ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
Настоящая заявка является частичным продолжением и притязает на приоритет согласно 35 U.S.C 120 заявки № 10/682764, поданной 9 октября 2003 года, по которой испрашивается приоритет согласно 35 U.S.C. 119(e) на основании предварительной заявки 60/417282, поданной 9 октября 2002 года.
ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Настоящее изобретение относится к области клонирующих векторных плазмид и к применению клонирующих векторных плазмид для создания конструкций ДНК или трансгенов.
УРОВЕНЬ ТЕХНИКИ
Основой молекулярной биологии является технология рекомбинантной ДНК, которую можно в настоящем описании кратко представить как модификацию и репродукцию нуклеиновых кислот в целях исследования структуры и функции нуклеиновых кислот и их белковых продуктов.
Отдельные гены, области регуляции генов, подгруппы генов и, конечно, целые хромосомы, в которых они находятся, состоят из двунитевых антипараллельных последовательностей нуклеотидов аденина, тимина, гуанина и цитозина, обычно обозначаемых буквами A, T, G и C соответственно. Указанные последовательности ДНК, а также последовательности кДНК, которые представляют собой двунитевые копии ДНК, полученные на основе молекул мРНК (матричной РНК), можно расщепить на отдельные фрагменты, выделить и встроить в вектор, такой как бактериальная плазмида, чтобы исследовать генные продукты. Плазмида является внехромосомной частью ДНК, которую исходно получили из бактерий и которая может быть подвергнута обработке и снова введена в бактерию-хозяина в целях исследования или получения генного продукта. ДНК плазмиды сходна со всей хромосомной ДНК тем, что она состоит из таких же нуклеотидов A, T, G и C, кодирующих гены и области регуляции генов, однако ДНК плазмиды представляет собой относительно небольшую молекулу, состоящую менее, чем примерно из 30000 пар оснований или 30 тысяч пар оснований (т.п.о.). Кроме того, пары нуклеотидных оснований двунитевой плазмиды образуют непрерывную кольцевую молекулу, что также отличает плазмидную ДНК от хромосомной ДНК.
Плазмиды усиливают быстрый обмен генетического материала между бактериальными организмами и обеспечивают быструю адаптацию к изменениям окружающей среды, таким как изменения температуры, пищевых ресурсов или другие сигналы. Любая приобретенная плазмида должна экспрессировать ген или гены, которые вносят вклад в жизнеспособность хозяина, или же она будет разрушена или отвергнута организмом, так как сохранение ненужных плазмид было бы неэкономичным использованием ресурсов. Клональная популяция клеток содержит идентичный генетический материал, включая любые плазмиды, которые они могут нести. Применение клонирующей векторной плазмиды со вставкой ДНК в такой клональной популяции клеток-хозяев будет увеличивать количество доступной представляющей интерес ДНК. Клонированную таким образом ДНК затем можно выделить и извлечь для последующей обработки, используя стадии, необходимые для создания конструкции ДНК. Таким образом, понятно, что клонирующие векторные плазмиды являются полезными средствами исследования функции генов, обеспечивающие возможность быстро получать большие количества представляющей интерес вставки ДНК.
В то время как некоторые элементы, выявляемые в плазмидах, являются природными, другие были сконструированы, чтобы расширить применимость плазмид в качестве ДНК-векторов. Такие элементы, наряду с прочим, включают гены резистентности к антибиотикам или химическим веществам и сайт множественного клонирования (MCS). Каждый из таких элементов играет определенную роль согласно настоящему изобретению, а также согласно известному уровню техники. При описании роли каждого элемента на первый план будут выделены ограничения предшествующего уровня техники и показана полезность настоящего изобретения.
Особенно полезным находящимся в плазмиде геном, который может быть приобретен хозяином, является ген, которые может придавать резистентность к антибиотикам. В повседневной практике технологии рекомбинантной ДНК гены резистентности к антибиотикам используют в качестве элементов для позитивной или негативной селекции, чтобы предпочтительно усилить культивирование и амплификацию требуемой плазмиды по сравнению с другими плазмидами.
Чтобы плазмида поддерживалась бактерией-хозяином, она также должна содержать участок последовательностей, который побуждает хозяина к удвоению плазмиды. Последовательности, известные как элемент начала репликации (ORI), заставляют хозяина использовать свои клеточные ферменты для получения копий плазмиды. Когда такая бактерия делится, каждая дочерняя клетка будет иметь копию или копии любой такой плазмиды. Были получены некоторые штаммы бактерий E. coli, которые позволяют максимизировать такую дупликацию, создавая более 300 копий на бактерию. Таким образом, культивирование требуемой плазмиды может быть усилено.
Другим необходимым элементом в любом клонирующем векторе является место для встраивания представляющих интерес генетических материалов. Такой элемент является синтетическим элементом, который был сконструирован в плазмидах «дикого типа», чтобы таким образом сделать их применимыми в качестве клонирующего вектора. Любая обычная коммерчески доступная векторная плазмида для клонирования содержит по меньшей мере одну такую область, известную как сайт множественного клонирования (MCS). MCS обычно содержит нуклеотидные последовательности, которые могут быть расщеплены одним или группой эндонуклеазных ферментов рестрикции (в дальнейшем называемых «ферментами рестрикции»), каждая из которых имеет отдельный сайт рестрикции эндонуклеазой и характерную картину расщепления. Такие сайты для эндонуклеаз или сайты рестрикции эндонуклеазами (в дальнейшем называемые «сайтами рестрикции»), кодируемые в молекуле ДНК, обычно содержат двунитевую палиндромную последовательность. Для некоторых ферментов рестрикции достаточно всего 4-6 нуклеотидов, чтобы обеспечить сайт рестрикции, тогда как некоторые ферменты рестрикции требуют наличия сайта рестрикции из 8 или более нуклеотидов. Например, фермент EcoR1 узнает гексануклеотидную последовательность: 5'G-A-A-T-T-C3', где 5' указывает конец молекулы, по договоренности названный «апстрим» концом, а 3' подобным образом указывает «даунстрим» конец. Комплементарная нить сайта рестрикции должна быть его антипараллельной нитью, 3'G-A-A-T-T-C5'. Таким образом, двунитевой сайт рестрикции может присутствовать в более крупной двунитевой молекуле, в которой он встречается в виде:
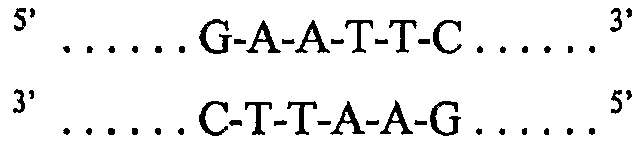
Подобно многим другим ферментам рестрикции, EcoR1 не расщепляет точно по оси симметрии двух нитей, а расщепляет в положениях, отстоящих друг от друга на четыре нуклеотида в двух нитях ДНК, между нуклеотидами, указанными «/»:
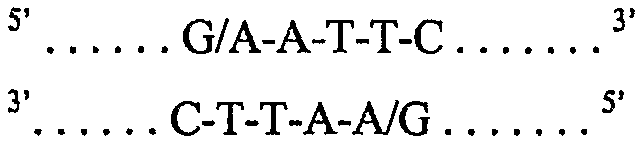
так что двунитевая молекула ДНК расщепляется и в результате имеет конфигурацию нуклеотидов на новообразованных «концах»:
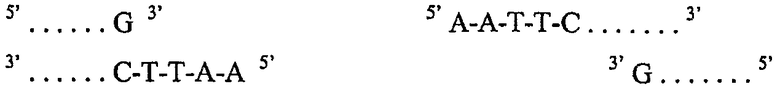
Такое ступенчатое расщепление дает фрагменты ДНК с выступающими 5'-концами. Поскольку пары A-T и G-C образуются спонтанно, если нуклеотиды находятся рядом друг с другом, то выступающие концы, такие как указанные выше, называют способными к связыванию или «липкими» концами. Любой из таких концов может образовывать водородные связи с любыми другими комплементарными концами, расщепленными таким же ферментом рестрикции. Так как любая ДНК, которая содержит специфичный сайт рестрикции, будет разрезана таким же образом, как и любая другая ДНК, содержащая такую же последовательность, то такие расщепленные концы будут комплементарными. Поэтому концы любых молекул ДНК, разрезанных одним и тем же ферментом рестрикции, «совпадают» друг с другом подобно «совпадению» расположенных рядом кусочков мозаики и могут быть связаны вместе ферментами. Именно такое свойство обеспечивает возможность образования рекомбинантных молекул ДНК и позволяет вводить фрагменты чужеродной ДНК в бактериальные плазмиды или в любую другую молекулу ДНК.
Следующий общий принцип, который учитывается при создании рекомбинантных молекул ДНК, заключается в том, что все сайты рестрикции, встречающиеся в молекуле, будут разрезаться конкретным ферментом рестрикции, но прежде всего представляющий интерес сайт. Чем крупнее молекула ДНК, тем более вероятно, что любой сайт рестрикции будет встречаться повторно. При условии, что любые сайты рестрикции распределены случайным образом вдоль молекулы ДНК, тетрануклеотидный сайт будет встречаться в среднем один раз на каждые 44 (то есть 256) нуклеотидов, тогда как гексануклеотидный сайт будет встречаться один раз на каждые 46 (то есть 4096) нуклеотидов, а октануклеотидные сайты будут встречаться один раз на каждые 48 (то есть 114688) нуклеотидов. Таким образом, легко можно понять, что более короткие сайты рестрикции будут встречаться часто, тогда как более длинные сайты рестрикции будут встречаться редко. При планировании конструкции трансгена или другой молекулы рекомбинантной ДНК, это является важной проблемой, так как такой проект часто требует сборки нескольких фрагментов ДНК разных размеров. Чем крупнее такие фрагменты, тем более вероятно, что сайты, которые собираются использовать, встречаются в нескольких фрагментах компонентов ДНК, что в лучшем случае затрудняет процесс обработки.
Часто встречающиеся сайты рестрикции в настоящем описании названы широко распространенными сайтами рестрикции, и эндонуклеазы, которые расщепляют такие сайты, названы часто щепящими ферментами рестрикции. Ферменты рестрикции, которые узнают последовательности длиной более 6 нуклеотидов, называют редко щепящими ферментами рестрикции, а узнаваемые ими сайты - редкими сайтами рестрикции. Однако существуют некоторые сайты рестрикции из 6 п.н., которые встречаются более редко, чем можно ожидать статистически, и такие сайты и эндонуклеазы, которые их расщепляют, также называют редкими. Таким образом, обозначения «редкий» и «широко распространенный» не относятся к относительной численности или доступности какого-либо конкретного фермента рестрикции, а относятся к частоте встречаемости последовательности нуклеотидов, которая составляет узнаваемый им сайт рестрикции в какой-либо молекуле ДНК или выделенном фрагменте молекулы ДНК или любом гене или в его последовательности ДНК.
Недавно выделен второй класс ферментов рестрикции, называемых ферментами хоминг-эндонуклеазами (HE). Ферменты HE имеют большие асимметричные сайты рестрикции (12-40 пар оснований). Сайты рестрикции HE чрезвычайно редко встречаются. Например, HE, известная как I-SceI, имеет сайт рестрикции длиной 18 п.о. (5'...TAGGGATAACAGGGTAAT...3'), который согласно расчетам встречается только один раз на каждые 7×1010 пар оснований случайной последовательности. Такая частота встречаемости эквивалентна только одному сайту на 20 геномов, имеющих размер генома млекопитающего. Редкая природа сайтов HE в значительной степени увеличивает вероятность того, что генный инженер может вырезать конечный трансгенный продукт, не нарушая целостность трансгена, если сайты HE были введены в соответствующие положения в векторной плазмиде для клонирования.
Так как молекула ДНК из любого выбранного в качестве источника организма будет разрезаться идентичным образом узнающим ее ферментом рестрикции, то чужеродные фрагменты ДНК любого вида могут быть вырезаны ферментом рестрикции, встроены в бактериальный плазмидный вектор, который был расщеплен таким же ферментом рестрикции, и амплифицированы в подходящей клетке-хозяине. Например, ген человека может быть вырезан в 2 местах ферментом рестрикции, известным как EcoR1, и требуемый фрагмент с EcoR1-концами может быть выделен и смешан с плазмидой, которую также разрезали EcoR1, с образованием смеси, обычно известной как реакционная смесь для лигирования или смесь для лигирования. В соответствующих условиях в смеси для лигирования некоторые из выделенных фрагментов гена человека будут соответствовать концам плазмидных молекул. Такие заново соединенные концы могут связываться вместе и посредством ферментов снова образовывать кольцевую плазмиду, теперь содержащую свою новую вставку ДНК. Смесь для лигирования затем вводят в E. coli или другого подходящего хозяина, и заново сконструированные плазмиды будут амплифицироваться по мере деления бактерий. Таким образом, относительно большое количество копий гена человека может быть получено и собрано из бактерий. Такие копии гена затем могут быть подвергнуты дальнейшей обработке в целях исследования, анализа или получения белкового продукта гена.
Технологию рекомбинантной ДНК часто осуществляют при создании так называемых «трансгенов». Трансгены часто содержат разнообразные генетические материалы, которые получены из одного или нескольких донорных организмов и введены в организм хозяина. Обычно трансген конструируют, используя клонирующий вектор в качестве начальной точки или «остова» конструкции, и планируют серию сложных стадий клонирования, чтобы осуществить сборку конечного продукта в таком векторе. Элементы трансгена, содержащие нуклеотидные последовательности, включают без ограничения 1) регуляторные промоторные и/или энхансерные элементы, 2) ген, который будет экспрессироваться в виде молекулы мРНК, 3) ДНК-элементы, которые обеспечивают стабилизацию первичного транскрипта мРНК, 4) нуклеотидные последовательности, имитирующие интронные области генов млекопитающих, и 5) сигналы для процессинга мРНК, такие как поли-A-хвост, добавляемый к концу природных мРНК. В некоторых случаях экспериментальная конструкция может требовать добавления сигнала локализации, чтобы обеспечить транспорт генного продукта в конкретное субклеточное положение. Каждый из таких элементов представляет собой фрагмент более крупной молекулы ДНК, которая вырезается из генома донора или в некоторых случаях синтезируется в лаборатории. Каждый участок подвергается сборке вместе с другими участками в определенном порядке и в 5'-3'-ориентации в векторной плазмиде для клонирования.
Промотор любого гена может быть выделен в виде фрагмента ДНК и помещен в синтетическую молекулу, такую как плазмида, чтобы управлять экспрессией требуемого гена, при условии, что необходимые условия для стимуляции представляющего интерес промотора могут быть обеспечены. Например, последовательности промотора гена инсулина могут быть выделены, помещены в векторную плазмиду для клонирования наряду с репортерный геном и использованы для исследования условий, необходимых для экспрессии гена инсулина в соответствующем типе клеток. Альтернативно промотор гена инсулина может быть связан с кодирующей белок последовательностью любого представляющего интерес гена в клонирующей векторной плазмиде и использован для управления экспрессией представляющего интерес гена в экспрессирующих инсулин клетках, при условии, что все необходимые элементы присутствуют в ДНК-трансгене, сконструированном таким образом.
Репортерный ген является особенно полезным компонентом некоторых типов трансгенов. Репортерный ген содержит нуклеотидные последовательности, кодирующие белок, которые будут экспрессироваться под контролем конкретного представляющего интерес промотора, с которым они связаны в трансгене, обеспечивая измеряемый биохимический ответ активности промотора. Репортерный ген обычно легко выявлять или измерять на фоне эндогенных клеточных белков. Широко используемые репортерные гены включают без ограничения LacZ, белок с зеленой флуоресценцией и люциферазу и другие репортерные гены, многие из которых хорошо известны специалистам в данной области.
Интроны, которые представляют собой некодирующие области генов млекопитающих, не встречаются в бактериальных геномах, но требуются для правильного образования молекул мРНК в клетках млекопитающих. Поэтому любая конструкция ДНК для применения в системах млекопитающих должна иметь по меньшей мере один интрон. Интроны могут быть выделены из любого гена млекопитающих и встроены в конструкцию ДНК наряду с подходящими сигналами сплайсинга, которые позволяют клеткам млекопитающих вырезать интрон и соединять оставшиеся концы мРНК вместе.
Элементом стабилизации мРНК является последовательность ДНК, которую распознают связывающие белки, которые защищают некоторые мРНК от расщепления. Введение элемента стабилизации мРНК часто будет повышать уровень экспрессии гена по сравнению с уровнем мРНК в некоторых типах клеток млекопитающих, и таким образом может быть полезным в некоторых конструкциях ДНК или трансгенах. Элемент стабилизации мРНК может быть выделен из природных ДНК или РНК или получен синтетически для введения в конструкцию ДНК.
Сигналом локализации является последовательность ДНК, которая кодирует белковый сигнал для определения маршрута представляющего интерес белка в клетке. Например, сигнал ядерной локализации будет направлять белок в ядро; сигнал локализации в плазматической мембране будет направлять его к плазматической мембране и т.д. Таким образом, сигнал локализации может быть введен в конструкцию ДНК, чтобы стимулировать перемещение белкового продукта в требуемое место в клетке.
Последовательность метки может быть закодирована в конструкции ДНК для того, чтобы белковый продукт имел уникальную связанную с ним область. Такая уникальная область служит в качестве метки белка, по которой его можно отличить от его эндогенной копии. Альтернативно, такая уникальная область может служить в качестве опознавательного признака, который может быть выявлен различными способами, хорошо известными в данной области, включая без ограничения ОТ-ПЦР, иммуногистохимию или гибридизацию in situ.
В случае сложного трансгена или трансгена, который содержит особенно большие области ДНК, существует повышенная вероятность того, что будут множественные сайты рестрикции в таких участках ДНК. Напомним, что сайты рестрикции, кодирующие любой один гексануклеотидный сайт, встречаются на каждые 4096 п.о. (46). Если последовательность промотора длиной 3000 п.о. и представляющий интерес ген длиной 1500 п.о. собраны в клонирующем векторе длиной 3000 п.о., то статистически очень вероятно, что многие сайты из 6 или меньшего количества нуклеотидов не будут полезными, так как любые пригодные сайты должны встречаться только в двух участках. Кроме того, сайты должны встречаться в подходящих областях соответствующих молекул, которые необходимо собрать вместе. Кроме того, для большинства клонирующих конструкций будет необходимо добавление дополнительных элементов ДНК, что увеличивает сложность растущей молекулы и вероятность ненужного повторения какого-либо конкретного сайта. Так как любой фермент рестрикции будет расщеплять все свои сайты рестрикции в молекуле ДНК, если сайт рестрикции ферментом повторно встречается, то все неподходящие сайты будут расщепляться наряду с нужными сайтами, нарушая целостность молекулы. Таким образом, каждая стадия клонирования должна быть тщательно спланирована так, чтобы не нарушить растущую молекулу расщеплением ферментом рестрикции, который уже был использован для введения предшествующего элемента. И наконец, когда исследователь хочет ввести законченный трансген в организм млекопитающего, полностью собранную трансгенную конструкцию часто необходимо линеаризовать в уникальном сайте рестрикции по меньшей мере с одного конца трансгена, что таким образом требует еще одного уникального сайта, нигде больше не встречающегося в конструкции. Так как большинство конструкций ДНК предназначено для одной цели, мало думают о том, что любые модификации, которые вероятно могут понадобиться в будущем, дополнительно усложнят будущие экспериментальные изменения.
Традиционно на дизайн и конструирование трансгена тратится достаточное количество времени и энергии по нескольким причинам, включая следующие:
1. Существует широкое множество доступных ферментов рестрикции и ферментов HE, которые будут создавать серию концов; однако большинство из них не совместимы друг с другом. Многие ферменты рестрикции, такие как EcoR1, образуют фрагменты ДНК с выступающими липкими 5'-концами или «хвостами»; другие (например, Pst1) образуют фрагменты с выступающими 3'-хвостами, тогда как следующие (например, Bal1) расщепляют по оси симметрии, давая фрагменты с тупыми концами. Некоторые из них будут совместимы с концами, образованными при расщеплении другими ферментами рестрикции и/или ферментами HE, но большинство из них не будут совместимыми. Концы, которые могут быть образованы при каждой процедуре выделения фрагментов ДНК, должны тщательно учитываться при проектировании конструкции ДНК.
2. Фрагменты ДНК, необходимые для сборки конструкции ДНК или трансгена должны сначала быть выделены из геномов, которые являются их источником, помещены в плазмидные клонирующие векторы и амплифицированы, чтобы получить приемлемые количества. Стадию можно осуществлять с использованием ряда коммерчески доступных или индивидуально измененных клонирующих векторов. Каждую из различных коммерчески доступных клонирующих векторных плазмид в основном разрабатывали независимо, и поэтому они содержат разные последовательности и сайты рестрикции для фрагментов ДНК представляющих интерес генов или генетических элементов. Поэтому гены необходимо подгонять индивидуально, чтобы приспособить к каждому из таких векторов, которые необходимы для любой данной серии экспериментов. Одинаковые фрагменты ДНК часто необходимо изменять дополнительно для последующих экспериментов или клонирования в других комбинациях для новых конструкций ДНК или трансгенов. Так как каждую конструкцию ДНК или трансген готовят на заказ для конкретного применения, не думая или не зная, как она будет использована в следующий раз, то ее зачастую необходимо «обратно подгонять» для последующих применений.
3. Кроме того, последовательность ДНК любого данного гена или генетического элемента варьирует и может содержать внутренние сайты рестрикции, которые делают ее несовместимой с доступными в настоящее время векторами, тем самым осложняя обработку. Особенно это верно в случае сборки нескольких фрагментов ДНК в одной конструкции ДНК или трансгене.
Хотя для обработки генетического материала можно использовать ферменты рестрикции, известно, что MCS или другие компоненты трансгена могут быть созданы посредством синтеза de novo, рекомбинацией и/или клонированием с использованием ПЦР с праймерами, имеющими выступающие концы с терминаторами. Один такой способ синтеза составляющих элементов трансгена описан Jarrell et al. в патенте США № 6358712. Хотя Jarrell описывает способ «сварки» элементов трансгена вместе, в известном уровне техники не описан способ или методика, позволяющая «не сваривать» и повторно собирать такие элементы после того, как они были собраны.
Поэтому было бы полезно разработать методику сборки каждого компонента трансгена с другими компонентами в точно установленном порядке и в 5'-3'-ориентации в векторной плазмиде для клонирования. Также существует потребность в системе, которая давала бы возможность пользователю быстро собирать несколько фрагментов ДНК в одну молекулу, несмотря на избыточное количество сайтов рестрикции, встречающихся на концах и в каждом из таких фрагментов ДНК. Также может быть полезной разработка простой методики быстрого изменения концов выбранных фрагментов ДНК, чтобы добавить к ним сайты рестрикции. Введение единичных или противостоящих пар сайтов рестрикции HE может повысить вероятность присутствия уникальных сайтов для клонирования. Система, которая позволила бы также легко заменять или удалять один или несколько фрагментов и могла бы повысить эксплуатационную гибкость, в настоящее время не доступна для пользователей. Поэтому «модульная» система, позволяющая встраивать или удалять фрагменты ДНК из областей «кассеты», фланкированных постоянными редкими сайтами рестрикции, в клонирующем векторе, может быть особенно полезной и долгожданной в области технологии рекомбинантной ДНК.
Соответственно один аспект настоящего изобретения относится к способу быстрой сборки конструкции ДНК или трансгена с использованием векторной плазмиды для клонирования. Другой аспект относится к последовательному введению фрагментов ДНК, также называемых «вставками» или «модулями», таких как промотор, экспрессируемая нуклеотидная последовательность и 3'-регуляторная нуклеотидная последовательность, в клонирующую векторную плазмиду.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к структуре и способам создания модульных клонирующих векторов для синтеза модуля генетического домена, трансгена PE3 и других сложных конструкций ДНК посредством создания остова в модульных клонирующих векторах, имеющего специальные точки стыковки, известные как генные шарниры.
Настоящее изобретение относится к вектору для стыковки доменных модулей, состоящему из клонирующего ДНК-вектора, содержащего модуль множественного клонирования (MC) для субклонирования представляющего интерес генетического материала в модуле MC, при этом модуль MC содержит: a) первый генный шарнир (GP), содержащий по меньшей мере два невариабельных редких сайта рестрикции, которые можно подвергать обработке для определения 5'-части модуля MC; b) последовательность нуклеиновой кислоты, содержащую сайт множественного клонирования (MCS), содержащий множество сайтов рестрикции, выбранных из общепринятых сайтов рестрикции, которые являются уникальными в векторе для стыковки доменных модулей, чтобы обеспечить клонирующие сайты для клонирования представляющего интерес генетического материала в модуле MC; и c) второй генный шарнир, содержащий по меньшей мере два невариабельных редких сайта рестрикции, которые можно подвергать обработке для определения 3'-части модуля MC.
Настоящее изобретение также относится к вектору доменного модуля, состоящему из клонирующего ДНК-вектора, содержащего доменный модуль, который содержит: a) первый генный шарнир, содержащий по меньшей мере два невариабельных редких сайта рестрикции, которые можно подвергать обработке для определения 5'-части доменного модуля; b) представляющий интерес генетический модуль, состоящий из последовательности нуклеиновой кислоты, содержащей представляющий интерес генетический материал; и c) второй генный шарнир, содержащий по меньшей мере два невариабельных редких сайта рестрикции, которые можно подвергать обработке для определения 3'-части доменного модуля.
Настоящее изобретение также относится к стыковочному вектору PE3, состоящему из клонирующего ДНК-вектора, содержащего модуль клонирования PE3, который содержит по меньшей мере один клонирующий модуль, скомпонованный для клонирования по меньшей мере модуля первого домена в модуле клонирования PE3, при этом модуль клонирования PE3 содержит: a) первый генный шарнир, содержащий по меньшей мере два невариабельных редких сайта рестрикции, которые при клонировании можно подвергать обработке для определения 5'-части по меньшей мере модуля первого домена; b) модуль вкладыша, состоящий из первой последовательности нуклеиновой кислоты, содержащей вкладыш, который при клонировании заменяется модулем первого домена; и c) второй генный шарнир, содержащий по меньшей мере два невариабельных редких сайта рестрикции, которые при клонировании можно подвергать обработке для определения 3'-части по меньшей мере модуля первого домена.
Настоящее изобретение также относится к стыковочному вектору PE3, состоящему из клонирующего ДНК-вектора, содержащего модуль клонирования PE3, который содержит множество клонирующих модулей, скомпонованных для клонирования множества доменных модулей в модуле клонирования PE3, при этом модуль клонирования PE3 содержит: a) первый генный шарнир, содержащий по меньшей мере два невариабельных редких сайта рестрикции, которые при клонировании можно подвергать обработке для определения 5'-части модуля первого домена; b) модуль первого вкладыша, состоящий из первой последовательности нуклеиновой кислоты, содержащей вкладыш, который при клонировании заменяется модулем первого домена; c) второй генный шарнир, содержащий по меньшей мере два невариабельных редких сайта рестрикции, которые при клонировании можно подвергать обработке для определения области соединения в одно целое между 3'-частью модуля первого домена и 5'-частью модуля второго домена; d) модуль второго вкладыша, состоящий из второй последовательности нуклеиновой кислоты, содержащей вкладыш, который при клонировании заменяется модулем второго домена; e) третий генный шарнир, содержащий по меньшей мере два невариабельных редких сайта рестрикции, которые при клонировании можно подвергать обработке для определения области соединения в одно целое между 3'-частью модуля второго домена и 5'-частью модуля третьего домена; f) модуль третьего вкладыша, состоящий из третьей последовательности нуклеиновой кислоты, содержащей вкладыш, который при клонировании заменяется модулем третьего домена; и g) четвертый генный шарнир, содержащий по меньшей мере два невариабельных редких сайта рестрикции, которые можно подвергать обработке для определения 3'-части модуля третьего домена.
Настоящее изобретение, кроме того, относится к стыковочному вектору PE3 для множественного клонирования (MC), состоящему из клонирующего ДНК-вектора, содержащего модуль клонирования PE3, скомпонованный для клонирования по меньшей мере трех доменных модулей в модуле клонирования PE3, при этом модуль клонирования PE3 содержит: a) первый генный шарнир, содержащий по меньшей мере два невариабельных редких сайта рестрикции, которые при клонировании можно подвергать обработке для определения 5'-части модуля первого домена; b) первую последовательность нуклеиновой кислоты; c) второй генный шарнир, содержащий по меньшей мере два невариабельных редких сайта рестрикции, которые при клонировании можно подвергать обработке для определения области соединения в одно целое между 3'-частью модуля первого домена и 5'-частью модуля второго домена; d) вторую последовательность нуклеиновой кислоты; e) третий генный шарнир, содержащий по меньшей мере два невариабельных редких сайта рестрикции, которые при клонировании можно подвергать обработке для определения области соединения в одно целое между 3'-частью модуля второго домена и 5'-частью модуля третьего домена; f) третью последовательность нуклеиновой кислоты; и g) четвертый генный шарнир, содержащий по меньшей мере два невариабельных редких сайта рестрикции, которые при клонировании можно подвергать обработке для определения области соединения 3'-части модуля третьего домена; при этом по меньшей мере одна первая, вторая и третья последовательность нуклеиновой кислоты является модулем множественного клонирования, содержащим сайт множественного клонирования (MCS), содержащий множество сайтов рестрикции, выбранных из широко распространенных сайтов рестрикции, которые являются уникальными в стыковочном векторе PE3, обеспечивающими клонирующие сайты для клонирования представляющего интерес генетического материала в модуле множественного клонирования, а остальные последовательности нуклеиновой кислоты являются последовательностями вкладышей.
Настоящее изобретение, кроме того, относится к вектору PE3, состоящему из клонирующего ДНК-вектора, содержащего модуль PE3, который содержит модуль промотора, модуль экспрессии и 3'-регуляторный модуль, при этом модуль PE3 содержит: a) первый генный шарнир, содержащий по меньшей мере два невариабельных редких сайта рестрикции, которые можно подвергать обработке для определения 5'-части модуля промотора; b) модуль промотора; c) второй генный шарнир, содержащий по меньшей мере два невариабельных редких сайта рестрикции, которые можно подвергать обработке для определения области соединения в одно целое между 3'-частью модуля промотора и 5'-частью модуля экспрессии; d) модуль экспрессии; e) третий генный шарнир, содержащий по меньшей мере два невариабельных редких сайта рестрикции, которые можно подвергать обработке для определения области соединения в одно целое между 3'-частью модуля экспрессии и 5'-частью 3'-регуляторного модуля; f) 3'-регуляторный модуль; и g) четвертый генный шарнир, содержащий по меньшей мере два невариабельных редких сайта рестрикции, которые можно подвергать обработке для определения 3'-части 3'-регуляторного модуля.
В настоящем изобретении также предполагается, что любой из векторов: стыковочный вектор PE3, вектор PE3 и стыковочный вектор PE3 для множественного клонирования (MC), может содержать элементы для высвобождения модуля PE3, обычно содержащего модули домена промотора, домена экспрессии и 3'-регуляторного домена, фланкированные генными шарнирами, из вектора PE3 для встраивания в стыковочный вектор нескольких генов. Один элемент для высвобождения и встраивания модуля клонирования PE3 содержит пару сайтов хоминг-эндонуклеаз (HE), расположенных в стыковочном векторе PE3 или стыковочном векторе PE3 MC, которые фланкируют последовательность нуклеиновой кислоты, которая содержит модуль клонирования PE3. После встраивания или клонирования домена промотора, домена экспрессии и 3'-регуляторных доменов в модуле клонирования PE3, пара HE обеспечивает одно из средств вырезания модуля PE3 из вектора PE3 для встраивания в совместимое положение для стыковки в мультигенном векторе, при этом совместимое положение для стыковки обычно определяется такой же парой HE. Хотя можно использовать любые хоминг-эндонуклеазы либо в первом положении (на 5'-конце вырезаемого модуля PE3), либо во втором положении (на 3'-конце вырезаемого модуля PE3), предпочтительная пара HE состоит из I-Ceu I в первом положении и обратного ISce-I во втором положении. В других вариантах стыковочного вектора PE3 или стыковочного вектора PE3 MC можно использовать другие средства для вырезания и встраивания вместо HE. Такие другие средства для вырезания и встраивания могут включать пару одного или нескольких невариабельных редких сайтов рестрикции, включая отдельную пару редких сайтов рестрикции из сайтов рестрикции в генных шарнирах или сами фланкирующие генные шарниры (GP1 и GP4). При таких инсерциях используется рекомбинация, обсуждаемая ниже.
Настоящее изобретение относится к способу конструирования модульного вектора PE3, включающему стадии: a) получения клонирующего вектора PE3, содержащего модуль клонирования PE3, при этом модуль клонирования PE3 содержит последовательно расположенные: первый генный шарнир, содержащий по меньшей мере два невариабельных редких сайта рестрикции, по меньшей мере модуль первого вкладыша, состоящий из последовательности нуклеиновой кислоты, содержащей вкладыш, и второй генный шарнир; b) получения по меньшей мере модульного вектора первого домена, содержащего последовательно расположенные: первый генный шарнир, представляющий интерес генетический модуль, состоящий из последовательности нуклеиновой кислоты, содержащей представляющий интерес генетический материал, и второй генный шарнир; c) получения первого фермента рестрикции, узнающего один из редких сайтов рестрикции первого генного шарнира, и второго фермента рестрикции, узнающего один из редких сайтов рестрикции во втором генном шарнире; d) вырезания и выделения представляющего интерес генетического модуля из модульного вектора первого домена с использованием первого и второго узнающих ферментов рестрикции; e) вырезание модуля первого вкладыша из модуля клонирования PE3 клонирующего вектора PE3 с использованием первого и второго узнающих ферментов рестрикции; и f) лигирование представляющего интерес генетического модуля в модуль клонирования PE3. Способ также обеспечивает встраивание второго представляющего интерес генетического модуля в PE3 с использованием третьего генного шарнира и узнающих ферментов рестрикции для вырезания и лигирования второго представляющего интерес генетического модуля в модуль клонирования PE3. Подобным образом, представляющий интерес третий генетический модуль может быть встроен в PE3 с использованием третьего генного шарнира и узнающих ферментов рестрикции для вырезания и лигирования второго представляющего интерес генетического модуля в модуль клонирования PE3. Способ обеспечивает последовательное расположение представляющих интерес генетических модулей в клонирующем векторе PE3. Обычно представляющие интерес первый, второй и третий генетические модули соответствуют модулю промотора, модулю экспрессии и 3'-регуляторному модулю, соответственно.
Другой вариант осуществления изобретения относится к способу получения трансгена, включающему стадии получения клонирующей векторной плазмиды, содержащей остов, при этом остов имеет по меньшей мере первую, вторую, третью и четвертую точку стыковки, точки стыковки расположены последовательно в 5'-3'-направлении, и каждая имеет по меньшей мере один невариабельный редкий сайт рестрикции, можно подвергать обработке для расщепления ферментом рестрикции, расщепления первой точки стыковки первым ферментом рестрикции, который соответствует одному из по меньшей мере одного невариабельного редкого сайта рестрикции первой точки стыковки, оставляя при этом расщепленную первую точку стыковки с незащищенным 3'-концом, расщепления второй точки стыковки вторым ферментом рестрикции, который соответствует одному из по меньшей мере одного невариабельного редкого сайта рестрикции второй точки стыковки, оставляя при этом расщепленную вторую точку стыковки с незащищенным 5'-концом, получения первой вставки, содержащий 5'-конец, представляющую интерес нуклеотидную последовательность и 3'-конец, где 5'-конец первой вставки совместим с незащищенным 3'-концом расщепленной первой точки стыковки, а 3'-конец первой вставки совместим с незащищенным 5'-концом расщепленной второй точки стыковки, и помещения первой вставки и расщепленной клонирующей векторной плазмиды в подходящую реакционную смесь, чтобы вызвать лигирование и самоориентацию первой вставки в остове между первой точкой стыковки и второй точкой стыковки, в которой остов подвергается повторной сборке.
В данном варианте вторую точку стыковки затем можно расщепить вторым ферментом рестрикции, оставляя расщепленную вторую точку стыковки с незащищенным 3'-концом, третью точку стыковки можно расщепить третьим ферментом рестрикции, который соответствует одному из по меньшей мере одного невариабельного редкого сайта рестрикции третьей точки стыковки, оставляя расщепленную третью точку стыковки с незащищенным 5'-концом, с последующими стадиями получения второй вставки, содержащей 5'-конец, представляющую интерес нуклеотидную последовательность и 3'-конец, при этом 5'-конец второй вставки совместим с незащищенным 3'-концом расщепленной второй точки стыковки, а 3'-конец второй вставки совместим с незащищенным 5'-концом расщепленной третьей точки стыковки, и помещения второй вставки и расщепленной клонирующей векторной плазмиды в подходящую реакционную смесь, чтобы вызвать лигирование и самоориентацию второй вставки в остове между второй точкой стыковки и третьей точкой стыковки, в которой остов подвергается повторной сборке.
Кроме того, данный вариант осуществления может включать стадии дальнейшего расщепления остова в третьей точке стыковки третьим ферментом рестрикции, оставляя при этом расщепленную третью точку стыковки с незащищенным 3'-концом, расщепления четвертой точки стыковки четвертым ферментом рестрикции, который соответствует одному из по меньшей мере одного невариабельного редкого сайта рестрикции четвертой точки стыковки, оставляя при этом расщепленную четвертую точку стыковки с незащищенным 5'-концом, получения третьей вставки, содержащий 5'-конец, представляющую интерес нуклеотидную последовательность и 3'-конец, при этом 5'-конец третьей вставки совместим с незащищенным 3'-концом расщепленной третьей точки стыковки, а 3'-конец третьей вставки совместим с незащищенным 5'-концом расщепленной четвертой точки стыковки, и помещения третьей вставки и расщепленной клонирующей векторной плазмиды в подходящую реакционную смесь, чтобы вызывать лигирование и самоориентацию третьей вставки в остове между четвертой точкой стыковки и второй точкой стыковки, в которой остов подвергается повторной сборке.
Еще один вариант осуществления изобретения относится к способу получения модульной клонирующей векторной плазмиды для синтеза трансгена или другой сложной конструкции ДНК, при этом способ включает стадии получения клонирующей векторной плазмиды, содержащей остов, где остов имеет по меньшей мере первую и вторую точки стыковки, каждая из точек стыковки имеет по меньшей мере один невариабельный редкий сайт рестрикции, который можно подвергнуть обработке, чтобы расщепить ферментом рестрикции, расщепления первой точки стыковки первым ферментом рестрикции, который соответствует одному из по меньшей мере одного невариабельного редкого сайта рестрикции первой точки стыковки, оставляя при этом расщепленную первую точку стыковки с незащищенным 3'-концом и расщепленный остов с незащищенным 5'-концом, получения первой вставки, содержащий 5'-конец, представляющую интерес нуклеотидную последовательность и 3'-конец, при этом 5'-конец первой вставки совместим с незащищенным 3'-концом расщепленной первой точки стыковки, а 3'-конец первой вставки можно подвергать обработке при объединении с незащищенным 5'-концом расщепленного остова с образованием третьей точки стыковки, имеющей по меньшей мере один невариабельный редкий сайт рестрикции, который можно подвергать обработке для расщепления третьим ферментом рестрикции, помещения первой вставки и расщепленной клонирующей векторной плазмиды в подходящую реакционную смесь, чтобы вызывать лигирование и самоориентацию первой вставки в остове между первой точкой стыковки и третьей точкой стыковки и апстрим от второй точки стыковки, при этом остов подвергается повторной сборке, последующего расщепления третьей точки стыковки третьим ферментом рестрикции, оставляя при этом расщепленную третью точку стыковки с незащищенным 3'-концом и расщепленный остов с незащищенным 5'-концом, получения второй вставки, содержащей 5'-конец, представляющую интерес нуклеотидную последовательность и 3'-конец, при этом 5'-конец второй вставки совместим с незащищенным 3'-концом расщепленной третьей точки стыковки, а 3'-конец второй вставки можно подвергать обработке при объединении с незащищенным 5'-концом расщепленного остова с образованием четвертой точки стыковки, содержащей по меньшей мере один невариабельный редкий сайт рестрикции, который можно подвергать обработке для расщепления четвертым ферментом рестрикции, помещения второй вставки и расщепленной клонирующей векторной плазмиды в подходящую реакционную смесь, чтобы вызывать лигирование и самоориентацию второй вставки в остове между третьей точкой стыковки и четвертой точкой стыковки и апстрим от второй точки стыковки, при этом остов подвергается повторной сборке, последующего расщепления четвертой точки стыковки четвертым ферментом рестрикции, оставляя при этом расщепленную четвертую точку стыковки с незащищенным 3'-концом, расщепления второй точки стыковки вторым ферментом рестрикции, соответствующим одному из по меньшей мере одного невариабельного редкого сайта рестрикции второй точки стыковки, оставляя при этом расщепленную вторую точку стыковки с незащищенным 5'-концом, получения третьей вставки, содержащей 5'-конец, представляющую интерес нуклеотидную последовательность и 3'-конец, при этом 5'-конец третьей вставки совместим с незащищенным 3'-концом расщепленной четвертой точки стыковки, а 3'-конец третьей вставки совместим с незащищенным 5'-концом расщепленной второй точки стыковки, и помещения третьей вставки и расщепленной клонирующей векторной плазмиды в подходящую реакционную смесь, чтобы вызывать лигирование и самоориентацию третьей вставки в остове между четвертой точкой стыковки и второй точкой стыковки, при этом остов подвергается повторной сборке.
Дальнейшее понимание природы и преимуществ настоящего изобретения будет более полным при обращении к следующим чертежам и подробному описанию.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Прилагаемые чертежи, которые включены и составляют часть настоящего описания, иллюстрируют варианты осуществления изобретения и вместе с общим описанием изобретения, приведенным выше, и подробным описанием, приведенным ниже, служат для объяснения принципов изобретения.
Фиг.1 является схематичной иллюстрацией стыковочного вектора для доменных модулей, содержащего сайт множественного клонирования в генетическом модуле.
Фиг.2 является схематичной иллюстрацией стыковочного вектора PE3, содержащего клонирующий домен модуль для клонирования доменного модуля.
Фиг.3 является схематичной иллюстрацией альтернативного варианта стыковочного вектора для множественного клонирования PE3, содержащего сайт множественного клонирования для клонирования в генетическом модуле.
Фиг.4 является схематичной иллюстрацией мультигенного вектора, содержащего элементы для клонирования в векторе PE3.
Фиг.5 является другой схематичной иллюстрацией модульной концепции изобретения.
Фиг.6 является картой стыковочной плазмиды.
Фиг.7 является линейной картой рестрикции, иллюстрирующей пример сайтов для ферментов рестрикции, которые могут быть включены в MCS стыковочной плазмиды.
Фиг.8 является картой мультигенной (или основной) стыковочной плазмиды.
Фиг.9 является линейной картой рестрикции, иллюстрирующей пример сайтов для ферментов рестрикции, которые могут быть включены в MCS мультигенной стыковочной плазмиды.
Фиг.10 является картой плазмидного челночного вектора P («SVP»).
Фиг.11 является линейной картой сайтов рестрикции, иллюстрирующей пример сайтов для ферментов рестрикции, которые могут быть включены в MCS челночного вектора P.
Фиг.12 является картой плазмидного челночного вектора E («SVE»).
Фиг.13 является линейной картой сайтов рестрикции, иллюстрирующей пример сайтов для ферментов рестрикции, которые могут быть включены в MCS челночного вектора E.
Фиг.14 является картой челночного вектора 3' («SV3»).
Фиг.15 является линейной картой сайтов рестрикции, иллюстрирующей пример сайтов для ферментов рестрикции, которые могут быть включены в MCS челночного вектора 3'.
Фиг.16 является схематичной иллюстрацией мультигенного стыковочного вектора, полученного и используемого с модулями PE3 согласно изобретению.
Фиг.17 является схематичной иллюстрацией встраивания модуля PE3 в мультигенный стыковочный вектор с использованием генных шарниров, чтобы получить мультигенный вектор с модулем PE3.
Фиг.18 является схематичной иллюстрацией встраивания модуля PE3 в мультигенный стыковочный вектор с использованием сайта множественного клонирования в мультигенном векторе.
Фиг.19 является схематичной иллюстрацией встраивания модуля PE3 в мультигенный стыковочный вектор с использованием сайтов рестрикции BstX-1 в мультигенном векторе.
Фиг.20 является схематичной иллюстрацией встраивания модуля PE3 в мультигенный стыковочный вектор с использованием хоминг-эндонуклеазы в мультигенном векторе.
Фиг.21 является схематичной иллюстрацией встраивания модуля PE3 в мультигенный стыковочный вектор с использованием рекомбинации между вектором PE3 и мультигенным вектором.
КРАТКОЕ ОПИСАНИЕ СПИСКА ПОСЛЕДОВАТЕЛЬНОСТЕЙ
Прилагаемые списки последовательностей, которые включены и составляют часть настоящего описания, иллюстрируют варианты осуществления изобретения и вместе с общим описанием изобретения, приведенным выше, и подробным описанием, приведенным ниже, служат для объяснения принципов изобретения.
SEQ:ID 01 является примером нуклеотидной последовательности стыковочной плазмиды PE3 MCS.
SEQ:ID 02 является примером нуклеотидной последовательности стыковочной плазмиды PE3.
SEQ:ID 03 является примером нуклеотидной последовательности основной стыковочной плазмиды MCS.
SEQ:ID 04 является примером нуклеотидной последовательности основной стыковочной плазмиды.
SEQ:ID 05 является примером нуклеотидной последовательности плазмидного челночного вектора P MCS.
SEQ:ID 06 является примером нуклеотидной последовательности плазмидного челночного вектора P.
SEQ:ID 07 является примером нуклеотидной последовательности плазмидного челночного вектора E MCS.
SEQ:ID 08 является примером нуклеотидной последовательности плазмидного челночного вектора E.
SEQ:ID 09 является примером нуклеотидной последовательности плазмидного челночного вектора 3 MCS.
SEQ:ID 10 является примером нуклеотидной последовательности плазмидного челночного вектора 3.
ОПРЕДЕЛЕНИЯ ТЕРМИНОВ, ИСПОЛЬЗУЕМЫХ ДЛЯ ОПИСАНИЯ ИЗОБРЕТЕНИЯ
В целях настоящего изобретения следующие термины определяют следующим образом.
«Домен модификации хроматина» (CMD) относится к нуклеотидным последовательностям, которые взаимодействуют с различными белками, связанными с поддержанием и/или изменением структуры хроматина.
«Клонирование» относится к процессу лигирования молекулы ДНК в плазмиду и ее перенос в подходящую клетку-хозяина для удвоения при размножении хозяина.
Термин «клонирующий вектор» относится к кольцевой молекуле ДНК, минимально содержащей начало репликации, средства для позитивной селекции клеток-хозяев, несущих вектор, такой как ген резистентности к антибиотикам; и сайт множественного клонирования. Клонирующий вектор может состоять из остова вектора на основе плазмиды, космиды, искусственной хромосомы (BAC, PAC, YAC и другие) или вируса.
«Узнаваемая последовательность» или «узнаваемые последовательности» относятся к минимальной цепочке нуклеотидов, необходимой для того, что фермент рестрикции связался и расщепил молекулу ДНК или ген.
«Широко распространенный» относится к любому сайту рестрикции, который встречается в геноме относительно часто.
«Совместимый с» относится к концу, либо 5', либо 3', нити ДНК, который может образовывать водородные связи с любым комплементарным концом, либо расщепленным таким же ферментом рестрикции, либо созданным каким-либо другим способом. Так как любая ДНК, которая содержит специфичный сайт рестрикции для фермента рестрикции, будет разрезаться таким же образом, что и любая другая ДНК, содержащая такую же последовательность, полученные при расщеплении концы будут комплементарными и, следовательно, совместимыми. Поэтому концы любых молекул ДНК, разрезанных одним и тем же ферментом рестрикции, «совпадают» друг с другом подобно «совпадению» расположенных рядом кусочков мозаики и могут быть связаны вместе ферментами. Совместимые концы будут образовывать сайт рестрикции для конкретного фермента рестрикции при их объединении.
«Синтез de novo» относится к способу синтеза двунитевых молекул ДНК любой длины посредством связывания комплементарных однонитевых молекул ДНК, совместимые выступающие концы которых представляют собой подпоследовательности общей требуемой молекулы ДНК.
«Конструкция ДНК» относится к молекуле ДНК, синтезированной с использованием последовательных стадий клонирования, в клонирующей векторной плазмиде, и конструкцию ДНК обычно используют для управления генной экспрессией в любой подходящей клетке-хозяине, например, в культивируемых клетках in vitro, или в трансгенной мыши in vivo. Трансген, используемый для получения такой мыши, также можно назвать конструкцией ДНК, особенно во время конструирования и синтеза трансгена.
«Фрагмент ДНК» относится к любой изолированной молекуле ДНК, включая без ограничения кодирующую белок последовательность, репортерный ген, промотор, энхансер, интрон, экзон, поли-A-хвост, сайт множественного клонирования, сигнал ядерной локализации или сигнал стабилизации мРНК или любую другую природную или синтетическую молекулу ДНК. Альтернативно фрагмент ДНК может быть полностью синтетического происхождения, получаемым in vitro. Кроме того, фрагмент ДНК может содержать любую комбинацию изолированных природных и/или синтетических фрагментов.
«Стыковочная плазмида» относится к специализированной клонирующей векторной плазмиде, используемой в изобретении для сборки фрагментов ДНК в конструкцию ДНК.
«Эндонуклеаза» или «эндонуклеазный фермент», также обычно называемый в настоящем описании «ферментом рестрикции», относится к представителю или представителям классификации каталитических молекул, которые связывают сайт рестрикции, кодируемый в молекуле ДНК, и расщепляет молекулу ДНК в точном положении в последовательности или рядом с ней.
Термины «сайт рестрикции эндонуклеазой» или «сайт рестрикции» (а также «узнаваемая последовательность» или «узнаваемые последовательности», указанные выше) относятся к минимальной цепочке нуклеотидов, необходимой для того, чтобы фермент рестрикции связывался и расщеплял молекулу ДНК или ген.
«Энхансерная область» относится к нуклеотидной последовательности, которая не требуется для экспрессии гена-мишени, но будет увеличивать уровень экспрессии гена в подходящих условиях.
«Ген селекции экспрессии генов хозяина» (GEH-S) относится к генетическому элементу, который придает организму хозяина признак, который можно отобрать, проследить или выявить оптическими датчиками, ПЦР-амплификацией, биохимическими анализами или анализами жизнеспособности клетки/организма (резистентность или токсичность, выявляемая на клетках или организмах при обработке соответствующим антибиотиком или химическим средством).
Термины «промотор гена» или «промотор» (P) могут быть использованы взаимозаменяемо и относятся к нуклеотидной последовательности, необходимой для экспрессии гена.
Термины «вставка» и «модуль» по существу взаимозаменяемы только с тонким различием, которое заключается в том, что «вставку» встраивают в вектор, а после того, как она встроена, ее в большинстве случаев называют «модуль». Затем модуль может быть удален из вектора. Также термин вставка обычно используют в отношении изолированного модуля, используемого в качестве вставки в вектор, являющийся акцептором модуля.
«Интрон» относится к нуклеотидным последовательностям не кодирующей белок области гена, находящейся между двумя кодирующими белок областями или экзонами.
«Сигнал локализации» (LOC) относится к нуклеотидным последовательностям, кодирующим сигнал для определения маршрута представляющего интерес белка в клетке.
«Сайт множественного клонирования» (MCS) относится к нуклеотидным последовательностям, содержащим по меньшей мере один уникальный сайт рестрикции и более обычно группу уникальных сайтов рестрикции для целей клонирования фрагмента ДНК в клонирующей векторной плазмиде.
Термин «элемент стабилизации мРНК» относится к последовательности ДНК, которую узнают связывающие белки, которые, как полагают, защищают некоторые мРНК от расщепления.
Термин «действуют, определяя» в случае применения по отношению к группе сайтов рестрикции означает, что в том случае, когда один любой из сайтов рестрикции в группе разрезается одним узнающим ферментом рестрикции, выступающий остаток представляет собой 5'-часть по отношению к нуклеотидным последовательностям, расположенным даунстрим от группы сайтов рестрикции.
Термин «начало репликации» (ORI) относится к нуклеотидным последовательностям, которые управляют репликацией или удвоением плазмиды в клетке-хозяине.
«Методика клонирования с использованием ПЦР с праймерами, имеющими выступающие концы с терминаторами» относится к способу амплификации генетических модулей с использованием полимеразной цепной реакции с однонитевыми ДНК-праймерами с защищенными 5'-выступающими нуклеотидами, которые могут служить в качестве сайтов связывания с комплементарными выступающими концами ДНК.
«Поли-A-хвост» относится к последовательности адениновых (A) нуклеотидов, обычно встречающихся на конце молекул матричной РНК (мРНК). Сигнал поли-A-хвоста вводят в 3'-концы конструкций ДНК или трансгенов, чтобы облегчить экспрессию представляющего интерес гена.
«Сайт праймера» относится к нуклеотидным последовательностям, которые служат в качестве ДНК-матриц, на которых могут отжигаться однонитевые олигонуклеотиды ДНК в целях инициации секвенирования ДНК, ПЦР-амплификации и/или транскрипции РНК.
Термин «pUC19» относится к плазмидному клонирующему вектору, хорошо известному специалистам в данной области, и может быть найден в базе данных NCBI Genbank с номером доступа L09137.
Термин «случайные нуклеотидные последовательности» относится к любой комбинации нуклеотидных последовательностей, которые не идентичны последовательностям, кодирующим другие элементы, охарактеризованные как компоненты той же самой молекулы. Количество нуклеотидов, необходимых в случайных последовательностях, зависит от требований ферментов рестрикции, которые фланкируют случайные последовательности. Для большинства эндонуклеаз требуется минимум 2-4 дополнительных случайных последовательности, чтобы стабилизировать связывание ДНК. Предпочтительно количество нуклеотидов в случайных последовательностей должно быть кратно 3, что соответствует количеству нуклеотидов, составляющих кодон. Поэтому предпочтительное минимальное количество нуклеотидов в случайных последовательностях составляет 6, однако можно использовать меньшее или большее количество нуклеотидов.
«Редкий» относится к сайту рестрикции, который встречается в геноме относительно нечасто.
«Плечо рекомбинации» относится к нуклеотидным последовательностям, которые способствуют гомологичной рекомбинации между трансгенной ДНК и геномной ДНК. Для успешной рекомбинации требуется присутствие левого плеча рекомбинации (LRA) и правого плеча рекомбинации (RRA), фланкирующих область трансгенной ДНК, которую необходимо включить в геном хозяина посредством гомологичной рекомбинации.
«Рекомбинация» относится к процессу использования случайных или сайт-специфичных ферментов рекомбиназ наряду с последовательностями ДНК, которые могут быть подвергнуты воздействию ферментов рекомбиназ для транслокации части генетического материала из одной молекулы ДНК в другую молекулу ДНК.
«Репортерный ген» относится к нуклеотидным последовательностям, кодирующим белок, применимый для регистрации активности конкретного представляющего интерес промотора.
«Эндонуклеаза рестрикции» или «фермент рестрикции» относится к представителю или представителям в классификации каталитических молекул, которые связывают узнаваемую ими последовательность ДНК и расщепляют молекулу ДНК в точном положении в данной последовательности.
«Челночный вектор» относится к специализированному клонирующему плазмидному вектору, применяемому в изобретении для получения промежуточной молекулы, которая будет модифицировать концы фрагмента ДНК.
«Последовательность метки» (TAG) относится к нуклеотидным последовательностям, кодирующим уникальную область белка, которая позволяет его выявлять или, в некоторых случаях, отличать от любого эндогенного аналога.
«Нетранслируемая область» (UTR) относится к нуклеотидным последовательностям, охватывающим не кодирующую белок область молекулы мРНК. Указанные нетранслируемые области могут находиться на 5'-конце (5'-UTR) или 3'-конце (3'-UTR) молекулы мРНК.
«Уникальный» относится к любому сайту для эндонуклеаз рестрикции или HE, который не встречается в другом месте в конкретной молекуле ДНК.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к структурам и способам создания мультигенного стыковочного вектора, обычно плазмидного вектора, также называемого модульным клонирующим вектором, для синтеза трансгена PE3 или другой сложной конструкции ДНК посредством получения остова, имеющего точки клонирования модулей или точки стыковки. Изобретение применимо для сборки различных фрагментов ДНК в конструкции ДНК de novo или трансгене с использованием клонирующих векторов, оптимизированных для уменьшения количества зачастую необходимых манипуляций. Вектор трансгена PE3, называемый в настоящем описании стыковочным вектором или стыковочной плазмидой PE3, обычно содержит по меньшей мере один сайт множественного клонирования (MCS) и разнообразные группы редких сайтов рестрикции и/или уникальных сайтов хоминг-эндонуклеаз («HE»), расположенных в линейном порядке. Такое расположение определяет модульную структуру, которая дает возможность пользователю помещать доменные модули в виде вставок в трансгенную конструкцию PE3, не нарушая целостность элементов ДНК, уже включенных в стыковочную плазмиду на предыдущих стадиях клонирования.
Хотя настоящее изобретение раскрывает и иллюстрирует модульные векторы и способы создания модулей генетических доменов в модулях трансгена PE3 и модулей трансгена PE3 для встраивания в мультигенные векторы, которые представляют собой плазмидные клонирующие векторы, сходные способы могут быть использованы для создания модулей генетических доменов и модулей трансгена PE3 в более крупных внехромосомных молекулах ДНК, таких как космиды или искусственные хромосомы, включая бактериальные искусственные хромосомы (BAC). Широкое множество генетических элементов, которые могут быть включены в плазмидные клонирующие векторы, также позволяют переносить конечные трансгенные продукты PE3 в широкое множество организмов-хозяев с помощью небольшой или без дополнительной обработки.
Настоящее изобретение предполагает, что оба конца, 5'- и 3'-концы, каждой из точек стыковки (генных шарниров) и каждого из доменных модулей или вставок, все совместимы с соответствующим концом другой точки стыковки или вставки. Например, если первая точка стыковки содержит сайт рестрикции для невариабельного редкого фермента рестрикции, такого как SgrAI, и такая точка стыковки затем расщепляется, то первая вставка, предназначенная для встраивания на 3'-конце расщепленной первой точки стыковки, будет содержать совместимый 5'-конец для создания сайта рестрикции для SgrAI при объединении вставки с первой точкой стыковки. Вторая точка стыковки в плазмиде может, например, иметь сайт рестрикции для невариабельного фермента рестрикции, такого как SwaI. Любая вторая вставка будет иметь на своем 3'-конце совместимую нуклеотидную последовательность для объединения с расщепленным 5'-концом расщепленной второй точки стыковки, чтобы создать сайт рестрикции для SwaI. Кроме того, 3'-конец первой вставки и 5'-конец второй вставки для того, чтобы их можно было соответствующим образом встроить в модульную клонирующую векторную плазмиду, а также впоследствии удалить в той же самой точке, должны содержать совместимые концы для создания третьего сайта рестрикции для третьего невариабельного редкого фермента рестрикции, такого как, например, PacI или SalI.
Векторы для стыковки доменных модулей согласно изобретению, также известные как «челночные векторы», для перемещения доменных модулей в стыковочный вектор PE3 содержат сайт множественного клонирования с широко распространенными сайтами рестрикции, которые фланкированы по меньшей мере редкими сайтами рестрикции и необязательно сайтами HE. Челночные векторы конструируют для клонирования фрагментов ДНК в широко распространенных сайтах рестрикции между редкими сайтами рестрикции. Клонированные фрагменты затем можно высвобождать расщеплением в редком сайте или сайтах рестрикции (или HE) и вводить в стыковочный вектор PE3 с использованием такого же редкого сайта или сайтов рестрикции и/или сайтов HE, встречающихся в челночных векторах.
Таким образом, в отличие от обычных клонирующих векторов конструирование MCS позволяет встраивать доменные модули («кассеты» или модули фрагментов ДНК) в модульные области стыковочного вектора (стыковочной плазмиды) PE3, используя определенные генные шарниры, содержащие группу редких сайтов рестрикции. Подобным образом, каждый из них может быть легко удален с использованием таких же редких ферментов рестрикции и/или ферментов HE и заменен любым другим представляющим интерес фрагментом ДНК. Указанная особенность дает возможность пользователю быстро и легко изменять направление экспериментального исследования без необходимости заново конструировать полную конструкцию ДНК. Поэтому доменный модуль и клонирующие векторы PE3 согласно настоящему изобретению обеспечивают пользователю возможность клонировать фрагмент ДНК в промежуточном векторе доменного модуля с использованием широко распространенных сайтов рестрикции, создавать модуль, который является акцептором кассеты, и затем переносить указанный модульный фрагмент в требуемое место для модуля в конечной конструкции с помощью редких сайтов рестрикции. Кроме того, это позволяет в будущем осуществлять изменения в молекуле, чтобы заменить отдельные модули в стыковочной плазмиде PE3 другими кассетными модулями. В приведенном ниже описании основное место уделено отличиям настоящего изобретения от известного уровня техники.
Каждая точка стыковки (определенная как генный шарнир) представляет собой область, в которой предпочтительно присутствуют по меньшей мере два постоянных невариабельных редких сайта рестрикции и более предпочтительно постоянная группировка по меньшей мере из трех невариабельных редких сайтов рестрикции, и наиболее предпочтительно постоянная группировка не более, чем из 4 невариабельных редких сайтов рестрикции. Конкретный сайт рестрикции каждой точки стыковки расщепляют узнающим его ферментом рестрикции. При этом будет создаваться либо требуемый 5'-конец, либо 3'-конец, который совместим с комплементарным 5'- или 3'-концом одной из предварительно сконструированных вставок, содержащих выбранную нуклеотидную последовательность, такую как промотор, экспрессируемая нуклеотидная последовательность или регуляторная нуклеотидная последовательность. По меньшей мере две вставки, каждая из которых имеет 5'- и 3'-концы, которые совместимы с представляющей интерес расщепленной точкой стыковки, могут быть добавлены вместе с расщепленной клонирующей векторной плазмидой в подходящую реакционную смесь, и при условии наличия надлежащей термодинамической среды, вставки могут одновременно, то есть в одну стадию, интегрироваться в клонирующую векторную плазмиду. В ходе такого единственного добавления и реакции лигирования снова образуются точки стыковки, и клонирующая векторная плазмида становится модульной, так как точки стыковки и соединение между двумя модулями могут быть снова расщеплены подходящими ферментами рестрикции. Впоследствии модуль может быть удален, а новый модуль может быть помещен на его место.
Хотя можно использовать один сайт рестрикции в шарнире, фланкирующей модули, существует явная возможность того, что один «редкий» сайт рестрикции довольно часто встречается в данной конкретной молекуле ДНК. Напомним, что частота сайтов для ферментов рестрикции является функцией 4n, как объясняется выше. Например, конкретный модуль промотора в клонирующей векторной плазмиде может содержать редкий сайт рестрикции в последовательности ДНК, который также присутствует в шарнире, следовательно предпочтительным является наличие более одного сайта рестрикции в шарнирах. Таким образом, предпочтительно использование более одного редкого сайта рестрикции в одном шарнире, поскольку статистическая вероятность наличия более одного сайта рестрикции в последовательности ДНК представляющего интерес модуля намного ниже. Однако, также предпочтительно, чтобы не более трех или четырех сайтов рестрикции располагались в одном шарнире, или в противном случае «группирование вместе» сайтов рестрикции начнет влиять на транскрипцию/трансляцию молекулы-мишени. При таком количестве комбинация сайтов рестрикции, сгруппированных вместе, будет длиной не больше 18-24 п.н.
Векторы доменных модулей
На фиг.1 приведено упрощенное представление первого варианта осуществления настоящего изобретения, то есть вектора 1 для стыковки доменных модулей. Вектор 1 содержит нить ДНК, имеющую сайт 2 множественного клонирования, и обычно представляет собой плазмиду. Вектор для стыковки доменных модулей содержит модуль множественного клонирования (модуль MC), состоящий из пяти сайтов клонирования, расположенных в следующей последовательности, MC-1, MC-2, MC-3, MC-4 и MC-5. Сайт множественного клонирования содержит множество сайтов рестрикции, которые независимо выбраны из широко распространенных сайтов рестрикции, которые указаны в настоящем описании ниже. Два сайта рестрикции определяют положение стыковки для представляющего интерес генетического материала, показанного в виде представляющего интерес гена 3. Модуль MC позволяет субклонировать представляющий интерес генетический материал между двумя сайтами рестрикции в сайте множественного клонирования модуля MC. Представляющий интерес ген 3 обычно высвобождают из вектора, несущего представляющий интерес ген (не показан), и он содержит пару сайтов для клонирования 4a и 4b, показанных в виде MC-1 и MC-3.
Вектор для стыковки доменных модулей также содержит пару сайтов рестрикции, называемых генными шарнирами 5 и 6, которые фланкируют сайт 2 множественного клонирования. Каждый из генных шарниров содержит по меньшей мере два невариабельных редких сайта рестрикции, которые определены в настоящем описании ниже. Генные шарниры 5 и 6 работают, определяя 5'- и 3'-части соответственно модуля MC.
Во втором варианте осуществления изобретения узнающие ферменты рестрикции MC-1 и MC-3 (не показаны) могут вырезать представляющий интерес ген из его вектора и открывать модуль MC в обоих MC-сайтах MC-1 и MC-3, таким образом обеспечивая возможность лигирования представляющего интерес гена между MC-сайтами MC-1 и MC-3 с образованием вектора 7 доменного модуля. В показанном варианте представляющий интерес ген содержит домен экспрессии, так что доменный модуль является модулем экспрессии, а вектор доменного модуля более предпочтительно является вектором модуля экспрессии. Вектор модуля экспрессии содержит модуль 8 экспрессии, который содержит первый и второй генные шарниры 5 и 6, которые фланкируют последовательность нуклеиновой кислоты, содержащую субклонируемый представляющий интерес ген 3, который включает домен экспрессии.
В показанном варианте представляющий интерес ген содержит домен экспрессии, в котором первый генный шарнир (или 5'-часть домена экспрессии) в дальнейшем называют GP2, а второй генный шарнир (или 3'-часть домена экспрессии) называют GP3. Можно понять, что в альтернативных вариантах представляющий интерес ген может содержать домен промотора или 3'-регуляторный домен, при этом их субклонирование дает модуль промотора в векторе модуля промотора и 3'-регуляторный модуль в векторе 3'-регуляторного модуля, соответственно. В случае стыковочных векторов для модуля промотора первый генный шарнир (или 5'-часть домена промотора) в дальнейшем называют GP1, а второй генный шарнир (или 3'-часть домена промотора) называют GP2. В случае стыковочных векторов для 3'-регуляторного модуля первый генный шарнир (или 5'-часть 3'-регуляторного домена) в дальнейшем называют GP3, а второй генный шарнир (или 3'-часть 3'- регуляторного домена) называют GP4.
Генные шарниры в случае любого вектора для стыковки доменных модулей, включая генные шарниры GP1, GP2, GP3 и GP4, могут содержать редкие сайты рестрикции, выбранные из группы, состоящей из AsiS I, Pac I, Sbf I, Fse I, Asc I, Mlu I, SnaB I, Not I, Sal I, Swa I, Rsr II, BSiW I, Sfo I, Sgr AI, AflIII, Pvu I, Ngo MIV, Ase I, Flp I, Pme I, Sda I, Sgf I, Srf I и Sse8781 I, и обычно выбранные из группы, состоящей из AsiS I, Pac I, Sbf I, Fse I, Asc I, Mlu I, SnaB I, Not I, Sal I, Swa I, Rsr II, BSiW I. Обычные варианты генного шарнира содержат серию по меньшей мере из 3 и не более 4 редких сайтов рестрикции, и обычно любой генный шарнир не содержит редкого сайта рестрикции, имеющегося в любом другом генном шарнире.
В предпочтительном варианте генный шарнир GP1 выбран из группы, состоящей по меньшей мере из AsiS I, Pac I и Sbf I, генный шарнир GP2 выбран из группы, состоящей по меньшей мере из Fse I, Asc I и Mlu I, генный шарнир GP3 выбран из группы, состоящей по меньшей мере из SnaB I, Not I и Sal I, и генный шарнир GP4 выбран из группы, состоящей по меньшей мере из Swa I, Rsr II и BSiW I. В конкретном предпочтительном варианте генный шарнир GP1 состоит из AsiS I, Pac I и Sbf I в указанном порядке; генный шарнир GP2 состоит из Fse I, Asc I и Mlu I в указанном порядке; генный шарнир GP3 состоит из SnaB I, Not I и Sal I в указанном порядке; и генный шарнир GP4 состоит из Swa I, Rsr II и BSiW I в указанном порядке.
Также будет понятно, что сайт множественного клонирования может содержать другие комбинации сайтов рестрикции с большим или меньшим количеством сайтов, в обратном порядке (например, MC-5, MC-4, MC-3, MC-2 и MC-1), и можно использовать другие типы сайтов клонирования. Сайты множественного клонирования могут варьировать, чтобы приспособить пару сайтов клонирования для конкретного представляющего интерес гена.
Также в альтернативном варианте предусмотрена библиотека векторов для стыковки доменных модулей, которую можно использовать для простого субклонирования широкого множества представляющих интерес генов. Библиотека векторов для стыковки доменных модулей может быть иметь такую конфигурацию, чтобы служить в качестве специализированного вектора для модуля, содержащего домен промотора, вектора для модуля, содержащего домен экспрессии, или вектора для 3'-регуляторного модуля, в зависимости от типа генных шарниров, выбранных для конкретного вектора для стыковки доменных модулей. Каждая подбиблиотека векторов для модуля домена промотора, домена экспрессии и 3'-регуляторного домена имеет свою собственную специальную пару генных шарниров (а именно, GP1 и GP2, GP2 и GP3 и GP3 и GP4), чтобы обеспечить правильное клонирование соответствующих доменных модулей в стыковочных векторах PE3 более высокого порядка, которые описаны далее.
Стыковочные векторы PE3 и стыковочные векторы PE3 MC
На фиг.2 показано упрощенное представление третьего варианта осуществления настоящего изобретения, первого стыковочного вектора 10 PE3. Стыковочный вектор 10 PE3 состоит из нити ДНК, имеющей по меньшей мере первый модуль 11b клонирования, и обычно представляет собой плазмиду. По меньшей мере первый модуль клонирования содержит по меньшей мере первый и второй генные шарниры, показанные как GP2 и GP3, которые фланкируют последовательность нуклеиновой кислоты, которая содержит вкладыш 18b. Домен вкладыша ДНК представляет собой случайную нуклеотидную последовательность, которая не кодирует сайт рестрикции или какую-либо другую биологическую функцию, присущую стыковочному вектору PE3. ДНК вкладыша служит для повышения эффективности разрезающей активности ферментов рестрикции посредством обеспечения более длинных участков ДНК, с которыми фермент рестрикции может связываться. Это имеет важное значение, поскольку многие ферменты рестрикции не могут связываться и разрезать узнаваемые ими сайты, если длина ДНК ограничена. Первый и второй генные шарниры определены в описании выше. Первый модуль 11b клонирования имеет конфигурацию для клонирования по меньшей мере первого доменного модуля в модуле клонирования PE3. В показанном варианте, где первый модуль клонирования является модулем клонирования домена экспрессии, первый и второй генные шарниры указаны в виде GP2 и GP3, которые определены в описании выше. Первый модуль 11b клонирования, содержащий вкладыш, также называемый модулем вкладыша для встраивания домена экспрессии, обеспечивает место стыковки при встраивания модуля домена экспрессии, содержащего совместимые генные шарниры. Совместимым генным шарниром является шарнир, который имеет идентичные сайты рестрикции или который содержит по меньшей мере один уникальный редкий сайт рестрикции, такой как и в генном шарнире в модуле клонирования.
Также показано, что второй модуль 11a клонирования является модулем вкладыша для встраивания промотора, имеющим первый генный шарнир 12 в виде GP1 и второй генный шарнир 13 в виде GP2, который является областью соединения в одно целое с 5'-частью модуля 11b клонирования. Модуль 11a вкладыша для встраивания промотора содержит вкладыш и обеспечивает место стыковки при встраивании модуля домена промотора, имеющего совместимые генные шарниры, который заменяет модуль вкладыша для встраивания промотора при клонировании. Третий модуль 11c клонирования представляет собой модуль вкладыша для встраивания 3'-регуляторного домена, имеющий первый генный шарнир 14 в виде GP3, который является областью соединения в одно целое с 3'-частью модуля 11b клонирования, и второй генный шарнир 15 в виде GP4. Модуль 11c вкладыша для встраивания 3'-регуляторного домена содержит вкладыш и обеспечивает место стыковки при встраивании модуля 3'-регуляторного домена, имеющего совместимые генные шарниры, который заменяет модуль вкладыша 3'-регуляторного домена при клонировании.
В четвертом варианте осуществления изобретения ферменты рестрикции, узнающие один из редких сайтов рестрикции, находящихся в каждом генном шарнире для одного из доменных модульных векторов, показанного модуля 7 экспрессии, могут отрезать редкий сайт рестрикции от остальной части вектора и подобным образом раскрывать стыковочный вектор PE3 в таких же редких сайтах рестрикции в соответствующих генных шарнирах GP2 и GP3, таким образом заменяя модуль вкладыша для встраивания модуля экспрессии модулем экспрессии. В отдельном событии клонирования подходящие ферменты рестрикции, узнающие генные шарниры модуля промотора (не оказан) и соответствующие генные шарниры 12 и 13 модуля 11a вкладыша для встраивания промотора, можно использовать для того, чтобы лигировать модуль промотора в модуль PE3, таким образом заменяя модуль вкладыша для встраивания промотора модулем промотора. Подобным образом, 3'-регуляторный модуль может быть лигирован в модуль PE3.
В настоящем изобретении по меньшей мере два и более, обычно по меньшей мере три редких сайта рестрикции используют в качестве стыковочных шарниров или генных шарниров. Редкость встречаемости редкого сайта рестрикции в генетических последовательностях делает маловероятным событие, когда оба редких сайта рестрикции в содержащем два сайта генном шарнире могут встречаться в стыковочном векторе, и очень невероятным событие, когда все три редких сайта рестрикции в содержащем три сайта генном шарнире могут встречаться в стыковочном векторе. В случае когда обнаружено, что клонирующий ДНК вектор содержит один из редких сайтов рестрикции генного шарнира, специалист может разрезать генный шарнир любым другим узнающим редким ферментом рестрикции. Как показано, если генный шарнир GP2 состоит из Fse I, Asc I и Mlu I, а стыковочный вектор PE3 или домен E сам по себе имеет Asc I в последовательности, то специалист может использовать либо Fse I, либо Mlu I для расщепления генных шарниров.
Клонирующий ДНК вектор в случае вектора PE3 также обычно содержит средства для высвобождения модуля клонирования PE3 из стыковочного вектора PE3 для клонирования в мультигенном стыковочном векторе, который будет описан далее.
На фиг.3 показан альтернативный вариант клонирования доменных модулей в стыковочном векторе 20 для множественного клонирования (MC) PE3. Первый стыковочный вектор PE3 MC, называемый стыковочным вектором 20a PE3 MC-промотор, состоит из нити ДНК, имеющей по меньшей мере первый модуль 21a клонирования и обычно представляет собой плазмиду. Первый модуль 21a клонирования содержит по меньшей мере первый и второй генные шарниры, показанные в виде GP1 и GP2, которые фланкируют последовательность нуклеиновой кислоты, которая содержит сайт множественного клонирования (MCS), содержащий множество сайтов рестрикции, независимо выбранных из широко распространенных сайтов рестрикции, которые описаны ниже. В показанном варианте стыковочный вектор 20 PE3 MC-промотор имеет три модуля 21a, 21b и 21c клонирования, каждый из которых связан генными шарнирами GP1, GP2, GP3 и GP4, которые описаны в настоящем описании. Второй и третий модули клонирования показаны в виде модулей вкладышей, которые содержат последовательность нуклеиновой кислоты, которая содержит вкладыш.
Сайт множественного клонирования состоит из пяти сайтов клонирования, расположенных в следующем порядке: MC-1, MC-2, MC-3, MC-4, MC-5 и MC-6. Любые два сайта рестрикции могут определять место стыковки для представляющего интерес генетического материала. В стыковочном векторе 20a PE3 MC представляющий интерес генетический материал обычно представляет собой домен промотора. Выбор двух представляющих интерес сайтов клонирования может быть основан на понимании того, что такие сайты рестрикции являются уникальными в стыковочном векторе 20a PE3 MC.
Альтернативные варианты включают стыковочный вектор PE3 MC-экспрессия, в котором второй модуль 21b клонирования содержит сайт множественного клонирования, при этом остальные первый и третий модули 21a и 21c клонирования содержат вкладыш, и стыковочный вектор PE3 MC-3-регуляторный, в котором третий модуль 21c клонирования содержит сайт множественного клонирования, при этом остальные первый и второй модули 21a и 21b клонирования содержат вкладыш. В других альтернативных вариантах два из трех модулей клонирования могут содержать сайт множественного клонирования.
Субклонирование сайта множественного клонирования с представляющим интерес генетическим материалом и клонирование доменных модулей в модулях вкладышей в стыковочных векторах PE3 можно осуществить как описано выше.
Генные шарниры GP1, GP2, GP3 и GP4 описаны выше при описании положения соответствующих модулей в модулях домена промотора, домена экспрессии и 3'-регуляторного домена.
Настоящее изобретение также относится к получению векторов PE3 из модульных доменных векторов и клонирующего модульного вектора PE3. Первый способ конструирования модульного вектора PE3 включает стадии:
a) получения клонирующего вектора PE3, содержащего модуль клонирования PE3, при этом модуль клонирования PE3 содержит в следующем порядке: первый генный шарнир, содержащий по меньшей мере два невариабельных редких сайта рестрикции, по меньшей мере модуль первого вкладыша, состоящий из последовательности нуклеиновой кислоты, содержащей вкладыш, и второй генный шарнир;
b) получения по меньшей мере первого доменного модульного вектора, содержащего в указанном порядке: первый генный шарнир, представляющий интерес генетический модуль, состоящий из последовательности нуклеиновой кислоты, содержащей представляющий интерес генетический материал; и второй генный шарнир;
c) предоставления первого фермента рестрикции, узнающего один из редких сайтов рестрикции первого генного шарнира, и второго фермента рестрикции, узнающего один из редких сайтов рестрикции второго генного шарнира;
d) вырезания и выделения представляющего интерес генетического модуля из первого доменного модульного вектора с использованием первого и второго узнающих ферментов рестрикции;
e) вырезания модуля первого вкладыша из модуля клонирования PE3 клонирующего вектора PE3 с использованием первого и второго узнающих ферментов рестрикции; и
f) лигирования представляющего интерес генетического модуля в модуль клонирования PE3.
Кроме того, также предлагается способ, при котором получаемый модуль клонирования PE3 дополнительно содержит в указанном порядке после второго генного шарнира модуль второго вкладыша и третий генный шарнир, с использованием следующих дополнительных стадий:
g) получения второго доменного модульного вектора, содержащего в указанном порядке второй генный шарнир, второй представляющий интерес генетический модуль, состоящий из последовательности нуклеиновой кислоты, содержащей второй представляющий интерес генетический материал; и третий генный шарнир;
h) предоставления третьего фермента рестрикции, узнающего один из редких сайтов рестрикции третьего генного шарнира;
i) вырезания и выделения второго представляющего интерес генетического модуля из второго доменного модульного вектора с использованием второго и третьего узнающих ферментов рестрикции;
j) вырезания модуля второго вкладыша из модуля клонирования PE3 клонирующего вектора PE3 с использованием второго и третьего узнающих ферментов рестрикции; и
k) лигирования второго представляющего интерес генетического модуля в модуль клонирования PE3.
Подобным образом третий представляющий интерес генетический модуль может быть встроен в PE3 с использованием третьего генного шарнира и узнающих ферментов рестрикции для вырезания и лигирования второго представляющего интерес генетического модуля в модуль клонирования PE3. Способ обеспечивает последовательное расположение представляющих интерес генетических модулей в клонирующем векторе PE3. Обычно первый, второй и третий представляющие интерес генетические модули соответствуют модулю промотора, модулю экспрессии и 3'-регуляторному модулю, соответственно. В типичном варианте первым генным шарниром является GP1, вторым генным шарниром является GP2, третьим генным шарниром является GP3 и четвертым генным шарниром является GP4, которые описаны выше.
В другом варианте предлагается способ получения клонирующего вектора PE3, при этом способ включает стадии:
a) получения клонирующего вектора PE3, имеющего остов, при этом остов содержит по меньшей мере первый, второй, третий и четвертый генные шарниры, указанные генные шарниры расположены последовательно в 5'-3'-направлении и каждый из них имеет по меньшей мере два невариабельных редких сайта рестрикции, которые можно подвергнуть обработке, чтобы расщепить узнающим их ферментом рестрикции;
b) расщепления первого генного шарнира первым ферментом рестрикции, узнающим один из невариабельных редких сайтов рестрикции первого генного шарнира, оставляющим расщепленный первый генный шарнир с незащищенным 3'-концом;
c) расщепления второго генного шарнира вторым ферментом рестрикции, узнающим один из невариабельных редких сайтов рестрикции второго генного шарнира, оставляющим расщепленный второй генный шарнир с незащищенным 5'-концом;
d) получения модуля первого домена, содержащего 5'-конец, первый представляющий интерес генетический материал и 3'-конец, при этом 5'-конец модуля первого домена совместим с незащищенным 3'-концом расщепленного первого генного шарнира, а 3'-конец первого домена совместим с незащищенным 5'-концом расщепленного второго генного шарнира; и
e) помещения модуля первого домена и расщепленного клонирующего вектора PE3 в подходящую реакционную смесь, чтобы вызывать лигирование и самоориентацию модуля первого домена в остове между первым генным шарниром и вторым генным шарниром, в которой остов подвергается повторной сборке.
Способ дополнительно может включать стадии:
f) дальнейшего расщепления второго генного шарнира вторым узнающим ферментом рестрикции, оставляющим расщепленный второй генный шарнир с незащищенным 3'-концом;
g) расщепления третьего генного шарнира третьим ферментом рестрикции, узнающим один из редких сайтов рестрикции третьего генного шарнира, оставляющим расщепленный третий генный шарнир с незащищенным 5'-концом;
h) получения модуля второго домена, содержащего 5'-конец, первый представляющий интерес генетический материал и 3'-конец, при этом 5'-конец модуля второго домена совместим с незащищенным 3'-концом расщепленного второго генного шарнира, а 3'-конец модуля второго домена совместим с незащищенным 5'-концом расщепленного третьего генного шарнира; и
i) помещения модуля второго домена и расщепленного клонирующего вектора PE3 в подходящую реакционную смесь, чтобы вызывать лигирование и самоориентацию модуля второго домена в остове между вторым генным шарниром и третьим генным шарниром, в которой остов подвергается повторной сборке.
Способ также дополнительно может включать стадии:
j) дальнейшего расщепления остова в третьем генном шарнире третьим узнающим ферментом рестрикции, оставляющим расщепленный третий генный шарнир с незащищенным 3'-концом;
k) расщепления четвертого генного шарнира четвертым ферментом рестрикции, узнающим один из редких сайтов рестрикции четвертого генного шарнира, оставляющим расщепленный четвертый генный шарнир с незащищенным 5'-концом;
l) получения модуля третьего домена, содержащего 5'-конец, третий представляющий интерес генетический материал и 3'-конец, при этом 5'-конец модуля третьего домена совместим с незащищенным 3'-концом расщепленного третьего генного шарнира, а 3'-конец модуля третьего домена совместим с незащищенным 5'-концом расщепленного четвертого генного шарнира; и
m) помещения модуля третьего домена и расщепленного клонирующего вектора PE3 в подходящую реакционную смесь, чтобы вызывать лигирование и самоориентацию модуля третьего домена в остове между третьим генным шарниром и четвертым генным шарниром, в которой остов подвергается повторной сборке.
Обычно первый, второй и третий представляющие интерес генетические модули соответствуют модулю промотора, модулю экспрессии и 3'-регуляторному модулю, соответственно. В типичном варианте первым генным шарниром является GP1, вторым генным шарниром является GP2, третьим генным шарниром является GP3 и четвертым генным шарниром является GP4, которые описаны выше.
Мультигенные клонирующие векторы
Стыковочные векторы PE3 и стыковочные векторы MC согласно настоящему изобретению обеспечивают возможности простой и быстрой сборки одного или нескольких представляющих интерес трансгенов PE3. Изобретение также относится к способам быстрого и простого встраивания одного или нескольких модулей PE3 в модульный мультигенный вектор. На фиг.4 показано простое схематичное встраивание модуля PE3 в мультигенный стыковочный вектор, который содержит один или (как показано) несколько модулей вкладышей PE3, которые можно легко удалять, чтобы обеспечить инсерцию одного или нескольких модулей PE3. Мультигенные векторы более подробно описаны далее и на фиг.16-21.
Сайты рестрикции, используемые в изобретении, выбирают согласно уровню встречаемости. Чтобы определить частоту встречаемости сайтов рестрикции, информацию о последовательностях ДНК, соответствующих девятнадцати разным генам, анализировали, используя компьютерную программу Vector NTI. Такой поиск охватывал всего 110530 нуклеотидов последовательности ДНК. Результаты таких анализов рассчитывали на основании количества случаев встречаемости сайта рестрикции среди анализируемых 110530 нуклеотидов. Затем сайтам рестрикции давали иерархическое обозначение в соответствии с четырьмя уровнями классификации, при этом «широко распространенные» сайты встречаются более 25 раз на 110530 нуклеотидов, «менее часто встречающиеся сайты из 6 п.н.» встречаются 6-24 раза на 110530 нуклеотидов, и «редкие» сайты встречаются 0-5 раз на 110530 нуклеотидов. Неполный список «подходящих» ферментов приведен в настоящем описании согласно классификации встречаемости:
Широко распространенные сайты рестрикции:
Подходящие широко распространные ферменты рестрикции могут включать без ограничения Ase I, BamH I, Bgl II, Blp I, BstX I, EcoR I, Hinc II, Hind III, Nco I, Pst I, Sac I, Sac II, Sph I, Stu I, Xba I.
Подходящие сайты рестрикции, которые являются сайтами узнавания из 6 п.н., но имеют более низкую частоту встречаемости, могут включать без ограничения Aar I, Aat II, Afl II, Age I, ApaL I, Avr II, BseA I, BspD I, BspE I, BstB I, Cla I, Eag I, Eco0109 I, EcoR V, Hpa I, Kpn I, Mfe I, Nar I, Nde I, NgoM IV, Nhe I, Nsi I, Pml I, SexA I, Sma I, Spe I, Xho I.
Редкие сайты рестрикции:
Подходящие редкие ферменты могут включать без ограничения Acl I, Nru I, Pac I, Pme I, Sbf I, Sfi I, PI-Sce I, I-Sce I, I-Ceu I, PI-Psp I, I-Tli I, Fse I, Sfo I, Asi SI, Sgr AI, Asc I, Mlu I, Sna BI, Not I, Sal I, Swa I, Rsr II, Bsi WI, AflIII, Pvu I, Ngo MIV, Ase I, Flp I, Pme I, Sda I, Sgf I, Srf I и Sse8781 I.
Хоминг-эндонуклеазы:
Подходящие хоминг-эндонуклеазы могут включать без ограничения I-SceI, PI-Sce I, I-Ceu I, PI-Psp I, I-Chu I, I-Cmoe I, I-Cpa I, I-Cpa II, I-Cre I, I-Cvu I, I-Dmo I, I-Lla I, I-Mso I, I-Nan I, I-Nit I, I-Nja I, I-Pak I, I-Por I, I-Ppo I, I-Sca I, I-Sce II, I-Sce III, I-Sce IV, I-Sce V, I-Sce VI, I-Ssp68031, I-Tev I, I-Tev II, I-Two I, PI-Mga I, PI-Mtu I, PI-Pfu I, PI-Pfu II, PI-Pko I, PI-Pko II, PI-Tfu I, PI-Tfu II, PI-Thy I и PI-Tli.
ПОДРОБНЫЕ ВАРИАНТЫ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Отдельные компоненты вектора PE3 или трансгена представляют собой модуль промотора-энхансера (обозначаемый «P»), модуль экспрессируемого белка («E») и/или модуль 3'-регуляторной области («3»), и компоненты могут быть собраны в виде модулей, перенесенных из векторов, несущих модули доменов (или «челночных» векторов) в стыковочный вектор PE3 (который также можно называть стыковочным узлом). На фиг.5 показан модуль промотора в векторе промотора, который может быть вырезан и встроен в стыковочный вектор PE3 в предварительно определяемое положение для стыковки. Также на фиг.5 показано, что если необходима сложность более высокого порядка, то собираемые модульные трансгены PE3 или другие нуклеотидные последовательности затем могут быть перенесены в мультигенный стыковочный вектор (также называемый основным плазмидным стыковочным узлом) или в стыковочный вектор, направленный в определенный локус. Каждый из пяти типов клонирующих векторных плазмид (векторы модуля промотора, модуля экспрессии и 3'-регуляторного модуля, стыковочные векторы PE3 и мультигенные векторы) будет объяснен более подробно, чтобы показать компоненты, включенные в каждую плазмиду.
Векторы для стыковки доменных модулей, также известные как «челночные векторы», которые содержат сайт множественного клонирования, обычно содержащий широко распространенные сайты рестрикции и фланкированный генным шарниром, обычно содержащим редкий сайт рестрикции и/или HE-сайты. Вектор для стыковки доменных модулей конструируют из остова pUC19, и он имеет указанные далее модификации остова pUC19, где последовательности пронумерованы в соответствии с файлом последовательностей в Cenbank для pUC19, № доступа L09137:
1. В стыковочной плазмиде используют только последовательности от 806 до 2617 (Afl3-Aat2).
2. Сайт BspH1 в положении 1729 в pUC19 подвергают мутации от TCATGA до GCATGA.
3. Сайт Acl1 в положении 1493 в pUC19 подвергают мутации от AACGTT до AACGCT.
4. Сайт Acl1 в положении 1120 в pUC19 подвергают мутации от AACGTT до CACGCT.
5. Сайт Ahd1 в pUC19 подвергают мутации от GACNNNNNGTC до CACNNNNNGTC.
Последовательности, кодирующие BspH1/I-Ppo 1/BspH1, встраивают только в оставшийся сайт BspH1 в pUC19 после стадии мутирования 2 в указанном выше списке.
Три отдельных челночных вектора согласно настоящему изобретению, стыковочный вектор модуля промотора, стыковочный вектор модуля экспрессии и стыковочный вектор 3'-регуляторного модуля, описаны в конкретных вариантах и указаны как челночный вектор промотора/интрона («SVP»), челночный вектор экспрессии («SVE») и челночный вектор 3'-регуляторной области («SV3»), соответственно. Каждый вектор более полно описан ниже.
Челночный вектор P
SVP показан на фиг.10, и он представляет собой клонирующую векторную плазмиду, которую можно использовать для получения промотора и интронных последовательностей для сборки в стыковочном векторе PE3 (конструкции трансгена), который описан выше.
На фиг.11 показан пример плазмиды SVP, содержащей следующие последовательные элементы в MCS в порядке перечисления:
1. Два невариабельных и уникальных широко распространенных сайта рестрикции, которые определяют 5'-сайт встраивания для мутантного вектора pUC19, описанного выше (например, AatII и BlpI);
2. Сайт праймера T7;
3. Невариабельный и уникальный широко распространенный сайт рестрикции, который обеспечивает эффективное клонирование модуля челночного вектора даунстрим от сайта праймера T7 (например, Eco0109I);
4. Постоянную группу невариабельных редких сайтов рестрикции, которые определяют 5'-часть модуля промотора (например, AsiSI и SgrAI);
5. Вариабельный MCS, содержащий любую группу широко распространенных или редких сайтов рестрикции, которые являются уникальными для челночного вектора (например, серию сайтов рестрикции, показанных на фиг.11);
6. Постоянную группу невариабельных редких сайтов рестрикции, которые определяют 3'-часть модуля промотора (например, PacI, AscI и MluI);
7. Невариабельный и уникальный широко распространенный сайт рестрикции, который обеспечивает эффективное клонирование модуля челночного вектора апстрим от сайта праймера T3 (например, BspEI);
8. Сайт праймера в обратной ориентации T3; и
9. Два невариабельных и уникальных широко распространенных сайта рестрикции, которые определяют 3'-сайт встраивания для мутантного вектора pUC19, описанного выше (например, PmeI и SapI).
В альтернативном примере постоянная группа невариабельных редких сайтов рестрикции, которые определяют 5'-часть модуля промотора (генный шарнир GP1), включает AsiS I, Pac I и Sbf I, и редкие сайты рестрикции, которые определяют 3'-часть модуля промотора (генный шарнир GP2), включают Fse I, Asc I и Mlu I. В более конкретном примере генный шарнир GP1 состоит из последовательности AsiS I, Pac I и Sbf I, а генный шарнир GP2 состоит из последовательности Fse I, Asc I и Mlu I.
Челночный вектор E (SVE)
SVE, показанный на фиг.12, является клонирующей векторной плазмидой, которая может быть использована для получения последовательностей, экспрессируемых трансгеном, для модульной сборки в векторе PE3 (конструкции трансгена), который описан ранее.
На фиг.13 показан пример плазмиды SVE, содержащий следующие последовательно расположенные в порядке перечисления элементы в MCS:
1. Два невариабельных и уникальных широко распространенных сайтов рестрикции, которые определяют 5'-сайт встраивания для мутантного вектора pUC19, описанного выше (например, BlpI);
2. Сайт праймера T7;
3. Невариабельный и уникальный широко распространенный сайт рестрикции, который обеспечивает эффективное клонирование модуля челночного вектора даунстрим от сайта праймера T7 (например, Eco0109I);
4. Постоянную группу невариабельных редких сайтов рестрикции, которые определяют 5'-часть модуля экспрессии (например, PacI, AscI и MluI);
5. Вариабельный MCS, состоящий из любой группы широко распространенных или редких сайтов рестрикции, которые являются уникальными в челночном векторе (например, ряд сайтов рестрикции, показанных на фиг.13);
6. Постоянную группу невариабельных редких сайтов рестрикции, которые определяют 3'-часть модуля экспрессии (например, SnaBI, NotI и SalI);
7. Невариабельный и уникальный широко распространенный сайт рестрикции, который обеспечивают эффективное клонирование модуля челночного вектора апстрим от сайта праймера T3 (например, BspEI);
8. Сайт праймера T3 в обратной ориентации; и
9. Два невариабельных и уникальных широко распространенных сайтов рестрикции, которые определяют 3'-сайт встраивания мутантного вектора pUC19, описанного выше (например, PmeI).
В альтернативном примере постоянная группа невариабельных редких сайтов рестрикции, которые определяют 5'-часть модуля экспрессии (генный шарнир GP2), включает Fse I, Asc I и Mlu I, и редкие сайты рестрикции, которые определяют 3'-часть модуля экспрессии (генный шарнир GP3) включают SnaB I, Not I и Sal I. В более конкретном примере генный шарнир GP2 состоит из последовательности Fse I, Asc I и Mlu I, а генный шарнир GP3 состоит из последовательности SnaB I, Not I и Sal I.
Челночный вектор 3 (SV3)
SV3, показанный на фиг.14, представляет собой клонирующую векторную плазмиду, которую можно использовать для получения 3'-регуляторных последовательностей для сборки в векторе PE3 (конструкции трансгена), который описан выше.
На фиг.15 пример плазмиды SV3 может содержать следующие элементы в MCS в порядке перечисления:
1. Два невариабельных и уникальных широко распространенных сайта рестрикции, которые определяют 5'-сайт встраивания для мутантного вектора pUC19. описанного выше (например, BlpI);
2. Сайт праймера T7;
3. Невариабельный и уникальный широко распространенный сайт рестрикции, который обеспечивает эффективное клонирование модуля челночного вектора даунстрим от праймера T7 (например, Eco0109I);
4. Постоянную группу невариабельных редких сайтов рестрикции, которые определяют 5'-часть 3'-регуляторного модуля (например, SnaBI, NotI и SalI);
5. Вариабельный MCS, состоящий из любой группы широко распространенных или редких сайтов рестрикции, которые являются уникальными в челночном векторе (например, ряд сайтов рестрикции, показанных на фиг.15);
6. Постоянную группу невариабельных редких сайтов рестрикции, которые определяют 3'-часть 3'-регуляторного модуля (например, SwaI, RsrII и BsiWI);
7. Невариабельный и уникальный нередкий сайт рестрикции, который обеспечивает эффективное клонирование модуля челночного вектора апстрим от сайта праймера T3 (например, BspEI);
8. Сайт праймера T3 в обратной ориентации; и
9. Два невариабельных и уникальных нередких сайта рестрикции, которые определяют 3'-сайт встраивания для мутантного вектора pUC19, описанного выше (например, PmeI).
В альтернативном примере постоянная группа невариабельных редких сайтов рестрикции, которые определяют 5'-часть модуля экспрессии (генный шарнир GP3), включает SnaB I, Not I и Sal I, и редкие сайты рестрикции, которые определяют 3'-часть модуля экспрессии (генный шарнир GP4), включают Swa I, Rsr II и BsiW I. В более конкретном примере генный шарнир GP3 состоит из последовательности SnaB I, Not I и Sal I, и генный шарнир GP4 состоит из последовательности Swa I, Rsr II и BsiW I.
Стыковочный вектор PE3 (стыковочная плазмида PE3), показанный на фиг.6, содержит остов pUC19, модифицированный как описано в примере выше.
Сайт множественного клонирования (MCS) в стыковочной плазмиде PE3, показанный на фиг.7, содержит следующие последовательные элементы в порядке перечисления:
1. Три невариабельных и уникальных широко распространенных сайта рестрикции, которые определяют 5'-сайт встраивания для мутантного вектора pUC19, описанного выше (например, Aat II, Blp I и Eco0109 I);
2. Сайт праймера T7;
3. Первый уникальный сайт HE (например, I-SceI (в этом месте в прямой ориентации));
4. Пару невариабельных и уникальных широко распространенных сайтов рестрикции, фланкирующих случайные нуклеотидные последовательности, которые могут служить в качестве акцепторного модуля домена модификации хроматина (RNAS-CMD-I) (например, Kpn I и Avr II);
5. Постоянную группу невариабельных редких сайтов рестрикции, которые определяют 5'-часть модуля промотора. Примеры невариабельных редких сайтов рестрикции для применения в качестве шарниров на 5'-конце модуля промотора, предпочтительно включают AsiS I, Pac 1 и Sfo I, но также могут включать PvuI, AsiSI и SgrA I.
6. Случайные нуклеотидные последовательности, которые могут служить в качестве акцепторного модуля промотора/интрона (RNAS-P);
7. Постоянную группу невариабельных редких сайтов рестрикции, которые определяют область соединения в одно целое между 3'-частью модуля промотора/интрона и 5'-частью модуля экспрессии. Примеры невариабельных редких сайтов рестрикции для применения в качестве шарниров на 3'-конце модуля промотора предпочтительно включают Fse1, AscI и MluI, но также могут включать Pac1;
8. Случайные нуклеотидные последовательности, которые могут служить в качестве акцепторного модуля экспрессии (RNAS-E);
9. Постоянную группу невариабельных редких сайтов рестрикции, которые определяют область соединения 3'-части модуля экспрессии и 5'-части 3'-регуляторного модуля (например, SnaB I, Not I и Sal I);
10. Случайные нуклеотидные последовательности, которые могут служить в качестве акцепторного модуля 3'-регуляторного домена (RNAS-3);
11. Постоянную группу невариабельных редких сайтов рестрикции, которые определяют 3'-часть 3'-регуляторного модуля (например, Swa I, Rsr II и BsiW I);
12. Пару невариабельных и уникальных широко распространенных сайтов рестрикции, фланкирующих случайную нуклеотидную последовательность ДНК, которые могут служить в качестве акцепторного модуля домена модификации хроматина (RNAS-CMD-2) (например, Xho I и Nhe I);
13. Второй уникальный сайт HE (в данном месте идентичный сайту, указанному в пункте 3 выше (ISce1), а также в обратной ориентации). Однако недавно обнаружено, что если два уникальных сайта HE идентичны друг другу и находятся в обратной ориентации, то может происходить неблагоприятное событие рекомбинации. Поэтому предпочтительным может быть применение второго уникального сайта HE, который не является идентичным первому уникальному сайту HE;
14. Сайт праймера T3 в обратной ориентации; и
15. Четыре невариабельных и уникальных широко распространенных сайтов рестрикции, которые определяют 3'-сайт встраивания для мутантного вектора pUC19, описанного выше (например, BspE I, Pme I, Sap I и BspH I).
В альтернативном примере постоянная группа невариабельных редких сайтов рестрикции, которые определяют 5'-часть модуля промотора (генный шарнир GP1), включает в себя AsiS I, Pac I и Sbf I; редкие сайты рестрикции, которые определяют 3'-часть модуля промотора и 5'-часть модуля экспрессии (генный шарнир GP2), включают Fse I, Asc I и Mlu I; редкие сайты рестрикции, которые определяют 3'-часть модуля экспрессии и 5'-часть 3'-регуляторного модуля (генный шарнир GP3), включают SnaB I, Not I и Sal I; и редкие сайты рестрикции, которые определяют 3'-часть 3'-регуляторного модуля (генный шарнир GP4), включают Swa I, Rsr II и BsiW I.
В более конкретном примере генный шарнир GP1 состоит из последовательности AsiS I, Pac I и Sbf I, генный шарнир GP2 состоит из последовательности Fse I, Asc I и Mlu I, генный шарнир GP3 состоит из последовательности SnaB I, Not I и Sal I, и генный шарнир GP4 состоит из последовательности Swa I, Rsr II и BsiW I.
Мультигенный стыковочный вектор
На фиг.16 показан мультигенный стыковочный вектор, который содержит остов pUC19, идентичный остову стыковочного вектора PE3 за исключением введения дополнительных сайтов рестрикции для создания границ модулей, чтобы фланкировать модули в остове вектора. Такие модули остова включают модуль селективного гена репликации хозяина (RHS) и генный модуль начала репликации (ORI). RHS характеризуется Aat II шарнира векторного остова (VB), последовательностью ДНК, кодирующей селективный ген репликации хозяина, и двумя дополнительными шарнирами векторного остова Age I и Avr II. Модуль RHS на фиг.16 характеризуется шарниром векторного остова VB-1=Aat II, модулем RHS=ген резистентности к ампициллину и шарнирами остова вектора VB-2=Age I и VB-3=Avr II. ORI характеризуется шарнирами векторного остова Age I и Avr II, последовательностью ДНК, кодирующей начало репликации, и шарниром векторного остова Pme I. Модуль ORI на фиг.16 характеризуется шарнирами векторного остова VB-2=Age I и VB-3=Avr II, модулем ORI=начало репликации pUC10 и шарнирами векторного остова VB-4=Pme I.
Мультигенный стыковочный вектор характеризуется точным расположением случайных нуклеотидных последовательностей и неслучайных нуклеотидных последовательностей, определяющих широко распространенные, редкие сайты и сайты хоминг-эндонуклеаз в целях создания разнообразного ряда акцепторных областей для доменов. Такие модульные акцепторы делятся на следующие классы: GHS, BRD, SRS, CMD, (CMD/BRD), вкладыш PE3, PE3 MCS. Указанные модули представляют собой:
A. GHS=селективный ген экспрессии генома хозяина (например: NEO или PURO).
B. BRD=Неслучайная последовательность нуклеиновой кислоты, которую можно использовать в качестве сайта гомологичной рекомбинации в «компетентных по рекомбинации» линиях бактериальных клеток.
C. SRS=Неслучайная последовательность нуклеиновой кислоты, которую можно использовать в качестве сайта сайт-специфичной гомологичной рекомбинации, опосредованной продуктом гена сайт-специфичной рекомбиназы (например, Cre/Lox или Flp/Frt).
D. CMD=Неслучайная последовательность нуклеиновой кислоты, которая может изменять структуру хроматина генома хозяина (например, инсулятор бета-глобина цыплят HS4).
E. CMD/BRD=Неслучайная последовательность нуклеиновой кислоты, кодирующая домен модификации хроматина, которая может служить в качестве последовательности ДНК для гомологичной рекомбинации в «компетентных по рекомбинации» линиях бактериальных клеток.
F. Вкладыш PE3=случайная нуклеотидная последовательность, которая не кодирует сайт рестрикции или какую-либо другую биологическую функцию, имеющуюся в мультигенном стыковочном векторе.
G. PE3 MCS=неслучайная нуклеотидная последовательность, состоящая из множества сайтов рестрикции, уникальных для мультигенного стыковочного вектора.
Указанные модули фланкированы сайтами рестрикции, которые можно использовать для получения модульных стыковочных векторов. Такие связанные с модулями сайты рестрикции можно охарактеризовать согласно следующей классификации:
A. VB=сайты рестрикции, определяющие границы модуля векторного остова.
B. ES=сайты рестрикции, определяющие границы модуля селекции экспрессии генома хозяина.
C. TX=синтетические сайты BstX I, создающие выступающие концы, совместимые со специфичными редкими сайтами или сайтами хоминг-эндонуклеаз (например, I-Ceu I, I-Sce I, Sbf I).
D. CM=сайты рестрикции, определяющие границы модуля модификации хроматина.
E. SB=сайты рестрикции, определяющие границы модуля сайт-специфичной рекомбинации.
F. BL=сайты рестрикции, которые дают тупые концы.
Один вариант осуществления изобретения относится к нуклеотидной последовательности, расположенной между шарнирами векторного остова VB-1 и VB-5, где VB-1=Aat II, а VB-5=BspE I.
1. VB-1 представляет собой Aat II.
2. Нет последовательности или случайная нуклеотидная последовательность.
3. ES-I=SexA I.
4. Вкладыш случайной нуклеотидной последовательности (или модуль GHS).
5. ES-2=Srf I.
6. Вкладыш случайной нуклеотидной последовательности (или модуль BRD).
7. HE-3=I-Ppo I.
8. Вкладыш случайной нуклеотидной последовательности (или модуль SRS).
9. Нет последовательности или вкладыш случайной нуклеотидной последовательности.
10. CM-1=Kpn I.
11. Случайная нуклеотидная последовательность (или модуль CMD).
12. CM-2=Sac I.
13. Нет последовательности или вкладыш случайной нуклеотидной последовательности.
14. TX-1=Bst X I (I-Ceu I) в прямой ориентации.
15. Вкладыш случайной нуклеотидной последовательности (или модуль PE3).
16. TX-2=Bst X I (I-Sce I) в обратной ориентации.
17. Нет последовательности или вкладыш случайной нуклеотидной последовательности.
18. CM-3=Mfe I.
19. Вкладыш случайной нуклеотидной последовательности (или модуль CMD).
20. CM-4=Sac II.
21. Нет последовательности или вкладыш случайной нуклеотидной последовательности.
22. GP-1=(AsiS I/Pac I/Sbf I).
23. Вкладыш случайной нуклеотидной последовательности (или модуль PE3).
24. G-4/CM-5=(Swa I/Rsr II/BsiW I).
25. Вкладыш случайной нуклеотидной последовательности (или модуль CMD).
26. CM-6=Nar I, который имеет такой же мотив, как Kas I.
27. Нет последовательности или вкладыш случайной нуклеотидной последовательности.
28. SB-1=Nsi I, липкий выступающий конец совместим с Sbf I.
29. Нет последовательности или вкладыш случайной нуклеотидной последовательности.
30. BL-1=Sfo I.
31. Нет последовательности или вкладыш случайной нуклеотидной последовательности.
32. BL-2=Pvu II.
33. Нет последовательности или вкладыш случайной нуклеотидной последовательности.
34. BL-3=Nru I.
35. Нет последовательности или вкладыш случайной нуклеотидной последовательности.
36. SB-2=BsrG I, липкий выступающий конец совместим с BsiW I.
37. Нет последовательности или вкладыш случайной нуклеотидной последовательности.
38. CM-7=Spe I.
39. Вкладыш случайной нуклеотидной последовательности (или модуль CMD).
40. CM-8=Sph I.
41. Вкладыш случайной нуклеотидной последовательности (или модуль SRS).
42. HE-4=PI-Sce I (в прямой ориентации).
43. Вкладыш случайной нуклеотидной последовательности (или модуль BRD).
44. VB-5=BspE I.
На фиг.17, 18 и 19 показано, как мультигенный стыковочный вектор, представленный на фиг.16, может служить в качестве остова ДНК, с использованием обычных методик субклонирования, чтобы создать мультигенный вектор, имеющий два или более модулей PE3. В данном примере создания мультигенного вектора с тремя отдельными модулями PE3 используют предпочтительный порядок событий субклонирования на основании статистической вероятности того, что шарниры, необходимые для создания акцепторного домена PE3 в стыковочном векторе, не будут присутствовать в каком-либо модуле (модулях) PE3, уже включенных в остов мультигенного вектора.
Следующий порядок субклонирования модулей PE3 в мультигенном стыковочном векторе, показанном на фиг.16, является предпочтительным:
1. Модуль PE3 сначала необходимо субклонировать в домене вкладыша PE3-2, и данный модуль PE3 следует выбирать в соответствии с отсутствием по порядку следующих сайтов:
i. Nsi I или BsrG I
ii. Sfo I, Pvu II или Nru I
iii. BstX I
2. Второй модуль PE3 должен быть субклонирован в домене PE3-3 следующим, и данный модуль PE3 следует выбирать в соответствии с отсутствием по порядку следующих сайтов:
iv. Sbf I или BsiW I
v. Sbf I отдельно
vi. BsiW I
vii. BstX I
3. Третий Модуль PE3 должен быть субклонирован в домене PE3-1.
Модуль PE3, который удовлетворяет критериям, перечисленным в пункте 1, может быть субклонирован в мультигенном стыковочном векторе посредством независимой подготовки остова вектора и модуля PE3 расщеплением двумя ферментами рестрикции, при этом один фермент узнает один сайт рестрикции в домене GP-1, а второй фермент узнает один сайт рестрикции в домене GP-4. Выбранные ферменты не должны разрезать модуль PE3. Пример такого способа показан на фиг.17. Один способ субклонирования модуля PE3-A, показанного на фиг.17, в домене вкладыша PE3-2 требует получения линеаризованного векторного остова, получаемого расщеплением мультигенного стыковочного вектора ферментами AsiS I и Rsr II. Указанный молекулярно-биологический протокол дает точки стыковки ДНК с AsiS I-специфичными и Rsr II-специфичными липкими выступающими концами. Линейный фрагмент ДНК, содержащий требуемый модуль PE3-A, также может быть получен расщеплением ферментами AsiS I и Rsr II. Полученные в результате векторный остов и модуль PE3-A могут быть биохимически связаны вместе с использованием фермента ДНК-лигазы с образованием мультигенного вектора, содержащего модуль PE3-A.
На фиг.18 помещено изображение одного способа, посредством которого модуль PE3, который удовлетворяет критериям, указанным в пункте 2, может быть субклонирован в мультигенном стыковочном векторе, содержащем модуль PE3-A, в сайте множественного клонирования PE3-3 (PE3-3 MCS). В данном примере модуль PE3-A не содержит ни сайта Nsi I, ни сайта BsrG I (обозначены на фиг.16 как SB-1 и SB-2, соответственно), и модуль PE3-B не содержит сайта Sbf I или BsiW I. Как изображено на фиг.18, линеаризованный модуль PE3-B получают расщеплением вектора модуля PE3-B ферментами Sbf I и BsiW I. Линеаризованный остов мультигенного вектора получают расщеплением ферментами Nsi I и BsrG I. Комплементарные липкие концы в модуле PE3-B и остове мультигенного вектора могут быть связаны вместе биохимически с использованием фермента ДНК-лигазы. Получаемая в результате молекула ДНК состоит из мультигенного вектора, в котором модули PE3-A и PE3-B теперь являются частью непрерывной молекулы ДНК.
На фиг.18 представлена только одна из многочисленных методик субклонирования модуля PE3 в домене вкладыша PE3-3 в мультигенном стыковочном векторе. Другие методики субклонирования включают получение линеаризованных модулей PE3 и линеаризованных мультигенных стыковочных векторов, имеющих липкие/тупые, тупые/липкие или тупые/тупые точки стыковки. Предполагается, что такие методики субклонирования показывают меньшую долю успешных попыток создания лигированных продуктов, чем методика на основе липких/липких точек стыковки. Указанные альтернативные методики могут быть выбраны, если структура модулей PE3-A или PE3-B не удовлетворяет одному или нескольким критериям, перечисленным в пункте 2.
На фиг.19 показано, как модуль PE3-C может быть субклонирован в мультигенном векторе, содержащем модули PE3-A и PE3-B. Такая методика требует, чтобы ни PE3-A, ни PE3-B не содержали сайта BstX I. В данной методике мультигенный стыковочный вектор линеаризуют расщеплением BstX I. Модуль PE3-C линеаризуют расщеплением хоминг-эндонуклеазами I-Ceu I и I-Sce I. Получаемые в результате линейные модуль PE3-C и мультигенный стыковочный вектор связывают вместе биохимически, используя фермент ДНК-лигазу. Получаемая молекула ДНК состоит из мультигенного вектора, в котором модули PE3-A, PE3-B и PE3-C теперь являются частью непрерывной молекулы ДНК.
На фиг.21 показано, как модуль PE3, фланкированный либо доменами модификации хроматина (CMD), либо доменами бактериальной рекомбинации (BRD), может быть введен в мультигенный стыковочный вектор с использованием методики бактериальной рекомбинации. В данном примере модуль PE3 заменяет домен вкладыша PE3-1 посредством действия гомологичной рекомбинации между доменами CMD/BRD, фланкирующими как модуль PE3 в векторе модуля PE3, так и домен вкладыша PE3-1 в мультигенном стыковочном векторе. Указанная методика требует использования вспомогательных ДНК-векторов или специальных штаммов бактерий, в которых экспрессия ферментов рекомбинации может быть индуцирована изменениями окружающей среды (химическими, температурными или метаболическими).
Другой вариант осуществления изобретения, показанного на фиг.16, можно охарактеризовать нуклеотидной последовательностью, присутствующей между шарнирами векторного остова VB-1 и VB-5, где VB-1=Aat II, а VB-5=BspE I.
1. VB-1 представляет собой Aat II.
2. Нет последовательности или случайная нуклеотидная последовательность.
3. ES-1=SexA I.
4. Вкладыш случайной нуклеотидной последовательности (или модуль GHS).
5. ES-2=Srf I.
6. Вкладыш случайной нуклеотидной последовательности (или модуль BRD).
7. HE-3=I-Ppo I.
8. Вкладыш случайной нуклеотидной последовательности (или модуль SRS).
9. Нет последовательности или вкладыш случайной нуклеотидной последовательности.
10. CM-1=Kpn I.
11. Случайная нуклеотидная последовательность (или модуль CMD).
12. CM-2=Sac I.
13. Нет последовательности или вкладыш случайной нуклеотидной последовательности.
14. HE-1=I-Ceu I в прямой ориентации.
15. Вкладыш случайной нуклеотидной последовательности (или модуль PE3).
16. HE-2=I-Sce I в обратной ориентации.
17. Нет последовательности или вкладыш случайной нуклеотидной последовательности.
18. CM-3=Mfe I.
19. Вкладыш случайной нуклеотидной последовательности (или модуль CMD).
20. CM-4=Sac II.
21. Нет последовательности или вкладыш случайной нуклеотидной последовательности.
22. GP-1=(AsiS I/Pac I/Sbf I).
23. Вкладыш случайной нуклеотидной последовательности (или модуль PE3).
24. G-4/CM-5=(Swa I/Rsr II/BsiW I).
25. Вкладыш случайной нуклеотидной последовательности (или модуль CMD).
26. CM-6=Nar I, который имеет такой же мотив, как Kas I.
27. Нет последовательности или вкладыш случайной нуклеотидной последовательности.
28. TX-1=Bst X I (I-Ceu I) в прямой ориентации.
29. SB-1=Nsi I, липкий выступающий конец совместим с Sbf I.
30. Нет последовательности или вкладыш случайной нуклеотидной последовательности.
31. BL-1=Sfo I.
32. Нет последовательности или вкладыш случайной нуклеотидной последовательности.
33. BL-2=Pvu II.
34. Нет последовательности или вкладыш случайной нуклеотидной последовательности.
35. BL-3=Nru I.
36. Нет последовательности или вкладыш случайной нуклеотидной последовательности.
37. SB-2=BsrG I, липкий выступающий конец совместим с BsiW I.
38. Нет последовательности или вкладыш случайной нуклеотидной последовательности.
39. TX-2=Bst X I (I-Sce I) в обратной ориентации.
40. Нет последовательности или вкладыш случайной нуклеотидной последовательности.
41. CM-7=Spe I.
42. Вкладыш случайной нуклеотидной последовательности (или модуль CMD).
43. CM-8=Sph I.
44. Вкладыш случайной нуклеотидной последовательности (или модуль SRS).
45. HE-4=PI-Sce I (в прямой ориентации).
46. Вкладыш случайной нуклеотидной последовательности (или модуль BRD).
47. VB-5=BspE I.
На фиг.20 показано, как могут быть использованы хоминг-эндонуклеазы для введения модуля PE3 в мультигенный стыковочный вектор, описанный выше. В данном примере модуль PE3 и мультигенный стыковочный вектор линеаризуют расщеплением хоминг-эндонуклеазами I-Ceu I и I-Sce I. Полученный в результате линейный модуль PE3 может быть связан с линейным мультигенным стыковочным вектором биохимически с использованием фермента ДНК-лигазы. Указанная методика субклонирования может быть использована, если модуль PE3 в мультигенном стыковочном векторе или вводимый в стыковочный вектор не удовлетворяет одному или нескольким критериям, перечисленным в пунктах 1-3.
Мультигенный стыковочный вектор (основная стыковочная плазмида), показанный на фиг.8, можно применять для сборки двух полных трансгенов PE3, которые сначала конструируют в плазмидах стыковочных узлов PE3, или двух гомологичных плеч, необходимых для конструирования направленного к определенному гену трансгена, или для введения двух типов элементов для позитивной или негативной селекции.
Неограничивающий пример сайта множественного клонирования (MCS) в мультигенной (или основной) стыковочной плазмиде показан на фиг.9 и содержит следующие последовательные элементы в порядке перечисления:
1. Два невариабельных и уникальных широко распространенных сайта рестрикции, которые определяют 5'-сайт встраивания для мутантного вектора pUC19, описанного выше (например, Aat II и Blp I).
2. Сайт обратного праймера M13.
3. Пару уникальных сайтов HE в противоположной ориентации, фланкирующих случайную нуклеотидную последовательность ДНК, которая может служить в качестве акцепторного модуля гена селекции экспрессии генома хозяина (RNAS-GEH-S1) (например, PI-SceI (в прямой ориентации) и PI-SceI (в обратной ориентации)).
4. Невариабельный и уникальный широко распространенный сайт рестрикции, который обеспечивает клонирование модуля челночного вектора даунстрим от пары HE (например, Eco0109I).
5. Постоянную группу невариабельных редких сайтов рестрикции, которые определяют 5'-часть модуля левого плеча рекомбинации (например, SgrA I и AsiS I).
6. Случайные нуклеотидные последовательности, которые могут служить в качестве акцепторного модуля левого плеча рекомбинации (RNAS-LRA).
7. Постоянную группу невариабельных редких сайтов рестрикции, которые определяют 3'-часть акцепторного модуля левого плеча рекомбинации (например, PacI, MluI и AscI).
8. Уникальный сайт HE (например, I-Ceu I (в прямой ориентации)).
9. Пару невариабельных и уникальных широко распространенных сайтов рестрикции, фланкирующих случайную нуклеотидную последовательность ДНК, которая может служить в качестве акцепторного модуля домена модификации хроматина (RNAS-CMD-1) (например, Kpn I и Avr II).
10. Сайт праймера T7.
11. Пару уникальных сайтов BstX I в противоположной ориентации (при этом вариабельная область нуклеотидов в сайте рестрикции BstX I характеризуется нуклеотидами, идентичными некомплементарным хвостам, образованным благодаря расположению двух идентичных сайтов рестрикции HE, находящихся в обратно-комплементарной ориентации; например, PI-SceI (в прямой ориентации) и PI-SceI (в обратной ориентации)), фланкирующие случайную нуклеотидную последовательность ДНК, которая может служить в качестве акцепторного модуля сложного трансгена (RNAS-PE3-1).
12. Пару уникальных сайтов HE в противоположной ориентации, фланкирующих случайную нуклеотидную последовательность ДНК, которая может служить в качестве акцепторного модуля сложного трансгена (RNAS-PE3-2) (например, I-SceI (в прямой ориентации) и I-SceI (в обратной ориентации)).
13. Сайт праймера T3 в обратно ориентации.
14. Пару невариабельных и уникальных широко распространенных сайтов рестрикции, фланкирующих случайную нуклеотидную последовательность ДНК, которая может служить в качестве акцепторного модуля домена модификации хроматина (RNAS-CMD-2) (например, Xho I и Nhe I).
15. Уникальный сайт HE (в данном случае идентичный сайту, указанному в пункте 8 выше (ISce1), а также в обратной ориентации).
16. Постоянную группу невариабельных редких сайтов рестрикции, которые определяют 5'-часть модуля правого плеча рекомбинации (например, SnaB I, Sal I и Not I).
17. Случайные нуклеотидные последовательности, которые могут служить в качестве акцепторного модуля правого плеча рекомбинации (RNAS-RRA).
18. Постоянную группу невариабельных редких сайтов рестрикции, которые определяют 3'-часть акцепторного модуля правого плеча рекомбинации (например, Rsr II, Swa I и BsiW I).
19. Невариабельный и уникальный широко распространенный сайт рестрикции, который обеспечивает клонирование модуля челночного вектора апстрим от пары HE (например, BspE I).
20. Пару уникальных сайтов HE в противоположной ориентации, фланкирующих случайную нуклеотидную последовательность ДНК, которая может служить в качестве акцепторного модуля гена селекции экспрессии генома хозяина (RNAS-GEH-S2) (например, PI-Psp I (в прямой ориентации) и PI-Psp I (в обратной ориентации)).
21. Сайт прямого праймера M13, помещенный в обратной ориентации.
22. Три невариабельных и уникальных широко распространенных сайтов рестрикции, которые определяют 3'-сайт встраивания для мутантного вектора pUC19, описанного выше (например, Pme I, Sap I и BspH I).
В качестве примера способа практического осуществления настоящего изобретения может быть сконструирован трансген, содержащий указанные элементы:
1. Нуклеотидные последовательности промотора поверхностно-активного белка C человека (SP-C).
2. Последовательности, кодирующие белковый продукт гена рецептора бета-c колониестимулирующего фактора гранулоцитов макрофагов мышей (GMRβc).
3. Последовательности интрона бета-глобина кролика, и
4. Сигнал поли-A SV40.
Последовательность SP-C содержит внутренние сайты BamH1 и могут быть высвобождены из исходной плазмиды только с использованием Not1 и EcoR1. GMRβc имеет внутренний сайт Not1 и может быть вырезана из исходной плазмиды с использованием BamH1 и Xho1. Интронные последовательности бета-глобина кролика могут быть вырезаны из исходной плазмиды с использованием EcoR1. Поли-A-хвост SV-40 может быть вырезан из исходной плазмиды с использованием Xho1 и Sac1. Вследствие избыточности нескольких сайтов рестрикции ни одна из исходных плазмида не может быть использована для сборки всех необходимых фрагментов.
Следующие стадии использовали для создания требуемого трансгена в стыковочной плазмиде PE3 согласно изобретению:
1. Так как Not1 и PspOM1 создают совместимые липкие концы, последовательности промотора SP-C человека вырезают ферментами Not1 и EcoR1 и клонируют в сайтах PspOM1 и EcoR1 челночного вектора P. Продукт такой реакции назван pSVP-SPC.
2. После стадий размножения и извлечения, хорошо известных специалистам в данной области, интронные последовательности бета-глобина кролика клонировали в сайте EcoR1 pSVP-SPC. Ориентацию интрона в полученной в результате промежуточной конструкции подтверждали секвенированием продукта, называемого pSVP-SPC-rβG.
3. Промотор и интрон вырезали и выделяли в виде одного непрерывного фрагмента из pSVP-SPC-rβG, используя AsiS1 и Asc1. Одновременно стыковочную плазмиду PE3 разрезали AsiS1 и Asc1 при подготовке для лигирования с участком промотор/интрон. Фрагмент промотор/интрон лигировали в стыковочную плазмиду, размножали и выделяли.
4. Сайт Xho 1 фрагмента GMRβc заполняют для создания тупого 3'-конца, используя способы, хорошо известные специалистам в данной области. Затем его клонируют в сайте BamH1 и сайте Pvu2 с тупыми концами плазмиды pSVP-SPC-rβG. Полученную плазмиду (pDP- SPC-GMRβc-rβG) размножают и извлекают.
5. Конечная стадия клонирования представляет собой добавление поли-A-хвоста SV-40. Фрагмент SV40-поли-A вырезают, используя Xho1 и Sac1, так же как реципиентный вектор pDS1-SPC- GMRβc-rβG. Оба фрагмента ДНК очищают в геле и выделяют. Готовят смесь для лигирования с молярным соотношением 10:1 SV-40-поли-A к pDS1-SPC-GMRβc-rβG. Продукты лигирования размножают и собирают.
Новая плазмида, pDS1-SPC-GMRβc-rβG-pA содержит все элементы, необходимые для трансгена, включая уникальный сайт рестрикции на 3'-конце, с использованием которого полную плазмиду pDS1-SPC- GMRβc-rβG-pA можно линеаризовать для трансфекции в эукариотические клетки или микроинъекции в пронуклеус оплодотворенной яйцеклетки.
Что касается сайтов HE, то обычно по меньшей мере два сайта рестрикции HE, каждый из которых может быть расщеплен по меньшей мере одним ферментом рестрикции HE, помещают, фланкируя области модулей, в целях создания акцепторного сайта генной кассеты, который не может самоотжигаться. Кроме того, при необходимости можно, но необязательно, помещать такие сайты HE в противоположной ориентации по отношению друг к другу. То есть, поскольку сайты HE асимметричны и не являются палиндромными, то можно создать некомплементарные выступающие липкие 3'-хвосты, помещая два сайта рестрикции HE в противоположной ориентации. Например, HE I-SceI разрезает узнаваемый ею сайт рестрикции, как показано «/»:
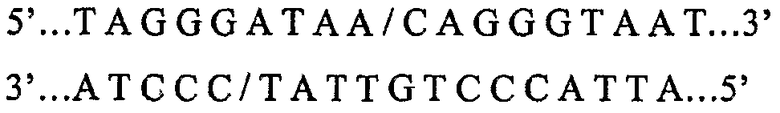
Обратное расположение второго сайта в MCS может создать два некомплементарных липких выступающих конца:

Указанное особенно полезно, когда в вектор требуется ввести большие трансгены. Из-за большого размера вставки термодинамически более выгоден для вектора самоотжиг, а не акцепция крупной вставки. Присутствие некомплементарных хвостов, создаваемых размещением сайтов рестрикции HE, обеспечивает благоприятные химические условия для противодействия термодинамически обусловленной склонности к самолигированию.
Кроме того, асимметричная природа большинства выступающих HE-хвостов также создает эффективное средство клонирования при использовании в комбинации с сайтом фермента рестрикции BstX I (5' CCANNNNN/NTGG 3'). Нейтральный в отношении последовательности домен BstX I («N» может быть любым нуклеотидом) можно использовать для создания совместимых липких концов для двух обратно ориентированных выступающих HE-хвостов, при этом предотвращая самоотжиг.
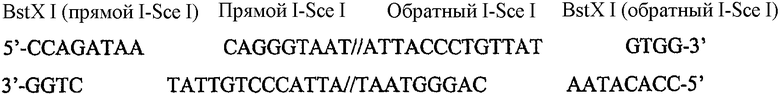
Также можно использовать другие эндонуклеазы, не включенные в приведенное перечисление, сохраняя такую же функциональность и сущность и цель изобретения.
Легко можно понять, что к числу многочисленных преимуществ настоящего изобретения относится то, что можно быстро собирать трансгены, обычно содержащие модули промотора, экспрессии и 3'-регуляторной области, в очень короткий период времени, а также быстро и легко варьировать или реконструировать заново собранный трансген. Обычные попытки осуществлять изменения собранного трансгена с использованием известных способов могут, как правило, занимать год или более времени работы лаборатории. Применяя способы согласно настоящему изобретению, можно получить требуемые трансгены в течение нескольких дней или недель и затем провести требуемое тестирование каждой, тем самым экономя значительное время и средства исследователя. Могут быть взяты челноки, которые исходно были созданы способами синтеза de novo, рекомбинации и способами клонирования на основе ПЦР с использованием выступающих концов с терминаторами, и использованы с применением способа, основанного на точках стыковки, согласно настоящему изобретению, чтобы быстро собрать такие предварительно полученные элементы в большое количество трансгенов. Трансгены PE3, полученные с применением изобретения, можно использовать в одном организме или множестве организмов, включая бактерии, дрожжи, мышей и других эукариот с небольшой модификацией или без дополнительной модификации.
Хотя настоящее изобретение проиллюстрировано описанием вариантов его осуществления, и хотя варианты были описаны достаточно подробно, не подразумевается, что объем прилагаемой формулы изобретения сужен или иным образом ограничен такими подробностями. Дополнительные преимущества и модификации легко смогут понять специалисты в данной области. Поэтому изобретение в его более широких аспектах не ограничено конкретными подробностями, типичными способами и структурами и показанными и описанными иллюстративными примерами. Следовательно, можно осуществлять отступления от таких подробностей, не выходя за рамки объема заявленного изобретения.




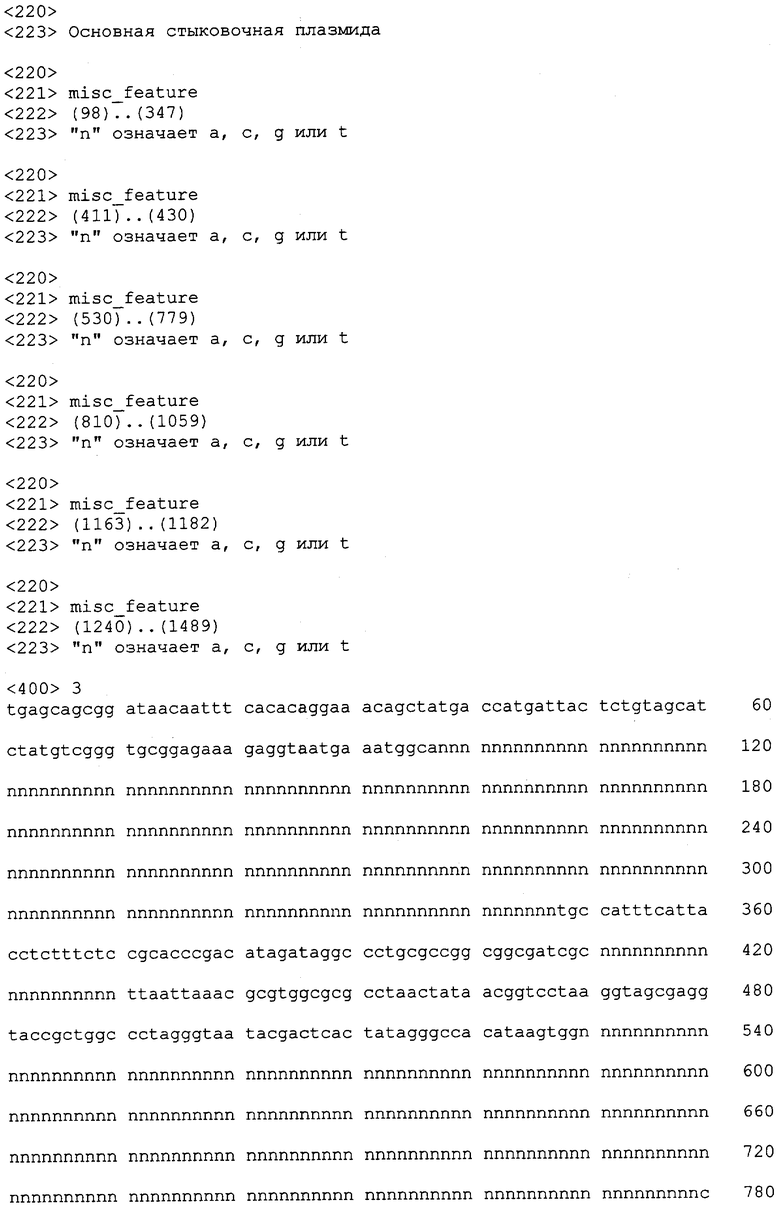








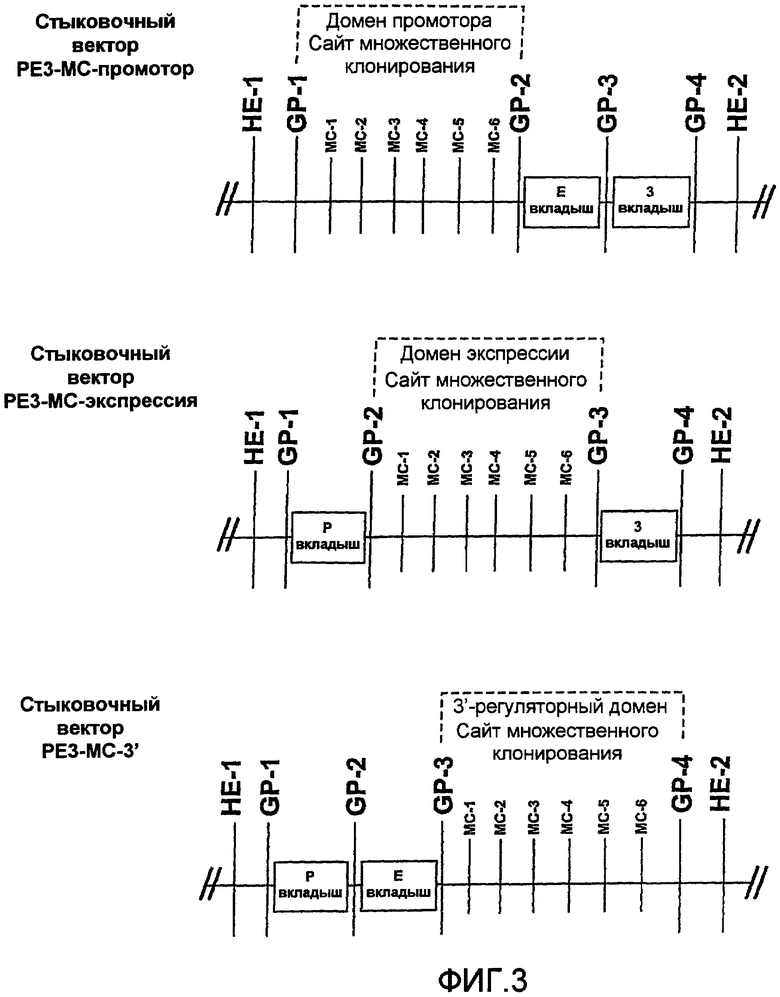
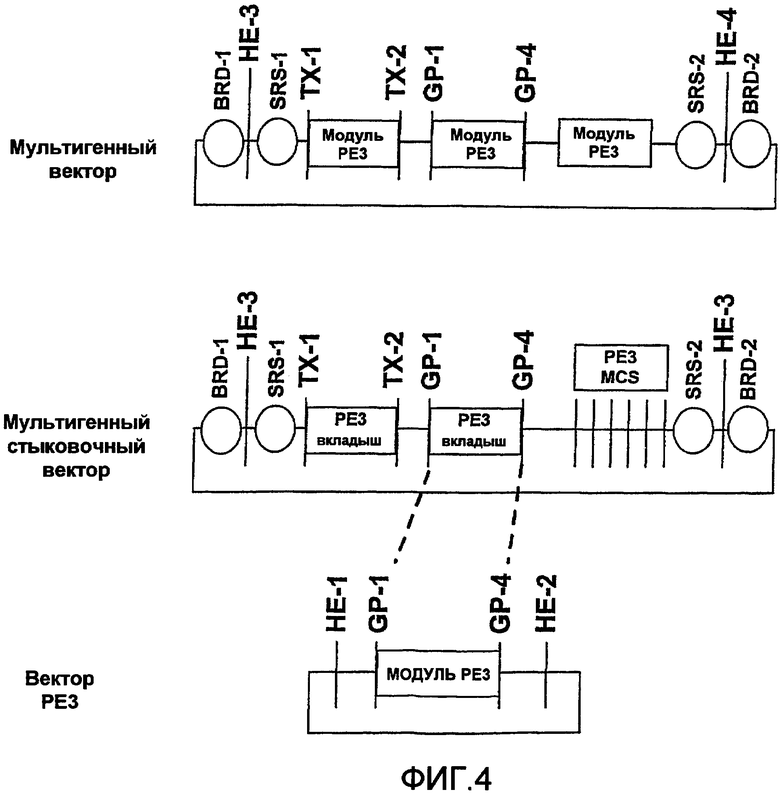
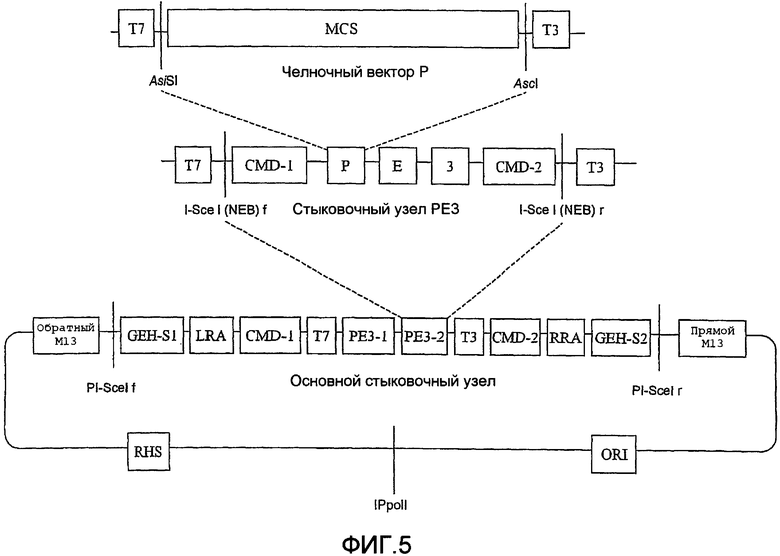
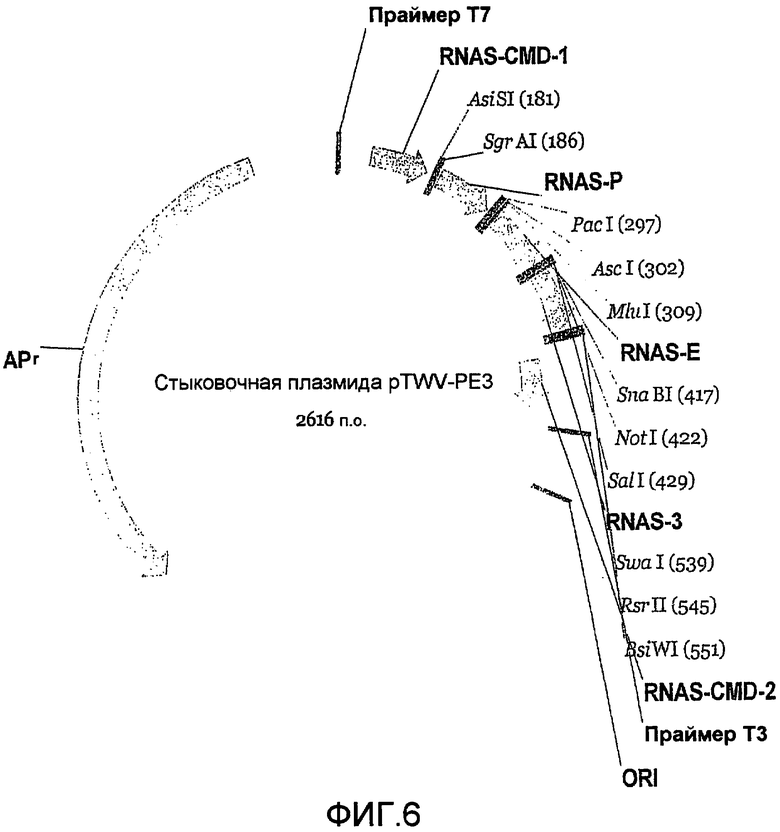
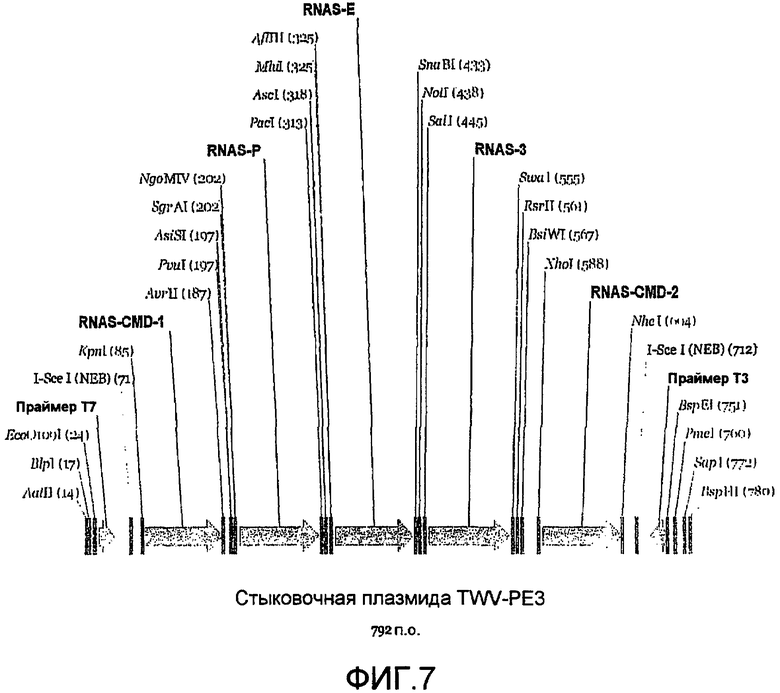
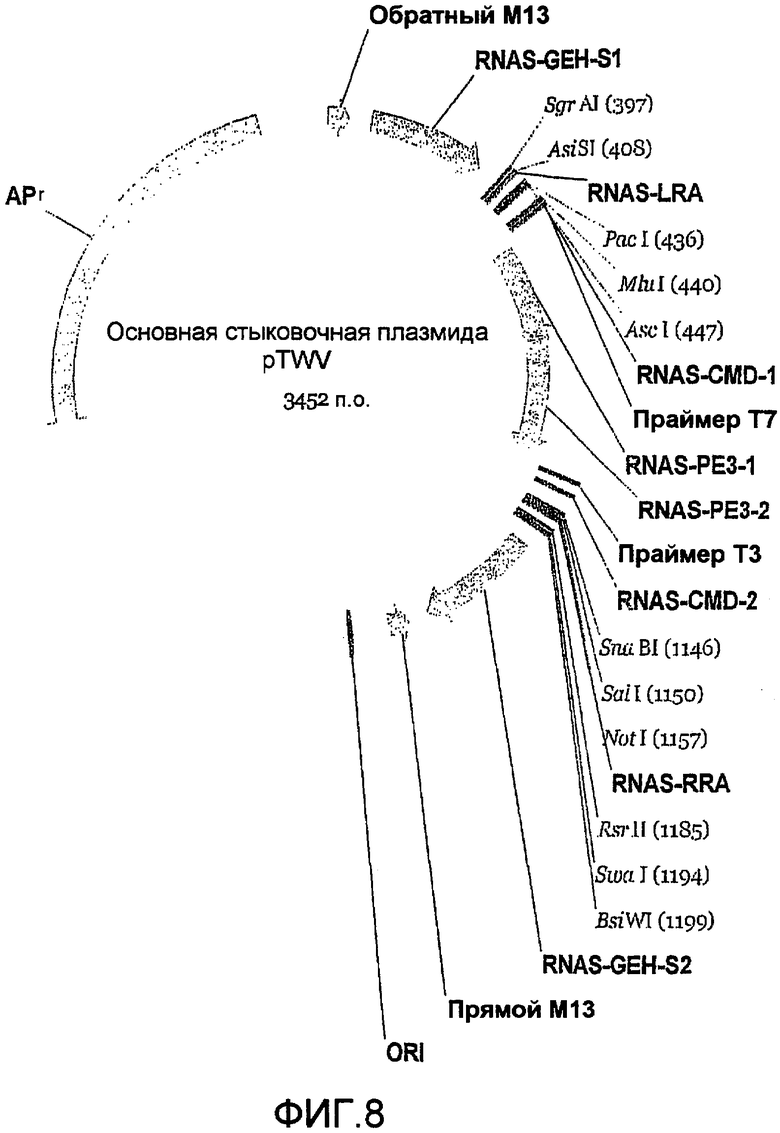
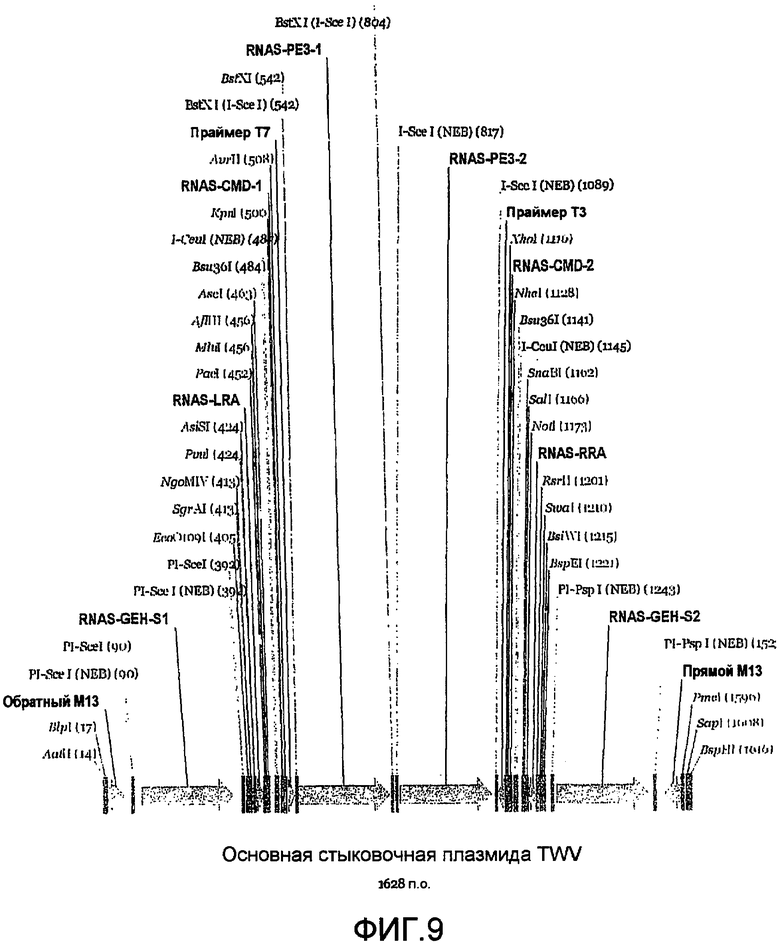
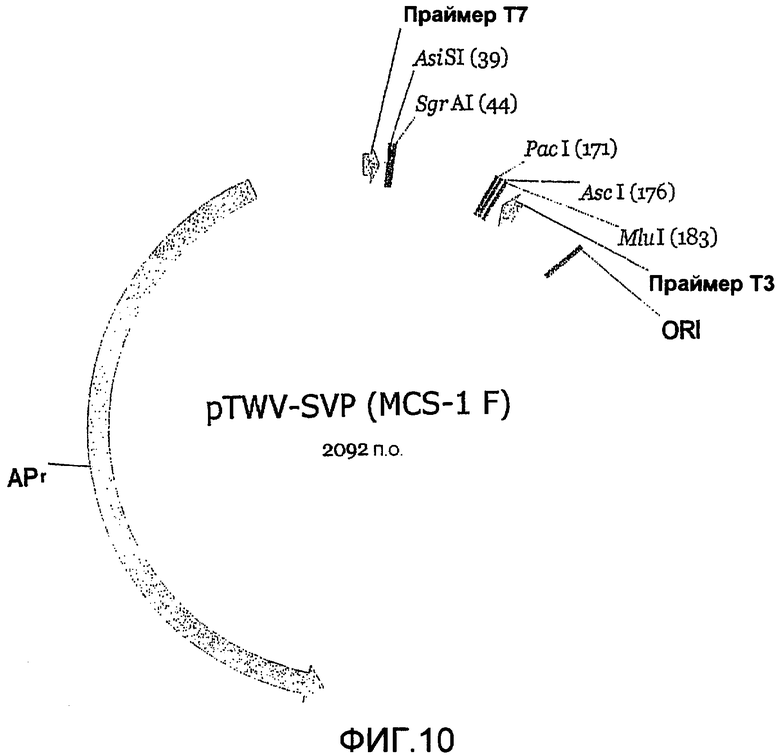
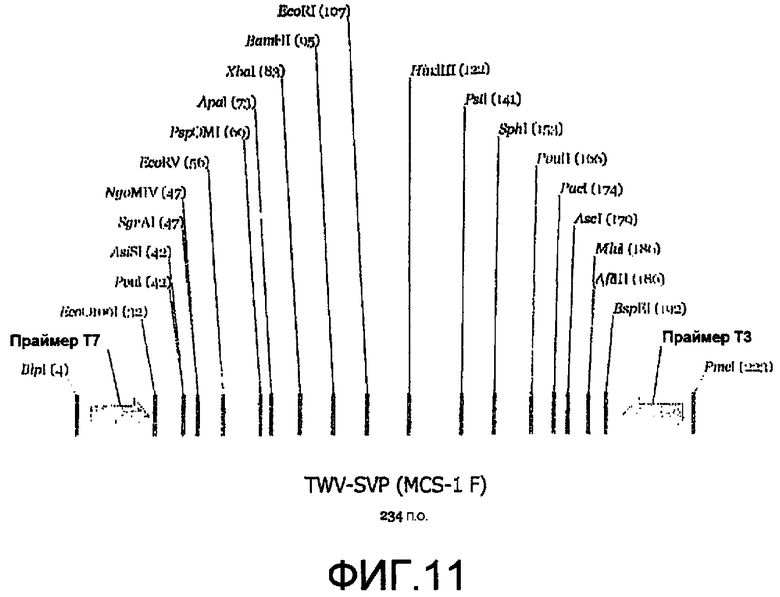
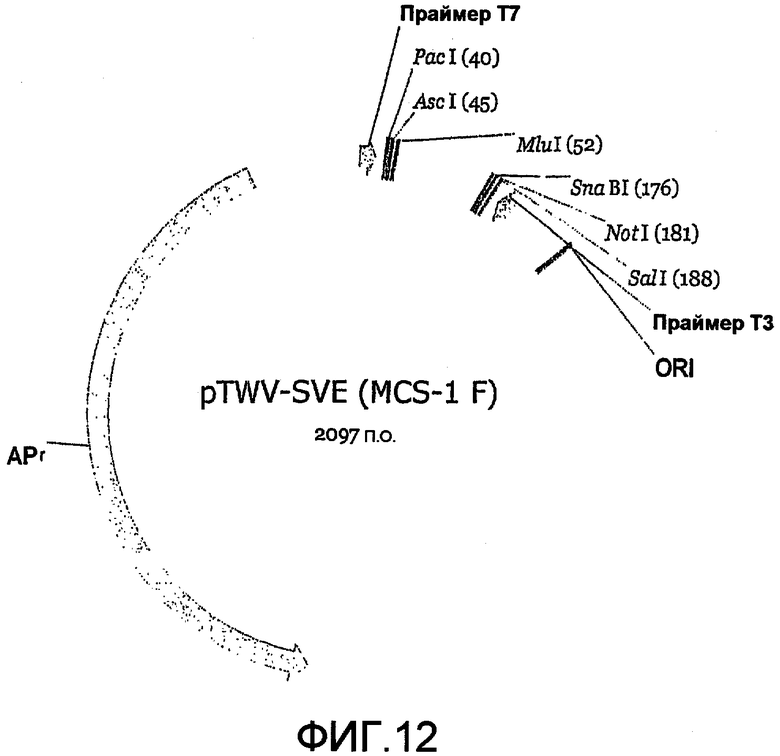
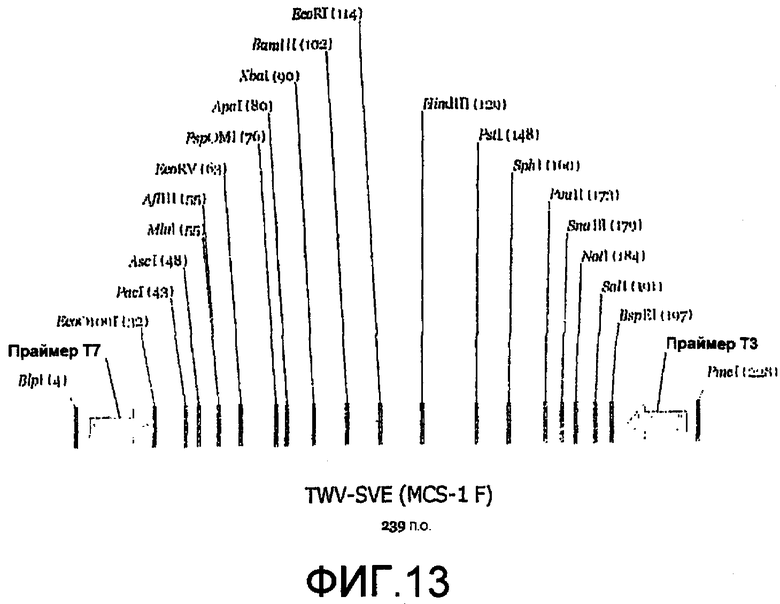
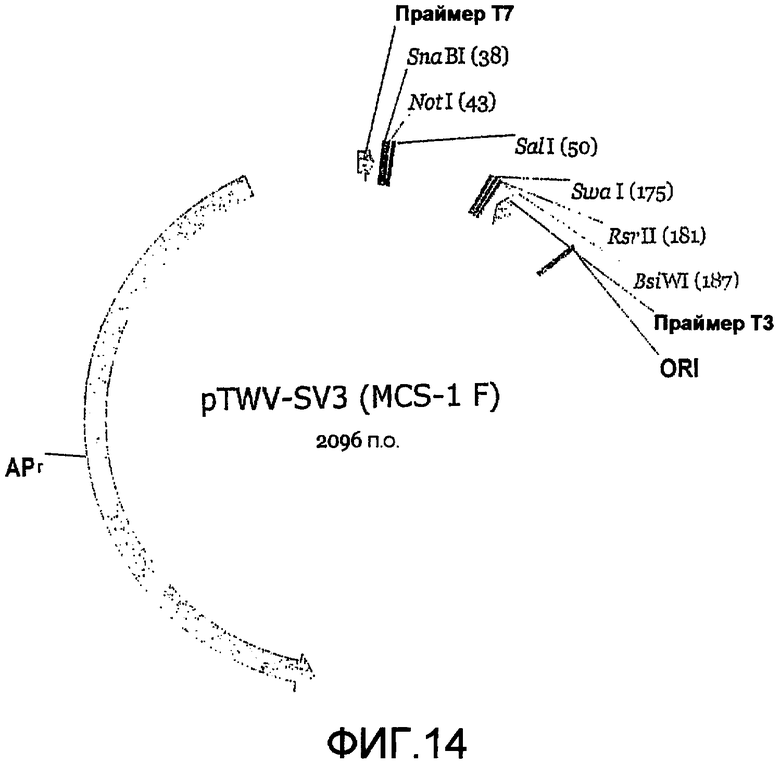
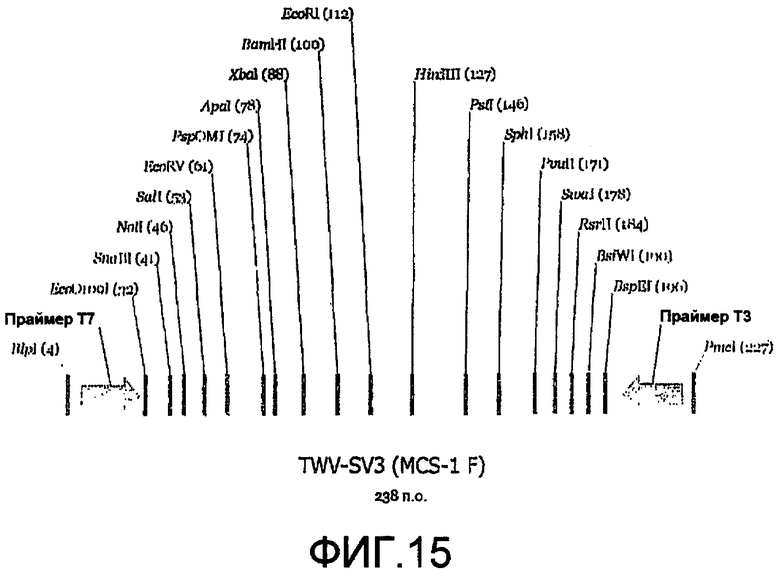
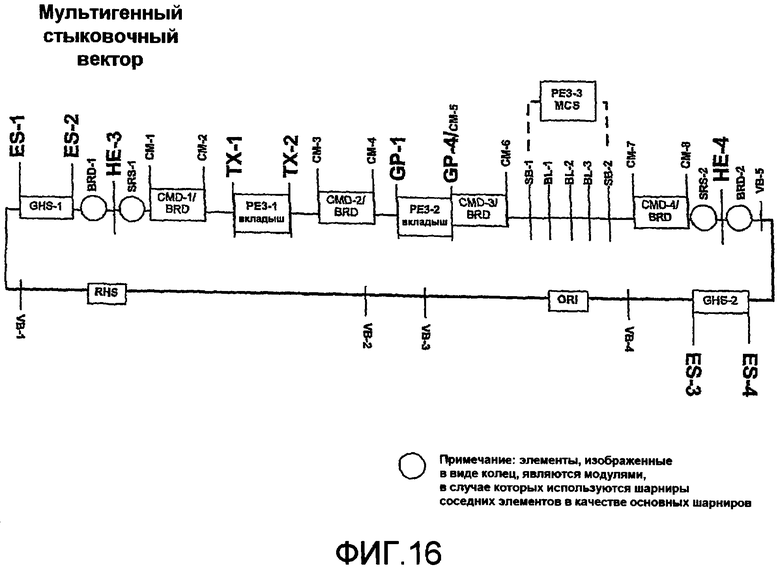
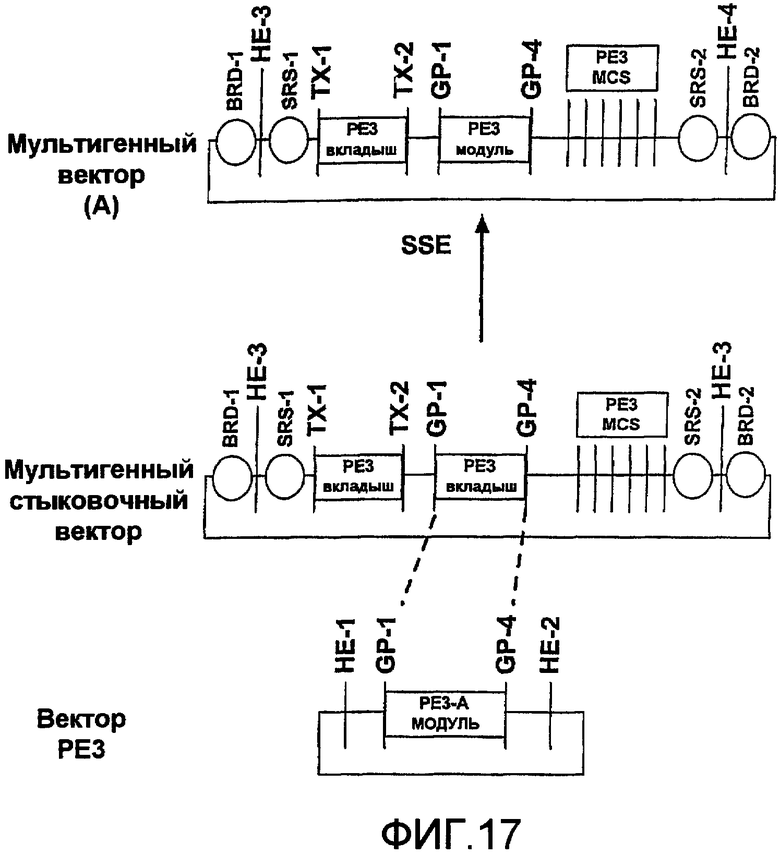
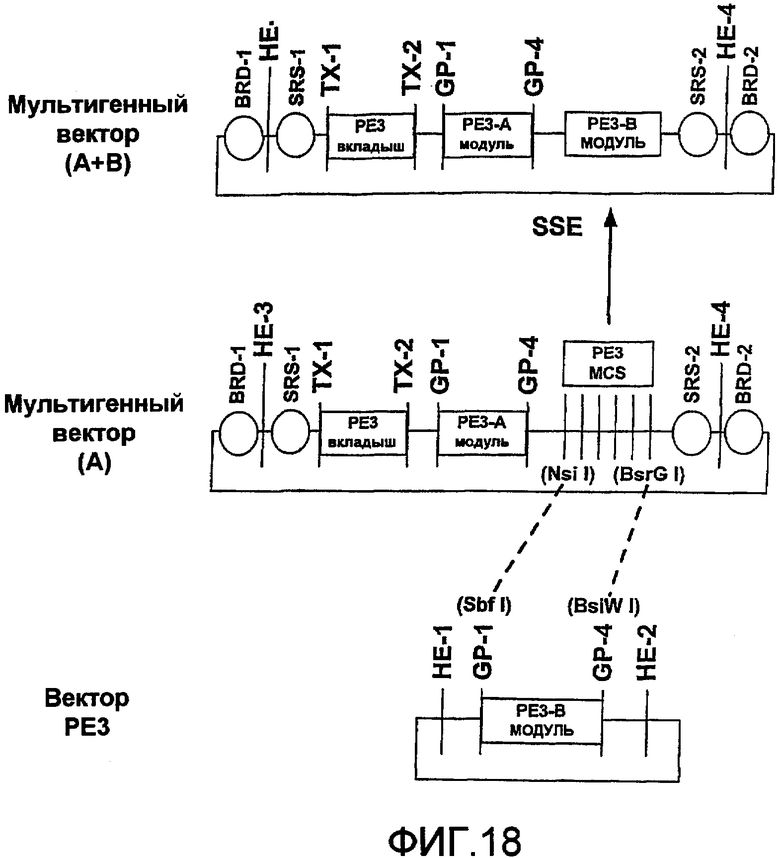
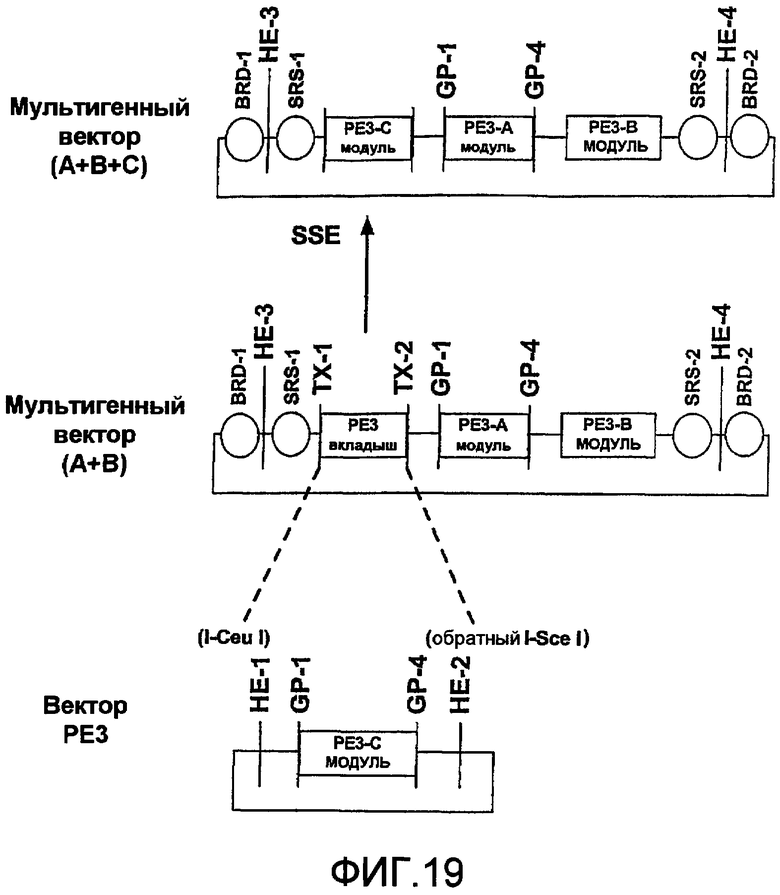
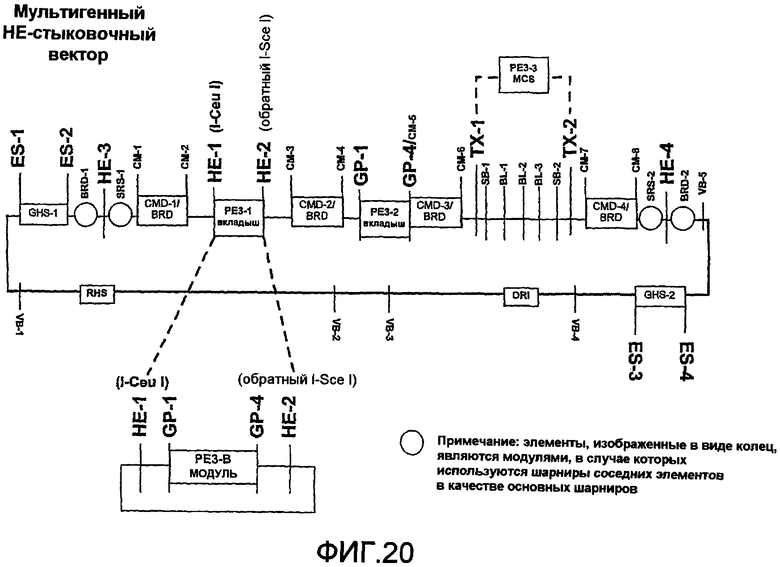
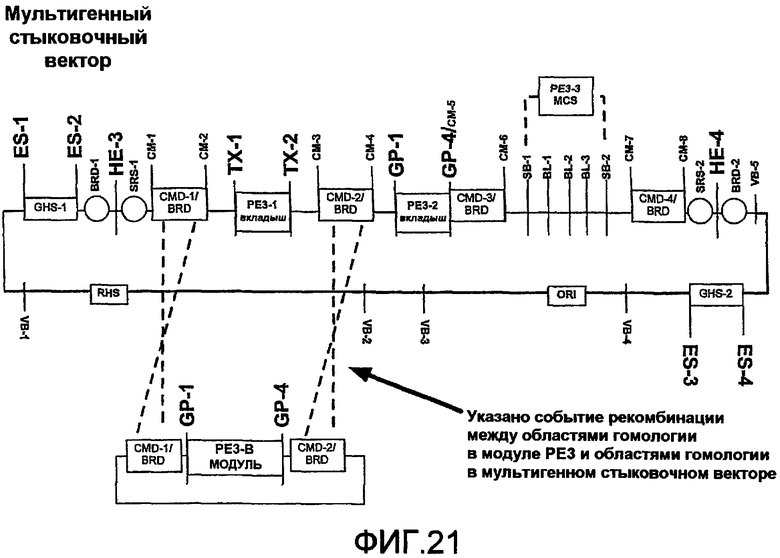
Изобретение относится к области генной инженерии и может быть использовано в биотехнологических процессах в качестве средства для объединения фрагментов ДНК и получения сложных ДНК-конструкций. Предложены векторные плазмиды («стыковочные векторы») для синтеза трансгенов (РЕ3), представляющие собой клонирующий вектор, в который включен модуль клонирования, состоящий из четырех генных шарниров (ГШ 1-4) и расположенных между ними трех нуклеотидных последовательностей (НП 1-3), где каждый из ГШ содержит от 2 до 4 невариабельных редких сайтов рестрикции из более чем 6 нуклеотидов, а нуклеотидные последовательности между ГШ включают «вкладыши», которые при клонировании заменяются в НП 1 - модулем промотора; в НП 2 - модулем экспрессии, а в НП 3 - 3'-регуляторным модулем. При этом либо ГШ 1, либо ГШ 2 независимо содержат 3-4 невариабельных редких сайта рестрикции более чем из 6 нуклеотидов. Предлагается также вариация стыковочного вектора РЕ3 по изобретению, предназначенная для множественного клонирования (МС) и отличающаяся тем, что, по меньшей мере, одна из трех НП является модулем множественного клонирования с сайтом множественного клонирования, включающим распространенные сайты рестрикции, которые являются уникальными в стыковочном векторе РЕ3. 2 н. и 4 з.п. ф-лы, 21 ил.
1. Стыковочный вектор РЕ3, содержащий модуль клонирования РЕ3, где модуль клонирования РЕ3 содержит:
a) первый генный шарнир, содержащий два, три или четыре невариабельных сайта рестрикции из более чем шести нуклеотидов, которые при клонировании можно подвергать обработке для определения 5'-части модуля промотора;
b) первую последовательность нуклеиновой кислоты, содержащую вкладыш, который при клонировании заменяют модулем промотора;
c) второй генный шарнир, содержащий два, три или четыре невариабельных сайта рестрикции из более чем шести нуклеотидов, которые при клонировании можно подвергать обработке для определения области соединения в одно целое между 3'-частью модуля промотора и 5'-частью модуля экспрессии;
d) вторую последовательность нуклеиновой кислоты, содержащую вкладыш, который при клонировании заменяют модулем экспрессии;
e) третий генный шарнир, содержащий два, три или четыре невариабельных сайта рестрикции из более чем шести нуклеотидов, которые при клонировании можно подвергать обработке для определения области соединения в одно целое между 3'-частью модуля экспрессии и 5'-частью 3'-регуляторного модуля;
f) третью последовательность нуклеиновой кислоты, содержащую вкладыш, который при клонировании заменяют 3'-регуляторным модулем; и
g) четвертый генный шарнир, содержащий два, три или четыре невариабельных сайта рестрикции, которые при клонировании могут быть подвергнуты обработке для определения 3'-части 3'-регуляторного модуля,
где либо первый генный шарнир, либо второй генный шарнир независимо содержат по меньшей мере три и не более чем четыре невариабельных сайта рестрикции из более чем шести нуклеотидов.
2. Стыковочный вектор PE3 по п.1, в котором невариабельные редкие сайты рестрикции из более чем шести нуклеотидов выбраны из группы, состоящей из AsiS I, Рас I, Sbf I, Fse I, Asc I, Not I, Swa I, Rsr II, Sgr AI, Flp I, Pme I, Sda I, Sgf I, Srf I и Sse8781 I.
3. Стыковочный вектор PE3 для множественного клонирования (МС), состоящий из клонирующего ДНК-вектора, содержащего модуль клонирования РЕЗ, скомпонованный для клонирования модулей промотора, экспрессии и 3'-регуляторной области в модуле клонирования PE3, где модуль клонирования PE3 содержит:
a) первый генный шарнир, содержащий два, три или четыре невариабельных сайта рестрикции из более чем шести нуклеотидов, которые при клонировании могут быть подвергнуты обработке для определения 5'-части модуля промотора;
b) первую последовательность нуклеиновой кислоты;
c) второй генный шарнир, содержащий два, три или четыре невариабельных сайта рестрикции из более чем шести нуклеотидов, которые при клонировании могут быть подвергнуты обработке для определения области соединения в одно целое между 3'-частью модуля промотора и 5'-частью модуля экспрессии;
d) вторую последовательность нуклеиновой кислоты;
e) третий генный шарнир, содержащий два, три или четыре невариабельных сайта рестрикции из более чем шести нуклеотидов, которые при клонировании могут быть подвергнуты обработке для определения области соединения в одно целое между 3'-частью модуля экспрессии и 5'-частью 3'-регуляторного модуля;
f) третью последовательность нуклеиновой кислоты; и
g) четвертый генный шарнир, содержащий два, три или четыре невариабельных сайта рестрикции из более чем шексти нуклеотидов, которые при клонировании могут быть подвергнуты обработке для определения 3'-части 3'-регуляторного модуля;
где либо первый генный шарнир, либо второй генный шарнир независимо содержат по меньшей мере три и не более чем четыре невариабельных сайта рестрикции из более чем шести нуклеотидов и в котором по меньшей мере одна из первой, второй и третьей последовательностей нуклеиновых кислот является модулем множественного клонирования, содержащим сайт множественного клонирования (MCS), включающий множество сайтов рестрикции, выбранных из широко распространенных сайтов рестрикции, которые являются уникальными в стыковочном векторе РЕ3, обеспечивая сайты клонирования для клонирования представляющего интерес генетического материала в модуле множественного клонирования, а остальные последовательности нуклеиновых кислот представляют собой вкладыши.
4. Стыковочный вектор PE3 (МС) по п.3, в котором вторая последовательность нуклеиновой кислоты является модулем множественного клонирования, сайты рестрикции во втором генном шарнире содержат Fse I и Asc I, а сайты рестрикции в третьем генном шарнире содержат Not I.
5. Стыковочный вектор РЕ3 (МС) по п.3, в котором третья последовательность нуклеиновой кислоты является модулем множественного клонирования; сайты рестрикции в третьем генном шарнире содержат NotI, а сайты рестрикции в четвертом генном шарнире содержат Swa I и Rsr II.
6. Стыковочный вектор PE3 (МС) по п.3, в котором сайты рестрикции в первом генном шарнире состоят из AsiS I, Pac I и Sbf I в указанном порядке, сайты рестрикции во втором генном шарнире содержат Fse I и Asc I в указанном порядке, сайты рестрикции в третьем генном шарнире состоит содержат Not I и сайты рестрикции в четвертом генном шарнире содержат Swa I и Rsr II в указанном порядке.
| US 20050176099, 11.08.2005 | |||
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |

