Группа изобретений относится к биотехнологии, в частности к биосинтезу ксилоглюканазы, и представляет собой рекомбинантный штамм Escherichia coli, способный синтезировать рекомбинантную ксилоглюканазу и полученный путем трансформации штамма Escherichia coli BL21-CODONPLUS (DE3)-RIPL плазмидой рЕТ-AsCeGH12b, содержащей ген β-1,4-гликозилгидролазы класса GH12 AsCeGH12b из Aspergillus cervinus ВКПМ F-612 под контролем промотора T7lac.
Растительная биомасса является дешевым и возобновляемым источником сахаров для нужд промышленной биотехнологии. Ферментативный гидролиз - эффективный и экологически безопасный способ конверсии полисахаридов растительной биомассы в сахара. Химическое разнообразие растительных полисахаридов обуславливает необходимость использования широкого спектра ферментов для гидролиза.
Ксилоглюкан - один из основных полисахаридов первичной клеточной стенки двудольных и многих однодольных растений (Hayashi, 1989). Молекулы ксилоглюкана состоят из мономеров 1,4-β-D-глюкотетрозы, в которой три из четырех глюкозных остатка связаны α-1,6 связями с остатками D-ксилозы. Некоторые из остатков глюкозы, в свою очередь, могут быть связаны β-1,2 связями с остатками D-галактозы или L-арабинозы. Остатки D-галактозы могут быть связаны α-1,2 связями с остатками L-фукозы. Модификация остатков ксилозы видоспецифична, что определяет множество типов ксилоглюкана в растениях. В клеточной стенке растений ксилоглюкан выполняет структурную функцию. Ксилоглюкан выступает связующим гликаном, взаимодействуя как с молекулами целлюлозы, так и между собой и формирует прочную полисахаридную сеть. Также ксилоглюкан может играть роль запасного полисахарида, например, в случае с тамариндом индийским Tamarindus indica ксилоглюкан выступает единственным запасным полисахаридом семян (Shankaracharya, 1998).
Ферменты, способные специфически гидролизовать β-1,4-связи глюканового остова молекулы ксилоглюкана, называются ксилоглюканазами. Также известны гликозилгидролазы, которые могут гидролизовать различные связи в молекуле ксилоглюкана: β-галактозидазы, α-ксилозидазы, β-глюкозидазы и α-фукозидазы. Конечными продуктами гидролиза ксилоглюкана ксилоглюканазами является смесь олигосахаридов различного состава и строения (Grishutin et al., 2004). Ксилоглюканазы классифицируются по механизму действия на ксилоглюкановые олигосахариды: экзо-ксилоглюканазы (ЕС 3.2.1.155 и ЕС 3.2.1.150) гидролизуют олигосахариды с редуцирующего конца цепи; эндо-ксилоглюканазы (ксилоглюкан-специфические эндо-β-1,4-глюканазы, ЕС 3.2.1.151) гидролизуют только гликозидную связь у незамещенного остатка глюкозы в молекуле ксилоглюкана и ксилоглюкановых олигосахаридах (Yaoi and Mitsuishi, 2004). Согласно данным базы CAZy (http://www.cazy.org/) ферменты с ксилоглюканазной активностью найдены среди семейств гликозил-гидролаз GH5, GH9, GH12, GH16, GH26, GH44, GH74. Ферменты, специфически гидролизующие ксилоглюкан, также могут проявлять побочную активность на других β-1,4-/1,3-глюканах: растворимой и нерастворимой целлюлозе, β-глюкане, лихенане и ламиранане.
На основе ксилоглюканаз созданы ферментные препараты для ферментативного гидролиза растительных полисахаридов, в особенности биомассы молодых побегов (RU 2358756, RU 2361918). Полный ферментативный гидролиз целлюлозы с помощью только целлюлаз затруднен без предварительного гидролиза связующего ксилоглюкана. Также ксилоглюканазы находят применение как модифицирующие агенты для изменения физических и биохимических свойств растительных волокон (Park et al., 2004), для легирования химически модифицированных олигосахаридов к молекуле ксилоглюкана для изменения функциональности целлюлозных фибрилл (Brumer et al., 2004). Также отмечается возрастающий интерес к применению ксилоглюканаз в пищевой и текстильной индустрии и в фармацевтике (Shankaracharya, 1998; Miyazaki et al., 2003).
В источниках информации имеются сведения о грибах - природных продуцентах ксилоглюканаз: Geotrichum sp. (Yaoi and Mitsuishi, 2004), Aspergillus japonicus, Chrysosporium lucknowense, Trichoderma reesei (Grishutin et al., 2004; Markov et al., 2005; Qi et al., 2013), Phanerochaete chrysosporium (Ishida et al., 2007). Методами селекции и радиационного мутагенеза получены штаммы, синтезирующие повышенные количества ксилоглюканаз: Penicillium verruculosum (RU 2361918), Penicillium funiculosum (RU 2323254), Aspergillus aculeatus (RU 2303057).
Рекомбинантные продуценты ксилоглюканаз обладают рядом несомненных преимуществ, основным из которых является повышенная продуктивность рекомбинантных штаммов по целевому белку. Для гетерологичной экспрессии грибных ксилоглюканаз в качестве штаммов-реципиентов применяют различные виды бактерий, в частности, Acidothermus cellulolyticus использован для гетерологичной экспрессии ксилоглюканаз из Acremonium sp., Cladorrhinum foecundissimum, Humicola insolens, Thielavia terrestris, Trichoderma reesei, Aspergillus aculeatus, Aspergillus aculeatus, Aspergillus fumigatus, Myceliophthora thermophila (Vlasenko et al., 2010). Описаны полученные методами генной инженерии штаммы Е. coli - продуценты рекомбинантных ксилоглюканаз из Geotrichum sp. M128 (US 2004038367; Yaoi and Mitsuishi, 2002), Fusarium graminearum (Habrylo et al., 2012), Rhizomucor miehei (Song et al., 2013).
Аналогом заявляемого штамма является штамм Е. coli BL21-CodonPlus (DE3)-RP содержащий на плазмиде рЕТ-29а ген ксилоглюканазы класса GH74 из Geotrichum sp. strain M128 под контролем промотора Т7 и экспрессирующий гликозил-гидролазу класса GH74, специфически гидролизующую ксилоглюкан (US 20040038367). Фермент из Geotrichum sp. strain M128 имеет большую молекулярную массу (96 кДа), что затрудняет получение физиологически активного фермента с помощью бактериального синтеза. Кроме того фермент из Geotrichum sp. strain M128 обладает дополнительной ферментативной активностью, гидролизуя вторую β-гликозидную связь с редуцированного конца олигосахарида. Способ микробиологического синтеза на основе штамма ближайшего аналога приводит к образованию преимущественно нерастворимых телец включения, что требует проведения стадии рефолдинга для получения физиологически активного фермента.
Задачи заявляемой группы изобретений состоят в расширении арсенала рекомбинантных ксилоглюканаз и в разработке способа микробиологического синтеза заявляемой ксилоглюканазы.
Задачу решают путем:
- конструирования рекомбинантного штамма Escherichia coli ВКПМ В-12683 - продуцента ксилоглюканазы семейства GH12, содержащего ген AsCeGH12b (SEQ ID NO 1), клонированный из гриба Aspergillus cervinus ВКПМ F-612;
- разработки способа микробиологического синтеза ксилоглюканазы, предусматривающего культивирование рекомбинантного штамма бактерии Escherichia coli, содержащего ген ксилоглюканазы, в аэробных условиях в подходящей питательной среде до максимального накопления целевого продукта, отличающийся тем, что в качестве продуцента используют рекомбинантный штамм Escherichia coli ВКПМ В-12683, а целевым продуктом является ксилоглюканаза семейства GH12.
Ген ксилоглюканазы семейства GH12 из Aspergillus cervinus ВКПМ F-612 выделен и клонирован впервые. Последовательность нуклеотидов генома Aspergillus cervinus ВКПМ F-612 неизвестна.
Процесс получения заявляемого штамма состоит из нескольких этапов:
- получения полной нуклеотидной последовательности гена, кодирующего ксилоглюканазу семейства GH12, синтезируемую штаммом Aspergillus cervinus ВКПМ F-612,
- конструирования плазмидной ДНК (плазмиды) pET-AsCeGH12b, содержащей структурную часть гена ксилоглюканазы семейства GH12 из Aspergillus cervinus ВКПМ F-612,
- конструирования заявляемого рекомбинантного штамма Escherichia coli ВКПМ В-12683.
Этап 1. Получение полной нуклеотидной последовательности гена, кодирующего ксилоглюканазу семейства GH12, синтезируемую штаммом Aspergillus cervinus ВКПМ F-612
Штамм Aspergillus cervinus ВКПМ F-612 секретирует в культуральную среду ферменты, способные гидролизовать ксилоглюкан. Методом зимографии определено, что культуральная жидкость штамма Aspergillus cervinus ВКПМ F-612 содержит ксилоглюканазу с молекулярной массой 25 кДа. Известно, что ксилоглюканазную активность у видов царства грибов проявляют ферменты семейств GH12 и GH74, которые отчетливо различают по молекулярному весу (около 25-30 кДа и около 75-85 кДа соответственно). Для получения и клонирования нуклеотидной последовательности гена гликозил-гидролазы класса GH12 из Aspergillus cervinus ВКПМ F-612 проводят амплификацию консервативных участков гена с помощью вырожденных праймеров и методами прогулки по хромосоме получают фланкирующие консервативный участок гена последовательности ДНК (Green and Sambrook, 2012). Вырожденные праймеры подобраны на консервативные мотивы в известных аминокислотных последовательностях ксилоглюкан-специфичных гликозил-гидролаз класса GH12 представителей рода Aspergillus. Для определения фланкирующих последовательностей применяют два метода прогулки по хромосоме: FPNI-PCR (Fusion primer and nested integrated PCR) для определения последовательности геномной ДНК и RACE (Rapid Amplification of cDNA Ends) для определения последовательности матричной РНК (Wang et al., 2012). В результате получают последовательность нуклеотидов гена ксилоглюканазы класса GH12 из Aspergillus cervinus ВКПМ F-612, обозначенную как AsCeGH12b.
Этап 2. Конструирование плазмидной ДНК (плазмиды) pET-AsCeGH12b, содержащей процессированный вариант гена ксилоглюканазы AsCeGH12b
Плазмиду pET-AsCeGH12b конструируют путем клонирования полученного амплификацией фрагмента ДНК, содержащего структурную часть гена AsCeGH12b, в вектор рЕТ-24а, разработанный для индуцируемой высокоэффективной экспрессии гомологичных и гетерологичных белков в Escherichia coli. Вектор рЕТ-24а размером 5310 пар оснований содержит промотор фага Т7; терминатор фага Т7; сайт множественного клонирования с кодирующей полигистидиновый хвост последовательностью на 3'-конце; кодирующую область гена lacI; кодирующую область гена kan, обеспечивающего устойчивость штаммов Е. coli к канамицину; репликон ColE1 (pBR-322).
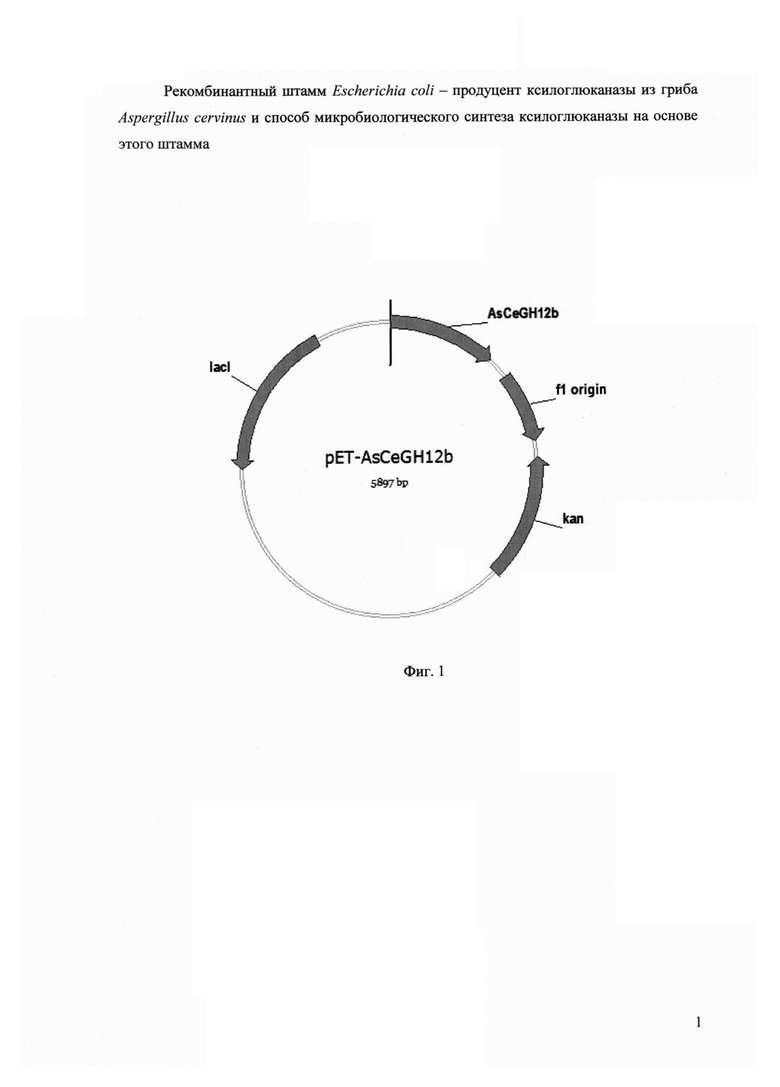
Сконструированная плазмида pET-AsCeGH12b, размером 5897 пар оснований, наряду с генами вектора рЕТ-24а, содержит кодирующую область гена AsCeGH12b ксилоглюканазы под контролем промотора T7lac (Фиг. 1).
Этап 3. Получение заявляемого рекомбинантного штамма Escherichia coli ВКПМ-12683.
В качестве штамма-реципиента используют штамм Escherichia coli BL21-CODONPLUS (DE3)-RIPL (Agilent Technologies). Компетентные клетки данного штамма трансформируют плазмидой pET-AsCeGH12b. В результате получают заявляемый рекомбинантный штамм Escherichia coli BL21-CODONPLUS (DE3)-RIPL//pET-AsCeGH12b, способный синтезировать ксилоглюканазу семейства GH12 из Aspergillus cervinus ВКПМ F-612.
Заявляемый штамм Escherichia coli депонирован во Всероссийской коллекции промышленных микроорганизмов как Escherichia coli ВКПМ В-12683.
Морфологические и физиолого-биохимические характеристики заявляемого штамма.
Заявляемый штамм Escherichia coli ВКПМ В-12683 имеет следующие морфологические и физиолого-биохимические характеристики:
Морфологические признаки: Клетки палочковидной формы, грамотрицательные, неспороносные.
Культуральные признаки: клетки хорошо растут на обычно используемых питательных средах. На агаризованной LB-среде (вес.%: триптон - 1, дрожжевой экстракт - 0,5, хлорид натрия - 1, агар - 1,5, остальное - вода) образуются гладкие, серые, блестящие, круглые с ровным краем колонии. Рост в жидкой среде LB характеризуется равномерным помутнением среды.
Физиолого-биохимические признаки: клетки штамма продуцента растут в диапазоне температур 20-42°С, оптимум составляет 37°С. Наиболее благоприятные для роста значения pH находятся в интервале 6,8-7,2. При росте в аэробных условиях культура может усваивать азот как органических соединений (пептон, триптон, аминокислоты, дрожжевой экстракт), так и аммонийных и нитратных солей. Углерод усваивается в форме углеводов и аминокислот. Устойчивость к антибиотикам. Клетки проявляют устойчивость к канамицину (до 50 мкг/мл), обусловленную наличием в плазмиде pET-AsCeGH12b гена kan, а также тетрациклину (до 20 мкг/мл), хлорамфениколу (до 34 мкг/мл) и стрептомицину (до 25 мкг/мл).
Штамм Escherichia coli ВКПМ В-12683 синтезирует рекомбинантную ксилоглюканазу AsCeGH12b.
Способ микробиологического синтеза ксилоглюканазы AsCeGH12b в общем виде.
Посевной материал, представляющий собой клетки рекомбинантного штамма-продуцента Escherichia coli ВКПМ В-12683, подготавливают путем инкубации в течение 16-18 часов при температуре 30-37°C с аэрацией на среде LB, содержащей 25 мг/мл канамицина, 34 мкг/мл хлорамфеникола и 25 мкг/мл стрептомицина. Выросшую культуру переносят в соотношении 1:100 (по объему) в среду LB, содержащую 25 мг/мл канамицина, 34 мкг/мл хлорамфеникола и 25 мкг/мл стрептомицина. Процесс культивирования ведут при температуре 30-37°C с аэрацией. Синтез целевого белка индуцируют путем добавления индуктора изопропил-β-D-1-тиогалактопиранозида (ИПТГ) до конечной концентрации 0,1 мМ при OD600=0,8-1,0 ОЕ. Биомассу клеток отделяют центрифугированием при об/мин и дважды промывают натрий-фосфатный буфером (вес. %: NaCl - 0,8, KCl - 0,02, Na2HPO4 - 0,15, KH2PO4 - 0,024, остальное - вода). Выделение ксилоглюканазы проводят методом металл-афинной хроматографии (Green and Sambrook, 2012). Уровень синтеза целевого продукта определяют с помощью денатурирующего электрофореза в полиакриламидном геле (Green and Sambrook, 2012). Количество целевого белка составляет: в виде телец включения - 10 и более процентов от суммарного белка клетки в зависимости от количества индуктора, и не менее 330 единиц активности физиологически активного белка на 1 грамм влажной биомассы.
Фигура 1. Рекомбинантная плазмида pET-AsCeGH12b.
Пример 1. Определение полной нуклеотидной последовательности гена ксилоглюканазы AsCeGH12b
Для определения полной нуклеотидной последовательности гена гликозил-гидролазы класса GH12 из Aspergillus cervinus ВКПМ F-612 проводят амплификацию консервативных участков последовательности гена с помощью вырожденных праймеров. Методами прогулки по хромосоме получают фланкирующие консервативный участок гена последовательности ДНК. Вырожденные праймеры подбирают на консервативные мотивы в аминокислотных последовательностях белков класса GH12, охарактеризованных как ксилоглюкан-специфичные гликозил-гидролазы, описанные у представителей рода Aspergillus: A. clavatus, A. fumigatus, A. aculeatus, A. niger, A.flavus, А. terreus, A. nidulans. Амплификацию проводят методом ПЦР с помощью высокоточной полимеразы Phusion (Thermo Scientific) по протоколам производителя. Геномную ДНК выделяют по стандартной методике (Green and Sambrook, 2012). В качестве матрицы используют геномную ДНК штамма Aspergillus cervinus ВКПМ F-612. Амплификация с применением полученных вырожденных праймеров: прямого - F_140_gr.2 - GCYYTGGGCGGYGCKGGHCC, и обратного - R_230_gr.2 - GAAKGGCTCRGTWCCGGCCTG, приводит к получению продукта ПЦР с длиной 280 пар оснований. Полученный фрагмент очищают с помощью гель-электрофореза в 1,8% агаровом геле с последующим выделением нужной полосы с помощью набора для выделения ДНК из агарозных гелей (Биосилика, Россия). Полученный амплификат секвенируют методом Сэнгера на генетическом анализаторе ABI 3500 (Life technology, США) и проверяют последовательность на соответствие ожидаемому классу ферментов поиском по гомологии с помощью BLAST.
На основе полученной последовательности синтезируют серию праймеров, элонгация которых приводит к амплификации ДНК фланговых последовательностей, лежащих за пределами полученного фрагмента. Для определения фланкирующих последовательностей применяют два метода прогулки по хромосоме: FPNI-PCR для определения последовательности геномной ДНК и RACE для определения последовательности матричной РНК (Wang et al., 2012; Lukyanov et al., 1999; Matz et al., 1999). Синтезируемые праймеры подбирают с таким расчетом, чтобы соответствовать требованиям обоих методов: длина 20-25 пар оснований, температура плавления 58-62 С, GC состав не более 50%. Получают праймеры: 3'for_1 - AGCATTGCAGGGCAGACGTGG, 3'for_2 - CACGACGGTGTACAGTTTCGTGG, 3'for_3 - AGCTTCTCGGCTGATCTGATGG, 5'rev_1 - ACGTGTAGAAGTCCATCAGATCAGCC, 5'rev_2 - CACGAAACTGTACACCGTCGTGG, 5'rev_3 - CGAGTACAGTTTCCACGTCTGCC. Метод FPNI-PCR проводят на матрице геномной ДНК штамма Aspergillus cervinus ВКПМ F-612 согласно методике (Wang et al., 2012). В результате получают продукты амплификации, приходящиеся на фланкирующие последовательности геномной ДНК в направлении 3' начала гена и 5' конца гена. Метод RACE проводят на матрице кДНК, полученной с генетического материала культуры штамма Aspergillus cervinus ВКПМ F-612, индуцированной добавлением ксилоглюкана в культуральную среду. Метод RACE осуществляют с использованием набора RACE Primer Set (Евроген, Россия) по протоколу производителя. кДНК получают синтезом одно- и двухцепочечных ДНК с матрицы тотальной РНК методом обратной транскрипции с использованием набора реагентов Mint (Евроген, Россия) по протоколам производителя. Выделение тотальной РНК осуществляют с помощью реагента ExtractRNA (Евроген, Россия) по протоколам производителя. Амплификацию проводят методом ПЦР с помощью высокоточной полимеразы Phusion (Thermo Scientific) по протоколам производителя. В результате получают продукты амплификации, приходящиеся на фланкирующие последовательности матричной РНК в направлении 3' начала гена и 5' конца гена. Полученные методами FPNI-PCR и RACE продукты амплификации секвенируют методом Сэнгера на генетическом анализаторе ABI 3500 (Life technology, США) и собирают единую последовательность ДНК. В результате получена последовательность ДНК длиной 908 пар оснований, которая содержит ген ксилоглюканазы класса GH12, синтезируемую штаммом Aspergillus cervinus. Ген ксилоглюканазы класса GH12 длиной 768 пар оснований назван AsCeGH12b и содержит один интрон длиной 53 пары оснований и два экзона длиной 314 и 400 пар оснований. Процессированный транскрипт гена AsCeGH12b длиной 714 пар нуклеотидов кодирует белок ASCEGH12B (SEQ ID NO 1). Белок ASCEGH12B состоит из 238 аминокислотных остатков и содержит на N-конце предсказанный программой SignalP 4.1 (http://www.cbs.dtu.dk/services/SignalP/) сигнальный пептид длиной 17 аминокислотных остатков (SEQ ID NO 2). Процессированный секретируемый зрелый белок имеет массу 24,5 кДа. Поиск по гомологии аминокислотной последовательности с помощью программы BLAST выявляет ближайшего гомолога белка AsCeGH12b - вероятную ксилоглюкан-специфичную эндо-бета-1,4-глюканазу из Aspergillus lentulus с идентичностью 72%.
Пример 2. Конструирование рекомбинантной плазмидной ДНК рЕТ-AsCeGH12b, содержащей процессированный вариант гена ксилоглюканазы AsCeGH12b
Все стандартные генно-инженерные и микробиологические манипуляции проводят по известным методикам (Green and Sambrook, 2012).
Для получения полноразмерной, не содержащей интронов и сигнального пептида, кодирующей части гена AsCeGH12b проводят амплификацию на матрице кДНК, полученной с генетического материала культуры штамма Aspergillus cervinus ВКПМ F-612, индуцированной добавлением ксилоглюкана в культуральную среду. кДНК получают синтезом одно- и двухцепочечных ДНК с матрицы тотальной РНК методом обратной транскрипции с использованием набора реагентов Mint (Евроген, Россия) по протоколам производителя. Выделение тотальной РНК производят с помощью реагента ExtractRNA (Евроген, Россия) по протоколам производителя. Амплификацию фрагмента, содержащего ген AsCeGH12b, проводят методом ПЦР с помощью высокоточной полимеразы Phusion (Thermo Scientific) по протоколам производителя с использованием следующих праймеров: прямого - SP_AsCeGH12B_NdeI - TTACATATGGCGACTACGTACTGCGGCCA, и обратного - ASP_AsCeGH12B_XhoI_his_tag - TAACTCGAGAGCCACACTGGCCGAGTACG. В прямой праймер был введен сайт NdeI, а в обратный - сайт XhoI. Фрагмент конструируют с таким расчетом, чтобы рамка считывания продолжилась полигистидиновой последовательностью на С-конце рекомбинантного белка, кодируемого последовательностью на векторе рЕТ-24а. Очистку полученного амплификацией фрагмента ДНК проводят с помощью гель-электрофореза в 1,8% агаровом геле с последующим выделением нужной полосы с помощью набора для выделения ДНК из агарозных гелей (Биосилика, Россия).
Плазмиду pET-AsCeGH12b конструируют путем клонирования фрагмента, содержащего ген AsCeGH12b в вектор рЕТ-24а.
Гидролиз плазмиды рЕТ-24а и фрагмента ДНК, содержащего ген AsCeGH12b, проводят рестриктазами NdeI и XhoI (Thermo scientific, EU). Очистку полученных фрагментов ДНК проводят с помощью гель-электрофореза в 1,8% агаровом геле с последующим выделением нужной полосы с помощью набора для выделения ДНК из агарозных гелей (Биосилика, Россия). Легирование проводят в реакционной смеси объемом 20 мкл, содержащей 20 нг ДНК вектора, 20 нг ДНК фрагмента и 5 ед. Т4 ДНК лигазы (СибЭнзим, Россия), согласно методике производителя. Полученной лигазной смесью в количестве 5 мкл трансформируют компетентные клетки штамма Escherichia coli XL1 blue MRF' (Stratagene, США) (генотип - Δ(mcrA)183 Δ(mcrCB-hsdSMR-mrr)173 endA1 supE44 thi-1 recA1 gyrA96 relA1 lac [F' proAB lacIqZΔM15 Tn10 (Tetr)]). Плазмидную ДНК полученных трансформантов анализируют путем гидролиза рестриктазами NdeI и XhoI. В результате отбирают клоны, содержащие NdeI/XhoI фрагменты размером, равным исходному фрагменту вставки. Правильность клонирования подтверждают секвенированием вставки методом Сэнгера на генетическом анализаторе ABI 3500 (Life technology, США).
В конечном итоге получают рекомбинантную плазмиду pET-AsCeGH12b размером 5897пар оснований, содержащую наряду с генами вектора рЕТ-24а, также ген AsCeGH12b, кодирующий ксилоглюканазу семейства GH12 из Aspergillus cervinus ВКПМ F-612 (SEQ ID NO 3) (Фиг. 1).
Пример 3. Конструирование заявляемого штамма-продуцента ксилоглюканазы AsCeGH12b
С целью получения рекомбинантного штамма Escherichia coli BL21-CODONPLUS (DE3)-RIPL//pET-AsCeGH12b продуцента ксилоглюканазы, клетки штамма Escherichia coli BL21-CODONPLUS (DE3)-RIPL трансформируют плазмидой pET-AsCeGH12b. Трансформацию проводят 10 нг плазмидной ДНК методом химической трансформации по стандартной методике (Green and Sambrook, 2012). Селекцию трансформантов проводят на LB-агаре, содержащем канамицин (25 мг/л), хлорамфеникол (34 мкг/мл) и стрептомицин (25 мкг/мл). В результате получают штамм Escherichia coli, синтезирующий ксилоглюканазу AsCeGH12b, модифицированную на N-конце полигистидиновым «хвостом». His-Tag с целью последующей очистки методом металл-афинной хроматографии.
Штамм Escherichia coli продуцент ксилоглюканазы AsCeGH12b депонирован во Всероссийской коллекции промышленных микроорганизмов как Escherichia coli ВКПМ В-12683.
Пример 4. Микробиологический синтез рекомбинантной ксилоглюканазы AsCeGH12b с использованием заявляемого штамма
Исходным посевным материалом служит культура Escherichia coli ВКПМ В-12683, выращенная при температуре 30°С в течение 18 часов на среде LB, содержащей 25 мг/мл канамицина, 34 мкг/мл хлорамфеникола и 25 мкг/мл стрептомицина. Процесс биосинтеза ведут к колбах Эрленмейера объемом 750 мл, содержащих 100 мл среды LB с канамицином (25 мг/л), хлорамфениколом (34 мкг/мл) и стрептомицином (25 мкг/мл). Посевной материал вносят в ферментационную среду в количестве, необходимом для создания его концентрации около 1 об.%. Процесс культивирования продуцента ведут при температуре 30°С на круговой качалке со скоростью вращения 130 об/мин до достижения оптической OD600=0,8-1,0 ОЕ (около 3 часов культивирования), после чего в культуральную жидкость добавляют ИПТГ до концентрации 0,1 мМ. Ферментацию продолжают при тех же условиях в течение еще 5 часов. Биомассу клеток отделяют центрифугированием при 3000 об/мин, промывают 0,1М натрий-фосфатным буфером и вновь центрифугируют. Контроль экспрессии осуществляют с помощью денатурирующего электрофореза в полиакриламидном геле.
Полученную биомассу ресуспендируют в 50 мМ фосфатном буфере, pH 8,0, содержащем 150 мМ хлорида натрия и 10 мМ имидазола. Клетки разрушают обработкой ультразвуком, проводя процедуру на льду. Целые клетки и нерастворимую фракцию отделяют центрифугированием (16000 об/мин, 25 мин). К супернатанту добавляют равный объем 50% суспензии Ni-NTA агарозы (Qiagen, EU), предварительно уравновешенной вышеуказанным буфером и интенсивно перемешивают в течение 1 часа. Колонку заполняют полученной суспензией, промывают промывочным буфером (50 мМ фосфатный буфер, 150 мМ хлорида натрия, 20 мМ имидазола) и элюируют целевой белок с колонки буфером для смыва (50 мМ фосфатный буфер, 150 мМ хлорида натрия, 400 мМ имидазола). Полученные фракции, содержащие ксилоглюканазу AsCeGH12b, объединяют и подвергают диализу против дистиллированной воды. Активность ферментных препаратов, содержащих ксилоглюканазу, определяют по скорости гидролиза ксилоглюкана в следующих условиях: pH 4,0, температура 50 С, концентрация субстрата - 0,5 г/л. Оптимум активности рекомбинантной ксилоглюканазы, продуцируемой штаммом Escherichia coli ВКПМ В-12683 - 55°С, pH 5.0. При инкубации препарата рекомбинантного фермента в течении часа при 50°С и pH 5.0 сохраняется 95% ферментативной активности. Количество полученного целевого белка в виде нерастворимых телец включения составляет 11% от суммарного белка клетки. Количество полученного целевого физиологически активного белка составляет 343 единицы активности на 1 грамм влажной биомассы.
Таким образом:
- впервые определена нуклеотидная и аминокислотная последовательности ксилоглюканазы семейства GH12 из Aspergillus cervinus,
- сконструирована плазмида для гетерологичной экспрессии гена AsCeGH12b в бактерии Escherichia coli,
- сконструирован рекомбинантный штамм Escherichia coli - продуцент ксилоглюканазы AsCeGH12b, обладающей молекулярной массой 25 кДа,
- разработан способ микробиологического синтеза ксилоглюканазы AsCeGH12b на основе штамма Escherichia coli и не уступающий по продуктивности ближайшему аналогу.
Источники информации
1. Brumer Н., Zhou Q., Baumann M.J., Carlsson K., Teeri Т.Т. // Activation of crystalline cellulose surfaces through the chemoenzymatic modification of xyloglucan // J. Am. Chem. Soc, 2004.
2. Green M.R., Sambrook J. (2012) Molecular Cloning: a Laboratory Manual. (Fourth Edition) Cold Spring Harbor Laboratory Press.
3. Grishutin S.G., Gusakov A.V., Markov A.V., Ustinov B.B., Semenova M.V., Sinitsyn A.P. // Specific xyloglucanases as a new class of polysaccharide-degrading enzymes. // Biochim Biophys Acta, 2004.
4. Habrylo O., Song X., Forster A., Jeltsch J.M., Phalip V. // Characterization of the four GH12 Endoxylanases from the plant pathogen Fusarium graminearum. // J. Microbiol Biotechnol., 2012.
5. Hayashi T. // Xyloglucans in the primary-cell wall // Annu. Rev. Plant Physiol. Plant Mol. Biol., 1989.
6. Ishida Т., Yaoi K., Hiyoshi A., Igarashi K., Samejima M. // Substrate recognition by glycoside hydrolase family 74 xyloglucanase from the basidiomycete Phanerochaete chrysosporium. // FEBS J., 2007.
7. Miyazaki S., Endo K., Kawasaki N., Kubo W., Watanabe H., Attwood D. // Oral sustained delivery of paracetamol from in situ gelling xyloglucan formulations // Drug Dev. Ind. Pharm., 2003.
8. Matz M., Shagin D., Bogdanova E., Britanova O., Lukyanov S., Diatchenko L., Chenchik A. // Amplification of cDNA ends based on template-switching effect and step-out PCR // Nucleic Acids Research, 1999.
9. Lukyanov K., Gurskaya N., Bogdanova E., Lukyanov S. // Selective suppression of polymerase chain reaction // Bioorganicheskaya Khimiya (Russ), 1999.
10. Shankaracharya N.B. // Tamarind - chemistry, technology and uses - a critical approach // J. Food. Sci. Technol., 1998.
11. Park Y.W., Baba K., Furuta Y., Iida I., Sameshima K., Arai M., Hayashi T. // Enhancement of growth and cellulose accumulation by overexpression of xyloglucanase in poplar // FEBS Lett., 2004.
12. Markov A.V., Gusakov A.V., Kondratyeva E.G., Okunev O.N., Bekkarevich A.O., Sinitsyn A.P. // New effective method for analysis of the component composition of enzyme complexes from Trichoderma reesei. // Biochemistry (Mosc)., 2005.
13. Qi H., Bai F., Liu A. // Purification and characteristics of xyloglucanase and five other cellulolytic enzymes from Trichoderma reesei QM9414. // Biochemistry (Mosc), 2013.
14. Wang Z., Ye S., Li J., Zheng В., Bao M., Ning G. // Fusion primer and nested integrated PCR (FPNI-PCR): a new high-efficiency strategy for rapid chromosome walking or flanking sequence cloning // BMC Biotechnol., 2011.
15. Yaoi K., Mitsuishi Y. // Purification, characterization, cloning, and expression of a novel xyloglucan-specific glycosidase, oligoxyloglucan reducing end-specific cellobiohydrolase. // J Biol Chem., 2002.
16. Yaoi K., Mitsuishi Y. // Purification, characterization, cDNA cloning, and expression of a xyloglucan endoglucanase from Geotrichum sp. M128. // FEBS Lett., 2004.
17. Song S., Tang Y., Yang S., Yan Q., Zhou P., Jiang Z. // Characterization of two novel family 12 xyloglucanases from the thermophilic Rhizomucor miehei. // Appl Microbiol Biotechnol., 2013.
18. Vlasenko E.,  M., Cherry J., Xu F. // Substrate specificity of family 5, 6, 7, 9, 12, and 45 endoglucanases. // Bioresour Technol., 2010.
M., Cherry J., Xu F. // Substrate specificity of family 5, 6, 7, 9, 12, and 45 endoglucanases. // Bioresour Technol., 2010.
19. RU 2358756 // Способ получения ферментного препарата для расщепления гемицеллюлозных гетерополисахаридов клеточной стенки растений и ферментный препарат (варианты) // Дата приоритета: 26.11.2007.
20. RU 2303057 // Штамм мицелиального гриба Aspergillus aculeatus - продуцент комплекса карбогидраз, содержащего ксиланазы, бета-глюканазы, пектиназы и ксилоглюканазы // Дата приоритета: 14.11.2005.
21. RU 2323254 // Штамм мицелиального гриба Penicillium funiculosum - продуцент комплекса карбогидраз, содержащего целлюлазы, -глюканазы, -глюкозидазы, ксиланазы и ксилоглюканазы, и способ получения ферментного препарата комплекса карбогидраз для осахаривания лигноцеллюлозных материалов // Дата приоритета: 04.04.2006.
22. RU 2361918 // Штамм мицелиального гриба Penicillium verruculosum - продуцент комплекса целлюлаз, ксиланазы и ксилоглюканазы и способ получения ферментного препарата комплекса целлюлаз, ксиланазы и ксилоглюканазы для гидролиза целлюлозы и гемицеллюлозы // Дата приоритета: 26.02.2008.
23. US 2004038367 // Novel xyloglucan oligosaccharide-degrading enzyme, polynucleotide encoding the enzyme, and method of preparing the enzyme // Priority date: JP 20020083433 20020325.





Группа изобретений относится к биотехнологии. Предложен рекомбинантный штамм Escherichia coli ВКПМ В-12683, продуцент ксилоглюканазы семейства GH12, кодируемой геном AsCeGH12b (SEQ ID NO 1), клонированным из Aspergillus cervinus ВКПМ F-612. Штамм Escherichia coli ВКПМ В-12683 получен путем трансформации штамма Escherichia coli BL21-CODONPLUS (DE3)-R1PL плазмидой pET-AsCeGH12b (SEQ ID NO 3). Предложен также способ микробиологического синтеза ксилоглюканазы семейства GH12, кодируемой геном AsCeGH12b, клонированным из Aspergillus cervinus ВКПМ F-612, путем культивирования штамма Escherichia coli ВКПМ В-12683, в аэробных условиях в подходящей питательной среде до максимального накопления целевого продукта. 2 н.п. ф-лы, 1 ил., 4 пр.
1. Рекомбинантный штамм Escherichia coli ВКПМ В-12683 - продуцент ксилоглюканазы семейства GH12, кодируемой геном AsCeGH12b (SEQ ID NO 1), клонированным из Aspergillus cervinus ВКПМ F-612, и полученный путем трансформации штамма Escherichia coli BL21-CODONPLUS (DE3)-R1PL плазмидой pET-AsCeGH12b (SEQ ID NO 3).
2. Способ микробиологического синтеза ксилоглюканазы, предусматривающий культивирование рекомбинантного штамма бактерии Escherichia coli, содержащего ген ксилоглюканазы, в аэробных условиях в подходящей питательной среде до максимального накопления целевого продукта, отличающийся тем, что в качестве продуцента используют рекомбинантный штамм по п. 1, а целевым продуктом является ксилоглюканаза семейства GH12, кодируемая геном AsCeGH12b, клонированным из Aspergillus cervinus ВКПМ F-612.
| US 20040038367 A1, 26.02.2004 | |||
| FURTADO G | |||
| ET AL | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| // Appl | |||
| Microbiol | |||
| Biotechnol., 2015, Jun;99(12):5095-107 | |||
| TISHKOV V.I | |||
| ET AL | |||
| Способ гальванического снятия позолоты с серебряных изделий без заметного изменения их формы | 1923 |
|
SU12A1 |
| Многоступенчатая активно-реактивная турбина | 1924 |
|
SU2013A1 |
| ШТАММ БАКТЕРИИ Esherichia coli EX pQE30, ПРОДУЦЕНТ ЭНДОКСИЛАНАЗЫ БАКТЕРИИ Geobacillus stearothermophillus 22 | 2013 |
|
RU2542486C1 |

