применению фармацевтической композиции по четвертому аспекту изобретения в производстве лекарственного средства для лечения и/или профилактики заболевания.
Заболевание может являться злокачественным новообразованием.
В восьмом аспекте настоящее изобретение относится к способу получения клетки по первому аспекту изобретения, включающему стадию встраивания конструкции нуклеиновой кислоты по второму аспекту изобретения, вектора или набора векторов по третьему аспекту изобретения в клетку.
Клетку можно получать из образца, выделенного из индивидуума.
В первом дополнительном аспекте настоящее изобретение также относится к слитому белку, содержащему: (i) домен SH2 ZAP70 или PTPN6; и (ii) гетерологичный домен.
Слитый белок может содержать домен SH2 ZAP70 и ITAM-содержащий домен. ITAM-содержащий домен может являться эндодоменом CD3 дзета или содержать его.
Слитый белок может содержать домен SH2 PTPN6 и ITIM-содержащий домен. ITIM-содержащий домен может являться эндодоменом из PDCD1, BTLA4, LILRB1, LAIR1, CTLA4, KIR2DL1, KIR2DL4, KIR2DL5, KIR3DL1 или KIR3DL3 или содержать его.
Слитый белок может содержать домен SH2 PTPN6, и его можно подвергать слиянию с киназным доменом ZAP70.
Слитый белок может содержать домен SH2 ZAP70, слитый с киназным доменом PTPN6.
Слитый белок может содержать: (i) домен SH2 ZAP70 или PTPN6; и (ii) гетерологичный сигнальный домен.
Гетерологичный сигнальный домен можно получать из сигнальной молекулы, как правило, не активированной ITAM-содержащим рецептором. Гетерологичный сигнальный домен может являться или содержать эндодомен CD28, 41BB или OX40. Гетерологичный сигнальный домен может являться эндодоменом ICOS, CD27, BTLA, CD30, GITR или HVEM или содержать его.
Слитый белок может содержать: (i) домен SH2 ZAP70 или PTPN6; и (ii) киназный домен.
Киназный домен может являться киназным доменом AKT или JAK или содержать его.
Слитый белок может содержать: (i) домен SH2 ZAP70 или PTPN6; и (ii) протеазный домен.
Протеазный домен может являться протеазой вируса гравировки табака (TeV) или содержать ее.
Во втором дополнительном аспекте настоящее изобретение относится к укороченному белку, содержащему домен SH2 ZAP70, но в котором отсутствует киназный домен ZAP70.
В третьем дополнительном аспекте настоящее изобретение относится к укороченному белку, содержащему домен SH2 PTPN6, но в котором отсутствует киназный домен PTPN6.
Настоящее изобретение также относится к сигнальной системе, содержащей:
(i) рецептор, содержащий антигенсвязывающий домен, трансмембранный домен и внутриклеточный сигнальный домен, содержащий эндодомен CD3 дзета; и
(ii) слитый белок по первому дополнительному аспекту изобретения, содержащий домен SH2 ZAP70; или укороченный белок по второму дополнительному аспекту изобретения;
где связывание антигена с антигенсвязывающим доменом приводит к связыванию между эндодоменом CD3 дзета и слитым/укороченным белком.
Настоящее изобретение также относится к сигнальной системе, содержащей:
(i) рецептор, содержащий антигенсвязывающий домен, трансмембранный домен и внутриклеточный сигнальный домен, содержащий связывающий домен PTPN6; и
(ii) слитый белок по первому дополнительному аспекту изобретения, содержащий домен SH2 PTPN6; или укороченный белок по третьему дополнительному аспекту изобретения,
где связывание антигена с антигенсвязывающим доменом приводит к связыванию между связывающим доменом PTPN6 и слитым/укороченным белком.
Рецептор может являться T-клеточным рецептором (TCR) или химерным антигенным рецептором (CAR).
В четвертом дополнительном аспекте настоящее изобретение относится к нуклеиновой кислоте, кодирующей слитый белок по первому дополнительному аспекту настоящего изобретения или укороченный белок по второму или третьему дополнительным аспектам настоящего изобретения.
В пятом дополнительном аспекте настоящее изобретение относится к конструкции нуклеиновой кислоты, содержащей последовательность нуклеиновой кислоты, кодирующую слитый белок, содержащий (i) домен SH2 ZAP70 или PTPN6; и (ii) протеазный домен (например, TeV-домен), и последовательность нуклеиновой кислоты, кодирующую мембраносвязанный фактор транскрипции, содержащий:
(i) мембранную связь;
(ii) участок распознавания протеазы; и
(iii) фактор транскрипции.
В шестом дополнительном аспекте настоящее изобретение относится к конструкции нуклеиновой кислоты, содержащей
(a) последовательность нуклеиновой кислоты, кодирующую слитый белок по первому дополнительному аспекту настоящего изобретения, содержащий домен SH2 PTPN6, или укороченный белок по третьему дополнительному аспекту настоящего изобретения; и
(b) последовательность нуклеиновой кислоты, кодирующую рецептор, содержащий ITIM-содержащий эндодомен.
В седьмом дополнительном аспекте настоящее изобретение относится к конструкции нуклеиновой кислоты, содержащей последовательность нуклеиновой кислоты, кодирующую слитый белок, содержащий (i) домен SH2 ZAP70 или PTPN6; и (ii) протеазный домен (например, TeV-домен), и последовательность нуклеиновой кислоты, кодирующую рецептор, содержащий участок расщепления протеазой.
В восьмом дополнительном аспекте настоящее изобретение относится к конструкции нуклеиновой кислоты, содержащей:
(a) последовательность нуклеиновой кислоты, кодирующую слитый белок, содержащий (i) домен SH2 PTPN6; и (ii) протеазный домен (например, TeV-домен);
(b) последовательность нуклеиновой кислоты, кодирующую рецептор, содержащий участок расщепления протеазой; и
(c) последовательность нуклеиновой кислоты, кодирующую рецептор, содержащий ITIM-содержащий эндодомен.
Рецептор может являться T-клеточным рецептором (TCR) или химерным антигенным рецептором (CAR).
Соответственно, в конструкции нуклеиновой кислоты по восьмому аспекту настоящего изобретения последовательность нуклеиновой кислоты (b) может являться T-клеточным рецептором (TCR) или химерным антигенным рецептором (CAR), содержащим:
(i) участок расщепления протеазой между трансмембранным доменом и активирующим эндодоменом; или
(ii) активирующий эндодомен, слитый с ингибиторным эндодоменом посредством участка расщепления протеазой.
В девятом дополнительном аспекте настоящее изобретение относится к вектору, содержащему нуклеиновую кислоту по четвертому дополнительному аспекту настоящего изобретения или конструкцию нуклеиновой кислоты по любому из дополнительных аспектов настоящего изобретения с пятого по девятый.
Вектор может являться ретровирусным вектором или лентивирусным вектором.
В десятом дополнительном аспекте настоящее изобретение относится к клетке, содержащей слитый белок по первому дополнительному аспекту настоящего изобретения или укороченный белок по второму или третьему дополнительным аспектам настоящего изобретения.
В одиннадцатом дополнительном аспекте настоящее изобретение относится к клетке, содержащей (a) слитый белок по первому дополнительному аспекту настоящего изобретения, содержащий домен SH2 PTPN6, или укороченный белок по третьему дополнительному аспекту настоящего изобретения; и (b) рецептор, содержащий ITIM-содержащий эндодомен.
Клетка может являться иммунной клеткой, такой как T-клетка или естественный киллер (NK).
В двенадцатом дополнительном аспекте настоящее изобретение относится к клетке, содержащей слитый белок, содержащий (i) домен SH2 ZAP70 или PTPN6; и (ii) протеазный домен (например, TeV-домен) и рецептор, содержащий участок расщепления протеазой.
В тринадцатом дополнительном аспекте настоящее изобретение относится к клетке, содержащей:
(a) слитый белок, содержащий (i) домен SH2 PTPN6; и (ii) протеазный домен (например, TeV-домен);
(b) рецептор, содержащий участок расщепления протеазой; и
(c) рецептор, содержащий ITIM-содержащий эндодомен.
Рецептор может являться T-клеточным рецептором (TCR) или химерным антигенным рецептором (CAR).
Рецептор (b) может являться T-клеточным рецептором (TCR) или химерным антигенным рецептором (CAR), содержащим:
(i) участок расщепления протеазой между трансмембранным доменом и активирующим эндодоменом; или
(ii) активирующий эндодомен, слитый с ингибиторным эндодоменом посредством участка расщепления протеазой.
В четырнадцатом дополнительном аспекте настоящее изобретение относится к клетке, содержащей нуклеиновую кислоту по четвертому дополнительному аспекту настоящего изобретения или конструкцию нуклеиновой кислоты по любому из дополнительных аспектов настоящего изобретения с пятого по девятый.
В пятнадцатом дополнительном аспекте настоящее изобретение относится к фармацевтической композиции, содержащей множество клеток по любому из дополнительных аспектов настоящего изобретения с десятого по четырнадцатый.
В шестнадцатом дополнительном аспекте настоящее изобретение относится к фармацевтической композиции по пятнадцатому дополнительному аспекту настоящего изобретения для применения в лечении и/или профилактики заболевания.
В семнадцатом дополнительном аспекте настоящее изобретение относится к способу лечения и/или профилактики заболевания, включающему стадию введения индивидууму фармацевтической композиции по пятнадцатому дополнительному аспекту.
Способ может включать следующие стадии:
(i) выделение образца, содержащего T-клетки или NK-клетки, из индивидуума;
(ii) трансдукция или трансфекция T-клеток или NK-клеток с использованием нуклеиновой кислоты по любому из дополнительных аспектов настоящего изобретения с четвертого по девятый или вектора по десятому дополнительному аспекту настоящего изобретения; и
(iii) введение индивидууму T-клеток или NK-клеток по (ii).
В восемнадцатом дополнительном аспекте настоящее изобретение относится к применению фармацевтической композиции по пятнадцатому дополнительному аспекту настоящего изобретения в производстве лекарственного средства для лечения и/или профилактики заболевания.
Заболевание может являться злокачественным новообразованием.
В девятнадцатом дополнительном аспекте настоящее изобретение относится к набору, содержащему нуклеиновую кислоту по четвертому дополнительному аспекту настоящего изобретения, или конструкцию нуклеиновой кислоты по любому из дополнительных аспектов настоящего изобретения с пятого по восьмой, или вектор по девятому дополнительному аспекту настоящего изобретения.
В двадцатом дополнительном аспекте настоящее изобретение относится к набору, содержащему клетку по любому из дополнительных аспектов настоящего изобретения с десятого по четырнадцатый.
В двадцать первом дополнительном аспекте настоящее изобретение относится к способу получения клетки по любому из дополнительных аспектов настоящего изобретения с десятого по четырнадцатый, включающему стадию встраивания последовательности нуклеиновой кислоты по любому из дополнительных аспектов настоящего изобретения с четвертого по восьмой или вектора по девятому дополнительному аспекту настоящего изобретения в клетку.
Клетку можно получать из образца, выделенного из индивидуума.
Дополнительные аспекты по изобретению суммированы в следующих параграфах:
A1. Укороченный белок, содержащий домен SH2 из белка, связывающегося с фосфорилированным иммунорецепторным тирозиновым активирующим мотивом (ITAM), но в котором отсутствует киназный домен.
A2. Укороченный белок по п.A1, содержащий домен SH2 ZAP70, но в котором отсутствует киназный домен ZAP70.
B1. Укороченный белок, содержащий домен SH2 из белка, связывающегося с фосфорилированным иммунорецепторным тирозиновым ингибирующим мотивом (ITIM), но в котором отсутствует фосфатазный домен.
B2. Укороченный белок по п.B1, содержащий домен SH2 PTPN6, но в котором отсутствует фосфатазный домен PTPN6.
B3. Укороченный белок по п.B1, содержащий домен SH2 SHP-2, но в котором отсутствует фосфатазный домен SHP-2.
C1. Слитый белок, содержащий (i) домен SH2 из белка, связывающегося с фосфорилированным иммунорецепторным тирозиновым активирующим мотивом (ITAM); и (ii) фосфатазный домен.
C2. Слитый белок по п.C1, содержащий домен SH2 ZAP70.
C3. Слитый белок по п.C1 или C2, содержащий фосфатазный домен PTPN6 или SHP-2.
D1. Слитый белок, содержащий (i) домен SH2 из белка, связывающегося с фосфорилированным иммунорецепторным тирозиновым ингибирующим мотивом (ITIM); и (ii) киназный домен.
D2. Слитый белок по п.D1, содержащий домен SH2 из PTPN6 или SHP-2.
D3. Слитый белок по п.D1 или D2, содержащий киназный домен Zap70.
D4. Слитый белок по п.D1 или D2, содержащий киназный домен AKT или JAK.
E1. Слитый белок, содержащий (i) домен SH2 из белка, связывающегося с фосфорилированным иммунорецепторным тирозиновым активирующим мотивом (ITAM), или из белка, связывающегося с фосфорилированным иммунорецепторным тирозиновым ингибирующим мотивом (ITIM); и (ii) гетерологичный сигнальный домен.
E2. Слитый белок по п.E1, содержащий домен SH2 из ZAP70, PTPN6 или SHP-2.
E3. Слитый белок по п.E1 или E2, где гетерологичный сигнальный домен получают из сигнальной молекулы, как правило, не активированной ITAM- или ITIM-содержащим рецептором.
E4. Слитый белок по п.E1, E2 или E3, где гетерологичный сигнальный домен является костимуляторным доменом.
E5. Слитый белок по п.E4, содержащий костимуляторный домен CD28, OX40 или 41BB.
E6. Слитый белок по п.E1, E2 или E3, где костимуляторный домен является ингибиторным доменом.
E7. Слитый белок по п.E6, где ингибиторный домен содержит эндодомен CD148 или CD45.
E8. Слитый белок по п.E6, где гетерологичный сигнальный домен является или содержит эндодомен ICOS, CD27, BTLA, CD30, GITR или HVEM.
F1. Слитый белок, содержащий (i) домен SH2 из белка, связывающегося с фосфорилированным иммунорецепторным тирозиновым активирующим мотивом (ITAM); и (ii) ITAM-содержащий домен.
F2. Слитый белок по п.F1, содержащий домен SH2 ZAP70.
F3. Слитый белок по п.F1 или F2, где ITAM-содержащий домен является эндодоменом CD3 дзета или содержит его.
G1. Слитый белок, содержащий (i) домен SH2 из белка, связывающегося с фосфорилированным иммунорецепторным тирозиновым ингибирующим мотивом (ITIM); и (ii) ITIM-содержащий домен.
G2. Слитый белок по п.G1, содержащий домен SH2 из PTPN6 или SHP-2.
G3. Слитый белок по п.G1 или G2, где ITIM-содержащий домен является эндодоменом из PD1, PDCD1, BTLA4, LILRB1, LAIR1, CTLA4, KIR2DL1, KIR2DL4, KIR2DL5, KIR3DL1 или KIR3DL3 или содержит его.
H1. Слитый белок, содержащий (i) домен SH2 из белка, связывающегося с фосфорилированным иммунорецепторным тирозиновым ингибирующим мотивом (ITIM), или из белка, связывающегося с фосфорилированным иммунорецепторным тирозиновым ингибирующим мотивом (ITIM); и (ii) протеазный домен.
H2. Слитый белок по п.H1, содержащий домен SH2 из ZAP70, PTPN6 или SHP-2.
H3. Слитый белок по п.H1 или H2, где протеазный домен является протеазой вируса гравировки табака (TeV) или содержит его.
I1. Последовательность нуклеиновой кислоты, кодирующая укороченный белок по любому из пп.A или B, или слитый белок по любому из пп.C, D, E, F, G или H.
J1. Конструкция нуклеиновой кислоты, содержащая последовательность нуклеиновой кислоты по п.I и последовательность нуклеиновой кислоты, кодирующую химерный антигенный рецептор
J2. Конструкция нуклеиновой кислоты, содержащая последовательность нуклеиновой кислоты по п.I и последовательность нуклеиновой кислоты, кодирующую мембраносвязанный фактор транскрипции, имеющий участок расщепления протеазой.
J3. Конструкция нуклеиновой кислоты, содержащая последовательность нуклеиновой кислоты по п.I и последовательность нуклеиновой кислоты, кодирующую целевой CAR, содержащий внутриклеточный участок расщепления протеазой.
K1. Вектор, содержащий последовательность нуклеиновой кислоты по п.I или конструкцию нуклеиновой кислоты по п.J.
L1. Клетка, содержащая укороченный белок по любому из пп.A или B или слитый белок по любому из пп.C, D, E, F, G или H.
M1. Клетка, содержащая слитый белок по любому из пп.H и мембраносвязанный фактор транскрипции с участком расщепления протеазой.
M2. Клетка по п.M1, где расщепление по участку расщепления протеазой приводит к высвобождению фактора транскрипции, что приводит к повышенной экспрессии гена-мишени.
M3. Клетка по п.M2, где ген-мишень кодирует цитокин.
M4. Клетка по п.M3, где цитокин выбран из следующей группы: ИЛ-2, ИЛ-7, ИЛ-15 и ИЛ-12.
M5. Клетка, содержащая слитый белок по любому из пп.H и целевой рецептор (CAR), содержащий внутриклеточный участок расщепления протеазой.
M6. Клетка по п.M5, где целевой CAR содержит активирующий или костимуляторный эндодомен, и расщепление по участку расщепления протеазой приводит к удалению эндодомена из целевого CAR.
M7. Клетка по п.M5, где целевой CAR содержит ингибиторный эндодомен, и расщепление по участку расщепления протеазой приводит к удалению ингибиторного эндодомена из целевого CAR.
M8. Клетка по п.M7, где ингибиторный эндодомен содержит эндодомен CD148 или CD45.
M9. Клетка, содержащая слитый белок по любому из пп.H и два CAR: активирующий CAR, содержащий ITAM-содержащий эндодомен; и целевой CAR по любому из пп.M5-M8.
M10. Клетка, содержащая слитый белок по любому из пп.H и два CAR: ингибиторный CAR, содержащий ITIM-содержащий эндодомен; и целевой CAR по любому из пп.M5-M8.
Аспекты настоящего изобретения, описанные выше, позволяют модулировать T-клеточные пути передачи сигнала и изменять их посредством, например, механизмов, описанных в таблице 1.
Таблица 1: Использование модуляции сигнала
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фигура 1 (a) - Диаграмма прямых путей активации T-клеток. Активация T-клеточного рецептора приводит к фосфорилированию ITAM. Фосфорилированные ITAM распознаются доменами SH2 ZAP70. После распознавания ZAP70 рекрутируется к околомембранной области, а затем его киназный домен фосфорилирует LAT. Затем фосфорилированный LAT распознается доменами SH2 GRAP, GRB2 и PLC-γ. (b) -Диаграмма прямых путей ингибирования T-клеток. Активация ингибиторного иммунного рецептора, такого как PD1, приводит к фосфорилированию доменов ITIM. Они распознаются доменами SH2 PTPN6. После распознавания PTPN6 рекрутируется к околомембранной области, а затем его фосфатазный домен дефофсфорилирует домены ITAM, ингибирующие иммунную активацию.
Фигура 2 - Диаграмма системы блокирующих сигналов - a) Гиперэкспрессируется укороченный ZAP70, не содержащий киназный домен. Таким образом, он конкурирует с полноразмерным ZAP70 за ITAM и снижает передачу сигнала ITAM. (b) Гиперэкспрессируется укороченный PTPN6, не содержащий фосфатазный домен, конкурируя за полноразмерный PTPN6, снижающий передачу сигнала ITIM.
Фигура 3 - Диаграмма системы перекрестных сигналов: (a) SH2 ZAP70 подвергают слиянию с фосфатазой PTPN6, таким образом, он действует, снижая фосфорилирование ITAM; (b) SH2 PTPN6 подвергают слиянию с киназой ZAP70, что приводит к парадоксальной активации в ответ на ингибиторный сигнал.
Фигура 4 - Диаграмма системы усиленных сигналов: (a) Полноразмерный ZAP70 содержит эндодомен CD3 дзета, прикрепленный к его амино-концу, таким образом, составляется каскад ITAM. (b) Полноразмерный PTPN6 содержит эндодомен PD1, прикрепленный к его амино-концу, таким образом, составляется каскад ITIM.
Фигура 5 - Диаграмма примеров системы обходных сигналов: (a) ZAP70, слитый с эндодоменом CD28; (b) ZAP70, слитый с эндодоменом 41BB; (c) ZAP70, слитый с киназой AKT; (d) домен SH2 PTPN6 подвергают слиянию с эндодоменом 41BB.
Фигура 6 - Диаграмма иллюстративной системы транскрипционных сигналов: a) Слитый белок ZAP-TeV коэкспрессируется с мембраносвязанным фактором транскрипции, который может высвобождаться от мембраны посредством расщепления его мотива распознавания TeV. Это показано посредством коэкспрессии с CAR CD19. Таким образом, после распознавания CD19 на клетке-мишени T-клетка становится активированной, и, кроме того, фактор транскрипции становится активным. (b) Альтернативная система с использованием слитого белка PTPN6-TeV вместо него. В этом случае CAR состоит из ITIM-несущего эндодомена. Таким образом, после распознавания CD19 посредством CAR фактор транскрипции становится активным, но не зависит от активации T-клеток.
Фигура 7 - Диаграмма системы стерилизующих сигналов: представлены два CAR - один из которых распознает CD19 и является активирующим, и один из которых распознает CD33 и является ингибирующим - эти признаки являются исключительно иллюстративными (a) Элемент "И НЕТ" стерилизующего сигнала; в этом случае слитый белок SH2-Tev рекрутируется к активированному ITIM CAR после его активации. Это приводит к отщеплению ITAM от активирующего CAR, сконструированного таким образом, что участок расщепления TeV соединяет трансмембранный домен с доменом ITAM. Таким образом, ингибируют активирующий CAR. (b) Элемент "И" стерилизующего сигнала: В этом случае слитый белок SH2-Tev рекрутируется к ITIM CAR после его активации. Это приводит к высвобождению фосфатазного домена из активирующего CAR, сконструированного таким образом, что фосфатазу соединяют с его карбокси-концом посредством домена расщепления TeV. Это приводит к конститутивному ингибированию, что позволяет CAR активироваться в присутствие когнатного антигена.
Фигура 8 - Конструировали несколько слитых белков различных доменов SH2 и киназного домена AKT: ZAP-AKT, GRAP-AKT, GRB-AKT и PLC-γ.
Фигура 9 - (a) Окрашивание на фосфо-AKT T-клеток, трансдуцированных с использованием различных слитых белков SH2/AKT с активацией митогенным антителом OKT3 и без нее. (b) Окрашивание на фосфо-AKT T-клеток, трансдуцированных с использованием слитого белка ZAP-AKT, улучшенного слитого белка ZAP-AKT, где ZAP и AKT соединяют гибким линкером, и контрольного ZAP-AKT, где замена R190K приводит к устранению способности ZAP связываться с ITAM. T-клетки стимулировали OKT3 или не стимулировали OKT3. Графики FACS наложены на графики для нетрансдуцированных T-клеток.
Фигура 10 - (a) Блоттинг фосфо-AKT T-клеток, активированных повышающимися количествами OKT3. (b) Микроскопия ZAP-AKT или контрольных T-клеток, нестимулированных, стимулированных OKT3 или стимулированных OKT3 и ИЛ-2. T-клетки ZAP-AKT, стимулированные только OKT3, схожи с нетрансдуцированными T-клетками, стимулированными OKT3 и ИЛ-2.
Фигура 11 - (a) Вариант осуществления прямого транскрипционного переключателя TeV. Эндодомен CAR CD19 заменяют протеазой TeV. Также коэкспрессируется мембраносвязанный фактор транскрипции VP16/GAL4. С помощью люциферазного репортера определяют активность VP16/GAL5. (b) Вариант осуществления с ZAP-TeV. Стандартный CAR CD19 коэкспрессируется со слитым белком ZAP-TeV вместе с мембраносвязанным фактором транскрипции.
Фигура 12 - Активность транскрипционных переключателей на основе ZAP-TeV и контроль экспрессии T-клетки после воздействия CD19-отрицательных (слева) или CD19-положительных (справа) мишеней. Активность измеряют по светоотдаче после добавления люциферазы. Порядок тестируемых условий: (a) CAR против CD19, коэкспрессируемый с ZAP-TeV; (b) CAR против CD19, коэкспрессируемый с неактивным (R190K); (c) CAR против CD19, коэкспрессируемый с ZAP-TeV и мембраносвязанным фактором транскрипции; (d) CAR против CD19, коэкспрессируемый с неактивным (R190K) ZAP-TEV, коэкспрессируемым с мембраносвязанным фактором транскрипции; (e) слитый белок CAR против CD19/TeV, коэкспрессируемый с мембраносвязанным фактором транскрипции; (f) конститутивно активный фактор транскрипции GAL4/VP16.
Фигура 13 - (a) Общая структура CAR: Связывающий домен распознает антиген; спейсер отделяет связывающий домен от поверхности клетки; трансмембранный домен заякоривает белок в мембране, и эндодомен передает сигнал. (b)-(d): различные варианты и перестановки эндодоменов CAR: (b) в исходном дизайне сигналы ITAM передаются в отдельности через FcεR1-γ или эндодомен CD3ζ, в то время как в последнем дизайне передаются дополнительные (c) один или (d) два костимуляторных сигнала в цис-положении.
Фигура 14 - Иллюстративные белковые последовательности по настоящему изобретению.
Фигура 15 - Блокада сигнала PD-1 с использованием укороченного SHP-1 (PTPN6) или укороченного SHP-2.
Клетки PBMC контрансдуцировали с использованием PD1 и CAR в отдельности (FMC63); или использовали бицистронную конструкцию, содержащую CAR и укороченный SHP-1 или CAR и укороченный SHP-2. Эти клетки сокультивировали в течение 48 часов с клетками SupT1, трансдуцированными с CD19, PDL1 или и того и другого, и высвобождение ИФНγ измеряли посредством ELISA.
Фигура 16 - Перехват сигнала PD-1 с использованием слияния доменов SH2 SHP-2 и киназы Zap70.
Клетки PBMC котрансдуцировали с использованием PD1 и CAR в отдельности (FMC63); или использовали бицистронную конструкцию, содержащую CAR и слитый белок, содержащий домены SH2 SHP-2 и киназу ZAP70. Эти клетки сокультивировали в соотношении 1:1 в течение 24 часов с клетками SupT1, трансдуцированными с использованием CD19 или PDL1. Высвобождение ИФНγ измеряли посредством ELISA (A) и уничтожение клеток SupT1 количественно оценивали с помощью FACS (B).
ПОДРОБНОЕ ОПИСАНИЕ
БЕЛОК
Настоящее изобретение относится к укороченному белку, содержащему домен SH2.
Настоящее изобретение также относится к слитому белку, содержащему (i) домен SH2 и (ii) гетерологичный домен.
Домен SH2 можно получать из белка, связывающегося с фосфорилированным иммунорецепторным тирозиновым активирующим мотивом (ITAM), или из белка, связывающегося с фосфорилированным иммунорецепторным тирозиновым ингибирующим мотивом (ITIM).
Примером белка, связывающегося с ITAM, является ZAP70. Примеры белков, связывающихся с ITIM, включают PTPN6 и SHP-2
Таким образом, слитый белок по изобретению содержит домен SH2 и по меньшей мере один дополнительный домен, не присутствующий в белке дикого типа, из которого получали домен SH2.
ДОМЕН ГОМОЛОГИИ SRC 2 (SH2)
Внутриклеточные пути передачи сигнала инициируются и контролируются обратимой посттрансляционной модификацией белков, включая фосфорилирование, убиквитинилирование и ацетилирование.
Домены SH2 являются модульными белковыми доменами, служащими в качестве адаптеров и опосредующими белок-белковые взаимодействия посредством связывания с фосфорилированными пептидами в соответствующих белковых партнерах по связыванию, зачастую рецепторами поверхности клетки. Как правило, домены SH2 связываются с фосфорилированными остатками тирозина в контексте более длинного пептидного мотива в белке-мишени, и домены SH2 представляют собой самый большой класс известных pTyr-распознающих доменов.
Хотя у доменов SH2 отсутствует какая-либо присущая им каталитическая активность, они часто соединены с независимыми каталитическими доменами и, таким образом, в ответ на специфический входной сигнал действуют, локализуя эти каталитические домены в конкретные субклеточные локации или вблизи подходящих субстратов, активаторов или ингибиторов. Кроме того, домены SH2 также можно обнаружить связанными с адаптерными белковыми доменами, и, таким образом, они могут участвовать в образовании крупных мультибелковых комплексов.
ДЗЕТА-ЦЕПЬ-АССОЦИИРОВАННАЯ ПРОТЕИНКИНАЗА 70 (ZAP70)
ZAP70 является белком, как правило, экспрессирующимся вблизи поверхностной мембраны T-клеток и естественных киллеров. Он является частью T-клеточного рецептора (TCR) и играет критическую роль в передаче T-клеточного сигнала. Его молекулярная масса составляет 70 кДа, и он состоит из 2 N-концевых доменов SH2 и C-концевого киназного домена. Он является частью семейства протеин-тирозинкиназ.
Самой ранней стадией активации T-клеток является распознавание комплекса пептид-MHC на клетке-мишени посредством TCR. Это исходное событие вызывает связывание киназы Lck с цитоплазматическим хвостом CD3 дзета в комплексе TCR. Затем Lck фосфорилирует остатки тирозина в цитоплазматическом хвосте CD3 дзета, что делает возможным рекрутирование ZAP70. ZAP70 является SH2-содержащей киназой, играющей ключевую роль в активации T-клеток после вовлечения TCR. Тандемные домены SH2 в ZAP70 связываются с фосфорилированным CD3, что приводит к фосфорилированию и активации ZAP70 посредством Lck или другими молекулами ZAP70 в транс-положении. Затем активный ZAP70 способен фосфорилировать нижележащие мембранные белки, ключевым среди которых является линкер активированных T-клеток (LAT). LAT является каркасным белков, и его фосфорилирование по множеству остатков позволяет ему взаимодействовать с некоторыми другими белками, содержащими домен SH2, включая Grb2, PLC-g и Grap, распознающими фосфорилированные пептиды в LAT и передающими сигнал активации T-клеток ниже, что в конечном итоге приводит к диапазону T-клеточных ответов. Этот процесс представлен на фигуре 1.
Белок ZAP70 человека имеет регистрационный номер UniProtKB P43403. Эта последовательность составляет аминокислот 619 в длину и представлена как SEQ ID NO: 1.
Аминокислотная последовательность ZAP70 (SEQ ID NO: 1)
MPDPAAHLPFFYGSISRAEAEEHLKLAGMADGLFLLRQCLRSLGGYVLSLVHDVRFHHFPIERQLNGTYAIAGGKAHCGPAELCEFYSRDPDGLPCNLRKPCNRPSGLEPQPGVFDCLRDAMVRDYVRQTWKLEGEALEQAIISQAPQVEKLIATTAHERMPWYHSSLTREEAERKLYSGAQTDGKFLLRPRKEQGTYALSLIYGKTVYHYLISQDKAGKYCIPEGTKFDTLWQLVEYLKLKADGLIYCLKEACPNSSASNASGAAAPTLPAHPSTLTHPQRRIDTLNSDGYTPEPARITSPDKPRPMPMDTSVYESPYSDPEELKDKKLFLKRDNLLIADIELGCGNFGSVRQGVYRMRKKQIDVAIKVLKQGTEKADTEEMMREAQIMHQLDNPYIVRLIGVCQAEALMLVMEMAGGGPLHKFLVGKREEIPVSNVAELLHQVSMGMKYLEEKNFVHRDLAARNVLLVNRHYAKISDFGLSKALGADDSYYTARSAGKWPLKWYAPECINFRKFSSRSDVWSYGVTMWEALSYGQKPYKKMKGPEVMAFIEQGKRMECPPECPPELYALMSDCWIYKWEDRPDFLTVEQRMRACYYSLASKVEGPPGSTQKAEAACA
Слитый белок по изобретению может содержать домен SH2 ZAP70. Укороченный белок по изобретению может содержать домен SH2 ZAP70 или состоять из нее. В связи с этим, слитый или укороченный белок может содержать последовательность, представленную как SEQ ID NO: 2, или состоять из нее.
Полный домен SH2 ZAP70 (SEQ ID NO: 2)
MPDPAAHLPFFYGSISRAEAEEHLKLAGMADGLFLLRQCLRSLGGYVLSLVHDVRFHHFPIERQLNGTYAIAGGKAHCGPAELCEFYSRDPDGLPCNLRKPCNRPSGLEPQPGVFDCLRDAMVRDYVRQTWKLEGEALEQAIISQAPQVEKLIATTAHERMPWYHSSLTREEAERKLYSGAQTDGKFLLRPRKEQGTYALSLIYGKTVYHYLISQDKAGKYCIPEGTKFDTLWQLVEYLKLKADGLIYCLKEACPNSSASNASGAAAPTLPAHPSTLTHP
ZAP70 содержит два домена SH2 на N-конце последовательности, остатки 10-102 и 163-254 последовательности, представленной как SEQ ID NO: 1. Укороченный белок или слитый белок по изобретению может содержать одну или обе последовательности, представленные как SEQ ID NO: 3 и 4.
SH2 1 ZAP70 (SEQ ID NO: 3)
FFYGSISRAEAEEHLKLAGMADGLFLLRQCLRSLGGYVLSLVHDVRFHHFPIERQLNGTYAIAGGKAHCGPAELCEFYSRDPDGLPCNLRKPC
SH2 2 ZAP70 (SEQ ID NO: 4)
WYHSSLTREEAERKLYSGAQTDGKFLLRPRKEQGTYALSLIYGKTVYHYLISQDKAGKYCIPEGTKFDTLWQLVEYLKLKADGLIYCLKEAC
Слитый белок может содержать вариант SEQ ID NO: 2, 3 или 4, имеющий по меньшей мере 80, 85, 90, 95, 98 или 99% идентичности последовательности, при условии, что вариант последовательности является последовательностью домена SH2, имеющей необходимые свойства. Другими словами, вариант последовательности должен быть способен связываться с фосфорилированными остатками тирозина в цитоплазматическом хвосте CD3 дзета, что делает возможным рекрутирование ZAP70.
Способы выравнивания последовательностей хорошо известны в этой области, и их осуществляют с использованием подходящих программ для выравнивания. % идентичности последовательности относится к процентной доли остатков аминокислот или нуклеотидов, являющихся идентичными в двух последовательностях, когда они оптимально выровнены. Гомологию или идентичность нуклеотидных и белковых последовательностей можно определять с использованием стандартных алгоритмов, таких как программа BLAST (Basic Local Alignment Search Tool от National Center for Biotechnology Information) с использованием параметров по умолчанию, общедоступная на http://blast.ncbi.nlm.nih.gov. Другие алгоритмы для определения идентичности или гомологии последовательности включают: LALIGN (http://www.ebi.ac.uk/Tools/psa/lalign/ и http://www.ebi.ac.uk/Tools/psa/lalign/nucleotide.html), AMAS (Analysis of Multiply Aligned Sequences на http://www.compbio.dundee.ac.uk/Software/Amas/amas.html), FASTA (http://www.ebi.ac.uk/Tools/sss/fasta/), Clustal Omega (http://www.ebi.ac.uk/Tools/msa/clustalo/), SIM (http://web.expasy.org/sim/), и EMBOSS Needle (http://www.ebi.ac.uk/Tools/psa/emboss_needle/nucleotide.html).
В определенных вариантах осуществления слитый белок может содержать домен SH2 ZAP70 и киназный домен ZAP70. Например, слитый белок может содержать последовательность, представленную как SEQ ID NO: 1 или ее вариант, имеющий по меньшей мере 80, 85, 90, 95, 98 или 99% идентичности последовательности.
ТИРОЗИН-ПРОТЕИНФОСФАТАЗА НЕРЕЦЕПТОРНОГО ТИПА 6 (PTPN6)
PTPN6 также известен как фосфатаза-1, содержащая домен области гомологии Src 2 (SHP-1). Он является членом семейства белковых тирозинфосфатаз.
N-концевая область PTPN6 содержит два тандемных домена SH2, опосредующих взаимодействие PTPN6 и его субстратов. C-концевая область содержит тирозин-протеинфосфатазный домен.
PTPN6 способен связываться и распространять сигналы от ряда ингибиторных иммунных рецепторов или ITIM-содержащих рецепторов. Неограничивающие примеры таких рецепторов включают PD1, PDCD1, BTLA4, LILRB1, LAIR1, CTLA4, KIR2DL1, KIR2DL4, KIR2DL5, KIR3DL1 и KIR3DL3.
Белок PTPN6 человека имеет регистрационный номер UniProtKB P29350. Эта последовательность составляет 595 аминокислот в длину и представлена как SEQ ID NO: 5.
Аминокислотная последовательность PTPN6 (SEQ ID NO: 5)
MVRWFHRDLSGLDAETLLKGRGVHGSFLARPSRKNQGDFSLSVRVGDQVTHIRIQNSGDFYDLYGGEKFATLTELVEYYTQQQGVLQDRDGTIIHLKYPLNCSDPTSERWYHGHMSGGQAETLLQAKGEPWTFLVRESLSQPGDFVLSVLSDQPKAGPGSPLRVTHIKVMCEGGRYTVGGLETFDSLTDLVEHFKKTGIEEASGAFVYLRQPYYATRVNAADIENRVLELNKKQESEDTAKAGFWEEFESLQKQEVKNLHQRLEGQRPENKGKNRYKNILPFDHSRVILQGRDSNIPGSDYINANYIKNQLLGPDENAKTYIASQGCLEATVNDFWQMAWQENSRVIVMTTREVEKGRNKCVPYWPEVGMQRAYGPYSVTNCGEHDTTEYKLRTLQVSPLDNGDLIREIWHYQYLSWPDHGVPSEPGGVLSFLDQINQRQESLPHAGPIIVHCSAGIGRTGTIIVIDMLMENISTKGLDCDIDIQKTIQMVRAQRSGMVQTEAQYKFIYVAIAQFIETTKKKLEVLQSQKGQESEYGNITYPPAMKNAHAKASRTSSKHKEDVYENLHTKNKREEKVKKQRSADKEKSKGSLKRK
Слитый белок по изобретению может содержать домен SH2 PTPN6. Укороченный белок по изобретению может содержать домен SH2 PTPN6 или состоять из него. В связи с этим, слитый или укороченный белок может содержать последовательность, представленную как SEQ ID NO: 6, или состоять из нее.
Полный домен SH2 PTPN6 (SEQ ID NO: 6)
MVRWFHRDLSGLDAETLLKGRGVHGSFLARPSRKNQGDFSLSVRVGDQVTHIRIQNSGDFYDLYGGEKFATLTELVEYYTQQQGVLQDRDGTIIHLKYPLNCSDPTSERWYHGHMSGGQAETLLQAKGEPWTFLVRESLSQPGDFVLSVLSDQPKAGPGSPLRVTHIKVMCEGGRYTVGGLETFDSLTDLVEHFKKTGIEEASGAFVYLRQPYY
PTPN6 содержит два домена SH2 на N-конце последовательности, остатки 4-100 и 110-213 последовательности, представленной как SEQ ID NO: 5. Укороченный белок или слитый белок по изобретению может содержать одну или обе последовательности, представленные как SEQ ID NO: 3 и 4.
SH2 1 PTPN6 (SEQ ID NO: 7)
WFHRDLSGLDAETLLKGRGVHGSFLARPSRKNQGDFSLSVRVGDQVTHIRIQNSGDFYDLYGGEKFATLTELVEYYTQQQGVLQDRDGTIIHLKYPL
SH2 2 PTPN6 (SEQ ID NO: 8)
WYHGHMSGGQAETLLQAKGEPWTFLVRESLSQPGDFVLSVLSDQPKAGPGSPLRVTHIKVMCEGGRYTVGGLETFDSLTDLVEHFKKTGIEEASGAFVYLRQPY
Слитый белок может содержать вариант SEQ ID NO: 6, 7 или 8, имеющий по меньшей мере 80, 85, 90, 95, 98 или 99% идентичности последовательности, при условии, что вариант последовательности является последовательностью домена SH2, имеющей необходимые свойства. Другими словами, вариант последовательность должен быть способен связываться с фосфорилированными остатками тирозина в цитоплазматическом хвосте по меньшей мере одного из PD1, PDCD1, BTLA4, LILRB1, LAIR1, CTLA4, KIR2DL1, KIR2DL4, KIR2DL5, KIR3DL1 или KIR3DL3, что делает возможным рекрутирование PTPN6.
В определенных вариантах осуществления слитый белок может содержать домен SH2 PTPN6 и фосфатазный домен PTPN6. Например, слитый белок может содержать последовательность, представленную как SEQ ID NO: 5 или ее вариант, имеющий по меньшей мере 80, 85, 90, 95, 98 или 99% идентичности последовательности.
SHP-2
SHP-2, также известный как PTPN11, PTP-1D и PTP-2C, является членом семейства белковых тирозинфосфатаз (PTP). Подобно PTPN6, SHP-2 имеет доменную структуру, состоящую из двух тандемных доменов SH2 на его N-конце, за которыми следует домен белковой тирозинфосфатазы (PTP). В неактивном состоянии N-концевой домен SH2 связывается с доменом PTP и блокирует доступ потенциального субстрата к активному центру. Таким образом, SHP-2 самоингибируется. После связывания с целевыми фосфо-тирозильными остатками, N-концевой домен SH2 высвобождается из домена PTP, каталитически активируя фермент, облегчая аутоингибирование.
SHP-2 человека имеет регистрационный номер UniProtKB P35235-1. Эта последовательность составляет 597 аминокислот в длину и представлена как SEQ ID NO: 9.
Аминокислотная последовательность SHP-2 (SEQ ID NO: 9)
MTSRRWFHPNITGVEAENLLLTRGVDGSFLARPSKSNPGDFTLSVRRNGAVTHIKIQNTGDYYDLYGGEKFATLAELVQYYMEHHGQLKEKNGDVIELKYPLNCADPTSERWFHGHLSGKEAEKLLTEKGKHGSFLVRESQSHPGDFVLSVRTGDDKGESNDGKSKVTHVMIRCQELKYDVGGGERFDSLTDLVEHYKKNPMVETLGTVLQLKQPLNTTRINAAEIESRVRELSKLAETTDKVKQGFWEEFETLQQQECKLLYSRKEGQRQENKNKNRYKNILPFDHTRVVLHDGDPNEPVSDYINANIIMPEFETKCNNSKPKKSYIATQGCLQNTVNDFWRMVFQENSRVIVMTTKEVERGKSKCVKYWPDEYALKEYGVMRVRNVKESAAHDYTLRELKLSKVGQALLQGNTERTVWQYHFRTWPDHGVPSDPGGVLDFLEEVHHKQESIVDAGPVVVHCSAGIGRTGTFIVIDILIDIIREKGVDCDIDVPKTIQMVRSQRSGMVQTEAQYRFIYMAVQHYIETLQRRIEEEQKSKRKGHEYTNIKYSLVDQTSGDQSPLPPCTPTPPCAEMREDSARVYENVGLMQQQRSFR
Слитый белок по изобретению может содержать домен SH2 SHP-2. Укороченный белок по изобретению может содержать домен SH2 SHP-2 или состоять из него. В связи с этим, слитый или укороченный белок может содержать первый домен SH2 SHP-2, например, содержащий аминокислоты 6-102 SEQ ID NO: 9, или второй домен SH2 SHP-2, например, содержащий аминокислоты 112-216 SHP-2, или состоять из него. Слитый или укороченный белок может содержать последовательность, представленную как SEQ ID NO: 10, 11 или 12, или состоять из нее.
Первый домен SH2 SHP-2 (SEQ ID NO: 10)
WFHPNITGVEAENLLLTRGVDGSFLARPSKSNPGDFTLSVRRNGAVTHIKIQNTGDYYDLYGGEKFATLAELVQYYMEHHGQLKEKNGDVIELKYPL
Второй домен SH2 SHP-2 (SEQ ID NO: 11)
WFHGHLSGKEAEKLLTEKGKHGSFLVRESQSHPGDFVLSVRTGDDKGESNDGKSKVTHVMIRCQELKYDVGGGERFDSLTDLVEHYKKNPMVETLGTVLQLKQPL
Оба домена SH2 SHP-2 (SEQ ID NO: 12)
WFHPNITGVEAENLLLTRGVDGSFLARPSKSNPGDFTLSVRRNGAVTHIKIQNTGDYYDLYGGEKFATLAELVQYYMEHHGQLKEKNGDVIELKYPLNCADPTSERWFHGHLSGKEAEKLLTEKGKHGSFLVRESQSHPGDFVLSVRTGDDKGESNDGKSKVTHVMIRCQELKYDVGGGERFDSLTDLVEHYKKNPMVETLGTVLQLKQPL
Слитый белок может содержать вариант SEQ ID NO: 10, 11 или 12, имеющий по меньшей мере 80, 85, 90, 95, 98 или 99% идентичности последовательности, при условии, что вариант последовательность является последовательностью домена SH2, способной связываться с ITIM-содержащим доменом. Например, вариант последовательности может быть способен связываться с фосфорилированными остатками тирозина в цитоплазматическом хвосте PD1, PDCD1, BTLA4, LILRB1, LAIR1, CTLA4, KIR2DL1, KIR2DL4, KIR2DL5, KIR3DL1 или KIR3DL3.
ГЕТЕРОЛОГИЧНЫЙ ДОМЕН
В рамках изобретения термин "гетерологичный домен" относится к любому белковому домену, не присутствующему в:
i) ZAP70 дикого типа (см. SEQ ID NO: 1) в случае слитых белков, содержащих домен SH2 ZAP70;
ii) PTPN6 дикого типа (см. SEQ ID NO: 5) в случае слитых белков, содержащих домен SH2 PTPN6; или
iii) дикого типа SHP-2 (см. SEQ ID NO: 9) в случае слитых белков, содержащих домен SH2SHP-2.
Гетерологичный домен может являться другим белком из ZAP70, SHP-2 или PTPN6 или его можно получать из них (например, он может являться их частью).
Альтернативно, слитый белок может содержать продукт слияния домена SH2 ZAP70 и домена из PTPN6, такого как киназный домен PTPN6. К тому же, слитый белок может содержать продукт слияния домен SH2 PTPN6 и домена из ZAP70, такого как киназный домен ZAP70.
УСИЛЕННЫЙ СИГНАЛ
Настоящее изобретение относится к слитому белку, содержащему: домен SH2 из ITAM-связывающего белка; и ITAM-содержащий домен.
Настоящее изобретение также относится к слитому белку, содержащему: домен SH2 из ITIM-связывающего белка; и ITIM-содержащий домен.
Эти "усиленные" сигнальные молекулы будут усиливать возбуждающий или ингибиторный сигнал внутри иммунной клетки, такой как T-клетка.
Как показано на фигуре 4, наличие таких молекул будет приводить к каскаду ITAM или ITIM, что приводит к усиленному активирующему или ингибиторному сигналу, соответственно.
Амплификация активирующего сигнала применима в ситуациях, когда желательно повышать чувствительность иммунной клетки (такой как CAR-T-клетка) к антигену. Это может иметь место, когда, например, антиген-мишень экспрессируется на клетках-мишенях на низких уровнях.
Амплификация ингибиторных систем применима в ситуациях, когда желательно снижать или предотвращать активацию T-клеток. В WO2015/075469 описывают панель "логических элементов" пар химерных антигенных рецепторов, которые при экспрессии клеткой, такой как T-клетка, способны определять конкретный паттерн экспрессии по меньшей мере двух антигенов-мишеней A и B). "Элемент И НЕТ", описываемый в настоящей заявке, содержит пару CAR таким образом, что T-клетка запускается, только когда антиген A, но не антиген B присутствует на клетке-мишени. В этом элементе "И НЕТ" один CAR (распознающий антиген A) содержит активирующий эндодомен, содержащий и ITAM, в то время как другой CAR (распознающий антиген B) содержит ингибиторный эндодомен, который может содержать ITIM. В присутствие антигена A в отдельности наличия нелигированного ингибиторного CAR недостаточно для предотвращения активации T-клеток, таким образом, происходит активация. Однако в присутствие обоих антигенов образуются области мембраны с высокими концентрациями активирующих и ингибиторных CAR. Т.к. оба эндодомена сконцентрированы, происходит предотвращение или снижение активации T-клеток.
Амплификацию ингибиторного сигнала с использованием усиленной сигнальной молекулы по настоящему изобретению можно использовать в элементе "И" для снижения или устранения любой остаточной передачи сигнала, происходящей в присутствие обоих антигенов, т.е. при неполном ингибировании активирующего CAR ингибиторным CAR.
ITAM-СОДЕРЖАЩИЙ ДОМЕН
В одном из вариантов осуществления слитый белок содержит домен SH2 ZAP70 и домен, содержащий иммунорецепторный тирозиновый активирующий мотив (ITAM).
Слияние полноразмерного ZAP70 с ITAM-содержащим доменом приводит к получению структуры, усиливающей активирующий иммунный сигнал. В этом случае слитый белок рекрутируется к иммунорецепторному эндодомену фосфо-ITAM. ZAP70, как правило, функционирует, распространяя сигнал, но также обеспечивает наличие другого набора ITAM, становящихся фосфорилированными и рекрутирующими больше ZAP70. Это может быть применимо для повышения силы сигнала и может повышать чувствительность, например, к антигенам низкой плотности. В некоторых вариантах осуществления продукт слияния может включать только домен SH2 ZAP70 с ITAM-содержащим эндодоменом (например, продукт слияния не содержит киназный домен ZAP70). В других вариантах осуществления соотношение каталитических доменов (киназных доменов) ZAP70 и ITAM можно варьировать, чтобы повлиять на кинетику активации в ответ на динамику взаимодействий активирующего рецептора с когнатной мишенью.
ITAM является консервативной последовательностью из четырех аминокислот, повторяющихся дважды в цитоплазматических хвостах конкретных белков поверхности клеток иммунной системы. Мотив содержит тирозин, отделенный от лейцина или изолейцина любыми двумя другими аминокислотами, образуя сигнатуру YxxL/I. Две из этих сигнатур, как правило, отделены 6-8 аминокислотами в хвосте молекулы (YxxL/Ix(6-8)YxxL/I).
ITAM важны для передачи сигнала в иммунных клетках. Таким образом, их обнаруживают в хвостах важных сигнальных молекул клетки, таких как CD3 и ζ-цепи комплекса T-клеточного рецептора, CD79-альфа- и -бета-цепи комплекса B-клеточного рецептора и конкретные Fc-рецепторы. Остатки тирозина в этих мотивах становятся фосфорилированными после взаимодействия рецепторных молекул с их лигандами и образуют участки докинга для других белков, вовлеченных в клеточные пути передачи сигнала.
Известно, что несколько белков содержат эндодомены с одним или более мотивами ITAM. Примеры таких белков включают, среди прочего, эпсилон-цепь CD3, гамма-цепь CD3 и дельта-цепь CD3. Мотив ITAM легко может распознаваться в качестве тирозина, отделенного от лейцина или изолейцина любыми двумя другими аминокислотами, образуя сигнатуру YxxL/I. Как правило, но не всегда, два из этих мотивов разделены 6-8 аминокислотами в хвосте молекулы (YxxL/Ix(6-8)YxxL/I). Таким образом, специалист в этой области легко может находить существующие белки, содержащие один или более ITAM для передачи активационного сигнала. Кроме того, учитывая, что мотив является простым, и сложная вторичная структура не нужна, специалист в этой области может конструировать полипептиды, содержащие искусственные ITAM для передачи активационного сигнала (см. WO 2000063372, относящийся к синтетическим сигнальным молекулам).
ITAM-содержащий домен может являться эндодоменом CD3 дзета или содержать его. Соответственно, ITAM-содержащий домен может содержать последовательность, представленную как SEQ ID NO: 13 или ее вариант, имеющий по меньшей мере 80, 85, 90, 95, 98 или 99% идентичности последовательности, сохраняющий способность фосфорилироваться и рекрутировать ZAP70.
SEQ ID NO: 13 (эндодомен CD3 дзета)
RVKFSRSADAPAYQQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPR
В качестве примера, слитый белок может являться последовательностью, представленной как SEQ ID NO: 14, содержащей домен SH2ZAP70, слитый с эндодоменом CD3 дзета, или содержать ее.
SEQ ID NO: 14
MRRVKFSRSADAPAYQQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPRSGGGGSGGGGSGGGGSGGGGSMPDPAAHLPFFYGSISRAEAEEHLKLAGMADGLFLLRQCLRSLGGYVLSLVHDVRFHHFPIERQLNGTYAIAGGKAHCGPAELCEFYSRDPDGLPCNLRKPCNRPSGLEPQPGVFDCLRDAMVRDYVRQTWKLEGEALEQAIISQAPQVEKLIATTAHERMPWYHSSLTREEAERKLYSGAQTDGKFLLRPRKEQGTYALSLIYGKTVYHYLISQDKAGKYCIPEGTKFDTLWQLVEYLKLKADGLIYCLKEACPNSSASNASGAAAPTLPAHPSTLTHPQRRIDTLNSDGYTPEPARITSPDKPRPMPMDTSVYESPYSDPEELKDKKLFLKRDNLLIADIELGCGNFGSVRQGVYRMRKKQIDVAIKVLKQGTEKADTEEMMREAQIMHQLDNPYIVRLIGVCQAEALMLVMEMAGGGPLHKFLVGKREEIPVSNVAELLHQVSMGMKYLEEKNFVHRDLAARNVLLVNRHYAKISDFGLSKALGADDSYYTARSAGKWPLKWYAPECINFRKFSSRSDVWSYGVTMWEALSYGQKPYKKMKGPEVMAFIEQGKRMECPPECPPELYALMSDCWIYKWEDRPDFLTVEQRMRACYYSLASKVEGPPGSTQKAEAACA
Соответственно, слитый белок может содержать последовательность, представленную как SEQ ID NO: 14 или ее вариант, имеющий по меньшей мере 80, 85, 90, 95, 98 или 99% идентичности последовательности.
ITIM-СОДЕРЖАЩИЙ ДОМЕН
В одном из вариантов осуществления слитый белок содержит домен SH2 PTPN6 и домен, содержащий иммунорецепторный тирозиновый ингибирующий мотив (ITIM).
Слияние полноразмерного PTPN6 с ITIM-содержащим доменом приводит к образованию структуры, усиливающей ингибиторный иммунный сигнал. В этом случае слитый белок рекрутируется к иммунорецепторному эндодомену фосфо-ITIM. PTPN6, как правило, функционирует, распространяя сигнал, но также обеспечивает наличие другого набора ITIM, становящихся фосфорилированными и рекрутирующими больше PTPN6. В некоторых вариантах осуществления продукт слияния может включать только домен SH2 PTPN6 с ITIM-содержащим эндодоменом (например, продукт слияния не содержит фосфатазный домен PTPN6). В других вариантах осуществления соотношение каталитических доменов (фосфатазных доменов) PTPN6 с ITIM можно варьировать, чтобы влиять на кинетику активации в ответ на динамику ингибиторных рецепторных взаимодействий с когнатной мишенью.
ITIM является консервативной последовательностью аминокислот (S/I/V/LxYxxI/V/L), обнаруживаемой в цитоплазматических хвостах многих ингибиторных рецепторов иммунной системы. После взаимодействия ITIM-содержащих ингибиторных рецепторов с их лигандом их мотив ITIM фосфорилируется ферментами из киназ Src, что позволяет им рекрутировать PTPN6 посредством взаимодействий между доменом SH2 PTPN6 и фосфорилированными доменами ITIM.
ITIM-содержащие эндодомены включают эндодомены, например, из CD22, LAIR-1, семейства рецепторов подавления цитотоксичности (KIR), LILRB1, CTLA4, PD-1, BTLA.
Эндодомены ITIM из PDCD1, BTLA4, LILRB1, LAIR1, CTLA4, KIR2DL1, KIR2DL4, KIR2DL5, KIR3DL1 и KIR3DL3 представлены как SEQ ID NO: 15-24, соответственно
SEQ ID NO: 15 эндодомен PDCD1
CSRAARGTIGARRTGQPLKEDPSAVPVFSVDYGELDFQWREKTPEPPVPCVPEQTEYATIVFPSGMGTSSPARRGSADGPRSAQPLRPEDGHCSWPL
SEQ ID NO: 16 BTLA4
KLQRRWKRTQSQQGLQENSSGQSFFVRNKKVRRAPLSEGPHSLGCYNPMMEDGISYTTLRFPEMNIPRTGDAESSEMQRPPPDCDDTVTYSALHKRQVGDYENVIPDFPEDEGIHYSELI
QFGVGERPQAQENVDYVILKH
SEQ ID NO: 17 LILRB1
LRHRRQGKHWTSTQRKADFQHPAGAVGPEPTDRGLQWRSSPAADAQEENLYAAVKHTQPEDGVEMDTRSPHDEDPQAVTYAEVKHSRPRREMASPPSPLSGEFLDTKDRQAEEDRQMDTEAAASEAPQDVTYAQLHSLTLRREATEPPPSQEGPSPAVPSIYATLAIH
SEQ ID NO: 18 LAIR1
HRQNQIKQGPPRSKDEEQKPQQRPDLAVDVLERTADKATVNGLPEKDRETDTSALAAGSSQEVTYAQLDHWALTQRTARAVSPQSTKPMAESITYAAVARH
SEQ ID NO: 19 CTLA4
FLLWILAAVSSGLFFYSFLLTAVSLSKMLKKRSPLTTGVYVKMPPTEPECEKQFQPYFIPIN
SEQ ID NO: 20 KIR2DL1
GNSRHLHVLIGTSVVIIPFAILLFFLLHRWCANKKNAVVMDQEPAGNRTVNREDSDEQDPQEVTYTQLNHCVFTQRKITRPSQRPKTPPTDIIVYTELPNAESRSKVVSCP
SEQ ID NO: 21 KIR2DL4
GIARHLHAVIRYSVAIILFTILPFFLLHRWCSKKKENAAVMNQEPAGHRTVNREDSDEQDPQEVTYAQLDHCIFTQRKITGPSQRSKRPSTDTSVCIELPNAEPRALSPAHEHHSQALMGSSRETTALSQTQLASSNVPAAGI
SEQ ID NO: 22 KIR2DL5
TGIRRHLHILIGTSVAIILFIILFFFLLHCCCSNKKNAAVMDQEPAGDRTVNREDSDDQDPQEVTYAQLDHCVFTQTKITSPSQRPKTPPTDTTMYMELPNAKPRSLSPAHKHHSQALRGSSRETTALSQNRVASSHVPAAGI
SEQ ID NO: 23 KIR3DL1
KDPRHLHILIGTSVVIILFILLLFFLLHLWCSNKKNAAVMDQEPAGNRTANSEDSDEQDPEEVTYAQLDHCVFTQRKITRPSQRPKTPPTDTILYTELPNAKPRSKVVSCP
SEQ ID NO: 24 KIR3DL3
KDPGNSRHLHVLIGTSVVIIPFAILLFFLLHRWCANKKNAVVMDQEPAGNRTVNREDSDEQDPQEVTYAQLNHCVFTQRKITRPSQRPKTPPTDTSV
ITIM-содержащий домен может являться эндодоменом PDCD1, BTLA4, LILRB1, LAIR1, CTLA4, KIR2DL1, KIR2DL4, KIR2DL5, KIR3DL1 или KIR3DL3 или содержать его. Соответственно, ITIM-содержащий домен может содержать последовательность, представленную как любая из SEQ ID NO: 15-24 или их вариант, имеющий по меньшей мере 80, 85, 90, 95, 98 или 99% идентичности последовательности, сохраняющий способность быть фосфорилированной киназами Src и усиливать ингибиторный иммунный сигнал.
В качестве примера, слитый белок может являться последовательностью, представленной как SEQ ID NO: 25, содержащей домен SH2 PTPN6, слитый с эндодоменом PD1, или содержать ее.
SEQ ID NO: 25
MTGQPLKEDPSAVPVFSVDYGELDFQWREKTPEPPVPCVPEQTEYATIVFPSGMGTSSPARRGSADGPRSAQPLRPEDGHCSWPLSGGGGSGGGGSGGGGSGGGGSMVRWFHRDLSGLDAETLLKGRGVHGSFLARPSRKNQGDFSLSVRVGDQVTHIRIQNSGDFYDLYGGEKFATLTELVEYYTQQQGVLQDRDGTIIHLKYPLNCSDPTSERWYHGHMSGGQAETLLQAKGEPWTFLVRESLSQPGDFVLSVLSDQPKAGPGSPLRVTHIKVMCEGGRYTVGGLETFDSLTDLVEHFKKTGIEEASGAFVYLRQPYYATRVNAADIENRVLELNKKQESEDTAKAGFWEEFESLQKQEVKNLHQRLEGQRPENKGKNRYKNILPFDHSRVILQGRDSNIPGSDYINANYIKNQLLGPDENAKTYIASQGCLEATVNDFWQMAWQENSRVIVMTTREVEKGRNKCVPYWPEVGMQRAYGPYSVTNCGEHDTTEYKLRTLQVSPLDNGDLIREIWHYQYLSWPDHGVPSEPGGVLSFLDQINQRQESLPHAGPIIVHCSAGIGRTGTIIVIDMLMENISTKGLDCDIDIQKTIQMVRAQRSGMVQTEAQYKFIYVAIAQFIETTKKKLEVLQSQKGQESEYGNITYPPAMKNAHAKASRTSSKHKEDVYENLHTKNKREEKVKKQRSADKEKSKGSLKRK
Соответственно, слитый белок может содержать последовательность, представленную как SEQ ID NO: 25 или ее вариант, имеющий по меньшей мере 80, 85, 90, 95, 98 или 99% идентичности последовательности.
ПЕРЕКРЕСТНЫЙ СИГНАЛ
Настоящее изобретение относится к слитому белку, содержащему: домен SH2 из ITAM-связывающего белка и фосфатазный домен.
Настоящее изобретение также относится к слитому белку, содержащему домен SH2 из ITIM-связывающего белка и киназный домен.
Эти "перекрестные" сигнальные молекулы будут реверсировать возбуждающий или ингибиторный сигнал внутри иммунной клетки, такой как T-клетка. Если T-клетка получает возбуждающий сигнал, например, после распознавания антигена-мишени посредством CAR, или MHC:пептид посредством TCR, наличие первого типа перекрестной молекулы будет приводить к тому, что клетка будет интерпретировать возбуждающий сигнал как ингибиторный сигнал.
Снижение или изменение активации T-клеток может быть применимо во множестве ситуаций, например, его можно использовать для CAR-экспрессирующих T-клеток, в которых есть высокий уровень экспрессии антигена-мишени на клетке-мишени. Его можно использовать для предотвращения гиперактивации T-клеток, что может приводить к истощению T-клеток и/или гибели клеток, индуцируемой активацией. Предотвращая то, чтобы T-клетка становилась активированной слишком сильно или слишком быстро, также можно предотвращать или снижать патологические побочные эффекты лечения CAR-T-клетками, такими как синдром высвобождения цитокинов (CRS).
Обратная ситуация происходит тогда, когда T-клетка получает ингибиторный сигнал, например, после лигирования PD1, и наличие второго типа перекрестной молекулы приводит к тому, что клетка интерпретирует ингибиторный сигнал как возбуждающий сигнал.
Снижение или реверсирование ингибирования T-клеток будет помогать клетке преодолевать ингибиторные стимулы в неблагоприятном опухолевом микроокружении и должно, таким образом, повышать персистирование и экспансию T-клеток in vivo.
В одном из вариантов осуществления слитый белок содержит домен SH2 PTPN6 и киназный домен ZAP70. В другом варианте осуществления слитый белок по настоящему изобретению содержит домен SH2 ZAP70, слитый с киназным доменом PTPN6.
В вариантах осуществления, относящихся к домену SH2 ZAP70, слитому с фосфатазным доменом из PTPN6, когда T-клетка получает возбуждающий сигнал, она интерпретирует его как ингибиторный сигнал, т.к. фосфатазный домен PTPN6 рекрутируется к активированному ITAM посредством домена SH2 ZAP70.
В вариантах осуществления, относящихся к домену SH2 PTPN6, слитому с киназным доменом из ZAP70, когда T-клетка получает ингибиторный сигнал, она интерпретирует его как возбуждающий сигнал, т.к. киназный домен ZAP70 рекрутируется к активированному ITIM посредством домена PTPN6. Слияние между доменом SH2 PTPN6 и киназным доменом ZAP70 будет приводить к конкуренции за фосфорилированные ITIM посредством блокирования ингибиторных сигналов PTPN6 дикого типа, но, кроме того, будет передавать парадоксальный активационный сигнал. Это может быть применимо в преодолении сигналов блокады контрольных точек в опухолевом микроокружении.
Последовательность киназного домена ZAP70 человека, фосфатазного домена PTPN6 и фосфатазного домена SHP-2 представлены как SEQ ID NO: 26, 27 и 28, соответственно.
SEQ ID NO: 26 - киназный домен ZAP70
DPEELKDKKLFLKRDNLLIADIELGCGNFGSVRQGVYRMRKKQIDVAIKVLKQGTEKADTEEMMREAQIMHQLDNPYIVRLIGVCQAEALMLVMEMAGGGPLHKFLVGKREEIPVSNVAELLHQVSMGMKYLEEKNFVHRDLAARNVLLVNRHYAKISDFGLSKALGADDSYYTARSAGKWPLKWYAPECINFRKFSSRSDVWSYGVTMWEALSYGQKPYKKMKGPEVMAFIEQGKRMECPPECPPELYALMSDCWIYKWEDRPDFLTVEQRMRACYYSLASKVEGPPGSTQKAEAACA
SEQ ID NO: 27 - фосфатазный домен PTPN6
FWEEFESLQKQEVKNLHQRLEGQRPENKGKNRYKNILPFDHSRVILQGRDSNIPGSDYINANYIKNQLLGPDENAKTYIASQGCLEATVNDFWQMAWQENSRVIVMTTREVEKGRNKCVPYWPEVGMQRAYGPYSVTNCGEHDTTEYKLRTLQVSPLDNGDLIREIWHYQYLSWPDHGVPSEPGGVLSFLDQINQRQESLPHAGPIIVHCSAGIGRTGTIIVIDMLMENISTKGLDCDIDIQKTIQMVRAQRSGMVQTEAQYKFIYVAIAQFIETTKKKL
SEQ ID NO: 28 - фосфатазный домен SHP-2
WEEFETLQQQECKLLYSRKEGQRQENKNKNRYKNILPFDHTRVVLHDGDPNEPVSDYINANIIMPEFETKCNNSKPKKSYIATQGCLQNTVNDFWRMVFQENSRVIVMTTKEVERGKSKCVKYWPDEYALKEYGVMRVRNVKESAAHDYTLRELKLSKVGQALLQGNTERTVWQYHFRTWPDHGVPSDPGGVLDFLEEVHHKQESIMDAGPVVVHCSAGIGRTGTFIVIDILIDIIREKGVDCDIDVPKTIQMVRSQRSGMVQTEAQYRFIYMA
Киназный домен ZAP70, фосфатазный домен PTPN6 или фосфатазный домен SHP-2 может являться последовательностью, представленной как SEQ ID NO: 26, SEQ ID NO: 27 или SEQ ID NO: 28, соответственно, или ее вариантом, имеющим по меньшей мере 80, 85, 90, 95, 98 или 99% идентичности последовательности, сохраняющим способность фосфорилировать или дефосфорилировать нижележащие белки тем же образом, что и киназный/фосфатазный домены дикого типа, или содержать их.
Примеры слитого белка, содержащего домен SH2 PTPN6, слитый с киназным доменом ZAP70; домен SH2 ZAP70, слитый с киназным доменом PTPN6; и домен SH2 SHP-2, слитый с киназным доменом ZAP70, представлены как SEQ ID NO: 29, SEQ ID NO: 30 и SEQ ID NO: 61, соответственно.
SEQ ID NO: 29 - Продукт слияния домена SH2 PTPN6:киназного домена ZAP70
MVRWFHRDLSGLDAETLLKGRGVHGSFLARPSRKNQGDFSLSVRVGDQVTHIRIQNSGDFYDLYGGEKFATLTELVEYYTQQQGVLQDRDGTIIHLKYPLNCSDPTSERWYHGHMSGGQAETLLQAKGEPWTFLVRESLSQPGDFVLSVLSDQPKAGPGSPLRVTHIKVMCEGGRYTVGGLETFDSLTDLVEHFKKTGIEEASGAFVYLRQPYYSGGGGSDPEELKDKKLFLKRDNLLIADIELGCGNFGSVRQGVYRMRKKQIDVAIKVLKQGTEKADTEEMMREAQIMHQLDNPYIVRLIGVCQAEALMLVMEMAGGGPLHKFLVGKREEIPVSNVAELLHQVSMGMKYLEEKNFVHRDLAARNVLLVNRHYAKISDFGLSKALGADDSYYTARSAGKWPLKWYAPECINFRKFSSRSDVWSYGVTMWEALSYGQKPYKKMKGPEVMAFIEQGKRMECPPECPPELYALMSDCWIYKWEDRPDFLTVEQRMRACYYSLASKVEGPPGSTQKAEAACA
SEQ ID NO: 30 - Продукт слияния домена SH2 ZAP70:фосфатазного домена PTPN6
MPDPAAHLPFFYGSISRAEAEEHLKLAGMADGLFLLRQCLRSLGGYVLSLVHDVRFHHFPIERQLNGTYAIAGGKAHCGPAELCEFYSRDPDGLPCNLRKPCNRPSGLEPQPGVFDCLRDAMVRDYVRQTWKLEGEALEQAIISQAPQVEKLIATTAHERMPWYHSSLTREEAERKLYSGAQTDGKFLLRPRKEQGTYALSLIYGKTVYHYLISQDKAGKYCIPEGTKFDTLWQLVEYLKLKADGLIYCLKEACPNSSASNASGAAAPTLPAHPSTLTHPSGGGGSGGGGSGGGGSGGGGSFWEEFESLQKQEVKNLHQRLEGQRPENKGKNRYKNILPFDHSRVILQGRDSNIPGSDYINANYIKNQLLGPDENAKTYIASQGCLEATVNDFWQMAWQENSRVIVMTTREVEKGRNKCVPYWPEVGMQRAYGPYSVTNCGEHDTTEYKLRTLQVSPLDNGDLIREIWHYQYLSWPDHGVPSEPGGVLSFLDQINQRQESLPHAGPIIVHCSAGIGRTGTIIVIDMLMENISTKGLDCDIDIQKTIQMVRAQRSGMVQTEAQYKFIYVAIAQFIETTKKKL
SEQ ID NO: 61 - двойные домены SH2 из SHP-2, слитые с киназным доменом ZAP70
WFHPNITGVEAENLLLTRGVDGSFLARPSKSNPGDFTLSVRRNGAVTHIKIQNTGDYYDLYGGEKFATLAELVQYYMEHHGQLKEKNGDVIELKYPLNCADPTSERWFHGHLSGKEAEKLLTEKGKHGSFLVRESQSHPGDFVLSVRTGDDKGESNDGKSKVTHVMIRCQELKYDVGGGERFDSLTDLVEHYKKNPMVETLGTVLQLKQPLNTTRINPNSSASNASGAAAPTLPAHPSTLTHPQRRIDTLNSDGYTPEPARITSPDKPRPMPMDTSVYESPYSDPEELKDKKLFLKRDNLLIADIELGCGNFGSVRQGVYRMRKKQIDVAIKVLKQGTEKADTEEMMREAQIMHQLDNPYIVRLIGVCQAEALMLVMEMAGGGPLHKFLVGKREEIPVSNVAELLHQVSMGMKYLEEKNFVHRDLAARNVLLVNRHYAKISDFGLSKALGADDSYYTARSAGKWPLKWYAPECINFRKFSSRSDVWSYGVTMWEALSYGQKPYKKMKGPEVMAFIEQGKRMECPPECPPELYALMSDCWIYKWEDRPDFLTVEQRMRACYYSL
Слитый белок может являться последовательностью, представленной как SEQ ID NO: 29, SEQ ID NO: 30 или SEQ ID NO: 61 или ее вариантом, имеющим по меньшей мере 80, 85, 90, 95, 98 или 99% идентичности последовательности, или содержать их.
ГЕТЕРОЛОГИЧНЫЙ СИГНАЛЬНЫЙ ДОМЕН
Слитый белок по настоящему изобретению может содержать (i) домен SH2 ZAP70, PTPN6 или SHP-2; и (ii) гетерологичный сигнальный домен.
В рамках изобретения термин "гетерологичный сигнальный домен" относится к сигнальному домену, не присутствующем в белке ZAP70, PTPN6 или SHP-2 дикого типа. В связи с этим, если слитый белок содержит домен SH2 ZAP70, он содержит сигнальный домен, не являющийся киназным доменом ZAP70. Альтернативно, если слитый белок содержит домен SH2 PTPN6, он содержит сигнальный домен, не являющийся фосфатазным доменом PTPN6.
ОБХОДНОЙ СИГНАЛ
Гетерологичный сигнальный домен можно получать из сигнальной молекулы, как правило, не активированной ITAM-содержащим рецептором. Другими словами, гетерологичный сигнальный домен можно получать из сигнальной молекулы, не участвующей в распространении иммунологического сигнала 1 после связывания антигена с TCR. Иммунологического сигнала 1 достаточно для запуска уничтожения T-клетками когнатных клеток-мишеней, но он не активирует T-клетку полностью для пролиферации и выживания.
В одном из вариантов осуществления этого аспекта изобретения настоящее изобретение относится к слитому белку, содержащему (i) домен SH2 из белка, связывающегося с ITAM, и (ii) гетерологичный сигнальный домен.
Продукт слияния между, например, ZAP70 и другой сигнальной молекулой, как правило, не активированной ITAM-содержащим рецептором, может действовать, приводя к переходу сигнала с одного пути на другой. Одним из примеров является костимуляция. Продукт слияния между ZAP70 и эндодоменом CD28 может передавать костимуляторный сигнал CD28, а также ITAM-активирующий сигнал. Аналогично, продукт слияния между ZAP70 и эндодоменом 41BB или OX40 может передавать костимуляторный сигнал 41BB или OX40. Также могут рекрутироваться другие пути, например, продукт слияния между ZAP70 и киназным доменом AKT может приводить к передаче сигнала AKT после фосфорилирования ITAM. Другие примеры могут включать киназный домен из JAK. Таким образом, T-клетка может интерпретировать простой сигнал распознавания антигена как передачу костимуляторного сигнала или даже сигнала цитокинового типа.
Такие слитые белки могут быть применимы, например, в подходах, когда повторные стимуляции T-клеток ex vivo могут приводить к образованию популяций, в которых отсутствуют костимуляторные поверхностные антигены, и имеющих ограниченную пролиферативную способность in vivo, приводящую к ограниченному персистированию и эффективности. Утрата костимуляторных поверхностных антигенов, приводящая к активации T-клеток исключительно через TCR, связана с более высокой степенью индуцируемой активацией гибели клеток, которая будет отрицательно влиять на эффективность и персистирование in vivo. Эффект можно реверсировать посредством активации экспрессируемых на поверхности 4-1BB и OX40, что свидетельствует о том, что костимуляция может предотвращать индуцируемую активацией гибель клеток и может поддерживать более сильную экспансию опухолеспецифических T-клеток.
В другом варианте осуществления этого аспекта изобретения настоящее изобретение относится к слитому белку, содержащему (i) домен SH2 из белка, связывающегося с ITIM; и (ii) гетерологичный сигнальный домен.
Например, домен SH2 PTPN6 или домен SH2 SHP-2 можно подвергать слиянию с костимуляторным эндодоменом таким образом, что T-клетка интерпретирует ингибиторный сигнал как костимуляторный сигнал.
Гетерологичный сигнальный домен можно получать, например, из CD28, 41BB или OX40.
CD28 обеспечивает мощный костимуляторный сигнал, а именно иммунологический сигнал 2, запускающий пролиферацию T-клеток. CD28 является рецептором для белков CD80 (B7.1) и CD86 (B7.2).
41BB (CD137) является трансмембранным гликопротеином типа 2, принадлежащим к суперсемейству ФНО, экспрессируемым на активированных T-клетках. Перекрестная сшивка 41BB повышает пролиферацию T-клеток, секрецию ИЛ-2, выживание и цитолитическую активность.
OX40 (CD134) является вторичной костимуляторной молекулой, экспрессируемой через 24-72 часа после активации; его лиганд OX40L также не экспрессируется на покоящихся антигенпрезентирующих клетках, но экспрессируется после их активации. Экспрессия OX40 зависит от полной активации T-клетки; без CD28 экспрессия OX40 замедляется и находится на уровне в четыре раза ниже. Передача сигнала через OX40 необходима для пролонгированного выживания T-клеток после исходной активации и пролиферации.
Сигнальные домены (эндодомены) CD28, 41BB и OX40 представлены как SEQ ID NO: 31, 32 и 33, соответственно.
SEQ ID NO: 31 - эндодомен CD28
MRSKRSRLLHSDYMNMTPRRPGPTRKHYQPYAPPRDFAAYRS
SEQ ID NO: 32 - эндодомен 41BB
MKRGRKKLLYIFKQPFMRPVQTTQEEDGCSCRFPEEEEGGCEL
SEQ ID NO: 33 - эндодомен OX40
MRDQRLPPDAHKPPGGGSFRTPIQEEQADAHSTLAKI
Гетерологичный сигнальный домен может являться последовательностью, представленной как SEQ ID NO: 31; SEQ ID NO: 32 или SEQ ID NO: 33, соответственно, или ее вариантом, имеющим по меньшей мере 80, 85, 90, 95, 98 или 99% идентичности последовательности, или содержать их.
Гетерологичный сигнальный домен может являться ингибиторным сигнальным доменом или содержать его.
Например, ингибиторный сигнальный домен может содержать эндодомен CD148 или CD45. Показано, что CD148 и CD45, как правило, действуют на фосфорилированные тирозины выше в каскаде передачи сигнала TCR.
CD148 является рецептор-подобной белковой тирозинфосфатазой, отрицательно регулирующей передачу сигнала TCR, противодействуя фосфорилированию и функционированию PLCγ1 и LAT.
CD45 присутствует на всех гемопоэтических клетках, является белковой тирозинфосфатазой, способной регулировать передачу сигнала и функциональные ответы, снова, посредством фосфорилирования PLC γ1.
Ингибиторный сигнальный домен может содержать всю рецептор-подобную тирозинфосфатазу или ее часть. Фосфатаза может противодействовать фосфорилированию и/или функционированию элементов, участвующих в передаче T-клеточного сигнала, таких как PLCγ1 и/или LAT.
Ингибиторный сигнальный домен может являться эндодоменом ICOS, CD27, BTLA, CD30, GITR или HVEM или содержать его.
Ингибиторный сигнальный домен может содержать последовательность, представленную как SEQ ID NO: 34-39 или их вариант, имеющий по меньшей мере 80% идентичности последовательности.
SEQ ID NO: 34 - эндодомен ICOS
CWLTKKKYSSSVHDPNGEYMFMRAVNTAKKSRLTDVTL
SEQ ID NO: 35 - эндодомен CD27
QRRKYRSNKGESPVEPAEPCHYSCPREEEGSTIPIQEDYRKPEPACSP
SEQ ID NO: 36 - эндодомен BTLA
RRHQGKQNELSDTAGREINLVDAHLKSEQTEASTRQNSQVLLSETGIYDNDPDLCFRMQEGSEVYSNPCLEENKPGIVYASLNHSVIGPNSRLARNVKEAPTEYASICVRS
SEQ ID NO: 37 - эндодомен CD30
HRRACRKRIRQKLHLCYPVQTSQPKLELVDSRPRRSSTQLRSGASVTEPVAEERGLMSQPLMETCHSVGAAYLESLPLQDASPAGGPSSPRDLPEPRVSTEHTNNKIEKIYIMKADTVIVGTVKAELPEGRGLAGPAEPELEEELEADHTPHYPEQETEPPLGSCSDVMLSVEEEGKEDPLPTAASGK
SEQ ID NO: 38 - эндодомен GITR
QLGLHIWQLRSQCMWPRETQLLLEVPPSTEDARSCQFPEEERGERSAEEKGRLGDLWV
SEQ ID NO: 39 - эндодомен HVEM
CVKRRKPRGDVVKVIVSVQRKRQEAEGEATVIEALQAPPDVTTVAVEETIPSFTGRSPNH
Вариант последовательности может иметь по меньшей мере 80%, 85%, 90%, 95%, 98% или 99% идентичности последовательности по отношению к SEQ ID NO: 34-39, при условии, что последовательность обеспечивает эффективный внутриклеточный сигнальный домен.
Соответственно, слитый белок может являться или содержать любую из последовательностей, представленных как SEQ ID NO: 40-45.
(SEQ ID NO: 40) - эндодомен CD28, слитый с амино-концом полноразмерного ZAP
MRSKRSRLLHSDYMNMTPRRPGPTRKHYQPYAPPRDFAAYRSSGGGGSGGGGSGGGGSGGGGSMPDPAAHLPFFYGSISRAEAEEHLKLAGMADGLFLLRQCLRSLGGYVLSLVHDVRFHHFPIERQLNGTYAIAGGKAHCGPAELCEFYSRDPDGLPCNLRKPCNRPSGLEPQPGVFDCLRDAMVRDYVRQTWKLEGEALEQAIISQAPQVEKLIATTAHERMPWYHSSLTREEAERKLYSGAQTDGKFLLRPRKEQGTYALSLIYGKTVYHYLISQDKAGKYCIPEGTKFDTLWQLVEYLKLKADGLIYCLKEACPNSSASNASGAAAPTLPAHPSTLTHPQRRIDTLNSDGYTPEPARITSPDKPRPMPMDTSVYESPYSDPEELKDKKLFLKRDNLLIADIELGCGNFGSVRQGVYRMRKKQIDVAIKVLKQGTEKADTEEMMREAQIMHQLDNPYIVRLIGVCQAEALMLVMEMAGGGPLHKFLVGKREEIPVSNVAELLHQVSMGMKYLEEKNFVHRDLAARNVLLVNRHYAKISDFGLSKALGADDSYYTARSAGKWPLKWYAPECINFRKFSSRSDVWSYGVTMWEALSYGQKPYKKMKGPEVMAFIEQGKRMECPPECPPELYALMSDCWIYKWEDRPDFLTVEQRMRACYYSLASKVEGPPGSTQKAEAACA
(SEQ ID NO: 41) - эндодомен 41BB, слитый с амино-концом полноразмерного ZAP
MKRGRKKLLYIFKQPFMRPVQTTQEEDGCSCRFPEEEEGGCELSGGGGSGGGGSGGGGSGGGGSMPDPAAHLPFFYGSISRAEAEEHLKLAGMADGLFLLRQCLRSLGGYVLSLVHDVRFHHFPIERQLNGTYAIAGGKAHCGPAELCEFYSRDPDGLPCNLRKPCNRPSGLEPQPGVFDCLRDAMVRDYVRQTWKLEGEALEQAIISQAPQVEKLIATTAHERMPWYHSSLTREEAERKLYSGAQTDGKFLLRPRKEQGTYALSLIYGKTVYHYLISQDKAGKYCIPEGTKFDTLWQLVEYLKLKADGLIYCLKEACPNSSASNASGAAAPTLPAHPSTLTHPQRRIDTLNSDGYTPEPARITSPDKPRPMPMDTSVYESPYSDPEELKDKKLFLKRDNLLIADIELGCGNFGSVRQGVYRMRKKQIDVAIKVLKQGTEKADTEEMMREAQIMHQLDNPYIVRLIGVCQAEALMLVMEMAGGGPLHKFLVGKREEIPVSNVAELLHQVSMGMKYLEEKNFVHRDLAARNVLLVNRHYAKISDFGLSKALGADDSYYTARSAGKWPLKWYAPECINFRKFSSRSDVWSYGVTMWEALSYGQKPYKKMKGPEVMAFIEQGKRMECPPECPPELYALMSDCWIYKWEDRPDFLTVEQRMRACYYSLASKVEGPPGSTQKAEAACA
(SEQ ID NO: 42) - эндодомен OX40, слитый с амино-концом полноразмерного ZAP
MRDQRLPPDAHKPPGGGSFRTPIQEEQADAHSTLAKISGGGGSGGGGSGGGGSGGGGSMPDPAAHLPFFYGSISRAEAEEHLKLAGMADGLFLLRQCLRSLGGYVLSLVHDVRFHHFPIERQLNGTYAIAGGKAHCGPAELCEFYSRDPDGLPCNLRKPCNRPSGLEPQPGVFDCLRDAMVRDYVRQTWKLEGEALEQAIISQAPQVEKLIATTAHERMPWYHSSLTREEAERKLYSGAQTDGKFLLRPRKEQGTYALSLIYGKTVYHYLISQDKAGKYCIPEGTKFDTLWQLVEYLKLKADGLIYCLKEACPNSSASNASGAAAPTLPAHPSTLTHPQRRIDTLNSDGYTPEPARITSPDKPRPMPMDTSVYESPYSDPEELKDKKLFLKRDNLLIADIELGCGNFGSVRQGVYRMRKKQIDVAIKVLKQGTEKADTEEMMREAQIMHQLDNPYIVRLIGVCQAEALMLVMEMAGGGPLHKFLVGKREEIPVSNVAELLHQVSMGMKYLEEKNFVHRDLAARNVLLVNRHYAKISDFGLSKALGADDSYYTARSAGKWPLKWYAPECINFRKFSSRSDVWSYGVTMWEALSYGQKPYKKMKGPEVMAFIEQGKRMECPPECPPELYALMSDCWIYKWEDRPDFLTVEQRMRACYYSLASKVEGPPGSTQKAEAACA
(SEQ ID NO: 43) - эндодомен CD28, слитый с амино-концом домена SH2 PTPN6.
MRSKRSRLLHSDYMNMTPRRPGPTRKHYQPYAPPRDFAAYRSSGGGGSGGGGSGGGGSGGGGSMVRWFHRDLSGLDAETLLKGRGVHGSFLARPSRKNQGDFSLSVRVGDQVTHIRIQNSGDFYDLYGGEKFATLTELVEYYTQQQGVLQDRDGTIIHLKYPLNCSDPTSERWYHGHMSGGQAETLLQAKGEPWTFLVRESLSQPGDFVLSVLSDQPKAGPGSPLRVTHIKVMCEGGRYTVGGLETFDSLTDLVEHFKKTGIEEASGAFVYLRQPY
(SEQ ID NO: 44) - эндодомен 41BB, слитый с амино-концом домена SH2 PTPN6
MKRGRKKLLYIFKQPFMRPVQTTQEEDGCSCRFPEEEEGGCELSGGGGSGGGGSGGGGSGGGGSMVRWFHRDLSGLDAETLLKGRGVHGSFLARPSRKNQGDFSLSVRVGDQVTHIRIQNSGDFYDLYGGEKFATLTELVEYYTQQQGVLQDRDGTIIHLKYPLNCSDPTSERWYHGHMSGGQAETLLQAKGEPWTFLVRESLSQPGDFVLSVLSDQPKAGPGSPLRVTHIKVMCEGGRYTVGGLETFDSLTDLVEHFKKTGIEEASGAFVYLRQPY
(SEQ ID NO: 45) - эндодомен OX40, слитый с амино-концом домена SH2 PTPN6
MRDQRLPPDAHKPPGGGSFRTPIQEEQADAHSTLAKISGGGGSGGGGSGGGGSGGGGSMVRWFHRDLSGLDAETLLKGRGVHGSFLARPSRKNQGDFSLSVRVGDQVTHIRIQNSGDFYDLYGGEKFATLTELVEYYTQQQGVLQDRDGTIIHLKYPLNCSDPTSERWYHGHMSGGQAETLLQAKGEPWTFLVRESLSQPGDFVLSVLSDQPKAGPGSPLRVTHIKVMCEGGRYTVGGLETFDSLTDLVEHFKKTGIEEASGAFVYLRQPY
Соответственно, слитый белок может содержать последовательность, представленную как любая из SEQ ID NO: 40-45 или их вариант, имеющий по меньшей мере 80, 85, 90, 95, 98 или 99% идентичности последовательности.
КИНАЗНЫЙ ДОМЕН
Гетерологичный сигнальный домен может являться киназным доменом. Например, гетерологичный сигнальный домен может содержать киназный домен AKT или киназный домен JAK.
Akt, также известный как протеинкиназа B (PKB), является серин/треонин-специфической протеинкиназой, играющей ключевую роль во множестве клеточных процессов, таких как метаболизм глюкозы, апоптоз, пролиферация клеток, транскрипция и миграция клеток.
После активации TCR T-клетки секретируют ИЛ-2, поддерживающий выживание и пролиферацию. Однако эта секреция является транзиторной, и T-клетки, активированные и подвергнутые экспансии in vitro, начинают зависеть от экзогенного ИЛ-2 для выживания. Повышая фосфорилирование AKT после фосфорилирования ITAM, ассоциированного с активацией TCR или CAR, можно снижать или устранять зависимость активированных T-клеток от экзогенного ИЛ-2 и повышать их пролиферацию и выживание.
Киназный домен Akt представлен как SEQ ID NO: 46.
SEQ ID NO: 46 - киназный домен Akt
AEEMEVSLAKPKHRVTMNEFEYLKLLGKGTFGKVILVKEKATGRYYAMKILKKEVIVAKDEVAHTLTENRVLQNSRHPFLTALKYSFQTHDRLCFVMEYANGGELFFHLSRERVFSEDRARFYGAEIVSALDYLHSEKNVVYRDLKLENLMLDKDGHIKITDFGLCKEGIKDGATMKTFCGTPEYLAPEVLEDNDYGRAVDWWGLGVVMYEMMCGRLPFYNQDHEKLFELILMEEIRFPRTLGPEAKSLLSGLLKKDPKQRLGGGSEDAKEIMQHRFFAGIVWQHVYEKKLSPPFKPQVTSETDTRYFDEEFTAQMITITPPDQDDSMECVDSERRPHFPQFSYSASGTA
Гетерологичный сигнальный домен может являться последовательностью, представленной как SEQ ID NO: 46 или ее вариант, имеющий по меньшей мере, 80, 85, 90, 95, 98 или 99% идентичности последовательности, или содержать ее, при условии, что последовательность обеспечивает эффективный киназный домен.
В качестве примера, слитый белок может являться любой из последовательностей, представленных как SEQ ID NO: 47-49 и 62, содержащую домен SH2 ZAP70, слитый непосредственно с киназным доменом Akt, домен SH2 ZAP70, слитый с киназным доменом Akt через линкер, ZAP70, подвергнутый мутагенезу для придания нефункциональности и слитый с киназным доменом Akt; и двойной домен SH2 SHP-2, слитый с киназным доменом Akt, соответственно, или содержать их.
SEQ ID NO: 47 - домен SH2 ZAP70, слитый напрямую с киназным доменом Akt
MPDPAAHLPFFYGSISRAEAEEHLKLAGMADGLFLLRQCLRSLGGYVLSLVHDVRFHHFPIERQLNGTYAIAGGKAHCGPAELCEFYSRDPDGLPCNLRKPCNRPSGLEPQPGVFDCLRDAMVRDYVRQTWKLEGEALEQAIISQAPQVEKLIATTAHERMPWYHSSLTREEAERKLYSGAQTDGKFLLRPRKEQGTYALSLIYGKTVYHYLISQDKAGKYCIPEGTKFDTLWQLVEYLKLKADGLIYCLKEACPNSSASNASGAAAPTLPAHPSTLTHPAEEMEVSLAKPKHRVTMNEFEYLKLLGKGTFGKVILVKEKATGRYYAMKILKKEVIVAKDEVAHTLTENRVLQNSRHPFLTALKYSFQTHDRLCFVMEYANGGELFFHLSRERVFSEDRARFYGAEIVSALDYLHSEKNVVYRDLKLENLMLDKDGHIKITDFGLCKEGIKDGATMKTFCGTPEYLAPEVLEDNDYGRAVDWWGLGVVMYEMMCGRLPFYNQDHEKLFELILMEEIRFPRTLGPEAKSLLSGLLKKDPKQRLGGGSEDAKEIMQHRFFAGIVWQHVYEKKLSPPFKPQVTSETDTRYFDEEFTAQMITITPPDQDDSMECVDSERRPHFPQFSYSASGTA
SEQ ID NO: 48 - домен SH2 ZAP70, слитый с киназным доменом Akt через линкер
MPDPAAHLPFFYGSISRAEAEEHLKLAGMADGLFLLRQCLRSLGGYVLSLVHDVRFHHFPIERQLNGTYAIAGGKAHCGPAELCEFYSRDPDGLPCNLRKPCNRPSGLEPQPGVFDCLRDAMVRDYVRQTWKLEGEALEQAIISQAPQVEKLIATTAHERMPWYHSSLTREEAERKLYSGAQTDGKFLLRPRKEQGTYALSLIYGKTVYHYLISQDKAGKYCIPEGTKFDTLWQLVEYLKLKADGLIYCLKEACPNSSASNASGAAAPTLPAHPSTLTHPSGGGGSGGGGSGGGGSGGGGSAEEMEVSLAKPKHRVTMNEFEYLKLLGKGTFGKVILVKEKATGRYYAMKILKKEVIVAKDEVAHTLTENRVLQNSRHPFLTALKYSFQTHDRLCFVMEYANGGELFFHLSRERVFSEDRARFYGAEIVSALDYLHSEKNVVYRDLKLENLMLDKDGHIKITDFGLCKEGIKDGATMKTFCGTPEYLAPEVLEDNDYGRAVDWWGLGVVMYEMMCGRLPFYNQDHEKLFELILMEEIRFPRTLGPEAKSLLSGLLKKDPKQRLGGGSEDAKEIMQHRFFAGIVWQHVYEKKLSPPFKPQVTSETDTRYFDEEFTAQMITITPPDQDDSMECVDSERRPHFPQFSYSASGTA
SEQ ID NO: 49 - ZAP70, подвергнутый мутагенезу для придания нефункциональности и слитый с киназным доменом Akt
MPDPAAHLPFFYGSISRAEAEEHLKLAGMADGLFLLRQCLRSLGGYVLSLVHDVRFHHFPIERQLNGTYAIAGGKAHCGPAELCEFYSRDPDGLPCNLRKPCNRPSGLEPQPGVFDCLRDAMVRDYVRQTWKLEGEALEQAIISQAPQVEKLIATTAHERMPWYHSSLTREEAERKLYSGAQTDGKFLLKPRKEQGTYALSLIYGKTVYHYLISQDKAGKYCIPEGTKFDTLWQLVEYLKLKADGLIYCLKEACPNSSASNASGAAAPTLPAHPSTLTHPAEEMEVSLAKPKHRVTMNEFEYLKLLGKGTFGKVILVKEKATGRYYAMKILKKEVIVAKDEVAHTLTENRVLQNSRHPFLTALKYSFQTHDRLCFVMEYANGGELFFHLSRERVFSEDRARFYGAEIVSALDYLHSEKNVVYRDLKLENLMLDKDGHIKITDFGLCKEGIKDGATMKTFCGTPEYLAPEVLEDNDYGRAVDWWGLGVVMYEMMCGRLPFYNQDHEKLFELILMEEIRFPRTLGPEAKSLLSGLLKKDPKQRLGGGSEDAKEIMQHRFFAGIVWQHVYEKKLSPPFKPQVTSETDTRYFDEEFTAQMITITPPDQDDSMECVDSERRPHFPQFSYSASGTA
SEQ ID NO: 62 - двойной домен SH2 SHP-2, слитый с киназным доменом Akt
WFHPNITGVEAENLLLTRGVDGSFLARPSKSNPGDFTLSVRRNGAVTHIKIQNTGDYYDLYGGEKFATLAELVQYYMEHHGQLKEKNGDVIELKYPLNCADPTSERWFHGHLSGKEAEKLLTEKGKHGSFLVRESQSHPGDFVLSVRTGDDKGESNDGKSKVTHVMIRCQELKYDVGGGERFDSLTDLVEHYKKNPMVETLGTVLQLKQPLNTTRINAEEMEVSLAKPKHRVTMNEFEYLKLLGKGTFGKVILVKEKATGRYYAMKILKKEVIVAKDEVAHTLTENRVLQNSRHPFLTALKYSFQTHDRLCFVMEYANGGELFFHLSRERVFSEDRARFYGAEIVSALDYLHSEKNVVYRDLKLENLMLDKDGHIKITDFGLCKEGIKDGATMKTFCGTPEYLAPEVLEDNDYGRAVDWWGLGVVMYEMMCGRLPFYNQDHEKLFELILMEEIRFPRTLGPEAKSLLSGLLKKDPKQRLGGGSEDAKEIMQHRFFAGIVWQHVYEKKLSPPFKPQVTSETDTRYFDEEFTAQMITITPPDQDDSMECVDSERRPHFPQFSYSASGTA
Слитый белок может содержать последовательность, представленную как любая из SEQ ID NO: 47-49 или 62 или ее вариант, имеющий по меньшей мере 80, 85, 90, 95, 98 или 99% идентичности последовательности.
Киназа Janus (JAK) относится к семейству внутриклеточных, нерецепторных тирозинкиназ, передающих цитокин-опосредованные сигналы через путь JAK-STAT. Четырьмя членами семейства JAK являются: киназа Janus 1 (JAK1); киназа Janus 2 (JAK2); киназа Janus 3 (JAK3) и тирозинкиназа 2 (TYK2).
SEQ ID NO: 50 - Киназа, содержащая домен JAK2
RNEDLIFNESLGQGTFTKIFKGVRREVGDYGQLHETEVLLKVLDKAHRNYSESFFEAASMMSKLSHKHLVLNYGVCVCGDENILVQEFVKFGSLDTYLKKNKNCINILWKLEVAKQLAWAMHFLEENTLIHGNVCAKNILLIREEDRKTGNPPFIKLSDPGISITVLPKDILQERIPWVPPECIENPKNLNLATDKWSFGTTLWEICSGGDKPLSALDSQRKLQFYEDRHQLPAPKWAELANLINNCMDYEPDFRPSFRAIIRDLNSLFTPDYELLTENDMLPNMRIGALGFSGAFEDRDPTQFEERHLKFLQQLGKGNFGSVEMCRYDPLQDNTGEVVAVKKLQHSTEEHLRDFEREIEILKSLQHDNIVKYKGVCYSAGRRNLKLIMEYLPYGSLRDYLQKHKERIDHIKLLQYTSQICKGMEYLGTKRYIHRDLATRNILVENENRVKIGDFGLTKVLPQDKEYYKVKEPGESPIFWYAPESLTESKFSVASDVWSFGVVLYELFTYIEKSKSPPAEFMRMIGNDKQGQMIVFHLIELLKNNGRLPRPDGCPDEIYMIMTECWNNNVNQRPSFRDLALRVDQIRDNM
ПРОТЕАЗНЫЙ ДОМЕН
Настоящее изобретение также относится к слитому белку, содержащему (i) домен SH2 из белка, связывающегося с ITAM- или ITIM-содержащим белком, и (ii) протеазный домен.
Протеазный домен может представлять собой любую протеазу, способную расщеплять по специфической последовательности распознавания. В связи с этим, протеазный домен может представлять собой любую протеазу, делающую возможной расщепление одного полипептида-мишени на два разных полипептида посредством расщепления по специфической последовательности-мишени.
Протеазный домен может являться протеазным доменом вируса гравировки табака (TeV).
Протеаза TeV является высокоспецифичной в отношении последовательности цистеиновой протеазой, являющейся химотрипсин-подобной протеазой. Она очень специфична для своего целевого участка расщепления, и, таким образом, ее часто используют для контролируемого расщепления слитых белков in vitro и in vivo. Консенсусным участком расщепления TeV является ENLYFQ\S (где "'\'" означает расщепляемую пептидную связь). Клетки млекопитающих, такие как клетки человека, не экспрессируют эндогенную протеазу TeV.
Таким образом, участок расщепления TeV представлен как SEQ ID NO: 51.
SEQ ID NO: 51 - участок расщепления Tev
ENLYFQS
Протеазный домен TeV представлен как SEQ ID NO: 52.
SEQ ID NO: 52
SLFKGPRDYNPISSTICHLTNESDGHTTSLYGIGFGPFIITNKHLFRRNNGTLLVQSLHGVFKVKNTTTLQQHLIDGRDMIIIRMPKDFPPFPQKLKFREPQREERICLVTTNFQTKSMSSMVSDTSCTFPSSDGIFWKHWIQTKDGQCGSPLVSTRDGFIVGIHSASNFTNTNNYFTSVPKNFMELLTNQEAQQWVSGWRLNADSVLWGGHKVFMSKPEEPFQPVKEATQLMNELVYSQ
Таким образом, протеазный домен может являться последовательностью, представленной как SEQ ID NO: 52 или ее вариант, имеющий по меньшей мере 80, 85, 90, 95, 98 или 99% идентичности последовательности, или содержать ее, при условии, что последовательность обеспечивает эффективную протеазную функцию.
В качестве примера, слитый белок может являться последовательностью, представленной как SEQ ID NO: 53 или 54, содержащую домен SH2 ZAP70, слитый с последовательностью протеазы TEV, или домен SH2 PTPN6, слитый с последовательностью протеазы TEV; соответственно, или содержать ее.
SEQ ID NO: 53
MPDPAAHLPFFYGSISRAEAEEHLKLAGMADGLFLLRQCLRSLGGYVLSLVHDVRFHHFPIERQLNGTYAIAGGKAHCGPAELCEFYSRDPDGLPCNLRKPCNRPSGLEPQPGVFDCLRDAMVRDYVRQTWKLEGEALEQAIISQAPQVEKLIATTAHERMPWYHSSLTREEAERKLYSGAQTDGKFLLRPRKEQGTYALSLIYGKTVYHYLISQDKAGKYCIPEGTKFDTLWQLVEYLKLKADGLIYCLKEACPNSSASNASGAAAPTLPAHPSTLTHPSGGGGSGGGGSGGGGSGGGGSSLFKGPRDYNPISSTICHLTNESDGHTTSLYGIGFGPFIITNKHLFRRNNGTLLVQSLHGVFKVKNTTTLQQHLIDGRDMIIIRMPKDFPPFPQKLKFREPQREERICLVTTNFQTKSMSSMVSDTSCTFPSSDGIFWKHWIQTKDGQCGSPLVSTRDGFIVGIHSASNFTNTNNYFTSVPKNFMELLTNQEAQQWVSGWRLNADSVLWGGHKVFMSKPEEPFQPVKEATQLMNELVYSQ
SEQ ID NO: 54
MVRWFHRDLSGLDAETLLKGRGVHGSFLARPSRKNQGDFSLSVRVGDQVTHIRIQNSGDFYDLYGGEKFATLTELVEYYTQQQGVLQDRDGTIIHLKYPLNCSDPTSERWYHGHMSGGQAETLLQAKGEPWTFLVRESLSQPGDFVLSVLSDQPKAGPGSPLRVTHIKVMCEGGRYTVGGLETFDSLTDLVEHFKKTGIEEASGAFVYLRQPYYSGGGGSSLFKGPRDYNPISSTICHLTNESDGHTTSLYGIGFGPFIITNKHLFRRNNGTLLVQSLHGVFKVKNTTTLQQHLIDGRDMIIIRMPKDFPPFPQKLKFREPQREERICLVTTNFQTKSMSSMVSDTSCTFPSSDGIFWKHWIQTKDGQCGSPLVSTRDGFIVGIHSASNFTNTNNYFTSVPKNFMELLTNQEAQQWVSGWRLNADSVLWGGHKVFMSKPEEPFQPVKEATQLMNELVYSQ
Слитый белок может содержать последовательность, представленную как SEQ ID NO: 53 или 54 или ее вариант, имеющий по меньшей мере 80, 85, 90, 95, 98 или 99% идентичности последовательности.
Домен SH2 и гетерологичный домен слитого белка можно разделять линкером для пространственного разделения домена SH2 и гетерологичного домена.
ТРАНСКРИПЦИОННЫЙ СИГНАЛ
Слитый белок, содержащий протеазу, как описано в предыдущем разделе, может коэкспрессироваться в клетке с мембраносвязанным белком, имеющим участок расщепления протеазой. Расщепление мембраносвязанного белка по участку расщепления протеазой будет высвобождать дистальную по отношению к мембране часть белка.
Мембраносвязанный белок может являться, например, мембраносвязанным фактором транскрипции. Когда происходит расщепление, фактор транскрипции высвобождается от своей связи и свободен для транспорта в ядро.
Слияние между SH2 ZAP70 или доменом SH2 PTPN6 и протеазным доменом будет приводить к проксимальному в отношении мембраны рекрутированию протеазы после фосфорилирования ITAM или ITIM, соответственно.
Фосфорилирование доменов ITAM или ITIM приводит к рекрутированию SH2 ZAP70 или SH2 PTPN6, слитого с протеазным доменом, соответственно, в проксимальной по отношению к мембране области. Это приводит к отщеплению фактора транскрипции от его связи и транспорту в ядро. Это может иметь различное применение: например, после активации T-клетку можно программировать для экспрессии факторов транскрипции, действующих, предотвращая дифференцировку T-клетки. Например, после активации T-клетку можно программировать для экспрессии цитокина, такого как ИЛ-2, ИЛ-7 или ИЛ-15, который может стимулировать пролиферацию или выживание T-клетки, или ИЛ-12, который может преобразовывать неблагоприятное опухолевое микроокружение в то, которое будет более благоприятно для иммунного отторжения опухоли.
В частности, настоящее изобретение относится к клетке, коэкспрессирующей:
(i) слитый белок, содержащий домен SH2 из белка, связывающегося с фосфорилированным ITAM; и
(ii) мембраносвязанный фактор транскрипции
где фактор транскрипции при высвобождении из мембранной связи повышает экспрессию ИЛ-2, ИЛ-7 и/или ИЛ-15 в клетке.
Настоящее изобретение также относится к клетке, коэкспрессирующей:
(i) слитый белок, содержащий домен SH2 из белка, связывающегося с фосфорилированным ITIM; и
(ii) мембраносвязанный фактор транскрипции
где фактор транскрипции при высвобождении из мембранной связи повышает экспрессию ИЛ-12 в клетке.
Участок распознавания протеазой
Участок распознавания протеазой может являться любой аминокислотной последовательностью, позволяющей протеазному домену слитого белка специфически расщеплять мембраносвязанный фактор транскрипции между мембранной связью и фактором транскрипции. Например, в одном из вариантов осуществления протеазный домен является протеазным доменом TeV, и участок распознавания протеазой является участком распознавания протеазой TeV.
Мембранная связь
Мембранная связь может являться любой последовательностью, сигналом или доменом, способным локализовать фактор транскрипции и участок распознавания протеазой проксимально к мембране. Например, мембранная связь может являться сигналом миристоилирования или трансмембранным доменом.
Соответственно, трансмембранный домен может являться любой белковой структурой, термодинамически стабильной в мембране. Как правило, он является альфа-спиралью, состоящей из нескольких гидрофобных остатков. Для получения трансмембранной части можно использовать трансмембранный домен из любого трансмембранного белка. Наличие и длину трансмембранного домена белка могут определять специалисты в этой области с использованием алгоритма TMHMM (http://www.cbs.dtu.dk/services/TMHMM-2.0/). Кроме того, учитывая, что трансмембранный домен белка является относительно простой структурой, т.е. полипептидной последовательностью, которая, как прогнозируют, будет образовывать гидрофобную альфа-спираль достаточной длины, чтобы перекрывать мембрану, также можно использовать искусственно сконструированный домен TM (в патенте США № 7052906 B1 описывают синтетические трансмембранные компоненты).
Трансмембранный домен можно получать из CD28, что приводит к хорошей стабильности.
Фактор транскрипции
Фактор транскрипции может являться любым фактором транскрипции, выбранным для стимуляции желаемого ответа после фосфорилирования соответствующих мотивов ITAM или ITIM.
Транскрипционный фактор может являться природным или искусственным. Искусственные факторы транскрипции можно получать, например, из TALEN, структур с цинковыми пальцами или CrispR/CAS9, последний из которых коэкспрессируется с гидовой мРНК.
Предпочтительно, фактор транскрипции будет содержать сигнал ядерной локализации, чтобы облегчить его транспортировку к нуклеазе после расщепления протеазным доменом.
В качестве примера, последовательность нуклеиновой кислоты (ii) (кодирующая белок, содержащий мембраносвязанный фактор транскрипции, содержащий: (i) мембранную связь; (ii) участок распознавания протеазой и (iii) фактор транскрипции) может кодировать белок, состоящий из последовательности, представленной как SEQ ID NO: 55, содержащей домен RQR8; трансмембранный домен CD4-Endotox1, участок распознавания протеазой TEV и фактор транскрипции VP16-GAL4, или содержащий ее.
SEQ ID NO: 55
MGTSLLCWMALCLLGADHADACPYSNPSLCSGGGGSELPTQGTFSNVSTNVSPAKPTTTACPYSNPSLCSGGGGSPAPRPPTPAPTIASQPLSLRPEACRPAAGGAVHTRGLDFACDMALIVLGGVAGLLLFIGLGIFFCVRCRHRRRQAERMAQIKRVVSEKKTAQAPHRFQKTCSPISGGGGSENLYFQMPKKKRKVAPPTDVSLGDELHLDGEDVAMAHADALDDFDLDMLGDGDSPGPGFTPHDSAPYGALDMADFEFEQMFTDALGIDEYGGSGGGSMQILVASDATMKLLSSIEQACDICRLKKLKCSKEKPKCAKCLKNNWECRYSPKTKRSPLTRAHLTEVESRLERLEQLFLLIFPREDLDMILKMDSLQDIKALLTGLFVQDNVNКДАVTDRLASVETDMPLTLRQHRISATSSSEESSNKGQRQLTV
Соответственно, белок может содержать последовательность, представленную как SEQ ID NO: 55 или ее вариант, имеющий по меньшей мере 80, 85, 90, 95, 98 или 99% идентичности последовательности.
РЕЦЕПТОР
Настоящее изобретение дополнительно относится к конструкции нуклеиновой кислоты, содержащей (a) последовательность нуклеиновой кислоты, кодирующую слитый белок по первому аспекту настоящего изобретения, содержащий домен SH2 PTPN6, или укороченный белок по третьему аспекту настоящего изобретения; и (b) последовательность нуклеиновой кислоты, кодирующую рецептор, содержащий ITIM-содержащий эндодомен.
СТЕРИЛИЗУЮЩИЙ СИГНАЛ
Слитый белок, содержащий протеазу, как описано выше, может коэкспрессироваться в клетке с рецептором-мишенью, содержащим внутриклеточный участок расщепления протеазой. Расщепление рецептора-мишени по участку распознавания протеазой будет высвобождать внутриклеточную, дистальную по отношению к мембране часть рецептора-мишени.
Рецептор-мишень, например, может являться T-клеточным рецептором (TCR) или химерным антигенным рецептором (CAR).
Рецептор может содержать активирующий или костимуляторный эндодомен, расположенный на конце внутриклеточной части белка. Затем расщепление по участку расщепления протеазой приводит к удалению активирующего или костимуляторного эндодомена из целевого CAR, снижению или предотвращению опосредуемой рецептором-мишенью активации T-клеток.
Альтернативно, рецептор-мишень может содержать ингибиторный эндодомен, расположенный на конце внутриклеточной части белка. Затем расщепление по участку расщепления протеазой приводит к удалению ингибиторного эндодомена из целевого CAR, "включению" потенциала для опосредуемой рецептором-мишенью активации T-клеток.
Ингибиторный эндодомен, например, может содержать эндодомен CD148 или CD45 или ITIM-содержащий эндодомен из белка, такого как PD1, PDCD1, BTLA4, LILRB1, LAIR1, CTLA4, KIR2DL1, KIR2DL4, KIR2DL5, KIR3DL1 или KIR3DL3.
В качестве примера, рецептор-мишень может содержать последовательность, представленную как SEQ ID NO: 56, содержащую CAR против CD33, содержащий эндодомен ITIM из PD-1.
SEQ ID NO: 56
MAVPTQVLGLLLLWLTDARCDIQMTQSPSSLSASVGDRVTITCRASEDIYFNLVWYQQKPGKAPKLLIYDTNRLADGVPSRFSGSGSGTQYTLTISSLQPEDFATYYCQHYKNYPLTFGQGTKLEIKRSGGGGSGGGGSGGGGSGGGGSRSEVQLVESGGGLVQPGGSLRLSCAASGFTLSNYGMHWIRQAPGKGLEWVSSISLNGGSTYYRDSVKGRFTISRDNAKSTLYLQMNSLRAEDTAVYYCAAQDAYTGGYFDYWGQGTLVTVSSMDPATTTKPVLRTPSPVHPTGTSQPQRPEDCRPRGSVKGTGLDFACDIYVGVVGGLLGSLVLLVWVLAVICSRAARGTIGARRTGQPLKEDPSAVPVFSVDYGELDFQWREKTPEPPVPCVPEQTEYATIVFPSGMGTSSPARRGSADGPRSAQPLRPEDGHCSWPL
Соответственно, белок может содержать последовательность, представленную как SEQ ID NO: 56 или ее вариант, имеющий по меньшей мере 80, 85, 90, 95, 98 или 99% идентичности последовательности.
Если рецептор содержит участок расщепления протеазой между трансмембранным доменом и активирующим эндодоменом, слитый белок со стерилизующим сигналом можно использовать для ингибирования рецептора. Например, первый CAR можно конструировать таким образом, что его эндодомен отделен от трансмембранного домена участком расщепления протеазой. Второй CAR, распознающий другой антиген, может содержать ITIM-содержащий эндодомен. Распознавание когнатного антигена второго рецептора будет приводить к рекрутированию слитого белка со стерилизующим сигналом к мембране и последующему расщеплению по участку распознавания протеазой. Такое расщепление будет отделять активирующий эндодомен от первого рецептора и предотвращать активацию и распространение сигнала от указанного рецептора.
Это будет приводить к логическому элементу типа "И НЕТ", когда поддерживаемый сигнал будет передаваться, только если первый CAR активирован в изоляции (т.е. если первый CAR связан со своим когнатным антигеном, но второй CAR не связывается со своим когнатным антигеном). Такие "логические элементы" могут быть полезны, например, т.к. это относительно редко встречается, чтобы наличие (или отсутствие) одного антигена эффективно определяло злокачественное новообразование, что может приводить к отсутствию специфичности. Направленное воздействие на экспрессию антигена на нормальных клетках приводит к специфической для мишени, внеопухолевой токсичности. При некоторых злокачественных новообразованиях опухоль лучше всего определяется наличием одного антигена, (как правило, тканеспецифического антигена) и отсутствием другого антигена, присутствующего на нормальных клетках. Например, клетки острого миелолейкоза (AML) экспрессируют CD33. Нормальные стволовые клетки экспрессируют CD33, но также экспрессируют CD34, в то время как клетки AML, как правило, являются CD34-отрицательными. Направленное воздействие CD33 в отдельности для лечения AML ассоциировано со значительной токсичностью, т.к. оно истощает нормальные стволовые клетки. Однако подвергаемые специфическому воздействию клетки, являющиеся CD33-положительными, но не CD34-положительными, будут избегать этой значительной неспецифической для мишени токсичности.
Потенциальные пары антигенов для такого элемента "И НЕТ" представлены в таблице 2.
Таблица 2
В качестве примера, рецептор, содержащий участок расщепления протеазой, между трансмембранным доменом и активирующим эндодоменом может являться последовательностью, представленной как SEQ ID NO: 57, содержащей CAR против CD19 с расщепляемым эндодоменом CD3 дзета.
SEQ ID NO: 57
MSLPVTALLLPLALLLHAARPDIQMTQTTSSLSASLGDRVTISCRASQDISKYLNWYQQKPDGTVKLLIYHTSRLHSGVPSRFSGSGSGTDYSLTISNLEQEDIATYFCQQGNTLPYTFGGGTKLEITKAGGGGSGGGGSGGGGSGGGGSEVKLQESGPGLVAPSQSLSVTCTVSGVSLPDYGVSWIRQPPRKGLEWLGVIWGSETTYYNSALKSRLTIIKDNSKSQVFLKMNSLQTDDTAIYYCAKHYYYGGSYAMDYWGQGTSVTVSSDPTTTPAPRPPTPAPTIASQPLSLRPEACRPAAGGAVHTRGLDFACDIFWVLVVVGGVLACYSLLVTVAFIIFWVRCRHRRRQAERMAQIKRVVSEKKTAQAPHRFQKTCSPISGGGGSENLYFQMRRVKFSRSADAPAYQQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPR
Соответственно, рецептор может содержать последовательность, представленную как SEQ ID NO: 57 или ее вариант, имеющий по меньшей мере 80, 85, 90, 95, 98 или 99% идентичности последовательности.
Если рецептор содержит активирующий эндодомен, слитый с ингибиторным эндодоменом через участок расщепления протеазой, слитый белок со стерилизующим сигналом можно использовать для активации искусственных сигнальных доменов. Например, первый CAR можно конструировать таким образом, чтобы его эндодомен содержал активирующий эндодомен, слитый с ингибиторным эндодоменом через участок расщепления протеазой. Второй CAR, распознающий другой антиген, может содержать ITIM-содержащий эндодомен. Распознавание когнатного антигена второго рецептора будет приводить к рекрутированию слитого белка со стерилизующим сигналом к мембране и последующему отщеплению ингибиторного эндодомена от активирующего эндодомена первого рецептора. Расщепление и, таким образом, отделение ингибиторного домена от активирующего домена сделает возможной активацию первого CAR после связывания антигена и, таким образом, активации передачи сигнала через первый рецептор.
Это будет приводить к получению логического элемента CAR типа "И", в котором продуктивная передача сигнала будет происходить, только если оба, первый и второй, рецепторы активированы. Такие "логические элементы" являются полезными, например, т.к. большинство злокачественных новообразований нельзя отличить от нормальных тканей с учетом одного антигена. Таким образом, имеет место значительная "специфическая в отношении мишени, внеопухолевая" токсичность, в результате чего нормальные ткани повреждаются при терапии. В случае некоторых злокачественных новообразований направленное воздействие при наличии двух антигенов злокачественных опухолей может быть более селективным и, таким образом, эффективным, чем направленное воздействие на один. Например, хронический B-клеточный лимфоцитарный лейкоз (B-CLL) является распространенным лейкозом, который в настоящее время лечат посредством направленного воздействия на CD19. Таким образом лечат лимфому, но это также истощает весь B-клеточный компартмент таким образом, что лечение имеет значительный токсический эффект. B-CLL имеет необычный фенотип, в котором коэкспрессируются CD5 и CD19. При направленном воздействии только на клетки, экспрессирующие CD5 и CD19, можно будет значительно снижать специфическую в отношении мишени, внеопухолевую токсичность.
Потенциальные пары антигенов для такого логического элемента "И" представлены в таблице 3.
Таблица 3
В качестве примера, рецептор, содержащий активирующий эндодомен, слитый с ингибиторным эндодоменом через участок расщепления протеазой, может являться последовательностью, представленной как SEQ ID NO: 58, содержащей CAR против CD19 с эндодоменом CD3 дзета и расщепляемым эндодоменом CD148.
SEQ ID NO: 58
MSLPVTALLLPLALLLHAARPDIQMTQTTSSLSASLGDRVTISCRASQDISKYLNWYQQKPDGTVKLLIYHTSRLHSGVPSRFSGSGSGTDYSLTISNLEQEDIATYFCQQGNTLPYTFGGGTKLEITKAGGGGSGGGGSGGGGSGGGGSEVKLQESGPGLVAPSQSLSVTCTVSGVSLPDYGVSWIRQPPRKGLEWLGVIWGSETTYYNSALKSRLTIIKDNSKSQVFLKMNSLQTDDTAIYYCAKHYYYGGSYAMDYWGQGTSVTVSSDPTTTPAPRPPTPAPTIASQPLSLRPEACRPAAGGAVHTRGLDFACDIFWVLVVVGGVLACYSLLVTVAFIIFWVRRVKFSRSADAPAYQQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPRENLYFQMAVFGCIFGALVIVTVGGFIFWRKKRКДАKNNEVSFSQIKPKKSKLIRVENFEAYFKKQQADSNCGFAEEYEDLKLVGISQPKYAAELAENRGKNRYNNVLPYDISRVKLSVQTHSTDDYINANYMPGYHSKKDFIATQGPLPNTLKDFWRMVWEKNVYAIIMLTKCVEQGRTKCEEYWPSKQAQDYGDITVAMTSEIVLPEWTIRDFTVKNIQTSESHPLRQFHFTSWPDHGVPDTTDLLINFRYLVRDYMKQSPPESPILVHCSAGVGRTGTFIAIDRLIYQIENENTVDVYGIVYDLRMHRPLMVQTEDQYVFLNQCVLDIVRSQKDSKVDLIYQNTTAMTIYENLAPVTTFGKTNGYIA
Соответственно, рецептор может содержать последовательность, представленную как SEQ ID NO: 58 или ее вариант, имеющий по меньшей мере 80, 85, 90, 95, 98 или 99% идентичности последовательности.
БЛОКИРУЮЩИЙ СИГНАЛ
Настоящее изобретение относится к укороченному белку, содержащему домен SH2 из белка, связывающегося с фосфорилированным иммунорецепторным тирозиновым активирующим мотивом (ITAM), но в котором отсутствует киназный домен
Например, укороченный белок может содержать домен SH2 ZAP70, но в нем отсутствует киназный домен ZAP70. Другими словами, настоящее изобретение относится к укороченному белку, который: (i) содержит последовательность, представленную как SEQ ID NO: 2, но не содержащую последовательность, представленную как SEQ ID NO: 26.
Гиперэкспрессия домена SH2 ZAP70 приводит к конкуренции с полноразмерным ZAP70/ZAP70 дикого типа. Т.к. укороченный ZAP70 не может распространять сигналы, передача сигнала снижена пропорционально соотношению ZAP70 дикого типа и укороченного белка. Полезным может являться снижение силы активации T-клеток, например, для предотвращения гиперактивации T-клеток, которая может приводить к истощению T-клеток, индуцируемой активацией гибели клеток и, в клинических условиях, к цитокиновым бурям.
Настоящее изобретение также относится к укороченному белку, содержащему домен SH2 из белка, связывающегося с фосфорилированным иммунорецепторным тирозиновым ингибирующим мотивом (ITIM), но в котором отсутствует фосфатазный домен
Например, укороченный белок может содержать домен SH2 PTPN6, но в нем отсутствует фосфатазный домен PTPN6. Другими словами, настоящее изобретение относится к укороченному белку, который: (i) содержит последовательность, представленную как SEQ ID NO: 6, но не содержащую последовательность, представленную как SEQ ID NO: 27.
В этом случае, передачу сигнала ITIM можно снижать пропорционально соотношению PTPN6 дикого типа и укороченного белка. Полезным может являться снижение ингибиторных сигналов, таких как сигнал PD1. Это может быть применимым, когда T-клетки направленно воздействуют на опухоль, гиперэкспрессирующую PDL1 (или схожие ингибиторные рецепторы) для избегания иммунного отторжения.
Использование блокирующего сигнала или перекрестного сигнала, как описано выше, имеет значительное преимущество по сравнению с общепринятыми подходами блокады иммунных контрольных точек, когда, как правило, блокируют одно взаимодействие лиганд/рецептор, такое как PD-L1/PD1, с использованием антитела. Как описано выше, класс ингибиторных иммунных рецепторов содержит множество членов с избыточностью и профилями экспрессии, колеблющимися в зависимости от состояния T-клеток. Использование антитела или рекомбинантного лиганда/рецептора может эффективно блокировать один ингибиторный рецептор, но не будет влиять на ингибиторные сигналы, передаваемые от остальных. Геномное редактирование отдельных ингибиторных рецепторов (Menger et al, Cancer Res. 2016 Apr 15;76(8):2087-93) обладает схожими ограничениями. Стратегии слияния между отдельными ингибиторными рецепторами и костимуляторными доменами также обладает схожими ограничениями (Liu et al, Cancer Res. 2016 Mar 15;76(6):1578-90).
С помощью способа по настоящему изобретению будут блокировать (и в зависимости от стратегии реинтерпретировать) ингибиторные сигналы, передаваемые через ITIM. Таким образом, модулируют целый класс ингибиторных сигналов. Список ингибиторных рецепторов, передающих сигнал через ITIM, представлен в таблице II в Odorizzi and Wherry (2012) J. Immunol. 188:2957-2965. Они включают: PD1, BTLA, 2B4, CTLA-4, GP49B, Lair-1, Pir-B, PECAM-1, CD22, Siglec 7, Siglec 9, KLRG1, ILT2, CD94-NKG2A и CD5.
НУКЛЕИНОВАЯ КИСЛОТА
В одном из аспектов настоящее изобретение относится к нуклеиновой кислоте, кодирующей слитый белок или укороченный белок по настоящему изобретению.
В рамках изобретения термины "полинуклеотид", "нуклеотид" и "нуклеиновая кислота" должны быть синонимичными друг другу.
Специалисту в этой области будет понятно, что многочисленные различные полинуклеотиды и нуклеиновые кислоты могут кодировать один полипептид в результате вырожденности генетического кода. Кроме того, специалистам в этой области понятно, что с использованием рутинных способов можно осуществлять замены нуклеотидов, не влияющих на последовательность полипептида, кодируемую описанными полинуклеотидами, для отражения использования кодонов любого конкретного организма-хозяина, в котором полипептиды должны экспрессироваться.
Нуклеиновые кислоты по изобретению могут содержать ДНК или РНК. Они могут являться одноцепочечными или двухцепочечными. Они также могут являться полинуклеотидами, включающими синтетические или модифицированные нуклеотиды. В этой области известен ряд различных типов модификаций олигонуклеотидов. Они включают метилфосфонатные и фосфотиоатные остовы, добавление акридина или полилизиновых цепей на 3'- и/или 5'-концы молекулы. В целях использования, как представлено в настоящем описании, следует понимать, что полинуклеотиды можно модифицировать любым способом, доступным в этой области. Такие модификации можно осуществлять для повышения активности in vivo или продолжительности жизни интересующих полинуклеотидов.
Термины "вариант", "гомолог" или "производное" в отношении нуклеотидной последовательности включают любую замену, вариант, модификацию, делецию или добавление одной (или более) нуклеиновой кислоты из последовательности или в нее.
КОНСТРУКЦИЯ НУКЛЕИНОВОЙ КИСЛОТЫ
В одном из аспектов настоящее изобретение относится к конструкции нуклеиновой кислоты, коэкспрессирующей укороченный белок или слитый белок по настоящему изобретению с другим белком. Конструкция нуклеиновой кислоты может содержать: последовательность нуклеиновой кислоты, кодирующую укороченный белок или слитый белок по настоящему изобретению; и нуклеиновую кислоту, кодирующую другой белок.
Настоящее изобретение относится к конструкции нуклеиновой кислоты, коэкспрессирующей укороченный белок или слитый белок по настоящему изобретению с химерным антигенным рецептором. Конструкция нуклеиновой кислоты может содержать: (i) последовательность нуклеиновой кислоты, кодирующую укороченный белок или слитый белок по настоящему изобретению; и (ii) нуклеиновую кислоту, кодирующую химерный антигенный рецептор.
Химерный антигенный рецептор (CAR) может являться активирующим CAR, содержащим ITAM-содержащий эндодомен, такой как CD3 дзета. CAR может являться ингибиторным CAR, содержащим эндодомен "без лигирования", как описано в WO2015/075469, который может содержать весь или часть эндодомена из рецептор-подобной тирозинфосфатазы, такой как CD148 или CD45. CAR может являться ингибиторным CAR, содержащим эндодомен "с лигированием", как описано в WO2015/075470, который может содержать домен ITIM.
Слитые белки и укороченные белки по изобретению можно использовать вместе с клеткой, экспрессирующей "логический элемент"-комбинацию двух или более CAR. Логический элемент "ИЛИ" содержит два активирующих CAR, как описано в WO2015/075468. Логический элемент "И" содержит активирующий CAR и ингибиторный CAR "без лигирования", как описано в WO2015/075469. "И" не содержит активирующий CAR и ингибиторный CAR "с лигированием", как описано в WO2015/075470.
Таким образом, настоящее изобретение относится к конструкции нуклеиновой кислоты, содержащей:
(i) последовательность нуклеиновой кислоты, кодирующую укороченный белок или слитый белок по изобретению;
(ii) первый химерный антигенный рецептор (CAR); и
(iii) второй химерный антигенный рецептор.
Что касается аспекта транскрипционного сигнала по изобретению, настоящее изобретение относится к конструкции нуклеиновой кислоты, содержащей (i) последовательность нуклеиновой кислоты, кодирующую слитый белок, содержащий домен SH2; и протеазный домен; и (ii) последовательность нуклеиновой кислоты, кодирующую мембраносвязанный фактор транскрипции, содержащий: мембранную связь; участок распознавания протеазой и фактор транскрипции.
Что касается аспекта стерилизующего сигнала по изобретению, настоящее изобретение относится к конструкции нуклеиновой кислоты, содержащей (i) последовательность нуклеиновой кислоты, кодирующую слитый белок, содержащий домен SH2 и протеазный домен (например, TeV-домен); и (ii) последовательность нуклеиновой кислоты, кодирующую рецептор, содержащий участок расщепления протеазой.
Например, настоящее изобретение относится к конструкции нуклеиновой кислоты, содержащей: (a) последовательность нуклеиновой кислоты, кодирующую слитый белок, содержащий (i) домен SH2 PTPN6; и (ii) протеазный домен (например, TeV-домен); (b) последовательность нуклеиновой кислоты, кодирующую рецептор, содержащий участок расщепления протеазой; и (c) последовательность нуклеиновой кислоты, кодирующую рецептор, содержащий ITIM-содержащий эндодомен.
Рецептор может являться T-клеточным рецептором (TCR) или химерным антигенным рецептором (CAR).
Соответственно, белок, кодируемый последовательностью нуклеиновой кислоты (b), может являться T-клеточным рецептором (TCR) или химерным антигенным рецептором (CAR), содержащим: (i) участок расщепления протеазой между трансмембранным доменом и активирующим эндодоменом; или (ii) активирующий эндодомен, слитый с ингибиторным эндодоменом через участок расщепления протеазой.
Если конструкция нуклеиновой кислоты по изобретению приводит к образованию отдельных полипептидов, как в том случае, когда она коэкспрессирует слитый белок по изобретению и CAR, она также может содержать последовательность нуклеиновой кислоты, делающую возможной экспрессию обоих белков. Например, она может содержать последовательность, кодирующую участок расщепления между двумя последовательностями нуклеиновой кислоты. Участок расщепления может являться саморасщепляющимся, таким образом, что если продуцируется выделяющийся полипептид, он незамедлительно расщепляется на два белка без необходимости какой-либо внешней расщепляющей активности.
Известны различные саморасщепляющиеся участки, включая саморасщепляющийся пептид 2a вируса ящура (FMDV), имеющий представленную последовательность:
SEQ ID NO: 59
RAEGRGSLLTCGDVEENPGP
или
SEQ ID NO: 60
QCTNYALLKLAGDVESNPGP
Коэкспрессируемая последовательность может являться участком внутренней посадки рибосомы (IRES). Коэкспрессируемая последовательность может являться внутренним промотором.
ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР (CAR)
CAR, схематически представленные на фигуре 13, являются химерными трансмембранными белками типа I, соединяющими внеклеточный антигенраспознающий домен (связывающее средство) с внутриклеточным сигнальным доменом (эндодоменом). Связывающее средство, как правило, является одноцепочечным вариабельным фрагментом (scFv), полученным из моноклонального антитела (mAb), но оно может быть основано на других форматах, содержащих антитело-подобный антигенсвязывающий участок. Спейсерный домен, как правило, необходим для изоляции связывающего средства от мембраны, и чтобы оно приняло подходящую ориентацию. Распространенным используемым спейсерным доменом является Fc IgG1. Может быть достаточно более компактных спейсеров, например, стеблевой области из CD8α и даже просто шарнирной области IgG1 в отдельности в зависимости от антигена. Трансмембранный домен заякоривает белок в мембране клетки и соединяет спейсер с эндодоменом.
Ранние дизайны CAR содержали эндодомены, полученные из внутриклеточных частей γ-цепи FcεR1 или CD3ζ. Таким образом, эти рецепторы первого поколения передавали иммунологический сигнал 1, достаточный для уничтожения T-клетками когнатных клеток-мишеней, но не способный полностью активировать T-клетку для пролиферации и выживания. Для преодоления этого ограничения конструировали составные эндодомены: слияние внутриклеточной части костимуляторной молекулы T-клетки с таковой из CD3ζ приводит к получению рецепторов второго поколения, которые могут передавать активирующий и костимуляторный сигнал одновременно после распознавания антигена. Наиболее общеупотребительным костимуляторным доменом является домен CD28. Он обеспечивает наиболее мощный костимуляторный сигнал, а именно иммунологический сигнал 2, запускающий пролиферацию T-клеток. Также описаны некоторые рецепторы, включающие эндодомены семейства рецепторов ФНО, таких как близкородственные OX40 и 41BB, передающие сигналы выживания. Описаны даже более мощные CAR третьего поколения, содержащие эндодомены, способные передавать сигналы активации, пролиферации и выживания.
CAR-кодирующие нуклеиновые кислоты можно переносить в T-клетки с использованием, например, ретровирусных векторов. Можно использовать лентивирусные векторы. Таким образом, можно получать большое количество опухолеспецифических T-клеток для адоптивного переноса клеток. Когда CAR связывается с антигеном-мишенью, это приводит к передаче активирующего сигнала в T-клетку, на которой он экспрессируется. Таким образом, CAR направляет специфичность и цитотоксичность T-клетки на опухолевые клетки, экспрессирующие антиген-мишень.
Таким образом, CAR, как правило, содержат: (i) антигенсвязывающий домен; (ii) спейсер; (iii) трансмембранный домен и (iii) внутриклеточный домен, содержащий сигнальный домен или связанный с ним.
АНТИГЕНСВЯЗЫВАЮЩИЙ ДОМЕН
Антигенсвязывающий домен является частью CAR, распознающей антиген. В этой области известно множество антигенсвязывающих доменов, включая домены на основе антигенсвязывающего участка антитела, миметики антитела и T-клеточные рецепторы. Например, антигенсвязывающий домен может содержать: одноцепочечный вариабельный фрагмент (scFv), полученный из моноклонального антитела; природный лиганд антигена-мишени; пептид с достаточной аффинностью к мишени; однодоменное антитело; искусственное одиночное связывающее средство, такое как дарпин (сконструированный белок с анкириновыми повторами); или одноцепочечное средство, полученное из T-клеточного рецептора.
Антигенсвязывающий домен может содержать домен, не основанный на антигенсвязывающем участке антитела. Например, антигенсвязывающий домен может содержать домен на основе белка/пептида, являющегося растворимым лигандом для рецептора поверхности опухолевой клетки (например, растворимым пептидом, таким как цитокин или хемокин); или внеклеточным доменом заякоренного в мембране лиганда или рецептора, партнер по связыванию которого экспрессируется на опухолевой клетке.
Антигенсвязывающий домен может быть основан на природном лиганде антигена.
Антигенсвязывающий домен может содержать аффинный пептид из комбинаторной библиотеки или сконструированный de novo аффинный белок/пептид.
СПЕЙСЕРНЫЙ ДОМЕН
CAR содержат спейсерную последовательность для соединения антигенсвязывающего домена с трансмембранным доменом и пространственного разделения антигенсвязывающего домена из эндодомена. Гибкий спейсер позволяет антигенсвязывающему домену ориентироваться в различных направлениях для облегчения связывания.
В аспектах по настоящему изобретению, когда необходимы два CAR, первый и второй CAR могут содержать различные спейсерные молекулы. Например, спейсерная последовательность может содержать Fc-область IgG1, шарнирную область IgG1 или стеблевую область CD8 человека или стеблевую область CD8 мыши. Альтернативно, спейсер может содержать альтернативную линкерную последовательность, имеющую схожую длину и/или свойства пространственного положения домена с таковыми у Fc-области IgG1, шарнирной области IgG1 или стеблевой области CD8. Спейсер IgG1 человека можно изменять для удаления Fc-связывающих мотивов.
Все спейсерные домены, упомянутые выше, образуют гомодимеры. Однако механизм не ограничен использованием гомодимерных рецепторов и должен работать с мономерными рецепторами при условии, что спейсер является достаточно жестким. Примером такого спейсера является CD2 или укороченный CD22.
Т.к. CAR, как правило, являются гомодимерами (см. фигуру 13a), перекрестное спаривание может приводить к образованию гетеродимерного химерного антигенного рецептора. Это нежелательно по различным причинам, например: (1) эпитоп может не находиться на том же "уровне" на клетке-мишени таким образом, что перекрестно спариваемый CAR может быть способен связываться лишь с одним антигеном; (2) VH и VL из двух разных scFv могут менять местами и не распознавать мишень или хуже распознавать неожиданный и непрогнозируемый антиген. В случае описываемых выше логических элементов "И" и "И НЕТ", спейсер первого CAR может достаточно отличаться от спейсера второго CAR во избежание перекрестного спаривания, но быть достаточно схожим для колокализации. Можно использовать пары ортологичных спейсерных последовательностей. Примерами являются стеблевые области CD8 мыши и человека или, альтернативно, спейсерные домены, являющиеся мономерными, например, эктодомен CD2.
Примеры колокализующихся пар спейсеров представлены в следующей таблице:
Трансмембранный домен
Трансмембранный домен является последовательностью CAR, пересекающей мембрану.
Трансмембранный домен может являться любой белковой структурой, термодинамически стабильной в мембране. Как правило, он является альфа-спиралью, содержащей несколько гидрофобных остатков. Для получения трансмембранной части по изобретению можно использовать трансмембранный домен из любого трансмембранного белка. Наличие и длину трансмембранного домена белка могут определять специалисты в этой области с использованием алгоритма TMHMM (http://www.cbs.dtu.dk/services/TMHMM-2.0/). Кроме того, учитывая, что трансмембранный домен белка является относительно простой структурой, т.е. полипептидной последовательностью, которая, как прогнозируют, будет образовывать гидрофобную альфа-спираль достаточной длины, чтобы пересекать мембрану, также можно использовать искусственно сконструированный TM домен (в патенте США № 7052906 B1 описывают синтетические трансмембранные компоненты).
Трансмембранный домен можно получать из CD28, что обеспечивает хорошую стабильность рецептора.
АКТИВИРУЮЩИЙ ЭНДОДОМЕН
Эндодомен является частью CAR, передающей сигнал. Он может являться частью или быть связанным с внутриклеточным доменом CAR. После распознавания антигена, рецепторы кластеризуются, нативные CD45 и CD148 исключаются из синапса, и сигнал передается в клетку. Наиболее общеупотребительным эндодоменным компонентом является таковой из CD3 дзета, содержащий 3 ITAM. Он передает активационный сигнал в T-клетку после связывания антигена. CD3 дзета может не обеспечивать полностью надлежащий активационный сигнал, и может потребоваться дополнительный костимуляторный сигнал. Например, можно использовать химерный CD28 и OX40 с CD3 дзета для передачи пролиферативного сигнала/сигнала выживания или все три можно использовать совместно.
Если CAR содержит активирующий эндодомен, он может содержать только эндодомен CD3 дзета, эндодомен CD3 дзета с таковым из CD28 или OX40 или эндодомен CD28 и эндодомен OX40 и CD3 дзета.
Любой эндодомен, содержащий мотив ITAM, может действовать как активационный эндодомен по настоящему изобретению. Подходящие эндодомены, содержащие мотив ITAM, представлены в настоящем описании.
ИНГИБИТОРНЫЙ ДОМЕН
В вариантах осуществления, обозначенных выше как логический элемент "И", первый CAR может содержать активирующий эндодомен, слитый с ингибиторным эндодоменом через участок расщепления протеазой. В связи с этим, ингибиторный эндодомен ингибирует активацию T-клетки первым CAR в отсутствие активации второго CAR. После активации второго CAR ITIM в эндодомене второго CAR фосфорилируется, и слитый белок PTPN6/протеазный домен рекрутируется к мембране. Это приводит к расщеплению первого CAR между активирующим эндодоменом и ингибиторным эндодоменом, таким образом, делая возможной активацию T-клеток.
Ингибиторные эндодомены могут содержать любую последовательность, ингибирующую передачу сигнала T-клеток активирующим CAR, когда он находится в том же эндодомене.
Ингибиторный эндодомен может являться тирозинфосфатазой, такой как рецептор-подобная тирозинфосфатаза, или содержать ее. Ингибиторный эндодомен может являться любой тирозинфосфатазой, способной ингибировать передачу сигнала TCR при колокализации с активирующим эндодоменом CAR, или содержать ее. Ингибиторный эндодомен может являться любой тирозинфосфатазой с достаточно большой каталитической скоростью для фосфорилированных ITAM, способной ингибировать передачу сигнала TCR при колокализации с активирующим эндодоменом CAR, или содержать ее.
ВЕКТОР
Настоящее изобретение также относится к вектору или набору векторов, содержащих одну более последовательностей нуклеиновой кислоты или конструкций по настоящему изобретению. Такой вектор можно использовать для встраивания последовательностей нуклеиновой кислоты или конструкций в клетку-хозяина таким образом, что она экспрессирует белки, кодируемые последовательностью нуклеиновой кислоты или конструкцией.
Вектор может являться, например, плазмидой или вирусным вектором, таким как ретровирусный вектор или лентивирусный вектор, или вектором на основе транспозона или синтетической мРНК.
С помощью вектора можно трансфицировать или трансдуцировать T-клетку.
КЛЕТКА
Настоящее изобретение также относится к иммунной клетке, содержащей слитый белок, укороченный белок, нуклеиновую кислоту и/или конструкцию нуклеиновой кислоты по настоящему изобретению.
Клетка может являться цитолитической иммунной клеткой.
Цитолитические иммунные клетки могут являться T-клетками или T-лимфоцитами, представляющими собой тип лимфоцитов, играющий центральную роль в клеточном иммунитете. Их можно отличать от других лимфоцитов, таких как B-клетки и естественные киллеры (NK-клетки), по наличию T-клеточного рецептора (TCR) на поверхности клетки. Существуют различные типы T-клеток, как описано ниже.
T-хелперные клетки (TH-клетки) помогают другим лейкоцитам в иммунологических процессах, включая созревание B-клеток в плазматические клетки и B-клетки памяти и активацию цитотоксических T-клеток и макрофагов. TH-клетки экспрессируют CD4 на своей поверхности. TH-клетки становятся активированными, когда молекулы MHC класса II презентируют им пептидные антигены на поверхности антигенпрезентирующих клеток (APC). Эти клетки могут дифференцироваться в несколько субпопуляций, включая TH1, TH2, TH3, TH17, Th9 или TFH, секретирующие различные цитокины для стимуляции различных типов иммунных ответов.
Цитолитические T-клетки (TC-клетки или CTL) разрушают инфицированные вирусом клетки и опухолевые клетки и также участвуют в отторжении трансплантата. CTL экспрессируют CD8 на своей поверхности. Эти клетки распознают свои мишени посредством связывания с антигеном, связанным с MHC класса I, присутствующим на поверхности всех ядросодержащих клеток. С помощью ИЛ-10, аденозина и других молекул, секретируемых регуляторными T-клетками, CD8+ клетки могут инактивироваться до анергического состояния, что предотвращает аутоиммунные заболевания, такие как экспериментальный аутоиммунный энцефаломиелит.
T-клетки памяти представляют собой субпопуляцию антиген-специфических T-клеток, персистирующих в течение длительного времени после разрешения инфекции. Они быстро проходят экспансию до больших количеств эффекторных T-клеток после повторного воздействия когнатного антигена, таким образом, снабжая иммунную систему "памятью" в отношении последних инфекции. T-клетки памяти включают три субпопуляции: T-клетки центральной памяти (TCM-клетки) и два типа эффекторных T-клеток памяти (TEM-клетки и TEMRA-клетки). Клетки памяти могут являться CD4+ или CD8+. T-клетки памяти, как правило, экспрессируют белок поверхности клетки CD45RO.
Регуляторные T-клетки (Treg-клетки), ранее известные как супрессорные T-клетки, являются ключевыми для поддержания иммунологической толерантности. Их основной ролью является выключение T-клеточного иммунитета для окончания иммунной реакции и супрессия аутореактивных T-клеток, избежавших отрицательной селекции в тимусе.
Описано два основных класса CD4+ Treg-клеток - природные Treg-клетки и адаптивные Treg-клетки.
Природные Treg-клетки (также известные как CD4+CD25+FoxP3+ Treg-клетки) возникают в тимусе и связаны с взаимодействиями между развивающимися T-клетками с миелоидными (CD11c+) и плазмацитоидными (CD123+) дендритными клетками, активированными TSLP. Природные Treg-клетки можно отличать от других T-клеток по наличию внутриклеточной молекулы под названием FoxP3. Мутации гена FOXP3 могут предотвращать развитие регуляторных T-клеток, вызывая фатальное аутоиммунное заболевание IPEX.
Адаптивные Treg-клетки (также известные как Tr1-клетки или Th3-клетки) могут возникать в течение нормального иммунного ответа.
Естественные киллеры (или NK-клетки) являются типом цитолитических клеток, образующих часть врожденной иммунной системы. NK-клетки обеспечивают быстрые ответы на врожденные сигналы от инфицированных вирусом клеток MHC-независимым образом.
NK-клетки (принадлежащие к группе врожденных лимфоидных клеток) определяют как большие гранулярные лимфоциты (LGL), и они составляют третий тип клеток, дифференцирующихся из общего лимфоидного предшественника с образованием B- и T-лимфоцитов. Известно, что NK-клетки дифференцируются и созревают в костном мозгу, лимфоузлах, селезенке, миндалинах и тимусе, откуда они затем могут попадать в кровоток.
Клетки по изобретению могут относиться к любому из упомянутых выше типов клеток.
T- или NK-клетки, экспрессирующие молекулы по изобретению, можно получать ex vivo из собственной периферической крови пациента (1-ая партия), или в условиях трансплантации гемопоэтических стволовых клеток из периферической крови донора (2-ая партия), или периферической крови несвязанного донора (3-я партия).
Альтернативно, T- или NK-клетки, экспрессирующие молекулы по изобретению, можно получать при дифференцировке ex vivo индуцибельных клеток-предшественников или эмбриональных клеток-предшественников в T-клетки. Альтернативно, можно использовать линию иммортализованных T-клеток, сохраняющих свою литическую функцию и способных действовать в качестве терапевтического средства.
Во всех этих вариантах осуществления клетки получают посредством встраивания ДНК или РНК, кодирующей рецепторный компонент и сигнальный компонент, одним из множества способов, включая трансдукцию с использованием вирусного вектора, трансфекцию с использованием ДНК или РНК.
Клетка по изобретению может являться T- или NK-клеткой ex vivo из индивидуума. T- или NK-клетку можно получать из образца мононуклеарных клеток периферической крови (PBMC). T- или NK-клетки можно активировать и/или подвергать экспансии перед трансдукцией с использованием нуклеиновой кислоты по изобретению, например, посредством обработки моноклональным антителом против CD3.
T- или NK-клетку по изобретению можно получать посредством:
(i) выделения образца, содержащего T- или NK-клетку из индивидуума или других указанных выше источников; и
(ii) трансдукции или трансфекции T- или NK-клеток с использованием одной более последовательностей нуклеиновой кислоты по изобретению.
Затем T- или NK-клетки можно очищать, например, выбирать с учетом экспрессии антигенсвязывающего домена антигенсвязывающего полипептида.
Настоящее изобретение также относится к клетке, содержащей слитый белок или укороченный белок по изобретению и химерный антигенный рецептор (CAR).
Химерный антигенный рецептор (CAR) может являться активирующим CAR, содержащим ITAM-содержащий эндодомен, такой как CD3 дзета. CAR может являться ингибиторным CAR, содержащим эндодомен "без лигирования", как описано в WO2015/075469, который может содержать весь или часть эндодомена из рецептор-подобной тирозинфосфатазы, такой как CD148 или CD45. CAR может являться ингибиторным CAR, содержащим эндодомен "с лигированием", как описано в WO2015/075470, который может содержать домен ITIM.
Слитые белки и укороченные белки по изобретению можно использовать вместе с клеткой, экспрессирующей комбинацию "логических элементов" из двух или более CAR. Логический элемент "ИЛИ" содержит два активирующих CAR, как описано в WO2015/075468. Логический элемент "И" содержит активирующий CAR и ингибиторный CAR "без лигирования", как описано в WO2015/075469. Элемент "И НЕТ" содержит активирующий CAR и ингибиторный CAR "с лигированием", как описано в WO2015/075470.
Таким образом, настоящее изобретение относится к клетке, содержащей:
(i) последовательность нуклеиновой кислоты, кодирующую укороченный белок или слитый белок по изобретению;
(ii) первый химерный антигенный рецептор (CAR); и
(iii) второй химерный антигенный рецептор.
Что касается аспект транскрипционного сигнала по изобретению, настоящее изобретение относится к клетке, содержащей (i) слитый белок, содержащий домен SH2 и протеазу; и (ii) мембраносвязанный фактор транскрипции, содержащий: мембранную связь, участок распознавания протеазой и фактор транскрипции.
Что касается аспекта стерилизующего сигнала по изобретению, настоящее изобретение относится к клетке, содержащей (i) слитый белок, содержащий домен SH2 и протеазу; и (ii) рецептор, содержащий участок расщепления протеазой.
Рецептор может являться, например, T-клеточным рецептором (TCR) или химерным антигенным рецептором (CAR), содержащим: (i) участок расщепления протеазой между трансмембранным доменом и активирующим эндодоменом; или (ii) активирующий эндодомен, слитый с ингибиторным эндодоменом через участок расщепления протеазой.
КОМПОЗИЦИЯ
Настоящее изобретение также относится к фармацевтической композиции, содержащей множество клеток по изобретению. Фармацевтическая композиция может дополнительно содержать фармацевтически приемлемый носитель, дилюент или эксципиент. Фармацевтическая композиция, необязательно, может содержать один более дополнительных фармацевтически активных полипептидов и/или соединений. Такой состав может находиться, например, в форме, подходящей для внутривенной инфузии.
СПОСОБ ЛЕЧЕНИЯ
Клетки по настоящему изобретению могут быть способны уничтожать клетки-мишени, такие как злокачественные клетки.
Клетки по настоящему изобретению можно использовать для лечения инфекции, такой как вирусная инфекция.
Клетки по изобретению также можно использовать для контроля патогенных иммунных ответов, например, при аутоиммунных заболеваниях, аллергиях и реакции "трансплантат против хозяина".
Клетки по изобретению можно использовать для лечения злокачественного заболевания, такого как рак мочевого пузыря, рак молочной железы, рак толстого кишечника, рак эндометрия, рак почки (почечноклеточный), лейкоз, рак легких, меланома, неходжкинская лимфома, рак поджелудочной железы, рак предстательной железы и рак щитовидной железы.
Клетки по изобретению можно использовать для лечения: злокачественных новообразований полости рта и глотки, включающих рак языка, рта и глотки; злокачественных новообразований пищеварительной системы, включающих рак пищевода, рак желудка и колоректальный рак; злокачественных новообразований печени и желчных протоков, включающих печеночноклеточные карциномы и холангиокарциномы; злокачественных новообразований дыхательной системы, включающие бронхогенные злокачественные новообразования и злокачественные новообразования гортани; злокачественных новообразований костей и суставов, включающих остеосаркому; злокачественных новообразований кожи, включающих меланому; рак молочной железы; злокачественных новообразований половой системы, включающих рак матки, яичников и шейки матки у женщин, рак предстательной железы и яичка у мужчин; злокачественных новообразований выделительной системы, включающих почечноклеточную карциному и переходноклеточные карциномы мочеточника или мочевого пузыря; злокачественных новообразований головного мозга, включая глиомы, мультиформную глиобластому и медуллобластомы; злокачественных новообразований эндокринной системы, включая рак щитовидной железы, аденокарциному и злокачественные новообразования, ассоциированные с синдромами множественных эндокринных неоплазий; лимфом, включая лимфому Ходжкина и неходжкинскую лимфому; множественной миеломы и плазмоцитом; лейкозов, острых и хронических, миелоидных или лимфоидных; и злокачественных новообразований других и неуказанных мест, включая нейробластому.
Лечение клетками по изобретению может способствовать профилактике избегания или высвобождения опухолевых клеток, часто возникающих при использовании стандартных подходов.
Изобретение далее будет дополнительно описано с помощью примеров, которые должны помочь специалисту в этой области в осуществлении изобретения, но не предназначены для какого-либо ограничения объема изобретения.
ПРИМЕРЫ
Пример 1 - Подчинение пути активации T-клеток повышенным или нефизиологическим сигналам
Ряд доменов SH2, участвующих в ранней активации T-клеточного сигнала тестировали для определения того, могут ли сигналы активации T-клеток подчиняться или "быть захваченными", таким образом, что когда T-клетка активирована, сигнал можно модулировать или повторно передавать.
Авторы настоящего изобретения получали несколько химерных конструкций AKT, соединяя киназный домен AKT с доменами SH2 из Zap70, Grap, Grb2 и PLCγ (фигура 8).
В нетрансдуцированных (NT) T-клетках определяют очень низкие уровни фосфорилирования эндогенного AKT после обработки OKT3 для индукции перекрестной сшивки и активации TCR (фигура 10b: верхняя панель). Однако в клетках, экспрессирующих конструкцию Zap-AKT, наблюдали значительные уровни фосфо-AKT (фигура 10b: нижняя панель).
Линкер для активации T-клеток (LAT) является нижележащей мишенью ZAP70 и связан несколькими SH2-содержащими белками, такими как Grb2, Grap и PLCγ. Предполагают, что домены SH2 из каждого из этих LAT-связывающих средств также будут делать возможным захват активационного сигнала от CD3 дзета. Однако этого не было.
Не наблюдали TCR-зависимого фосфорилирования киназного домена AKT выше уровней, наблюдаемых для NT T-клеток, если киназный домен AKT соединяли с доменами SH2 из Grb2, Grap или PLCγ (фигура 9a).
Это свидетельствует о том, что для этой системы захвата T-клеточного сигнала конкретно необходим тандемный домен SH2 из очень ранней T-клеточной сигнальной молекулы, такой как Zap70 или тирозин-протеинфосфатаза нерецепторного типа 6 (PTPN6).
Пример 2 - Транскрипционный контроль
Протеазу TeV подвергали слиянию с доменом SH2 Zap70. Мембраносвязанный фактор транскрипции также получали следующим образом: RQR8 клонировали в рамке считывания с фактором транскрипции VP16/GAL4, разделенным участком расщепления TeV. Этот слитый белок делает возможным высвобождение фактора транскрипции VP16/GAL4 (содержащего сигнал ядерной локализации) после расщепления TeV.
Оба этих белка экспрессировались в T-клетке, которая также экспрессировала CD19-специфический химерный антигенный рецептор. Для демонстрации того, что необходим подход с использованием ZAP70-TeV, фактор транскрипции коэкспрессировали с CD19 CAR, эндодомен которого заменяли TeV (фигура 11).
T-клетки подвергали воздействию CD19-отрицательных и положительных мишеней. Активацию транскрипции измеряли с помощью люциферазной кассеты, отвечающей на GALv/VP16. Только условия, в которых стандартный CD19 CAR коэкспрессировали с ZAP-TeV и мембраносвязанным фактором транскрипции, приводили к селективной активации транскрипции после распознавания CD19. CD19 CAR, слитый непосредственно с TeV, вызывал конститутивную активацию транскрипции (фигура 12).
Пример 3 - Блокада сигнала PD-1 с использованием укороченного SHP-1 (PTPN6) или укороченного SHP-2
Клетки PBMC трансдуцировали, как показано в следующей таблице:
и pDual-PD1-GFP
Клетки сокультивировали в течение 48 часов с клетками SupT1, трансдуцированными с использованием CD19, PDL1 или и того, и другого, и измеряли высвобождение ИФНγ, измеряемое с помощью ELISA. Результаты представлены на фигуре 15.
Наличие PDL1 на клетках-мишенях SupT1 вызывало снижение высвобождения ИФНγ. Наблюдали повышенное высвобождение ИФНγ при использовании PBMC, экспрессировавших конструкцию CAR вместе с укороченным SHP-1 или укороченным SHP-2, по сравнению с теми, которые экспрессировали только CAR. Это свидетельствует о том, что конструкции укороченного SHp-1 и SHP-2 успешно ингибировали ингибиторный сигнал PDL1 от клеток-мишеней.
Пример 4 - Перехват сигнала PD-1 с использованием продукта слияния доменов SH2 SHP-2 и киназы Zap70
Клетки PBMC трансдуцировали, как показано в следующей таблице:
Клетки сокультивировали в соотношении 1:1 в течение 24 часов с клетками SupT1, трансдуцированными с использованием CD19 или PDL1. Высвобождение ИФНγ измеряли с помощью ELISA (фигура 16A). Повышение продукции ИФНγ наблюдали в совместных культурах T-клетках, трансдуцированных с использованием CAR-SHP2.Zap70 +PD1, с клетками-мишенями PDL1 SupT1 по сравнением с T-клетками, трансдуцированными с использованием CAR +PD1.
Также осуществляли анализ цитотоксичности, в котором уничтожение клеток SupT1 количественно анализировали с помощью FACS (фигура 16B). Наблюдали почти полное уничтожение мишеней PDL1 SupT1 в совместных культурах PDL1-положительных клеток-мишеней с T-клетками, трансдуцированными с использованием CAR-SHP2.Zap70+PD1. В отличие от этого, не наблюдали уничтожение при использовании конструкции CAR+PD1 в отдельности. Это свидетельствует о том, что замена фосфатазного домена SHP2 киназным доменом Zap70 успешно преобразует ингибиторный сигнал PD1 в активирующий сигнал. Таким образом, слитый белок SHP-2-киназа Zap70 успешно перехватывал ингибиторный сигнал PDL1-PD1 и преобразовывал его в сигнал активации T-клеток.
Все публикации, упомянутые в представленном выше описании, включены в настоящее описание в качестве ссылок. Различные модификации и варианты описанных способов и системы по изобретению будут очевидны специалистам в этой области без отклонения от объема и сущность изобретения. Хотя изобретение описано в отношении конкретных предпочтительных вариантов осуществления, следует понимать, что изобретение не должно быть ненадлежаще ограничено такими конкретными вариантами осуществления. Фактически, различные модификации описанных способов осуществления изобретения, очевидные специалистам в молекулярной биологии, клеточной иммунологии или родственных областях, предназначены для включения в объем следующей формулы изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| КЛЕТКА | 2014 |
|
RU2732236C2 |
| КЛЕТКА | 2014 |
|
RU2717984C2 |
| ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР | 2014 |
|
RU2684713C2 |
| КЛЕТКА | 2015 |
|
RU2768019C2 |
| ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР | 2016 |
|
RU2752880C2 |
| ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР | 2015 |
|
RU2685479C2 |
| ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ И СПОСОБЫ ИХ ПОЛУЧЕНИЯ | 2015 |
|
RU2753965C2 |
| СИГНАЛЬНАЯ СИСТЕМА | 2015 |
|
RU2731638C2 |
| ХИМЕРНЫЙ ЦИТОКИНОВЫЙ РЕЦЕПТОР | 2016 |
|
RU2826122C1 |
| ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР | 2019 |
|
RU2810092C2 |
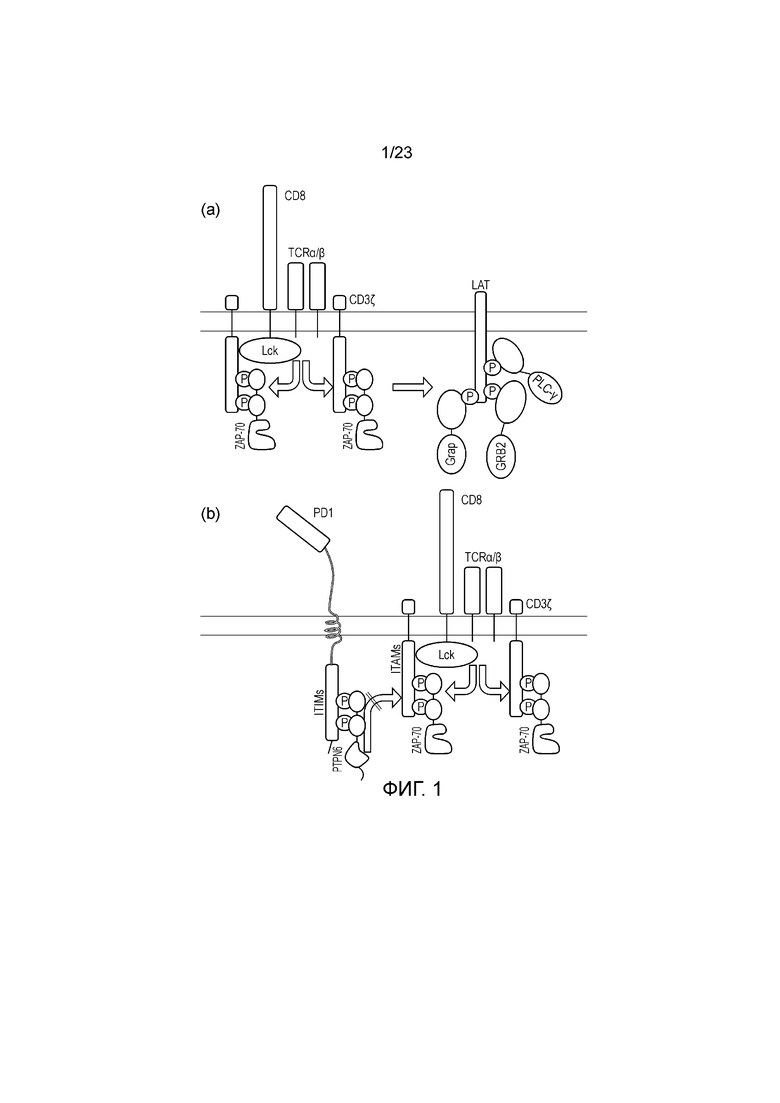
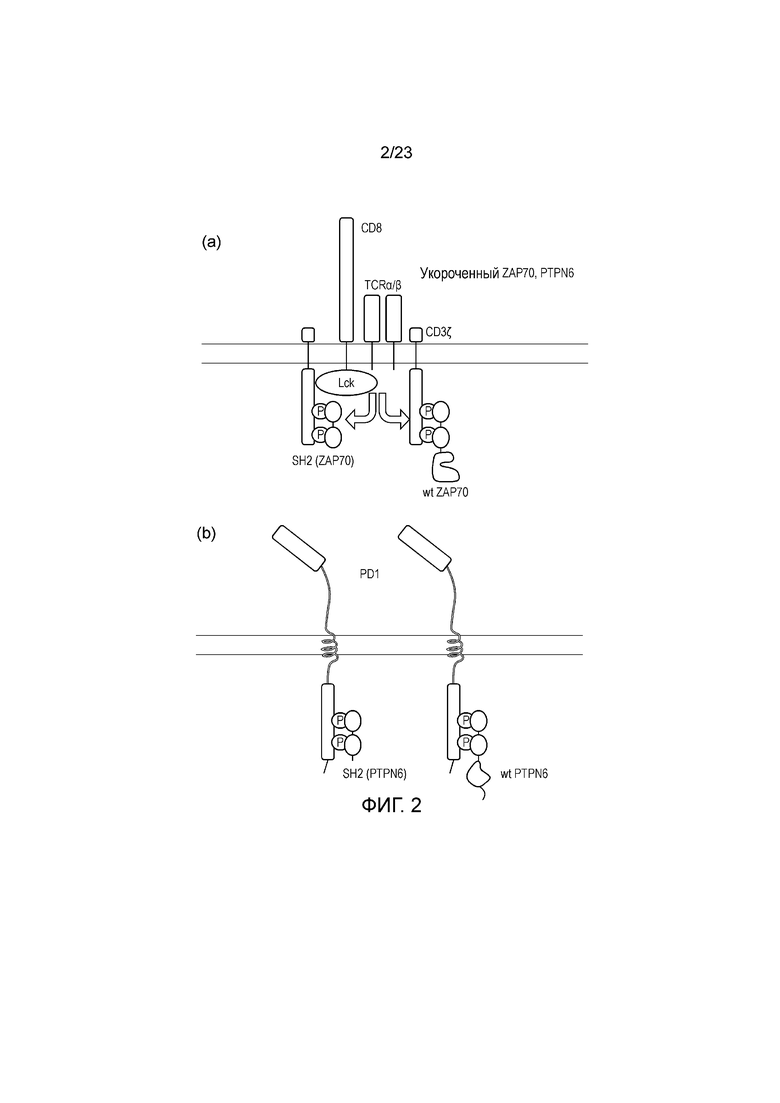
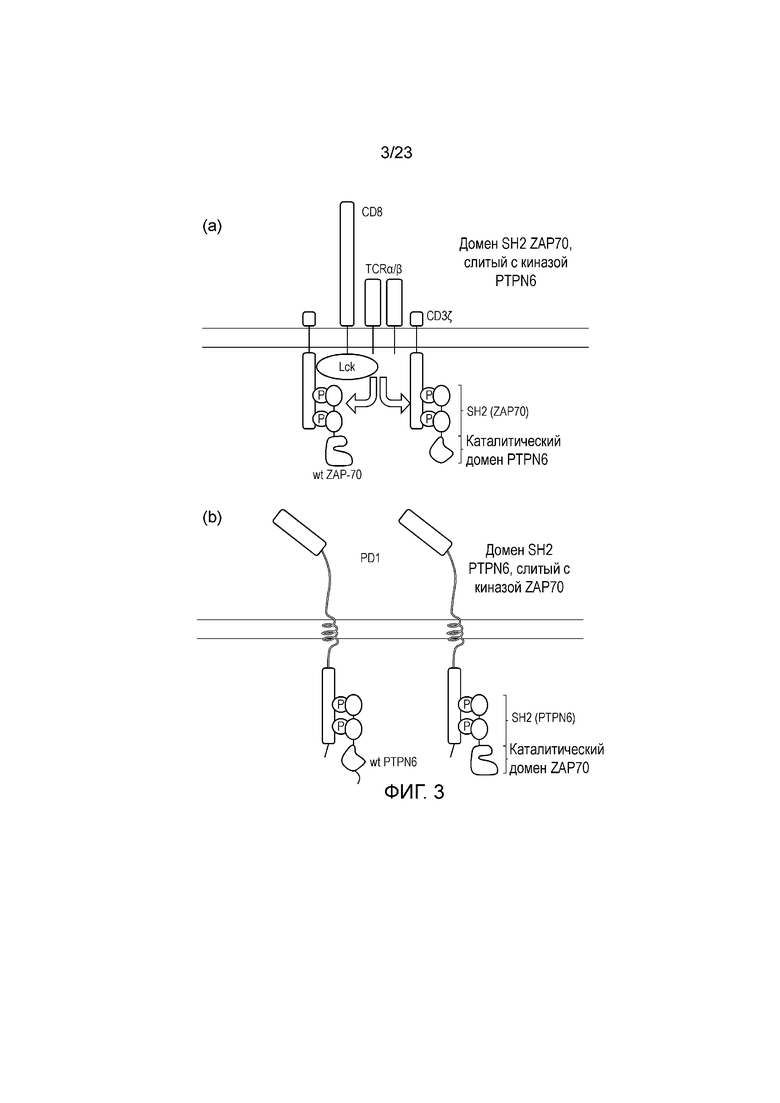
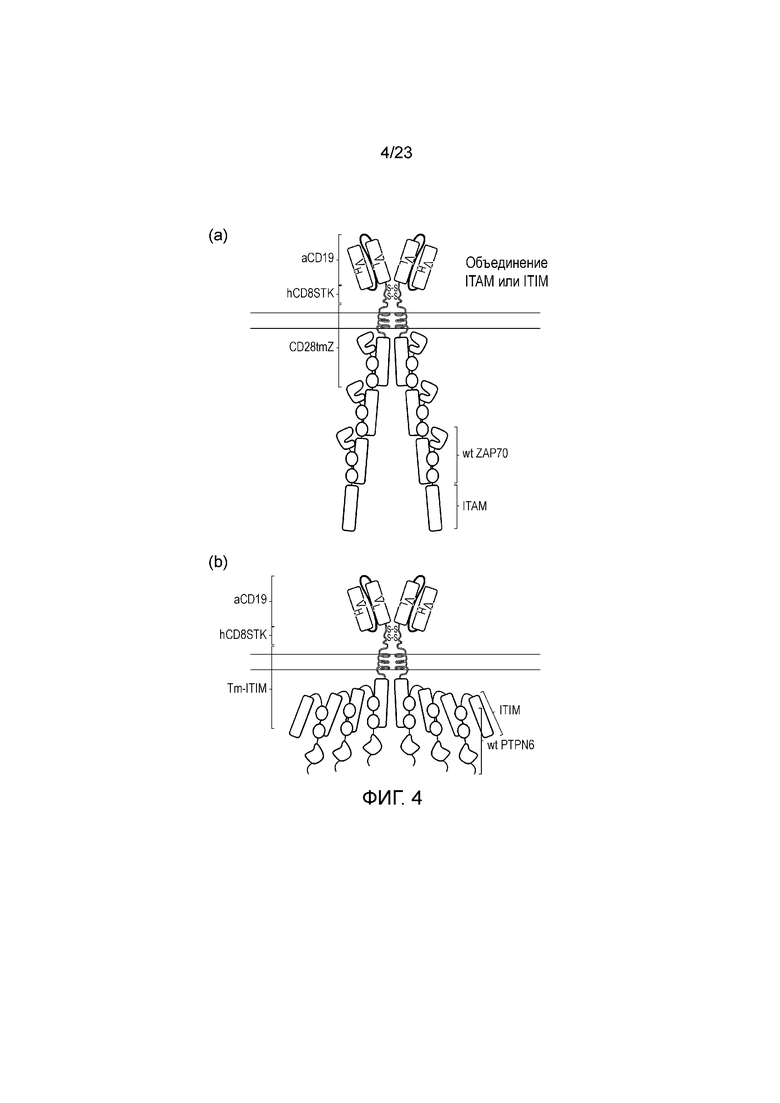
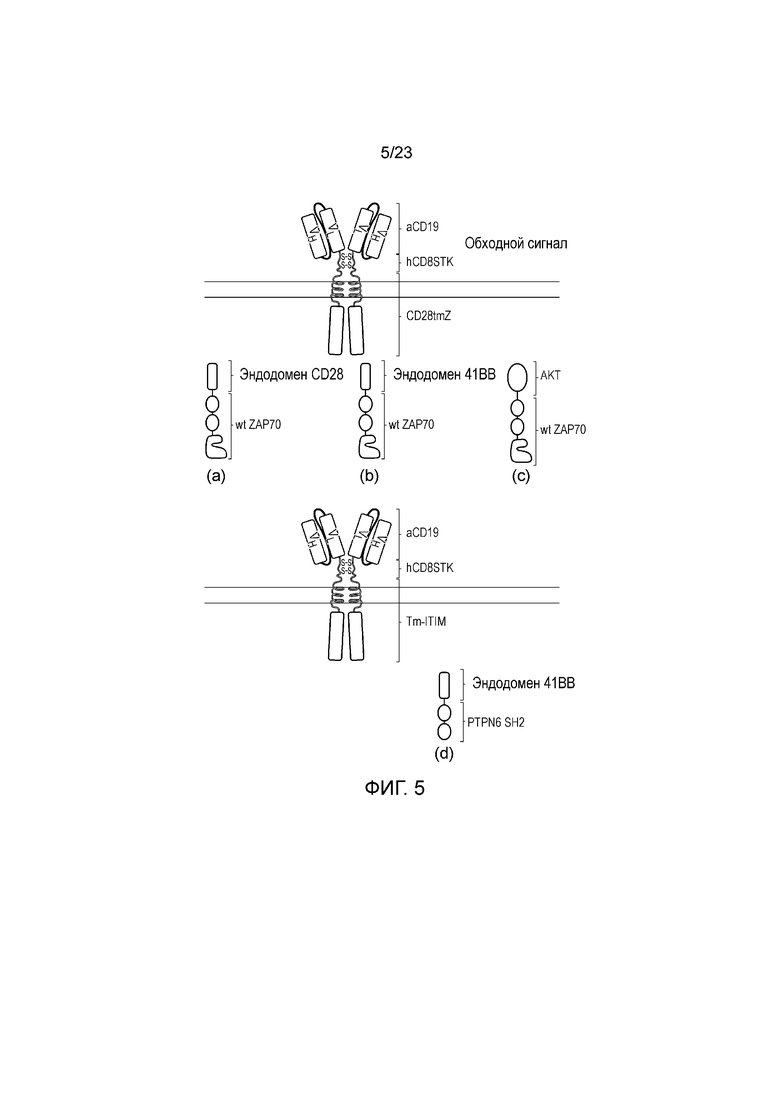
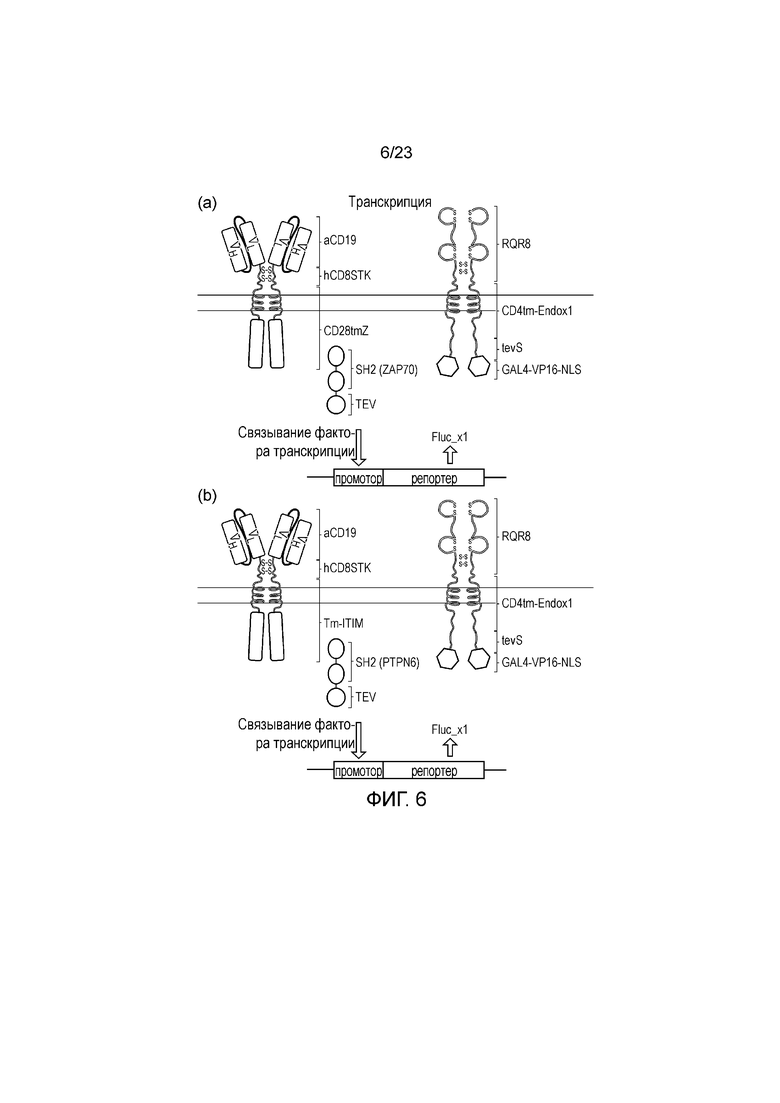
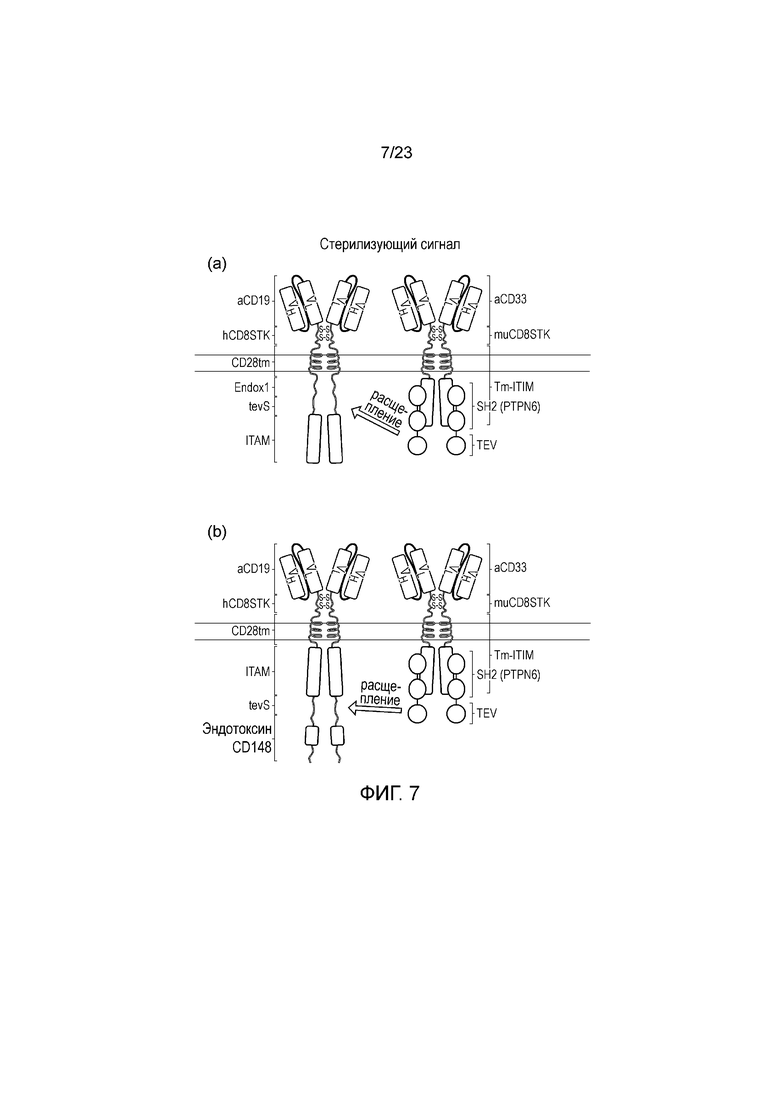
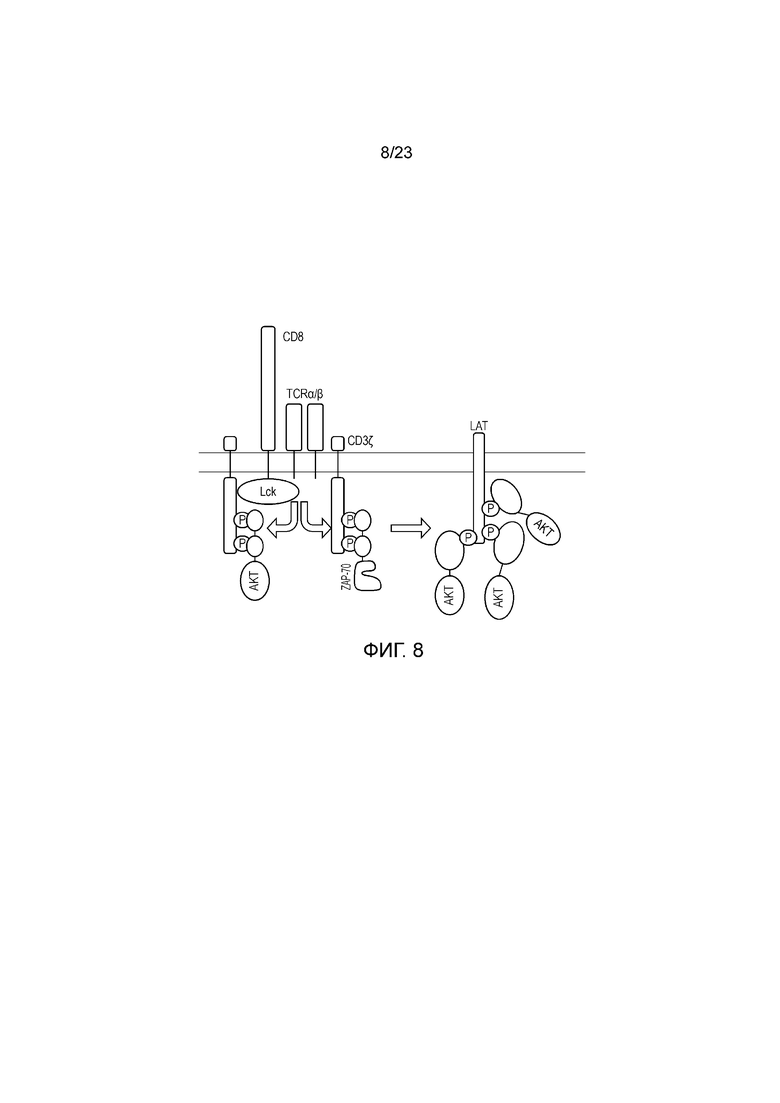
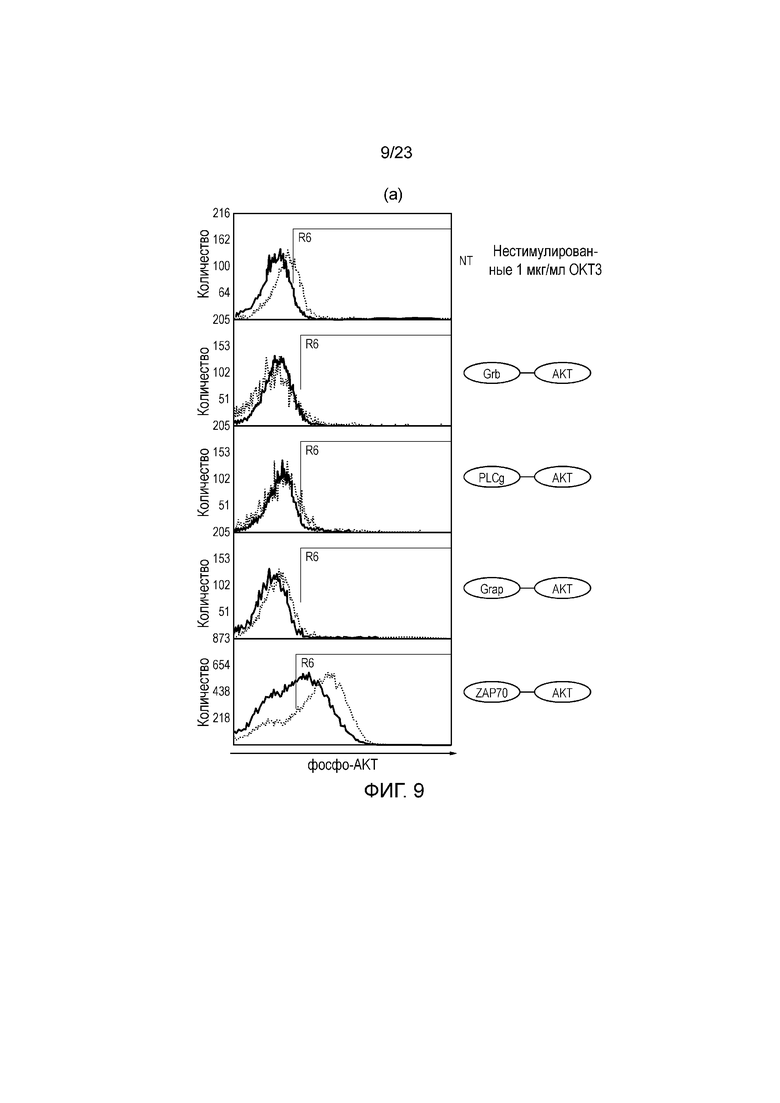
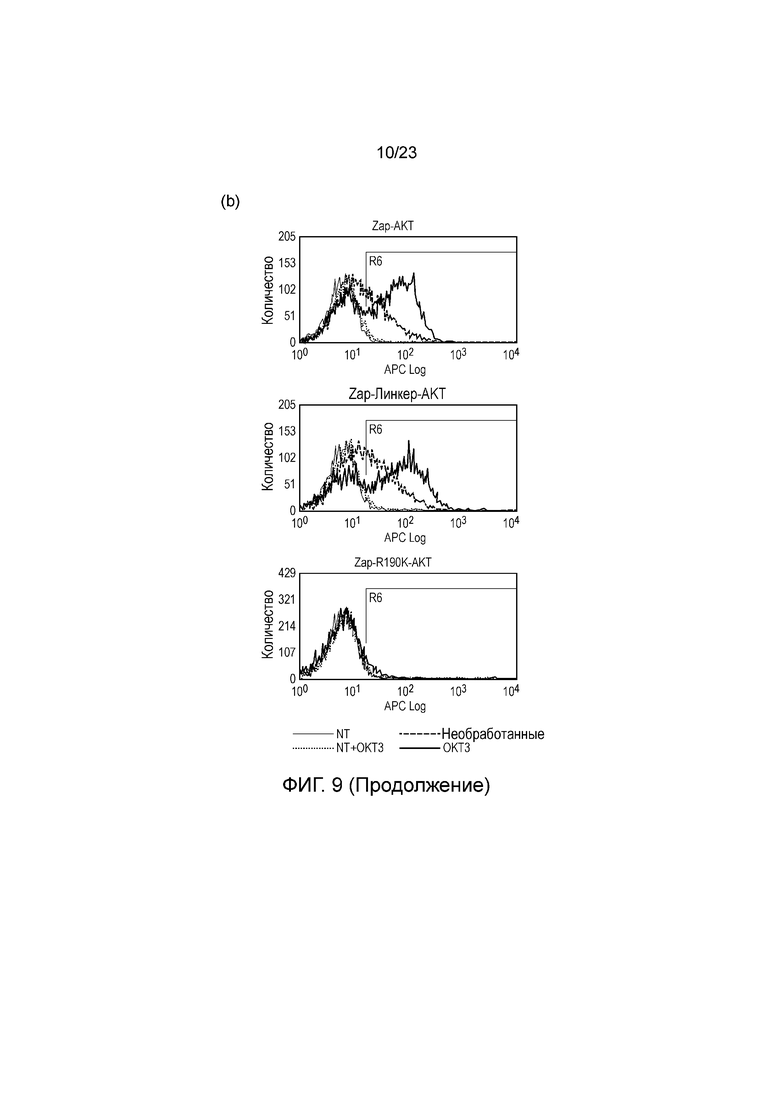
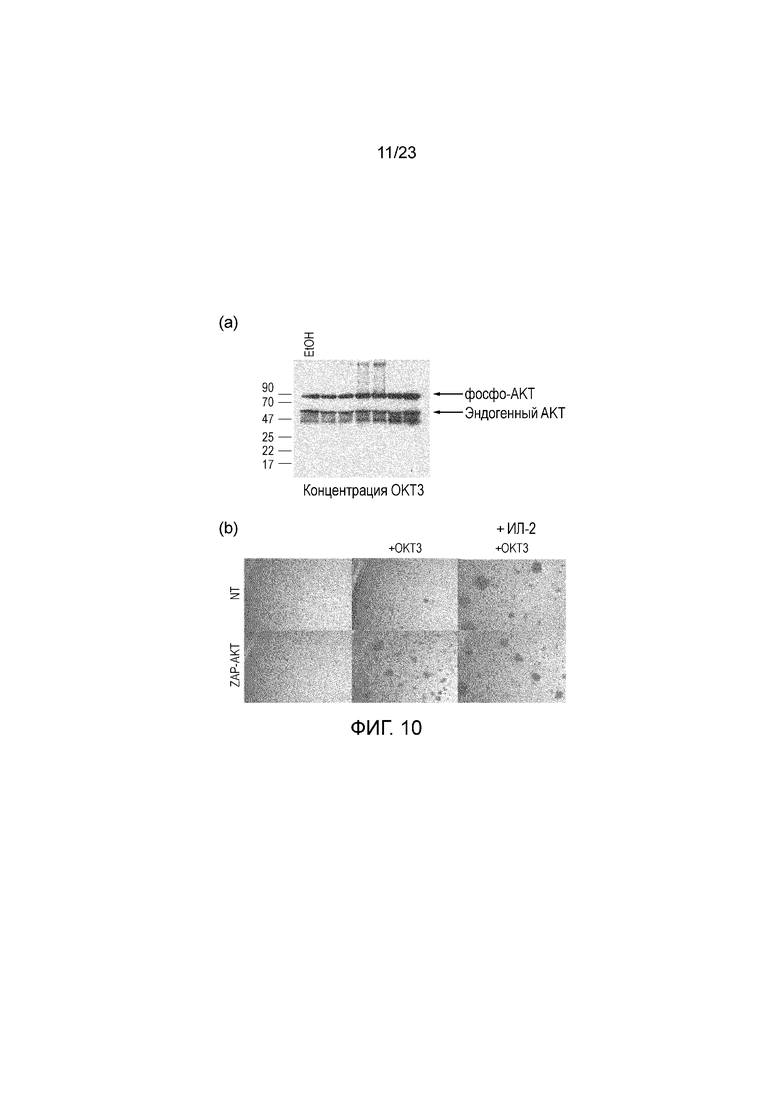
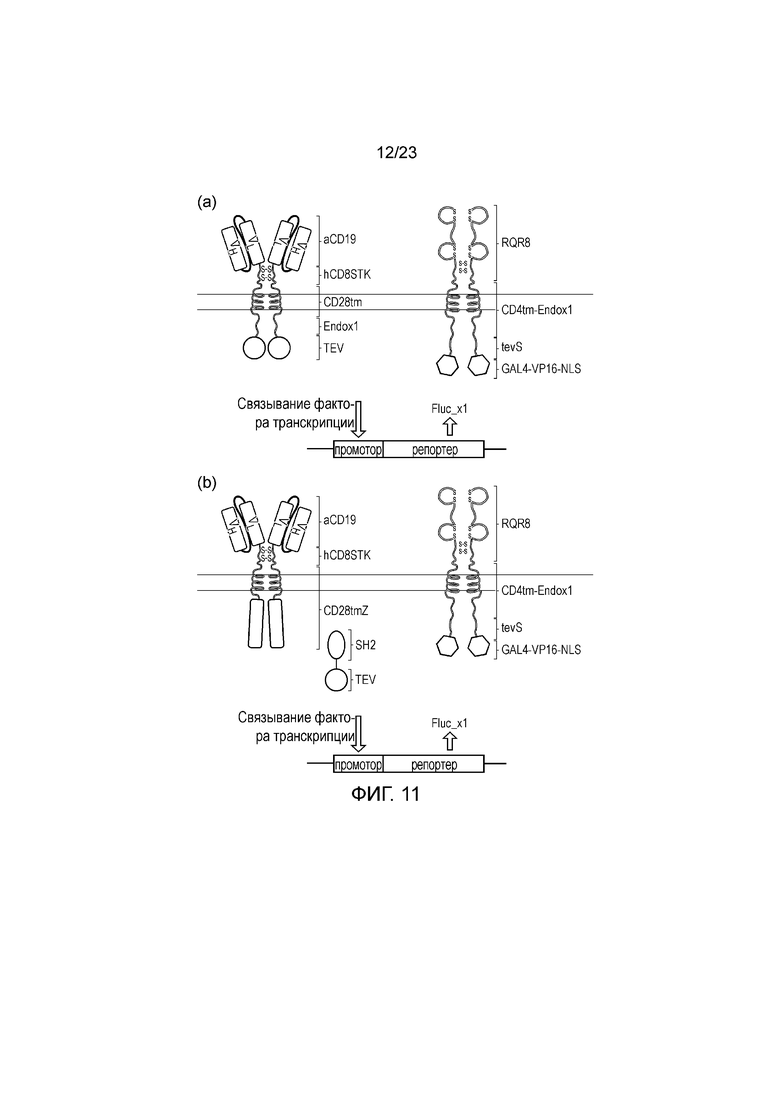
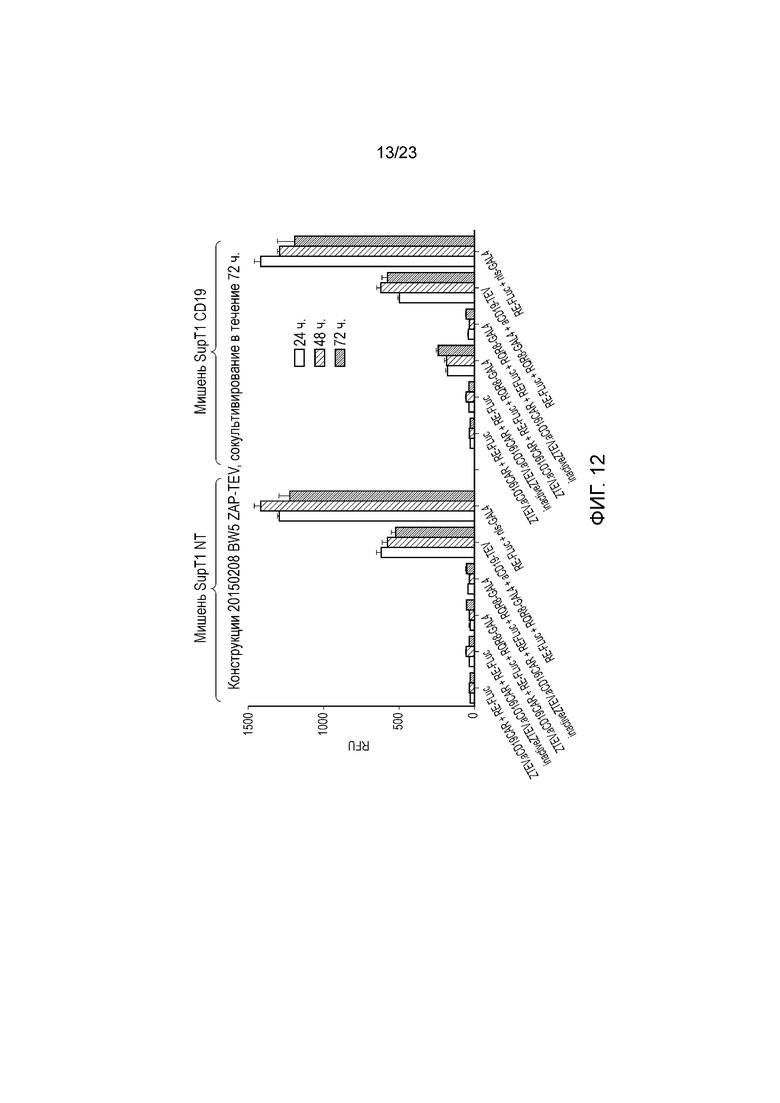
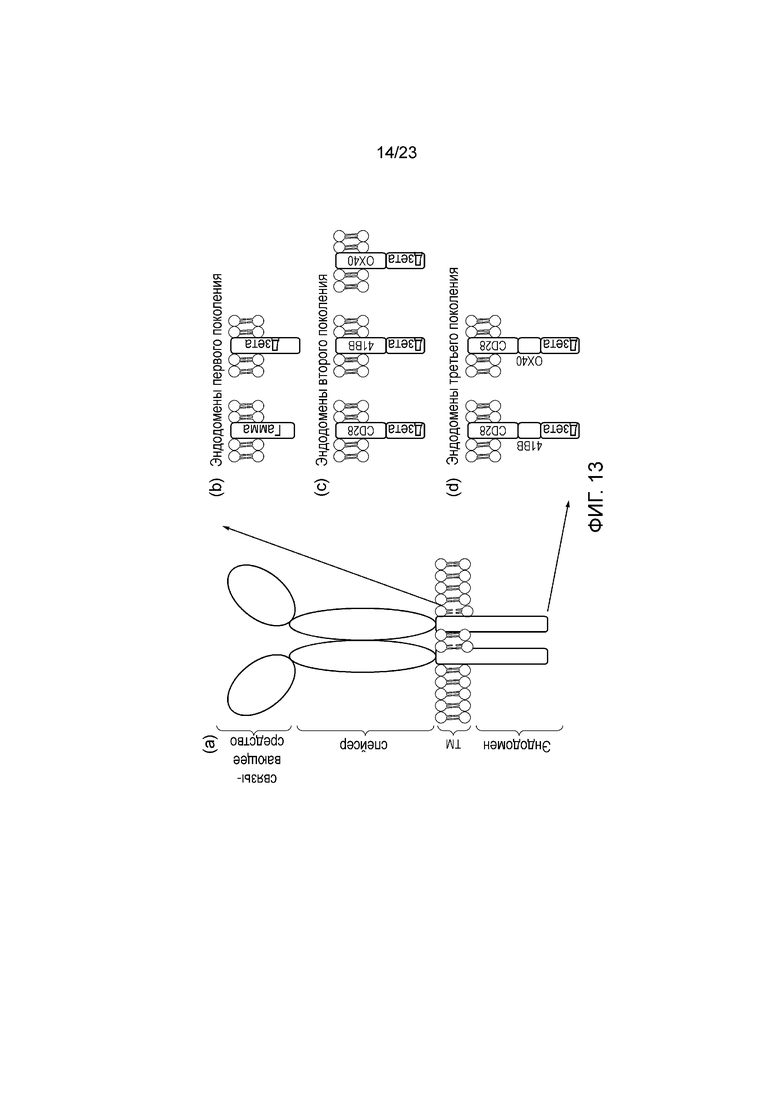







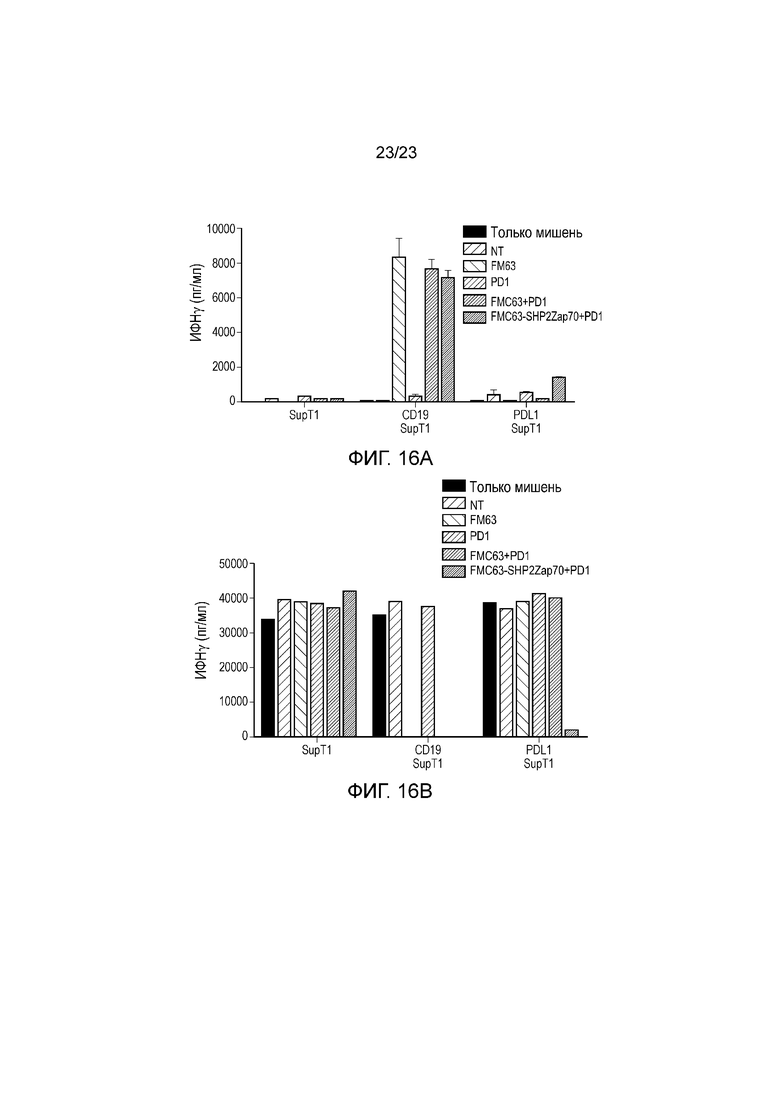
Изобретение относится к области биохимии, в частности к цитолитической иммунной клетке, способной уничтожать клетки-мишени, экспрессирующие антиген-мишень, а также к способу ее получения. Также раскрыта композиция, содержащая множество вышеуказанных клеток, а также ее применение в способе лечения злокачественного новообразования. Изобретение также относится к конструкции нуклеиновой кислоты, содержащей: первую последовательность нуклеиновой кислоты, кодирующую химерный антигенный рецептор, который связывает указанный антиген-мишень; и вторую последовательность нуклеиновой кислоты, кодирующую укороченный белок PTPN6 или SHP-2, который содержит один или оба домена SH2, но в котором отсутствует фосфатазный домен, а также к вектору, содержащему вышеуказанную конструкцию. Изобретение позволяет эффективно осуществлять эффективное лечение злокачественных новообразований с длительным сроком действия. 9 н. и 3 з.п. ф-лы, 16 ил., 6 табл., 4 пр.
1. Цитолитическая иммунная клетка, способная уничтожать клетки-мишени, экспрессирующие антиген-мишень, где указанная иммунная клетка содержит: химерный антигенный рецептор (CAR), который связывает указанный антиген-мишень; и укороченный белок PTPN6 или SHP-2, который содержит один или оба домена SH2, но в котором отсутствует фосфатазный домен.
2. Конструкция нуклеиновой кислоты для получения клетки по п.1, содержащая:
первую последовательность нуклеиновой кислоты, кодирующую химерный антигенный рецептор, который связывает указанный антиген-мишень; и
вторую последовательность нуклеиновой кислоты, кодирующую укороченный белок PTPN6 или SHP-2, который содержит один или оба домена SH2, но в котором отсутствует фосфатазный домен.
3. Вектор для получения клетки по п.1, содержащий конструкцию нуклеиновой кислоты по п.2.
4. Вектор по п.3, который представляет собой ретровирусный или лентивирусный вектор.
5. Набор векторов для получения клетки по п.1, содержащий первый вектор, который содержит первую последовательность нуклеиновой кислоты, кодирующую химерный антигенный рецептор; и второй вектор, который содержит вторую последовательность нуклеиновой кислоты, кодирующую укороченный белок PTPN6 или SHP-2, который содержит один или оба домена SH2, но в котором отсутствует фосфатазный домен.
6. Набор векторов по п.5, в котором указанные первый и второй векторы представляют собой ретровирусные или лентивирусные векторы.
7. Фармацевтическая композиция для лечения злокачественного новообразования, содержащая множество клеток по п.1, которые содержат CAR, связывающий антиген-мишень, который экспрессирует клетка злокачественного новообразования, и фармацевтически приемлемый носитель, дилюент или эксципиент.
8. Способ лечения злокачественного новообразования, включающий стадию введения индивидууму фармацевтической композиции по п.7.
9. Способ лечения злокачественного новообразования, включающий следующие стадии:
(i) отбор образца, содержащего цитолитическую иммунную клетку, у индивидуума;
(ii) трансдукцию или трансфекцию указанных иммунных клеток с использованием конструкции нуклеиновой кислоты по п.2, вектора по п.3 или 4 или набора векторов по п.5 или 6, где указанный антиген-мишень представляет собой антиген, который экспрессирует клетка злокачественного новообразования; и
(iii) введение указанному индивидууму иммунных клеток из (ii).
10. Применение фармацевтической композиции по п.7 в производстве лекарственного средства для лечения злокачественного новообразования.
11. Способ получения клетки по п.1, включающий стадию введения конструкции нуклеиновой кислоты по п.2, вектора по п.3 или 4 или набора векторов по п.5 или 6 в указанную клетку.
12. Способ по п.11, согласно которому указанная клетка получена из образца, выделенного из индивидуума.
| WO 2015075468 A1, 28.05.2015 | |||
| WO 9640199 A1, 19.12.1996 | |||
| WO 9731113 A1, 28.08.1997 | |||
| NORTHROP J P ET AL, Characterization of the roles of SH2 domain-containing proteins in T-lymphocyte activation by using dominant negative SH2 domains, MOLECULAR AND CELLULAR BIOLOGY, 1996, vol | |||
| Устройство для электрической сигнализации | 1918 |
|
SU16A1 |
| Кипятильник для воды | 1921 |
|
SU5A1 |
| Т-КЛЕТКИ С ТРАНСДУЦИРОВАННЫМ В НИХ АНТИГЕНОМ, ПРИМЕНЯЕМЫЕ В КАЧЕСТВЕ СИСТЕМЫ ДОСТАВКИ АНТИГЕНОВ | 2003 |
|
RU2330884C2 |

