Область техники
Группа заявляемых изобретений относится к области биотехнологии, в частности, к биосинтезу ксилоглюканазы, и представляет собой рекомбинантную термостабильную ксилоглюканазу, имеющую последовательность SEQ ID NO 1, которая соответствует последовательности ксилоглюканазы семейства 12 из Aspergillus cervinus ВКПМ F-612 с заменой (Y37V), направленной на повышение термостабильности фермента, рекомбинантный штамм Escherichia coli, способный синтезировать заявленную ксилоглюканазу и полученный путем трансформации штамма Escherichia coli BL21 (DE3) GOLD плазмидой pET-AsCeGH12b-mut, содержащей мутантный ген ксилоглюканазы из Aspergillus cervinus ВКПМ F-612 под контролем промотора T7lac, а также способ микробиологического синтеза термостабильной ксилоглюканазы на основе заявленного штамма.
Уровень техники
Растительная биомасса является дешевым возобновляемым источником ферментируемых сахаров для нужд промышленной биотехнологии и сельского хозяйства. Ферментативный гидролиз - эффективный и экологически безопасный способ конверсии полисахаридов растительной биомассы в сахара. Химическое разнообразие растительных полисахаридов обуславливает необходимость использования широкого спектра ферментов для гидролиза.
Ксилоглюкан - один из основных полисахаридов первичной клеточной стенки двудольных и многих однодольных растений [Hayashi Т. Xyloglucans in the primary-cell wall. Annu. Rev. Plant Physiol. Plant Mol. Biol, 1989, 40, 139-168]. Молекулы ксилоглюкана состоят из (1,4)-β-связанных олигосахаридов на основе целлотетраозы, в которой три из четырех глюкозных остатков связаны (1,6)-α-связями с остатками D-ксилозы. Некоторые из остатков ксилозы, в свою очередь, могут быть связаны (1,2)-β-связями с остатками D-галактозы или L-арабинозы. Остатки D-галактозы могут быть связаны (1,2)-α-связями с остатками L-фукозы. Модификация остатков ксилозы видоспецифична, что определяет множество типов ксилоглюкана в растениях. В клеточной стенке растений ксилоглюкан выполняет структурную функцию и является связующим гликаном. Молекулы ксилоглюкана формируют прочную полисахаридную сеть, взаимодействуя с молекулами целлюлозы, гемицеллюлоз и белков. Также ксилоглюкан может играть роль запасного полисахарида, как, например, в семенах Tamarindus indica [Shankaracharya N.B. Tamarind - chemistry, technology and uses - a critical approach. J. Food. Sci. Technol, 1998, 35, 193-208].
Поскольку ксилоглюкан является основным связующим гликаном в составе первичной клеточной стенки всех двудольных и многих однодольных растений, полный ферментативный гидролиз растительной биомассы с помощью только целлюлаз затруднен без предварительного гидролиза ксилоглюкана. Ферменты, способные специфически гидролизовать (1,4)-β-связи глюканового остова молекулы ксилоглюкана, называются ксилоглюканазами [Grishutin S.G., Gusakov A.V., Markov A.V., Ustinov B.B., Semenova M.V., Sinitsyn A.P. Specific xyloglucanases as a new class of polysaccharide-degrading enzymes. Biochim. Biophys. Acta., 2004, 1674(3), 268-281]. Целлобиогидролазы, специфичные к редуцирующим концам олигоксилоглюканов (ЕС 3.2.1.150, OXG-RCBH), отрезают целлобиозу с редуцирующего конца основной цепи ксилоглюкана или ксилоглюкан-олигосахаридов, в случае, если остаток глюкозы на конце цепи незамещен; эндо-ксилоглюканазы (ксилоглюкан-специфические эндо-(1,4)-β-глюканазы, ЕС 3.2.1.151, XEG) гидролизуют только (1,4)-β-гликозидные связи у незамещенных остатков глюкозы в молекуле ксилоглюкана и ксилоглюкановых олигосахаридах [Yaoi K., Mitsuishi Y. Purification, characterization, cDNA cloning, and expression of a xyloglucan endoglucanase from Geotrichum sp. M128. FEBS Lett., 2004, 560(1-3), 45-50]. Согласно данным базы CAZy (http://www.cazy.org/) ферменты с ксилоглюканазной активностью найдены среди семейств гликозил-гидролаз GH5, GH9, GH12, GH16, GH26, GH44, GH74. Ферменты, специфически гидролизующие ксилоглюкан, также могут проявлять побочную активность на растворимой и нерастворимой целлюлозе и (1,4/1,3)-β-глюканах: β-глюкане злаков и лихенане.
Комплексные ферментные препараты, содержащие ксилоглюканазы, используются в качестве кормовых добавок для повышения пищевой и энергетической ценности кормов [Завьялов А.В., Рыков С.В., Лунина Н.А., Сушкова В.И., Яроцкий С.В., Березина О.В. Растительный полисахарид ксилоглюкан и ферменты, его гидролизующие (Обзор литературы). Химия растительного сырья., 2018, 4, 43-61). На основе ксилоглюканаз созданы ферментные препараты для гидролиза растительных полисахаридов, в особенности биомассы молодых побегов [RU 2358756; RU 2361918 С1]. Также ксилоглюканазы находят применение как модифицирующие агенты для изменения физических и биохимических свойств растительных волокон [Park Y.W., Baba K., Furuta Y., Iida I., Sameshima K., Arai M., Hayashi T. Enhancement of growth and cellulose accumulation by overexpression of xyloglucanase in poplar. FEBS Lett., 564 (1-2), 183-187], для лигирования химически модифицированных олигосахаридов с молекулой ксилоглюкана, для изменения функциональности целлюлозных фибрилл [Brumer Н., Zhou Q., Baumann М.J., Carlsson K., Teeri Т.Т. Activation of crystalline cellulose surfaces through the chemoenzymatic modification of xyloglucan. J. Am. Chem. Soc, 2004, 126 (18), 5715-5721]. Ксилоглюканазы часто применяют в пищевой и текстильной индустрии и в фармацевтике [Shankaracharya N.B. Tamarind -chemistry, technology and uses - a critical approach. J. Food. Sci. Technol, 1998, 35, 193-208; Miyazaki S., Endo K., Kawasaki N., Kubo W., Watanabe H., Attwood D. Oral sustained delivery of paracetamol from in situ gelling xyloglucan formulations. Drug Dev. Ind. Pharm., 2003, 29 (2), 113-119].
Технологические процессы с участием ферментов часто протекают при повышенной температуре [RU 2481000]. Например, зерновые корма для сельскохозяйственных животных предварительно запаривают горячей водой. В связи с этим возникает необходимость использования термостабильных ферментов, сохраняющих активность при высоких температурах. Продуцентами термостабильных ферментов являются главным образом термофильные, но в ряде случаев и мезофильные бактерии и грибы. Удельная активность у бактериальных ферментов обычно ниже, чем у грибных [Viikari L., Alapuranen М., Puranen Т., Vehmaanperä J., Siika-Aho M. Thermostable enzymes in lignocellulose hydrolysis. Adv. Biochem. Eng. BiotechnoL, 2007, 108, 121-145].
В источниках информации имеются сведения о микроорганизмах -природных продуцентах ксилоглюканаз: Geotrichum sp. [Yaoi K., Mitsuishi Y. Purification, characterization, cDNA cloning, and expression of a xyloglucan endoglucanase from Geotrichum sp. M128. FEBS Lett, 2004, 560 (1-3), 45-50], Aspergillus japonicus, Chrysosporium lucknowense, Trichoderma reesei [Grishutin S.G., Gusakov A.V., Markov A.V., Ustinov B.B., Semenova M.V., Sinitsyn A.P. Specific xyloglucanases as a new class of polysaccharide-degrading enzymes. Biochim. Biophys. Acta., 2004, 1674 (3), 268-281; Markov A.V., Gusakov A.V., Kondratyeva E.G., Okunev O.N., Bekkarevich A.O., Sinitsyn A.P. New effective method for analysis of the component composition of enzyme complexes from Trichoderma reesei. Biochemistry (Mosc)., 2005, 70 (6), 657-663; Qi H., Bai F., Liu A. Purification and characteristics of xyloglucanase and five other cellulolytic enzymes from Trichoderma reesei QM9414. Biochemistry (Mosc)., 2013, 78 (4), 424-30; Lopes D.C.B., Carraro C.B., Silva R.N., de Paula R.G. Molecular characterization of xyloglucanase cel74a from Trichoderma reesei. Int J Mol Sci., 2021, 22(9), 4545), Phanerochaete chrysosporium [Ishida Т., Yaoi K., Hiyoshi A., Igarashi K., Samejima M. Substrate recognition by glycoside hydrolase family 74 xyloglucanase from the basidiomycete Phanerochaete chrysosporium. FEBS J., 2007, 274 (21), 5727-5736], Cellvibho japonicus [Attia M., Stepper J., Davies G.J., Brumer H. Functional and structural characterization of a potent GH74 endo-xyloglucanase from the soil saprophyte Cellvibrio japonicus unravels the first step of xyloglucan degradation. FEBS J., 2016, 283 (9):1701-1719), Aspergillus clavatus [Damásio A.R., Rubio M.V., Oliveira L.C., Segato F., Dias B.A., Citadini A.P., Paixão D.A., Squina F.M. Understanding the function of conserved variations in the catalytic loops of fungal glycoside hydrolase family 12. Biotechnol Bioeng., 2014, 111 (8), 1494-1505], Sporotrichum thermophile, Myceliophthora thermophila и Sporotrichum pruinosum [Крестьянова И.Н., Сахибгараева Л.Ф., Березина O.B., Рыков С.В., Завьялов А.В., Зверлов В.В., Яроцкий С.В. Характеристика грибных штаммов-продуцентов термостабильных ксилоглюканаз из всероссийской коллекции промышленных микроорганизмов. Мол. генетика, микробиол., вирусол., 2016, 3, 109-114], Paenibacillus odorifer [Gusakov A.V., Uporov I.V., Sinitsyna O.A. Molecular dynamics simulations of two GH74 endo-processive xyloglucanases and the mutated variants to understand better the mechanism of the enzyme action. Biochim. Biophys. Acta Gen. Subj., 2020, 1864(12), 129721), Dichomitus squalens и Pleurotus ostreatus [Sun P., Li X., Dilokpimol A., Henrissat В., de Vries R.P., Kabel M.A., Mäkelä M.R. Fungal glycoside hydrolase family 44 xyloglucanases are restricted to the phylum Basidiomycota and show a distinct xyloglucan cleavage pattern. Science, 2021, 25(1), 103666], Bacteroides ovatus и В. uniformis [Grondin J.M., Dejean G., van Petegem F., Brumer H. Cell surface xyloglucan recognition and hydrolysis by the human gut commensal Bacteroides uniformis. Appl. Microbiol. Biotechnol., 2022, 88(1), e0156621], Trichoderma longibrachiatum и Thermothelomyces thermophiles [Contato A.G., de Oliveira Т.В., Aranha G.M., de Freitas E.N., Vici A.C., Nogueira K.M.V., de Lucas R.C., Scarcella A.S. d. A., Buckeridge M.S., Silva R.N., Polizeli M. d. L.T. d. M. Prospection of fungal lignocellulolytic enzymes produced from jatoba (Hymenaea courbaril) and tamarind (Tamarindus indica) seeds: scaling for bioreactor and saccharification profile of sugarcane bagasse. Microorganisms., 2021, 9(3), 533]. Методами селекции и радиационного мутагенеза получены штаммы с повышенным уровнем синтеза ксилоглюканаз: Penicillium verruculosum [RU 2361918 C1], Penicillium funiculosum [RU 2323254], Aspergillus aculeatus [RU 2303057]. Подобным образом был получен высокоактивный штамм мицелиального гриба Aspergillus foetidus - продуцента комплекса карбогидраз, содержащего ксилоглюканазу [RU 2323973]. В последовательностях гликозил-гидролаз из Humicola grisea, Hypocrea jecorina и Hypocrea schweinitzii с помощью сайт-направленного мутагенеза проведены аминокислотные замены с целью повышения термостабильности ферментов [Sandgren М., Stahlberg J., Mitchinson С. Structural and biochemical studies of GH family 12 cellulases: improved thermal stability, and ligand complexes. Prog. Biophys. Mol. Biol, 2005., 89(3), 246-91; US 8008056 B2; US 20140302585 A1].
Рекомбинантные продуценты ксилоглюканаз обладают рядом несомненных преимуществ, основным из которых является повышенная продуктивность рекомбинантных штаммов по целевому белку.
В качестве штаммов-реципиентов часто выступают микроскопические грибы родов Aspergillus, Penicillium, Fusarium. Например, в Aspergillus oryzae экспрессированы ксилоглюканазы из Malbranchea cinnamomea [US 6500658], в Fusarium venenatum - ксилоглюканаза из Trichoderma reesei [US 2004067569], в Penicillium canescens - ксилоглюканаза из Penicillium canescens [RU 2358756], в Aspergillus niger - ксилоглюканаза из Thielavia australiensis и собственная ксилоглюканаза [WO 2014138983; Hasper A.A., Dekkers E., van Mil M., van de Vondervoort P.J., de Graaff L.H. EglC, a new endoglucanase from Aspergillus niger with major activity towards xyloglucan. Appl Environ Microbiol., 2002, 68(4), 1556-60), в Aspergillus nidulans - ксилоглюканаза из Aspergillus terreus (Vitcosque GL, Ribeiro LF, de Lucas RC, da Silva TM, Ribeiro LF, de Lima Damasio AR, Farinas CS, Goncalves AZ, Segato F, Buckeridge MS, Jorge JA, Polizeli ML. The functional properties of a xyloglucanase (GH12) of Aspergillus terreus expressed in Aspergillus nidulans may increase performance of biomass degradation. Appl. Microbiol. Biotechnol., 2016, 100(21), 9133-9144]. К недостаткам штаммов-продуцентов на основе микроскопических грибов относят сложности с проведением генетических манипуляций на штаммах вследствие нестандартизированного и недостаточно разработанного генетического инструментария (Demain AL, Vaishnav P. Production of recombinant proteins by microbes and higher organisms. Biotechnol Adv., 2009, 27(3), 297-306).
Методами генной инженерии получены штаммы дрожжей Pichia pastoris -продуценты рекомбинантных ксилоглюканаз из Phanerochaete chrysosporium [Ishida Т., Yaoi K., Hiyoshi A., Igarashi K., Samejima М. Substrate recognition by glycoside hydrolase family 74 xyloglucanase from the basidiomycete Phanerochaete chrysosporium. FEBS J., 2007, 274(21), 5727-5736], Aspergillus cervinus ВКПМ F-612 [RU 2639248], Myceliophthora thermophila ВКПМ F-244 [RU 2605629] и Penicillium oxalicum [CN 104388406; Xian L., Wang F., Yin X., Feng J.-X. Identification and characterization of an acidic and acid-stable endoxyloglucanase from Penicillium oxalicum. Int J Biol Macromol., 2016, 86, 512-518). Недостатком штаммов-продуцентов на основе дрожжей является их способность осуществлять посттрансляционные модификации, в частности, различные типы гликозилирования, что мешает получению белков в гомогенной форме, подходящей для биохимическихи структурных исследований.
Описаны полученные методами генной инженерии штаммы Escherichia coli - продуценты рекомбинантных ксилоглюканаз из Geotrichum sp. M128 [US 2004038367; Yaoi K., Mitsuishi Y. Purification, characterization, cloning, and expression of a novel xyloglucan-specific glycosidase, oligoxyloglucan reducing end-specific cellobiohydrolase. J Biol Chem., 2002, 277(50), 48276-48281], Fusarium graminearum [Habrylo O., Song X., Forster A., Jeltsch J.M., Phalip V. Characterization of the four GH12 endoxylanases from the plant pathogen Fusarium graminearum. J. Microbiol Biotechnol, 2012, 22(8), 1118-1126], Rhizomucor miehei [Song S., Tang Y., Yang S., Yan Q., Zhou P., Jiang Z. Characterization of two novel family 12 xyloglucanases from the thermophilic Rhizomucor miehei. Appl. Microbiol. Biotechnol., 2013, 97(23), 10013-10024], Aspergillus cervinus [RU 2625013], Xanthomonas citri [Feng Т., Yan K.P., Mikkelsen M.D., Meyer A.S., Schols H.A., Westereng В., Mikkelsen J.D. Characterisation of a novel endo-xyloglucanase (XcXGHA) from Xanthomonas that accommodates a xylosyl-substituted glucose at subsite -1. Appl. Microbiol. Biotechnol., 2014, 98(23), 9667-9679], Xanthomonas campestris [de Araujo E.A., Tomazini A.Jr., Kadowaki M.A., Murakami M.T., Polikarpov I. Crystallization and preliminary X-ray diffraction analysis of a new xyloglucanase from Xanthomonas campestris pv. campestris. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun., 2013, 69(Pt 6), 676-678], Bacteroides uniformis [Grondin J.M., Dejean G., van Petegem F., Brumer H. Cell surface xyloglucan recognition and hydrolysis by the human gut commensal Bacteroides uniformis. Appl. Microbiol. Biotechnol., 2022, 88(1), e0156621], Cellvibrio japonicus [Attia M.A., Brumer H. New family of carbohydrate-binding modules defined by a galactosyl-binding protein module from a Cellvibrio japonicus endo-xyloglucanase. Appl. Environ. Microbiol, 2021, 87(5), е02634-20]. Иногда для гетерологичной экспрессии грибных ксилоглюканаз применяют отличные от Е. coli бактериальные реципиенты, например, Acidothermus cellulolyticus использован для гетерологичной экспрессии ксилоглюканаз из Acremonium sp., Cladorrhinum foecundissimum, Humicola insolens, Thielavia terrestris, Trichoderma reesei, Aspergillus aculeatus, Aspergillus aculeatus, Aspergillus fumigatus, Myceliophthora thermophila [Vlasenko E., Schülein M., Cherry J., Xu F. Substrate specificity of family 5, 6, 7, 9, 12, and 45 endoglucanases. Bioresour Technol., 2010, 101(7), 2405-2411).
В отличие от гетерологичного синтеза ферментов в грибных продуцентах, биосинтез в Е. coli позволяет получить рекомбинантный белок в негликозилированном виде, подходящем для биохимических исследований и кристаллизации.
Ближайшим аналогом заявляемого штамма является рекомбинантный штамм Escherichia coli BL21-CODONPLUS (DE3)-RIPL - продуцент ксилоглюканазы семейства GH12 из Aspergillus cervinus ВКПМ F-612, кодируемой геном AsCeGH12b, интегрированным в рекомбинантную плазмиду pET-AsCeGH12b под контроль промотора Т71ас [RU 2625013] Рекомбинантный фермент AsCeGH12b является ксилоглюкан-специфической эндо-(1,4)-β-глюканазой семейства 12 гликозил-гидролаз (GH12).
Ближайшим аналогом заявляемой ксилоглюканазы является рекомбинантная ксилоглюканаза AsCeGH12b семейства GH12 (ЕС 3.2.1.151). Фермент проявляет максимум активности при 55°С, рН 5.0 и практически полностью инактивируется при 65°С [RU 2625013; Rykov S.V., Kornberger P., Herlet J., Tsurin N.V., Zorov I.N., Zverlov V.V., Liebl W., Schwarz W.H., Yarotsky S.V., Berezina O.V. Novel endo-(1,4)-β-glucanase Bgh12A and xyloglucanase Xgh12B from Aspergillus cervinus belong to GH12 subgroup I and II, respectively. Appl. Microbiol. Biotechnol., 2019, 103(18), 7553-7566].
Ближайшим аналогом заявляемого способа микробиологического синтеза ксилоглюканазы является способ синтеза рекомбинантной ксилоглюканазы AsCeGH12b при ферментации штамма Escherichia coli BL21-CODONPLUS (DE3)-RIPL [RU 2625013].
Технической проблемой, на решение которой направлена заявляемая группа изобретений, является расширение арсенала рекомбинантных ксилоглюканаз, штаммов-продуцентов и способов микробиологического синтеза рекомбинантных ксилоглюканаз.
Раскрытие сущности изобретения
Техническим результатом заявляемого изобретения является получение рекомбинантной мутантной ксилоглюканазы из Aspergillus cervinus, которая отличается от фермента дикого типа более высокими температурным максимумом активности и более высокой термостабильностью.
Технический результат достигнут тем, предложена рекомбинантная мутантная ксилоглюканаза AsCeGH12b-mut, имеющая SEQ ID NO 1, которая соответствует аминокислотной последовательности ксилоглюканазы семейства 12 из Aspergillus cervinus ВКПМ F-612 являющаяся ферментом дикого типа имеющая последовательность SEQ ID NO 3, с заменой природного остатка тирозина в положении 37 остатком валина, и обладающая повышенными термостабильностью и температурным максимумом активности по сравнению с ферментом дикого типа.
Кроме того, предложен рекомбинантный штамм Escherichia coli ВКПМ В-14321 -продуцент ксилоглюканазы AsCeGH12b-mut по п. 1, имеющий SEQ ID NO 1 и кодируемый геном AsCeGH12b-mut, имеющим SEQ ID NO 2.
Кроме того, предложен способ микробиологического синтеза рекомбинантной мутантной ксилоглюканазы AsCeGH12b-mut по п. 1, предусматривающий культивирование рекомбинантного штамма бактерии Escherichia coli по п. 2 в аэробных условиях в подходящей питательной среде до максимального накопления целевого продукта.
Краткое описание чертежей
Изобретение проиллюстрировано следующими фигурами графических изображений:
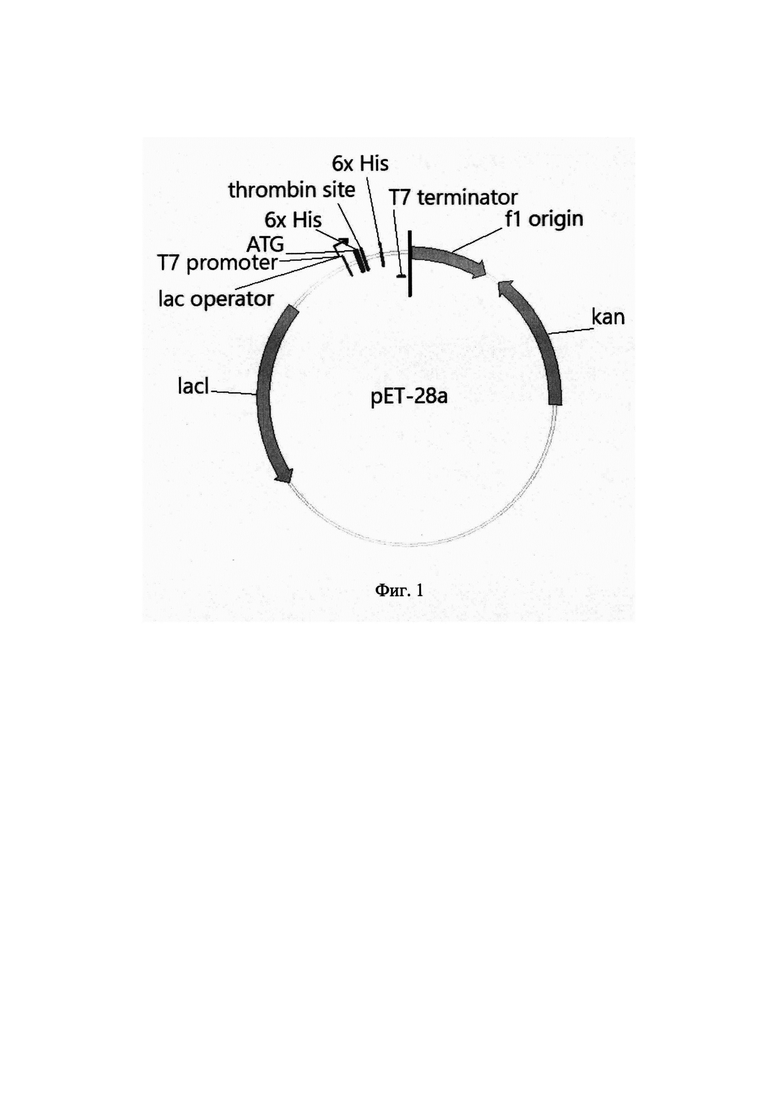
На Фиг. 1 изображена схема плазмидной ДНК рЕТ-28а. В состав плазмидного вектора рЕТ-28а входят: промотор фага T7lac; последовательность, кодирующая полигистидиновый хвост; тромбиновый сайт; последовательность, кодирующая полигистидиновый хвост; терминатор фага Т7; кодирующая область гена kan, обеспечивающая устойчивость штаммов Е. coli к канамицину; кодирующая область гена lacI; репликон ColE1 (pBR-322).
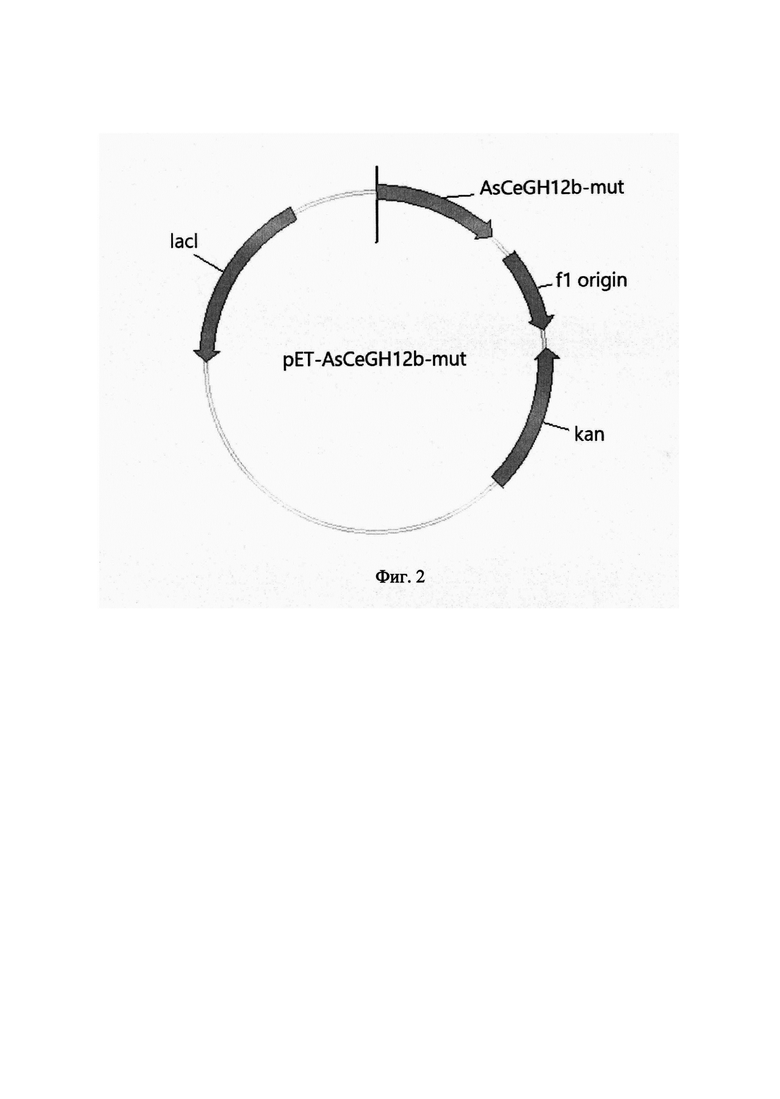
На Фиг. 2 изображена схема рекомбинантной плазмидной ДНК рЕТ-AsCeGH12b-mut. В состав плазмидного вектора pET-AsCeGH12b-mut входят: кодирующая часть гена AsCeGH12b-mut с полигистидиновой последовательностью на 5'-конце; кодирующая область гена кап, обеспечивающая устойчивость штаммов Е. coli к канамицину; кодирующая область гена lacI; репликон ColE1 (pBR-322).
Осуществление изобретения
Процесс получения заявляемого штамма состоит из нескольких этапов:
- конструирования мутантного гена AsCeGH12b-mut (SEQ ID NO 2), кодирующего ксилоглюканазу AsCeGH12b-mut (SEQ ID NO 1).
- конструирования плазмидной ДНК (плазмиды) pET-AsCeGH12b-mut (SEQ ID NO 4), содержащей структурную часть мутантного гена AsCeGH12b-mut.
- конструирования заявляемого рекомбинантного штамма Escherichia coli ВКПМ В-14321 Е. colli /pET-AsCeGH12b-mut.
Этап 1. Конструирование гена AsCeGH12b-mut, кодирующего мутантную форму ксилоглюканазы AsCeGH12b из Aspergillus cervinus ВКПМ F-612
Ксилоглюканаза AsCeGH12b из Aspergillus cervinus ВКПМ F-612, имеющая последовательность SEQ ID NO 3, проявляет максимальную активность на ксилоглюкане при 55°С, рН 5.0. AsCeGH12b сохраняет 100% активности после инкубации в течение 2 ч при 50°С, рН 5.0, время полуинактивации при 55°С, рН 5.0, составляет 158 мин, при 60°С - менее 5 мин.
Для получения более термостабильного варианта данного фермента конструируют мутантную форму AsCeGH12b-mut (SEQ ID NO 1), осуществляя в последовательности AsCeGH12b замену остатка тирозина в положении 37 остатком валина путем сайт-направленного мутагенеза. Дизайн нуклеотидной последовательности AsCeGH12b-mut разрабатывают на основании гена AsCeGH12b (GenBank: MF527262.1), кодирующего ксилоглюканазу AsCeGH12b семейства GH12 из Aspergillus cervinus ВКПМ F-612. Введение мутаций в ген AsCeGH12b осуществляли методом перекрывающихся ПЦР-продуктов (Overlap PCR) с использованием праймеров и плазмиды pPIC-AsCeGH12b, содержащей ген AsCeGH12b в качестве матрицы [RU 2639248]. Для проведения мутагенеза используют следующие прайм еры:
AsCeXgh_for_BamHI - ATAGGATCCGCGACTACGTACTGC,
AsCeXgh_rev_Sal - ATAGTCGACTTAAGCCACACTGGCCGAG,
AsCeXgh_for_Y37V - AGTGCTTCACGGTCAATTCTCTCTC,
AsCeXgh_rev_Y37V - AGAGAATTGACCGTGAAGCACTG.
В результате получают фрагмент ДНК, содержащий мутированный ген AsCeGH12b-mut. Корректность введенной мутации подтверждают при помощи секвенирования методом Сэнгера.
Этап 2. Конструирование плазмидной ДНК (плазмиды) pET-AsCeGH12b-mut, содержащей структурную часть мутантного гена ксилоглюканазы семейства GH12 из Aspergillus cervinus ВКПМ F-612
Плазмиду pET-AsCeGH12b-mut (SEQ ID NO 4) конструируют путем клонирования полученного амплификацией фрагмента ДНК, содержащего структурную часть гена AsCeGH12b-mut, в вектор рЕТ-28а, предназначенный для индуцируемой высокоэффективной экспрессии гомологичных и гетерологичных белков в Е. coli. Вектор рЕТ-28а размером 5369 пар оснований содержит промотор фага T7lac; последовательность, кодирующую полигистидиновый хвост: тромбиновый сайт; последовательность, кодирующую полигистидиновый хвост; терминатор фага Т7; кодирующую область гена кап, обеспечивающего устойчивость штаммов Е. coli к канамицину; кодирующую область гена lacI; репликон ColE1 (pBR-322) (Фиг. 1). Сконструированная плазмида рЕТ-AsCeGH12b-mut, размером 6007 пар оснований, наряду с генами вектора рЕТ-28а, содержит кодирующую область мутантного гена AsCeGH12b-mut. Со стороны 5'-конца гена AsCeGH12b-mut в той же рамке считывания расположена последовательность, кодирующая полигистидиновый хвост, находящаяся под контролем промотора T7lac (Фиг. 2).
Этап 3. Конструирование заявляемого рекомбинантного штамма Escherichia coli ВКПМ В-14321 (Е. coli/pET-AsCeGH12b-muf)
В качестве штамма-реципиента используют штамм Escherichia coli BL21 (DE3) GOLD (Novagen, США). Компетентные клетки данного штамма трансформируют плазмидой pET-AsCeGH12b-mut. В результате получают заявляемый рекомбинантный штамм Escherichia coli BL21 (DE3) GOLD/pET-AsCeGH12b-mut, способный синтезировать мутантную форму AsCeGH12b-mut ксилоглюканазы семейства GH12 из A. cervinus ВКПМ F-612 с повышенной по сравнению с ферментом дикого типа термостабильностью.
Заявляемый штамм Е. coli депонирован во Всероссийской коллекции промышленных микроорганизмов как Escherichia coli ВКПМ В-14321.
Морфологические и физиолого-биохимические характеристики заявляемого штамма.
Заявляемый штамм Е. coli ВКПМ В-14321 имеет следующие морфологические и физиолого-биохимические характеристики:
Морфологические признаки: Клетки палочковидной формы, грамотрицательные, неспороносные.
Культуральные признаки: клетки хорошо растут на обычно используемых питательных средах. На агаризованной LB-среде (вес. %: триптон - 1, дрожжевой экстракт - 0,5, хлорид натрия - 1, агар - 1,5, остальное - вода) образуются гладкие, серые, блестящие, круглые с ровным краем колонии. Рост в жидкой среде LB характеризуется равномерным помутнением среды.
Физиолого-биохимические признаки: клетки штамма продуцента растут в диапазоне температур 20-42°С, оптимум составляет 37°С. Наиболее благоприятные для роста значения рН находятся в интервале 6,8-7,2. При росте в аэробных условиях культура может усваивать азот как органических соединений (пептон, триптон, аминокислоты, дрожжевой экстракт), так и аммонийных и нитратных солей. Углерод усваивается в форме углеводов и аминокислот. Клетки проявляют устойчивость к канамицину (до 50 мкг/мл), обусловленную наличием в плазмиде рЕТ-28а гена kan.
Штамм Е. coli ВКПМ В-14321 синтезирует рекомбинантную ксилоглюканазу AsCeGH12b-mut.
Способ микробиологического синтеза ксилоглюканазы AsCeGH12b-mut в общем виде
Посевной материал, представляющий собой клетки рекомбинантного штамма - продуцента Е. coli ВКПМ В-14321, подготавливают путем инкубации в течение 16-18 часов при температуре 30-37°С с аэрацией на среде LB, содержащей 25 мкг/мл канамицина. Выросшую культуру переносят в соотношении 1:100 (по объему) в среду LB, содержащую 25 мкг/мл канамицина. Процесс культивирования ведут при температуре 37°С с аэрацией. Синтез целевого белка индуцируют путем добавления индуктора изопропил-β-D-1-тиогалактопиранозида (ИПТГ) до конечной концентрации 0,1 мМ при OD600=0,6-0,8 ОЕ. Клетки разрушают обработкой ультразвуком, проводя процедуру на льду. Биомассу клеток отделяют центрифугированием при 3000 об/мин и дважды промывают натрий-фосфатным буфером (вес. %: NaCl - 0,8, KCl - 0,02, Na2HPO4 - 0,15, KH2PO4 - 0,024, остальное - вода). Выделение ксилоглюканазы из клеточного экстракта проводят методом металл-афинной хроматографии (Green and Sambrook, 2012). Уровень синтеза целевого продукта определяют с помощью денатурирующего электрофореза в полиакриламидном геле (Green and Sambrook, 2012), а также путем определения удельной активности ксилоглюканазы (Ед/г сырой биомассы клеток). Количество целевого белка составляет: в виде телец включения - 10 и более процентов от суммарного белка клетки в зависимости от количества индуктора, и не менее 500 единиц активности физиологически активного белка на 1 грамм сырой биомассы, измеренной при 50°С на ксилоглюкане.
Пример 1. Конструирование рекомбинантной плазмидной ДНК рЕТ-AsCeGH12b-mut содержащей мутантный ген AsCeGH12b-mut, кодирующий ксилоглюканазу AsCeGH12b-mut
Все стандартные генно-инженерные и микробиологические манипуляции проводят по известным методикам (Green and Sambrook, 2012).
Мутантный ген AsCeGH12b-mut (SEQ ID NO 2) получают путем введения мутаций в ген AsCeGH12b (GenBank: MF527262.1) методом перекрывающихся ПЦР-продуктов (Overlap PCR) с использованием плазмиды pPIC-AsCeGH12b, содержащей ген AsCeGH12b в качестве матрицы [RU 2639248]. Для этого используют следующие праймеры:
прямой - AsCeXgh_for_BamHI - ATAGGATCCGCGACTACGTACTGC и обратный - AsCeXgh_rev_Y37V - AGAGAATTGACCGTGAAGCACTG для получения первого фрагмента размером 576 п.о.;
прямой - AsCeXgh_rev_Sal - ATAGTCGACTTAAGCCACACTGGCCGAG и обратный - AsCeXgh_for_Y37V - AGTGCTTCACGGTCAATTCTCTCTC для получения второго фрагмента размером 129 п.о.
В праймер AsCeXgh for BamHI был введен сайт BamHI, а в праймер AsCeXgh_rev_Sal - сайт SalI. Фрагмент конструируют с таким расчетом, чтобы рамка считывания начиналась с полигистидиновой последовательности.
Плазмиду pET-AsCeGH12b-mut конструируют путем клонирования фрагмента, содержащего ген AsCeGH12b-mut в вектор рЕТ-28а (Фиг. 1).
Гидролиз плазмиды рЕТ-28а и фрагмента ДНК, содержащего ген AsCeGH12b-mut, проводят рестриктазами BamHI и SalI (Thermo Fisher Scientific, США). Очистку полученных фрагментов ДНК проводят с помощью гель-электрофореза в 1,8% агарозном геле с последующим выделением нужной полосы с помощью набора для выделения ДНК из агарозных гелей (Евроген, Россия) согласно инструкции производителя. Лигирование проводят в реакционной смеси объемом 20 мкл, содержащей 20 нг ДНК вектора, 1,5 нг ДНК фрагмента и 5 ед. Т4 ДНК лигазы (Thermo Fisher Scientific, США), согласно методике производителя. Полученной лигазной смесью в количестве 2,5 мкл трансформируют компетентные клетки штамма Е. coli ТОР10 (Invitrogen, США) (генотип - F- mcrA Δ(mrr-hsdRMS-mcrBC) ϕ80lacZAM15 ΔlacX74 nupG recA1 araD139 Δ(ara-leu)7697 galE15 galK16 rpsL(StrR) end Al λ-). Плазмидную ДНК полученных трансформантов анализируют путем гидролиза рестриктазами BamHI и SalI. В результате отбирают клоны, содержащие BamHI/SalI фрагменты размером, равным исходному фрагменту вставки. Правильность клонирования подтверждают секвенированием каждой из рекомбинантных вставок методом Сэнгера на генетическом анализаторе ABI 3500 (Life technology, США). В результате получают рекомбинантную плазмиду рЕТ-AsCeGH12b-mut размером 6007 п. о., содержащую наряду с генами вектора рЕТ-28а, также мутированный ген AsCeGH12b-mut, кодирующий рекомбинантную ксилоглюканазу AsCeGH12b-mut (SEQ ID NO 1) (Фиг. 2).
Пример 2. Конструирование заявляемого штамма-продуцента ксилоглюканазы AsCeGH12b-mut
С целью получения рекомбинантного штамма Е. coli BL21 (DE3) GOLD/pET-AsCeGH12b-mut, продуцента ксилоглюканазы AsCeGH12b-mut, клетки штамма Е. coli BL21 (DE3) GOLD трансформируют плазмидой pET-AsCeGH12b-mut. Трансформацию проводят 2,5 нг плазмидной ДНК методом химической трансформации по стандартной методике [Green M.R., Sambrook J. Molecular Cloning: a Laboratory Manual. (Fourth Edition). Cold Spring Harbor Laboratory Press, 2012, 2028 р.]. Селекцию трансформантов проводят на LB-arape, содержащем канамицин (25 мг/л). В результате получают штамм Е. coli, синтезирующий ксилоглюканазу AsCeGH12b-mut, модифицированную на N-конце полигистидиновой последовательностью с целью последующей очистки методом металл-афинной хроматографии.
Штамм Е. coli продуцент ксилоглюканазы AsCeGH12b-mut депонирован во Всероссийской коллекции промышленных микроорганизмов как Escherichia coli ВКПМ В-14321.
Пример 3. Микробиологический синтез рекомбинантной термостабильной ксилоглюканазы AsCeGH12b-mut с использованием заявляемого штамма
Исходным посевным материалом служит культура Е. coli ВКПМ В-14321, выращенная при температуре 37°С в течение 18 часов на среде LB, содержащей 25 мг/л канамицина. Процесс биосинтеза ведут в колбах Эрленмейера объемом 750 мл, содержащих 100 мл среды LB с канамицином (25 мг/л). Посевной материал вносят в ферментационную среду в количестве, необходимом для создания его концентрации около 1 об. %. Процесс культивирования продуцента ведут при температуре 30°С на круговой качалке со скоростью вращения 130 об/мин до достижения оптической OD600=0,6-0,8 ОЕ (около 4 часов культивирования), после чего в культуральную жидкость добавляют ИПТГ до концентрации 0,1 мМ. Ферментацию продолжают при 90 об/мин и 29°С в течение 12 часов. Биомассу клеток отделяют центрифугированием при 3000 об/мин, промывают 0,1 М натрий-фосфатным буфером и вновь центрифугируют. Контроль экспрессии рекомбинантного гена осуществляют с помощью денатурирующего электрофореза в полиакриламидном геле.
Полученную биомассу ресуспендируют в 50 мМ фосфатном буфере, рН 8.0, содержащем 150 мМ хлорида натрия и 10 мМ имидазола. Клетки разрушают обработкой ультразвуком, проводя процедуру на льду. Целые клетки и нерастворимую белковую фракцию отделяют центрифугированием (16000 об/мин, 25 мин). К нерастворимой фракции, содержащей тельца включения, добавляют 50 мМ фосфатный буфер, рН 8.0, содержащий 150 мМ хлорида натрия и 10 мМ имидазола, ресуспендируют и инкубируют сутки при 4°С для перевода рекомбинантного белка, содержащегося в тельцах включения, в растворимую форму. Нерастворимую фракцию отделяют центрифугированием (16000 об/мин, 25 мин). Супернатанты, полученные после разрушения клеток и отмывки телец включения, объединяют, к раствору белка добавляют равный объем 50% суспензии Ni-NTA агарозы (Qiagen, EU), предварительно уравновешенной вышеуказанным буфером, и интенсивно перемешивают в течение 1 часа. Хроматографическую колонку объемом 10 мл заполняют полученной суспензией, промывают буфером (50 мМ фосфатный буфер, 150 мМ хлорида натрия, 20 мМ имидазола) и элюируют целевой белок с колонки буфером (50 мМ фосфатный буфер, 150 мМ хлорида натрия, 400 мМ имидазола). Активность в полученных образцах определяют с помощью метода с динитросалициловым реактивом [ГОСТ 31662-2012. Препараты ферментные. Методы определения ферментативной активности целлюлазы. Москва, 2012. 9 с.] по скорости гидролиза ксилоглюкана в следующих условиях: рН 5.0, температура 50°С, концентрация субстрата - 0,5 г/л. Рекомбинантная ксилоглюканаза AsCeGH12b-mut, продуцируемая штаммом Е. coli ВКПМ В-14321, проявляет максимум активности при 60°С, рН 5.0. Дикий вариант фермента AsCeGH12b, сконструированный и очищенный аналогичным образом, а также ближайший аналог - рекомбинантная ксилоглюканаза AsCeGH12b [RU 2625013; Rykov S.V., Kornberger P., Herlet J., Tsurin N.V., Zorov I.N., Zverlov V.V., Liebl W., Schwarz W.H., Yarotsky S.V., Berezina O.V. Novel endo-(1,4)-β-glucanase Bgh12A and xyloglucanase Xgh12B from Aspergillus cervinus belong to GH12 subgroup I and II, respectively. Appl. Microbiol. Biotechnol, 2019, 103(18), 7553-7566], имеют более низкий максимум активности - 55°С, рН 5.0. При инкубации препарата фермента AsCeGH12b-mut в течение 10 минут при 65 и 70°С рН 5.0 сохраняется 80% и 28% ферментативной активности соответственно. В таких же условиях у дикого варианта фермента AsCeGH12b, сконструированного и очищенного аналогичным образом, сохраняется 30% и 12% активности, соответственно. Полученный фермент AsCeGH12b-mut является более термостабильным и имеет более высокий максимум активности по сравнению с диким вариантом AsCeGH12b, сконструированным и очищенным аналогичным образом, и ближайшим аналогом.
Таким образом, получен рекомбинантный штамм Escherichia coli, способный к гетерологичному биосинтезу ксилоглюканазы семейства GH12 из A. cervinus, кодируемой мутантным геном. Получена рекомбинантная ксилоглюканаза AsCeGH12b-mut, которая отличается от фермента дикого типа более высокими температурным максимумом активности и более высокой термостабильностью в интервале температур 60-70°С. При культивировании заявляемым способом уровень синтеза целевого продукта у заявляемого штамма (≥500 Ед/г сырой биомассы) превышает таковой у ближайшего аналога (343 Ед/г сырой биомассы). Также, в отличие от ближайшего аналога, продуцируемый фермент имеет более высокий температурный максимум активности и более эффективно гидролизует ксилоглюкан при температурах 55-70°С. Разработан способ микробиологического синтеза ксилоглюканазы AsCeGH12b-mut на основе штамма Escherichia coli, отличающийся тем, что в качестве продуцента используют заявляемый рекомбинантный штамм, а целевым продуктом является мутантная ксилоглюканаза AsCeGH12b-mut, обладающая повышенной по сравнению с ферментом дикого типа термостабильностью.
--->
<?xml version="1.0" encoding="UTF-8"?>
<!DOCTYPE ST26SequenceListing PUBLIC "-//WIPO//DTD Sequence Listing
1.3//EN" "ST26SequenceListing_V1_3.dtd">
<ST26SequenceListing dtdVersion="V1_3" fileName="МУТАНТНАЯ ФОРМА
КСИЛОГЛЮКАНАЗЫ ИЗ ASPERGILLUS CERVINUS C ПОВЫШЕННОЙ ПО СРАВНЕНИЮ С
ФЕРМЕНТОМ ДИКОГО ТИПА ТЕРМОСТАБИЛЬНОСТЬЮ, РЕКОМБИНАНТНЫЙ ШТАММ
ESCHERICHIA COLI.xml" softwareName="WIPO Sequence"
softwareVersion="2.2.0" productionDate="2023-01-25">
<ApplicationIdentification>
<IPOfficeCode>RU</IPOfficeCode>
<ApplicationNumberText>000001</ApplicationNumberText>
<FilingDate>2022-12-07</FilingDate>
</ApplicationIdentification>
<ApplicantFileReference>000001</ApplicantFileReference>
<ApplicantName languageCode="ru">Национальный исследовательский
центр «Курчатовский институт» </ApplicantName>
<ApplicantNameLatin>National Research Center Kurchatov
Institute</ApplicantNameLatin>
<InventorName languageCode="ru">Селимзянова Алина
Ильдаровна</InventorName>
<InventorNameLatin>Selimzanova Alina Ildarovna</InventorNameLatin>
<InventionTitle languageCode="ru">МУТАНТНАЯ ФОРМА КСИЛОГЛЮКАНАЗЫ ИЗ
ASPERGILLUS CERVINUS C ПОВЫШЕННОЙ ПО СРАВНЕНИЮ С ФЕРМЕНТОМ ДИКОГО
ТИПА ТЕРМОСТАБИЛЬНОСТЬЮ, РЕКОМБИНАНТНЫЙ ШТАММ ESCHERICHIA COLI,
ПРОДУЦИРУЮЩИЙ ЭТУ КСИЛОГЛЮКАНАЗУ, И СПОСОБ МИКРОБИОЛОГИЧЕСКОГО
СИНТЕЗА МУТАНТНОЙ КСИЛОГЛЮКАНАЗЫ ИЗ ASPERGILLUS CERVINUS НА ОСНОВЕ
ЭТОГО ШТАММА</InventionTitle>
<SequenceTotalQuantity>4</SequenceTotalQuantity>
<SequenceData sequenceIDNumber="1">
<INSDSeq>
<INSDSeq_length>250</INSDSeq_length>
<INSDSeq_moltype>AA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..250</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>protein</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q2">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>MGSSHHHHHHSSGLVPRGSHMENLYFQSGSATTYCGQWDSVTEGNYILY
NDLWGESAATSGSQCFTVNSLSGNTLSWSTSWTWAGGSSSVKSFANAALQFTPKQLSSISSIDTTWTWSY
TGTSIVADVAYDMFLAASASGSSEYEIMVWLAALGGAGPISSTGSTIATPSIAGQTWKLYSGPNGATTVY
SFVAESETTSFSADLMDFYTYLIDNQGLSSALYLTNVQAGTEPFTGSNAVLTVSSYSASVA</INSDSeq
_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="2">
<INSDSeq>
<INSDSeq_length>753</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..753</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q4">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>atgggcagcagccatcatcatcatcatcacagcagcggcctggtgccgc
gcggcagccatatggagaatctttattttcagtctggatccgcgactacgtactgcggccaatgggactc
agtgactgagggcaactacatcctctataatgacctctggggcgaatccgctgctaccagcggctcccag
tgcttcacggtcaattctctctccggaaacacactctcctggagtacctcctggacctgggccggaggct
caagcagcgtgaagagctttgccaatgccgctcttcagtttaccccgaaacagttgagcagtattagtag
tattgatactacgtggacgtggagctacaccggcacctccattgtcgccgacgtcgcctatgacatgttc
ctagccgctagcgcaagcggatctagcgagtacgaaatcatggtctggcttgctgcgttgggcggcgcag
gcccgatctcgtctaccggctcgaccatcgcgactcccagcattgcagggcagacgtggaaactgtactc
gggcccgaacggcgccacgacggtgtacagtttcgtggcagagtctgagacgacgagcttctcggctgat
ctgatggacttctacacgtatttgattgataatcaggggctgtcgtcggcgttgtatttgactaatgtgc
aggcgggtacggagccatttaccggaagcaatgcggtgctgacggtgtcatcgtactcggccagtgtggc
ttaa</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="3">
<INSDSeq>
<INSDSeq_length>220</INSDSeq_length>
<INSDSeq_moltype>AA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..220</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>protein</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q6">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>Aspergillus
cervinus</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>ATTYCGQWDSVTEGNYILYNDLWGESAATSGSQCFTYNSLSGNTLSWST
SWTWAGGSSSVKSFANAALQFTPKQLSSISSIDTTWTWSYTGTSIVADVAYDMFLAASASGSSEYEIMVW
LAALGGAGPISSTGSTIATPSIAGQTWKLYSGPNGATTVYSFVAESETTSFSADLMDFYTYLIDNQGLSS
ALYLTNVQAGTEPFTGSNAVLTVSSYSASVA</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="4">
<INSDSeq>
<INSDSeq_length>6007</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..6007</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q8">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>tggcgaatgggacgcgccctgtagcggcgcattaagcgcggcgggtgtg
gtggttacgcgcagcgtgaccgctacacttgccagcgccctagcgcccgctcctttcgctttcttccctt
cctttctcgccacgttcgccggctttccccgtcaagctctaaatcgggggctccctttagggttccgatt
tagtgctttacggcacctcgaccccaaaaaacttgattagggtgatggttcacgtagtgggccatcgccc
tgatagacggtttttcgccctttgacgttggagtccacgttctttaatagtggactcttgttccaaactg
gaacaacactcaaccctatctcggtctattcttttgatttataagggattttgccgatttcggcctattg
gttaaaaaatgagctgatttaacaaaaatttaacgcgaattttaacaaaatattaacgtttacaatttca
ggtggcacttttcggggaaatgtgcgcggaacccctatttgtttatttttctaaatacattcaaatatgt
atccgctcatgaattaattcttagaaaaactcatcgagcatcaaatgaaactgcaatttattcatatcag
gattatcaataccatatttttgaaaaagccgtttctgtaatgaaggagaaaactcaccgaggcagttcca
taggatggcaagatcctggtatcggtctgcgattccgactcgtccaacatcaatacaacctattaatttc
ccctcgtcaaaaataaggttatcaagtgagaaatcaccatgagtgacgactgaatccggtgagaatggca
aaagtttatgcatttctttccagacttgttcaacaggccagccattacgctcgtcatcaaaatcactcgc
atcaaccaaaccgttattcattcgtgattgcgcctgagcgagacgaaatacgcgatcgctgttaaaagga
caattacaaacaggaatcgaatgcaaccggcgcaggaacactgccagcgcatcaacaatattttcacctg
aatcaggatattcttctaatacctggaatgctgttttcccggggatcgcagtggtgagtaaccatgcatc
atcaggagtacggataaaatgcttgatggtcggaagaggcataaattccgtcagccagtttagtctgacc
atctcatctgtaacatcattggcaacgctacctttgccatgtttcagaaacaactctggcgcatcgggct
tcccatacaatcgatagattgtcgcacctgattgcccgacattatcgcgagcccatttatacccatataa
atcagcatccatgttggaatttaatcgcggcctagagcaagacgtttcccgttgaatatggctcataaca
ccccttgtattactgtttatgtaagcagacagttttattgttcatgaccaaaatcccttaacgtgagttt
tcgttccactgagcgtcagaccccgtagaaaagatcaaaggatcttcttgagatcctttttttctgcgcg
taatctgctgcttgcaaacaaaaaaaccaccgctaccagcggtggtttgtttgccggatcaagagctacc
aactctttttccgaaggtaactggcttcagcagagcgcagataccaaatactgtccttctagtgtagccg
tagttaggccaccacttcaagaactctgtagcaccgcctacatacctcgctctgctaatcctgttaccag
tggctgctgccagtggcgataagtcgtgtcttaccgggttggactcaagacgatagttaccggataaggc
gcagcggtcgggctgaacggggggttcgtgcacacagcccagcttggagcgaacgacctacaccgaactg
agatacctacagcgtgagctatgagaaagcgccacgcttcccgaagggagaaaggcggacaggtatccgg
taagcggcagggtcggaacaggagagcgcacgagggagcttccagggggaaacgcctggtatctttatag
tcctgtcgggtttcgccacctctgacttgagcgtcgatttttgtgatgctcgtcaggggggcggagccta
tggaaaaacgccagcaacgcggcctttttacggttcctggccttttgctggccttttgctcacatgttct
ttcctgcgttatcccctgattctgtggataaccgtattaccgcctttgagtgagctgataccgctcgccg
cagccgaacgaccgagcgcagcgagtcagtgagcgaggaagcggaagagcgcctgatgcggtattttctc
cttacgcatctgtgcggtatttcacaccgcatatatggtgcactctcagtacaatctgctctgatgccgc
atagttaagccagtatacactccgctatcgctacgtgactgggtcatggctgcgccccgacacccgccaa
cacccgctgacgcgccctgacgggcttgtctgctcccggcatccgcttacagacaagctgtgaccgtctc
cgggagctgcatgtgtcagaggttttcaccgtcatcaccgaaacgcgcgaggcagctgcggtaaagctca
tcagcgtggtcgtgaagcgattcacagatgtctgcctgttcatccgcgtccagctcgttgagtttctcca
gaagcgttaatgtctggcttctgataaagcgggccatgttaagggcggttttttcctgtttggtcactga
tgcctccgtgtaagggggatttctgttcatgggggtaatgataccgatgaaacgagagaggatgctcacg
atacgggttactgatgatgaacatgcccggttactggaacgttgtgagggtaaacaactggcggtatgga
tgcggcgggaccagagaaaaatcactcagggtcaatgccagcgcttcgttaatacagatgtaggtgttcc
acagggtagccagcagcatcctgcgatgcagatccggaacataatggtgcagggcgctgacttccgcgtt
tccagactttacgaaacacggaaaccgaagaccattcatgttgttgctcaggtcgcagacgttttgcagc
agcagtcgcttcacgttcgctcgcgtatcggtgattcattctgctaaccagtaaggcaaccccgccagcc
tagccgggtcctcaacgacaggagcacgatcatgcgcacccgtggggccgccatgccggcgataatggcc
tgcttctcgccgaaacgtttggtggcgggaccagtgacgaaggcttgagcgagggcgtgcaagattccga
ataccgcaagcgacaggccgatcatcgtcgcgctccagcgaaagcggtcctcgccgaaaatgacccagag
cgctgccggcacctgtcctacgagttgcatgataaagaagacagtcataagtgcggcgacgatagtcatg
ccccgcgcccaccggaaggagctgactgggttgaaggctctcaagggcatcggtcgagatcccggtgcct
aatgagtgagctaacttacattaattgcgttgcgctcactgcccgctttccagtcgggaaacctgtcgtg
ccagctgcattaatgaatcggccaacgcgcggggagaggcggtttgcgtattgggcgccagggtggtttt
tcttttcaccagtgagacgggcaacagctgattgcccttcaccgcctggccctgagagagttgcagcaag
cggtccacgctggtttgccccagcaggcgaaaatcctgtttgatggtggttaacggcgggatataacatg
agctgtcttcggtatcgtcgtatcccactaccgagatatccgcaccaacgcgcagcccggactcggtaat
ggcgcgcattgcgcccagcgccatctgatcgttggcaaccagcatcgcagtgggaacgatgccctcattc
agcatttgcatggtttgttgaaaaccggacatggcactccagtcgccttcccgttccgctatcggctgaa
tttgattgcgagtgagatatttatgccagccagccagacgcagacgcgccgagacagaacttaatgggcc
cgctaacagcgcgatttgctggtgacccaatgcgaccagatgctccacgcccagtcgcgtaccgtcttca
tgggagaaaataatactgttgatgggtgtctggtcagagacatcaagaaataacgccggaacattagtgc
aggcagcttccacagcaatggcatcctggtcatccagcggatagttaatgatcagcccactgacgcgttg
cgcgagaagattgtgcaccgccgctttacaggcttcgacgccgcttcgttctaccatcgacaccaccacg
ctggcacccagttgatcggcgcgagatttaatcgccgcgacaatttgcgacggcgcgtgcagggccagac
tggaggtggcaacgccaatcagcaacgactgtttgcccgccagttgttgtgccacgcggttgggaatgta
attcagctccgccatcgccgcttccactttttcccgcgttttcgcagaaacgtggctggcctggttcacc
acgcgggaaacggtctgataagagacaccggcatactctgcgacatcgtataacgttactggtttcacat
tcaccaccctgaattgactctcttccgggcgctatcatgccataccgcgaaaggttttgcgccattcgat
ggtgtccgggatctcgacgctctcccttatgcgactcctgcattaggaagcagcccagtagtaggttgag
gccgttgagcaccgccgccgcaaggaatggtgcatgcaaggagatggcgcccaacagtcccccggccacg
gggcctgccaccatacccacgccgaaacaagcgctcatgagcccgaagtggcgagcccgatcttccccat
cggtgatgtcggcgatataggcgccagcaaccgcacctgtggcgccggtgatgccggccacgatgcgtcc
ggcgtagaggatcgagatctcgatcccgcgaaattaatacgactcactataggggaattgtgagcggata
acaattcccctctagaaataattttgtttaactttaagaaggagatataccatgggcagcagccatcatc
atcatcatcacagcagcggcctggtgccgcgcggcagccatatggagaatctttattttcagtctggatc
cgcgactacgtactgcggccaatgggactcagtgactgagggcaactacatcctctataatgacctctgg
ggcgaatccgctgctaccagcggctcccagtgcttcacggtcaattctctctccggaaacacactctcct
ggagtacctcctggacctgggccggaggctcaagcagcgtgaagagctttgccaatgccgctcttcagtt
taccccgaaacagttgagcagtattagtagtattgatactacgtggacgtggagctacaccggcacctcc
attgtcgccgacgtcgcctatgacatgttcctagccgctagcgcaagcggatctagcgagtacgaaatca
tggtctggcttgctgcgttgggcggcgcaggcccgatctcgtctaccggctcgaccatcgcgactcccag
cattgcagggcagacgtggaaactgtactcgggcccgaacggcgccacgacggtgtacagtttcgtggca
gagtctgagacgacgagcttctcggctgatctgatggacttctacacgtatttgattgataatcaggggc
tgtcgtcggcgttgtatttgactaatgtgcaggcgggtacggagccatttaccggaagcaatgcggtgct
gacggtgtcatcgtactcggccagtgtggcttaagtcgacaagcttgcggccgcactcgagcaccaccac
caccaccactgagatccggctgctaacaaagcccgaaaggaagctgagttggctgctgccaccgctgagc
aataactagcataaccccttggggcctctaaacgggtcttgaggggttttttgctgaaaggaggaactat
atccggat</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
</ST26SequenceListing>
<---
Изобретение относится к биотехнологии. Предложена мутантная ксилоглюканаза AsCeGH12b-mut, имеющая аминокислотную последовательность SEQ ID NO 1, которая соответствует аминокислотной последовательности ксилоглюканазы семейства 12 дикого типа из Aspergillus cervinus ВКПМ F-612, с заменой природного остатка тирозина в положении 37 остатком валина, и обладающая повышенными термостабильностью и температурным максимумом активности по сравнению с ферментом дикого типа. Также предложены рекомбинантный штамм Escherichia coli ВКПМ В-14321, продуцирующий указанную ксилоглюканазу AsCeGH12b-mut, и способ микробиологического синтеза указанной ксилоглюканазы с использованием указанного штамма. Изобретение обеспечивает расширение арсенала рекомбинантных ксилоглюканаз, штаммов-продуцентов и способов микробиологического синтеза рекомбинантных ксилоглюканаз. 3 н.п. ф-лы, 2 ил., 3 пр.
1. Рекомбинантная мутантная ксилоглюканаза AsCeGH12b-mut, имеющая аминокислотную последовательность SEQ ID NO 1, которая соответствует аминокислотной последовательности ксилоглюканазы семейства 12 дикого типа из Aspergillus cervinus ВКПМ F-612, с заменой природного остатка тирозина в положении 37 остатком валина, и обладающая повышенными термостабильностью и температурным максимумом активности по сравнению с ферментом дикого типа.
2. Рекомбинантный штамм Escherichia coli ВКПМ В-14321 - продуцент ксилоглюканазы AsCeGH12b-mut по п. 1, имеющей SEQ ID NO 1 и кодируемой геном AsCeGH12b-mut, имеющим SEQ ID NO 2.
3. Способ микробиологического синтеза рекомбинантной мутантной ксилоглюканазы AsCeGH12b-mut по п. 1, предусматривающий культивирование рекомбинантного штамма бактерии Escherichia coli по п. 2 в аэробных условиях в подходящей питательной среде до максимального накопления целевого продукта.
| Рекомбинантный штамм Escherichia coli - продуцент ксилоглюканазы из гриба Aspergillus cervinus и способ микробиологического синтеза ксилоглюканазы на основе этого штамма | 2016 |
|
RU2625013C1 |
| Рельсовое стыковое скрепление | 1922 |
|
SU461A1 |
| Способ гальванического снятия позолоты с серебряных изделий без заметного изменения их формы | 1923 |
|
SU12A1 |
| RYKOV S.V | |||
| ET AL | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |

