Изобретение относится к медицине (более точно - травматологии, космической медицине, а также к физиологии) и может найти применение при устранении негативного последствия гипокинезии или гравитационной разгрузки.
При длительной гипокинезии, гравитационной разгрузке, иммобилизации конечности, а также при длительном лишении человека обычной двигательной активности скелетные мышцы подвергаются атрофии, происходит снижение максимальной силы и скорости мышечного сокращения, а также работоспособности. Параллельно с развитием атрофических процессов при функциональной разгрузке в мышцах снижается содержание митохондриальных белков и ДНК (Cannavino, Brocca et al. 2014, Leermakers, Kneppers et al. 2019, Fernandez-Gonzalo, Tesch et al. 2020), а также происходит изменение паттерна экспрессии миозиновых генов, что ведет к повышению доли «быстрых» гликолитических волокон, обладающих слабой устойчивостью к утомлению (Stevens, Sultan et al. 1999, Pette and Staron 2000, Trappe, Costill et al. 2009, Desaphy, Pierno et al. 2010). В результате происходит снижение работоспособности мышц.
При ряде моделей функциональной разгрузки скелетных мышц было обнаружено повышение уровня кальция в миоплазме (Booth and Giannetta 1973, Ingalls, Warren et al. 1999, Tomiya, Tamura et al. 2019). Этот эффект связывают с нарушением функций Na, K-ATФазы и деполяризацией сарколеммы на ранних этапах разгрузки, которое приводит к активации дигидропиридиновых каналов и входу кальция в миоплазму через рианодиновые рецепторы (Krivoi, Kravtsova et al. 2008). Высокий уровень кальция в миоплазме может способствовать активации кальций-зависимых протеаз кальпаинов, что ведет к усилению протеолиза, (Turner, Westwood et al. 1988, Shenkman, Belova et al. 2015) а также провоцировать нарушение функций митохондрий (Matuz-Mares, Gonzalez-Andrade et al. 2022). Нарушение кальциевого гомеостаза в скелетной мышце также приводит к изменениям в экспрессии генов, включая маркеры протеолиза и гены, определяющие тип мышечного волокна («быстрый или «медленный») (Berchtold, Brinkmeier et al. 2000, Shimizu, Langenbacher et al. 2017).
Известно, что ключевую роль в удалении кальция из миоплазмы играет SERCA (Са2+-АТФаза сарко/эндоплазматического ретикулума), которая откачивает ионы кальция из миоплазмы в саркоплазматический ретикулюм (СПР). В условиях функциональной разгрузки скелетных мышц функция SERCA нарушается. Обнаружено снижение интенсивности закачивания ионов кальция в СПР на фоне иммобилизации мышц и денервации (Midrio, Danieli-Betto et al. 1997, Thom, Thompson et al. 2001). Ранее также показано, что в условиях функциональной разгрузки мышц наблюдается изменение паттерна экспрессии изоформ SERCA, что может приводить к изменениям в перераспределении ионов кальция между СПР и миоплазмой (Schulte, Peters et al. 1994, Mukhina, Altaeva et al. 2006, Altaeva, Ogneva et al. 2010).
Избыточное накопление кальция в миоплазме способно также приводить к накоплению активных форм кислорода (АФК), которые, в свою очередь, нарушают функционирование рианодиновых каналов, что ведет к истощению кальциевого депо СПР и повышению утомляемости мышц: данный механизм был описан для условий старения, а также для трансгенных животных (Andersson, Betzenhauser et al. 2011, Andersson, Meli et al. 2012, Umanskaya, Santulli et al. 2014, Qaisar, Bhaskaran et al. 2019, Qaisar, Pharaoh et al. 2020). Поскольку в условиях функциональной разгрузки скелетных мышц также наблюдается избыточное накопление кальция и АФК в миоплазме (Ingalls, Warren et al. 1999, Hord, Garcia et al. 2021), нельзя исключать, что кальций-зависимые процессы вносят вклад в повышение утомляемости и в этих условиях. Это подтверждается и недавней работой, где обнаружено функциональное взаимодействие между SERCA и митохондриальным окислительным фосфорилированием (Petrick, Brownell et al. 2022). В ней показано предотвращение развития утомления m.soleus при введении мышам пищевого нитрата в качестве активатора SERCA.
Мы предположили, что при функциональной разгрузке в камбаловидной мышце происходит снижение функций SERCA, что ведет к избыточному накоплению ионов кальция в миоплазме. В свою очередь, кальций активирует сигнальные пути, запускающие процессы протеолиза и трансформации миозинового фенотипа в «быструю» сторону, и способствует снижению устойчивости мышцы к утомлению. Камбаловидная мышца принимает участие в ходьбе и беге, у человека отвечает за поддержание вертикального положения тела в пространстве, и является особенно подверженной негативным изменениям, происходящим при функциональной разгрузке, поэтому она была выбрана для исследования в данной работе (Shenkman В.S. 2004)
Для проверки гипотезы о роли снижения активности SERCA при ограничении функциональной активности мышц в регуляции ее сократительных характеристик был применен CDN1163 (специфический аллостерический активатор насоса SERCA) в модели вывешивания крыс. Такой подход раньше не применялся. Специфический активатор SERCA появился недавно и используется впервые при моделировании функциональной разгрузки крыс. Мы впервые показали, что при применении активатора SERCA при недельной функциональной разгрузке soleus полностью предотвращается увеличение ее утомления. Происходит это за счет того, что введение активатора SERCA CDN1163 на фоне 7-суточной функциональной разгрузки приводит к снижению уровня окисления тропомиозина (показателя окислительного стресса), способствует сохранению доли «медленных» устойчивых к утомлению мышечных волокон и предотвращению увеличения «быстрых» мышечных волокон, а также снижения митохондриальной ДНК и белка. Все это предотвращает повышение утомляемости камбаловидной мышцы на фоне вывешивания без эффекта предотвращения атрофии.
В результате проведенных исследований было обнаружено, что данный способ предотвращения атрофии основан на том, что при разгрузке активируются рецепторы SERCA.
Таким образом, заявляемое изобретение основано на применении селективного активатора рецепторов SERCA CDN1163 на фоне 7-суточной функциональной разгрузки химическое наименование: 4-(1-Methylethoxy)-N-(2-methyl-8-quinolinyl)benzamide;
O=C(NC1=C2N=C(C)C=CC2=CC=C1)C3=CC=C(OC(C)C)C=C3
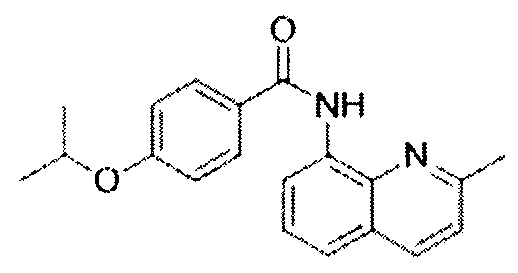
для предотвращения повышения утомления скелетных мышц при их функциональной разгрузке.
Наиболее близким способом (прототипом), где показано предотвращение развития утомления m.soleus при введении мышам пищевого нитрата, предполагающегося авторами в качестве активатора SERCA (Petrick, Brownell et al. 2022) (Petrick H.L., Brownell S., Vachon В., Brunetta H.S., Handy R.M., van Loon L.J.C, Murrant C.L., Holloway G.P. Dietary nitrate increases submaximal SERCA activity and ADP transfer to mitochondria in slow-twitch muscle of female mice. Am J Physiol Endocrinol Metab. 323(2): E171-E184. 2022). Ранее не показано, что 1 mM sodium nitrate (NaNO3) является непосредственным прямым активатором SERCA, но по ряду косвенных признаков авторы сделали такое предположение. Авторы отметили известное свойство диетических нитратов снижать затраты АТФ на сокращение, что увеличивает выработку силы и увеличивает цитозольный Са2+, что, по-видимому, требует большей потребности в Са2+ -АТФазе саркоплазматического ретикулума (SERCA) для снижения Са2+ в саркоплазматическом ретикулуме (CP) во время расслабления. В случае камбаловидной мышцы авторы обнаружили, что пищевой нитрат увеличивал выработку силы при всех тестируемых частотах стимуляции и в течение 25-минутного теста на утомление.
Техническим результатом заявленного изобретения является предотвращение повышения утомляемости мышцы путем активирования SERCA.
Этот технический результат достигается тем, что в известном способе предотвращения развития утомления m.soleus мы вводили активатор рецепторов SERCA CDN1163 на фоне 7-суточной функциональной разгрузки (химическое наименование: 4-(1-Methylethoxy)-N-(2-methyl-8-quinolinyl)benzamide;
O=C(NC1=C2N=C(C)C=CC2=CC=C1)C3=CC=C(OC(C)C)C=C3),
который приводит к предотвращению развития утомления мышцы m.soleus, сохранению доли «медленных» устойчивых к утомлению мышечных волокон и предотвращению увеличения «быстрых» мышечных волокон, а также снижению митохондриальной ДНК и белка.
В отличие от предыдущей работы мы использовали селективный активатор фермента SERCA, имеющий известное прямое действие на фермент. Препарат вводили ежедневно на протяжении 7 суток эксперимента, внутрибрюшинно, в дозировке 50 мг/кг в 10% DMSO, 10% Tween 80 на 0.9% растворе NaCl. Животным из контрольной группы и группы «чистой» 7-суточной функциональной разгрузки вводили плацебо (10% DMSO, 10% Tween 80 на 0.9% растворе NaCl).
Гипотеза была проверена с помощью активирования фермента SERCA при вывешивании крыс.
Возможные механизмы действия метода.
SERCA при функциональной разгрузке мышцы становится неактивной, в результате чего перестает выполнять функцию обратного закачивания ионов Са в саркоплазматический ретикулум из миоплазмы. Активатор SERCA CDN1163 предотвращает падение активности фермента и ионы кальция из миоплазмы активно поступают обратно в саркоплазматический ретикулум.
Это приводит к предотвращению накопления кальция в миоядрах, что способствует (возможно, с другими сигнальными каскадами) к предотвращению активации факторов транскрипции (ФТ), приводящих к экспрессии или репрессии генов, вовлеченных в фенотип мышечных клеток, ухудшающих устойчивость к утомлению при функциональной разгрузке. Материалы, подтверждающие воспроизводимость способа Функциональная разгрузка моделировалась вывешиванием задних конечностей по стандартной методике Ильина-Новикова в модификации Morey-Holton. Вывешивание проводилось так, что задние конечности крыс не касались пола, а передние свободно опирались на пол и животные благодаря уникальной конструкции экспериментального стенда свободно передвигались. Крысы были подвешены на специальных мягких шинках. Пищу и воду животные получали ad libitum. Для проведения запланированного эксперимента потребовалось 24 самцов крыс линии Wistar массой 180-200 грамм в возрасте 2,5 месяца. Препарат вводился в дозе 50 мг/кг в 10% DMSO, 10% Tween 80 in 0.9% NaCl) согласно данным литературы (Kang, Dahl et al. 2016).
Животные были распределены на 3 группы - виварный контроль с введением плацебо (С, n=8), группа вывешивания с введением плацебо (7HS, n=8) и группа вывешивания с введением CDN1163 (7HS+CDN, n=8). Через 7 дней эксперимента крыс наркотизировали авертином (5 мл/кг веса 10% раствора), выделяли m.soleus, одну из них немедленно замораживали в жидком азоте и хранили в кельвинаторе при -85°С до проведения анализов, вторую использовали для определения утомляемости в ex vivo тесте. Эвтаназия крыс осуществлялась введением сверхдозы авертина.
Эксперимент был одобрен комиссией по биомедицинской этике ГНЦ РФ ИМБП РАН (протокол №584).
Определение уровня утомляемости камбаловидной мышцы
Камбаловидные мышцы экспериментальных животных были помещены в охлажденный раствор Рингера-Кребса с постоянным продуванием 95% карбогеном (O2+CO2) и инкубированы в течении 15 минут. В это время на сухожилиях были завязаны узлы, необходимые для крепления мышцы. После 15 минут мышца была помещена в ванночку для теста, с фиксированной температурой 37°С, прикреплена за сухожилия к датчику силы с одного конца и к фиксированному крючку с другого, после чего была определена оптимальная длина мышцы с помощью подачи короткого электрического импульса (супра-максимальные квадратные импульсы 0.5 мс, 20 В) и замера силы сокращения мышцы (наибольшая сила одиночного сокращения соответствует оптимальной длине мышцы (L0) (Gordon, Huxley et al. 1966). L0 была измерена с помощью цифрового штангенциркуля. После одиночных сокращений был проведен тест на тетаническое изометрическое сокращение. Мышца была установлена на длину L0, после чего была начата стимуляция (1 мс квадратные импульсы, 40 Гц, продолжительность стимуляции 2 секунды), в процессе чего фиксировалась максимальная сила тетанического сокращения. Для тестирования индекса утомления камбаловидной мышцы проводилась серия из 20 тетанических сокращений за одну минуту, с перерывом в 1 секунду между сокращениями. За индекс утомления брали силу сокращения после 20 повторений, деленную на максимальную силу сокращения, измеренную в течении всего теста (Burke, Levine et al. 1974, Roy, Zhong et al. 2002). Показатели нормировали на физиологическое поперечное сечение мышцы (pCSA). Измерение силовых показателей проводили с помощью Aurora Scientific Dual Mode Lever System 305C-LR (Aurora Scientific, США) с частотой сбора данных 10 кГц. Обработку данных проводили с помощью программного обеспечения Aurora Scientific 615А Analysis Software Suite.
Анализ содержания белков-маркеров в m.soleus проводился с помощью электрофореза с последующим Вестерн-блоттингом.
Для этого с каждого из образцов m.soleus были сделаны срезы толщиной 20 мкм (40-60 мг) на микротоме-криостате фирмы Leica. Для выделения тотальной фракции белка с образца m.soleus были сделаны срезы толщиной 20 мкм (10-15 мг) и немедленно прогомогенизированы в шариковом гомогенизаторе TissueLyser LT (QIAGEN, Германия) в течение 25 минут в 100 мкл лизирующего буфера RIPA (Santa-Cruz, USA), содержащего 50 mM Tris (рН 7.4), 150 mM NaCl, 0.1% Triton Х-100, 0.1% SDS, 5 mM EDTA (рН 8.0) 1 mM DTT, 1 mM PMSF, 1 мМ Na3VO4, 1 mM PMSF, апротинин (10 μg/ml), леупептин (10 μg/ml), пепстатин А (10 μg/ml), протеазный ингибиторный коктейль (Santa-Cruz, USA) и фосфотазный ингибиторный коктейль (Santa-Cruz, USA). Затем образцы центрифугировали при 20000xg в течение 15 минут. Супернатант отбирали, разаликвочивали и убирали на хранение при -85°С. Для определения уровня окисленного и тотального тропомиозина проводили выделение белка с помощью реагента RIPA фирмы Cell signaling Technology с добавлением 1 mM PMSF, протеазного ингибиторного коктейля (Santa-Cruz, USA) и фосфотазного ингибиторного коктейля (Santa-Cruz, USA).
Часть мышечных лизатов отбирали для определения концентрации общего белка с помощью реактива Бредфорда (Bio-Rad Laboratories, США). Определения проводились на планшетном фотометре Epoch при длине волны 595 нм. Пробы для нанесения разводились в 2х-кратном Laemly-буфере для образцов (5,4 мМ Tris-HCl (рН 6,8), 4%-ный Ds-Na, 20%-ный глицерин, 10%-ный β-меркаптоэтанол, 0,02%-ный бромфеноловый синий), за исключением проб для анализа уровня окисленного тропомиозина, которые разводили в 2х-кратном Laemly-буфере без добавления -меркаптоэтанола. Заливку и подготовку ПААГ гелей проводили с помощью заливочных столов фирмы и стекол «Bio-Rad Laboratories)). Гели устанавливали в камеры mini-Protean 3 Cell «Bio-Rad Laboratories)). Электрофорез проводили в 10%-ном разделяющем ПААГ (0,2%-ный метилбисакриламид, 0,1%-ный Ds-Na, 375 мМ Tris-HCl (рН 8,8), 0,05%-ный персульфат аммония, 0,1%-ный ТЕМЕД) и в 5%-ном концентрирующем ПААГ (0,2%-ный метилбисакриламид, 0,1%-ный Ds-Na, 125 мМ Tris-HCl (рН 6,8), 0,05% аммоний персульфат, 0,1%-ный ТЕМЕД). Для проведения электрофореза был использован трис-глициновый буфер (192 мМ Tris-глицин (рН 8,6), 0,1%-ный Ds-Na). Образцы каждой группы загружались на один гель с контрольными образцами и маркерами молекулярных весов. Образцы загружались из расчета 20 мкг общего белка на дорожку и нормировались относительно уровня GAPDH, содержащегося в той же пробе. Электрофорез проводился при 15 мА на гель в мини-системе («Bio-Rad Laboratories))) при комнатной температуре. После электрофореза гели переносились в установку для электропереноса белков на мембрану. Электроперенос проводился в буфере (25 мМ Tris (рН 8,3), 192 мМ глицин, 20%-ный этанол, 0,04%-ный Ds-Na) на нитроцеллюлозную мембрану при 100 V при температуре 4°С в системе mini Trans-Blot («Bio-Rad Laboratories))) в течение 2 часов. После электропереноса НЦ-мембраны инкубировались в течение 5 минут в 0,3% растворе Ponceau Red в 5%-ой уксусной кислоте, затем отмывались в PBS (Биолот) с 0,1% Tween 20 (PBST) до появления четких белковых полос на мембране. Этот этап проводился для контроля эффективности переноса; а также для того, чтобы убедиться, что количества общего белка, внесенного в каждую дорожку, было одинаковым. Мембраны блокировались в растворе 5% сухого молока («Bio-Rad Laboratories))) в PBST 1 час при комнатной температуре, затем помещались в раствор первичных антител на ночь на +4°С. Условия работы антител проверялись и подбирались экспериментально.
Для выявления белковых полос были использованы первичные антитела против тропомиозина (sc-58868) фирмы Santa Cruz ВТ, и против ТОМ20 (4F3, BioRad, Германия, 1:500) На следующий день мембрана отмывалась от первичных антител в PBST 3 раза по 5 минут на шейкере и инкубировалась 1 час со вторичными антителами goat-anti-rabbit (1: 30000, «Jackson Immuno Research)), США) или goat-anti-mouse (1:20000 «Bio-Rad Laboratories)), США). Потом мембрана отмывалась от вторичных антител в PBST 3 раза по 5 минут на шейкере. Выявление проводилось с помощью Clarity Western ECL Substrate (Bio-Rad Laboratories, США). Хемилюминисцентный сигнал детектировался с помощью сканера C-DiGit Blot Scanner (LI-COR, США). Белковые полосы были анализированы с использованием Image Studio Software (LI-COR).
Данный прибор и его аналоги вместе с программным обеспечением широко используются во всем мире для детекции сигнала в иммуноблотинге, делая эту методику гораздо производительнее и экономичнее. Для каждого параметра электрофорез с последующим иммуноблоттингом был повторен не менее Зх раз. Затем полученные на сканере изображения обрабатывались с помощью прилагаемого программного обеспечения Image Studio Software (LI-COR) для получения количественных данных для анализа. Хемилюминесцентный сигнал полосы контрольной группы на анализируемой мембране принимали за 100%, а сигнал полос других групп сравнивали с сигналом полос контрольной группы, расположенных на одной и той же мембране.
ПЦР в реальном времени
Для исследования экспрессии генов и содержания митохондриальной ДНК методом ПЦР в реальном времени проводили выделение РНК и ДНК из образцов мышечной ткани. Для выделения РНК из скелетных мышц был использован набор HiPure Fibrous DNA/RNA Kit (Magen, Китай) согласно рекомендациям производителя.
Определение концентрации мРНК и ДНК определялось по поглощению раствора мРНК выполнялось при помощи спектрофотометра NanoPhotometer IMPLEN. Снимались показания в диапазоне от 200 до 320 нм. Измерение каждой пробы проводилось не менее 3 раз. Чистота образцов была оценена исходя из соотношений показателей поглощения при различных длинах волн. Соотношение А260/А230 экспериментальных образцов было>2,0, что указывает на то, что они являлись достаточно чистыми от углеводов, пептидов, фенолов или ароматических соединений (Watson M.L., 2018). Для проведения обратной транскрипции были использованы реагенты фирмы «Синтол» (Россия). Для подготовки кДНК водный раствор, содержащий 1 мкг тотальной РНК, 30 мкМ случайных гексануклеотидов и 17,4 мкМ олиго-d(Т)15 инкубировался 3 минуты при 70°С, и немедленно переносился на лед. Далее к смеси было добавлено 11,5 мкл мастер-микса (1,3 мМ дНТФ, 0,02 ед./мкл ингибитора РНКазы, 6 ед./мкл M-MLV-ревертазы, 4 мкл 5х-буфера для М-MLV-ревертазы, «Синтол»). После этого пробы были помещены в амплификатор (iQ5 Multicolor Real-Time PCR Detection System, «Bio-Rad Laboratories))) для проведения обратной транскрипции: 10 мин при 25°С, 60 мин при 37°С, 5 мин при 95°С, 30 мин при 4°С. После проведения реакции образцы, содержащие кДНК, хранились при -25°С. Праймеры сконструированы с помощью программы Primer3 v.0.4.0, находящейся в свободном доступе (http://frodo.wi.mit.edu/primer3/). Для проведения ПЦР в реальном времени смешивалось 2 мкл кДНК или ДНК, 2 мкл праймеров с концентрацией 10 мкМ и 21 мкл мастер-микса (0,3 мМ дНТФ, 3 мМ MgCl2, 2,5 мкл 10х ПЦР-буфера Б (рН 8,8), 0,06 ед./мкл Taq ДНК-полимеразы, «Синтол»). Затем пробы были помещены в амплификатор для проведения реакции: 1 цикл: 5 мин при 95°С; 2 цикл по 44 повтора: 15 сек при 93°С, 15 сек при Tm (табл. 2), 15 сек при 72°С; 3 цикл: 6 мин 45 сек при 72°С, 30 сек при 65°С; 4 цикл по 61 повтору: 15 сек при 65°С; 5 цикл: 10 мин 15°С.
Для гена мишени ПЦР-реакция проводилась не менее 3-х раз.
Используемые в работе праймеры:
Ген mtDNA. Прямой сиквенс 5'-ATTGGAGGCTTCGGGAACTG-3'- прямой
Обратный сиквенс 5'-AGATAGAAGACACCCCGGCT-3'
Все праймеры были синтезированы фирмой «Синтол)) (Россия). Для анализа полученных с помощью ПЦР в реальном времени данных применялось относительное количественное определение исследуемого гена, нормализованное к референсному, метод 2-ΔΔCt (метод Ливака). В качестве референсного гена был использован RPL19, экспрессия которого постоянна в т.soleus в условиях эксперимента.
С помощью криомикротома изготавливали поперечные срезы замороженной мышцы толщиной 9 мкм. Срезы высушивали на воздухе и хранили на -20°. Перед окрашиванием срезы оттаивали и регидратировали при комнатной температуре в фосфатно-буферном растворе (PBS) в течение 20 минут, а затем инкубировали с антителами против тяжелых цепей миозина быстрого или медленного типов (MHCI и MHCII, Sigma, США) 1:400 в PBS во влажной камере при 37° в течение часа (или при+4°на ночь). Затем антитела отмывали в PBS 3x5 минут. Инкубацию со вторичными антителами, конъюгированными с AlexaFluor, 1:500 в PBS проводили в течение 40 мин при комнатной температуре. После отмывки вторичных антител срезы заключали в среду, стабилизирующую флуоресцентную метку. Срезы анализировали с использованием флуоресцентного микроскопа LeicaQ500MC с встроенной цифровой фотокамерой (ТСМ 300F, Leica, Германия), с увеличением х200. Анализ изображений проводился с помощью программы ImageJ. Измеряли площадь поперечного сечения, по крайней мере, 100 волокон, подсчитывали число волокон медленного и быстрого типов и их количественное соотношение.
Данные проверяли на нормальность распределения по критерию Колмогорова-Смирнова, после чего проводили анализ ANOVA с post hoc анализом Тьюки на множественные сравнения. Достоверными считали отличия при р<0.05. Данные представлены в виде средних значений±стандартная ошибка среднего.
Вес m.soleus обеих вывешенных групп (7HS и 7HS+CDN) не различался между собой и был достоверно ниже, чем в группе контроля (67,5±2.9 мг и 65,3±4.2 соответственно мг против 109,4±4.26 мг в группе контроля, р<0,05).
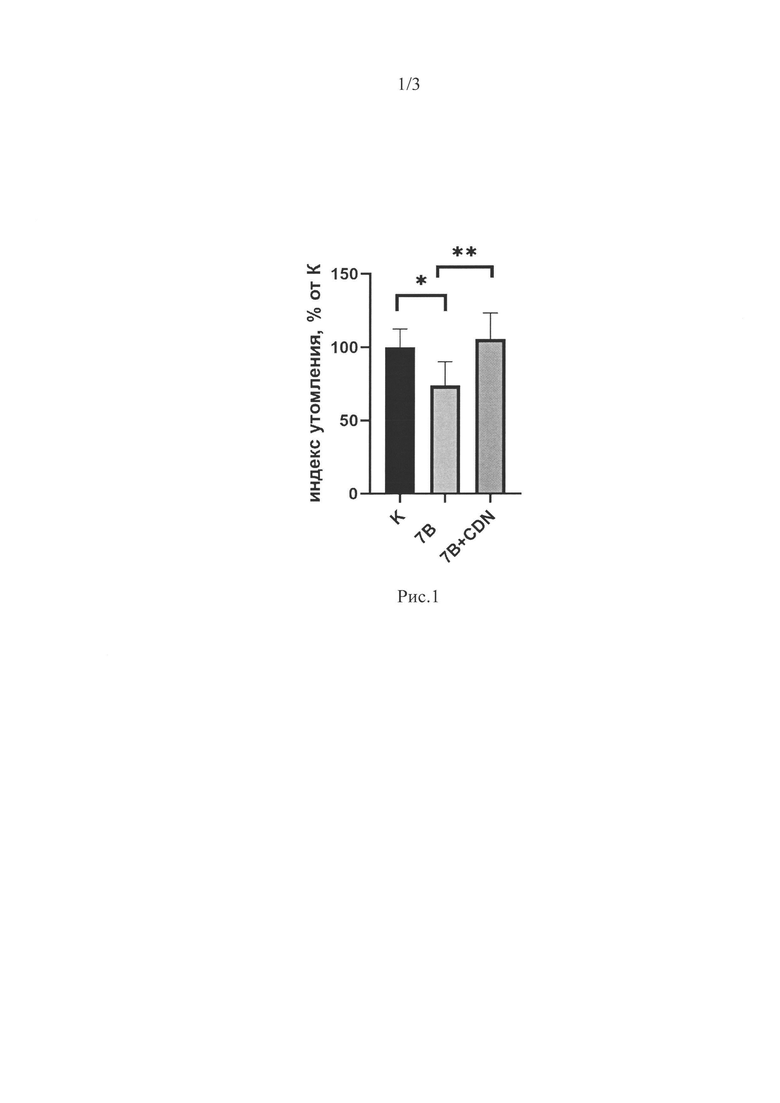
Мы наблюдали достоверное снижение индекса утомления в группе 7HS на 17% относительно группы контроля. Это снижение отсутствовало в группе 7HS+CDN(РИС 1).
Влияние активатора SERCA на миозиновый фенотип
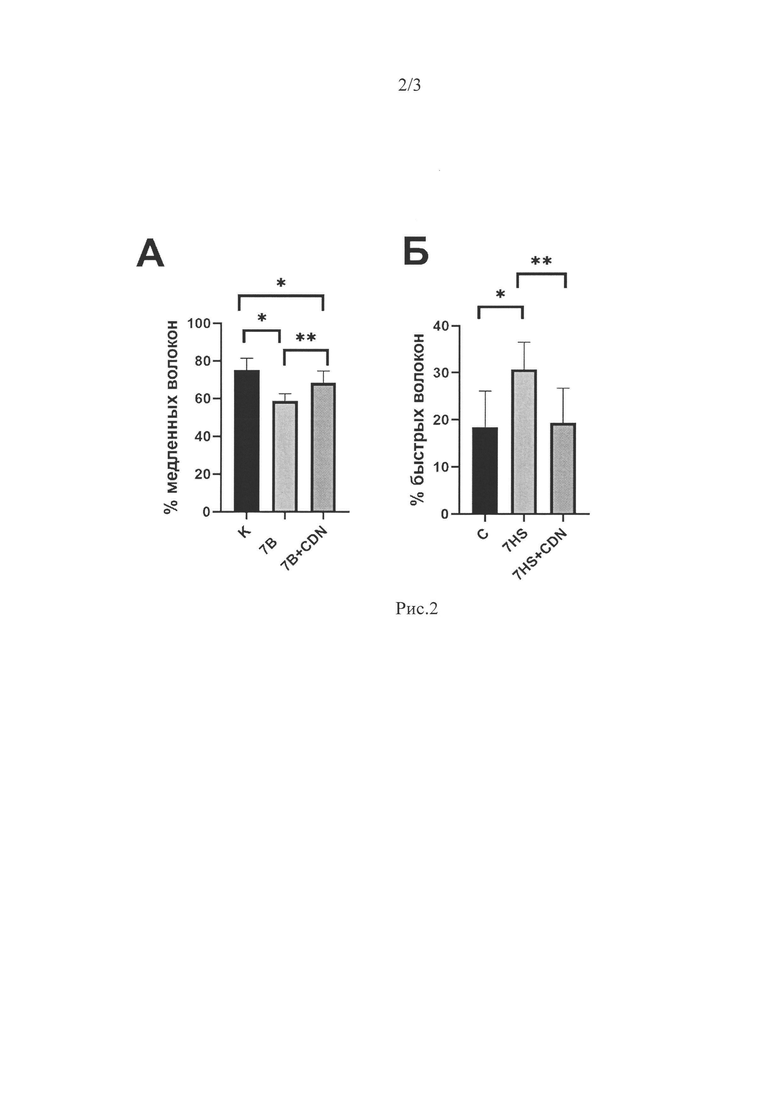
После 7 суток вывешивания процент мышечных волокон медленного типа в группе 7HS был ниже, а процент «быстрых» мышечных волокон существенно выше по сравнению с группой С на 22% и 60% соответственно (Рис 2 А, Б) (р<0.05). Введение активатора SERCA предотвратило эти изменения. В группе 7HS+CDN процент «быстрых» мышечных волокон был достоверно ниже чем в группе 7HS и не имел достоверных отличий от группы С, а процент «медленных» мышечных волокон был достоверно выше, чем в группе 7HS, но несколько отличался от группы С (Рис 2 А, Б) (р<0.05).
Влияние активатора SERCA на окисление тропомиозина и параметры биогенеза митохондрий в камбаловидной мышце
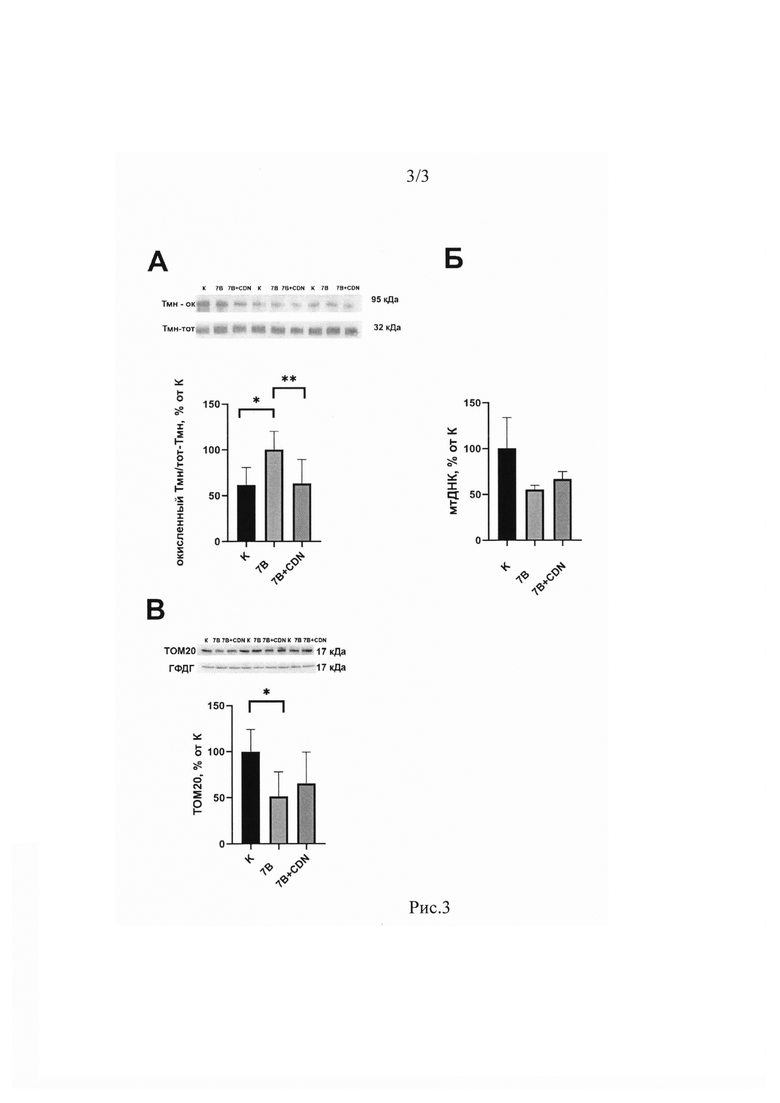
Уровень окисленного тропомиозина в группе 7HS достоверно вырос на 55% в сравнении с группой С, тогда как в группе 7HS+CDN уровень окисленного тропомиозина составлял 93% от значений группы С и был достоверно ниже значений группы 7HS (Рис. 3А).
Содержание митохондриальной ДНК в группе 7HS было на 55% ниже, чем в группе С (р<0,05). В группе 7HS+CDN содержание митохондриальной ДНК не имело достоверных отличий ни от группы С, ни от группы 7HS (Рис. 3Б). Содержание структурного белка митохондрий ТОМ20 в группе 7HS составляло 52% от гр. С (р<0,05), тогда как в группе 7HS+CDN содержание ТОМ20 не имело достоверных отличий от других экспериментальных групп (Рис. 3В).
Мы не обнаружили предотвращение атрофии m soleus в обеих вывешенных группах. 7-суточная функциональная разгрузка m soleus крыс привела к достоверному росту утомляемости m soleus. Аналогичные результаты получены ранее при разгрузке мышц человека и животных (Kubis, Scheibe et al. 2002, Lomonosova, Turtikova et al. 2016). Введение активатора SERCA на фоне 7-суточного вывешивания предотвратило рост утомляемости т soleus. Аналогичные эффекты CDN1163 на функциональные характеристики скелетных мышц, были получены ранее на мышах Sod1-/- и мышах с саркопенией (Qaisar, Bhaskaran et al. 2019, Qaisar, Pharaoh et al. 2020). Увеличение субмаксимальной активности SERCA при введении нитрата также предотвращало развитие утомления в т soleus (Petrick, Brownell et al. 2022).
Причины, предотвращающие снижение утомляемости m soleus: 1. Наблюдаемые эффекты 7-суточного вывешивания на изменение долей «быстрых» и «медленных» мышечных волокон в группе 7HS согласуются с ранее полученными данными (Sharlo, Paramonova et al. 2021). Вклад в увеличение устойчивости m soleus к утомлению в группе 7HS+CDN может вносить предотвращение снижения медленных мышечных волокон и увеличения количества быстрых мышечных волокон в этой группе.
2. Введение активатора SERCA предотвратило увеличение доли окисленного тропомиозина (Рис. 3А). Этот белок миофиламентов ранее был использован как показатель индекса окислительного стресса карбоксилирования/декарбоксилирования мышечных белков и маркера окислительного повреждения тканей (van der Velden 2006, Jackson 2016, Lechado, Vitadello et al. 2018). Эти результаты согласуются с данными о снижении уровня окислительного стресса при введении CDN1163 на мышах Sod1-/- (Qaisar, Bhaskaran et al. 2019) и в тучных клетках (Hunter, Crozier et al. 2023). Эта модификация тропомиозина может также изменять чувствительность к кальцию и максимальную способность миофиламентов генерировать силу (van der Velden 2006). Известен прямой эффект действия избыточного миоплазматического кальция на повышение утомляемости, связанный с усилением генерации активных форм кислорода митохондриями, которое приводит к окислению рианодиновых рецепторов, утечке (leakage) кальция из СПР и истощению кальциевого депо саркоплазматического ретикулума (СПР) (Andersson, Betzenhauser et al. 2011, Yutaka Kano 2012). Введение CDN1163 может предотвращать повышение утомляемости m soleus за счет предотвращения накопления избытка кальция и активных форм кислорода (АФК) в миоплазме.
3. Вклад в увеличение устойчивости т soleus к утомлению в группе 7HS+CDN может вносить предотвращение снижения в ней содержания митохондриальных белков и ДНК. Снижение содержания митохондриальной ДНК и белка ТОМ20, являющегося маркером митохондрий (Wei, Xiao et al. 2015, Buso, Comelli et al. 2019) согласуется с характерными для 7-суточного вывешивания показателями ухудшения состояния митохондрий (Wagatsuma, Kotake et al. 2011, Liu, Peng et al. 2012, Theeuwes, Gosker et al. 2017, Sharlo, Lvova et al. 2022). Влияние CDN1163 в группе 7HS+CDN на содержание митохондриальных белков, их мРНК и ДНК согласуется с данными об активации им утилизации жирных кислот и биогенеза митохондрий (Kang, Dahl et al. 2016, Mengeste, Lund et al. 2021). Положительный эффект CDN1163 в группе 7HS+CDN может быть связан со снижением уровня активных форм кислорода, вызывающих митохондриальный стресс (Heher, Ganassi et al. 2022). Другие авторы также обнаружили увеличение скорости дыхания митохондрий и снижение утомления т soleus при введении пищевого нитрата в качестве активатора SERCA (Petrick, Brownell et al. 2022). Предотвращение снижения содержания мтДНК и белка в группе 7HS+CDN может вносить вклад в увеличение устойчивости к утомлению т soleus.
Итак, введение активатора SERCA CDN1163 на фоне 7-суточной функциональной разгрузки привело к снижению уровня окисления тропомиозина (показателя окислительного стресса), способствовало сохранению доли «медленных» устойчивых к утомлению мышечных волокон, предотвратило увеличение количества «быстрых» волокон, а также митохондриальной ДНК и белка, и предотвратило повышение утомляемости камбаловидной мышцы.
Краткое описание рисунков
Рис. 1. Индекс утомления камбаловидных мышц экспериментальных животных. Данные представлены в виде средних значений±стандартная ошибка среднего. К - контрольная группа, 7 В - группа 7-суточного вывешивания, 7B+CDN - группа 7-суточного вывешивания с введением активатора SERCA. * - достоверные отличия от группы К, ** - достоверные отличия от группы 7В.
Рис. 2. Процент мышечных волокон «медленного» типа (А) и «быстрого» типа (Б) в камбаловидных мышцах экспериментальных животных по данным иммуногистохимического анализа. Данные представлены в виде средних значений±стандартная ошибка среднего. К - контрольная группа, 7В - группа 7-суточного вывешивания, 7B+CDN - группа 7-суточного вывешивания с введением активатора SERCA. * - достоверные отличия от группы К, ** - достоверные отличия от группы 7В.
Рис. 3. Соотношение окисленного тропомиозина к общему тропомиозину (А), содержание митохондриальной ДНК (Б) и содержание белка ТОМ20 (В) в камбаловидных мышцах экспериментальных животных по данным Вестерн-блоттинга. Данные представлены в виде средних значений±стандартная ошибка среднего. К - контрольная группа, 7В - группа 7-суточного вывешивания, 7B+CDN - группа 7-суточного вывешивания с введением с введением активатора SERCA. * - достоверные отличия от группы К, ** - достоверные отличия от группы 7В.
| название | год | авторы | номер документа |
|---|---|---|---|
| Способ предотвращения атрофии, повышенной утомляемости медленных мышечных волокон и их трансформации в быстрые мышечные волокна в камбаловидной мышце | 2024 |
|
RU2834246C1 |
| Способ поддержания силы и жесткости постуральных мышц млекопитающих при их функциональной разгрузке | 2022 |
|
RU2800317C1 |
| Способ предотвращения атрофии скелетных мышц при их функциональной разгрузке | 2022 |
|
RU2797216C1 |
| СПОСОБ ПРЕДОТВРАЩЕНИЯ АТРОФИИ СКЕЛЕТНЫХ МЫШЦ ПРИ ИХ ФУНКЦИОНАЛЬНОЙ РАЗГРУЗКЕ | 2012 |
|
RU2481105C1 |
| СПОСОБ ПРОФИЛАКТИКИ И СНИЖЕНИЯ ДЕСТРУКЦИИ ЦИТОСКЕЛЕТНЫХ БЕЛКОВ СКЕЛЕТНЫХ МЫШЦ ПРИ ИХ АТРОФИИ, ВЫЗВАННОЙ ГИПОКИНЕЗИЕЙ И/ИЛИ ГРАВИТАЦИОННОЙ РАЗГРУЗКОЙ | 2010 |
|
RU2444354C2 |
| СРЕДСТВО ДЛЯ ПРОФИЛАКТИКИ И СНИЖЕНИЯ ДЕСТРУКЦИИ БЕЛКОВ СКЕЛЕТНЫХ МЫШЦ ПРИ ИХ АТРОФИИ, ВЫЗВАННОЙ ГИПОКИНЕЗИЕЙ И/ИЛИ ГРАВИТАЦИОННОЙ РАЗГРУЗКОЙ | 2013 |
|
RU2517576C1 |
| СПОСОБ ПРОФИЛАКТИКИ И СНИЖЕНИЯ ДЕСТРУКЦИИ БЕЛКОВ СКЕЛЕТНЫХ МЫШЦ ПРИ ИХ АТРОФИИ, ВЫЗВАННОЙ ГИПОКИНЕЗИЕЙ И/ИЛИ ГРАВИТАЦИОННОЙ РАЗГРУЗКОЙ | 2013 |
|
RU2517259C1 |
| ПРИМЕНЕНИЕ VX 745 ДЛЯ ПРЕДОТВРАЩЕНИЯ АТРОФИИ СКЕЛЕТНЫХ МЫШЦ ПРИ ИХ ФУНКЦИОНАЛЬНОЙ РАЗГРУЗКЕ | 2019 |
|
RU2714923C1 |
| ИНГИБИТОР ПРОТЕОЛИЗА ЦИТОСКЕЛЕТНЫХ БЕЛКОВ СКЕЛЕТНЫХ МЫШЦ ПРИ ФИЗИЧЕСКОЙ НАГРУЗКЕ | 2008 |
|
RU2389486C1 |
| СПОСОБ ПРОФИЛАКТИКИ РАЗРУШЕНИЯ ЦИТОСКЕЛЕТНЫХ БЕЛКОВ СКЕЛЕТНЫХ МЫШЦ ПРИ ЭКСЦЕНТРИЧЕСКОЙ НАГРУЗКЕ | 2008 |
|
RU2391968C1 |
Изобретение относится к медицине, а именно к экспериментальной медицине и травматологии, и может быть использовано для предотвращения развития утомления скелетной мышцы. Способ включает инъекционное введение в организм на фоне 7-суточной функциональной разгрузки селективного активатора рецепторов SERCA (Са2+-АТФаза сарко/эндоплазматического ретикулума) - CDN1163 в дозировке 50 мг/кг ежедневно на протяжении 7 суток. Использование изобретения позволяет предотвратить повышение утомляемости мышцы путем активирования рецепторов SERCA и снижения уровня окисления тропомиозина (показателя окислительного стресса), сохранения доли «медленных» устойчивых к утомлению мышечных волокон, предотвращения увеличение количества «быстрых» волокон, а также митохондриальной ДНК и белка. 3 ил.
Способ предотвращения развития утомления скелетной мышцы, характеризующийся тем, что в организм на фоне 7-суточной функциональной разгрузки инъекционно вводят селективный активатор рецепторов SERCA CDN1163 в дозировке 50 мг/кг ежедневно на протяжении 7 суток.
| ПРИМЕНЕНИЕ VX 745 ДЛЯ ПРЕДОТВРАЩЕНИЯ АТРОФИИ СКЕЛЕТНЫХ МЫШЦ ПРИ ИХ ФУНКЦИОНАЛЬНОЙ РАЗГРУЗКЕ | 2019 |
|
RU2714923C1 |
| СРЕДСТВО ДЛЯ ПРОФИЛАКТИКИ И СНИЖЕНИЯ ДЕСТРУКЦИИ БЕЛКОВ СКЕЛЕТНЫХ МЫШЦ ПРИ ИХ АТРОФИИ, ВЫЗВАННОЙ ГИПОКИНЕЗИЕЙ И/ИЛИ ГРАВИТАЦИОННОЙ РАЗГРУЗКОЙ | 2013 |
|
RU2517576C1 |
| ПРОИЗВОДНЫЕ ТЕТРАГИДРОИЗОХИНОЛИНА | 2017 |
|
RU2743424C2 |
| Автомобиль-сани, движущиеся на полозьях посредством устанавливающихся по высоте колес с шинами | 1924 |
|
SU2017A1 |
| MENGESTE A | |||
| M | |||
| et al | |||
| The small molecule SERCA activator CDN1163 increases energy metabolism in human skeletal muscle cells//Current Research in Pharmacology and Drug Discovery, Volume 2, 2021,100060,ISSN 2590-2571, | |||

