Область техники, к которой относится изобретение
Настоящее изобретение относится к антагонистам CXCL13 и способу применения антагонистов CXCL13 для лечения легочных нарушений, симптомов и состояний, а также родственных заболеваний и состояний. Более конкретно, изобретение относится к способам лечения таких заболеваний путем применения антагонистов CXCL13 отдельно или вместе с антагонистами TNFα, таких как интерферирующие РНК, ДНКзимы и антитела против CXCL13, включая определенные участки или варианты, специфические, по меньшей мере, в отношении одного белка или его фрагмента, в количестве, эффективном для ингибирования активности CXCL13. Настоящее изобретение также относится к способу применения антагонистов CXCL13 и антагонистов TNFα для лечения животного с другими воспалительными заболеваниями, такими как системная красная волчанка.
Уровень техники
Астма представляет собой комплексное, хроническое заболевание с генетическим и природным компонентом (1). Оно характеризуется обратимой обструкцией дыхательных путей, гиперреактивностью дыхательных путей, воспалением и ремоделированием дыхательных путей (2). Установлено, что астмой страдает 15 миллионов американцев, и заболеваемость и смертность, ассоциированные с ней, возрастают в промышленно развитых странах (3, 4). Воспаление дыхательных путей у астматиков-аллергиков связано с инфильтрацией на слизистой субпопуляции CD4+ Т-клеток, относящихся к Т-хелперным клеткам (Th)2, и эозинофилов (5, 6). Взаимодействие между данными клетками приводит к продукции различных медиаторов провоспаления, вовлеченных в патогенез астмы (7, 8). Другими формами астмы являются такие формы, которые возникают в результате больших нагрузок, воздействия вирусов, приема аспирина и у людей определенных профессий. Несмотря на то, что в механизме, ответственном за развитие данных форм астмы, могут принимать участие Th2-лимфоциты и цитокины, он может «запускаться» различным образом (9-12). Многие цитокины и хемокины вовлечены в патогенез астмы (13, 14). В частности, цитокины Th2-клеток (интерлейкин 4, 5, 9 и 13) играют важную роль в развитии аллергических заболеваний, включая астму.
Хроническое обструктивное заболевание легких (COPD) является хроническим воспалением легких, характеризующимся инфильтрацией нейтрофилов, макрофагов, В- и Т-клеток. Данные иммунокомпетентные клетки активируются под воздействием различных цитокинов и хемокинов, которые высвобождаются в легких в ответ на длительное воздействие токсических газов и частиц (15). Бронхит и эмфизема, наряду с необратимой обструкцией дыхательных путей, являются клиническими проявлениями заболевания. Отсутствуют известные средства, которые замедляли бы быструю потерю функции легких, характерную для COPD.
Недавно было установлено, что прогрессирование COPD тесно связано с паренхимальной инфильтрацией естественными и адаптивными иммунными клетками воспаления, образующими эктопические лимфоидные фолликулы, содержащие герминативный центр. Наличие лимфоидных фолликулов связано с процессом ремодуляции, в результате которого утолщаются стенки дистальных малых отделов дыхательных путей (16). На основании этих данных можно с уверенностью предположить о возможной патологической роли эктопических лимфоидных фолликулов в развитии COPD.
При попытке выявить новые гены, вовлекаемые в патогенез астмы, исследователи использовали технологию ДНК-микроматриц для определения профиля генов, различным образом экспрессирующихся на животных моделях астмы (17, 18). Технология микроматриц представляет собой эффективный инструмент, поскольку она позволяет провести анализ экспрессии тысяч генов одновременно, а также ее можно автоматизировать с получением высокопроизводительного формата. При многофакторных заболеваниях, таких как астма, результаты анализа с помощью микроматриц могут обеспечить профиль экспрессии генов, который может оказаться очень полезным при разработке новых лекарственных препаратов. Также было показано, что она является очень действенной для идентификации новых генов и получения информации для генов с неизвестной функцией (19).
CXCL13 (также известный как BLC (хемокин хоминга В-клеток) или ВСА-1 (хемокин 1-аттрактант В-клеток) или Angie 2)) представляет собой хемотактический фактор, который является наиболее сильным и избирательным аттрактантом В-клеток. Он также способствует миграции некоторых Т-клеток и макрофагов через рецептор CXCR5 (20). CXCL13 экспрессируется в фолликулах пейеровских бляшек, селезенки и лимфатических узлов, и полагают, что он имеет значение для развития и гомеостаза фолликулов (21).
В течение ряда лет наблюдали, что в местах хронического воспаления структура воспалительного инфильтрата (Т-, В-клетки и стромальные клетки) имеет общие структурные признаки с лимфоидной тканью, при этом образуются так называемые эктопические лимфоидные фолликулы (21). Кроме того, высокая эктопическая продукция CXCL13 ассоциирована со скоплением лимфоцитов и образованием эктопических лимфоидных фолликулов при хронических воспалительных заболеваниях, таких как ревматоидный артрит (21), синдром Шенгрена (22), различные формы волчанки, такие как системная красная волчанка (23, 24), язвенный колит (25, 26), рассеянный склероз (27-29), диабет 1 типа (30-32) и аутоиммунные заболевания щитовидной железы (33, 34). Несмотря на то, что точная патогенетическая роль эктопических лимфоидных фолликулов не ясна, имеются данные, на основании которых можно предположить о значении последних в переходе из острого в хроническое, устойчивое воспаление за счет локального скопления лимфоцитов в воспаленной ткани (35). Следовательно, разрушение или элиминация эктопических лимфоидных фолликулов будет обеспечивать новый лечебный подход в отношении воспалительных заболеваний. CXCL13 представляет идеальную модель для терапии за счет его высокой экспрессии в эктопических лимфоидных фолликулах и его роли в поддержании микроструктуры и привлечении В-клеток.
Системная красная волчанка (SLE или волчанка) представляет собой хроническое аутоиммунное заболевание, которое потенциально является разрушающим и в некоторых случаях фатальным, поскольку иммунная система атакует собственные клетки и ткани организма, приводя к развитию воспаления и повреждению тканей. SLE может поражать любой орган в организме, но чаще всего страдают сердце, суставы, кожа, легкие, кровеносные сосуды, печень, почки и нервная система.
Ген CXCL13 (номер доступа в GenBank No. NM_006419, SEQ ID NO:1) находится в хромосоме 4q21 человека. CXCL13 относится к семейству хемокинов СХС. CXCL13 имеет ключевое значение для формирования/развития лимфоидных органов, образования фолликулов из В-клеток и рекрутинга В-клеток. Он эктопически продуцируется на высоком уровне в воспаленных тканях при многих хронических воспалительных заболеваниях, и полагают, что он играет важную роль в поддержании локальной активации В- и Т-клеток и воспаления.
Экспрессию генов можно модулировать несколькими различными путями, включая применение миРНК (siRNA), мшРНК (shRNA), антисмысловых молекул и ДНКзимов. Как миРНК, так и мшРНК функционируют через путь, связанный с РНКи (RNAi), и их успешно используют для подавления экспрессии генов. Вначале РНКи открыли в гельминтах, и сообщение Fire и Mello о явлении «сайленсинга» гена, связанного с дсРНК (dsRNA), первоначально касалось растений, и полагали, что оно представляет путь борьбы с инфекцией, вызванной РНК-содержащими вирусами, у растительных клеток. На данном пути длинный вирусный продукт дсРНК процессируется в более мелкие фрагменты размером 21-25 п.о. под действием DICER-подобного фермента, и затем двухцепочечная молекула разворачивается и превращается в РНК-индуцированный сайленсинг-комплекс (RISC). Аналогичный путь был выявлен в клетках млекопитающих с заметной разницей, заключающейся в том, что размер молекул дсРНК должен быть менее чем 30 п.о., чтобы избежать индукции так называемого ответа интерферона, который не является ген-специфическим и приводит к полному прекращению синтеза белка в клетке.
Можно сконструировать синтетические миРНК, конкретно нацеленные на один ген, и их можно легко ввести в клетки in vitro или in vivo. мшРНК являются ДНК-эквивалентами молекул миРНК, и они обладают тем преимуществом, что включаются в геном клеток и затем реплицируются во время каждого митотического цикла.
ДНКзимы также использовали для модуляции экспрессии гена. ДНКзимы представляют собой каталитические молекулы ДНК, расщепляющие одноцепочечную РНК. Они являются высоко избирательными для последовательности РНК-мишени, и в результате этого их можно использовать для снижения экспрессии определенных генов посредством направленного воздействия на матричную РНК.
Следовательно, имеется потребность в идентификации и характеристике новых способов диагностики и лечения, связанных с CXCL13, для легочных нарушений, таких как астма, и связанных с заболеваниями и состояниями. Кроме того, существует потребность в идентификации и характеристике новых способов лечения заболеваний, таких как системная красная волчанка.
Сущность изобретения
Настоящее изобретение относится к агонистам и/или антагонистам CXCL13 или его рецептора, CXCR5, и/или одному или обоим их действиям (далее «антагонисты CXCL13») и способу применения антагонистов CXCL13, включая антитела против CXCL13 и их определенные участки или варианты, специфические, по меньшей мере, для одного белка CXCL13 или его фрагмента, для лечения связанных с легкими заболеваний. Данные антагонисты CXCL13 можно вводить вместе с антагонистами TNFα, такими как антитела к TNFα, например, инфликсимаб и другие. Антагонист CXCL13, такой как моноклональное антитело, подавляет локальный рекрутинг В- и Т-клеток и последующую активацию с обеспечением новой стратегии для борьбы с хроническими воспалительными заболеваниями, опосредованными иммунной системой.
В одном варианте осуществления антагонист CXCL13 представляет собой антитело, которое специфически связывается с CXCL13 или его рецептором. Конкретное преимущество таких антител заключается в том, что они способы связываться с CXCL13 или его рецептором таким образом, что предотвращается их действие. Таким образом, в способе по настоящему изобретению используются антитела, обладающие желательным нейтрализующим свойством, которое делает их идеально подходящими для медикаментозной и профилактической терапии болезненных состояний, ассоциированных с различными нарушениями, связанными с легкими, у человека и не относящихся к человеку пациентов. Следовательно, настоящее изобретение относится к способу лечения связанного с легкими заболевания или состояния у пациента, нуждающегося в таком лечении, который включает введение пациенту количества нейтрализующего CXCL13 антитела для подавления связанного с легкими заболевания или состояния.
В другом аспекте изобретение относится к способам модуляции активности CXCL13 или его рецептора, включающим контактирование клетки со средством (например, антагонистом или агонистом), которое модулирует (ингибирует или стимулирует) активность или экспрессию CXCL13 или его рецептора таким образом, что активность или экспрессия в клетке модулируется. В предпочтительном варианте осуществления таким средством является антитело, которое специфически связывается с CXCL13 или его рецептором. В других вариантах осуществления модулятором является пептид, пептидомиметик или другая малая молекула.
В другом варианте осуществления настоящее изобретение относится к способу лечения связанного с легкими заболевания или состояния у пациента, нуждающегося в таком лечении, который включает введение пациенту количества нейтрализующего CXCL13 антитела или другого антагониста вместе с одним или несколькими антагонистами TNFα для ингибирования связанного с легкими заболевания или состояния.
Настоящее изобретение также относится к способам лечения субъекта, страдающего легочным или родственным заболеванием, в которых заболевание можно купировать модуляцией количества или активности CXCL13. Также настоящее изобретение относится к способам лечения субъекта, страдающего заболеванием, характеризующимся аномальной активностью CXCL13 или кодирующего его полинуклеотида, путем введения субъекту средства, которое является модулятором активности CXCL13 или модулятором экспрессии CXCL13.
В одном варианте осуществления модулятором является полипептид или соединение с малой молекулой. В другом варианте осуществления модулятором является полинуклеотид. В конкретном варианте осуществления антагонист CXCL13 представляет молекулу миРНК, молекулу мшРНК или ДНКзим, способные предупреждать продукцию CXCL13 клетками.
Другим аспектом изобретения является способ лечения животного с системной красной волчанкой, включающий введение антагониста CXCL13 животному и введение антагониста TNF-альфа животному; в котором каждый антагонист вводят в количестве, эффективном для того, чтобы вызвать ослабление симптома системной красной волчанки у животного. В одном варианте осуществления данного способа антагонист CXCL13 представляет собой связывающееся с CXCL13 антитело или связывающийся с CXCL13 фрагмент антитела, и антагонист TNF-α представляет собой связывающееся с TNF-α антитело или связывающийся с TNF-α фрагмент антитела.
В другом варианте осуществления данного способа животное представляет собой млекопитающее. В другом варианте осуществления данного способа млекопитающим является человек. В еще одном варианте осуществления данного способа вводимое количество каждого антитела или связывающего фрагмента антитела составляет от примерно 25 мг на кг массы тела животного до примерно 40 мг на кг массы тела животного. В еще одном варианте осуществления данного способа симптомом системной красной волчанки является количество периартериальных очагов лимфоцитарных инфильтратов, обнаруженных при исследовании тканей почек. В еще одном варианте осуществления данного способа симптомом системной красной волчанки является соотношение общего белка в моче к общему креатинину в моче. В еще одном варианте осуществления данного способа антагонист CXCL13 представляет собой связывающееся с CXCL13 антитело или связывающийся с CXCL13 фрагмент антитела, и антагонист TNF-α представляет собой связывающееся с TNF-α антитело или связывающийся с TNF-α фрагмент антитела. В еще одном варианте осуществления данного способа антагонист CXCL13 представляет собой связывающееся с CXCL13 антитело или связывающийся с CXCL13 фрагмент антитела, и антагонист TNF-α представляет собой связывающееся с TNF-α антитело инфликсимаб.
Настоящее изобретение дополнительно обеспечивает любое изобретение, описанное в данном документе.
Краткое описание фигур
На фигуре 1 показано, что уровни транскриптов мРНК CXCL13 повышены в пораженных тканях легких.
На фигуре 2 показано, что совместное лечение моноклональными антителами, специфичными для TNF-α и CXCL13, ослабляет симптомы заболевания легких, как определено по размеру эктопических фолликулов в пораженных тканях легких.
На фигуре 3 показано, что совместное лечение моноклональными антителами, специфичными для TNF-α и CXCL13, уменьшает инфильтрацию В-клеток, экспрессирующих CD22.2, в ткани легких.
На фигуре 4 показано, что совместное лечение моноклональными антителами, специфичными для TNF-α и CXCL13, уменьшает инфильтрацию В-клеток, экспрессирующих CD19, в ткани легких.
На фигуре 5 показано, что совместное лечение моноклональными антителами, специфичными для TNF-α и CXCL13, уменьшает инфильтрацию В-клеток, экспрессирующих CD45R/B220, в ткани легких.
На фигуре 6 показано, что совместное лечение моноклональными антителами, специфичными для TNF-α и CXCL13, уменьшает инфильтрацию В-клеток, экспрессирующих CD4, в ткани легких.
На фигуре 7 показано, что совместное введение mAb, специфичных для CXCL13 и TNF-α, снижает связанное с гломерулонефритом содержание белка в моче мышей NZB/W F1 с симптомами системной красной волчанки (SLE) до уровней ниже, чем у не подвергавшихся лечению контрольных мышей NZB/W F1, которые получали только забуференный фосфатом физиологический раствор (PBS).
На фигуре 8 показано, что совместное введение mAb, специфичных для CXCL13 и TNF-α, снижает тяжесть заболевания почек, ассоциированного с системной красной волчанкой (SLE), у мышей NZB/W F1 с симптомами SLE до уровня ниже, чем у не подвергавшихся лечению контрольных мышей NZB/W F1, которые получали только раствор PBS.
Подробное описание изобретения
Определения
Последующие определения приведены для иллюстрации и определения значения и объема различных терминов, используемых для описания настоящего изобретения.
«Активность», биологическая активность и функциональная активность полипептида относится к активности, проявляемой CXCL13 или его рецептором в ответ на специфическое взаимодействие с другим белком или молекулой, как определено in vivo, in situ или in vitro, согласно стандартным методикам. Такие виды активности могут быть прямой активностью, такой как ассоциация с ферментативной активностью или ферментативная активность на втором белке, или опосредованная активность, такая как клеточный процесс, опосредованный взаимодействием белка со вторым белком или серией взаимодействий в виде внутриклеточной передачи сигналов или каскада процесса коагуляции.
«Антитело» включает любой полипептид или пептид, содержащий молекулу, которая включает, по меньшей мере, область молекулы иммуноглобулина, такую как, без ограничения, по меньшей мере, одна определяющая комплементарность область (CDR) тяжелой или легкой цепи, или его связывающую лиганд область, вариабельную область тяжелой или легкой цепи, константную область тяжелой или легкой цепи, каркасную область, или любой их участок, фрагмент или вариант. Термин «антитело» дополнительно предназначен для включения антител, фрагментов, полученных в результате расщепления, их определенных участков и вариантов, включающих миметики антител или содержащих участки антител, которые имитируют структуру и/или функцию антитела или его специфического фрагмента или участка, включая одноцепочечные антитела, антитела с одним доменом и их фрагменты. Например, фрагменты антител включают, но не ограничиваются ими, Fab (например, полученные путем расщепления папаином), Fab' (например, полученные путем расщепления пепсином и неполного восстановления) и F(ab')2 (например, полученные путем расщепления пепсином), facb (например, полученные путем расщепления плазмином), pFc' (например, полученные путем расщепления пепсином или плазмином), Fd (например, полученные путем расщепления пепсином, неполного восстановления и реагрегации), Fv или scFv (например, полученные методами молекулярной биологии) (см., например, Colligan et al., eds. Current Protocols in Immunology, John Wiley & Sons, Inc., NY (1994-2001); Colligan et al., eds. Current Protocols in Polypeptide Science, John Wiley & Sons, Inc., NY (1997-2001)).
«Химерные» или «слитые» молекулы представляют нуклеиновые кислоты или полипептиды, которые получены объединением, например, одного или нескольких антагонистов CXCL13 (или их фрагментов) с дополнительной нуклеиновокислотной последовательностью(ями). Такие комбинированные последовательности можно ввести в соответствующий вектор и экспрессировать с получением химерного или слитого полипептида.
«Комплемент» нуклеиновокислотной последовательности или «комплементарный» нуклеиновокислотной последовательности по изобретению относится к полинуклеотидной молекуле, имеющей комплементарную последовательность основания и обратную ориентацию по сравнению с первым полинуклеотидом.
«Фрагмент» является вариантом полипептида с аминокислотной последовательностью, которая полностью является такой же, как часть, но не вся аминокислотная последовательность антагониста CXCL13, или вариантом полинуклеотида с нуклеиновокислотной последовательностью, которая полностью является такой же, как часть, но не вся нуклеиновокислотная последовательность полинуклеотида антагониста CXCL13. Фрагменты могут включать, например, усеченные полипептиды или их варианты, такие как непрерывные ряды остатков, которые включают гетерологичную амино- и/или карбоксиконцевую аминокислотную последовательность. Также сюда входят антагонисты CXCL13, полученные в результате деградации, с использованием клетки-хозяина или в клетке-хозяине. Другие приведенные в качестве примеров фрагменты характеризуются структурными или функциональными признаками, такие как фрагменты, содержащие альфа-спираль или области, образующие альфа-спираль, бета-складки или области, образующие бета-складки, витки или области, образующие витки, спирали или области, образующие спирали, гидрофильные области, гидрофобные области, альфа-амфипатические области, бета-амфипатические области, гибкие области, образующие поверхность области, связывающие субстрат области, внеклеточные области и области с высоким показателем антигенности.
«Идентичность», как это известно в данной области, представляет взаимосвязь между двумя или более полипептидными последовательностями, или двумя или более полинуклеотидными последовательностями, как определено путем сравнения последовательностей. В данной области «идентичность» также означает степень сходства последовательностей между полипептидными или полинуклеотидными последовательностями, как определено по совпадению между полосами таких последовательностей. «Идентичность» и «гомологию» можно легко рассчитать известными способами, включая, но не ограничиваясь ими, способы, описанные в Computational Molecular Biology, Lesk, A.M., ed., Oxford University Press, New York, 1988; Biocomputing: Informatics and Genome Projects, Smith, D.W., Academic Press, New York, 1993; Computer Analysis of Sequence Data, Part I, Griffin, A.M. and Griffin, H.G., eds., Humana Press, New Jersey, 1994; Sequence Analysis in Molecular Biology, von Heinje, G.,, Academic Press, 1987; и Sequence Analysis Primer, Gribskov, M. and Devereux, J., M Stockton Press, New York, 1991; и Carillo, H. and Lipman, D., Siam J. Applied Math., 48:1073 (1988). Кроме того, можно установить процентную идентичность по данным выравнивания аминокислотной и нуклеотидной последовательности, полученных с использованием дефолтовых параметров для компонента AlignX Vector NTI Suite 8.0 (Informax, Frederick, MD).
Разработаны предпочтительные способы определения идентичности с получением максимального совпадения между тестируемыми последовательностями. Способы определения идентичности и гомологии кодифицированы в широко доступных компьютерных программах. Предпочтительные компьютерные программы для методов определения идентичности и гомологии между двумя последовательностями включают, но не ограничиваются ими, программу GCG (Devereux, J., et al., Nucleic Acids Research 12(1):387 (1984)); BLASTP, BLASTN и FASTA (Atschul, S.F. et al., J. Molec. Biol., 215:403-410 (1990)). Программа BLAST Х является общедоступной от NCBI и других источников (BLAST Manual, Altschul, S., et al., NCBINLM NIH Bethesda, Md. 20894: Altschul, S., et al., J. Mol. Biol., 215:403-410 (1990). Для определения идентичности можно использовать хорошо известный алгоритм Smith Waterman.
Предпочтительные параметры сравнения полипептидных последовательностей включают следующие:
(1) Алгоритм: Needleman and Wunsch, J. Mol. Biol., 48:443-453 (1970) Матрица сравнения: BLOSSUM62 от Hentikoff and Hentikoff, Proc. Natl. Acad. Sci, USA, 89:10915-10919 (1992)
Гэп штраф: 12
Длина штрафуемого гэпа: 4
Программа, пригодная для данных параметров, является широкодоступной в виде «гэп» программы Genetics Computer Group, Madison Wis. Указанные выше параметры являются дефолтовыми параметрами для сравнений пептидных последовательностей (с отсутствием штрафов для концевых гэпов).
Предпочтительные параметры сравнения полинуклеотидных последовательностей включают следующие:
(1) Алгоритм: Needleman and Wunsch, J. Mol. Biol. 48:443-453 (1970)
Матрица сравнения: совпадения=+10, ошибочные совпадения=0
Гэп штраф: 50
Длина штрафуемого гэпа: 3
Доступна в виде: «гэп» программы Genetics Computer Group, Madison Wis. Это дефолтовые параметры для сравнений нуклеиновокислотных последовательностей.
В качестве примера, полинуклеотидная последовательность может быть идентичной последовательности, которая на 100% является идентичной, или она может содержать определенное целое число изменений нуклеотидов по сравнению со ссылочной последовательностью. Такие изменения выбраны из группы, состоящей из делеции, замены, включая транзицию и трансверсию, или вставки, по меньшей мере, одного нуклеотида, и где изменения могут иметь место в 5'- или 3'-концевых положениях ссылочной нуклеотидной последовательности, или где угодно между данными концевыми положениями, расположенные либо индивидуально среди нуклеотидов в ссылочной последовательности, либо в одной или нескольких смежных группах в ссылочной последовательности. Число нуклеотидных изменений определяют умножением общего числа нуклеотидов в последовательности на численный процент соответствующей процентной идентичности (деленный на 100) и вычитанием полученного производного из общего числа нуклеотидов в последовательности, или:
n.sub.n.ltorsim.x.sub.n-(x.sub.n.y.),
где n.sub.n. представляет число нуклеотидных изменений, x.sub.n представляет общее число нуклеотидов в последовательности, и y равно, например, 0,70 для 70%, 0,80 для 80%, 0,85 для 85%, 0,90 для 90%, 0,95 для 95% и т.д., и где любое производное x.sub.n, не равное целому числу, и y округляют до наиболее близкого целого числа перед вычитанием из x.sub.n.
Изменения полинуклеотидной последовательности, кодирующей последовательность, могут привести к несмысловой, ошибочно смысловой мутации или мутации со сдвигом рамки считывания в данной кодирующей последовательности и, тем самым, к изменению полипептида, кодированного полинуклеотидом, после таких изменений. Аналогично, полипептидная последовательность может быть идентична ссылочной последовательности, которая на 100% является идентичной, или она может включать определенное целое число аминокислотных изменений по сравнению со ссылочной последовательностью, так что процентная идентичность составляет менее 100%. Такие изменения выбраны из группы, состоящей из делеции, замены, включая консервативную и неконсервативную замену, или вставки, по меньшей мере, одной аминокислоты, и где изменения могут иметь место в амино- или карбоксиконцевых положениях ссылочной полипептидной последовательности, или где угодно между данными концевыми положениями, расположенные либо индивидуально среди аминокислот в ссылочной последовательности, либо в одной или нескольких смежных группах в ссылочной последовательности. Число аминокислотных изменений для данной % идентичности определяют умножением общего числа аминокислот в последовательности на численный процент соответствующей процентной идентичности (деленный на 100) и вычитанием полученного производного из общего числа аминокислот в последовательности, или:
n.sub.а.ltorsim.x.sub.а-(x.sub.а.y.),
где n.sub.а. представляет число аминокислотных изменений, x.sub.а представляет общее число аминокислот в последовательности, и y равно, например, 0,70 для 70%, 0,80 для 80%, 0,85 для 85% и т.д., и где любое производное x.sub.а, не равное целому числу, и y округляют до наиболее близкого целого числа перед вычитанием из x.sub.а.
«Нуклеиновые кислоты» являются полимерами нуклеотидов, в которых нуклеотид содержит основание, связанное с сахаром, где сахара, в свою очередь, связаны друг с другом посредством, по меньшей мере, бивалентной молекулы, такой как фосфорная кислота. В природных нуклеиновых кислотах сахар представляет собой или 2'-дезоксирибозу (ДНК), или рибозу (РНК). Поли- или олигонуклеотиды, не относящиеся к природным, содержат модифицированные основания, сахара или связывающие молекулы, но, как правило, представляют собой миметики комплементарной природы естественных нуклеиновых кислот, по аналогии с которыми они сконструированы. Примером олигонуклеотида, не относящегося к природному, является антисмысловая молекула, которая имеет фосфоротиоатный остов. Как правило, «олигонуклеотид» относится к молекуле нуклеиновой кислоты, содержащей менее чем 30 нуклеотидов.
«Полипептид» является полимером из аминокислотных остатков, соединенных пептидными связями, и пептид, как правило, относится к аминокислотным полимерам из 12 или менее остатков. Пептидные связи могут быть получены естественным путем, т.е. с использованием нуклеиновокислотной матрицы, или синтетически с использованием способов, известных в данной области.
«Белок» является макромолекулой, содержащей одну или несколько полипептидных цепей. Белок может дополнительно содержать заместители, присоединенные к боковым группам аминокислот, но не участвующие в образовании пептидных связей. Обычно, белки, экспрессированные эукариотическими клетками, также содержат углеводы. Белки определяют в данном документе по их аминокислотной последовательности или остова, и заместители не указывают, известные или нет.
Термин «рецептор» означает молекулу, обладающую способностью оказывать влияние на биологическую активность, например, в клетке, в результате взаимодействия со специфическим лигандом или партнером по связыванию. Для связанных с клеточной мембраной рецепторов характерно наличие внеклеточного связывающего лиганд домена, одного или нескольких мембранных пространственных или трансмембранных доменов и внутриклеточного домена-эффектора, который, как правило, вовлечен в сигнальную трансдукцию. Связывание лиганда с клеточными мембранными рецепторами вызывает изменения во внеклеточном домене, которые передаются по клеточной мембране, прямое или опосредованное взаимодействие с одним или несколькими внутриклеточными белками, и изменяет свойства клеток, такие как ферментная активность, форма клетки или профиль экспрессии генов. Рецепторы также могут быть не связаны с клеточной поверхностью и могут быть цитозольными, ядерными или высвобождаемыми из клетки в целом. Не связанные с клеткой рецепторы называются растворимыми рецепторами.
Все публикации или патенты, цитируемые в данном документе, полностью включены в описание путем ссылки, специально или нет обозначенные соответственно, поскольку они демонстрируют уровень техники на время создания настоящего изобретения и/или обеспечивают описание и представление настоящего изобретения. Публикации относятся к любым научным или патентным публикациям, или любой другой информации, доступной в любом медийном формате, включая все записанные, электронные или печатные форматы. Последующие источники полностью включены в данный документ путем ссылки: Ausubel, et al., ed., Current Protocols in Molecular Biology, John Wiley & Sons, Inc., NY (1987-2001); Sambrook, et al., Molecular Cloning: A Laboratory Manual, 2nd Edition, Cold Spring Harbor, NY (1989); Harlow and Lane, antibodies, a Laboratory Manual, Cold Spring Harbor, NY (1989); Colligan, et al., eds., Current Protocols in Immunology, John Wiley & Sons, Inc., NY (1984-2001); Colligan et al., Current Protocols in Protein Science, John Wiley & Sons, NY (1987-2001).
Биологическая функция CXCL13
Были выявлены новая экспрессия и новая функция CXCL13, и его биологическая функция в формировании/развитии лимфоидных органов, образовании В-клеточных фолликулов и рекрутинге В-клеток при различных заболеваниях человека и на моделях животных. Впервые была установлена связь эктопической экспрессии с образованием лимфоидных фолликулов, ассоциированных с заболеваниями легких, в частности, но без ограничения, COPD.
Композиции CXCL13 могут содержать одну или несколько изоформ белка, их иммуногенные участки или полинуклеотиды, кодирующие такие участки. Альтернативно, терапевтическая композиция может содержать клетку, которая экспрессирует белок CXCL13, или Т-клетку, которая является специфической для клеток, экспрессирующих полипептид, кодированный геном, или другой тип агонистов; и антагонисты, такие как нейтрализующие моноклональные антитела (mAb), лекарственные препараты на основе нуклеиновых кислот, или соединения с малой молекулой к любому участку ДНК, РНК или белка CXCL13. Данные композиции можно применять, например, для профилактики и лечения ряда опосредованных иммунной системой воспалительных заболеваний. Также раскрываются диагностические и прогностические способы, основанные на детектировании белка CXCL13 или мРНК, кодирующей такой белок, в пробе.
CXCL13 и его рецепторные белки, полипептиды и молекулы нуклеиновых кислот, кодирующие их, включают семейство молекул с определенными консервативными структурными и функциональными свойствами. Каждая из этих молекул включена в определение CXCL13. Используемый в данном документе термин «семейство» относится к двум или более белкам или молекулам нуклеиновой кислоты, имеющим общую или аналогичную структуру доменов и обладающим достаточной идентичностью аминокислотных и нуклеотидных последовательностей, определенных в данном документе. Члены семейства могут происходить от одного или различных видов. Например, семейство может включать два или более белков человека, или может включать один или несколько белков человека, или один или несколько белков, не относящихся к человеческим.
Домен, который может присутствовать в белках CXCL13, представляет сигнальную последовательность. Используемый в данном документе термин «сигнальная последовательность» включает пептид длиной, по меньшей мере, примерно из 10 аминокислотных остатков, который находится на аминоконце связанных с мембраной белков и который содержит, по меньшей мере, примерно 45% гидрофобных аминокислотных остатков, таких как аланин, лейцин, изолейцин, фенилаланин, пролин, тирозин, триптофан или валин. В предпочтительном варианте осуществления сигнальная последовательность содержит, по меньшей мере, примерно от 10 до 35 аминокислотных остатков, предпочтительно примерно от 10 до 20 аминокислотных остатков, и имеет, по меньшей мере, примерно 35-60%, более предпочтительно 40-50% и еще более предпочтительно, по меньшей мере, примерно 45% гидрофобных остатков. Сигнальная последовательность служит для направления белка, содержащего такую последовательность, к липидному бислою. Таким образом, в одном варианте осуществления белок CXCL13 может содержать сигнальную последовательность. Данная сигнальная последовательность отщепляется во время процессинга зрелого белка.
Белки CXCL13 содержат внеклеточный домен. Используемый в данном документе термин «внеклеточный домен» относится к участку белка, который находится не с цитоплазматической стороны липидного бислоя клетки, когда нуклеиновая кислота, кодирующая белок, экспрессируется в клетке.
Кроме того, белок CXCL13 содержит трансмембранный домен. Используемый в данном документе термин «трансмембранный домен» относится к аминокислотной последовательности длиной, по меньшей мере, примерно из 15 аминокислотных остатков и которая содержит, по меньшей мере, примерно 65-70% гидрофобных аминокислотных остатков, таких как аланин, лейцин, фенилаланин, пролин, тирозин, триптофан или валин (Eric, et al., Proc. of Sixth Int. Conf. on Intelligent Systems for Molecular Biology, p. 175-182). В предпочтительном варианте осуществления трансмембранный домен содержит примерно 15-30 аминокислотных остатков, предпочтительно примерно 20-25 аминокислотных остатков, и имеет, по меньшей мере, примерно 60-80%, более предпочтительно 65-75% и еще более предпочтительно, по меньшей мере, примерно 70% гидрофобных остатков.
Белки CXCL13 содержат цитоплазматический домен, в частности, включая белки с карбоксиконцевым цитоплазматическим доменом. Используемый в данном документе термин «цитоплазматический домен» относится к участку белка, который находится с цитоплазматической стороны липидного бислоя клетки, когда нуклеиновая кислота, кодирующая белок, экспрессируется в клетке. Белки CXCL13 обычно содержат различные сайты потенциальных посттрансляционных модификаций (часто они находятся во внеклеточном домене).
Антагонисты CXCL13
Используемый в данном документе термин «антагонисты CXCL13» относится к соединениям, которые ингибируют или нейтрализуют биологическую активность CXCL13 или его рецептора, CXCR5. Такие антагонисты осуществляют это действие различными путями. Одна группа антагонистов CXCL13 связывается с белком CXCL13 с достаточной аффинностью и специфичностью для нейтрализации биологических эффектов CXCL13. В данную группу молекул входят антитела и фрагменты антител (например, такие молекулы, как F(ab) или F(ab')2). Другая группа антагонистов CXCL13 включает фрагменты белка CXCL13, мутеины или малые органические молекулы, т.е. пептидомиметики, которые связываются с CXCL13 или партнерами по связыванию с CXCL13, тем самым, подавляя биологическую активность CXCL13. Антагонист CXCL13 может относиться к любой из данных групп, при условии, что он является соединением, ингибирующим биологическую активность CXCL13. Антагонисты CXCL13 включают антитело к CXCL13, антитело к рецептору CXCL13, модифицированный CXCL13 и неполные пептиды CXCL13. Другая группа антагонистов CXCL13 включает миРНК (siRNA), мшРНК (shRNA), антисмысловые молекулы и ДНКзимы с направленным действием к последовательности гена CXCL13, известные в данной области и раскрытые в данном документе.
Следовательно, используемые в данном документе термины «антитело СХСL13», «антитело против CXCL13», «участок антитела против CXCL13» или «фрагмент антитела против CXCL13» и/или «вариант антитела против CXCL13» и тому подобное включают любой белок или полипептид, содержащий молекулу, которая включает, по меньшей мере, участок молекулы иммуноглобулина, например, но не ограничиваясь ими, по меньшей мере, одну определяющую комплементарность область (CDR) тяжелой или легкой цепи, или его связывающую лиганд область, вариабельную область тяжелой цепи или легкой цепи, константную область тяжелой цепи или легкой цепи, каркасную область, или любой их участок, или, по меньшей мере, один фрагмент связывающего CXCL13 белка, полученный из белка или пептида CXCL13, который может быть включен в антитело для применения в настоящем изобретении. Такое антитело необязательно дополнительно воздействует на специфический лиганд, например, но не ограничиваясь этим, когда такое антитело модулирует, снижает, повышает, оказывает противодействие, оказывает агонистическое действие, ослабляет, облегчает, блокирует, ингибирует, отменяет и/или нарушает активность CXCL13 in vitro, in situ и/или in vivo. В качестве не ограничивающего примера, подходящее антитело против CXCL13, определенный участок или вариант настоящего изобретения могут связываться, по меньшей мере, с одним белком или пептидом CXCL13, или его определенными участками, вариантами или доменами. Подходящее антитело против CXCL13, определенный участок или вариант оказывают влияние на функцию CXCL13 различными путями, например, но не ограничиваясь этим, влияют на синтез РНК, ДНК или белка, высвобождение CXCL13, передачу сигналов с участием CXCL13, связывание CXCL13, продукцию и/или синтез CXCL13.
Антитела могут содержать один или несколько из следующих элементов: по меньшей мере, одну CDR, по меньшей мере, одну вариабельную область, по меньшей мере, одну константную область, по меньшей мере, одну тяжелую цепь (например, g1, g2, g3, g4, m, a1, a2, d, e), по меньшей мере, одну легкую цепь (например, κ или l) или их любой участок или фрагмент, и могут дополнительно содержать дисульфидные связи между и внутри цепей, шарнирные области, сайты гликозилирования, которые могут быть разделены шарнирной областью, а также тяжелые цепи и легкие цепи. Как правило, легкие цепи имеют молекулярную массу примерно 25 кДа, а молекулярная масса тяжелых цепей находится в пределах 50-77 кДа. Легкие цепи могут находиться в двух различных формах или изотипах каппа (κ) и лямбда (l), которые могут быть объединены с любым из типов тяжелой цепи. Все легкие цепи содержат, по меньшей мере, одну вариабельную область и, по меньшей мере, одну константную область. Считают, что IgG-антитело имеет типичную структуру антитела и содержит две внутрицепочечные дисульфидные связи в легкой цепи (одну в вариабельной области и одну в константной области), четыре в тяжелой цепи и посредством таких связей образуется пептидная петля примерно из 60-70 аминокислот, включающая «домен» примерно из 110 аминокислот в цепи. IgG-антитела можно разделить на четыре класса: IgG1, IgG2, IgG3 и IgG4. Каждый класс иммуноглобулинов имеет ряд различных функций. В таблице 1 приведены данные по физико-химическим свойствам каждого класса и подкласса иммуноглобулинов.
В таблице 2 приведены не ограничивающие примеры эффекторных функций антитела для классов и подклассов человеческих антител.
Следовательно, можно выбрать тип антитела или его фрагмента для применения по настоящему изобретению, основываясь на желаемых характеристиках и функциях, которые желательны для конкретного терапевтического или диагностического применения, таких как, но не ограничиваясь ими, период полужизни, внутрисосудистое распределение, связывание комплемента и т.д.
Выделенный полипептид CXCL13 или его фрагмент можно использовать в качестве иммуногена для стимуляции выработки антител с использованием стандартных методик получения поликлональных и моноклональных антител. Можно использовать полноразмерный полипептид или белок, или, альтернативно, изобретение предусматривает антигенные пептидные фрагменты для применения в качестве иммуногенов. Антигенный пептид белка CXCL13 содержит, по меньшей мере, 8 (предпочтительно 10, 15, 20 или 30 или более) аминокислотных остатков и включает эпитоп белка, так что антитело, продуцированное против пептида, образует специфический иммунный комплекс с белком.
Как правило, иммуноген используют для получения антител иммунизацией подходящего (т.е. иммунокомпетентного) животного, такого как кролик, коза, мышь или другое млекопитающее или позвоночное животное. Подходящий иммуногенный препарат может содержать, например, экспрессированный рекомбинантным путем или синтезированный химическим путем полипептид. Препарат может дополнительно содержать адъювант, такой как полный или неполный адъювант Фрейнда, или аналогичный иммуностимулирующий агент.
Клетки, продуцирующие антитела, можно получить из периферической крови или, предпочтительно, из селезенки или лимфатических узлов людей или других подходящих животных, которые были иммунизированы представляющим интерес иммуногеном. Также может быть использована любая другая подходящая клетка-хозяин для экспрессии гетерологичной или эндогенной нуклеиновой кислоты, кодирующей антитело, его определенный фрагмент или вариант по настоящему изобретению. Слитые клетки (гибридомы) или рекомбинантные клетки можно выделить, используя селективные культуральные условия или другие подходящие известные способы, и клонировать ограничивающим разведением или клеточным сортингом, или другими известными способами. Клетки, которые продуцируют антитела с желаемой специфичностью, можно отобрать с использованием подходящего анализа (например, ELISA).
При одном подходе гибридому получают слиянием клеток подходящей иммортальной линии (например, миеломной клеточной линии, включая, но не ограничиваясь этим, Sp2/0, Sp2/0-AG14, NSO, NS1, NS2, AE-1, L.5, >243, P3X63Ag8.653, Sp2 SA3, Sp2 MAI, Sp2 SS1, Sp2 SA5, U937, MLA 144, ACT IV, MOLT4, DA-1, JURKAT, WEHI, K-562, COS, RAJI, NIH 3T3, HL-60, MLA 144, NAMALWA, NEURO 2A или тому подобное) или гетеромиелом, их слитых продуктов, или любой клетки или слитой клетки, полученной из них, или любой другой подходящей клеточной линии, известной в данной области (см., например, www.atcc.org, www.lifetech.com и тому подобное), с продуцирующими антитела клетками, такими как, но не ограничиваясь ими, выделенные или клонированные клетки селезенки, периферической крови, лимфатических узлов, миндалин, или другие иммунные или В-клетки, или любые другие клетки, экспрессирующие последовательности константных или вариабельных, или каркасных областей, или CDR тяжелой или легкой цепи, или в виде эндогенной или гетерологичной нуклеиновой кислоты, в виде рекомбинантной или эндогенной, вирусов, бактерий, дрожжей, прокариотов, амфибий, насекомых, рептилий, рыб, млекопитающих, грызунов, лошадей, овец, коз, овец, приматов, эукариотов, геномной ДНК, кДНК, рДНК, митохондриальной ДНК или РНК, ДНК или РНК хлоропластов, гяРНК (hnRNA), мРНК (mRNA), тРНК (tRNA), одноцепочечной, двухцепочечной или трехцепочечной, гибридизованной и тому подобное, или их любой комбинации. См., например, Ausubel, выше, и Colligan, Immunology, выше, глава 2, полностью включенные в данный документ путем ссылки.
Могут быть использованы другие подходящие способы получения или выделения антител с необходимой специфичностью, включая, но не ограничиваясь ими, способы, с помощью которых рекомбинантное антитело выбирают из пептидной или полипептидной библиотеки (например, но не ограничиваясь ими, библиотеки бактериофагов, рибосом, олигонуклеотидов, РНК, кДНК или тому подобное, библиотека фагового дисплея; например, доступная от Cambridge Antibody Technologies, Cambridgeshire, UK; MorphoSys, Martinsreid/Planegg, DE; Biovation, Aberdeen, Scotland, UK; BioInvent, Lund, Sweden; Dyax Corp., Enzon, Affymax/Biosite; Xoma, Berkeley, CA; Ixsys. См., например, публикации ЕР № 368894, PCT/GB91/01134; PCT/GB92/01755; PCT/GB92/002240; PCT/GB92/00883; PCT/GB93/00605; патент США № 5962255; PCT/GB94/01422; PCT/GB94/02662; PCT/GB97/01835; (CAT/MRC); WO90/14443; WO90/14424; WO90/14430; PCT/US94/1234; WO92/18619; WO96/07754; (Scripps); EP 614989 (MorphoSys); WO95/16027 (BioInvent); WO88/06630; WO90/3809 (Dyax); патент США № 4704692 (Enzon); PCT/US91/02989 (Affymax); WO89/06283; EP 371998; EP 550400; (Xoma); EP 229046; PCT/US91/07149 (Ixsys); или стохастически полученные пептиды или полипептиды - патенты США №№ 5723323; 5763192; 5814476; 5817483; 5824514 и 5976862; WO 86/05803; ЕР 590689 (Ixsys, в настоящее время Applied Molecular Evolution (AME)), каждый из этих источников полностью включен в данный документ путем ссылки), или которые основаны на иммунизации трансгенных животных (например, мышей SCID, см. Nguyen et al., Microbiol. Immunol. 41:901-907 (1997); Sandhu et al., Crit. Rev. Biotechnol., 16:95-118 (1996); Eren et al., Immunol. 93:154-161 (1998), каждый из этих источников полностью включен в данный документ путем ссылки, а также родственные патенты и заявки), которые способны продуцировать репертуар человеческих антител, как это известно в данной области и/или описано в данном документе. Такие способы включают, но не ограничиваются ими, рибосомный дисплей (Hanes et al., Proc. Natl. Acad. Sci. USA, 94:4937-4942 (May 1997); Hanes et al., Proc. Natl. Acad. Sci. USA, 95:14130-14135 (Nov. 1998); технологии получения антитела из одной клетки (например, метод отобранных лимфоцитарных антител («SLAM») (патент США № 5627052; Wen et al.,J. Immunol., 17:887-892 (1987); Babcook et al., Proc. Natl. Acad. Sci. USA 93:7843-7848 (1996)); гелевый капельный микроанализ и проточная цитометрия (Powell et al., Biotechnol., 8:333-337 (1990); One Cell Systems, Cambridge, MA; Gray et al., J. Imm. Meth., 182:155-163 (1995); Kenny et al., Bio/Technol., 13:787-790 (1995)); В-клеточный отбор (Steenbakkers et al., Molec. Biol. Reports 19:125-134 (1994); Jonak et al., Progress Biotech., Vol. 5, In Vitro Immunization in Hybridoma Technology, Borrebaeck, ed., Elsevier Science Publishers B.V., Amsterdam, Netherlands (1988)).
Также можно использовать методы генной инженерии и получения гуманизированных антител, не относящихся к человеческим, и они хорошо известны в данной области. Как правило, гуманизированные или генно-инженерные антитела содержат один или несколько аминокислотных остатков животного, не относящегося к человеку, например, но не ограничиваясь ими, мыши, крысы, кролика, примата, не относящегося к человеку, или другого млекопитающего. Аминокислотные остатки человека обычно относят к «импортным» остаткам, которые, как правило, взяты из «импортного» вариабельного, константного или другого домена известной человеческой последовательности. Известные человеческие Ig-последовательности раскрыты, например, на сайтах:
www.ncbi.nlm.nih.gov/entrez/query.fcgi;
www.ncbi.nih.gov/igblast;
www.atcc.org/phage/hdb.html;
www.mrc-cpe.cam.ac.uk/ALIGNMENTS.php;
www.kabatdatabase.com/top.html;
ftp.ncbi.nih.gov/repository/kabat;
www.sciquest.com;
www.abcam.com;
www.antibgodyresource.com/onlinecomp.html;
www.public.iastate.edu/~pedro/research_tools.html;
www.whfreeman.com/immunolgy/CH05/kuby05.htm;
www.hhmi.org/grants/lectures/1996/vlab;
www.path.cam.ac.uk/~mrc7/mikeimages.html;
mcb.harvard.edu/BioLinks/Immunology.html;
www.immunologylink.com;
pathbox.wustl.edu/~hcenter/index.html;
www.appliedbiosystems.com;
www.nal.usda.gov/awic/pubs/antibody;
www.m.ehime-u.ac.jp/~yasuhito/Elisa.html;
www.biodesign.com;
www.cancerresearchuk.org;
www.biotech.ufl.edu;
www.isac-net.org;
baserv.uci.kun.nl/~jraats/links1.html;
www.recab.unihd.de/immuno.bme.nwu.edu;
www.mrc-cpe.cam.ac.uk;
www.ibt.unam.mx/vir/V_mice.html;
http://www.bioinf.org.uk/abs;
antibody.bath.ac.uk;
www.unizh.ch;
www.cryst.bbk.ac.uk/~ubcg07s;
www.nimr.mrc.ac.uk/CC/ccaewg/ccaewg.html;
www.path.cam.ac.uk/~mrc7/humanisation/TAHHP.html;
www.ibt.unam.mx/vir/structure/stat_aim.html;
www.biosci.missouri.edu/smithgp/index.html;
www.jerini.de;
Kabat et al., Sequences of Proteins of Immunological Interest, US Dept. Health (1983); каждый из этих источников полностью включен в данный документ путем ссылки.
Такие импортированные последовательности можно использовать для снижения иммуногенности или снижения, усиления или модификации связывания, аффинности, включения, выключения, авидности, специфичности, периода полужизни или любой другой подходящей характеристики, известной в данной области. Как правило, не относящиеся к человеку или человеческие последовательности CDR, частично или полностью, сохраняются, в то время как не относящиеся к человеку последовательности вариабельной и константной областей замещаются на человеческие или другие аминокислоты. Также антитела можно необязательно гуманизировать с сохранением высокой аффинности для антигена и других ценных биологических свойств. Для достижения этой цели гуманизированные антитела можно необязательно получить проведением анализа исходных последовательностей и различных концептуальных гуманизированных продуктов, используя трехмерные модели исходных и гуманизированных последовательностей. Трехмерные модели иммуноглобулина обычно являются доступными и они знакомы специалистам в данной области. Имеются компьютерные программы, которые иллюстрируют и показывают возможные трехмерные конформационные структуры выбранных последовательностей-кандидатов иммуноглобулина. Исследование данных дисплеев позволяет анализировать возможную роль остатков в функционировании иммуноглобулиновой последовательности-кандидата, т.е. анализировать остатки, которые оказывают влияние на способность иммуноглобулина-кандидата связываться с антигеном. При данном подходе можно выбрать остатки FR и объединить их с консенсусными и импортными последовательностями таким образом, чтобы добиться желательной характеристики антитела, такой как повышенная аффинность для антигена(ов)-мишени. Как правило, остатки CDR непосредственно и наиболее существенно оказывают влияние на связывание антигена. Гуманизацию или конструирование антител по настоящему изобретению можно осуществлять с использованием любого известного способа, включая, но не ограничиваясь ими, способы, описанные Winter (Jones et al., Nature 321:522 (1986); Reichmann et al., Nature 332:323 (1988); Verhoeyen et al., Science 239:1534 (1988); Sims et al., J. Immunol. 151:2296 (1993); Clothia and Lesk, J. Mol. Biol. 196:901 (1987); Carter et al., Proc. Natl. Acad. Sci. U.S.A. 89:4285 (1992); Presta et al., J. Immunol. 151:2623 (1993); патенты США №№: 5723323; 5976862; 5824514; 5817483; 5814476; 5763192; 5723323; 5768886; 5714352; 6204023; 6180370; 5693762; 5530101; 5585089; 5225539 и 4816567; PCT/US98/16280; US96/18978; US91/09630; US91/05939; US94/01234; GB89/01334; GB91/01134; GB92/01755; WO90/14443; WO90/14424 и WO90/14430; ЕР 229246; каждый из этих источников полностью включен в данный документ путем ссылки, в том числе источники, цитированные в них.
Антитело CXCL13 можно также необязательно получить иммунизацией трансгенного животного (например, мыши, крысы, хомяка, примата, не относящегося к человеку, и тому подобное), способного продуцировать репертуар человеческих антител, как здесь описано и/или известно в данной области. Клетки, продуцирующие человеческие антитела CXCL13, можно выделить у таких животных и подвергнуть иммортализации с использованием подходящих способов, таких как способы, описанные в данном документе.
Трансгенные мыши, которые могут продуцировать репертуар человеческих антител, которые связываются с человеческими антигенами, могут быть получены известными способами (например, но не ограничиваясь ими, способы, описанные в патентах США №№: 5770428; 5569825; 5545806; 5625126; 5625825; 5633425; 5661016 и 5789650, выданные Lonberg et al.; Jakobovits et al. WO 98/50433; Jakobovits et al. WO 98/24893; Lonberg et al. WO 98/24884; Lonberg et al. WO 97/13852; Lonberg et al. WO 94/25585; Kucherlapate et al. WO 96/34096; Kucherlapate et al. EP 0463151 B1; Kucherlapate et al. EP 0710719 A1; Surani et al. патент США № 5545807; Bruggemann et al. WO 90/04036; Bruggemann et al. EP 0438474 B1; Lonberg et al. EP 0814259 A2; Lonberg et al., GB 2272440 A, Lonberg et al. Nature 368:856-959 (1994); Taylor et al., Int. Immunol. 6(4) 579-591 (1994); Green et al., Nature Genetics 7:13-21 (1994); Mendez et al., Nature Genetics 15:146-156 (1997); Taylor et al., Nucleic Acids Research 20(23):6287-6295 (1992); Tuaillon et al., Proc. Natl. Acad. Sci. USA 90(8):3720-3724 (1993); Lonberg et al., Int. Rev. Immunol. 13(1):65-93 (1995) и Fishwald et al., Nat. Biotechnol. 14(7):845-851 (1996), каждый из этих источников полностью включен в данный документ путем ссылки). Как правило, эти мыши включают, по меньшей мере, один трансген, содержащий ДНК, по меньшей мере, из одного локуса человеческого иммуноглобулина, который функционально реаранжирован, или который может претерпевать функциональную реаранжировку. Эндогенные локусы иммуноглобулина у таких мышей могут быть разрушены или удалены для элиминации способности животного продуцировать антитела, кодированные эндогенными генами.
Антитела по настоящему изобретению также могут быть получены в молоке введением, по меньшей мере, одной нуклеиновой кислоты, кодирующей антитело против CXCL13, трансгенным животным или млекопитающим, таким как козы, коровы, лошади, овцы и тому подобное, которые продуцируют антитела в их молоке. Такие животные могут быть получены с использованием известных способов. См., например, но не ограничиваясь ими, патенты США №№ 5827690; 5849992; 4873316; 5849992; 5994616; 5565362; 5304489 и тому подобное; каждый из этих источников полностью включен в данный документ путем ссылки. Антитела по настоящему изобретению могут быть дополнительно получены с использованием, по меньшей мере, одной нуклеиновой кислоты, кодирующей CXCL13 антитело, с получением трансгенных растений и культивированных растительных клеток (например, но не ограничиваясь ими, табака и кукурузы), которые продуцируют такие антитела, их определенные участки или варианты в частях растений или в клетках, культивированных из них.
Антитела по изобретению могут связываться с человеческим CXCL13 с аффинностью, находящейся в широком ряду значений (Kd). В предпочтительном варианте осуществления, по меньшей мере, одно человеческое mAb по настоящему изобретению может необязательно связываться с человеческим CXCL13 с высокой аффинностью. Например, человеческое mAb может связываться с человеческим CXCL13 с Kd, равной или составляющей менее чем примерно 10-7 М, например, но не ограничиваясь этим, 0,1-9,9 (или в любом интервале и при любом значении в этом интервале) × 10-7, 10-8, 10-9, 10-10, 10-11, 10-12, 10-13 или в любом интервале и при любом значении в этом интервале.
Аффинность или авидность антитела для антигена можно определить с использованием любого подходящего способа (См., например, Berzofsky, et al., “Antibody-Antigen Interactions” в Fundamental Immunology, Paul, W.E., Ed., Raven Press: New York, NY (1984); Kuby, Janis Immunology, W. H. Freeman and Company: New York, NY (1992); и способы, описанные в них). Измеренная аффинность взаимодействия конкретного антитела-антигена может варьировать при оценке в различных условиях (например, в зависимости от концентрации соли, рН). Таким образом, предпочтительно проводить определение аффинности и других параметров связывания антигена (например, KD, Ka, Kd) с использованием стандартизированных растворов антитела и антигена, и стандартизированного буфера, такого как буфер, описанный в данном документе.
Антагонист CXCL13 (например, моноклональное антитело) можно использовать для выделения полипептида CXCL13 с использованием обычных методов, таких как аффинная хроматография или иммунопреципитация. Кроме того, такое антитело можно использовать для детектирования белка (например, в клеточном лизате или клеточном супернатанте) для оценки избыточности и характера экспрессии полипептида. Антитела можно также использовать в диагностических целях для определения уровней белка в ткани в качестве составной части процедуры клинического тестирования, например, для определения эффективности конкретной схемы лечения. Детектирование можно облегчить сочетанием антитела с детектируемым соединением. Примеры детектируемых соединений включают различные ферменты, простетические группы, флуоресцентные вещества, люминесцентные вещества, биолюминесцентные вещества и радиоактивные вещества. Примеры подходящих ферментов включают пероксидазу хрена, щелочную фосфатазу, β-галактозидазу и ацетилхолинэстеразу; примеры подходящих простетических групп-комплексов включают стрептавидин/биотин и авидин/биотин; примеры подходящих флуоресцентных веществ включают умбеллиферон, флуоресцеин, флуоресцеин изотиоцианат, родамин, дихлортриазиниламин флуоресцеин, дансилхлорид или фикоэритрин; примеры люминесцентного вещества включают люминол; примеры биолюминесцентных веществ включают люциферазу, люциферин и экворин; и примеры подходящих радиоактивных веществ включают 125I, 131I, 35S или 3Н.
Термин «антитело» дополнительно предназначен для включения антител, фрагментов, полученных в результате расщепления, их специфических участков и вариантов, в том числе, миметиков антител или участков антител, которые имитируют структуру и/или функцию антитела или его специфического фрагмента или участка, в том числе одноцепочечных антител и их фрагментов. Функциональные фрагменты включают антигенсвязывающие фрагменты, которые связываются с CXCL13 млекопитающих. Например, фрагменты антител, способные связываться с CXCL13, или их участки, включая, но не ограничиваясь ими, фрагменты Fab (например, полученные путем расщепления папаином), Fab' (например, полученные путем расщепления пепсином и неполного восстановления) и F(ab')2 (например, полученные путем расщепления пепсином), facb (например, полученные путем расщепления плазмином), pFc' (например, полученные путем расщепления пепсином или плазмином), Fd (например, полученные путем расщепления пепсином, неполного восстановления и реагрегации), Fv или scFv (например, полученные методами молекулярной биологии), входят в объем изобретения (см., например, Colligan et al., Immunology, выше).
Такие фрагменты можно получить ферментативным расщеплением, синтетическими или рекомбинантными методами, известными в данной области и/или описанными в данном документе. Антитела можно также получить в виде различных усеченных форм с использованием генов антител, в которых один или несколько стоп-кодонов введены слева от природного стоп-сайта. Например, можно сконструировать ген, кодирующий тяжелую цепь F(ab')2, с включением последовательностей ДНК, кодирующих СН1 домен и/или шарнирную область тяжелой цепи. Различные области антител можно соединить вместе химическим путем с использованием обычных методик, или можно получить в виде сцепленного белка с использованием методов генной инженерии.
Антитело против CXCL13 может представлять собой антитело примата, грызуна или человека, или может быть химерным или гуманизированным антителом. Используемый в данном документе термин «человеческое антитело», относится к антителу, в котором по существу каждый фрагмент белка (например, CDR, каркасная область, домены CL, CH (например, СН1, СН2, СН3), шарнирная область, (VL, VH)) является по существу неиммуногенным у людей, только с минимальными изменениями или вариациями последовательностей, и/или оно сконструировано для, получено из или содержит известные компоненты человеческого антитела. Аналогично, антитела, сконструированные как антитела приматов (обезьяны, павиана, шимпанзе и т.д.), грызунов (мыши, крысы, кролика, морской свинки, хомяка и тому подобное) и других млекопитающих, означают антитела, специфичные для вида, подрода, рода, подсемейства, семейства. Кроме того, химерные антитела по изобретению могут включать любую комбинацию вышеуказанного. Такие изменения или вариации необязательно и предпочтительно приводят к сохранению или снижению иммуногенности у людей или других видов по сравнению с немодифицированными антителами. Таким образом, человеческое антитело отличается от химерного или гуманизированного антитела. Необходимо подчеркнуть, что человеческое антитело можно получить с использованием животного, отличного от человека, или прокариотической или эукариотической клетки, способной экспрессировать функционально реаранжированные гены иммуноглобулинов (например, тяжелой цепи и/или легкой цепи). Кроме того, когда человеческое антитело является одноцепочечным антителом, оно может содержать линкерный пептид, который не обнаружен в нативных человеческих антителах. Например, Fv может содержать линкерный пептид, такой как состоящий из 2 до примерно 8 глицинов или других аминокислотных остатков, который связывает вариабельную область тяжелой цепи и вариабельную область легкой цепи. Считается, что такие линкерные пептиды должны быть человеческого происхождения.
Также можно использовать биспецифические, гетероспецифические, гетероконъюгатные или подобные антитела, которые являются моноклональными, предпочтительно человеческими или гуманизированными антителами, которые обладают специфичностью связывания, по меньшей мере, для двух различных антигенов. В данном случае, один из типов специфичности связывания соответствует, по меньшей мере, одному белку CXCL13, другой тип специфичности - любому другому антигену, например, рецептору CXCL13 или TNFα. В данной области известны способы получения биспецифических антител. Обычно рекомбинантная продукция биспецифических антител основана на коэкспрессии двух пар тяжелой цепи-легкой цепи иммуноглобулина, где две тяжелые цепи обладают различными типами специфичности (Milstein and Cuello, Nature 305:537 (1983)). За счет произвольного ассортимента тяжелой и легкой цепей иммуноглобулина данные гибридомы (квадромы) продуцируют потенциальную смесь 10 различных молекул антител, из которых только одно имеет правильную биспецифическую структуру. Выделение правильной молекулы, которое обычно проводят методом аффинной хроматографии, является довольно громоздким, и выход продукта довольно низкий. Аналогичные способы описаны, например, в WO 93/08829, патентах США №№ 6210668; 6193967; 6132992; 6106833; 6060285; 6037453; 6010902; 5989530; 5959084; 5959083; 5932448; 5833985; 5821333; 5807706; 5643759; 5601819; 5582996; 5496549; 4676980; WO 91/00360; WO 92/00373; ЕР 03089; Traunecker et al., EMBO J. 10:3655 (1991); Suresh et al., Methods in Enzymology 121:210 (1986), каждый из этих источников полностью включен в данный документ путем ссылки.
Антитела против CXCL13, пригодные для способов и композиций по настоящему изобретению, могут необязательно характеризоваться высокой аффинностью связывания с CXCL13 и необязательно и предпочтительно обладают низкой токсичностью. В частности, антитело, определенный фрагмент или вариант по изобретению, где отдельные компоненты, такие как вариабельная область, константная область и каркасная область, индивидуально и/или вместе, необязательно и предпочтительно обладают низкой иммуногенностью, пригодны для настоящего изобретения. Антитела, которые могут быть использованы в изобретении, необязательно характеризуются возможностью применять их для лечения пациентов в течение продолжительного периода времени с ощутимым ослаблением симптомов и низкой и/или приемлемой токсичностью. Низкая или приемлемая иммуногенность и/или высокая аффинность, а также другие подходящие свойства, могут вносить свой вклад в достигаемые результаты лечения. «Низкая иммуногенность» определяют в данном документе как индукцию сильных ответных реакций HAHA, HACA или НАМА у менее чем примерно 75%, или предпочтительно у менее чем примерно 50% пациентов, подвергавшихся лечению, и/или индукцию низких титров у пациентов, подвергавшихся лечению (менее чем примерно 300, предпочтительно менее чем примерно 100 по данным двойного иммуноферментного анализа с использованием антигена) (Elliott et al., Lancet 344:1125-1127 (1994), этот источник полностью включен в данный документ путем ссылки).
Подходящие антитела включают антитела, которые конкурируют за связывание с человеческим CXCL13 с моноклональными антителами, которые блокируют активацию CXCL13.
Антагонисты CXCL13 в виде siRNA (миРНК), shRNA (мшРНК), антисмысловых молекул, рибозимов и ДНКзимов
Терапевтическая направленность индуктора CXCL13 может обеспечить лучшие шансы для успеха. Экспрессию гена можно модулировать несколькими различными путями, включая применение миРНК, мшРНК, антисмысловых молекул, рибозимов и ДНКзимов. Можно сконструировать синтетические миРНК, мшРНК, рибозимы и ДНКзимы для определенного нацеливания на один или несколько генов, и их можно легко ввести в клетки in vitro или in vivo.
Настоящее изобретение включает антисмысловые нуклеиновокислотные молекулы, т.е. молекулы, которые являются комплементарными к смысловой нуклеиновой кислоте, кодирующей полипептид CXCL13, например, комплементарными к кодирующей цепи молекулы двухцепочечной кДНК или комплементарными к последовательности мРНК. Соответственно, антисмысловая нуклеиновая кислота может связываться за счет образования водородных связей со смысловой нуклеиновой кислотой. Антисмысловая нуклеиновая кислота может быть комплементарной к целой кодирующей цепи или только к ее фрагменту, например, ко всей или части кодирующей белок области (или открытой рамке считывания). Антисмысловая нуклеиновая кислота может быть антисмысловой ко всей или части некодирующей области кодирующей цепи нуклеотидной последовательности, кодирующей полипептид CXCL13. Некодирующие области («5' и 3' нетранслируемые области») представляют собой 5'- и 3'-последовательности, которые фланкируют кодирующую область и не транслируются в аминокислоты.
Антисмысловой олигонуклеотид может, например, иметь в длину примерно 5, 10, 15, 20, 25, 30, 35, 40, 45 или 50 или более нуклеотидов. Антисмысловую нуклеиновую кислоту по изобретению можно сконструировать с использованием реакций химического синтеза и ферментативного лигирования, следуя методикам, известным в данной области. Например, антисмысловую нуклеиновую кислоту (например, антисмысловой олигонуклеотид) можно синтезировать химическим путем с использованием природных нуклеотидов или различных модифицированных нуклеотидов, сконструированных с целью повышения биологической стабильности молекул или повышения физической стабильности дуплекса, образованного антисмысловой и смысловой нуклеиновыми кислотами, например, можно использовать фосфоротиоатные производные, пептидные нуклеиновые кислоты (PNA) и замещенные акридином нуклеотиды. Примеры модифицированных нуклеотидов, которые могут быть использованы для получения антисмысловой нуклеиновой кислоты, включают 5-фторурацил, 5-бромурацил, 5-хлорурацил, 5-йодурацил, гипоксантин, ксантин, 4-ацетилцитозин, 5-(карбоксигидроксиметил)урацил, 5-карбоксиметиламинометил-2-тиоуридин, 5-карбоксиметиламинометилурацил, дигидроурацил, бета-D-галактозилквеозин, инозин, N6-изопентениладенин, 1-метилгуанин, 1-метилинозин, 2,2-диметилгуанин, 2-метиладенин, 2-метилгуанин, 3-метилцитозин, 5-метилцитозин, N6-аденин, 7-метилгуанин, 5-метиламинометилурацил, 5-метоксиаминометил-2-тиоурацил, бета-D-маннозилквеозин, 5'-метоксикарбоксиметилурацил, 5-метоксиурацил, 2-метилтио-N6-изопентениладенин, урацил-5-оксиуксусную кислоту (v), вибутоксозин, псевдоурацил, квеозин, 2-тиоцитозин, 5-метил-2-тиоурацил, 2-тиоурацил, 4-тиоурацил, 5-метилурацил, метиловый эфир урацил-5-оксиуксусной кислоты, урацил-5-оксиуксусную кислоту (v), 5-метил-2-тиоурацил, 3-(3-амино-3-N-2-карбоксипропил)урацил, (acp3)w и 2,6-диаминопурин. Альтернативно, антисмысловую нуклеиновую кислоту можно получить биологическим путем с использованием экспрессирующего вектора, в который субклонируют нуклеиновую кислоту в антисмысловой ориентации (т.е. РНК, транскрибируемая со вставленной нуклеиновой кислоты, будет находиться в антисмысловой ориентации по отношению к интересующей нуклеиновой кислоте-мишени, описанной дополнительно в следующем подразделе).
Молекулы антисмысловых нуклеиновых кислот по изобретению обычно вводят субъекту или воспроизводят in situ таким образом, чтобы они гибридизовались или связывались с клеточной мРНК и/или геномной ДНК, кодирующей выбранный полипептид CXCL13, тем самым ингибируя экспрессию, например, подавлением транскрипции и/или трансляции. Гибридизация может быть основана на обычной комплементарности нуклеотидов с образованием стабильного дуплекса или, например, в случае молекулы антисмысловой нуклеиновой кислоты, которая связывается с ДНК-дуплексами, путем специфических взаимодействий в большой бороздке двойной спирали. Примером пути введения молекул антисмысловых нуклеиновых кислот по изобретению является прямая инъекция в участок ткани. Альтернативно, молекулы антисмысловых нуклеиновых кислот могут быть модифицированы для направленного транспорта к выбранным клеткам-мишеням и затем введены системно. Например, для системного введения антисмысловые молекулы можно модифицировать таким образом, чтобы они специфически связывались с рецепторами или антигенами, экспрессированными на поверхности выбранной клетки, например, путем связывания молекул антисмысловых нуклеиновых кислот с пептидами или антителами, которые связываются с рецепторами клеточной поверхности или антигенами. Молекулы антисмысловых нуклеиновых кислот также могут быть доставлены в клетки с использованием векторов, описанных в данном документе. Для достижения достаточных внутриклеточных концентраций антисмысловых молекул предпочтительными являются векторные конструкции, в которых молекула антисмысловой нуклеиновой кислоты помещена под контролем сильного промотора pol II или pol III.
Молекула антисмысловой нуклеиновой кислоты по изобретению может быть молекулой α-аномерной нуклеиновой кислоты. Молекула α-аномерной нуклеиновой кислоты образует специфические двухцепочечные гибриды с комплементарной РНК, в которых, в противоположность обычным α-единицам, цепи расположены параллельно по отношению друг к другу (Gaultier et al. (1987) Nucleic Acids Res. 15:6625-6641). Молекула антисмысловой нуклеиновой кислоты также может содержать 2'-о-метилрибонуклеотид (Inoue et al. (1987) Nucleic Acids Res., 15:6131-6148) или химерный аналог РНК-ДНК (Inoue et al. (1987) FEBS Lett. 215:327-330).
Изобретение также включает рибозимы. Рибозимы представляют собой каталитические молекулы РНК с рибонуклеазной активностью, которые способны расщеплять одноцепочечную нуклеиновую кислоту, такую как мРНК, к которой они имеют комплементарную область. Таким образом, рибозимы (например, рибозимы акулы-молота, описанные Haselhoff and Gerlach (1988) Nature 334:585-591), можно использовать для каталитического расщепления транскриптов мРНК с подавлением, тем самым, трансляции белка, кодированного мРНК. Можно сконструировать рибозим, обладающий специфичностью для молекулы нуклеиновой кислоты, кодирующей полипептид CXCL13, на основе нуклеотидной последовательности кДНК, раскрытой в данном документе. Например, можно получить производное РНК Tetrahymena L-19 IVS, в котором нуклеотидная последовательность активного сайта комплементарна нуклеотидной последовательности, предназначенной для расщепления, Cech et al., патент США № 4987071; и Cech et al., патент США № 5116742. Альтернативно, можно использовать мРНК, кодирующую полипептид CXCL13, для выбора каталитической РНК со специфической рибонуклеазной активностью из пула молекул РНК. См., например, Haselhoff and Gerlach, выше; Bartel and Szostac (1993) Science 261:1411-1418).
Изобретение также включает молекулы рибонуклеиновой кислоты, которые являются комплементарными, антисмысловыми, двухцепочечными гомологами, миРНК (siRNA), или являются последовательность-специфическими одноцепочечными РНК, которые образуют короткие структуры в форме «шпильки», мшРНК (shRNA) (в общем называемые интерферирующими РНК), которые можно использовать для снижения специфической экспрессии генов, в данном случае CXCL13, и, следовательно, для ингибирования экспрессии белка и подавления их соответствующих биологических функций. (Fire, A. et al. (1998) Nature 391:806-811; Paddison, P.J. et al. (2002) Genes Develop. 16:948-958).
Изобретение дополнительно включает ДНКзимы, способные расщеплять молекулы РНК (Breaker and Joyce, 1994; Santoro and Joyce, 1997) или ДНК (Carmi et al., 1996). Скорость каталитического расщепления таких нуклеиновокислотных ферментов зависит от присутствия и концентрации ионов бивалентных металлов, таких как Ba2+, Sr2+, Mg2+, Ca2+, Ni2+, Co2+, Mn2+, Zn2+ и Pb2+ (Santoro and Joyce, 1998; Brown et al., 2003).
Каталитические ДНКзимы, такие как ДНКзимы 10:23 и 8:17, имеют много доменов. Они содержат консервативный каталитический домен (каталитическое ядро), фланкированный двумя неконсервативными субстрат-связывающими доменами (гибридизирующими плечами), которые являются областями последовательности, специфически связывающимися с субстратом. ДНКзимы 10:23 и 8:17 способны расщеплять нуклеиновокислотные субстраты по специфическим фосфодиэфирным связям РНК (Santoro and Joyce, 1997). ДНКзим 10:23 содержит каталитический домен из 15 дезоксинуклеотидов, фланкированных двумя субстрат-распознающими плечами. ДНКзим 8:17 имеет аналогичный размер.
Каталитическая нуклеиновая кислота может расщеплять нуклеиновокислотный субстрат с последовательностью-мишенью, которая должна соответствовать минимальным требованиям. Последовательность субстрата должна быть по существу комплементарной гибридизирующим плечам каталитической нуклеиновой кислоты, и субстрат должен содержать специфическую последовательность в сайте расщепления. Специфические требования для последовательности в сайте расщепления включают, например, наличие последовательности из пуриновых:пиримидиновых нуклеотидов для расщепления ДНКзимом 10:23 (Santoro and Joyce, 1997).
В различных вариантах осуществления молекулы нуклеиновой кислоты по изобретению можно модифицировать по группе основания, группе сахара или фосфатному остову для повышения, например, стабильности, гибридизации или растворимости молекулы. Например, природные нуклеотиды можно заменить на аналоги нуклеотидов, описанные выше.
В другом примере дезоксирибозофосфатный остов нуклеиновых кислот можно модифицировать с получением пептидных нуклеиновых кислот (см. Hyrup et al. (1996) Bioorganic & Medicinal Chemistry 4(1):5-23). Используемые в данном документе термины «пептидные нуклеиновые кислоты» или “PNA” относятся к миметикам нуклеиновых кислот, например, миметикам ДНК, в которых дезоксирибозофосфатный остов заменен на псевдопептидный остов и остаются только четыре природных нуклеиновых оснований. Было показано, что нейтральный остов PNA обеспечивает возможность специфической гибридизации ДНК и РНК в условиях низкой ионной силы. Синтез олигомеров PNA можно проводить с использованием обычных протоколов твердофазного пептидного синтеза, описанных Hyrup et al. (1996), выше; Perry-O'Keefe et al. (1996) Proc. Natl. Acad. Sci. USA 93:14670-675.
PNA можно использовать в терапевтических и диагностических целях. Например, PNA могут быть использованы в качестве антисмысловых или антигенных агентов для последовательность-специфической модуляции экспрессии генов, например, путем индукции остановки транскрипции или трансляции или подавления репликации. Также PNA можно использовать, например, при анализе мутаций по одной паре оснований в гене, например, PNA-направленной ПЦР фиксацией; в качестве искусственных рестриктаз при использовании в комбинации с другими ферментами, например, S1 нуклеазами (Hyrup (1996), выше); или в качестве зондов или праймеров для последовательности ДНК и гибридизации (Hyrup (1996), выше; Perry-O'Keefe et al. (1996) Proc. Natl. Acad. Sci. USA 93:14670-675).
В другом варианте осуществления PNA могут быть модифицированы, например, с целью повышения их стабильности или захвата клетками, путем присоединения к PNA липофильных или других хелперных групп с образованием химерных конструкций PNA-ДНК, или использования липосом или других способов доставки лекарственных препаратов, известных в данной области. Например, можно получить химерные конструкции PNA-ДНК, которые могут объединять полезные свойства PNA и ДНК. Такие химеры позволяют ферментам с функцией распознавания ДНК, например, РНКазе Н и ДНК-полимеразе, взаимодействовать с ДНК-фрагментом, в то время как PNA-фрагмент будет обеспечивать высокую аффинность и специфичность. Химеры PNA-ДНК можно получить с использованием линкеров соответствующей длины, выбранной с учетом «стэкинга» оснований, числа связей между нуклеиновыми основаниями и ориентации (Hyrup (1996), выше). Синтез химерных конструкций PNA-ДНК можно проводить, как описано Hyrup (1996), выше, и Finn et al. (1996) Nucleic Acids Res. 24(17):3357-63. Например, цепь ДНК можно синтезировать на твердой подложке с использованием обычного фосфорамидитного метода химического синтеза и модифицированных нуклеозидных аналогов. В качестве линкера между PNA и 5'-концом ДНК можно использовать такие соединения, как фосфорамидит 5'-(4-метокситритил)амино-5'дезокситимидина (Mag et al. (1989) Nucleic Acids Res. 17:5973-88). Затем мономеры PNA постадийно сочетают с получением химерной молекулы с 5'-PNA сегментом и 3'-ДНК сегментом (Finn et al. (1996) Nucleic Acids Res. 24(17):3357-63). Альтернативно, химерные молекулы можно синтезировать с 5'-ДНК сегментом и 3'-PNA сегментом (Peterser et al. (1975) Bioorganic Med. Chem. Lett. 5:1119-1124).
В других вариантах осуществления олигонуклеотид может включать другие присоединенные группы, такие как пептиды (например, для придания направленности к рецепторам клеток-хозяев in vivo), или агенты, которые облегчают транспорт через клеточную мембрану (см., например, Letsinger et al. (1989) Proc. Natl. Acad. Sci. USA, 86:6553-6556; Lemaitre et al. (1987) Proc. Natl. Acad. Sci. USA, 84:648-652; публикацию РСТ № WO 88/09810), или гематоэнцефалический барьер (см., например, публикацию РСТ № WO 89/10134). Кроме того, олигонуклеотиды можно модифицировать агентами расщепления, «запускаемого» гибридизацией (см., например, Krol et al. (1988) Bio/Techniques 6:958-976), или интеркалирующими агентами (см., например, Zon (1988) Pharm. Res. 5:539-549). Наконец, олигонуклеотид может быть конъюгирован с другой молекулой, например, пептидом, агентом перекрестного сшивания, «запускаемого» гибридизацией, агентом для транспорта, агентами расщепления, «запускаемого» гибридизацией и т.д.
Белки
Изобретение также относится к химерным и слитым белкам. Используемые в данном документе термины «химерный белок» или «слитый белок» включают весь полипептид или фрагмент (предпочтительно биологически активный) полипептида CXCL13, функционально связанный с гетерологичным полипептидом (т.е. полипептидом иным, чем аналогичный полипептид CXCL13). В отношении слитого белка, термин «функционально связанный» предназначен для указания того, что полипептид CXCL13 и гетерологичный полипептид слиты друг с другом в рамке считывания. Гетерологичный полипептид может быть слит с аминоконцом или карбоксиконцом полипептида CXCL13. В другом варианте осуществления полипептид CXCL13 или домен, или его активный фрагмент может быть слит с последовательностью гетерологичного белка или его фрагмента с образованием химерного белка, где полипептиды, домены или фрагменты не слиты от конца к концу, а разделены в остове гетерологичным белком.
Одним из пригодных слитых белков является слитый GST белок, в котором полипептид CXCL13 слит с карбоксильным концом последовательности GST. Такие слитые белки могут облегчить очистку рекомбинантного полипептида CXCL13.
В другом варианте осуществления слитый белок содержит гетерологичную сигнальную последовательность на своем аминоконце. Например, нативная сигнальная последовательность полипептида CXCL13 может быть удалена и заменена на сигнальную последовательность из другого белка. Например, можно использовать секреторную последовательность gp67 белка оболочки бакуловирусов в качестве гетерологичной сигнальной последовательности (Current Protocols in Molecular Biology, Ausubel et al., eds., John Wiley & Sons, 1992). Другие примеры эукариотических гетерологичных сигнальных последовательностей включают секреторные последовательности мелиттина и человеческой плацентарной щелочной фосфатазы (Stratagene; La Jolla, Calif). В еще одном примере подходящие прокариотические гетерологичные сигнальные последовательности включают секреторный сигнал phoA (Sambrook et al., выше) и секреторный сигнал белка А (Pharmacia Biotech; Piscataway, NY).
В еще одном варианте осуществления слитый белок представляет собой иммуноглобулиновый слитый белок, в котором весь полипептид CXCL13 или его фрагмент слит с последовательностями, полученными от члена семейства белков-иммуноглобулинов. Иммуноглобулиновые слитые белки по изобретению могут быть включены в фармацевтические композиции и введены субъекту для ингибирования взаимодействия между лигандом (растворимым или связанным с мембраной) и белком на поверхности клетки (рецептором) с подавлением, тем самым, сигнальной трансдукции in vivo. Иммуноглобулиновый слитый белок можно использовать для воздействия на биодоступность лиганда полипептида CXCL13. Ингибирование взаимодействия лиганд/рецептор может быть полезным в терапевтических целях как для лечения заболеваний, связанных с нарушением пролиферации и дифференцировки клеток, так и для модуляции (например, стимуляции или подавления) выживаемости клеток. Предпочтительным вариантом осуществления иммуноглобулинового химерного белка является иммуноглобулин с делетированным доменом СН1 или «миметитело» с активным полипептидным фрагментом, вставленным в модифицированную каркасную область, как описано в совместной заявке PCT WO/04002417. Кроме того, иммуноглобулиновые слитые белки по изобретению можно использовать в качестве иммуногенов для получения антител, направленных против полипептида CXCL13 у субъекта, для выделения лигандов и при проведении скринингового анализа для идентификации молекул, которые ингибируют взаимодействие рецепторов с лигандами.
Химерные и слитые белки по изобретению могут быть получены обычными методами рекомбинантной ДНК. В еще одном варианте осуществления слитый ген можно синтезировать с использованием обычных методов, включая автоматические синтезаторы ДНК. Альтернативно, можно провести амплификацию фрагментов гена с помощью ПЦР, используя «якорные» праймеры для индукции комплементарных выступов между двумя последовательными фрагментами гена, которые затем можно подвергнуть отжигу и повторной амплификации с получением последовательности химерного гена (см., например, Ausubel et al., выше). Кроме того, коммерчески доступными являются многие экспрессирующие векторы, которые уже кодируют слитый фрагмент (например, полипептид GST). Можно клонировать нуклеиновую кислоту, кодирующую полипептид CXCL13, в подобный экспрессирующий вектор таким образом, чтобы слитый фрагмент был связан в рамке считывания с полипептидом CXCL13.
Сигнальную последовательность полипептида CXCL13 можно использовать для облегчения секреции и выделения секретированного белка или других представляющих интерес белков. Как правило, для сигнальных последовательностей характерно ядро из гидрофобных аминокислот, которые обычно отщепляются от зрелого белка во время секреции в одном или нескольких случаях отщепления. Такие сигнальные пептиды содержат сайты процессинга, которые обеспечивают возможность отщепления сигнальной последовательности от зрелых белков по мере их прохождения по секреторному пути. Таким образом, изобретение относится к описанным полипептидам с сигнальной последовательностью, а также к самой сигнальной последовательности и к полипептиду без сигнальной последовательности (например, продуктам расщепления). В одном варианте осуществления нуклеиновокислотная последовательность, кодирующая сигнальную последовательность по изобретению, может быть функционально связана в экспрессирующем векторе с представляющим интерес белком, таким как белок, который обычно не секретируется или который по иным причинам трудно выделить. Сигнальная последовательность направляет секрецию белка, например, из эукариотического хозяина, в который трансформирован экспрессирующий вектор, и затем или одновременно сигнальную последовательность отщепляют. Затем белок можно легко выделить из внеклеточной среды с использованием способов, известных в данной области. Альтернативно, сигнальную последовательность можно связать с интересующим белком, используя последовательность, которая облегчает выделение, такую как домен GST.
В еще одном варианте осуществления сигнальные последовательности по настоящему изобретению можно использовать для идентификации регуляторных последовательностей, например, промоторов, энхансеров и/или репрессоров. Поскольку сигнальные последовательности в своем большинстве являются аминоконцевыми последовательностями пептида, то нуклеиновые кислоты, фланкирующие сигнальную последовательность по ее аминоконцевой стороне, вероятно, являются регуляторными последовательностями, которые оказывают влияние на транскрипцию. Таким образом, нуклеотидную последовательность, которая кодирует всю сигнальную последовательность или ее фрагмент, можно использовать в качестве зонда для идентификации и выделения сигнальных последовательностей и их фланкирующих областей, и данные фланкирующие области можно исследовать для идентификации в них регуляторных элементов.
Настоящее изобретение также относится к вариантам полипептидов CXCL13, и они могут включать одну или несколько аминокислотных замен, делеций и вставок, полученных в результате естественных мутаций или манипуляций человека, как это описано в данном документе. Такие мутации или замены могут включать мутеины, мутации в которых являются достаточными для изменения свойств пептида без изменения биологической активности пептида в отношении ингибирования связывания человеческого CXCL13 с его лигандом. Конечно, специалисту в данной области понятно, что количество аминокислотных замен будет зависеть от многих факторов, включая описанные выше. В некоторых вариантах осуществления количество аминокислотных замен, вставок или делеций для данного полипептида CXCL13, его фрагмента или варианта будет составлять не более 1-5, или представлять любой диапазон или значение в этом диапазоне, как это указано в данном документе.
Полипептиды CXCL13 также могут включать модифицированные, отличные от природных и необычные аминокислоты, замещенные или добавленные в их аминокислотные последовательности. Перечень приведенных в качестве примера модифицированных, отличных от природных и необычных аминокислот приведен ниже.
Аминокислоты в полипептиде CXCL13, которые являются важными для функции, могут быть идентифицированы способами, известными в данной области, такими как сайт-направленный мутагенез или аланин-сканирующий мутагенез (например, Ausubel, выше, главы 8, 15; Cunningham and Wells, Science 244:1081-1085 (1989)). В последнем способе используют единичные аланиновые мутации в каждом остатке молекулы. Затем полученные мутантные молекулы тестируют на биологическую активность, например, но не ограничиваясь этим, нейтрализующую активность, по меньшей мере, для одного CXCL13.
Такие варианты имеют измененную аминокислотную последовательность и могут функционировать в качестве или агонистов (миметиков), или антагонистов. Варианты могут быть получены путем мутагенеза, например, дискретной точечной мутацией или усечением. Агонист может сохранять по существу одну и ту же или несколько типов биологической активности, характерных для природной формы белка. Антагонист белка может ингибировать одну или несколько типов активности природной формы белка, например, конкурентным связыванием с компонентом справа и слева каскада передачи сигналов в клетке, который включает вызывающий интерес белок. Таким образом, специфическое биологическое действие можно обеспечить лечением с использованием варианта с ограниченной функцией. Лечение субъекта вариантом, обладающим несколькими типами биологической активности природной формы белка, может привести к меньшему проявлению побочных эффектов у субъекта по сравнению с лечением природной формой белка.
Варианты полипептида CXCL13, которые функционируют в качестве агонистов (миметиков) или в качестве антагонистов, могут быть идентифицированы путем скрининга комбинаторных библиотек мутантов, например, усеченных мутантов, белка по изобретению на агонистическую или антагонистическую активность. В одном варианте осуществления разнообразную библиотеку вариантов получают комбинаторным мутагенезом на уровне нуклеиновых кислот и кодируют созданной библиотекой генов. Разнообразную библиотеку вариантов можно получить, например, ферментативным лигированием смеси синтетических олигонуклеотидов в последовательности генов таким образом, чтобы вырождающийся ряд потенциальных белковых последовательностей экспрессировался в виде отдельных полипептидов или, альтернативно, в виде ряда более крупных слитых белков (например, для фагового дисплея). Имеются разнообразные методы, которые могут быть использованы для получения библиотек потенциальных вариантов полипептида CXCL13 из вырожденной олигонуклеотидной последовательности. Методы синтеза вырожденных олигонуклеотидов известны в данной области (см., например, Narang (1983) Tetrahedron 39:3; Itakura et al. (1984) Annu. Rev. Biochem. 53:323; Itakura et al. (1984) Science 198:1056; Ike et al. (1983) Nucleic Acid Res. 11:477).
Кроме того, библиотеки фрагментов кодирующей последовательности полипептида CXCL13 можно использовать для получения разнообразной популяции полипептидов для скрининга и последующего отбора вариантов. Например, библиотеку фрагментов кодирующей последовательности можно получить обработкой двухцепочечного фрагмента ПЦР представляющей интерес кодирующей последовательности нуклеазой в условиях, при которых расщепление происходит только примерно один раз на молекулу, денатурацией двухцепочечной ДНК, ренатурацией ДНК с образованием двухцепочечной ДНК, которая может содержать смысловую/антисмысловую пару из различных расщепленных продуктов, удалением одноцепочечных фрагментов из вновь образованных дуплексов путем обработки нуклеазой S1 и лигированием полученной библиотеки фрагментов в экспрессирующий вектор. С использованием данного способа можно получить экспрессирующую библиотеку, которая кодирует аминоконцевые и внутренние фрагменты представляющего интерес белка различных размеров.
В данной области известно несколько способов для скрининга генных продуктов комбинаторных библиотек, полученных точечными мутациями или усечением, и для скрининга библиотек кДНК для генных продуктов с выбранным свойством. Наиболее часто используемыми способами являются способы, которые можно преобразовать в высокопроизводительный формат анализа, для скрининга крупных библиотек генов, которые, как правило, включают клонирование библиотеки генов в реплицируемые экспрессирующие векторы, трансформацию соответствующих клеток с помощью полученной библиотеки векторов и экспрессирование комбинаторных генов в условиях, при которых детектирование желаемой активности облегчает выделение вектора, кодирующего ген, продукт которого детектируется. Можно использовать рекурсивный согласованный мутагенез (REM), метод, который повышает частоту образования функциональных мутантов в библиотеках, в комбинации со скринингом для идентификации вариантов антагониста полипептида CXCL13 (Arkin and Yourvan (1992) Proc. Natl. Acad. Sci. USA 89:7811-7815; Delgrave et al. (1993) Protein Engineering 6(3):327-331).
Композиции и их применения
В соответствии с изобретением нейтрализующие антагонисты CXCL13, такие как моноклональные антитела, описанные в данном документе, можно использовать для ингибирования активности CXCL13. Кроме того, такие антагонисты можно использовать для купирования связанных с CXCL13 воспалительных заболеваний, поддающихся такому лечению, которые включают, но не ограничиваются этим, заболевания легких. Индивидуум, подвергающийся лечению, может представлять собой любое млекопитающее и предпочтительно является приматом, комнатным животным, которое является млекопитающим, и наиболее предпочтительно является человеком. Количество вводимого антагониста будет варьировать в зависимости от цели, для достижения которой он применяется, и способа введения.
Антагонисты CXCL13 можно вводить любым путем, который приводит к проявлению эффекта в ткани, в которой желательно предотвратить или отменить активность CXCL13. Кроме того, необязательно, чтобы антагонисты CXCL13 присутствовали в месте проявления эффекта в отношении активности CXCL13; следовательно, их можно вводить многими путями, при условии, чтобы при этом достигался доступ к любому органу или жидкости организма, содержащим CXCL13. В случае воспаленных, злокачественных или иначе пораженных тканей данные способы могут включать непосредственное применение композиции, содержащей антагонисты. Такие способы включают внутривенное введение жидкой композиции, трансдермальное введение жидкой или твердой композиции, пероральное, местное применение или интерстициальное или интероперативное введение. Введение можно проводить при имплантации устройства, основной функцией которого может не быть его использование в качестве носителя лекарственного препарата.
Введение также может быть пероральным или представлять собой местную инъекцию непосредственно в опухоль или ткань, но, как правило, моноклональное антитело вводят внутривенно. Обычно дозы колеблются в пределах примерно от 0,05 мг/кг до примерно 12,0 мг/кг. Введение может быть в виде болюса или в виде медленной или непрерывной инфузии, при этом введение можно контролировать с помощью микропроцессора и запрограммированного насосного устройства.
Альтернативно, ДНК, кодирующая предпочтительно фрагмент моноклонального антитела, может быть выделена из гибридомных клеток и введена млекопитающему. ДНК можно вводить в чистой форме или вставить в рекомбинантный вектор, например, вакцинный вирус, таким образом, что это приведет к экспрессии ДНК в клетках пациента и доставке антитела.
Моноклональное антитело, используемое в способе по настоящему изобретению, можно представить в композиции любым из известных способов приготовления фармацевтических композиций, например, как это описано в Remington's Pharmaceutical Sciences, 1985. Для простоты введения, как правило, моноклональное антитело объединяют с фармацевтически приемлемым носителем. Такие носители включают воду, физиологический раствор или масла.
Композиции, подходящие для парентерального введения, включают водные и неводные стерильные растворы для инъекций, которые могут содержать антиоксиданты, буферы, бактериостаты и растворенные вещества, которые делают композицию изотоничной с кровью предполагаемого реципиента; и водные и неводные стерильные суспензии, которые могут содержать суспендирующие вещества и загустители. Можно использовать любую обычную среду в любой из композиций, за исключением случая, когда она несовместима с активным ингредиентом и его намеченным применением.
Композиции могут быть представлены в разовой дозе или в виде множества доз в контейнере, например, в запаянных ампулах и флаконах, и они могут храниться в высушенном вымораживанием (лиофилизированном) состоянии, для которого требуется только добавление стерильного жидкого носителя, например, воды для инъекций, непосредственно перед применением.
Молекулы нуклеиновой кислоты, полипептиды и антитела антагониста CXCL13 могут быть включены в фармацевтические композиции, подходящие для введения. Как правило, такие композиции содержат молекулу нуклеиновой кислоты, белок или антитело и фармацевтически приемлемый носитель. Используемый в данном документе термин «фармацевтически приемлемый носитель» включает любой или все растворители, дисперсионную среду, оболочки, антибактериальные и противогрибковые средства, изотонические и замедляющие всасывание вещества и тому подобное, совместимые с фармацевтическим введением. Применение такой среды и агентов для фармацевтически активных соединений хорошо известно в данной области. Можно использовать любую обычную среду или любой агент в композициях, за исключением случая, когда они несовместимы с активным соединением. В композиции также могут быть включены дополнительные активные соединения.
В другом аспекте изобретение относится к антагонистам CXCL13, описанным в данном документе, которые модифицированы ковалентным присоединением группы. Подобные модификации могут привести к получению антагониста CXCL13 с улучшенными фармакокинетическими свойствами (например, увеличенным периодом полужизни в сыворотке крови in vivo). Органическая группа может представлять собой линейную или разветвленную гидрофильную полимерную группу, жирную кислоту или эфир жирной кислоты. В конкретных вариантах осуществления гидрофильная полимерная группа может иметь молекулярную массу примерно от 800 до примерно 120000 дальтон, и может представлять собой полиалкиленгликоль (например, полиэтиленгликоль (ПЭГ), полипропиленгликоль (ППГ)), углеводный полимер, аминокислотный полимер или поливинилпирролидон, и группы жирной кислоты или эфира жирной кислоты могут содержать примерно от 8 до примерно 40 атомов углерода. Используемый в данном документе термин «жирная кислота» включает монокарбоновые кислоты и дикарбоновые кислоты. Жирные кислоты и эфиры жирных кислот, подходящие для модификации антител по изобретению, могут быть насыщенными или могут содержать одну или несколько ненасыщенных связей. Жирные кислоты, которые подходят для модификации антител по изобретению, включают, например, н-додеканоат (С12 лаурат), н-тетрадеканоат (С14 миристат), н-октадеканоат (С18 стеарат), н-эйкозаноат (С20 арахидат), н-докозаноат (С22 бегенат), н-триаконтаноат (С30), н-тетраконтаноат (С40), цис-дельта 9-октадеканоат (С18 олеат), полностью цис-дельта 5,8,11,14-эйкозатетраеноат (С20 арахидонат), октандикарбоновую кислоту, тетрадекандикарбоновую кислоту, октадекандикарбоновую кислоту, докозандикарбоновую кислоту и тому подобное. Подходящие эфиры жирных кислот включают моноэфиры дикарбоновых кислот, которые содержат линейную или разветвленную низшую алкильную группу. Низшая алкильная группа может содержать от 1 до 12, предпочтительно от 1 до 6 атомов углерода.
Модифицированные человеческие полипептиды и антитела можно получить с использованием подходящих способов, таких как взаимодействие с одним или несколькими модифицирующими агентами. Используемый в данном документе термин «модифицирующий агент» относится к подходящей органической группе (например, гидрофильному полимеру, жирной кислоте, эфиру жирной кислоты), которая содержит активирующую группу. «Активирующая группа» является химической группой или функциональной группой, которая в определенных условиях взаимодействует со второй химической группой, образуя, тем самым, ковалентную связь между модифицирующим агентом и второй химической группой. Например, амин-реактивные активирующие группы включают электрофильные группы, такие как тозилат, мезилат, галоген (хлор, бром, фтор, йод), N-гидроксисукцинимидиловые эфиры (NHS) и тому подобное. Активирующие группы, которые могут взаимодействовать с тиолами, включают, например, малеимид, йодацетил, акрилолил, пиридилдисульфиды, 5-тиол-2-нитробензойную кислоту-тиол (TNB-тиол) и тому подобное. Альдегидную функциональную группу можно сочетать с амин- или гидразидсодержащими молекулами, и азидная группа может взаимодействовать с трехвалентным фосфором с образованием фосфорамидатных или фосфоримидных связей. В данной области известны подходящие способы введения активирующих групп в молекулы (см., например, Hermanson G.T., Bioconjugate Techniques, Academic Press: San Diego, CA (1996)).
Изобретение включает способы получения фармацевтических композиций для модуляции экспрессии или активности полипептида, нуклеиновой кислоты или антитела CXCL13. Такие способы включают объединение фармацевтически приемлемого носителя со средством, которое модулирует экспрессию или активность полипептида, нуклеиновой кислоты или антитела CXCL13. Подобные композиции могут дополнительно содержать дополнительные активные соединения. Таким образом, изобретение дополнительно включает способы получения фармацевтической композиции объединением фармацевтически приемлемого носителя со средством, которое модулирует экспрессию или активность полипептида, нуклеиновой кислоты или антитела CXCL13, и одного или нескольких дополнительных активных соединений.
Средство, которое модулирует экспрессию или активность, например, может представлять собой малую молекулу. Например, такие малые молекулы включают пептиды, пептидомиметики, аминокислоты, аналоги аминокислот, полинуклеотиды, аналоги полинуклеотидов, нуклеотиды, аналоги нуклеотидов, органические или неорганические соединения (т.е. включая гетероорганические и металлорганические соединения) с молекулярной массой менее чем примерно 10000 г на моль, органические или неорганические соединения с молекулярной массой менее чем примерно 5000 г на моль, органические или неорганические соединения с молекулярной массой менее чем примерно 1000 г на моль, органические или неорганические соединения с молекулярной массой менее чем примерно 500 г на моль и соли, сложные эфиры и другие фармацевтически приемлемые формы таких соединений.
Следует понимать, что соответствующие дозы средств с малой молекулой и белковых или полипептидных средств зависят от ряда факторов, известных врачам, ветеринарным врачам или исследователям в данной области. Доза(ы) данных средств будет варьировать, например, в зависимости от идентичности, размера и состояния субъекта или образца, подлежащего лечению, дополнительно в зависимости от пути, которым вводят композицию, если он применим, и эффекта, который практикующий врач желает получить при введении полипептида, нуклеиновой кислоты или антитела CXCL13. Примерные дозы малых молекул включают количества в мг или мкг на кг массы субъекта или образца (например, примерно от 1 мкг на кг до примерно 500 мг на кг, примерно от 100 мкг на кг до примерно 5 мг на кг или примерно от 1 мкг на кг до примерно 50 мкг на кг). Примерные дозы белка или полипептида включают количества в г, мг или мкг на кг массы субъекта или образца (например, примерно от 1 мкг на кг до примерно 5 г на кг, примерно от 100 мкг на кг до примерно 500 мг на кг или примерно от 1 мг на кг до примерно 50 мг на кг). Кроме того, необходимо понимать, что соответствующие дозы одного из данных средств зависят от эффективности средства в отношении экспрессии или активности, подвергаемых модуляции. Такие соответствующие дозы могут быть определены с использованием тестов, описанных в данном документе.
В том случае, если одно или несколько средств вводят животному (например, человеку) для модуляции экспрессии или активности полипептида, нуклеиновой кислоты или антитела CXCL13, то врач, ветеринарный врач или исследователь могут, например, прописать вначале использовать относительно низкую дозу с последующим повышением дозы до получения соответствующей ответной реакции. Кроме того, следует понимать, что конкретная доза для любого конкретного животного будет зависеть от различных факторов, включая активность конкретного используемого средства, возраста, массы тела, общего состояния здоровья, пола и рациона субъекта, времени введения, пути введения, скорости выделения и комбинации лекарственных средств, а также степени экспрессии или активности, подвергаемых модуляции.
Фармацевтическую композицию по изобретению готовят так, чтобы она была совместимой с предназначаемым путем введения. Примеры путей введения включают парентеральный, например, внутривенный, внутрикожный, подкожный, пероральный (например, ингаляционный или буккальный), трансдермальный (местный), введение через слизистую и ректальное введение. Растворы или суспензии, используемые для парентерального, внутрикожного или подкожного введения, могут включать следующие компоненты: стерильный разбавитель, такой как вода для инъекций, физиологический раствор, жирные масла, полиэтиленгликоли, глицерин, пропиленгликоль или другие синтетические растворители; антибактериальные средства, такие как бензиловый спирт или метилпарабены; антиоксиданты, такие как аскорбиновая кислота или бисульфит натрия; хелатообразующие соединения, такие как этилендиаминтетрауксусная кислота; буферы, такие как ацетаты, цитраты или фосфаты, и вещества для регулирования тоничности, такие как хлорид натрия или декстроза. Значение рН можно регулировать с помощью кислот или оснований, таких как соляная кислота или гидроксид натрия. Препарат для парентерального введения может быть заключен в ампулы, одноразовые шприцы или флаконы с множеством доз, изготовленные из стекла или пластика.
Фармацевтические композиции, подходящие для инъекционного введения, включают стерильные водные растворы (в случае, когда соединение растворимо в воде) или дисперсии, и стерильные порошки для экстемпорального приготовления стерильных растворов или дисперсий для инъекций. Для внутривенного введения подходящие носители включают физиологический раствор, бактериостатическую воду, Cremophor EL (BASF; Parsippany, N.J.) или забуференный фосфатом физиологический раствор (PBS). Во всех случаях композиция должна быть стерильной и должна быть текучей, чтобы ее можно было легко набирать шприцом. Она должна быть стабильной в условиях производства и хранения и должна содержать консерванты во избежание загрязнения микроорганизмами, такими как бактерии и грибы. Носителем может быть растворитель или дисперсионная среда, например, вода, этанол, полиол (например, глицерин, пропиленгликоль и жидкий полиэтиленгликоль и тому подобное) и их подходящие смеси. Необходимую текучесть можно поддерживать, например, путем использования оболочки, такой как лецитин, путем сохранения необходимого размера частиц в случае дисперсии и путем применения поверхностно-активных веществ. Во многих случаях предпочтительно включать в композицию изотонические агенты, например, сахара, полиспирты, такие как маннит, сорбит, или хлорид натрия. Пролонгированного всасывания композиций для инъекций можно добиться включением в композицию вещества, которое замедляет всасывание, например, моностеарата алюминия и желатина. Фармацевтические эксципиенты и добавки, пригодные для стабилизации настоящей композиции, включают, но не ограничиваются ими, полипептиды, пептиды, аминокислоты, липиды и углеводы (например, сахара, включая моносахариды, ди-, три-, тетра- и олигосахариды; дериватизированные сахара, такие как алдитолы, альдоновые кислоты, этерифицированные сахара и тому подобное; и полисахариды или полимеры сахаров), которые могут присутствовать отдельно или в комбинации, составляя по отдельности или в комбинации 1-99,99% по массе или объему. Приводимые в качестве примера, но не ограничивающиеся ими, полипептидные эксципиенты включают сывороточный альбумин, такой как человеческий сывороточный альбумин (HAS), рекомбинантный человеческий альбумин (rHA), желатин, казеин и тому подобное. Показательные примеры аминокислот, которые также могут действовать как соединения, обладающие буферирующей способностью, включают аланин, глицин, аргинин, бетаин, гистидн, глутаминовую кислоту, аспарагиновую кислоту, цистеин, лизин, лейцин, изолейцин, валин, метионин, фенилаланин, аспартам и тому подобное. Одной из предпочтительных аминокислот является глицин.
Стерильные инъекционные растворы можно приготовить включением активного соединения (например, полипептида или антитела) в необходимом количестве в соответствующий растворитель с одним ингредиентом или комбинацией ингредиентов, перечисленных выше, при необходимости, с последующей стерилизацией фильтрованием. Как правило, дисперсии готовят включением активного соединения в стерильный растворитель, который содержит основную дисперсионную среду, и затем включением необходимых других ингредиентов из перечисленных выше. В случае стерильных порошков для последующего получения стерильных инъекционных растворов, предпочтительными способами приготовления являются вакуумная сушка и лиофилизация, которые обеспечивают порошок активного ингредиента плюс любой другой дополнительный желательный ингредиент из его предварительно стерилизованного фильтрованием раствора. Не ограничивающие примеры и способы получения таких стерильных растворов хорошо известны в данной области, например, описаны, но не ограничены этим, Gennaro, ed., Remington's Pharmaceutical Sciences, 18th Edition, Mack Publishing Co., (Easton, PA) 1990.
Композиции для приема внутрь обычно содержат инертный разбавитель или пищевой носитель. Они могут быть заключены в желатиновые капсулы или спрессованы в таблетки. Для перорального терапевтического введения активное соединение можно включать с эксципиентами и использовать в виде таблеток, пастилок или капсул. Композиции для перорального введения также можно приготовить с использованием жидкого носителя для применения в качестве жидкости для полоскания рта, где соединение в жидком носителе применяют перорально в виде полосканья и выплевывают или проглатывают.
Фармацевтически совместимые связующие средства и/или адъюванты можно включать в качестве части композиции. Таблетки, пилюли, капсулы, пастилки и тому подобное могут содержать любой из следующих ингредиентов или соединений аналогичной природы: связующее вещество, такое как микрокристаллическая целлюлоза, трагакантовая камедь или желатин; эксципиент, такой как крахмал или лактоза, разрыхлитель, такой как альгиновая кислота, примогель или кукурузный крахмал; смазывающее вещество (лубрикант), такое как стеарат магния или стеротес; вещество, способствующее скольжению, такое как коллоидный диоксид кремния; подсластитель, такой как сахароза или сахарин; или вкусовая добавка, такая как перечная мята, метилсалицилат или вещество со вкусом и запахом апельсина.
Для ингаляционного введения соединения вводят в форме аэрозольных частиц из находящегося под давлением контейнера или распылительного устройства, содержащего подходящий пропеллент, например, газ, такой как диоксид углерода, или из пульверизатора. Альтернативно, композиции, приготовленные в виде частиц, можно распылять с помощью электростатических, механических средств, включая вибрационные или ультразвуковые средства, описанные в патентах США №№: 4530464; 4533082; 5838350; 6113001; 6514496; 5518179; 5152456; 5261601 и 4605167.
Системное введение также можно осуществлять через слизистую или через кожу. Для введения через слизистую или через кожу в композиции используют соединения, подходящие для проникновения через барьер. Такие проникающие соединения обычно известны в данной области, и они включают, например, для введения через слизистую, детергенты, желчные соли и производные фусидовой кислоты. Введение через слизистую может быть выполнено посредством использования назальных спреев или суппозиториев. Для введения через кожу активные соединения предоставляют в виде мазей, бальзамов, гелей или кремов, как правило, известных в данной области.
Соединения можно также приготовить в форме суппозиториев (например, с использованием обычных основ для суппозиториев, таких как масло какао и другие глицериды) или удерживающих клизм для ректального введения.
В одном варианте осуществления активные соединения получают с носителями, которые будут защищать соединение от быстрого выделения из организма, в виде композиции с контролируемым высвобождением, включая имплантаты и микроинкапсулированные системы доставки. Можно использовать биодеградируемые, биосовместимые полимеры, такие как этиленвинилацетат, полиангидриды, полигликолевая кислота, коллаген, полиортоэфиры и полимолочная кислота. Способы получения таких композиций известны специалистам в данной области. Также можно использовать липосомальные суспензии (включая липосомы, содержащие введенные в них или на них моноклональные антитела) в качестве фармацевтически приемлемых носителей. Особенно предпочтительные композиции и способы описаны в патентах США №№ 5891468 и 6316024.
Особенно предпочтительно получать композиции для приема внутрь или парентерального введения в виде единичной лекарственной формы для простоты введения и однородности дозировки. Используемый в данном документе термин «единичная лекарственная форма» относится к физически дискретным единицам, подходящим для использования в качестве единичных лекарственных форм для субъекта, подвергаемого лечению; каждую единицу, содержащую заранее определенное количество активного соединения, рассчитывают для обеспечения желаемого терапевтического эффекта в сочетании с необходимым фармацевтическим носителем. Подробное определение единичных лекарственных форм по изобретению диктуется и непосредственно зависит от уникальных характеристик активного соединения и конкретного терапевтического эффекта, которого желательно достичь, и ограничений, известных в данной области, для получения препаратов такого активного соединения для лечения индивидуумов.
Для антител предпочтительная доза составляет примерно от 0,1 мг/кг до 100 мг/кг массы тела (как правило, примерно от 10 мг/кг до 20 мг/кг). Если антитело должно проявлять свое действие в мозге, то, как правило, подходящей является доза примерно от 50 мг/кг до 100 мг/кг. Как правило, не полностью человеческие антитела и полностью человеческие антитела имеют более длинный период полужизни в организме человека по сравнению с другими антителами. Следовательно, возможно применение более низких доз и меньшей кратности введения. Можно использовать модификации, такие как липидизация, для стабилизации антител и повышения захвата и проникновения в ткани (например, в мозг). Способ липидизации антител описан Cruikshank et al. ((1997) J. Acquired Immune Deficiency Syndromes and Human Retrovirology 14:193).
Молекулы нуклеиновой кислоты антагониста CXCL13 можно вставить в векторы и использовать в качестве векторов для генной терапии. Векторы для генной терапии можно вводить субъекту, например, с помощью внутривенной инъекции, местного применения (патент США № 5328470) или стереотаксической инъекции (см., например, Chen et al. (1994) Proc. Natl. Acad. Sci. USA 91:3054-3057). Фармацевтический препарат вектора для генной терапии может содержать вектор для генной терапии в приемлемом разбавителе или может содержать матрицу для медленного высвобождения, в которую включен носитель для доставки гена. Альтернативно, в тех случаях, когда полный вектор для доставки гена можно получить интактным из рекомбинантных клеток, например ретровирусные векторы, фармацевтический препарат может содержать одну или несколько клеток, продуцирующих систему доставки генов.
Фармацевтические композиции можно поместить в контейнер, упаковку или распределительное устройство вместе с инструкциями по применению.
Фармакогеномика
Средства или модуляторы, обладающие стимулирующим или ингибирующим действием на активность или экспрессию полипептида CXCL13, идентифицированные методом скринингового анализа, описанного в данном документе, можно вводить индивидуумам для лечения (профилактического или медикаментозного) нарушений, связанных с аномальной активностью полипептида. В связи с таким лечением может быть полезным учитывать фармакогеномику субъекта (т.е. изучение взаимосвязи между генотипом субъекта и ответной реакцией субъекта на введение чужеродного соединения или лекарственного средства). Различия в метаболизме лекарственных препаратов могут привести к проявлению тяжелых токсических реакций или отсутствию терапевтического эффекта в результате изменения взаимосвязи между дозой и концентрацией фармакологически активного лекарственного средства в крови. Таким образом, данные по фармакогеномике индивидуума позволяют выбрать эффективные средства (например, лекарственные средства) для профилактического или медикаментозного лечения с учетом генотипа индивидуума. Такие данные по фармакогеномике можно дополнительно использовать для определения соответствующих доз и терапевтических схем. Соответственно, можно определить активность полипептида CXCL13, экспрессию нуклеиновой кислоты CXCL13 или мутации гена CXCL13 у индивидуума для выбора, тем самым, соответствующего средства(в) для медикаментозного или профилактического лечения индивидуума.
Фармакогеномика связана с клинически значимыми наследственными вариациями в ответной реакции на лекарственные средства за счет изменений в распределении лекарственного средства и аномального действия у больных людей. См., например, Linder (1997) Clin. Chem. 43(2):254-266. В основном, можно выделить два типа фармакогенетических признаков. Генетические признаки, передаваемые в виде одного фактора, приводящего к изменению действия лекарственных средств на организм, относятся к «измененному действию лекарственного средства». Генетические признаки, передаваемые в виде единичных факторов, приводящих к изменению влияния организма на лекарственные средства, относятся к «измененному метаболизму лекарственного средства». Эти фармакогенетические признаки могут иметь место в виде редких аномалий или в виде полиморфизмов. Например, дефицит глюкозо-6-фосфатдегидрогеназы (G6PD) представляет собой распространенную наследуемую энзимопатию, основным клиническим проявлением которой является гемолиз после приема лекарственных средств со свойствами окислителей (противомалярийные препараты, сульфонамиды, анальгетики, нитрофураны) и потребления бобов.
В качестве иллюстративного варианта осуществления, активность ферментов, метаболизирующих лекарственное средство, является основным определяющим фактором как интенсивности, так и продолжительности действия лекарственного средства. Открытие генетического полиморфизма ферментов, метаболизирующих лекарственное средство (например, N-ацетилтрансферазы 2 (NAT 2) и ферментов цитохрома Р450 CYP2D6 и CYP2D19) предоставило объяснение того, почему у некоторых пациентов не проявляется ожидаемое действие лекарственного средства или у некоторых пациентов развивается сильная ответная реакция и тяжелые токсические эффекты после приема обычной и безопасной дозы лекарственного средства. Данный полиморфизм проявляется в наличии двух фенотипов в популяции, «быстрых метаболизаторов» (EM) и «медленных метаболизаторов» (РМ). Преобладание РМ различно среди различных групп населения. Например, ген, кодирующий CYP2D6, является высоко полиморфным, и у PM было выявлено наличие нескольких мутаций, которые все приводят к отсутствию функционального CYP2D6. У «медленных метаболизаторов» CYP2D6 и CYP2D19 довольно часто наблюдают сильную ответную реакцию на лекарственное средство и побочные эффекты при приеме обычных доз. Если метаболит принадлежит к активной терапевтической группе, то у РМ будет отсутствовать терапевтическая реакция, как показано на примере анальгетического действия кодеина, опосредуемого его метаболитом морфином, который образуется при участии CYP2D6. Другими экстремальными субъектами являются так называемые «сверхбыстрые метаболизаторы», которые не реагируют на стандартные дозы. Недавно было установлено, что молекулярной основой сверхбыстрого метаболизма является амплификация гена CYP2D6.
Таким образом, активность полипептида CXCL13, экспрессию нуклеиновой кислоты, кодирующей полипептид, или мутацию в гене, кодирующем полипептид у индивидуума, можно определить при выборе средства(в) для медикаментозного или профилактического лечения индивидуума. Кроме того, результаты фармакогенетических исследований можно использовать для генотипирования полиморфных аллелей, кодирующих ферменты, метаболизирующие лекарственное средство, для идентификации фенотипа реактивности индивидуумов на лекарственное средство. Используя эти данные при выборе дозы и лекарственного средства, можно избежать побочных реакций или отсутствия эффективности лечения и, таким образом, повысить терапевтическую или профилактическую эффективность при лечении субъекта модулятором активности или экспрессии полипептида, такого как модулятор, идентифицированный в одном из приведенных в качестве примера скрининговых анализов, описанных в данном документе.
Способы лечения
Настоящее изобретение относится к профилактическим и терапевтическим способам лечения субъекта при риске развития (или расположенного к) нарушения или страдающего нарушением, связанным с аномальной экспрессией или активностью полипептида CXCL13, и/или в развитии которого принимает участие CXCL13.
Настоящее изобретение относится к способу модуляции или лечения, по меньшей мере, одного связанного с CXCL13 заболевания или состояния, на уровне клетки, ткани, органа, организма животного или пациента, известного в данной области или описанного в данном документе, с использованием, по меньшей мере, одного антагониста CXCL13.
Композиции антагонистов CXCL13 могут найти терапевтическое применение при лечении заболеваний, связанных с легкими, таких как астма, эмфизема, COPD и хроническое заболевание легких у новорожденных.
Настоящее изобретение также относится к способу модуляции или лечения, по меньшей мере, одного связанного с иммунной системой заболевания, на уровне клетки, ткани, органа, организма животного или пациента, включая, но не ограничиваясь ими, ревматоидный артрит, ювенильный ревматоидный артрит, системный ревматодный артрит с началом в юношеском возрасте, псориатический артрит, анкилозирующий спондилит, язву желудка, серонегативные артропатии, остеоартрит, воспалительное заболевание кишечника, язвенный колит, системную красную волчанку, антифосфолипидный синдром, иридоциклит/увеит/неврит зрительного нерва, идиопатический фиброз легких, системный васкулит/грануломатоз Вегенера, саркоидоз, орхит/полную вазэктомию, аллергические/атопические заболевания, аллергический ринит, экзему, аллергический контактный дерматит, аллергический конъюнктивит, пневмонию на фоне гиперчувствительности, трансплантаты, отторжение трансплантатов органов, синдром трансплантат против хозяина, синдром системной воспалительной ответной реакции, сепсис, сепсис, вызванный грамположительными бактериями, сепсис, вызванный грамотрицательными бактериями, культуральный отрицательный сепсис, сепсис, вызванный грибками, нейтропеническую лихорадку, уросепсис, менингококцемию, травму/кровотечение, ожоги, воздействие ионизирующей радиации, острый панкреатит, респираторный дистресс-синдром взрослых, ревматоидный артрит, вызванный алкоголем гепатит, хронические воспалительные патологии, саркоидоз, болезнь Крона, серповидно-клеточную анемию, диабет, нефроз, атопические заболевания, реакции гиперчувствительности, аллергический ринит, сенную лихорадку, перениальный ринит, конъюнктивит, эндометриоз, крапивницу, системную анафилаксию, дерматит, пернициозную анемию, гемолитическое заболевание, тромбоцитопению, отторжение трансплантата любого органа или ткани, отторжение трансплантата почек, отторжение трансплантата сердца, отторжение трансплантата печени, отторжение трансплантата поджелудочной железы, отторжение трансплантата легких, отторжение трансплантата костного мозга (BMT), отторжение кожного аллотрансплантата, отторжение хрящевого трансплантата, отторжение костного трансплантата, отторжение трансплантата тонкого кишечника, отторжение имплантата тимуса плода, отторжение трансплантата паращитовидной железы, отторжение ксенотрансплантата любого органа или ткани, отторжение аллотрансплантата, реакции антирецепторной гиперчувствительности, болезнь Граве, болезнь Рейно, инсулинрезистентный диабет типа В, злокачественную миастению, антитело-опосредованную цитотоксичность, реакции гиперчувствительности типа III, системную красную волчанку, синдром POEMS (полиневропатию, органомегалию, эндокринопатию, моноклональную гаммопатию и синдром кожных изменений), антифосфолипидный синдром, пемфигус, склеродерму, смешанное заболевание соединительной ткани, идиопатическую болезнь Аддисона, сахарный диабет, хронический активный гепатит, первичный цирроз желчного пузыря, витилиго, васкулит, пост-MI кардиотомический синдром, гиперчувствительность типа IV, контактный дерматит, пневмонию на фоне гиперчувствительности, отторжение аллотрансплантата, гранулему, вызванную внутриклеточными микроорганизмами, чувствительность к лекарственным средствам, метаболическую/идиопатическую болезнь, болезнь Вильсона, гематохроматоз, дефицит альфа-1-антитрипсина, диабетическую ретинопатию, тиреоидит Хашимото, остеопороз, нарушения оси гипоталамус-гипофиз-надпочечники, первичный билиарный цирроз, тиреоидит, энцефаломиелит, кахексию, муковисцидоз, семейный гемофагоцитарный лимфогистиоцитоз, дерматологические заболевания, псориаз, алопецию, нефротический синдром, нефрит, гломерулярный нефрит, острую почечную недостаточность, гемодиализ, уремию, токсичность, преэклампсию, okt3 терапию, анти-cd3 терапию, терапию цитокинами, химиотерапию, последствия лучевой терапии (например, включая, но не ограничиваясь ими, тоастению, анемию, кахексию и тому подобное), хроническую интоксикацию салицилатами и тому подобное. См., например, Merck Manual, 12th-17th Editions, Merck & Company, Rahway, NJ (1972, 1977, 1982, 1987, 1992, 1999), Pharmacotherapy Handbook, Wells et al., eds., Second Edition, Appleton and Lange, Stamford, Conn. (1998, 2000), каждый из этих источников включен в данный документ путем ссылки.
Настоящее изобретение также относится к способу модуляции или лечения, по меньшей мере, одного злокачественного заболевания на уровне клетки, ткани, органа, организма животного или пациента, включая, но не ограничиваясь ими, по меньшей мере, одно из следующих заболеваний: лейкемия, острая лимфобластная лейкемия (ALL), В-клеточная, Т-клеточная или FAB ALL, острая миелоидная лейкемия (AML), острая промиелоцитарная лейкемия (APL), хроническая миелоцитарная лейкемия (СML), хроническая лимфоцитарная лейкемия (СLL), волосатоклеточный лейкоз, миелодиспластический синдром (MDS), лимфома, болезнь Ходжкина, злокачественная лимфома, неходжкинская лимфома, лимфома Беркитта, множественная миелома, саркома Капоши, карцинома прямой и ободочной кишки, карцинома поджелудочной железы, карцинома носоглотки, злокачественный гистиоцитоз, паранеопластический синдром/гиперкальцемия при злокачественном заболевании, солидные опухоли, аденокарциномы, саркомы, злокачественная меланома, гемангиома, метастазы, резорбция костей, связанная со злокачественным заболеванием, костные боли, связанные со злокачественной опухолью, и тому подобное.
Нарушения, характеризующиеся аномальной экспрессией или активностью полипептидов CXCL13, дополнительно описаны в различных разделах данной заявки.
1. Профилактические способы
В одном аспекте изобретение относится к способу, по меньшей мере, существенной профилактики у субъекта заболевания или состояния, связанного с аномальной экспрессией или активностью полипептида CXCL13, введением субъекту средства, которое модулирует экспрессию или, по меньшей мере, один тип активности полипептида. Субъектов с риском заболевания, которое вызывается или в развитие которого вносит свой вклад аномальная экспрессия или активность полипептида CXCL13, можно выявить, например, любым одним из или комбинацией диагностических или прогностических тестов, описанных в данном документе. Введение профилактического средства можно провести до проявления симптомов, характерных для аномалии, так чтобы предотвратить заболевание или нарушение, или, альтернативно, замедлить его прогрессирование. В зависимости от типа аномалии можно использовать, например, агонист или антагонист для лечения субъекта. Подходящее средство можно определить, основываясь на результатах скрининговых анализов, описанных в данном документе.
2. Терапевтические способы
Другой аспект изобретения относится к способам модуляции экспрессии или активности полипептида CXCL13 в терапевтических целях. Способ модуляции по изобретению включает контактирование клетки со средством, которое модулирует один или несколько типов активности полипептида. Средство, которое модулирует активность, может быть средством, описанным в данном документе, таким как нуклеиновая кислота или белок, природный лиганд полипептида, пептид, пептидомиметик или другая малая молекула. В одном варианте осуществления средство стимулирует один или несколько типов биологической активности полипептида. В другом варианте осуществления средство ингибирует один или несколько типов биологической активности полипептида. Примеры таких ингибиторов включают молекулы антисмысловых нуклеиновых кислот и антитела, и другие способы описаны в данном документе. Данные способы модуляции можно проводить in vitro (например, культивированием клетки со средством) или, альтернативно, in vivo (например, введением средства субъекту). При этом настоящее изобретение относится к способам лечения индивидуума, страдающего заболеванием или нарушением, характеризующимся аномальной экспрессией или активностью полипептида CXCL13. В одном варианте осуществления способ включает введение средства (например, средства, выявленного по результатам скринингового анализа, описанного в данном документе) или комбинации средств, которые модулируют (например, повышают или снижают) экспрессию или активность. Ингибирование активности желательно в ситуациях, при которых активность или экспрессия являются аномально высокими или повышенными, и/или при которых снижение активности, возможно, будет оказывать положительное действие.
Способ по настоящему изобретению включает введение эффективного количества композиции или фармацевтической композиции, содержащей, по меньшей мере, один антагонист CXCL13 или его специфический участок или вариант, в клетку, ткань, орган, животному или пациенту, нуждающемуся в такой модуляции, лечении или терапии. Такой способ может необязательно дополнительно включать совместное введение или проведение комбинированной терапии для лечения таких иммунных заболеваний, где введение указанного, по меньшей мере, одного антагониста CXCL13, его специфического участка или варианта дополнительно включает введение до, одновременно и/или после, по меньшей мере, одного соединения, выбранного из глутирамера ацетата (например, Сорахоnе), циклофасфамида, азатиоприна, глюкокортикостероидов, метотрексата, паклитаксела, 2-хлордезоксиаденозина, митоксантрона, IL-10, TGBb, CD4, CD52, антегрена, CD11, CD18, TNF-альфа, IL-1, IL-2 и/или СD4 антитела или слитого антитела против рецептора, интерферона альфа, иммуноглобулина, лисмида (Requinimax™), инсулиноподобного фактора-1 роста (IGF-1), элпродила, пирфенидона, миелина для перорального введения, или соединений, которые воздействуют на одно или более, по меньшей мере, из следующего: аутоиммунную супрессию деструкции миелина, иммунную регуляцию, активацию, пролиферацию, миграцию и/или супрессорную функцию Т-клеток, ингибирование взаимодействия рецептора Т-клеток/пептида/МНС-II, индукцию анергии Т-клеток, истощение аутореактивных Т-клеток, снижение проникновения через гематоэнцефалический барьер, изменение баланса провоспалительных (Th1) и иммуномодуляторных (Th2) цитокинов, подавление ингибиторов матричной металлопротеазы, нейропротекцию, ослабление глиоза, стимуляцию ремиелинизации), антагонистов TNF (например, но не ограничиваясь ими, TNF Ig белка или фрагмента, растворимого рецептора TNF или фрагмента, его слитых белков или антагониста TNF с малой молекулой), противоревматического средства, мышечного релаксанта, наркотического средства, нестероидного противовоспалительного препарата (НПВС), анальгетика, анестетика, седативного средства, местного анестетика, нейромышечного блокатора, противомикробного препарата (например, аминогликозида, противогрибкового препарата, противопаразитарного препарата, противовирусного препарата, карбапенема, цефалоспорина, фторхинолона, макролида, пенициллина, сульфонамида, тетрациклина, другого противомикробного препарата), средства против псориаза, кортикостероида, анаболического стероида, минерального вещества, питательного вещества, препарата для лечения заболеваний щитовидной железы, витамина, гормона, связанного с обменом кальция, средства против диареи, противокашлевого средства, противорвотного препарата, противоязвенного препарата, слабительного, антикоагулянта, эритропоэтина (например, эпоэтина альфа), филграстина (например, G-CSF, Neupogen), сарграмостима (GM-CSF, Leukine), препарата для иммунизации, иммуноглобулина, иммуносупрессора (например, базиликсимаба, циклоспорина, даклизумаба), гормона роста, препарата для заместительной гормональной терапии, модулятора рецептора эстрогенов, мидриатического средства, циклоплегического препарата, алкилирующего агента, антиметаболита, ингибитора митоза, радиоактивного препарата, антидепрессанта, препарата против маний, антипсихотического средства, анксиолитика, гипнотического препарата, симпатомиметика, стимулятора, донепезила, такрина, препарата для лечения астмы, бета-агониста, стероида для ингаляционного введения, ингибитора лейкотриенов, метилксантина, кромолина, эпинефрина или его аналога, дорназы альфа (Pulmozyme), цитокина или антагониста цитокина. Подходящие дозы хорошо известны в данной области. См., например, Wells et al., eds., Pharmacotherapy Handbook, 2nd Edition, Appleton and Lange, Stamford, CT (2000); PDR Pharmacopoeia, Tarascon Pocket Pharmacopoeia 2000, Deluxe Edition, Tarascon Publishing, Loma Linda, CA (2000), каждый из этих источников полностью включен в данный документ путем ссылки.
Антагонисты TNF, подходящие для композиций, комбинированной терапии, совместного введения, устройств и/или способов по настоящему изобретению (дополнительно содержащие, по меньшей мере, одно антитело, специфический участок или его вариант по настоящему изобретению), включают, но не ограничиваются ими, анти-TNF-Ig белки, их антигенсвязывающие фрагменты и рецепторные молекулы, которые специфически связываются с TNF; соединения, которые предупреждают и/или ингибируют синтез TNF, высвобождение TNF или его воздействие на клетки-мишени, такие как талидомид, тенидап, ингибиторы фосфодиэстеразы (например, пентоксифиллин или ролипрам), агонисты рецептора аденозина A2b и энхансеры рецептора аденозина A2b; соединения, которые предупреждают и/или ингибируют сигнальный путь с участием рецептора TNF, такие как ингибиторы активированной митогеном протеинкиназы (МАР); соединения, которые блокируют и/или ингибируют мембранное расщепление TNF, такие как ингибиторы металлопротеиназы; соединения, которые блокируют и/или ингибируют активность TNF, такие как ингибиторы ангиотензин-конвертирующего фермента (АСЕ) (например, каптоприл); и соединения, которые блокируют и/или ингибируют продукцию и/или синтез TNF, такие как ингибиторы МАР киназы.
Используемые в данном документе термины «фактор некроза опухолей Ig белок», «TNF Ig белок», «TNFα Ig белок» или его фрагмент и тому подобное относятся к факторам, которые снижают, блокируют, ингибируют, отменяют или нарушают активность TNFα in vitro, in situ и/или предпочтительно in vivo. Например, подходящий человеческий TNF Ig белок по настоящему изобретению может связывать TNFα и он включает анти-TNF Ig белки, их антигенсвязывающие фрагменты и специфические мутанты или домены, которые специфически связываются с TNFα. Подходящее антитело против TNF или его фрагмент также может снижать, блокировать, отменять, нарушать, предупреждать и/или ингибировать синтез РНК, ДНК или белка TNF, высвобождение TNF, сигнальный путь с участием рецептора TNF, мембранное расщепление TNF, активность TNF, продукцию и/или синтез TNF.
Химерный Ig белок сА2 состоит из высокоаффинной антигенсвязывающей вариабельной области нейтрализующего мышиного антитела против человеческого TNFα IgG1 Ig белка, обозначенного А2, и константных областей человеческого IgG1, каппа иммуноглобулина. Fc область человеческого IgG1 повышает эффекторную функцию аллогенного Ig белка, повышает период полужизни в сыворотке крови и снижает иммуногенность Ig белка. Авидность и специфичность эпитопа химерного Ig белка сА2 происходит от вариабельной области мышиного Ig белка А2. В конкретном варианте осуществления предпочтительным источником нуклеиновых кислот, кодирующих вариабельную область мышиного Ig белка А2, является линия клеток гибридомы А2.
Химерный А2 (сА2) нейтрализует цитотоксическое действие природного и рекомбинантного TNFα в зависимости от дозы. По данным теста связывания химерного Ig белка сА2 и рекомбинантного человеческого TNFα показано, что константа аффинности химерного Ig белка сА2 составляет 1,04×1010 М-1. Предпочтительные методы определения специфичности и аффинности моноклонального Ig белка конкурентным ингибированием можно найти у Harlow et al., Ig derived proteins: A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, 1988; Colligan et al., eds., Current protocols in Immunology, Greene Publishing Assoc. and Wiley Interscience, New York (1992-2003); Kozbor et al., Immunol. Today, 4:72-79 (1983); Ausubel et al., eds., Current Protocols in Мolecular Biology, Wiley Interscience, New York (1987-2003); и Muller, Meth. Enzymol., 92:589-601 (1983), каждый из этих источников полностью включен в данный документ путем ссылки.
В конкретном варианте осуществления мышиный моноклональный Ig белок А2 получают с использованием клеточной линии, обозначенной с134А. Химерный Ig белок сА2 получают с использованием клеточной линии, обозначенной с168А.
Дополнительные примеры моноклональных анти-TNF-Ig белков, которые можно использовать в настоящем изобретении, описаны в данной области (см., например, патент США № 5231024; Möller A. et al., Cytokine 2(3):162-169 (1990); заявка на патент США № 07/943852 (подана 11 сентября 1992); Rathjen et al., международная заявка WO 91/02078 (опубликована 21 февраля, 1991); Rubin et al., EPO публикация патента № 0218868 (опубликована 22 апреля 1987); Yone et al., EPO публикация патента № 0288088 (опубликована 26 октября 1988); Liang et al., Biochem. Biophys. Res. Comm. 137:847-854 (1986); Meager et al., Hybridoma 6:305-311 (1987); Fendly et al., Hybridoma 6:359-369 (1987); Bringman et al., Hybridoma 6:489-507 (1987); и Hirai et al., J. Immunol. Meth. 96:57-62 (1987), каждый из этих источников полностью включен в данный документ путем ссылки.
Рецепторные молекулы TNF
Предпочтительными молекулами рецепторов TNF, пригодными для настоящего изобретения, являются те, которые связываются с TNFα с высокой аффинностью (см., например, Feldmann et al., международная публикация № WO 92/07076 (опубликована 30 апреля 1992); Schall et al., Cell 61:361-370 (1990); и Loetscher et al., Cell 61:351-359 (1990), каждый из этих источников полностью включен в данный документ путем ссылки) и необязательно обладают низкой иммуногенностью. В частности, пригодными для настоящего изобретения являются клеточные поверхностные рецепторы TNF с молекулярной массой 55 кДа (p55 TNF-R) и 75 кДа (p75 TNF-R). Также подходящими для настоящего изобретения являются усеченные формы данных рецепторов, содержащие внеклеточные домены (ECD) рецепторов или их функциональные участки (см., например, Corcoran et al., Eur. J. Biochem. 223:831-840 (1994)). Усеченные формы TNF-рецепторов, содержащие ECD, были обнаружены в моче и сыворотке крови в виде ингибирующих связывающих белков TNFα с молекулярной массой 30 кДа и 40 кДа (Engelmann H. et al., J. Biol. Chem. 265:1531-1536 (1990)). Мультимерные молекулы рецептора TNF и слитые молекулы иммунорецептора TNF и их производные и фрагменты или участки являются дополнительными примерами молекул TNF-рецепторов, которые пригодны для способов и композиций по настоящему изобретению. Молекулы TNF-рецепторов, которые можно использовать в настоящем изобретении, характеризуются их способностью применяться для лечения пациентов в течение продолжительного периода времени с хорошим или превосходным ослаблением симптомов и низкой токсичностью. Низкая иммуногенность и/или высокая аффинность, а также другие неопределенные свойства, могут вносить свой вклад в обеспечение достигаемых результатов терапии.
Мультимерные молекулы TNF-рецепторов, пригодные для настоящего изобретения, содержат весь или функциональный участок ECD двух или более рецепторов TNF, связанных посредством одного или нескольких полипептидных линкеров или других непептидных линкеров, таких как полиэтиленгликоль (ПЭГ). Мультимерные молекулы могут дополнительно содержать сигнальный пептид секретированного белка для прямой экспрессии мультимерной молекулы. Данные мультимерные молекулы и способы их получения описаны в заявке на патент США № 08/437533 (подана 9 мая 1995), содержание которой полностью включено в данный документ путем ссылки.
Слитые молекулы TNF-иммунорецепторов, пригодные для способов и композиций по настоящему изобретению, содержат, по меньшей мере, один участок одной или нескольких иммуноглобулиновых молекул и весь или функциональный участок одного или нескольких TNF-рецепторов. Данные иммунорецепторные слитые молекулы можно «собрать» в виде мономеров или гетеро- или гомомультимеров. Иммунорецепторные слитые молекулы могут быть моновалентными или мультивалентными. Примером такой иммунорецепторной слитой молекулы является слитый белок TNF-рецептор/IgG. Иммунорецепторные слитые молекулы TNF и способы их получения описаны в данной области (Lesslauer et al., Eur. J. Immunol. 21:2883-2886 (1991); Ashkenazi et al., Proc. Natl. Acad. Sci. USA 88:10535-10539 (1991); Peppel et al., J. Exp. Med. 174:1483-1489 (1991); Kolls et al., Proc. Natl. Acad. Sci. USA 91:215-219 (1994); Butler et al., Cytokine 6(6):616-623 (1994); Baker et al., Eur. J. Immunol. 24:2040-2048 (1994); Beutler et al., патент США № 5447851; и заявка на патент США № 08/442133 (подана 16 мая 1995), каждый из этих источников полностью включен в данный документ путем ссылки). Способы получения иммунорецепторных слитых молекул также можно найти у Capon et al., патент США № 5116964; Capon et al., патент США № 5225538; и Capon et al., Nature 337:525-531 (1989), каждый из этих источников полностью включен в данный документ путем ссылки.
Функциональный эквивалентный вариант, производное, фрагмент или область молекулы TNF-рецептора относится к участку молекулы TNF-рецептора или участку последовательности молекулы TNF-рецептора, кодирующей молекулу TNF-рецептора, которая имеет достаточный размер и последовательности, которые в функциональном отношении «напоминают» молекулы TNF-рецептора, которые можно использовать в настоящем изобретении (например, они связываются с TNFα с высокой аффинностью и обладают низкой иммуногенностью). Функциональный эквивалент молекулы TNF-рецептора также включает молекулы модифицированного TNF-рецептора, которые функционально «напоминают» молекулы TNF-рецептора, и их можно использовать в настоящем изобретении (например, они связываются с TNFα с высокой аффинностью и обладают низкой иммуногенностью). Например, функциональный эквивалент молекулы TNF-рецептора может содержать кодон «SILENT» или одну или несколько аминокислотных замен, делеций или добавлений (например, замену одной кислой аминокислоты на другую кислую аминокислоту; или замену одного кодона, кодирующего одну и ту же или отличную гидрофобную аминокислоту на другой кодон, кодирующий гидрофобную аминокислоту). См. Ausubel et al., Current Protocols in Molecular Biology, Greene Publishing Assoc. and Wiley-Interscience, New York (1987-2003).
Несмотря на описание изобретения в общем плане, ниже будут дополнительно описаны варианты осуществления в последующих примерах, которые не следует рассматривать в качестве ограничивающих объем формулы изобретения.
Пример 1
Уровни транскриптов мРНК CXCL13 повышены в пораженных тканях легких
Уровни транскриптов мРНК, кодирующих белок CXCL13, повышены в тканях легких у не подвергавшихся обработке мышей SP-C/TNFα и мышей SP-C/TNFα, обработанных специфическими mAb против TNFα, по сравнению с контрольными мышами C57BL6 (фиг.1). Мыши SP-C/TNFα являются трансгенными мышами, полученными из штамма мышей C57BL6, которые конститутивно сверхэкспрессируют мышиный TNFα в тканях легких. Сверхэкспрессия TNFα в легких (у трансгенных мышей SP-C/TNFα) вызывает много патологических изменений, аналогичных обнаруженным у пациентов с COPD, включая воспаление легких, эмфизему, фиброз легких [16] и образование эктопических лимфоидных фолликулов. Мыши SP-C/TNFα являются моделью таких заболеваний легких, как воспалительная эмфизема легких, фиброз легких и застойное обструктивное заболевание легких (COPD). Образование эктопических лимфоидных фолликулов в легких является патологией, связанной с каждым из этих заболеваний легких. Эктопические лимфоидные фолликулы в легких могут образоваться посредством инфильтрации В-клеток в легкие и ткани других дыхательных органов. Результаты, приведенные на фиг.1, показывают, что повышенные уровни белка CXCL13 в тканях легких мышей SP-C/TNFα вносят свой вклад в образование эктопических лимфоидных фолликулов в легких и начало развития связанных с легкими заболеваний у данных животных.
В данном опыте 12-недельным контрольным мышам C57BL6 и необработанным мышам SP-C/TNFα вводили внутрибрюшинно 200 мкл PBS каждый понедельник и пятницу в течение 6 недель. Мышам SP-C/TNFα вводили 0,5 мг специфических mAb cV1q против TNFα в 200 мкл PBS внутрибрюшинно каждый понедельник и пятницу в течение 6 недель. mAb cV1q представляют собой моноклональные мышиные антитела изотипа IgG1, которые связываются с мышиным TNFα. mAb cV1q получали обычными способами. Через 6 недель после начала обработок мышей подвергали эвтаназии в соответствии с правилами по уходу и проведению работ с животными.
Легкие извлекали из убитых животных, гомогенизировали и экстрагировали и выделяли мРНК с использованием стандартных методов. Также проводили ОТ-ПЦР для мРНК CXCL13 и мРНК GAPDH, следуя стандартным методикам. Затем количественно определяли уровни мРНК CXCL13 и GAPDH с использованием стандартных методов и уровни мРНК CXCL13 нормализовали до уровней мРНК гена «домашнего хозяйства» GAPDH. Данные представлены в виде среднего значения ± стандартное отклонение нормализованных уровней мРНК CXCL13 для 5 различных мышей (фиг.1).
Пример 2
Совместная обработка моноклональными антителами, специфическими для TNFα и CXCL13, ослабляет симптомы заболевания легких
Совместная обработка моноклональными антителами, специфическими для TNFα и CXCL13, ослабляет симптомы заболевания легких у мышей SP-C/TNFα (фиг.2). Размер эктопических фолликулов был существенно ниже у мышей SP-C/TNFα, совместно обработанных mAb, специфическими против TNFα и CXCL13, по сравнению с контрольными необработанными животными (фиг.2). У трансгенных мышей SP-C/TNFα, обработанных mAb против TNFα в течение 8 недель, была существенно снижена инфильтрация дыхательных путей нейтрофилами, однако обработка оказывала незначительное влияние на эктопические лимфоидные фолликулы. Данный факт указывает на то, что образовавшись, эктопические лимфоидные фолликулы сохраняют свой гомеостаз независимо от TNFα.
В данном опыте 12-недельным контрольным мышам SP-C/TNFα проводили внутрибрюшинные инъекции каждый понедельник и пятницу в течение 6 недель. Контрольные мыши получали 200 мкл только PBS. Животным, которых обрабатывали антителами против TNFα, вводили 0,5 мг специфических mAb cV1q против TNFα в 200 мкл PBS. Животным, которых обрабатывали антителами против CXCL13, вводили 0,5 мг специфических mAb МАВ4701 против CXCL13 в 200 мкл PBS. Животные с совместной обработкой получали 0,5 мг специфических mAb cV1q против TNFα и 0,5 мг специфических mAb МАВ4701 против CXCL13 в 200 мкл PBS. МАВ4701 представляют собой моноклональные крысиные антитела изотипа IgG1, которые связываются с мышиным CXCL13. mAb МАВ4701 получали с использованием стандартных способов, и они являются коммерчески доступными (R&D Systems, Inc., Minneapolis, MN). Через 6 недель после начала инъекций мышей подвергали эвтаназии в соответствии с правилами по уходу и проведению работ с животными. Легкие извлекали из убитых животных, готовили срезы и препараты и проводили гистологические исследования для количественной оценки размера и образования эктопических фолликулов с использованием стандартных методов. Для проведения микроскопических исследований использовали микроскоп Phase 3 Imaging System (Glen Mills, PA) и соответствующее аналитическое программное обеспечение и определяли размер 10-20 фолликулов на препарат на мышь. Приведенные данные выражали в виде среднего значения ± стандартное отклонение, которое получали от 5 различных мышей из каждой опытной группы (фиг.2).
Была выдвинута гипотеза, что результаты, описанные выше, указывают на то, что монотерапия CXCL13 может быть пригодной для лечения связанных с дыхательной системой заболеваний. Поскольку на данной модели мышей имеет место повышенная экспрессия TNFα, то ингибирование TNFα вместе с терапией антителами против CXCL13 может быть необходимо для демонстрации ослабления симптомов заболевания на данной модели. На моделях, где отсутствует сверхэкспрессия TNFα, ингибирование CXCL13 может быть достаточным для ослабления заболевания. Кроме того, на модели, описанной выше, экспрессия TNF имеет место у трансгенных животных и она не зависит от прогрессирования заболевания. При реальной патологии, например, при лечении пациента, экспрессия TNF, возможно, будет снижаться по мере купирования воспаления, таким образом, если антагонист CXCL13 может привести к снижению миграции и накопления продуцирующих TNFα клеток, то он один может быть эффективным для лечения. Это проверяется in vivo на моделях заболеваний.
Пример 3
Совместная обработка моноклональными антителами, специфическими для TNFα и CXCL13, снижает инфильтрацию В-клеток в ткани легких
Совместное введение mAb, специфических для TNFα и CXCL13, снижает инфильтрацию В-клеток в ткани легких (фиг.3, 4, 5 и 6). Инфильтрацию В-клеток анализировали методом проточной цитометрии по детектированию В-клеточных маркеров соответственно CD22.2, CD19, CD45R/B220 и CD4. Инфильтрация В-клеток в тканях легких существенно снижалась у мышей SP-C/TNFα, которых совместно обрабатывали mAb, специфическими для TNFα и CXCL13, по сравнению с необработанными контрольными животными (фиг.3, 4, 5 и 6).
Подготовку животных и обработки проводили, как описано в примере 2, приведенном выше. Через 6 недель после начала инъекций мышей подвергали эвтаназии в соответствии с правилами по уходу и проведению работ с животными. Затем ткани легких измельчали и переваривали коллагеназой VII (1500 МЕ/мл) при 37°С в течение 45 мин для высвобождения отдельных клеток. Затем клеточные препараты инкубировали с коммерчески доступными, меченными CD22.2 (фиг.3), CD19 (фиг.4), CD45R/B220 (фиг.5) и CD4 (фиг.6) специфическими mAb с использованием стандартных методов. Полученные В-клетки в клеточной популяции несли один из маркеров CD22.2, CD19, CD45R/B220 или CD4 и становились мечеными после инкубации с коммерчески доступными CD22.2, CD19, CD45R/B220 или CD4 mAb. Данное мечение является основой для идентификации В-клеток методом проточной цитометрии. Анализ методом проточной цитометрии клеточных популяций проводили с использованием стандартных методов и затем определяли процент В-клеток, присутствующих в клеточной популяции, для каждого маркера с использованием стандартных методов. Приведенные данные (фиг.3, 4, 5 и 6) представляют средние значения ± стандартное отклонение по результатам анализа клеточных препаратов легких от 5 различных мышей из каждой опытной группы.
Последовательности CXCL13, CXCR5 и TNFα, описанные в данном документе, являются следующими:
Пример 4
Совместное введение моноклональных антител, специфических для TNFα и CXCL13, лечит патологии почек, связанные с системной красной волчанкой
Совместное введение mAb, специфических для CXCL13 и TNFα, лечит патологии почек, связанные с системной красной волчанкой (SLE), на мышиной модели SLE. Гломерулонефрит представляет собой воспаление внутренних структур почек, называемых клубочками, и является признаком SLE. Протеинурия представляет собой избыточное выделение сывороточных белков с мочой и является признаком заболевания почек, возникшего в результате ассоциированного с SLE гломерулонефрита. Гистологические изменения в ткани почек, такие как образование периартериальных очагов лимфоцитарных инфильтратов, являются еще одним признаком заболевания почек, возникшего в результате ассоциированного с SLE гломерулонефрита.
Совместное введение mAb, специфических для CXCL13 и TNFα, снижало уровень белка, ассоциированного с гломерулонефритом, в моче мышей NZB/W F1 с симптомами SLE до уровня ниже такового у необработанных контрольных мышей NZB/W F1, которые получали только один растворитель PBS (фиг.7).
Кроме того, совместное введение mAb, специфических для CXCL13 и TNFα снижало тяжесть (в баллах) ассоциированного с SLE заболевания почек у мышей NZB/W F1 с симптомами SLE до уровня ниже такового у необработанных контрольных мышей NZB/W F1, которые получали только один растворитель PBS (фиг.8). Тяжесть ассоциированного с SLE заболевания почек у мышей NZB/W F1 определяли с использованием оценочной шкалы. Баллы основывались на количестве периартериальных очагов лимфоцитарных инфильтратов, обнаруженных при гистологическом исследовании ткани почек, и значениях ассоциированного с гломерулонефритом содержания белка в тканях почек у мышей NZB/W F1 из каждой опытной группы (фиг.8).
В данном опыте 12-недельных мышей NZB/W F1 получали из питомника Jackson Labs (Bar Harbor, ME). По мере роста у мышей NZB/W F1 проявляются признаки, подобные таковым при системной красной волчанке (SLE), и они являются моделью SLE. На 0 сутки опытных животных произвольно разделяли на контрольные или опытные группы (n=15/группу). Животным внутрибрюшинной инъекцией вводили растворитель PBS (контрольные животные), mAb против CXCL13 (1 мг на мышь в PBS), mAb против TNFα (1 мг на мышь в PBS) или оба вида mAb против CXCL13 (1 мг на мышь в PBS) и mAb против TNFα (1 мг на мышь в PBS) раз в неделю, начиная с возраста 18 недель до возраста 38 недель. mAb против CXCL13 представляют собой нейтрализующие крысиные mAb против CXCL13 (R&D Systems, Inc., Minneapolis, MN). mAb против TNFα представляют собой химерные крысиные mAb против TNFα от Centocor, Inc., которые получали с использованием стандартных способов. Состояние животных оценивали раз в неделю. Периодически собирали мочу свободным отбором и хранили при -80°С. Животных в конце опыта подвергали эвтаназии и отобранные почки помещали в соответствующие буферы для хранения до анализа. Уход, обращение и убой животных проводили в соответствии с утвержденными правилами по уходу и проведению работ с животными.
Общий белок и креатинин в собранной моче определяли с использованием стандартных методов и на анализаторе Ace Analyzer (Alpha Wasserman, West Caldwell, NJ). Общий белок в моче определяли в неразбавленных пробах мочи объемом 100 мкл и креатинин определяли в разбавленных деионизированной Н2О 1:10 пробах мочи объемом 100 мкл. Затем эти значения использовали для расчета полученного соотношения общий белок/креатинин в моче. Соотношение общий белок/креатинин в моче выражали в виде среднего значения ± стандартное отклонение и статистически достоверные различия между пробами определяли с использованием двухстороннего вариационного анализа со стандартным t-критерием при р <0,05.
Проводили гистологическое исследование почек, отобранных у животных в возрасте 38 недель. Отобранные почки сразу же помещали в 0,7% буфер периодат натрия-лизин-параформальдегид (PLP), состоящий из 0,1 М фосфатного буфера, 0,7% параформальдегиа, 75 мМ L-лизина и 10 мМ NaIO4. Почки заливали в парафиновые блоки после фиксации в буфере PLP течение ночи. Для получения срезов толщиной 7 мкМ и окрашивания гематоксилин-эозином использовали стандартные методы. Затем препараты исследовали и оценивали в баллах на тяжесть проявления ассоциированного с SLE заболевания почек при «слепом» исследовании образцов. Оценка в баллах основывалась на количестве периартериальных очагов лимфоцитарных инфильтратов, обнаруженных при гистологическом исследовании препаратов ткани почек, и значений ассоциированного с гломерулонефритом содержания белка в тканях почек у мышей NZB/W F1 из каждой опытной группы. Для определения статистической достоверности различий между опытными группами проводили обработку данных с использованием вариационного анализа рангов Kruskal-Wallis с поправочным коэффициентом Дана для множественных сравнений и значении p <0,05.
Несмотря на то, что настоящее изобретение было раскрыто и описано выше со ссылкой на некоторые конкретные варианты осуществления, оно не предназначено ограничиваться приведенными деталями. В большей степени настоящее изобретение относится к антагонистам CXCL13 полипептидам, полинуклеотидам, антителам, устройствам и наборам, раскрытым в данном документе, и их областям применения и способам регуляции уровня CXCL13, необязательно наряду с уровнем TNFα, и могут быть разработаны различные модификации в деталях в объеме и в пределах эквивалентов формулы изобретения, не отступая от сущности изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| ЛИГАНД GITR И СВЯЗАННЫЕ С ЛИГАНДОМ GITR МОЛЕКУЛЫ И АНТИТЕЛА И ВАРИАНТЫ ИХ ПРИМЕНЕНИЯ | 2004 |
|
RU2369636C2 |
| АНТАГОНИСТЫ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2005 |
|
RU2401842C2 |
| ГЕТЕРОДИМЕРНЫЕ ПОЛИПЕПТИДЫ IL-17 A/F И ВОЗМОЖНОСТИ ИХ ЛЕЧЕБНОГО ПРИМЕНЕНИЯ | 2007 |
|
RU2440134C2 |
| НОВЫЕ ИЗОФОРМЫ ИНГИБИТОРА РОСТА ВАСКУЛЯРНЫХ ЭНДОТЕЛИАЛЬНЫХ КЛЕТОК | 2002 |
|
RU2316591C2 |
| ЛЕЧЕНИЕ АУТОИММУННЫХ ЗАБОЛЕВАНИЙ У ПАЦИЕНТА С НЕАДЕКВАТНЫМ ОТВЕТОМ НА ИНГИБИТОР TNF-АЛЬФА | 2004 |
|
RU2358762C9 |
| ПРИМЕНЕНИЕ АНТИТЕЛА ДЛЯ ЛЕЧЕНИЯ АУТОИММУННЫХ ЗАБОЛЕВАНИЙ У ПАЦИЕНТА С НЕАДЕКВАТНЫМ ОТВЕТОМ НА ИНГИБИТОР TNF-АЛЬФА | 2008 |
|
RU2489166C2 |
| ГУМАНИЗИРОВАННЫЕ АНТИТЕЛА К CXCR5, ИХ ПРОИЗВОДНЫЕ И ИХ ПРИМЕНЕНИЕ | 2008 |
|
RU2571515C2 |
| ВЫДЕЛЕННОЕ АНТИТЕЛО ЧЕЛОВЕКА ИЛИ ЕГО АНТИГЕН-СВЯЗЫВАЮЩИЙ ФРАГМЕНТ (ВАРИАНТЫ), СПОСОБ ЛЕЧЕНИЯ ЗАБОЛЕВАНИЙ, ОПОСРЕДОВАННЫХ hTNFα С ИХ ПОСРЕДСТВОМ (ВАРИАНТЫ), ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, РЕКОМБИНАНТНОЕ АНТИТЕЛО ЧЕЛОВЕКА ИЛИ ЕГО АНТИГЕН-СВЯЗЫВАЮЩИЙ ФРАГМЕНТ, ВЫДЕЛЕННАЯ НУКЛЕИНОВАЯ КИСЛОТА (ВАРИАНТЫ) | 2005 |
|
RU2458704C9 |
| ХИМЕРНЫЙ БЕЛОК, СОСТАВЛЕННЫЙ ИЗ ДОМЕНА АНТАГОНИСТА NGF И ДОМЕНА АНТАГОНИСТА TNFα | 2015 |
|
RU2678810C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ РЕГУЛЯЦИИ РАЗВИТИЯ СОСУДОВ | 2005 |
|
RU2365382C2 |
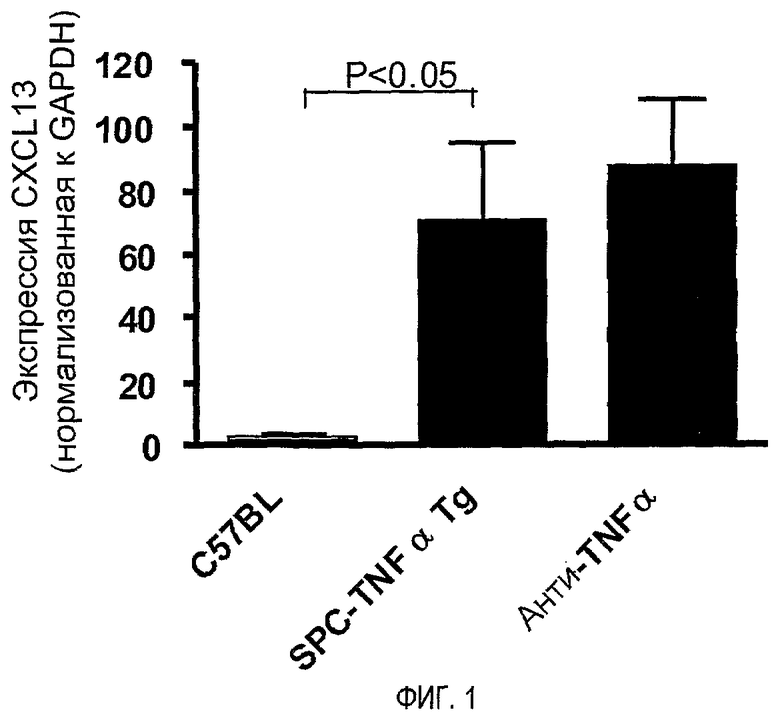
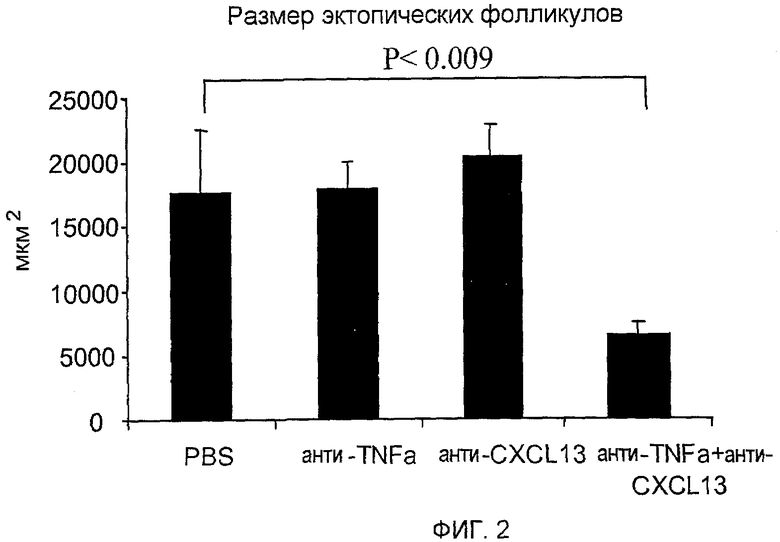
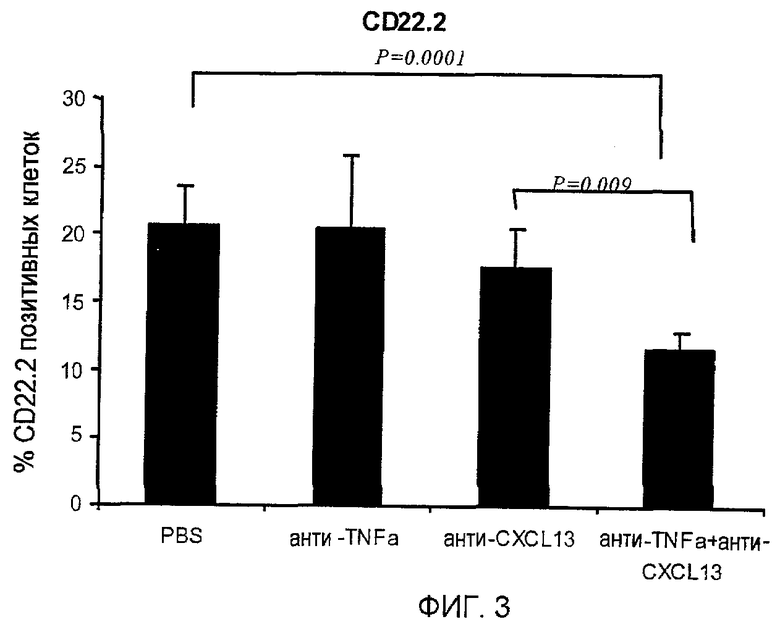
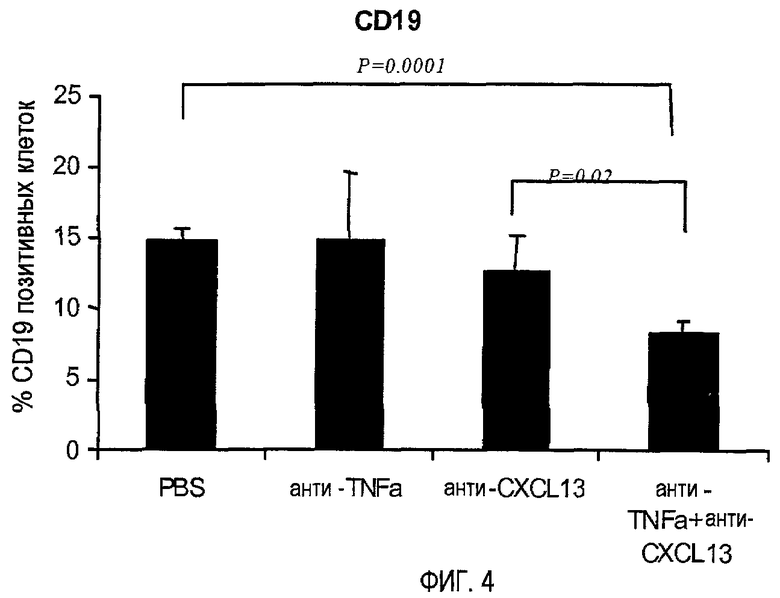
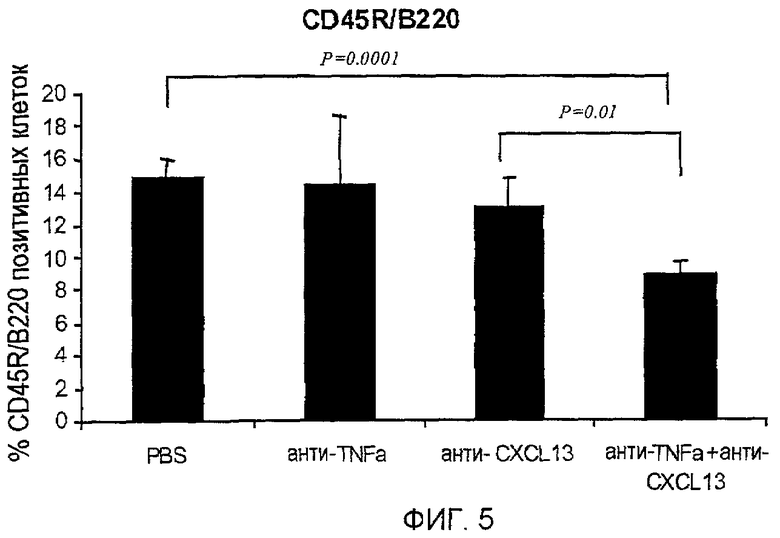
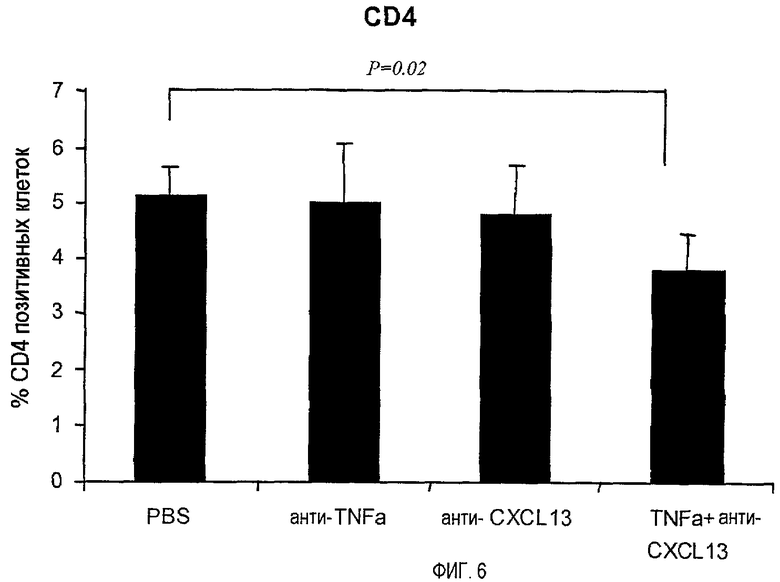
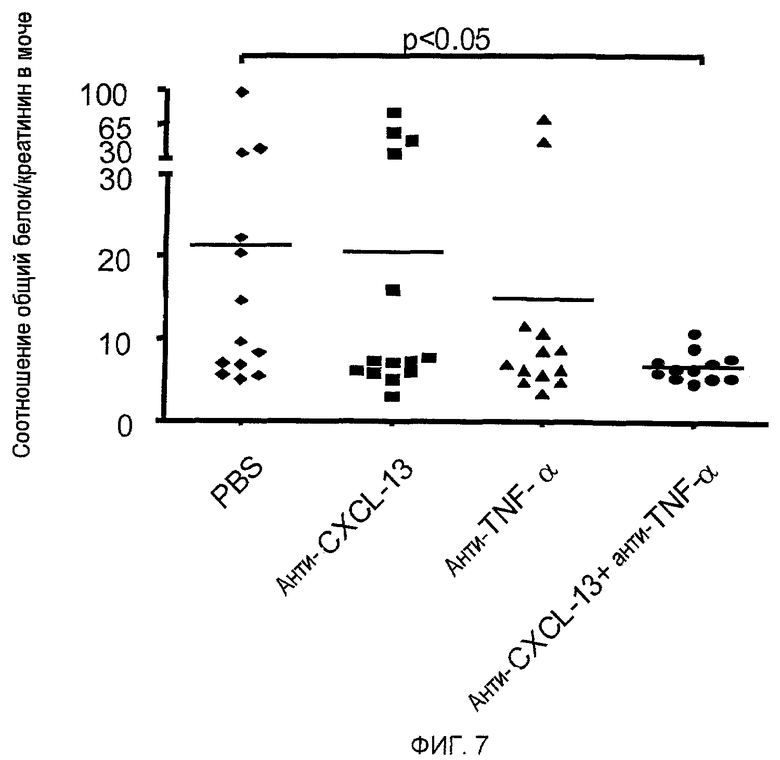
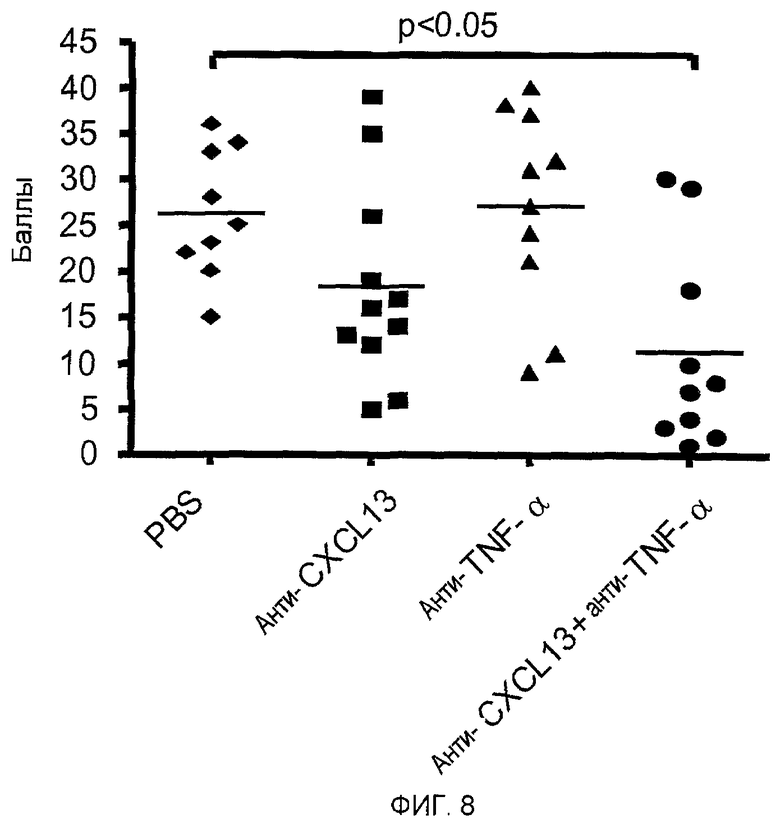
Группа изобретений относится к медицине, а именно к терапии, и может быть использовано для лечения нарушений, связанных с активностью CXCL13 в клетке, ткани, органе или организме животного, где нарушения представляют собой хроническое обструктивное заболевание легких (COPD) или системную красную волчанку. Для этого в клетку, ткань, орган или организм животного вводят антагонист CXCL13 в количестве, эффективном для ингибирования активности CXCL13 совместно с антагонистом TNFα в количестве, эффективном для ингибирования активности TNFα в указанной клетке, ткани, органе или организме животного. Группа изобретений позволяет лечить хроническое обструктивное заболевание легких и системную красную волчанку за счет совместного использования моноклональных антител, специфических для CXCL13 и TNFα, что ведет к уменьшению инфильтрации В-клеток, экспрессирующих CD4, а также снижает связанное с гломерулонефритом содержание белка в моче у животного с симптомами системной красной волчанки. 2 н. и 17 з.п.ф-лы, 8 ил., 3 табл., 4 пр.
1. Способ лечения связанного с активностью CXCL13 нарушения в клетке, ткани, органе или организме животного, включающий:
введение в клетку, ткань, орган или организм животного антагониста CXCL13 в количестве, эффективном для ингибирования активности CXCL13 в указанной клетке, ткани, органе или организме животного; и введение в клетку, ткань, орган или организм животного антагониста TNFα в количестве, эффективном для ингибирования активности TNFα в указанной клетке, ткани, органе или организме животного, где связанное с активностью CXCL13 нарушение представляет собой хроническое обструктивное заболевание легких (COPD) или системную красную волчанку.
2. Способ по п.1, в котором антагонист CXCL13 представляет собой связывающее CXCL13 моноклональное антитело или его фрагмент, и антагонист TNFα представляет собой связывающее TNFα моноклональное антитело или его фрагмент.
3. Способ по п.2, в котором фрагмент антитела представляет собой фрагмент Fab, Fab' или P(ab')2 или его производное.
4. Способ по п.2, в котором животное является млекопитающим.
5. Способ по п.4, в котором, по меньшей мере, одно моноклональное антитело или фрагмент вводят, по меньшей мере, одним путем, выбранным из парентерального, подкожного, внутримышечного, внутривенного, внутриартикулярного, внутрибронхиального, внутриабдоминального, внутрикапсульного, внутрихрящевого, внутриполостного введения, внутриклеточного, внутримозжечкового, интрацеребровентрикулярного пути введения, введения в толстый кишечник, интрацервикального, внутрижелудочного пути введения, введения в печень, внутримиокардиального, внутрикостного, внутритазового, внутриперикардиального, внутрибрюшинного, внутриплеврального, интрапростатического, внутрилегочного, интраректального пути введения, введения в почку, интраретинального, интраспинального, внутрисуставного, интраторакального пути введения, введения в матку, внутрипузырного пути введения, введения в область поражения, болюсного введения, интравагинального, ректального, буккального, сублингвального, интраназального и трансдермального пути введения.
6. Способ по п.5, в котором, по меньшей мере, одно моноклональное антитело или фрагмент вводят в количестве примерно от 0,05 мг/кг до примерно 30,0 мг/кг массы тела указанного млекопитающего.
7. Способ по п.5, в котором млекопитающее представляет собой пациента-человека.
8. Способ по п.5, в котором, по меньшей мере, одно моноклональное антитело или фрагмент вводят внутрибрюшинно.
9. Способ по п.5, в котором, по меньшей мере, одно моноклональное антитело или фрагмент вводят в виде дозы-болюса с последующей инфузией указанного антитела.
10. Способ лечения животного с системной красной волчанкой, включающий:
a) введение антагониста CXCL13 животному, и
b) введение антагониста TNFα животному;
в котором каждый антагонист находится в количестве, эффективном для того, чтобы вызвать ослабление симптома системной красной волчанки у животного.
11. Способ по п.10, в котором антагонист CXCL13 представляет собой связывающее CXCL13 антитело или связывающий CXCL13 фрагмент антитела, и антагонист TNFα представляет собой связывающее TNFα антитело или связывающий TNFα фрагмент антитела.
12. Способ по п.11, в котором животное является млекопитающим.
13. Способ по п.12, в котором млекопитающее является человеком.
14. Способ по п.11, в котором количество каждого антитела или связывающего фрагмента антитела вводят в количестве примерно от 0,05 мг на кг до примерно 50,0 мг на кг массы тела животного.
15. Способ по п.14, в котором количество каждого антитела или связывающего фрагмента антитела вводят в количестве примерно от 25 мг на кг массы тела животного до примерно 40 мг на кг массы тела животного.
16. Способ по п.10, в котором симптомом системной красной волчанки является число периартериальных очагов лимфоцитарных инфильтратов, обнаруживаемых при исследовании ткани почек.
17. Способ по п.10, в котором симптомом системной красной волчанки является соотношение общий белок мочи/общий креатинин мочи.
18. Способ по п.16 или 17, в котором антагонист CXCL13 представляет собой связывающее CXCL13 антитело или связывающий CXCL13 фрагмент антитела, и антагонист TNFα представляет собой связывающее TNFα антитело или связывающий TNFα фрагмент антитела.
19. Способ по п.16 или 17, в котором антагонист CXCL13 представляет собой связывающее CXCL13 антитело или связывающий CXCL13 фрагмент антитела, и антагонист TNFα представляет собой связывающее TNFα антитело ифликсимаб или его фрагмент.
| СПОСОБ ВЫРАБОТКИ КОНСЕРВИРОВАННОГО ПРОДУКТА "СИЧЕНИКИ РЫБНЫЕ УКРАИНСКИЕ" | 2013 |
|
RU2501422C1 |
| НОВЫЕ ИММУНОТЕРАПЕВТИЧЕСКИЕ СОЕДИНЕНИЯ, СОДЕРЖАЩАЯ ИХ ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ И СПОСОБ СНИЖЕНИЯ УРОВНЕЙ ФДЭ, TNFα И NFκB | 1997 |
|
RU2188819C2 |
| Промежуточное звено для соединения нескольких трубчатых ветвей, в частности перегревательных трубок, в одну общую трубку | 1925 |
|
SU2272A1 |
| Способ обработки целлюлозных материалов, с целью тонкого измельчения или переведения в коллоидальный раствор | 1923 |
|
SU2005A1 |
| Прибор, замыкающий сигнальную цепь при повышении температуры | 1918 |
|
SU99A1 |
| БЕЛОУСОВА Е.А | |||
| и др | |||
| Инфликсимаб (Ремикейд) в лечении рефрактерных форм болезни Крона | |||
| - Русский медицинский журнал, 2005, №1 [найдено 05.05.2011 на сайте http://www.rmj.ru/articles_5417.htm] Yang N | |||
| et | |||

