Настоящее изобретение относится к плазмидным векторам, содержащим p185neu-кодирующие последовательности, и к их применению в ДНК-вакцинации против опухолей. Плазмиды согласно изобретению содержат последовательности, кодирующие различные фрагменты человеческого или крысиного онкогенного белка p185neu, и способны вызвать гуморальный или клеточный иммунный ответ против опухолей, экспрессирующих онкогены семейства ErbB.
Изобретение также относится к фармацевтическим композициям, содержащим указанные плазмиды и к их применению для профилактики или терапии p185neu-экспрессирующих опухолей.
Предпосылки создания изобретения
Белок p185neu, один из наиболее изученных антигенов опухоли, представляет большой интерес в качестве мишени для иммунотерапии рака из-за его наличия на мембране клеток некоторых из самых распространенных человеческих карцином.
p185neu - мембранный рецептор, кодируемый в крысе протоонкогеном Her-2/neu и принадлежащий к семейству рецепторов тирозин-киназы класса I (RTK), который также включает рецептор эпидермального фактора роста EGF-R (ErbB-1) и другие родственные им рецепторы (ErbB-3, ErbB-4). Эти рецепторы участвуют в пролиферации и дифференцировке клеток (Hynes and Stern, 1994 BBA 1198:165) и поэтому привлекают большой биологический и клинический интерес. Рецептор состоит из трех различающихся доменов: внеклеточного, трансмембранного и интрацитоплазматического доменов. p185neu участвует в сложной сети механизмов внутриклеточной трансдукции сигнала и внутриклеточной коммуникации, которые регулируют процессы пролиферации и дифференцировки (Boyle 1992 Curr. Op. Oncol. 4:156). Онкоген neu назван в честь химически-индуцированной нейроглиобластомы крысы, из которой он был впервые выделен. Эта активированная форма neu имеет единственную точечную мутацию, которая приводит к замене "A" на "T" и, вследствие этого, к замене остатка валина в положении 664 p185neu на глутаминовый кислотный остаток (Val664Glu) (Bargmann et al. 1986, Cell 45:649).
Также гомолог человеческого neu, ErbB-2, был изолирован и охарактеризован, и было показано, что и крысиный рецептор HER2/neu и человеческий ErbB2 имеют значительную гомологию с EGFR (Coussens et al. 1985, Sciente 230:1132; Yamamoto et al. 1986, Nature 319:230). В то время как генетическая мутация в крысиной последовательности является причиной конститутивной активации рецептора посредством димеризации, в ErbB-2-положительных человеческих опухолях наблюдается аберрантная экспрессия онкогена (Di Marco et al. 1990, Mol. Cell. Biol. 10:3247; Klapper et al., 2000, Adv Cancer Res, 77:25), даже при том, что, в редких случаях, были найдены активирующие точечные мутации и патологические механизмы сплайсинга (Kwong et al., 1998, Mol Carcinog, 23:62; Xie et al., 2000, J Natl Cancer Inst, 92:412). Полный эффект является подобным: амплификация гена и увеличение уровня транскрипции определяют избыток мембранного рецептора p185neu, соответственно, с увеличением активных димеров, внутриклеточно преобразовывающих сигналы роста независимым от лиганда способом. Кристаллическая структура внеклеточной области человеческого и крысиного p185neu, о которой недавно было сообщено, показывает, что этот белок характеризуется жесткой конформацией, которая позволяет взаимодействовать с другими рецепторами ErbB без непосредственного связывания с лигандами, и вызывать трансдукцию сигнала пролиферации (Cho HS et al. 2003, Nature 421:756).
При нормальных обстоятельствах человеческий p185neu участвует в органогенезе и эпителиальном росте; он экспрессируется в высоких уровнях в течение формирования плаценты и развития плода, тогда как присутствует в очень низких уровнях во взрослых тканях (Press et al. 1990, Oncogene 5:953).
Некоторые исследования показали, что сверхэкспрессия человеческого p185neu связана с опухолевым процессом и с уровнем агрессии опухоли. Сверхэкспрессия p185neu была описана в аденокарциномах легкого (Kern et al. 1986, Cancer Res. 50:5184), ободочной кишки (Cohen et al. 1989, Oncogene 4:81), яичника (Slamon et al. 1989, Science 244:707) и в большом количестве разновидностей человеческого рака груди (Slamon et al. 1989, Science 244:707; Jardines et al. 1993, Pathobiology 61:268).
Фундаментальными свойствами, которые делают p185neu оптимальной мишенью для плазмидной вакцинации, являются a) его прямое участие в клеточном росте и канцерогенезе, поэтому клонированные варианты, в силу генетической нестабильности опухоли, теряют способность к экспрессии этого антигена, а также теряют онкогенные свойства; b) его экспрессия на плазматической мембране, которая делает его распознаваемым антителами даже в опухолевых клетках, утративших способность к экспрессии системы главного комплекса гистосовместимости (Lollini P. and Forni G. 2003, Trends Immunol. 24:62).
Исследования на мышах, трансгенных в отношении активированного крысиного онкогена Her-2/neu (в которых спонтанно развиваются p185neu-положительные грудные опухоли), и на мышиных моделях с использованием p185neu-положительных трансплантируемых линий опухоли, показали возможность профилактики и лечения предраковых поражений. Что касается в особенности профилактики грудных опухолей у мышей, трансгенных в отношении активированного крысиного Her-2/neu, авторы показали, что плазмида, кодирующая внеклеточные и трансмембранные домены крысиного p185neu, способна индуцировать защиту in vivo с большей эффективностью, чем плазмида, кодирующая полноразмерный крысиный p185neu или только внеклеточный домен (секретируемый антиген) (Amici A. et al. 2000, Gene Ther., 7:703; Rovero S. et al. 2000, J. of Immunol., 165:5133). О подобных результатах сообщил Chen et al. (1998, Cancer Res 58:1965). Другие авторы демонстрировали, что плазмиды, кодирующие p185neu - неизмененный или мутированный таким образом, чтобы лишить его тирозин-киназной активности - являются эффективными в профилактике начала развития опухолей после инокуляции p185neu-положительных клеток (Wei WZ et al. 1999, Int. J. Cancer 81:748). Кроме того, плазмиды, лишенные сигнала, ответственного за процессинг в эндоплазматической сети (лидер), который определяет эндоплазматическую локализацию антигена p185neu, оказались столь же эффективными. Защита, индуцируемая различными плазмидами, была главным образом опосредуемой гуморальным иммунным ответом в случае мембранной экспрессии p185neu, и опосредована T-лимфоцитарным иммунным ответом в случае эндоплазматической локализации (Pilon SA et al. 2001, J. of Immunol. 167:3201). Однако комбинированная вакцинация плазмидами, вызывающими суперэкспрессию p185neu как в цитоплазме, так и в мембране, была более эффективна в защите против роста опухоли (Piechocki MP et al. 2001, J. Immunol. 167:3367).
Таким образом, баланс между различными механизмами иммунного ответа мог бы быть особенно важен (Reilly et al., 2001, Cancer Res. 61:880). Кроме того, наблюдалось, что вакцинация плазмидами, кодирующими внеклеточные и трансмембранные домены крысиного p185neu, может ликвидировать массы опухоли с диаметром 2 мм при инокуляции клеток, осуществляющих сверхэкспрессию p185neu, посредством множества различных механизмов иммунной системы (T-хелперы и T-киллеры, антитела, макрофаги, нейтрофилы, NK-клетки, рецепторы Fc, гамма-интерферон и перфорины), которые совместно действуют на устранение опухоли (Curcio C. et al. 2003, J. Clin. Invest. 111:1161).
Описание изобретения
Различные конструкции, кодирующие человеческий или человеческий/крысиный химерный p185 белок, были встроены в плазмидные векторы и использовались в экспериментах по иммунизации, направленных на профилактику развития опухоли. Для конструирования плазмиды фрагменты человеческого p185neu белка, содержащего трансмембранный домен и части внеклеточного домена меньшей длины, были получены от последовательности онкогена ErbB2, или его части были заменены гомологичными последовательностями кДНК крысиного Her-2/neu, чтобы получить химерные плазмиды.
Таким образом, полученные плазмиды были оценены в экспериментах по вакцинации мышей, инокулированных опухолевыми клетками, осуществляющих сверхэкспрессию человеческого p185neu. Плазмиды, содержащие обрезанные формы p185neu, индуцировали противоопухолевую реактивность, опосредуемую хелперными и киллерными Т-лимфоцитами, в то время как химерные плазмиды индуцировали гуморальный иммунный ответ как против человеческого, так и против крысиного p185neu.
На основании результатов экспериментов in vivo были отобраны плазмиды, содержащие p185neu последовательности, приведенное в конце описания, способные вызывать сильный иммунный ответ как клеточного, так и гуморального типа. Эти плазмиды, являющиеся объектом настоящего изобретения, содержат последовательность, кодирующую фрагмент p185neu, выбранный из группы, состоящей из SEQ ID NO:1-14 (последовательности человеческого и крысиного p185neu доступны в Банке генов под номерами доступа M11730 и X03362 соответственно).
Согласно изобретению p185neu-кодирующие последовательности могут быть встроены в любые плазмидные векторы, подходящие для введения человеку. Помимо кодирующих последовательностей, плазмиды могут содержать функциональные элементы для контроля транскрипции, в частности промотор, размещенный выше кодирующей последовательности, предпочтительно, промотор CMV, элементы начала и остановки транскрипции, селекционные маркерные гены, такие как гены резистентности к ампициллину или канамицину, звенья CpG, сайты полиаденилирования или активаторы транскрипции. Элементы контроля транскрипции должны быть пригодны для использованием вектора на людях. В предпочтительном варианте осуществления плазмиды по изобретению содержат по меньшей мере 4 звена CpG, предпочтительно по меньшей мере 8, максимум 80. Звенья CpG (ATAATCGACGTTCAA) бактериального происхождения индуцируют в макрофагах секрецию IL-12, которые, в свою очередь, вызывают секрецию гамма IFN NK-клетками, таким образом активизируя опосредуемую Т-хелперным лимфоцитом реакцию (Chu R.S. et al. 1997, J. Exp. Med., 186:1623). Поэтому вставка звеньев CpG в последовательности плазмиды усиливает иммунный ответ.
В следующем варианте осуществления изобретение относится к фармацевтической композиции, содержащей одну или более различных плазмид, как определено выше, в комбинации с фармацевтически приемлемыми носителями и эксципиентами. Фармацевтические композиции в форме, подходящей для парентерального введения, предпочтительно в форме инъецируемого раствора, удобны в использовании для ДНК-вакцинации. Принципы и способы ДНК-вакцинации известны специалисту и описаны, например, Liu MA 2003; J Int Med 253:402.
В другом варианте осуществления изобретение относится к комбинированному препарату, содержащему по меньшей мере два, предпочтительно по меньшей мере четыре, более предпочтительно по меньшей мере восемь различных плазмид для одновременного, последовательного или раздельного введения пациенту.
Плазмиды, композиции и препараты согласно изобретению используются в профилактическом или терапевтическом лечении пациентов с риском развития p185neu-положительных опухолей, или пациентов с первичными опухолями, метастазами или рецидивами p185neu-положительных опухолей. Профилактика может быть первичной, когда опухоль не проявляется, вторичной, когда опухоль находится в начальных фазах, таких как предраковые поражение, или третичной, в случае рецидива опухоли или метастатического процесса. Опухолями, для которых может быть показано лечение плазмидами по изобретению, являются опухоли эпителиального происхождения, в частности аденокарциномы легкого, яичника и молочной железы и, в более общем смысле, опухоли, экспрессирующие белок p185neu.
Подробное описание изобретения
Конструирование основы плазмиды pCMV3.1
Для конструирования плазмид, кодирующих фрагменты человеческого p185neu, и химерных плазмид использовали основу pCMV3.1. Фрагменты, происходящие от кДНК человеческого протоонкогена ErbB-2 и от кДНК крысиного протоонкогена Her-2/neu, были вставлены в pCMV3.1 (Invitrogen, Milano, Italia), с удалением с помощью рестрикционных ферментов DraIII (nt1531) и BsmI (nt3189) фрагмента длиной 1658 пар оснований, содержащего сайт начала репликации f1, сайт начала репликации и ранний промотор SV40, ген, кодирующий устойчивость к неомицину и сигналы полиаденилирования SV40. Полученная модифицированная плазмида (pCMV3.1) представляет некоторые преимущества по сравнению с нативной пкДНК3.1. Фактически уменьшение размера до 3900 пар оснований и удаление ненужных последовательностей способствует увеличению эффективности трансфекции in vivo.
Конструирование плазмиды pCMV3.1 erbB2
Человеческая кДНК ErbB2, полученная от плазмиды pSVerbB2, была встроена в множественный сайт клонирования pCMV3.1 с сайтами рестрикции HindIII 10 и XbaI. Эта плазмида используется для конструирования плазмид, экспрессирующих усеченный p185neu и химерных плазмид.
Конструирование плазмид, содержащих последовательность 4XCpG: pCMV3.1hECD-TM-4CpG и pCMV3.1hECD-TM-4noCpG
После удаления последовательности, кодирующей интрацитоплазматический домен, из плазмиды pCMV3.1-erbB2 были получены две плазмиды, кодирующие внеклеточные и трансмембранные области протоонкогена ErbB2. Процедура включала сначала рестрикционный анализ, чтобы идентифицировать уникальные сайты, присутствующие в нуклеотидной последовательности кДНК ErbB2. Был идентифицирован уникальный сайт, распознаваемый ферментном AccIII (nt 2195), размером приблизительно 20 пар оснований ниже конца трансмембранного домена.
Цитоплазматический домен был удален с использованием фермента AccIII, присутствующего в качестве уникального сайта рестрикции, и фермента XbaI. Для повторного встраивания на 3' концах ДНК ErbB2 ECD-TM триплета нуклеотидов TAA, распознаваемого как сигнал остановки трансляции, авторы использовали две синтетических последовательности, состоящие из двух олигонуклеотидов: смыслового (олигонуклеотид #1, #3) и антисмыслового (олигонуклеотид #2, #4), содержащих сайты рестрикции AccIII и XbaI на своих концах. В этих синтетических последовательностях есть также четыре повторяющихся последовательности CpG и noCpG. Последний используется как отрицательный контроль. Эти две новых плазмиды назвали pCMV3.1hECD-ТМ-4CpG и pCMV3.1hECD-ТМ-4noCpG.
Конструирование плазмид, содержащих последовательность 8XCpG: pCMV3.1H/NhECD-TM-8CpG и pCMV3.1H/NhECD-ТМ-8noCpG
Чтобы добавить далее неспецифические иммунные стимулы, авторы создавали новую основу плазмиды, содержащую 4 иммуностимулирующих последовательности СрС, названные pCMV3.1H/N-4CpG. С этой целью авторы модифицировали pCMV3.1, чтобы удалить один из двух сайтов рестрикции для фермента PmeI и инвертировать сайты рестрикции для HindIII и NneI, присутствующие в множественном сайте клонирования, посредством синтетической последовательности, состоящей из двух олигонуклеотидов: смыслового (олигонуклеотид #5) и антисмыслового (олигонуклеотид #6). В эту новую плазмиду, названную pCMV3.1 H/N, были вставлены две синтетических последовательности, состоящие из двух олигонуклеотидов: смыслового (олигонуклеотид #7, #9) и антисмыслового (олигонуклеотид #8, #10), содержащих четыре повтора для CpG и noCpG последовательностей в уникальных сайтах рестрикции XbaI и PmeI, с получением, таким образом, pCMV3.1 H/N-4CpG и 4noCpG. После этого ДНК-фрагменты hECD-TM-4CpG и hECD-TM-4noCpG были вставлены в pCMV3.1 H/N-4CpG и в pCMV3.1 H/N-4noCpG соответственно, с получением, таким образом, двух новых плазмид, названных pCMV3.1H/N-hECD-TM-8CpG и pCMV3.1H/N-hECD-TM-8noCpG.
Конструирование плазмиды, содержащей последовательность второго домена цистеина и трансмембранного домена человеческого p185neu: pCMV3.1H/Nh2°cysECD-TM-8CpG
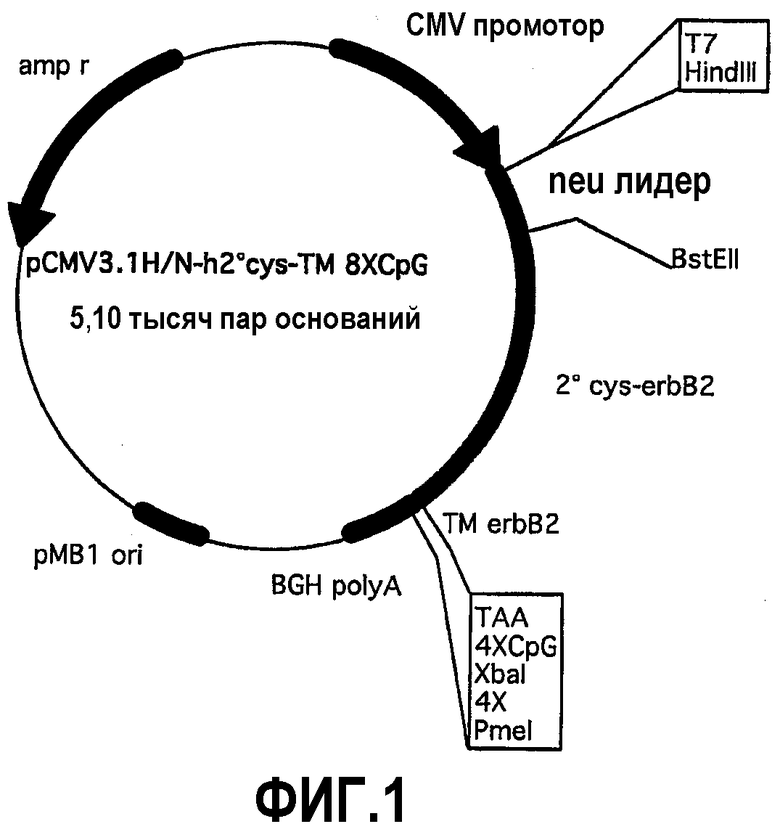
Человеческий p185neu внеклеточный домен характеризуется двумя областями, богатыми цистеинами, известными как 1-й и 2-ой субдомен цистеина (1-й cys и 2-ой cys). В отличие от крысиной кДНК последовательности, содержащей только один сайт BstEII (nt1250) во внеклеточном домене, расположенном в нуклеотидной области, которая отделяет 1-ый cys от 2-ого cys, кДНК последовательность внеклеточного домена ErbB2 имеет два сайта рестрикции для BstEII: в дополнение к сайту в том же самом положении, как в случае крысиного (nt1372), другой сайт BstEII (nt963) присутствует в части, кодирующей 1-ый cys внеклеточного домена. Расщеплением плазмиды pCMV3.1H/NhECD-TM-8CpG с использованием HindIII и BstEII получали фрагмент ДНК, состоящий из 2-ого cys от внеклеточного домена, трансмембранного домена, последовательности 8CpG, и плазмиду pCMV3.1H/N. Затем ферментативной амплификацией ДНК (ПЦР) встраивали сигнал для секреции крысиного p185neu через эндоплазматическую сеть, используя смысловой олигонуклеотид, состоящий из праймера T7 (олигонуклеотид # 11), который распознает Т7 РНК-полимеразу, расположенного в начале pCMV3.1H/N множественного сайта клонирования, и антисмысловой олигонуклеотид (олигонуклеотид #12), имеющий на конце сайт BstEII. После очистки, ферментативного расщепления амплифицированного фрагмента с ферментами рестрикции HindIII и BstEII и последующего клонирования получили pCMV3.1H/Nh2°cys-ТМ-8CpG (фиг.1). Эта плазмида использовалась в экспериментах по вакцинации для сравнения ее с pCMV3.1 H/NhECD-TM-8CpG. После этого получали химерную кДНК, кодирующую слитый белок между 2-ым cys и трансмембранным доменом (nt 1372 - nt 2204) человеческой последовательности и 1-м cys (nt 1 - nt 1250) крысиной последовательности. Восстановление всей последовательности белка слиянием частей, происходящих от крысиной и человеческой кДНК соответственно, позволяет усиливать иммунный ответ.
Конструирование химерной плазмиды, содержащей последовательность первого домена цистеина крысиного p185neu и второго домена цистеина и трансмембранного домена человеческого (nt 1 - nt 1250): pCMV3.1H/N-r1°cysh2°cysTM-8CpG
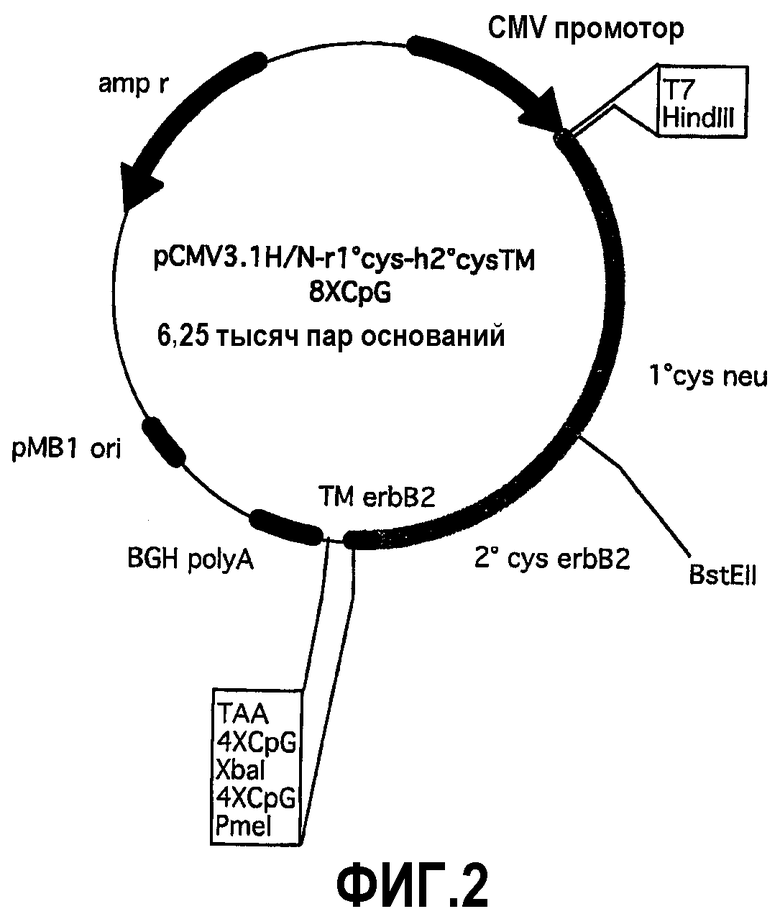
В отличие от крысиной кДНК-последовательности, содержащей только BstEII (nt1250) сайт во внеклеточном домене, расположенном в нуклеотидной области, которая разделяет первую и вторую области, богатые цистеинами, кДНК последовательность внеклеточного домена Erb2 имеет два сайта рестрикции для BstEII: один в положении 1372 (nt), как в крысиной последовательности, и другой в положении 963 (nt), то есть в части последовательности, кодирующей 1-ый cys внеклеточный домен. Наличие сайта BstEII в том же самом положении как в домене крысиной кДНК (1250nt), так и в человеческой кДНК (1372nt), позволило получить плазмиду, способную кодировать продукт слияния между крысиным 1-ым cys и человеческим 2-ым cys. Фактически расщепление pCMV3.1H/N-h2°cysTM-8CpG с помощью ферментов рестрикции HindIII и BstEII позволило заменить фрагмент ДНК, кодирующий сигнал секреции крысиного p185neu нуклеотидной последовательностью, кодирующей крысиный 1-ый cys, полученной расщеплением pCMV3.1rECD-TM4CpG теми же самыми ферментами. Продукт плазмиды pCMV3.1H/N-r1°cys-h2°cysTM-8CpG (фиг.2) состоит из части 412 аминокислот крысиного p185neu, и части 274 аминокислот человеческого p185neu. Эта новая плазмида, pCMV3.1H/Nr1°cysh2°cysTM-8CpG, была использована в экспериментах по вакцинации с использованием pCMV3.1H/N-hECD-TM-8CpG для сравнения. Неожиданно, плазмида, кодирующая химерный белок, вызывает в мышах полную защиту против опухолей, экспрессирующих человеческий p185neu (таблица). Эта защита подобна индуцируемой pCMV3.1H/N-hECD-TM-8CpG. Более того, анализ сыворотки мышей, привитых обеими плазмидами, доказал сходный титр антител к человеческому p185neu.
Плазмиды, способные кодировать уменьшенные фрагменты внеклеточного и трансмембранного домена человеческого p185neu
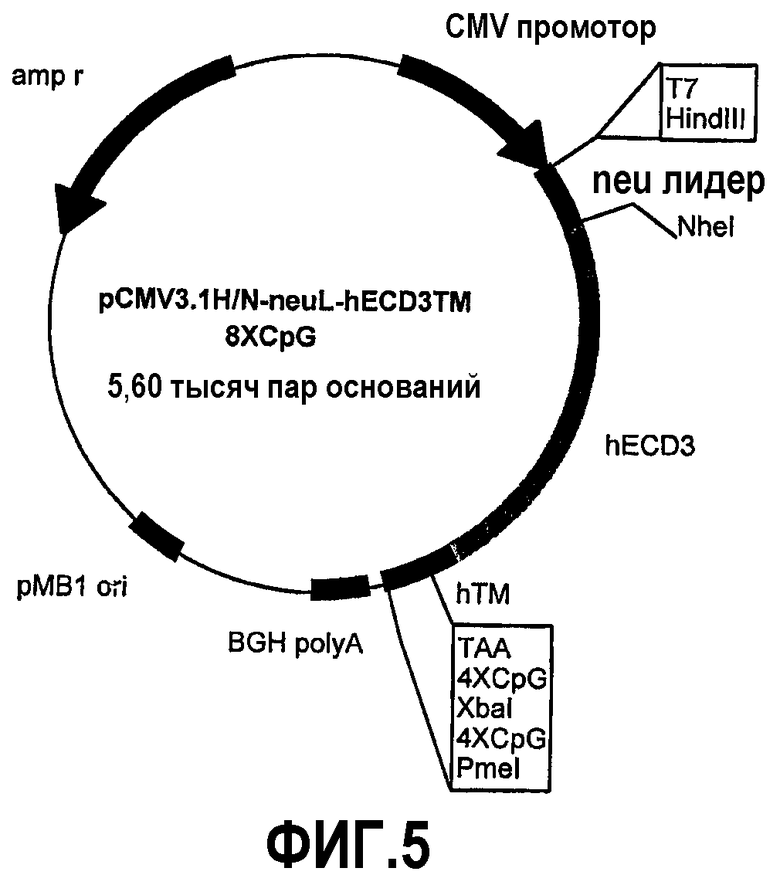
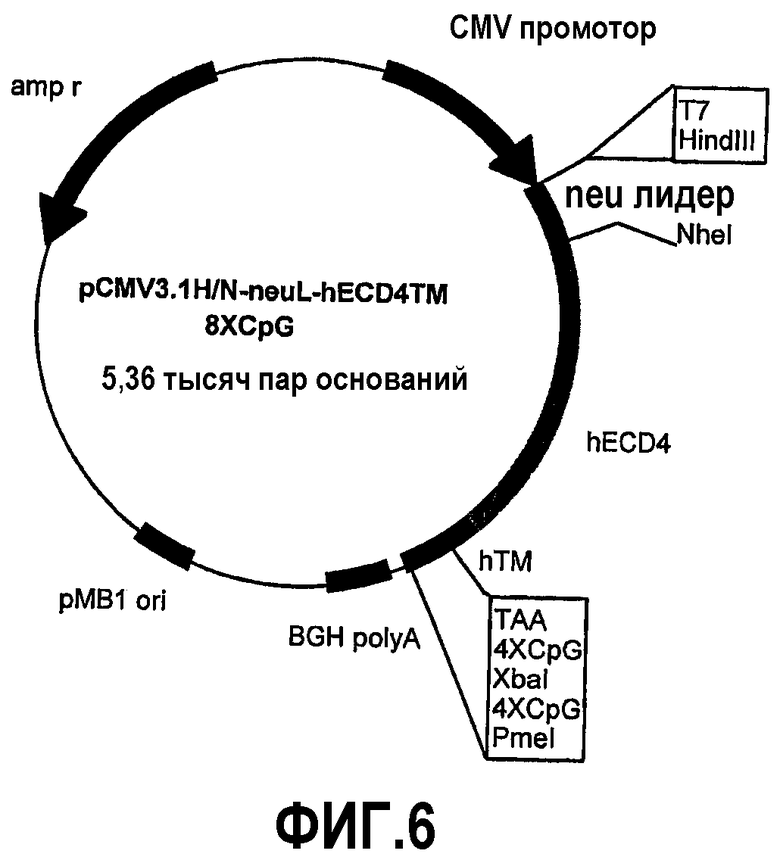
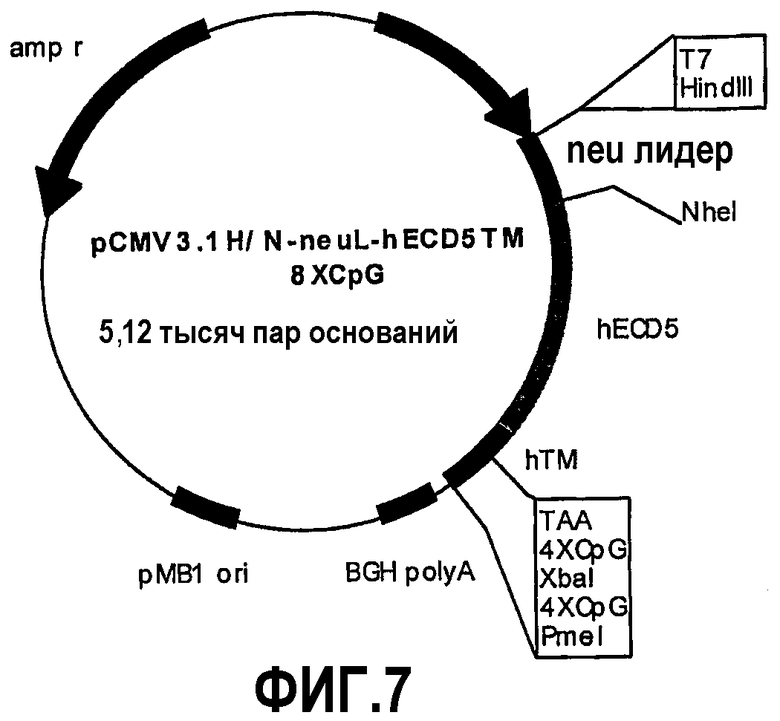
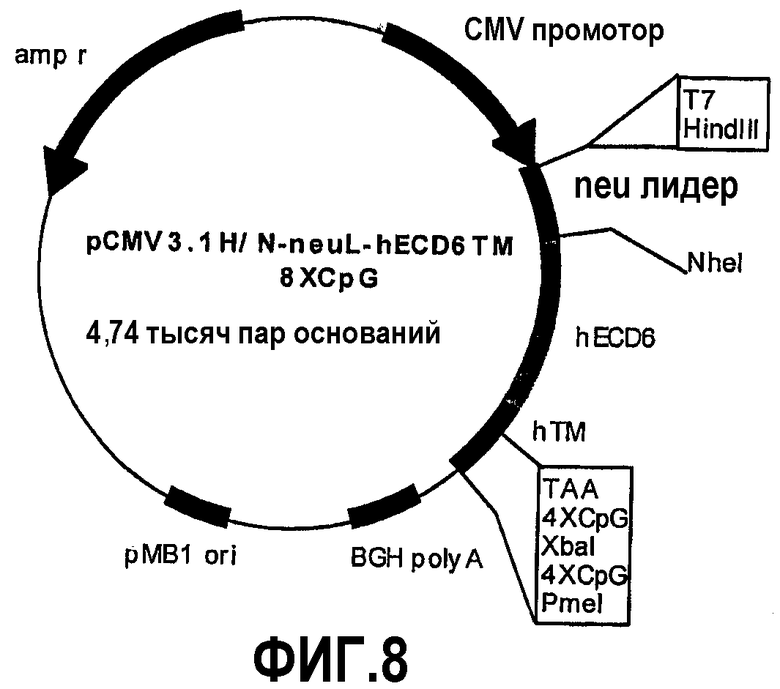
Конструкция семи плазмид, которые кодируют уменьшенные фрагменты внеклеточного и трансмембранного домена человеческого p185neu, а именно: pCMV3.1H/NhECD1-TM-8CpG (-70 аминокислот), pCMV3.1H/NhECD2-TM-8CpG (-150 аминокислот), pCMV3.1H/NhECD3-TM-8CpG (-230 аминокислот), pCMV3.1H/NhECD4-ТМ-8CpG (-310 аминокислот), pCMV3.1H/NhECD5-TM-8CpG (-390 аминокислот), pCMV3.1H/NhECD6-TM-8CpG (-470 аминокислот) и pCMV3.1H/NhECD7-TM-8CpG (-550 аминокислот).
Фрагмент, кодируемый первым из этих фрагментов, короче на 70 аминокислот (делеция 360 пар оснований). Все другие короче на 80 аминокислот (делеция 240 пар оснований).
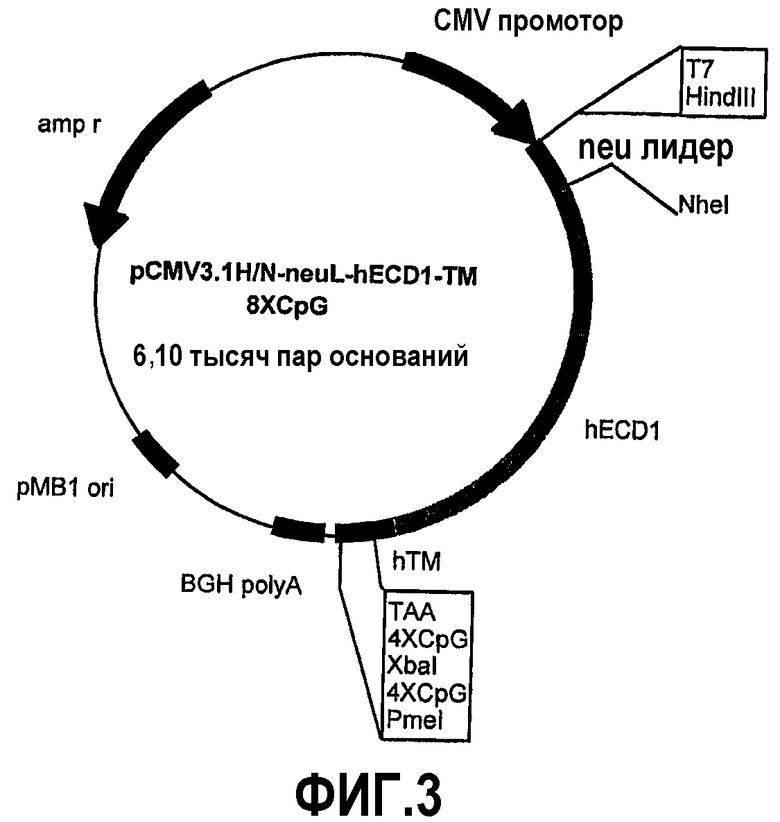
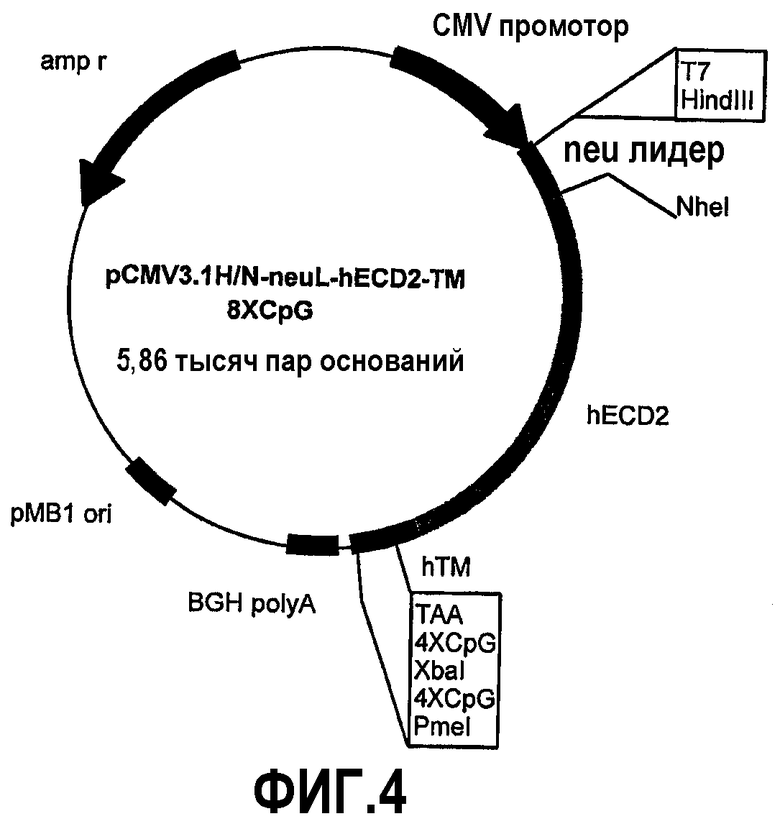
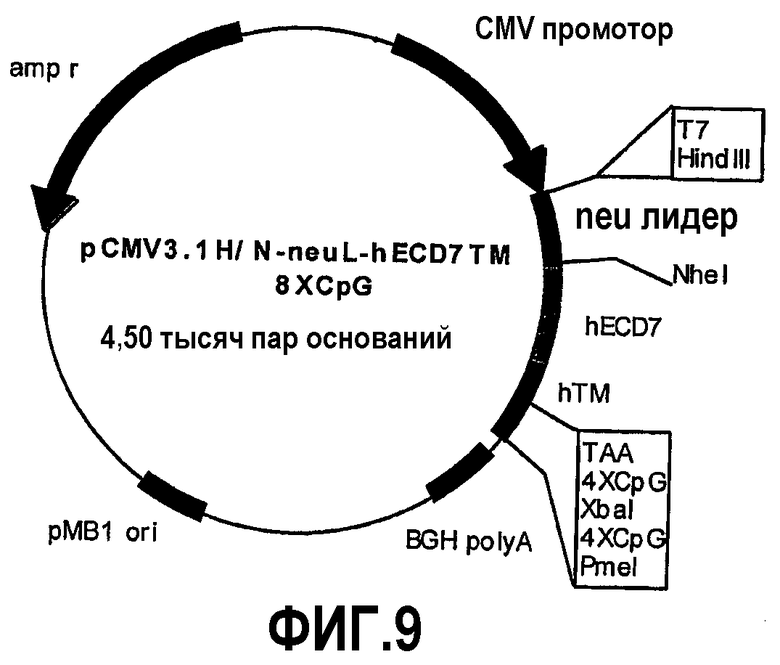
Эти фрагменты были получены ферментативной амплификацией ДНК с использованием семи различных смысловых олигонуклеотидов с сайтом рестрикции NheI (олигонуклеотиды #13-#19) на конце и антисмыслового олигонуклеотида (олигонуклеотид #20), способного распознавать сайт, называемый "пкДНК3.1/BGH Reverse priming site" (830-850 nt), находящийся на 3'-конце полилинкера pCMV3.1. После ферментативного расщепления ферментами рестрикции NheI и PmeI продукты амплификации клонировали в pCMV3.1H/N-neu лидере, предварительно полученном встраиванием сигнала секреции в эндоплазматической сети крысиного p185neu в сайтах рестрикции. Фрагмент ДНК сигнала секреции крысиного p185neu был получен ферментативной амплификацией ДНК с использованием праймера T7 (олигонуклеотид #11) в качестве смыслового нуклеотида и антисмыслового нуклеотида (олигонуклеотид #21) с сайтом NheI на конце. Амплифицированный фрагмент после очистки и рестрикции HindIII и NheI клонировали в плазмиде pCMV3.1H/N, расщепляли теми же самыми ферментами, получая таким образом pCMV3.1H/N-neu лидер. Мембранная экспрессия различных усеченных форм человеческого p185neu ожидается ввиду наличия в эндоплазматической сети сигнала секреции крысиного p185neu. Плазмиды, кодирующие усеченные формы pCMV3.1H/NhECD1-TM-8CpG (фиг.3), pCMV3.1H/NhECD2-ТМ-8CpG (фиг.4), pCMV3.1H/NhECD3-ТМ-8CpG (фиг.5), pCMV3.1H/NhECD4-TM-8CpG (фиг.6), а также контрольная плазмида pCMV3.1H/NhECD-TM-8CpG, обеспечивают у 100% вакцинированных мышей защиту против летальной инокуляции опухолевых клеток, экспрессирующих человеческий p185neu (таблица). Плазмида pCMV3.1H/NhECD5-TM-8CpG (фиг.7) обеспечивает защиту 60% животных (таблица), в то время как плазмиды pCMV3.1H/NhECD6-TM-8CpG и pCMV3.1H/NhECD7-TM-8CpG (фиг.8, 9) не имеют защитного эффекта в отношении летальной инокуляции опухолевых клеток, экспрессирующих человеческий p185neu (таблица). Белковые продукты, экспрессируемые этими различными плазмидами, не секретируются через эндоплазматический ретикулум. Отсутствие консенсусных последовательностей, необходимых для гликозилирования и для их процессинга в эндоплазматической сети, или конформационные изменения из-за делеции аминокислот на NH2-конце, может объяснить отсутствие белковых продуктов в мембране. Поэтому, чтобы далее проверить, правильно ли экспрессируются различные усеченные формы внеклеточного и трансмембранного домена человеческого p185neu, были получены новые плазмиды, кодирующие слитые белки, характеризующиеся эпитопом myc на NH2-конце. Эти рекомбинантные белки распознаются anti-myc моноклональным антителом, поэтому софокусной микроскопией возможно анализировать их экспрессию и локализацию.
Сначала получали новую плазмиду, кодирующую сигнал секреции в крысиной эндоплазматической сети (neu лидер) и myc эпитоп. Проводили клонирование, используя синтетическую последовательность, состоящую из смыслового (олигонуклеотид #22) и антисмыслового (олигонуклеотид #23), имеющую на обоих концах сайт NheI. Сайт NheI в 5'-положении был мутирован так, чтобы при правильном лигировании он не распознавался ферментом. Таким образом получили pCMV3.1H/Nneuleader-myc эпитоп. С помощью этой плазмиды в сайтах рестрикции NheI и PmeI клонировали последовательности, кодирующие усеченные формы человеческого p185neu. Затем 3T3 фибробласты NIH были трансфицированы in vitro плазмидами с использованием липофектамина 2000 (Invitrogen, Милан, Италия). Через 48 часов трансфицированные клетки анализировали софокусной микроскопией, используя FITC-сопряженное анти-myc моноклональное антитело (Sigma-Aldrich Srl, Милан, Италия). Таким образом, было показано, что все кодируемые плазмидами усеченные формы локализованы в цитоплазме. 3T3 фибробласты NIH были трансфицированы параллельно плазмидой pCMV3.1H/NhECD-TM-8CpG и анализировались софокусной микроскопией с использованием c-erbB2/c-neu моноклонального антитела Ab-3 (Oncogene, Бостон, MA) в качестве первичного антитела и FITC-конъюгированного вторичного антимышиного антитела (PharMigen, Сан-Диего, Калифорния). Таким образом, наблюдалось, что человеческий ECD-TM экспрессируется в мембране. Результаты, полученные с использованием первых четырех плазмид, описанных выше (pCMV3.1H/NhECD1-ТМ-8CpG, pCMV3.1H/NhECD2-ТМ-8CpG, pCMV3.1H/NhECD3-TM-8CpG, pCMV3.1H/NhECD4-TM-8CpG), показывают, что клеточного ответа достаточно для противоопухолевой профилактики. Однако известно, что одновременная активация клеточного и гуморального ответа необходима для более эффективной терапии (Rielly et al., 2001, Cancer Res 61:880). Как уже описано в предыдущем параграфе, химерный белок, кодируемый плазмидой pCMV3.1H/N-r1°cys-h2°cysTM-8CpG, в состоянии защитить 100% вакцинированных животных и в состоянии вызвать сильный гуморальный ответ у мышей.
Химерные плазмиды, способные кодировать пять различных химерных крыса-человек p185neu
Для конструирования плазмид, кодирующих химерные белки, авторы выбрали pCMV3.1H/NhECD1-TM-8CpG, pCMV3.1H/NhECD2-ТМ-8CpG, pCMV3.1H/NhECD3-TM-8CpG и pCMV3.1H/NhECD4-TM-8CpG. Эти четыре плазмиды защищают 100% привитых мышей против летальной инокуляции опухолевых клеток, экспрессирующих человеческий p185neu. Также была отобрана плазмида pCMV3.1H/NhECD5-TM-8CpG, хотя она защищает только 60% привитых мышей, потому что кодируемый белок отличается только 17 аминокислотами от кодируемого pCMV3.1H/Nh2°cysECD-TM-8CpG (275 аминокислот), который защищает 20% привитых мышей. Можно выдвинуть гипотезу, что пептидная последовательность 17 аминокислот соответствует эпитопу, важному для индукции эффективного иммунного ответа.
ДНК фрагменты, кодирующие части крысиного p185neu, были получены ферментативной амплификацией ДНК. Для амплификации этих кДНК фрагментов использовали шесть олигонуклеотидов, имеющих одинаковую ориентацию, а именно ориентацию праймера T7 (олигонуклеотид #11), в то время как пять антисмысловых были предназначены для распознавания крысиной кДНК в правильных положениях и имеют сайт рестрикции для NheI на своих концах (олигонуклеотиды #24-#28). После очистки и расщепления рестрикционными ферментами HiNdIII и NheI, амплифицированные фрагменты встраивали в соответствующие плазмиды (pCMV3.1H/NhECD1-ТМ-8CpG, pCMV3.1H/NhECD2-ТМ-8CpG, pCMV3.1H/NhECD3-ТМ-8CpG, pCMV3.1H/NhECD4-ТМ-8CpG, pCMV3.1H/NhECD5-TM-8CpG) и расщепляли теми же самыми ферментами рестрикции. Таким образом, получили пять новых плазмид, способных кодировать химерные белки из 689 аминокислот, из которых 2 (Val-Ser) относятся к сайту рестрикции NheI, используемому для конъюгирования крысиной и человеческой ДНК. Наличие этих двух аминокислот делает гетеролитическими как человеческие, так и крысиные части.
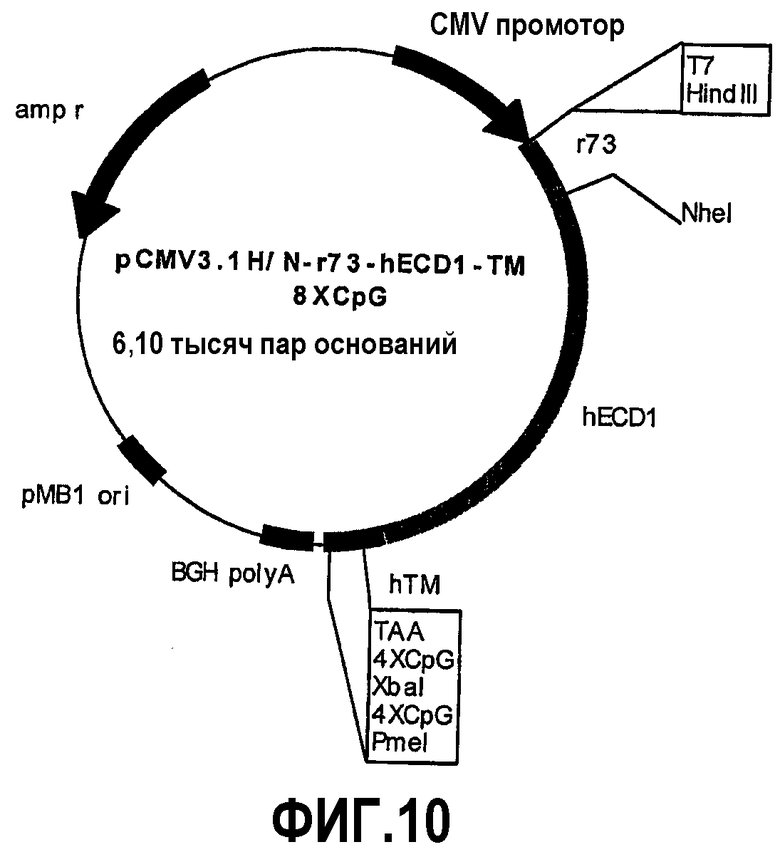
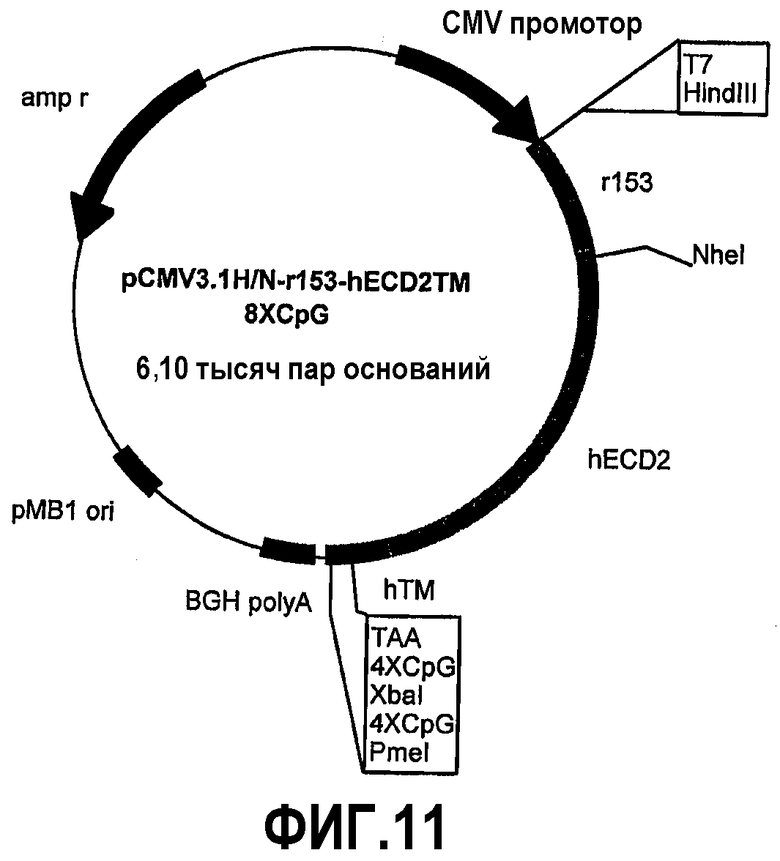
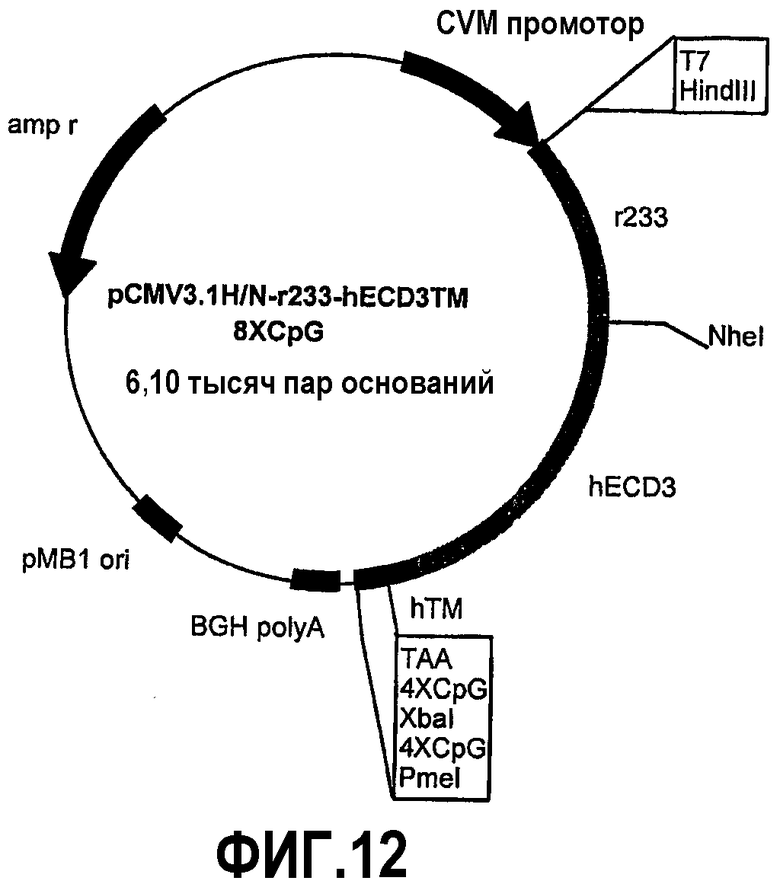
Химерные белки отличаются для уменьшающихся частей человеческого p185neu и увеличивающихся частей крысиного p185neu. Плазмида pCMV3.1H/Nr73-hECD1-TM-8CpG (фиг.10) кодирует 73 аминокислоты внеклеточного домена крысиного p185neu и 614 аминокислот человеческого p185neu. Плазмида pCMV3.1H/Nr153-hECD2-TM-8CpG (фиг.11) кодирует 153 аминокислоты внеклеточного домена крысиного p185neu и 534 аминокислоты человеческого p185neu. Плазмида pCMV3.1H/Nr233-hECD3-TM-8CpG (фиг.12) кодирует 233 аминокислоты внеклеточного домена крысиного p185neu и 454 аминокислоты человеческого p185neu. Плазмида pCMV3.1H/Nr313-hECD4-TM-8CpG (фиг.13) кодирует 313 аминокислот внеклеточного домена крысиного p185neu и 374 аминокислоты человеческого p185neu. Плазмида pCMV3.1H/Nr393-hECD5-TM-8CpG (фиг.14) кодирует 393 аминокислоты внеклеточного домена крысиного p185neu и 294 аминокислоты человеческого p185neu. Косвенное доказательство мембранной экспрессии человеческого/крысиного химерного p185neu, кодируемого этими плазмидами, было получено путем иммунизации мышей пятью новыми плазмидами и pCMV3.1H/N-r1°cys-h2°cysTM-8CpG в качестве положительного контроля. Сыворотка всех привитых мышей содержит специфические антитела против человеческого p185neu. Кроме того, животные, вакцинированные плазмидами, кодирующими пять различных химерных белков, также защищены от летальной инокуляции опухолевых клеток, экспрессирующих человеческий p185neu.
ПРИМЕРЫ
Пример 1
Конструирование плазмиды pCMV3.1H/N-r1°cys-h2°cysTM-8CpG
Для получения химерной плазмиды pCMV3.1H/N-r1°cys-h2°cysTM-8CpG использовали плазмиду pCMV-ECD-TM, которая экспрессирует внеклеточный и трансмембранный домен крысиного p185neu (Amici et al. 2000, Gene Ther., 7:703). pCMV-ECD-TM расщепляли ферментами рестрикции HindIII и XbaI (BioLabs, Beverly, MA), чтобы отделить вставку от основы плазмиды.
Рестрикция ферментом HindIII:
Смесь культивировали при 37°C в течение 4 ч, и продукт рестрикции контролировали электрофорезом на 1%-ном агарозном геле, используя маркер молекулярной массы и нерасщепленную плазмиду как контроль.
После подтверждения линеаризации плазмиды ДНК осаждали, добавляя 1/10 объема 3М ацетата натрия с рН 5,2 и 2 объема холодного абсолютного этанола.
Образец сохраняли на льду в течение 20 мин, затем центрифугировали с использованием миницентрифуги при 14000 об/мин в течение 12 мин. Осадок был промыт три раза 1 мл 70%-ного холодного этанола, высушен под вакуумом в течение 5 мин, затем повторно суспендирован в 84 мкл H2О и ферментативно расщеплен ферментом рестрикции XbaI.
Рестрикция ферментом XbaI:
Смесь культивировали при 37°C в течение 4 ч, и продукт расщепления осаждали и высушивали, как описано выше. ДНК ресуспендировали в 30 мкл H2О.
Два фрагмента ДНК, соответствующие основе плазмиды (pCMV, длиной 4400 пар оснований) и вставке (ECD-TM, длиной 2100 пар оснований), разделяли электрофорезом на 1%-ном агарозном геле.
Полосу, соответствующую вставке, удаляли, и ДНК элюировали с геля, используя набор Qiaquick gel extraction (Qiagen Италия).
Параллельно, новую основу плазмиды (pCMV3.1H/N-4CpG) с фрагментом ДНК, соответствующим крысиному p185 ECD-TM, расщепляли теми же самыми ферментами рестрикции и элюировали на агарозном геле.
Фрагменты ДНК, соответствующие крысиному ECD-TM и линеаризованной плазмиде pCMV3.1H/N-4CpG, использовались для получения pCMV3.1H/N-rECD-TM-4CpG реакцией лигирования.
Реакция лигирования
Реакцию лигирования инкубировали при 16°C в течение 4 ч.
Продукт лигирования затем использовали для трансформации бактериального штамма DH5 α E. coli. Бактериальные клетки делали компетентными методом CaCl2.
Трансформация бактериального штамма DH5 α:
Для проникновения плазмидной ДНК в компетентные клетки, они сохранялись на льду в течение 40 мин и подвергались тепловому шоку (1,5 мин при 42°C и затем 2 мин на льду).
После добавления 1 мл среды роста LB трансформированные бактериальные клетки культивировали при 37°C в течение 1 ч, чтобы восстановить их физиологическое состояние.
Затем суспензию клеток центрифугировали при 6000 об/мин в течение 1 мин, и осадок ресуспендировали в 100 мкл LB.
Клетки высевали в чашки Петри, содержащие селективную твердую среду (LB с агар-агаром + ампициллин 100 мкг/мл) и выращивали при 37°C в течение 1 ночи. Ампициллин обеспечивает рост клеток, содержащих плазмиду pCMV3.1H/N-rECD-ТМ-4CpG, которая придает устойчивость к ампициллину.
Полученные клоны анализировали щелочным лизисом, чтобы выбрать те, которые содержат рекомбинантную плазмиду pCMV3.1H/N-rECD-TM-4CpG.
Чтобы получить химерную плазмиду pCMV3.1H/N-r1°cys-h2°cysTM-8CpG, плазмиду pCMV3.1H/N-rECD-ТМ-4CpG расщепляли ферментами рестрикции, BstEII и XbaI, чтобы удалить второй домен цистеина вместе с трансмембранным доменом крысиного p185neu. В то же самое время, плазмиду pCMV3.1hECD-TM-4CpG расщепляли теми же самыми ферментами, чтобы выделить фрагмент ДНК, соответствующий второму цистеиновому субдомену и трансмембранному домену человеческого гена.
Расщепление с использованием BstEII:
Смесь культивировали при 60°C в течение 4 ч.
Рестрикция с использованием XbaI, восстановление фрагментов, которые используются для клонирования, реакции лигирования и трансформации компетентных клеток были описаны выше.
Полученная химерная плазмида pCMV3.1H/N-r1°cys-h2°cysTM-8CpG была секвенирована с использованием автоматического ABI PRISM 310 Genetic Analyzer (Applied Biosystem), для подтверждения правильной вставки фрагмента, соответствующего второму субдомену цистеина и трансмембранному домену человеческого гена.
Пример 2
in vivo тест
Животные
Для всех экспериментов использовались самки мышей Balb/cAnCr (H-2d) в возрасте приблизительно семи недель.
Животные, поставляемые Charles River Laboratories (Calco, MI, Италия), выращены в асептических условиях и в соответствии с European Community guidelines.
Внутримышечное введение, сопровождаемое in vivo электропорацией
Во избежание нежелательных сокращений большеберцовой мышцы, каждая мышь была анестезирована i.p. введением 300 мкл авертина, полученного из 0,58 г трибромэтанола (Sigma-Aldrich) и 310 мкл трет-амилового спирта (Aldrich), разведенных в 39,5 мл деионизированной H2О. Все мыши были обриты в области большеберцовой мышцы для инокуляции.
Животных вакцинировали путем введения в обе переднебольшеберцовые мышцы 40 мкл раствора, содержащего 50 мкг ДНК.
Содержащая ДНК смесь была получена незадолго до использования, в соответствии с указаниями Dr. F. Pericle (Valentis, Inc., The Woodlands, Texas, USA). Этот раствор содержит плазмидную ДНК в количестве 1,25 мг/мл, 6 мг/мл поли-L-глутамата натриевой соли (Sigma-Aldrich, S.r.l., Милан, Италия), 150 мМ хлорида натрия (Fluka, BioChemika, Buchs, Швейцария) и дистиллированную воду, свободную от эндотоксинов (Nucleare Free Water, Promega Corporation) на конечный объем 1 мл.
Приблизительно после 5 мин после инокуляции, обработанная область была подвергнута электропорации, с применением двух электрических импульсов, имеющих интенсивность 375 V/cm2, каждая длительностью 25 мс, с использованием электропоратора Electro Square Porator (T820, BTX, Сан-Диего, Калифорния, США). Чрескожные электрические импульсы были применены при помощи двух квадратных стальных электродов, помещенных в 3-х мм друг от друга, около каждой лапы. Генная иммунизация электропорацией была выполнена дважды для каждого животного за 21 и 7 дней до инокуляции опухолевых клеток.
Инокуляция опухолевых клеток
Мышам вводили суспензию, содержащую 2×105 клеток D2F2/E2. Эти клетки происходят от грудной опухоли, спонтанно генерируемой в гиперплазированном альвеолярном узле мыши BALB/c и экспрессируют высокие уровни человеческого p185.
In vivo оценка роста опухоли
Рост опухоли оценивали еженедельно пальпацией, и размеры опухолей были измерены по двум перпендикулярным диаметрам с калибром. Опухолевые массы, насчитывающие более 3 мм, рассматривают как опухоли.
Рост опухоли отслеживали в течение 100 дней от инокуляции опухоли или до достижения опухолью диаметра более 10 мм, после чего животных умерщвляли.
Мыши: самки BALB/c
Опухоль: D2F2-E2, экспрессирующая человеческий p185neu
Список олигонуклеотидов, синтезируемых и используемых для конструкции плазмиды




























Изобретение относится к биотехнологии и представляет собой вектор переноса ДНК, который содержит последовательность, кодирующую p185neu -фрагмент. Данный вектор входит в состав фармацевтической композиции для использования в профилактическом или терапевтическом лечении пациентов с риском развития р185neu-положительных опухолей или пациентов, имеющих первичные опухоли, метастазы или рецидивы p185neu-экспрессирующих опухолей. Изобретение позволяет получать новые более эффективные фармацевтические композиции для профилактики и лечения р185neu-положительных опухолей. 3 н. и 8 з.п. ф-лы, 14 ил., 1 табл.
| AMICI A et | |||
| al | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Gene Ther | |||
| ЩИТОВОЙ ДЛЯ ВОДОЕМОВ ЗАТВОР | 1922 |
|
SU2000A1 |
| CHEN Y et | |||
| al | |||
| DNA vaccines encoding full-length or truncated Neu induce protective immunity against Neu-expressing mammary tumors | |||
| Cancer Res | |||
| Способ и аппарат для получения гидразобензола или его гомологов | 1922 |
|
SU1998A1 |
| ВЫДЕЛЕННАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ ДНК, ВЕКТОР, СПОСОБ ПОЛУЧЕНИЯ ГОМОГЕННОГО БЕЛКА gp350, ГОМОГЕННЫЙ БЕЛОК gp350, ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ ЕВV-СВЯЗАННОГО ЗАБОЛЕВАНИЯ ИЛИ СОСТОЯНИЯ | 1995 |
|
RU2178807C2 |

