ОБЛАСТЬ ТЕХНИКИ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к рекомбинантному аденовирусу, который содержит рибозим, опосредующий транс-сплайсинг, и противораковый терапевтический ген, а также к применению указанного аденовируса, более конкретно, к рекомбинантному аденовирусу, содержащему полинуклеотид, кодирующий комплекс рибозима, опосредующего транс-сплайсинг, и HSVtk, который действует против рак-специфического гена, и противораковый терапевтический ген, к фармацевтической композиции для профилактики или лечения рака, которая содержит рекомбинантный аденовирус в качестве активного ингредиента, и к способу лечения рака, который включает введение пациенту рекомбинантного аденовируса или фармацевтической композиции.
УРОВЕНЬ ТЕХНИКИ ИЗОБРЕТЕНИЯ
Рак является основной причиной смерти в Корее и представляет собой неизлечимую болезнь, которая еще не побеждена, несмотря на многочисленные исследования, проводимые в этом направлении. Традиционные методы противораковой терапии включают хирургию, химиотерапию и радиотерапию. Однако каждый из указанных методов имеет много ограничений и в связи с этим в последние годы проводятся исследования по разработке других терапевтических методов, в основе которых лежит концепция, отличающаяся от концепции вышеуказанных методов. Среди новых методов активно разрабатывается метод генной терапии.
В рамках изобретения термин “генная терапия” относится к способу лечения рекомбинантными методами наследственных или приобретенных генетических аномалий, которые плохо поддаются лечению традиционными методами. А именно, генная терапия включает в себя введение в организм человека генетических материалов, таких как ДНК и РНК, обеспечивающих экспрессию терапевтических белков или ингибирование экспрессии конкретных белков, с целью лечения или профилактики наследственных или приобретенных генетических дефектов, вирусных заболеваний или хронических заболеваний, таких как раковые или сердечно-сосудистые заболевания. Генная терапия позволяет фундаментально излечивать заболевания путем анализа их причин на генетическом уровне и, следовательно, предположительно может использоваться для лечения неизлечимых болезней и в качестве альтернативы традиционным терапевтическим методам.
Противораковые методы генной терапии можно подразделить на иммунологический метод генной терапии, который индуцирует иммунные ответы в организме человека, и метод генной терапии, основанный на использовании генов, способных непосредственно разрушать или уничтожать раковые клетки. В последнем случае важную роль играют векторы, обеспечивающие доставку генов в клетки и экспрессию указанных генов. Аденовирусный вектор известен как один из векторов, представляющих наибольший интерес с точки зрения генной терапии, поскольку он обеспечивает высокоэффективную доставку генов и обладает способностью доставлять гены в недифференцированные клетки, а также вследствие простоты получения исходного вирусного препарата с высоким титром.
Аденовирусные векторы, обычно используемые в генной терапии, конструируют путем делеции ряда генов, необходимых для репликации, и введения промотора цитомегаловируса (CMV), или вируса саркомы Рауса (RSV), обладающих высокой промоторной активностью, позволяющей обеспечить высокоэффективную экспрессию терапевтического белка in vivo.
В последние годы разрабатывается метод терапии, направленный на раковые ткани, который позволяет уменьшить побочные эффекты, связанные с тем, что ряд целевых генов, используемых в генной терапии, также экспрессируется в нормальных клетках, характеризующихся интенсивным клеточным делением (Fukuzawa et al., Cancer Res 64: 363-369, 2004). В указанном методе терапии предлагается использовать тканеспецифичный промотор вместо промотора CMV или RSV, однако, несмотря на повышение специфичности данный метод не находит практического применения из-за низкой терапевтической эффективности.
Чтобы преодолеть вышеуказанный недостаток, недавно были предприняты исследования с целью разработки тканеспецифичного аденовируса для противораковой терапии с использованием факторов, отличных от тканеспецифичного промотора. Например, были разработаны методы с использованием рибозима, опосредующего транс-сплайсинг, и т.п.
В процессе исследований по разработке тканеспецифичного аденовируса для противораковой терапии с использованием рибозима, опосредующего транс-сплайсинг, было обнаружено, что рибозим интрона группы I из Tetrahymena thermophila может соединять два отдельных транскрипта друг с другом посредством реакции транс-сплайсинга не только in vitro, но и в бактериальных и человеческих клетках.
А именно, рибозим, опосредующий транс-сплайсинг и расположенный на указанном интроне группы I, может быть направлен на транскрипт связанного с заболеванием гена, или на конкретную РНК, специфически экспрессирующуюся в пораженных клетках, инициируя перепрограммирование РНК с восстановлением до нормальной РНК, или замену генного транскрипта на новый транскрипт терапевтического гена. Таким образом, можно ожидать что метод, основанный на применении рибозима, опосредующего транс-сплайсинг, может представлять собой болезнь-специфичный и безопасный метод генной терапии. Кроме того, рибозим, опосредующий транс-сплайсинг, может значительно повысить терапевтический эффект, поскольку он может удалять болезнь-специфическую РНК и индуцировать экспрессию целевого терапевтического генного продукта.
Поскольку известно, что рибозим, опосредующий транс-сплайсинг и направленный на hTERT (обратная транскриптаза теломеразы человека), способен специфически воздействовать на раковую ткань, в недавних исследованиях были предприняты попытки разработать противораковые терапевтические средства на основе данного рибозима, опосредующего транс-сплайсинг, однако положительные результаты еще не опубликованы.
Известные терапевтические гены, подходящие для использования в генной терапии, включают гены тимидинкиназы простого герпеса (далее обозначаемой HSVtk), цитозиндеаминазы E. coli (CD) и пуриннуклеозидфосфорилазы E. coli (далее обозначаемой PNP). В методах генной терапии, основанных на применении указанных генов и называемых “методы ген-направленной ферментно-пролекарственной терапии” (GDEPT), обычно используют пролекарства. Подходящие для использования в GDEPT пролекарства включают в себя ганцикловир (GCV), 5-фторурацил (5-FU) и 6-метилпурин-2-дезоксирибозид (6-MeP-dR), которые используют вместе с HSV-TK, CD и PNP, соответственно. Нетоксичные пролекарства превращаются в цитотоксические средства в результате введения генов и активируются, в основном, путем фосфорилирования. Преимуществом генной терапии, использующей суицидные гены, является заметное проявление так называемого эффекта ″свидетеля″, включающего в себя уничтожение соседних клеток под действием лекарств, активируемых в клетках, в которые введены указанные гены. Указанная генная терапия, возможно, является лучшей стратегией, которая может применяться в состоянии, характеризующемся низкой эффективностью введения генов.
Хорошо известно, что HSVtk можно использовать в указанном методе генной терапии. А именно, среди методов, основанных на применении суицидных генов, наиболее широко используется метод, основанный на применении системы HSVtk/GCV, раскрытый в WO 90/07936, US 5837510, US 5861290, WO 98/04290, WO 97/37542 и US 5631236. Клетки, которые экспрессируют ген HSVtk, могут фосфорилировать ганцикловир (GCV), в результате чего индуцируется гибель клетки, обусловленная нарушением репликации ДНК. В настоящее время данный способ участвует в 30 или более видах клинических испытаний методов генной терапии, направленных на разные виды раковых заболеваний человека. Однако данный способ также имеет недостатки, заключающиеся в том, что уничтожаться могут только пролиферирующие клетки, эффект ″свидетеля″ не является достаточным и может возникать проблема с цитотоксичностью, если пролекарство используется в больших количествах. В связи с этим активно проводятся исследования способов лечения рака, в которых используются два или более пролекарств, что позволяет повысить эффект уничтожения клеток и эффект ″свидетеля″.
Между тем, PD-1 (белок программируемой гибели 1) представляет собой трансмембранный белок типа I размером 55 кДа, кодируемый геном суперсемейства Ig, и известный как молекула, оказывающая со-ингибиторное действие на T-клетки. Другими словами, PD-1 является одной из со-ингибиторных молекул семейства рецепторов CD28 (например, CD28, CTLA-4, ICOS и BTLA), которые экспрессируются на активированных B-клетках, T-клетках и клетках костного мозга. Лиганды PD-1 включают в себя PD-L1 и PD-L2, которые, как известно, осуществляют понижающую регуляцию активации T-клеток после связывания с PD-1. PD-1 не экспрессируется на обычных наивных T-клетках, однако после активации T-клеток наблюдается индуцируемая экспрессия PD-1. Кроме того, PD-L1 обнаруживается на высоком уровне при разных раковых заболеваниях человека, а взаимодействие PD-1 и PD-L1 приводит к генерированию сигналов, оказывающих стимуляторное или ингибиторное действие на T-клетки. Другими словами, взаимодействие PD-1 с PD-L1 вызывает уменьшение уровня опухоль-инвазивных лимфоцитов и пролиферации, опосредованной T-клеточным рецептором, а также способствует уклонению опухолевых клеток от иммунологического надзора. Как следствие, в последние годы проводятся исследования по лечению рака путем блокирования сигнального пути PD-1 или PD-L1, приводящего к эффективной индукции противораковых иммунных ответов.
ОПИСАНИЕ
ТЕХНИЧЕСКАЯ ЗАДАЧА
Авторы настоящего изобретения провели обширные исследования с целью разработки метода генной терапии для лечения рака, который характеризуется повышенной тканевой специфичностью и улучшенной терапевтической эффективностью. В результате авторы настоящего изобретения обнаружили, что рекомбинантный аденовирус, содержащий полинуклеотид, который кодирует специфичный к раковой ткани комплекс рибозима, опосредующего транс-сплайсинг, и HSVtk (тимидинкиназы вируса простого герпеса человека), и противораковый терапевтический ген sPD-1, может оказывать превосходный терапевтический эффект, специфичный для раковой ткани, и, кроме того, его применение может значительно уменьшить побочные эффекты, вызываемые генной терапией, и тем самым завершили настоящее изобретение.
ТЕХНИЧЕСКОЕ РЕШЕНИЕ
Целью настоящего изобретения является получение рекомбинантного аденовируса, содержащего: полинуклеотид, кодирующий комплекс рибозима, опосредующего транс-сплайсинг, и HSVtk, который действует против рак-специфического гена; и противораковый терапевтический ген.
Другой целью настоящего изобретения является получение фармацевтической композиции для профилактики или лечения рака, которая содержит рекомбинантный аденовирус в качестве активного ингредиента.
Следующей целью настоящего изобретения является разработка способа лечения рака у животных, в том числе, у человека, где указанный способ включает введение пациенту рекомбинантного аденовируса или фармацевтической композиции.
БЛАГОПРИЯТНЫЕ ЭФФЕКТЫ
Рекомбинантный аденовирус согласно настоящему изобретению обладает селективностью в отношении раковых клеток, обусловленную рибозимом, опосредующим транс-сплайсинг, который действует против рак-специфического гена, и повышенной противораковой активностью, обусловленной противораковым терапевтическим геном. Следовательно, его можно эффективно использовать для профилактики и лечения рака.
ОПИСАНИЕ ЧЕРТЕЖЕЙ
На фиг.1 приведено схематическое изображение рибозима, опосредующего транс-сплайсинг, (mTERT-TR)-HSVtk, специфичного к мышиному TERT. Участок, определяющий специфичность к транскрипту мышиного TERT (mTERT), указан как последовательность вблизи участка сплайсинга. Рибозим, опосредующий транс-сплайсинг, специфически распознает РНК mTERT посредством антисмысловой последовательности РНК mTERT и расщепляет РНК mTERT по 3′-концу IGS (внутренней адапторной последовательности). 5′-Конец HSVtk-кодирующей РНК рибозима, опосредующего транс-сплайсинг, соединяется с расщепленным концом РНК mTERT. Возможные нуклеотидные связи между mTERT-специфичной мРНК и рибозимом указаны вертикальными линиями.
На фиг.2 показано влияние мышиного TERT-TR на регуляцию экспрессии аденовирусного гена HSVtk. На фиг.2(a) показаны результаты экспрессии HSVtk аденовирусом Ad5mTR, содержащим делецию E1/E3, под контролем мышиного TERT-TR. Ad5MOCK представляет собой регуляторный элемент аденовируса, содержащего делецию E1/E3. На фиг.2(b) показаны результаты, полученные путем инокуляции клеток CT26 (3×103) в 96-луночный планшет с последующим воздействием на клетки разных MOI Ad5MOCK или Ad5mTR в присутствии 100 мкМ GCV. На фигуре также показаны результаты измерения цитотоксичности с использованием набора для анализа пролиферации клеток через 3 дня после инфицирования. Данные выражают в виде среднего значения ± стандартное отклонение для трех независимых экспериментов. На фиг.2(c) показаны результаты, полученные путем введения 5×108 БОЕ Ad5MOCK или Ad5mTR мышам BALB/c, введения GCV (75 мг/кг) внутрибрюшинно мышам два раза в день и инокуляции мышам клеток CT26 (1×106) подкожно в оба бока. Данные выражают в виде среднего значения ± SEM (n=5).
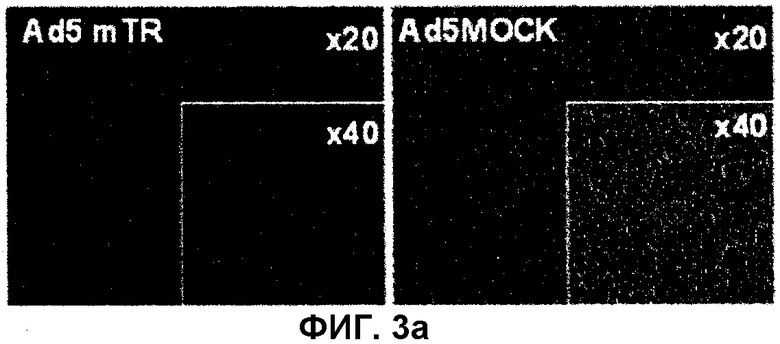
На фиг.3 показан улучшенный DC-опосредованный антиген-презентирующий эффект, индуцированный HSVtk. На фиг.3(a) Ad5MOCK или Ad5mTR вводят в опухоль E.G7, присутствующую у мышей C57/BL6. Через 4 дня опухоли собирают и подвергают гистологическому анализу путем окрашивания H&E. Фиг.3(b): через 2 дня после введения аденовируса DC (5×104) собирают из дренированных лимфоузлов, находящихся вблизи опухоли E.G7, методом хроматографии MACS с использованием микрогранул против CD11c и затем культивируют совместно с OVA-специфичными T-клетками CD8, полученными из трансгенных мышей OT-1. Через 72 часа культуральный супернатант собирают и измеряют в нем количество IFN-γ. DC-Ad5MOCK обозначает DC, собранные у мышей, которым был введен Ad5MOCK, и CD-Ad5mTR обозначает DC, собранные у мышей, которым был введен Ad5mTR. Данные выражают в виде среднего значения ± стандартное отклонение.
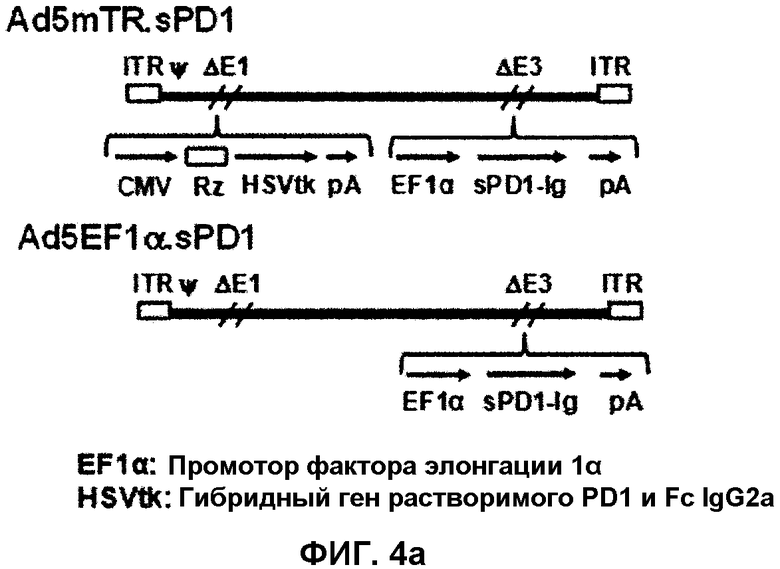
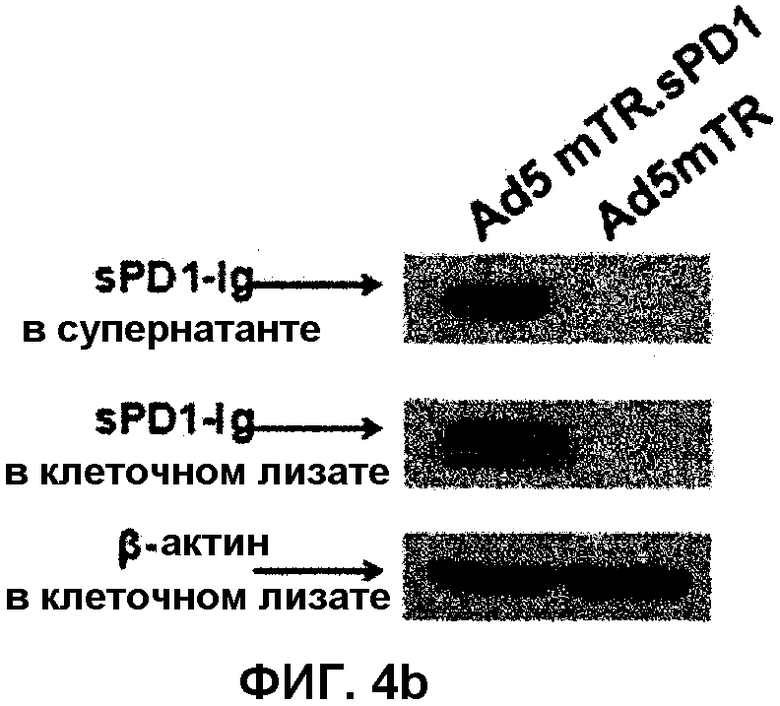
На фиг.4 показан способ получения двухмодульного аденовируса, который экспрессирует mTERT-TR-контролируемый HSVtk и sPD1-Ig. Фиг.4(a): Ad5mTR.sPD1 кодирует mTERT-TR-HSVtk под контролем промотора CMV в участке E1 и sPD1-Ig под контролем EF1α в участке E3. Фиг.4(b): клетки HEK293 (4×105) инфицируют 2 MOI аденовируса в течение 48 часов, разрушают буфером RIPA и затем анализируют методом иммуноблоттинга. Экспрессию sPD1-Ig с использованием Ad5mTR.sPD1 (80 мкг белка на линию) анализируют с помощью антитела против PD1. Фиг.4(c): в клетках CT26 анализируют ферментативную активность HSVtk в фосфорилированном GCV путем измерения накопления меченного радиоизотопом PCV (1 мкКи/мл). Данные выражают в виде среднего значения ± стандартное отклонение для трех независимых экспериментов. Фиг.4(d): супернатант, собранный из культуры клеток, инфицированных 10 MOI аденовируса, смешивают с совместной культурой клеток MC38/OVA (1×104) и OVA-специфичных T-клеток CD8 (1×106), полученных из мышей OT-1. Через 72 часа культуральный супернатант собирают и измеряют в нем количество IFN-γ. Данные выражают в виде среднего значения ± стандартное отклонение.
На фиг.5 показана экспрессия PD-L1 на поверхности клеток рака ободочной и прямой кишки мыши CT26. Фиг.5(a): из клеток каждого типа получают общую РНК и детектируют транскрипт PD-L1 методом RT-PCR. В качестве контроля используют мышиные клетки рака печени (Hepa-1). Фиг.5(b): белок PD-L1, экспрессирующийся на поверхности клеток каждого типа, анализируют методом проточной цитометрии.
На фиг.6 показана противоопухолевая активность Ad5mTR.sPD1 in vivo и in vitro. Фиг.6(a): клетки CT26 инфицируют Ad5MOCK, Ad5mTR или Ad5mTR.sPD1 с разными значениями MOI в присутствии 100 мкМ GCV. Через 3 дня измеряют цитотоксичность с помощью метода, показанного на фиг.1(d). Данные выражают в виде среднего значения ± стандартное отклонение для трех независимых экспериментов. Фиг.6(b): мышам BALB/c подкожно инокулируют клетки CT26 (1×106) с обоих боков. Опухоль обрабатывают 5×108 PFU Ad5MOCK, Ad5EF1α.sPD1, Ad5mTR или Ad5mTR.sPD1. Мышам внутрибрюшинно вводят GCV (75 мг/кг) два раза в день. Объем опухоли измеряют в указанные моменты времени и регистрируют. Данные выражают в виде среднего значения ± SEM (n=10).
На фиг.7 показано, что ингибирование роста опухоли под действием sPD1-Ig опосредуется T-клетками CD8. Фиг.7(a): в подкожную опухоль CT26 мышей BALB/c вводят Ad5MOCK, Ad5mTR или Ad5mTR.sPD1 (левая панель). Чтобы определить участие T-клеток CD8 в противоопухолевой активности sPD1-Ig, в подкожную опухоль CT26 мышей BALB/c вводят Ad5MOCK, Ad5mTR или Ad5mTR.sPD1 (правая панель). За 2 дня до введения вируса и с 5-дневными интервалами после введения вируса внутривенно вводят антитело против CD8 2.43α (500 мкг), чтобы удалить T-клетки CD8 (правая панель). Стрелкой указано время введения антитела. Объем опухоли измеряют в указанные моменты времени и регистрируют. Фиг.7(b): ингибирование роста опухоли в отсутствие или присутствии антитела против CD8 2.43α рассчитывают относительно значения, полученного с использованием Ad5MOCK примерно через 12 дней. Данные выражают в виде среднего значения ± SEM.
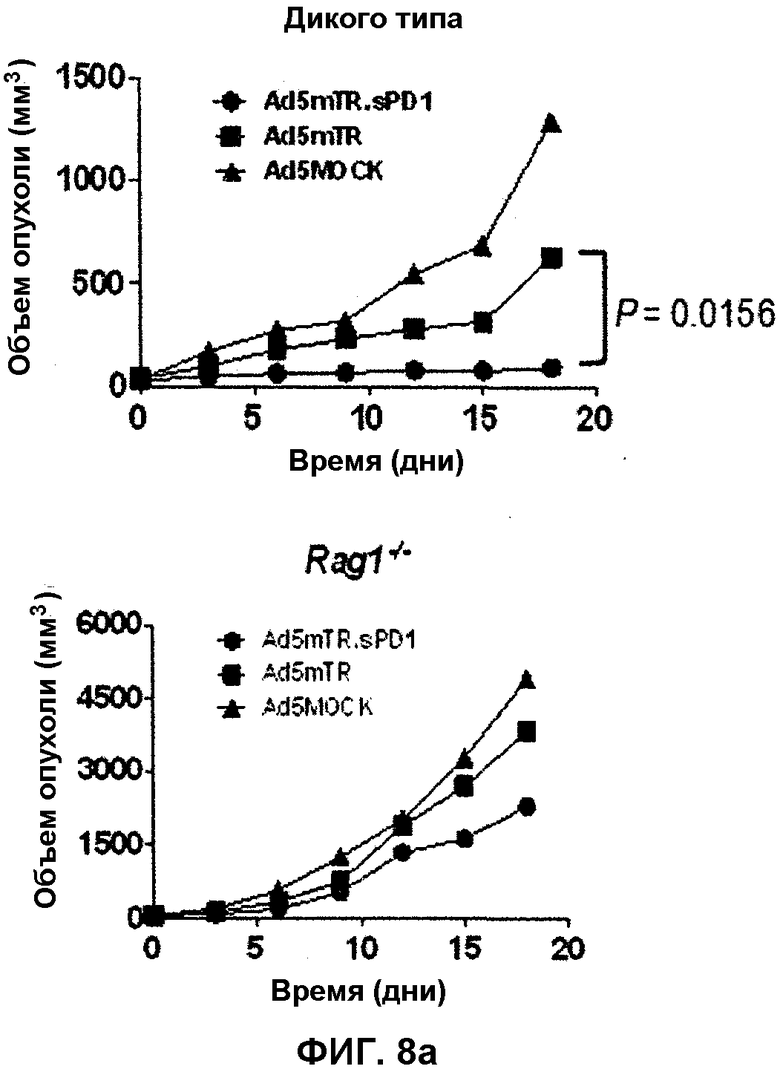
Фиг.8 демонстрирует ингибиторный эффект sPD1-Ig на рост опухоли у мышей Rag1-/-, истощенных по T- и B-клеткам. Фиг.8(a): в опухоль E.G7, введенную подкожно мышам C57/BL6, вводят Ad5MOCK, Ad5mTR или Ad5mTR.sPD1 (левая панель). Чтобы измерить вклад лимфоцитов в противоопухолевую активность sPD1-Ig, в опухоль E.G7, введенную подкожно мышам Rag1-/-, вводят Ad5MOCK, Ad5mTR или Ad5mTR.sPD1 (правая панель). Объем опухоли измеряют в указанные моменты времени и регистрируют. Фиг.8(b): уменьшение объема опухоли у мышей Rag1-/- или у мышей дикого типа рассчитывают относительно значения, полученного с использованием Ad5MOCK через 12 дней. Фиг.8(c): в опухоль E.G7, присутствующую у мышей Rag1-/- (1×106 клеток), вводят 5×108 БОЕ Ad5MOCK, Ad5mTR или Ad5mTR.sPD1 два раза с интервалом в 7 дней. T-клетки CD8 (5×104), полученные от мышей OT-1, вводят внутривенно одновременно с первой инъекцией аденовируса. Объем опухоли измеряют в указанные моменты времени и регистрируют. Данные выражают в виде среднего значения ±SEM (n=6). Фиг.8(d): чтобы провести in vitro анализ иммунных клеток, клетки опухоли E.G7 мыши Rag1-/- культивируют совместно с T-клетками CD8 мыши OT-1 и подходящим аденовирусом. Через 8 дней после первого введения аденовируса PBMC (мононуклеарные клетки периферической крови) собирают из глазных вен мышей. Методом проточной цитометрии определяют число T-клеток CD8 OT-1 в общей массе жизнеспособных клеток и отношение TCR Vα2+, Vβ5+, T-клеток (клеток OT-1).
Фиг.9 демонстрирует ингибиторное действие Ad5mTR.sPD1 на вторичную опухоль. Фиг.9(a): опухоль CT26 трансплантируют подкожно мышам BALB/c, после чего мышам вводят 5×108 БОЕ Ad5mTR.sPD1 три раза с интервалами в 3 дня. Через 2 недели после первой инъекции 1×106 опухолевых клеток вводят в противоположный бок. Через 1 день после первой инъекции вируса в течение 12 дней интраперитонеально вводят GCV (75 мг/кг). Объем опухоли измеряют с 3-дневными интервалами. Фиг.9(b): опухоль E.G7 трансплантируют подкожно мышам C57/BL6 и затем проводят процедуру, описанную выше для фиг.9(a). Фиг.9(c): через 7 дней после инокуляции вторичной опухоли измеряют объем опухоли у мышей, обработанных Ad5mTR.sPD1, и у необработанных мышей. Данные выражают в виде среднего значения ± стандартное отклонение.
НАИЛУЧШИЙ СПОСОБ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
В одном аспекте настоящее изобретение относится к рекомбинантному аденовирусу, содержащему: полинуклеотид, кодирующий комплекс рибозим, опосредующий транс-сплайсинг-HSVtk (тимидинкиназа вируса простого герпеса человека), который действует против мРНК TERT (обратная транскриптаза теломеразы) (SEQ ID NO: 1), представляющий собой рак-специфический ген; и противораковый терапевтический ген.
В рамках изобретения термин "рак-специфический ген" относится к гену, который экспрессируется конкретно в раковых клетках, или к гену, который характеризуется специфической экспрессией в раковых клетках или значительной сверхэкспрессией в раковых клетках. Рак-специфический ген может иметь характеристики, на которые может действовать рибозим согласно настоящему изобретению. Такой рак-специфический ген может включать, без ограничения, мРНК TERT (обратной транскриптазы теломеразы), мРНК AFP (альфа-фетопротеина), мРНК CEA (карциноэмбрионального антигена), мРНК PSA (простатоспецифического антигена) или мРНК CKAP2 (цитоскелет-ассоциированного белка 2).
В рамках изобретения термин "TERT (обратная транскриптаза теломеразы)" относится к одному из наиболее важных ферментов, который регулирует иммортализацию и способность к пролиферации раковых клеток, а также образует теломеры, ингибирующие старение клеток путем защиты хромосомных концов. В нормальных клетках длина теломеров понемногу уменьшается по мере деления клеток, в результате чего генетический материал утрачивается и клетки умирают. Однако в раковых клетках указанный фермент непрерывно наращивает теломеры и, следовательно, клетки не умирают. Кроме того, этот фермент известен как существенное препятствие для лечения рака, поскольку он непосредственно участвует в иммортализации раковых клеток. Активность теломеразы в зародышевых клетках, гематопоэтических клетках и раковых клетках составляет 80-90%, тогда как в нормальных клетках, окружающих раковые клетки, теломеразная активность отсутствует. В свете вышесказанного, TERT может быть непосредственно направлен, без ограничения, на противораковый терапевтический ген согласно настоящему изобретению.
В рамках изобретения термин "рибозим" относится к ферментативно активной молекуле РНК, способной осуществлять транс-сплайсинг и аутосплайсинг. В целях настоящего изобретения рибозим можно использовать для ингибирования активности рак-специфического гена посредством реакции транс-сплайсинга, что обуславливает селективный противораковый эффект рибозима. Кроме того, он может экспрессироваться в сочетании с противораковым терапевтическим геном, активируя противораковый терапевтический ген. Следовательно, в настоящем изобретении можно использовать любой рибозим, при условии, что он способен инактивировать рак-специфический ген и активировать противораковый терапевтический ген. Предпочтительно, он может включать, без ограничения, либо рибозим Rib67, который представляет собой hTERT-специфический рибозим группы I, опосредующий транс-сплайсинг, способность которого к транс-сплайсингу подтверждается распознаванием рак-специфичной TERT (обратной транскриптазы теломеразы), либо рибозим, который кодируется полинуклеотидом, имеющим нуклеотидную последовательность SEQ ID NO: 2.
В рамках изобретения термин "HSVtk (тимидинкиназа вируса простого герпеса)" относится к тимидинфосфорилазе, полученной из вируса простого герпеса. Ген этого фермента представляет собой типичный лекарство-активирующий ген, обеспечивающий превращение нетоксичного пролекарства в токсичную форму, способную уничтожить клетки, трансфицированные указанным геном. В целях настоящего изобретения ген HSVtk можно использовать в качестве противоракового терапевтического гена, который экспрессируется в сочетании с рибозимом, проявляя противораковую активность. Указанный ген HSVtk предпочтительно может иметь нуклеотидную последовательность, описанную в SEQ ID NO: 3, и может быть выбран из генов, описанных в Genbank под номерами доступа AAP13943, P03176, AAA45811, P04407, Q9QNF7, KIBET3, P17402, P06478, P06479, AAB30917, P08333, BAB84107, AAP13885, AAL73990, AAG40842, BAB11942, NP_044624, NP_044492, CAB06747 и т.п., но не ограничивается ими.
В рамках изобретения термин "противораковый терапевтический ген" относится к полинуклеотидной последовательности, кодирующей полипептид, оказывающий терапевтический эффект при экспрессии в раковых клетках. В соответствии с настоящим изобретением, противораковая активность противоракового терапевтического гена проявляется при экспрессии этого гена отдельно или в сочетании с рибозимом. Примеры противоракового терапевтического гена, подходящие для применения в настоящем изобретении, включают, без ограничения, лекарство-активирующий ген, проапоптотический ген, цитостатический ген, цитотоксический ген, опухоль-супрессорный ген, антигенный ген, ген цитокина, анти-ангиогенный ген и т.п. В настоящем изобретении указанные противораковые терапевтические гены могут использоваться по отдельности или в сочетании из двух или более генов.
В рамках изобретения термин “лекарство-активирующий ген” относится к гену фермента, который превращает нетоксичное пролекарство в токсичную форму. Предпочтительно указанный ген включает суицидный ген, поскольку клетки, трансфицированные таким геном, погибают. То есть, при системном введении пролекарства, которое является нетоксичным для нормальных клеток, оно превращается в токсичные метаболиты только в раковых клетках под действием лекарство-активирующего гена, изменяющего чувствительность к лекарству и таким образом уничтожающего раковые клетки. Типичные примеры лекарство-активирующего гена, подходящие для применения в настоящем изобретении, включают, без ограничения, ген HSV-tk (тимидинкиназы вируса простого герпеса), ганцикловир, ген цитозиндеаминазы (CD) E. coli, 5-фторцитозин (5-FC) и др.
В рамках изобретения термин "проапоптотический ген" относится к нуклеотидной последовательности, которая при экспрессии индуцирует программируемую гибель клетки. Примеры проапоптотического гена, подходящие для применения в настоящем изобретении, включают, без ограничения, p53, аденовирус E3-11.6K (полученный из Ad2 и Ad5) или аденовирус E3-10.5K (полученный из Ad), ген аденовируса E4, ген пути p53 и ген, кодирующий каспазу.
В рамках изобретения термин “цитостатический ген” относится к нуклеотидной последовательности, которая экспрессируется в клетках, останавливая клеточный цикл. Типичные примеры цитостатического гена, подходящие для применения в настоящем изобретении, включают, без ограничения, p21, ген ретинобластомы, гибридный ген белка E2F-Rb, гены, кодирующие циклин-зависимый ингибитор киназы (например, p16, p15, p18 и p19), гены гомеобокса, блокирующие рост (GAX) и т.п.
В рамках изобретения термин “цитотоксический ген” относится к нуклеотидной последовательности, которая при экспрессии в клетках оказывает цитотоксический эффект. Примеры цитотоксического гена, подходящие для применения в настоящем изобретении, включают, без ограничения, нуклеотидные последовательности, кодирующие экзотоксин Pseudomoas, лизиновый токсин, токсин дифтерии и т.п.
В рамках изобретения термин “опухоль-супрессорный ген” относится к нуклеотидной последовательности, которая может экспрессироваться в клетках-мишенях, ингибируя опухолевый фенотип или вызывая гибель клеток. Примеры опухоль-супрессорного гена, подходящие для применения в настоящем изобретении, включают, без ограничения, ген фактора некроза опухоли α (TNF-α), ген p53, ген APC, ген DPC-4/Smad4, ген BRCA-1, ген BRCA-2, ген WT-1, ген ретинобластомы, ген MMAC-1, ген белка аденоматозного полипоза толстой кишки, удаленный ген карциномы толстой кишки (DCC), ген MMSC-2, ген NF-1, опухоль-супрессорный ген ENT, расположенный в хромосоме 3p21.3, ген MTS1, ген CDK4, ген NF-1, ген NF-2, ген VHL, ген sPD-1 (растворимого белка программируемой гибели 1) и др.
В рамках изобретения термин "sPD-1 (растворимый белок программируемой гибели 1)" относится к внеклеточному домену PD-1 (белка программируемой гибели 1), известному как соингибиторная молекула T-клеток, которая является членом семейства молекул иммуноглобулинов. Как правило, взаимодействие PD-1 и PD-L1 индуцирует уменьшение уровня опухоль-инвазивных лимфоцитов и пролиферации, опосредованной T-клеточным рецептором, а также способствует уклонению опухолевых клеток от иммунологического надзора. Однако проведенные в последнее время исследования показали, что sPD-1, который является растворимой формой PD-1, может эффективно индуцировать противораковые иммунные ответы путем ингибирования иммунного надзора, индуцируемого взаимодействием между PD-1 и PD-L1. Следовательно, в соответствии с настоящим изобретением ген sPD-1 можно конъюгировать с рак-специфическим геном посредством транс-сплайсинга, опосредуемого рибозимом согласно настоящему изобретению, и использовать в качестве терапевтического гена, направленного против раковых клеток. Предпочтительно, ген sPD-1 может содержать, без ограничения, нуклеотидную последовательность, описанную в SEQ ID NO: 4.
В рамках изобретения термин "антигенный ген" относится к нуклеотидной последовательности, которая экспрессируется в клетках-мишенях, продуцируя клеточный поверхностный антигенный белок, который может распознаваться иммунной системой. Примеры антигенного гена, подходящие для применения в настоящем изобретении, включают, без ограничения, гены карциноэмбрионального антигена (CEA), p53 и т.п.
В рамках изобретения термин "ген цитокина" относится к нуклеотидной последовательности, которая экспрессируется в клетках, продуцируя цитокин. Примеры гена цитокина, подходящие для применения в настоящем изобретении, включают, без ограничения, гены GM-CSF, интерлейкинов (IL-1, IL-2, IL-4, IL-12, IL-10, IL-19 и IL-20), интерферонов α, β и γ (интерферон α-2b), а также гибридов, таких как интерферон α-2α-1.
В рамках изобретения термин "антиангиогенный ген" относится к нуклеотидной последовательности, которая при экспрессии в клетках обеспечивает высвобождение из них антиангиогенных факторов. Примеры антиангиогенного гена, подходящие для применения в настоящем изобретении, включают, без ограничения, ген ангиостатина, гены ингибиторов фактора роста эндотелия сосудов (VEGF), ген эндостатина и т.п.
В рамках изобретения термин "аденовирус" имеет такое же значение, как и аденовирусный вектор, и относится к члену семейства Adenoviridae. Adenoviridae включает в себя все аденовирусы животных рода Mastadenovirus. В частности, человеческие аденовирусы включают подрода A-F и их отдельные серотипы. Подрода A-F включают, без ограничения, аденовирусы человека, относящиеся к типам 1, 2, 3, 4, 4a, 5, 6, 7, 8, 9, 10, 11 (Ad11A и Ad11P), 12, 13, 14, 15, 16, 17, 18, 19, 19a, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 34a, 35, 35p, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48 и 91.
В другом аспекте настоящее изобретение относится к фармацевтической композиции для профилактики или лечения рака, которая содержит вышеописанный рекомбинантный аденовирус в качестве активного ингредиента.
Рекомбинантный аденовирус в соответствии с настоящим изобретением содержит полинуклеотид, кодирующий комплекс рибозима, опосредующего транс-сплайсинг, и HSVtk, который действует против рак-специфического гена TERT, и противораковый терапевтический ген. Следовательно, при введении рекомбинантного аденовируса в раковые клетки, в которых экспрессируется TERT, HSVtk может высвобождаться из комплекса рибозима, опосредующего транс-сплайсинг, и HSVtk, под действием TERT, после чего HSVtk может проявлять цитотоксичность и индуцировать активность противоракового терапевтического гена против раковых клеток. Таким образом, рекомбинантный аденовирус может функционировать как противораковое терапевтическое средство. Однако если рекомбинантный аденовирус попадает в нормальные клетки, в которых TERT не экспрессируется, HSVtk не проявляет цитотоксичность, поскольку HSVtk не высвобождается из комплекса рибозима, опосредующего транс-сплайсинг, и HSVtk. Соответственно, рекомбинантный аденовирус согласно настоящему изобретению обладает высокой селективностью в отношении раковых клеток и, следовательно, может использоваться для противораковой терапии в безопасном способе.
В рамках изобретения термин "рак" относится к клеткам или тканям, которые характеризуются аномальной пролиферацией клеток вследствие нарушений механизмов регуляции деления, дифференциации и смерти, а также способностью проникать в окружающие ткани и органы, образуя массу, с разрушением или изменением существующих структур. Примеры указанного рака включают, без ограничения, рак поджелудочной железы, рак молочной железы, опухоли мозга, карциному головы и шеи, меланому, миелому, лейкоз, лимфому, рак печени, рак желудка, рак толстой кишки, рак кости, рак матки, рак яичника, рак прямой кишки, рак пищевода, рак тонкого кишечника, рак анального отверстия, карциному фаллопиевых труб, карциному эндометрия, рак шейки матки, карциному влагалища, рак вульвы, болезнь Ходжкина, рак мочевого пузыря, рак почки, рак мочеточника, почечноклеточную карциному, карциному почечной лоханки и опухоли центральной нервной системы.
В рамках изобретения термин “профилактика” относится к любому действию, которое приводит к ингибированию рака или к замедлению развития рака путем введения рекомбинантного аденовируса или композиции согласно настоящему изобретению.
В рамках изобретения термин “лечение” относится к любому действию, которое приводит к облегчению или благоприятному изменению течения ракового заболевания путем введения рекомбинантного аденовируса или композиции согласно настоящему изобретению.
Кроме того, фармацевтическая композиция согласно настоящему изобретению может дополнительно содержать фармацевтически приемлемый носитель, наполнитель или разбавитель.
Примеры фармацевтически приемлемых носителей, наполнителей или разбавителей, подходящих для применения в фармацевтической композиции согласно настоящему изобретению, включают такие средства, как лактоза, декстроза, сахароза, сорбит, маннит, ксилит, эритритол, мальтитол, крахмал, аравийская камедь, альгинат, желатин, фосфат кальция, силикат кальция, карбонат кальция, целлюлоза, метилцеллюлоза, поливинилпирролидон, вода, метилгидроксибензоат, пропилгидроксибензоат, тальк, стеарат магния, минеральное масло и др.
Фармацевтическую композицию согласно настоящему изобретению можно получить с помощью традиционных способов в виде пероральных лекарственных форм, включающих в себя порошки, гранулы, таблетки, капсулы, суспензии, эмульсии, сиропы и аэрозоли, препаратов для наружного применения, свечей и стерильных растворов для инъекций. Композиция согласно настоящему изобретению может содержать традиционно используемые разбавители или среды для лекарств, такие как наполнители, сухие разбавители, связующие средства, увлажняющие средства, дезинтегрирующие средства, поверхностно-активные вещества и др. Твердые композиции для перорального введения включают таблетки, пилюли, порошки, гранулы, капсулы и т.п., причем такие твердые композиции могут содержать, помимо рекомбинантного аденовируса, по меньшей мере один наполнитель, такой как крахмал, карбонат кальция, сахароза, лактоза или желатин. Кроме простых наполнителей также можно использовать смазывающие средства, такие как стеарат магния или тальк. Жидкие композиции для перорального введения включают суспензии, растворы, эмульсии и сиропы, и могут содержать разные наполнители, такие как увлажняющие средства, ароматизирующие средства, душистые вещества и консерванты, помимо воды и жидкого парафина, которые представляют собой часто используемые простые разбавители. Композиции для парентерального введения включают стерильные водные растворы, неводные растворы, суспензии, эмульсии, высушенные из замороженного состояния препараты и свечи. В качестве неводных растворителей или суспендирующих средств можно использовать пропиленгликоль, полиэтиленгликоль, растительные масла, такие как оливковое масло, инъецируемые сложные эфиры, такие как этилолеат и т.п. В качестве основы для свечей можно использовать витепсол, макрогол, твин 61, масло какао, лауриновый жир, глицерожелатин и т.п.
В другом аспекте настоящее изобретение относится к способу лечения рака, который включает введение пациенту фармацевтически эффективного количества вышеописанных рекомбинантного аденовируса или фармацевтической композиции.
В рамках изобретения термин "фармацевтически эффективное количество" в применении к любому лекарственному средству относится к количеству, достаточному для лечения заболеваний при приемлемом соотношении польза/риск. Уровень эффективной дозы композиции зависит от вида пациента, тяжести заболевания, возраста и пола пациента, типа инфицирующего вируса, активности лекарственного средства, чувствительности к лекарственному средству, времени введения, способа введения, скорости выведения, продолжительности лечения, лекарственных средств, используемых в сочетании с композицией, и других факторов, известных в области медицины. Фармацевтическую композицию согласно настоящему изобретению можно вводить отдельно или в сочетании с другими терапевтическими средствами, причем ее можно вводить последовательно или одновременно с обычными терапевтическими средствами. Композицию согласно настоящему изобретению можно вводить в виде разовой или многократной дозировочной формы. Композицию нужно вводить в минимальном количестве, которое, с учетом всех вышеуказанных факторов, может обеспечить максимальный эффект, не вызывая побочных эффектов, и которое может быть легко определено специалистом в данной области. Фармацевтическую композицию согласно настоящему изобретению предпочтительно вводят перорально или внутривенно.
В рамках изобретения термин "ведение" относится к введению заранее определенного вещества животному с помощью любого подходящего метода. Фармацевтическую композицию согласно настоящему изобретению можно вводить с помощью любого общего способа, при условии, что он обеспечивает достижение ткани-мишени. Кроме того, фармацевтическую композицию согласно настоящему изобретению можно вводить с использованием любой системы, способной доставлять активный ингредиент к клеткам-мишеням.
Предпочтительная доза композиции в соответствии с настоящим изобретением может варьировать в зависимости от состояния и массы тела пациента, тяжести заболевания, типа композиции, способа введения и продолжительности лечения, и устанавливается специалистом в данной области в соответствии с указанными факторами. Однако для достижения желательных эффектов композицию согласно настоящему изобретению можно вводить в суточной дозе, составляющей от 1 до 10 мг/кг, предпочтительно от 1 до 5 мг/кг. Суточную дозу можно вводить в один примем, или ее можно разделить на несколько приемов.
Фармацевтическую композицию согласно настоящему изобретению можно вводить отдельно или в комбинации с вспомогательными терапевтическими методами, такими как хирургическая терапия. Химиотерапевтические средства, которые могут быть использованы с композицией согласно настоящему изобретению, включают, без ограничения, цисплатин, карбоплатин, прокарбазин, меклоретамин, циклофосфамид, ифосфамид, мелфалан, хлорамбуцил, бисульфан, нитрозомочевину, дактиномицин, даунорубицин, доксорубицин, блеомицин, пликомицин, митомицин, этопозид, тамоксифен, таксол, трансплатин, 5-фторурацил, винкристин, винбластин, метотрексат и т.п. Кроме того, композицию согласно настоящему изобретению можно использовать вместе с радиотерапевтическими методами, которые включают, без ограничения, облучение рентгеновскими лучами и облучение γ-лучами.
В одном примере авторы настоящего изобретения соединяют рибозим, опосредующий транс-сплайсинг, который распознает и расщепляет человеческую TERT (hTERT), с мРНК HSVtk и расщепленной по 3′-концу мРНК hTERT (фиг.1). Кроме того, чтобы подтвердить эффект, оказываемый рибозимом, опосредующим транс-сплайсинг, на мышей, авторы настоящего изобретения конструируют подобную систему на основе рибозима, опосредующего транс-сплайсинг, направленную на мРНК мышиной TERT (mTERT-TR), и сконструированный рибозим, опосредующий транс-сплайсинг, вставляют в геном аденовируса с делецией E1/E3, получая аденовирус (Ad5mTR), содержащий mTERT-TR-контролируемый HSVtk (mTERT-TR-HSVtk) (фиг.2a). Сконструированный аденовирус вводят в клетки рака толстой и прямой кишки CT26, полученные от мышей BALB/c, и в результате обнаруживают, что, если клетки инфицируют аденовирусом с MOI 2,5, большая часть клеток погибает (фиг.2b), и что, если Ad5mTR вводят в подкожную опухоль CT26 мышей BALB/c, рост опухоли значительно ингибируется, что находится в соответствии с результатами анализа цитотоксичности in vitro (фиг.2c). Кроме того, авторы настоящего изобретения анализируют способность Ad5mTR вызывать гибель клеток опухоли E.G7, введенной подкожно мышам B6, и в результате обнаруживают, что после введения вируса высвобождается опухолевый антиген (фиг.3a), и что DC, полученные от Ad5mTR-обработанных мышей, обеспечивают уровень антигена OVA, достаточный для стимуляции OVA-специфичных T-клеток (фиг.3b).
Между тем, авторы настоящего изобретения конструируют двухмодульный аденовирус (Ad5mTR.sPD1), содержащий HSVtk, который контролируется mTER-TR в участке E1 аденовирусного генома, и sPD1-Ig, присутствующим в участке E3 (фиг.4a), и анализируют активность двухмодульного аденовируса. В результате обнаруживают, что секреция IFN-γ антиген-специфичными T-клетками в ответ на атаку опухолевых клеток повышается в присутствии супернатанта, содержащего sPD1-Ig (фиг.4d), и что PD-L1 экспрессируется на поверхности опухолевых клеток, что подтверждают методом проточной цитометрии (фиг.5). Кроме того, показано, что клетки CT26, инфицированные Ad5mTR.sPD1, обладают цитотоксичностью in vitro, подобной цитотоксичности клеток CT26, инфицированных Ad5mTR (фиг.6a), и что Ad5mTR.sPD1 значительно инициирует регрессию опухоли по сравнению с Ad5mTR и приводит к почти полной регрессии опухоли (фиг.6b). В соответствии с настоящим изобретением полагают, что HSVtk стимулирует реагирование антиген-специфических T-клеток CD8 на DC (фиг.3b), а sPD1-Ig стимулирует отвечаемость противоопухолевых T-клеток CD8 во внеклеточной среде (фиг.4d).
Повышенное ингибиторное действие указанного двухмодульного Ad5mTR.sPD1 на рост опухоли практически отсутствует у мышей, истощенных по T-клеткам CD8 (фиг.7a), причем влияние истощения по CD8 на противоопухолевую активность у Ad5mTR.sPD1 выше, чем у Ad5mTR (1.9 раз по сравнению с 4,76 раз) (фиг.7b). Кроме того, у модели CT26 эффект, оказываемый Ad5mTR.sPD1, превышает эффект Ad5mTR (1,61 раз по сравнению с 6,58 раз) (фиг.8b), причем, если антиген-специфичный T-клеточный ответ формируется T-клетками OT-I, введенными внутривенно мышам Rag1-/-, содержащим E.G7, противоопухолевый эффект, обусловленный введением аденовируса, сохраняется, причем данный эффект является более очевидным при введении Ad5mTR.sPD1, чем при введении Ad5mTR (фиг.8c). Кроме того, число T-клеток OT-I в крови значительно увеличивается при введении Ad5mTR.sPD1 по сравнению с введением Ad5mTR (фиг.8d).
Указанный эффект аденовируса анализируют in vivo и обнаруживают, что у мышей, которым вводят Ad5mTR.sPD1, пролиферация первичной опухоли минимизируется, а вторичная опухоль не развивается (фиг.9a и 9c), причем такие же результаты наблюдаются у мышей B6, несущих опухоль E.G7 (фиг.9b и 9c).
Хотя данный эффект рекомбинантного аденовируса согласно настоящему изобретению зарегистрирован у мышей, рекомбинантный аденовирус согласно настоящему изобретению также можно использовать для лечения рака человека, поскольку TERT, против которой направлено действие аденовируса, экспрессируется не только в мышиных опухолях, но и в опухолях человека.
СПОСОБ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Далее настоящее изобретение описывается в деталях со ссылкой на предпочтительные примеры. Однако следует понимать, что указанные примеры приведены только для иллюстрации и не предназначены для ограничения объема согласно настоящему изобретению.
Пример 1: Материалы и методы
Пример 1-1: Клетки и мыши
Все клетки культивируют в среде RPMI, содержащей 10% FBS (фетальная бычья сыворотка) и 1% пенициллина/стрептомицина. Самок мышей BALB/c и C57/BL6 возрастом 6 недель получают от SLC (Japan). Мышей OT-1 (фон B6) и мышей Rag1-/- (фон B6) were получают от Jackson Laboratory. Все исследования с применением животных проводят в соответствии с Руководством по содержанию и использованию лабораторных животных Национального онкологического центра (Корея).
Пример 1-2: Конструирование аденовируса
Чтобы получить Ad5mTR, содержащий мышиный ген TERT-TR-HSVtk под контролем промотора CMV, используют системы AdenoZAPTM и AdenoQuickTM. pAVQ-CMV-mTERT AS100 Rib(+67) TK обрабатывают SpeI/SacII с получением фрагмента ДНК, который вставляют в участок расщепления SpeI/EcoRV pZAP1.1 и получают pZAP1.1.CMV.mTR.HSVtk. pZAP1.1.CMV.mTR.HSVtk расщепляют PacI/DraIII, лигируют с использованием RightZAP1.2 и вводят в клетки HEK293. Чтобы сконструировать sPD1-Ig, внеклеточный домен PD-1 амплифицируют методом ПЦР с использованием следующих праймеров:
Прямой праймер: 5′-CCG CTC GAG CTC ACC ATG TGG GTC CGG CAG GTA CCC TGG-3′ (SEQ ID NO: 5)
Обратный праймер: 5′-AGA TCT TCC TCC TCC TCC TTG AAA CCG GCC TTC TGG TTT GGG-3′ (SEQ ID NO: 6)
Продукт амплификации вставляют в участок XhoI/BglII вектора pFUSE-mIgG2A.Fc1 (Invitrogen, San Diego, CA) с получением конструкции pFUSE-mIgG2A.Fc1.EF1.sPD-1. EF1.sPD1-Ig, полученный из pFUSE-mIgG2A.Fc1.EF1.sPD1-Ig вставляют в участок EcoRI/SwaI вектора pE3.1 (OD260) с получением конструкции pE3.1.EF1.sPD1-Ig. Фрагмент CMV.mTR.HSVtk конструкции pZAP1.1.CMV.mTR.HSVtk вставляют в участок BamHI/SpeI вектора pE1.2 (OD260) с получением конструкции pE1.2.CMV.mTR.HSVtk. Чтобы сконструировать Ad5mTR.sPD1, pE1.2.CMV.mTR.HSVtk и pE3.1.EF1.sPD1-Ig расщепляют рестриктазами DraIII/PflMI, лигируют с использованием AdenoQuick13.1 и затем вводят в клетки HEK293. Чтобы сконструировать Ad5EF1.sPD1 аналогично Ad5mTR.sPD1, pE3.1.EF1.sPD1-Ig и пустой вектор pE1.2 расщепляют рестриктазами DraIII/PflMI, лигируют с использованием AdenoQuick13.1 и затем вводят в клетки HEK293.
Пример 1-3: абсорбция GCV и анализ цитотоксичности во внеклеточной среде
Ферментативную активность HSVtk определяют путем измерения накопления фосфорилированного GCV в клетках. Анализ пролиферации клеток (Dojindo Laboratories, Rockville, MD) проводят путем оценки цитотоксичности аденовируса с помощью стандартного метода. Затем клетки (3×103) вносят в 96-луночный планшет и культивируют в течение ночи при 37°С. Культивируемые клетки инфицируют аденовирусом при разных значениях MOI (множественность заражения). Через 1 день к клеткам добавляют GCV до конечной концентрации 200 мкМ и затем измеряют пролиферацию клеток в течение 3 дней. Все эксперименты повторяют три раза.
Пример 1-4: Антитело и реагент
Чтобы определить экспрессию белка sPD1-Ig на основе Ad5CMV.mTR.sPD1, клетки HEK293 (4×105) инфицируют аденовирусом с MOI 2. Через 24 часа после заражения культуральный супернатант и клеточный остаток (80 мкг) анализируют методом иммуноблотинга с использованием антитела против Pdcd-1 (Santa Cruz Biotechnology, Santa Cruz, CA). Чтобы удалить T-клетки CD8, внутрибрюшинно вводят 500 мкг антитела против CD8 2.43α за 2 дня до введения аденовируса, после чего антитело вводят внутрибрюшинно с 5-дневными интервалами в течение 15 дней.
Пример 1-5: Анализ продукции IFN-γ с использованием совместной культуры T-клеток CD8/DC
Полученные из дренированных лимфоузлов DC (DLN-DCs) разделяют в градиенте 17,5% Nycodenz и очищают на колонке MACS с использованием микрогранул против CD11c (Miltenyi Biotec, Auburn, CA). Чистые T-клетки CD8, полученные из мышей OT-1, разделяют с использованием микрогранул против CD8. DLN-DCs (5×104) совместно культивируют с чистыми T-клетками CD8 (1×105) в 96-луночном планшете в течение 72 часов. Через 72 часа культуральный супернатант собирают и измеряют в нем содержание IFN-γ методом ELISA (eBioscience, San Diego, CA).
Пример 1-6: Анализ активации T-клеток OT-1 во внеклеточной среде
Клетки рака ободочной и прямой кишки мыши MC38/OVA (фон B6), стабильно экспрессирующие овальбумин (OVA), культивируют в 6-луночном планшете. sPD1-Ig собирают из супернатанта культуры клеток HEK293, инфицированных аденовирусом с MOI 10 в течение 24 часов, затем добавляют к клеткам MC38/OVA (1×104), содержащим T-лимфоциты CD8 OT-1 (1×106), и культивируют в течение 72 часов. Чистые T-лимфоциты CD8+, полученные из мышей OT-1, выделяют с использованием колонки MACS, как описано выше. Количество IFN-γ, продуцируемое T-лимфоцитами CD8, измеряют с помощью набора CBA для анализа мышиного IFN-γ (BD bioscience, San Jose, CA).
Пример 1-7: Исследование животных in vivo и анализ in vitro
1×106 клеток CT26 инокулируют подкожно самкам мышей BALB/c возрастом 6 недель. Когда размер опухоли достигает измеряемой величины (через 7 дней), в нее вводят 5×108 БОЕ аденовируса и внутривенно вводят GCV (75 мг/кг) два раза в день в течение 14 дней. Аденовирус вводят два раза в день с 7-дневными интервалами. Объем опухоли измеряют по следующему уравнению:
Длина × ширина2 × 0,5236
Подобные процедуры получения подкожной опухоли и введения вируса проводят в эксперименте с истощением по T-клеткам CD8. Клетки E.G7 (фон C57/BL6; 1×106) вводят мышам Rag1-/- или мышам C57/BL6, после чего проводят обработку аденовирусом и GCV по описанному выше способу. В анализе in vitro опухоль E.G7 мыши Rag1-/- обрабатывают Ad5mTR.sPD1 в соответствии со способом, в котором внутривенно вводят T-клетки CD8, выделенные из мышей OT-1. Через 8 дней после инъекции из крови собирают PBMC (мононуклеарные клетки периферической крови) и анализируют их методом проточной цитометрии.
Пример 2: Результаты
Пример 2-1: Конструирование аденовируса, содержащего мышиный TERT-TR-HSVtk
Авторы настоящего изобретения разработали новую стратегию экспрессии HSVtk с использованием известного опухолевого маркера TERT и опухоль-специфичного аденовируса. В данной стратегии рибозим, который распознает и расщепляет человеческий TERT (hTERT), доставляют в опухолевые клетки с помощью рекомбинантного аденовируса. Кроме того, рибозим конструируют так, чтобы он был связан с мРНК HSVtk и расщеплял мРНК hTERT по 3′-концу. Он индуцирует новую трансляцию мРНК HSVtk в опухолевых клетках. Данный вид рибозима определяют как рибозим, опосредующий транс-сплайсинг (фиг.1). Теоретически экспрессия HSVtk, которая регулируется рибозимом, опосредующим транс-сплайсинг, ограничивается опухолевыми клетками, экспрессирующими мРНК hTERT на высоком уровне. Поэтому указанная система предлагается в качестве эффективного способа доставки HSVtk в опухолевые клетки, не оказывающего влияние на нормальные клетки окружающих тканей, в которых hTERT экспрессируется на высоком уровне. Фактически данная стратегия обеспечивает значительный эффект против клеток рака ободочной и прямой кишки человека, трансплантированных мышиной модели ксенотрансплантата. Недостатком данной системы с точки зрения иммунологического анализа является то, что эксперимент с использованием клеток опухоли человека проводят на иммунодефицитных мышах, у которых подавляют реакцию отторжения ксенотрансплантата. Поэтому данная система не подходит для анализа влияния HSVtk на противоопухолевый иммунный ответ. Чтобы преодолеть данный недостаток, авторы настоящего изобретения разработали подобную систему, в которой используется рибозим, опосредующий транс-сплайсинг и специфичный к мРНК мышиной TERT (mTERT-TR), а иммунологический эффект TERT-TR-контролируемой HSVtk анализируют с использованием модели мышиной опухоли. Последовательность распознавания TERT расположена вблизи HSVtk-кодирующей последовательности, а экспрессия HSVtk зависит от присутствия транскриптов mTERT (фиг.1). Указанную экспрессионную кассету, которая находится под контролем промотора CMV, вставляют в геном аденовируса, содержащего делецию E1/E3, и получают аденовирус (Ad5mTR), содержащий mTERT-TR-контролируемый HSVtk (mTERT-TR-HSVtk) (фиг.2a).
В качестве контроля используют аденовирус (Ad5MOCK), который не содержит экспрессионную кассету. Чтобы определить, действительно ли HSVtk, экспрессируемый из Ad5mTR, характеризуется цитотоксичностью в отношении клеток мышиной опухоли, авторы настоящего изобретения используют клетки рака ободочной и прямой кишки CT26, полученные из мышей BALB/c, причем такие клетки экспрессируют мРНК mTERT на высоком уровне. Клетки CT26 обрабатывают Ad5mTR с разными значениями MOI. 2,5 MOI Ad5mTR достаточно, чтобы уничтожить большую часть клеток, тогда как Ad5MOCK не проявляет цитотоксичности даже при 50 MOI (фиг.2b). Если Ad5mTR вводят в подкожную опухоль CT26, присутствующую у мышей BALB/c, рост опухоли значительно ингибируется, что согласуется с результатами анализа цитотоксичности in vitro (фиг.2c).
Таким образом, можно видеть, что Ad5mTR оказывает противоопухолевый эффект подобно аденовирусу, содержащему человеческий TERT-TR-HSVtk, и может использоваться для генерирования иммунного ответа у мышей.
Пример 2-2: Повышение DC-опосредованной презентации антигена под действием HSVtk
Известно, что экспрессия HSVtk в опухолевых клетках в результате введения ДНК или инфекции аденовируса стимулирует антигенный ответ цитотоксических T-клеток CD8. Поскольку экспрессия HSVtk в присутствии GCV in vivo может индуцировать гибель клеток в значительной части объема опухоли, опухолевый антиген, присутствующий в уничтоженных клетках, можно детектировать с использованием APC, таких как DC, причем указанные клетки мигрируют к лимфоузлам, которые отвечают на T-клетки, специфичные к опухолевому антигену. Хотя данная модель описана в литературе, авторы настоящего изобретения анализируют такую возможность с использованием определенных антиген-специфичных T-клеток. В данном анализе авторы настоящего изобретения используют линию клеток мышиной опухоли E.G7 (полученную из клеток EL4, которые стабильно экспрессируют овальбумин) в качестве модельной антигенной опухоли, а очищенные T-клетки, полученные из трансгенных мышей, несущих OVA-специфичный T-клеточный рецептор, используют в качестве колонии T-клеток CD8, специфичных к модельному опухолевому антигену (OVA). Все T-клетки CD8, выделенные из мышей OT-1, представляют собой OVA-специфичные цитотоксичные T-клетки. Вначале авторы настоящего изобретения тестируют способность Ad5mTR индуцировать гибель клеток опухоли E.G7, введенных подкожно мышам B6. После выделения опухоли и введения Ad5mTR проводят гистологический анализ, который демонстрирует значительный уровень гибели клеток, позволяя предположить, что опухолевый антиген высвобождается после инфекции вируса (фиг. 3a). Затем авторы настоящего изобретения выделяют DC из опухолевых дренированных лимфоузлов и определяют, действительно ли OVA осуществляют продукцию IFN-γ, путем анализа совместной культуры DC/T-клеток CD8. T-клетки OT-I, культивированные и очищенные с DC, полученными из опухоль-несущих мышей, обработанных Ad5mTR, продуцируют большое количество IFN-γ по сравнению с T-клетками OT-I, культивированными с DC, полученными из опухоль-несущих мышей, обработанных контрольным вирусом. Этот результат позволяет предположить, что DC, полученные из мышей, обработанных Ad5mTR, могут продуцировать уровень антигена OVA, достаточный для стимуляции OVA-специфичных T-клеток (фиг.3b). Таким образом, можно видеть, что экспрессия HSVtk в опухоли и обусловленная ею GCV-индуцированная гибель клеток являются результатом высвобождения опухолевых антигенов, и что указанные антигены эффективно улавливаются DC, которые могут стимулировать опухолевый антиген-специфичные цитотоксичные T-клетки.
На основе полученных результатов можно сделать вывод, что опухоль-специфичная экспрессия HSVtk может эффективно стимулировать противоопухолевую активность T-клеток, опосредованную DC, и непосредственно индуцировать цитотоксичность в опухолевых клетках.
Пример 2-3: Конструирование аденовируса, содержащего mTERT-TR-HSVtk и sPD1-Ig
Ad5mTR-стимулированный ответ противоопухолевых T-клеток CD8 вызывает повышенный интерес. Сочетание указанных стратегий с другой стратегией, включающей в себя инактивацию опухоль-индуцированной иммунной устойчивости, дополнительно усиливает отвечаемость противоопухолевых T-клеток. PD-L1 представляет собой известный иммунный супрессор, который экспрессируется на поверхности опухолевых клеток. Авторы настоящего изобретения конструируют sPD1 (растворимая форма рецептора PD-L1, которая нейтрализует PD-L1), чтобы нейтрализовать PD-L1-опосредованную супрессию T-клеток. Чтобы повысить стабильность sPD1 in vivo, sPD1 гибридизуют с участком Fc IgG2a, получая sPD1-Ig. Авторы настоящего изобретения конструируют двухмодульный аденовирус (Ad5mTR.sPD1), содержащий HSVtk, который контролируется mTERT-TR в участке E1 генома аденовируса genome, и sPD1-Ig, находящийся в участке E3 (фиг.4a). Клетки HEK293 инфицируют Ad5mTR.sPD1, sPD1-Ig, после экспрессии и секреции во внеклеточную среду, анализируют методом иммуноблоттинга (фиг.4b). Уровень экспрессии HSVtk в Ad5mTR.sPD1-инфицированных клетках определяют путем измерения ферментативной активности и сравнивают с уровнем, наблюдающимся в Ad5mTR-инфицированных клетках (фиг.4c). Кроме того, авторы настоящего изобретения определяют, действительно ли sPD1-Ig, секретированный в окружающую среду in vitro, может повышать отвечаемость противоопухолевых T-клеток путем нейтрализации PD-L1 на поверхности опухолевых клеток. Совместную культуру OVA-экспрессирующих опухолевых клеток и OVA-специфичных T-клеток OT-I добавляют к супернатанту культуры Ad5EF1α.sPD1-инфицированных клеток 293HEK и измеряют отвечаемость T-клеток по секреции IFN-γ. Как и ожидалось, секреция IFN-γ антиген-специфичными T-клетками в ответ на атаку опухолевых клеток повышается в присутствии sPD1-Ig-содержащего супернатанта (фиг. 4d). Экспрессию PD-L1 на поверхности опухолевых клеток подтверждают методом проточной цитометрии (фиг.5). Полученные результаты позволяют предположить, что двойной модель может повышать отвечаемость противоопухолевых T-клеток в микроокружении опухоли in vivo.
Пример 2-4: Эффективное ингибирование опухолевого роста под действием Ad5mTR.sPD1
Чтобы подтвердить, действительно ли sPD1-Ig может повышать противоопухолевую активность HSVtk in vivo, используют модель рака ободочной и прямой кишки CT26 (фиг.2c). Выбор данной модели обусловлен тем, что клетки CT26 экспрессируют на своей поверхности PD-L1 на высоком уровне (фиг.5). Предполагается, что клетки CT26, инфицированные Ad5mTR.sPD1, будут проявлять цитотоксичность, подобную цитотоксичности клеток CT26, инфицированных Ad5mTR, и что sPD1-Ig не являются непосредственной причиной цитотоксичности, индуцируемой HSVtk (фиг.6a). В отличие от этого, Ad5mTR.sPD1 вызывает значительную регрессию опухоли при сравнении с Ad5mTR, причем в случае Ad5mTR.sPD1 наблюдается почти полная регрессия опухоли (фиг.6b). В данной модели контрольный аденовирус (Ad5EF1α.sPD1), содержащий только sPD1-Ig, не проявляет какого-либо противоопухолевого эффекта, позволяя предположить, что экспрессия sPD1-Ig в опухолевой ткани не является достаточной для индукции регрессии опухоли, и что антигенный ответ, обусловленный HSVtk in vivo является необходимым для проявления эффекта sPD1-Ig.
Пример 2-5: Ингибиторное действие sPD1-Ig на рост опухоли опосредуется T-клетками CD8
HSVtk стимулирует ответ антиген-специфичных T-клеток CD8 на DC (фиг.3b), а sPD1-Ig стимулирует отвечаемость противоопухолевых T-клеток CD8 во внеклеточной среде (фиг.4d). Такие результаты позволяют предположить, что повышенная отвечаемость T-клеток CD8 in vivo обусловлена повышенным противоопухолевым эффектом Ad5mTR.sPD1. Чтобы подтвердить данное предположение, мышам, несущим опухоль CT26, вводят антитело против CD8, чтобы удалить T-клетки CD8. Повышенное ингибиторное влияние двухмодульного Ad5mTR.sPD1 на рост опухоли практически отсутствует у мышей, истощенных по T-клеткам CD8 (фиг.7a). Интересно, что противоопухолевая активность Ad5mTR также уменьшается, позволяя предположить, что эффект Ad5mTR у мышиной модели опухоли CT26 зависит от противоопухолевых иммунных ответов, опосредованных T-клетками CD8. Однако влияние удаления CD8 на противоопухолевую активность у Ad5mTR.sPD1 выше, чем у Ad5mTR (1,9 раз и 4,76 раз) (фиг.7b). Для более непосредственной оценки функции опухоль-специфичных T-клеток CD8 используют модели опухоли E.G7 и OVA-специфичные T-клетки OT-I. Введение Ad5mTR.sPD1 в инъецированную подкожно мышам B6 опухоль E.G7 приводит к регрессии опухоли, как и у модели CT26. В отличие от данных результатов, проведение такого же эксперимента на мышах Rag1-/-, истощенных по лимфоцитам, приводит к значительному снижению противоопухолевых эффектов двух типов вирусов (фиг.8a). Подобно эффекту удаления T-клеток CD8, степень уменьшения опухолевого роста у модели CT26 выше при использовании Ad5mTR.sPD1, чем Ad5mTR (1,61 раз и 6,58 раз) (фиг.8b). Если ответ антиген-специфических T-клеток формируется в присутствии T-клеток OT-I, введенных внутривенно мышам Rag1-/-, несущим E.G7, противоопухолевый эффект введения аденовируса сохраняется, причем в случае введения Ad5mTR.sPD1 данный эффект более выражен, чем в случае введения Ad5mTR (фиг.8c). Кроме того, число T-клеток OT-I в крови значительно повышается при введении Ad5mTR.sPD1 по сравнению с введением Ad5mTR (фиг.8d). Такие результаты позволяют предположить, что Ad5mTR.sPD1 стимулирует регрессию опухоли путем непосредственного усиления функции опухоль-специфичных T-клеток CD8.
Пример 2-6: Противораковый эффект обработки Ad5mTR.sPD1
В крови мышей, которым был введен Ad5mTR.sPD1, обладающий повышенной способностью к ингибированию роста вторичной опухоли в участке, удаленном от места локализации первичной опухоли, инфицированной вирусом, число рак-специфичных T-клеток увеличивается. Ad5mTR.sPD1 вводят в опухоль CT26 мышей BALB/c три раза с интервалами в 3 дня. Через 14 дней после первого введения индуцируют образование вторичной опухоли CT26 с противоположного бока тех же мышей. В данном случае, у мышей, которым был введен Ad5mTR.sPD1, пролиферация первичной опухоли минимизируется, тогда как у мышей, которым был введен Ad5MOCK, образуется первичная опухоль большого размера (>600 мм3). Новую группу нормальных мышей BALB/c используют в качестве контрольной группы для получения вторичной опухоли. Вторичная опухоль благополучно растет в контрольной группе, но не растет у мышей, которым был введен Ad5mTR.sPD1 (фиг.9a и 9c). Такой же результат наблюдается у мышей B6, несущих опухоль E.G7 (фиг.9b и 9c). Таким образом, можно сделать вывод, что введение двухмодульного аденовируса обеспечивает противораковый эффект путем усиления противоракового иммунитета.
| название | год | авторы | номер документа |
|---|---|---|---|
| ПРОТИВОРАКОВАЯ КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ ОПУХОЛЕСПЕЦИФИЧЕСКИЙ ОНКОЛИТИЧЕСКИЙ АДЕНОВИРУС И ИНГИБИТОР КОНТРОЛЬНОЙ ТОЧКИ ИММУННОГО ОТВЕТА | 2018 |
|
RU2777523C2 |
| ПРОТИВОРАКОВАЯ КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ ОПУХОЛЕСПЕЦИФИЧЕСКИЙ ОНКОЛИТИЧЕСКИЙ АДЕНОВИРУС И ИНГИБИТОР КОНТРОЛЬНОЙ ТОЧКИ ИММУННОГО ОТВЕТА | 2018 |
|
RU2740713C1 |
| КОМБИНИРОВАННАЯ ТЕРАПИЯ ДЛЯ ЛЕЧЕНИЯ РАКА С ПОМОЩЬЮ РЕКОМБИНАНТНОГО ПОКСВИРУСА, ЭКСПРЕССИРУЮЩЕГО ОПУХОЛЕВЫЙ АНТИГЕН, И АНТАГОНИСТА ИЛИ АГОНИСТА МОЛЕКУЛЫ ИММУННОЙ КОНТРОЛЬНОЙ ТОЧКИ | 2015 |
|
RU2724433C2 |
| МНОГОПРОФИЛЬНЫЙ ПРОМОТОР, ЭКСПРЕССИРУЮЩИЙ ВЕКТОР И СПОСОБ ИЗБИРАТЕЛЬНОГО УБИЙСТВА РАКОВЫХ КЛЕТОК С ИХ ИСПОЛЬЗОВАНИЕМ | 2012 |
|
RU2476596C1 |
| УНИВЕРСАЛЬНЫЕ РАКОВОСПЕЦИФИЧНЫЕ ПРОМОТОРЫ И ИХ ИСПОЛЬЗОВАНИЕ В ПРОТИВООПУХОЛЕВОЙ ТЕРАПИИ | 2013 |
|
RU2539764C2 |
| СЛИТЫЙ БЕЛОК ТЕЛОМЕРАЗНОЙ ОБРАТНОЙ ТРАНСКРИПТАЗЫ, КОДИРУЮЩИЕ ЕГО НУКЛЕОТИДЫ И ИХ ПРИМЕНЕНИЕ | 2007 |
|
RU2473691C2 |
| ПРОТИВОРАКОВАЯ КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ РЕКОМБИНАНТНЫЙ АДЕНОВИРУС, ЭКСПРЕССИРУЮЩИЙ РАЗРУШАЮЩИЙ ФАКТОР ДЛЯ ВНЕКЛЕТОЧНОГО МАТРИКСА | 2017 |
|
RU2742726C2 |
| ФАРМАКОЛОГИЧЕСКАЯ КОМБИНАЦИЯ ПОЛИКАТИОННОГО НОСИТЕЛЯ ПЭГ-ПЭИ-ТАТ С ЗАКЛЮЧЕННОЙ В НЕМ ПЛАЗМИДОЙ, НЕСУЩЕЙ ТЕРАПЕВТИЧЕСКИЕ ГЕНЫ HSVtk И GM-CSF ДЛЯ ЦЕЛЕЙ ГЕНОТЕРАПИИ ОПУХОЛЕВЫХ ЗАБОЛЕВАНИЙ | 2013 |
|
RU2575077C2 |
| ПРОТИВООПУХОЛЕВАЯ КОМПОЗИЦИЯ | 2019 |
|
RU2768287C1 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ TUSC2-ИММУНОТЕРАПИИ | 2017 |
|
RU2755903C2 |























Предложены рекомбинантный аденовирус, фармацевтическая композиция, содержащая такой вирус, и способ лечения рака с использованием такого вируса или композиции. Охарактеризованный рекомбинантный аденовирус содержит полинуклеотид, кодирующий комплекс рибозима, опосредующего транс-сплайсинг, и HSVtk (тимидинкиназы вируса простого герпеса человека), где рибозим, опосредующий транс-сплайсинг, представляет собой рибозим Rib67, и где мишенью рибозима, опосредующего транс-сплайсинг, является мРНК гена рак-специфичной TERT (обратной транскриптазы теломеразы) и противораковый терапевтический ген, кодирующий sPD-1 (растворимый белок программируемой гибели 1). Рекомбинантный аденовирус обладает селективностью в отношении раковых клеток, обусловленной рибозимом, опосредующим транс-сплайсинг, и повышенной противораковой активностью, обусловленной противораковым терапевтическим геном. Представленные изобретения могут быть использованы для эффективных профилактики и лечения рака. 3 н. и 2 з.п. ф-лы, 23 ил., 13 пр.
1. Рекомбинантный аденовирус для генной терапии рака, содержащий: полинуклеотид, кодирующий комплекс рибозима, опосредующего транс-сплайсинг, и HSVtk (тимидинкиназы вируса простого герпеса человека), где рибозим, опосредующий транс-сплайсинг, представляет собой рибозим Rib67, кодируемый нуклеотидной последовательностью SEQ ID NO: 2, и HSVtk кодируется нуклеотидной последовательностью SEQ ID NO: 3, и где мишенью рибозима, опосредующего транс-сплайсинг, является мРНК гена рак-специфичной TERT (обратной транскриптазы теломеразы), состоящая из последовательности ДНК SEQ ID NO: 1; и противораковый терапевтический ген, состоящий из нуклеотидной последовательности SEQ ID NO: 4, кодирующей sPD-1 (растворимый белок программируемой гибели 1).
2. Рекомбинантный аденовирус по п. 1, где полинуклеотид, кодирующий комплекс, представляет собой чужеродную последовательность ДНК, встроенную в область гена Е1 аденовируса; и
где противораковый терапевтический ген, состоящий из нуклеотидной последовательности SEQ ID NO: 4, представляет собой чужеродную последовательность ДНК, встроенную в область гена Е3 аденовируса.
3. Фармацевтическая композиция для профилактики или лечения рака, содержащая в качестве активного ингредиента рекомбинантный аденовирус по любому из пп. 1 и 2.
4. Композиция по п. 3 дополнительно содержащая фармацевтически приемлемый носитель, эксципиент или разбавитель.
5. Способ лечения рака, включающий введение пациенту фармацевтически приемлемого количества рекомбинантного аденовируса по любому из пп. 1 и 2.
| M-S SONG et al., Validation of tissue-specific promoter-driven tumor-targetingtrans-splicing ribozyme system as a multifunctional cancer genetherapy device in vivo, Cancer Gene Therapy, 2009, Vol.16, 113-125; | |||
| WO 2010024483 A1, 04.03.2010 | |||
| SILKE SCHEPELMANN et al, Suicide Gene Therapy of Human Colon Carcinoma Xenografts Using an Armed Oncolytic |

