Область техники
Онколитические вирусы представляют собой класс новых терапевтических веществ, используемых для лечения рака, обладающих уникальной способностью к опухоле-зависимому самообеспечению (HERMISTON. A demand for next-generation oncolytic adenoviruses. Current opinion in molecular therapeutics. 2006, vol. 8, no. 4, p.322-30.). Онколитические вирусы способны к селективной репликации в злокачественных клетках и поэтому предлагают уровни силы действия и специфичности, которые потенциально намного превышают стандартные противораковые терапии (FISHER. Striking out at disseminated metastases; the systemic delivery of oncolytic viruses. Current opinion in molecular therapeutics. 2006, vol. 8, no. 4, p.301-13.). Преимущество использования этих вирусов состоит в том, что при своей репликации они лизируют своих клеток-хозяев. Раковые клетки - идеальные хозяева для многих вирусов, потому у них инактивирован антивирусный интерфероновый путь или мутированы гены-супрессоры опухоли, которые позволяют беспрепятственно протекать вирусной репликации (CHERNAJOVSKY, et al. Fighting cancer with oncolytic viruses. British medical journal. 2006, vol. 332, no. 7534, p.170-2.). Некоторые вирусы по природе способны селективно реплицироваться в опухолевых клетках, но онколитические вирусы могут также быть получены путем модификации природных вирусов. С этой целью главные стратегии, используемые в настоящее время для модификации вирусов, включают: функциональные делеции в существенных вирусных генах; опухоль- или ткане-специфичные промотеры, используемые для контроля экспрессии этих вирусных генов; и модификация тропизма, чтобы перенаправить аденовирус на поверхность раковой клетки. В ближайшем будущем необходимо оптимизировать онколитические аденовирусы, чтобы полностью реализовать их потенциал как критических противораковых инструментов и, таким образом, улучшить прогноз для пациентов со злокачественными глиомами (JIANG, et al. Oncolytic adenoviruses as antiglioma agents. Expert review of anticancer therapy. 2006, vol. 6, no. 5, p.697-708.).
Например, ONYX-015, селективно модифицированный аденовирус для репликации и уничтожения клеток, которые имеют р53 мутации, разрабатывается компанией «Onyx Pharmaceuticals» для потенциального лечения различных солидных опухолей, включая опухоли головы и шеи, желудочно-кишечные и панкреатические опухоли. Он представляет собой рекомбинантный аденовирус, который несет мутацию «потери функции» в локусе Е1В, продуктом которой является 55 кДа белок, который связывается с р53 опухолевым белком-супрессором и инактивирует его. Таким образом, аденовирус ONYX-015, как предполагается, не поражает нормальные клетки. Мутации в р53 гене-супрессоре опухоли представляют собой наиболее распространенный тип генетической патологии при раке, который встречается в более чем половине всех главных типов рака. Таким образом, эти клетки восприимчивы к вирусу, который будет легко реплицироваться и вызывать смерть клетки. Продолжаются исследования фазы III применения ONYX-015 относительно лечения рецидивирующего рака головы и шеи, исследования фазы II - относительно лечения колоректальной опухоли, опухолей яичника, поджелудочной железы и ротовой полости, и исследования фазы I - относительно болезней органов пищеварения, опухолей пищевода и печени (COHEN, et al. ONYX-015. Onyx Pharmaceuticals. Current opinion in investigational drugs. 2001, vol. 2, no. 12, p.1770-5.). Природные онколитические вирусы представляют собой репликация-компетентные вирусы, обладающие врожденной способностью селективно заражать и уничтожать клетки опухоли. Несмотря на то, что они использовались в оригинальных попытках лечить рак живыми вирусами пять десятилетий назад, интерес к природным онколитическим вирусам отстал от поддержки созданных генной инженерией аденовирусов и вирусов герпеса в качестве терапии рака. Однако, недавно был возобновлен интерес к высокой силе действия и селективности этих природных агентов (ROBERTS, et al. Naturally oncolytic viruses. Current opinion in molecular therapeutics. 2006, vol. 8, no. 4, p.314-21.).
Среди природных онколитических вирусов, вирусы осповакцины (Poxviridae) обладают многими из ключевых признаков, необходимых для идеального вирусного каркаса для использования в онкодитической виротерапии. Они включают короткий период полувыведения, с быстрым межклеточным распространением, сильную литическую способность, большую способность к клонированию и четкую молекулярную биологию. Кроме того, хотя они способны к репликации в клетках человека, их не считают природной проблемой для здоровья и особенно хорошо характеризуются тем, что их вводили миллионам людей во время кампании по уничтожению оспы. Ранние клинические результаты, использующие или штаммы вакцины или генетически модифицированные штаммы осповакцины продемонстрировали противоопухолевые эффекты (THORNE, et al. Vaccinia virus and oncolytic virotherapy of cancer. Current opinion in molecular therapeutics. 2005, vol. 7, no. 4, p.359-65.).
Напротив, поксвирус миксомы является новым онколитическим кандидатом, у которого нет никакой истории непосредственного использования у людей, поскольку он обладает отличительным и абсолютным тропизмом к виду-хозяину по отношению к лагоморфам (кроликам). Вирус миксомы, как недавно показано, может также селективно заражать и уничтожать человеческие опухолевые клетки, уникальный тропизм, который связан с дисрегулированными внутриклеточными сигнальными путями, обнаружен в большинстве человеческих раковых образований. Этот обзор обрисовывает в общих чертах существующие данные относительно тропизма вируса миксомы к человеческим раковым клеткам, а также доклинические данные, показывающие его способность заражать и уничтожать опухоли на животных моделях рака (STANFORD, et al. Myxoma virus and oncolytic virotherapy: a new biologic weapon in the war against cancer. Expert opinion on biological therapy. 2007, vol. 7, no. 9, p.1415-25.).
Техническая проблема
Инъекция высоких доз поксвирусов, необходимая для достижения противоопухолевого эффекта, вызвала проблемы, связанные с токсичностью. Большинство неблагоприятных явлений представляют собой незначительные неблагоприятные реакции, которые обычно связаны с вирусом осповакцины, ограничены и включают лихорадку, головную боль, усталость, миальгию, озноб, местные кожные реакции, неспецифичную сыпь, мультиформную эритему, увеличение лимфатических узлов и боль в участке прививки. Другие реакции могли бы потребовать дополнительных методов лечения (например, VIG, терапия первой линии и цидофовир, терапия второй линии). Неблагоприятные реакции, которые могли бы потребовать дальнейшей оценки или терапии, включают неумышленную инокуляцию, генерализированную коровью оспу (GV), экзему после прививки (EV), прогрессирующую коровью оспу (PV), поствакцинальную болезнь центральной нервной системы, и эмбриональную коровью оспу (CONO, et al. Smallpox vaccination and adverse reactions. Guidance for clinicians. MMWR. Recommendations and reports: Morbidity and mortality weekly report. Recommendations and reports / Centers for Disease Control. 2003, vol. 52, no. RR-4, p.1-28.).
Таким образом, существует потребность в более безопасных поксвирусах с такой же онколитической активностью, как и у их природных копий.
Предпосылки создания изобретения
В US 5364773 (VIROGENETICS CORPORATION (TROY, NY)) 15/11/1994 описывают модифицированный рекомбинантный поксвирус, в особенности вирус коровьей оспы, имеющий инактивированные несущественные закодированные вирусом генетические функции таким образом, чтобы рекомбинантный поксвирус обладал уменьшенной токсичностью и повышенной безопасностью. В частности, генетические функции инактивированы путем делеции открытой рамки считывания, кодирующей фактор вирулентности, или инсерционной инактивацией открытой рамки считывания, кодирующей фактор вирулентности. Более подробно, этот патент описывает вирус осповакцины, в котором открытая рамка считывания для J2R, B13R+B14R, A26L, A56R, C7L - K1L, и I4L была инактивирована. Этот вирус (NYVAC) может быть спроектирован как вектор для чужеродной нуклеиновой кислоты и использоваться в качестве вакцины для того, чтобы вызвать иммунологический ответ в животном-хозяине. Однако, NYVAC неспособен эффективно реплицироваться в большинстве клеток млекопитающих и не может использоваться как онколитический вирус (XIANGZHI, et al. Vaccinia virus K1L protein supports viral replication in human and rabbit cells through a cell-type-specific set of its ankyrin repeat residues that are distinct from its binding site for ACAP2. Journal of virology. 2006, vol. 353, no. 1, p.220-233.).
WO 2004/014314 (KIRN DAVID (US)) 19/02/2004 описывает измененный вирус осповакцины, который включает одну или несколько мутаций в его вирусном геноме. Описанные мутации находятся в одном или нескольких из следующих классов полипептидов: 1) интерферон-модулирующий полипептид; 2) комплемент-контрольный полипептид; 3) TNF или хемокин-модулирующий полипептид; 4) ингибитор сериновой протеазы; 5) IL-1p-модулирующий полипептид; 6) неинфекционные EEV формы полипептидов; и 7) вирусный полипептид, который действует для ингибирования высвобождения инфекционного вируса из клеток (противоинфекционная вирусная форма полипептида). Кроме того, также раскрыты мутации в вирусе осповакцины A41L или C11R.
Участки генома осповакцины, такие как A34R, A41L, A53R, B5R, B7R, B8R, B13R, B15R, B18R, B22R, B28R, B29R, CUR, E3L, K2L, NIL, vC12L, vCKBP более подробно описаны в этой заявке. Способы изобретения вовлекают использование любого из поксвирусов, обсужденных авторами. Изобретатели также раскрывают способы для лечения рака путем введения в раковую клетку или пациенту эффективного количества этого измененного вируса осповакцины.
Раскрытие изобретения
Изобретатели к удивлению обнаружили, что поксвирусы, включающие дефектный I4L и/или F4L ген, имеют улучшенный профиль безопасности, но сохраняют эквивалентную онколитическую активность (по сравнению с их естественной копией). Настоящее изобретение касается поксвируса, включающего дефектный I4L и/или F41 ген с условием, что указанный поксвирус не представляет собой NYVAC.
Как используются по всей заявке термины в единственном числе используются в смысле, что они подразумевают "по крайней мере один", "по крайней мере первый", "один или несколько" или "множество" компонентов или шагов, на которые ссылаются, если контекст ясно не указывает иначе. Например, термин "клетка" включает множество клеток, включая их смеси.
Термин "и/или", используемый авторами, включает значение "и", "или" и "все или любая другая комбинация элементов, связанных указанным термином".
Термин "около" или "приблизительно", используемый авторами, обозначает в пределах 20%, предпочтительно в пределах 10%, и более предпочтительно в пределах 5% данного значения или диапазона.
Используемые авторами термины "включающие" и "включают" означают, что продукты, композиции и способы включают компоненты или стадии, на которые ссылаются, но исключают другие. "Состоящий по существу из", используемый для определения продуктов, композиций и способов, должен означать исключение других компонентов или стадий любого существенного значения. Таким образом, композиция, состоящая по существу из перечисленных компонентов, не исключает загрязнители в следовых количествах и фармацевтически приемлемые носители. "Состоящий из" должен означать исключение больше чем элементы в следовых количествах других компонентов или стадий.
Используемый авторами термин "поксвирус, включающий дефектный ген" относится к поксвирусу, включающему делеции, замены или дополнения одной или нескольких нуклеиновых кислот дефектного гена, или любой комбинации этих возможностей, причем указанные модификации приводят к неспособности для вируса продуцировать белок, имеющий активность белка, произведенного немодифицированным геном. В предпочтительном варианте осуществления изобретения поксвирус, включающий дефектный ген, касается поксвируса, в котором была удалена целая генная последовательность. Мутация может быть осуществлена многими способами, известными квалифицированным специалистам в данной области техники, используя рекомбинантные методы. Методы для модификации генома поксвируса доступны в данной области. Например, методы, раскрытые в MCCART, et al. Systemic cancer therapy with a tumor selective vaccinia virus mutant lacking thymidine kinase and vaccinia growth factor genes.. Cancer res.. 2001, no. 61, p.8751-57., KIM, et al. Systemic armed oncolytic ans immunologic therapy for cancer with JX-594, a targeted poxvirus expressing GM-CSF. Molecular Therapeutic. 2006, no. 14, p.361-70., WO 2004/014314 (KIRN DAVID (US)) 19/02/2004 и US 5364773 (VIROGENETICS CORPORATION (TROY, NY)) 15/11/1994 могут использоваться, чтобы произвести поксвирус изобретения. Способы, раскрытые в примере данной заявки, особенно относятся к получению поксвируса согласно изобретению. Последовательности генома различных поксвирусов доступны в данной области, например, геномы вируса осповакцины, вируса коровьей оспы, вируса Canarypox, вируса Ectromelia, вируса миксомы, доступны в Genbank (инвентарный номер NC_006998, NC_003663, NC_005309, NC_004105, NC_001132, соответственно).
Используемый авторами термин "поксвирус" относится к вирусу, принадлежащему к семейству Poxviridae. Согласно предпочтительному варианту осуществления, поксвирус согласно изобретению принадлежит к подсемейству Chordopoxvirinae, более предпочтительно роду Orthopoxvirus и еще более предпочтительно к виду вируса Vaccinia.
Например, могут использоваться штаммы вируса осповакцины Dairen I, IHD-J, L-IPV, LC16M8, LC16MO, Lister, LIVP, Tashkent, WR 65-16, Wyeth, Ankara, Copenhagen, Tian Tan и WR. Согласно особенно предпочтительному варианту осуществления, поксвирус согласно изобретению представляет собой штаммы Copenhagen вируса осповакцины. Поксвирус осповакцины содержит большой двойной геном ДНК (187 пар килооснований) и является членом единственного известного семейства ДНК вирусов, которое реплицируется в цитоплазме инфицированных клеток. Поскольку инфицированная клетка должна поставить большое количество предшественников ДНК в цитоплазматические участки репликации, вирус кодирует и экспрессирует много ферментативных активностей, требуемых для метаболизма и синтеза ДНК, включая рибонуклеотидредуктазу и дезоксиуридин 5'-трифосфат-нуклеотидогидролазу (dUTPase).
Рибонуклеотидредуктаза (ЕС 1.17.4.1) катализирует восстановление рибонуклеотидов до дезоксирибонуклеотидов, реакция, которая представляет первый фиксированный шаг в биосинтезе ДНК. Вирусный фермент подобен по структуре субъединице фермента млекопитающих, состоящему из двух гетерологичных субъединиц под названием R1 и R2. Гены, которые кодируют субъединицы вирусной рибонуклеотидредуктазы, были секвенированы и локализованы в положениях на геноме осповакцины, отделенном 35 килооснованиями (SLABAUGH, et al.. Journal of virology. 1988, vol. 62, p.519-27.; TENGELSEN, et al. Virology. 1988, no. 164, p.121-31.; SCHMITT, et al. Journal of virology. 1988, no. 62, p.1889-97.). Мономеры большой субъединицы вируса осповакцины (под названием R1, закодированные геном I4L) представляют собой 86-кДа полипептиды, и содержат связывающие участки для нуклеотидных субстратов и аллостерических эффекторов (SLABAUGH, et al.. Journal of virology. 1984, no. 52, p.507-14.; SLABAUGH, et al.. Journal of virology. 1984, no. 52, p.501-6.). Маленькая субъединица (под названием R2, закодированная геном F4L) представляет собой гомодимер, включающий два 37-кДа полипептида; каждый полипептид содержит стабилизированный железом основанный на белке свободный радикал, который требуется для катализа (HOWELL, et al.. Journal of Biological Chemistry. 1992, no. 267, p.1705-11.). Последовательности для генов I4L и F4L и их местоположения в геноме различных поксвирусов доступны в общественных базах данных, например, по инвентарному номеру DQ437594, DQ437593, DQ377804, АН015635, AY313847, AY313848, NC_003391, NC_003389, NC_003310, M35027, AY243312, DQ011157, DQ011156, DQ011155, DQ011154, DQ011153, Y16780, Х71982, AF438165, U60315, AF410153, AF380138, U86916, L22579, NC_006998, DQ121394 и NC_008291.
Генная номенклатура, используемая в данном описании, является номенклатурой штамма Copenhagen осповакцины, и используется также для гомологичных генов других poxviridae, если не указано иначе. Однако, генная номенклатура может отличаться в зависимости от штамма оспы. Для получения информации, соответствия между генами Copenhagen и MVA, см. Таблицу I ANTOINE. Virology. 1998, no. 244, p.365-396.
Согласно предпочтительному варианту осуществления, поксвирус данного изобретения дополнительно включает дефектный ген J2R.
Ген J2R кодирует тимидинкиназу (ТК), которая являются частью "реутилизационного" пути для синтеза дезоксирибонуклеотида пиримидина. Реакция, катализируемая ТК, вовлекает перенос γ-фосфорилостатка из АТР на 2'дезокси-тимидин (dThd), чтобы произвести тимидин 5'-монофосфат (dTMP). ТК вируса осповакцины имеет тип 2. ТК типа 2 имеют меньшую полипептидную цепь по сравнению с типом 1, ~25 кДа, но формируют гомотетрамеры. Они чувствительны к ингибиторам обратной связи dTDP или dTTP, которые генерируются в конце метаболического пути. У ТК типа 2 более узкая специфичность к субстрату по сравнению с ТК типа 1 и только фосфорилат 2'дезоксиуридин (dU) и/или dThd (EL ОМАМ, et al. Structure of vaccinia virus thymidine kinase in complex with dTTP: insights for drug design. BMC structural biology. 2006, no. 6, p.22.).
Поксвирусы, дефектные на участке J2R, и методы их получения доступны в данной области техники. Например, руководство MCCART, et al. Systemic cancer therapy with a tumor-selective vaccinia virus mutant lacking thymidine kinase and vaccinia growth factor genes, cancer research, 2001, vol. 61, no. 24, p.8751-7., PUHLMANN, et al. Vaccinia as a vector for tumor-directed gene therapy: biodistribution of a thymidine kinase-deleted mutant. Cancer gene therapy. 2000, vol. 7, no. 1, p.66-73., GNANT, et al. Systemic administration of a recombinant vaccinia virus expressing the cytosine deaminase gene and subsequent treatment with 5-fluorocytosine leads to tumor-specific gene expression and prolongation of survival in mice. Cancer Research, 1999, vol. 59, no. 14, p.3396-403 может использоваться для получения поксвирусов с делецией участка J2R.
Согласно предпочтительному варианту осуществления поксвирус изобретения дополнительно включает дефектный ген F2L.
Дезоксиуридин 5'-трифосфатнуклеотидогидролаза (dUTPase, EC 3.6.1.23) катализирует гидролиз dUTP до dUMP и пирофосфата в присутствии ионов Mg (2+). dUTPаза, при удалении dUTP из пула dNTP и производства dUMP, вовлечена и в поддержание преданности репликации ДНК и в предоставление предшественника для производства ТМР тимидилатсинтазой. dUTPаза осповакцины представляет собой 15-кДа белок, закодированный геном F2L (MCGEOGH.. Nucleic Acids Research. 1990, no. 18, p.4105-10; BROYLES.. Virology. 1993, no. 195, p.863-5.). Последовательность гена F2L вируса осповакцины доступна в генном банке по инвентарному номеру М25392, последовательности и местоположения гена F2L в различных геномах поксвирусов также доступны в генном банке, например, по инвентарному номеру NC_006998, DQ121394, NC_001611, AY689436, AY689437, NC_008291, DQ437594, DQ437593, AY313847, AY313848, NC_006966, NC_005309, NC_003391, NC_003389, NC_001132, NC_003310, NC_002188, M35027, AY243312, AF170726, DQ011157, DQ011156, DQ011155, DQ011154, DQ011153, X94355, Y16780, AY318871, U94848, AF198100 и M34368.
Согласно предпочтительному варианту воплощения, поксвирус согласно изобретению дополнительно включает целевую нуклеиновую кислоту.
В предпочтительном варианте воплощения целевая нуклеиновая кислота содержит по крайней мере одну целевую последовательность, кодирующую генный продукт, который является терапевтической молекулой (то есть терапевтическим геном). "Терапевтическая молекула" является молекулой, обладающей фармакологической или защитной активностью при надлежащем введении пациенту, особенно пациенту, страдающему от болезненного состояния или болезни или тому, кого следует защитить от этой болезни или состояния. Такая фармакологическая или защитная активность представляет собой активность, которая, как ожидают, будет связана с благоприятным воздействием на ход или симптом указанной болезни или указанного состояния. Когда квалифицированный специалист выбирает в ходе существующего изобретения ген, кодирующий терапевтическую молекулу, он вообще связывает свой выбор с ранее полученными результатами, и может разумно ожидать, без чрезмерного эксперимента, кроме осуществления изобретения согласно формуле, получить такое фармакологическое свойство. Согласно изобретению, целевая последовательность может быть гомологичной или гетерологичной мишеневым клеткам, в которые она вводится. Предпочтительно указанная целевая последовательность кодирует весь или часть полипептида, особенно терапевтический или профилактический полипептид, дающий терапевтическое или профилактическое свойство. Полипептид, как понимают, является любым трансляционным продуктом полинуклеотида независимо от размера, и независимо от гликозилирования, и включает пептиды и белки. Терапевтические полипептиды включают как первичный пример те полипептиды, которые могут компенсировать дефектные или несовершенные белки в животном или человеческом организме, или те, которые действуют через токсичные эффекты для ограничения или удаления вредных клеток из организма. Они могут также быть придающими иммунность полипептидами, которые действуют как эндогенный антиген, чтобы вызвать гуморальный или клеточный ответ, или и тот, и другой.
Примеры полипептидов, закодированных терапевтическим геном, включают гены, кодирующие цитокин (альфа, бета или гамма интерферон, интерлейкин, в особенности IL-2, IL-6, IL-10 или IL-12, фактор некроза опухоли (TNF), колоние-стимулирующий фактор GM-CSF, C-CSF, M-CSF…), иммуностимуляторный полипептид (В7.1, В7.2 и т.п.), фактор коагуляции (FVIII, FIX…), фактор роста (трансформирующий фактор роста TGF, фактор роста фибробластов FGF и т.п.), фермент (уреаза, ренин, тромбин, металлопротеиназа, синтаза оксида азота NOS, SOD, каталаза…), ингибитор фермента (альфа 1-антитрипсин, антитромбин III, вирусный ингибитор протеазы, ингибитор активатора плазминогена PAI-1), CFTR (регулятор трансмембранной проводимости кистозного фиброза) белок, инсулин, дистрофии, антиген МНС класса I или II, полипептид, который может модулировать/регулировать экспрессию клеточных генов, полипептид, способный к ингибированию бактериальной, паразитной или вирусной инфекции или ее развития (антигенные полипептиды, антигенные эпитопы, трансдоминантные варианты, ингибирующие действие нативного белка путем конкуренции…), индуктор или ингибитор апоптоза (Вах, Вс12, Вс1Х…), цитотоксический агент (р21, р16, Rb…), аполипопротеин (ApoAI, ApoAIV, ApoE…), ингибитор ангиогенеза (ангиостатин, эндостатин…), ангиогенный полипептид (семейство сосудистых эндотелиальных факторов роста VEGF, семейство FGF, семейство CCN, включая CTGF, Cyr61 и Nov), поглотитель кислородных радикалов, полипептид, имеющий антиопухолевый эффект, антитело, токсин, иммунотоксин и маркер (бета-галактозидаза, люцифераза…) или любые другие целевые гены, признанные в данной области техники как полезные для лечения или предотвращения клинического состояния.
Подходящие противоопухолевые гены включают, кроме прочих, гены, которые кодируют гены-супрессоры опухоли (например, Rb, p53, DCC, NF-1, опухоль Вильма, NM23, BRUSH-1, p16, р21, р56, р73, а также их соответствующие мутанты), продукты суицидального гена, антитела, полипептиды, ингибирующие клеточное деление или сигналы трансдукции.
Согласно особенно предпочтительному варианту воплощения, поксвирус настоящего изобретения дополнительно включает суицидальный ген.
Суицидальный ген касается гена, кодирующего белок, способный преобразовать предшественник лекарственного препарата в цитотоксичное соединение.
Суицидальные гены включают, кроме прочих, гены, кодирующие белок, имеющий цитозин-дезаминазную активность, тимидин-киназную активность, урацил-фосфорибозильную трансферазную активность, пуриннуклеозид-фосфорилазную активность и/или тимидилат-киназную активность.
Примеры суицидальных генов и соответствующие предшественники лекарственного препарата, включающего один остаток нуклеооснования, раскрыты в следующей таблице:
Согласно предпочтительному варианту воплощения изобретения, суицидальный ген кодирует белок, имеющий по крайней мере активность СОазы. СОаза вовлечена в пиримидиновый метаболический путь, по которому экзогенный цитозин преобразовывается в урацил посредством гидролитического дезаминирования. В то время как активности СБазы были продемонстрированы в прокариотах и низших эукариотах (JUND, et al.. Journal of Bacteriology. 1970, no. 102, p.607-15.; BECK, et al.. Journal of Bacteriology. 1972, no. 110, p.219-28.; HOEPRICH, et al.. Journal of Infectious Diseases. 1974, no. 130, p.112-18.; ESDERS, et al.. J. biol. chem.. 1985, no. 260, p.3915-22.), они не присутствуют у млекопитающих (KOECHLIN, et al.. Biochemical pharmacology. 1966, no. 15, p.435-46.; POLAK, et al.. Chemotherapy. 1976, no. 22, p.137-53.).
CDаза также дезаминирует аналог цитозина, то есть 5-фторцитозин (5-FC), таким образом формируя 5-фтороурацил (5-FU), который представляет собой соединение, являющееся очень цитотоксичным при его преобразовании в 5-фтор-UMP (5-FUMP). Клетки, которые не имеют активности СDазы, или из-за мутации, которая инактивирует ген, кодирующий фермент, или потому что у них природно нет этого фермента, как у клеток млекопитающих, являются резистентными к 5-FC (JUND, et al. Journal of Bacteriology. 1970, no. 102, p.607-15.; KILLSTRUP, et al.. Journal of Bacteriology. 1989, no. 171, p.2124-2127.). В отличие от этого, клетки млекопитающих, в которые были перенесены последовательности, кодирующие активность СDазы, стали чувствительными к 5-FC (HUBER, et al. Cancer Research. 1993, no. 53, p.4619-4626.; MULLEN, et al.. Proceedings of the National Academy of Sciences of the United States of America. 1992, no. 89, p.33-37.; WO 93/01281 (US HEALTH)). Кроме того, соседние, непреобразованные клетки также становятся чувствительными к 5-FC (HUBER, et al.. Proceedings of the National Academy of Sciences of the United States of America. 1994, no. 91, p.8302-6.). Это явление, которое называют эффектом «свидетеля», происходит из-за клеток, которые экспрессируют активность СDазы, секретирующей 5-FU, который интоксицирует соседние клетки путем прямой диффузии через плазматическую мембрану. Это свойство 5-FU относительно пассивной диффузии представляет преимущество по сравнению с tk/GCV эталонной системой, в которой эффект «свидетеля» требует контакта с клетками, которые экспрессируют tk (MESNIL, et al. Proceedings of the National Academy of Sciences of the United States of America. 1996, no. 93, p.1831-35.). Все преимущества, которые CDаза предлагает в пределах контекста генной терапии, в особенности противораковой генной терапии, могут поэтому быть легко поняты.
Гены codA Saccharomyces cerevisiae (S.cerevisiae) FCY1, Candida Albicans FCA1 и Е.coli, которые соответственно кодируют CDasy этих двух организмов, известны, и их последовательности были опубликованы (SEQ ID N°:4; SEQ ID N°:5; SEQ ID N°:6 соответственно).
В этом отношении согласно более предпочтительному варианту воплощения настоящего изобретения, ген, кодирующий белок, имеющий активность CDазы, является FCY1, FCA1 или CodA или их аналогом. Аналоги этих генов касаются гена, имеющего последовательность нуклеиновой кислоты, у которой степень идентичности, по крайней мере, больше чем 70%, преимущественно больше чем 80%, предпочтительно больше чем 90%, и наиболее предпочтительно больше чем 95% с последовательностью нуклеиновой кислоты родительского гена.
Патент WO 2005/007857 описывает ген, кодирующий белок, имеющий улучшенную активность CDазы. Эти полипептиды получены из нативной CDазы путем дополнения аминокислотной последовательности. Согласно другому предпочтительному варианту воплощения данного изобретения, белок, имеющий активность CDазы, представляет собой полипептид, раскрытый в WO 2005/007857, и более предпочтительно полипептид FCU1-8, представленный в идентификаторе последовательности SEQ ID N°:2 и их аналоги.
В прокариотах и низших эукариотах, урацил преобразовывается в UMP под действием урацилфосфорибозилтрансферазы (UPRTазы). Этот фермент превращает 5-FU в 5-FUMP. Согласно другому предпочтительному варианту воплощения настоящего изобретения, суицидальный ген кодирует белок, имеющий активность UPRTазы.
Рассматриваемая UPRTаза может иметь любое происхождение, в особенности прокариотическое происхождение, грибковое или дрожжевое происхождение. Посредством иллюстрации, последовательности нуклеиновой кислоты, кодирующие UPRTазы из Е.coli (ANDERSEN, et al. Characterization of the upp gene encoding uracil phosphoribosyltransferase of Escherichia coli K12. European Journal of Biochemistry. 1992, no. 204, p.51-56.), из Lactococcus lactis (MARTINUSSEN, et al. Cloning and-characterization of upp, a gene encoding uracil phosphoribosyltransferase from Lactococcus lactis. Journal of Bacteriology. 1994, vol. 176, no. 21, p.6457-63.), из Mycobacterium bovis (KIM, et al. Complete sequence of the UPP gene encoding uracil phosphoribosyltransferase from Mycobacterium bovis BCG. Biochemistry and molecular biology international. 1997, vol.41, no. 6, p.1117-24.) и из Bacillus subtilis (MARTINUSSEN, et al. Two genes encoding uracil phosphoribosyltransferase are present in Bacillus subtilis. Journal of Bacteriology. 1995, vol. 177, no. 1, p.271-4.) может использоваться в контексте изобретения. Однако, больше всего особенно предпочтительно использовать дрожжевую UPRTазу и в особенности закодированную S.cerevisiae FUR1 геном, последовательность которого раскрыта в KERN, et al. The FUR1 gene of Saccharomyces cerevisiae: cloning, structure and expression of wild-type and mutant alleles. (Gene. 1990, vol. 88, no. 2, p.149-57), который вводится авторами посредством ссылки. В качестве руководства последовательности генов и последовательности соответствующих UPR-Таз могут быть найдены в литературе и банках данных специалистов (SWISSPROT, EMBL, Genbank, Medline и т.п.).
Заявка ЕР 0998568 А описывает ген FUR1, не имеющий 105 нуклеотидов в 5' кодирующей части, позволяющей синтез UPRTазы, из которой были удалены 35 первых остатков в N-терминальном положении и начиная с метионина в положении 36 в нативном белке. Продукт экспрессии гена-мутанта под названием FUR1A105, способен к комплементированию fur1 мутанта S.cerevisiae. Кроме того, усеченный мутант показывает более высокую активность UPRТазы, чем таковая для нативного фермента. Таким образом, согласно особенно предпочтительному варианту воплощения настоящего изобретения, суицидальный ген кодирует мутант делеции нативной UPRТазы. Делеция предпочтительно расположена в N-терминальной области оригинальной UPRTазы. Она может быть полной (затрагивающей все остатки указанной N-терминальной области) или частичной (воздействующей на один или несколько непрерывных или прерывистых остатков в первичной структуре). Вообще, полипептид состоит из N-терминальной, центральной и C-терминальной частей, каждая из которых представляет приблизительно одну треть молекулы. Например, поскольку UPRTаза S.cerevisiae имеет 251 аминокислоту, ее N-терминальная часть состоит из первых 83 остатков, начинающихся с так называемого инициатора метионина, расположенного в первом положении нативной формы. Что касается UPRTазы E.coli, ее N-терминальная часть охватывает положения 1-69.
Предпочтительный белок, имеющий активность UPRTазы, включает аминокислотную последовательность практически такую как представлена в идентификаторе последовательности SEQ ID N°:1 из ЕР 0998568 А, начинающаяся с остатка Met в положении 1 и заканчивающаяся остатком Val в положении 216. Термин "практически" относится к степени идентичности с указанной последовательностью SEQ ID N°:1 ЕР 0998568 больше чем 70%, преимущественно больше чем 80%, предпочтительно больше чем 90%, и наиболее предпочтительно больше чем 95%. Все еще более предпочтительно она включает аминокислотную последовательность, представленную в идентификаторе последовательности SEQ ID N°:1 ЕР 0998568 А. Как упомянуто выше, она может включать дополнительные мутации. В особенности, может быть упомянута замена остатка серина в положении 2 (положение 37 в нативной UPRTазе) на аланиновый остаток.
Согласно другому предпочтительному варианту воплощения настоящего изобретения, суицидальный ген кодирует белок, имеющий по крайней мере одну активность СDазы и одну активность UPRTазы. Патентные заявки WO 96/16183 и ЕР 0998568 А описывают использование слитого белка, кодирующего фермент с двумя доменами, имеющими активности СDазы и UPRTазы, и демонстрируют, что перенос гибридного гена codA::upp или FCY1::FUR1 или FCY1::FUR1Δ105 (т.е. FCU1), который несет плазмида экспрессии, увеличивает чувствительность трансфецированных В 16 клеток к 5-FC. Согласно более предпочтительному варианту воплощения настоящего изобретения, суицидальный ген кодирует полипептид, включающий аминокислотную последовательность, практически такую как представлена в идентификаторе последовательности SEQ ID N°:3 (coda::upp), SEQ ID N°:1 (FCU1) или FCY1::FUR1. Термин "практически" относится к степени идентичности с указанной последовательностью, больше чем 70%, преимущественно больше чем 80%, предпочтительно больше чем 90%, и наиболее предпочтительно больше чем 95%. Все еще более предпочтительно, она включает аминокислотную последовательность, практически такую как представлена в идентификаторе последовательности SEQ ID N°:3 (coda::upp), SEQ ID N°:1 (FCU1) или FCY1::FUR1. Как упомянуто выше, она может включать дополнительные мутации.
Нуклеиновокислотные последовательности могут быть легко получены путем клонирования, ПЦР или химическим синтезом согласно обычным используемым способам. Они могут быть нативными генами или генами, полученными из них путем мутации, делеции, замены и/или дополнения одного или нескольких нуклеотидов. Кроме того, их последовательности широко описаны в литературе, к которой могут обратиться специалисты, квалифицированные в данной области техники.
Специалисты, квалифицированные в данной области техники, способны клонировать последовательности CDазы или UPRTазы на основании опубликованных данных и выполнить возможные мутации, тестировать ферментативную активность мутантных форм в безклеточной или клеточной системе согласно технологии из предшествующего уровня техники или основанные на протоколе, обозначенном в заявке ЕР 0998568 А, и сплавить, в особенности в фазе, полипептиды с активностью СDазы и UPRTазы, и следовательно со всеми или частью соответствующих генов.
Согласно более предпочтительному варианту воплощения, поксвирус изобретения дополнительно включает нуклеиновокислотную последовательность, включающую ген, кодирующий пермеазу.
Пермеаза касается трансмембранного белка, вовлеченного в перенос лекарственного препарата, включающего один остаток нуклеооснования, или его предшественника через клеточную мембрану.
Пермеаза включает, но ограничения перечисленными, пуринпермеазу, цитозинпермеазу и транспортеры нуклеозида. Согласно предпочтительному варианту воплощения данного изобретения, пермеаза представляет собой пурин или цитозинпермеазу S.Cerevisiae. Транспортеры нуклеооснования S.cerevisiae состоят из пурин-цитозин пермеазы, известной как FCY2, и урацилпермеазы, известной как FUR4. Пурин-цитозин пермеаза, FCY2 опосредует симпорт протонов и аденина, гуанина, гипоксантина и цитозина через плазматическую мембрану дрожжей (Grenson 1969, Jund and Lacroute 1970, Polak and Grenson 1973, Chevallier et al. 1975, Hopkins et al. 1988). Белок FCY2 опосредует также транспорт 5-фторцитозина, аналога цитозина (Grenson 1969, Jund and Lacroute 1970). FCY2 ген кодирует белок из 533 аминокислот (58 кДа), как изначально предполагалось, имеет 10-12 трансмембранных вращающихся доменов (Weber et al. 1990), девять из которых на сегодняшний день одобрены (Ferreira et al. 1999). FCY2 показывает подобные сродства для пуриновых нуклеооснований и цитозина (Brethes et al. 1992). Захват урацила в S. cerevisiae опосредуется урацилпермеазой, FUR4 (Jund and Lacroute 1970, Jund et al. 1977). FUR4 представляет собой урацил-протонный симпортер (Hopkins et al. 1988), который по-видимому является белком из 633 аминокислот (71,7 кДа) с 10 трансмембранными доменами и длинными цитоплазматическими гидрофильными N- и C-терминальными хвостами (Jund et al. 1988, Gamier et al. 1996). Белок FUR4 может также опосредовать транспорт 5-фтороурацила, аналога урацила (Jund and Lacroute 1970).
Аминокислотные последовательности FCY2 и Fur4 особенно доступны в базе данных swissprot (инвентарный номер Р17064 и Р05316, соответственно). Предпочтительно, пермеаза имеет аминокислотную последовательность, выбранную из группы, включающей аминокислотную последовательность SEQ ID NO:1 и SEQ ID NO:2, как раскрыто в патентной заявке WO 2006/048768.
В этом отношении согласно предпочтительному варианту воплощения данного изобретения, пермеаза выбирается из группы, включающей FCY2 и Fur4 и их аналоги. Аналоги Fur4 и FCY2 касаются полипептида, имеющего аминокислотную последовательность, которая имеет степень идентичности, по крайней мере, больше чем 70%, преимущественно больше чем 80%, предпочтительно больше чем 90%, и наиболее предпочтительно больше чем 95% с аминокислотной последовательностью родительского белка как описано авторами выше и которые сохраняют способность транспортировать лекарственный препарат, включающий один остаток нуклеооснования через клеточную мембрану.
Специалист, квалифицированный в данной области техники, может выбрать пермеазу, которая будет связана с препаратом или предшественником препарата, включающего один остаток нуклеооснования. Например, FCY2 и Fur4 предпочтительно связываются с 5-фторцитозином (5-FC).
В соответствии с более предпочтительным вариантом воплощения, поксвирус изобретения может дополнительно включать элементы, необходимые для экспрессии целевой нуклеиновой кислоты.
В соответствии с более предпочтительным вариантом воплощения, поксвирус изобретения может дополнительно включать элементы, необходимые для экспрессии нуклеиновокислотной последовательности, включающей ген, кодирующий пермеазу.
Эти элементы, необходимые для экспрессии целевой нуклеиновой кислоты и/или нуклеиновокислотной последовательности, включающей ген, кодирующий пермеазу, включали элементы, требуемые для транскрипции указанной ДНК в мРНК и, в случае необходимости, для трансляции мРНК в полипептид. Транскрипционные промотеры, подходящие для использования в различных системах позвоночных, широко описаны в литературе. Например, среди подходящих промотеров вирусные промотеры, такие как RSV, MPSV, SV40, CMV или 7,5k, промотер осповакцины, индуцибельные промотеры, и т.д. Предпочтительные промотеры изолируются из поксвирусов, например, 7.5K, H5R, TK, р28, р11 или K1L вируса осповакцины. Альтернативно, можно использовать синтетический промотер, такой как описан в CHAKRABARTI.. Biotechniques. 1997, no. 23, p.1094-97., HAMMOND, et al.. Journal of Virological Methods. 1997, no. 66, p.135-38. и KUMAR.. Virology. 1990, no. 179, p.151-8, а также химерные промотеры между ранними и поздними поксвирусными промотерами.
Целевая нуклеиновокислотная последовательность и нуклеиновокислотная последовательность, включающая ген, кодирующий пермеазу, могут дополнительно включать дополнительные функциональные элементы, такие как последовательности интрона, нацеливающие последовательности, транспортные последовательности, сигнал секреции, ядерный сигнал локализации, IRES, поли А последовательности терминации транскрипции, трехсторонние лидерные последовательности, последовательности, вовлеченные в репликацию или интеграцию. Об указанных последовательностях сообщалось в литературе, и они могут быть легко получены квалифицированными специалистами в данной области техники.
Изобретение также касается способа для получения поксвируса в соответствии с изобретением, в котором способ:
(i) поксвирус согласно изобретению вводят в клетку,
(ii) указанную клетку культивируют при условиях, которые являются соответствующими для того, чтобы позволить продукцию указанного поксвируса, и
(iii) указанный поксвирус выделяют из клеточной культуры.
В то время как поксвирус может, конечно, быть выделен из супернатанта культуры, он может также быть выделен из клеток. Один из обычно используемых способов состоит в разложении клеток посредством последовательных циклов замораживания/размораживания, чтобы собрать вирионы в лизисный супернатант. Затем эти вирионы могут быть амплифицированы и очищены с использованием способов из данной области техники (хроматографический метод, метод ультрацентрифугирования, в особенности через градиент хлорида цезия, и т.д.).
Настоящее изобретение также касается композиции, которая включает поксвирус в соответствии с изобретением в комбинации с фармацевтически приемлемым вспомогательным веществом.
Композиция в соответствии с изобретением более определенно предназначена для профилактической или излечивающей терапии болезней посредством генной терапии и более специфично нацелена на пролиферативные болезни (раковые образования, опухоли, рестеноз, и т.д.) или нацелены на болезни, связанные с увеличенной активностью остеокластов (например, ревматоидный артрит, остеопороз).
Композиция в соответствии с изобретением может быть сделана традиционно в целях ее введения локально, парентерально или пищеварительным путем. В частности, терапевтически эффективное количество рекомбинантного вектора или поксвируса данного изобретения комбинируют с фармацевтически приемлемым вспомогательным веществом. Возможно предусмотреть большое количество путей введения. Примеры, которые могут быть упомянуты, включают внутрижелудочный, подкожный, внутрисердечный, внутримышечный, внутривенный, внутрибрюшинный, внутриопухолевый, интраназальный, внутрилегочный и внутритрахеальный пути. В случае этих трех последних вариантов воплощения предпочтительно, чтобы введение происходило с помощью аэрозоля или с помощью инстилляции. Введение может происходить в виде единственной дозы или дозы, которая повторяется в одном или нескольких случаях после определенного временного интервала. Соответствующий путь введения и дозировки изменяется в зависимости от множества параметров, например, таких как пациент, болезнь, которую будут лечить, или целевой ген(ы), который(ые) будет(будут) перенесен(ы). Препараты, основанные на вирусных частицах согласно изобретению, могут быть сформулированы в форме доз, составляющих от 104 до 1014 pfu (бляшкообразующая единица), преимущественно от 105 до 1013 pfu, предпочтительно от 106 до 1012 pfu, более предпочтительно от 106 до 107.
Композиция может также включать растворитель, адъювант или вспомогательное вещество, которое является приемлемым с фармацевтической точки зрения, а также солюбилизирующее, стабилизирующее средство и консервант. В случае инъекционного введения предпочтение отдается композиции в водном, неводном или изотоническом растворе. Она может быть представлена в виде однократной дозы или мультидозы, в жидком или сухом виде (порошок, лиофилизат и т.д.), которая может быть приготовлена во время использования с помощью соответствующего растворителя.
Настоящее изобретение также касается использования поксвируса или композиции согласно изобретению для приготовления лекарственного средства, которое предназначено для лечения организма человека или животных генной терапией. Это лекарственное средство можно вводить непосредственно in vivo (например, внутривенная инъекция, в доступную опухоль, в легкие с помощью аэрозоля, в сосудистую систему, используя соответствующий катетер, и т.д.). Предпочтительное использование состоит в лечении или предотвращении раковых образований, опухолей и болезней, которые следуют из нежелательной пролиферации клеток. Мыслимые применения, которые могут быть упомянуты, включают раковые образования молочной железы, матки (в особенности, вызванные вирусами папилломы), предстательной железы, легкого, мочевого пузыря, печени, кишечника, поджелудочной железы, желудка, пищевода, гортани, центральной нервной системы (например, глиобластома) и крови (лимфомы, лейкемия, и т.д.). Другое предпочтительное использование состоит в лечении или предотвращении ревматоидного артрита, остеопороза и других болезней, связанных с увеличенной активностью остеокластов. Она может также использоваться в контексте сердечнососудистых болезней, например, для ингибирования или задержки пролиферации клеток гладких мышц стенки кровеносного сосуда (рестеноз). Наконец, в случае инфекционных болезней, возможно применять лекарственное средство при СПИДе.
Когда поксвирус, композиция или способ изобретения используются для лечения рака, предпочтительный путь введения представляет собой системный путь, так как поксвирус согласно изобретению в состоянии специфично нацеливаться на опухолевые клетки.
Изобретение также охватывает метод для лечения болезни, характеризующийся тем, что поксвирус, композицию в соответствии с изобретением вводят в организм хозяина или клетку, которая нуждается в таком лечении.
Согласно выгодному варианту воплощения, терапевтическое использование или способ лечения также включает дополнительную стадию, в которой фармацевтически приемлемые количества пролекарства, преимущественно аналога цитозина, в особенности 5-FC, вводят в организм хозяина или клетку. Посредством иллюстрации возможно использовать дозу от 50 до 500 мг/кг/сутки, при этом предпочтительна доза 200 мг/кг/ сутки или 100 мг/кг/сутки. В пределах контекста настоящего изобретения пролекарство вводят в соответствии со стандартной практикой (например, перорально, систематически).
Предпочтительно, введение, происходящее после введения терапевтического агента в соответствии с изобретением, предпочтительно по крайней мере спустя 3 дня, более предпочтительно по крайней мере 4 дня и еще более предпочтительно по крайней мере 5 дней после введения терапевтического агента. В соответствии с даже более предпочтительным вариантом воплощения данного изобретения, введение пролекарства имеет место спустя 7 дней после введения терапевтического агента. Предпочтителен пероральный путь. Возможно вводить однократную дозу пролекарства или дозами, которые повторяются какое-то время, которых достаточно, чтобы позволить выработку токсичного метаболита в пределах организма хозяина или клетки. Кроме того, композиция или способ согласно изобретению могут быть комбинированы с одним или несколькими веществами, которые потенциируют цитотоксический эффект 5-FU. В особенности можно упомянуть лекарственные препараты, которые ингибируют ферменты пути для de novo биосинтеза пиримидинов (например, упомянутые ниже), лекарственные препараты, такие как лейковорин (Waxman et al., 1982, Eur. J. Cancer Clin. Oncol. 18, 685-692), который, в присутствии продукта метаболизма 5-FU (5-FdUMP), увеличивает ингибирование тимидилатсинтазы, приводя к уменьшению пула dTMP, который требуется для репликации, и наконец, лекарственные препараты, такие как метотрексат (Cadman et al., 1979, Science 250, 1135-1137), который, ингибируя дигидрофолатредуктазу и увеличивая пул PRPP (фосфорибозилпирофосфат), вызывает увеличение внедрения 5-FU в клеточную РНК.
Согласно настоящему изобретению, лекарственные препараты, которые ингибируют ферменты пути для de novo биосинтеза пиримидинов, предпочтительно выбираются из группы, состоящей из PALA (N-(фосфоноацетил)-L-аспартата; Moore et al., 1982, Biochem. Pharmacol. 31, 3317-3321), лефлуномида, А771726 (активный метаболит лефлуномида; Davis et al., 1996, Biochem. 35, 1270-1273) и бреквинара (Chen et al., 1992, Cancer Res. 52, 3251-3257).
Композиция или способ в соответствии с изобретением могут быть комбинированы с одним или несколькими веществами, эффективными в противораковой терапии. Среди фармацевтических веществ, эффективных в противораковой терапии, которые могут использоваться в ассоциации или в комбинации с композициями согласно изобретению, можно упомянуть алкилирующие агенты, такие как, например, митомицин С, циклофосфамид, бусульфан, ифосфамид, изосфамид, мелфалан, гексаметилмеламин, тиотепа, хлорамбуцил, или дакарбазин; антиметаболиты такие как, например, гемцитабин, капецитабин, 5-фтороурацил, цитарабин, 2-фтордезоксицитидин, метотрексат, идатрексат, томудекс или триметрексат; ингибиторы топоизомеразы II, такие как, например, доксорубицин, эпирубицин, этопозид, тенипозид или митоксантрон; ингибиторы топоизомеразы I, такие как, например, иринотекан (СРТ-11), 7-этил-10-гидрокси-камптотецин (SN-38) или топотекан; антимитотические препараты, такие как, например, паклитаксель, доцетаксель, винбластин, винкристин или винорелбин; и производные платины, такие как, например, цисплатин, оксалиплатин, спироплатин или карбоплатин.
Композиции или способы в соответствии с изобретением могут также быть использованы в комбинации с лучевой терапией.
Композиции или способы в соответствии с изобретением могут также быть использованы в комбинации с одним или несколькими другими агентами, включая, кроме прочих, иммуномодуляторные агенты, такие как, например, альфа, бета или гамма интерферон, интерлейкин (в особенности, IL-2, IL-6, IL-10 или IL-12) или фактор некроза опухоли; агенты, которые влияют на регулирование рецепторов клеточной поверхности, такие как, например, ингибиторы рецептора эпидермального фактора роста (в особенности, цетуксимаб, панитумумаб, залутумумаб, нимотузумаб, матузумаб, гефитиниб, эрлотиниб или лапатиниб) или ингибиторы рецептора человеческого эпидермального фактора роста 2 (в особенности, транстузумаб); и агенты, которые влияют на ангиогенез, такие как, например, ингибитор сосудистого эндотелиального фактора роста (в особенности, бевацизумаб или ранибизумаб).
Краткое описание Фигур в чертежах
Фигура 1. Чувствительность in vitro к 5-FC из инфицированных вирусом осповакцины человеческих клеток колоректальной опухоли (LoVo). Клетки LoVo, инфицированные при MOI 0,0001 указанными вирусами (пример (•) VVTK-/FCU1 (■) или VVTK-I4L-/FCU1 (Δ)), подвергают действию различной концентрации 5-FC. Выживание клеток определяют через 5 дней после инфицирования. Результаты выражают в процентах клеточной жизнеспособности в присутствии или в отсутствии лекарственных препаратов. Значения представляют в виде среднее значение ± SD трех индивидуальных определений без смертности клеток из-за репликации вирусов.
Фигура 2. Чувствительность in vitro к 5-FC из инфицированных вирусом осповакцины человеческих клеток колоректальной опухоли (LoVo). Клетки LoVo, инфицированные при MOI 0,0001 указанными вирусами (пример (•) VVTK-/FCU1 (■) или VVTK-F4L-/FCU1(◊) подвергают действию различной концентрации 5-FC. Выживание клеток определяют через 5 дней после инфицирования. Результаты выражают в процентах клеточной жизнеспособности в присутствии или в отсутствии лекарственных препаратов. Значения представляют в виде среднее значение ± SD трех индивидуальных определений без смертности клеток из-за репликации вирусов.
Фигура 3. Эффективность репликации in vitro VVTK-/FCU1 и VVTK-I4L-/PCU1 в LoVo, инфицированных при MOI 0,0001 указанными вирусами в день 5 после инфицирования. Значения представляют в виде среднее значение ± SD трех индивидуальных определений.
Фигура 4. Эффективность репликации in vitro VVTK-/FCU1 и VVTK-F4L-/FCU1 в LoVo, инфицированных при MOI 0,0001 указанными вирусами в день 5 после инфицирования. Значения представляют в виде среднее значение ± SD трех индивидуальных определений.
Фигура 5. Средний объем опухоли ± SEM п/к LoVo у швейцарских голых мышей после в/в инъекции вируса. Спустя 7 дней после инокуляции опухоли (пальпируемая опухоль), мышей лечат с помощью 107 pfu буфера + солевой раствор (◊), буфер + 5-FC (♦), VVTK-I4L-/FCU1 + солевой раствор (Δ) или VVTK-I4L-/FCU1 + 5-FC (▲). Животных лечат солевым раствором или 5-FC при 100 мг/кг/j два раза в день чреззондовым кормлением, спустя 7 дней после вирусной инъекции в течение 3 недель. Два раза в неделю определяют объем опухоли.
Фигура 6. Средний объем опухоли ± SEM п/к LoVo у швейцарских голых мышей после в/в инъекции вируса. Спустя 7 дней после инокуляции опухоли (пальпируемая опухоль), мышей лечат с помощью 107 pfu буфера + солевой раствор (◊), буфер + 5-FC (♦), VVTK-F4L-/FCU1 + солевой раствор (□) или VVTK-F4L-/FCU1 + 5-FC (■). Животных лечат солевым раствором или 5-FC при 100 мг/кг/j два раза в день чреззондовым кормлением, спустя 7 дней после вирусной инъекции в течение 3 недель. Два раза в неделю определяют объем опухоли.
Фигура 7. Средний объем опухоли ± SEM п/к LoVo у швейцарских голых мышей после в/в инъекции вируса. Спустя 11 дней после инокуляции опухоли (пальпируемая опухоль), мышей лечат буфером + H2O (◊), буфер + 5-FC (♦), или одна инъекция 107 pfu VVTK-I4L-/FCU1 + H2O (○), или одна инъекция 107 pfu VVTK-I4L-/FCU1 + 5-FC (5-FC, который вводят спустя 7 дней после инъекции вируса и в течение 3 недель) (•), или две инъекции (день 11 и день 33) 107 pfu VVTK-I4L-/FCU1 + H2O (□), или две инъекции (день 11 и день 33) 107 pfu VVTK-I4L-/FCU1 + 5-FC (5-FC, который вводят с дня 18 до дня 32 и с дня 40 до дня 54) (■). Животных лечат 5-FC при 100 мг/кг два раза в день чреззондовым кормлением. Два раза в неделю определяют объем опухоли.
Фигура 8. Средний объем опухоли ± SEM п/к U87-MF (опухолевые клетки глиобластомы) у швейцарских голых мышах после в/в инъекции вируса. Спустя 11 дней после инокуляции опухоли (пальпируемая опухоль), мышей лечат с помощью буфера + H2O (◊), или буфера + 5-FC (♦), или 107 pfu VVTK-I4L-/FCU1 + Н2О (○), или 107 pfu VVTK-I4L-/FCU1 + 5-FC (•). Животных лечат с помощью 5-FC при 100 мг/кг два раза в день чреззондовым кормлением, спустя 7 дней после инъекции вируса и в течение 3 недель. Два раза в неделю определяют объем опухоли.
Фигура 9. Соотношение выхода вируса в делящихся клетках в сравнении со сливающимися клетками. Клетки PANC1 (опухоль поджелудочной железы человека), Н1299 (опухоль легких человека) или U118MG (опухоль глиомы человека) инфицируют с помощью 100 pfu (■) VVTK-/FCU1 или (□) VVTK-I4L-/FCU1. Спустя 48 часов после инфицирования определяют вирусные титры. Значения представляют собой соотношение между выходами вируса в делящихся клетках в сравнении со сливающимися клетками.
Фигура 10. Соотношение выхода вируса в делящихся клетках в сравнении со сливающимися клетками. Клетки PANC1 (опухоль поджелудочной железы человека), Н1299 (опухоль легких человека) или U118MG (опухоль глиомы человека) инфицируют с помощью 100 pfu (■) VVTK-/FCU1 или (□) VVTK-F4L-/FCU1. Спустя 48 часов после инфицирования определяют вирусные титры. Значения представляют собой соотношение между выходами вируса в делящихся клетках в сравнении со сливающимися клетками.
Фигура 11. Вирусные титры (pfu/мг ткани) в органах или опухолях в день 6 и день 21 после в/в инфекции у швейцарских голых мышей, носящих подкожные человеческие опухоли с 1×106 PFU VVTK-/FCU1 (■) или VVTK-I4L-/FCU1 (□).
Фигура 12. Вирусные титры (pfu/мг ткани) в органах или опухолях в день 6 и день 21 после в/в инфекции у швейцарских голых мышей, носящих подкожные человеческие опухоли с 1×106 PFU VVTK-/FCU1 (■) или VVTK-F4L-/FCU1 (□).
Фигура 13. Выживание швейцарских голых мышей после лечения с помощью 1×108 pfu VVTK-/FCU1 (■) или VVTK-I4L-/FCU1 (○) путем в/в инъекции.
Фигура 14. Выживание иммунокомпетентных мышей B6D2 после лечения с помощью 1×107 pfu (А) или 1×108 pfu (В) VVTK-/FCU1 (■) или VVTK-I4L-/FCU1 (◊) путем в/в инъекции.
Фигура 15. Среднее количество оспин на хвостах после в/в инъекции 1×106 pfu VVTK-/FCU1 или VVTK-I4L-/FCU1 у швейцарских голых мышей в день 13 после инфицирования и в день 34 после инфицирования.
Фигура 16. Среднее количество оспин на хвостах после в/в инъекции 1×106 pfu VVTK-/FCU1 или VVTK-F4L-/FCU1 у швейцарских голых мышей в день 13 после инфицирования и в день 34 после инфицирования.
Фигура 17. Среднее количество оспин на хвостах после в/в инъекции 1×107 pfu VVTK-/FCU1 или WTK-I4L-/FCU1 у швейцарских голых мышей в день 15 после инфицирования и в день 31 после инфицирования.
Фигура 18. Среднее количество оспин на хвостах после в/в инъекции 1×107 pfu VVTK-/FCU1 или VVTK-F4L-/FCU1 у швейцарских голых мышей в день 15 после инфицирования и в день 31 после инфицирования.
Способ(ы) для осуществления изобретения
Примеры
Конструкция векторных плазмид
Строят челночную плазмиду для удаления I4L с использованием ДНК штамма Copenhagen вируса осповакцины (инвентарный номер М35027), удаленной на гене Тимидинкиназы и экспрессирующей ген FCU1 под контролем синтетического промотера осповакцины p11K.7.5. Фланкирующие участки ДНК I4L амплифицируют с помощью ПЦР. Праймеры нисходящего фланкирующего участка I4L представляют собой 5'-ТСС ССС GGG ТТА АСС ACT GCA TGA TGT ACA-3' (SEQ ID N°:7; участок SmaI подчеркнут), и 5'-GCC GAG CTC GAG GTA GCC GTT TGT AAT TCT -3' (SEQ ID N°:8; участок SacI подчеркнут). Праймеры нисходящего участка представляют собой 5'-GCC TGG ССА ТАА CTC CAG GCC GTT -3' (SEQ ID N°:9; участок MscI подчеркнут), и 5'-GCC CAG CTG АТС GAG CCG ТАА CGA ТТТ ТСА -3' (SEQ ID N°:10; участок PvuII подчеркнут). Амплифицированный фрагмент ДНК расщепляют ферментом рестрикции SmaI/SacI или MscI/PvuII и лигируют в соответствующие участки в плазмиде PpolyIII. Повторный участок нисходящего фланкирующего участка I4L амплифицируют спомощью ПЦР с использованием праймера 5'-GCC GCA TGC АТС СТТ GAA САС САА ТАС CGA -3' (SEQ ID N°:11; участок SphI подчеркнут), и 5'-GCT СТА GAG AGG TAG CCG ТТТ GTA АТС TG -3' (SEQ ID N°:12; участок XbaI подчеркнут) и вставляют в плазмиду PpolyIII. Повторный участок используют для устранения кассеты селекции во время производства удаленных вирусов. Кассету селекции, соответствующую слитому гену GFP/GPT при контроле pH5R промотера осповакцины, вставляют в участок SacI/SphI в плазмиде PpolyIII. Полученная плазмида представляет собой рекомбинантную челночную плазмиду под названием pΔI4L для удаления гена I4L.
Строят челночную плазмиду для удаления F4L с использованием ДНК штамма Copenhagen вируса осповакцины (инвентарный номер М35027). Фланкирующие участки ДНК F4L амплифицируют с помощью ПЦР. Праймеры нисходящего фланкирующего участка F4L представляют собой 5'-CGC GGA TCC TTT GGT АСА GTC TAG TAT CCA -3' (SEQ ID N°:13; участок BamHI подчеркнут), и 5'-TCC ССС GGG ТТА ТАА CAG ATG CAG ТАТ CCA -3' (SEQ ID N°:14; участок SmaI подчеркнут). Праймеры восходящего участка представляют собой 5'-GCC CAG CTG ТТС ААТ GGC CAT CTG AAA TCC -3' (SEQ ID N°:15; участок PvuII подчеркнут) и 5'-GAA GAT СТА GTA TCG CAT СТА AAA GAT GG -3' (SEQ ID N°:16; участок BglII подчеркнут). Амплифицированный фрагмент ДНК расщепляют ферментом рестрикции BamHI/SmaI или BglII/PvuII и лигируют в соответствующие участки в плазмиде PpolyIII. Повторный участок нисходящего фланкирующего участка I4L амплифицируют с помощью ПЦР с использованием праймеров 5'-GCC GAG СТС АСС САС ACG TTT ТТС GAA AAA -3' (SEQ ID N°:17; участок SacI подчеркнут), и 5'-GCC GCA TGC ТТА ТАА CAG ATG CAG TAT CAA -3' (SEQ ID N°:18; участок SphI подчеркнут) и вставляют в плазмиду PpolyIII. Повторный участок используют для устранения кассеты селекции во время производства удаленных вирусов. Кассету селекции, соответствующую слитому гену GFP/GPT при контроле pH5R промотера осповакцины, вставляют в участок SacI/SphI в плазмиде PpolyIII. Полученная плазмида представляет собой рекомбинантную челночную плазмиду под названием pAF4L для удаления гена F4L.
Получение рекомбинантных вирусов осповакцины.
Клетки СЕР инфицируют штаммом Copenhagen VVTK-FCU1 (вирус Vaccinia, дефектный на ген киназы J2R, экспрессирующий ген FCU1 под контролем синтетического промотера p11k7.5) при MOI 0,1 и инкубируют при 37°С в течение 2 ч., затем трансфецируют с копреципитатом CaCl2 рекомбинантной челночной плазмиды (0,2 мкг). Клетки инкубируют в течение 48 ч. при 37°С. Полученные разведения вируса используют, чтобы инфицировать клетки CEF в селективной среде, содержащей гипоксантин при конечной концентрации 15 мкг/мл, ксантин при конечной концентрации 250 мкг/мл и микофеноловую кислоту при конечной концентрации 250 мкг/мл. Флуоресцентные (GFP) и позитивные (GPT селекция) бляшки изолируют и отбирают для нескольких раундов селекции в клетках CEF в присутствии селективной среды GPT. Присутствие или отсутствие VVTK-FCU1 определяют 40 циклами ПЦР с праймерами в участке делеции. После устранения родительского вируса используют двойной удаленный вирус для инфицирования CEF без селективной среды GPT, чтобы устранить кассету селекции. Нефлуоресцентные бляшки изолируют и отбирают для 2 циклов в CEF. Заключительные рекомбинантные вирусы VV амплифицируют в СЕР, очищают и вирусные исходные растворы титруют на CEF с помощью теста на бляшки.
Клеточная чувствительность in vitro к 5-FC.
Человеческие клетки опухоли преобразовывают соответствующим рекомбинантным VV при MOI 0,0001. В общей сложности 3×105 клеток/лунку помещают на 6-луночные чашки для культуры в 2 мл среды, содержащей различные концентрации 5-FC. Затем клетки культивируют при 37°С в течение 5 дней, и жизнеспособные клетки подсчитывают с помощью исключения трипаном синим. Результаты, изображенные на фигурах 1, 2, 3 и 4, показывают, что активность FCU1 эквивалентна у вирусов, дефектных на ген J2R, чем у вирусов, дефектных на ген I4L и J2R или у вирусов, дефектных на ген F4L и J2R.
In vitro репликация в культивированных клетках.
Делящиеся клетки или сливающиеся клетки инфицируют, в 6-лунковых бляшках, при 100 PFU вирусов (почти MOI 0,0005). Добавляют 2 мл среды с 10% FCS для делящихся клеток и без добавок для сливающихся клеток. Клетки собирают через 48 часов после инфицирования. Клетки хранят при -20°С и обрабатывают ультразвуком для высвобождения вируса, вирус также определяют количественно титрованием бляшек на клетках CEF. Соотношение между репликацией в делящихся клетках и сливающихся клетках подобно во всех клетках. Оба вируса VVTK-/FCU1, VVTK-I4L-/FCU1 и VVTK-F4L-/FCU1 реплицируются больше в делящихся клетках, чем в сливающихся клетках.
Как косвенное значение для количественного определения вирусной специфичности к репликации, определяют выход вируса, полученный в делящихся клетках по сравнению со сливающимися опухолевыми клетками (человеческая опухоль поджелудочной железы PANC1; человеческая опухоль легкого Н1299; человеческая опухоль глиомы U118MG). Сливающиеся клетки помещают на планшеты при 1×106 клеток/лунку и культивируют в полных средах в течение 7 дней, затем за 1 день до инфицирования клетки промывают и культивируют в среде без сыворотки. Делящиеся клетки помещают на планшет при 3×105 клеток/лунку на один день перед инфицированием. Чтобы оценить уровень клеточного деления, количество оттитрованного тимидина, включенного в нуклеиновую кислоту, определяют спустя 5 часов, 24 часа и 48 часов после культивирования клеток на планшете. Во время этого периода включение тимидина является относительно постоянным в сливающихся клетках, тогда как в делящихся клетках увеличение включения замечают со временем. Тогда клетки инфицируют с помощью 100 pfu вирусов, и спустя 48 часов после инфицирования соотношение между выходом вируса, произведенного в делящихся опухолевых клетках и в сливающихся опухолевых клетках, определяют титрованием бляшек на СЕР. Результаты, изображенные на фигурах 9 и 10, показывают, что оба вируса VVTK-/FCU1, VVTK-I4L-/FCU1 и VVTK-F4L-/FCU1 больше реплицируются в делящихся клетках, чем в сливающихся клетках. Результаты, изображенные на фигурах 9 и 10, кроме того, показывают увеличение соотношения во всех различных типах клеток для обоих вирусов VVTK-I4L-/FCU1 и VVTK-F4L-/FCU1 в сравнении с VVTK-/FCU1. Это увеличение соотношения во всех различных типах клеток происходит из-за более низкой репликации обоих вирусов VVTK-I4L-/FCU1 и VVTK-F4L-/FCU1 в сливающихся клетках. Эти результаты демонстрируют, что оба вируса VVTK-I4L-/FCU1 и VVTK-F4L-/FCU1 показывают увеличенную специфичность по отношению к делящимся клеткам в сравнении с VVTK-/FCU1.
Подкожная модель опухоли.
Самок швейцарских голых мышей получают из Charles River Laboratories. Животные, используемые в исследованиях, являются однородными по возрасту (6 недель), а массы тела колеблются от 23 до 26 г. Швейцарским голым мышам вводят подкожно (п/к) в бок 5×106 клеток LoVo. Когда опухоли достигают диаметра 50-70 мм3, мышей рандомизируют произвольным способом и обрабатывают указанными векторами для экспериментов in vivo.
Биораспределение вируса.
Присутствие различных вирусов оценивают вирусным титрованием в образцах органов и опухолей. 1×106 PFU VV-FCU1 или VVTK-I4L-/FCU1 или VVTK-F4L-/FCU1 вводят внутривенной (в/в) инъекцией в хвостовую вену голым мышам с заданными п/к опухолями LoVo. Мышей умертвляют в определенные периоды времени, и опухоли и другие органы собирают и взвешивают. Опухоли и органы гомогенизируют в PBS, и титры определяют на CEF, как описано ранее. Вирусные титры стандартизируют к миллиграмму ткани, Вирусные титры стандартизируют к миллиграмму ткани. Результаты, изображенные в Таблице 2, 3, 4 и 5 (диапазон вирусных титров представлен в pfu/мг ткани), показывают, что после 14 дней вирус согласно изобретению главным образом обнаруживается в опухоли. Результаты, изображенные на фигурах 11 и 12, показывают, что оба вируса VVTK-/FCU1, WTK-I4L-/FCU1 и VVTK-F4L-/FCU1 нацеливаются на опухоль приблизительно в 1000-10000 раз больше вируса в опухоли, чем в других органах, проанализированных за исключением хвостов в случае VVTK-/FCU1. Небольшое количество VVTK-/FCU1 обнаружено в легких, селезенке, почке и лимфоузлах (меньше чем 10 pfu/мг) и больше в коже, хвосте и костном мозге в день 6, и коже и хвосте в день 21. Напротив, и WTK-I4L-/FCU1 и VVTK-F4L-/FCU1 имеют более высокую специфичность к опухоли с только небольшим количеством в лимфоузлах и хвосте в день 6, и только в опухоли в день 21.
Противоопухолевая активность поксвируса изобретения на п/к модели опухоли.
Голые мыши с установленными п/к опухолями LoVo (50-70 мм3) получают лечение один раз внутривенно (хвостовая вена) указанными векторами в дозе 1.107 PFU, соответственно. Начиная с дня 7 после вирусной инъекции, 5-FC вводится перорально через зонд при 100 мг/кг (0,5 мл 5-FC 0,5% в воде) два раза в день в течение 3 недель. Размер опухоли определяют дважды в неделю с использованием кронциркуля. Объем опухоли вычисляют в мм3 с использованием формулы (р/6) (длина × ширина2). Результаты, изображенные на фигуре 5 и 6, показывают, что у разных вирусов есть подобная эффективность с онколитической активностью (p<0,05), способная контролировать рост опухоли, и объединенная активность (онколитическая вируса и терапевтическая гена FCU1) с введением 5-FC, который может дополнительно улучшить контроль роста опухоли (p<0,01).
Голые мыши с установленными п/к опухолями LoVo (50-70 мм3) также получают лечение один раз внутривенно (хвостовая вена) указанными векторами в дозе 1.107 PFU в соответствии со следующей схемой: спустя 11 дней после инокуляции опухоли (пальпируемая опухоль) мышей лечат: буфер + H2O, или буфер + 5-FC, или одна инъекция 107 pfu VVTK-I4L-/FCU1 + Н2О, или одна инъекция 107 pfu VVTK-I4L-/FCU1+5-FC (5-FC вводится спустя 7 дней после вирусной инъекции и в течение 3 недель), или две инъекции (день 11 и день 33) 107 pfu VVTK-I4L-/FCU1 + Н2О, или две инъекции (день 11 и день 33) 107 pfu VVTK-I4L-/FCU1+5-FC (5-FC вводится со дня 18 до дня 32 и со дня 40 до дня 54). Животных лечат 5-FC при 100 мг/кг два раза в день перорально через зонд. Размер опухоли определяют дважды в неделю с использованием кронциркуля. Объем опухоли вычисляют в мм3 с использованием формулы (р/6) (длина × ширина2). Результаты, изображенные на фигуре 7, показывают, что не отмечается никакой антиопухолевой активности одного только вируса после одной или двух инъекций. Добавление лечения с помощью 5-FC показывает статистически существенное ингибирование роста опухоли (p<0,05) при сравнении с группами носителя и только вируса (без 5-FC) до дня 50. Как и в случае с однократной инъекцией, две в/в инъекции VVTK-I4L-/FCU1 + 5-FC демонстрируют существенную антиопухолевую активность при сравнении с группами носителя и двумя инъекциями одного только вируса (без 5-FC) (p<0,05). Кроме того, существенное различие в развитии опухоли наблюдается со дня 56 между одной и двумя инъекциями вируса в комбинации с лечением 5-FC (p<0,05).
Голые мыши с установленными п/к U87-MG (опухолевые клетки глиобластомы) получают лечение внутривенно (хвостовая вена) указанными векторами в дозе 1.107 PFU в соответствии со следующей схемой: спустя 11 дней после инокуляции опухоли (пальпируемая опухоль) мышей лечат: буфер + Н2О, или буфер + 5-FC, или 107 pfu VVTK-I4L-/FCU1 + H2O, или 107 pfu VVTK-I4L-/FCU1 + 5-FC. Животных лечат с помощью 5-FC при 100 мг/кг два раза в день перорально через зонд, спустя 7 дней после вирусной инъекции и в течение 3 недель. Размер опухоли определяют дважды в неделю с использованием кронциркуля. Объем опухоли вычисляют в мм3 с использованием формулы (π/6) (длина × ширина2). Результаты, изображенные на фигуре 8, показывают высокую онколитическую активность VVTK-I4L-/FCU1 на клетках U87-MG, которые приводят к сильной противоопухолевой активности (p<0,0001). Комбинированная активность с добавлением 5-FC перорально через зонд приводит к подобной активности (p<0,0001).
Вирусная патогенность.
Вирусную патогенность оценивают исследованиями выживания, которые проводят и на швейцарских голых мышах (фигура 13) и на иммунокомпетентных мышах B6D2 (фигура 14). Мышам в/в вводят 1.107 или 1.108 PFU всех VVTK-/FCU1 и VVTK-I4L-/FCU1 в 100 мкл буфера на мышь. За мышами наблюдают ежедневно по ходу эксперимента. У швейцарских голых мышей (фигура 13) инъекция 1×108 PFU VVTK-/FCU1 приводит к смерти 40% животных спустя 3 дня после инфицирования. Остающиеся мыши умирают между днем 50 и днем 80 после инфицирования. Введение VVTK-I4L-/FCU1 является менее патогенным, большинство животных умирает между днем 65 и 140 (p<0,01). Не наблюдается никаких свидетельств токсичности с обоими вирусами при 107 pfu (фигура 14 (А)). Все мыши умирают после в/в инъекции 108 pfu VVTK-/FCU1 (фигура 14 (В). Группа с лечением VVTK-I4L-/FCU1 имеет значительно более длительное выживание до 70% по сравнению с VVTK-/FCU1 инфицированными мышами (фигура 14 (В). Поэтому, этот результат демонстрирует уменьшение токсичности с двойным удаленным вирусом VVTK-I4L-/FCU1.
Модель поражения хвоста оспинами.
Швейцарским голым мышам в/в вводят 1.106 (фигуры 15 и 16) или 1.107 (фигуры 17 и 18) PFU каждого вируса. Поражения хвоста высчитывают один раз в неделю. У мышей, которым вводят 1.106 PFU VVTK-I4L-/FCU1 или VVTK-F4L-/FCU1, отмечается меньше чем 1 оспина/мышь по сравнению с мышами, которым вводят VVTK-/FCU1 со средним числом, равным 8 оспин на мышь в день 13 после инфицирования (p<0,001) как показано на фигуре 15 (А) и фигуре 16 (А). Результаты подобны в день 34 после инфицирования со средним числом, равным 4 оспинам с VVTK-/FCU1 по сравнению с почти 1 для VVTK-I4L-/FCU1 или VVTK-F4L-/FCU1 (p<0,0001) как показано на фигуре 15 (В) и фигуре 16 (В). У мышей, которым вводят 1.107 PFU VVTK-I4L-/FCU1 или VVTK-F4L-/FCU1, отмечается соответственно среднее число, равное 3 оспины/мышь и 2 оспины/мышь по сравнению с мышами, которым вводят 1.10 PFU VVTK-/FCU1, имеющими в среднем 10 оспин/мышу в день 15 после инфицирования (фигура 17 (А) и фигура 18 (А)). В день 31 после инфицирования у мышей, которым вводят VVTK-I4L-/FCU1 или VVTK-F4L-/FCU1, отмечается соответственно в среднем 1,5 оспин/мышь и 2 оспин/мышь по сравнению с мышами, которым вводят VVTK-/FCU1, имеющими в среднем 7 оспин/мышь (фигура 17 (В) и фигура 18 (В)). Различие в количестве оспин между VVTK-/FCU1 и VVTK-I4L-/FCU1 и VVTK-F4L-/FCU1 является статистически существенным (p<0,01). Формирование оспин коррелируют с репликацией вируса в хвосте и также с вирулентностью и токсичностью. Инъекция в/в VVTK-I4L-/FCU1 или VVTK-F4L-/FCU1 является менее токсичной, чем с однократным удаленным вирусом TK.
Статистический анализ.
Проводят статистические исследования, используя непараметрический U тест Mann-Whitney и программное обеспечение STATISTICA 7.1 (StatSoft, Inc). Р<0,05, как полагают, является статистически существенным.
Ссылки
- US 5364773 (VIROGENETICS CORPORATION (TROY, NY)) 15.11.1994
- WO 2004/014314 (KIRN DAVID (US)) 19.02.2004
- WO 2004/014314 (KIRN DAVID (US)) 19.02.2004
- US 5364773 (VIROGENETICS CORPORATION (TROY, NY)) 15.11.1994
- WO 93/01281 (US HEALTH)
- WO 2005/007857
- WO 2005/007857
- EP 0998568 A
- EP 0998568 A
- EP 0998568 A
- ЕР 0998568 А
- WO 96/16183
- ЕР 0998568 А
- ЕР 0998568 А
- WO 2006/048768
- HERMISTON. A demand for next-generation oncolytic adenovirases. Current opinion in molecular therapeutics. 2006, vol.8, no.4, p.322-30.
- FISHER. Striking out at disseminated metastases: the systemic delivery of oncolytic viruses. Current opinion in molecular therapeutics. 2006, vol.8, no.4, p.301-13.
- CHERNAJOVSKY, et al. Fighting cancer with oncolytic viruses. British medical journal. 2006, vol.332, no.7534, p.170-2.
- JIANG, et al. Oncolytic adeno viruses as antiglioma agents. Expert review of anticancer therapy. 2006, vol.6, no,5, p.697-708.
- COHEN, et al. ONYX-015. Onyx Pharmaceuticals. Current opinion in investigational drugs. 2001, vol.2, no.12, p.1770-5.
- ROBERTS, et al. Naturally oncolytic viruses. Current opinion in molecular therapeutics. 2006, vol.8, no.4, p.314-21.
- THORNE, et al. Vaccinia virus and oncolytic virotherapy of cancer. Current opinion in molecular therapeutics. 2005, vol.7, no.4, p.359-65.
- STANFORD, et al. Myxoma virus and oncolytic virotherapy: a new biologic weapon in the war against cancer. Expert opinion on biological therapy. 2007, vol.7, no.9, p.1415-25.
- CONO, et al. Smallpox vaccination and adverse reactions. Guidance for clinicians. MMWR. Recommendations and reports: Morbidity and mortality weekly report. Recommendations and reports / Centers for Disease Control. 2003, vol.52, no.RR-4, p.1-28.
- XIANGZHI, et al. Vaccinia virus K1L protein supports viral replication in human and rabbit cells through a cell-type-specific set of its ankyrin repeat residues that are distinct from its binding site for ACAP2. Journal of virology. 2006, vol.353, no.1, p.220-233.
- MCCART, et al. Systemic cancer therapy with a tumor selective vaccinia virus mutant lacking thymidine kinase and vaccinia growth factor genes.. Cancer res.. 2001, no.61, p.8751-57.
- KIM, et al. Systemic armed oncolytic ans immunologic therapy for cancer with JX-594, a targeted poxvirus expressing GM-CSF. Molecular Therapeutic. 2006, no.14, p.361-70.
- SLABAUGH, et al.. Journal of virology. 1988, vol.62, p.519-27.
- TENGELSEN, et al.. Virology. 1988, no.164, p.121-31.
- SCHMITT, et al.. Journal of virology. 1988, no.62. p.1889-97.
- SLABAUGH, et al.. Journal of virology. 1984, no.52, p.507-14.
- SLABAUGH, et al.. Journal of virology. 1984, no.52, p.501-6.
- HOWELL, et al.. Journal of Biological Chemistry. 1992, no.267, p.1705-11.
- ANTOINE.. Virology. 1998, no.244, p.365-396.
- EL OMARI, et al. Structure of vaccinia virus thymidine kinase in complex with dTTP: insights for drug design. BMC structural biology. 2006, no.6, p.22.
- MCCART, et al. Systemic cancer therapy with a tumor-selective vaccinia virus mutant lacking thymidine kinase and vaccinia growth factor genes, cancer research. 2001, vol.61, no.24, p.8751-7.
- PUHLMANN, et al. Vaccinia as a vector for tumor-directed gene therapy: biodistribution of a thymidine kinase-deleted mutant. Cancer gene therapy. 2000, vol.7, no.1, p.66-73.
- GNANT, et al. Systemic administration of a recombinant vaccinia virus expressing the cytosine deaminase gene and subsequent treatment with 5-fluorocytosine leads to tumor-specific gene expression and prolongation of survival in mice. Cancer Research. 1999, vol.59, no.14, p.3396-403.
- MCGEOGH.. Nucleic Acids Research. 1990, no.18, p.4105-10.
- BROYLES.. Virology. 1993, no.195, p.863-5.
- JUND, et al.. Journal of Bacteriology. 1970, no.102, p.607-15.
- BECK, et al.. Journal of Bacteriology. 1972, no.110, p.219-28.
- HOEPRICH, et al.. Journal of Infectious Diseases. 1974, no.130, p.112-18.
- ESDERS, et al.. J. biol. chem.. 1985, no.260, p.3915-22.
- KOECHLIN, et al.. Biochemical pharmacology. 1966, no.15, p.435-46.
- POLAK, et al.. Chemotherapy. 1976, no.22, p.137-53.
- JUND, et al.. Journal of Bacteriology. 1970, no.102, p.607-15.
- KILLSTRUP, et al.. Journal of Bacteriology. 1989, no.171, p.2124-2127.
- HUBER, et al.. Cancer Research. 1993, no.53, p.4619-4626.
- MULLEN, et al.. Proceedings of the National Academy of Sciences of the United States of America. 1992, no.89, p.33-37.
- HUBER, et al.. Proceedings of the National Academy of Sciences of the United States of America. 1994, no.91, p.8302-6.
- MESNIL, et al.. Proceedings of the National Academy of Sciences of the United Slates of America. 1996, no.93, p.1831-35.
- ANDERSEN, et al. Characterization of the upp gene encoding uracil phosphoribosyltransferase of Escherichia coli K12. European Journal of Biochemistry. 1992, no.204, p.51-56.
- MARTINUSSEN, et al. Cloning and characterization of upp, a gene encoding uracil phosphoribosyltransferase from Lactococcus lactis. Journal of Bacteriology. 1994, vol.176, no.21, p.6457-63.
- KIM, et al. Complete sequence of the UPP gene encoding uracil phosphoribosyltransferase from Mycobacterium bovis BCG. Biochemistry and molecular biology international. 1997, vol.41, no.6, p.1117-24.
- MARTINUSSEN, et al. Two genes encoding uracil phosphoribosyltransferase are present in Bacillus subtilis. Journal of Bacteriology. 1995, vol.177, no.1, p.271-4.
- KERN, et al. The FUR1 gene of Saccharomyces cerevisiae: cloning, structure and expression of wild-type and mutant alleles. Gene. 1990, vol.88, no.2, p.149-57.
- CHAKRABARTI.. Biotechniques. 1997, no.23, p.1094-97.
- HAMMOND, et al.. Journal of Virological Methods. 1997, no.66, p.135-38.
- KUMAR.. Virology. 1990, no.179, p.151-8.
| название | год | авторы | номер документа |
|---|---|---|---|
| ПОКСВИРУСНЫЕ ОНКОЛИТИЧЕСКИЕ ВЕКТОРЫ | 2008 |
|
RU2508401C2 |
| ПОКСВИРУСНЫЕ ОНКОЛИТИЧЕСКИЕ ВЕКТОРЫ | 2008 |
|
RU2557312C2 |
| M2-ДЕФЕКТНЫЙ ПОКСВИРУС | 2019 |
|
RU2819245C2 |
| КОМБИНАЦИЯ ОНКОЛИТИЧЕСКОГО ВИРУСА С МОДУЛЯТОРАМИ ИММУНОЛОГИЧЕСКИХ КОНТРОЛЬНЫХ ТОЧЕК | 2015 |
|
RU2705780C2 |
| ОНКОЛИТИЧЕСКИЙ ВИРУС ДЛЯ ЭКСПРЕССИИ МОДУЛЯТОРОВ ИММУНОЛОГИЧЕСКИХ КОНТРОЛЬНЫХ ТОЧЕК | 2015 |
|
RU2696312C2 |
| НОВЫЕ АНТИТЕЛА И НУКЛЕОТИДНЫЕ ПОСЛЕДОВАТЕЛЬНОСТИ И ИХ ПРИМЕНЕНИЕ | 2019 |
|
RU2802450C2 |
| ПЕРСОНАЛИЗИРОВАННАЯ ВАКЦИНА | 2018 |
|
RU2779987C2 |
| Рекомбинантный штамм VV-NS1-dGF вируса осповакцины, продуцирующий белок NS1 парвовируса H-1 и обладающий онколитической активностью в отношении глиобластомы человека | 2018 |
|
RU2692628C1 |
| КОМБИНИРОВАННАЯ ТЕРАПИЯ ДЛЯ ЛЕЧЕНИЯ РАКА ПУТЕМ ВНУТРИВЕННОГО ВВЕДЕНИЯ РЕКОМБИНАНТНОГО MVA И АНТИТЕЛА | 2018 |
|
RU2795103C2 |
| ВИРУСНЫЕ ВАКЦИНЫ | 2017 |
|
RU2778312C2 |













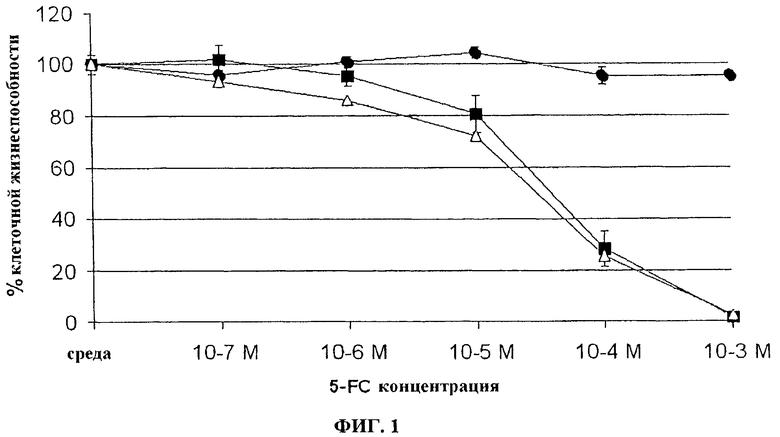
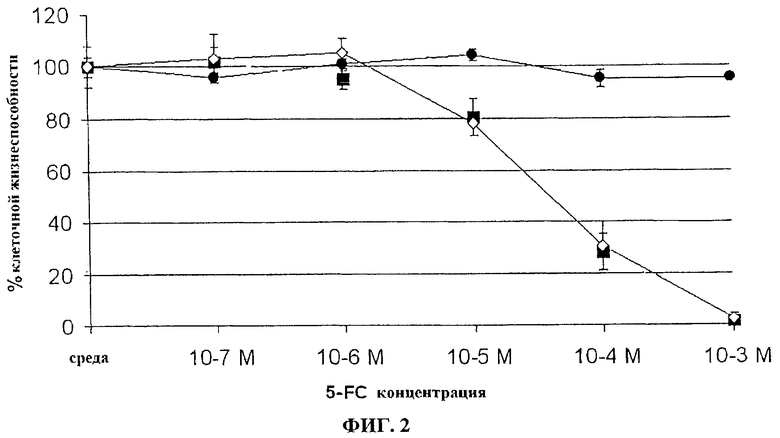
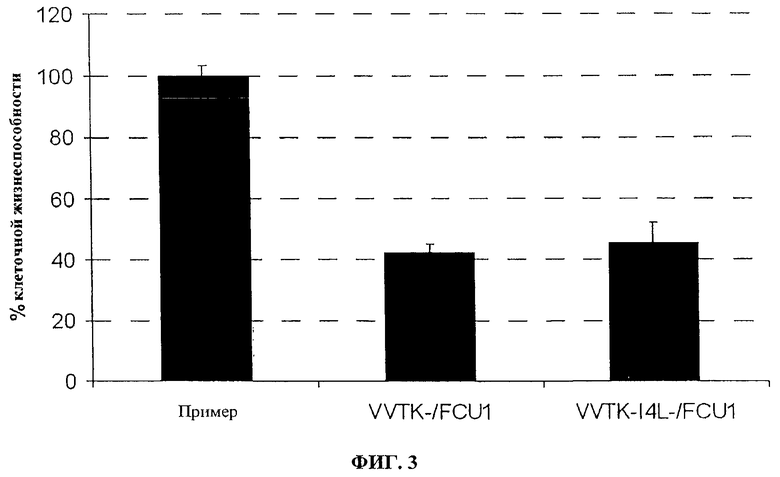
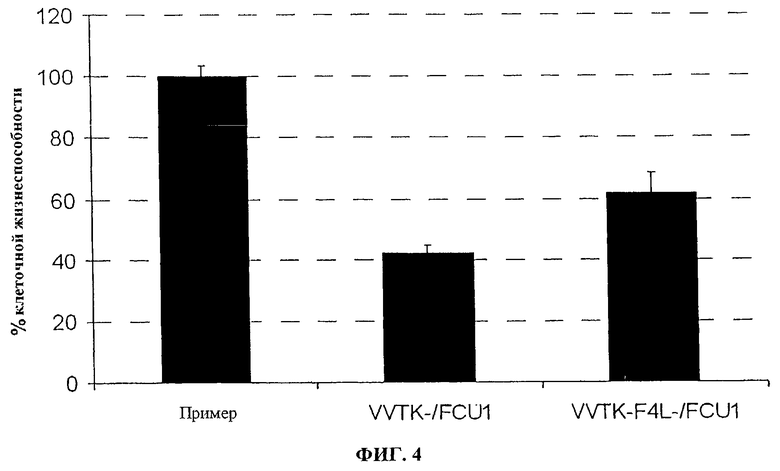
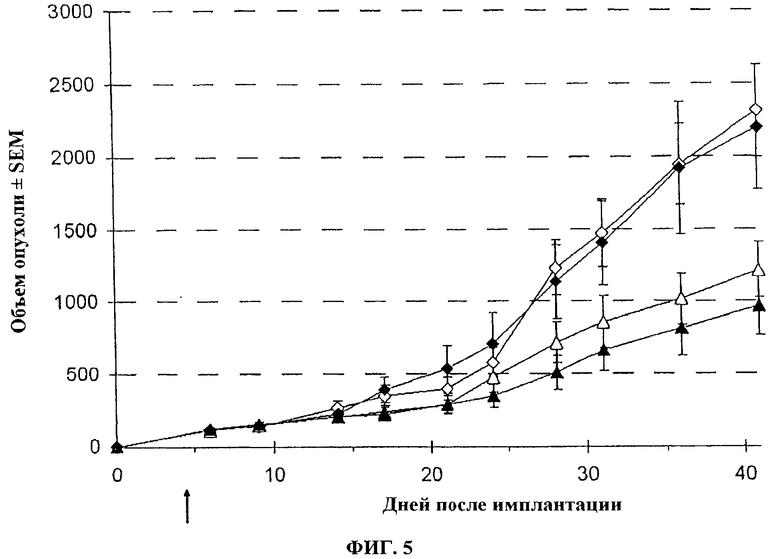
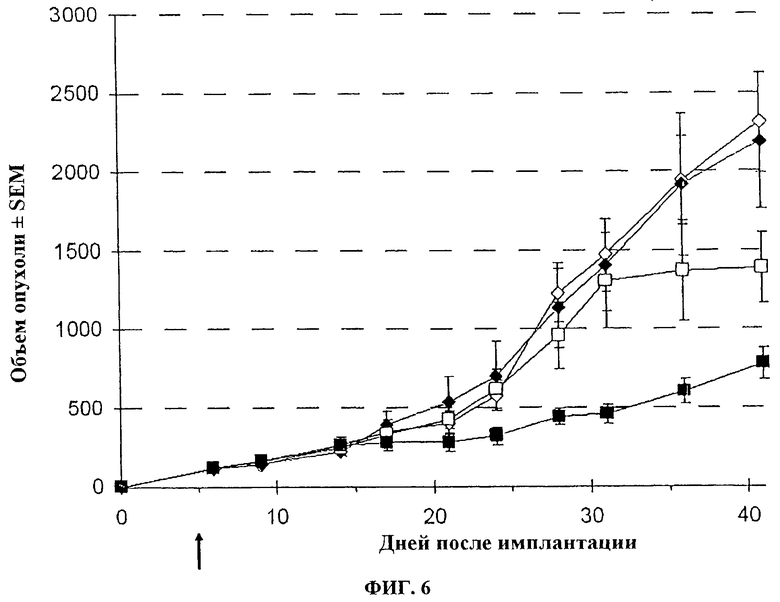
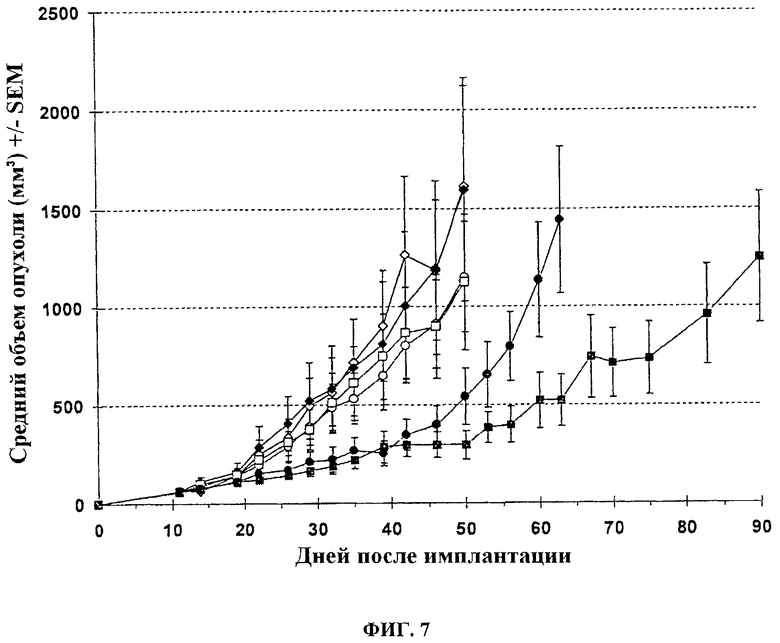
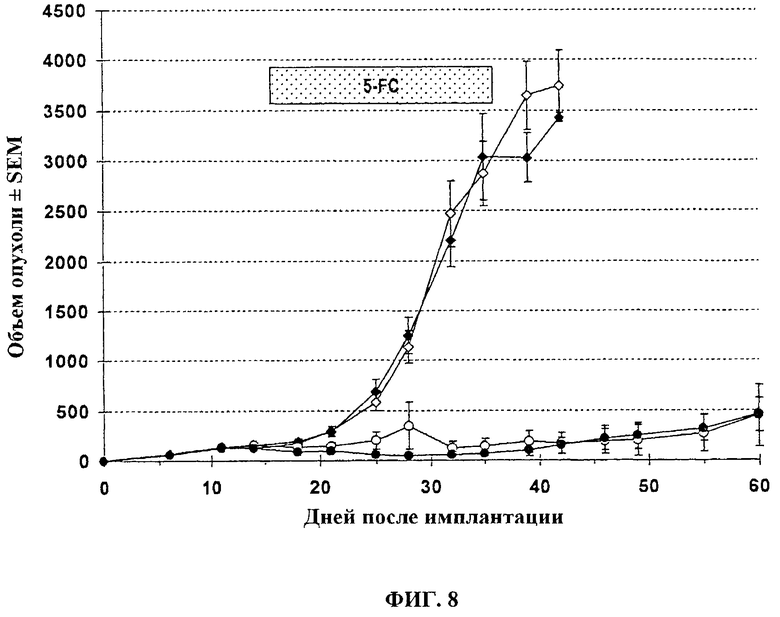
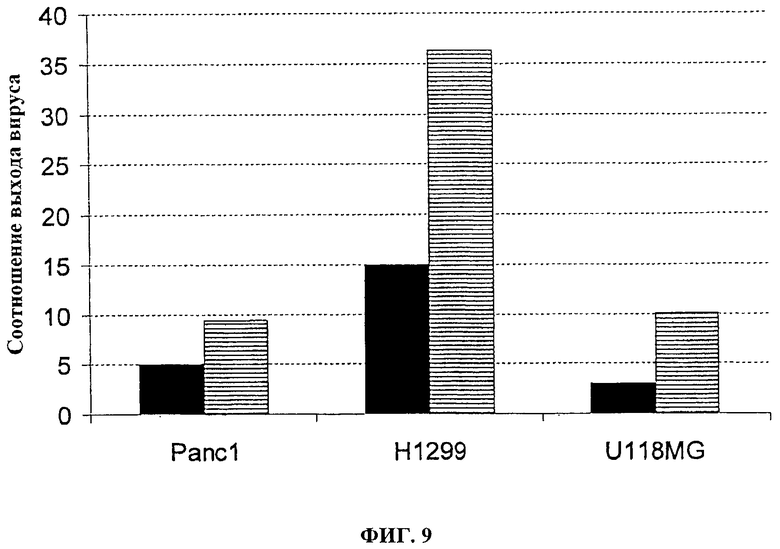
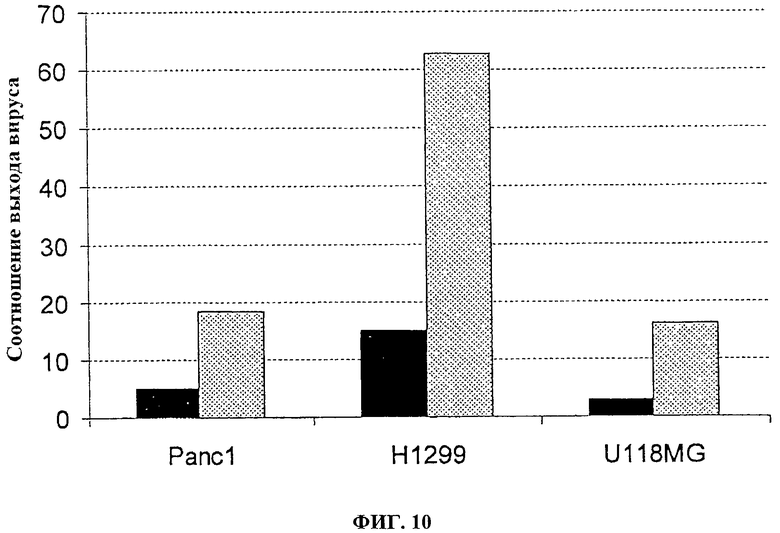
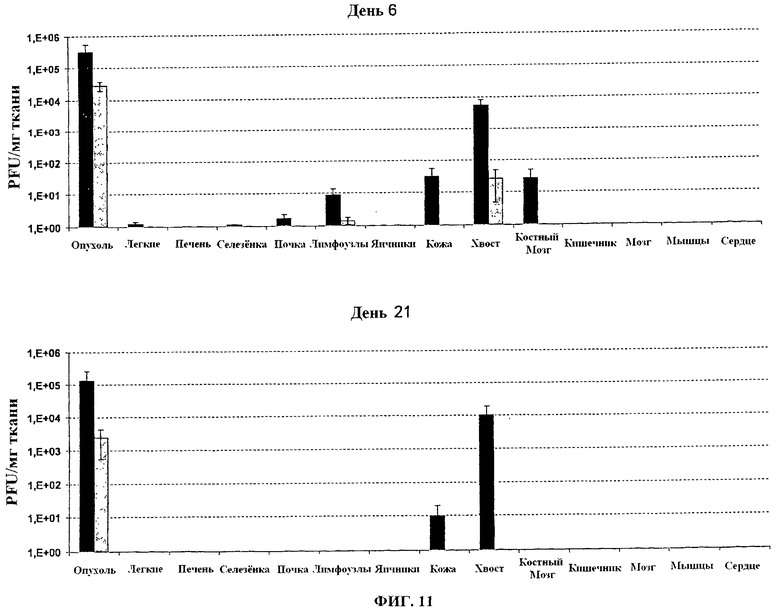
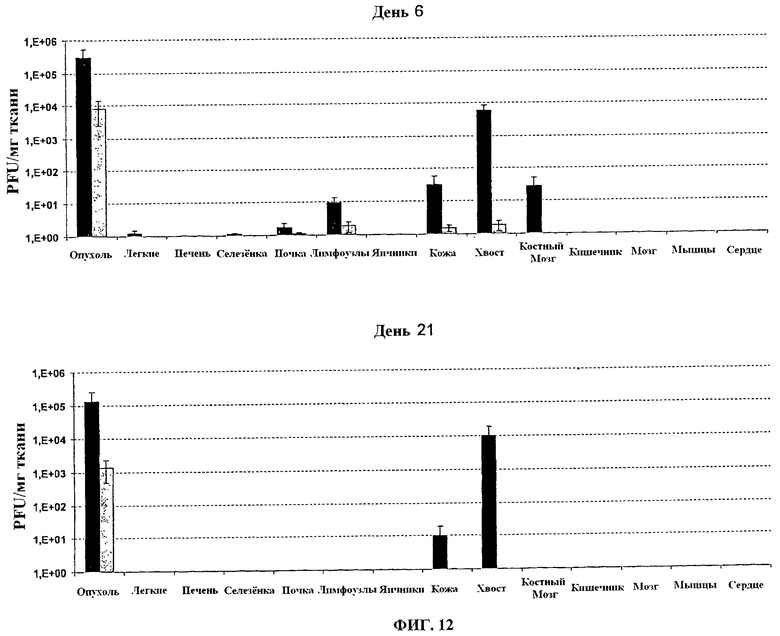
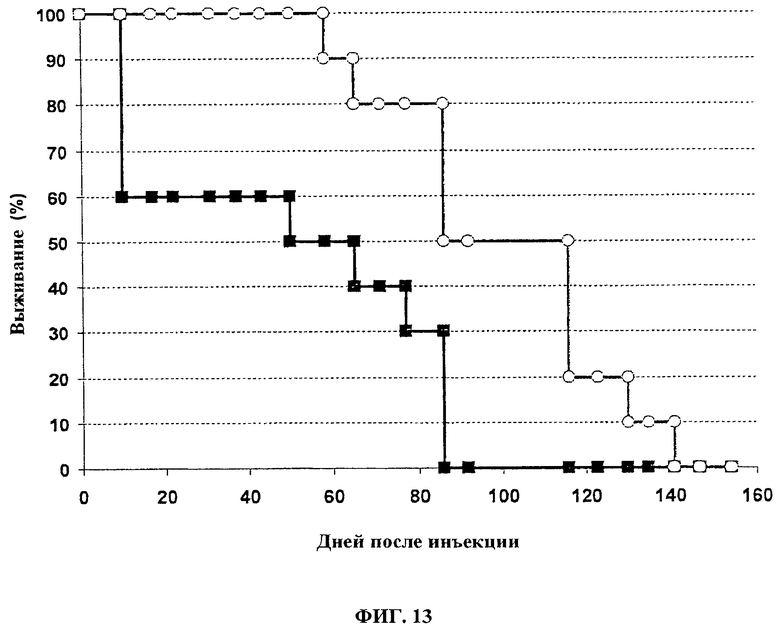
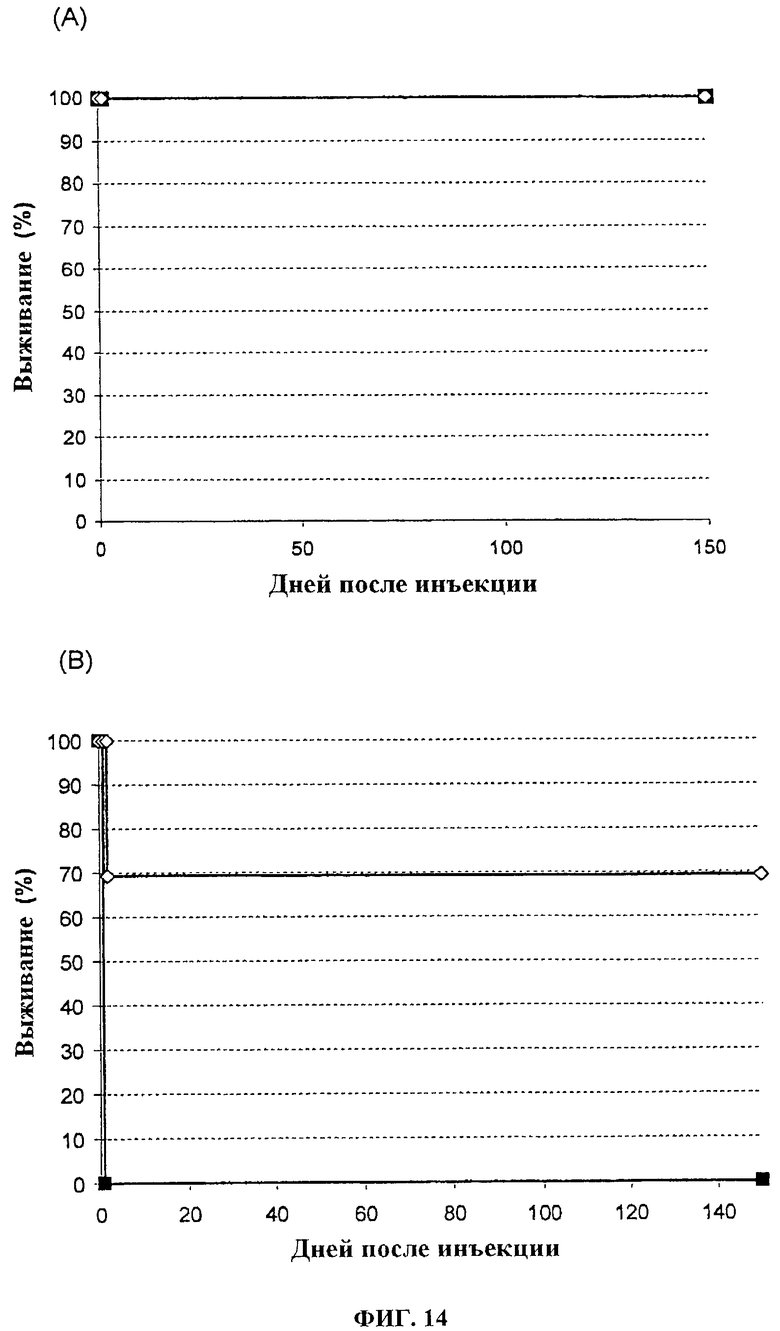
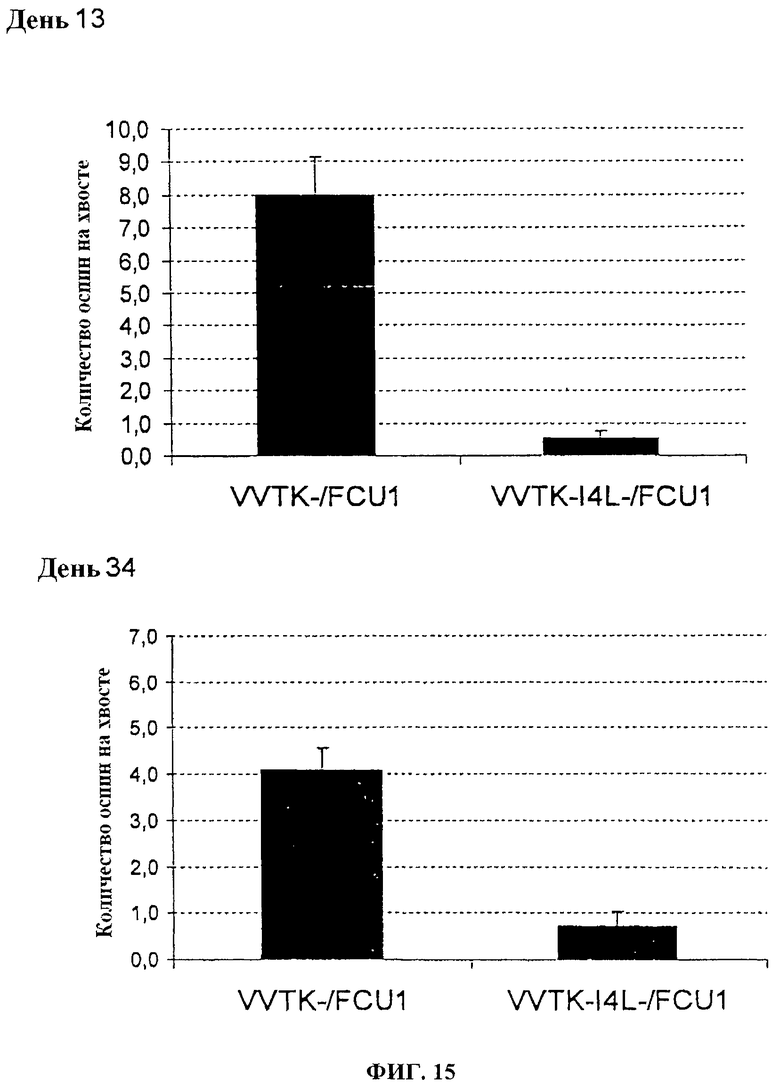
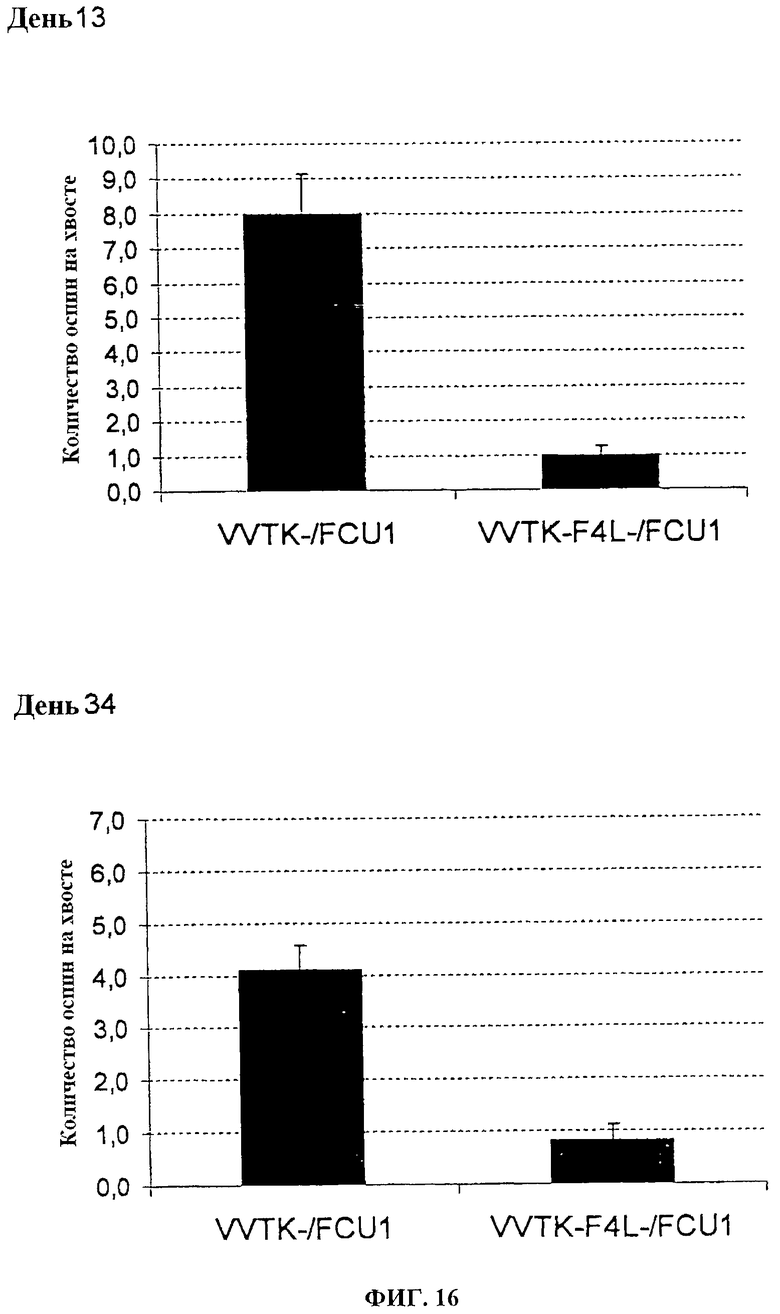
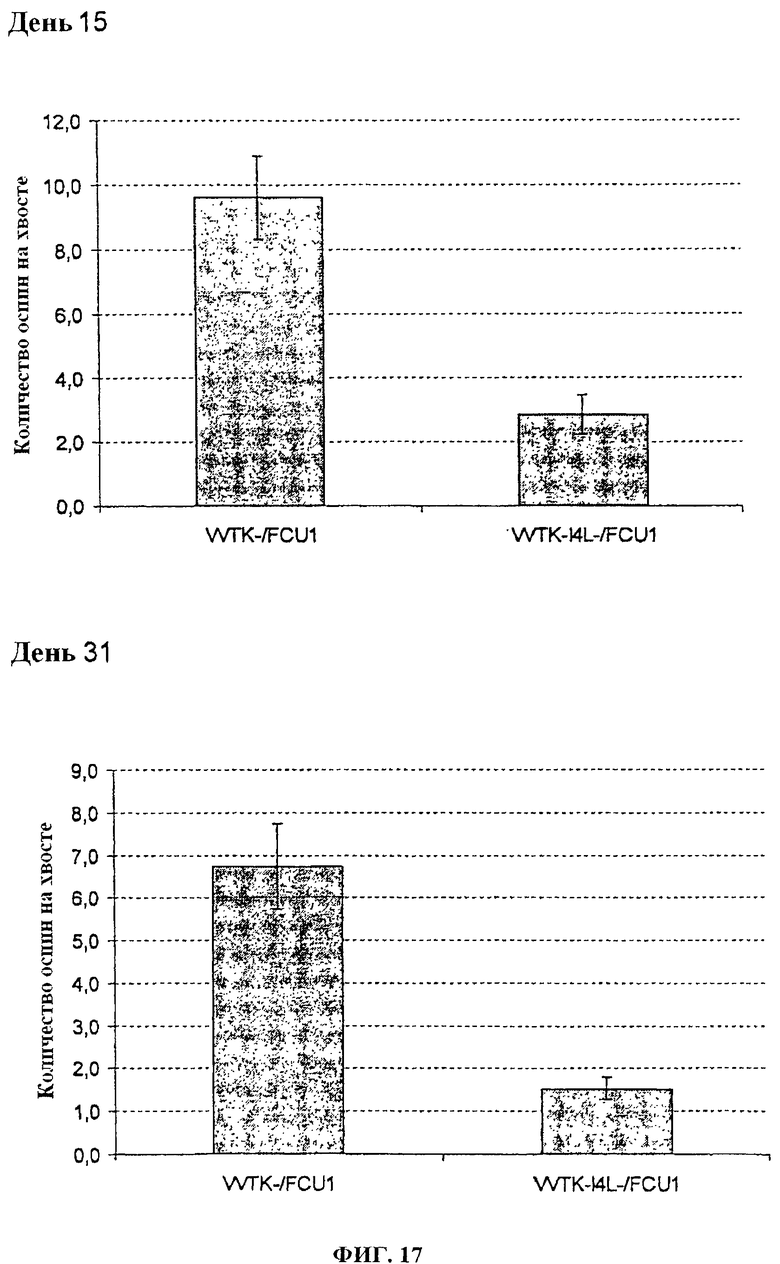
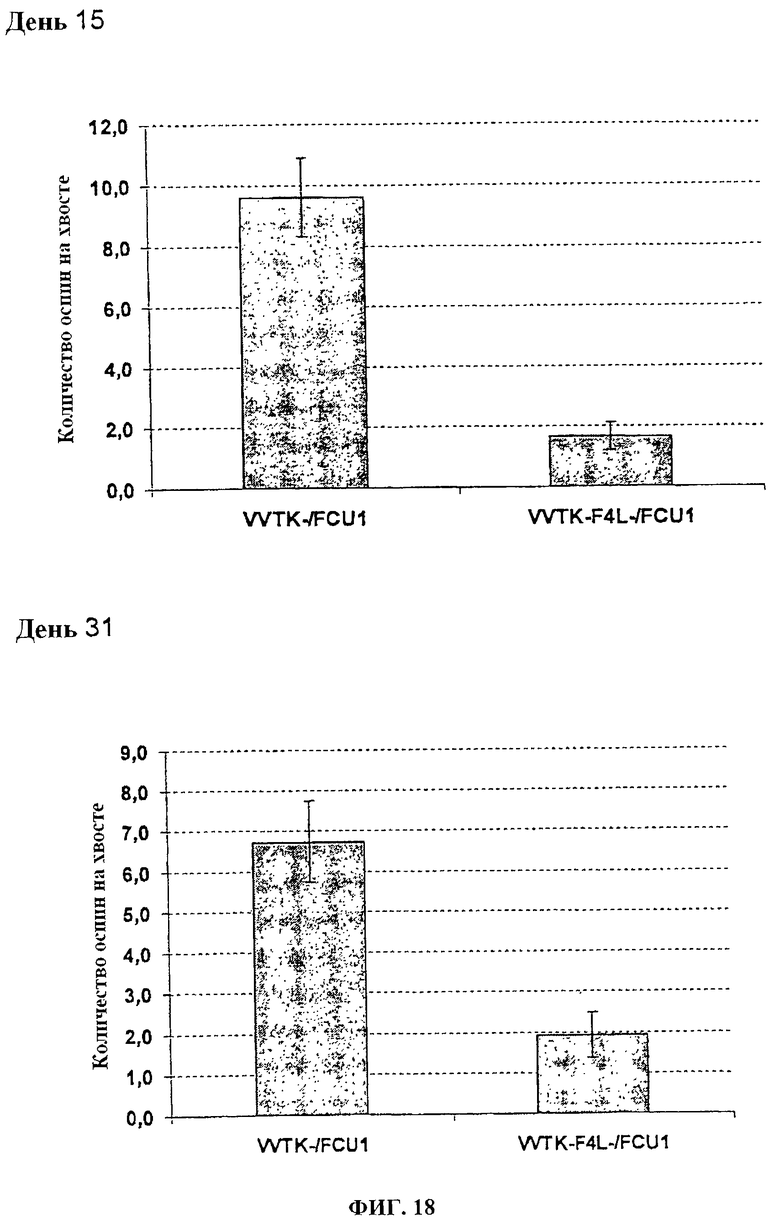
Изобретение относится к области биотехнологии и вирусологии. Предложен поксвирус осповакцины, который обладает онколитической активностью. Поксвирус включает дефектный I4L и/или F4L ген и целевую нуклеиновую кислоту, содержащую суицидальный ген. Также описаны композиция, включающая такой поксвирус, его применение для получения лекарственного средства и способ лечения пролиферативного заболевания или заболевания с повышенной активностью остеокластов с его использованием. Предложенная группа изобретений может быть использована в медицине. 6 н. и 28 з.п. ф-лы, 18 ил., 5 табл.
1. Поксвирус осповакцины, обладающий онколитической активностью и включающий дефектный I4L и/или F4L ген и целевую нуклеиновую кислоту, содержащую суицидальный ген, причем дефектный ген I4L и/или F4L приводит к неспособности вируса к продукции какого-либо белка, обладающего активностью белка, продуцируемого немодифицированным геном.
2. Поксвирус по п.1, где указанный поксвирус дополнительно включает дефектный ген J2R.
3. Поксвирус по п.1 или 2, где указанный поксвирус дополнительно включает дефектный ген F2L.
4. Поксвирус по п.1, где указанный поксвирус принадлежит к подсемейству Chordopoxvirinae.
5. Поксвирус по п.4, где указанный поксвирус принадлежит к виду вируса Vaccinia.
6. Поксвирус по п.5, где указанный поксвирус представляет собой штамм Copenhagen вируса Vaccinia.
7. Поксвирус по п.5, где указанный поксвирус представляет собой штамм WR вируса Vaccinia.
8. Поксвирус по п.1 или 5, в котором указанный суицидальный ген кодирует белок, имеющий по крайней мере активность цитозиндезаминазы.
9. Поксвирус по п.8, в котором указанный суицидальный ген представляет собой FCY1, FCA1 или CodA или их аналог.
10. Поксвирус по п.8, в котором указанный белок, имеющий по крайней мере активность цитозиндезаминазы, является полипептидом FCU1-8, представленным в идентификаторе последовательности SEQ ID N°:2, и его аналоги.
11. Поксвирус по п.1 или 5, в котором указанный суицидальный ген кодирует белок, имеющий по крайней мере одну активность цитозиндезаминазы и одну урацилфосфорибозилтрансферазы.
12. Поксвирус по п.11, в котором указанный суицидальный ген кодирует полипептид, включающий аминокислотную последовательность практически такую, как представлена в идентификаторе последовательности SEQ ID N°:3 (coda::upp), SEQ ID N°:1 (FCU1) или аминокислотную последовательность FCY1: FUR1.
13. Поксвирус по п.1 или 5, в котором указанный поксвирус дополнительно включает нуклеиновокислотную последовательность, включающую ген, кодирующий пермеазу.
14. Поксвирус по п.13, в котором пермеаза является пурин- или цитозинпермеазой S.Cerevisiae.
15. Поксвирус по п.14, в котором пермеазу выбирают из группы, включающей FCY2 и Fur4 и их аналоги.
16. Поксвирус по п.1 или 5, в котором указанный поксвирус дополнительно включает элементы, необходимые для экспрессии целевой нуклеиновой кислоты.
17. Поксвирус по п.13, в котором указанный поксвирус дополнительно включает элементы, необходимые для экспрессии последовательности нуклеиновой кислоты, включающей ген, кодирующий пермеазу.
18. Способ воспроизведения поксвируса по п.1, где в способе:
поксвирус по п.1 вводят в клетку;
указанную клетку культивируют при условиях, которые являются соответствующими для того, чтобы позволить продуцировать указанный поксвирус, и;
указанный поксвирус выделяют из клеточной культуры.
19. Композиция, обладающая онколитической активностью и включающая поксвирус по п.1 в комбинации с фармацевтически приемлемым вспомогательным веществом.
20. Применение поксвируса по п.1 для получения лекарственного средства лечения пролиферативного заболевания или заболевания с повышенной активностью остеокластов.
21. Применение по п.20 для получения лекарственного средства для лечения рака.
22. Способ лечения пролиферативного заболевания или заболевания с повышенной активностью остеокластов, отличающийся тем, что поксвирус по п.1 вводят в организм-хозяин или клетку, которые нуждаются в таком лечении, в терапевтически эффективном количестве.
23. Способ по п.22, где заболеванием является рак или рестеноз.
24. Способ по п.22, где заболеванием является ревматоидный артрит или остеопороз.
25. Способ по п.22, в котором указанный поксвирус или композицию вводят системным путем.
26. Способ по п.22, включающий дополнительную стадию, в которой фармацевтически приемлемые количества пролекарства вводят в указанный организм-хозяин или клетку.
27. Способ по п.24, в котором введение указанного пролекарства проводят предпочтительно спустя по крайней мере 3 дня, более предпочтительно по крайней мере 4 дня и еще более предпочтительно по крайней мере 5 дней после введения указанного поксвируса или композиции.
28. Способ по п.25, в котором введение указанного пролекарства проводят спустя 7 дней после введения указанного поксвируса или композиции.
29. Способ по п.28, где указанный поксвирус или композицию вводят в комбинации с одним или несколькими веществами, которые потенциируют цитотоксический эффект 5-фторцитозина.
30. Способ по п.29, в котором указанные вещества, потенциирующие цитотоксический эффект 5-фторцитозина, представляют собой лекарственные препараты, которые ингибируют ферменты пути для de novo биосинтеза пиримидинов, предпочтительно выбранных из группы, состоящей из PALA, лефлуномида и А771726.
31. Способ по п.29, в котором указанное вещество, потенциирующее цитотоксический эффект 5-фторцитозина, представляет собой метотрексат.
32. Применение поксвируса, включающего дефектный I4L и/или F4L ген, для получения лекарственного средства для лечения пролиферативного заболевания или заболевания с повышенной активностью остеокластов.
33. Применение по п.32, в котором указанный поксвирус дополнительно включает целевую нуклеиновую кислоту.
34. Применение по п.33, в котором целевая нуклеиновая кислота содержит по крайней мере одну целевую последовательность, кодирующую терапевтическую молекулу.
| US 5759553 А, 02.06.1998 | |||
| US 5453364 А, 26.09.1995 | |||
| HOWLEY P.M | |||
| et al., "A vaccinia virus transfer vector using a GUS reporter gene inserted in to the I4L locus" | |||
| Gene, 1996 Jun 26; 172(2):233-237 | |||
| US 6596279 B1, 22.07.2003 | |||
| RU 2006100679 A, 27.07.2007. |

