По данной заявке испрашивается приоритет по предварительной заявке на патент США №61/001602, поданной 2 ноября 2007 г, описание которой включено в данное описание путем ссылки.
Данная работа была поддержана грантом №1K23CA12007501A1 от Национальных Институтов Здоровья. Правительство США имеет определенные права на данное изобретение.
Область, к которой относится изобретение
Настоящее изобретение относится в целом к стимуляции иммунных ответов и, в частности, к применению угольной ангидразы IX (CA9) для стимуляции иммунного ответа, направленного против антигена.
Предшествующий уровень техники
Экспрессия CA9 связана с опухолевым ростом и коррелировалась с прогнозом при различных видах рака мозга, легких, молочной железы, шейки матки, почек, желудочно-кишечного тракта и головы и шеи (1-6). CA9 экспрессируется в ответ на снижение парциального давления кислорода (7). Считается, что это адаптивный ответ, который приводит к увеличенной доставке кислорода и питательных веществ к опухоли и может содействовать прогрессированию заболевания и, в конечном счете, метастазированию раковых клеток. Экспрессия СА9 увеличивается при многих солидных злокачественных опухолях, причем наибольшая экспрессия имеется в опухолевых клетках, непосредственно прилегающих к областям некроза (1, 7). СА9 также является маркером гипоксии в нормальных клетках и большинстве злокачественных опухолей. При всех злокачественных новообразованиях, где сообщалось о СА9 как о прогностическом маркере, за исключением прозрачно-клеточной почечноклеточной карциномы (RCC), увеличенная экспрессия CA9 прогнозировала худший прогноз (1-4, 6). Экспрессия в другой нормальной ткани ограничивается базальными клетками волосяных фолликулов, эпителием гонад, паутинным сплетением и некоторыми отделами слизистой оболочки желудочно-кишечного тракта (8). Однако при прозрачно-клеточной RCC экспрессия CA9 не регулируется парциальным давлением кислорода. CA9 присутствует более чем в 80% первичных и метастатических RCC, и СА9 присутствует в 95-100% прозрачно-клеточного варианта, наиболее распространенного гистологического типа RCC. CA9 не экспрессирован в нормальной почечной ткани (8, 9). Представляется, что для прозрачно-клеточной RCC СА9 является превосходным маркером, который способствует установлению диагноза, определению прогноза, прогнозированию реакции на лечение и служит в качестве мишени для лечения. В частности, высокая экспрессия CA9 является независимым прогностическим показателем более длительного специфичного для заболевания выживания у пациентов с метастатической RCC (5, 10) и улучшенного выживания у пациентов с локализованной RCC (11, 12). Однако, несмотря на широко распространенный интерес к CA9 и лучшее понимание молекулярного дефекта, ведущего к избыточной экспрессии при прозрачно-клеточной RCC, непонятен механизм, связывающий экспрессию CA9 с улучшенным прогнозом и реакцией на лечение у пациентов с RCC, в то же время в иных отношениях являющийся показателем неблагоприятного прогноза. Поэтому, существует потребность в разработке способов и композиций с использованием CA9 для применения в качестве терапевтического средства.
Сущность изобретения
Настоящее изобретение основано на неожиданно обнаруженной у CA9 функции шаперона, выполняемой подобно белку теплового шока (HSP). В частности, в данном описании раскрывается, что CA9, а также растворимая форма CA9, которая отделяется с поверхности определенных клеток и приблизительно на 4 кДа меньше, чем CA9 полной длины, может ингибировать вызванную нагреванием агрегацию белка и может содействовать сворачиванию белка. Однако авторы показывают, что в отличие от HSP, CA9 может образовывать иммуногенные комплексы с антигенами при комнатной температуре (например, приблизительно 20°C) и при температуре тела человека (например, 37°C). В данном описании также показано, что CA9 интернализуется представляющими антиген клетками и перерабатывается в первую очередь по протеосомальному пути, который считается важным для активации опосредованных клетками иммунных ответов. Кроме того, авторы продемонстрировали на мышиной модели меланомы, что способ по изобретению эффективен для стимуляции антиген-специфического противоопухолевого ответа, который является более выраженным, чем ответ, генерируемый антигеном, в отсутствие белка CA9. Соответственно, изобретение относится к способу стимуляции иммунного ответа на антиген, включающему введение композиции по изобретению индивидууму в количестве, эффективном для стимуляции иммунного ответа на антиген, где иммунный ответ на антиген является более выраженным, чем иммунный ответ, стимулированный антигеном, в отсутствие белка CA9. Композиции по изобретению содержат антиген и изолированный белок CA9. Изолированный белок CA9 и антиген предоставлены в комплексе, причем комплекс может представлять собой нековалентную связь между изолированным CA9 и антигеном или может представлять собой химически конъюгированный белок CA9 с антигеном, или может представлять собой слитый белок CA9/антигена. Иммунный ответ, стимулированный введением композиции, может быть профилактическим или терапевтическим и может включать гуморальный или клеточно-опосредованный иммунный ответ, или их комбинацию. Изобретение может использоваться для стимуляции иммунного ответа против широкого разнообразия пептидных и белковых антигенов, но белки теплового шока не включены в объем антигенов, используемых в композициях и способах по настоящему изобретению. В одном варианте осуществления антиген представляет собой опухолевый антиген.
Краткое описание фигур
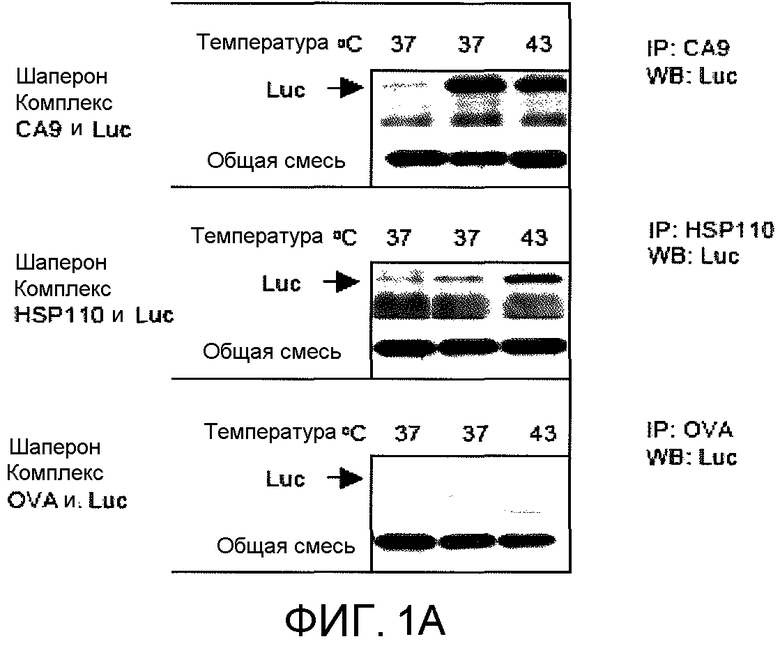
На фиг.1А в виде фотографии представлены результаты прерывистого анализа, использованного для того, чтобы показать, что CA9 способен связывать белки-клиенты. Люцифераза (Luc) служила в качестве белка-клиента, и CA9 образовывал комплекс с Luc при 37°C и 43°C. HSP110 служил в качестве положительного контроля и образовывал комплекс с люциферазой при 43°C, но не при 37°C. Овальбумин (OVA) служил в качестве отрицательного контроля и не обладал способностью выполнять функции шаперона. Полосы без шаперона (1ая полоса) были включены в качестве контролей. Иммунопреципитацию (IP) выполняли, используя антитела против CA9, HSP110 или OVA с последующим вестерн-блоттингом (WB) с антителом против люциферазы. На нижней панели WB выполняли с анти-люциферазой для подтверждения того, что прерывистый анализ OVA был успешным.
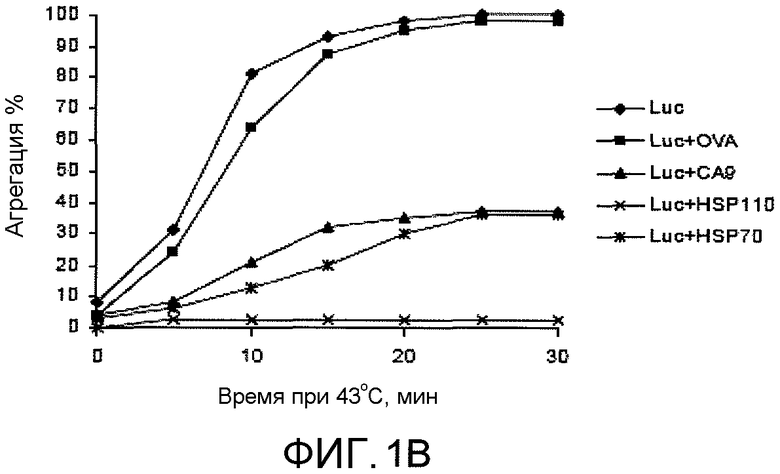
На фиг.1B графически представлены данные, демонстрирующие, что рекомбинантный CA9, так же как HSP70, эффективен в предотвращении агрегации люциферазы при 43°C. OVA служил в качестве контрольного белка. Мониторинг агрегации люциферазы в динамике во времени осуществляли измерением оптической плотности при 320 нм.
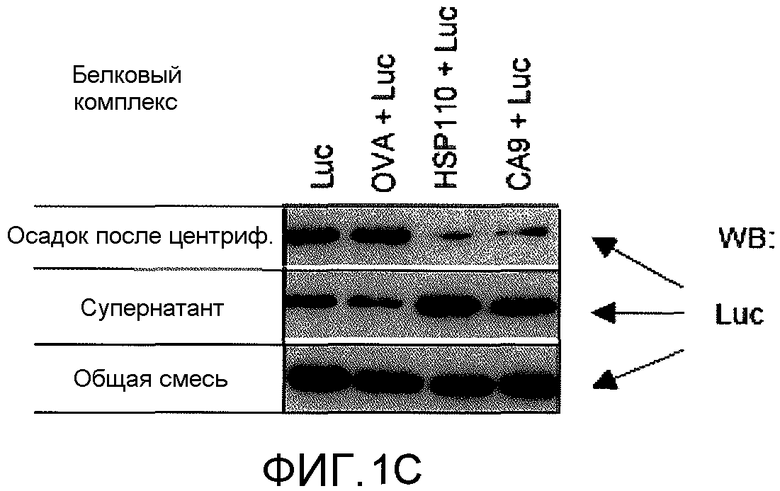
На фиг.1C в виде фотографии представлены результаты вестерн-блоттинга, демонстрирующие, что CA9 способен предотвращать агрегацию люциферазы и удерживать ее в растворе. Люциферазу нагревали до 43°C в течение 30 мин с белком шаперона (CA9, HSP110, OVA) или без него. Всю реакционную смесь центрифугировали при 16000×g в течение 15 мин для отделения супернатанта и осадка после центрифугирования. В контрольных ответах (полосы 1 и 2) большая часть люциферазы агрегировалась и была обнаружена в осадке после центрифугирования. CA9 и HSP110 были способны удерживать люциферазу в растворе при температуре теплового шока. Вся реакционная смесь служила в качестве контроля загрузки.
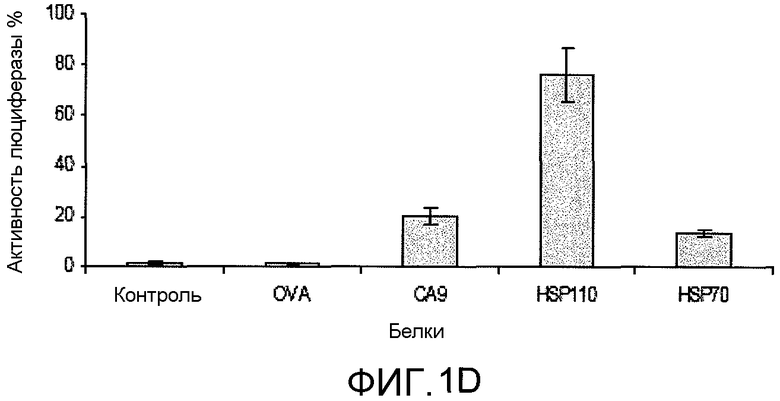
На фиг.1D представлено графическое изображение данных, демонстрирующих, что изолированный белок CA9 и классические белки теплового шока представляют собой белки, денатурированные шапероном, и обеспечивают возможность повторного сворачивания. Люциферазу денатурировали при 43°C в течение 30 мин в присутствии CA9, HSP110, HSP70 или OVA (отрицательный контроль). Добавляли лизат кроличьих ретикулоцитов (RRL), и повторное сворачивание оценивали мониторингом ферментной активности люциферазы. Молярное отношение белка шаперона к люциферазе составляло 20:1. Способность повторного сворачивания коррелировалась со способностью предотвращать агрегацию белков-клиентов; CA9 и HSP70 были одинаковы по их способности предотвращения агрегации и повторного сворачивания люциферазы. Для экспериментов, выполненных в трех повторах, представлена средняя величина±SEM (стандартная ошибка среднего).
На фиг.2A представлено графическое изображение данных, показывающих, что CA9 связывает DC (дендритные клетки) насыщаемым образом, что указывает на связывание, опосредованное рецепторами. Полученные из костного мозга DC (1×106/мл) инкубировали при 4°C с CA9, меченным FITC (флюоресцеин-изоцианатом) или BSA, меченным FITC, в течение 30 мин и дважды промывали 1% BSA/PBS (бычий сывороточный альбумин/забуференный фосфатом солевой раствор). Связывание CA9 оценивали измерением средней интенсивности флюоресценции (MFI) с использованием проточной цитометрии.
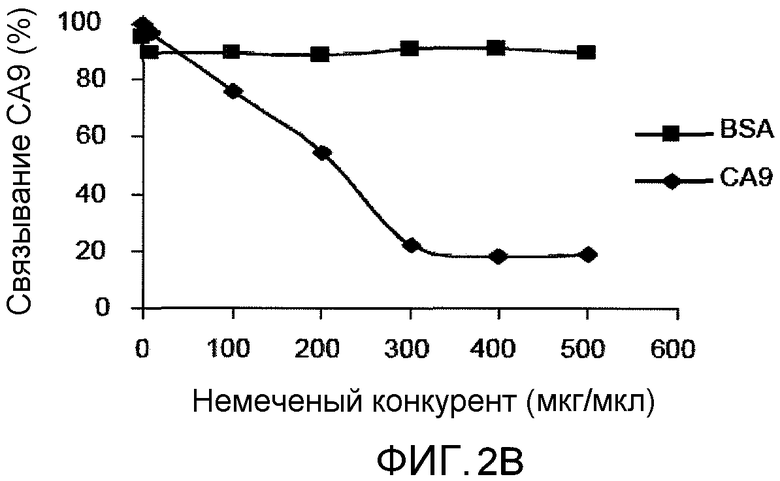
На фиг.2B представлено графическое изображение данных, показывающих конкурентный анализ связывания CA9 с DC. Связывание CA9 с DC ингибировалось немеченым CA9. DC, полученные из костного мозга (1×106/мл), инкубировали при 4°C с немеченым CA9 или BSA в указанных концентрациях, промывали и затем инкубировали с 200 мкг/мл CA9, меченного FITC.
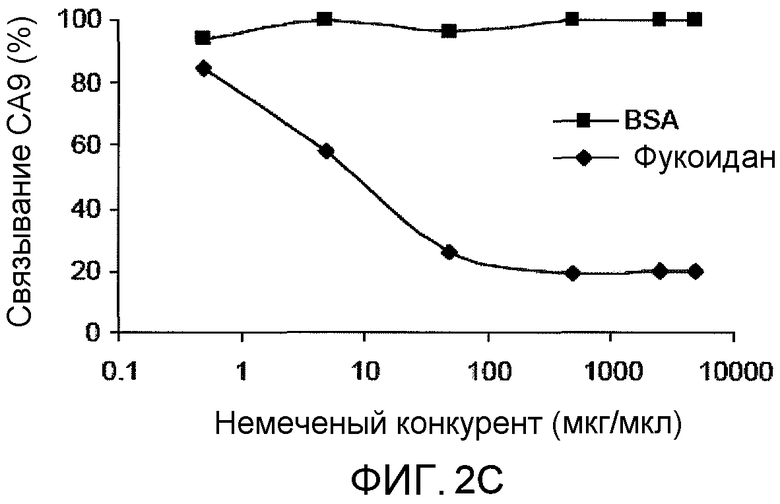
На фиг.2C представлено графическое изображение данных, показывающих, что связывание CA9 с DC ингибируется лигандом акцепторного рецептора. DC, полученные из костного мозга (1×106/мл), инкубировали при 4°C с фукоиданом или BSA в указанных концентрациях, промывали и затем инкубировали с 200 мкг/мл CA9, меченного FITC.
На фиг.2D представлено графическое изображение данных, показывающих, что связывание CA9 было уменьшено в DC из акцепторного рецептора А (SRA) от нокаутированных мышей (мышей SRA-KO), по сравнению с DC от мышей дикого типа (WT), указывая на то, что связывание CA9 частично опосредуется SRA. DC, полученные из костного мозга (1×106/мл), инкубировали при 4°C с BSA, CA9 или фукоиданом в концентрации 500 мкг/мл перед промыванием и затем инкубировали с 200 мкг/мл CA9, меченного FITC. «Усы» указывают SEM для экспериментов, выполненных в трех повторах.
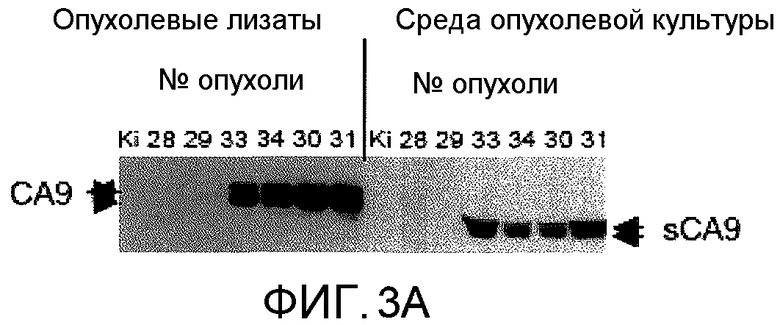
На фиг.3А представлено графическое изображение данных, показывающих, что опухолевые клетки сбрасывают растворимую форму CA9. Клеточные лизаты из человеческих почечных опухолей зондировали антителом против CA9, демонстрирующим экспрессию CA9 полной длины. Такие же опухоли, выращенные в кратковременных культурах, сбрасывали растворимую Форму CA9 (sCA9), которая имела размер, приблизительно на 4 кДа меньше, чем CA9 полной длины. Прозрачно-клеточные опухоли (опухоли 30-34) экспрессировали и сбрасывали CA9. Здоровые почки (Ki) и 2 папиллярные почечные опухоли (опухоли 28 и 29) не экспрессировали или не сбрасывали CA9.
На фиг.3B представлено графическое изображение данных, показывающих, что сброшенный CA9 обладает свойствами, подобно шаперону. SCA9 был так же эффективен как CA9 полной длины в ингибировании агрегации люциферазы при 43°C. Мониторинг агрегации люциферазы с течением времени проводили измерением оптической плотности при 320 нм.
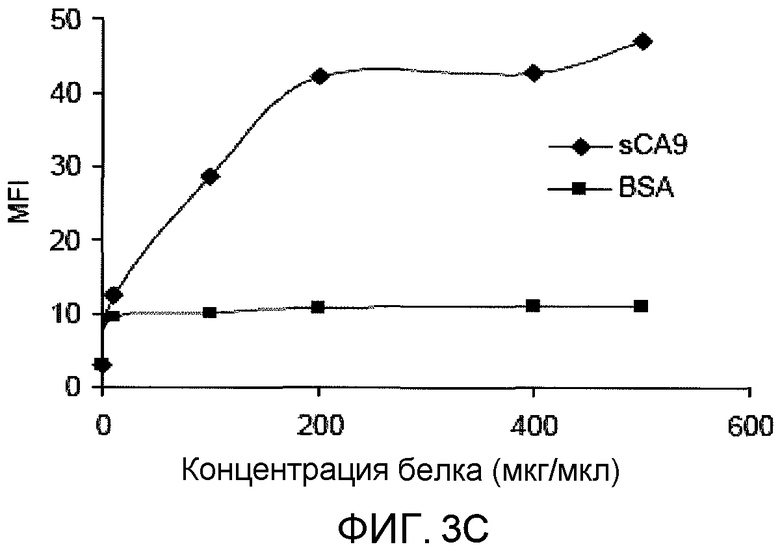
На фиг.3C представлено графическое изображение данных, показывающих, что растворимый CA9 связывает DC насыщаемым образом. Клетки DC, полученные из костного мозга (1×106/мл), инкубировали при 4°C с sCA9 или BSA, меченым FITC, в течение 30 мин и дважды промывали 1% BSA/PBS (слева). Как и с CA9, связывание sCA9 с DC ингибировалось немеченым CA9 и фукоиданом, свидетельствуя о том, что CA9 и sCA9 связывают акцепторные рецепторы на DC (справа). Клетки DC, полученные из костного мозга (1×106/мл), инкубировали при 4°C с BSA, CA9 или фукоиданом при 500 мкг/мл перед промыванием и инкубировали с 200 мкг/мл меченого FITC CA9. «Усы» указывают SEM для экспериментов, выполненных в трех повторах.
На фиг.3D представлено графическое изображение данных, показывающих, что растворимый CA9 содействует повторному сворачиванию денатурированного белка. Люцифераза денатурировалась при 43°C в течение 30 мин в присутствии CA9, sCA9 и HSP110. Добавляли лизат кроличьих ретикулоцитов, и повторное сворачивание оценивали мониторингом ферментной активности люциферазы. Молярное отношение белка шаперона к люциферазе составляло 20:1. CA9 и sCA9 были одинаковы в их способности повторно сворачивать люциферазу. Средняя величина±SEM представлены для экспериментов, выполненных в трех повторах.
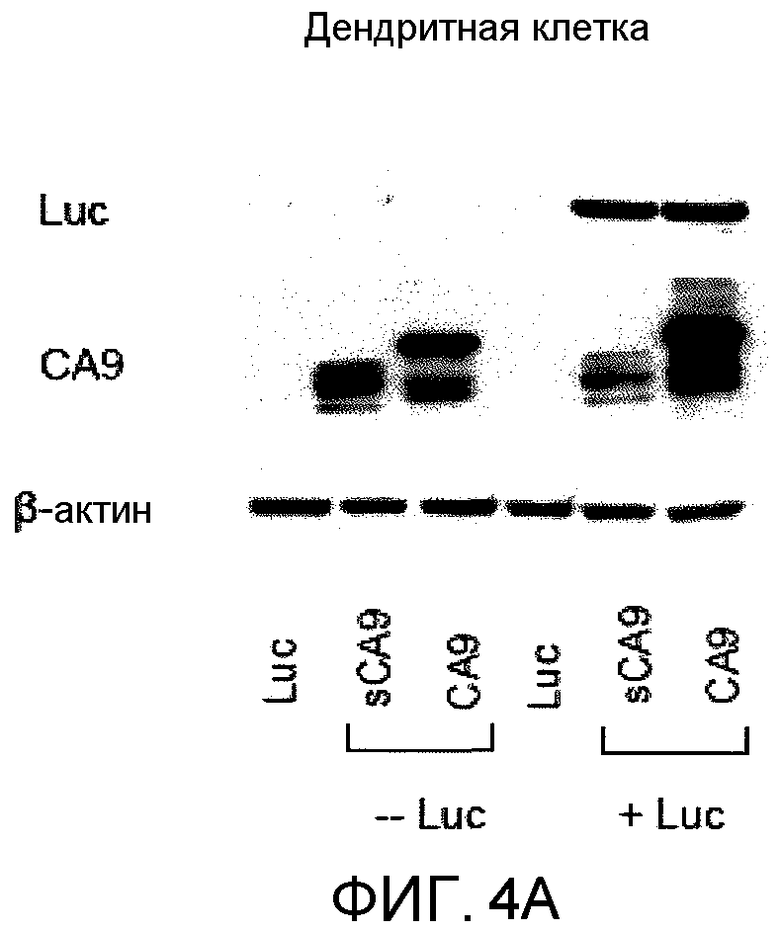
На фиг.4A представлено графическое изображение данных, показывающих, что и CA9, и сброшенный CA9 (sCA9) способны связывать и нести люциферазу к DC. CA9 и sCA9 образовывали комплексы с люциферазой при 37°C в течение 30 мин и инкубировали с DC, полученными из костного мозга при 4°C в течение 2 ч. CA9 и sCA9 без люциферазы и одну люциферазу также инкубировали с DC и служили в качестве контролей. DC дважды промывали для удаления несвязанного CA9, лизировали в буфере RIPA и зондировали вестерн-блоттингом антителами против люциферазы и против CA9.
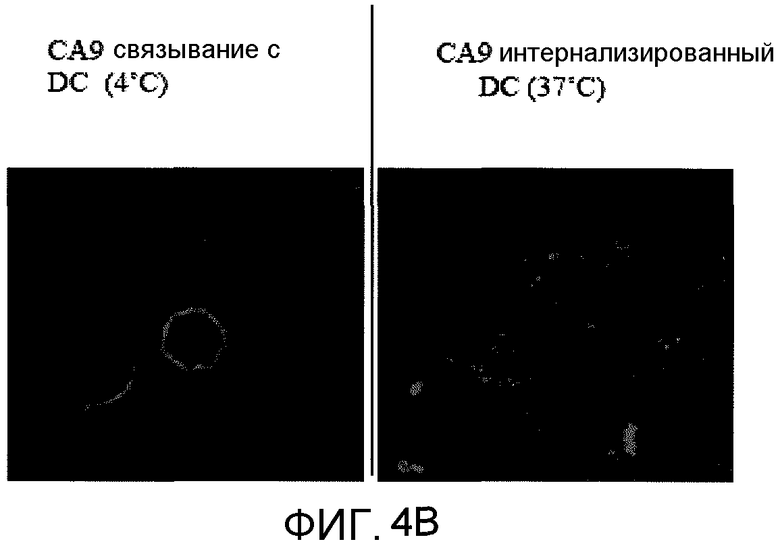
На фиг.4B представлена фотография, полученная при выполнении анализа с использованием конфокальной микроскопии, демонстрирующая, что CA9 способна связывать DC и быть интернализированным. В частности, конфокальная микроскопия показала CA9, меченный FITC, связанный с поверхностью полученных из костного мозга DC при 4°C. DC промывали для удаления несвязанного CA9 и инкубировали при 37°C в течение 2 часов, что приводило к интернализации CA9, меченного FITC. Проводили контрокрашивание ядра DAPI (диамидин-2-фенилиндолом).
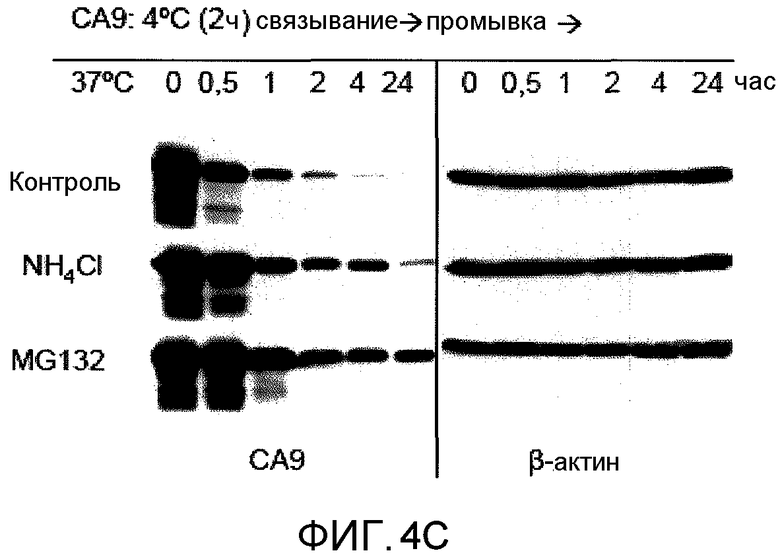
На фиг.4C представлена фотография, полученная при выполнении вестерн-блоттинга, показывающая, что DC способны интернализировать и перерабатывать CA9. CA9 давали возможность связывать DC в течение 2 ч при 4°C. DC дважды промывали и инкубировали в свежей среде при 37°C в течение указанного времени. CA9 клеточной поверхности промывали, и мониторинг внутриклеточного CA9 проводили вестерн-блоттингом для выявления CA9. β-актин служил в качестве контроля загрузки. В необработанной группе (контроль), обработка CA9 приводила к уменьшению внутриклеточного CA9, выявленного вестерн-блоттингом. Предварительная обработка DC MGl32 (ингибитор протеосомы) или NH4Cl (ингибитор лизосомы) в течение 30 мин при 37°C показала, что CA9 обрабатывался обоими путями; однако протеосомальный путь был преобладающим.
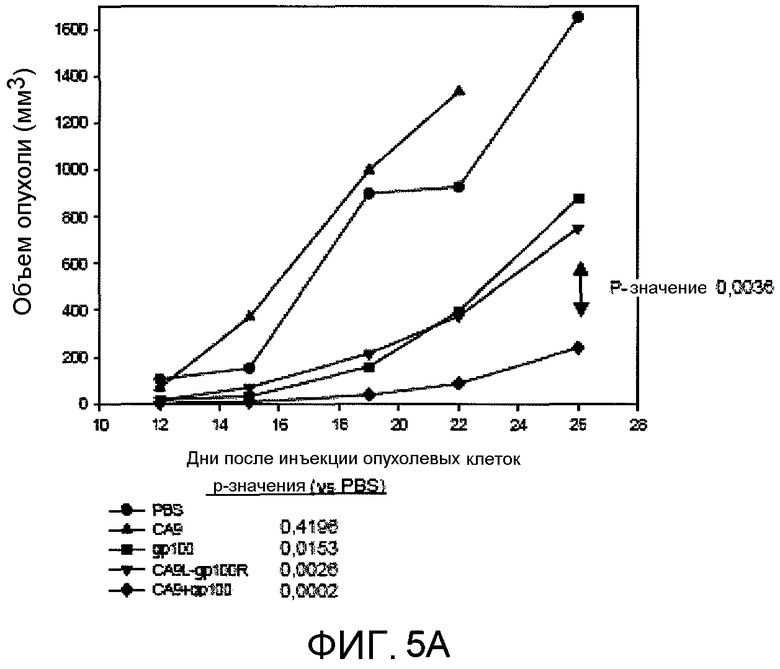
На фиг.5А представлено графическое изображение результатов, полученных иммунизацией мышей C57/BL6 (5 на группу) комплексом CA9-gp100 (CA9+gp100), которое демонстрирует стимуляцию противоопухолевого эффекта против опухолевых клеток B16-gp100. Опухолевые клетки B16-gp100 (2×105) инъецировали интрадермально через 7 дней после 3 иммунизаций, проводимых с интервалом 7 дней. Величины P на основании повторных измерений с использованием ANOVA (вариационный анализ), который учитывает вариабельность внутри каждой группы, представлены в таблице, сравнивающей каждую группу с контрольными мышами, которым инъецировали PBS. Скорость роста опухолей значимо отличалась при сравнении мышей, иммунизированных CA9+gp100, и мышей в любой контрольной группе; например, величина p составила 0,0036 при сравнении с группой CA9+gρ100, и мышами, иммунизированными инъекцией CA9 и gp100 в отдельные бока (CA9L-gp100R). Одинаковые результаты были получены в 3 отдельных экспериментах.
На фиг.5B представлено графическое изображение результатов, полученных иммунизацией мышей комплексом CA9+gp100, который стимулировал gp100-специфический ответ IFN-γ (интерферон-гамма) из клеток CD8+, измеренный с использованием анализа иммуноферментного анализа ELISPOT. Средняя величина±SEM представлены для экспериментов, выполненных в трех повторах.
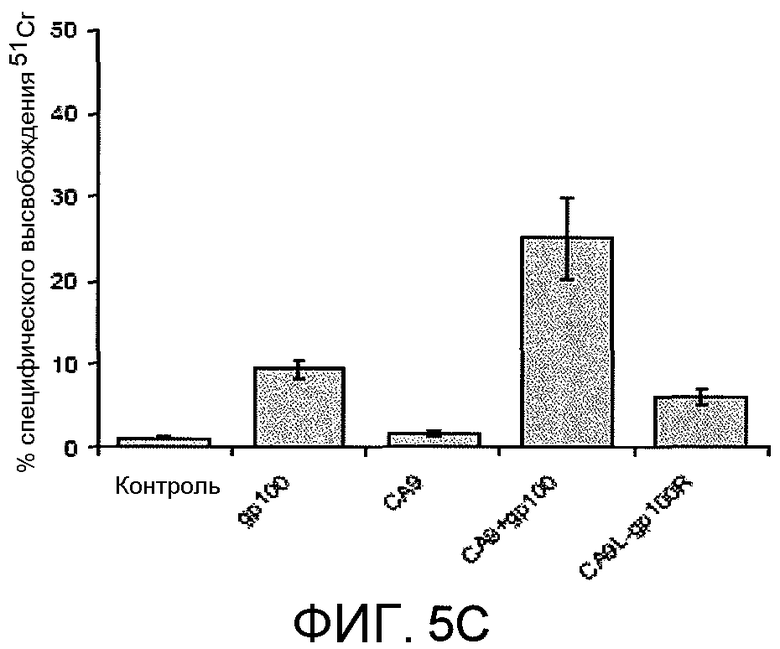
На фиг.5C представлено графическое изображение результатов, полученных иммунизацией мышей комплексом CA9+gp100, который стимулировал специфическую для опухоли Т-клеточный ответ, измеренный с использованием анализа высвобождения 51Cr. Средняя величина±SEM представлены для экспериментов, выполненных в трех повторах.
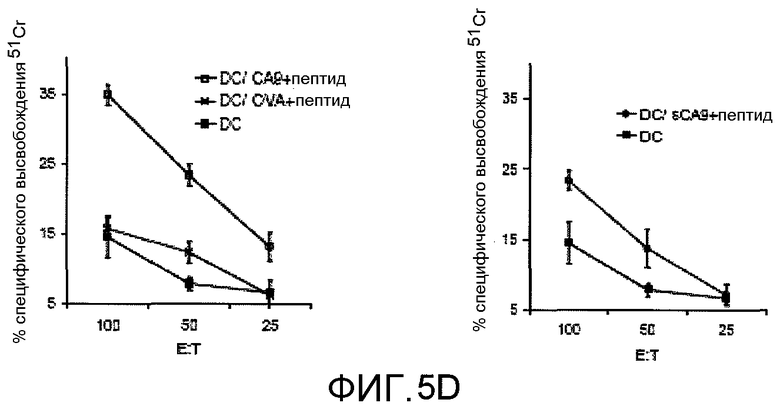
На фиг.5D представлено графическое изображение результатов, показывающих, что сброшенный CA9 способен стимулировать специфический клеточный иммунный ответ против мышиного пептида gp100 (EGSRNQDWL (SEQ ID NO:2)). Иммунизация DC, обработанными комплексом из CA9 и пептида gp100, вызывала пептид-специфический ответ цитотоксических Т-клеток, измеренный с использованием анализа высвобождения 51Cr. Мышей C57/BL6 (3 на группу) иммунизировали 3 раза с интервалом 7 дней 2×106 DC, полученными из костного мозга, обработанными указанными белок-пептидными комплексами в концентрации 10 мкг/мл и активировали LPS (липополисахаридом). Средняя величина±SEM представлены для экспериментов, выполненных в трех повторах.
На фиг.6A представлена фотография, на которой показаны результаты вестерн-блоттинга, полученные при анализе эффекта цитокинов на отделение и экспрессию CA9. Экспрессия CA9 линией клеток R6 RCC была увеличена в ответ на среды, кондиционированные человеческими WBC, обработанными IL2 (интерлейкин 2) и IFN-α (интерферон-альфа). WBC выделяли из крови человека и обрабатывали IL2, IFN-α, IFN-γ или не обрабатывали ничем (контроль) в течение 24 ч для получения кондиционированных сред (CM). Клетки R6 обрабатывали CM в течение 48 ч перед лизисом и зондированием антителом против CA9. CM использовали, поскольку цитокины обеспечивают терапевтическое благоприятное действие стимуляцией иммунных клеток, а не нацеливаясь непосредственно на опухоли.
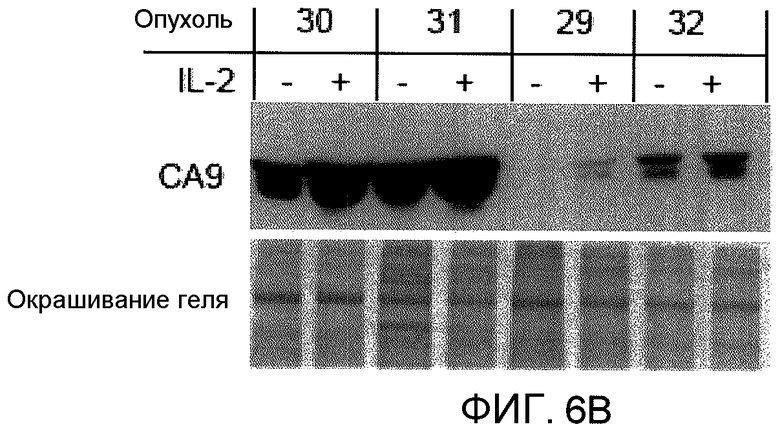
На фиг.6B представлена фотография, фотография, на которой показаны результаты вестерн-блоттинга, свидетельствующие о том, что сброс CA9 первичными опухолевыми эксплантатами в кратковременной культуре увеличивается в ответ на действие IL2. Первичные почечные опухоли культивировали с IL2 или без него. Культуральные среды зондировали антителом против CA9. Культуральные среды разделяли электрофорезом и окрашивали кумасси голубым, и они служили в качестве контроля. IL2 наносили непосредственно на хирургические образцы, которые содержат как опухолевые клетки, так и иммунные клетки.
Описание изобретения
Настоящее изобретение относится к композициям и способам стимуляции иммунного ответа на антиген у индивидуума. Композиция содержит комплекс изолированного белка CA9 и антигена. Способ включает введение композиции индивидууму в количестве, эффективном для стимуляции иммунного ответа на антиген, где стимулированный иммунный является более выраженным, чем иммунный ответ, стимулированный одним антигеном. Стимулированный иммунный ответ может оказывать терапевтический или профилактический эффект и может включать клеточно-опосредованный и/или гуморальный ответ или его комбинацию.
Настоящее изобретение основано на неожиданно обнаруженной у CF9 новой функции шаперона, и это считается открытием первого белка клеточной поверхности, который может функционировать в качестве шаперона. «Шаперон» в целом считается в данной области белком, который может связывать белки-клиенты, предотвращать агрегацию белков-клиентов при температурах, вызывающих тепловой шок, таких как температуры приблизительно 42-43°C, и обеспечивать возможность повторного свертывания денатурированных белков. Однако тепловое повреждение белков, необходимое для образования комплексов с белками-шаперонами, может происходить при температурах, достигающих приблизительно 55°C. Белки теплового шока (HSP) являются хорошо известными, вездесущими внутриклеточными шаперонами, экспрессия которых запускается индуцированными тепловым шоком факторами транскрипции, включая HSFl, HSF2, HSF2BP, HSF4, HSF5, HSFXl, HSFX2, HSFYl, HSFY2. Авторы продемонстрировали, что CA9 выполняет много функций, обычно относимых к белкам теплового шока HSP, и что растворимая форма CA9, которая отделяется от поверхности клеток, также обладает свойствами шаперона HSP. В частности, авторы продемонстрировали, что CA9 ингибирует вызванную нагреванием агрегацию белка, обеспечивает возможность повторного сворачивания денатурированного белка и восстанавливает ферментативную функцию белков, с которыми он образует комплексы. Авторы продемонстрировали, что CA9 связывается с дендритными клетками (DC) специфичным для рецептора образом, и показали, что связанный CA9 интернализуется DC и перерабатывается в первую очередь по протеосомальному пути, который, как считают, необходим для представления экзогенных антигенов на MHC I (главном комплексе тканевой совместимости) и активации Т лимфоцитов CD8+. Считается также, что такое представление антигена является важным для стимуляции вакциной иммунного ответа, который может ингибировать рост опухолей. Таким образом, приведенные результаты совместно указывают на то, что CA9, вероятно, играет непосредственную роль в стимуляции адаптивного иммунного ответа. В этом отношении, авторы также продемонстрировали на модели рака в виде мышиной меланомы, что введение композиции, содержащей CA9 и антиген, может стимулировать специфический для антигена противоопухолевый ответ, который является более выраженным, чем ответ, генерируемый одним антигеном.
Несмотря на указанные выше сходства между CA9 и HSP, CA9 имеет несколько необычных и желаемых признаков, по сравнению с HSP, и поэтому не считается одним из HSP. Например, HSP индуцируются нагреванием и, в соответствии с их ролью в обеспечении термической устойчивости, HSP связывают антигены-мишени при температурах, вызывающих тепловой шок, например, 42-43°C. Напротив, CA9 индуцируется не только нагреванием (но и гипоксией), и авторы продемонстрировали, что CA9 эффективно связывает антигены-мишени при 37°C. Кроме того, CA9 может также образовывать иммуногенные комплексы при комнатной температуре. Таким образом, такое свойство CA9 может содействовать более эффективному получению композиций на основе CA9 для применения в способе по изобретению. Кроме того, HSP являются внутриклеточными шаперонами и в целом высвобождаются во внеклеточную среду только после нарушения клеточной целостности. С другой стороны, CA9 представляет собой белок клеточной поверхности и также может сбрасываться во внеклеточную среду жизнеспособными клетками. Кроме того, в клетках с интактной реакцией на гипоксию, CA9 экспрессируется в ответ на уменьшение парциального давления кислорода. Поэтому и без связи с какой-либо конкретной теорией, полагают, что настоящее изобретение выявляет, что экспрессия и сброс CA9 может представлять собой общий механизм вовлечения иммунной системы в нацеливание на внеклеточные антигены в ответ на гипоксический стресс, и может объяснить благоприятный прогноз в случаях RCC, где растворимый CA9 сбрасывается из опухолей. Кроме того, клетки, экспрессирующие CA9 в организме хозяина, могут представлять непрерывный источник для продукции иммунного адъюванта, который мог бы служить содействию иммуностимулирующим эффектам настоящего изобретения.
Белок CA9 полной длины имеет следующую аминокислотную последовательность:
1 maplcpspwl pllipapapg ltvqlllsll llvpvhpgrl prmqedsplg ggssgeddpl
61 geedlpseed spreedppge edlpgeedlp geedlpevkp kseeegslkl edlptveapg
121 dpqepqnnah rdkegddqsh wryggdppwp rvspacagrf qspvdirpql aafcpalrpl
181 ellgfqlppl pelrlrnngh svqltlppgl emalgpgrey ralqlhlhwg aagrpgseht
241 veghrfpaei hvvhlstafa rvdealgrpg glavlaafle egpeensaye qllsrleeia
301 eegsetqvpg ldisallpsd fsryfqyegs lttppcaqgv iwtvfnqtvm lsakqlhtls
361 dtlwgpgdsr lqlnfratqp lngrvieasf pagvdsspra aepvqlnscl aagdilalvf
421 gllfavtsva flvqmrrqhr rgtkggvsyr paevaetga. (SEQ ID NO:1).
кДНК человеческого CA9 известна в данной области и кодирует белок из 459 аминокислот с N-концевой внеклеточной частью из 414 аминокислот, связанной посредством прогнозированной гидрофобной трансмембранной области (ТМ) из 20 аминокислот с прогнозированным С-концевым внутриклеточным хвостом (IC) из 25 аминокислот. Прогнозированная последовательность ТМ представлена в SEQ ID NO:1 в виде аминокислот 415-434. Прогнозированная последовательность IC представлена в SEQ ID NO:1 в виде аминокислот 435-459. Таким образом, прогнозированная последовательность сброшенного белка CA9 представлена в SEQ ID NO:1 в виде аминокислот 1-414.
CA9 полной длины имеет прогнозированную молекулярную массу 49,7 кДа, но он может быть выявлен на вестерн-блотах 54 кДа и 58 кДа. Сброшенная форма CA9 представляет собой растворимую форму CA9, которая высвобождается в культуральную среду и в биологические жидкости, наиболее вероятно, путем протеолитического отщепления внеклеточной части от трансмембранной и внутриклеточной последовательностей. Сброшенный CA9 на 4 кДа меньше чем CA9 полной длины, по данным выявления на вестерн-блотах и, таким образом, мигрирует в виде сдвоенных полос 50/54 кДа (Zavada et al., Br J Cancer. 2003 Sep 15;89(6):1067-71), но имеет прогнозированную молекулярную массу 45,7 кДа.
Композиции по настоящему изобретению содержат изолированный белок CA9. Под термином «изолированный белок CA9» подразумевается, что белок выделен из его естественной среды. Считается, что описанные в настоящем изобретении комплексы CA9/антигена содержат изолированный белок CA9.
Белок CA9 для применения в композициях и способах по изобретению может быть выделен из клеток, которые экспрессируют белок CA9. Некоторые неограничивающие примеры таких клеток включают клетки, которые эндогенно экспрессируют белок CA9 из геномных кодирующих последовательностей, и клетки, которые были подвергнуты манипуляция генной инженерии для экспрессии рекомбинантного белка CA9. Такая рекомбинантная экспрессия белка CA9 может быть достигнута с использованием широкого разнообразия обычных методик и систем рекомбинантной экспрессии белка, известных в данной области. Альтернативно, сброшенный белок CA9 может быть выделен с использованием известных способов из сред для клеточных культур, в которых культивируются клетки, экспрессирующие CA9 посредством эндогенной генной экспрессии или генной инженерии. В случае пептидных антигенов, пептиды могут быть химически синтезированы при помощи любой из разнообразных хорошо известных методик или могут быть получены в результате протеолитического расщепления более крупных белков.
Изолированный белок CA9 необязательно должен представлять собой очищенный белок. Однако изолированный белок CA9 может, тем не менее, быть очищен до любой желаемой степени чистоты. Способы очистки белка хорошо известны в данной области и могут применяться для получения очищенного белка СА9 для использования в настоящем изобретении.
Предусматривается, что настоящее изобретение может применяться для стимуляции иммунного ответа на любой белок, полипептид или пептидный антиген, за исключением того, что используемый в данном описании термины «антиген» и «антигены» не включают HSP. Примеры HSP включают hsp40, калретикулин, hsp60, hsp70, hsp90, HSP110. Таким образом, за исключением иных случаев, описанных в данном описании, антигены, подходящие для использования в настоящем изобретении, включают, но без ограничения, антигены, экспрессируемые раковыми клетками или инфекционными агентами. Антиген может быть хорошо охарактеризован или может быть неизвестен, кроме как известностью его присутствия, например, в лизате из определенного типа клеток, такого как опухоль или бактериальная культура.
В одном варианте осуществления антиген, присутствующий в изолированных комплексах CA9/антиген по изобретению, представляет собой опухолевый антиген. Опухолевые антигены могут быть получены при помощи обычных методик, таких как получение лизатов опухолевых клеток повторным замораживанием и оттаиванием опухолевых клеток/тканей в забуференном фосфатом солевом растворе, содержащем лейпептин и апротинин (полученный из свежей биопсии ткани опухоли или из опухолевых клеток, генерированных in vitro тканевой культурой). Такое замораживание и оттаивание приводит к лизису клеток. Опухолевой лизат может быть получен путем центрифугирования и сбора надосадочной жидкости. Лизаты опухолевых клеток могут применяться сразу или замораживаться и храниться, например, при -70°C до готовности к использованию.
В различных вариантах осуществления антиген может представлять собой антиген, экспрессируемый раковыми клетками, конкретные примеры которых включают, но без ограничения, фибросаркому, миксосаркому, липосаркому, хондросаркому, остеогенную саркому, хордому, ангиосаркому, эндотелиосаркому, лимфангиосаркому, псевдомиксому брюшины, лимфангиоэндотелиосаркому, синовиому, мезотелиому, опухоль Юинга, лейомиосаркому, рабдомиосаркому, карциному ободочной кишки, рак поджелудочной железы, рак молочной железы, рак яичника, рак предстательной железы, плоскоклеточную карциному, базальноклеточную карциному, аденокарциному, карциному потовых желез, карциному сальных желез, папиллярную карциному, папиллярные аденокарциномы, цистаденокарциному, медуллярную карциному, бронхогенную карциному, почечноклеточную карциному, гепатому, карциному желчных протоков, хориокарциному, семиному, эмбриональную карциному, опухоль Вильнса, рак шейки матки, рак семенников, карциному легких, мелкоклеточную карциному легких, карциному мочевого пузыря, эпителиальную карциному, глиому, астроцитому, медуллобластому, краниофарингому, эпендимому, пинеалому, гемангиобластому, нейрому слухового нерва, олигодендроглиому, менингиому, меланому, нейробластому, ретинобластому, лейкоз, лимфому, множественную миелому, макроглобулинемию Вальденстрома и болезнь тяжелых цепей. В одном варианте осуществления антиген представляет собой gp 100.
В других вариантах осуществления, антигены, используемые в изобретении, могут представлять собой такие, которые экспрессируются инфекционными агентами. Примеры таких инфекционных агентов включают, но без ограничения, вирусы, бактерии, грибы и другие паразиты. Примеры вирусов включают, но без ограничения, вирусы гепаптита типа В или типа С, гриппа, ветряной оспы, аденовирус, вирус простого герпеса типа I или типа II, чумы рогатого скота, риновирус, эховирус, тотавирус, респираторный синцитиальный вирус, вирус папилломы, цитомегаловирус, эхиновирус, арбовирус, гантавирус, коксаки вирус, вирус эпидемического паротита, вирус кори, вирус краснухи, вирус полиомиелита, вирус иммунодефицита человека типа I или типа II. Примеры бактерий включают, но без ограничения, M. tuberculosis, микобактерию, микоплазму, Neisseria и Legionella. Примеры других паразитов включают, но без ограничения, рикеттсии и хламидии.
Без намерения быть связанными какой-либо конкретной теорией, считают, что для применения в способе по изобретению, изолированный белок СА9 и антиген должны присутствовать в комплексе и, таким образом, соединены друг с другом химическим связыванием, таким как ковалентные связи, ионные связи, водородные связи и/или связи Ван дер Ваальса или их комбинации. Таким образом, в одном варианте осуществления белок СА9 и антиген могут присутствовать в комплексе, где белок СА9 и антиген ковалентно не связаны друг с другом. Способы образования комплексов белка/антигена без ковалентного связывания известны в данной области и могут использоваться для образования комплексов между изолированным белком СА9 и одним или более антигенами. Комплексы по изобретению могут содержать белок СА9 и антиген или могут состоять по существу из белка СА9 и антигена, или могут состоять из белка СА9 и антигена.
В одном варианте осуществления комплекс, содержащий изолированный СА9 и нековалентно связанный антиген, может быть образован в подходящем буфере при комнатной температуре (например, приблизительно 20°C), или при более высокой температуре, например, 37°C, но ниже более высоких температур, обычно требуемых для того, чтобы HSP образовывали комплекс с белком, например, 43°C. Соответственно комплексы CA9/антигена по настоящему изобретению могут быть образованы, например, при температуре в диапазоне от 20°C до 42°C, включительно, и включая все целые числа от 20°C до 42°C. Комплексы могут также быть образованы при более высоких температурах. В одном варианте осуществления комплекс CA9/антиген образуется при 37°C. Подходящие молярные отношения CA9 к антигену могут быть определены специалистами в данной области, с учетом благоприятного эффекта, раскрытого в данном описании. В одном варианте осуществления используется отношение CA9/антигена 1:1.
В одном варианте осуществления для использования в настоящем способе эндогенный CA9 может быть изолирован из биологического образца, содержащего клетки, такого как биологический образец, полученный из опухоли, и выделенный белок CA9, таким образом, может уже образовывать комплексы с опухолевыми антигенами, против которых надлежит стимулировать иммунный ответ.
В другом варианте осуществления изолированный белок CA9 может быть ковалентно связан с антигеном, например, химическим конъюгированием белка CA9 с антигеном для образования комплекса конъюгата изолированного белка CA9/антигена. Подходящие способы химического конъюгирования белков известны в данной области и могут использоваться для образования конъюгатов изолированного белка CA9/антигена для композиций и способов по изобретению. Кратко, CA9 могут быть ковалентно конъюгированы с антигеном посредством поперечной сшивки аминокислотных остатков CA9 с аминокислотными остатками антигена. Аминокислотные остатки могут быть конъюгированы, например, поперечной сшивкой амино, сульфгидрильных или групп карбоновых кислот с использованием широкого разнообразия хорошо известных реагентов и методик.
В еще одном варианте осуществления комплекс CA9/антигена может быть получен в виде слитого белка CA9/антигена. Кратко, для получения такого слитого белка, последовательности ДНК, кодирующие белок CA9, и антиген могут быть сконструированы с использованием обычных методик и экспрессированы в подходящем типе клеток с использованием любого целесообразного вектора экспрессии. Затем слитый белок может быть экспрессирован в клетках и изолирован с использованием любого способа, известного специалистам в данной области. В одном варианте осуществления слитый белок CA9/антигена может быть отделен линкерной последовательностью.
В еще одном варианте осуществления изолированный белок CA9 может быть смешан с клеточным материалом, таким как опухолевый лизат, для обеспечения возможности связывания CA9 с одним или более опухолевыми антигенами, присутствующими в лизате, образуя, таким образом, гетерогенную смесь изолированных комплексов CA9/антигена, которая может использоваться в способе по изобретению в качестве мультивалентной противораковой вакцины. Таким образом, будет понятно, что могут быть предоставлены изолированные белки CA9 для применения в изобретении с тем, чтобы дискретные изолированные белки CA9 образовывали комплексы с различными антигенами.
В композициях и способах по изобретению могут использоваться любые из различных средств доставки. Подходящие средства доставки включают, но без ограничения, представляющие антиген клетки (APC), такие как дендритные клетки, макрофаги, B клетки, моноциты и другие клетки, которые могут методами генной инженерии сделаны эффективными APC.
В одном варианте осуществления изобретение относится к композиции и способу стимуляции у индивидуума иммунного ответа на антиген, включающему введение индивидууму композиции, содержащей APC, такие как дендритные клетки, которые вступили в контакт с комплексом, содержащим изолированный белок CA9 и антиген. Дендритные клетки могут содержать комплекс белка CA9/антигена во время, когда дендритные клетки вводятся индивидууму. Это может быть достигнуто, например, предварительной загрузкой дендритных клеток комплексом или трансфекцией клеток ДНК, кодирующей белок CA9, и антигеном в виде или различных белков, или в виде слитого белка.
Когда используются APC, то APC, такие как дендритные клетки, могут быть сначала выделены у индивидуума и подготовлены для воздействия комплекса CA9/антиген с использованием обычных методик. Дендритные клетки могут быть выделены у индивидуума, у которого желателен стимулированный иммунный ответ на антиген, или могут быть выделены у другого индивидуума.
В одном варианте осуществления изобретение относится к по существу очищенной популяции дендритных клеток, которые контактировали с комплексом CA9/антиген.
Для применения в композициях и способах по изобретению, может быть использован любой подходящий носитель, известный специалистам в данной области. Тип носителя варьируется в зависимости от пути введения. Композиции по настоящему изобретению могут составляться для любого целесообразного пути введения, включая, например, местное, пероральное, интраназальное, внутривенное, внутричерепное, внутрибрюшинное, подкожное или внутримышечное введение. Для парентерального введения, такого как подкожная инъекция, носитель может содержать воду, солевой раствор, спирт, жир, воск или буфер. Для перорального введения, можно использовать любой из указанных выше носителей или твердый носитель, такой как маннит, лактоза, крахмал, стеарат магния, сахарин натрия, тальк, целлюлоза, глюкоза, сахароза и карбонат магния. Композиции могут также содержать буферы (например, нейтральный забуференный солевой раствор или забуференный фосфатом солевой раствор), углеводы (например, глюкозу, маннозу, сахарозу или декстраны), манит, антиоксиданты, хелатообразующие агенты, такие как EDTA или глютатион, адъюванты (например, гидроксид алюминия) и/или консерванты.
Композиции по изобретению могут вводиться с использованием любого подходящего пути введения. Некоторые неограничивающие примеры включают пероральный, парентеральный, подкожный, внутрибрюшинный, внутрилегочный и интраназальный. Парентеральные вливания включают внутримышечное, внутривенное, внутриартериальное, внутрибрюшинное и подкожное введение.
Введение композиций по изобретению может выполняться в сочетании с обычными видами лечения, которые предназначены для лечения заболевания или расстройства, связанного с антигеном. Например, композиция может вводиться перед, одновременно или после обычных противораковых видов лечения. Такие виды лечения могут включать, но без ограничения, виды химиотерапии, хирургические вмешательства и лучевую терапию.
Как правило, соответствующая дозировка и схема лечения предоставляет композицию в количестве, эффективном для стимуляции иммунного ответа, который обеспечивает терапевтический и/или профилактический благоприятный эффект. Мониторинг такого ответа может осуществляться улучшенным клиническим исходом, например, ингибированием роста опухоли и/или метастазирования, повышенной устойчивостью к инфекции, улучшенной активацией иммунных клеток и/или другими параметрами, которые очевидны для специалистов в данной области, в зависимости от подвергаемого лечению состояния.
Пути и частота введения терапевтических композиций, раскрытых в данном описании, а также дозировка будут варьироваться от индивидуума к индивидууму и могут быть легко установлены с использованием стандартных методик. В целом, количество каждого комплекса CA9/антиген, присутствующего в дозе, может находиться в диапазоне примерно от 100 мкг до 5 мг на кг массы тела хозяина. Подходящие объемы будут варьироваться в зависимости от размеров тела пациента, но обычно будут находиться в диапазоне примерно от 0,1 мл до примерно 5 мл. В одном варианте осуществления, композиция по изобретению может вводиться один раз в неделю всего в количестве 3 доз.
Пример 1
В данном примере представлено описание материалов и способов, которые использовались при создании изобретения.
Мыши, клеточные линии и конструкты ДНК
Самок мышей C57/BL6 в возрасте 6-8 недель закупали в NCI (Frederick, MD) и содержали в условиях, лишенных болезнетворных микроорганизмов. Мышей SR-A null, подвергнутых обратному бридингу в исходный фенотип C57BL/6J, получали у B. Berwin (Dartmouth University) в виде щедрого дара господ T. Kodama из Токийского университа (Tokyo University) и M.W. Freeman (Massachusetts General Hospital, NHLBI Program in Genomics Applications). Клетки B16, трансдуцированные человеческим gp100 (B16-gp100), были любезно предоставлены доктором Alexander Rakhmilevich (University of Wisconsin, Madison, WI). R6, линия клеток человеческой RCC, которая экспрессирует CA9, была даром доктора Arie Belldegrun (UCLA, Los Angeles). Клетки RENCA и RENCA, устойчиво трансдуцированные для экспрессии человеческого CA9 (RENCA-CA9) были даром доктора Arie Belldegrun (UCLA, Los Angeles, CA). Эти клетки поддерживали в среде RPMI 1640 с добавлением 10% инактивированной нагреванием фетальной телячьей сыворотки (FBS; Life Technologies, Grand Island, NY), 2 ммоль/л L-глутамина, 100 единиц/мл пенициллина и 100 мкг/мл стрептомицина.
кДНК для мышиного HSP110, мышиного HSP70, человеческого CA9 и человеческого gp100 (дар доктора Nicholas Restifo, National Cancer Institute, Bethesda, MD) клонировали в вектор pBacPAK-his (BD Biosciences Clontech, Palo Alto, CA), трансформировали в монослой клеток Sf21 с использованием вируса с дефектной репликацией, и экспрессировали, используя систему бакуловируса BacPAK. Белки очищали с использованием никелевой колонки с нитрилуксусной кислотой-агарозой (Qiagen, Valencia, CA). Концентрации белка измеряли с использованием набора Protein Assay (Bio-Rad, Hercules, CA). Чистоту белка оценивали с использованием электрофореза в додецилсульфате натрия-полиакриламидном геле (SDS-PAGE) и окрашиванием кумасси голубым. Уровни эндотоксина в рекомбинантных белках оценивали, используя набор лизата Limulus Amebocyte (Biowhittaker, Walkersville, MD) и составляли 10-25 эндотоксиновых единиц/мг белка.
Образование комплекса шаперон-антиген
Для образования комплекса между шапероном (т.е. HSP110, CA9, OVA) и антигеном (т.е. gp100 и люциферазой), 2 белка объединяли в молярном соотношении 1:1 и инкубировали в течение 30 мин при 37°C или при температурах теплового шока 43°C, как описано ранее (27). Комплекс предварительно обрабатывали 30 мкл зерен белка G и подвергали иммунопреципитации с использованием мышиного моноклонального антитела против человеческого CA9 (дар доктора Egbert Oosterwijk, University of Nijmegen, Nijmegen, Netherlands), ранее описанного кроличьего антитела против HSP110 (28) или мышиного антитела против OVA (Sigma, St. Louis, MO). После электрофореза SDS-PAGE (10%), выполняли вестерн-блоттинг с использованием антитела против люциферазы (Promega, Madison, WI) или антитела против gp100 (Santa Cruz Biotechnology, Santa Cruz, CA).
Агрегация люциферазы и анализ повторного сворачивания
Для анализа агрегации люциферазы, 0,15 мкМ люциферазы (Sigma, St Louis, MO) и белок шаперон (т.е. CA9, HSP110, HSP70) инкубировали в 25 мМ Hepes (pH 7,9), 5 мМ стеарата магния, 50 мМ KCl и 5 мМ β-меркаптоэтанола при 43°C в течение 30 мин. OVA служил в качестве контроля для белков-шаперонов. Мониторинг агрегации белков осуществляли измерением оптической плотности при 320 нм. Для подтверждения того, что белки-шапероны предотвращали агрегацию, растворы центрифугировали при 16000×g в течение 15 мин, и растворимую и осажденную фракции разделяли, проводили SDS-PAGE и подвергали вестерн-блот анализу с антителом против люциферазы (Promega, Madison, WI).
Для анализа повторного сворачивания люциферазы, люциферазу и белок-шаперон нагревали в буфере повторного сворачивания (25 мМ Hepes, pH 7,6, 5 мМ MgCl2, 2 мМ дитиотрейтол и 2 мМ АТФ) при 43°C в течение 30 мин. Нагретую люциферазу разводили в 100 раз в буфере повторного сворачивания, содержащем 60% лизат кроличьих ретикулоцитов (Promega, Madison, WI) и инкубировали при 30°C в течении 2 ч. Для измерения активности люциферазы, раствор дополнительно разводили в 5 раз в 25 мМ Hepes (pH 7,6), 1 мг/мл бычьем сывороточном альбумине; 10 мкл добавляли к 100 мкл раствора для анализа люциферазы (Promega, Madison, WI). Количественное определение активности люциферазы проводили с использованием люминометра Lumat LB9501 (Berthoid, Bad. Wildbad, Germany).
Предотвращение опухоли и иммунный мониторинг
Самок мышей C57/BL6 (NCI, Frederick, MD) в возрасте 6-8 недель (5 на группу) иммунизировали 3 раза с интервалом 7 дней 100 мкл вакцины. У мышей проводили антигенную стимуляцию 2×105 B16-gp100 клетками путем интрадермальной инъекции, через 7 дней после последней иммунизации. Опухоли измеряли через каждые 3 дня с использованием электронного циркуля и рассчитывали объем опухоли [(самый маленький диаметр2×самый большой диаметр)/2]. Полный набор экспериментов повторяли 3 раза. Были описаны анализы ELISPOT и высвобождения 51Cr (27). Кратко, лимфатические узлы (LN) и селезенку извлекали через 2 недели после иммунизации. Т клетки CD8+ изолировали отрицательным отбором с использованием колонок обогащения CD8+ клетками (Cedarlane Laboratories, Ontario, Canada). Обогащенные CD8+ T клетки (5×104/лунка) инкубировали с клетками DC 1.2, предварительно подвергнутыми импульсному воздействию gp100 (10 мкг/мл) при 37°C в течение 48 ч. Пятна IFN-γ подсчитывали с использованием устройства KS Elispot System (версия 4.3.56) от компании Zeiss Microscopy (Oberkochen, Germany).
Для анализа высвобождения 51Cr, спленоциты и LN извлекали через 2 недели после иммунизацию и стимулировали in vitro клетками B16-gp100, предварительно обработанными митомицином (50 мкг/мл) в течение 5 дней. Затем спленоциты серийно разводили в 96-луночном планшете, содержащем меченные 51Cr опухолевые клетки (1×104 клеток/лунка) в трех повторах при изменяющемся соотношении E:T. После 8 ч инкубации при 37°C, супернатант анализировали на радиоактивность с использованием гамма счетчика (Packard, Downers Grove, IL).
Для анализа CTL (цитотоксических Т лимфоцитов) in vivo, спленоциты брали у интактных мышей для получения клеток-мишеней. Эритроциты лизировали и на одноклеточные суспензии спленоцитов, 1×107 клеток/мл, импульсно обрабатывали в течение 30 мин при 37°C 10 мкМ пептида или без него в среде DMEM (модифицированной Дульбекко среда Игла), содержащей 10% FBS. Двухклеточные популяции в концентрации 2×107 клеток/мл (в PBS/0,1% BSA) метили различными концентрациями CFSE (0,5 или 12,5 мкМ). Мечение CFSE прекращали добавлением равного объема FBS в течение 1 мин и промыванием 3 раза полной средой RPMI 1640. 5×106 клеток из каждой популяции, подвергнутой импульсной обработке пептидом и не подвергнутой такой обработке, смешивали и инъецировали внутривенно иммунизированным и неиммунизированным мышам.
Мышей умерщвляли через 16 ч. Одноклеточные суспензии спленоцитов анализировали проточной цитометрией. Специфический лизис флюоресцентных донорских спленоцитов в процентах у иммунизированных мышей рассчитывали следующим образом:
[(число не подвергнутых импульсному воздействию мишеней×A-число подвергнутых импульсному воздействию мишеней)/число не подвергнутых импульсному воздействию мишеней×A]×100,
где A=[число подвергнутых импульсному воздействию мишеней/число не подвергнутых импульсному воздействию мишеней] у неиммунизированных мышей-реципиентов.
Генерирование иммунного ответа sCA9
Мышей (5 на группу) иммунизировали 3 раза с интервалами 7 дней вакцинами на основе DC. Группы вакцинации включали DC, обработанные комплексом CA9 и мышиным пептидом gp100 (EGSRNQDWL с чистотой >99% по ВЭЖХ, синтезированным Alpha Diagnostic international, San Antonio, TX) (CA9+пептид), HSP110+пептид и sCA9+пептид. Необработанные DC и обработанные OVA+пептид служили в качестве отрицательного контроля. Для образования белково-пептидных комплексов, 2 мкг пептида инкубировали в течение 30 мин с 20 мкг белков (OVA при 43°C, CA9 при 37°C, sCA9 при 37°C или HSP110 при 43°C). Белково-пептидные комплексы добавляли к DC, полученным из костного мозга.
Для генерирования DC, образцы костного мозга брали из бедренных и большеберцовых костей мышей и обрабатывали буфером для лизиса эритроцитов, промывали и высевали при плотности 1×106 клеток на 1 мл в 12-луночных планшетах в среде RPMI-1640, содержащей 10% FBS и 10 нг/мл рекомбинантного фактора, стимулирующего колонии мышиных гранулоцитов-моноцитов (GM-CSF) (eBioscience, San Diego CA). Клетки подавали каждый 2 день и собирали между 7 и 9 днями. Культуры состояли из 75-90% клеток CD11C+. Для генерирования вакцин, культивированные клетки подвергали импульсной обработке в течение 4-6 часов 10 мкг/мл белково-пептидного комплекса и обрабатывали 100 нг/мл LPS в течение 16 часов. 2×106 клеток инъецировали мыши подкожно. Через 7 дней после последней иммунизации, лимфатические узлы и спленоциты собирали для анализов CTL реакции CA9 на цитокины in vivo и in vitro.
Ответ CA9 на цитокины
Мониторинг экспрессии CA9 осуществляли зондированием лизатов клеток R6 антителами против CA9 после обработки кондиционированными средами (CM) в концентрации 200 мкл/мл в течение 48 ч. WBC отделяли от цельной крови, полученной от здоровых людей, и культуральные среды от WBC, обработанные 100 нг/мл цитокинами или без обработки (контроль) в течение 24 ч служили в качестве CM. Для мониторинга сбрасывания CA9 из краткосрочных культур эксплантатов RCC, фрагменты опухоли, разрезанные на кусочки по 1 мм (33 мг/мл) споласкивали бессывороточной средой DMEM, культивировали с IL2 (100 нг/мл) или без него в DMEM с 10% FBS в 24-луночном планшете и инкубировали при 37°C в инкубаторе с 5% CO2 в течение 3 дней. Для количественного анализа экспрессии CA9, фрагменты опухоли оценивали вестерн-блоттингом, используя антитело против CA9.
Связывание комплекса шаперона-антигена дендритными клетками (DC)
К меченному FITC CA9 или BSA (контрольный белок), добавляли FITC (Sigma, St. Louis, MO) при 20M избытке в 0,1M буфере бикарбонат/карбонат натрия. Свободный FITC удаляли на колонке с Sephadex G-25 (Pharmacia, Piscataway, NJ). Белки подвергали SDS-PAGE для подтверждения конъюгации FITC.
Для оценки связывания с DC, 10 мкг/мл конъюгированных с FITC белков инкубировали в течение 20 мин на льду с DC, полученными из мышиного костного мозга (см. подраздел «Генерирование иммунного ответа sCA9») в концентрации 1×106 клеток/мл в 100 мкл PBS, содержащем 1% BSA. Для исследования конкуренции связывания, немеченый CA9 или фукоидан добавляли в изменяющихся концентрациях к 1×106 DC/млл при 4°C в течение 20 мин. DC промывали 3 раза 1% BSA/PBS и затем инкубировали с 200 мкг/мл FITC-CA9. Проводили контрокрашивание ядра DAPI.
Клетки фиксировали 1% параформальдегидом (Fisher, Fair Lawn, NJ) и исследовали конфокальной микроскопией (Bio-Rad 600, Hercules, CA) и анализировали проточной цитометрией (Becton Dickenson, La Jolla, CA).
Обработка комплекса шаперон-антиген DC
DC выращивали до 90% слияния, обрабатывали 10 мкМ MGl32 или 10 мМ NH4Cl в течение 2 ч при 37°C. Необработанные клетки служили в качестве контролей. Клетки охлаждали до 4°C в течение 30 мин перед добавлением CA9 в концентрации 10 мкг/мл. Клетки выдерживали при 4°C в течении еще одного часа и затем промывали охлажденной полной средой RPMI 1640. Затем клетки нагревали до 37°C, и собирали в точки времени 0, 0,5, 1, 2, 4 и 24 ч, промывали и обрабатывали буфером для анализа радиоиммунной защиты (RIPA) (Sigma, St. Louis, MO) в течение 15 мин на льду для лизиса клеток. 20 мкг лизата подвергали вестерн-блоттингу. Блоты зондировали мышиным антителом против человеческого CA9.
Кратковременная культура почечной опухоли из хирургических образцов
Свежие опухоли почек человека получали от учрежденной службы заготовки тканей в соответствии с протоколом, утвержденным IRB (153605). Опухолевые ткани разрезали на кусочки по 1 мм, споласкивали бессывороточной средой RPMI 1640, суспендировали в DMEM с 10% FBS и инкубировали в 100 мм чашках Петри при 37°C в инкубаторе с 5% CO2. Для количественного определения CA9, сбрасываемого из опухолей, культуральную среду собирали из суспензионной культуры через 2 дня. Для количественного определения уровней CA9 в опухоли, небольшие фрагменты опухоли обрабатывали буфером RIPA. CA9 из культуральной среды или клеточного экстракта анализировали вестерн-блоттингом.
SCA9 концентрировали из клеток RENCA-CA9, сбрасывающие растворимую форму CA9, которая на 4 кДа меньше, чем CA9. Клетки RENCA-CA9 культивировали в 20 мл среды RPCI 1640 с 10% FBS. Клетки, выращенные до 100% слияния, культивировали в течение еще 24 часов в бессывороточной среде RPMI 1640. Культуральную среду диализировали в течение 24 ч с PBS и центрифугировали при 4000 об/мин в течение 1 ч в концентрирующей пробирке. Концентрацию sCA9 измеряли вестерн-блоттингом, используя очищенный CA9 в качестве стандарта. Среду от материнских клеток RENCA, которые не экспрессируют CA9, концентрировали, используя такой же протокол, и они служили в качестве контроля.
Анализ данных
Планки погрешности показаны ±SEM для экспериментов, выполненных в трех повторах. Различия роста опухолей оценивали с использованием повторных мер ANOVA. Величина P<0,05 считалась статистически значимой. Статистический анализ выполняли с использованием программного обеспечения Stata 8.2 (StataCorp, College Station, Texas).
Пример 2
В данном примере описана способность изолированного белка CA9 действовать в качестве шаперона.
Для оценки наличия функции шаперона у CA9, люциферазу использовали в качестве белка-репортера (фиг.1А). HSP110, который представляет собой белок теплового шока с хорошо охарактеризованной функцией шаперона (29), служил в качестве положительного контроля, и овальбумин (OVA) служил в качестве отрицательного контроля. CA9, HSP110 или OVA смешивали с люциферазой в соотношении 1:1. Иммунопреципитацию выполняли антителами против CA9, HSP110 или OVA, и комплекс зондировали антителами против люциферазы. HSP110 эффективно и необратимо образовывал комплекс с люциферазой при 43°C. CA9 был более эффективен в образовании комплекса с люциферазой при комнатной температуре, чем при 43°C.
Для тестирования того, защищает ли люциферазу от агрегации образование комплекса между CA9 и люциферазой, CA9 и люциферазу смешивали в молярных соотношениях 1:1 и нагревали до 43°C (фиг.1B). Мониторинг агрегации белка осуществляли с течением времени оптической денсометрией. HSP70 представляет собой еще один хорошо охарактеризованный белок теплового шока. HSP110 и HSP70 включены в качестве положительных контролей. HSP110 был способен полностью предотвратить агрегацию люциферазы, и HSP70 и CA9 были одинаково эффективны в ингибировании агрегации люциферазы. Овальбумин (OVA) был отрицательным контролем и не был способен предотвратить агрегацию люциферазы. В подтверждающем эксперименте, CA9 был способен удерживать люциферазу в растворе при 43°C (фиг.1C). Смесь CA9 и люциферазы нагревали до 43°C и центрифугировали. Как CA9, так и HSP110 были эффективны в удерживании большей части люциферазы в супернатанте и вне осадка после центрифугирования.
HSP содействуют сворачиванию вновь синтезированных белков и повторному сворачиванию денатурированных белков. Для анализа на наличие этой функции, денатурированный нагреванием фермент может быть объединен с шаперонами в лизате кроличьих ретикулоцитов, и может быть проведен мониторинг восстановления ферментативной активности. Для оценки способности CA9 повторно сворачивать денатурированный белок, люциферазу использовали в качестве фермента-репортера (фиг.1D). HSP110 был самым эффективным шапероном для обеспечения возможности повторного сворачивания денатурированной нагреванием люциферазы. CA9 и HSP70 были одинакового эффективны. OVA служил в качестве отрицательного контроля и не содействовал повторному сворачиванию.
Пример 3
Данный пример демонстрирует, что изолированный CA9 может связываться с клетками, представляющими антиген.
Авторы продемонстрировали, что CA9 связывается с DC насыщаемым образом, указывающим на специфическое рецепторное связывание (фиг.2A). Связывание CA9 блокировали немеченым CA9 (фиг.2B) и фукоиданом (фиг.2C), который является лигандом для акцепторных рецепторов. Акцепторный рецептор А представляет собой один из многих акцепторных рецепторов на DC. Связывание CA9 уменьшалось, когда анализ связывания выполняли с использованием полученных из костного мозга DC, полученных у нокаутированных акцепторным рецептором А мышей (фиг.2D).
Пример 4
Данный пример демонстрирует, что сброшенный CA9 действует в качестве шаперона.
В предыдущих сообщениях описана растворимая форма сброшенного CA9, сбрасываемая с поверхности RCC (25, 26). Эти сообщения были подтверждены блоттингом клеточной культуральной среды для CA9 (фиг.3A). Растворимая форма CA9 (sCA9), которая была приблизительно на 4 кДа меньше, чем CA9 полной длины, сбрасывался из краткосрочной культуры прозрачно-клеточных опухолей почек. Однако здоровые почки и папиллярные опухоли почек не сбрасывали CA9. SCA9 и CA9 были одинаково эффективны в предотвращении агрегации люциферазы при 43°C (фиг.3B). Ключевые исследования связывания повторяли с использованием sCA9 с результатами, идентичными результатам, полученным для CA9. SCA9 также связывал DC насыщаемым образом (фиг.3C), и связывание sCA9 ингибировалось немеченым CA9 и фукоиданом (фиг.3D).
Пример 5
Данный пример иллюстрирует опосредованную CA9 доставку антигена к DC и переработку CA9.
Доставка антигенов к DC представляет собой раннюю стадию генерирования адаптивного иммунного ответа. Как CA9, так и sCA9 были способны связывать люциферазу и доставлять ее к свежим мышиным DC (фиг.4A). Комплекс sCA9 и люциферазы добавляли к DC при 4°C. Вестерн-блот анализ показал, что люцифераза связывалась с DC при образовании комплексов с sCA9 или CA9, но не когда одну люциферазу добавляли к DC. CA9, меченный FITC, связывался с поверхностью DC при 4°C, как показано конфокальной микроскопией (фиг.4B). Когда клетки нагревали до 37°C, меченый CA9 интернализировался DC.
После интернализации CA9, следующая стадия в активации адаптивного иммунного ответа представляет собой обработку CA9 DC. Мониторинг связывания с клеточной поверхностью проводили при 4°C. Для мониторинга статуса внутреннего CA9, клетки инкубировали при 37°C для обеспечения возможности протекания внутриклеточных процессов. CA9 на клеточной поверхности промывали и внутриклеточный CA9 измеряли зондированием клеточного лизата. При 37°C внутриклеточное содержание CA9 быстро увеличивалось, но он почти не выявлялся в пределах 4 часов (фиг.4C). Для оценки пути переработки CA9, мониторинг содержания внутриклеточного CA9 проводили в присутствии NH4Cl и MGl32, которые ингибируют соответственно лизосомы и протеосомы. Хотя и NH4Cl, и MGl32 ингибировали переработку CA9, MGl32 был более эффективен в ингибировании переработки CA9. Поэтому, внутриклеточный CA9 перерабатывается в первую очередь протеосомами, которые перерабатывают антигены для перекрестного представления.
Пример 6
Данный пример демонстрирует, что введение композиции, содержащей комплекс изолированного CA9 и антигена, стимулирует иммунный ответ на антиген in vivo, который является более выраженным, чем ответ, стимулированный композицией, содержащей антиген, но не содержащей CA9.
Для тестирования того, может ли CA9 стимулировать иммунный ответ, использовали модель мышиной меланомы для нацеливания на антиген меланомы, gp100. Рекомбинантный CA9 и gp100 подвергали образованию комплекса in vitro (CA9+gp100) и использовали для иммунизации мышей C57/BL6. Мышей подвергали антигенной стимуляции сингенными опухолями B16, устойчиво трансдуцированными gp100. У мышей, иммунизированных CA9+gp100, имелся значительно более медленный рост опухоли (фиг.5A) и более длительное выживание, по сравнению с любой из контрольных групп (p<0,05, данные не показаны). Иммунизация CA9+gp100 вызывала gp100-специфический ответ IFN-γ, измеренный с использованием анализа ELISPOT (фиг.5B), и ответ специфичных для опухоли цитотоксических Т клеток, измеренный с использованием анализа высвобождения 51Cr (фиг.5C). Таким образом, иммунный мониторинг продемонстрировал, что CA9 способен вызывать gp100-специфический клеточный иммунный ответ.
Подобно CA9 полной длины, sCA9 был способен стимулировать специфический иммунный ответ.На модели мышиной меланомы, описанной на фиг.5A-C, человеческий gp100 использовали в качестве мишени вакцины. Поэтому, сам gp100 вызывал незначительный противоопухолевый иммунный ответ; однако такая модель эффективно демонстрирует, что иммунность gp100 усиливается CA9. В подтверждающем исследовании, мышиный пептид gp100 (pep) оценивали в качестве мишени для генерирования специфического цитотоксического Т-клеточного ответа. Мышей иммунизировали DC, обработанными комплексом CA9 и пептидом (CA9+пептид) или sCA9 и пептидом (sCA9+пептид). У иммунизированных мышей развились pep-специфические цитотоксические T-лимфоциты (CTL), как определено с использованием анализа высвобожден 51Cr (фиг.5D).
Пример 7
Данный пример иллюстрирует экспрессию и сбрасывание CA9 в ответ на цитокины.
Сообщалось, что экспрессия CA9 в первичной почечной опухоли прогнозирует ответ на лечение IL2. В качестве скринингового исследования, мониторинг экспрессии CA9 в линии человеческих клеток R6 RCC после добавления кондиционированных сред (CM) из WBC, обработанных различными цитокинами (фиг.6A). CM использовали, поскольку цитокины обеспечивают терапевтическую выгоду стимуляцией иммунных клеток, а непосредственно не нацеливаясь на опухоли. Экспрессия CA9 увеличивалась в ответ на IL2 и INF-α, но не INF-γ.
Поскольку сообщалось, что CA9 прогнозирует ответ на лечение IL2, авторы задались вопросом, увеличивается ли сбрасывание CA9 кратковременно культурой эксплантатов RCC в ответ на IL2 (фиг.6B). Во всех 3 исследованных прозрачно-клеточных RCC, сброс CA9 увеличился в ответ на IL2. Одна папиллярная опухоль с отсутствием исходной экспрессии CA9 (опухоль 29) сбрасывала низкие уровни CA9 после обработки IL2. IL2 непосредственно наносили на хирургические образцы, которые содержали как опухолевые клетки, так и иммунные клетки.
Пример 8
В данном примере представлено подтверждающее доказательство того, что CA9 представляет собой прогностический маркер у пациентов с почечно-клеточной карциномой.
Экспрессию CA9 количественно определяли с использованием RT-PCR в реальном масштабе времени и замороженных RCC от 46 пациентов с клинически локализованными RCC, которые были подвергнуты хирургической резекции с намерением излечения. В таблице 1 суммированы клинические и патоморфологические признаки. При средней продолжительности наблюдения 13,7 месяцев, у пациентов с низкой экспрессией CA9 была значительно больше вероятность рецидива метастатического заболевания (p=0,0326).
| название | год | авторы | номер документа |
|---|---|---|---|
| АГЕНТЫ И СПОСОБЫ, ОСНОВАННЫЕ НА ПРИМЕНЕНИИ ДОМЕНА EDA ФИБРОНЕКТИНА | 2006 |
|
RU2430738C2 |
| Покрытые онколитические аденовирусы для противораковых вакцин | 2015 |
|
RU2695375C2 |
| ИММУНОСТИМУЛИРУЮЩИЕ КОМПОЗИЦИИ | 2016 |
|
RU2732118C2 |
| РЕКОМБИНАНТНЫЙ АДЕНОВИРУС, КОТОРЫЙ СОДЕРЖИТ РИБОЗИМ, ОПОСРЕДУЮЩИЙ ТРАНС-СПЛАЙСИНГ, И ПРОТИВОРАКОВЫЙ ТЕРАПЕВТИЧЕСКИЙ ГЕН, И ЕГО ПРИМЕНЕНИЕ | 2012 |
|
RU2575620C2 |
| КОМПОЗИЦИИ ВАКЦИН И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2010 |
|
RU2600798C2 |
| ТЕРАПИЯ ДЛЯ ЛЕЧЕНИЯ РАКА С ПОМОЩЬЮ ВНУТРИОПУХОЛЕВОГО И/ИЛИ ВНУТРИВЕННОГО ВВЕДЕНИЯ РЕКОМБИНАНТНОГО MVA, КОДИРУЮЩЕГО 4-1BBL (CD137L) И/ИЛИ CD40L | 2019 |
|
RU2830601C2 |
| ДОСТАВКА БИОМОЛЕКУЛ В КЛЕТКИ ИММУННОЙ СИСТЕМЫ | 2015 |
|
RU2739794C2 |
| ОНКОЛИТИЧЕСКИЕ АДЕНОВИРУСЫ, КОДИРУЮЩИЕ БИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА, А ТАКЖЕ СПОСОБЫ И ПРИМЕНЕНИЯ, СВЯЗАННЫЕ С НИМИ | 2016 |
|
RU2725799C2 |
| КОМБИНАЦИЯ МОДУЛЯТОРА ИММУННЫХ КОНТРОЛЬНЫХ ТОЧЕК И КОМПЛЕКСА, СОДЕРЖАЩЕГО ПРОНИКАЮЩИЙ В КЛЕТКУ ПЕПТИД, КАРГО-МОЛЕКУЛУ И ПЕПТИДНЫЙ АГОНИСТ TLR, ДЛЯ ПРИМЕНЕНИЯ В МЕДИЦИНЕ | 2017 |
|
RU2769314C1 |
| ВЕКТОР, КОЭКСПРЕССИРУЮЩИЙ МОЛЕКУЛЫ ДЛЯ ВАКЦИНАЦИИ И КОСТИМУЛИРУЮЩИЕ МОЛЕКУЛЫ | 2016 |
|
RU2714157C2 |









Группа изобретений относится к медицине, а именно к иммунологии, и может быть использована для стимуляции иммунного ответа. Для этого индивидууму вводят композицию, содержащую комплекс изолированного белка угольной ангидразы IX (СА9) и антигена, где изолированный белок СА9 и антиген нековалентно связаны друг с другом. Группа изобретений также относится к композиции, содержащей очищенную популяцию дендритных клеток, которые вступали в контакт с вышеуказанной композицией. Группа изобретений обеспечивает стимуляцию иммунного ответа путем усиления иммуногенности антигена за счет включения в комплекс белка СА9, который может функционировать в качестве шаперона. 3 н. и 15 з.п. ф-лы, 1 табл., 6 ил., 8 пр.
1. Способ стимуляции иммунного ответа против антигена у индивидуума, включающий введение индивидууму композиции, содержащей комплекс изолированного белка угольной ангидразы IX (СА9) и антигена, где изолированный белок СА9 и антиген нековалентно связаны друг с другом.
2. Способ по п.1, где комплекс образуется при температуре от 30°С до 42°С включительно.
3. Способ по п.1, где комплекс образуется при 37°С.
4. Способ по п.1, где антиген представляет собой опухолевый антиген.
5. Способ по п.1, где изолированный белок СА9 получают из среды для культуры клеток, в которой культивируются клетки, экспрессирующие белок СА9.
6. Способ по п.1, где опухолевый антиген и изолированный белок СА9 присутствуют в комплексе, образованном смешиванием изолированного белка СА9 с лизатом опухоли.
7. Способ по п.1, где композиция дополнительно содержит представляющие антиген клетки, которые вступали в контакт с комплексом перед введением индивидууму.
8. Способ по п.1, где иммунный ответ против антигена, стимулированный введением комплекса, включает клеточно-опосредованный иммунный ответ против антигена.
9. Способ по п.1, где антиген представляет собой антиген меланомы.
10. Способ по п.9, где антиген меланомы представляет собой gp 100.
11. Композиция для стимуляции иммунного ответа, содержащая комплекс изолированного белка угольной ангидразы IX (СА9) и антигена, где изолированный белок СА9 и антиген нековалентно связаны друг с другом.
12. Композиция по п.11, где антиген представляет собой опухолевый антиген.
13. Композиция по п.12, где опухолевый антиген и изолированный белок СА9 присутствуют в комплексе, образованном смешиванием изолированного белка СА9 с лизатом опухоли.
14. Композиция по п.11, где комплекс образуется при температуре от 30°С до 42°С включительно.
15. Композиция по п.11, дополнительно содержащая клетки, представляющие антиген.
16. Композиция по п.11, где антиген представляет собой антиген меланомы.
17. Композиция по п.16, где антиген меланомы представляет собой gp 100.
18. Композиция для стимуляции иммунного ответа, содержащая очищенную популяцию дендритных клеток, которые вступали в контакт с композицией по п.11.
| US 2005112134 A1, 26.05.2005 | |||
| KIM H.L | |||
| et al | |||
| Прибор, автоматически записывающий пройденный путь | 1920 |
|
SU110A1 |
| HERNANDEZ J.M | |||
| et al | |||
| Novel kidney cancer immunotherapy based on the granulocyte-macrophage colony-stimulating factor and carbonic | |||

