Изобретение относится к области молекулярной генетики растений, а именно к способу генотипирования растений ржи в отношении мутантной аллели vi4 и аллели дикого типа Vi4 гена транскрипционного фактора Мус-типа, влияющего на развитие признака окраски зерна в фиолетовый цвет. Изобретение может найти применение для нужд сельского хозяйства и растениеводства при диагностике селекционного материала ржи.
Молекулярная диагностика признака окраски зерна в фиолетовый цвет важна для селекции ржи, поскольку, в зависимости от целей создания сорта, необходим отбор растений с окрашенным или неокрашенным зерном. Для создания хлебопекарных сортов ржи необходим отбор с неокрашенным зерном, для сортов, предназначенных для создания функциональных продуктов здорового питания, необходим целенаправленный отбор форм с окрашенным зерном, поскольку такие зерна содержат большое количество окрашенных и неокрашенных флавоноидов, положительно влияющих на профилактику заболеваний человека. Возможность молекулярной идентификации доминантной или рецессивной аллели гена Vi4, который является транскрипционным фактором, контролирующим синтез флавоноидов в зернах ржи, сокращает время отбора необходимых растений, поскольку снимает необходимость получения взрослых растений с зернами. Аллели гена Vi4 отличаются наличием сайта для рестриктазы XbaI (TCTAGA) (доминантная аллель Vi4, приводящая к формированию окрашенного зерна) и отсутствием этого сайта (ТСТАТА) для рецессивной аллели vi4 - отсутствие окраски зерна. Аллели отличаются однонуклеотидной заменой (SNP) G/T.
Известен способ определения аллельного состояния гена Vi4, влияющего на окрашенность зерна по морфологии растения (фенотипу) [1], сущность его состоит оценке степени окрашенности зерна. Основным недостатком этого способа является трудоемкость, связанная с необходимостью выращивания растений до момента проявления признака. Кроме того, этим методом не отличить доминантную гомозиготу от гетерозиготы, а степень проявления признака зависит от климатических условий.
Методы молекулярного ДНК-маркирования, в частности CAPS- маркеры позволяют четко описать аллельное состояние гена на любой стадии развития растения за короткий срок. Это возможно благодаря амплификации на матрице ДНК различных генотипов их полиморфных фрагментов методом ПЦР и их аллель-специфическому разрезанию рестриктазой XbaI, сайт узнавания которой присутствует у аллели дикого типа и отсутствует у мутантной аллели (SNP G/T, позиция 443 в ПЦР-продукте). В результате рестрикции образуются фрагменты, которые можно различить по их электрофоретической подвижности.
На настоящий момент на основе CAPS-маркеров разработано огромное количество диагностических систем различного назначения, в том числе для нужд сельского хозяйства и растениеводства [3-6], которые можно считать аналогами нашего изобретения. Однако, все описанные наборы относятся к различным конкретным видам растений и конкретным аллелям их генов. Диагностические системы для описания полиморфизма гена Vi4 - транскрипционного фактора, контролирующего синтеза флавоноидов, из литературных и других источников информации не известны.
Заявителю известно, что работа над созданием подобных диагностических систем строится обычно следующим образом.
1. Выбор фрагментов, соответствующих полиморфному участку исследуемого гена, с помощью открытых и коммерческих баз данных последовательностей ДНК растений.
2. Подбор последовательностей олигонуклеотидов, используемых для проведения ПЦР-реакции (2 праймера) на основании выбранного участка генома с помощью специального программного обеспечения. На данном этапе работа заключается в создании выравнивания нуклеотидных последовательностей и выборе консервативных участков последовательности ДНК, расположенных с разных сторон от SNP на расстоянии порядка 100-500 нуклеотидных пар.
3 Выбор рестриктазы, сайт которой перекрывает зону SNP.
4 Изготовление праймеров.
5. Доказательство с помощью практических экспериментов пригодности подобранных последовательностей для конкретных целей (например, для выявления конкретных аллелей гена).
Наиболее близким способом по достигаемому техническому результату является способ диагностики аллельного состояния гена Myb113-подобного транскрипционного фактора, контролирующего синтез антоцианов (однонуклеотидная инсерция Т1227 приводящая к образованию стоп-кодона) [3], принятый в качестве прототипа. Сущность его состоит в проведении ПЦР с использованием специфических праймеров комплиментарных участкам Myb113-подобного гена. Ключевым элементом заявляемого способа является использование рестриктазы AccI. В пределах амплифицированного фрагмента аллели дикого типа, приводящего к формировании окраски плодов, отсутствует сайт рестрикции. После его обработки ферментом AccI сохраняется интактный ПЦР-продукт 589 п.н. Указанная выше мутация находится в 393 положении продукта амплификации и приводит к образовании сайта рестрикции, в результате чего у неокрашенных форм образуются фрагменты 393 и 195 п.н. Их идентифицируют после электрофоретического разделения относительно маркера молекулярного веса. Таким образом, по количеству и размеру фрагментов судят о присутствии аллелей в каждом из исследуемых генотипов.
Недостатком известного способа, принятого в качестве прототипа, является невозможность его применения для диагностики других аллелей у других культур, в том числе, ржи.
Технический результат изобретения состоит в возможности применения диагностикума для описания аллельного состояния гена Vi4, контролирующего синтез флавоноидов в зернах ржи.
Указанный технический результат достигается тем, что в способе диагностики присутствия в генотипе растения мутантной аллели (vi4) и аллели дикого типа Vi4 гена Vi4, заключающемся в выделении из растений ДНК и проведении на ней полимеразной цепной реакции с последующей рестрикцией и разделением фрагментов электрофоретически, в соответствии с изобретением после выделения из образцов ДНК в концентрации 50-100 нг/мкл на ее основе составляют реакционную смесь, содержащую праймеры

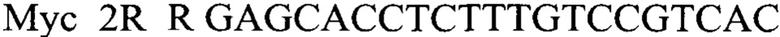 .
.
1х буфер для полимеразы, катионы Mg 2+ в концентрации 0,1-0,3 мМ, нуклеотиды в концентрации 2-10 мкМ, Taq полимеразу (или другую термостабильную полимеразу, обладающую 5' экзонуклеазной активностью), 0,1-0,3 ед.
Полимеразную цепную реакцию проводят с температурой отжига праймеров 60°С при 42 циклах, далее к амплификату добавляют рестриктазы и инкубируют смесь при 37°С в течение часа, а затем разделяют продукты электрофоретически. Кроме этого, указанный технический результат достигается тем, что в качестве термостабильной полимеразы с активностью 0,1-0,3 ед. на 1 мкл смеси берут Taq. Вместе с тем указанный технический результат достигается тем, что в качестве термостабильной полимеразы с активностью 0,1-0,3 ед. на 1 мкл смеси берут Tth.
Указанный технический результат достигается тем, что в качестве эндонуклеаз рестрикции используют ферменты RsaI, XbaI с активностью 0,05 ед. на 1 мкл смеси каждого фермента.
Сущность изобретения состоит в том, что для диагностики аллельного состояния гена Vi4 ржи проводят ПЦР с праймерами к гену Vi4, а затем при помощи рестриктаз RsaI, XbaI выявляют полиморфизм в нем.
Например, при сравнении уровня техники:
Известна идентификация ДНК-маркеров генов биосинтеза антоцианов у перца и баклажанов [3].
При схожей технологии детекции в данной статье описывается детекция полиморфизма другого гена у другой культуры сходным методом.
Существенными признаками изобретения являются:
из анализируемого образца выделяют ДНК чистоты, достаточной для проведения ферментативных реакций, после этого составляют реакционную смесь, содержащую, по меньшей мере: праймеры

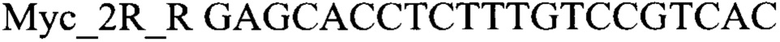
1х буфер для полимеразы, катионы Mg 2+ в концентрации 0,1-0,3 мМ, нуклеотиды в концентрации 2-10 мкМ, Taq полимеразу (или другую термостабильную полимеразу, обладающую 5' экзонуклеазной активностью), 0,1-0,3 ед.
Полимеразную цепную реакцию проводят в амплификаторе (термоциклире) при температуре отжига праймеров 60°С при 42 циклах, далее проводят рестрикцию амплификата ферментами RsaI, XbaI в течение 1 часа при 37°С.
Наличие после электрофоретического разделения фрагментов размером 400 н. и 500 н. свидетельствует о присутствии в генотипе только аллели дикого типа, отсутствие фрагментов размером 400 н. и 500 н. свидетельствует о присутствии в генотипе только аллели мутантного типа.
Реализация заявленного способа.
Для обнаружения заявленным способом аллелей гена Vi4 требуется:
1. Выделить ДНК из образца СТАБ-методом. Для этого нужно гомогенезировать образец в СТАВ-буфере (NaCl 1,4М, Трис HCl (рН 8) ОДМ, ЭДТА (рН 8) 20 мМ, СТАВ 2% m/v), инкубировать полученную смесь 1 ч при 56°С, 40-60 мин экстрагировать хлороформом, разделить фракции центрифугированием, верхнюю (водную), содержащую ДНК фракцию переосадить этанолом, ДНК растворить в воде.
2. Приготовить реакционную смесь для ПЦР, которая должна содержать 1х буфер для полимеразы, катионы Mg 2+ в концентрации 0,1-0,3 мМ, нуклеотиды в концентрации 2-10 мкМ, Taq полимеразу (или другую термостабильную полимеразу, обладающую 5' экзонуклеазной активностью), 0,1-0,3 ед. активности на 1 мкл смеси, ДНК матрицу исследуемого образца 10-100 нг/мкл, праймеры, следующего состава:

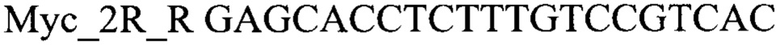
3. Провести полимеразную цепную реакцию с температурой отжига праймеров 60°С при 40 циклах. Пример программы: 95°С - 2 мин, 42 цикла (95°С - 30 с, 60°С - 30 с, 72°С - 40 с), 72°С - 2 мин.
4. Обработать рестриктазами RsaI, XbaI в течение 1 часа при 37°С.
5. Провести электрофоретическое разделение фрагментов в присутствии маркера молекулярного веса (размерный ряд фрагментов от 100 н. до 1000 н., определить их размеры.
При обнаружении фрагментов размером 400 и 500 н. делают вывод о присутствии в генотипе только аллели дикого типа, отсутствие фрагментов размером 400 и 500 н. свидетельствует о присутствии в генотипе только мутантной аллели.
Заявленный способ апробирован на лабораторной базе Санкт-Петербургского государственного университета в рамках НЦМУ «Агротехнологии будущего» и поясняется конкретным примером.
. Пример конкретной реализации.
ДНК выделяли из двух линий ржи (дикого типа и мутанта по гену Vi4) / Соответственно их генотипы по исследуемому признаку Vi4Vi4 и vi4vi4)
Ткани проростков исследуемых линий ржи перетирали в ступке в СТАВ - буфере, инкубировали полученную смесь 1 час при 56°С, 40 минут экстрагировали хлороформом, разделяли фракции центрифугированием, верхнюю (водную), содержащую ДНК фракцию переосадили этанолом, ДНК растворили в воде, определяли концентрацию спектрофотометрически. Далее в работе использовали ДНК в концентрации 10-100 нг/мкл.
Постановка ПЦР
В ходе апробации заявленного способа была приготовлена реакционная смесь для ПЦР, содержавшая 1х буфер для полимеразы, катионы Mg 2+ в концентрации 0,2 мМ, нуклеотиды в концентрации 2 мкМ, Taq полимеразу, 0,2 ед. активности на 1 мкл смеси, ДНК матрицу исследуемого образца 10-100 нг/мкл, праймеры, следующего состава:
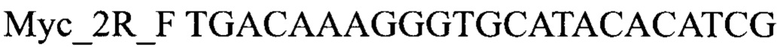

Полимеразную цепную реакцию проводили по программе:
95°С - 2 мин, 42 цикла (95°С - 30 с, 60°С - 30 с, 72°С - 40 с), 72°С - 2 мин. в термоциклере, Терцик (ДНК-Технология, Россия)
В ходе ПЦР на ДНУ обоих генотипов были наработаны фрагменты размером 1670 п.н. (Фиг. 1). На дорожки геля нанесены: 1 - маркер молекулярного веса ДНК с размерами фрагментов 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1500, 2000, 3000, 4000, 5000, 6000 п.н., 2- пустая дорожка, 3,4 продукты ПЦР с праймерами Myc2R на ДНК мутантной линии ржи (vi4vi4); 5, 6 - продукты ПЦР с праймерами Myc2R на ДНК нормальной линии ржи (Vi4Vi4).
Далее проводили рестрикцию этих фрагментов рестриктазами RsaI, XbaI в течение 1 часа при 37°С, а далее проводили электрофорез продуктов рестрикции в агарозном геле. Полученные результаты рестрикции представлены на Фиг. 2., где изображены паттерны фрагментов, характерные для образцов гомозиготных по мутантной аллели (дорожки 3, 4) и гомозигот по аллели дикого типа (дорожки 5, 6). Маркер молекулярного веса и порядок нанесения полностью соответствуют таковому на фиг. 1.
Во всех образцах мы наблюдаем фрагменты размером 300 н.п.и менее. Они образуются в результате работы короткощепящей рестриктазы RsaI и свидетельствуют об успешном прохождении рестрикции в каждой из рестрикционных смесей. В то же время, у растений с доминантной аллелью Vi4 присутствует сайт для рестриктазы XbaI, что приводит к появлению 2 полос размером 400 и 500 н. (дорожки 5, 6), рецессивные аллели связаны с мутацией в сайте рестрикции, что приводит к отсутствию фрагментов размеров 400 и 500 н. (дорожки 3, 4).
Таким образом, можно сделать вывод о успешной работе диагностической системы при использовании ДНК заявленных концентраций.
Технико-экономическая эффективность изобретения состоит в том, что предлагаемый новый способ диагностики растительного материала на предмет аллельного состояния гена Vi4 (SNP G/T, позиция 443) направлен на решение проблемы быстрой диагностики растительного материала (растения и продукты их переработки), т.е., на эффективное и оперативное выявление наличия в исследуемых материалах аллелей гена Vi4. Заявленный способ по достигаемому техническому результату отличается от известных в данной области аналогов: позволяет применить технологи caps-маркирования для исследования аллелей гена Vi4 ржи, контролирующего антоциановую пигментацию в зернах.
Источники информации
1. Lykholay AN, Vladimirov IA, Andreeva EA, Smirnov VG, Voylokov AV. [Genetics of anthocyaninless rye]. Genetika. 2014 Oct; 50(10): 1245-9. Russian. PMID: 25720256.
2. Falentin C, Brégeon M, Lucas MO, Renard M (2007) Genetic markers for high oleic content in plants. International Patent Application Publication, WO 2007/138444.
3. Babak O, Nikitinskaya T, Nekrashevich N, Yatsevich K, Kilchevsky A. Identification of DNA Markers of Anthocyanin Biosynthesis Disorders Based on the Polymorphism of Anthocyanin 1 Tomato Ortholog Genes in Pepper and Eggplant. Crop Breed Genet Genom. 2020; 2(3): e200011. https://doi.org/10.20900/cbgg20200011 (прототип).
4. Hu C.Y., Tsai Y.Z., Lin S.F. Development of STS and CAPS markers for variety identification and genetic diversity analysis of tea germplasm in Taiwan. Botanical Studies. 2014; 55(1): 12. DOI: 10.1186/1999-3110-55-12.
5. Bang H., Kim S., Park S.O., Yooa K.S., Patil B.S. Development of a codominant CAPS marker linked to the Ms locus controlling fertility restoration in onion (Allium сера L.). Sci. Hortic. 2013; 153: 42-49. DOI: 10.1016/j.scienta.2013.01.020.
6. Грушецкая З.E., Лемеш B.A., Мозгова Г.В., Пилюк Я.Э., Бакановская А.В. "Способ идентификации генов FAE1.1, контролирующих содержание эруковой кислоты в масле семян рапса, с помощью dCAPS-маркеров", 2012. Патент РФ №2486254.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ОПРЕДЕЛЕНИЯ ПОЛИМОРФИЗМА ГЕНЕТИЧЕСКИХ МАРКЕРОВ МОЛОЧНОЙ ПРОДУКТИВНОСТИ КРУПНОГО РОГАТОГО СКОТА | 2022 |
|
RU2782833C1 |
| CAPS-маркер для отбора гибридов ячменя, не накапливающих проантоцианидины в зерне | 2023 |
|
RU2809369C1 |
| СПОСОБ ИДЕНТИФИКАЦИИ ПОЛИМОРФИЗМА R287Q В 8 ЭКЗОНЕ ГЕНА ЦИТОЗОЛЬНОЙ ЭПОКСИДГИДРОЛАЗЫ У ЧЕЛОВЕКА | 2006 |
|
RU2346053C2 |
| СПОСОБ ГЕНОТИПИРОВАНИЯ ПОЛИМОРФИЗМА rs17522918 ГЕНА ПЕРОКСИРЕДОКСИНА-1 У ЧЕЛОВЕКА | 2011 |
|
RU2458145C1 |
| СПОСОБ ДИАГНОСТИКИ ПРЕДРАСПОЛОЖЕННОСТИ К ГЕМОХРОМАТОЗУ | 1999 |
|
RU2144566C1 |
| СПОСОБ ИДЕНТИФИКАЦИИ ПОЛИМОРФИЗМА rs1128446 ГЕНА ТИОРЕДОКСИНРЕДУКТАЗЫ-1 У ЧЕЛОВЕКА | 2011 |
|
RU2458146C1 |
| ДНК-маркер для селекции гибридов сорго на основе цитоплазматической мужской стерильности А1-типа | 2021 |
|
RU2792135C1 |
| СПОСОБ ГЕНОТИПИРОВАНИЯ ПОЛИМОРФИЗМА rs2551715 ГЕНА ГЛУТАТИОНРЕДУКТАЗЫ У ЧЕЛОВЕКА | 2013 |
|
RU2549688C1 |
| СПОСОБ ИДЕНТИФИКАЦИИ ГЕНОВ FAE1.1, КОНТРОЛИРУЮЩИХ СОДЕРЖАНИЕ ЭРУКОВОЙ КИСЛОТЫ В МАСЛЕ СЕМЯН РАПСА, С ПОМОЩЬЮ dCAPS-МАРКЕРОВ | 2012 |
|
RU2486254C1 |
| БИОЛОГИЧЕСКИЙ ДНК МАРКЕР ДЛЯ ОПРЕДЕЛЕНИЯ СОРТОВ КАРТОФЕЛЯ, НАБОР И СПОСОБ СОРТОВОЙ ИДЕНТИФИКАЦИИ КАРТОФЕЛЯ | 2009 |
|
RU2413774C1 |

Изобретение относится к биотехнологии и может найти применение для нужд сельского хозяйства и растениеводства при диагностике аллельного состояния гена Vi4 ржи, контролирующего антоциановую пигментацию зерновок. Кроме этого изобретение может быть использовано в пищевой промышленности при анализе сырья растительного происхождения. Заявленный способ позволяет существенно сократить время диагностирования в сроки до одного рабочего дня и осуществлять одному исследователю параллельно анализ сотен испытуемых образцов, а также позволяет проводить раннюю диагностику признака окраски зерна в фиолетовый цвет до его проявления на уровне фенотипа; он является дешевым, достоверным и экологически чистым. 2 ил., 1 пр.
Способ диагностики аллельного состояния гена Vi4 ржи, отвечающего за антоциановую пигментацию зерновок, заключающийся в выделении из растительных тканей ржи ДНК и проведение на ней полимеразной цепной реакции с последующей рестрикцией продуктов и анализом ее результатов методом электрофореза, отличающийся тем, что после выделения из образцов ДНК в концентрации 10-100 нг/мкл на основе ее составляют реакционную смесь, содержащую праймеры Myc_2R_F TGACAAAGGGTGCATACACATCG
Myc_2R_R GAGCACCTCTTTGTCCGTCAC, буфер для полимеразы, катионы Mg 2+ в концентрации 0,1-0,3 mM, нуклеотиды в концентрации 2-10 мкМ, термостабильную полимеразу 0,1-0,3 ед. активности на микролитр смеси, ПЦР проводят с температурой отжига праймеров 60°С при 42 циклах, далее обрабатывают амплифицированный фрагмент рестриктазами RsaI и XbaI, с активностью 0,05 ед. на 1 мкл смеси каждого фермента в 1х буфере, после чего разделяют фрагменты электрофоретически и определяют их длины, относительно маркера молекулярного веса с размерным рядом от 100 нуклеотидов до 5000 нуклеотидов, при этом присутствие фрагментов длиной 400 и 500 нуклеотидов свидетельствует о наличии аллели Vi4, отсутствие фрагментов длиной 400 и 500 нуклеотидов свидетельствует о наличии аллели vi4.
| Lap B., Rai M., Tyagi W | |||
| Playing with colours: genetics and regulatory mechanisms for anthocyanin pathway in cereals //Biotechnology and Genetic Engineering Reviews | |||
| Способ регенерирования сульфо-кислот, употребленных при гидролизе жиров | 1924 |
|
SU2021A1 |
| - Т | |||
| Пишущая машина | 1922 |
|
SU37A1 |
| - No | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| - С | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Zykin P | |||
| A | |||
| et al | |||
| Anthocyanin composition and content in rye plants with different grain color //Molecules | |||
| Способ получения цианистых соединений | 1924 |
|
SU2018A1 |
| - Т | |||
| Прибор для равномерного смешения зерна и одновременного отбирания нескольких одинаковых по объему проб | 1921 |
|
SU23A1 |
| - No | |||
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |
| - С | |||
| СОСТАВНАЯ РАМА ПОВОРОТНОГО КРУГА ИЛИ ПЕРЕДВИЖНОЙ ТЕЛЕЖКИ | 1924 |
|
SU948A1 |

