Изобретение относится к области селекции растений, а именно маркерсопутствующему отбору. Создание на основании генетических карт специфических молекулярно-генетических маркеров к отдельным хозяйственно-ценным генам дает возможность проводить маркерный отбор (MAS - Marker Assisted Selection). Маркерный, или маркерсопутствующий отбор - сравнительно новый подход в селекции растений. Его применение позволяет избежать трудоемкой или дорогостоящей оценки агрономически ценного признака, по которому ведется селекция.
В частности, изобретение относится к главному методологическому подходу в подобного рода исследованиях - применению молекулярных маркеров для идентификации желательных признаков в селекционном материале.
Изобретение может использоваться для геном-специфической идентификации генов FAE1.1, отвечающих за синтез эруковой кислоты у рапса. Использование подобного подхода позволяет независимо проанализировать аллели, локализованные в А геноме рапса, и установить их гомо- либо гетерозиготное состояние, сокращая таким образом количество поколений и, соответственно, количество лет, необходимое для выравнивания данного признака в селекционном материале.
Для селекции рапса одной из существенных проблем является аллополиплоидный характер генома. В формировании селекционно ценного признака участвуют оба родительских генома, причем аллели, определяющие признак, могут быть практически идентичны. В этой связи возможным решением проблемы выявления доли участия определенного генома в развитии селекционно цепного признака может быть разработка геном-специфических ДНК-маркеров.
Уровень содержания эруковой кислоты у рапса определяется двумя аллелями, локализованными в А- и С-геномах [1,2]. Функция гена FAE1 в синтезе эруковой кислоты у рапса была доказана путем генетической трансформации низкоэруковых сортов [3]. Были охарактеризованы оба гомолога гена FAE1 (Bn-FAE1.1 и Bn-FAE1.2). Они проявляют сходство нуклеотидов на 99,4%. Выявлено, что к формированию растений рапса типа «канола» с практически нулевым содержапием эруковой кислоты приводит замена одной пары оснований гена FAE1.1 генома А, результатом которой является замена аминокислоты серии на фенилаланин [4].
Аналогами патентуемого способа являются методы, основанные на биохимическом определении жирнокислотного состава масла в семенах рапса. Подобные традиционные методы оценки признаков осуществимы только на определенной стадии развития растения - после обмолота семян. При наличии в гетерозиготном состоянии нежелательных генов, определяющих синтез эруковой кислоты, распространение этих генов происходит при опылении и фенотипически проявляется в следующем поколении.
Прототипом по отношению к заявляемому способу являются специфические ДНК-маркеры к генам FAE1.1, основанные на детекции однонуклеотидной замены в геноме А рапса. Суть такого подхода заключается в определении однонуклеотидной замены с помощью метода SNaPshot[5].
Недостатком подобного анализа являются:
1) Высокая себестоимость, а также необходимость использования специализированного дорогостоящего оборудования (анализ возможен только на генетических анализаторах Applied Biosystems с помощью набора ABI SNaPshot kit).
2) Возможность появления рекомбинаций между аллелями А и С геномов рапса, что приводит к появлению SNP замены в 282 положении в С геноме и возникновению делеции в геноме А рапса [6,7]. В связи с этим использование прямой детекции мутаций без учета геном-специфичности может привести к некорректному анализу селекционного материала.
Задачей предлагаемого изобретения являлось создание способа геном-специфической и аллель-специфической идентификации гена FAE1.1, контролирующего синтез эруковой кислоты в рапсовом масле. Сохраняя достоинства метода-прототипа, как-то прямую детекцию однонуклеотидной замены в геноме и определение гомо- и гетерозиготного состояния локуса, предлагаемый способ лишен его недостатков и позволяет избежать использования специализированного оборудования и реактивов, и ниже прототипа по себестоимости.
Поставленная задача решается путем разработки системы двухступенчатой геном-специфичной идентификации аллелей гена FAE1.1, отвечающего за синтез эруковой кислоты в геноме А рапса. Данные маркеры позволяют достоверно различать гены FAE1, принадлежащие С-геному рапса, полученному от капусты, от генов FAE1, принадлежащих А-геному рапса, источником которого является сурепица.
На первом этапе для специфической амплификации фрагмента генома А рапса, содержащего ген FAE1.1, используется 50-100 нг тотальной ДНК рапса. Реакционная смесь объемом 25 мкл содержит 200 мкМ каждого dATP, dCTP, dGTP и dTTP, 1,5 мМ
MgCl2, 1,5 единицы Taq-полимеразы в инкубационном буфере и по 0,25 мкМ праймеров следующего нуклеотидного состава:
ЕrА F 5'-CTCATTCCCGAGAAACACTGA-3'
ЕrА R 5'-CTACGATCTCCAGGCTTGTT-3'
Условия проведения реакции: денатурация - 4 мин при 94°С; циклы 2 - 35: 30 сек при 94°С, 30 сек при 55°С и 2 мин при 72°С; цикл 36-10 мин при 72°С.
Продукты реакции разделяются электрофорезом в 2% агарозном геле с добавлением этидиум бромида.
В результате амплификации разработанных для предлагаемого способа праймеров с тотальной ДНК рапса может быть получен ПЦР-фрагмент размером 980 пар нуклеотидов, который используется для последующей идентификации аллелей гена FAE1.1.
Геном-специфичность данных праймеров доказывается их избирательной амплификацией с ДНК капусты огородной и сурепицы. Как показано на фигуре 1, маркер к А-геному размером 980 пар нуклеотидов амплифицируется с ДНК рапса и сурепицы (2 и 3 дорожки), с ДНК капусты амплификация отсутствует (1 дорожка).
Полученный ПЦР-фрагмент используется в качестве ДНК-матрицы для последующей ген-специфичной амплификации. Для этого 2 мкл ПЦР-смеси, полученной в результате 1 этапа и содержащей 50-100 нг ПЦР-фрагмента генома А, добавляется в реакционную смесь, включающую 200 мкМ каждого dATP, dCTP, dGTP и dTTP, 1,5 мМ MgCl2, 1,5 единицы Taq-полимеразы в инкубационном буфере и по 0,25 мкМ праймеров следующего нуклеотидного состава:
SATF 5'-GCTCTTGTGGTGAGCACAGAGAACA-3'
SATR 5'-TCTACGATCTCCAGGCTTGTCG-3'
Условия проведения реакции: денатурация - 4 мин при 94°С; циклы 2 - 35: 30 сек при 94°С, 30 сек при 56°С и 30 сек при 72°С; цикл 36-10 мин при 72°С.
В результате амплификации разработанных для предлагаемого способа праймеров с ПЦР-фрагментом А генома рапса, амплифицированного на предыдущем этапе, могут быть получены ПЦР-фрагменты размером 135 пар нуклеотидов, содержащие однонуклеотидную замену в случае наличия мутантной аллели на расстоянии в 2 нуклеотида от 3'-конца SATR праймера.
Праймер SATR разработан таким образом, чтобы при наличии SNP-замены возникал искусственный сайт рестрикции для рестриктазы TaqI. Полученный ПЦР-фрагмент размером 135 нуклеотидов подвергается рестрикции при 65°С в течение 3 часов. Условия проведения реакции: 1X буфер TaqI:10 mM Tris-HCl (рН 8.0), 5 mМ MgCl2, 100 mМ NaCl и 0.1 mg/ml BSA, и 2 единицы TaqI в инкубационном буфере.
Продукты рестрикции разделяются в неденатурирующем 8%-ном полиакриламидном геле [8] и визуализируются с помощью этидиум бромида.
Как показано на фигуре 2, в результате рестрикции ампликона, полученного с помощью праймеров SAT, могут быть получены фрагменты следующего размера:
Гомозиготный ген FAE1.1 дикого типа - 135 п.н.
Гомозиготный мутантный аллель FAE1.1 - 113 п.н.
Гетерозигота по гену FAE1.1 - 135 и 113 п.н. Разработанные авторами геном-специфические ДНК-маркеры к генам FAE1, контролирующим синтез эруковой кислоты в семенах рапса, имеют ряд преимуществ:
- возможность прямой идентификации генов FAE1 в селекционном материале на ранней стадии развития растения, что позволяет в начале вегетации выбраковать нежелательный для селекционера материал и исключить проведение биохимического анализа жирнокислотного состава масла в селекционных образцах после уборки и обмолота семян. Тем самым экономятся время и средства, затрачиваемые на уход за посевами, уборку и послеуборочную обработку селекционного материала,
- независимое тестирование генов FAE1, расположенных в А и С геномах рапса, что позволяет вдвое сократить сроки создания сортов,
- низкая себестоимость анализа и возможность использования реактивов белорусского производства.
Предлагаемый способ идентификации может быть проведен на ранней стадии развития растения, что позволяет в начале вегетации выбраковать нежелательный для селекционера материал и исключить проведение биохимического анализа жирнокислотного состава масла в селекционных образцах после уборки и обмолота семян. Тем самым экономится время и средства, затрачиваемые на уход за посевами, уборку и послеуборочную обработку селекционного материала.

| название | год | авторы | номер документа |
|---|---|---|---|
| Способ диагностики аллельного состояния гена Vi4, контролирующего фиолетовую окраску зерна у ржи | 2021 |
|
RU2787110C1 |
| Молекулярно-генетический маркер типа CAPS у растений гороха посевного Pisum sativum L. и способ идентификации растений, гомозиготных по аллели, обуславливающей формирование повышенной специфичности по отношению к симбиотическим клубеньковым бактериям | 2023 |
|
RU2815453C1 |
| СПОСОБ ИДЕНТИФИКАЦИИ ПОЛИМОРФИЗМА rs1128446 ГЕНА ТИОРЕДОКСИНРЕДУКТАЗЫ-1 У ЧЕЛОВЕКА | 2011 |
|
RU2458146C1 |
| СПОСОБ ДИАГНОСТИКИ НАСЛЕДСТВЕННОЙ ПРЕДРАСПОЛОЖЕННОСТИ К ТРОМБОФИЛИИ | 2007 |
|
RU2352641C1 |
| CAPS-маркер для отбора гибридов ячменя, не накапливающих проантоцианидины в зерне | 2023 |
|
RU2809369C1 |
| СПОСОБ ИДЕНТИФИКАЦИИ ПОЛИМОРФИЗМА R287Q В 8 ЭКЗОНЕ ГЕНА ЦИТОЗОЛЬНОЙ ЭПОКСИДГИДРОЛАЗЫ У ЧЕЛОВЕКА | 2006 |
|
RU2346053C2 |
| СПОСОБ ОБНАРУЖЕНИЯ МУТАЦИИ А40Т В ГЕНЕ ВНЕКЛЕТОЧНОЙ СУПЕРОКСИДДИСМУТАЗЫ | 2006 |
|
RU2333964C1 |
| Способ оценки генетического потенциала овец породы манычский меринос на основе молекулярно-генетических маркеров | 2021 |
|
RU2776044C1 |
| Способ идентификации полиморфизмов Cys1079Gly и Cys1079Phe медь-транспортной АТФ-азы Вильсона | 2020 |
|
RU2756112C1 |
| ГЕН И ВАРИАЦИИ, СВЯЗАННЫЕ С ФЕНОТИПОМ BM1, МОЛЕКУЛЯРНЫЕ МАРКЕРЫ И ИХ ПРИМЕНЕНИЕ | 2012 |
|
RU2617958C2 |
Изобретение относится к области биотехнологии, а именно к способу идентификации генов FAE1.1, контролирующих содержание эруковой кислоты в семенах рапса. Способ включает последовательную геном-специфическую и аллель-специфическую ПЦР-реакцию. Проводят с помощью рестриктазы TaqI рестрикцию ампликона, полученного путем последовательной ПЦР с помощью прямого SATF 5'-GCTCTTGTGGTGAGCACAGAGAACA-3' и обратного SATR 5'-TCTACGATCTCCAGGCTTGTCG-3' праймеров. Получают фрагменты ДНК определенного размера. При этом гомозиготному состоянию гена FAE1.1 дикого типа соответствует фрагмент 135 п.н., гомозиготному мутантному аллелю FAE1.1 соответствует фрагмент 113 п.н., а гетерозиготе по гену FAE1.1 соответствуют фрагменты 135 и 113 п.н. Предложенное изобретение позволяет осуществлять геном-специфичную идентификацию генов FAE1.1, контролирующих содержание эруковой кислоты в масле семян рапса, на любой стадии развития растения, и позволяет исключить стадию биохимического анализа жирнокислотного состава масла. 2 ил., 1 пр.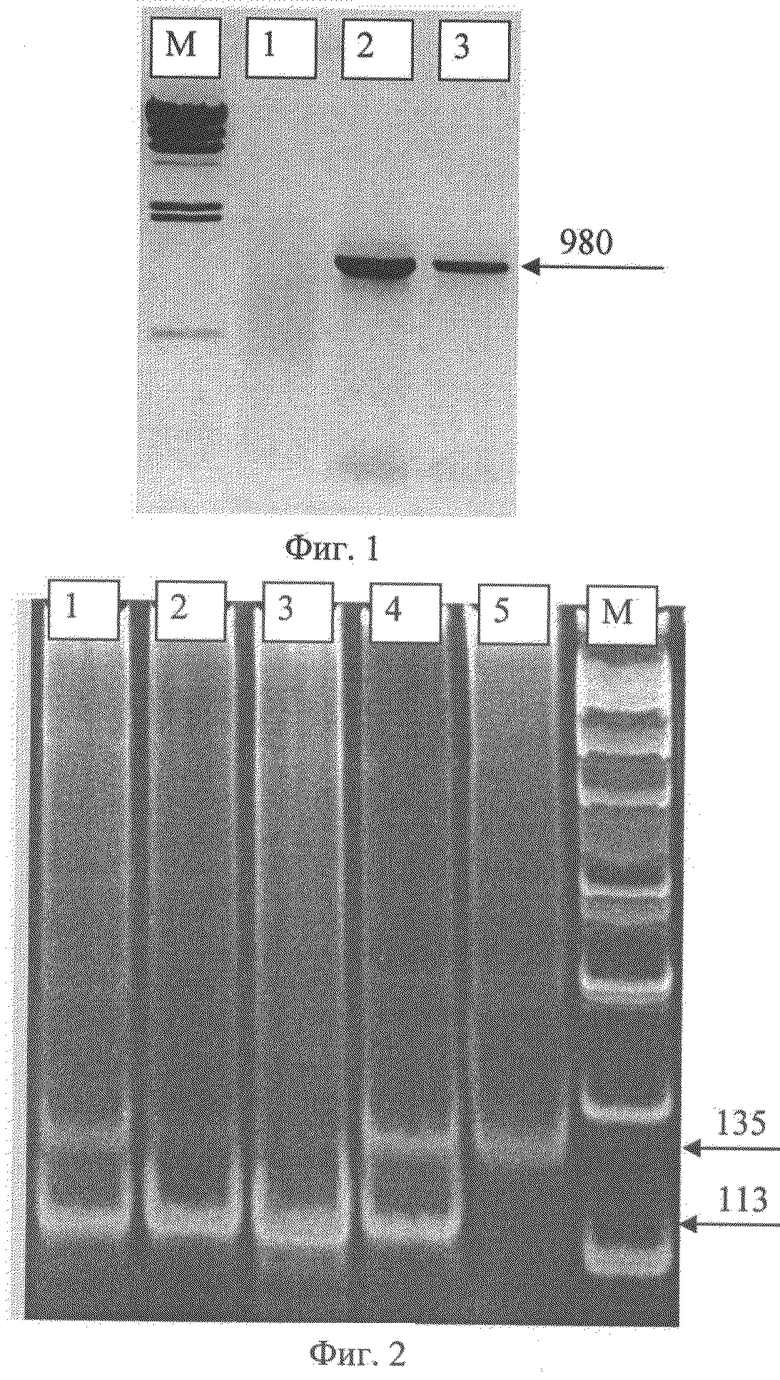
Способ идентификации генов FAE1.1, контролирующих содержание эруковой кислоты в семенах рапса, с помощью dCAPS-маркеров, включающий последовательную геном-специфическую и аллель-специфическую ПЦР-реакцию, отличающийся тем, что с помощью рестриктазы TaqI проводится рестрикция ампликона, полученного путем последовательной ПЦР с помощью прямого (SATF 5'-GCTCTTGTGGTGAGCACAGAGAACA-3') и обратного (SATR 5'-TCTACGATCTCCAGGCTTGTCG-3') праймеров, в результате чего получают фрагменты ДНК определенного размера, соответствующие гомозиготному состоянию гена FAE1.1 дикого типа (135 п.н.), гомозиготному мутантному аллелю FAE1.1 (113 п.н.) и гетерозиготе по гену FAE1.1 (135 и 113 п.н.).
| RAHMAN М | |||
| et al | |||
| High throughput genome-specific and gene-specific molecular markers for erucic acid genes in Brassica napus (L.) for marker-assisted selection in plant breeding, Theor Appi Genet, 2008, v.117, no.6, p.895-904 | |||
| US 0007745131 B2, 29.06.2010 | |||
| WANG N et al | |||
| Assessment ofFAEl polymorphisms in three Brassica species using EcoTILLING and |

