Изобретение относится к медицине, а именно к биотехнологии, иммунологии и аллергологии, и касается получения рекомбинантного полипептида для аллерген-специфической иммунотерапии, способного индуцировать блокирующие IgG-антитела к аллергену пыльцы березы и к перекрестным пищевым аллергенам.
Основной механизм в аллерген-специфической иммунотерапии (AIT) - индукция аллерген-специфических антител класса IgG, которые блокируют связывание антител класса IgE с сенсибилизирующим агентом у пациентов [1]. Аллерген-специфические блокирующие IgG-антитела, индуцированные в ходе AIT, уменьшают симптомы аллергии, блокируя перекрестное связывание IgE-антител на тучных клетках и базофилах [2]. К тому же, индуцированные в ходе AIT аллерген-специфические IgG-антитела способны ингибировать IgE-обусловленную презентацию аллергена Т-клетками и, таким образом, уменьшать позднюю стадию аллергической реакции [3]. Наконец, аллерген-специфические IgG-антитела способны связывать аллерген, предотвращая усиление аллерген-специфического IgE-ответа, у пациентов с аллергией при контакте с аллергеном. Предположительно этот механизм, приводит к долгосрочному снижению уровня аллерген-специфических IgE-антител у пациентов, проходящих долговременный курс AIT [4].
Аллергия на пыльцу березы часто встречается у людей на территории России и других северных стран. Она зачастую ассоциирована с оральным аллергическим синдромом (OAS) и возникает при употреблении пищи растительного происхождения, поскольку IgE-антитела, направленные против основного аллергена березы - Bet v 1, так же перекрестно реагируют со структурно родственными аллергенами других растений [5]. В частности, такая кросс-реактивность Bet v 1 имеет место с аллергеном яблока - Mal d 1, что приводит к появлению OAS у пациентов с аллергией на березу при употреблении в пищу яблок. В ходе аллерген-специфичной иммунотерапии с применением экстрактов, содержащих Bet v 1 или рекомбинантных производных аллергена Bet v 1 происходит индукция не только протективных IgG-антител к Bet v 1, но также блокирующих IgG-антител к аллергену яблока Mal d 1 [6]. Поэтому AIT, направленная на терапию аллергии на пыльцу березы, частично защищает от появления OAS, но эта защита не стойкая [7]. Следовательно, существует потребность в разработке вакцин для AIT, способных индуцировать блокирующие IgG-антитела не только к аллергену пыльцы березы Bet v 1, но и к перекрестным пищевым аллергенам.
Разработана фармацевтическая композиция для специфической иммунотерапии аллергий, в инициацию которых вовлечен основной аллерген пыльцы березы Bet v 1, включающая гипоаллергенный основной аллерген пыльцы березы Bet v 1 и/или его фармацевтически приемлемые производные. Гипоаллергенные основные аллергены пыльцы березы отличаются отсутствием или снижением связывания иммуноглобулина Е с одновременным сохранением терапевтически релевантной стимуляции Т-клеток (WO 03/072601 (МЕРК ПАТЕНТ ГМБХ), 04.09.2003).
Известны рекомбинантные аллергены Bet v 1, использование которых позволяет снизить специфическую IgE-связывающую способность по сравнению с IgE-связывающей способностью аллергена природного происхождения. Рекомбинантный аллерген Bet v 1 является мутантом аллергена Bet v 1 природного происхождения, где мутантный аллерген Bet v 1 имеет по меньшей мере четыре мутации, где каждая мутация является заменой одного экспонированного на поверхности аминокислотного остатка другим остатком, который не встречается в том же положении в аминокислотной последовательности любого известного гомологичного белка в пределах таксономического вида (WO 02/40676 (АЛЬК-АБЕЛЛО), 23.05.2002).
Согласно WO 2013/017591 (LOFARMA SPA), 07.02.2013 получен гибридный белок для применения в иммунотерапии пациентов, подверженных аллергии к пыльце Malus domectica и/или Betula verrucosa, содержащий белок, который представляет собой гипоаллергенный мутант главного аллергена Malus domectica, и белок, который представляет собой гипоаллергенный мутант главного аллергена Bet v 1 из пыльцы Betula verrucosa, разделенные линкером. Данная технология создания гипоаллергенных вакцин примечательна тем, что в последовательность ДНК аллергенов вводиться сайт-направленные мутации. В результате чего полипептидная последовательность аллергена также меняется, приводя к снижению аллергенности. Такой подход позволил авторам изобретения снизить IgE-реактивность химерного мутантного белка. Недостатком полученного химерного мутантного белка является его остаточная IgE-реактивность. Кроме того, он содержит в своем составе Т-клеточные эпитопы. т.е. имеется возможность активации Т-клеток, продуцирующих провоспалительные факторы, что в свою очередь снижает безопасность данного белка.
В настоящее время создано новое поколение молекулярных вакцин для AIT, способных связать IgG-антитела с сайтами IgE-антител на аллергенах [8].
Показано, что связывание IgG-антитела с сайтами IgE-антител на аллергенах достигается путем слияния не аллергенных пептидов, полученных из участков IgE-связывания аллергенов с PreS-белком вируса гепатита В, выступающего в качестве носителя. Этот новый тип молекулярных аллерговакцин обладает множеством преимуществ перед традиционными вакцинами на основе экстрактов аллергенов, как по способу производства, безопасности и удобству клинического применения [9]. Технология на основе белка-носителя PreS, так же используется для создания гипоаллергенных вакцин для лечения пациентов с аллергией на пыльцу березы [10]. В частности, один из кандидатов, названный 2PAPB-PreS, обозначаемый также так же B-Pres, при иммунизации кроликов лучше индуцирует протективные IgG-антитела, чем нативный рекомбинантный аллерген rBetv 1 [10].
Известно получение гипоаллергенного белка, который состоит из одной гипоаллергенной молекулы, происходящей из аллергена, слитой со вторым не аллергенным белком, происходящим из патогена. Аллерген выбран из группы, состоящий из основных аллергенов пыльцы березы, в частности Bet v 1 и Bet v 4, основные аллергенов пыльцы тимофеевки луговой, основных аллергенов клеща домашней пыли, основных аллергенов кошек Fel din Fel d 2, основных аллергенов пчел, основных аллергенов ос, аллергенов амброзии (WO 2007/140505 (BIOMAY AG), 13.12.2007).
Наиболее близким аналогом изобретения является вакцинная композиция, предназначенная для лечения или предотвращении аллергии на аллерген пыльцы березы Bet v 1, содержащая эффективное количество аллергена, в частности, аллерген пыльцы березы Bet v 1, и белок-носитель, который является полипептидом PreS вируса гепатита В (WO 2012/168487 (BIOMAY AG), 13.12.2012). Оказалось, что аллерген-специфические IgG антитела, индуцированные иммунизацией подобранными, происходящими от аллергена пептидными фрагментами, соединенными с полипептидом PreS вируса гепатита В, лучше нацеливаются на IgE эпитопы аллергена, в то время как иммунизация аллергеном дикого типа дает в результате IgG, направленный на все части аллергена - также и те, которые не являются IgE-реактивными.
Однако, данная вакцинная композиция не обладает индуцированием блокирующих IgG-антител перекрестным пищевым аллергенам.
Целью, лежащей в основе данного изобретения, является получение вакцины для аллерген-специфической иммунотерапии, способной индуцировать блокирующие IgG-антитела к аллергену пыльцы березы и к перекрестным пищевым аллергенам, в частности, к аллергену яблока Mal d 1.
Решение этой проблемы обеспечено получением рекомбинантного полипептида для лечения или профилактики аллергии на аллерген пыльцы березы и аллерген яблока, содержащего пептидные фрагменты аллергена дикого типа пыльцы березы, и яблока, соединенные с поверхностным полипептидом PreS вируса гепатита В, и имеющий аминокислотную последовательность SEQ ID NO 2.
Рекомбинантный полипептид содержит, по меньшей мере, четыре пептидных фрагмента аллергена дикого типа пыльцы березы Bet v 1, и, по меньшей мере, два пептидных фрагмента аллергена дикого типа яблока Mal d 1.
Аллерген дикого типа выбирают из группы, состоящей из основных аллергенов пыльцы березы, в частности Bet v 1, где два пептидных фрагмента имеют аминокислотную последовательность SEQ ID NO 5 и другие два пептидных фрагмента имеют аминокислотную последовательность SEQ ID NO 6.
Также аллерген дикого типа выбирают из группы, состоящей из основных аллергенов яблока, в частности Mal d 1, имеющих один пептидный фрагмент с аминокислотной последовательностью SEQ ID NO 3 и другой пептидный фрагмент с аминокислотной последовательностью SEQ ID NO 4.
Композиция вакцины для лечения или профилактики аллергии на аллерген пыльцы березы и яблока содержит эффективное количество рекомбинантного полипептида, имеющего аминокислотную последовательность SEQ ID NO 2 и фармацевтически приемлемый наполнитель. Композиция дополнительно содержит по меньшей мере один адъювант и/или консервант.Адъювант представляет собой гидроксид алюминия.
Техническими результатами заявленного изобретения является получение рекомбинантного полипептида AB-PreS (SEQ ID NO 2), содержащего эпитопы, необходимые для активации защитных антител против Bet v 1 и Mal d 1 в одном белке, что позволяет получить однокомпонентную вакцину, вместо того, чтобы использовать вакцины, содержащие два или более производных аллергенов.
У полученного рекомбинантного полипептида AB-PreS, который содержит пептиды из аллергена яблока и березы, слитые с белком PreS, полностью отсутствует IgE-реактивность, что свидетельствует о большей безопасности данного белка. Кроме того, в состав AB-PreS входят пептиды аллергена яблока и березы, имитирующие В-, но не Т-клеточные эпитопы указанных аллергенов. Элиминация Т-клеточных эпитопов из состава AB-PreS позволяет добиться более существенной безопасности конструкта, т.к. исключена возможность активации Т-клеток, продуцирующих провоспалительные факторы.
Согласно настоящему изобретению, полученный рекомбинантный полипептид АВ-PreS (SEQ ID NO 2), кроме пептидных производных от аллергена дикого типа березы Bet v 1 (SEQ ID NO 5 и SEQ ID NO 6) содержит пептидные производные от аллергена яблока Mal d 1 (SEQ ID NO 3 и SEQ ID NO 4), которые находятся между пептидами аллергена березы и поверхностным полипептидом PreS гепатита В (SEQ ID NO 7), что позволяет данной рекомбинантной конструкции вызвать повышенный иммунный ответ против аллергенных молекул, и индуцировать блокирующие IgG-антитела к аллергену пыльцы березы Bet v 1 и к аллергену яблока Mal d 1.
Таким образом, полученный рекомбинантный белок, представленный SEQ ID NO 2, ингибирует IgE-связывание с Mal d 1 значительно более выраженное, чем ингибирование такими же дозами B-PreS полипептида. В итоге вакцина от аллергии на пыльцу березы на основе AB-PreS полипептида индуцирует более высокий уровень проективных антител, блокирующих IgE-связывание с Bet v 1 и Mal d 1 по сравнению с другими протестированными вакцинами.
Краткое описание графических материалов/фигур.
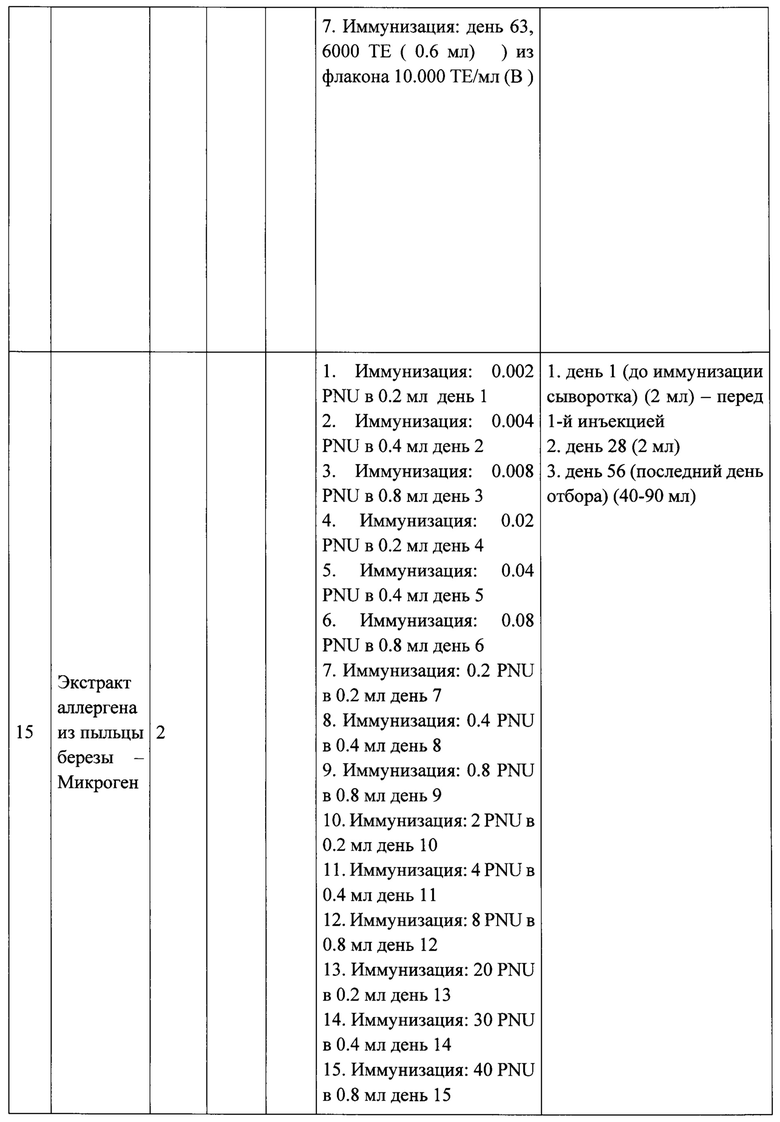
ФИГ. 1. Множественное выравнивание последовательностей аллергена Bet v 1 и сходных аллергенов.
Аминокислотную последовательность Bet v 1 выравнивали совместно с родственными аллергенами из фруктов, овощей и пыльцы. Идентичные аминокислоты обозначены точками, пробелы обозначены как тире, количество аминокислот указано в правой части поля.
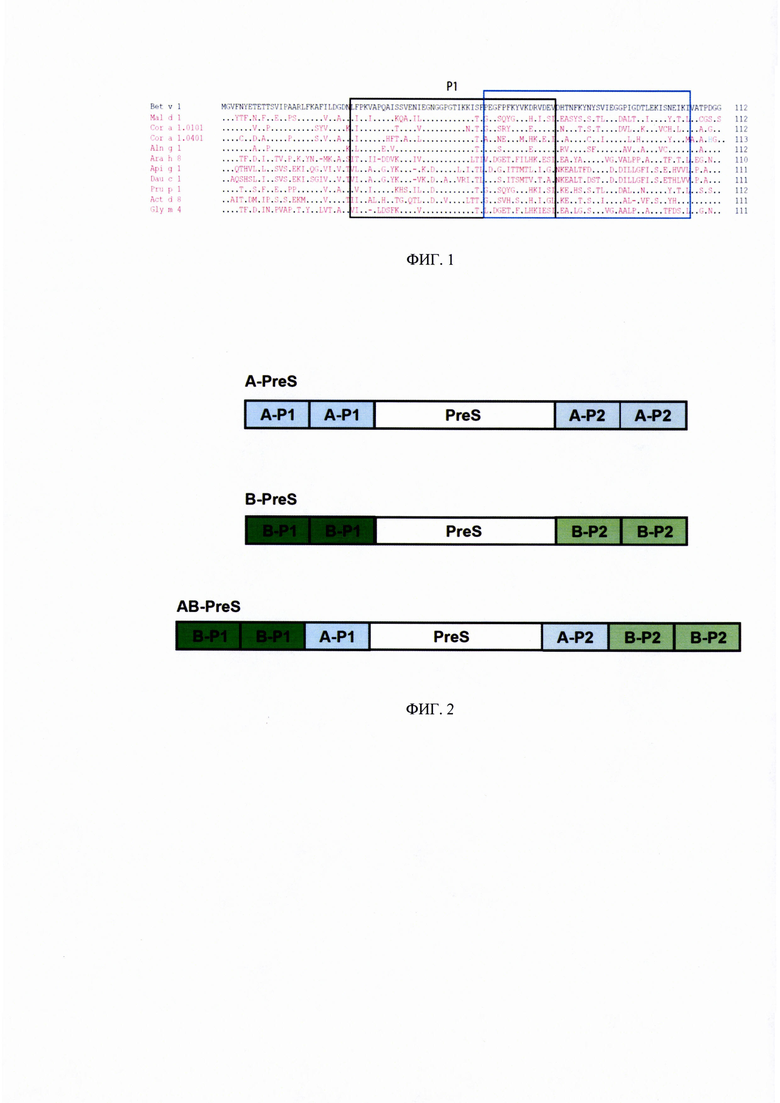
ФИГ. 2. Схематичное изображение полипептидов на основе PreS-белка.
Конструкция A-PreS состоит из двух копий каждого из двух пептидов, производных от Mal d 1 (А-Р1 и А-Р2), объединенных с PreS-белком. Конструкции B-PreS состоит из двух копий каждого из двух пептидов, производных от Bet v 1 (В-Р1 и В-Р2), объединенных с PreS-белком. Конструкция AB-PreS сходна по общей конструкции B-PreS, с каждым из двух пептидов от производных Mal d 1, помещенных между последовательностями PreS-белка и пептидов из аллергена березы.
ФИГ. 3. Последовательность ДНК, кодирующая полипептид AB-PreS и схематическое изображение регионов, кодирующих основные элементы полипептида.
ФИГ. 4. Последовательность аминокислот полипептида AB-PreS и схематическое изображение основных элементов.
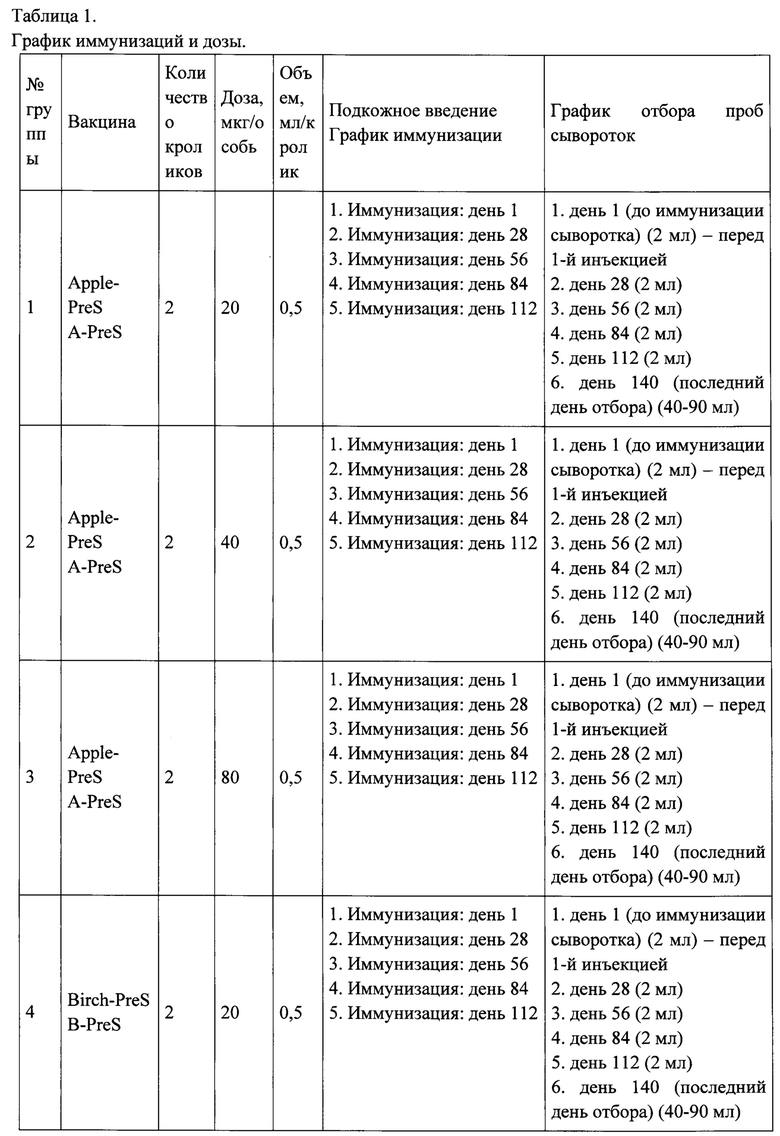
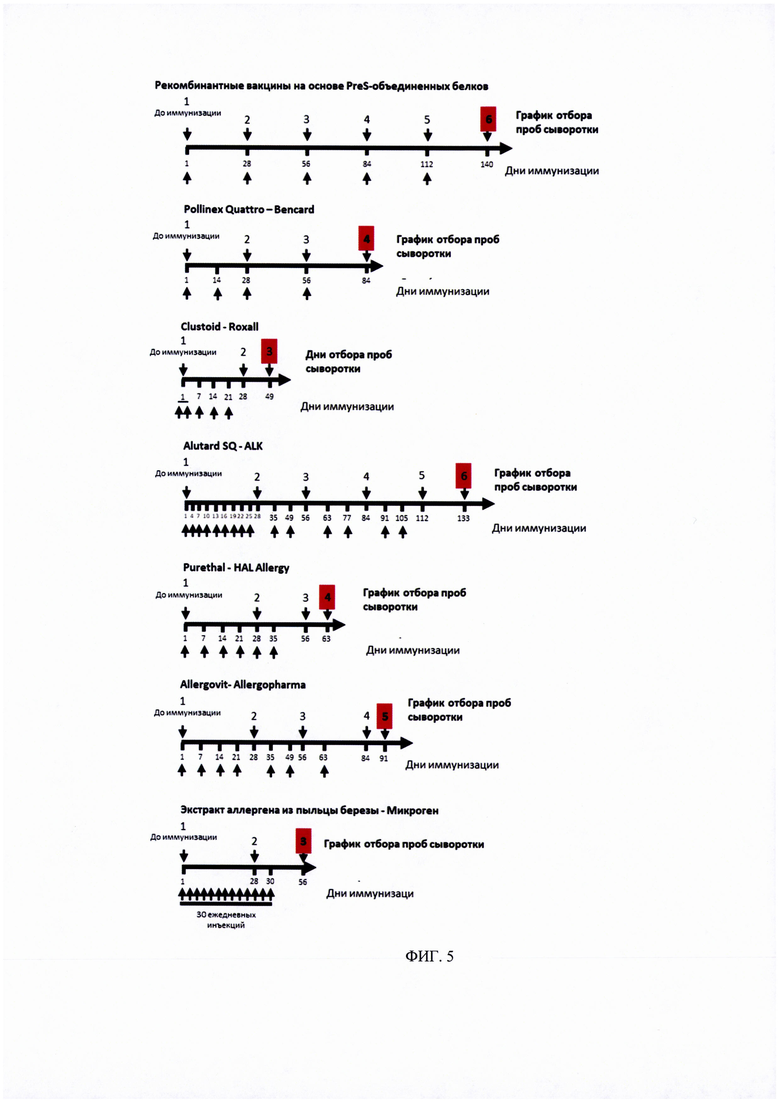
ФИГ. 5. Протокол иммунизации кроликов рекомбинантными вакцинами на основе PreS-белка и коммерческими вакцинами на основе экстрактов аллергенов.
Заключительный отбор образцов сыворотки крови отмечен красным квадратом.
ФИГ. 6. Характеристика рекомбинантных белков.
SDS-PAGE, окрашенные раствором кумасси, на котором видны очищенные PreS-белки (B-PreS, A-PreS и AB-PreS) и маркер молекулярной массы (М).
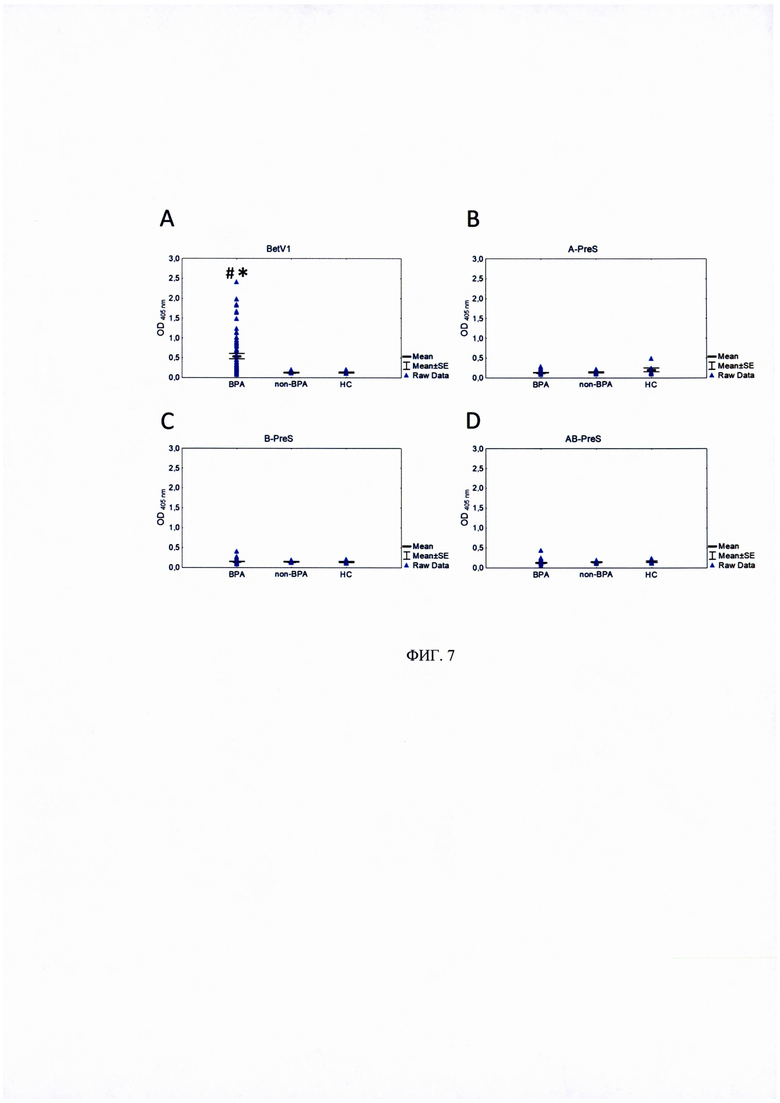
ФИГ. 7. IgE-связывающая способность PreS-белков.
Рекомбинантный Bet v 1 (А) и PreS-белки A-PreS (В), B-Pres (С) и AB-PreS (D) абсорбировались на ИФА-плейте с дальнейшей инкубацией с сыворотками пациентов с аллергией на пыльцу березы (ВРА), без аллергии на пыльцу березы (non-ВРА) и здоровых добровольцев (НС) в качестве контроля.
Данные представлены как среднее значение оптической плотности (OD) при длине волны 405 нм ± стандартная ошибка. Статистический анализ проводился с использованием программы Statistica 8.0 в соответствии с U-критерием Манна-Уитни.
* - статистически значимо отличается от «НС».
# - статистически значимо по сравнению с «non-ВРА».
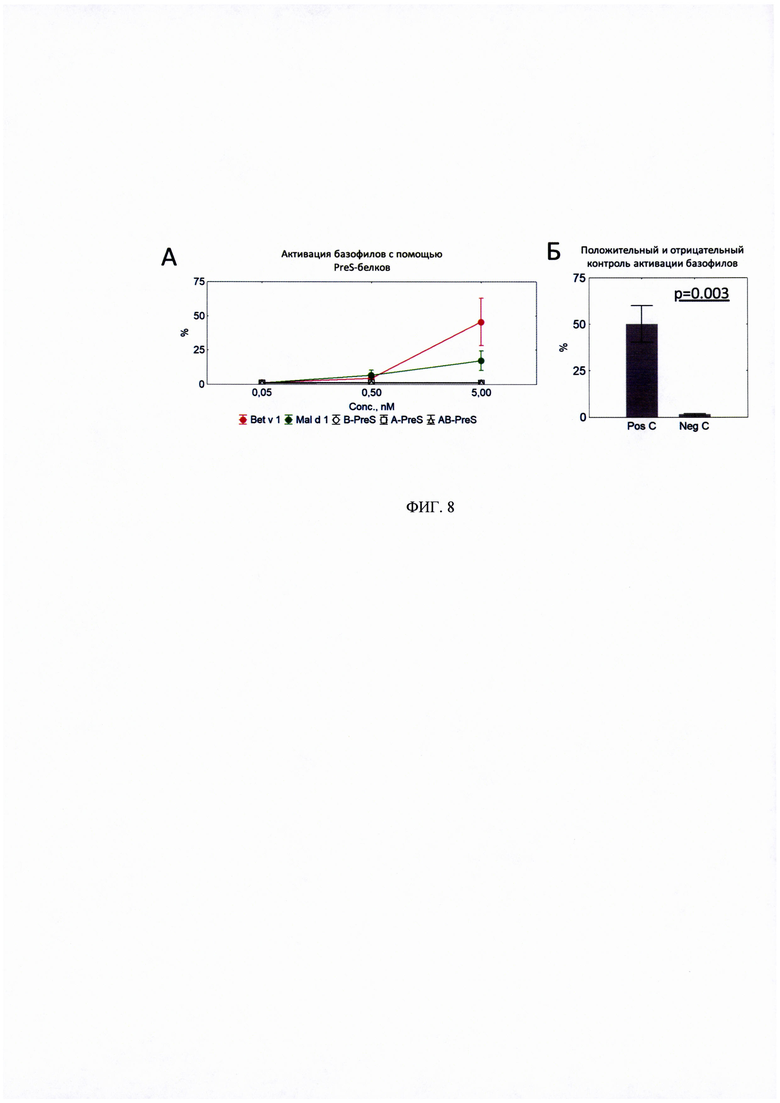
ФИГ. 8. Сравнение A-PreS, B-PreS и AB-PreS с rBet v 1 и rMal d 1 по способности активировать базофилы крови.
Показаны процентные содержания активированных базофилов в крови у четырех пациентов с аллергией на пыльцу березы (В1-В4) после стимуляции различными концентрациями (0,05-5 Нм) Bet v 1 и Mal d 1, эквимолярными концентрациями B-PreS, А-PreS и AB-PreS (А), антитела против IgE использованы в качестве положительного контроля (Pos С), буферный раствор в качестве отрицательного контроля (Neg С) (В).
Данные представлены в виде среднего значения процента активированных базофилов±стандартная ошибка (N=4).
Статистический анализ проводился с использованием программы Statistica 8.0 в соответствии с U-критерием Манна-Уитни.
* - статистически значимо отличается от «Neg С»
× - статистически значимо отличается от «Bet v 1» и «Mal d 1».
ФИГ. 9. Ингибирование связывания IgE-антител пациентов с аллергеном Bet vie использованием сыворотки кроликов, иммунизированных различными дозами рекомбинантных вакцин на основе PreS-белка.
Процент ингибирования IgE-связывания с rBet v 1 определяли для сывороток от 10 пациентов с аллергией на пыльцу березы.
Предварительная инкубация Bet vie PBS была принята за 0% ингибирования. Данные отображены в виде диаграмм, для которых 50% значений находятся внутри прямоугольной фигуры; горизонтальные линии обозначают медианные значения. Показаны существенные различия в ингибировании, рассчитанные с использованием t-критерия Стьюдента.
× - статистически значимо по сравнению с сывороткой не иммунизированных кроликов «Normal».
* - статистически значимо по сравнению с сывороткой от кроликов, иммунизированных A-PreS.
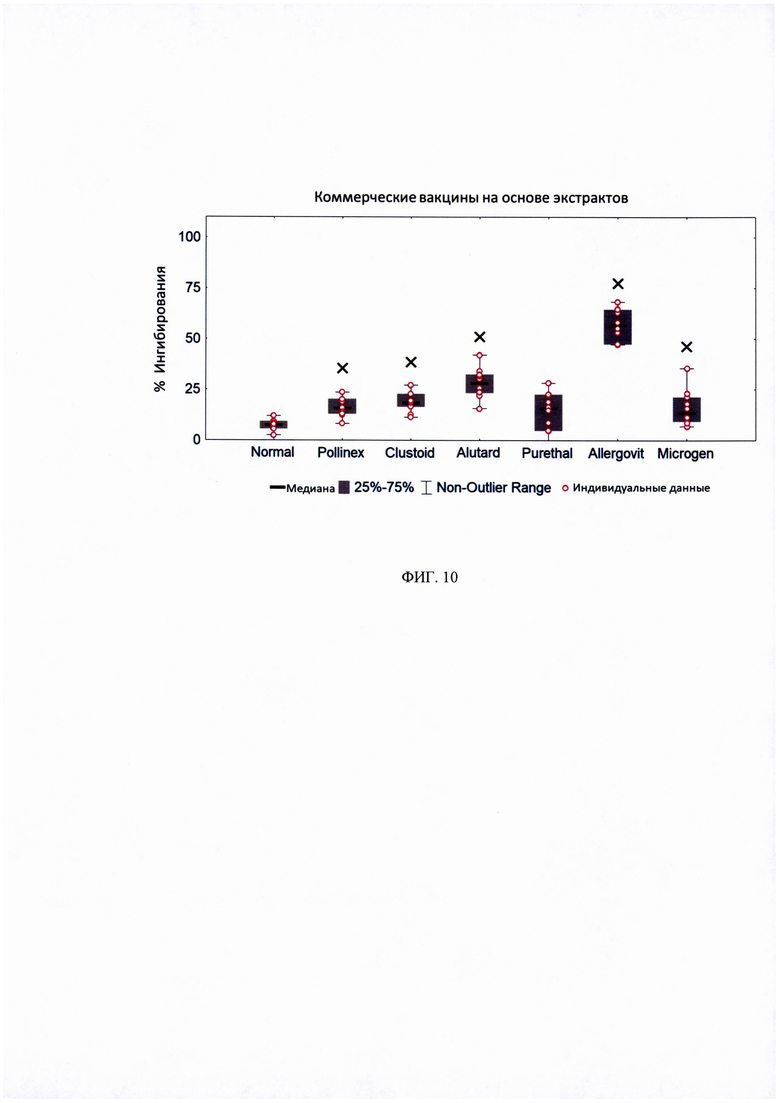
ФИГ. 10. Ингибирование IgE-связывания с Bet v 1 сыворотками кроликов, иммунизированных коммерческими вакцинами на основе экстрактов аллергенов, через один месяц после последней иммунизации.
Процент ингибирования IgE-связывания с rBet v 1 определяли для сывороток от 10 пациентов с аллергией на пыльцу березы. Предварительная инкубация Bet vie PBS принята за 0% ингибирования. Данные отображены в виде диаграмм, для которых 50% значений находятся внутри прямоугольной фигуры; горизонтальные линии обозначают медианные значения. Показаны существенные различия в ингибировании, рассчитанные с использованием t-критерия Стьюдента.
× - статистически значимо по сравнению с сывороткой не иммунизированных кроликов «Normal».
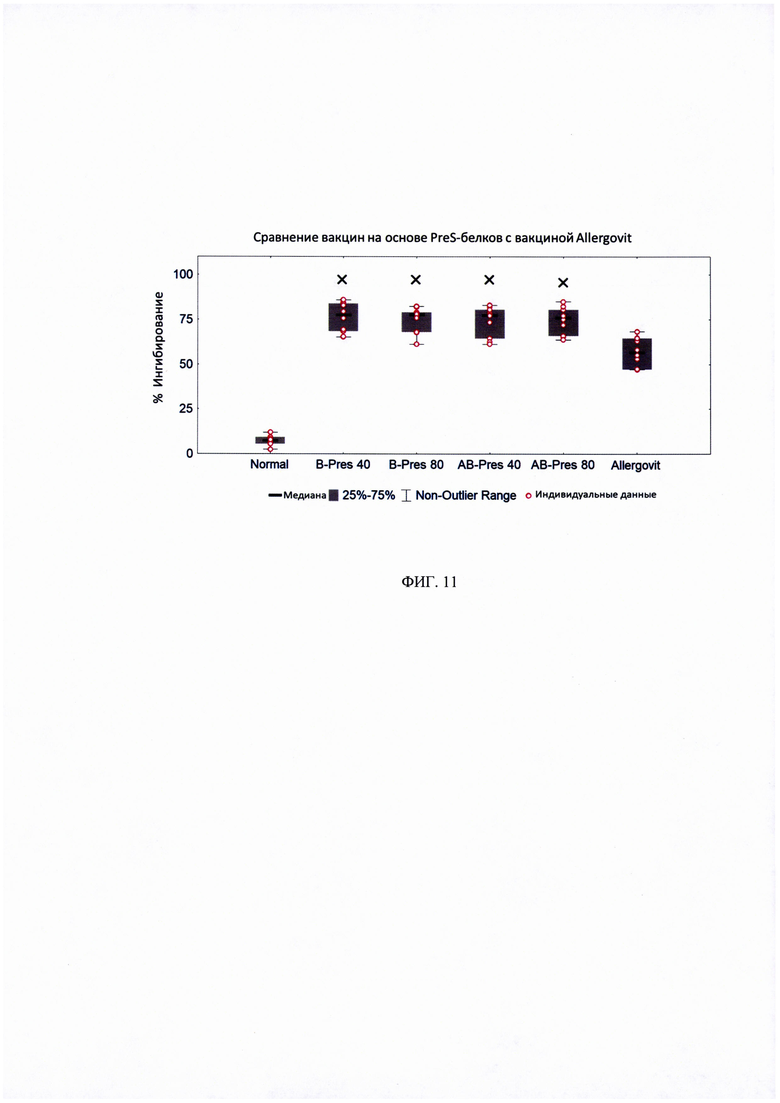
ФИГ. 11. Ингибирование связывания IgE-антител пациентов с Bet vie использованием сывороток кроли кроликов, иммунизированных рекомбинантными вакцинами на основе PreS-белка, по сравнению с Allergovit.
Процент ингибирования IgE-связывания с rBet v 1 определяли для сывороток от 10 пациентов с аллергией на пыльцу березы. Предварительная инкубация Bet vie PBS принята за 0% ингибирования. Данные отображены в виде диаграмм, для которых 50% значений находятся внутри прямоугольной фигуры; горизонтальные линии обозначают медианные значения. Показаны существенные различия в ингибировании, рассчитанные с использованием t-критерия Стьюдента.
× - статистически значимо по сравнению с сывороткой кроликов, иммунизированных кроликов «Allergovit».
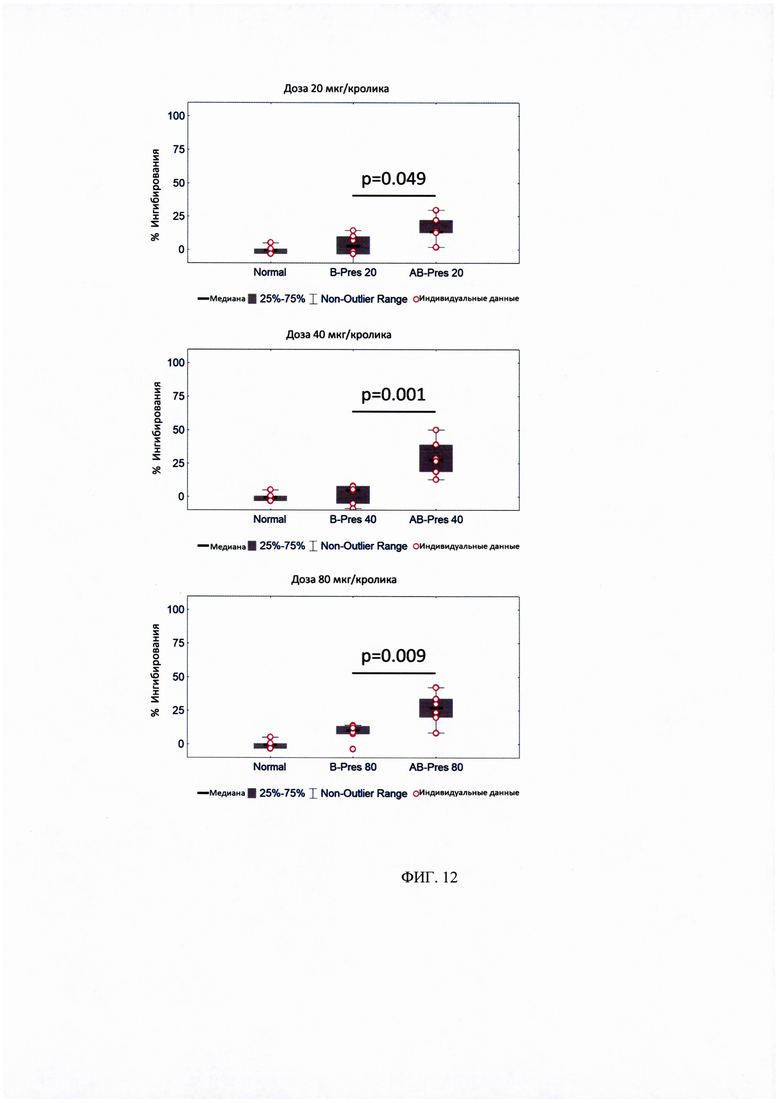
ФИГ. 12. Ингибирование IgE-связывания с аллергеном Mal die использованием сывороток кроликов, получавших курс AIT различными дозами PreS-белков.
Процент ингибирования IgE-связывания с rMal d 1 определяли для сывороток от 10 пациентов с аллергией на пыльцу березы. Предварительная инкубация Bet vie PBS принята за 0% ингибирования. Данные отображены в виде диаграмм, для которых 50% значений находятся внутри прямоугольной фигуры; горизонтальные линии обозначают медианные значения. Показаны существенные различия в ингибировании, рассчитанные с использованием t-критерия Стьюдента.
× - статистически значимо по сравнению с сывороткой кроликов, иммунизированных кроликов «Allergovit».
Подробное описание изобретения
Чтобы исследовать, может ли включение в вакцину B-PreS пептидов из аллергена Mal d 1 иметь преимуществе в части индукции протективных IgG-антител к аллергену яблока дополнительно разработано две вакцины на основе PreS-белка. Вакцина, содержащая пептиды, производные от Mal d 1, (A-PreS) и пептиды производные от Bet v 1 и Mal d 1 (АВ-PreS), и сравнили их действие с B-PreS и несколькими другими коммерческими вакцинами на основе экстрактов аллергена пыльцы березы. При этом оценивали способность вакцин индуцировать антитела классе IgG, ингибирующих связывание IgE-антител пациентов с аллергенами Bet v 1 и Mal d 1.
ПРИМЕР 1. Проектирование вакцинных конструкций.
На рисунке 1 изображено множественное выравнивание последовательности Bet vie родственными PR10-аллергенами из пыльцы, фруктов, овощей, в которых можно выделить два пептида размером 45 аминокислотных остатков (P1, Р2). Эти два пептида представляют собой производные от Bet v 1 из ранее описанной рекомбинантной вакцины, созданной на основе PreS-белка, которая способна индуцировать IgG-антитела, блокирующие IgE пациентов с аллергией на пыльцу березы эффективнее в сравнении с рекомбинантным аллергеном Bet v 1 дикого типа [10].
Были спроектированы три рекомбинантных конструкции на основе PreS-белка. В-PreS (Рисунок 2) был идентичен ранее описанной конструкции 2PAPB-PreS, которая, как было обнаружено, активирует антитела IgG, способные блокировать IgE пациентов с аллергией на пыльцу березы, эффективнее чем рекомбинантный Bet v 1 дикого типа [10]. Так же, были подготовлены две дополнительные конструкции, одна из которых напоминает конструкцию B-PreS, но вместо пептидных производных от Bet v 1, в нее были добавлены пептиды из Mal d 1 (Рисунок 2). Третья конструкция AB-PreS кроме пептидных производных от Bet v 1 содержит вставленный в B-PreS копию двух пептидных производных от Mal d 1, А-Р1 и А-Р2, которые находятся между пептидами аллергена березы и PreS последовательностью (Рисунок 2).
Конструкции для экспрессии рекомбинантных антигенов были сделаны следующим образом: ДНК, кодирующие His-меченые фьюжн-белки (рисунок 3) состоящие из PreS-белка, (SEQ ID NO 7) объединенного с пептидами, производными Bet v 1 или Mal d 1, или объединенные с пептидами производных Bet v 1 или Mal d 1 (Mal d 1: А-Р1(30-74): LIPKIAPQAIKQAEILEGNGGPGTIKKITFGEGSQYGYVKHRIDS (SEQ ID NO 3), A-P2 (60-104): GEGSQYGYVKHRIDSIDEASYSYSYTLIEGDALTDTIEKISYETK (SEQ ID NO 4) Bet v 1: B-P1 (aa 30-74): LFPKVAPQAISSVENIEGNGGPGTIKKISFPEGFPFKYVKDRVDE (SEQ ID NO 5), B-P2 (aa 60-104): PEGFPFKYVKDRVDEVDHTNFKYNYSVIEGGPIGDTLEKISNEIK (SEQ ID NO 6)) были оптимизированы для экспрессии в Escherichia coli, синтезированы (конструкция Mal d 1 -PreS / A-P2-PreS; конструкция Bet v 1 / конструкция Mal d 1-PreS: 2B-P1 / B-P2+1A-P1 / A-P2-PreS - PreS) (ATG:biosynthetics GmBH, Merzhausen, Германия) (Рисунок 2) и вставлены по сайтам рестрикции Ndel / Xhol плазмиды pET-27b (Novagen, Darmstadt, Германия). Правильность последовательности ДНК подтверждали секвенированием обеих цепей рекомбинантной плазмиды (ATG:biosynthetics GmBH, Merzhausen, Германия).
Разработан дизайн рекомбинантных слитых PreS-белков, содержащих не аллергенные пептиды из участков связывания IgE основного аллергена пыльцы березы Bet v 1 и основного аллергена яблока Mal d 1.
На рисунке 1 было показано выравнивание пептидных последовательностей Bet v 1, использованных для создания вакцин, с гомологичными PR10-аллергенами из растительной пищи и пыльцы. Пептиды Р1 и Р2, производные от Bet v 1, были идентичны ранее описанной конструкции 2PAPB-PreS, которая, как было установлено, активирует антитела IgG, блокирующие IgE пациентов с аллергией на пыльцу березы эффективнее, чем рекомбинантный Bet v 1 дикого типа [10]. Однако, эта вакцина не сравнивалась с вакцинами на основе экстрактов аллергенов в отношении способности индуцировать IgG-антитела, блокирующих связывание IgE пациентов с аллергией к аллергену Bet v 1 и Mal d 1, последний из который является одним из наиболее важных перекрестно-реактивных пищевых аллергенов для пациентов с аллергией на пыльцу березы. Таким образом, A-PreS был разработан аналогично B-PreS на основе гомологичных пептидов А-Р1 и А-Р2 (рисунок 2). В химерную конструкцию AB-PreS были включены пептиды из аллергена березы и яблока А-Р1 и А-Р2 (конечный белковый продукт имел прогнозируемый размер около 50 кДа). Белковая последовательность AB-PreS представлена на рисунке 4.
Таким образом, в состав AB-PreS входят пептиды аллергена яблока и березы, имитирующие В-, но не Т-клеточные эпитопы указанных аллергенов.
ПРИМЕР 2. Экспрессия, очистка и биохимическая характеристика рекомбинантных полипептидов.
Рекомбинантные PreS-фьюжн-белки, A-PreS, B-PreS и AB-PreS экспрессировали в штамме Е. Coli BL21 (DE3) (Strategene, La Jolla, СА). Трансформированные клетки наращивали в жидкой среде Луция-Бертани (Luria-Bertani), в состав которой входило 25 мг/мл канамицина до оптической плотности OD=0.6-0.8. Индукцию экспрессии белка осуществляли путем добавления изопропил-β-О-тиогалактопирозид до конечной концентрации 1 ммоль/л, клетки культивировали в течении 3 часов при температуре 37 С, после центрифугировали при 4000 об/мин в течении 20 минут.
Получившиеся бактериальные осадки были ресуспендированны в 8 М мочевине, 10 Мм NaH2PO4 и 10 Мм Трис (8 Рн), содержащей в себе смесь ингибиторов протеаз (Sigma, Aldrich) и 5 мкг/мл ДНКазы I (Sigma, Aldrich), после гомогенизированы в Ultra Тurrах 3 раза по 5 минут (IKA, Stauffen, Германия). Осадок гомогенизированных клеток лизировали путем перемешивания в течение 2 часов при 4С. Лизаты клеток очищали центрифугированием при 14000 об/мин в течение 15 минут при 4С, после чего рекомбинантные белки очищали с помощью аффинной хроматографии с никелем (Qiagen, Hilden, Германия). Очищенные фьюжн-белки элюировали с помощью 8 М мочевины, 100Мм NaH2PO4 и 10 Мм Трис (Рн 4.5), диализовали 10 Мм NaH2PO4, для A-PreS Ph 4.8 Ph 7.8, AB-PreS Ph и для B-PreS Ph 7.4, после чего концентрировали ультрафильтрацией (Amicon Ultra 15; Merck KgaA, Darmstadt, Германия) до значений 0.75-10 мг/мл. Значение определяли с помощью набора ВСА Protein Assay Kit (Thermo Scientific, IL).
На рисунке 6, показан окрашенный красителем Кумасси синим SDS-PAGE, который содержит очищенные рекомбинантные белки B-PreS, A-PreS и AB-PreS. Белки хорошо очищены примерно на 95%, содержатся в значительном для детекции количестве, примерно 10 мг/л культуры, без каких-либо признаков распада. Очистка проводилась по протоколу, описанному в разделе методы. Наблюдаемая молекулярная масса для трех белков в SDS-PAGE соответствовала прогнозируемой, которая была рассчитана в соответствии с их аминокислотной последовательностью и наблюдаемыми массами, которые мы определяли с помощью масс-спектрометрии (Рисунок 6). Анализ методом кругового дихроизма (КД) показал, что белки не были свернуты.
ПРИМЕР 3. Получение вакцинной композиции.
Каждый из рекомбинантных очищенных белков был растворен в изотоническом буфере, содержащем 0,9% хлорид натрия и 10 мМ фосфат натрия, и к каждому раствору белка было добавлено соответствующее количество гидроксида алюминия. Была приготовлена смесь, содержащая равные части четырех полученных суспензий, и поделена на аликвоты в стерильных условиях в запаянные флаконы. Инъекционная композиция, полученная с помощью этого метода, содержала 0,16 мг/мл каждого A-PreS, B-PreS и АВ-PreS.
Состав вакцин с адсорбированным гидроксидом алюминия, содержащих A-PreS, В-PreS и AB-PreS, выполняли, как описано в публикации [11]. Состав вакцинной композиции на 1 мл стерильной воды:
1. AB-PreS - 0,16 мг.
2. А1(ОН)3 - 1,25 мг.
3. NaH2PO4 - 1,2 мг (конечная концентрация 10 мМ).
4. NaCl - 9 мг (конечная концентрация 0,9%).
Данный состав необходим для достижения максимальной дозы 80 мкг AB-PreS на особь в объеме 0,5 мл/особь. Остальные дозы готовились путем разбавления исходной максимальной дозы соответствующим объемом буфера (10 мМ NaH2PO4+0,9% NaCl).
ПРИМЕР 4. Иммунологическая характеристика рекомбинантных белковых вакцин in vitro.
Для оценки способности белков слитых с PreS взаимодействовать с аллерген-специфическими IgE использовали тест ИФА. Планшеты покрывали рекомбинантными аллергенами rBet vl и rMal dl или PreS-гибридными белками (A-PreS, B-PreS или AB-PreS) в концентрации 2 мкг/мл при 4С в течение ночи. Затем промывали и блокировали в течение ночи с помощью 2% раствора BSA. После чего планшет инкубировали в течение ночи при 4С с разведенными 1:10 сыворотками пациентов с аллергией на пыльцу березы (группа «ВРА»), пациентов, не страдающих аллергией на пыльцу березы (группа «без ВРА»), или здоровых добровольцев (группа «НС»). Связавшиеся IgE-антитела человека детектировали с помощью АР-меченных козьих антител против человеческого IgE (Invitrogen, USA) в разведении 1:2500. После стадии промывки вносили раствор субстрат ABTS (10 мг/мл). Оптическую плотность измеряли при 405 нм.
4.1. Оценка гипоаллергенных свойств рекомбинантных белков B-PreS, A-PreS и AB-PreS.
Возможная аллергенная активность трех рекомбинантных PreS-белков была исследована в экспериментах по связыванию IgE и в экспериментах по активации базофилов. В первую серию экспериментов было включено 84 добровольца (таблица 2). IgE-связывающая способность B-PreS, A-PreS и AB-PreS сравнивалась с таковой для rBet v 1 с помощью теста ИФА с использованием сывороток от 70 пациентов с аллергией на пыльцу березы (рисунок 7). Мы обнаружили, что ни один из трех рекомбинантных белков не продемонстрировал какой-либо реактивности с IgE-антителами из сывороток пациентов с аллергией на пыльцу березы. В то же время сыворотки этих пациентов значительно реагировали с нативным аллергеном rBet v 1. Также для B-PreS, A-PreS и AB-PreS не выявлено IgE-реактивности с сыворотками пациентов с аллергией на другие аллергены (не пыльцу березы) (n=7) и здоровых добровольцев (n=7) (рисунок 7).
4.2. Тест на активацию базофилов крови (ВАТ-тест).
Процентное содержание активных базофилов в образцах цельной крови оценивали с помощью набора для определения аллергенности (ВАТ-тест) (Beckman Coulter, Fullerton, СА, USA) в соответствии с инструкцией производителя. ВАТ-тест выполняли с использованием двух очищенных рекомбинантных аллергенов Bet v 1 и Mal d 1 в трех концентрациях для каждого аллергена: 1 нг / мл, 10 нг/мл, 100 нг/мл и с эквимолярными концентрациями белковых конструкций B-PreS и A-PreS (2,1 нг/мл, 21 нг/мл, 210 нг/мл) и AB-PreS (2,7 нг/мл, 26,6 нг/мл, 266 нг/мл), что соответствует молярной концентрации 0,05, 0,5 и 5 Нм. Положительным контролем были анти-IgE-антитела (0,01 мг / мл), а отрицательным контролем (растворитель PBS). Контроли готовили для каждого пациента.
Образцы крови собирали в полипропиленовые пробирки, содержащие КЗ EDTA (S-Monovette, Sarstedt, Германия), и использовали для ВАТ-теста сразу после сбора. Использовали 100 мкл цельной крови, 20 мкл тестируемого аллергена / контроля и 20 мкл трехцветных антител (CRTH2-FITC, CD203c-PE, CD3-PC7) и 100 мкл раствора для активации. Смешивали указанные компоненты в пластиковых пробирках и инкубировали в течение 15 минут при 37°С на водяной бане. После инкубации, реакцию останавливали добавлением 100 мкл стоп-раствора и 2 мл раствора для фиксации и лизиса. После остановки реакции образцы инкубировали при комнатной температуре, в темном месте, в течение 10 минут и центрифугировали 5 минут при 200 g. После удаления супернатанта добавляли по 3 мл PBS в каждую пробирку и центрифугировали образцы в течение 5 мин при 200 g. Эритроциты лизировали, а лейкоциты фиксировали добавлением 0,5 мл 0,1% раствора формальдегида в PBS.
Проточную цитометрию проводили на приборе BD FACS Canto II (BD Biosciences, США). Компенсация была установлена в BD FACSDiva Software (BD Biosciences). Результаты анализировали с помощью программного обеспечения Flow Jo ™ версии 10.6.2 (Tree Star). Результаты представлены в виде процентного содержания активированных базофилов. Активированные базофилы были идентифицированы как CRTH2 pos CD203c pos CD3 neg. Изотипические контроли (крысиный IgG2a, изотипический контроль каппа (Ebr2a), FITC, eBioscience ™, мышиный IgG1, (РЕ-Су7), MG112, Thermofisher) отрицательный (PBS) и положительный контроль (антитела анти-IgE) использовали для гейтирования активированных базофилов.
Затем, мы провели эксперименты с участием четырех пациентов с аллергией на пыльцу березы в тесте активации базофилов крови. Полученные данные показали, что активация базофилов с помощью rBet v 1 в концентрации 5 Мм была выраженной, тогда как ни один из трех рекомбинантных PreS-белков не индуцировал активацию этих клеток (Рисунок 8А). rMal d 1 также индуцировал активацию базофилов в концентрации 5 Мм у двух пациентов с аллергией на пыльцу березы (B1, В4) (Рисунок 8А). Так же были проведены эксперименты по активации базофилов положительным контролем (антителами против IgE), при этом не было обнаружено активации базофилов при использовании только буферного раствора (отрицательный контроль) (Рисунок 8 В).
ПРИМЕР 5. Ингибирование связывания IgE-антител пациентов с рекомбинантными аллергенами rBet v 1 и rMal die использованием антител кроликов.
Животные
Сорок кроликов (линия Новозеландские белые кролики, самки) массой 2,2-2,5 кг, приобретены в компании «КролИнфо» (Россия). Перед первой иммунизацией у каждого кролика собирали образцы сыворотки объемом 0,5-1,0 мл и анализировали с помощью метода ИФА, для определения фонового уровня IgG-антител против Bet v 1 и Mal d 1. В исследование включали кроликов с низким естественным фоновым уровнем этих антител.
Протокол вакцинации
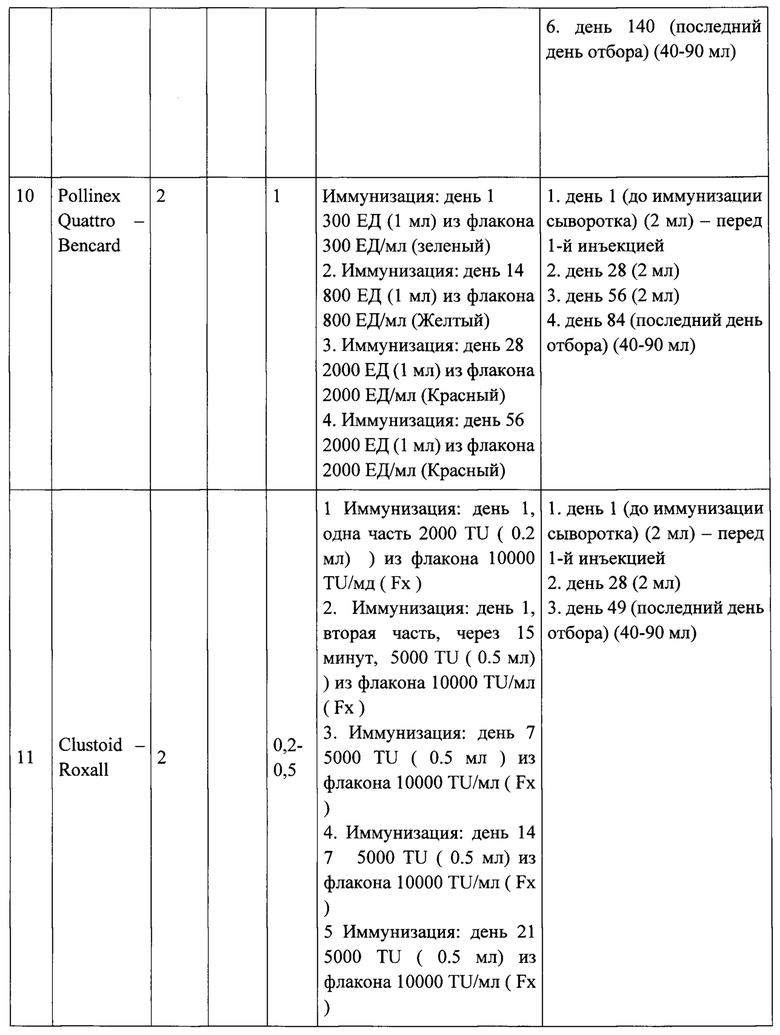
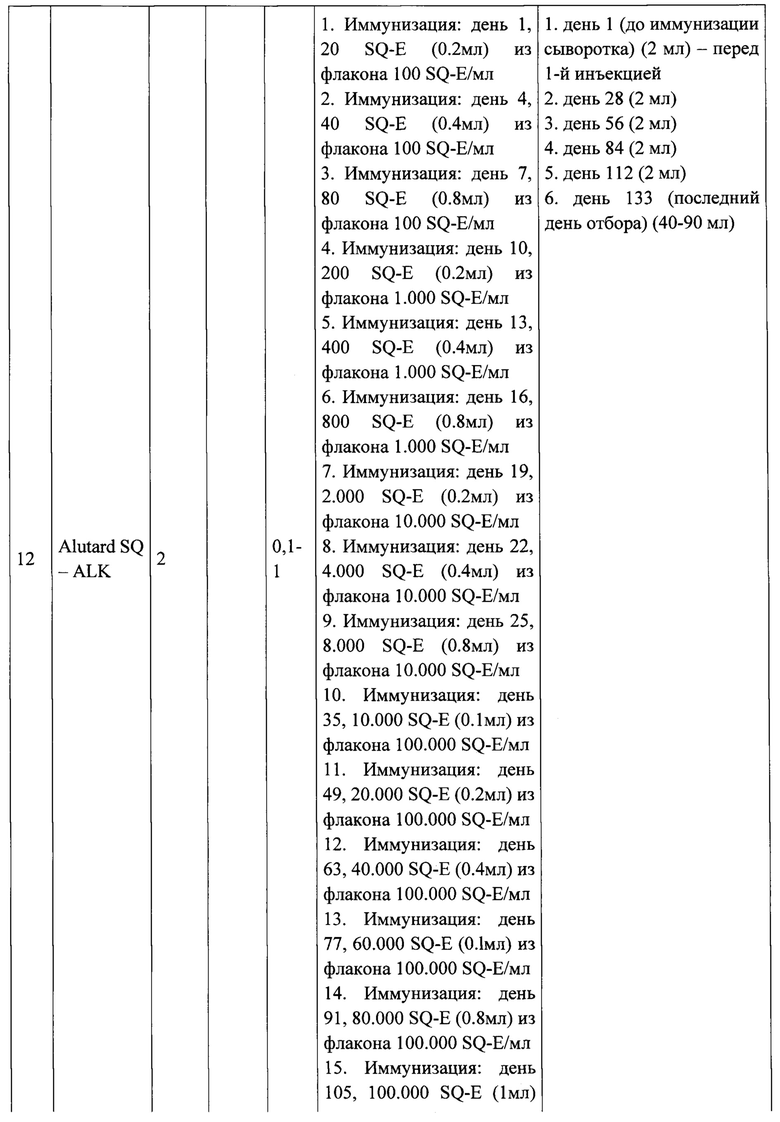
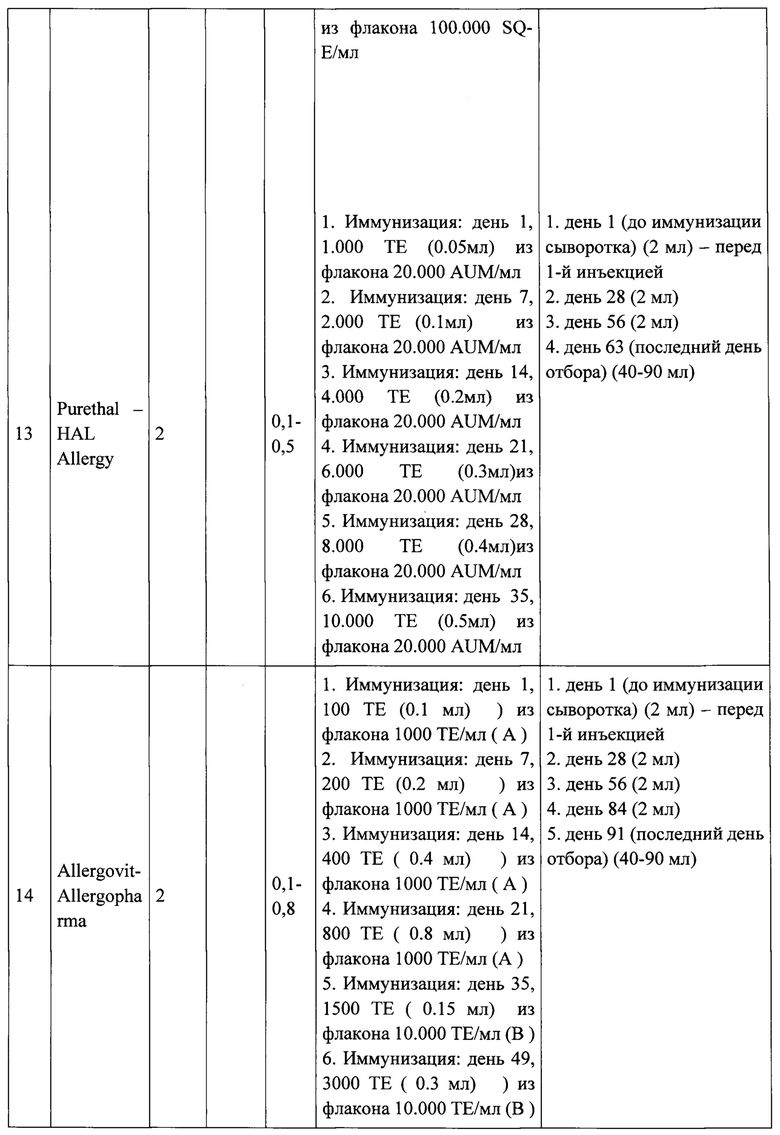
Тридцать кроликов с низким уровнем IgG-антител к Bet v 1 и Mal d 1 были включены в исследование после рандомизированы на 15 групп по 2 кролика в каждой. Каждую группу кроликов индивидуально иммунизировали 3 рекомбинантными вакцинами (A-PreS, B-PreS, AB-PreS) в трех дозах каждая и 6 другими коммерческими вакцинами на основе экстрактов аллергенов:
1. Pollinex Quattro - Bencard
2. Clustoid - Roxall
3. AlutardSQ - ALK
4. Purethal - HAL Allergy
5. Allergovit - Allergopharma
6. Экстракт аллергена из пыльцы березы - Birch Pollen Allergen Extract (Микроген), Россия.
Иммунизация коммерческими вакцинами на основе экстрактов аллергенов проводилась в соответствии рекомендациями производителей для пациентов с аллергией. Порядок инъекций и время отбора образцов крови показаны на Рисунке 5 и в Таблице 1, также в них предоставлена дополнительная информация о введенных дозах и количестве взятой крови.
Рекомбинантные вакцины получали путем адсорбции трех доз (20,40, 80 мкг каждого из рекомбинантных белков (Apple-Pres: A-Pres; Birch-Pres: B-Pres; Birch / Apple-Pres: AB-Pres) на гидроксиде алюминия, как описано для рекомбинантной вакцины против аллергии на пыльцу трав ВМ32, созданной также на основе PreS-белка [11]. Каждому кролику делали 5 подкожных инъекций, общим объемом введения 0.5 мл, с 4-недельными интервалами времени (Таблица 1, Рисунок 5).
Выполняли отбор образцов сыворотки крови (примерно 2 мл от каждого кролика) в день первой иммунизации и с 4-недельными интервалами, как указано в таблице 1. Сыворотки хранили при -20°С до анализа.
Ингибирование связывания IgE пациентов с аллергией на аллерген пыльцы березы rBet v 1 и яблока rMal d 1 определяли с помощью конкурентного ингибирования ИФА. Планшеты сорбировали rBet v 1 или rMal die концентрации 1 мкг/мл и инкубировали ночь при 4°С. После промывки и блокирования в течение ночи 2% BSA, планшеты инкубировали в течение 2 часов при 37°С после чего в течение 1 часа при 4°С с сывороткой кроликов, получавших курсы AIT, в разведении 1:10. Для контроля использовали сыворотку от не иммунизированных кроликов и сравнивали ее с сывороткой, полученной от кроликов через 1 месяц после окончания курса AIT. Сыворотку кроликов предварительно инкубировали в течение 1 ч при 56°С, чтобы инактивировать IgE, но сохранить другие изотипы антител, в частности IgG-антитела. Так же использовали дополнительный отрицательный контроль с помощью фосфатно-солевого буфера (PBS), который вносили в лунки планшета вместо сыворотки кроликов. После стадии отмывки, добавляли сыворотку пациентов с аллергией на пыльцу березы в разведении 1:5 и инкубировали ночь при 4°С, затем детектировали связавшиеся IgE-антитела человека с помощью меченых козьих антител против человеческого IgE, конъюгированных с HRP (Invitrogen, США) в разведении 1:2000. Ингибирование связывания IgE с аллергенами rBet v 1 или rMal d 1 после предварительной инкубации с сывороткой иммунизированных кроликов в процентном отношении рассчитывали для каждой сыворотки пациента следующим образом:
процент ингибирования = 100 - (Odi/OdPBSxl00),
где Odi - оптическая плотность после предварительной инкубации с сывороткой от иммунизированных кроликов, ODPBS - оптическая плотность после предварительной инкубации с PBS.
Т-критерий Стьюдента (STATISTICA 8.0 StatSoft Inc) использовался для сравнения степени ингибирования после различных курсов иммунизации. Значение р≤0,05 считалось статистически значимым.
ПРИМЕР 6. Определение ингибирования связывания IgE с Bet v 1 при иммунизации B-Pres и AB-PreS.
Пример 6.1. Сравнительный анализ ингибирования связывания IgE с Bet v 1 при иммунизации AB-PreS, B-Pres и A-PreS.
Степень ингибирования связывания IgE-антител пациентов с аллергеном Bet v 1 сыворотками кроликов, получивших курсы AIT рекомбинантными вакцинами B-PreS, А-PreS и AB-PreS (в трех дозах 20 мкг, 40 мкг или 80 мкг) по сравнению с сывороткой не иммунизированных кроликов и кроликов показаны на рисунке 10. Среднее ингибирование IgE-связывания с Bet v 1 с помощью антител кроликов, получавших A-Pres, всегда было менее 50%, тогда как сыворотка кроликов, иммунизированных B-PreS- и AB-PreS приводила к ингибированию приблизительно на 75%, более того, все три дозы значительно лучше ингибировали IgE-связывание, чем A-PreS (Рисунок 9). Не было выявлено значимых отличий в степени ингибирования IgE-связывания сыворотками кроликов, получавших курсы AIT вакцинами B-PreS- и AB-PreS (Рисунок 9).
Таким образом, иммунизация B-Pres и AB-PreS индуцирует антитела, которые, ингибируют связывание IgE с Bet v 1 значительно сильнее, чем антитела, индуцированные A-PreS.
Пример 6.2. Сравнительный анализ связывания IgE с Bet v 1 при иммунизации AB-PreS, B-Pres и коммерческих вакцин на основе экстрактов аллергенов березы.
На рисунке 10 представлены данные, демонстрирующие, что антитела, индуцированные у кроликов всеми вакцинами (кроме Purethal), значительно ингибировали связывание IgE-антител пациентов с аллергеном Bet v 1. Самая высокая степень ингибирования IgE-связывания была получена с антителами кроликов, индуцированными вакциной на основе экстрактов аллергенов Allergovit.
Стоит отметить, что рекомбинантные вакцины B-PreS и AB-PreS индуцируют более значимый протективный эффект в сравнении с коммерческой Allergovit (рисунок 10). Таким образом, иммунотерапевтические вакцины на основе B-PreS и AB-PreS превосходят все существующие коммерческие вакцины на основе экстрактов аллергенов в отношении активации антител, блокирующих связывание IgE-антител пациентов с аллергеном Bet v 1.
Для большинства коммерческих вакцин, на основе экстрактов аллергенов было проведено большее количество инъекций, чем для рекомбинантных вакцин на основе PreS-белка (Allergovit: 7 инъекций; Alutard: 15 инъекций; экстракт аллергена из пыльцы березы (Микроген): 30 инъекций; Purethal: 7 инъекций) (Рисунок 5, Таблица 1). Только для вакцин Pollinex и Clustoid, было сделано сопоставимое количество инъекций, четыре и пять, соответственно (рисунок 5, таблица 1). Однако, антитела активируемые двумя последними вакцинами, только в незначительной степени ингибировали связывание IgE-антител пациентов с аллергенов пыльцы березы Bet v 1 (рисунок 10).
Что касается режима введения тестируемых коммерческих вакцин, необходимо иметь в виду, что вакцины Pollinex, Clustoid, Purethal и экстракт аллергена из пыльцы березы компании Микроген должны рассматриваться, как способ предсезонного лечения, что означает необходимость каждый год делать одинаковое количество инъекций перед сезоном цветения березы. В итоге, для полного курса лечения вакциной Allergovit потребуется ввести 14 инъекций в течение 2 лет.Вакцину Alutard также можно вводить каждый год, в качестве предсезонного лечения, которое сводится к 30 инъекциям в течение двухлетнего периода лечения. Кроме того, существует возможность осуществлять ежемесячные инъекции, для которых также потребуется 15 инъекций, из которых 9 инъекций осуществляются в первый год с момента начала лечения и 12 инъекций в дополнительный год лечения.
Еще большим затруднением является лечение с помощью сублингвальной иммунотерапии (SLIT), требующей ежедневного введения экстракта аллергена, что является одной из причин низкого спроса такой терапии. Кроме того, SLIT индуцирует очень низкие уровни аллерген-специфических IgG-антител, и пока неясно, достаточно ли активируется IgG-антител после такой терапии, чтобы блокировать связывание IgE у пациентов с аллергией.
В отличие от приведенных выше особенностей применения вакцин на основе экстрактов аллергенов, схема лечения рекомбинантными вакцинами на основе PreS-белка, будет представлять собой предсезонный курс из 3-5 инъекций с последующими 1 или 2 предсезонными инъекциями в следующем году. Таким образом максимальный курс будет составлять 7 инъекций на два года лечения. Предположение о том, что для повышения уровня аллерген-специфических IgG-антител после базового курса иммунизации необходимы только 1 или 2 дополнительные инъекции, подтверждается результатами, полученными в клинических испытаниях вакцины против аллергии на пыльцу трав ВМ32, созданной на основе PreS-белка. В проведенном клиническом исследовании было обнаружено, что одной инъекции достаточно, для восстановления аллерген-специфических антител IgG до уровней, полученных через месяц после основного курса иммунизации [12, 13]. Таким образом, лечение вакцинами на основе рекомбинантных PreS-белков будет намного удобнее, чем любая существующая в настоящее время форма AIT.
Иммунизация B-Pres и AB-PreS индуцирует протективные антитела, которые ингибируют связывание IgE с аллергеном Bet v 1 значительно сильнее, чем антитела, индуцированные коммерческими вакцинами на основе экстрактов аллергенов.
Пример 6.3. Сравнительный анализ связывания IgE с аллергеном яблока Mal d 1 при иммунизации AB-PreS и B-Pres.
Иммунизации двумя рекомбинантными вакцинами на основе PreS, B-Pres и AB-Pres, индуцировали сходный уровень протективных антител, которые ингибировали IgE-связывание с аллергеном Bet v 1 (рисунок 11). Обе эти вакцины, в отношении активации антител, блокирующих связывание IgE с Bet v 1, оказались эффективнее остальных вакцин, что мы протестировали (Рисунок 9 и 11). Поэтому B-PreS и AB-PreS сравнивали по их способности индуцировать протективные антитела, блокирующие IgE-связывание с одним из наиболее важных перекрестно-реактивных пищевых аллергенов - аллергеном яблока Mal d 1. На рисунке 12 показано, что после иммунизации тремя дозами (20 мкг, 40 мкг, 80 мкг) AB-PreS, ингибирование IgE-связывания с Mal d 1 было значительно выраженнее, чем ингибирование такими же дозами B-PreS. В итоге рекомбинантная вакцина AB-PreS от аллергии на пыльцу березы на основе PreS-белка, индуцирует более высокий уровень проективных антител, блокирующих IgE-связывание с Bet v 1 и Mal d 1 по сравнению с другими протестированными вакцинами.
Антитела, индуцированные иммунизацией AB-Pres, значительно лучше ингибируют IgE-связывание с аллергеном яблока Mal d 1, в сравнении с B-Pres.
Таким образом, получение рекомбинантного полипептида AB-PreS (SEQ ID NO 2), содержащего эпитопы, необходимые для активации защитных антител против Bet v 1 и Mal d 1 в одном белке, позволяет получить однокомпонентную вакцину, вместо того, чтобы использовать вакцины, содержащие два или более производных аллергенов.
У полученного рекомбинантного полипептида AB-PreS, который содержит пептиды из аллергена яблока и березы, слитые с белком PreS, полностью отсутствует IgE-реактивность, что свидетельствует о большей безопасности данного белка. Кроме того, в состав AB-PreS входят пептиды аллергена яблока и березы, имитирующие В-, но не Т-клеточные эпитопы указанных аллергенов. Элиминация Т-клеточных эпитопов из состава AB-PreS позволяет добиться более существенной безопасности конструкта, т.к. исключена возможность активации Т-клеток, продуцирующих провоспалительные факторы.
Данная конструкция полипептида AB-PreS взывает повышенный иммунный ответ против аллергенных молекул, и индуцирует блокирующие IgG-антитела к аллергену пыльцы березы Bet v 1 и к аллергену яблока Mal d 1. При этом ингибирует IgE-связывание с Mal d 1 значительно сильнее, чем ингибирование такими же дозами B-PreS полипептида. В итоге вакцина от аллергии на пыльцу березы на основе AB-PreS полипептида индуцирует более высокий уровень проективных антител, блокирующих IgE-связывание с Bet v 1 и Mal d 1 по сравнению с другими коммерческими вакцинами.
Список источников
1. Dorofeeva Y. et al. Past, presence, and future of allergen immunotherapy vaccines // Allergy: European Journal of Allergy and Clinical Immunology. 2020.
2. Eckl-Dorna J. et al. Allergen-specific antibodies regulate secondary allergen-specific immune responses // Frontiers in Immunology. Frontiers Media S.A., 2019. Vol. 9, №3131. P. 1-15.
3. van Neerven R.J. et al. Blocking antibodies induced by specific allergy vaccination prevent the activation of CD4+T cells by inhibiting serum-IgE-facilitated allergen presentation. // J. Immunol. 1999.
4. Niederberger V. et al. Vaccination with genetically engineered allergens prevents progression of allergic disease // Proc. Natl. Acad. Sci. U. S. A. 2004. Vol. 101, №2. P. 14677-14682.
5. Elisyutina O. et al. Bet v 1-specific IgE levels and PR-10 reactivity discriminate silent sensitization from phenotypes of birch allergy // Allergy: European Journal of Allergy and Clinical Immunology. 2019.
6. Gadermaier E. et al. Analysis of the antibody responses induced by subcutaneous injection immunotherapy with birch and Fagales pollen extracts adsorbed onto aluminum hydroxide // Int. Arch. Allergy Immunol. 2010. Vol. 151, № LP. 17-27.
7. Bucher X. et al. Effect of tree pollen specific, subcutaneous immunotherapy on the oral allergy syndrome to apple and hazelnut // Allergy Eur. J. Allergy Clin. Immunol. 2004.
8. Valenta R., Campana R., Niederberger V. Recombinant allergy vaccines based on allergen-derived В cell epitopes // Immunology Letters. Elsevier B.V., 2017. Vol. 189. P. 19-26.
9. Valenta R. et al. Vaccine development for allergen-specific immunotherapy based on recombinant allergens and synthetic allergen peptides: Lessons from the past and novel mechanisms of action for the future // J. Allergy Clin. Immunol. Mosby Inc., 2016. Vol. 137, №2. P. 351-357.
10. Marth K. et al. A nonallergenic birch pollen allergy vaccine consisting of hepatitis PreS-fused Bet v 1 peptides focuses blocking IgG toward IgE epitopes and shifts immune responses to a tolerogenic and Thl phenotype // J. Immunol. The American Association of Immunologists, 2013. Vol.190, №7. P. 3068-3078.
11. Zieglmayer P. et al. Mechanisms, safety and efficacy of а В cell epitope-based vaccine for immunotherapy of grass pollen allergy // EBioMedicine. Elsevier B.V., 2016. Vol. 11. P. 43-57.
12. Niederberger V. et al. Safety and efficacy of immunotherapy with the recombinant B-cell epitope-based grass pollen vaccine BM32 // J. Allergy Clin. Immunol. Mosby Inc., 2018. Vol. 142, №2. P. 497-509.
13. Eckl-Dorna J. et al. Two years of treatment with the recombinant grass pollen allergy vaccine BM32 induces a continuously increasing allergen-specific IgG4 response // EBioMedicine. 2019. Vol. 50. P. 421-432.



Были собраны образцы сыворотки от пациентов с аллергией на пыльцу березы (ВРА), пациентов с аллергией, но без аллергии на пыльцу березы (non-ВРА) или сыворотки от пациентов, не страдающих аллергией - здоровые добровольцы (НС).
| название | год | авторы | номер документа |
|---|---|---|---|
| ГИПОАЛЛЕРГИЧЕСКИЕ ВАРИАНТЫ Mal d 1, ГЛАВНОГО АЛЛЕРГЕНА Malus domectica | 2012 |
|
RU2624030C2 |
| ГИБРИДНЫЕ БЕЛКИ-НОСИТЕЛИ ПЕПТИДОВ В КАЧЕСТВЕ ВАКЦИН ОТ АЛЛЕРГИИ | 2012 |
|
RU2630652C2 |
| НОВЫЕ МУТАНТНЫЕ АЛЛЕРГЕНЫ | 2001 |
|
RU2285042C2 |
| НОВЫЕ АЛЛЕРГЕНЫ ПШЕНИЦЫ | 2008 |
|
RU2502742C2 |
| КЛЕЩЕВОЙ АЛЛЕРГЕН ДОМАШНЕЙ ПЫЛИ | 2007 |
|
RU2570563C2 |
| Способ получения аллергоида на основе пчелиного яда для проведения аллерген-специфической иммунотерапии | 2019 |
|
RU2731509C1 |
| МИКРОАНАЛИЗ НА АЛЛЕРГЕНЫ | 2001 |
|
RU2276790C2 |
| СПОСОБ ПОЛУЧЕНИЯ ГИПОАЛЛЕРГЕННОГО ОСНОВНОГО АЛЛЕРГЕНА ПЫЛЬЦЫ БЕРЕЗЫ r Bet v 1 | 2003 |
|
RU2311424C2 |
| НОВЫЙ АЛЛЕРГЕН | 2011 |
|
RU2608494C2 |
| АЛЛЕРГЕНЫ, ПОЛУЧЕННЫЕ РЕКОМБИНАНТНЫМИ СПОСОБАМИ | 2010 |
|
RU2592674C2 |







Изобретение относится к рекомбинантному полипептиду для лечения и профилактики аллергии на пыльцу березы и яблоко, содержащему пептидные фрагменты аллергенов Bet vl и Mal d 1, слитые с полипептидом PreS вируса гепатита В, представленный аминокислотной последовательностью SEQ ID NO: 2, а также к композиции вакцины для лечения или профилактики аллергии на пыльцу березы и яблоко, содержащей эффективное количество вышеуказанного рекомбинантного полипептида. 2 н.п. ф-лы, 6 пр., 12 ил., 2 табл.
1. Рекомбинантный полипептид для лечения и профилактики аллергии на пыльцу березы и яблоко, содержащий пептидные фрагменты аллергенов Bet vl и Mal d 1, слитые с полипептидом PreS вируса гепатита В, представленный аминокислотной последовательностью SEQ ID NO: 2.
2. Рекомбинантный полипептид по п. 1, содержащий по меньшей мере четыре пептидных фрагмента аллергена дикого типа пыльцы березы Bet v 1 и по меньшей мере два пептидных фрагмента аллергена дикого типа яблока Mal d 1.
3. Рекомбинантный полипептид по п. 2, где аллерген дикого типа выбирают из группы, состоящей из основных аллергенов пыльцы березы, в частности Bet v 1, где два пептидных фрагмента имеют аминокислотную последовательность SEQ ID NO: 5 и другие два пептидных фрагмента имеют аминокислотную последовательность SEQ ID NO: 6.
4. Рекомбинантный полипептид по п. 2, где аллерген дикого типа выбирают из группы, состоящей из основных аллергенов яблока, в частности Mal d 1, имеющих один пептидный фрагмент с аминокислотной последовательностью SEQ ID NO: 3 и другой пептидный фрагмент с аминокислотной последовательностью SEQ ID NO: 4.
5. Композиция вакцины для лечения или профилактики аллергии на пыльцу березы и яблоко, содержащая эффективное количество рекомбинантного полипептида по п. 1 и фармацевтически приемлемый наполнитель.
6. Композиция по п. 5, отличающаяся тем, что дополнительно содержит по меньшей мере один адъювант и/или консервант.
| NIEDERBERGER V | |||
| et al., Vaccination with genetically engineered allergens prevents progression of allergic disease, Proceedings of the National Academy of Sciences of the United States of America, 2004, Vol.101, N2, pp.14677-14682 | |||
| CHAPMAN M.D | |||
| et al., Recombinant allergens for diagnosis and therapy of allergic disease, Journal of Allergy and |

