ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
[001] Настоящее изобретение относится к аттенуированной вакцине на основе поксвирусного вектора для защиты от чикунгуньи и натуральной оспы, вируса Зика и натуральной оспы и/или чикунгуньи, вируса Зика и натуральной оспы.
УРОВЕНЬ ТЕХНИКИ
[002] Библиографические сведения о ссылках в рассматриваемом описании изобретения приведены в конце описания изобретения.
[003] Ссылка в настоящем описании изобретения на любую предыдущую публикацию (или информацию, полученную из нее) или на любой известный объект не является и не должна восприниматься как подтверждение или допущение или любая форма предложения о том, что эта предыдущая публикация (или информация, полученная из нее) или известный объект являются частью общих знаний в области деятельности, к которой относится это описание изобретения.
[004] Все публикации, упомянутые в этом описании изобретения, полностью включены в настоящее описание посредством ссылки.
[005] Семейство поксвирусов включает два подсемейства, Chordopoxvirinae и Entomopoxvirinae. Chordopoxvirinae включает восемь родов, включая Orthopoxviridae, к которым относятся виды, которые инфицируют человека (например, вирус вариолы, возбудитель натуральной оспы, вирус коровьей оспы (из которого сформирована оригинальная противооспенная вакцина, о которой сообщил Дженнер в 1796 г.), вирус осповакцины (используемый в качестве противооспенной вакцины второго поколения) и вирус оспы обезьян), и вирусы Avipoxviridae, к которым относятся виды, которые инфицируют птиц, такие как вирусы оспы птиц и оспы канареек. Помимо их использования в качестве антигенов в противооспенных вакцинах, очень интересно использование рекомбинантных вирусов на основе вируса осповакцины и авипоксвирусов в качестве "базовых" векторов. Как внутрицитоплазматические векторы Orthopoxviridae могут, помимо прочего, доставлять чужеродные антигены в цитоплазму хозяина и пути процессирования антигена, в которых происходит процессирование антигенов в пептиды для презентации на клеточной поверхности. Такие векторы, экспрессирующие чужеродные антигены, используют при разработке вакцин для таких заболеваний, как СПИД, туберкулез, малярия и рак, которые трудно поддаются лечению с помощью других стратегий вакцинации.
[006] Chordopoxvirinae имеют линейные двухцепочечные ДНК-геномы с размерами в диапазоне от 130 п.о. у парапоксвирусов до более 300 п.о. у авипоксвирусов, и их жизненный цикл в хозяине проходит полностью в цитоплазме клетки-хозяина. Поксвирусы работают по существу независимо от их клетки-хозяина и молекул клетки-хозяина, особенно что касается процессов раннего синтеза мРНК. Однако оказывается, что молекулы хозяина используются для инициирования или прекращения средней и поздней вирусной транскрипции. Поксвирусы продуцируют структурно разнообразные "факторы круга хозяев", которые специфически воздействуют и управляют сигнальными путями хозяина для обеспечения клеточных условий, делающих возможной вирусную репликацию. Большинство поксвирусов могут связываться с клетками млекопитающих и инфицировать их, но то, является или нет последующее инфицирование пермиссивным (способным продуцировать инфекционные вирионы) или не пермиссивным (по существу неспособным продуцировать инфекционные вирионы), зависит от конкретного поксвируса и конкретного типа клеток. В настоящее время существует относительно слабое понимание взаимодействий поксвирус-хозяин на молекулярном уровне, в частности генов круга хозяев, и того, какие факторы необходимы для модуляции взаимосвязи для облегчения как вирусного, так и клеточного размножения. Для обзора генов круга хозяев можно сослаться на документ Werden et al. 2008, полностью включенный в настоящее описание.
[007] Наблюдения за штаммами вируса осповакцины в отношении их применения в качестве противооспенных вакцин, а затем и в качестве вирусных векторов, публиковались с начала 1960-х годов до настоящего времени. Некоторые штаммы вируса осповакцины, включая штаммы, используемые в качестве противооспенных вакцин, способны размножаться в человеческих клетках и поэтому представляют риски для здоровья, такие как развитие вирусного энцефалита. С целью разработки более безопасной вакцины штамм вируса осповакцины из Анкары (называемый "CVA") пассировали более 500 раз в нечеловеческих клетках. Во время этого процесса происходит существенное изменение генома вируса осповакцины, включая возникновение по меньшей мере шести значительных делеций по сравнению с исходным геномом CVA. Модифицированный вирус является менее патогенным для человека, но все же способен вызывать защитный иммунный ответ. Этот аттенуированный вирус осповакцины называется MVA (модифицированный вирус осповакцины Анкара) и также классифицируется по количеству пассажей, поскольку оказалось, что вирусы с различными количествами пассажей генетически и фенотипически отличаются. Однако было признано, что MVA515 с количеством пассажей 515 является генетически стабильным. В начале 1990-х годов обнаружили, что штаммы MVA, такие как MVA572 и его производное MVA F6, способны экспрессировать белки вируса осповакцины и гетерологичные (рекомбинантные) белки на высоком уровне в непермиссивных клетках (в которых вирус не размножается), что обеспечивает разработку MVA в качестве вектора для представляющих интерес гетерологичных молекул, таких как кодирующие антигены для доставки вакцин или препаратов.
[008] Недавно были предприняты попытки создания модифицированного вируса осповакцины с качествами MVA путем введения шести известных больших делеций MVA в CVA. Интересно, что это не привело к вирусу с аттенуированными качествами MVA. Было высказано предположение, что причиной наблюдаемой аттенуации может быть отсутствие генов круга хозяев, однако это не было подтверждено (смотри, например, Meyer et al., Journal of General Virology (1991) 72:1031-1038).
[009] Поксвирусы образуют большое семейство вирусов, отличающихся большим линейным дцДНК-геномом, цитоплазматическим местом размножения и сложной морфологией вириона. Вирус осповакцины является типичным вирусом этой группы вирусов и наиболее изученным в отношении вирусного морфогенеза. Вирионы вируса осповакцины выглядят как имеющие форму "бруска" или "яйца" мембрано-связанные частицы со сложной внутренней структурой с имеющим оболочку двояковогнутым кором, окаймленным "боковыми телами". Путь сборки вирионов включает производство мембраны, содержащей полумесяцы, которые развиваются в незрелые вирионы (IV), а затем эволюционируют в зрелые вирионы (MV). В вирионе вируса осповакцины содержится более 70 специфических генных продуктов, причем в настоящее время описано влияние мутаций на сборку вируса осповакцины в более чем 50 специфических генах.
[0010] Вирус осповакцины проникает в клетки путем слияния своих поверхностных мембран с плазматической мембраной клетки-хозяина, высвобождая кор (и боковые тела) в цитоплазму и активируя транскрипционную программу вируса. Коры вирионов содержат полный комплемент кодируемых вирусом ферментов, требующихся для синтеза и модификации ранней мРНК. Ранние гены кодируют ферменты, требующиеся для размножения ДНК, и поэтому при достижения пика экспрессии ранних генов в цитоплазматических сайтах, называемых "фабриками", происходит размножение вирусной ДНК. Ранние гены также кодируют средние факторы транскрипции, а средние гены в свою очередь кодируют поздние факторы транскрипции, так что средние и поздние гены экспрессируются последовательно, обязательно после начала размножения вирусной ДНК. Таким образом, полный комплемент вирусных генов транскрибируется во временном каскаде, причем ранние, средние и поздние классы различаются по класс-специфическим транскрипционным промоторам и кодируемым вирусом факторам транскрипции. Кроме того, только размноженные геномы являются компетентными матрицами для средней и поздней транскрипции. Эти два класса генов вместе кодируют структурные белки вирионов, ферменты вирионов и факторы сборки и необходимы для сборки новых дочерних вирусных частиц.
[0011] Вскоре после захвата и ранней экспрессии вируса внутри клетки образуются специфические для инфекции цитоплазматические домены, которые имеют однородную плотность и иногда окружены цистернами, происходящими из эндоплазматического ретикулума (ER), размер которых увеличивается со временем. Эти домены представляют участки размножения вирусной ДНК и часто называются "вирусными фабриками".
[0012] Вирусная сборка начинается с формирования жестких структур в виде полумесяцев (куполов в трех измерениях) внутри вирусных фабрик. На электронно-микроскопических снимках высокого разрешения внешний слой этих структур в виде полумесяцев состоит из регулярно расположенных выступов, называемых "спикулами". Полумесяцы заметно растут в длину, сохраняя ту же кривизну, пока не станут замкнутыми кругами (сферами в трех измерениях), называемыми незрелыми вирионами (IV). IV заполнены материалом "вироплазмой", который является однородным по плотности, но заметно более электронноплотным, чем окружающая фабрика. По мере формирования IV также происходит захват инкапсидированной ДНК: его видно на электронно-микроскопических снимках как электронноплотный круглый или яйцевидный субдомен внутри IV, называемый "нуклеоидом". IV, которые содержат нуклеоиды конденсированной ДНК, часто называют "IVN". Для морфогенеза IVN до зрелых вирионов (MV) требуется созревание нескольких предшественников вирионных белков посредством протеолитического расщепления. Большинство зрелых вирионов находятся за пределами фабрик и могут существовать в кластерах или на периферии фабрики, или явно отдельно, на значительном расстоянии от ближайшей фабрики.
[0013] Вирионы поксвирусов существуют в трех инфекционных формах: зрелые вирионы (MV), вирионы в оболочке (WV) и внеклеточные вирионы (EV). MV, простейшая форма вируса, представляют собой покрытые мембраной частицы, содержащие двояковогнутый ДНК-содержащий кор, окаймленный боковыми телами, которые заполняют вогнутости кора. MV обычно обнаруживаются исключительно внутри клеток и высвобождаются только путем лизиса клеток. WV состоят из MV, которые окружены двумя дополнительными липидными бислоями, происходящими из цистерн транс-Гольджи. WV, чьи наружные мембраны содержат характерные вирусные белки, являются предшественниками EV и также находятся внутри клетки. EV состоят из WV, которые подверглись экзоцитозу путем слияния самой внешней мембраны WV с плазматической мембраной, из-за чего MV оказываются завернуты в одну дополнительную мембрану. Фракция EV оказывается прикреплена к клеточной поверхности, тогда как ее часть свободно находится в внеклеточной среде. Считается, что EV важны для распространения вируса в организме.
[0014] Чикунгунья представляет собой инфекцию, вызываемую вирусом чикунгуньи. Она характеризуется внезапной лихорадкой, обычно продолжающейся от двух до семи дней и болями в суставах, обычно продолжающимися неделями или месяцами, а иногда годами. Смертность составляет чуть менее 1 на 1000, при этом чаще всего умирают пожилые люди.
[0015] Вирус передается человеку комарами. К резервуарным животным относятся обезьяны, птицы, крупный рогатый скот и грызуны. Этим от отличается от денге, для которого хозяевами являются только приматы.
[0016] Наилучшим средством профилактики является общий контроль численности комаров и предотвращение укусов комаров в странах, где распространено это заболевание. Однако вакцина, которая обеспечила бы защиту от этого заболевания, была бы очень желательна.
[0017] Вирус Зика (ZIKV) является членом рода Flavivurus семейства Flaviviridae и представляет собой положительный одноцепочечный РНК-вирус с плюс-цепью, который передается главным образом комаром Aedes aegypti, который также переносит вирусы денге и чикунгуньи. В меньшей степени вирус Зика может также распространять Aedes albopictus, распространение которого простирается далеко в зоны умеренного климата.
[0018] Вирус Зика был впервые обнаружен у обезьян в лесу Зика в Уганде в 1947 году, и до недавнего времени этот вирус представлял только научный интерес. По-видимому, около 80% инфицированных не проявляют никаких симптомов, а у остальных наблюдается не более чем легкое заболевание, которое проходит в течение нескольких дней. Первоначально проявлявшийся только небольшими вспышками в Африке и Азии, этот вирус распространился в Тихом океане в 2007 году на остров Яп в Микронезии. В 2013 году вспышка во Французской Полинезии вызвала 28000 заражений. Считается, что нынешняя вспышка в Америке происходит из Полинезии. Вирус, возможно, был завезен в Бразилию во время чемпионата мира FIFA 2014 года или гонки на каноэ, в которой участвовали полинезийские спортсмены. С тех пор этот вирус распространился по нескольким латиноамериканским странам, включая Колумбию, Мексику, Парагвай и Венесуэлу.
[0019] До сих пор были описаны две генетические линии ZIKV, африканская и азиатская. Штаммы, выделенные из образцов в Бразилии в период между 2015 и 2016 годами, напоминали азиатские штаммы, в частности штамм из Французской Полинезии [[Baronti et al., 2014, Complete coding sequence of Zika virus from a French Polynesia outbreak in 2013. Genome Announc 2: e00500-e00514; Brasil et al., 2016, Zika virus outbreak in Rio de Janeiro, Brazil: Clinical characterization, epidemiological and virological aspects. PLoS Negl Trop Dis 10:e0004636; Faria et al., 2016, Zika virus in the Americas: Early epidemiological and genetic findings. Science 352:345-349; Giovanetti et al., 2016, Zika virus complete genome from Salvador, Bahia, Brazil. Infect Genet Evol.].
[0020] Эталонный штамм ZIKV африканской линии MR766 отличается от азиатской линии тем, что в нем удален консервативный сайт гликозилирования в ASN 153/154 белка оболочки (смотри фиг. 3 в документе Sirohi et al 2016, The 3.8 Å resolution cryo-EM structure of Zika virus. Science, 352 (6284): 467-70), что приводит к негликозилированию белка E в штамме MR766. Такое отсутствие гликозилирования, по-видимому, не влияет на инфекционность вируса Зика, но может играть роль в вирулентности и может быть причиной того, что азиатские штаммы кажутся более вирулентными, чем MR766, однако на сегодняшний день это не доказано.
[0021] При разработке вакцин на основе SCV антигенные последовательности для этих вакцин получали на основе азиатской линии. В качестве вакцинного антигена вакцины использовали последовательность полипротеина PrME (prMembrane и оболочка).
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
[0022] Авторы настоящего изобретения обнаружили, что при использовании аттенуированного поксвируса, который сконструирован таким образом, что его геном содержит последовательность нуклеиновой кислоты, кодирующую субгеномный полипротеин 26S вируса чикунгуньи и/или PrME вируса Зика, может быть получена композиция, которая снижает риск инфицирования чикунгуньей и натуральной оспой, инфицирования вирусом Зика и натуральной оспой и/или инфицирования чикунгуньей, вирусом Зика и натуральной оспой.
[0023] Соответственно, в первом аспекте настоящее изобретение относится к композиции для повышения иммунного ответа у животного, которая снижает риск инфицирования чикунгуньей и натуральной оспой, причем данная композиция содержит фармацевтически приемлемый носитель и аттенуированный поксвирус, причем геном поксвируса содержит последовательность нуклеиновой кислоты, кодирующую субгеномный полипротеин 26S вируса чикунгуньи.
[0024] Во втором аспекте настоящее изобретение относится к композиции для повышения иммунного ответа у животного, которая снижает риск инфицирования вирусом Зика и инфицирования натуральной оспой, причем данная композиция содержит фармацевтически приемлемый носитель и аттенуированный поксвирус, причем геном поксвируса содержит последовательность нуклеиновой кислоты, кодирующую полипротеин PrME вируса Зика.
[0025] В третьем аспекте настоящее изобретение относится к композиции для повышения иммунного ответа у животного, которая снижает риск инфицирования чикунгуньей, вирусом Зика и натуральной оспой, причем данная композиция содержит фармацевтически приемлемый носитель и аттенуированный поксвирус, причем геном поксвируса содержит последовательность нуклеиновой кислоты, кодирующую субгеномный полипротеин 26S вируса чикунгуньи, и последовательность нуклеиновой кислоты, кодирующую полипротеин PrME вируса Зика.
[0026] В четвертом аспекте настоящее изобретение относится к способу индуцирования у пациента защитного иммунного ответа на инфицирование чикунгуньей и натуральной оспой, натуральной оспой и вирусом Зика и/или инфицирование чикунгуньей, натуральной оспой и вирусом Зика, включающему введение пациенту композиции в соответствии с первым, вторым или третьим аспектами настоящего изобретения.
[0027] В пятом аспекте настоящее изобретение относится к применению композиции в соответствии с первым, вторым или третьим аспектами настоящего изобретения в получении лекарственного средства для индуцирования у пациента защитного иммунного ответа на инфицирование чикунгуньей и натуральной оспой, натуральной оспой и вирусом Зика и/или инфицирование чикунгуньей, натуральной оспой и вирусом Зика.
КРАТКОЕ ОПИСАНИЕ ФИГУР
[0028] Фигура 1. Кассета гомологичной рекомбинации SCV301A.
[0029] Фигура 2. Мишени F1 и F2 для гомологичной рекомбинации в геноме VACV-COP.
[0030] Фигура 3. Сайт вставки CHIKV-26S в SCV301A.
[0031] Фигура 4. Удаление Ecogpt и EGFP с помощью гомологичной рекомбинации.
[0032] Фигура 5. Плазмидные карты pTC29.
[0033] Фигура 6. HR-кассета делеции.
[0034] Фигура 7. HR-кассета делеции B7R-B8R.
[0035] Фигура 8. Способ действия гомологичной рекомбинации с трансдоминантной селекцией.
[0036] Фигура 9. HR-кассета ZIKR.
ПОДРОБНОЕ ОПИСАНИЕ
[0037] Рассматриваемое изобретение не ограничено конкретными процедурами или средствами, конкретными композициями средств и различными медицинскими методологиями, так как они могут изменяться. Терминология, используемая в настоящем описании, предназначена только для описания конкретных вариантов осуществления и не предназначена для ограничения. Если не указано иное, все технические и научные термины, используемые в настоящем описании, имеют значения, хорошо понятные специалисту в области техники, к которой относится настоящее изобретение.
[0038] Все материалы и способы, аналогичные или эквивалентные описанным в настоящем описании, можно использовать для осуществления на практике или тестирования настоящего изобретения. Практикующим врачам следует конкретно рассмотреть Sambrook et al., (1989) Molecular Cloning: A Laboratory Manual, 2nd ed., Cold Spring Harbor Press, Plainsview, N.Y.; Ausubel et al. (1999) Current Protocols in Molecular Biology (Supplement 47), John Wiley & Sons, New York; Murphy et al. (1995) Virus Taxonomy Springer Verlag:79-87, Mahy Brian WJ and Kangro O Hillar (Eds): Virology Methods Manual 1996, Academic Press; Davison AJ and Elliott RM (Eds): Molecular Virology, A practical Approach 1993, IRL Press at Oxford University Press; Perkus et al., Virology (1990) 179(1):276-86 или определения и термины данной области техники и другие способы, известные специалисту в данной области техники.
[0039] Хотя для осуществления на практике или тестирования настоящего изобретения можно использовать любые способы и материалы, аналогичные или эквивалентные описанным в настоящем описании, описаны предпочтительные способы и материалы. Для целей настоящего изобретения следующие термины определены ниже.
[0040] Во всем тексте описания, если контекст не требует иного, слова "содержать", "содержит" и "содержащий" следует понимать как подразумевающие включение указанного этапа или элемента или группы этапов или элементов, но не исключение какого-либо другого этапа или элемента или группы этапов или элементов. Таким образом, использование термина "содержащий" и т.п. указывает на то, что перечисленные элементы являются необходимыми или обязательными, но другие элементы являются необязательными и могут присутствовать или не присутствовать. В контексте аттенуированных ортопоксвирусных векторов данные векторы модифицированы для аттенуации благодаря тому, что они содержат делецию незаменимого гена созревания или сборки, но также включены и дополнительные модификации, например направленные на антиген или другой белок.
[0041] "Состоит из" обозначает включает и ограничен тем, что следует за оборотом "состоит из". Таким образом, оборот "состоит из" указывает на то, что перечисленные элементы необходимы или обязательны, и что не могут присутствовать никакие другие элементы. "Состоит по существу из" обозначает включает любые элементы, перечисленные после данного оборота, и ограничен другими элементами, которые не препятствуют или не вносят вклад в активность или действие, указанные в раскрытии для перечисленных элементов. Таким образом, оборот "состоит по существу из" указывает на то, что перечисленные элементы необходимы или обязательны, но другие элементы необязательны и могут присутствовать или не присутствовать в зависимости от того, влияют ли они на активность или действие перечисленных элементов.
[0042] В рамках изобретения формы единственного числа включают множественное число, если контекст прямо не требует обратного. Таким образом, например, ссылка на "клетку" включает одну клетку, а также две или более клетки; ссылка на "организм" включает один организм, а также два или более организма; и т.д. В некоторых вариантах осуществления единственное число обозначает "один или больше один".
[0043] В рамках изобретения "и/или" относится и охватывает любые возможные комбинации одного или нескольких соответствующих перечисленных элементов, а также отсутствие комбинаций при альтернативной интерпретации (или).
[0044] "Аттенуация" или "аттенуированный", в рамках изобретения, обозначает снижение вирулентности вирусного вектора. Вирулентность определяется как способность вируса вызывать заболевание у конкретного хозяина. Поксвирусный вектор, который не способен продуцировать инфекционные вирусы, может первоначально инфицировать клетки, но по существу не способен полностью реплицироваться или размножаться в хозяине или вызывать состояние. Это желательно, поскольку вектор доставляет свой белок или нуклеиновую кислоту в цитоплазму клетки-хозяина, но не наносит вреда пациенту.
[0045] "Контрольный элемент" или "контрольная последовательность" означают последовательности нуклеиновой кислоты (например, ДНК), необходимые для экспрессии функционально связанной кодирующей последовательности в конкретном поксвирусе, векторе, плазмиде или клетке. Контрольные последовательности, которые подходят для эукариотических клеток, включают последовательности контроля транскрипции, такие как промоторы, сигналы полиаденилирования, энхансеры транскрипции, последовательности контроля трансляции, такие как энхансеры трансляции и внутренние сайты связывания рибосом (IRES), последовательности нуклеиновой кислоты, которые модулируют стабильность мРНК, а также направляющие последовательности, которые направляют продукт, кодируемый транскрибируемым полинуклеотидом, во внутриклеточный компартмент внутри клетки или во внеклеточную среду.
[0046] Там, где приводятся последовательности, включены соответствующие последовательности. "Соответствует", "соответствуя" или "соответствующий" означают последовательность нуклеиновой кислоты, которая демонстрирует существенную идентичность последовательности с эталонной последовательностью нуклеиновой кислоты (например, идентичность последовательности со всей или части эталонной последовательности нуклеиновой кислоты по меньшей мере приблизительно 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 97, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99% или даже вплоть до 100%), или аминокислотную последовательность, которая демонстрирует существенные сходство или идентичность последовательности с эталонной аминокислотной последовательностью (например, сходство или идентичность последовательности со всей или частью эталонной аминокислотной последовательности по меньшей мере 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 97, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99% или даже вплоть до 100%).
[0047] "Эффективное количество" в контексте лечения или профилактики некоторого состояния или для модулирования иммунного ответа на целевой антиген или организм обозначает введение количества средства (например, аттенуированного ортопоксвирусного вектора, как описано в настоящем описании) или содержащей его композиции нуждающемуся в таком лечении или профилактике индивидууму, или в виде одной дозы, или как части серии, которое эффективно для предупреждения возникновения симптома, контроля таких симптомов и/или лечения существующих симптомов этого состояния или для модулирования иммунного ответа на целевой антиген или организм. Эффективное количество будет варьироваться в зависимости от состояния здоровья и физического состояния подлежащего лечению индивидуума, таксономической группы подлежащего лечению индивидуума, состава композиции, оценки медицинской ситуации и других соответствующих факторов. Ожидается, что данное количество будет попадать в относительно широкий диапазон, который может быть определен с помощью рутинных испытаний.
[0048] В рамках изобретения, термины "кодируют", "кодирующий" и т.п. относятся к способности нуклеиновой кислоты предоставлять другую нуклеиновую кислоту или полипептид. Например, говорят, что последовательность нуклеиновой кислоты "кодирует" полипептид, если она может быть транскрибирована и/или транслирована с получением этого полипептида, или если она может быть процессирована в форму, которая может быть транскрибирована и/или транслирована с получением этого полипептида. Такая последовательность нуклеиновой кислоты может включать в себя кодирующую последовательность или как кодирующую последовательность, так и некодирующую последовательность. Таким образом, термины "кодируют", "кодирующий" и т.п. включают РНК-продукт, получаемый в результате транскрипции молекулы ДНК, белок, получаемый в результате трансляции молекулы РНК, белок, получаемый в результате транскрипции молекулы ДНК с образованием РНК-продукта и последующей трансляции РНК-продукта, или белок, получаемый в результате транскрипции молекулы ДНК с получением РНК-продукта, процессирования РНК-продукта с получением процессированного РНК-продукта (например, мРНК) и последующей трансляции процессированного РНК-продукта.
[0049] Термин "эндогенный" относится к гену или последовательности нуклеиновой кислоты или сегменту, которые обычно можно обнаружить в организме-хозяине.
[0050] Термины "экспрессируемый", "экспрессированный" и их варианты относятся к способности клетки транскрибировать нуклеотидную последовательность в РНК и, необязательно, транслировать мРНК для синтеза пептида или полипептида, которые обеспечивают биологическую или биохимическую функцию.
[0051] В рамках изобретения, термин "ген" включает молекулу нуклеиновой кислоты, которую можно использовать для получения мРНК, необязательно при добавлении элементов, способствующих этому процессу. Гены могут быть или не быть использованы для получения функционального белка. Гены могут включать в себя как кодирующие, так и некодирующие области (например, интроны, регуляторные элементы, промоторы, энхансеры, последовательности терминации и 5' и 3'-нетранслируемые области).
[0052] Термины "гетерологичная последовательность нуклеиновой кислоты", "гетерологичная нуклеотидная последовательность", "гетерологичный полинуклеотид", "чужеродный полинуклеотид", "экзогенный полинуклеотид" и т.п. используют взаимозаменяемо для обозначения любой нуклеиновой кислоты (например, нуклеотидной последовательности, содержащей IRES), которую вводят в геном организма с помощью экспериментальных манипуляций, и которая может включать в себя генные последовательности, обнаруживаемые в этом организме, при условии, что введенный ген содержит некоторую модификацию (например, точечную мутацию, делецию, замену или добавление по меньшей мере одного нуклеотида, наличие сайта расщепления эндонуклеазой, наличие сайта loxP и т.д.) по сравнению с вирусной геномной последовательностью до модификации.
[0053] Термины "гетерологичный полипептид", "чужеродный полипептид" и "экзогенный полипептид" используют взаимозаменяемо для обозначения любого пептида или полипептида, который кодируется "гетерологичной последовательностью нуклеиновой кислоты", "гетерологичной нуклеотидной последовательностью", "гетерологичным полинуклеотидом", "чужеродным полинуклеотидом" и "экзогенным полинуклеотидом", как определено выше.
[0054] Термин "защитный иммунный ответ" обозначает иммунный ответ, который предотвращает или снижает риск или тяжесть инфицирования чикунгуньей и/или натуральной оспой.
[0055] Поксвирусный вектор согласно настоящему изобретению, предпочтительно, размножается в клетке млекопитающего. Подробности о клетках млекопитающих, которые можно использовать в настоящем изобретении, представлены в документе PCT/AU2014/050330, раскрытие которого включено в настоящее описаниее посредством перекрестной ссылки.
[0056] В некоторых вариантах осуществления клетка млекопитающего представляет собой клетку человека, клетку примата, клетку хомяка или клетку кролика.
[0057] Клетки могут быть одноклеточными или могут быть выращены в тканевой культуре в виде жидких культур, монослоев и т.п. Клетки-хозяева также могут быть получены непосредственно или опосредованно из тканей или могут существовать в организме, включая животных.
[0058] Следует понимать, что "индуцирование" иммунного ответа, как рассматривается в настоящем описании, включает вызывание или стимуляцию иммунного ответа и/или усиление уже существующего иммунного ответа.
[0059] Термин "функционально соединенный" или "функционально связанный", в рамках изобретения, относится к сопоставлению, в котором компоненты, описанные таким образом, находятся во взаимосвязи, позволяющей им функционировать надлежащим образом. Например, контрольная последовательность транскрипции, "функционально связанная" с кодирующей последовательностью, относится к позиционированию и/или ориентации контрольной последовательности транскрипции по отношению к кодирующей последовательности для обеспечения экспрессии кодирующей последовательности в условиях, совместимых с контрольной последовательностью транскрипции. В другом примере, IRES, функционально соединенный с кодирующей последовательностью ортопоксвируса, относится к позиционированию и/или ориентации IRES по отношению к кодирующей последовательности ортопоксвируса для обеспечения кэп-независимой трансляции кодирующей последовательности ортопоксвируса.
[0060] В рамках изобретения, термины "открытая рамка считывания" и "ORF" используют взаимозаменяемо для обозначения аминокислотной последовательности, кодируемой между кодонами инициации и терминации трансляции кодирующей последовательности. Термины "инициирующий кодон" (например, ATG) и "терминирующий кодон" (например, TGA, TAA, TAG) относятся к блоку из трех смежных нуклеотидов ("кодону") в кодирующей последовательности, который определяет инициацию и терминацию цепи, соответственно, при синтезе белка (трансляция мРНК).
[0061] Термины "полинуклеотид", "полинуклеотидная последовательность", "нуклеотидная последовательность", "нуклеиновая кислота" или "последовательность нуклеиновой кислоты", в рамках изобретения, обозначают мРНК, РНК, кРНК, кДНК или ДНК. Этот термин обычно относится к полимерной форме нуклеотидов длиной по меньшей мере 10 оснований, или рибонуклеотидов, или дезоксинуклеотидов, или модифицированной формы нуклеотидов любого типа. Термин включает одно- и двухцепочечные формы РНК или ДНК.
[0062] "Полипептид", "пептид", "белок" и "белковая молекула" используют взаимозаменяемо в настоящем описании для обозначения молекулы, содержащей или состоящей из полимера аминокислотных остатков, и ее вариантов и синтетических аналогов. Таким образом, эти термины применяются к аминокислотным полимерам, в которых один или несколько аминокислотных остатков являются синтетическими не встречающимися в природе аминокислотами, такими как химический аналог соответствующей встречающейся в природе аминокислоты, а также к встречающимся в природе аминокислотным полимерам.
[0063] В рамках изобретения, термин "рекомбинантный" применительно к "молекулам нуклеиновой кислоты", "полинуклеотидам" и т.п. следует понимать как обозначающий искусственные структуры нуклеиновой кислоты (т.е. нереплицирующиеся кДНК или РНК; или репликоны, самореплицирующиеся кДНК или РНК), которые могут быть транскрибированы и/или транслированы в клетках-хозяевах или бесклеточных системах, описанных в настоящем описании. В вектор могут быть введены рекомбинантные молекулы нуклеиновой кислоты или полинуклеотиды. Можно использовать невирусные векторы, такие как плазмидные векторы экспрессии, или вирусные векторы. Специалисту известны тип векторов и методика введения конструкций нуклеиновой кислоты в соответствии с настоящим изобретением. Молекула нуклеиновой кислоты или полинуклеотид в соответствии с настоящим изобретением не встречаются в природе в конфигурации, описанной в настоящем изобретении. Другими словами, гетерологичная нуклеотидная последовательность не объединяется в природе с элементами родительского вирусного генома (например, промотором, ORF, сигналом полиаденилирования, рибозимом).
[0064] В рамках изобретения, термин "рекомбинантный вирус" следует понимать как ссылку на "родительский вирус", содержащий по меньшей мере одну гетерологичную последовательность нуклеиновой кислоты.
[0065] Термин "идентичность последовательностей", в рамках изобретения, относится к той степени, в которой последовательности идентичны по нуклеотидам или по аминокислотам в окне сравнения. Таким образом, "процентную величину идентичности последовательностей" рассчитывают путем сравнения двух оптимально выровненных последовательностей по окну сравнения, определения количества положений, в которых идентичное основание нуклеиновой кислоты (например, A, T, C, G, I) или идентичный аминокислотный остаток (например, Ala, Pro, Ser, Thr, Gly, Val, Leu, Ile, Phe, Tyr, Trp, Lys, Arg, His, Asp, Glu, Asn, Gln, Cys и Met) встречается в обеих последовательностях, для получения количества совпадающих положений, деления количества совпадающих положений на общее количество положений в окне сравнения (т.е. на размер окна) и умножения результата на 100 для получения процентной величины идентичности последовательностей. Для целей настоящего изобретения под "идентичностью последовательностей" следует понимать "процентную величину совпадений", рассчитанную с помощью компьютерной программы DNASIS (версия 25 для Windows; доступна от Hitachi Software engineering Co., Ltd., South San Francisco, Калифорния, США) с использованием стандартных значений по умолчанию, используемых в справочном руководстве, прилагаемом к программному обеспечению.
[0066] Термины "сигнальная последовательность" или "сигнальный пептид" относятся к короткому (длиной от приблизительно 3 до приблизительно 60 аминокислот) пептиду, который направляет ко- или посттрансляционный транспорт белка из цитозоля к определенным органеллам, таким как, например, ядро, митохондриальный матрикс и эндоплазматический ретикулум. Для белков с нацеленными на ER сигнальными пептидами сигнальные пептиды обычно отщепляются от формы-предшественника сигнальной пептидазой после того, как белки транспортируются в ER, и полученные белки перемещаются по секреторному пути к их внутриклеточному (например, аппарат Гольджи, клеточная мембрана или клеточная стенка) или внеклеточному местоположению. "Нацеленные на ER сигнальные пептиды", в рамках изобретения, включают аминоконцевые гидрофобные последовательности, которые обычно ферментативно удаляются после введения части или всего белка через мембрану ER в просвет ER. Таким образом, в данной области техники известно, что сигнальная форма-предшественник последовательности может присутствовать как часть формы-предшественника белка, но обычно отсутствует в зрелой форме белка.
[0067] "Сходство" относится к процентному количеству аминокислот, которые являются идентичными или представляют собой консервативные замены, как определено в таблице A ниже. Сходство можно определять с помощью программ сравнения последовательностей, таких как GAP (Deveraux et al. 1984, Nucleic Acids Research, 12: 387-395). Таким образом, последовательности, аналогичные или существенно отличающиеся по длине от указанных в настоящем описании, можно сравнивать путем вставки в выравнивание промежутков, причем такие промежутки определяют, например, с помощью алгоритма сравнения, используемого GAP.
Таблица A: Примеры консервативных аминокислотных замен
[0068] Оптимальное выравнивание последовательностей для выравнивания окна сравнения может быть проведено с помощью компьютерных реализаций алгоритмов (GAP, BESTFIT, FASTA и TFASTA в Wisconsin Genetics Software Package Release 70, Genetics Computer Group, 575 Science Drive Madison, WI, США) или путем рассмотрения и наилучшего выравнивания (т.е. приводящего к наивысшей процентной гомологии в окне сравнения), генерируемого с помощью любого из различных выбранных способов. Можно также упомянуть семейство программ BLAST, раскрытое, например, в документе Altschul et al., 1997, Nucl. Acids Res.25:3389. Подробное обсуждение анализа последовательности можно найти в разделе 19.3 документа Ausubel et al., "Current Protocols in Molecular Biology", John Wiley & Sons Inc, 1994-1998, Chapter 15.
[0069] Термины "пациент", "хозяин" или "индивидуум", используемые взаимозаменяемо в настоящем описании, относятся к любому пациенту, конкретно к пациенту-позвоночному, и даже более конкретно к пациенту-млекопитающему, для которого желательна терапия или профилактика. Подходящие позвоночные животные, которые подпадают под объем настоящего изобретения, включают, но без ограничения, всех членов подтипа Chordata, включая приматов (например, людей, мартышек и обезьян), и включают виды обезьян, например из рода Macaca (например, яванских макак, таких как Macaca fascicularis и/или макак-резус (Macaca mulatta)) и павиана (Papio ursinus), а также мармозеток (виды из рода Callithrix), беличьих обезьян (виды из рода Saimiri) и тамаринов (виды из рода Saguinus), а также такие виды обезьян, как шимпанзе (Pan troglodytes)), грызунов (например, мышей, крыс, морских свинок), зайцеобразных (например, кроликов, зайцев), бычьих (например, крупный рогатый скот), баранов (например, овец), козьих (например, коз), свиней (например, домашних свиней), лошадиных (например, лошадей), собачьих (например, собак), кошачьих (например, кошек), птиц (например, кур, индеек, уток, гусей, домашних птиц, таких как канарейки, волнистые попугайчики и т.д.), морских млекопитающих (например, дельфинов, китов), рептилий (змей, лягушек, ящериц и т.д.) и рыб. Предпочтительным пациентом является человек, нуждающийся в лечении или профилактике некоторого состояния. Однако следует понимать, что вышеупомянутые термины не подразумевают наличия симптомов.
[0070] Термин "трансген" используется в настоящем описании для описания генетического материала, который был или будет искусственно введен в геном организма-хозяина, и который передается потомству этого хозяина. В некоторых вариантах осуществления он придает желаемое свойство клетке млекопитающего или ортопоксвирусному вектору, в который он введен, или иным образом приводит к желаемому терапевтическому или диагностическому результату.
[0071] В рамках изобретения, термины "лечение", "лечить" и т.п. относятся к получению желаемого фармакологического и/или физиологического эффекта. Эффект может быть профилактическим в смысле полного или частичного предотвращения заболевания или его симптома и/или может быть терапевтическим в смысле частичного или полного излечения от заболевания и/или неблагоприятного эффекта, связанного с заболеванием. "Лечение", в рамках изобретения, охватывает любое лечение заболевания у млекопитающего, особенно у человека, и включает: (a) предотвращение возникновения заболевания у пациента, который может быть предрасположен к заболеванию, но у которого оно еще не было диагностировано; (b) ингибирование заболевания, т.е. прекращение его развития; и (c) облегчение заболевания, т.е. вызывание регресса заболевания.
[0072] Термины "дикого типа", "природный", "нативный" и т.п. по отношению к организму, полипептиду или последовательности нуклеиновой кислоты означают, что организм, полипептид или последовательность нуклеиновой кислоты встречаются в природе или доступны по меньшей мере в одном встречающемся в природе организме, который не был изменен, подвергнут мутации или иному воздействию человека.
[0073] Варианты включают молекулы нуклеиновой кислоты, в достаточной степени сходные с эталонной молекулой, или формы, комплементарные им полностью или частично, так что может быть достигнута селективная гибридизация в условиях умеренной или высокой жесткости, или те, которые имеют идентичность последовательности приблизительно от 60% до 90% или от 90 до 98% с нуклеотидными последовательностями, определяющими фактор круга хозяев эталонного поксвируса в окне сравнения, содержащем по меньшей мере приблизительно 15 нуклеотидов. Предпочтительно, длина области гибридизации составляет от приблизительно 12 до приблизительно 18 нуклеиновых оснований или больше. Предпочтительно, процентная идентичность между конкретной нуклеотидной последовательностью и эталонной последовательностью составляет по меньшей мере приблизительно 80% или 85%, или, более предпочтительно, сходство составляет приблизительно 90% или больше, например приблизительно 95%, 96%, 97%, 98%, 99% или больше. Включены значения процентной идентичности между 80% и 100%. Длина нуклеотидной последовательности зависит от ее предполагаемой функции. Включены гомологи. Термины "гомолог" "гомологичные гены" или "гомологи" в широком смысле относятся к функционально и структурно родственным молекулам, в том числе из других видов. Гомологи и ортологи являются примерами вариантов.
[0074] Идентичность последовательностей нуклеиновой кислоты может быть определена следующим образом. Рассматриваемая последовательность нуклеиновой кислоты используется для поиска в базе данных последовательностей нуклеиновой кислоты, такой как база данных GenBank (доступная на веб-сайте http://www.ncbi.nln.nih.gov/blast/), с использованием программы BLASTM версии 21 (на основании документа Altschul et al. (1997) Nucleic Acids Research 25:3389-3402). Программа используется в режиме без промежутков. Для удаления гомологий последовательностей из-за областей низкой сложности используется фильтрация по умолчанию. Используются параметры BLASTM по умолчанию.
[0075] Идентичность аминокислотных последовательностей can может быть определена следующим образом. Рассматриваемая полипептидная последовательность используется для поиска в базе данных полипептидных последовательностей, такой как база данных GenBank (доступная на веб-сайте http://www.ncbi.nln.nih.gov/blast/), с использованием программы BLASTP. Программа используется в режиме без промежутков. Для удаления гомологий последовательностей из-за областей низкой сложности используется фильтрация по умолчанию. Используются параметры BLASTP. Фильтрация для последовательностей низкой сложности может использовать программу SEG.
[0076] Предпочтительные последовательности будут гибридизоваться в жестких условиях с эталонной последовательностью или ее комплементом. Термин "гибридизоваться в жестких условиях" и его грамматические эквиваленты относятся к способности молекулы нуклеиновой кислоты гибридизоваться с целевой молекулой нуклеиновой кислоты (такой как целевая молекула нуклеиновой кислоты, иммобилизованная на ДНК или РНК-блоте, таком как Саузерн-блот или нозерн-блот) при определенных условиях температуры и концентрации соли. Что касается молекул нуклеиновой кислоты, длина которых превышает приблизительно 100 оснований, типичными жесткими условиями гибридизации является температура не более чем на 25°C-30°C (например, 10°C) ниже температуры плавления (Tm) нативного дуплекса (смотри в общем Sambrook et al., (выше); Ausubel et al., (1999)). Tm для молекул нуклеиновой кислоты длиннее приблизительно 100 оснований может быть рассчитана по формуле Tm=81,5+0,41% (G+C-log(Na+)). Что касается молекул нуклеиновой кислоты, имеющих длину менее 100 оснований, типичными жесткими условиями гибридизации является температура на 5°C-10°C ниже Tm.
[0077] Термин "делеция" в настоящем контексте означает удаление всей или части кодирующей области целевого гена. Термин также охватывает все формы мутаций или трансформаций, которые уничтожают экспрессию целевого гена или уничтожают или по существу понижающе регулируют уровень или активность кодируемого белка.
[0078] Ссылка на "ген" включает ДНК, соответствующую экзонам или открытой рамке считывания гена. В настоящем описании также считается, что ссылка на "ген" включает: классический геномный ген, состоящий из последовательностей регуляции транскрипции и/или трансляции, и/или кодирующей области, и/или нетранслируемых последовательностей (т.е. интронов, 5'- и 3'- нетранслируемых последовательностей); или мРНК или кДНК, соответствующие кодирующим областям (т.е. экзонам) и 5'- и 3'- нетранслируемым последовательностям гена.
[0079] "Регуляторный элемент" или "регуляторная последовательность" означают последовательности нуклеиновой кислоты (например, ДНК), необходимые для экспрессии функционально связанной кодирующей последовательности в конкретной клетке-хозяине. Регуляторные последовательности, которые подходят для прокариотических клеток, например, включают в себя промотор и, необязательно, цис-действующую последовательность, такую как операторная последовательность и сайт связывания рибосомы. Контрольные последовательности, которые подходят для эукариотических клеток, включают в себя промоторы, сигналы полиаденилирования, энхансеры транскрипции, энхансеры трансляции, лидерные или хвостовые последовательности, которые модулируют стабильность мРНК, а также направляющие последовательности, которые направляют продукт, кодируемый транскрибируемым полинуклеотидом, во внутриклеточный компартмент внутри клетки или во внеклеточную среду.
[0080] Химерные конструкции, подходящие для создания модифицированных клеток млекопитающих настоящего изобретения, содержат последовательность нуклеиновой кислоты, кодирующую фактор круга хозяев ортопоксвируса, который функционально связан с регуляторной последовательностью. Регуляторная последовательность соответствующим образом содержит контрольные последовательности транскрипции и/или трансляции, которые совместимы с экспрессией в клетке. Как правило, регуляторные контрольные последовательности транскрипции и/или трансляции включают в себя, но без ограничения, последовательность промотора, 5'-некодирующую область, цис-регуляторную область, такую как функциональный сайт связывания для транскрипционного регуляторного белка или трансляционного регуляторного белка, вышележащую открытую рамку считывания, связывающие рибосомы последовательности, сайт начала транскрипции, сайт начала трансляции и/или нуклеотидную последовательность, которая кодирует лидерную последовательность, терминирующий кодон, сайт окончания трансляции и 3'-нетранслируемую область. Рассматриваются конститутивные или индуцируемые промоторы, известные в данной области техники. Промоторы могут представлять собой или встречающиеся в природе промоторы, или гибридные промоторы, которые объединяют элементы более чем одного промотора.
[0081] Рассматриваемые последовательности промоторов могут быть нативными для клеток млекопитающих или могут быть получены из альтернативного источника, если данная область является функциональной в выбранном организме. Выбор промотора будет отличаться в зависимости от предполагаемой клетки-хозяина. Например, промоторы, которые можно использовать для экспрессии в клетках млекопитающих, включают промотор металлотионеинов, который можно индуцировать в ответ на тяжелые металлы, такие как кадмий, промотор β-актина, а также вирусные промоторы, такие как промотор большого Т-антигена SV40, промотор немедленно-ранних (IE) генов цитомегаловируса человека (CMV), LTR-промотор вируса саркомы Рауса, LTR-промотор вируса опухоли молочной железы мышей, главный поздний промотор аденовируса (Ad MLP), промотор вируса простого герпеса и промотор HPV, в частности верхней регуляторной области (URR) HPV, и другие. Все эти промоторы хорошо описаны и легко доступны в данной области техники.
[0082] В настоящем описании для повышения уровней экспрессии конструкций млекопитающих могут быть также использованы энхансерные элементы. Примеры включают в себя энхансер раннего гена SV40, как описано, например, в документе Dijkema et al. (1985) EMBO J. 4:761, энхансер/промотор из длинного концевого повтора (LTR) вируса саркомы Рауса, как описано, например, в документе Gorman et al., (1982) Proc. Natl. Acad. Sci. USA 79:6777, и элементы из CMV человека, как описано, например, в документе Boshart et al. (1985) Cell 41:521, такие как элементы, включенные в последовательность интрона A CMV.
[0083] Химерная конструкция может также содержать 3'-нетранслируемую последовательность. 3'-нетранслируемая последовательность относится к части гена, содержащей сегмент ДНК, который содержит сигнал полиаденилирования и любые другие регуляторные сигналы, способные вызывать процессирование мРНК или экспрессию гена. Сигнал полиаденилирования характеризуется тем, что добавляет тракты полиадениловой кислоты к 3'-концу мРНК-предшественника. Сигналы полиаденилирования обычно распознаются по наличию гомологии с канонической формой 5'-AATAAA-3', хотя не редки варианты. 3'-нетранслируемая регуляторная последовательность ДНК, предпочтительно, включает приблизительно от 50 до 1000 нуклеотидов и может содержать последовательности терминации транскрипции и трансляции в дополнение к сигналу полиаденилирования и любым другим регуляторным сигналам, способным вызывать процессирование мРНК или экспрессию гена.
[0084] В некоторых вариантах осуществления химерная конструкция дополнительно содержит селективный маркерный ген, обеспечивающий отбор клеток, содержащих данную конструкцию. Селективные гены хорошо известны в данной области техники и совместимы с экспрессией в представляющей интерес клетке.
[0085] В одном варианте осуществления экспрессия структурного гена или гена сборки ортопоксвируса находится под контролем промотора. В одном неограничивающем варианте осуществления изобретения промотор представляет собой клеточный конститутивный промотор, такой как человеческий EF1-альфа (промотор гена фактора элонгации-1-альфа человека), DHFR (промотор гена дигидрофолатредуктазы) или PGK (промотор гена фосфоглицераткиназы), который направляет экспрессию достаточного уровня CP77 для поддержания размножения вируса в отсутствие значительного токсического воздействия на клетку-хозяина. Промоторы также могут быть индуцируемыми, такими как индуцируемый клеточный промотор, вирусные промоторы MTH (из гена металлотионеина) также используются в клетках млекопитающих, таких как CMV, RSV, SV-40 и MoU3.
[0086] Настоящее изобретение относится к композиции, содержащей аттенуированный поксвирус, который может быть использован в качестве вакцины для индуцирования защиты от инфицирования вирусами чикунгуньи и натуральной оспы. Термины "аттенуация", "аттенуированный" и т.п., в рамках изобретения, означают снижение вирулентности вирусного вектора. Вирулентность обычно определяется как способность вируса вызывать заболевание у конкретного хозяина. Например, поксвирус, который не способен продуцировать инфекционные вирусы, может первоначально инфицировать клетки, но по существу не способен полностью реплицироваться или размножаться в хозяине или клетке-хозяине или вызывать заболевание или состояние. Это желательно, поскольку поксвирусный вектор может доставлять нуклеиновую кислоту хозяину или клетке-хозяину, но обычно не наносит вреда хозяину или клетке-хозяину.
[0087] Семейство поксвирусов включает два подсемейства, Chordopoxvirinae и Entomopoxvirinae. Chordopoxvirinae включает восемь родов, включая Orthopoxviridae, к которым относятся виды, которые инфицируют человека, тогда как Entomopoxvirinae инфицируют насекомых. Orthopoxviridae включает, например, вирус вариолы, который является возбудителем натуральной оспы, вирус коровьей оспы, из которого сформирована оригинальная противооспенная вакцина, о которой сообщил Дженнер в 1796 г., и вирус осповакцины, который используют в качестве противооспенной вакцины второго поколения. К вирусам Avipoxviridae относятся виды, которые инфицируют птиц, такие как вирусы оспы птиц и оспы канареек. Помимо их использования в качестве антигенов в противооспенных вакцинах, очень интересно использование рекомбинантных вирусов на основе вируса осповакцины и авипоксвирусов в качестве векторов для доставки и/или экспрессии представляющих интерес гетерологичных генов. Как внутрицитоплазматические векторы Orthopoxviridae могут доставлять чужеродные антигены в цитоплазму хозяина и пути процессирования антигена, в которых происходит процессирование антигенов в пептиды для презентации на клеточной поверхности. Такие векторы, экспрессирующие чужеродные антигены, подходят для применения в генной терапии и разработке вакцин для широкого диапазона состояний и заболеваний.
[0088] Поксвирусы образуют большое семейство вирусов, отличающихся большим линейным дцДНК-геномом, цитоплазматическим местом размножения и сложной морфологией вириона. Вирус осповакцины является типичным вирусом этой группы вирусов и одним из наиболее изученных в отношении вирусного морфогенеза. Вирионы вируса осповакцины выглядят как имеющие форму "бруска" или "яйца" мембрано-связанные частицы со сложной внутренней структурой с имеющим оболочку двояковогнутым кором, окаймленным "боковыми телами". Путь сборки вирионов включает производство мембраны, содержащей полумесяцы, которые развиваются в незрелые вирионы (IV), а затем эволюционируют в зрелые вирионы (MV). В вирионе вируса осповакцины содержится более 70 специфических генных продуктов, причем в настоящее время описано влияние мутаций на сборку вируса осповакцины в более чем 50 специфических генах.
[0089] Подходящие аттенуированные поксвирусы известны специалистам в данной области техники. Иллюстративные примеры включают аттенуированный модифицированный вирус осповакцины Анкара (MVA), NYVAC, авипоксвирус, вирус оспы канареек и вирус оспы птиц. Таким образом, в варианте осуществления, раскрытом в настоящем описании, аттенуированный поксвирус выбирают из группы, состоящей из модифицированного вируса осповакцины Анкара (MVA), NYVAC, авипоксвируса, вируса оспы канареек и вируса оспы птиц. В варианте осуществления аттенуированный поксвирус представляет собой аттенуированный вирус осповакцины. Иллюстративные примеры штаммов вируса осповакцины включают Copenhagen (COP), Western Reserve (WR), Wyeth, ACAM2000, LC16m8 и Connaught Laboratories (CL).
[0090] Специалистам в данной области техники понятно, что другие штаммы ортопоксвирусов могут быть модифицированы для получения аттенуированного поксвируса. В иллюстративном примере аттенуированный поксвирус может быть получен путем модификации (например, делеции, замены или иного нарушения функции) гена из генома поксвируса, который кодирует эндогенный незаменимый белок сборки или созревания. Таким образом, в варианте осуществления, раскрытом в настоящем описании, аттенуированный поксвирус представляет собой модифицированный ортопоксвирус, причем модификация содержит делецию гена, кодирующего эндогенный незаменимый белок сборки или созревания.
[0091] В варианте осуществления аттенуированный поксвирус представляет собой модифицированный вирус осповакцины, причем модификация содержит делецию гена генома вируса осповакцины, кодирующего эндогенный белок сборки или созревания (или иначе нарушает его функцию), и причем модификация трансформирует вектор на основе вируса осповакцины, который размножается (или который может размножаться) в клетке-хозяине (например, человеческой клетке), в аттенуированный вектор на основе вируса осповакцины, который по существу не реплицируется в клетке-хозяине. В варианте осуществления незаменимый эндогенный ген сборки или созревания выбирают из группы, состоящей из COP-A25L, COP-A3L, COP-A4L, COP-A7L, COP-A8R, COP-A9L, COP-A10L, COP-A11R, COP-A12L, COP-A13L, COP-A14L, COP-A145L, COP-A15L, COP-A16L, COP-A17L, COP-A21L, COP-A22R, COP-A26L, COP-A27L, COP-A28L, COP-A30L, COP-A32L, COP-D2L, COP-D3R, COP-D6R, COP-D8L, COP-D13L, COP-E8R, COP-E10R, COP-E11L, COP-F10L, COP-F17R, COP-G1L, COP-G3L, COP-G4L, COP-G5R, COP-G7L, COP-G7L, COP-H1L, COP-H2R, COP-H3L, COP-H4L, COP-H5R, COP-H6R, COP-I1L, COP-I2L, COP-I6L, COP-I7L, COP-I8R, COP-J1R, COP-J4R, COP-J6R, COP-L1R, COP-L3L, COP-L4R и COP-L5R.
[0092] В предпочтительном варианте осуществления модификация содержит делецию гена D13L, и/или гена K1L, и/или гена A39R, и/или генов B7R-B8R.
[0093] В еще одном предпочтительном варианте осуществления модификация содержит делецию гена D13L, гена A39R и генов B7R-B8R.
[0094] В первом аспекте настоящее изобретение относится к композиции для повышения иммунного ответа у животного, которая снижает риск инфицирования чикунгуньей и натуральной оспой, причем эта композиция содержит фармацевтически приемлемый носитель и аттенуированный поксвирус, причем геном поксвируса содержит последовательность нуклеиновой кислоты, кодирующую субгеномный полипротеин 26S вируса чикунгуньи.
[0095] Во втором аспекте настоящее изобретение относится к композиции для повышения иммунного ответа у животного, которая снижает риск инфицирования вирусом Зика и инфицирования натуральной оспой, причем эта композиция содержит фармацевтически приемлемый носитель и аттенуированный поксвирус, причем геном поксвируса содержит последовательность нуклеиновой кислоты, кодирующую полипротеин PrME вируса Зика.
[0096] В третьем аспекте настоящее изобретение относится к композиции для повышения иммунного ответа у животного, которая снижает риск инфицирования чикунгуньей, вирусом Зика и натуральной оспой, причем эта композиция содержит фармацевтически приемлемый носитель и аттенуированный поксвирус, причем геном поксвируса содержит последовательность нуклеиновой кислоты, кодирующую субгеномный полипротеин 26S вируса чикунгуньи, и последовательность нуклеиновой кислоты, кодирующую полипротеин PrME вируса Зика.
[0097] В четвертом аспекте настоящее изобретение относится к способу индуцирования у пациента защитного иммунного ответа на инфицирование чикунгуньей и натуральной оспой, натуральной оспой и вирусом Зика и/или инфицирование чикунгуньей, натуральной оспой и вирусом Зика, включающему введение пациенту композиции в соответствии с первым, вторым или третьим аспектами настоящего изобретения.
[0098] В пятом аспекте настоящее изобретение относится к применению композиции в соответствии с первым, вторым или третьим аспектами настоящего изобретения в получении лекарственного средства для индуцирования у пациента защитного иммунного ответа на инфицирование чикунгуньей и натуральной оспой, натуральной оспой и вирусом Зика и/или инфицирование чикунгуньей, натуральной оспой и вирусом Зика.
[0099] В предпочтительных формах настоящего изобретения аттенуированный поксвирус выбирают из группы, состоящей из вируса осповакцины, вируса коровьей оспы, модифицированного вируса осповакцины Анкара (MVA), NYVAC, авипоксвируса, вируса оспы канареек и вируса оспы птиц. Предпочтительно, чтобы аттенуированный поксвирус представлял собой модифицированный ортопоксвирус, причем модификация содержит делецию гена, кодирующего эндогенный незаменимый белок сборки или созревания. Также предпочтительно, чтобы модификация содержала делецию гена D13L и, предпочтительно, дополнительно содержала делецию гена K1L.
[0100] В некоторых вариантах осуществления фармацевтически приемлемый носитель содержит адъювант. Предпочтительно, чтобы адъювант был выбран из группы, состоящей из гидроксида алюминия, фосфата алюминия, сульфата алюминия-калия, гидроксида-фосфата кальция, полного адъюванта Фрейнда, Montanide®, неполного адъюванта Фрейнда, искомов, матрицы иском, адъюванта ISCOMATRIX™, адъюванта Matrix M™, адъюванта Matrix C™, адъюванта Matrix Q™, адъюванта AbISCO®-100, адъюванта AbISCO®-300, ISCOPREP™, производного ISCOPREP™, адъюванта, содержащего ISCOPREP™ или производное ISCOPREP™, QS-21, производного QS-21 и адъюванта, содержащего QS-21 или производное QS21.
[0101] Различные варианты осуществления, возможные в настоящем описании, далее описаны с помощью следующих неограничивающих примеров.
ПРИМЕР 1
Краткое изложение стратегий конструирования
[0102] Три вируса SCV были сконструированы до конструирования SCV1002, одновекторной вакцины от CHIK/ZIKA. SCV301C конструировали путем замены ORF A39R штамма Copenhagen вируса осповакцины на кассету экспрессии CHIKV-26S вместе с кассетами экспрессии EGFP и Ecogpt. SCV302 создавали путем удаления кассет EGFP и Ecogpt из SCV301C. Затем SCV305, вакцину от SCV-CHIK, создавали путем удаления ORF B7R-B8R и ORF D13L из SCV302. Наконец, SCV1002, вакцину от SCV-CHIK/ZIKA, создавали путем замены ORF B7R-B8R SCV302 на кассету экспрессии ZIKV-prME и удаления ORF D13L.
Конструирование SCV301C (VACV-CHIK)
Описание
[0103] Кассета экспрессии вакцинного антигена чикунгуньи в SCV301C (CHIKV-26S) состоит из следующих элементов в линейном расположении:
- ранний/поздний промотор вируса осповакцины,
- кодирующая белок CHIKV последовательность для субгеномного полипротеина 26S вирулентного реюньонского штамма 06_21 поксвируса
- и последовательность ранней остановки транскрипции поксвируса.
[0104] Субгеномный полипротеин 26S CHIKV при экспрессии процессируется в отдельные структурные гены, необходимые для формирования вирусных частиц. Поскольку весь геном CHIKV отсутствует, и субгеномная РНК не будет содержать 5'-НТО и 3'-НТО, транскрибируемая вирусная РНК будет упакована во вновь образованные вирусные частицы - по сути, экспрессия только субгеномного полипротеина 26S приведет к вирусоподобным частицам (VLP), лишенным вирусной геномной РНК. Экспрессия in vivo субгеномного полипротеина, приводящего к образованию VLP после вакцинации, будет благоприятной для нейтрализации стимуляции антител - ключевой коррелят иммунитета для профилактической вакцинации от CHIKV.
[0105] Аминокислотную последовательность субгеномного полипротеина 26S в кассете экспрессии CHIKV-26S брали из вирулентного реюньонского штамма, который был генетически изменен для расширения его круга хозяев для нового комариного вектора (Genbank, номер доступа: AM258992). Во время эпидемии в Реюньоне между 2005 и 2006 годами вирус мутировал с расширением круга хозяев его вектора до азиатского тигрового комара (Aedes albopictus). Азиатский тигровый комар является самым быстро распространяющимся комаром в мире. Чрезвычайно хорошо приспособлен для жизни среди людей и известен путешествиями в использованных шинах. Его находят в сельских и зеленых городских районах почти во всем мире, и он агрессивно кусает днем людей, домашних и диких животных и птиц. Этот комар обладает потенциалом распространения чикунгуньи по всему миру. Итоговую конструкцию нуклеотидной последовательности кодирующей последовательности белка CHIKV-26S подвергали скринингу в отношении раннего транскрипционного мотива поксвируса "TTTTTNT" - ничего не было найдено. Добавили последовательность ранней остановки транскрипции поксвируса TTTTTAT непосредственно после стоп-кодона. Последовательность кассеты экспрессии CHIKV-26S, описанная выше, приведена в SEQ ID NO:1.
[0106] Для создания SCV301C в GeneArt GmbH синтезировали кассету гомологичной рекомбинации, которая состояла из кассеты экспрессии CHIKV-26S, кассеты экспрессии усиленного зеленого флуоресцентного белка, кассеты экспрессии Ecogpt, причем все они фланкированы левым и правым плечами гомологичной рекомбинации, которые нацелены на вышележащую и нижележащую последовательности ORF A39R VACV-COP для гомологичной рекомбинации, как показано на фигурах 1 и 2 ниже. В таблице 1 приведены элементы кассеты гомологичной рекомбинации SCV301A, а последовательность подробно приведена в SEQ ID NO:2.
Таблица 1: Таблица элементов кассеты гомологичной рекомбинации SCV301C
[0107] SCV301A конструировали путем вставки кассеты гомологичной рекомбинации CHIKV-26S в штамм Copenhagen вируса осповакцины (VACV-COP) путем замены ORF A39R с помощью гомологичной рекомбинации между F1 и F2 и их гомологичными последовательностями в геноме VACV-COP, как показано на фигуре 2 выше, что приводит к конфигурации вставки, показанной на фигуре 3.
Методология
[0108] SCV301C (VACV-CHIKV) конструировали путем гомологичной рекомбинации для замены ORF A39R в VACV на кассету гомологичной рекомбинации CHIKV-26S, которая была синтезирована в GeneArt GmbH, содержащей поксвирусную кассету экспрессии, экспрессирующую структурный полипротеин CHIKV 26S (Genbank, номер доступа: AM258992) под контролем промоторов VACV (Chakrabarti etal, 1997) вместе с поксвирусной кассетой экспрессии для экспрессии гуанинфосфорибозилтрансферазы E. coli (Ecogpt). Гомологичную рекомбинация и положительную селекцию рекомбинантных вирусов, содержащих одновременную вставку кассеты экспрессии Ecogpt, проводили, как описано в протоколе 6 в документе Smith (1993) и опубликовано в документах Falkner et al (1988) и Boyle et al (1988), причем плечи гомологичной рекомбинации, фланкирующие кассеты экспрессии CHIKV-26S и Ecogpt, были сконструированы так, чтобы они были гомологичны с последовательностями, фланкирующими ORF A39R. Кратко говоря, гомологичную рекомбинацию проводили в клетках BHK21, которые были инфицированы VACV-COP при MOI 0,01 БОЕ на клетку в течение 1 часа с последующей трансфекцией кассетой гомологичной рекомбинации CHIKV-26S. Затем инфицированные/трансфицированные клетки инкубировали в течение 2-3 дней до тех пор, пока не наблюдали заметные цитопатические эффекты, за чем следовали сбор и клеточный лизис для получения вирусного экстракта. Осуществляли положительную селекцию SCV301C путем селекции Ecogpt по препарату с использованием обработки клеток BHK21 с помощью MXHAT, как описано в протоколе 6 документа Smith (1993), за исключением того, что были проведены 20 циклов очистки бляшек путем ограниченного разбавления в 48WP для устранения следов родительского VACV-COP по оценке с помощью специфического к A39R ПЦР-анализа. Затем клон-кандидат амплифицировали в клетках BHK21 без обработки MXHAT для получения вирусного посевного материала, из которого получали партии SCV301C для исследований вакцинации. Последовательность генома SCV301C приведена в SEQ ID NO:3.
Ссылки:
Chakrabarti, S, Sisler, JR, and Moss, B (1997). Compact, synthetic, vaccinia virus early/late promoter for protein expression. Biotechniques 23: 1094-1097.
Smith, GL (1993)In: Davison, AJ and Elliotand, RM (eds). Expression of genes by vaccinia virus vectors in "Molecular Virology a Practical Approach". IRL Press at Oxford University Press.
Falkner, FG, and Moss, B (1988). Escherichia coli gpt gene provides dominant selection for vaccinia virus open reading frame expression vectors. J Virol 62: 1849-1854.
Boyle, DB, and Coupar, BE (1988). A dominant selectable marker for the construction of recombinant poxviruses. Gene 65: 123-128.
Конструирование SCV305 (SCV-CHIK)
Удаление репортерной кассеты EGFP/Ecogpt для создания SCV302
Описание
[0109] Для конструирования SCV305 кассеты экспрессии EGFP и Ecogpt, которые должны быть удалены из SCV301C для создания SCV302, рекомбинантного штамма Copenhagen вируса осповакцины, который содержит только кассету экспрессии полипротеина чикунгуньи 26S, вставленную в ORF A39R. Этот вирус конструировали путем удаления репортерных кассет экспрессии Ecogpt/EGFP из SCV301C путем гомологичной рекомбинации с pTC29 и противоселекции вирусов с функциональной экспрессией Ecogpt, как показано ниже на фигуре 4.
Описание pTC29
[0110] pTC29 представляет собой клон плазмиды, который содержит плечи гомологичной рекомбинации, A39R-F1 и SCV301C-F2, требующиеся для удаления кассет экспрессии EGFP и Ecogpt из SCV301C путем гомологичной рекомбинации, как показано на фигуре 4. Последовательность pTC29 подробно приведена в SEQ ID NO:4, а карта плазмиды показана на фигуре 5.
Методология
[0111] Кассеты экспрессии Ecogpt и EGFP удаляли из SCV301C путем гомологичной рекомбинации и противоселекции экспрессии Ecogpt, как описано в протоколе 7 документа Smith (1993) и опубликовано в документе Isaacs et al (1990), причем плечи гомологичной рекомбинации были сконструированы так, чтобы они были гомологичны последовательностям, фланкирующим кассеты экспрессии Ecogpt и EGFP в рекомбинантном геноме VACV-CHIK. После гомологичной рекомбинации SCV302 дополнительно очищали от бляшек в клеточной линии hrpt- в присутствии 6-тиогуанина (6-TG) для противоселекции примесного вируса, экспрессирующего Ecogpt, т.е. исходного SCV301C.
Ссылки
Smith, G.L. (1993). Expression of genes by vaccinia virus vectors. In Molecular Virology a Practical Approach, A.J. Davison and R.M. Elliotand, eds. (IRL Press at Oxford University Press), pp. 257-283.
Isaacs, S.N., Kotwal, G.J., and Moss, B. (1990). Reverse guanine phosphoribosyltransferase selection of recombinant vaccinia viruses. Virology 178, 626-630.
Удаление B7R-B8R и D13L для создания SCV305
Описание
[0112] SCV305 конструировали путем одновременного удаления ORF D13L и B7R-B8R из SCV302 в одной реакции гомологичной рекомбинации с D13L-делеционной кассетой гомологичной рекомбинации (HR) (делеция D13L с трансдоминантной селекцией CP77/DsRed) и B7R-B8R-делеционной кассетой гомологичной рекомбинации (HR) (делеция B7R-B8R с трансдоминантной селекцией Cyan/зеоцин). Детали конфигурации каждой кассеты показаны на фигурах 6 и 7, а последовательность подробно приведена в SEQ ID NO:5 и SEQ ID NO:6.
Таблица 2: Таблица элементов D13L-делеционной HR-кассеты
Таблица 3: Таблица элементов B7R-B8R-делеционной HR-кассеты
[0113] Для селекции вируса с делецией D13L использовали клеточные линии, экспрессирующие белок D13: CHO+D13 (клеточная линия CHO, экспрессирующая белок D13) и CHO+D13+CP77 (клеточная линия CHO, экспрессирующий белок D13 и белок CP77). Причиной использования двух таких клеточных линий является то, что экспрессия CP77 вирусом обеспечивает размножение в клетках CHO, и поэтому ее использовали в качестве инструмента положительной селекции. Замена ORF D13L на кассету экспрессии CP77/DsRed обеспечивает амплификацию вируса с делецией D13L в клетках CHO-D13, что может быть подтверждено красной флуоресценцией инфицированных клеток. Для удаления репортерной кассеты CP77/DsRed после подтверждения делеции D13L была разработана внутримолекулярная рекомбинация для удаления этой кассеты, когда больше не требовалась вирусная экспрессия CP77. Это было сделано путем амплификации вируса с делецией D13L в CHO+D13+CP77, которые не только экспрессируют белок D13, но также белок CP77, что делает вирусно экспрессируемый CP77 избыточным, и позволяет его удаление путем внутримолекулярной рекомбинации, как показано на фигуре 4.
[0114] Для селекции вируса с делецией B7R-B8R использовали репортерную кассету резистентности к зеоцину и голубой флуоресценции для замены этих ORF путем гомологичной рекомбинации. Осуществляли положительную селекцию кассеты зеоцин/Cyan путем культивирования клеток в зеоцине во время инфицирования и селекции клеток с голубой флуоресценцией. После подтверждения делеции B7R-B8R инициировали удаление этого репортера путем внутримолекулярной рекомбинации в отсутствие зеоцина во время инфицирования, как показано на фигуре 8.
[0115] Экспериментальная стратегия одновременного удаления D13L и B7R-B8R была реализована путем (i) гомологичной рекомбинации SCV302 с D13L-делеционной HR-кассетой и B7R-B8R HR-кассетой в клетках CHO+D13, (ii) амплификации SCV305 в клетках CHO+D13, обработанных зеоцином, (iii) выделения индивидуально инфицированных клеток, которые одновременно давали голубую и красную флуоресценцию, до выделения чистого двойного нокаут-клона и затем (iv) инициирования удаления обеих репортерных кассет путем инфицирования клеток CHO+D13+CP77 без обработки зеоцином и выделения нефлуоресцентных (голубым и красным) инфицированных клеток.
[0116] Клоны, идентифицированые как свободные от примесей вирусных промежуточных продуктов с удаленными целевыми ORF, подтверждали с помощью ПЦР и секвенирования. Наличие кассеты экспрессии CHIKV-26S в ORF A39R дополнительно подтверждали с помощью ПЦР и секвенирования, а экспрессию белков CHIKV подтверждали с помощью вестерн-блота. Общую аттенуацию SCV305 верифицировали с помощью исследования инфекционности в пермиссивных к вирусу осповакцины клеточных линиях.
Методология
[0117] Как D13-делеционная HR, так и B7R-B8R-делеционная HR-кассеты были синтезированы GeneArt GmbH. Для создания SCV305 (SCV-CHIK) ORF D13L и ORF B7R-B8R удаляли из SCV302 путем гомологичной рекомбинации, при которой проводили положительную селекцию успешных делеций путем инфицирования клеток CHO, экспрессирующих только белок D13 (CHO-D13), в присутствии зеоцина. Поскольку CHO являются непермиссивными к инфицированию VACV, положительной селекции вируса с делецией D13L достигали путем замены ORF D13L на кассету экспрессии, кодирующую белок круга хозяев вируса коровьей оспы CHO CP77. Конструировали плазмиду гомологичной рекомбинации, которая состояла из кассеты экспрессии CP77 и кассеты экспрессии флуоресцентного белка dsRed, фланкированной левым и правым плечами рекомбинации, которые гомологичны последовательностям, фланкирующим ORF D13L в SCV302. Кассету экспрессии dsRed добавляли потому, что VACV, экспрессирующий CP77, не образует литических бляшек в CHO; поэтому инфицирование контролировали по наличию красной флуоресценции. Для помощи в удалении кассет экспрессии CP77 и dsRed, которые заменили ORF D13L, и кассеты экспрессии AmCyanZeo, которая заменила ORF B7R-B8R, последовательность повтора левого плеча гомологичной рекомбинации помещали ниже кассет экспрессии CP77 и dsRed и ниже кассеты экспрессии AmCyanZeo, но выше правого плеча гомологичной рекомбинации в обеих кассетах гомологичной рекомбинации. Гомологичную рекомбинацию проводили путем трансфекции удаляющих D13L и B7R-B8R кассет гомологичной рекомбинации в CHO-D13, ранее инфицированные VACV-CHIK при MOI 0,01 БОЕ на клетку. SCV305 обогащали после инфицирования гомологичной рекомбинации путем амплификации вируса в CHO-D13, обработанных зеоцином, с последующим инфицированием свежего набора клеток CHO-D13, обработанных зеоцином, амплифицированным вирусом. Эти инфицированные клетки извлекали и превращали в суспензию отдельных клеток с помощью расщепления TrypLE Select (Thermo Fisher Scientific), а затем поклеточно сортировали, так что одну клетку с красной и голубой флуоресценцией высевали в 1 лунку 96-луночного планшета, содержащего культивированные клетки CHO-D13, обработанные зеоцином, с использованием проточного цитометра FACSAria Fusion (BD Biosciences). Затем 96-луночный планшет, засеянный клетками с красной/голубой флуоресценцией, инкубировали при 37°C/5% CO2, пока не удавалось увидеть красные и голубые флуоресцентные очаги инфекции в какой-либо из лунок 96-луночного планшета. Лунки, содержащие один очаг инфицирования, собирали и ресуспендировали в виде суспензий отдельных клеток до поклеточной сортировки и высевания на 96-луночные планшеты со свежими клетками CHO-D13, обработанными зеоцином. Этот процесс поклеточной сортировки повторяли пять раз для удаления следовых примесей исходного SCV302 и получения клонального SCV305. Ряд клонально очищенных кандидатов SCV305 амплифицировали в клетках CHO-D13, обработанных зеоцином, и затем тестировали с помощью ПЦР-анализа на присутствие примесного SCV302 и для подтверждения замены D13L кассетами экспрессии CP77 и dsRed и замены B7R-B8R на кассету экспрессии AmCyanZeo. Затем наилучший клон амплифицировали в линии CHO+D13+CP77 (экспрессирующей как белок CP77, так и белок D13) для стимуляции внутримолекулярной рекомбинации между левыми последовательностями гомологичной рекомбинации и последовательностями повтора, поскольку зависимость от вирусной экспрессии CP77 больше не нужна, и необходимость в экспрессии белка резистентности к зеоцину больше не нужна в отсутствие зеоцина, что приводит к удалению из SCV305 кассет экспрессии CP77 и dsRed и кассеты экспрессии AmCyanZeo. Инфицированные клетки CHO+D13+CP77 вводили в суспензию отдельных клеток путем расщепления с помощью TrypLE Select, клетки сортировали с использованием проточного цитометра FACSAria Fusion, в котором нефлуоресцентные клетки были массово отсортированы, и сохраняли. Вирус SCV305 дополнительно амплифицировали для получения посевного материала путем инфицирования клеток CHO+D13+CP77 с последовательным масштабированием. Вся геномная последовательность SCV305 приведена в SEQ ID NO:7.
Конструирование SCV1002 (SCV-CHIK/ZIKA)
Описание
[0118] SCV1002 представляет собой одновекторную поливалентную вакцину от чикунгуньи и вируса Зика, которая имеет следующие особенности:
- Замена ORF A39R на кассету экспрессии SCV-CHIKV-(субгеномный 26S) (CHIKV-26S)
- Замена ORF B7R-B8R на кассету экспрессии SCV-ZIKV-prME
- Удаление ORF D13
[0119] SCV1002 конструировали путем замены ORF B7R-B8R SCV302 на кассету экспрессии полипротеина prME вируса Зика и удаления ORF D13L. Последовательность кассеты экспрессии prME вируса Зика подробно приведена в SEQ ID NO:8, причем взяли последовательность prME происхождением из бразильского штамма ZikaSPH2015 (Genbank: KU321639). Кассета гомологичной рекомбинации (HR) ZIKV-prME была синтезирована в GeneArt, как показано на фигуре 10, причем последовательность подробно приведена в SEQ ID NO:9.
Таблица 4: Таблица элементов HR-кассеты ZIKV prME
Методология
[0120] SCV1002 (SCV-CHIK/ZIKA) конструировали путем замены ORF B7R-B8R на поксвирусную кассету экспрессии для ZIKV prME (бразильский изолят ZikaSPH2015, Genbank: KU321639) и удаления ORF D13L путем гомологичной рекомбинации. Для вставки кассеты экспрессии ZIKV prME путем замены ORF B7R-B8R была синтезирована в GeneArt GmbH кассета гомологичной рекомбинации, состоящая из следующих элементов: (i) последовательности, нацеленные на гомологичную рекомбинацию F1 выше гена B7R, (ii) кассета экспрессии, состоящая из раннего/позднего промотора вируса осповакцины, функционально связанного с кодирующей белок последовательностью слияния синего флуоресцентного белка с белком резистентности к зеоцину (BFPzeo), и заканчивающаяся последовательностью ранней остановки транскрипции поксвируса, (iii) повтор плеча F1 гомологичной рекомбинации, (iv) кассета экспрессии, состоящая из раннего/позднего промотора вируса осповакцины, функционально связанного с кодирующей белок последовательностью ZIKV prME, за которой следует последовательность ранней остановки транскрипции поксвируса, и, наконец, (v) последовательности, нацеленные на гомологичную рекомбинацию F2 ниже гена B8R. ORF D13L удаляли, как описано в конструировании SCV305 (SCV-CHIK).
[0121] Гомологическую рекомбинацию проводили путем трансфекции как ZIKV prME HR-кассеты, так и HR-кассеты делеции D13L в клетки CHO+D13, ранее инфицированные SCV302 при MOI 0,01 БОЕ на клетку. Полученный SCV1002 обогащали после инфицирования гомологичной рекомбинации путем амплификации вируса в клетках CHO+D13 в присутствии зеоцина (ThermoFisher Scientific) с последующим вторым инфицированием свежего набора клеток CHO+D13 в присутствии зеоцина амплифицированным вирусом. Эти инфицированный клетки извлекали и превращали в суспензию отдельных клеток с помощью расщепления TrypLE Select, а затем поклеточно сортировали, так что одну клетку с синей и красной флуоресценцией высевали в одну лунку 96-луночного планшета, содержащего клетки CHO+D13, с использованием проточного цитометра FACSAria Fusion (BD Biosciences). После инкубации в присутствии зеоцина лунки, содержащие один синий и красный флуоресцентный очаг инфицирования, собирали и ресуспендировали в виде суспензий отдельных клеток до поклеточной сортировки и высевания на 96-луночные планшеты со свежими клетками CHO+D13. Эту поклеточную сортировку и культивирование в присутствии зеоцина повторяли пять раз для удаления следовых примесей исходного SCV302 и получения клонального SCV1002. Несколько клонально очищенных SCV1002 амплифицировали в клетках CHO+D13+CP77 в отсутствие зеоцина, а затем подвергали ПЦР-анализу для подтверждения вставки ZIKV prME в локус B7R-B8R, сохранения кассеты экспрессии CHIK в локусе A39R и отсутствия примесного SCV302. Клоны амплифицировали в клетках CHO+D13+CP77 в отсутствие зеоцина для стимуляции внутримолекулярной рекомбинации между последовательностью гомологичной рекомбинации F1 и последовательностью повтора F1, что приводит к удалению обеих кассет экспрессии BFPzeo и DsRed/CP77. Суспензии отдельных клеток этих культур массово сортировали (FACSAria), и сохраняли нефлуоресцентные клетки. ПЦР повторяли для подтверждения сохранения вставок и потери BFPzeo, DsRed/CP77 и удаления ORF D13L. Получали запасы вакцины SCV1002 в клетках CHO-D13+CP77, и титровали их. Последовательность всего генома SCV1002 приведена в SEQ ID NO:10.
Пример 2
Вакцинации SCV305 защищают от ECTV
[0122] Группы самок мышей C57BL/6 в возрасте 6-8 недель (n=5 мышей на группу) вакцинировали и/п SCV305 (105, 106, 107 БОЕ) или мнимо вакцинировали ФСБ. Уровни специфического IgG против вируса осповакцины определяли с помощью ELISA конечной точки. Было установлено, что вакцинация SC305 обеспечивает существенные титры специфического вирусного антитела против вируса осповакцины, которые были аналогичны получаемым при вакцинации аналогичной дозой вируса осповакцины (VACV).
[0123] Через четыре недели мышей заражали летальной дозой (50 LD50) вируса мышиной оспы, Ectromalia (ECTV). ECTV очень инфекционен для мышей и вызывает те же симптомы у мышей, что и натуральная оспа у людей.
[0124] Мыши, вакцинированные 107, 106, и 105 БОЕ SCV305 или 105 БОЕ VACV, демонстрировали аналогичные уровни защиты от потери веса, клинических симптомов и смертности. Распространение ECTV на несколько органов также было предотвращено у всех вакцинированных мышей. Эти результаты показывают, что однократная вакцинация SCV305 обеспечивает полную защиту от заражения смертельным поксвирусом, что позволяет предположить, что SCV сохраняет свою полезность в качестве противооспенной вакцины.
Однократная вакцинация SCV305 дает устойчивые и длительные ответы антител против вируса чикунгуньи
[0125] Антитела считаются существенно важными для защиты от вируса от чикунгуньи (CHIKV), причем человеческие и мышиные антитела направлены на поверхностный гликопротеин E2 CHIKV, который, как было показано, опосредует защиту. Мышам вводили однократную дозу вакцины (105, 106 и 107 БОЕ SCV-CHIK), и исследовали ответы специфических к E2 CHIKV IgG с помощью ELISA. Наблюдали зависящее от дозы и времени повышение уровней CHIKV-E2-специфических IgG, причем мыши, которые получили 107 БОЕ, демонстрировали кинетику и величину выработки антител, сходную с индуцированной у мышей, вакцинированных репликационно-компетентным VACV-CHIK. Это показывает, что дефектная по размножению вакцина SCV305 способна генерировать выработку антител на рекомбинантный иммуноген, сравнимую с компетентной по размножению вакциной VACV-CHIK.
[0126] Выработку нейтрализующих антител детектировали у 4/6 мышей, получавших 105 БОЕ вакцины SCV305, тогда как все 6 мышей, получавших 106 БОЕ и 107 БОЕ вакцины SCV305, вырабатывали нейтрализующие антитела.
[0127] Исследовали уровни CHIKV-специфических IgG1 и IgG2c в сыворотке мышей на 30 день после вакцинации для получения показателя "баланса Th1/Th2" выработки антител. При 107 БОЕ SCV305 стимулировала выработку IgG2c, сравнимую с индуцируемой VACV-CHIK. Однако SCV305 индуцировала более высокую выработку IgG1, чем VACV-CHIK, что говорит о генерации SCV-CHIK более сбалансированного по Th1 (IgG2c)/Th2 (IgG1) ответа, со смещением у VACV-CHIK с сторону Th1.
[0128] Для анализа долговечности выработки антител, индуцируемой вакциной от SCV-CHIK, исследовали выработку анти-CHIKV антител через 4 недели, 6 месяцев и 1 год после вакцинации. За это время наблюдали ограниченное падение уровней анти-CHIKV антител, причем уровни антител оставались сравнимыми у мышей, вакцинированных SCV305 и VACV-CHIK. Кроме того, детектировали секретирующие CHIKV E2-специфические антитела клетки (ASC) и продуцирующие CHIKV E2/E1 и капсид-специфический IFN-γ клетки через 1 год после вакцинации у вакцинированных SCV305 и VACV-CHIK мышей.
[0129] Модель вакцинации и заражения CHIKV была выведена из демонстрации того, что мыши, вакцинированные VACV-CHIK (107 БОЕ) и зараженные CHIKV (изолят острова Реюньон; LR2006-OPY1, 104 CCID50) были полностью защищены от виремии и артралгии. Поскольку вакцина от SCV-CHIK индуцировала выработку нейтрализующих антител, сравнимую с VACV-CHIK в той же дозе 107 БОЕ, защитную эффективность вакцины SCV305 оценивали при более низких дозах (105 и 106 БОЕ). Мыши, вакцинированные SCV305 при 106 БОЕ и зараженные через 40 дней (изолятом острова Реюньон; LR2006-OPY1, 104 CCID50), не продемонстрировали детектируемой виремии или припухлости стопы. Мыши, вакцинированные в 10 раз более низкой дозой (105 БОЕ) были практически защищены с уменьшением виремии приблизительно 4-log и значительно сниженной припухлостью стопы по сравнению с контрольными VACV-инфицированными мышами. Продолжительные исследования подтвердили, что вакцинированные SCV305 мыши защищены от заражения CHIKV через год после вакцинации.
[0130] Персистенцию геномной РНК CHIKV, в частности в тканях суставов, связывают с хроническим артритическим заболеванием после инфицирования CHIKV у людей и в модели на мышах. Через 30 дней после заражения мыши, вакцинированные 106 БОЕ вакцины SCV305, продемонстрировали значительное снижение уровня персистирующей вирусной РНК до фоновых уровней, что показывает, что вакцинация SCV305 может предотвращать сохранение персистирующей РНК CHIKV в тканях суставов.
[0131] В совокупности эти результаты демонстрируют способность вакцина SCV305 вызвать надежную, сбалансированную и продолжительную выработку CHIKV-специфических антител и обеспечить защиту от виремии, острого артрита и персистенции вирусной РНК после заражения CHIKV.
ПРИМЕР 3
Вакцинация SCV1002
[0132] При комбинировании различных живых аттенуированных вирусных вакцин наиболее часто наблюдаемой проблемой является конкуренция между вирусами. Ее можно обойти, увеличив дозу комбинированной вакцины или скорректировав дозировку каждого компонента вакцины, чтобы преодолеть конкуренцию со стороны доминирующих компонентов вакцины.
[0133] Сообщалось об конкуренции антигенов путем "иммунологического взаимовлияния" между компонентами трехвалентной вакцины против дифтерии, коклюша и столбняка, между бактеринами чумы собак и живым вирусом чумы собак и при использовании бордетеллы в качестве разбавителя для живых комбинированных вакцин от вируса чумы, аденовируса типа 2, парвовируса и вируса парагриппа (Hunt et al, 2001). В этих примерах прививка многокомпонентной вакциной вызывает меньше антител, чем когда компоненты вводят отдельно. В некоторых случаях ответ на один антиген доминирует, тогда как ответы на другие подавляются. В других случаях, если происходит взаимная конкуренция, уменьшается ответ на все компоненты.
[0134] Как было показано, степень антигенной конкуренции, зависит от ряда параметров вакцинации, включая относительные места прививки конкурирующих антигенов, интервал времени между введением антигенов и дозу доминирующего антигена по отношению к подавляемому антигену.
[0135] Несмотря на то, что вышеуказанное может быть решено путем экспрессии иммунизирующих антигенов нескольких вызывающих заболевание агентов из одного и того же вектора, что обеспечивает равное представление каждого иммунизирующего антигена иммунной системе, причем с одним вектором для доставки антигенов нужно рассматривать оптимальный способ вакцинации только для этого вектора, существует риск того, что может иметь место взаимовлияние антигенов путем захвата и ГКГ-презентации антигенпрезентирующими клетками, такими как B-клетки и дендритные клетки, преимущественного антигена. Антиген с доминантными В-клеточными и/или Т-клеточными эпитопами будет вызывать более сильный иммунный ответ, чем антиген, имеющий субоптимальные В-клеточные и/или Т-клеточные эпитопы. Следовательно, это может приводить к смещенному иммунному ответу на наиболее доминирующий антиген при вакцинации одним вектором, который экспрессирует множество антигенов от множества вызывающих заболевание агентов. Можно ожидать, что антиген, экспрессируемый из вектора, который также экспрессирует другие антигены от других заболеваний, например антиген чикунгуньи, экспрессируемый вместе с антигенами вируса Зика, может не давать столь же сильный иммунный ответ, что и иммунизирующий вектор, который экспрессирует только антиген чикунгуньи.
[0136] Было проведено исследование для определения, будет ли экспрессия нескольких доминирующих антигенов от нескольких вызывающих заболевание вирусов мешать иммунным ответам на них. Например, белок E2 чикунгуньи очень эффективен для стимуляции выработки очень хороших нейтрализующих антител, которые могут нейтрализовать инфицирование чикунгуньей. Аналогично, белок Е вируса Зика также стимулирует мощную выработку нейтрализующих антител для нейтрализации инфицирования вирусом Зика. Однако экспрессия этих двух доминирующих антигенов из одного и того же вектора может влиять на их способность стимулировать мощный иммунный ответ на их соответствующие вирусы, т.е. один доминирующий антиген может получить доминирование над другим.
[0137] Для определения, не является ли экспрессия нескольких доминирующих антигенов из одного и того же вектора вредной для стимуляции оптимальных иммунных ответов по сравнению с экспрессией каждого доминирующего антигена из отдельного вектора, провели исследование вакцинации на мышах со сравнением SCV1002 (вакцина от CHIK/ZIKA) с SCV305 (вакцина от CHIK) в отношении стимуляции специфических для чикунгуньи нейтрализующих антител.
Стратегия вакцинации:
[0138] Самок мышей C57BL/6 дикого типа и дефицитных по интерфероновому рецептору (IFNAR) вакцинировали однократно одновекторной вакциной от CHIKV/ZIKV (SCV1002), только от CHIKV (SCV305) или пустым контрольным вектором (SCV105) в группах по 6 мышей на лечбную группу. Все лечебные группы получали 106 БОЕ вакцины на мышь посредством интраперитонеальных инъекций, и через 2 и 4 недели после вакцинации у них брали кровь. Всех мышей заражали через 6 недель после вакцинации.
Анализ нейтрализации:
[0139] Уровни нейтрализующих антител часто используются в качестве коррелята защиты. Поэтому во всех группах вакцинации, ZIKV/CHIKV (SCV1002), только CHIKV (SCV305) или пустой контрольный вектор (SCV105), рассчитывали уровни нейтрализующих антител до заражения с использованием стандартного анализа микронейтрализации на клетках Vero на реюньонском штамме чикунгуньи. Кратко говоря, сыворотки инактивированли теплом (56°C в течение 30 мин), сыворотку от каждой мыши последовательной разводили в двух повторностях в 96-луночных планшетах и инкубировали с 100 CCID50 единицами вируса, инкубируя в течение 1 ч при 37°C. После этого этапа нейтрализации свежеразделенные клетки Vero накладывали (104 клеток на лунку) на смесь сыворотка/вирус и инкубировали в течение 5 дней до визуализации цитопатических эффектов под микроскопом. Разведение сыворотки, обеспечивающее 100% защиту от цитопатического эффекта, определяли с помощью окрашивания кристаллическим фиолетовым.
Результаты
[0140] Обе вакцины-кандидата, экспрессирующие антигены CHIKV, индуцировали нейтрализующие антитела к вирусу чикунгуньи после однократного введения вакцины.
[0141] Однако однократное введение одновекторной вакцины от CHIK/ZIKA индуцировало гораздо более высокие уровни CHIKV нейтрализующих антител, несмотря на эквивалентные дозы, используемые для вакцинации.
Вывод
[0142] Это исследование вакцинации продемонстрировало, что когда мышей вакцинируют однократным введением эквивалентных доз SCV305 (вакцина только от CHIK) и SCV1002 (одновекторная вакцина от CHIK/ZIKA), одновекторная вакцина (SCV1002) приводит к лучшей выработке чикунгунья-специфических нейтрализующих антител. Это было неожиданно, поскольку считалось, что экспрессия другого доминирующего антигена, т.е. белка E вируса Зика, будет создавать конкуренцию при иммунном ответе и, таким образом, ослабит или даже уничтожит иммунный ответ на доминирующие антигены как чикунгуньи (мишень для нейтрализующих антител антиген E2), так и вируса Зика (мишень нейтрализующих антител антиген E). Похоже, что это не так, и можно предположить, что экспрессия антигена вируса Зика имеет адъювантный эффект, помогая повысить иммунный ответ на антигены чикунгуньи, который ожидается для VLP чикунгуньи после вакцинации.
Защита плодов мышей от инфицирования вирусом Зика матерей, которые заранее были вакцинированы SCV1002 до беременности
[0143] Основными осложнениями продолжающейся вспышки вируса Зика в Америке и Азии являются врожденные дефекты, вызванные способностью вируса проникать через плаценту и инфицировать мозг плода. Setoh et al (2017, De Novo Generation and Characterization of New Zika Virus Isolate Using Sequence Data from a Microcephaly Case. mSphere 2:e00190-17. https://doi.org/101128/mSphereDirect.00190-17) описывает модель на мышах, в которой штамм ZIKV-Natal может быть использован для создания модели инфекции мозга плода у мышей IFNAR -/- (включая задержку внутриутробного развития), не вызывая симптоматических инфекций у самок.
[0144] Цель данного исследования состояла в том, чтобы показать, что предпринятая заранее вакцинация самок мышей SCV1002 (одновекторная от ZIKA/CHIK) до беременности может обеспечить защиту от инфицирования вирусом Зика их нерожденных плодов.
[0145] Это исследование было проведено путем вакцинации самок IFNAR -/- или SCV1002 (одновекторная вакцина от CHIK/ZIKA), или SCV105 (только вектор) с последующим скрещиванием с самцами мышей IFNAR. Беременных мышей затем инфицировали штаммом Natal вируса Зика, как описано Setoh et al (2017).
График вакцинации:
- Шести-восьминедельных дефицитных по рецептору интерферона самок мышей (IFNAR -/-) вакцинировали однократно внутримышечным путем или одновекторной вакциной от ZIKV/CHIKV (SCV1002), или пустой контрольной вакциной (SCV105) на неделе 0 по 106 на мышь
- У групп мышей брали кровь через 4 недели после вакцинации для проверки на сероконверсию к вакцине
- Через 6 недель после вакцинации были инициированы синхронные спаривания для вызывания беременности у вакцинированных мышей. Самок мышей ежедневно проверяли на наличие признаков успешной беременности (копулятивные пробки)
- В возрасте эмбрионов 6,5 дня беременных мышей инфицировали штаммом Zika Natal по 104 CCID50 единиц посредством подкожного инфицирования
- После инфицирования у беременных мышей брали кровь ежедневно с 1 по 5 день для проверки на виремию
- В возрасте эмбрионов 17,5 дня беременных мышей сортировали, материнскую плаценту и головки плода собирали для оценки на инфекционный ZIKV, как описано Setoh etal (2017).
Результаты
[0146] Беременные самки мышей, заранее вакцинированные SCV1002 (вакцина от CHIK/ZIKA) до того, как они забеременели, могли препятствовать репликации вируса Зика во время заражения штаммом Natal вируса Зика, что видно из отсутствия детектирования виремии после заражения. Однако, как и ожидалось, мыши, вакцинированные SCV105 (только вектор SCV), не могли препятствовать вирусной репликации вируса Зика, что видно из высокого уровня виремии, наблюдаемого в первые 5 дней после заражения.
[0147] Самки мышей, заранее вакцинированные одной дозой SCV1002 (вакцина от CHIK/ZIKA) до спаривания и беременности, не демонстрировали детектируемых уровней вируса Зика после заражения в своих плацентах и в головном мозге плодов. Вакцинация препятствовала инфицирование плаценты заражающим вирусом и тем самым блокировала дальнейшую передачу вируса Зика незащищенным плодам.
[0148] Однако все выглядело иначе у самок мышей, заранее вакцинированных только вектором SCV, у которых после заражения во время беременности некоторые плаценты были инфицированы, и происходила передача инфицирования вирусом Зика в головной мозг части плодов.
Выводы
[0149] Беременные самки мышей, заранее вакцинированные одной дозой одновекторной вакцины от CHIK/ZIKA, оказались защищены от заражения ZIKV по сравнению с вакцинированными контрольной вакциной, как показывают результаты виремии.
[0150] Вакцинация матерей до беременности обеспечивает защиту их нерожденных плодов посредством препятствования инфицирования материнской плаценты заражающим вирусом Зика и блокирования дальнейшей передачи в головной мозг плода.
[0151] Эти результаты демонстрируют в этой доклинической модели, что дальнейшая передача заражения вирусом Зика плодам может быть заблокирована путем предварительной вакцинации матери до беременности.
SCV1002 обеспечивает защиту мышей от смертельного заражения вирусом Росс-ривер
[0152] Чикунгунья (CHIKV) принадлежит к группе альфавирусов, а другим членом этой группы является вирус Росс-ривер (RRV). RRV эндемичен для северной и центральной частей Австралии, а также тихоокеанских регионов. RRV и CHIKV переносятся комарами и могут вызывать полиартрит и приводить к персистирующим инфекциям у людей. Однако комарами-переносчиками вируса чикунгуньи являются Aedes aegypti и Aedes albopictus, тогда как главными комарами-переносчиками вируса Росс-ривер являются два болотных комара Aedes camptorhynchus и Aedes vigilax и пресноводный комар Culex annulirostris.
[0153] Проводили исследование вакцинации на линии мышей (нокаут IRF3/7), которые летально восприимчивы к RRV, для определения, могут ли антигены вакцины от чикунгуньи из SCV1002 обеспечивать полную защиту от летального заражения RRV через шесть недель после вакцинации SCV1002 (вакцина от CHIK/ZIKA).
Схема эксперимента
[0154] Это исследование вакцинации и заражения проводили на мышах с нокаутом IRF3/7, которые восприимчивы к инфицированию как чикунгуньей, так и вирусом Росс-ривер, причем инфицирование этими вирусами летально для этой линии мышей.
[0155] Два группы мышей IRF3/7-/- вакцинировали или одной дозой 106 БОЕ на мышь SCV1002 (одновекторная вакцина от CHIK/ZIKA), или SCV105 (только вектор SCV) внутримышечным путем.
[0156] Через 6 недель после вакцинации всех мышей заряжали 104 CCID50 вируса Росс-ривер (штамм TT) подкожным путем в верхнюю часть стопы, как описано Rudd et al (2012).
[0157] За выживанием после заражения наблюдали в течение 30 дней после заражения. Мышей умерщвляли, когда клинические симптомы достигали этически определенных конечных точек (как описано в документе Rudd et al (2012) Interferon Response Factors 3 and 7 Protect against Chikungunya Virus Hemorrhagic Fever and Shock. J. Virol., 86 (18): 9888-9898)).
Результаты
[0158] Было обнаружено, что вакцинация IRF3/7-/- SCV1002, но не SCV105 обеспечивает полную защиту от летальности заражения RRV. Это демонстрирует, что вакцина от SCV-CHIK/ZIKV (SCV1002) обладает перекрестной защитой от других вирусов рода альфавирусов.
Вакцинированных SCV1002 мышей IFNAR-/- заражали ZIKV, а затем CHIKV
[0159] Группам по 6 взрослых самок мышей старше 6 недель инъецировали по 106 БОЕ на мышь SCV1002, SCZ305 или SCV105 (пустой вектор SCV) посредством интраперитонеальной инъекции. Мышей контролировали на предмет любых побочных эффектов после вакцинации.
[0160] Через шесть недель после вакцинации группы мышей заражали 103 CCID50 на мышь адаптированного для мышей африканского штамма вируса Зика MR766 посредством подкожной инъекции в основание хвоста.
[0161] Через шесть недель после заражения вирусом Зика выжившие группы мышей заражали 104 CCID50 на мышь реюньонского штамма вируса чикунгуньи (LR2006-OPY1) посредством подкожной инъекции в верхнюю часть стопы.
Результаты
[0162] Вакцинированные SCV1002 (одновекторная вакцина от CHIK+ZIK) мыши оказались полностью защищены от заражения вирусом Зика, однако все вакцинированные SCV305 (вакцина только от CHIK) и SCV105 (только вектор) мыши умерли от инфицирования вирусом Зика инфекция к 9 дню после инфицирования.
[0163] Вакцинированные SCV1002 (одновекторная вакцина от CHIK+ZIK) мыши IFNAR-/-, которые сначала были заражены ZIKV и выжили, были защищены частично, т.е. с эффективностью 33%, от чрезвычайно летального заражения CHIKV, которое оказалось способно убить всех невакцинированных мышей к 3 дню после заражения.
Выводы
[0164] Вакцинация мышей INFAR-/- однократной дозой SCV1002 по 106 БОЕ на мышь обеспечивала полную защиту от летального заражения адаптированным для мышей штаммом вируса Зика.
[0165] Последующее заражение выживших мышей чрезвычайно летальной дозой CHIKV показало частичную защиту, т.е. вакцина SCV1002 оказалась эффективна на 33% в этой модели на мышах в отношении инфицирования CHIKV в дозе 104 инфекционных единиц на мышь. Заражающая доза 104 инфекционных единиц оказалась слишком высокой для этой модели на мышах с ослабленным иммунитетом, у которых отсутствуют рецепторы интерферона, и поэтому они не могут реагировать на интерферон противовирусным образом. Продуцирование противовирусных интерферонов обычно контролирует первоначальную инфекцию, пока адаптивный иммунный ответ успевает отреагировать на инвазивную вирусную инфекцию. Ретроспективно, эта доза для заражения CHIKV в этой модели на мышах оказалась слишком высокой, поскольку она убила всех невакцинированных мышей за 3 дня - слишком короткий период времени для выработки противовирусный анамнестических антител, для того чтобы контролировать и устранить инвазивную вирусную инфекцию. В естественной ситуации противовирусные интерфероны будут контролировать первоначальную вирусную инфекцию достаточно долго для того, чтобы адаптивный иммунный ответ пришел им на смену и нейтрализовал и удалил инфекцию из хозяина. Несмотря на неестественно высокий уровень вирулентности CHIKV для мышей IFNAR, когда невакцинированные мыши умерли к 3 дню после инфицирования дозой 104 инфекционных единиц, вакцинация SCV1002 неожиданно смогла обеспечить некоторую защиту в этих экстремальных условиях, что подчеркивает высокую иммуногенную активность этой вакцины.
Инфицирование CHIKV после вакцинации SCV1002
[0166] Целью данного исследования была оценка антигенного взаимовлияния или антигенного стимула в контексте естественного инфицирования. Повлияет ли предыдущее естественное инфицирование вирусом чикунгуньи на индукцию иммунного ответа на вирус Зика, при том что имеет место индуцированый вакциной вторичный ответ на чикунгунью после вакцинации SCV1002 (вакцина от SCV-CHIK/ZIKA).
[0167] Мышей C57BL/6 дикого типа инфицировали вирусом чикунгуньи, допускали уничтожение вируса, а затем стимулировали одновекторной вакциной от CHIKV/ZIKV. У мышей брали кровь до введения вакцины и через 4 недели после вакцинации для оценки уровней выработки анти-CHIKV и анти-ZIKV антител.
[0168] Группам по 6 взрослых самок мышей старше 6 недель инъецировали по 104 CCID50 на мышь реюньонского штамма вируса чикунгуньи (LR2006-OPY1) посредством подкожной инфекции в верхнюю часть стопы. Мышей контролировали на предмет любых побочных эффектов после инфицирования и брали у них кровь через 8 недель после инфицирования до вакцинации.
[0169] Через восемь недель после инфицирования мышей стимулировали одновекторной вакциной от CHIKV/ZIKV (SCV1002) по 106 БОЕ на мышь, вводимой интраперитонеально. Мышей контролировали на предмет любых побочных эффектов после вакцинации и через четыре недели после вакцинации, у мышей снова брали кровь для сравнения уровней анти-CHIKV и анти-ZIKV антител до и после вакцинации.
[0170] Было обнаружено, что вакцинация SCV1002 мышей дикого типа, ранее подвергшихся воздействию инфекции CHIKV, индуцирует анти-ZIKV антитела на уровнях, аналогичных наблюдаемым ранее без инфицирования CHIKV.
[0171] Антигенный стимул представляет собой явление, при котором воздействие одного антигена может ингибировать способность второго антигена индуцировать защитный иммунный ответ. Предполагается, что в механизм, лежащий в основе этой концепции, включены B-клетки, примированные первым антигеном, которые запрограммированы отвечать только на первый антиген. Когда вводится второй антиген, эти B-клетки по существу удаляют второй антиген до того, как может возникнуть продуктивный иммунный ответ, что приводит к очень низкому или плохому иммунитету. В этом исследовании проверяли эту концепцию.
[0172] Однако поскольку мыши, ранее инфицированные вирусом чикунгуньи, а затем вакцинированные вакциной от CHIKV/ZIKV, способны индуцировать продуктивный иммунный ответ на CHIKV и ZIKV, аналогичный наблюдаемому без предварительного инфицирования CHIKV, антигенный стимул не играет роли в этих условиях. Это скорее всего обусловлено тем, что CHIKV и ZIKV являются очень разными вирусами, происходящими из двух разных семейств вирусов. Эти данные позволяют предположить, что двойная вакцина могла бы использоваться в странах с известным предшествующим распространением CHIKV.
Защита от инфицирования вирусом Зика у самцов мышей
[0173] Вирус Зика остается единственным вирусом, передаваемым комарами, который также может передаваться половым путем. Вирус Зика может передаваться от мужчины к женщине или от мужчины к мужчине, и было показано, что вирус сохраняется в сперме в течение нескольких месяцев после появления симптомов.
[0174] Цель данного исследования состояла в том, чтобы показать, что вакцинация SCV1002 (одновекторная от ZIKA/CHIK) может обеспечивать защиту от инфицирования вирусом Зика у самцов мышей.
[0175] Это исследование было проведено путем вакцинации самцов IFNAR-/- или SCV1002 (одновекторная вакцина от CHIK/ZIKA), или SCV105 (только вектор) с последующим инфицированием штаммом Natal вируса Зика, как описано в документе Setoh et al (2017) (De Novo Generation and Characterization of New Zika Virus Isolate Using Sequence Data from a Microcephaly Case. mSphere 2:e00190-17. https://doi.org/10.1128/mSphereDirect.00190-17).
[0176] Самцов дефицитных по рецептору интерферона мышей (IFNAR -/-) в возрасте 6-8 недель вакцинировали внутримышечным путем или одновекторной вакциной от ZIKV/CHIKV (SCV1002), или пустой контрольной вакциной (SCV105) на неделе 0 по 106 БОЕ на мышь.
[0177] У групп мышей брали кровь через 4 недели после вакцинации для проверки на сероконверсию к вакцине. Через 6 недель после вакцинации самцов мышей инфицировали штаммом Natal вируса Зика по 104 CCID50 единиц посредством подкожного инфицирования. После инфицирования у мышей брали кровь ежедневно с 1 по 5 день для проверки на виремию.
[0178] На 21 день после инфекции мышей сортировали, собирали семенники для оценки РНК ZIKV с использованием количественной ПЦР в реальном времени с использованием праймеров, нацеленных на ген prM. Экстрагировали РНК с использованием тризола (Life Technologies), следуя инструкциям производителя. Синтезировали кДНК с использованием Biorad Iscript Reverse Transcriptase Supermix, следуя инструкциям производителя. Количественный анализ в реальном времени проводили с использованием набора Biorad iTaq Universal Sybr green supermix. Результаты ПЦР в реальном времени определяли количественно по отношению к уровням конститутивного гена RPL13.
[0179] Самцы мышей, вакцинированные SCV1002 (вакцина от CHIK/ZIKA) не были способны к репликации вируса Зика после заражения штаммом Natal вируса Зика, что показывает минимальное детектирование виремии после заражения. Однако, как и ожидалось, вакцинированные SCV105 (только вектор SCV) мыши были способны к репликации вируса Зика, что показывает высокий уровень виремии, наблюдаемый в первые 3 дня после заражения.
[0180] Вакцинация SCV1002 (CHIK/ZIKA) также смогла снизить количество вирусной РНК, наблюдаемой в семенниках на 21 день, по сравнению мышами, вакцинированными контрольной вакциной SCV105.
СПИСОК ЛИТЕРАТУРЫ
Altschul et al. (1997) Nucleic Acids Research 25:3389-3402
Ausubel et al. (1999) Current Protocols in Molecular Biology (Supplement 47), John Wiley & Sons, New York
Boshart et al. (1985) Cell 41:521
Brooks et al. (1995) J. Virol. 69(12):7688-7698
Dijkema et al. (1985) EMBO J. 4:761
Drillien R, et al. (1978) J Virol. 28(3):843-50
Gorman et al., (1982) Proc. Natl. Acad. Sci. USA 79:6777
Ham RG. (1965) Proc. Natl. Acad. Sci. USA 53: 288-293
Hsiao JC, et al. (2006) J. Virol. 80(15):7714-28
Kibler et al. (2011) PLOSONE 6(11)
Meisinger-Henschel et al. (2007) J. Gen. Virol. 88(12):3249-3259
Murphy et al. (1995) Virus Taxonomy Springer Verlag:79-87
Puck TT, et al. (1958) J. Exp. Med. 108:945-956
Sambrook et al. (1989) Molecular Cloning: A Laboratory Manual, 2nd ed., Cold Spring Harbor Press, Plainsview, N.Y.
Shisler et al. (2004) J. Virol. 78(7):3553-3560
Spehner D, et al. (1988) J Virol., 62(4):1297-1304
Werden SJ, et al. (2008) Chapter 3: Poxvirus Host Range Genes. In: Advances in Virus Research, 71
| название | год | авторы | номер документа |
|---|---|---|---|
| ПОЛУЧЕНИЕ ВИРУСНОГО ВЕКТОРА | 2014 |
|
RU2658487C2 |
| НАБОР ОЛИГОНУКЛЕОТИДНЫХ ПРАЙМЕРОВ И ФЛУОРЕСЦЕНТНОМЕЧЕНЫХ ЗОНДОВ ДЛЯ ВИДОСПЕЦИФИЧНОЙ ЭКСПРЕСС-ИДЕНТИФИКАЦИИ ОРТОПОКСВИРУСОВ НА ОСНОВЕ МУЛЬТИПЛЕКСНОЙ ПЦР В РЕАЛЬНОМ ВРЕМЕНИ | 2010 |
|
RU2427648C1 |
| СРЕДСТВА И СПОСОБЫ ЛЕЧЕНИЯ ВИРУСА ГЕПАТИТА B | 2017 |
|
RU2740802C2 |
| ИММУНОГЕННЫЕ КОМПОЗИЦИИ, СОДЕРЖАЩИЕ СИЛИКАТИРОВАННЫЙ ВИРУС, И СПОСОБЫ ПРИМЕНЕНИЯ | 2014 |
|
RU2676083C2 |
| M2-ДЕФЕКТНЫЙ ПОКСВИРУС | 2019 |
|
RU2819245C2 |
| Живая аттенуированная культуральная вакцина для профилактики натуральной оспы и других ортопоксвирусных инфекций на основе вируса осповакцины и способы ее получения и применения | 2022 |
|
RU2781070C1 |
| КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ ПОВЫШЕНИЯ СТАБИЛЬНОСТИ ТРАНСГЕНОВ В ПОКСВИРУСАХ | 2017 |
|
RU2756534C2 |
| Рекомбинантный штамм VACΔ6 вируса осповакцины с нарушенными генами вирулентности C3L, N1L, J2R, A35R, A56R, B8R для получения живой культуральной аттенуированной вакцины против натуральной оспы и других ортопоксвирусных инфекций человека | 2016 |
|
RU2621868C1 |
| ПЕРСОНАЛИЗИРОВАННАЯ ВАКЦИНА | 2018 |
|
RU2779987C2 |
| РЕКОМБИНАНТНЫЙ ВИРУС СВИНОЙ ОСПЫ (SWINEPOX) И ВАКЦИНЫ | 2015 |
|
RU2727899C2 |

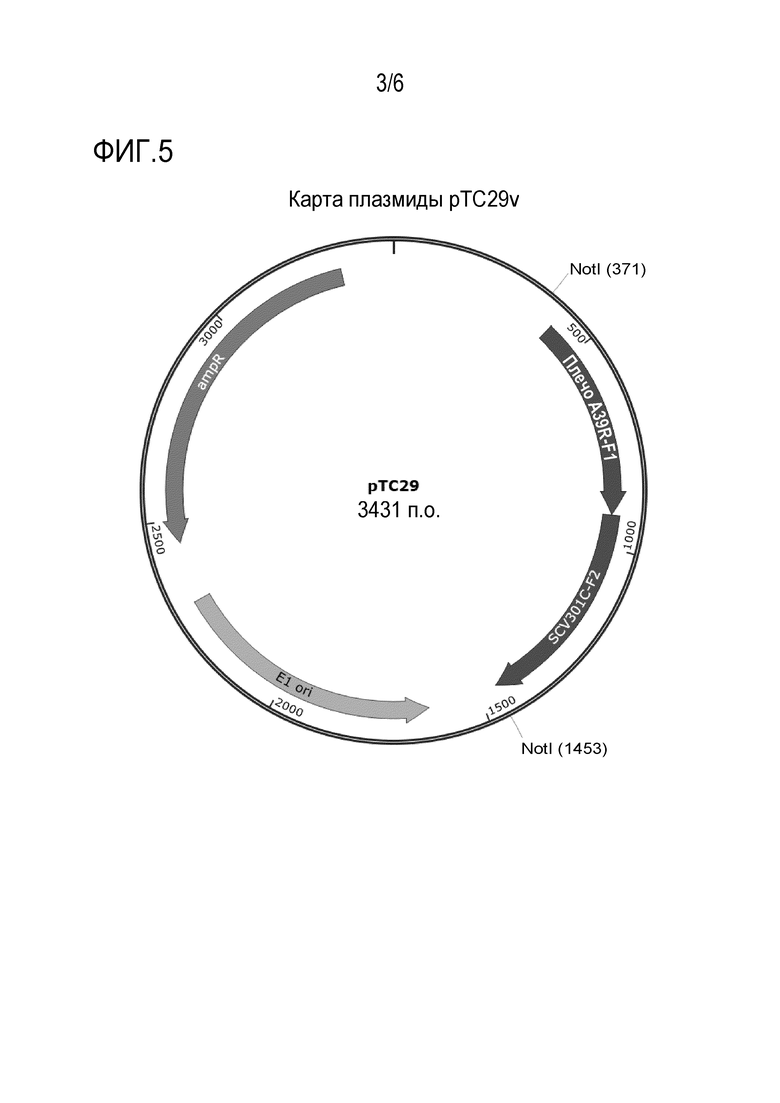
Группа изобретений относится к области медицины, а именно к инфектологии и иммунологии, и предназначена для индуцирования у пациента защитного иммунного ответа на инфицирование чикунгуньей, натуральной оспой и вирусом Зика. Композиция, которая индуцирует у пациента защитный иммунный ответ на инфицирование чикунгуньей, натуральной оспой и вирусом Зика, содержит фармацевтически приемлемый носитель и аттенуированный поксвирус. Геном поксвируса содержит последовательность нуклеиновой кислоты, кодирующую субгеномный полипротеин 26S вируса чикунгуньи, и последовательность нуклеиновой кислоты, кодирующую полипротеин PrME вируса Зика. Аттенуированный поксвирус представляет собой модифицированный поксвирус, причем модификация включает делецию по меньшей мере одного гена, кодирующего эндогенный незаменимый белок сборки или созревания. Также представлено применение указанной композиции для индуцирования у пациента защитного иммунного ответа на инфицирование чикунгуньей, натуральной оспой и вирусом Зика. Использование группы изобретений позволяет снизить риск инфицирования чикунгуньей, вирусом Зика и натуральной оспой. 2 н. и 9 з.п. ф-лы, 4 табл., 9 ил., 3 пр.
1. Композиция, которая индуцирует у пациента защитный иммунный ответ на инфицирование чикунгуньей, натуральной оспой и вирусом Зика, содержащая фармацевтически приемлемый носитель и аттенуированный поксвирус, причем геном поксвируса содержит последовательность нуклеиновой кислоты, кодирующую субгеномный полипротеин 26S вируса чикунгуньи, и последовательность нуклеиновой кислоты, кодирующую полипротеин PrME вируса Зика; и причем аттенуированный поксвирус представляет собой модифицированный поксвирус, причем модификация включает делецию по меньшей мере одного гена, кодирующего эндогенный незаменимый белок сборки или созревания.
2. Композиция по п. 1, в которой аттенуированный поксвирус выбирают из группы, состоящей из вируса осповакцины, модифицированного вируса осповакцины Анкара (MVA), NYVAC, авипоксвируса, вируса оспы канареек и вируса оспы птиц.
3. Композиция по п. 2, в которой аттенуированный поксвирус представляет собой модифицированный ортопоксвирус.
4. Композиция по любому из пп. 1-3, в которой модификация содержит делецию гена D13L.
5. Композиция по любому из пп. 1-4, в которой модификация содержит делецию гена K1L.
6. Композиция по любому из пп. 1-5, в которой модификация содержит делецию гена A39R.
7. Композиция по любому из пп. 1-6, в которой модификация содержит делецию генов B7R-B8R.
8. Композиция по любому из пп. 1-7, в которой модификация содержит делецию гена D13L, гена A39R и генов B7R-B8R.
9. Композиция по любому из пп. 1-8, в которой фармацевтически приемлемый носитель содержит адъювант.
10. Композиция по п. 9, в которой адъювант выбирают из группы, состоящей из гидроксида алюминия, фосфата алюминия, сульфата алюминия-калия, гидроксида-фосфата кальция, полного адъюванта Фрейнда, неполного адъюванта Фрейнда, искомов и искоматрикса.
11. Применение композиции по любому из пп. 1-10 для применения в индуцировании у пациента защитного иммунного ответа на инфицирование чикунгуньей, натуральной оспой и вирусом Зика.
| WO 2016086980 A1, 09.06.2016 | |||
| WO 2010037402 A1, 08.04.2010 | |||
| GARCIA-ARRIAZA J et al | |||
| A Novel Poxvirus-Based Vaccine, MVA-CHIKV, Is Highly Immunogenic and Protects Mice against Chikungunya Infection | |||
| JOURNAL OF VIROLOGY, 2014, vol | |||
| Шланговое соединение | 0 |
|
SU88A1 |
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| Водолазный аппарат | 1925 |
|
SU3527A1 |
| LAROCCA RA et al | |||
| Vaccine protection against Zika virus from Brazil | |||
| NATURE, 01.08.2016, | |||

