ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Изобретение относится к выделенному полипептиду, имеющему NADH-зависимую HMF-редуктазную активность (NADH - никотинамид-аденин-динуклеотид, HMF - 5-гидроксиметилфурфурол), где указанный полипептид демонстрирует 80% гомологию с аминокислотной последовательностью, показанной в SEQ ID NO: 2, и отличается от SEQ ID NO: 2 тем, что по меньшей мере имеет замены S117L и Y295 или S110, к нуклеотидной последовательности, кодирующей указанный полипептид, вектору, содержащему указанный полипептид или нуклеотидную последовательность, хозяину, содержащему указанную нуклеотидную последовательность или вектор, а также применению полипептида для восстановления фурановых или карбонильных соединений в лигноцеллюлозном материале или любом фуран- или карбонилсодержащем материале.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Получение биоэтанола из возобновляемого сырья с помощью пекарских дрожжей Saccharomyces cerevisiae стало привлекательной альтернативой ископаемому топливу. Однако ожидают, что доступность крахмального или сахарозного сырья, такого как кукурузные зерна или сахарный тростник, будет недостаточной, чтобы удовлетворить будущую всемирную потребность в биоэтаноле (Gray et al., 2006. Bioethanol. Current Opinion Chemical Biology. 10(2): 141-146). Предсказуемое решение представляет собой утилизацию лигноцеллюлозного сырья, такого как грубые корма из кукурузы, солома пшеницы, жмых сахарного тростника, древесина и так далее (Hahn-Hägerdal et al., 2006. Bioethanol - the fuel of tomorrow from the residues of today. Trends Biotechnol. 24(12): 549-556). Это требует решения новых проблем, связанных с утилизацией этих смешанных видов сырья.
Одна из этих проблем затрагивает присутствие ингибирующих соединений, таких как небольшие алифатические низкомолекулярные кислоты, производные фурана, карбонильные соединения и фенолы, которые высвобождаются во время предварительной обработки и гидролиза лигноцеллюлозного сырья (Almeida et al., 2007. Increased tolerance of inhibitors in lignocellulosic hydrolysates by Saccharomyces cerevisiae. J Chem Technol Biotechnol, 82: 340-349). Среди этих соединений присутствие производного фурана 5-гидроксиметилфурфурола (HMF), который образуется при дегидратации гексоз, приводит, как сообщают, к снижению выработки этанола во время ферментации лигноцеллюлозных гидролизатов дрожжами S.cerevisiae (Taherzadeh et al. 2000. Physiological effects of 5-hydroxymethyl furfural on Saccharomyces cerevisiae. Appl Microbiol Biotechnol. 53(6): 701-708). При сравнении в эквимолярных количествах с фурфуролом, другим ингибирующим производным фурана, найденным в гидролизатах, и объемная выработка этанола, и скорость потребления сахара клетками ниже в случае HMF (Larsson et al., 1999. The generation of fermentation inhibitors during dilute acid hydrolysis of softwood. Enzyme Microbial Technology 24: 151-159). Следовательно, удаление HMF и/или усиление клеточной HMF детоксикации являются ключевыми для промышленного процесса ферментации, основанного на лигноцеллюлозном сырье.
В WO 03072602 раскрыт полипептид, имеющий серин в положении 295. Однако у этого полипептида отсутствует лейцин в положении 117.
Nilsson et al., Applied and Environmetal Microbiology, Dec 2005, vol 71, page 7866-7871, обнаружили, что клеточные экстракты из устойчивого к лигноцеллюлозному гидролизату штамма ТМВ3000 показали ранее неизвестную NADH-зависимую HMF-редуктазную активность, которая отсутствовала у менее устойчивого штамма CBS 8066.
Абсолютно необходимое условие для развития процессов ферментации, основанных на лигноцеллюлозном сырье, представляет собой развитие и оптимизацию микроорганизмов, которые устойчивы к ингибирующим соединениям, упомянутым выше, или штамма, который может утилизировать/разлагать ингибирующие соединения. Установлено, что штаммы S.cerevisiae восстанавливают HMF и фурфурол (Larsson et al., 1999. The generation of fermentation inhibitors during dilute acid hydrolysis of softwood. Enzyme Microbial Technology 24: 151-159) до 2,5-диметанола (2,5-бис-гидроксиметилфурана) и 2-фуранметанола, соответственно, однако скорость восстановления мала и зависит от штамма. Штамм ТМВ3000 (Lindén et al., 1992. Isolation and characterization of acetic acid-tolerant galactose-fermenting strains of Saccharomyces cerevisiae from a spent sulfite liquor fermentation plant. Applied Environmental Microbiology. 58(5): 1661-1669) пока представляется наиболее устойчивым штаммом, который может расти на неразбавленных гидролизатах древесины (Brandberg et al., 2005. Continuous fermentation of undetoxified dilute acid lignocellulose hydrolysate by Saccharomyces cerevisiae ATCC 96581 using cell recirculation. Biotechnology progress, 21(4): 1093-1101).
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Изобретение относится к выделению нового полипептида, имеющего NADH-зависимую HMF-редуктазную активность, где указанный полипептид обладает способностью восстанавливать несколько соединений. Один пример полипептида содержит мутантную алкогольдегидрогеназу (ADH1) из Saccharomyces cerevisiae, которая в нативной форме не может восстанавливать HMF, а когда мутирована - может восстанавливать HMF. Полипептид по изобретению может быть использован в нескольких способах получения крупнотоннажных и платформных (platform) химикатов из лигноцеллюлозного материала, такого как лигноцеллюлозное сырье, где существует необходимость детоксикации HMF, или других фуранов либо их производных, или карбонильных соединений. Примеры биотоплива, крупнотоннажных и платформных химикатов включают этанол, бутанол, лактат, 1,4-дикарбоновые кислоты (янтарная, фумаровая, яблочная), глицерин, сорбит, маннит, ксилит/арабинит, L-аскорбиновая кислота, ксилит, газообразный водород, 2,5-фурандикарбоновая кислота, 3-гидроксипропионовая кислота, аспарагиновая кислота, глутаровая кислота, глутаминовая кислота, итаконовая кислота, левулиновая кислота и 3-гидроксибутиролактон, жирные кислоты, молекулы, являющиеся производными жирных кислот, изопреноиды, молекулы, являющиеся производными изопреноидов, алканы, изопентанол, изоамилацетат. При использовании нового полипептида вследствие более быстрого удаления из среды ингибирующих фурановых соединений и карбонильных соединений будет достигнута увеличенная специфичная производительность (грамм продукта от грамма клеток и за час).
В одном аспекте изобретение относится к выделенному полипептиду, имеющему NADH-зависимую HMF-редуктазную активность, где указанный полипептид демонстрирует 80% гомологию с аминокислотной последовательностью, показанной в SEQ ID NO: 2, и отличается от SEQ ID NO: 2 тем, что по меньшей мере имеет замену S117L и Y295 или S110. Путем включения единичной замены в SEQ ID NO: 2 и в то же время сохранения L в положении получают усовершенствованный полипептид, который эффективно восстанавливает HMF и фурфурол.
Во втором аспекте изобретение относится к выделенному полипептиду, описанному выше, где указанный полипептид отличается от SEQ ID NO: 2 тем, что по меньшей мере имеет замену Y295C, или Y295S, или Y295T, или S110P.
В третьем аспекте изобретение относится к нуклеотидной последовательности, кодирующей полипептид, описанный выше.
В четвертом аспекте изобретение относится к вектору, содержащему нуклеотидную последовательность.
В пятом аспекте изобретение относится к клетке-хозяину, содержащей нуклеотидную последовательность или вектор.
В шестом аспекте изобретение относится к применению полипептида, имеющего NADH-зависимую HMF-редуктазную активность, нуклеотидной последовательности, вектора или клетки-хозяина для получения крупнотоннажных химикатов из лигноцеллюлозного сырья, такого как сырье, упомянутое выше. При применении усовершенствованных полипептидов возрастают скорость потребления сахара и скорость роста, а также, если хозяина используют для получения этанола, возрастает скорость выработки этанола микроорганизмом по сравнению с применением нативного полипептида.
При предоставлении нового полипептида, как показано в SEQ ID NO: 2, имеющего NADH-зависимую HMF-редуктазную активность, впервые возможно восстанавливать фурановые соединения, или карбонильные соединения, или производные фурана, такие как 6-гидроксиметил-2-фуральдегид (HMF), используя NADH в качестве кофактора. Сообщали, что HMF уменьшает и рост клеток, и выработку этанола в пекарских дрожжах, в большинстве случаев используемых для промышленного производства этанола. Эти две проблемы будут уменьшены или устранены сейчас, когда новый выделенный полипептид будет включен в способ или новая выделенная нуклеотидная последовательность будет перенесена в клетку-хозяина, которую используют в способе, в котором обычно есть проблема с HMF.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
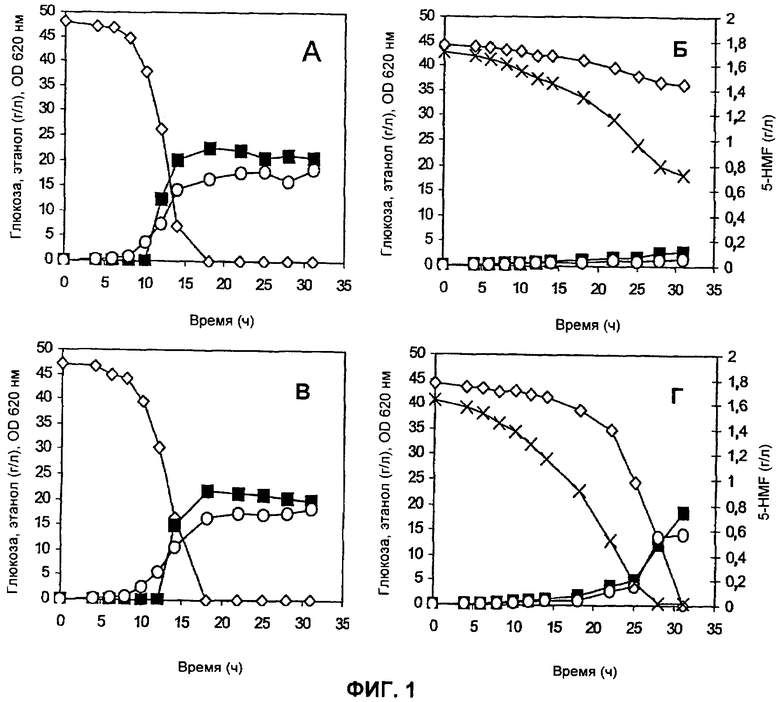
На Фиг.1 представлено потребление субстрата и HMF и образование основного продукта в штаммах ТМВ3280 (пустая плазмида) (А, Б) и ТМВ3206 (мутантная ADH1) (В, Г) в аэробных условиях. А, В = без добавления HMF; Б, Г = с добавлением 20 мМ HMF. Обозначения: ◇ глюкоза, ○ OD620 (OD - оптическая плотность), х HMF, ■ этанол.
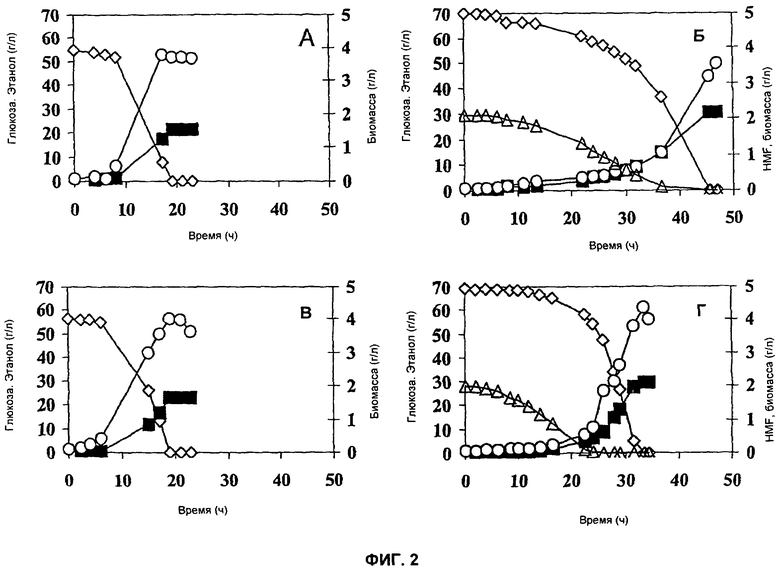
На Фиг.2 представлено потребление субстрата и HMF и образование основного продукта в штаммах ТМВ3280 (пустая плазмида) (А, Б) и ТМВ3206 (мутантная ADH1) (В, Г) в анаэробных условиях. А, В = без добавления HMF; Б, Г = с добавлением 20 мМ HMF. Обозначения: ◇ глюкоза, ○ OD620 (OD - оптическая плотность), Δ HMF, ■ этанол.
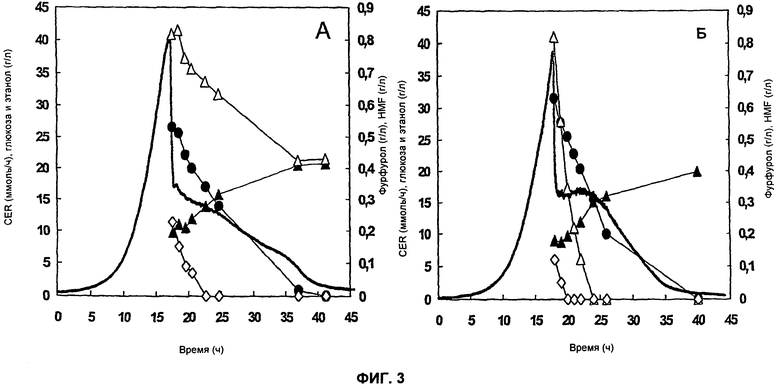
На Фиг.3 представлен ферментационный профиль контрольного штамма ТМВ3280 (А) и штамма ТМВ3206 с повышенной экспрессией мутантного гена ADH1 из ТМВ3000 (Б) в импульсном эксперименте с еловым гидролизатом. Штаммы сначала выращивают на питательной среде определенного состава, затем через 15-20 часов добавляют еловый гидролизат. Обозначения: Фурфурол (◇), HMF (Δ), Глюкоза (•), Этанол (▲), скорость выделения CO2, или CER (―).
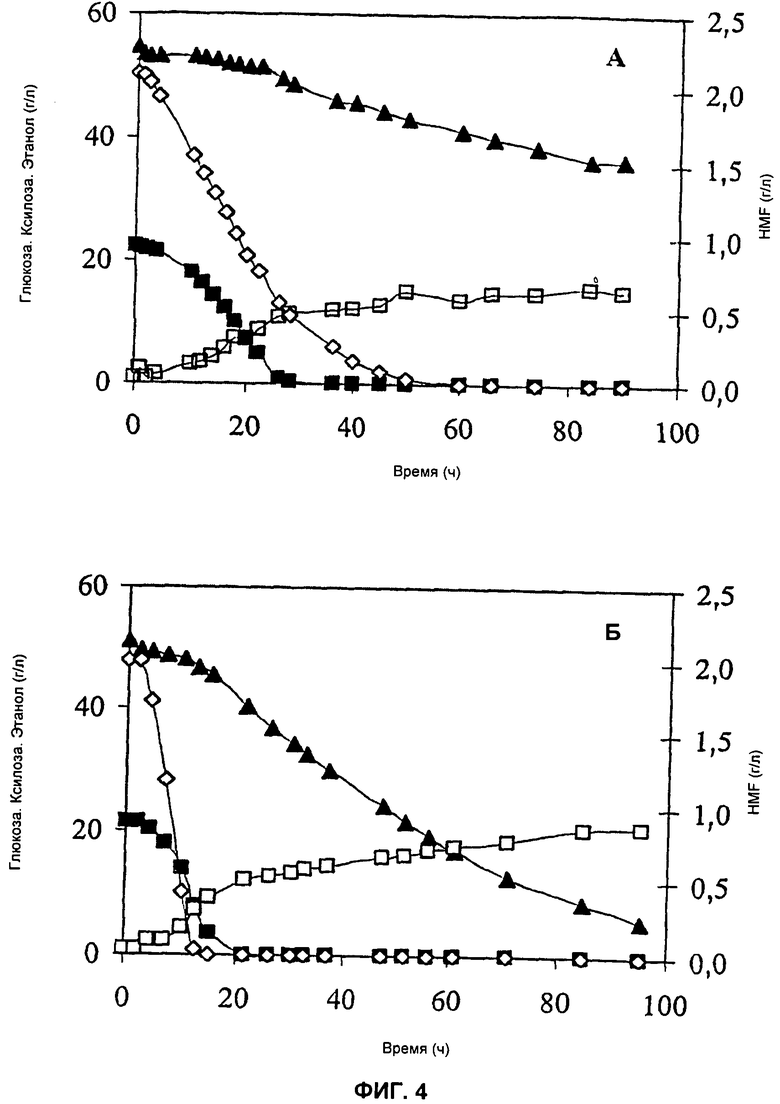
На Фиг.4 представлено потребление субстрата и HMF и образование этанола в штаммах ТМВ3290 (пустая плазмида) (А) и ТМВ3291 (ADH1-S110P-Y295C) (Б) в анаэробных условиях. Обозначения: (■) глюкоза, (▲) ксилоза, (◇) HMF, (□) этанол.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Определения
В контексте данной заявки и изобретения употребляются следующие определения.
Термин "нуклеотидная последовательность" обозначает последовательный отрезок из трех и более областей нуклеотидных последовательностей. Нуклеотиды могут быть из геномной ДНК, комплементарной ДНК (кДНК), РНК, полусинтетическими или синтетическими или их смесью. Термин включает одно- и двухцепочечные формы ДНК или РНК.
Термин "их аналог" обозначает, что часть или весь полипептид с SEQ ID NO: 2 основаны на небелковых аминокислотных остатках, таких как аминоизомасляная кислота (Aib), норвалин гамма-аминомасляная кислота (Abu) или орнитин. Примеры других небелковых аминокислотных остатков можно найти на http://www.hort.purdue.edu/rhodcv/hort640c/polyam/po00008.htm.
Термин "гомология" полипептида следует понимать как степень идентичности двух последовательностей, указывающая на происхождение первой последовательности от второй. Гомология может быть подходящим образом определена посредством компьютерных программ, известных в уровне техники, таких как GAP, входящих в программный пакет GCG (Program Manual for the Wisconsin Package, Version 8, August 1994, Genetics Computer Group, 575 Science Drive, Madison, Wis., USA 53711) (Needleman, S.B. and Wunsch, C.D., (1970), Journal of Molecular Biology, 48, 443-453. Используют следующие установки для сравнения аминокислотных последовательностей: штраф за создание пробела 3,0 и штраф за удлинение пробела 0,1. Релевантная для определения гомологии часть аминокислотной последовательности представляет собой зрелый полипептид.
Термин "вектор", как он использован здесь, относится к молекуле нуклеиновой кислоты или одно-, или двухцепочечной, которая выделена из встречающегося в природе гена или которая модифицирована так, что она содержит сегменты нуклеиновых кислот таким образом, что иначе не существует в природе. Термин "вектор" синонимичен термину "кассета экспрессии", когда вектор содержит регуляторные последовательности, необходимые для экспрессии кодирующей последовательности по настоящему изобретению. Термин "вектор" также синонимичен термину "интегрированная конструкция нуклеиновой кислоты" или "интегрированный фрагмент", когда конструкцию следует использовать для интеграции конструкции/фрагмента в геном хозяина.
Термин "регуляторные последовательности" определен здесь как включающий все компоненты, которые необходимы или полезны для экспрессии полинуклеотида, кодирующего полипептид по настоящему изобретению. Каждая регуляторная последовательность может быть нативной или чужеродной по отношению к нуклеотидной последовательности, кодирующей полипептид. Такие регуляторные последовательности включают последовательность полиаденилирования, пропептидную последовательность, промотор и терминатор транскрипции, но не ограничены ими. Как минимум регуляторные последовательности включают промотор и транскрипционные и трансляционные стоп-сигналы. Регуляторные последовательности могут быть обеспечены линкерами с целью введения специфических сайтов рестрикции, облегчающих лигирование регуляторных последовательностей с кодирующей областью нуклеотидной последовательности, кодирующей полипептид.
Термин "клетка-хозяин", как он использован здесь, включает любой тип клеток, который восприимчив к трансформации, трансфекции, трансдукции и подобному нуклеиновой кислотой.
В данном контексте названия аминокислот и названия атомов используют так, как определено Банком Данных Белка (PNB) (www.pdb.org). который основан на номенклатуре IUPAC (IUPAC Nomenclature and Symbolism for Amino Acids and Peptides (residue names, atom names etc.), Eur J Biochem., 138, 9-37 (1984) вместе с поправками в Eur J Biochem., 152, 1 (1985)). Термин "аминокислота" предназначен для обозначения аминокислоты из группы, состоящей из аланина (Ala или А), цистеина (Cys или С), аспарагиновой кислоты (Asp или D), глутаминовой кислоты (Glu или Е), фенилаланина (Phe или F), глицина (Gly или G), гистидина (His или Н), изолейцина (IIe или I), лизина (Lys или К), лейцина (Leu или L), метионина (Met или М), аспарагина (Asn или N), пролина (Pro или Р), глутамина (Gin или Q), аргинина (Arg или R), серина (Ser или S), треонина (Thr или Т), валина (Val или V), триптофана (Trp или W) и тирозина (Tyr или Y) или их производных.
Терминология, использованная для идентификации положений аминокислот, иллюстрирует следующее: S110 показывает, что положение 110 занято сериновым остатком в аминокислотной последовательности, показанной в SEQ ID NO: 2. S110P показывает, что сериновый остаток в положении 110 был заменен на пролиновый остаток.
Полипептид
Изобретение относится к новому полипептиду, имеющему NADH-зависимую HMF-редуктазную активность, где указанный полипептид демонстрирует 80% гомологию с аминокислотной последовательностью, показанной в SEQ ID NO: 2, по меньшей мере 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99%, или он такой же, как SEQ ID NO: 2. Полипептид по изобретению имеет несколько уникальных свойств, поскольку фермент использует NADH в качестве кофактора для восстановления HMF и других фурановых соединений и карбонильных соединений, таких как те, которые вырабатываются во время переработки лигноцеллюлозного материала.
Другое свойство выделенного полипептида состоит в том, что аминокислотный остаток в положении 295 или 110 SEQ ID NO: 2 заменен. Однако эта отдельная аминокислота может быть изменена, например, путем замены на другой аминокислотный остаток, такой как небольшой незаряженный аминокислотный остаток. Примеры представляют собой Y295C, или Y295S, или Y295T, или S110P.
В дополнение в полипептиде, показанном в SEQ ID NO: 2, аминокислотный остаток S117 должен быть заменен на L. Замена аминокислотного остатка, присутствующего в положении 295, важна и усиливает фурфуролредуктазу, а замещение обоих аминокислотных остатков, присутствующих в положениях 295 и 110, для получения этанола.
Один пример полипептида показан в SEQ ID NO: 2, в которой аминокислотные остатки 295 представляют собой С или S, 110 представляет собой Р. Другой конкретный пример представляет собой ситуацию, когда аминокислотный остаток 59 представляет собой Т, 210 представляет собой Р, 148 представляет собой Е, 152 представляет собой V и 295 представляет собой С или S в SEQ ID NO: 2. Полипептид, имеющий новую активность, то есть имеющий NADH-зависимую HMF-редуктазную активность, можно вследствие этого использовать в способе, в котором есть необходимость детоксикации фурановых соединений и карбонильных соединений. Также будет возросшей специфичная выработка вследствие того, что восстановление фуранов и карбонильных соединений, токсические эффекты которых замедляют весь процесс, будет более быстрым. Полипептид может быть синтетическим частично или полностью, пока сохранена активность.
Полипептид также может быть гибридным полипептидом. Термин "гибридный фермент" или "гибридный полипептид" предназначен для обозначения, например, тех полипептидов по изобретению, которые содержат первый набор аминокислотных последовательностей, содержащих аминокислотные остатки от приблизительно 110 до приблизительно 295, как показано в SEQ ID NO: 2, слитый/связанный со вторым набором аминокислотных остатков и, таким образом, создавая полностью синтетическую нуклеотидную последовательность, основанную на знании подходящих аминокислотных последовательностей, таких как линкеры, гомологичные аминокислотные последовательности и так далее.
Полипептид по изобретению клонировали из штамма дрожжей Saccharomyces cerevisiae TMB3000 (АТСС96581), у которого выявили наличие уникальной NADH-зависимой HMF-редуктазной активности. Во время исследования полипептида обнаружили, что этот полипептид похож на ADH1 полипептид, который обычно не может восстанавливать HMF. Единичное введение нескольких мутаций в этот полипептид изменило активность этого полипептида, и неожиданно обнаружили, что этот полипептид имел NADH-зависимую HMF-редуктазную активность. Однако полипептид можно получить из любого источника или даже создать путем синтеза при условии, что полипептид имеет свойства, как охарактеризованный полипептид.
Нуклеотидная последовательность
Также в задачу изобретения входит нуклеотидная последовательность, показанная в SEQ ID NO: 1, или нуклеотидная последовательность, имеющая по меньшей мере 60, 65, 70, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99% гомологию с нуклеотидной последовательностью, показанной в SEQ ID NO: 1, кодирующая полипептид, имеющий NADH-зависимую HMF-редуктазную активность. Нуклеотидную последовательность можно получить с помощью стандартных методов клонирования, используемых в генной инженерии для перемещения последовательности ДНК из ее естественного местоположения в другое, где она будет воспроизведена. Методы клонирования могут включать вырезание и выделение требуемого фрагмента ДНК, содержащего последовательность ДНК, кодирующую интересующий полипептид, вставку фрагмента в векторную молекулу и введение рекомбинантного вектора в клетку-хозяина, где будут реплицированы множественные копии или клоны последовательности ДНК. На выделенную последовательность ДНК можно воздействовать разными путями для обеспечения экспрессии интересующего полипептида. Манипуляции с последовательностью ДНК до ее включения в вектор могут быть желательны или необходимы в зависимости от вектора. Методики модифицирования последовательностей ДНК с использованием методов рекомбинантных ДНК хорошо известны в данной области техники.
Нуклеотидную последовательность для введения в ДНК клетки-хозяина можно интегрировать в векторы, содержащие нуклеотидную последовательность, функционально связанную с одной или более чем одной регуляторной последовательностью, которая управляет экспрессией кодирующей последовательности в подходящей клетке-хозяине в условиях, совместимых с регуляторными последовательностями. На нуклеотидную последовательность, кодирующую полипептид, можно воздействовать разными путями для обеспечения экспрессии полипептида.
Регуляторная последовательность может представлять собой подходящую промоторную последовательность, нуклеотидную последовательность, которая отвечает в клетке-хозяине за экспрессию нуклеотидной последовательности. Промоторная последовательность содержит транскрипционные регуляторные последовательности, которые опосредуют экспрессию полипептида. Промотор может представлять собой любую нуклеотидную последовательность, которая проявляет транскрипционную активность в предпочтительной клетке-хозяине, включающую нативные, мутантные, усеченные и гибридные промоторы, и может быть получена из генов, кодирующих внеклеточные и внутриклеточные полипептиды или гомологичные, или гетерологичные по отношению к клетке-хозяину. Промотор может быть слабым или сильным промотором, который является конститутивным или регулируемым в используемом хозяине.
Примеры подходящих промоторов для управления транскрипцией конструкций из нуклеиновых кислот в бактериях по настоящему изобретению описаны в "Useful proteins from recombinant bacteria" in Scientific American, 1980, 242: 74-94; и в Sambrook J et al., 1989 Molecular Cloning: A Laboratory Manuel. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, выше.
Примеры подходящих промоторов для управления транскрипцией конструкций из нуклеиновых кислот по настоящему изобретению в клетках-хозяевах нитчатых грибов представляют собой промоторы, полученные из генов ТАКА амилазы Aspergillus oryzae, аспарагиновой протеиназы Rhizomucor miehei, нейтральной альфа-амилазы Aspergillus niger, кислотоустойчивой альфа-амилазы Aspergillus niger, глюкоамилазы (glaA) Aspergillus niger или Aspergillus awamori, липазы Rhizomucor miehei, щелочной протеазы Aspergillus oryzae, триоза-фосфатизомеразы Aspergillus oryzae, ацетамидазы Aspergillus nidulans и трипсиноподобной протеазы Fusarium oxysporum (WO 96/00787), a также NA2-tpi промотор (гибрид промоторов из генов нейтральной альфа-амилаза Aspergillus niger и триоза-фосфатизомеразы Aspergillus oryzae) и мутантные, усеченные и гибридные промоторы.
В дрожжевом хозяине пригодные промоторы можно получить, например, из генов енолазы (ENO1) Saccharomyces cerevisiae, галактокиназы (GAL1) S.cerevisiae, алкогольдегидрогеназы 2 (ADH2), глицеральдегид-3-фосфат-дегидрогеназы (TDH1) S.cerevisiae, глицеральдегид-3-фосфат-дегидрогеназы (TDH3) S.cerevisiae (Bitter and Egan. Expression of heterologous genes in Saccharomyces cerevisiae from vectors utilizing the glyceraldehyde-3-phosphate dehydrogenase gene promoter. (1984) Gene 32: 263-274), алкогольдегидрогеназы 1 (ADH1) S.cerevisiae, 3-фосфоглицерат-киназы (PGK1) S.cerevisiae или цитохрома С (CYC1) S.cerevisiae (Karhumaa et al. Investigation of limiting metabolic steps in the utilization of xylose by recombinant Saccharomyces cerevisiae using metabolic engineering. (2005) Yeast 5: 359-68). Другой пример дрожжевого промотора представляет собой конститутивный усеченный промотор НХТ7 (Hauf et al. Enzym Microb Technol (2000) 26: 688-698). Другие подходящие векторы и промоторы для применения в дрожжевой экспрессии описаны в документе ЕР А-73657 (Hitzeman), который включен сюда посредством ссылки.
Настоящее изобретение также относится к векторам, как определено выше, которые могут содержать последовательность ДНК, кодирующую полипептид, промотор и транскрипционные и трансляционные стоп-сигналы, а также другие последовательности ДНК. Вектор содержит разнообразные ДНК и регуляторные последовательности, известные специалисту в данной области техники, которые можно соединять вместе для создания вектора, который может содержать один и более удобных сайтов рестрикции, чтобы сделать возможной вставку или замену последовательности ДНК, кодирующей полипептид, в таких сайтах. В качестве альтернативы последовательность ДНК по настоящему изобретению можно экспрессировать посредством вставки последовательности ДНК или конструкции ДНК, содержащей последовательность, в подходящий вектор экспрессии. При создании вектора кодирующая последовательность расположена в векторе так, что кодирующая последовательность функционально связана с подходящими регуляторными последовательностями для экспрессии и, возможно, секреции.
Вектор может представлять собой любой вектор (например, плазмидный, вирусный или интеграционный вектор), который можно легко подвергнуть методикам рекомбинантных ДНК и который может осуществить экспрессию последовательности ДНК. Выбор вектора обычно будет зависеть от совместимости вектора с клеткой-хозяином, в которую вводят вектор. Векторы могут быть линейными или замкнутыми кольцевыми плазмидами. Вектор может быть автономно реплицирующимся вектором, то есть вектором, который существует в виде внехромосомного объекта, репликация которого является независимой от репликации хромосомы, например плазмидой, внехромосомным элементом, минихромосомой, космидой или искусственной хромосомой. Вектор может содержать любые ресурсы, обеспечивающие саморепликацию. В качестве альтернативы вектор может быть одним из тех, которые, будучи введены в клетку-хозяина, интегрируются в геном и реплицируются вместе с хромосомой(ами), в которую интегрировались. Векторная система может быть единичным вектором или плазмидой или двумя и более векторами или плазмидами, которые вместе содержат всю ДНК, вводимую в геном клетки-хозяина, или транспозоном. Вектор также может быть интеграционным вектором, содержащим только ген или часть гена для интеграции.
Векторы по настоящему изобретению предпочтительно содержат один или более селектируемых маркеров, которые позволяют легко отобрать трансформированные клетки. Селектируемый маркер представляет собой ген, продукт которого обеспечивает биоцидную или вирусную устойчивость, устойчивость к тяжелым металлам, прототрофность для ауксотрофов и подобное.
Полезные для эукариотических хозяев векторы экспрессии включают, например, векторы, содержащие регуляторные последовательности из SV40 (вируса обезьян 40), вируса папилломы быка, аденовируса или цитомегаловируса. Специфичные векторы представляют собой, например, pCDNA3.1(+)Hyg (Invitrogen, Carlsbad, Calif, U.S.A.) и pCl-neo (Stratagene, La Jolla, Calif., U.S.A.). Полезные векторы экспрессии для дрожжевых клеток включают, например, 2µ (микрон) плазмиду и ее производные, векторы YIp, YEp и YCp, описанные Gietz и Sugino (1988, "New yeast vectors constructed with in vitro mutagenized yeast genes lacking six-base pair restriction sites", Gene 74: 527-534), векторы, описанные в Mumberg et al (Mumberg, Muller and Funk, 1995, "Yeast vectors for the controlled expression of heterologous proteins in different genetic backgrounds." Gene 156: 419-422), вектор YEplac-HXT (Karhumaa et al, 2005. Investigation of limiting metabolic steps in the utilization of xylose by recombinant Saccharomyces cerevisiae using metabolic engineering. Yeast. 22(5): 359-68)), вектор РОТ1 (патент США №4931373), вектор pJSO37, описанный в Okkels, Ann. New York Acad. Sci. 782, 202-207, 1996, векторы pPICZ А, В или С (Invitrogen). Полезные для клеток насекомых векторы включают pVL941, pBG311 (Gate et al., "Isolation of the Bovine and Human Genes for Mullerian Inhibiting Substance and Expression of the Human Gene in Animal Cells", Cell, 45, pp.685-98 (1986)), pBluebac 4.5 и pMelbac (оба доступны от Invitrogen). Полезные векторы экспрессии для бактериальных хозяев включают известные бактериальные плазмиды, такие как плазмиды из Е. coli, включая pBR322, рЕТ3а и рЕТ12а (обе от Novagen Inc., Wis., U.S.A.), плазмиды с более широким спектром хозяев, такие как RP4, ДНК фагов, например множество производных фага лямбда, например NM989, и других ДНК-фагов, таких как М13 и нитевидные одноцепочечные ДНК фаги. Примеры подходящих вирусных векторов представляют собой аденовирусные векторы, аденоассоциированные вирусные векторы, ретровирусные векторы, лентивирусные векторы, герпес-векторы и цитомегаловирусные векторы.
Векторы по настоящему изобретению могут содержать элемент(ы), который(е) дает(ют) возможность стабильной интеграции вектора в геном клетки-хозяина или автономной репликации вектора в клетке, независимой от генома клетки. Векторы по настоящему изобретению можно интегрировать в геном клетки-хозяина при введении в клетку-хозяина. Для интеграции вектор может быть основан на последовательности ДНК, кодирующей интересующий полипептид, или любом другом элементе вектора для стабильной интеграции вектора в геном с помощью гомологичной или негомологичной рекомбинации.
В качестве альтернативы вектор может содержать дополнительные последовательности ДНК для управления интеграцией в геном клетки-хозяина с помощью гомологичной рекомбинации. Дополнительные последовательности ДНК дают возможность вектору интегрироваться в геном клетки-хозяина в определенном(ых) месте(ах) в хромосоме(ах). Чтобы увеличить вероятность интеграции в определенное место, интеграционные элементы предпочтительно должны содержать достаточное количество нуклеотидов, такое как 100-1500 пар оснований, предпочтительно 400-1500 пар оснований и наиболее предпочтительно 800-1500 пар оснований, которые высоко гомологичны соответствующей последовательности-мишени, чтобы увеличить возможность гомологичной рекомбинации. Интеграционные элементы могут быть любой последовательностью, которая гомологична последовательности-мишени в геноме клетки-хозяина. Кроме того, интеграционные элементы могут быть некодирующими или кодирующими последовательностями ДНК. С другой стороны, вектор можно интегрировать в геном клетки-хозяина с помощью негомологичной рекомбинации. Эти последовательности ДНК могут быть любыми последовательностями, которые гомологичны последовательности-мишени в геноме клетки-хозяина, и, кроме того, могут быть некодирующими или кодирующими последовательностями. Более одной копии последовательности ДНК, кодирующей интересующий полипептид, можно ввести в клетку-хозяина, чтобы усилить экспрессию последовательности ДНК.
Выделенная клетка-хозяин
Изобретение также относится к выделенной клетке-хозяину, которая содержит нуклеотидную последовательность, как описано выше, или в векторе, таком как вектор экспрессии, или в качестве альтернативы имеет нуклеотидную последовательность, интегрированную в геном, например, с помощью гомологичной или гетерологичной рекомбинации. Нуклеотидная последовательность может быть представлена в виде одной копии или множества копий.
Клеткой-хозяином может быть любая подходящая прокариотическая или эукариотическая клетка, например бактериальная клетка, клетка нитчатых грибов, дрожжевая клетка, растительная клетка или клетка млекопитающего. Любую подходящую клетку-хозяина можно использовать для сохранения и получения вектора по изобретению, такую как эукариотическая или прокариотическая клетка, например бактериальная, грибная (включая дрожжи), растительная клетка, клетка насекомого, млекопитающего или другие подходящие клетки или линии клеток животных, а также трансгенные животные или растения. Клетка-хозяин может быть клеткой-хозяином, принадлежащей линии клеток, сертифицированной согласно Правилам организации производства и контроля качества лекарственных средств (GMP), такой как линия клеток млекопитающих.
Примеры бактериальных клеток-хозяев включают Escherichia coli, Zymomonas sp. и Klebsiella sp.
Примеры подходящих клеток-хозяев нитчатых грибов включают Aspergillus sp., например A.oryzae, A.niger или A.nidulans, Fusarium sp. или Hypocrea (ранее Trichoderma) sp.
Примеры подходящих дрожжевых клеток-хозяев включают Saccharomyces sp., например S.cerevisiae, S.bayanus или S.carlsbergensis, Schizosaccharomyces sp., такие как Sch. pombe, Kluyveromyces sp., такие как К.lactis, Pichia sp., такие как Р.stipitis, P.pastoris или Р.methanolica, Hansenula sp., такие как H.polymorpha, Candida sp., такие как С.Shehatae, или Yarrowia sp.. Примеры штаммов S.cerevisiae представляют собой DBY746, AH22, S150-2B, GPY55-15Bα, CEN.PK, USM21, TMB3500, TMB3400, VTT-A-63015, VTT-A-85068, VTT-c-79093) и их производные, а также Saccharomyces sp.1400, 424А (LNH-ST), 259A (LNH-ST) и их производные.
Примеры подходящих клеток-хозяев насекомых включают линию клеток Lepidoptora, таких как Spodoptera frugiperda (Sf9 или Sf21) или клетки Trichoplusioani (High Five) (патент США №5077214).
Примеры подходящих клеток-хозяев млекопитающих включают линии клеток яичников китайских хомячков (СНО) (например, СНО-К1; АТСС CCL-61 (АТСС - Американская коллекция типовых культур)), линии клеток зеленых мартышек (COS) (например, COS 1 (АТСС CRL-1650), COS 7 (АТСС CRL-1651)), клетки мышей (например, NS/O), линии клеток почки детенышей хомяков (ВНК) (например, АТСС CRL-1632 или АТСС CCL-10) и клетки человека (например, НЕК 293 (АТСС CRL-1573)), а также растительные клетки в культуре ткани.
Полипептид, нуклеотидную последовательность, вектор или клетку-хозяина по изобретению можно использовать в производстве биотоплива, крупнотоннажных и платформных химикатов, таких как этанол, бутанол, лактат, 1,4-дикарбоновые кислоты (янтарная, фумаровая, яблочная), глицерин, сорбит, маннит, ксилит/арабинит, L-аскорбиновая кислота, ксилит, газообразный водород, 2,5-фурандикарбоновая кислота, 3-гидроксипропионовая кислота, аспарагиновая кислота, глутаровая кислота, глутаминовая кислота, итаконовая кислота, левулиновая кислота и 3-гидроксибутиролактон, жирные кислоты, молекулы, являющиеся производными жирных кислот, изопреноиды, молекулы, являющиеся производными изопреноидов, алканы, изопентанол, изоамилацетат. Общая схема способа превращения лигноцеллюлозного сырья в крупнотоннажные химикаты, такие как этанол, может включать такие стадии, как предварительная обработка или стадия фракционирования, на которой измельченное сырье подвергают воздействию нейтрального, кислого или щелочного pH при высокой температуре с добавлением воздуха/кислорода или без него так, что фракция гемицеллюлозы частично гидролизуется до мономерных и олигомерных сахаров, приводя целлюлозную фракцию в состояние восприимчивости к гидролизу, или на которой измельченное сырье подвергают воздействию органического растворителя, такого как ацетон, этанол и подобные, при высокой температуре, так что фракция лигнина растворяется и ее экстрагируют, что приводит целлюлозную и гемицеллюлозную фракции в состояние восприимчивости к гидролизу. Гидролиз предварительно обработанного и фракционированного материала можно осуществлять с помощью концентрированных или разбавленных кислот или с помощью смесей целлюлозолитических и гемицеллюлозолитических ферментов.
Следующие примеры предназначены для иллюстрации, но не для ограничения изобретения любым способом или формой, явно или неявно.
ПРИМЕРЫ
ПРИМЕР 1
Идентификация NADH-зависимой HMF-редуктазы
Очистка NADH-зависимой HMF-редуктазы
Промышленный штамм ТМВ3000 (АТСС96581) (Lindén et al., 1992, Isolation and characterization of acetic acid-tolerant galactose-fermenting strains of Saccharomyces cerevisiae from a spent sulfite liquor fermentation plant. Applied Environmental Microbiology 58(5): 1661-1669) выращивали на богатой среде, содержащей 10 г/л экстракта дрожжей, 20 г/л пептона и 20 г/л глюкозы, дополненной 10% гидролизатом ели (Nilvebrant et al., 2003, Limits for alkaline detoxification of dilute-acid lignocellulose hydrolysates. Applied Biochemistry Biotechnology. 105-108: 615-628), доведенной до pH 5,5. Гидролизат центрифугировали в течение 10 минут при 5000 g и надосадочную жидкость пропускали через 2µ фильтр (Sarstedt, Nümbrecht, Germany) перед добавлением к автоклавированной среде. Рост происходил в 1 л качалочных колбах (без перегородки), содержащих 250 мл среды, при 200 об/мин и 30°C. Посев из 5 мл ночной предварительной культуры на богатую среду (без гидролизата) имел оптическую плотность при 620 нм (OD620), равную 0,1, и клетки собирали через 24 часа при OD620 примерно 4.
Клетки собирали и промывали два раза дважды дистиллированной водой перед ресуспендированием в детергенте Y-PER (Pierce, Rockford, IL) 1 мл на 0,6 г клеток. После легкого встряхивания при комнатной температуре в течение 50 минут суспензию центрифугировали в течение 20 минут при 15000 g. Надосадочную жидкость собирали и использовали для ферментативных анализов и дополнительных стадий очистки. Кумасси-реагент для анализов белков (Pierce, Rockford, IL) использовали в анализе по Брэдфорду для определения концентрации белка согласно рекомендациям производителя. Измерения NADH-зависимого восстановления фурана проводили согласно Wahlbom и Hahn-Hägerdal (2002, Furfural, 5-hydroxymethyl furfural, and acetoin act as external electron acceptors during anaerobic fermentation of xylose in recombinant Saccharomyces cerevisiae. Biotechnology and Bioengineering. 78(2): 172-178) со следующей модификацией: концентрацию NADH устанавливали 200 мкМ. Все измерения были выполнены при 30°C на спектрофотометре U-2000 (Hitachi, Tokyo, Japan).
Для идентификации фермента, ответственного за NADH-зависимое HMF-восстановление, белковый раствор фракционировали, используя осаждение сульфатом аммония, эксклюзионную и аффинную хроматографию согласно следующему протоколу: клеточный экстракт (15 мл) смешивали с 10 мл насыщенного раствора сульфата аммония до конечной концентрации 40%. После 1 час легкого встряхивания при 4°C смесь центрифугировали при 15000 g в течение 20 минут. Осадок хранили на льду для немедленного использования или при -20°C для более позднего использования (назвали 40% осадок), в то время как надосадочную жидкость смешивали с равным объемом так, что получился 70% раствор сульфата аммония. После 3 часов легкого встряхивания при 4°C повторили центрифугирование при 15000 g в течение 20 минут. Осадок снова хранили на льду или при -20°C (назвали 70% осадок), в то время как надосадочную жидкость (приблизительно 40 мл) диализировали в течение ночи против 12 л дистиллированной воды при 4°C, используя мембранные трубки Spectra/Рог (Номинальное Отсечение по Молекулярной Массе (MWCO) 12 кДа - 14 кДа) от Spectrum Laboratories, CA, USA. Эксклюзионную хроматографию осуществляли, используя колонку HiLoad 16/60 Superdex 200 (Amersham Pharmacia Biotech, Uppsala, Sweden). Буфер содержал 100 мМ Tris-HCl и 0,5 М NaCl (pH повышали до 6,7 NaOH), оба от Merck, Darmstadt, Germany. Скорость потока установили на 1 мл/мин, используя систему жидкостной экспресс-хроматографии белков (FPLC) (Amersham Pharmacia Biotech, Uppsala, Sweden), управляемую контроллером LCC-501 Plus. 50 мл набухшего красной сефарозы CL-6B поместили в колонку ХК50, оба от Amersham Pharmacia Biotech, Uppsala, Sweden. Связывающий буфер содержал 20 мМ Tris-HCl, pH 6,4, 5 мМ MgCl2, 0,4 мМ этилендиаминтетрауксусной кислоты (EDTA) и 2 мкМ β-меркаптоэтанола. Буфер для элюирования содержал в дополнение 10 мМ β-NAD+ (Sigma-Aldrich, St. Louis, МО, USA). Скорость потока установили на 5 мл/мин, используя систему FPLC, упомянутую выше.
Результаты стадий очистки представлены в таблице 1.
Очищенные фракции разгоняли методом электрофореза в полиакриламидном геле в присутствии додецилсульфата натрия (SDS-PAGE), используя Tris-HCl 4%-15% градиентный готовый гель и электрофоретический аппарат Mini-PROTEAN 3, оба от Bio-Rad (Hercules, CA, USA). Двойной краситель Precision Plus Protein (Bio-Rad, Hercules, CA, USA) использовали в качестве стандарта. Окрашивание геля проводили, используя 0,25% кумасси бриллиантовый голубой R-250 (ICN Biomedicals, Aurora, ОН, USA) в смеси H2O:МеОН:НАс 6:3:1. Все результаты масс-спектрометрии получали из центра ресурсов протеомики SWEGENE в Лунде, Швеция.
Конечная очищенная фракция показала одну полосу на SDS-PAGE, соответствующую молекулярной массе 37 кДа. Очищенный белок идентифицировали как алкогольдегидрогеназу 1 S.cerevisiae (Adh1), используя масс-спектрометрию с ионизацией электрораспылением (ESI-MS).
Клонирование гена ADH1 из S.cerevisiae TMB3000
Ген ADH1 из TMB3000 амплифицировали с помощью полимеразной цепной реакции (PCR) из геномной ДНК и клонировали в вектор YEplac-HXT, несущий сильный конститутивный усеченный промотор НХТ (Hauf et al,. 2000. Simultaneous genomic overexpression of seven glycolytic enzymes in the yeast Saccharomyces cerevisiae. Enzyme Microb Technol, 26: 688-698) (Karhumaa et al., 2005. Investigation of limiting metabolic steps in the utilization of xylose by recombinant Saccharomyces cerevisiae using metabolic engineering. Yeast. 22(5): 359-368).
Конкретнее, геномную ДНК TMB3000 экстрагировали, используя набор Y-DER (Pierce, Rockford, IL) согласно рекомендациям производителя. PCR для амплификации гена ADH1 из TMB3000 проводили, используя ДНК-полимеразу Pwo (Roche Diagnostics AB, Bromma, Sweden), с температурой отжига 42°C. Два прямых и два обратных праймера использовали во всех возможных комбинациях (сайты рестрикции для последующих расщепления и лигирования подчеркнуты): Прямой A-5'-GGGCGGATCCATACAATGTCTATCCCAGAAA-3', Прямой Б-5'-GGGGGGATCCATGTCTATCCCAGAAACTC-3', Обратный А-5'-CTTTAGATCTTTATTTAGAAGTGTCAACAACG-3', Обратный Б-5'-CTTTAGATCTGCTTATTTAGAAGTGTCAACA-3'. Очищенные ампликоны смешивали и расщепляли с помощью ферментов BamHI и Bg/II (Fermentas, Vilnius, Lithuania), а затем использовали для лигирования с плазмидой YEplac-HXT, которую предварительно расщепляли теми же ферментами рестрикции и дефосфорилировали. Продукт лигирования использовали для трансформации Escherichia coli DH5α (Life Technologies, Rockville, MD, USA) по способу Inoue et al. (1990, High efficiency transformation of Escherichia coli with plasmids. Gene 96: 23-28). Трансформированные клетки отбирали на чашках с агаровой средой Лурия-Бертани (LB) (Ausubel et al., 1995. Current protocols in molecular biology. Wiley, New York), содержащей ампициллин (50 мкг/мл). PCR осуществили на нескольких клонах, чтобы проверить правильность лигирования, и плазмиду выделяли, чтобы трансформировать штамм S. cerevisiae BY4741 ΔADH1 (Invitrogen, Groningen, the Netherlands), используя метод с ацетатом лития (Gietz et al., 1992. Improved method for high efficiency transformation of intact yeast cells. Nucleic Acids Research 20(6): 1425). Трансформированные клетки отбирали анаэробно на чашках со средой SD-ura (Ausubel et al., 1995. Current protocols in molecular biology. Wiley, New York), содержащей 20 мМ HMF (Sigma-Aldrich, location), 400 мкг/мл Tween 80 и 10 мкг/л эргостерина. Плазмиду выделяли из клонов, показывающих наивысшую NADH-зависимую HMF-редуктазную активность, выращивая клетки в течение ночи в 5 мл среды SD без урацила, промывая дважды дистиллированной водой и ресуспендируя в 1 мл раствора для протопластов (1,2 М сорбита, 100 мМ Tris pH 7,5, 10 мМ CaCl2, 4 Ед/мл зимолиазы, 0,5% β-меркаптоэтанола). После 30 минут встряхивания при 30°C клетки собирали и использовали для выделения плазмид с помощью набора QIAprep Spin Miniprep (Qiagen, Hilden, Germany). E.coli трансформировали плазмидой и отбирали на чашках с LB с 50 мкг/мл ампициллина. Колонии выращивали в 5 мл среды LB, содержащей ампициллин (50 мкг/мл), и использовали для выделения плазмид с помощью набора QIAprep Spin Miniprep (Qiagen, Hilden, Germany). Полученную плазмиду назвали YEplac-HXT-ADH1-mut.
Ген ADH1 из YEPlac-HXT-ADH1-mut секвенировали, используя набор Abi-Prism BigDye cycle sequencing (Applied Biosystems, Weiterstadt, Germany). Шесть мутаций, приводящих к аминокислотным заменам (таблица 2), обнаружили при сравнении секвенированного гена ADH1 из ТМВ3000 с нативным геном ADH1 (как сообщается в базе данных геномов Saccharomyces (SGD), www.yeastgenome.org. систематическое название YOL086C).
ПРИМЕР 2
In vivo поглощение HMF штаммом, экспрессирующим NADH-зависимую редуктазу
Сверхэкспрессия в S.cerevisiae CEN.PK 113-5D
Плазмиду YEplac-HXT-ADH1-mut, кодирующую мутантный ген ADH1 (как описано в примере 1), а также соответствующую пустую плазмиду YEplacHXT (Karhumaa et al., 2005, Investigation of limiting metabolic steps in the utilization of xylose by recombinant Saccharomyces cerevisiae using metabolic engineering. Yeast. 22(5): 359-368) использовали для трансформации S.cerevisiae CEN.PK 113-5D (SUC2, MAL2-8c, MEL, ura3), что привело к получению штаммов ТМВ3206 и ТМВ3280, соответственно. Как ожидали, NADH-зависимую HMF-редуктазную активность обнаружили только в штамме, несущем мутантный ген ADH1 (данные не показаны).
Аэробное восстановление HMF in vivo
Штаммы ТМВ3206 и ТМВ3280 предварительно выращивали в течение ночи в 5 мл питательной среды определенного состава (Verduyn et al., 1992. Effect of benzoic acid on metabolic fluxes in yeasts: a continuous-culture study on the regulation of respiration and alcoholic fermentation. Yeast, 8, 501-517) при 30°C и 200 об/мин. Клетки использовали для инокуляции до OD620 нм, равной 0,1, 100 мл питательной среды определенного состава с добавлением 40 г/л глюкозы в 1 л качалочных колбах с перегородками. Эксперименты, проведенные при 30°C и 200 об/мин, проводили с HMF (2 г/л) и без него, добавленным к обоим штаммам. Образцы отбирали каждые 2-4 часа.
Концентрацию клеток определяли по измерениям оптической плотности при длине волны 620 нм, калиброванной против измерений массы в сухом состоянии из повторных выборок. Для получения образцов массы в сухом состоянии 10 мл суспензии клеток фильтровали в вакууме через предварительно взвешенные фильтры Gelman (d=47 мм Supor-450, 0,45 мкм). Фильтры промывали водой и сушили. Образцы для анализа концентраций метаболитов регулярно отбирали из ферментатора. Образцы фильтровали через фильтры 0,2 мкм. Концентрации глюкозы, этанола и HMF измеряли на колонке Aminex HPX-87H (Bio-Rad, Hercules, California) при 65°C. Подвижная фаза представляла собой 5 мМ H2SO4 со скоростью потока 0,6 мл/мин. Все соединения обнаружили с помощью детектора показателя преломления, кроме HMF, который обнаружили с помощью ультрафиолетового (UV) детектора при длине волны 210 нм.
Характерный результат эксперимента показан на Фиг.1. Потребление глюкозы, образование этанола и продукция биомассы были аналогичны для обоих штаммов, когда в среду не добавляли HMF (Фиг.1А, 1В). Напротив, при добавлении в среду HMF наблюдали значительно более высокую скорость потребления глюкозы, продукции этанола и биомассы в штамме ТМВ3206, сверхэкспрессирующем мутантный ген ADH1, по сравнению с контрольным штаммом ТМВ3280 (Фиг.1Б, 1Г). Эксперимент подтвердил, что повышенная in vitro NADH-зависимая HMF-редуктазная активность коррелировала с повышенной in vivo скоростью восстановления HMF для штамма ТМВ3206, сверхэкспрессирующего мутантный ген ADH1.
Анаэробное восстановление HMF in vivo
Культуры инокулята штаммов ТМВ3280 (контрольный) и ТМВ3206 (сверхэкспрессирующий мутантный ген ADH1) выращивали в течение 24 часов при 30°C и 150 об/мин в 1 л качалочных колбах с 100 мл дважды концентрированной минеральной питательной среды определенного состава (как описано в Verduyn et al, 1992, Effect of benzoic acid on metabolic fluxes in yeasts: a continuous-culture study on the regulation of respiration and alcoholic fermentation. Yeast 8, 501-517), дополненной 40 г/л глюкозы и 200 мл/л фталатного буфера (10,2 г/л КН фталата, 2,2 г/л КОН). Такую же среду определенного состава с HMF (2 г/л) или без него использовали в следующей серии экспериментов. Среду дополняли 60 г/л глюкозы, эргостерином (0,075 г/л) и Tween 80 (0,84 г/л). Исходная OD620 в периодической ферментации составила 0,5. Ферментацию осуществляли в 1 л среды при 30°C в ферментаторе Braun Biotech. Анаэробные условия поддерживали постоянным барботированием 0,2 л/мин газообразного азота. pH 5,5 поддерживали с помощью 3 М КОН. Скорость перемешивания составила 200 об/мин.
Характерный результат эксперимента для двух штаммов в присутствии и в отсутствие HMF показан на Фиг.2.
Оба штамма демонстрировали более медленные рост, превращение глюкозы и продукцию этанола, когда в среду добавляли HMF. Однако штамм ТМВ3206, сверхэкспрессирующий мутантный ген ADH1, преобразовывал HMF намного быстрее, чем контрольный штамм ТМВ3280. Вследствие этого потребление глюкозы начиналось раньше в ТМВ3206 и полного превращения глюкозы в этанол, биомассу и другие второстепенные продукты достигали за 35 часов (Фиг.2Г). Напротив, контрольному штамму ТМВ3280 потребовалось более 45 часов, чтобы достичь подобного профиля (Фиг.2Б).
ПРИМЕР 3
Сайт-направленный мутагенез NADH-зависимой HMF-редуктазы
Плазмиду YEplacHXT-ADH1-mut, кодирующую мутантный ген ADH1 из ТМВ3000 (как описано в примере 1), а также плазмиду YEPlac-HXT, кодирующую нативный ген ADH1 из CEN.PK 113-5D (названную YEplacHXT-ADH1-nat), использовали для сайт-направленного мутагенеза, чтобы исследовать влияние различных мутаций аминокислот на NADH-зависимую фуран-редуктазную активность.
Сайт-направленный мутагенез осуществляли согласно следующему протоколу: ПЦР в две стадии использовали для создания гена ADH1 с выбранными заменами аминокислот. Разработали шесть праймеров для обратного мутагенеза мутантного гена ADH1 в положениях 110 (P110S), 117 (S117L) и 295 (C295Y) и четыре других праймера разработали для мутагенеза нативного гена ADH1 в положениях 110 (S110P) и 295 (Y295C) (таблица 3). Также создали два праймера для амплификации гена ADH1 (мутантного или нет): смысловой праймер ADH1 (5'-GGGGGGATCCATGTCTATCCCAGAAACTC-3'), несущий сайт BamHI, и антисмысловой праймер ADH1 (5'-CTTTAGATCTTTATTTAGAAGTGTCAACAACG-3'), несущий сайт Bg/II. Для обратной мутации гена ADH1 из ТМВ3000 плазмиду YEplacHXT-ADH1-25-mut использовали в качестве матрицы. Для мутации нативного гена ADH1 плазмиду YEplacHXT-ADH1-nat использовали в качестве матрицы.
В первом цикле ПЦР амплифицировали два фрагмента для каждой направленной мутации: один фрагмент, используя смысловой праймер ADH1 и антисмысловой праймер, соответствующие направленной мутации, и другой фрагмент, используя антисмысловой праймер ADH1 и смысловые праймеры, соответствующие направленной мутации. ДНК-полимеразу Pwo (Roche Diagnostics AB, Bromma, Sweden) использовали для амплификации при следующих условиях: первичная денатурация при 94°C в течение 5 минут, затем 30 циклов, состоящих из стадии денатурации при 94°C в течение 30 секунд, стадии отжига при 55°C в течение 30 секунд и стадии элонгации при 72°C в течение 1 минуты. Два амплифицированных фрагмента очищали и использовали во втором цикле ПЦР вместе со смысловым и антисмысловым праймерами ADH1, ДНК-полимеразой Pwo при следующих условиях: первичная денатурация при 94°C в течение 5 минут, затем 30 циклов, состоящих из стадии денатурации при 94°C в течение 30 секунд, стадии отжига при 55°C в течение 30 секунд и стадии элонгации при 72°C в течение 1 минуты. Амплифицированный фрагмент очищали и расщепляли с помощью BamHI и Bg/II (Fermentas, Vilnius, Lithuania). Расщепленный ПЦР-фрагмент лигировали с дважды расщепленной YEplacHXT, используя ДНК-лигазу Т4 (Fermentas, Vilnius, Lithuania). Продукт лигирования использовали для трансформации клеток Eschehchia coli DH5α согласно Inoue et al. (1990. High efficiency transformation of Escherichia coli with plasmids. Gene 96: 23-28). Трансформированные клетки отбирали на среде LB с 50 мкг/мл ампициллина и проверяли правильность мутации и последовательности с помощью рестрикционного анализа и секвенирования ДНК. Множественные мутации выполняли последовательно.
Плазмиду, несущую отдельные или комбинированные мутации в гене ADH1, вводили в штамм CEN.PK 113-5D с помощью трансформации литий-ацетатным способом (Gietz et al., 1992, Improved method for high efficiency transformation of intact yeast cells. Nucleic Acids Research 20:1425). Селекцию осуществляли на чашках SD без урацила. Полученные штаммы выращивали аэробно при 30°C в 25 мл среды SD без урацила (в 250 мл колбе с перегородкой).
Экстракты клеток готовили путем сбора клеток при OD620 7,5-8,5, ресуспендирования клеток в растворе Y-PER (Pierce, Rockford, IL, USA) 1 мл/0,6 г клеток, встряхивания при комнатной температуре в течение 50 минут. Надосадочную жидкость собирали после центрифугирования в течение 20 минут при 15000 g. In vitro NADH-зависимую фуран-редуктазную активность измеряли путем последующего окисления NADH при длине волны 340 нм на спектрофотометре Hitachi U-2000 (Hitachi, Tokyo, Japan). Образец состоял из 10 мМ фуральдегида (HMF или фурфурол), 200 мкМ NADH и экстракта клеток в 100 мМ фосфатного буфера (pH 6,7) при 30°C. Результаты представлены в таблице 4.
ПРИМЕР 4
Ферментация лигноцеллюлозного гидролизата штаммом, сверхэкспрессирующим NADH-зависимую HMF-редуктазу
Штаммы ТМВ3280 (контрольный) и ТМВ3206 (сверхэкспрессирующий мутантный ген ADH1 из ТМВ3000) сравнивали по анаэробной ферментации недетоксифицированного гидролизата в основном из древесины ели в способе двухступенчатого гидролиза в разбавленной кислоте, используя серную кислоту в качестве катализатора (Larsson et al. Development of a Saccharomyces cerevisiae strain with enhanced resistance to phenolic fermentation inhibitors in lignocellulose hydrolysates by heterologous expression of laccase. Appl Environ Microbiol 2001, 67: 1163-1170).
Эксперименты осуществляли путем добавления одной порции лигноцеллюлозного гидролизата с предварительно доведенным с помощью 6 М NaOH pH 5 к растущим в экспоненциальной фазе клеткам в пробе. Конкретнее, клетки предварительно выращивали в 300 мл качалочных колбах при 160 об/мин и 30°C в течение 24 часов. Объем жидкости в предварительных культурах составлял 100 мл с концентрацией глюкозы 15 г/л. Последующую ферментацию запускали с помощью 6 мл предварительной культуры и осуществляли в ферментаторах Belach BR 0.5 (Belach Bioteknik 5 AB, Solna, Sweden) при 30°C и скорости перемешивания 600 об/мин. Исходный рабочий объем составил 300 мл, и pH поддерживали постоянно на уровне 5,0 путем добавления 0,75 NaOH. Ферментатор постоянно барботировали газообразным азотом 300 мл/мин (содержащим менее 5 млн-1 кислорода), чтобы создать анаэробные условия, контролируемые массовым расходомером (Bronkhurst Hi-Тес, Ruurlo, The Netherlands).
Исходные концентрации компонентов среды были в 2,67 раза больше по сравнению с посевными культурами, чтобы компенсировать разведение. Эксперименты начинали с выращивания клеток на 30 г глюкозы в 300 мл среды для выращивания. Затем осуществляли единичное добавление 300 мл гидролизата, когда концентрация биомассы достигла приблизительно 4 г/л, за которой следили с помощью измерений OD620. За скоростью выделения углекислого газа следили в режиме реального времени путем измерения концентраций углекислого газа и кислорода в газе, выходящем из ферментатора, с помощью газового анализатора СР460 (Belach Bioteknik AB, Solna, Sweden). Газовый анализатор калибровали, используя газ, содержащий 20% кислорода и 5% углекислого газа. Концентрацию клеток определяли по измерениям оптической плотности при длине волны 610 нм, и измерения массы в сухом состоянии проводили по повторным выборкам 10 мл, которые центрифугировали, промывали дистиллированной водой и сушили в течение 24 часов при 105°C. Образцы для измерения метаболитов немедленно центрифугировали, фильтровали через 0,2 мкм фильтры и хранили при -20°C до проведения анализов. Концентрации глюкозы, маннозы, галактозы, HMF и фурфурола измеряли на колонке Aminex HPX-87P (Bio-Rad, USA) при 85°C, элюировали сверхчистой водой при скорости потока 0,6 мл/мин. Концентрации этанола измеряли на колонке Aminex HPX-87P (Bio-Rad, USA) при 60°C. Используемый элюент представлял собой 5 мМ H2SO4 при скорости потока 0,6 мл/мин. Соединения определяли с помощью детектора показателя преломления, кроме HMF и фурфурола, которые определяли с помощью УФ-детектора (210 нм).
Результаты представлены на Фиг.3. ТМВ3206 мог преобразовывать фурфурол и HMF приблизительно за 5 часов, тогда как в эксперименте с контрольным штаммом ТМВ3280 HMF по-прежнему находился в среде 20 часов после добавления гидролизата. ТМВ3206, который имел в 5 раз более высокую скорость поглощения HMF, показал более-менее постоянную CER (Фиг.3Б), в то время как CER постепенно снижалась у контрольного штамма (Фиг.3А). Специфичная выработка этанола также была ниже для контрольного штамма по сравнению с ТМВ3206, сверхэкспрессирующим мутантный ген ADH1.
ПРИМЕР 5
Изменение баланса кофактора и образования продукта в штамме, экспрессирующем NADH-зависимую HMF-редуктазу и ксилозный путь Pichia stipitis
Сверхэкспрессия в S.cerevisiae TMB3320
Плазмиду YEplac-HXT-ADH1-S110P-Y295C, кодирующую мутантный ген ADH1 (как описано в таблице 4), а также соответствующую пустую плазмиду YEplacHXT (Karhumaa et al., 2005, Investigation of limiting metabolic steps in the utilization of xylose by recombinant Saccharomyces cerevisiae using metabolic engineering. Yeast. 22(5): 359-368) использовали для трансформации потребляющего ксилозу штамма S.cerevisiae TMB3320, получая штаммы ТМВ3291 и ТМВ3290, соответственно. TMB3320 получили путем трансформации ТМВ3043 (leu2, ura3) плазмидой Yiplac128-XRXDH, которая регулирует экспрессию ксилоза-редуктазы и ксилит-дегидрогеназы Pichia stipitis. Как ожидали, NADH-зависимую HMF-редуктазную активность обнаружили только в штамме, несущем мутантный ген ADH1 (данные не показаны).
Изменения баланса кофактора в анаэробной непрерывной культуре с HMF
Штаммы ТМВ3291 и ТМВ3290 предварительно выращивали в течение ночи при 30°C и 200 об/мин в 1 л качалочных колбах с 100 мл питательной среды определенного состава (Verduyn et al., 1992. Effect of benzole acid on metabolic fluxes in yeasts: a continuous-culture study on the regulation of respiration and alcoholic fermentation. Yeast, 8, 501-517), дополненной 20 г/л глюкозы. Ту же питательную среду определенного состава, дополненную эргостерином (0,075 г/л) и Tween 80 (0,84 г/л), с HMF (2 г/л) или без него использовали в последующих экспериментах с непрерывной культурой. Исходная OD620 в ферментаторе составляла 0,2. Ферментацию осуществляли в 400 мл среды при 30°C в ферментаторах Belach Biotech со скоростью разведения 0,06 ч-1 и 0,12 ч-1. Анаэробные условия поддерживали путем постоянного барботирования 0,2 л/мин газообразного азота. Скорость перемешивания составляла 600 об/мин, и pH 5,5 поддерживали с помощью 0,75 М NaOH.
Концентрацию клеток определяли измерениями массы в сухом состоянии из повторных выборок. Для получения образцов массы в сухом состоянии 5 мл суспензии клеток фильтровали в вакууме через предварительно взвешенные фильтры Gelman (d=47 мм Supor-450, 0,45 мкм). Фильтры промывали водой и сушили. Образцы для анализа концентраций метаболитов отбирали при равновесном состоянии культуры, то есть выработка углекислого газа оставалась постоянной после приблизительно 5 изменений объема культуры. Образцы фильтровали через 0,2 мкм фильтры. Концентрации глюкозы, ксилозы, ксилита, глицерина, ацетата, этанола и HMF разделяли на колонке Aminex HPX-87H (Bio-Rad, Hercules, California) при 65°C. Подвижная фаза представляла собой 5 мМ H2SO4 со скоростью потока 0,6 мл/мин. Все соединения определяли с помощью детектора показателя преломления кроме HMF, который определяли с помощью УФ-детектора при длине волны 210 нм.
Поглощение глюкозы и ксилозы и образование продукта были сходными у обоих штаммов, когда в питательную среду не добавляли HMF. Напротив, при добавлении к питательной среде HMF происходили значительные изменения. Выработка ксилита уменьшалась для штамма ТМВ3291 (ADH1-S110P-Y295C), в то время как она увеличивалась для штамма ТМВ3290 (контрольный). Выработка глицерина была на 50% меньше для штамма ADH1-S110P-Y295C, в то время как она не менялась в контрольном штамме. Более того, выработка биомассы штаммом ADH1-S110P-Y295C увеличилась в присутствии HMF. Выработка ацетата увеличилась в обоих штаммах. При наивысшей скорости разбавления (0,12 ч-1) поглощение ксилозы уменьшилось на 50% для контрольного штамма, но не изменилось в штамме ADH1-S110P-Y295C. Более того, половина добавленного в питательную среду HMF все еще присутствовала в ферментаторе с контрольным штаммом, но HMF был полностью преобразован в ферментаторе со штаммом ADH1-S110P-Y295C. Выработка этанола в обоих штаммах не изменилась при добавлении HMF и была одинаковой для обоих штаммов.
Эксперимент подтвердил, что увеличенное in vivo NADH-зависимое восстановление HMF изменяет баланс кофактора и, следовательно, распределение побочных продуктов в штаммах S.cerevisiae.
ПРИМЕР 6
Улучшенное продуцирование этанола в штамме, экспрессирующем NADH-зависимую HMF-редуктазу и ксилозный путь Pichia stipitis
Анаэробное восстановление HMF in vivo
Посевные культуры штаммов ТМВ3290 (контрольный) и ТМВ3291 (сверхэкспрессирующий мутантный ген ADH1-S110P-Y295C) выращивали в течение ночи при 30°C и 150 об/мин в 1 л качалочных колбах с 100 мл дважды концентрированной минеральной питательной среды определенного состава (как описано в Verduyn et al, 1992, Effect of benzoic acid on metabolic fluxes in yeasts: a continuous-culture study on the regulation of respiration and alcoholic fermentation. Yeast 8, 501-517), дополненной 40 г/л глюкозы и 200 мл/л фталатного буфера (10,2 г/л КН фталата, 2,2 г/л КОН). Ту же питательную среду определенного состава с HMF (2 г/л) или без него использовали в последующей серии экспериментов. Среду дополняли 20 г/л глюкозы, 50 г/л ксилозы, эргостерином (0,075 г/л) и Tween 80 (0,84 г/л). Исходная OD620 в периодической ферментации составила 0,5. Ферментацию осуществляли в 1 л среды при 30°C в ферментаторах Braun Biotech. pH 5,5 поддерживали с помощью 3 М КОН. Скорость перемешивания составила 200 об/мин.
Показательные результаты экспериментов для двух штаммов в присутствии и в отсутствие HMF показаны на Фиг.4
Потребление глюкозы и ксилозы и образование продукта было сходным для обоих штаммов, когда в среду не добавляли HMF (данные не показаны). В присутствии HMF контрольный штамм (ТМВ3290) потреблял глюкозу и ксилозу медленнее, чем штамм ADH1-S110P-Y295C (ТМВ3291) (Фиг.4). Более того, к концу ферментации штамм ADH1-S110P-Y295C употребил в два раза больше ксилозы, чем контрольный штамм, что привело к большей конечной концентрации этанола. Результаты подтверждают, что преобразование HMF быстрее в штамме ADH1-S110P-Y295C (Фиг.4).
ПРИМЕР 7
Кинетическая характеристика ADH1 мутантов
Плазмиды, несущие отдельные или комбинированные мутации в гене ADH1, созданные сайт-направленным мутагенезом в примере 3, вводили в штамм BY4741 с помощью трансформации литий-ацетатным способом (Gietz et al., 1992, Improved method for high efficiency transformation of intact yeast cells. Nucleic Acids Research 20:1425). Отбор проводили на чашках SD без урацила. Полученные штаммы и соответствующие мутации обобщены в таблице 5.
Клетки выращивали аэробно при 30°C в 25 мл среды SD без урацила (в 250 мл колбе с перегородкой). Клеточные экстракты готовили, собирая клетки при OD620 7,5-8,5, ресуспендируя клетки в растворе Y-PER (Pierce, Rockford, IL, USA) 1 мл/0,6 г клеток, встряхивая при комнатной температуре в течение 50 минут. Надосадочную жидкость собирали после центрифугирования в течение 20 минут при 15000 g.
In vitro NADH-зависимую фуран-редуктазную активность измеряли путем последующего окисления NADH при длине волны 340 нм на спектрофотометре Hitachi U-2000 (Hitachi, Tokyo, Japan). NADH-зависимое восстановление альдегидов определяли (Wahlbom and Hahn-Hägerdal, 2002) с помощью 200 мкМ NADH. Кинетику восстановления определяли для ацетальдегида (интервал 500 мкМ - 100 мМ), фурфурола (100 мкМ - 20 мМ) и HMF (500 мкМ - 20 мМ). Для нативного Adh1 использовали фурфурол до 40 мМ.
Моделирование выполняли по уравнению Михаэлиса-Ментен с добавлением константы ингибирования субстрата: V=(Vmax·[S])/(Km+[S]+[S]2/Ki), V - скорость реакции, Vmax - максимальная скорость реакции, Km - константа сродства, Ki - константа ингибирования субстрата и [S] - концентрация субстрата. Оценку значений параметров проводили по методу наименьших квадратов, используя функцию решающей программы в Microsoft® Excel 2002. Параметры HMF-активности штамма BY474-ADH1-S117L оценивали без константы ингибирования субстрата, благодаря четкому различию модели ингибирования субстрата и фактических данных.
Результаты суммированы в таблице 6. Повышенные значения Km и Vmax разных мутантов ясно продемонстрировали важность мутации Y295C для восстановления HMF и фурфурола. Сравнение кинетических параметров штаммов BY474-ADH1-S117L и BY4741-ADH1-110P-L117S-295C показало, что L в положении 117 полезен, поскольку он значительно увеличивает активность по отношению к обоим фуральдегидам.
| название | год | авторы | номер документа |
|---|---|---|---|
| ФЕРМЕНТАТИВНОЕ ПОЛУЧЕНИЕ 1-БУТАНОЛА | 2006 |
|
RU2429295C2 |
| СПОСОБ МИКРОБИОЛОГИЧЕСКОГО СИНТЕЗА ЦЕЛЕВОГО СЕКРЕТИРУЕМОГО БЕЛКА В ДРОЖЖАХ Saccharomyces cerevisiae | 2012 |
|
RU2502805C1 |
| СПОСОБ ПОЛУЧЕНИЯ 2,4-ДИГИДРОКСИБУТИРАТА | 2013 |
|
RU2645260C2 |
| Способ получения 2,4-дигидроксимасляной кислоты | 2011 |
|
RU2626531C2 |
| МОЛЕКУЛА ДНК ДЛЯ ЭКСПРЕССИИ, СТИМУЛИРУЕМОЙ СОЛЯМИ ЖЕЛЧИ ЛИПАЗЫ (BSSL) | 1996 |
|
RU2157847C2 |
| ПОЛИПЕПТИД, ОБЛАДАЮЩИЙ АКТИВНОСТЬЮ D-ЛАКТАТДЕГИДРОГЕНАЗЫ, ПОЛИНУКЛЕОТИД, КОДИРУЮЩИЙ ЭТОТ ПОЛИПЕПТИД, И СПОСОБ ПОЛУЧЕНИЯ D-МОЛОЧНОЙ КИСЛОТЫ | 2010 |
|
RU2553563C2 |
| МИКРООРГАНИЗМ, ЭКСПРЕССИРУЮЩИЙ КСИЛОЗОИЗОМЕРАЗУ | 2009 |
|
RU2553537C2 |
| ФЕРМЕНТАТИВНОЕ ПОЛУЧЕНИЕ ЧЕТЫРЕХУГЛЕРОДНЫХ СПИРТОВ | 2006 |
|
RU2394913C2 |
| ГИБРИДНЫЙ БЕЛОК, ОБЛАДАЮЩИЙ ПРОЛОНГИРОВАННЫМ ДЕЙСТВИЕМ, НА ОСНОВЕ РЕКОМБИНАНТНОГО ИНТЕРФЕРОНА АЛЬФА-2 ЧЕЛОВЕКА (ВАРИАНТЫ), СПОСОБ ЕГО ПОЛУЧЕНИЯ И ШТАММ Saccharomyces cerevisiae ДЛЯ ОСУЩЕСТВЛЕНИЯ ЭТОГО СПОСОБА (ВАРИАНТЫ) | 2013 |
|
RU2515913C1 |
| ВАРИАНТ ГЛУТАМАТ-ЦИСТЕИНЛИГАЗЫ И СПОСОБ ПОЛУЧЕНИЯ ГЛУТАТИОНА С ЕГО ПРИМЕНЕНИЕМ | 2021 |
|
RU2809326C1 |



Изобретение относится к области биохимии. Представлен выделенный полипептид, имеющий NADH-зависимую HMF-редуктазную активность, где указанный полипептид на 80% гомологичен аминокислотной последовательностью SEQ ID NO: 2, имеющейся в описании, который имеет замены в положениях, соответствующих положениям a) Y295C, б) Y295C и S117L, в) Y295C и S110P, или г) Y295C и S110P и S117L в SEQ ID NO: 2. Описаны нуклеиновая кислота, кодирующая указанный полипептид, экспрессионный вектор, содержащий указанную нуклеиновую кислоту, и клетка-хозяин, содержащая указанный вектор и предназначенная для экспрессии полипептида с NADH-зависимой HMF-редуктазной активностью. Предложено применение указанных полипептида, нуклеиновой кислоты, вектора или клетки-хозяина в производстве крупнотоннажных химикатов из лигноцеллюлозного сырья и фурансодержащего материала. Изобретение позволяет получить NADH-зависимую HMF-редуктазу с увеличенной специфичной активностью. 5 н. и 13 з.п. ф-лы, 6 табл., 4 ил., 7 пр.
1. Выделенный полипептид, имеющий NADH-зависимую HMF-редуктазную активность (NADH - никотинамид-аденин-динуклеотид, HMF - 5-гидроксиметилфурфурол), где указанный полипептид демонстрирует 80%-ную гомологию с аминокислотной последовательностью, показанной в SEQ ID NO: 2, и который отличается от SEQ ID NO: 2 тем, что имеет замены(у)
a) Y295C,
б) Y295C и S117L,
в) Y295C и S110P, или
г) Y295C и S110P и S117L,
где положения аминокислот в полипептидной последовательности соответствуют их положениям в SEQ ID NO: 2.
2. Выделенный полипептид по п.1, где указанный полипептид отличается от SEQ ID NO: 2 тем, что имеет замену Y295C.
3. Выделенный полипептид по п.1, где указанный полипептид отличается от SEQ ID NO: 2 тем, что имеет замены Y295C и S117L.
4. Выделенный полипептид по п.1, где указанный полипептид отличается от SEQ ID NO: 2 тем, что имеет замены Y295C и S110P.
5. Выделенный полипептид по п.1, где указанный полипептид отличается от SEQ ID NO: 2 тем, что имеет замены S117L и Y295C и S110P.
6. Выделенный полипептид по любому из пп.1-5, где указанный полипептид имеет по меньшей мере 90%-ную гомологию с SEQ ID NO: 2.
7. Выделенный полипептид по п.6, где указанный полипептид имеет по меньшей мере 95%-ную гомологию с SEQ ID NO: 2.
8. Выделенный полипептид по п.7, где указанный полипептид имеет по меньшей мере 98%-ную гомологию с SEQ ID NO: 2.
9. Выделенный полипептид по п.8, где указанный полипептид имеет по меньшей мере 99%-ную гомологию с SEQ ID NO: 2.
10. Нуклеиновая кислота, кодирующая полипептид по любому из пп.1-9.
11. Вектор для экспрессии полипептида по любому из пп.1-9, содержащий нуклеотидную последовательность по п.10.
12. Клетка-хозяин для экспрессии полипептида по любому из пп.1-9, выбранная из группы, состоящей из бактерий, дрожжей и грибов, содержащая нуклеотидную последовательность по п.10 или вектор по п.11.
13. Клетка-хозяин по п.12, где указанные бактерии выбраны из группы, состоящей из Escherichia coli, Klebsiella sp. и Zymomonas mobilis, и указанные грибы представляют собой нитчатые грибы.
14. Клетка-хозяин по п.12, где указанная дрожжевая клетка выбрана из группы Saccharomyces sp., Schizosaccharomyces sp., Kluyveromyces sp., Pichia sp., Hansenula sp., Candida sp. и Yarrowia sp.
15. Клетка-хозяин по п.14, где указанная клетка-хозяин выбрана из группы, состоящей из Saccharomyces cerevisiae, Saccharomyces bayanus и Saccharomyces carlsbergensis.
16. Клетка-хозяин по п.15, где указанная клетка-хозяин представляет собой Saccharomyces cerevisiae.
17. Применение полипептида, имеющего NADH-зависимую HMF-редуктазную активность, по любому из пп.1-9, нуклеотидной последовательности по п.10, вектора по п.11 или клетки-хозяина по любому из пп.12-16 в производстве крупнотоннажных химикатов из лигноцеллюлозного сырья и фурансодержащего материала.
18. Применение по п.17, где указанный крупнотоннажный химикат выбран из группы, состоящей из этанола, бутанола, лактата, сукцината, глицерина, маннита, L-аскорбиновой кислоты, ксилита и газообразного водорода.
| WO 2005111214 A1, 24.11.2005 | |||
| ANNELI PETERSSON et al | |||
| Кипятильник для воды | 1921 |
|
SU5A1 |
| Пломбировальные щипцы | 1923 |
|
SU2006A1 |
| Способ восстановления хромовой кислоты, в частности для получения хромовых квасцов | 1921 |
|
SU7A1 |
| J.ZALDIVAR "Fuel ethanol production from lignocellulose: a challenge for metabolic engineering and process integration", Appl | |||

