ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение, в общем, относится к персонализированной противораковой вакцине, содержащей рекомбинантный поксвирус, кодирующий один или более чем один неопептид, или к композиции, содержащей такой рекомбинантный поксвирус и фармацевтически приемлемый носитель, а также к применению указанной персонализированной противораковой вакцины для лечения ракового субъекта, нуждающегося в этом. Конкретное воплощение направлено на способ предоставления такой вакцины или композиции, включающий стадию идентификации, включающую а) экстракцию ДНК из образца опухоли и неопухолевого образца, б) выбор областей-мишеней, предпочтительно полных кодирующих областей генома (экзом), в) секвенирование указанных областей-мишеней (например, экзома) из указанных экстрагированных ДНК и г) идентификацию одной или более чем одной опухолеспецифичной мутации посредством сравнения последовательностей ДНК, полученных из указанных опухолевых и неопухолевых образцов. Воплощения также включают способ лечения рака или предупреждения его рецидива, включающий введение такой персонализированной противораковой вакцины. Данное изобретение представляет очень специальный интерес в области персонализированной иммунотерапии, в частности, для стимуляции иммунного ответа Т-клеток.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
На протяжении последних десятилетий были получены многочисленные терапевтические вакцины, экспрессирующие опухолевые антигены, с целью стимуляции врожденных и специфичных иммунных ответов против опухолевых антигенов. Однако чаще всего идентифицируемые антигены, ассоциированные с опухолью (например, MUC-1, WT1, PSA, СЕА), селективно сверхэкспрессируются в опухоли, но могут иметь остаточную экспрессию в неопухолевых клетках. Таким образом, ожидается то, что эффективность данного традиционного подхода ограничивается аутотолерантностью против таких «своих» антигенов.
Кроме того, традиционная парадигма доставки того же самого набора раковых антигенов любому в популяции игнорирует индивидуальную изменчивость в ответах, связанных с риском заболевания, и иммунологических ответах. Важно, что не могло бы игнорироваться то, что люди по-разному отвечают на вакцины, и иммунный ответ хозяина значительно варьирует в популяции (Relman, 2008, J Infect Dis. 198(1):4-5; Plotkin, 2008, Clin Infect Dis.47(3):401-9). С недавним введением в клинике блокаторов иммунной контрольной точки, в самом деле, для медицинского персонала стало очевидным то, что некоторые лечения являются эффективными для некоторых пациентов, но не для других.
Успехи в иммунологии, генетике, молекулярной биологии и биоинформатике проторили путь к более персонализированным подходам. Технологические прорывы в области секвенирования генома (в частности, секвенирование следующего поколения (NGS)) теперь сделали возможным секвенирование полного генома или экзома (кодирующих областей генома) опухоли с беспрецедентными скоростью и затратами. Молекулярная характеристика опухолей продемонстрировала то, что во время процесса канцерогенеза и пролиферации опухолевых клеток мутации происходят вследствие высоких скоростей пролиферации, недостаточных механизмов репарации и клональной селекции. Накопление мутаций в геноме опухоли обычно приводит к экспрессии дефектных белковых соединений, которые являются специфичными для раковой ткани. Они называются неоантигенами. В отличие от самых обычных антигенов, ассоциированных с опухолью, опухолевые неоантигены присутствуют только в опухолевых клетках, но не в нормальных клетках, и не индуцируют устранение их антигенспецифичных Т-клеток в тимусе. Таким образом, ожидается то, что они индуцируют сильные иммунные ответы, не неся риска аутотолерантности и аутоиммунных реакций против собственных белков. Следовательно, опухолевые неоантигены могут быть идеальными мишенями для разработки терапевтических вакцин, специфично адаптированных к опухоли, хотя их принятие в установившиеся способы помощи требует преодоления целого ряда научных и технических вызовов. В частности, большинство раковых мутаций являются результатом стохастического явления и являются специфичными для каждого пациента.
Выбор подходящей вакцинной платформы является важным фактором для ее успеха в клинической ситуации. В настоящее время рассматриваются несколько технологических платформ для разработки вакцин, направленных на неоантигены, включая бактерии (как, например, вакцина ADXS-Neo на основе Listeria, разработанная Advaxis; WO 2016/207859; WO 2016/191545), вирусные векторы (как, например, лентивирусная система ZVex от Immune Design), вакцины на основе нуклеиновой кислоты (например, вакцина на основе кислой мРНК, инкапсулированная в наночастицы, в том виде, в котором она разработана Biontech; Kreiter et al., 2015, Nature 520 692-6; WO 2012/159754 - ДНК, кодирующая вакцинный комплекс, разработанная Vaccibody) и адъювантные пептиды (WO 2016/187508). Репрезентативные примеры пептидных вакцин включают, например, роlуIСLС(полиносиновая-полицитидильная кислота)-стабилизированные пептиды, разработанные в настоящее время NEON Therapeutics, при раковых заболеваниях мочевого пузыря, глиобластомах и немелкоклеточных раковых заболеваниях легкого (NSCLC), в комбинации с ниволумабом; gр96(96 кДа белок теплового шока)-адъювантные опухолевые неоантигены, разработанный Agenus; и инкапсулированные в липосомы пептиды от Школы медицины Университета Коннектикута). Другой недавней альтернативой является применение дендритных клеток (DC) в качестве вектора после воздействия антигена in vitro перед введением пациенту. DC подвергаются импульсному воздействию пептидами или лизатами опухоли (например, активированные DC вакцины - Sipuleucel-T, распространяемые на рынке Dendreon, DC, загруженные экзомами опухоли пациента, разработанные Exocyte Therapeutics; DC вакцина, разработанная OncoTherapeutics Science и Tella Inc).
Тем не менее, большинство инициатив в данной области должно удовлетворять главным вызовам перед выполнением их обещаний. Среди других тем, успешное претворение в жизнь подчиняется достижению эффективного способа изготовления для обеспечения быстрой доставки клинически достаточных количеств к койке пациента, поскольку идентификация опухолевых мутаций, конструирование неопептидов, включающих такие мутации, изготовление и тестирование персонализированной вакцины являются сопутствующими прогрессированию заболевания.
Таким образом, существует потребность в разработке хорошо налаженного способа с эффективным использованием времени, обеспечивающего идентификацию неоантигенов для каждой опухоли пациента и изготовление соответстующей персонализированной терапии со стандартами, установленными для изготовления лекарственного средства.
Рекомбинантный MVA (модифицированный вирус Анкара) служил в качестве эффективного вектора в предыдущих попытках разработки противораковых вакцин (Acres and Bonnefoy, 2008, Expert Review of Vaccines 7, 889-93). Вакцины на основе вируса осповакцины достигали результатов, когда их давали в виде монотерапии или в комбинации с химиотерапией, лучевой терапией или ингибитором иммунной контрольной точки (Farsaci et al., 2011, In Cancer Vaccines: From Research to Clinical Practice, Ed Bot; CRC Press, pp56-77; WO 2015/175340; WO 2015/175334). Например, TG4010 (или MVATG9931 с ее исследовательским названием) - противораковая вакцина на основе MVA, кодирующая ассоциированный с опухолью антиген MUC1 и человеческий интерлейкин 2 (IL-2) - демонстрировала эффективность в комбинации с химиотерапией стандарта помощи первой линии при прогрессирующем метастатическом NSCLC (немелкоклеточный рак легкого) (Quoix et al., 2011, The Lancet Oncology 12(12): 1125-33). Контроль антигенспецифичной опухоли посредством MVATG9931 в профилактической модели RMA-MUC1 явно зависит от временной экспрессии de novo MUC1 и Т-клеток CD8+ и CD4+. Другие поксвирусы и, в частности, вирусы осповакцины и вирусы оспы кур, также использовали в качестве противораковых векторов (Gulley et al., 2008, Clin Cancer Res 14(10): 3060-9).
Однако способность MVA или других поксвирусов индуцировать сильный иммунный ответ против раковых неоантигенов не тестировали, и ее остается установить. Кроме того, известные способы получения рекомбинантных MVA основаны на гомологичной рекомбинации, которая имеет низкую эффективность и, таким образом, является довольно длительной.
Однако авторы данного изобретения однаружили то, что рекомбинантный MVA, в самом деле, способен индуцировать иммунные ответы против нескольких раковых неоантигенов, и что персонализированный MVA, кодирующий несколько неопептидов, может быть быстро получен.
Данное раскрытие удовлетворяет вышеупомянутую потребность посредством предложения противораковых вакцин на основе MVA, экспрессирующих слияние противоопухлевых неопептидов, а также способа изготовления, который был конкретно разработан для максимизации образования рекомбинантного MVA и улучшения масштабируемости и времени до завершения способа.
Данная техническая проблема решается предложением воплощений, как определено в формуле изобретения.
Другие и дополнительные аспекты, характеристики и преимущества настоящего изобретения будут очевидными из следующего описания предпочтительных в настоящее время воплощений данного изобретения. Данные воплощения приводятся с целью раскрытия.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Настоящее изобретение, в общем, относится к персонализированной противораковой вакцине, содержащей рекомбинантный поксвирус, кодирующий один или более чем один неопептид.
В одном аспекте рекомбинантный поксвирус представляет собой вирус осповакцины, предпочтительно дефектный по репликации вирус осповакцины, такой как MVA.
В другом аспекте каждый из указанного одного или более чем одного неопептида, кодируемого рекомбинантным поксвирусом, содержит по меньшей мере одну опухолеспецифичную мутацию с предпочтением того, что по меньшей мере 60% указанных неопептидов содержат миссенс-мутацию или мутацию со сдвигом рамки считывания. Желательно один или более чем один неопептид имеет длину от 16 до 90 аминокислотных остатков, предпочтительно от 17 до 85 аминокислотных остатков и более предпочтительно от 18 до 80 аминокислотных остатков. В предпочтительных воплощениях неопептиды, содержащие миссенс-мутацию, имеют длину от 18 до 29 остатков, и неопептиды, содержащие мутацию со сдвигом рамки считывания, имеют длину от 30 до 80 аминокислотных остатков. По меньшей мере 80% неопептидов, имеющих миссенс-мутацию, несут замененную аминокислоту в центральном положении. В предпочтительном воплощении несколько и предпочтительно все неопептиды экспрессируются рекомбинантным поксвирусом в виде одного или более чем одного слияния. Данный рекомбинантный поксвирус может дополнительно кодировать один или более чем один терапевтический ген, предпочтительно выбранный из группы, состоящей из суицидных и иммуностимулирующих генов.
В другом аспекте согласно настоящему изобретению также предложен способ получения указанной персонализированной противораковой вакцины, включающий стадию идентификации одного или более чем одного неопептида, подходящего для кодирования персонализированной противораковой вакциной, где указанный один или более чем один неопептид содержит по меньшей мере одну опухолеспецифичную мутацию. Предпочтительный способ включает стадию идентификации, включающую следующие подстадии а) - г): а) экстрагирование ДНК из образца опухоли и неопухолевого образца, б) выбор областей-мишеней, предпочтительно всех кодирующих областей генома (экзом), в) секвенирование указанных областей-мишеней (например, экзома) из указанных экстрагированных ДНК и г) идентификация одной или более чем одной опухолеспецифичной мутации посредством сравнения последовательностей ДНК, полученных из указанных образцов опухоли и неопухолевых образцов. Такой способ может дополнительно включать по меньшей мере одну из следующих дополнительных подстадий д) - ж): д) ранжирование потенциального(ных) нуклеопептида(дов) по его(их) уровням экспрессии в опухоли либо на уровне транскрипции мРНК, либо на уровне трансляции белка; е) выбор неаутоэкспрессируемой(мых) опухолеспецифичной(ных) мутации(ций); ж) прогнозирование иммунного потенциала неоэпитопа(пов), содержащегося(сихся) в указанном(ных) неопептиде(дах).
Помимо стадии идентификации, данный способ также может включать стадию получения указанного рекомбинантного поксвируса. Предпочтительно указанная(ные) молекула(лы) нуклеиновой кислоты, кодирующая(щие) неопептид, подлежащая(щие) вставке в геном рекомбинантного поксвируса, организована(ны) в одной или более чем одной экспрессионной кассете под контролем подходящих регуляторных элементов, обеспечивающих экспрессию у данного субъекта. Предпочтительно в указанной стадии получения рекомбинантного поксвируса используется родительский поксвирус, содержащий ген флуоресцентного репортера, клонированный в месте вставки, которое выбрано для молекулы(кул) нуклеиновой кислоты, кодирующей(щих) неопептид, или экспрессионной(ных) кассеты(сет). В предпочтительном воплощении указанная стадия получения указанного рекомбинантного поксвируса включает стадию расщепления эндонуклеазой, способной генерировать по меньшей мере один двухнитевой разрыв в указанной нуклеотидной последовательности флуоресцентного репортера, но в которой указанная эндонуклеаза не расщепляет поксвирусный геном. Указанный способ также может включать стадию изготовления, где указанная стадия изготовления включает стадию амплификации в подходящей клетке-продуценте в подходящем масштабе и стадию выделения продуцированного рекомбинантного поксвируса из культуры клеток, и возможную стадию очистки выделенного рекомбинантного поксвируса.
В еще одном другом аспекте персонализированная противораковая вакцина служит для применения у субъекта, нуждающегося в этом, для лечения рака или предупреждения его рецидива у субъекта. Указанное раковое заболевание предпочтительно представляет собой солидную опухоль и, в частности, рак мозга или рак легкого. В предпочтительном воплощении персонализированная противораковая вакцина вводится в сочетании с одной или более чем одной дополнительной противораковой трапией, которая имеют пользу при лечении рака.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
На Фиг. 1 проиллюстрирована конструкция экспрессионной кассеты, кодируемой MVATG19022 и MVATG19023. «GS» представляет 10-аминокислотные линкеры, «Flag» - метку Flag, и «SR» представляет собой сокращение для сигнального пептида, полученного из гликопротеина вируса бешенства. FCU1, b-Gal, E16HPV16, E76HPV16 и MCU1 представляют пептиды длиной 27 аминокислот, полученные из данных антигенов.
На Фиг. 2 А-Д проиллюстрирован ELISpot (метод иммуноферментных пятен) после иммунизации мышей С57bI/6 (5 мышей на группу) векторами MVATG 19022 и MVATG19023, кодирующими пептид, по сравнению с векторами MVA, кодирующими целые антигены, соответственно, MUC1 (MVA-MUC1), FCU1 (MVA-FCU1), b-галактозидаза (MVA-bGal), Е7 HPV-16 (MVA-E7HPV16) и Е1HPV-16 (MVA-E1HPV16). Стимуляцию делали либо пептидом, специфичным для каждого экспрессируемого антигена/пептида (соответственно FCU1-специфичный EG15 (А), MUC1-специфичный L15L3 (Б), Е7-специфичный R9F (В), Е1-специфичный I8L (Г) и b-Gal-специфичный I8V5Е)), либо нерелевантным пептидом («CTRL» для контроля).
На Фиг. 3 проиллюстрированы аминокислотные последовательности первого и второго пентатопного слияний СТ26, экспрессируемых MVATG19030.
На Фиг. 4 проиллюстрирован ELISpot после иммунизации мышей Balb/c (5 мышей на группу) векторами MVATG19030 и MVATG19038. Ответ IFNγ (интерферон-гамма) оценивали после стимуляции либо пулом из десяти мутировавших пептидов СТ26, либо пулом пептидов СТ26 дикого типа (т.е. которые не содержат опухолеспецифичную мутацию) с примированием клеток CT26-MUC1 или без него.
На Фиг. 5 проиллюстрирован ответ IFNγ, генерированный против мутировавшего пептида СТ26 (ЕН27m) и немутировавшего пептида СТ26 (PE27wt), после иммунизации мышей Balb/c MVATG 19030 и MVATG 19038 (или буфером для приготовления вируса S08 в качестве негативного контроля) как в мышиной модели, примированной клетками CT26-MUC1, так и не примированной. Стимуляция культуральной средой проводится в качестве негативного контроля.
На Фиг. 6 проиллюстрирован ответ IFNγ, генерированный против мутировавшего пептида СТ26 (ЕН27m) и немутировавшего пептида СТ26 (PE27wt), после иммунизации мышей Balb/c MVATG 19030 и MVATG 19038 (или буфером для приготовления вируса S08 в качестве негативного контроля) как в мышиной модели, примированной клетками CT26-MUC1, так и не примированной. Ответ IFNγ оценивается в присутствии (плюс антитело против МНСII (главный комплекс гистосовместимости II)) или в отсутствие антител против мышиного МНСII. Стимуляция культуральной средой проводится в качестве негативного контроля.
На Фиг. 7 проиллюстрированы вырожденные нуклеотидные последовательности из 9 нуклеотидов, кодирующие линкеры GSG, GTS или GAS.
На Фиг. 8 проиллюстрирована процентная доля выделенных белых бляшек и число рекомбинантных (негативных в отношении mCherry и позитивных в отношении mCherry) или родительских (т.е. нерекомбинантных) вирусов, идентифицированных посредством ПЦР (полимеразная цепная реакция) после трансфекции клеток CEF (фибробласты куриных эмбрионов) плазмидами, показанное согласно числу кассет слияния, которые несет указанная конструкция (1, 2 или 3), числу неопептидов, содержащихся в каждом слиянии (Nbnео), присутствию или отсутствию ТМ (трансмембранный) сегмента (ТМ), баллу гидропатичности, рассчитанному для каждого слияния (Hydro), и присутствию и длине линкеров для каждой конструкции.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Общие определения
Если не определено иначе, все технические и научные термины, используемые в данном документе, имеют такое же значение, которое обычно понятно обычному специалисту в области, к которой относится данное изобретение.
Термин «один» относится к «одному» или к «более чем одному» грамматическому объекту предмета (т.е. по меньшей мере одному, включающему 2, 3, 4, 5 и т.д.), если контекст явно не диктует иное.
Термин «и/или» всякий раз, когда он используется в данном документе, включает значение «и», «или» и «все или любая другая комбинация элементов, соединенных указанным термином».
Термины «такой как», «например» в том виде, в котором они используются в данном документе, предназначены для иллюстративных целей и, таким образом, являются неограничивающими.
Термин «примерно» или «приблизительно» используется в данном документе для указания того, что значение или интервал, приведенный в данном документе, не является критическим и может варьировать в пределах 10%, предпочтительно в пределах 8% и более предпочтительно в пределах 5% от данного значения или интервала таким образом, чтобы включать подлинную вариацию или ошибку для используемого устройства или способа для определения такого значения или интервала, или вариацию, которая существует среди протестированных субъектов.
Термин «содержащий» (и любая форма содержащего, такая как «содержать» и «содержит»), «имеющий» (и любая форма имеющего, такая как «иметь» и «имеет»), «включающий» (и любая форма включающего, такая как «включает» и «включать») в том виде, в котором они используются в данном документе, при использовании для определения продуктов, композиций и способов, являются открытыми и не исключают дополнительных, неперечисленных элементов или стадий способа. Фраза «по существу состоящий из» означает исключение других компонентов или стадий, имеющих какую-либо существенную значимость. «Состоящий из» означает исключение более чем следовых элементов других компонентов или стадий.
Термины «полипептид», «пептид» и «белок» используются взаимозаменяемо для названия полимеров из аминокислотных остатков, содержащих по меньшей мере девять аминокислот, ковалентно связанных посредством пептидных связей. Полимер может быть линейным, разветвленным или циклическим и может содержать встречающиеся в природе аминокислоты и/или аналоги аминокислот, и он может прерываться неаминокислотами. Не накладывается ограничение на максимальное число аминокислот, содержащихся в полипептиде. В качестве общего указания, данный термин относится как к коротким полимерам (типично обозначенным в данной области как пептид), так и к более длинным полимерам (типично обозначенным в данной области как полипептид или белок). Данный термин охватывает, среди прочих, природные полипептиды, модифицированные полипептиды (также именуемые производные, аналоги, варианты или мутанты), фрагменты полипептидов, мультимеры полипептидов (например, димеры), рекомбинантные полипептиды, слитые полипептиды.
В контексте настоящего изобретения термины «нуклеиновая кислота», «молекула нуклеиновой кислоты», «полинуклеотид», «последовательность нуклеиновой кислоты» и «нуклеотидная последовательность» используются взаимозаменяемо и определяют полимер из по меньшей мере 9 нуклеотидных остатков либо в дезоксирибонуклеиновой кислоте (ДНК), либо в рибонуклеиновой кислоте (РНК), либо в смешанных полирибо-полидезоксирибонуклеотидах. Данные термины охватывают их одно- или двухцепочечные, линейные или кольцевые, природные или синтетические, немодифицированные или модифицированные версии (например, генетически модифицированные полинуклеотиды; оптимизированные полинуклеотиды), смысловые или антисмысловые полинуклеотиды, химерные смеси (например, гибриды РНК-ДНК). Типичные нуклеиновые кислоты ДНК включают, без ограничения, комплементарную ДНК (кДНК), геномную ДНК, плазмидную ДНК, векторы, вирусные ДНК (например, вирусные геномы, вирусные векторы), олигонуклеотиды, зонды, праймеры, кодирующую ДНК, некодирующую ДНК или ее любой фрагмент и т.д. Типичные нуклеиновые кислоты РНК включают, без ограничения, матричную РНК (мРНК), предшественник матричной РНК (пре-мРНК), кодирующую РНК, некодирующую РНК и т.д. Последовательности нуклеиновых кислот, описанных в данном документе, могут быть синтезированы стандартными способами, известными в данной области, например, посредством применения автоматического синтезатора ДНК (такого как синтезаторы, которые имеются в продаже у Biosearch, Applied Biosystems и т.д.), или получены из встречающегося в природе источника (например, геном, кДНК и т.д.), или искусственного источника (такого как имеющаяся в продаже библиотека, плазмида и т.д.) с использованием методик молекулярной биологии, хорошо известных в данной области (например, клонирование, ПЦР и т.д.).
В пределах контекста настоящего изобретения процент идентичности определяется на основе оптимального глобального выравнивания последовательностей, подлежащих сравнению, т.е. оптимального выравнивания последовательностей, взятых целиком по их полной длине с использованием любого алгоритма, хорошо известного специалисту в данной области, такого как алгоритм Needleman и Wunsch (1970). Это сравнение последовательностей может проводиться с использованием любой программы, хорошо известной специалисту в данной области, например, программы Needle, с использованием параметра «открыть пробел», равного 10,0, и параметра «удлинить пробел», равного 0,5, матрицы «Blosum 62», параметра «штраф за концевой пробел», соответствующего «неправильный», параметра «открыть концевой пробел», равного 10, и параметра «удлинить концевой пробел», равного 0,5. Программа Needle, например, доступна на https://www.ebi.ac.uk/Tools/psa/emboss_needle/.
Термины «вирус», «вирусная частица», «вирусный вектор» и «вирион» используются взаимозаменяемо, и их следует понимать широко, как означающие носитель, содержащий по меньшей мере один элемент вирусного генома дикого типа, который может быть упакован в вирусную частицу. Данный термин охватывает вирусный геном и вирусные частицы.
Термин «полученный из», «происходящий» или «происходить» используется для идентификации исходного источника компонента (например, неоэпитопа, неопептида, неоантигена, молекулы нуклеиновой кислоты, вируса и т.д.) или исходного источника образца (например, субъекта или группы субъектов), но подразумевается, что он не ограничивает способ, посредством которого получается компонент/образец, который может представлять собой, например, химический синтез или способы генной инженерии.
Термин «выделенный» в том виде, в котором он используется в данном документе, относится к компоненту (например, полипептиду, молекуле нуклеиновой кислоты, вектору и т.д.), который удаляется из его природного окружения (т.е., отделяется от по меньшей мере одного другого компонента, с которым он ассоциирован в природе или обнаруживается в природе). Более конкретно, он относится к компоненту, который очищается (частично или существенно). Например, молекула нуклеиновой кислоты является выделенной, когда она отделяется от последовательностей, обычно ассоциированных с ней в природе (например, отделяется от хромосомы или генома), но она может быть ассоциирована с гетерологичными последовательностями (например, в пределах рекомбинантного вектора). Синтетический компонент является выделенным по природе.
Термин «субъект», в общем, относится к позвоночному организму, для которого нужен или может быть полезен любой продукт или способы, раскрытые в данном изобретении. Типично организм представляет собой млекопитающее, в частности млекопитающее, выбранное из группы, состоящей из домашних животных, сельскохозяйственных животных, спортивных животных и приматов (человекообразных и нечеловекообразных). Термины «субъект» и «пациент», при отнесении к человеческому организму, могут использоваться взаимозаменяемо и охватывают мужчин и женщин, а также плоды, новорожденного, ребенка в возрасте до 2 лет, молодого взрослого, взрослого и пожилого.
Термин «опухоль» в том виде, в котором он используется в данном документе, может использоваться взаимозаменяемо с любым из терминов «рак», «злокачественное заболевание», «неоплазма» и охватывает любое заболевание или патологическое состояние, возникающее из-за неконтролируемого роста и распространения клеток. Подразумевается то, что данные термины включают любой тип ткани, органа или клетки, любую стадию злокачественного заболевания (например, от предпоражения до стадии IV). Типично опухоли, особенно злокачественные опухоли, демонстрируют частичное или полное отсутствие структурной организации и функциональной координации по сравнению с нормальной тканью, и обычно демонстрируют склонность ко вторжению в окружающие ткани (распространение) и/или к метастазированию до отдаленных сайтов. Настоящее изобретение предпочтительно разрабатывается для лечения солидных опухолей, как описывается в данном документе.
Термины «неопластическая клетка», «раковая клетка» или «опухолевая клетка» можно использовать взаимозаменяемо для названия клетки, которая делится с ненормальной (т.е. повышенной) скоростью.
Термин «лечение» (и любая форма лечения, такая как «осуществлять лечение», «лечить» и т.д.) в том виде, в котором он используется в данном документе, относится к профилактике и/или терапии. Типично «профилактика» относится к предупреждению, например, для предупреждения, задержки начала или уменьшения тяжести первого появления или рецидива по меньшей мере одного клинического или биохимического симптома (размер опухоли, уровень экспрессии ассоциированного биомаркера, прогрессирование стадии), тогда как терапия относится к патологическому состоянию с целью улучшения по меньшей мере одного клинического или биохимического симптома (размер опухоли, уровень экспрессии ассоциированного биомаркера) для замедления или контроля прогрессирования целевого патологического состояния, его симптома(мов) или вторичного состояния по отношению к патологическому состоянию у субъекта, которого лечили согласно настоящему изобретению.
Термин «осуществление введения» (или любая форма термина «введение», такая как «введенный» и т.д.) в том виде, в котором он используется в данном документе, относится к доставке субъекту компонента (например, по меньшей мере поксвируса, кодирующего неопептид) согласно способам, описанным в данном документе.
Персонализированная противораковая вакцина
В первом аспекте настоящее изобретение относится к персонализированной противораковой вакцине, содержащий рекомбинантный поксвирус, кодирующий один или более чем один неопептид.
Термин «персонализированная» в том виде, в котором он применяется в данном документе для характеристики противораковой вакцины по изобретению, относится либо к уровню индивида (конкретного субъекта), либо к уровню подпопуляции (малой группы людей, имеющих общую характеристику, например, имеющих конкретное заболевание, конкретную фенотипическую характеристику, или принимающих одинаковое лекарственное средство, или демонстрирующих одинаковую недостаточность, например, в иммунной системе).
Поксвирус
Термин «поксвирус» в том виде, в котором он используется в данном документе, относится к вирусу, принадлежащему к семейству Poxviridae, с предпочтением к подсемейству Chordopoxvirinae, направленному на хозяина-позвоночного, которое включает несколько родов, таких как Orthopoxvirus, Capri poxvirus, Avipoxvirus, Para poxvirus, Leporipoxvirus и Suipoxvirus. Предпочтительными в контексте настоящего изобретения являются ортопоксвирусы, а также авипоксвирусы, включая вирус оспы канареек (например, ALVAC) и вирус оспы кур (например, вектор FP9). В предпочтительном воплощении противораковая вакцина содержит поксвирусный вектор, принадлежащий к роду Orthopoxvirus, и даже более предпочтительно к виду вируса осповакцины (VV). В контексте настоящего изобретения можно использовать любой штамм вируса осповакцины, включающий, без ограничения, штаммы Western Reserve (WR), Copenhagen(Cop), Lister, LIVP, Wyeth, Tashkent, Tian Tan, Brighton, Ankara, MVA (модифицированный вирус осповакцины Ankara), LC16M8, LC16M0 и т.д., с конкретным предпочтением в отношении вирусов осповакцины WR, Copenhagen, Wyeth и MVA. Последовательности генома разных Poxviridae доступны в данной области в специализированных банках данных, таких как Genbank (например, номера доступа NC_006998, М35027, NC_005309, U94848 дают последовательности геномов WR, Copenhagen, Canarypoxvirus и MVA).
Другое подходящее воплощение направлено на противораковую вакцину, содержащую поксвирусный вектор, принадлежащий к роду Parapoxvirus. Подобно другим членам семейства Poxviridae, парапоксвирусы представляют собой относительно большие, оболочечные, двухцепочечные ДНК-вирусы с яйцевидными геометриями, которые могут инфицировать позвоночных, включая широкий набор млекопитающих и человека. Парапоксовирусы имеют уникальную спиральную оболочку, которая отличает их от других поксвирусов. Такой род охватывает ряд разных видов, включающих Parapoxvirus ovis (ORFV), вирус псевдооспы коров (PCPV), вирус папулезного стоматита коров и их разные штаммы (например, штаммы ORFV 01701, NZ2, NZ7 и OV-SA00, и штамм BV-AR02 вируса папулезного стоматита коров), которые могут иметь морфологические, структурные и/или генетические отличия друг от друга. В контексте настоящего изобретения предпочтение отдается видам PCPV. PCPV обладает геномом, который представляет собой линейный и двухцепочечный отрезок ДНК, типично размером 130-150 т.п.о.
В контексте настоящего изобретения можно использовать либо штамм дикого типа, либо его любое производное (т.е. поксвирус, который модифицируется по сравнению со штаммом дикого типа, например, посредством усечения, делеции, замены и/или вставки одного или более чем одного нуклеотида, смежного или несмежного в пределах вирусного генома). Модификация(ции) может находиться в пределах эндогенных вирусных генов (например, кодирующей и/или регуляторной последовательностей) и/или в пределах межгенных областей. Кроме того, модификация(ции) может(гут) быть молчащей(щими) или немолчащей(щими) (например, приводящей к модифицированному вирусному генному продукту). Модификацию(ции) можно делать целым рядом способов, известных специалистам в данной области, с использованием традиционных методик молекулярной биологии. Настоящее изобретение включает онколитические (например, генетически модифицированные для лучшей или селективной репликации в опухолевых клетках), а также дефектные по репликации поксвирусы.
Предпочтительно модификации, охватываемые настоящим изобретением, влияют, например, на вирулентность, токсичность, патогенность или репликацию вируса по сравнению с вирусом без таких модификаций. Типичные модификации, нацеленные на изменение вирусных генов, предпочтительно участвуют в метаболизме ДНК, вирулентности у хозяина или пути IFN (см., например, Guse et al., 2011, Expert Opinion Biol. Ther.11(5):595-608). Особенно подходящим геном, подлежащим разрушению, является ген, кодирующий тимидинкиназу (ТК) (локус J2R; номер доступа Genbank ААА48082). Фермент ТК участвует в синтезе дезоксирибонуклеотидов. ТК необходим для вирусной репликации в нормальных клетках, так как данные клетки обычно имеют низкую концентрацию нуклеотидов, тогда как он является необязательным в делящихся клетках, которые содержат высокую концентрацию нуклеотидов. Кроме того, известно то, что инактивация гена ТК увеличивает селективность к опухолевым клеткам. Альтернативно или в комбинации с, поксвирус для применения в данном докумете может быть модифицирован изменением по меньшей мере одного гена или обоих генов, кодирующих вирусную рибонуклеотидредуктазу. Данный вирусный фермент является аналогичным по субъединичной структуре ферменту млекопитающего, будучи состоящим из двух гетерологичных субъединиц, обозначенных R1 и R2, кодируемых, соответственно, локусом I4L и F4L. Последовательности генов I4L и F4L, и их положения в геноме разных поксвирусов доступны в публичных базах данных. Другие подходящие модификации включают модификации, изменяющие дезоксиуридинтрифосфатазу (F2L), вирусный гемагглютинин (A56R); ингибитор сериновой протеазы (B13R/B14R) и белок, связывающий комплемент 4b (C3L). Номенклатура генов, используемая в данном документе, представляет собой номенклатуру штамма вируса осповакцины Copenhagen. Она также используется в данном документе для гомологичных генов других представителей Poxviridae, если не указано иначе, и соответствие между Copenhagen и другими штаммами вируса осповакцины доступно специалисту. Для иллюстративных целей вирусы осповакцины (VV), дефектные в отношении ТК, ТК- и F2L, и ТК- и I4L, описываются в литературе (см., например, WO 2009/065547 и WO 2009/065546).
В предпочтительном воплощении поксвирус, содержащийся в противораковой вакцине по изобретению, представляет собой дефектный по репликации поксвирус и, предпочтительно, дефектный по репликации вирус осповакцины, что означает то, что он не может реплицироваться в какой-либо значимой степени в человеческих клетках. Поксвирусный вектор может быть сделан дефектным по репликации посредством частичной или полной делеции или инактивации критических для вирусной репликации областей, и ухудшение или дефектность функций репликации может оцениваться традиционными способами, как, например, посредством измерения синтеза ДНК и/или вирусного титра в непермиссивных клетках.
Особенно подходящим поксвирусным вектором для применения в контексте настоящего изобретения является MVA из-за его сильно ослабленного фенотипа (Mayr et al., 1975, Infection 3: 6-14; Sutter and Moss, 1992, Proc. Natl. Acad. Sci. USA 89: 10847-51) и более отчетливого ответа IFN-типа 1, генерируемого при инфекции, по сравнению с неослабленным поксвирусом. Для иллюстративных целей MVA был генерирован через серийные пассажи в фибробластах эмбрионов курицы. Анализ последовательности его генома показал то, что он потерял патогенность его родительского вируса - вируса осповакцины Ankara хориоаллантоиса - через изменения его генома. (Antoine et al., 1998, Virol. 244: 365-96 и номер доступа Genbank U94848). MVA безопасно и эффективно использовали для вакцинации против натуральной оспы у более чем сотни тысяч индивидов. Репликативный потенциал данного вируса в человеческих клетках также является дефектным, но не в клетках эмбрионов курицы. В данной области доступны разные клеточные системы для производства больших количеств вируса, а именно в способах изготовления на основе яиц (например, WO 2007/147528).
Термин «рекомбинантный» относится к поксвирусу, содержащему в виде вставки в его геном по меньшей мере одну экзогенную молекулу нуклеиновой кислоты (например, одну или более чем одну молекулу нуклеиновой кислоты, кодирующую неопептиды), как описано далее.
Тип и число неопептидов
Ради ясности, термин «антиген», в общем, относится к веществу (например, полипептиду), способному индуцировать гуморальный или специфичный ответ Т-клеток (или и тот, и другой) против антигена, включающий ответ Т-клеток CD4+ (например, Th1, Th2 и/или Th17) и/или CD8+ (например, ответ CTL (цитотоксические Т-лимфоциты)). В данной области доступно огромное разнообразие прямых или опосредованных биологических анализов для оценки иммуногенной природы антигена либо in vivo (животное или человек), либо in vitro (например, в биологическом образце), как описано в данном документе.
Для того чтобы быть выявляемыми иммунной системой, антигены подвергаются процессингу, и для данного процессинга требуется фрагментация антигена на пептиды, ассоциация пептидов с МНС (главный комплекс гистосовместимости; HLA - «человеческий лейкоцитарный антиген» у человека) и презентация комплекса пептид-МНС на поверхности клетки, где они могут распознаваться Т-клеткой через рецептор Т-клетки (TCR). Данные пептиды, происходящие из антигенов, включают минимальную иммунную детерминанту (т.е. эпитоп). Имеются два типа молекул МНС: МНС класса I и МНС класса II. Молекулы МНС класса I присутствуют на поверхности всех ядерных клеток млекопитающих субъектов, и комплексы пептид/МНС класса I запускают антивацию CTL CD8+. В отличие от этого, молекулы МНС класса II обычно присутствуют только на поверхности специализированных антигенпрезентирующих клеток, таких как дендритные клетки, одноядерные фагоциты, некоторые эндотелиальные клетки, эпителиальные клетки тимуса и В-клетки, и презентация пептида через молекулы МНС класса II обычно управляет ответом Т-клеток CD4+. Эпитопы для презентации посредством молекул МНС класса I обычно имеют длину по меньшей мере 8 аминокислот, предпочтительно 8, 9 или 10 аминокислот, тогда как эпитопы МНС класса II обычно длиннее (например, по меньшей мере 13 аминокислотных остатков (Rock et al., 2010, J. Immunol. 184(1): 9-15). В данной области существует большое число алгоритмов прогнозирования in silico связывания МНС класса I и класса II (см., например, Nielsen et al., 2010, Immunology 130(3): 319-28). Для иллюстративных целей можно процитировать SVMHC, NetMHCII, Tepitope/propped, syfpeithi, Epitollkit и т.д.
Термин «неоантиген» в том виде, в котором он используется в данном документе, относится к антигену, который возник во время процесса канцерогенеза в раковой клетке. В предпочтительном воплощении неоантиген содержит одну или более чем одну немолчащую мутацию аминокислотного(ных) остатка(тков) относительно соответствующего антигена дикого типа. Типично немолчащая мутация происходит на уровне нуклеотидов и транслируется в изменение на аминокислотном уровне. В другом предпочтительном воплощении неоантиген обнаруживается в раковых клетках или тканях, полученных от пациента, но не обнаруживается в образце нормальных клеток или тканей, полученных от пациента или здорового индивида.
Термин «неопептид» относится к фрагменту неоантигена, содержащему неоэпитоп, а также к фланкирующей(щим) последовательности(тям) на одной или обеих сторонах данного неоэпитопа. Фланкирующие последовательности представляют собой последовательности неоантигена, из которого взят неоэпитоп. Неопептид, таким образом, соответствует фрагменту неоантигена, который длиннее, чем сам неоэпитоп, который презентируется молекулами МНС. В неопептиде, таким образом, присутствует неоэпитоп, и он окружен (на одной из двух или на обеих сторонах фланкирующей(щими) последовательностью(тями), присутствующими в природе в его нормальном окружении. За исключением редких случаев, когда неопептид также присутствует в другом собственном белке, в большинстве случаев неопептид имеет несобственную природу (и может быть отобран как имеющий несобственную природу, см. ниже). Из-за этой несобственной природы ожидается то, что такой(кие) неопептид(ды) будет распознаваться опухолеспецифичными Т-лимфоцитами.
Термин «неоэпитоп» в том виде, в котором он используется в данном документе, относится к минимальной иммунной детерминанте неоантигена, которая способствует МНС-зависимому распознаванию Т-клетками (или который презентируется молекулами МНС на поврхности клеток субъекта) и содержит по меньшей мере одну немолчащую мутацию, идентифицированную в указанном неоантигене согласно способам, описанным в данном документе.
Термин «мутация» относится к по меньшей мере одному различию последовательности тестируемой последовательности и контрольной последовательности.
В одном воплощении каждый из одного или более чем одного неопептида, кодируемого реомбинантным поксвирусом, является специфичным для опухоли пациента и содержит по меньшей мере одну опухолеспецифичную мутацию из набора мутаций, описанных в данном документе. В контексте настоящего изобретения термин «опухолеспецифичная мутация» относится к мутации, которая генерируются вместе с неопластическим превращением и/или прогрессированием раковой клетки. Типично опухолеспецифичная мутация предпочтительно присутствует в ДНК, содержащейся в раковой клетке (например, в образце опухоли), но отсутствует в ДНК, содержащейся в нераковой клетке (например, в неопухолевом образце). В контексте настоящего изобретения опухолеспецифичная мутация, хотя и происходит на уровне нуклеотидов, не является молчащей и транслируется в изменение на аминокислотном уровне.
Настоящим изобретением охватываются несколько типов опухолеспецифичных мутаций, включающих миссенс-мутации, делеции, вставки, мутации со сдвигом рамки считывания и мутации в сайте сплайсинга. «Миссенс-мутация» возникает в результате замены одного нуклеоида другим нуклеотидом в пределах специфического кодона, которая влияет на закодированную аминокислотную последовательность, таким образом, приводя к замене одной аминокислоты. Другим способом влияния на последовательность белка является вставка или делеция одного или более чем одного нуклеотида (например, куска ДНК), которая изменяет число нуклеотидов в молекуле нуклеиновой кислоты. Мутации в виде вставки и делеции могут приводить к изменению рамки считывания (так называемая мутация со сдвигом рамки считывания), но не обязательно (например, вставка или делеция 3 нуклеотидов будет приводить к добавлению или устранению кодона). Почти весь полипетид может быть изменен, если данная мутация происходит рано в нуклеотидной последовательности. Мутация со сдвигом рамки считывания также может приводить к образованию стоп-кодона, который транслируется в стоп-сигнал, и образующийся белок будет тогда усеченным (подвергнутым делеции его части, находящейся ниже стоп-кодона, генерированного de novo). Миссенс-мутация также может располагаться в сайтах сплайсинга мРНК, приводя к непоавильному сплайсингу и, следовательно, к нарушенной последовательности белка. Термин «сплайсинг» относится к редактированию транскрипта растущего предшественника матричной РНК (пре- мРНК). После сплайсинга удаляются интроны, а экзоны соединяются друг с другом (лигируются). Термин «сайт сплайсинга» относится к коротким консервативным последовательностям, расположенным на концах интронов, которые являются решающими для распознавания интрона и для точности реакций сплайсинга.
Настоящее изобретение охватывает неопептиды, возникающие из всех классов белков, таких как ферменты, рецепторы, транскрипционные факторы и т.д. Хотя и может присутствовать любой тип мутаций, однако, опухолеспецифичная мутация предпочтительно представляет собой миссен-мутацию или мутацию со сдвигом рамки считывания. Преимущественно, по меньшей мере 60%, желательно по меньшей мере 65%, предпочтительно по меньшей мере 70%, более предпочтительно по меньшей мере 75%, по меньшей мере 80% (например, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91% и т.д.), по меньшей мере 85% (например, 85%, 86%, 87%, 88%, 89%, 90%, 91% и т.д.), по меньшей мере 90% (например, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или даже 100%), по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или даже 100% неопептидов для применения в данном документе содержат миссенс-мутацию или мутацию со сдвигом рамки считывания. Среди миссенс-мутаций и мутаций со сдвигом рамки считывания миссенс-мутаций являются более частыми. В некоторых воплощениях по меньшей мере 60%, желательно по меньшей мере 65%, предпочтительно по меньшей мере 70%, более предпочтительно по меньшей мере 75% и даже более предпочтительно по меньшей мере 80% (например, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91% и т.д.), по меньшей мере 85% (например, 85%, 86%, 87%, 88%, 89%, 90%, 91% и т.д.), по меньшей мере 90% (например, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или даже 100%), по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или даже 100% неопептидов для применения в данном документе содержат миссенс-мутацию. Однако при идентификации мутаций со сдвигом рамки считывания также является предпочтительным, чтобы один или более чем один неопептид, кодируемый рекомбинантным поксвирусом, содержал мутацию со сдвигом рамки считывания. Например, для 10 неопептидов, кодируемых рекомбинантным поксвирусом, предпочтительно по меньшей мере 6, по меньшей мере 7, по меньшей мере 8, по меньшей мере 9 или даже 10 неопептидов имеют миссенс-мутаций или мутации со сдвигом рамки считывания, и более предпочтительно по меньшей мере 6, по меньшей мере 7, по меньшей мере 8, по меньшей мере 9 или даже 10 неопептидов имеют миссенс-мутаций. Аналогичным образом, для 30 неопептидов, кодируемых рекомбинантным поксвирусом, предпочтительно по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26, по меньшей мере 27, по меньшей мере 28, по меньшей мере 29 или даже 30 неопептидов имеют миссенс-мутаций или мутации со сдвигом рамки считывания, и более предпочтительно по меньшей мере 18, по меньшей мере 19, по меньшей мере 20, по меньшей мере 21, по меньшей мере 22, по меньшей мере 23, по меньшей мере 24, по меньшей мере 25, по меньшей мере 26, по меньшей мере 27, по меньшей мере 28, по меньшей мере 29 или даже 30 неопептидов имеют миссенс-мутаций. Специалисты в данной области легко адаптируют данные примеры к поксвирусам, кодирующим другое число неопептидов.
Длина неопептидов, кодируемых рекомбинантным поксвирусом, обычно составляет от 13 аминокислотных остатков до примерно 151 аминокислотного остатка, но, естественно, данная длина может варьировать от одного неопетида до другого. Преимущественно каждый из неопептида(дов) для применения в данном документе имеет длину от 15 до 101 аминокислотного остатка, желательно от 16 до 90 аминокислотных остатков, предпочтительно от 17 до 85 аминокислотных остатков и более предпочтительно от 18 до 80 аминокислотных остатков. Преимущественно по меньшей мере 70% неопепидов для применения в данном документе имеют нечетное число аминокислот, так как, в случае одной миссенс-мутаций, это обеспечивает вставку в центре неопептида с одинаковым числом фланкирующих аминокислот на одной из двух сторон мутировавшей аминокислоты. Естественно, длина неопептида также может зависеть от типа мутации. Для иллюстративных, но не ограничивающих целей неопептиды, содержащие миссенс-мутацию, предпочтительно имеют длину от 18 до 29 остатков, тогда как неопептиды, содержащие мутацию со сдвигом рамки считывания, имеют длину от 30 до 80 аминокислотных остатков. В контексте настоящего изобретения по меньшей мере 60%, предпочтительно по меньшей мере 70% и более предпочтительно по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или даже 100% неопептидов, кодируемых противораковой вакциной на основе поксвируса по изобретению, предпочтительно имеют длину от 18 до 29 остатков. Особое предпочтение дается индивидуальным неопептидам из 19, 21, 26, 27 или 29 остатков. В особенно предпочтительном воплощении по меньшей мере 60%, предпочтительно по меньшей мере 70% и более предпочтительно по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или даже 100% неопептидов, кодируемых противораковой вакциной на основе поксвируса по изобретению, содержат миссенс-мутацию и имеют длину от 18 до 29 остатков, в частности, 19, 21, 26, 27 или 29 остатков.
Относительно положения мутации, в настоящем изобретении рассматривается любое положение в пределах неопептида. Центральное положение, однако, является особенно предпочтительным для миссенс-мутаций. Под «центральным положением» подразумевается то, что мутировавшая аминокислота располагается точно в середине неопептида (в случае, когда неопептид имеет нечетное число аминокислот) или в одном из двух центральных положений (в случае, когда неопептид имеет четное число аминокислот), или в любой из 2-5 аминокислот на каждой стороне от точно центрального положения, в зависимости от длины неопептида. Желательно по меньшей мере 80%, по меньшей мере 85%, более предпочтительно по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или даже 100% неопептидов, имеющих миссенс-мутацию, несут замененную аминокислоту в центральном положении, как определено выше. Особенно предпочтительными являются неопептиды из 19, 21, 27 или 29 аминокислот (даже более предпочтительно из 27 или 29 аминокислот), несущие миссенс-мутацию точно в центральном положении (например, в положении 10 для 19-мерного неопептида, в положении 11 для 21-мерного неопептида, в положении 14 для 27-мерного неопептида и в положении 15 для 29-мерного неопептида).
Данная мутация, однако, может, по меньшей мере в некоторых из неопептидов, располагаться близко к N-концу или С-концу, особенно для мутации со сдвигом рамки считывания, или при появлении миссенс-мутаций на или близко к N- или С-концу неоантигена.
В некоторых воплощениях также рассматривается экспрессия неопептидов рекомбинантным поксвирусом в виде одного или более чем одного слияния.
Термин «слияние» в том виде, в котором он используеся в данном документе, относится к комбинации двух или более чем двух неопептидов в одной полипептидной цепи. Данное слияние может быть прямым (т.е. без каких-либо дополнительных аминокислотных остатков между) или через линкер для улучшения доступности неопепидов. Помимо серии уникальных неопептидов, экспрессируемых индивидуально, настоящее изобретение, таким образом, также охватывает рекомбинантный поксвирус, экспрессирующий несколько неопептидов в виде слияния(ний) и некоторые другие индивидуально. В качестве альтернативы, все неопептиды, экспрессируемые поксвирусным вектором, крастеризуются в одном или более чем одном слиянии.
В некоторых воплощениях рассматривается присутствие сигнального пептида на N-конце закодированного(ных) неоэпитопа(пов) (или его слияния) для усиления процессинга посредством ЭР (эндоплазматический ретикулум) и/или секреции. Вкратце, сигнальные пептиды обычно содержат от 15 до 35 по существу гидрофобных аминокислот, вставляются на N-конце полипептида ниже кодона инициации трансляции и затем удаляются специфичной локализованной в ЭР эндопептидазой с получением зрелого полипептида. Заякоривание в мембране также может использоваться для улучшения презентации закодированных неопептидов (или их слияния) МНС класса I и/или МНС класса II посредством дополнительного включения последовательности заякоривания в мембране. Трансмембранные пептиды являются высокогидрофобными по природе и служат для заякоривания полипептидов в клеточной мембране. Они предпочтительно вставляются в С-концевую часть полипептида, предпочтительно непосредственно выше стоп-кодона. Подходящие трансмембранные и/или сигнальные пептиды известны в данной области. Они могут быть получены из клеточных или вирусных полипептидов, таких как полипепиды иммуноглобулинов, тканевой активатор плазминогена, инсулин, гликопротеин вируса бешенства, гликопротеин оболочки вируса ВИЧ (вирус иммунодефицита человека) или белок F вируса кори, или могут быть синтетическими. Предпочтительные сигнальные пептиды и заякоренные в мембране последовательности описываются в WO 2008/138649, с предпочтением для пептидов и последовательностей, происходящих из вируса бешенства или гликопротеина F вируса кори. Если в рекомбинантном поксвирусе подлежит применению больше чем один сигнальный пептид и/или трансмембранная последовательность, можно выбрать сигнальные пептиды и/или трансмембранные последовательности разного происхождения или выродить гомологичные последовательности, которые демонстрируют высокую степень идентичности последовательности (например, выше 75%) таким образом, чтобы ограничить события гомологичной рекомбинации, которые могут ухудшать процесс продукции.
Альтернативно или в комбинации с предыдущим воплощением, в настоящем изобретении рассматривается присутствие одного или более чем одного линкера (также именуемого спейсер), особенно в слиянии неопептидов. Типично линкеры представляют собой пептиды длиной 1-30 аминокислот, состоящие из таких аминокислотных остатков, как глицин, серии, треонин, аспарагин, аланин и/или пролин. Предпочтительные линкеры в контексте данного изобретения содержат от 2 до 15 аминокислот, с предпочтением в отношении 3, 5 или 10 аминокислот, главным образом, глицина и серина (например, GSG, GSGSG (SEQ ID NO: 66), SGSGS (SEQ ID NO: 67), GSGSGSGSGS (SEQ ID NO: 68)) или глицина, серина и треонина (например, GSTSG (SEQ ID NO: 69), SGTGS (SEQ ID NO: 70)), или глицина, серина и треонина и/или аланина (например, GAS, GTS). А именно, предпочтительные линкеры из 3 аминокислот включают линкеры последовательностей GSG, GAS и GTS. Оценка необходимости включения или невключения линкера между двумя слитыми неопептидами находится в пределах квалификации специалиста.
В предпочтительном воплощении неопептиды организуются в слиянии(ях) с линкерами между каждым неопептидом (например, между неопептидом 1 и неопептидом 2, между неопептидом 2 и неопептидом 3, и т.д.) и, возможно, на N-конце первого неопептида и/или на С-конце последнего неопептида. Когда в пределах одной конструкции подлежит применению несколько линкеров, можно варьировать аминокислотную последовательность или последовательность кодонов нуклеиновой кислоты для уменьшения процентной доли идентичности на уровне нуклеиновой кислоты (например, до меньше, чем 77% идентичности, желательно меньше, чем 75%, меньше, чем 70%, предпочтительно меньше, чем 60%, меньше, чем 50% и даже более предпочтительно меньше, чем 35%, особенно для линкеров из 10 аминокислот) и, таким образом, ограничения событий рекомбинации во время процесса продукции. В предпочтительных воплощениях последовательности нуклеиновой кислоты линкеров в одном рекомбинантном поксвирусе конструируются (посредством изменения аминокислотной последовательности или последовательности кодонов нуклеиновой кислоты) таким образом, что идентичные части последовательности нуклеиновой кислоты между любыми двумя из линкеров уменьшаются до максимум 8 смежных нуклеотидов (например, предпочтительно максимум 7 и более предпочтительно максимум 6 смежных нуклеотидов), таким образом, способствуя уменьшению идентичности последовательности нуклеиновой кислоты, как описано выше, и, таким образом, ограничению нежелательных событий рекомбинации во время процесса продукции. Как проиллюстрировано в разделе примеров, короткие линкеры из 2-5 аминокислотных остатков (например, 2, 3, 4 или 5 аминокислот) имеют преимущество, особенно когда рекомбинантный поксвирус кодирует 10 или более чем 10 неопептидов. Три аминокислотных линкера, таких как GSG, GAS и/или GTS, являются особенно предпочтительными в контексте данного изобретения для отделения неопептидов друг от друга. Типичные нуклеотидные последовательности, кодирующие 3 аминокислотных линкера, приведенные на Фиг. 7, подлежат применению индивидуально или в комбинации в слиянии(ниях) неопептидов, как описано в данном документе. Для иллюстративной цели получили 15 нуклеотидных последовательностей, кодирующих GSG, и сделали вырожденными, используя преимущество вырожденности кодонов (для G возможны 4 кодона и 6 - для S), для уменьшения риска гомологичной рекомбинации в рекомбинантном поксвирусе, которая может происходить между отрезками идентичных или сходных последовательностей. В то же самое время 10 и 5 вырожденных нуклеотидных последовательностей, кодирующих линкеры GTS и GAS, соответственно, приводятся на Фиг. 7.
В определенных воплощениях настоящего изобретения рассматривается присутствие метки для того, чтобы облегчать выявление экспрессии неопептидов (или их слияния(ний)) или инфицированных клеток-хозяев, экспрессирующих такие неопептиды или их слияние. Пептиды метки могут быть выявлены анализами иммуновыявления с использованием антител против метки. В контексте данного изобретения можно использовать широкий спектр пептидов меток, включая, без ограничения, метку РК, метку FLAG (DYKDDDK; SEQ ID NO: 6), метку МYС (EQKLISEEDL SEQ ID NO: 11), полигистидиновую метку (обычно отрезок из 5-10 остатков гистидина), метку НА (YPYDVPDYA; SEQ ID NO: 18), метку HSV (QPELAPEDPED; SEQ ID NO: 19) и метку VSV (YTDIEMNRLGK; SEQ ID NO: 20). Пептид метки может быть независимо расположен на N-конце неопептида или его слияния (метка-полипептид), или, в качестве альтернативы, на его С-конце (полипептид-метка), или, в качестве альтернативы, внутренне.
Число неопептидов, которые могут кодироваться рекомбинантным поксвирусом, не ограничивается, в зависимости от типа выбранной экспрессии (индивидуальная экспрессия, экспрессия в коротких или длинных слияниях, как описывается далее) и типа поксвируса. Для иллюстративных целей рекомбинантным поксвирусом может экспрессироваться от 1 до 50 неопептидов, более предпочтительно от 5 до 45 и даже более предпочтительно от 6 до 35, с предпочтением в отношении 10-30 неопептидов.
В предпочтительных воплощениях несколько и предпочтительно все неопептиды экспрессируются рекомбинантным поксвирусом в виде одного или более чем одного слияния. Предпочтительно поксвирус, содержащийся в персонализированной противораковой вакцине по изобретению, кодирует от 1 до 5 слияний 2 или более чем 2 неопептидов, предпочтительно от 1 до 4 слияний 2 или более чем 2 неопептидов и более предпочтительно от 1 до 3 слияний 2 или более чем 2 неопептидов, с особым предпочтением в отношении 2 или 3 слияний 2 или более чем 2 неопептидов. Более предпочтительно каждое слияние содержит от 2 до 15 неопептидов, предпочтительно от 3 до 12 неопептидов, более предпочтительно от 4 до 11 неопептидов и даже более предпочтительно от 5 до 10 неопептидов (например, 5, 6, 7, 8, 9 или 10). Число неопептидов может варьировать от одного слияния до другого слияния, кодируемого рекомбинантным поксвирусом. Однако в особенно предпочтительном воплощении поксвирус, содержащийся в персонализированной противораковой вакцине по изобретению, кодирует 2 или 3 слияния 5-10 неопептидов.
В контексте настоящего изобретения каждое слияние может быть сконструировано отлично от другого(гих), и они могут отличаться друг от друга присутствием и/или последовательностью, и/или числом, и/или расположением таких элементов, как пептидные сигналы, линкеры, метки и т.д. Согласно предпочтительному воплощению каждое слияние, однако, содержит а) сигнальный пептид на его N-конце, б) линкеры на N-конце первого неопептида, между каждыми неопептидами и на С-конце последнего неопептида, и в) метку на его С-конце.
Прогнозирование присутствия ТМ (трансмембранного) сегмента(тов) в неопептидах и их слиянии(ниях)
В одном воплощении аминокислотная последовательность одного или более чем одного неоэпитопа или его слияния, кодируемая рекомбинантным поксвирусом, не содержит какого-либо потенциального трансмембранного сегмента. ТМ сегмент может быть определен как представляющий собой короткую гидрофобную альфа-спираль из приблизительно 20 остатков (например, 18-30 остатков, с предпочтением в отношении 19-21 остатка). Для прогнозирования вероятности того, что данная последовательность (например, конкретный неопептид или слияние неопептидов) содержит или не содержит ТМ сегмент, можно использовать несколько средств прогнозирования, таких как ТМНММ (для Transmembrane Hidden Markov Model; Krogh et al., 2001, J. Mol. Biol. 305: 567-80) и DAS (Dense Alignment Surface). Например, алгоритм фильтра DAS-TM дает для запроса высокоточный профиль гидрофобности, от которого может быть получено расположение потенциальных трансмембранных сегментов.
В предпочтительном воплощении по меньшей мере 80% (например, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% и даже 100%) неопептидов, кодируемых поксвирусом, содержащимся в персонализированной противораковой вакцине по настоящему изобретению, не содержат потенциального(ных) ТМ сегмента(тов) (внутрипептидный ТМ). В данном контексте неопептиды, которые, как предсказывается, содержат ТМ сегмент, предпочтительно не выбираются.
Даже более предпочтительно прогнозирование присутствия ТМ сегмента также применяется к каждому слиянию неопептида. В самом деле, наблюдали то, что ТМ сегмент может быть получен соединением 2 конкретных неопептидов (внутрипептидный ТМ). В данном случае модифицирование порядка неопептидов в слиянии представляет собой опцию для устранения присутствия ТМ. Например, если ТМ сегмент возникает из-за слияния неопептида 1 на N-конце неопептида 2, обращение данных неопептидов (слияния неопептида 2 на N-конце неопептида 1) может устранять риск наличия ТМ сегмента. Другой возможностью может быть добавление подходящего линкера между 2 слитыми неопептидами.
Прогнозирование индексов гидрофобности и гидропатичности неопептидов и их слияния(ний)
В одном воплощении аминокислотная последовательность одного или более чем одного неоэпитопа или его слияния, кодируемая рекомбинантным поксвирусом, является гидрофильной по природе. Гидрофильную или гидрофобную природу данной последовательности можно легко определять посредством целого ряда способов и алгоритмов, доступных в данной области. Расчет индекса гидрофобности и/или индекса гидропатичности конкретной последовательности находится в пределах квалификации специалиста в данной области. Например, данные индексы могут быть определены с использованием способа Kyte-Doolittle (Kyte and Doolittle, 1982, J. Mol. Biol. 157: 105-32) или любого другого подходящего способа (например, Rose et al., 1993, Ann. Rev. Biomol. Struc. 22: 381-415; Kallol et al., 2003, J. Chromat. 1000: 637-55; Sweet et al., 1983, J. Mol. Biol. 171: 479-88; среди многих других) или алгоритма (например, ExPAsy Prot Scale Protein; графики гидрофобности белка, разработанные Colorado state или the World of Bioinformatics, и т.д.). Общим способом балл гидрофобности данной последовательности определяется суммой значения гидрофобности/гидрофильности каждого аминокислотного остатка, которое обобщается в следующей Таблице 1.
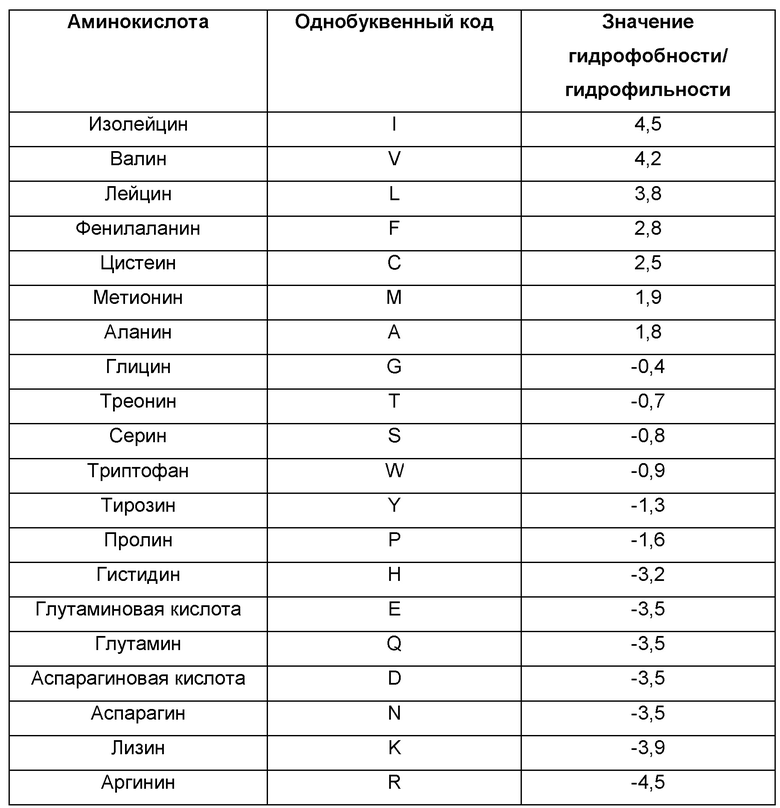
Как проиллюстрировано в Таблице 1, аминокислоты I, V, L, F, С, М и А демонстрируют положительный индекс гидрофобности (например, 4,5 для I), коррелирующий с гидрофобной природой данных остатков, тогда как G, Т, S, W, Y, Р, Н, Е, Q, D, N, K и R демонстрируют отрицательный индекс гидрофобности (например, -4,5 для R), коррелирующий с их гидрофильной природой.
Индекс гидрофобности данного неопептида определяется суммой значения гидрофобности/гидрофильности каждого аминокислотного остатка, содержащегося в указанном неопептиде, и индекс гидропатичности рассчитывается делением индекса гидрофобности, определенного для указанного неопептида, на число остатков, присутствующих в данном пептиде. Индекс гидрофобности данного слияния неопептидов соответствует сумме индексов гидрофобности, определенных для каждого неопептида, содержащегося в указанном слиянии. Индекс гидропатичности данного слияния неопептидов рассчитывается делением индекса гидрофобности, определенного для указанного слияния неопептидов, на число остатков, присутствующих в данном слиянии неопептидов.
В предпочтительном воплощении по меньшей мере 60% неопептидов, кодируемых рекомбинантным поксвирусом, содержащимся в персонализированной противораковой вакцине по настоящему изобретению, демонстрируют отрицательный индекс гидрофобности и/или индекс гидропатичности, равный или меньший 0,1 (например, 0,1; 0,09; 0,08 и т.д. или даже отрицательный индекс, например, -0,019, -1,5 и т.д.). В то же самое время, каждое слияние, кодируемое рекомбинантным поксвирусом, для применения в данном документе предпочтительно дает отрицательный индекс гидрофобности и/или равный или меньше 0,1 в отношении гидропатичности. Фраза «по меньшей мере 60%» охватывает по меньшей мере 65%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 98% и даже 100%.
Для иллюстративной цели неопептид последовательности GQSLPMTHSLKLSKTNRTLFLLGVTKY (SEQ ID NO: 58) имеет отрицательный индекс гидрофобности -3,6 (сложение -0,4 для G, -3,5 для Q, -0,8 для S,+3,8 для L, и т.д.) и индекс гидропатичности -0,13 (-3,6/29). Таким образом, данный пептид будет подходящим для экспрессии рекомбинантным поксвирусом, содержащимся в персонализированной противораковой вакцине по настоящему изобретению. В отличие от этого, неопептид GLMGIVVGTVFIIRGLRSVGASRHQGL (SEQ ID NO: 59) имеет положительный индекс гидрофобности (23,3) и имеет индекс гидропатичности 0,86. Таким образом, данный пептид не подходит для экспрессии рекомбинантным поксвирусом, содержащимся в персонализированной противораковой вакцине по настоящему изобретению, если другие неопептиды, содержащиеся в слиянии, не компенсируют обеспечение снижения общего индекса слияния до подходящего порога, описанного в данном документе.
В особенно предпочтительном воплощении рекомбинантный поксвирус, содержащийся в персонализированной противораковой вакцине по изобретению, кодирует 2 или 3 слияния, где:
• каждое слияние содержит от 5 до 10 неопептидов; имеет линкеры, предпочтительно длиной 3 аминокислоты, на N-конце первого неопептида указанного слияния и между каждым неопептидом (например, одним или более чем одним из неопептидов, показанных на Фиг. 7);
• каждое слияние содержит сигнальный пептид на N-конце указанного слияния;
• каждое слияние возможно содержит последовательность метки на С-конце указанного слияния; и
• каждое слияние:
ο демонстрирует общий отрицательный индекс гидрофобности и/или общий индекс гидропатичности, равный или меньший 0,1, и
ο не содержит какого-либо потенциального ТМ сегмента.
Идентификация неоантигенов и неопептидов, подлежащих кодированию персонализированной противораковой вакциной
Разработка персонализированной противораковой вакцины требует идентификации и селекции неопептидов на основе от пациента к пациенту.
Соответственно, настоящее изобретение относится к способу получения персонализированной противораковой вакцины, включающему стадию идентификации одного или более чем одного подходящего неопептида для кодирования рекомбинантным поксвирусом, где каждый из указанных неопептидов содержит по меньшей мере одну опухолеспецифичную мутацию.
Для данной цели от субъекта, подлежащего лечению, получают биологический образец опухоли, а также неопухолевый биологический образец. Термин «биологический образец» в том виде, в котором он используется в данном документе, относится к образцу, полученному от субъекта, который можно брать или отбирать in vivo. В контексте данного изобретения биологический(кие) образец(зцы), взятый(тые) или отобранный(ные) от субъекта, содержит(жат) ДНК и предпочтительно также содержит(жат) мРНК или белки.
В одном воплощении биологический образец получают из жидкости организма или из образца ткани. Типичные образцы жидкости организма включают кровь, сыворотку, плазму, мочу, кал, спинномозговую жидкость и бронхиальную жидкость, а также их любое производное (например, частично очищенная кровь, РВМС (одноядерные клетки периферической крови), но не ограничиваются ими. Образцы ткани могут представлять собой органы, ткани, части и клетки, выделенные из млекопитающего, из любого источника или типа клеток (например, образцы, содержащие компоненты полости рта, желудочно-кишечного тракта, кожи, легкого, головы и т.д.), но не ограничиваются ими. Типичные образцы ткани включают, например, клеточный лизат, культуры клеток, ткани, свежие замороженные ткани, цитологический материал (например, мазок из шейки матки, вагинальный смыв, бронхиальный соскоб, плевральные жидкости, колоскопию и т.д.), ткани, полученные после хирургического удаления ткани, а также биопсии тканей. В контексте настоящего изобретения биологический образец может находиться в суспензии или растворе, или все еще может быть присоединенным к твердой подложке (например, предметные стекла, шарики, чипы или наночастицы). Настоящее изобретение также охватывает «очищенные» или наполовину «очищенные» биологические образцы, например, выделенную РНК, кДНК, выделенный белок и т.д.
В одном и предпочтительном воплощении образец опухоли представляет собой биопсию опухоли, предпочтительно содержащую больше, чем 20% опухолевых клеток. Он может быть получен любым способом, включающим диссоциированные образцы тканей, как, например, аспираты тонкой иглой, или свежие солидные опухоли (которые могут быть ферментативно расщепленными), или микротомный срез биопсии, но не ограничивается ими. В другом воплощении образец опухоли представляет собой цитологический материал (например, смыв из шейки матки или влагалища).
В еще одном другом воплощении неопухолевый образец получают из биологической жидкости (например, крови с выделением РВМС или без него), цитологического материала (например, буккальный мазок), биопсии (например, биопсия здоровой ткани) или кусочка хирургической резекции, содержащего неопухолевые клетки. Данный неопухолевый образец может быть взят или отобран от того же самого пациента или от контрольного здорового субъекта, или от группы субъектов.
В случае когда образцы опухоли или неопухолевые образцы происходят в результате того же самого отбора образцов, специалист знает, как распознавать и отделять опухолевые клетки от неопухолевых клеток (например, посредством иммуногистологии).
В одном воплощении стадия идентификации включает:
а) экстрагирование ДНК из указанного образца опухоли и указанного неопухолевого образца,
б) выбор областей-мишеней, предпочтительно полных кодирующих областей генома (экзом),
в) секвенирование указанных областей-мишеней (например, экзома) из указанных экстрагированных ДНК и
г) идентификацию одной или более чем одной опухолеспецифичной мутации посредством сравнения последовательностей ДНК, полученных из указанных образцов опухоли и неопухолевых образцов.
Термин «экзом» относится к части генома, образованной экзонами (т.е. последовательностями, которые при транскрипции остаются в пределах мРНК после удаления интронов посредством сплайсинга РНК). Для иллюстративных целей экзом человеческого генома состоит из примерно 180000 экзонов, соответствующих примерно 1% всего генома. Методики секвенирования экзома хорошо известны в данной области (Anderson and Schrijver, 2010, Genes 1 (1): 38-69). В общем, способы включают двухэтапные подходы: сначала целевой захват кодирующих последовательностей с использованием гибридизации на основе чипа (имеется в продаже у Agilent technologies) или гибридизации на основе жидкости (имеется в продаже как NimbleGen от Roche; Bainbridge et al., 2010, Genome Biol. 11:R62); после стадии захвата следует стадия секвенирования (Ng S.B. et al, 2009, Nature 461: 272-6). Для секвенирования последовательностей ДНК были сделаны коммерчески доступными многие технические подходы, включая, среди прочих, пиросеквенирование (имеется в продаже как Roche 454), секвенирование с красителем (имеется в продаже посредством Illumina), цифровое секвенирование (имеется в продаже как секвенирование lonTorrent).
В то время как стадия идентификации опухолеспецифичных мутаций является обязательной, может использоваться дополнительное возможное фильтрование для отбора оптимальных неопептидов, подлежащих кодированию персонализированным рекомбинантным поксвирусом, и/или для их объединения в подходящих слияниях неопептидов. В самом деле, во время превращения и прогрессирования рака могут накапливаться многие опухолеспецифичные мутации, и некоторые могут иметь лучший потенциал для стимулирования иммунной системы субъекта, подлежащего лечению против его ракового заболевания. При использовании дополнительного фильтрования стадия идентификации предпочтительно включает, помимо подстадий а) - г), описанных выше, по меньшей мере одну из следующих дополнительных подстадий д) - и):
д) ранжирование потенциального(ных) неопептида(дов) по их уровням экспрессии в опухоли (или уровням экспрессии соответствующих неоантигенов) либо на уровне транскрипции мРНК, либо на уровне трансляции белка;
е) отбор неаутоэкспрессируемой(мых) опухолеспецифичной(ных) мутации(ций);
ж) прогнозирование иммунного потенциала неоэпитопа(пов), содержащегося(сихся) в неопептиде(дах) либо посредством прогнозирования in silico, либо посредством биологических анализов иммуногенности;
з) прогнозирование присутствия потенциального(ных) ТМ сегмента(тов) в самих неоэпитопах (внутрипептидный ТМ) и/или в слияниях неопептидов, содержащих несколько неопептидов (внутрипептидный ТМ в областях слияния, т.е. в областях, охватывающих С-конец первого неопептида и N-конец второго неопептида, и возможно линкер между первым и вторым неопептидами);
и) ранжирование потенциального неопептида(дов) и/или слияния(ний) неопептидов по степени гидрофобности и, в частности, отбор неопептида(дов) и/или слияния(ний) неопептидов таким образом, что указанный неопептид(ды) и/или слияние(ния) неопептидов демонстрирует(ют) отрицательный общий индекс гидрофобности и/или общий индекс гидропатичности, равный или меньший 0,1.
Как упомянуто выше, в стадии идентификации может присутствовать по меньшей мере одна дополнительная подстадия д) - и). Могут присутствовать несколько дополнительных подстадий д) - и). В частности, в предпочтительном воплощении стадия идентификации дополнительно включает, помимо подстадий а) - г), описанных выше, подстадий д) и е), приведенные выше (данные подстадий более подробно описываются ниже), и возможно также:
• подстадию ж) (которая также более подробно описывается ниже),
• подстадию з) (которая также более подробно описывается ниже),
• подстадию и) (которая также более подробно описывается ниже), или
• любую комбинацию подстадий ж), з) и и) (например, подстадий ж) и з), подстадий ж) и и), подстадий з) и и) или подстадий ж), з) и и)).
Подстадия д): может быть добавлено дополнительное фильтрование посредством ранжирования неоантигенов по их уровням экспрессии в опухоли. Таким образом, в предпочтительном воплощении стадия идентификации дополнительно включает подстадию д) отбора неопептидов, которые по меньшей мере экспрессируются и предпочтительно высоко экспрессируются в опухоли. Данная экспрессия может быть выявлена или количественно измерена на уровне мРНК или белка. В одном воплощении неопептиды могут быть отобраны на основе выявления того, что они по меньшей мере транскрибируются и, предпочтительно, высоко транскрибируются в опухоли, т.е. на основе экспрессируемой опухолеспецифичной мутации, которая обнаруживается в образце РНК опухоли и случается в ORF (открытая рама считывания) (при трансляции); (транскрипт RefSeq или Genbank). Для иллюстративных целей указанная(ные) экспрессируемая(мые) опухолеспецифичная(ные) мутация(ции) типично идентифицируется(ются) посредством секвенирования комплементарной ДНК (кДНК), генерированной из РНК, экстрагированной из указанного биологического образца опухоли, полученного от пациента, т.е. транскриптома. Термин «транскриптом» в том виде, в котором он используется в данном документе, относится к полному спектру матричной РНК (мРНК), экспрессируемой организмом. Методики секвенирования транскриптома хорошо известны в данной области (например, Chu and Corey, 2012, Nucleic Acid Ther. 22: 271-4; Wang et al., 2009, Nat Rev Genet. 10(1): 57-63). Предпочтительно мРНК, кодирующая мутировавший неоантиген, обильно экспрессируется в РНК, экстрагированной из образца опухоли. В то время как не возможно дать предпочтительное абсолютное значение экспрессии для отбора неопептидов, данный отбор является скорее относительным, причем экспрессия потенциальных неопептидов выявляется/количественно измеряется и ранжируется по порядку приоритета. Специалисты в данной области знают как сравнивать уровень экспрессии разных неопептидов-кандидатов в опухоли субъекта, подлежащего лечению, и ранжировать их по снижающемуся уровню экспрессии, причем высокоэкспрессируемые неопептиды ранжируются первыми, вплоть до неэкспрессируемых неопептидов в конце данного списка.
Альтернативно или в комбинации с экспрессией мРНК, отбор неопептидов может осуществляться на основе уровня экспрессии пептида, возникающего в результате опухолеспецифичной мутации, с использованием хорошо описанных протеомных технологий, таких как вестерн-блоттинг, иммуноанализы, масс-спектрометрия. Здесь также отбор является скорее относительным, экспрессия потенциальных неопептидов выявляется/количественно измеряется и ранжируется по порядку приоритета. Это также легко для специалистов в данной области.
Подстадия е): альтернативно или в комбинации с подстадией д), также можно добавлять дополнительное фильтрование посредством включения стадии отбора неаутологичных неопептидов, подразумевая неопептид, который не присутствует как таковой в рассматриваемом протеоме. В самом деле, в то время как присутствие опухолеспецифичной мутации обеспечивает то, что такой же пептид не присутствует в соответствующем белке дикого типа, экспрессируемом в здоровых клетках субъекта, целью подстадий е) является проверка того, что такой же пептид не является частью другого белка, экспрессируемого в здоровых клетках субъекта. Для иллюстративной цели полипептид, который идентифицируется как мутировавший, разрезается на множество минимальных иммунных детерминант (например, 9-меры, перекрывающиеся на одну аминокислоту) и выравнивается с использованием blastp (Altschul et al., 1990, J. Mol. Biol. 215(3):403-10) относительно протеома хозяина (например, человеческого протеома для человеческого субъекта), соответствуя белкам, которые экспрессируются у данного хозяина. Затем отбираются мутировавшие эпитопы, которые не являются общими с белками хозяина. Другими словами, аутологичные эпитопы (т.е. присутствующие в протеоме) устраняются для устранения проблем аутоиммунности и риска иммунной толерантности против собственного пептида, которая может уменьшать иммунный ответ.Как упомянуто выше, подстадию е) можно предпочтительно объединять с подстадией д), таким образом, приводя к отбору (высоко) экспрессируемых неаутологичных неопептидов.
Подстадия ж): альтернативно или в комбинации с подстадией д), подстадией е) и комбинацией подстадий д) и е), кроме того, может быть добавлено дополнительное фильтрование посредством прогнозирования иммунного потенциала отобранных неопептидов. Такой отбор, основанный на иммунном потенциале, может осуществляться или посредством прогнозирования in silico и/или с использованием биоаналитических подходов (например, посредством биологических анализов иммуногенности). Средства прогнозирования in silico строятся с использованием алгоритмов машинного обучения, таких как искусственные нейронные сети (ANN) или метод опорных векторов (SVM), и обучаются для идентификации высокоиммуногенных последовательностей. Данный алгоритм затем используется для классификации эпитопов на их аффинность в отношении белка HLA класса I и/или класса II в качестве индекса их потенциальной презентации. Для иллюстративных целей эта дополнительная и возможная стадия может включать стадию типирования HLA и прогнозирования связывания с МНС класса II (разрезание мутировавшего антигена на множество пептидов/эпитопов (обычно 8-10-меры для связывания с классом I и 10-20-меры для связывания с классом II с перекрыванием на одну аминокислоту) с использованием алгоритма обучения, такого как прогнозирование консенсусного связывания seq2HLA (Boegel et al., 2015, Methods Mol Biol 1310: 247-51), NetMHCIIpan (Andreatta et al., 2015, Immunogenetics 67(11-12): 641-50) и IEDB. Для иллюстративной цели предпочтительно отбираются мутировавшие пептиды, имеющие низкий балл. Другими словами, прогнозируется то, что такие пептиды имеют лучший балл связывания с классом МНС и, таким образом, лучшую вероятность генерации ответа Т-клеток.
Альтернативно или помимо прогнозирования in silico иммунного потенциала, иммуногенные неопептиды могут быть идентифицированы подходящими биоаналитическими анализами, хорошо известными в данной области, такими как масс-спектрометрия, анализы высвобождения цитокинов (например, ELIspot IFNγ в подходящей животной модели), анализы связывания с МНС класса I или класса II для идентификации мутаций, ограниченных МНС, проточная цитометрия, иммунофлуоресцентное окрашивание, гистохимия и т.д.
Как упомянуто выше, подстадия ж) предпочтительно может быть объединена с подстадией д), подстадией е) или обеими подстадиями д) и е), более предпочтительно с обеими подстадиями д) и е), затем приводя к отбору (высоко)экспрессируемых неаутологичных и иммуногенных неопептидов.
Подстадия з): альтернативно или в комбинации с подстадией д), подстадией е) или подстадией ж), или их любой комбинацией (например, подстадий д) и е) или подстадий е) и ж), или подстадий д) и ж), или подстадий д), е) и ж)), кроме того, может быть добавлено дополнительное фильтрование посредством прогнозирования присутствия потенциального ТМ сегмента в отобранных неопептидах (внутрипептидный ТМ) или в образующемся(щихся) слиянии(ниях) неопептидов (внутрипептидный ТМ). Как описано выше, предпочтительно, чтобы отобранные неопептиды, а также образующееся(щиеся) слияние(ния) неопептидов не содержали ТМ сегмента для облегчения получения рекомбинантного поксвируса и подходящей презентации неоэпитопов иммунной системе. Как упомянуто выше, для прогнозирования присутствия ТМ сегмента в данном пептиде можно использовать несколько средств прогнозирования, таких как ТМНММ (Krogh et al., 2001, J. Mol. Biol. 305: 567-80) и DAS (Dense Alignment Surface). В случае прогнозирования внутрипептидного ТМ в соединении двух неопептидов, обращение данных неопептидов может быть возможностью для устранения присутствия ТМ. Другой возможностью может быть добавление подходящего линкера между 2 неопептидами.
Подстадия и): альтернативно или в комбинации с подстадией д), подстадией е), подстадией ж) или подстадией з), или их любой комбинацией (например, подстадий д) и е) или подстадий е) и ж), или подстадий д) и ж), подстадий ж) и з) или подстадий д), е) и ж), или подстадий д), е), ж) и з)), кроме того, может быть добавлено дополнительное фильтрование посредством ранжирования неопептида(дов) и/или слияния(ний) неопептидов согласно их степени гидрофобности или гидрофильности. В самом деле, в то время как слияние(ния) неопептидов может(гут) переносить присутствие одного или более чем одного короткого отрезка гидрофобных аминокислотных остатков, предпочтительно, чтобы по меньшей мере 60% отобранных неопептидов были гидрофильными по природе, как описано выше, для обеспечения того, что образующееся слияние неопептидов имеет общий отрицательный индекс гидрофобности и/или общий индекс гидропатичности меньше 0,1. Такое прогнозирование может осуществляться традиционными способами и алгоритмами, известными в данной области, такими как процитированные выше (например, Kyte and Doolittle, 1982, J. Mol. Biol. 157: 105-32 или алгоритм ExPAsy Prot Scale Protein).
Получение рекомбинантного поксвируса
После стадии идентификации способ получения персонализированной противораковой вакцины согласно настоящему изобретению может включать стадию получения рекомбинантного поксвируса. Типично такая стадия включает получение синтетической нуклеиновой кислоты, кодирующей отобранный(ные) неопептид(ды), как описано в данном документе (например, организация в одном или более чем одном слиянии, возможно с пептидным сигналом, линкерами, меткой и т.д.), получение рекомбинантного поксвируса, экспрессирующего неопептид, и изготовление указанного рекомбинантного поксвируса.
Получение синтетической(ких) молекулы(кул) нуклеиновой кислоты Молекула(лы) нуклеиновой кислоты разрабатывается(ются) для кодирования отобранного(ных) неопептида(дов) с предпочтением в отношении организации в одном или более чем одном слиянии, возможно, с линкерами, метками и пептидными сигналами, как описано в данном документе. Ее предпочтительно получают химическим синтезом в автоматическом способе (например, собранная из перекрывающихся синтетических олигонуклеотидов или посредством прямого синтеза синтетического гена).
В некоторых воплощениях может быть полезным оптимизировать последовательность(ти) нуклеиновой кислоты для предоставления высокого уровня экспрессии у конкретного субъекта. Вкратце, молекула(лы) нуклеиновой кислоты может быть сконструирована с целью улучшения экспрессии посредством оптимизации кодонов, неотбора неоптимальных кодонов для данного организма-хозяина и/или устранения отрезков AT- или GC-богатых последовательностей, которые, как ожидается, отрицательно влияют на экспрессию. Последовательность TTTTTNT также избегается, так как она представляет собой ранний сигнал терминации транскрипции поксвируса.
Экспрессия молекулы(кул) синтетической нуклеиновой кислоты
Согласно настоящему изобретению молекула(лы) нуклеиновой кислоты, кодирующая(щие) неопептиды, подлежащая(щие) вставке в геном рекомбинантного поксвируса, функциональным образом связывается(ются) с подходящими регуляторными элементами. В контексте данного изобретения молекула(лы) нуклеиновой кислоты, кодирующая(щие) неопептиды, может(гут) быть организована(ны) в одной или более чем одной экспрессионной кассете. Типично «экспрессионная кассета» содержит молекулу нуклеиновой кислоты, кодирующую один или более чем один неопептид под контролем подходящих регуляторных элементов, обеспечивающих экспрессию у субъекта.
Термин «регуляторные элементы» в том виде, в котором он используется в данном документе, относится к любому элементу, который обеспечивает, способствует или модулирует экспрессию неопептида(дов), включая репликацию, дуликацию, транскрипцию, сплайсинг, трансляцию, стабильность и/или процесс транспорта. Термин «связанный функциональным образом» в том виде, в котором он используется в данном документе, означает то, что связанные элементы организуются таким образом, что они функционируют совместно для их намеченых целей.
Специалистам в данной области будет понятно то, что выбор регуляторных элементов может зависеть от таких факторов, как сами неопептиды, сам поксвирус, субъект, подлежащий лечению, желательный уровень экспрессии и т.д. Промотор является особенно важным. В контексте данного изобретения он может быть конститутивным, управляющим экспрессией во многих типах клеток, или специфичным для определенных клеток или типов клеток (например, опухолеспецифичные регуляторные последовательности), или регулируемым в ответ на специфические события или экзогенные факторы (например, посредством температуры, питательной добавки, гормона и т.д.), или согласно фазе вирусного цикла (например, поздней или ранней). Также можно использовать промоторы, которые репрессируются во время стадии продукции в ответ на специфические события или экзогенные факторы, для того, чтобы оптимизировать продукцию вируса и обойти потенциальную токсичность экспрессируемого(мых) полипептида(дов). Специалистам в данной области будет понятно то, что, кроме промотора, регуляторные элементы могут дополнительно содержать дополнительные элементы для правильной инициации, регуляции и/или терминации транскрипции (например, полиА последовательности терминации транскрипции), транспорта, процессинга и стабильности мРНК, трансляции (например, инициирующий Met, трехкомпонентные лидерные последовательности, сайты связывания рибосомы IRES, сигнальные пептиды и т.д.).
Промоторы вируса осповакцины являются особенно адаптированными для экспрессии в рекомбинантных поксвирусах. Типичные примеры включают, без ограничения, промоторы вируса осповакцины 7.5К, H5R, 11К7.5 (Erbs et al., 2008, Cancer Gene Ther. 15(1): 18-28), TK, p28, p11, B2R, A35R и K1L, синтетические промоторы, такие как промоторы, описанные в Chakrabarti et al. (1997, Biotechniques 23: 1094-7; Hammond et al, 1997, J. Virol Methods 66: 135-8 и Kumar and Boyle, 1990, Virology 179: 151-8), а также ранние/поздние химерные промоторы.
Когда рекомбинантный поксвирус содержит несколько экспрессионных кассет, кодирующих один или более чем один неопептид, экспрессия может контролироваться одинаковыми или отличными регуляторными элементами (например, промотором). Предпочтительно рекомбинантный поксвирус содержит от одной до 5 экспрессионных кассет, более предпочтительно каждую для экспрессии слияния неопептидов, как описано в данном документе. Особенно предпочтительное воплощение направлено на рекомбинантный MVA, содержащий от одной до трех кассет, каждую для экспрессии слияния 5-10 неопептидов под транскрипционным контролем промотора вируса осповакцины, выбранного из группы, состоящей из p11k7.5, pH5R и pB2R.
В альтернативном воплощении, если число экспрессионных кассет затрудняет конструирование рекомбинантного поксвируса (например, больше 3), можно продолжать посредством системы, содержащей более чем один уникальный рекомбинантный поксвирус, означая то, что персонализированная противораковая вакцина содержит больше, чем один рекомбинантный поксвирус (например, 2 или 3), экспрессирующий неопептиды.
Вставка молекулы(кул) нуклеиновой кислоты, кодирующей(щих) неопептиды, рекомбинантного поксвируса
Молекула(лы) нуклеиновой кислоты или экспрессионная(ные) кассета(ты), кодирующая(щие) один или более чем один неопептид для применения в данном документе, вставляется(ются) в геном родительского поксвируса с получением указанного рекомбинантного поксвируса. Последовательность(ти) нуклеиновой кислоты/экспрессионная(ные) кассета(ты), кодирующая(щие) неопептид, могут быть вставлены в любое место в пределах генома поксвируса, например, в пределах вирусного гена, межгенной области, в несущественный ген или область, или вместо вирусных последовательностей. Предпочтение отдается вставке в пределы поксвирусного генома в несущественный локус.Ген тимидинкиназы является особенно подходящим для вставки в вирус осповакцины Copenhagen и Western Reserve, а делеция II или III - для вставки в вектор на основе MVA (WO 97/02355; Meyer et al., 1991, J. Gen. Virol. 72: 1031-8). Предпочтительно, когда поксвирус представляет собой MVA, вставка молекулы(кул) нуклеиновой кислоты или экспрессионной(ных) кассеты(сет), кодирующей(щих) неопептид, делается в пределах делеции III MVA. Когда рекомбинантный поксвирус содержит несколько кассет, как описано выше, они могут быть вставлены в вирусный геном в то же самое или отличное место. Предпочтение отдается вставке всех экспрессионных кассет в то же самое место, особенно в локус ТК для рекомбинантного вируса осповакцины и в делецию III для рекомбинантного MVA.
Общие условия для конструирования рекомбинантных поксвирусов хорошо известны в данной области (см., например, WO 2007/147528; WO 2010/130753; WO 03/008533; US 6998252; US 5972597 и US 6440422). Типично молекула(лы) нуклеиновой кислоты/экспрессионная(ные) кассета(ты), подлежащая(щие) вставке, клонируется(ются) в плазмиду для переноса, окруженную двумя рекомбинантными ручками, соответствующими отрезкам поксвирусных последовательностей, гомологичных (например, на 90-100% идентичных) последовательностям, присутствующим в родительском геноме, на обеих сторонах сайта вставки. Длина данных рекомбинантных ручек в пределах плазмиды для переноса может варьировать. Желательно каждая из данных рекомбинантных ручек содержит по меньшей мере 150 п. о., предпочтительно по меньшей мере 200 п. о., более предпочтительно по меньшей мере 300 п. о., даже более предпочтительно от 300 до 600 п. о., с конкретным предпочтением в отношении 350-500 п. о. (например, приблизительно от 350 п. о. до 500 п. о.) или от 300 до 400 п. о. гомологичных поксвирусных последовательностей. Родительский поксвирус может представлять собой поксвирус дикого типа или модифицированный поксвирус (например, ослабленный, опухолеспецифичный и т.д.), как описано выше в связи с термином «поксвирус». Вставка затем проводится посредством гомологичной рекомбинации между отрезком гомологичных последовательностей, присутствующих и в родительском геноме, и в линеаризованной плазмиде для переноса, требующей трансфекции пермиссивных клеток линеаризованной плазмидой для переноса и инфекции родительским поксвирусом.
В некоторых воплощениях способа по настоящему изобретению стадия получения рекомбинантного поксвируса охватывает применение родительского поксвируса, содержащего ген-репортер и особенно флуоресцентный ген-репортер, клонированный в сайте вставки, который выбран для молекулы(кул) нуклеиновой кислоты/экспрессионной(ных) кассеты(сет), кодирующей(щих) неопептид. Предпочтительно данный ген-репортер ставится под транскрипционный контроль промотора, обеспечивающего его экспрессию в пермиссивных клетках, например, промотора вируса осповакцины. Данное воплощение облегчает отбор рекомбинантного поксвируса в отношении родительского поксвируса. Типичные примеры флуоресцентных репортеров, которые можно использовать в контексте настоящего изобретения, включают, без ограничения, GFP (зеленый флуоресцентный белок), eGFP (усиленный зеленый флуоресцентный белок), флуоресцентный белок AmCyan 1 и mCherry. Например, полагаясь на mCherry (мономерный флуоресцентный белок, который происходит из гриба Discosoma, с пиковым поглощением/испусканием при 587 нм и 610 нм), рекомбинантные вирусы, имеющие вставленную(ные) молекулу(лы) нуклеиновой кислоты или экспрессионную(ные) кассету(ты), кодирующую(щие) неопептид, вместо последовательностей, кодирующих mCherry, будут давать белые бляшки, тогда как родительские вирусы, сохраняющие экспрессионную кассету mCherry, будут давать красные бляшки. Отбор рекомбинантного поксвируса может осуществляться посредством прямой визуализации (белые бляшки) или также может облегчаться средствами сортировки, такими как FACS (флуоресцентная сортировка клеток), после мечения меченным АРС (аллофикоцианин) антителом против вируса осповакцины. Огромное число антител против вируса осповакцины доступно из коммерческих источников. Обычно один рекомбинант получают на 50-100 родительских вирусов, и весь способ от стадии вставки до получения рекомбинантного поксвируса занимает 5-6 недель.
Предпочтительное воплощение данного изобретения направлено на стадию получения рекомбинантного поксвируса, обеспечивающую увеличение эффективности гомологичной рекомбинации по меньшей мере в 10 раз и уменьшение необходимого времени для получения рекомбинантного поксвируса по меньшей мере в 1,5 раза. Как и в способе, описанном выше, нуклеиновая(вые) кислота(ты) или экспрессионная(ные) кассета(ты), подлежащая(щие) вставке в поксвирусный геном, клонируется(ются) в плазмиду для переноса, окруженную рекомбинантными ручками, и родительский вирус содержит ген-репортер (например, флуоресцентный репортер), клонированный в сайте вставки под контролем подходящего промотора. Предпочтительно указанный родительский вирус представляет собой MVA, содержащий ген-репортер, вставленный в делецию III (в частности, ген mCherry), под контролем промотора поксвируса. Однако стадия получения рекомбинантного посквируса включает дополнительную стадию расщепления эндонуклеазой, способной генерировать по меньшей мере один двухнитевой разрыв в нуклеотидной последовательности репортера (например, mCherry), но в которой указанная эндонуклеаза не расщепляет поксвирусный геном. Подходящая эндонуклеаза предпочтительно выбрана из группы, состоящей из нуклеаз с цинковыми пальцами (ZFN), эффекторных нуклеаз, подобных активатору транскрипции (TALEN), нуклеаз CRISPR (кластерные короткие палиндромные повторы, разделенные регулярными промежутками)/Саs9 и рестрикционных ферментов с уникальным расщеплением в пределах гена флуоресцентного репортера. Применение системы CRISPR/CAS9 для редактирования вируса описывается в данной области (Yuan et al., 2015, J. Virol 89, 5176-9; Yuan et al., 2016, Viruses 8, 72, doi:10.3390), требуя применения плазмиды, кодирующей Cas9 без сигнала ядерной локализации, а также подходящей РНК-проводника. При данном рассмотрении, помимо инфицирования родительским вирусом, пермиссивные клетки могут трансфицироваться плазмидой для переноса, плазмидой, экспрессирующей Cas9, и одной или более чем одной плазмидой, кодирующей РНК-проводник (например, РНК-проводник, нацеленную на mCherry). Отбор рекомбинантного поксвируса затем проводится визуально (прямое выделение белых бляшек, соответствующих рекомбинанту, тогда как окрашенные бляшки соответствуют родительскому вирусу, а цвет зависит от гена-репортера) или с использованием традиционных средств сортировки (FACS, возможно после стадии мечения подходящими антителами, как описано выше). С использованием данного способа один рекомбинант получается на 1-10 родительских вирусов, и весь способ от стадии вставки до получения рекомбинантного поксвируса занимает приблизиельно 3-4 недели. В предпочтительном воплощении стадия получения рекомбинантного поксвируса занимает самое большее 4 недели, предпочтительно самое большее 3 недели, с отношением рекомбинантного к родительскому, составляющим от 2 (2 к 1) до 0,1 (1 к 10), предпочтительно от 2 (2 к 1) до 0,05 (1 к 20). В конкретных воплощениях отношение рекомбинантный/родительский составляет 1 или больше 1, особенно когда две рекомбинантные ручки имеют приблизительно 350 п. о. в длину.
Особенно предпочтительное воплощение направлено на способ, обеспечивающий получение рекомбинантного поксвируса (например, вируса осповакцины и, более предпочтительно MVA), содержащего от одной до трех экспрессионных кассет, вставленных в пределы локуса ТК или делеции III MVA, причем каждая кодирует слияние из 5-10 неопептидов, помещенных под транскрипционный контроль промотора вируса осповакцины, выбранного из группы, состоящей из p11k7.5, р7.5К, pH5R и pB2R, предпочтительно из группы, состоящей из p11k7.5, pH5R и pB2R (предпочтительно каждое слияние находится под транскрипционным контролем отличного промотора). Настоящее изобретение также относится к персонализированной противораковой вакцине, содержащей такой рекомбинантный поксвирус.
Изготовление рекомбинантного поксвируса
Сразу после получения рекомбинантный поксвирус для применения в данном документе может быть продуцирован/амплифицирован с использованием традиционных методик. Следовательно, способ по настоящему изобретению может дополнительно включать стадию изготовления указанного рекомбинантного поксвируса. В предпочтительном воплощении указанная стадия изготовления включает стадию амплификации в подходящей клетке-продуценте в подходящем масштабе, стадию выделения продуцированного рекомбинантного поксвируса из культуры клеток и возможную стадию очистки выделенного рекомбинантного поксвируса.
Стадия амплификации включает культивирование (например, пермиссивной) продуцирующей клетки-хозяина, инфицирование культивируемых продуцирующих клеток-хозяев и культивирование инфицированных клеток-хозяев при подходящих условиях таким образом, чтобы обеспечивать продукцию рекомбинантного поксвируса (например, инфекционных вирусных частиц).
Выбор продуцирующей клетки зависит от типа рекомбинантного поксвируса, подлежащего амплификации. MVA является строго ограниченным в отношении хозяина и типично амплифицируется на клетках птиц: либо первичных клетках птиц (таких как фибробласты эмбриона курицы (CEF), полученные из эмбрионов курицы, полученных из оплодотворенных яиц), либо сделанных бессмертными линиях клеток птиц. Репрезентативные примеры подходящих линий клеток птиц для продукции MVA включают, без ограничения, линии клеток Cairina moschata, сделанные бессмертными с использованием утиного гена TERT (см., например, WO 2007/077256, WO 2009/004016, WO 2010/130756 и WO 2012/001075); линии клеток птиц, сделанные бессмертными с использованием комбинации вирусных и/или клеточных генов (см., например, WO 2005/042728); ставшие спонтанно бессмертными клетки (напримр, линия клеток курицы DF1, раскрытая в US 5879924); или сделанные бессмертными клетки, которые происходят от эмбриональных клеток посредством прогрессирующего отделения от факторов роста и фидерного слоя (например, линии клеток курицы Еbх, раскрытые в WO 2005/007840 и WO 2008/129058, такие как Еb66, описанная в Olivier et al., 2010, mAbs 2(4): 405-15).
Для других вирусов осповакцины или других штаммов поксвируса, помимо первичных клеток птиц (таких как CEF) и линий клеток птиц, доступны многие другие нептичьи линии клеток для продукции, включая человеческие линии клеток, такие как HeLa (ATCC-CRM-CCL-2™ или ATCC-CCL- 2.2™), MRC-5, НЕК-293; линии клеток хомяка, такие как ВНК-21 (АТСС CCL-10), и клетки Vero. В предпочтительном воплощении вирус осповакцины, не являющийся MVA, амплифицируется в клетках HeLa (см., например, WO 2010/130753).
Продуцирующие клетки предпочтительно культивируются в среде, не содержащей продуктов животного или человеческого происхождения, с использованием химически определенной среды без продукта животного или человеческого происхождения. В частности, в то время как могут присутствовать факторы роста, они предпочтительно продуцируются рекомбинантно, а не очищаются из животного материала. Подходящая среда, не содержащая животных компонентов, может быть легко выбрана специалистами в данной области, в зависимости от выбранных продуцирующих клеток. Такие среды имеются в продаже. В частности, при использовании CEF в качестве продуцирующих клеток, они могут культивироваться в среде для культивирования клеток VP-SFM (Invitrogen). Продуцирующие клетки предпочтительно культивируются при температуре, составляющей от 30°С до 38°С (более предпочтительно около 37°С), в течение от 1 до 8 суток (предпочтительно в течение 1-5 суток для CEF и 2-7 суток для клеток, сделанных бессмертными) перед инфицированием. Если необходимо, можно делать несколько пассажей от 1 до 8 суток для того, чтобы увеличивать общее число клеток. Например, после выделения белых бляшек амплификация рекомбинантного поксвируса (например, MVA), экспрессирующего неоэпитоп, может проводиться сначала в 6-12-луночных планшетах для культуры клеток перед переносом во флаконы для получения желательного качества вируса.
Инфекция продуцирующих клеток рекомбинантным поксвирусом осуществляется при подходящих условиях (в частности, с использованием подходящей множественности заражения (MOI)) для обеспечения продуктивной инфекции продуцирующих клеток. Подходящая MOI, подлежащая применению для амплификации поксвирусов, типично составляет от 0,001 до 1 (более предпочтительно примерно 0,05). Стадия инфекции также предпочтительно проводится в среде (которая может быть такой же или отличной от среды, используемой для культивирования продуцирующих клеток), не содержащей продуктов животного или человеческого происхождения, с использованием химически определенной среды без продукта животного или человеческого происхождения. Стадия инфекции обычно длится от одних до шести суток, более предпочтительно от двух до четырех суток и наиболее предпочтительно примерно 72 часа.
Инфицированные продуцирующие клетки затем культивируются при подходящих условиях, хорошо известных специалистам в данной области, пока не продуцируется вирусный вектор потомства (например, инфекционные вирусные частицы). Культивирование инфицированных продуцирующих клеток также предпочтительно проводится в среде (которая может быть такой же или отличной от среды, используемой для культивирования продуцирующих клеток и/или стадии инфицирования), не содержащей продуктов животного или человеческого происхождения (с использованием химически определенной среды без продукта животного или человеческого происхождения) при температуре от 30°С до 37°С в течение 1-5 суток.
Поксвирусные частицы могут быть отобраны из супернатанта культуры и/или продуцирующих клеток. Супернатант культуры клеток и продуцирующие клетки могут быть объединены или отобраны по-отдельности. Выделение из продуцирующих клеток (и возможно также из супернатанта культуры) может потребовать стадии, обеспечивающей разрушение мембраны продуцирующей клетки для обеспечения высвобождения вектора. Специалистам в данной области доступны разные методики, включающие: замораживание/оттаивание, гипотонический лизис, обработку ультразвуком, микропсевдоожижение или высокоскоростную гомогенизацию, но не ограничивающиеся ими. Согласно предпочтительному воплощению стадия выделения продуцированного рекомбинантного поксвируса включает стадию лизиса, в которой мембрана продуцирующей клетки разрушается, предпочтительно с использованием высокоскоростного гомогенизатора. Высокоскоростные гомогенизаторы имеются в продаже у Silverson Machines Inc (East Longmeadow, США) или Ika-Labotechnik (Staufen, Германия). Согласно особенно предпочтительному воплощению указанный высокоскоростной гомогенизатор представляет собой SILVERSON L4R.
Поксвирусные частицы могут быть затем дополнительно очищены с использованием стадий очистки, хорошо известных в данной области. Могут рассматриваться разные стадии очистки, включающие осветление, ферментативную обработку (например, эндонуклеаза, протеаза и т.д.), стадии хроматографии и фильтрования. В данной области описываются подходящие способы (например, WO 2007/147528; WO 2008/138533, WO 2009/100521, WO 2010/130753, WO 2013/022764). В предпочтительном воплощении стадия очистки включает стадию фильтрования тангенциальным потоком (TFF), которую можно использовать для отделения вируса от других биомолекул для концентрирования и/или обессоливания суспензии вируса. В данной области доступны разные системы и устройства TFF, в зависимости от объема, подлежащего фильтрованию, включающие, без ограничения, среди прочих, Spectrumlabs, Pall Corp, PendoTech и New Pellicon.
В предпочтительном воплощении в указанной стадии изготовления достигается продукция по меньшей мере 109 БОЕ (бляшкообразующая единица), желательно по меньшей мере 5×109 БОЕ и предпочтительно приблизительно 1010 БОЕ или более для персонализированной противораковой вакцины, подлежащей распространению в подходящих дозах для тестирования и лечения пациента (от которого происходит образец опухоли).
Рекомбинантные поксвирусы с ручками
В некоторых воплощениях данного изобретения также охватываются рекомбинантные поксвирусы, которые, кроме молекулы(кул) нуклеиновой кислоты, кодирующей(щих) неопептиды, содержат дополнительный(ные)
терапевтический(кие) ген(ны), вставленный(ные) в вирусный геном, например, для усиления противоопухолевого ответа. Можно рассматривать огромное число терапевтических генов, особенно генов, кодирующих полипептиды, способные потенцировать противоопухолевую эффективность вируса или усиливать иммунитет хозяина. Они могут быть человеческого происхождения или нет (например, бактериального, дрожжевого или вирусного происхождения) и представлять собой природный ген, его фрагмент или функциональный аналог (полученный из последнего посредством мутации, делеции, замены и/или присоединения одного или более чем одного нуклеотида). Такой аналог предпочтительно имеет нуклеотидную последовательность, имеющую степень идентичности по меньшей мере 70%, преимущественно по меньшей мере 80%, предпочтительно по меньшей мере 90% и наиболее предпочтительно по меньшей мере 95% с последовательностью нуклеиновой кислоты природного гена. Предпочтительные терапевтические гены выбраны из группы, состоящей из суицидных и иммуностимулирующих генов.
Иммуностимулирующие терапевтические гены
Иммуностимулирующие терапевтические гены кодируют полипептиды, способные стимулировать иммунную систему или эффекторные клетки специфичным или неспецифичным способом. Примеры подходящих иммуностимулирующих белков в контексте данного изобретения включают, без ограничения, цитокины, с особым предпочтением в отношении интерлейкинов (например, IL-2, IL-6, IL-12, IL-15, IL-24), хемокинов (например, CXCL10, CXCL9, CXCL11), интерферонов (например, IFNγ, Ральфа), фактора некроза опухолей (TNF), колониестимулирующих факторов (например, GM-CSF (гранулоцитарно-макрофагальный колониестимулирующий фактор), G-CSF (гранулоцитарный колониестимулирующий фактор), M-CSF (макрофагальный колониестимулирующий фактор)), белков, экспонированных АРС (антигенпрезентирующая клетка) (например, В7.1, В7.2 и тому подобные), факторов роста (трансформирующий фактор роста (TGF), фактор роста фибробластов (FGF), факторы роста эндотелия сосудов (VEGF) и тому подобные), компонентов главного комплекса гистосовместимости (МНС) класса I или II, индукторов или ингибитров апоптоза (например, Вах, Bcl2, ВсlХ…), цитостатических агентов (р21, р16, Rb…), иммунотоксинов, блокаторов механизмов ускользания иммунного ответа, таких как ингибиторы иммунных контрольных точек (указанный ингибитор может представлять собой блокирующие пептиды, антитела или родственные биологические соединения, такие как однодоменные антитела или фрагменты антител), cGAS и тому подобные.
Суицидные гены
Термин «суицидный ген» относится к гену, кодирующему белок, способный превращать предшественник лекарственного средства в цитотоксическое лекарственное средство. Типичные примеры подходящих суицидных генов включают тимидинкиназу (ТК), тимидилаткиназу, цитозиндезаминазу (ЦДаза) и урацилфосфорибозилтрансферазу (УФРТаза), но не ограничиваются ими.
ЦДаза и УФРТаза присутствуют у прокариот и низших эукариот (но не у млекопитающих). ЦДаза, посредством которой экзогенный цитозин превращается в урацил посредством гидролитического дезаминирования, участвует в метаболическом пути пиримидина. ЦДаза также дезаминирует аналог цитозина, т.е. пролекарство 5-фторцитозин (5-ФЦ), образуя, посредством этого, 5-фторурацил (5-ФУ) - соединение, которое является высокоцитотоксическим при его превращении УФРТазой в 5-фтор-УМФ (5-ФУМФ). Последовательности генов и закодированных ферментов являются общедоступными, например, в специализированных банках данных (SWISSPROT EMBL, Genbank, Medline и тому подобные). Особенно предпочтительными являются дрожжевые УФРТаза и ЦДаза, в частности, кодируемые Saccharomyces cerevisiae (ген FUR1; Kern et al., 1990, Gene 88: 149-57). Особый интерес представляет суицидный ген FCU1, кодирующий слияние ЦДазы S. cerevisia и усеченной УФРТазы, с делетированными первыми 35 остатками (слияние FCY1::FUR1[Дельта]105), причем данная аминокислотная последовательность описывается в идентификаторе последовательностей SEQ ID NO: 1 WO 2009/065546).
Терапевтический ген для применения в данном документе может быть независимо получен целым рядом способов, известных специалистам в данной области (например, клонирование, ПЦР-амплификация, перетасовка ДНК, химический синтез), и из любого доступного источника (например, биологических материалов, описанных в данной области, таких как кДНК, геномные библиотеки, вирусные геномы или вектор предшествующего уровня техники, для которого известно, что он содержит его) с использованием данных по последовательности, доступных специалисту, и информации по последовательности, предоставленной в данном документе, и затем подходящим образом вставлен в рекомбинантный поксвирус либо в то же самое место, что и молекула(лы) нуклеиновой кислоты, кодирующая(щие) неоэпитоп (например, в делецию III MVA), либо в другое место посредством традиционных методик молекулярной биологии.
Фармацевтическая композиция
В одном воплощении персонализированная противораковая вакцина по настоящему изобретению находится в форме композиции, содержащей терапевтически эффективное количество рекомбинантного поксвируса, описанного в данном документе (или полученного согласно способу, описанному в данном документе), и фармацевтически приемлемый носитель.
Подразумевается то, что термин «фармацевтически приемлемый носитель» включает любой и все носители, растворители, разбавители, эксципиенты, адъюванты, диспергирующие среды, покрытия, антибактериальные и противогрибковые средства, средства для поглощения и тому подобное, совместимые с введением млекопитающим и, в частности, человеческим субъектам.
«Терапевтически эффективное количество» соответствует количеству рекомбинантного поксвируса, требующемуся для продукции у субъекта, которого лечат согласно настоящему изобретению, наблюдаемого улучшения его клинического состояния, включая по меньшей мере одно из улучшений, упомянутых далее.
Такое терапевтически эффективное количество может варьировать, в зависимости от множества факторов, включающих характеристики поксвируса (включая тип вируса, биодоступность и дозы), тяжесть и ход заболевания (например, стадия рака), самого субъекта (включая возраст, пол, клиническую историю, общее физическое состояние и т.д.), природу фармацевтически приемлемого носителя или эксципиента в препарате вируса и способы лечения (путь введения, частоту введения, тип сопутствующего лекарственного средства и т.д.), но не ограничивающихся ими. Подходящая дозировка поксвируса может быть определена традиционным образом и адаптирована практикующим врачом в свете релевнтных обстоятельств, например, посредством отслеживания ответа субъекта на введение вируса и соответствующей корректировки дозировки (относительно дополнительного руководства, см. Remington,; The Science and Practice of Pharmacy; Gennaro ed., Pharmaceutical Press, London, UK; например, 22-ое и последующие издания).
Для иллюстративных целей подходящее терапевтически эффективное количество для индивидуальных доз может варьировать от приблизительно 105 до приблизительно 1013 вч (вирусные частицы), ие (инфекционная единица) или БОЕ (бляшкообразующая единица), в зависимости от использованных поксвируса и количественной методики. В качестве общего руководства, индивидуальные дозы от приблизительно 106 БОЕ до приблизительно 1011 БОЕ являются особенно подходящими в контексте настоящего изобретения, более предпочтительно - от приблизительно 107 БОЕ до приблизительно 5×109 БОЕ; даже более предпочтительно дозы от приблизительно 5×107 БОЕ до приблизительно 109 БОЕ (например, от 5×107 до 6×108, от 6×107 до 5×108, от 7×107 до 4×108, от 8×107 до 3×108, от 9×107 до 2×108 БОЕ) являются удобными для человеческого применения, с предпочтением для индивидуальных доз, содержащих приблизительно 108 рекомбинантного поксвируса. Индивидуальные дозы могут быть снижены в 2-20 раз для местного(ных) введения(ний), такого(ких) как внутриопухолевая инъекция. Количество вируса, присутствующего в образце, может быть определено традиционными методиками титрования, например, посредством подсчета числа бляшек после инфицирования пермиссивных клеток (например, ВНК-21 или CEF), иммуноокрашивания (например, с использованием антител против вируса; Caroll et al., 1997, Virology 238: 198-211), измерением поглощения А260 (титры вч), посредством количественной иммунофлуоресценции (титры ие) или посредством кПЦР (количественная полимеразная реакция) с использованием специфичных вирусных праймеров и зондов.
В контексте данного изобретения могут рассматриваться разные препараты: либо жидкие, либо лиофилизированные для обеспечения стабильности вируса при условиях изготовления и долговременного хранения (т.е. в течение по меньшей мере 6 месяцев) при замораживании (например, -70°С, -20°С), охлаждении (например, 4°С) или температуре окружающей среды (например, 20-25°С). Рекомбинантный поксвирус преимущественно помещается в подходящий для человеческого или животного применения разбавитель. Типичные примеры подходящего разбавителя включают стерильную воду, физиологический раствор (например, хлорид натрия), раствор Рингера, растворы глюкозы, трегалозы или сахарозы, раствор Хэнкса и другие водные физиологически сбалансированные солевые растворы.
Желательно композиция персонализированной противораковой вакцины подходящим образом буферизуется для человеческого применения. Для поддержания физиологического или слегка основного рН (например, от приблизительно рН 7 до приблизительно рН 9) особенно подходящими являются такие буферы, как TRIS (трис(гидроксиметил)метиламин), TRIS-HCl (трис(гидроксиметил)метиламин-НСl), HEPES (4-2-гидроксиэтил-1-пиперазинэтансульфоновая кислота), фосфатный буфер (например, PBS; смесь Na2HPO4 и КН2РO4; смесь Na2HPO4 и NaH2PO4), TEA (триэтаноламин), EPPS (N-(2-гидроксиэтил)-пиперазин-N'-3-пропансульфоновая кислота), TRIClNE (N-[Трис(гидроксиметил)-метил]-глицин) и бикарбонатные буферы. Данный буфер (например, TRIS-HCl) предпочтительно присутствует в концентрации от 10 до 50 мМ.
Также может быть полезным включение одновалентной соли таким образом, чтобы обеспечить подходящее осмотическое давление. А именно, указанная одновалентная соль может быть выбрана из NaCl и KCl, предпочтительно указанная одновалентная соль представляет собой NaCl, а именно: в концентрации от 10 до 500 мМ.
Если необходимо, композиция противораковой вакцины также может включать криопротектор таким образом, чтобы облегчать хранение при низкой температуре. Подходящие криопротекторы включают, без ограничения, сахарозу, трегалозу, мальтозу, лактозу, маннит, сорбит и глицерин, например, в концентрации, варьирующей от 0,5 до 20% (масса в г/объем в л - называется масс./об.) и предпочтительно от 5 до 15% (масс./об.), с предпочтением для примерно 10%. Присутствие высокомолекулярных полимеров, таких как декстран или поливинилпирролидон (PVP) особенно подходит для лиофилизированных препаратов для защиты рекомбинантного поксвируса во время стадий вакуумной сушки и лиофилизации (см., например, WO 03/053463; WO 2006/0850082; WO 2007/056847; WO 2008/114021; WO 2014/053571).
Для иллюстративной цели, буферизованные препараты, включающие NaCl и/или сахар, являются особенно адаптированными для консервации поксвирусов (например, 10 мМ Tris, рН 8, с 5% (масс./об.) сахарозой, 10 мМ глутамат натрия и 50 мМ NaCl; или фосфатно-солевой буферный раствор с глицерином (10%) и NaCl).
Терапевтические применения и способ лечения
Персонализированная противораковая вакцина и способ по настоящему изобретению особенно подходят для лечения раковых заболеваний и, в частности, метастатических раковых заболеваний и раковых заболеваний с высоким риском рецидива. Следовательно, согласно настоящему изобретению также предложены персонализированная противораковая вакцина (например, композиция поксвируса) для применения для лечения рака или предупреждения его рецидива у субъекта, а также применение данной персонализированной противораковой вакцины для таких терапевтических целей. Настоящее изобретение также относится к персонализированной противораковой вакцине или ее композиции, описанной в данном документе, для изготовления лекарственного средства для лечения рака или предупреждения его рецидива.
В другом аспекте настоящее изобретение также относится к способу лечения, включающему введение композиции персонализированной противораковой вакцины субъекту, нуждающемуся в этом, в достаточном количестве для лечения рака или предупреждения его рецидива у указанного субъекта.
Репрезентативные примеры раковых заболеваний, которые можно лечить в контексте настоящего изобретения, включают рак кости, рак печени, рак поджелудочной железы, рак желудка, рак толстой кишки, рак пищевода, рака рта и гортани, рак легкого, рак головы или шеи, рак кожи, меланому, рак матки, рак шейки матки, рак яичника, рак молочной железы, рак прямой кишки, рак анальной области, рак предстательной железы, лимфому, рак эндокринной системы, рак щитовидной железы, саркому мягкой ткани, хронические или острые лейкозы, рак мочевого пузыря, рак почки, новообразование центральной нервной системы (ЦНС), глиому и т.д. Настоящее изобретение особенно подходит для лечения солидных опухолей. Оно также является особенно полезным для лечения запущенных раковых заболеваний, включающих метастатические солидные раковые заболевания или раковые заболевания, ассоциированные с высоким риском рецидива. Предпочтительные раковые заболевания для лечения согласно способам, описанным в данном документе, включают рак мозга, такой как, например, астроцитомы, эмбриональные опухоли, герминомы, атипическая тератоидная/рабдоидная опухоль центральной нервной системы, краниофарингиома, эпендимома, глиома и глиобластома, а также рак головы и шеи. Другой тип раковых заболеваний для лечения согласно способам, описанным в данном документе, представляет собой раковые заболевания легкого и, в частности, NSCLC (немелкоклеточный рак легкого) с особым предпочтением для аденокарциномы, плоскоклеточной карциномы и крупноклеточной карциномы. Другим подходящим раковым заболеванием, подлежащим лечению персонализированной противораковой вакциной по данному изобретению, является рак яичника.
Полезные эффекты, предоставляемые применением способа с использованием персонализированной противораковой вакцины по настоящему изобретению, могут быть подтверждены по наблюдаемому улучшению клинического состояния относительно исходного состояния или относительно ожидаемого состояния без лечения согласно способам, описанным в данном документе. Улучшение клинического состояния может легко оцениваться посредством любого релевантного клинического измерения типично используемого лечащими врачами или другим квалифицированным персоналом системы здравоохранения. В контексте данного изобретения терапевтическая польза может быть кратковременной (в течение одного или двух месяцев после завершения введения) или длительной (в течение нескольких месяцев или лет). Поскольку естественное изменение клинического состояния может значительно варьировать от одного субъекта до другого, не требуется, чтобы терапевтическая польза наблюдалась у каждого субъекта, которого лечат, но у значительного числа субъектов (например, статистически значимые различия между двумя группами могут быть определены посредством любого статистического критерия, известного в данной области, такого как параметрический критерий Тьюки, критерий Крускала-Уоллиса, U-критерий согласно Манну и Уитни, t-критерий Стьюдента, критерий Уилкоксона и т.д.).
В конкретных воплощениях такая терапевтическая польза может коррелировать с одним или более чем одним из следующего: ингибирование или замедление роста опухоли, пролиферации и метастазов, предупреждение или задержка инвазии опухоли (распространения опухолевых клеток в соседние ткани), уменьшение числа опухолевых поражений; уменьшение размера опухоли, уменьшение числа или степени метастазов, обеспечение длительной общей выживаемости (OS), увеличение выживаемости без прогрессирования заболевания (PFS), увеличение длительности ремиссии, стабилизация (т.е. не ухудшение) состояния заболевания, предупреждение рецидива заболевания, обеспечение лучшего ответа на другое лечение и, в частности, другую иммунотерапию, улучшение качества жизни и/или индуцирование противоопухолевого ответа (например, неспецифичного (врожденного) и/или специфичного, такого как ответ цитотоксических Т-клеток) у субъекта, которого лечат согласно настоящему изобретению.
Подходящие измерения в установившейся практике проводятся в медицинских лабораториях и больницах, доступных для оценки клинической пользы, как, например, анализы крови, анализ биологических жидкостей и биопсий, а также методики медицинской визуализации, и большое число наборов имеется в продаже. Они могут проводиться до введения (исходный уровень) и в разные моменты времени во время лечения и после завершения лечения.
Введение персонализированной противораковой вакцины или ее композиции
Применим любой из традиционных путей введения, с предпочтением в отношении парентеральных путей. Парентеральные пути предназначены для введения в виде инъекции или инфузии и охватывают системные, а также местные пути. Особенно подходящие пути введения включают, без ограничения, внутривенный (в вену), внутрисосудистый (в кровеносный сосуд), внутриартериальный (в артерию), интрадермальный (в дерму), подкожный (под кожу), внутримышечный (в мышцу), внутрибрюшинный (в брюшину), внутримозговой (в мозг) и внутриопухолевый (в опухоль или рядом с ней) пути, а также скарификацию. Инфузии типично даются внутривенным путем или внутриопухолевым (в большую опухоль). Введения через слизистую также рассматриваются настоящим изобретением и включают, без ограничения, пероральный/алиментарный, интраназальный, внутритрахеальный, носоглоточный, внутрилегочный, интравагинальный или интраректальный путь. Местные введения наносились непосредственно на кожу или на поверхность тканей (например, глазные капли; ушные капли и т.д. Также может рассматриваться ингаляция, особенно когда опухоль, подлежащая лечению, находится в респираторном тракте и легких. Персонализированную противораковую вакцину или ее композицию предпочтительно вводят пациенту посредством внутривенной, подкожной, внутримышечной или внутриопухолевой инъекций.
При введениях могут использоваться стандартные иглы и шприцы или любое устройство, доступное в данной области, способное облегчать или улучшать доставку, включая, например, катетеры, электрический шприц, инъекционные иглы Quadrafuse, безыгольные инъекционные устройства (например, устройство Biojector ТМ), инфузионные насосы, спреи и т.д. Для облегчения внутримышечного введения также может применяться электропорация. Местное введение также может осуществляться с использованием чрескожных средств (например, пластырь, микроиглы и тому подобное).
Персонализированная противораковая вакцина может вводиться в одной дозе или, более желательно, во многих дозах в течение протяженного периода времени. В контексте настоящего изобретения можно идти посредством последовательных циклов введений, которые повторяются после периода отдыха. Интервалы между каждым введением вируса могут составлять от нескольких часов до 6 месяцев (например, 24 ч, 48 ч, 72 ч, неделя, 2 недели, 3 недели, месяц, 2 месяца и т.д.). Интервалы также могут быть регулярными или нет. Дозы могут варьировать для каждого введения в пределах интервала, описанного выше. С целью иллюстрации предпочтительная трапевтическая схема включает от одного до 10 (например, 2, 3, 4, 5, 6, 7, 8, 9, 10) введений от 107 до 5×109 БОЕ рекомбинантного MVA с приблизительно 1 или 2-недельным интервалом, пока не наблюдается клиническая польза, и каждые 1-6 месяцев позднее.
Комбинированные терапии
В других воплощениях способа и терапевтических применений, описанных в данном документе, персонализированная противораковая вакцина может вводиться в сочетании с одной или более чем одной дополнительной противораковой терапией/ями, которые имеют пользу в лечении вышеупомянутых раковых заболеваний. В частности, дополнительная(ные) противораковая(вые) терапия/пии выбрана(ны) из группы, состоящей из хирургии, лучевой терапии, химиотерапии, криотерапии, гормональной терапии, терапии токсином, иммунотерапии и терапии цитокинами. Такая дополнительная противораковая терапия/пии вводится(дятся) субъекту согласно стандартной практике до, после, по существу одновременно или вперемежку с персонализированной противораковой вакциной по настоящему изобретению.
В конкретных воплощениях способ или применение согласно данному изобретению может осуществляться в сочетании с хирургией. Например, персонализированная противораковая вакцина может вводиться после частичной или полной хирургической резекции опухоли (например, посредством местного нанесения в пределы вырезанной зоны).
В других воплощениях способ согласно данному изобретению может использоваться в ассоциации с лучевой терапией. Специалисты в данной области могут легко подготовить подходящие протоколы и параметры лучевой терапии (см., например, Perez and Brady, 1992, Principles and Practice of Radiation Oncology, 2nd Ed. JB Lippincott Co; используя подходящие адаптации и модификации, как будет совершенно очевидно специалистам в данной области). Типы излучения, которое можно использовать в лечении рака, хорошо известны в данной области и включают пучки электронов, высокоэнергетические протоны от линейного ускорителя или от радиоактивных источников, таких как кобальт или цезий, протоны и нейтроны. Интервалы дозировок для радиоизотопов широко варьируют и зависят от периода полужизни изотопа, силы и типа испускаемой радиации и поглощения неопластическими клетками. В настоящем изобретении рассматриваются регулярные дозы рентгеновских лучей в течение длительных периодов времени (от 3 до 6 недель) или высокие одиночные дозы.
В некоторых воплощениях данного изобретения персонализированная противораковая вакцина может использоваться в сочетании с химиотерапией, доступной в настоящее время для лечения рака. Репрезентативные примеры подходящих химиотерапевтических агентов включают, без ограничения, алкилирующие агенты, ингибиторы топоизомеразы I, ингибиторы топоизомеразы II, ингибиторы раrр (поли-(АДФ-рибоза)полимераза), производные платины, ингибиторы тирозинкиназных рецепторов, циклофосфамиды, антиметаболиты, агенты, повреждающие ДНК, и антимитотические агенты.
В других воплощениях персонализированная противораковая вакцина может использоваться в сочетании с иммунотерапевтическими средствами, такими как антинеопластические антитела, а также с миРНК (малая интерферирующая РНК) и антисмысловымии полинуклеотидами. Репрезентативные примеры включают, среди прочих, моноклональные антитела, блокирующие специфические иммунные контрольные точки, такие как антитела против PD-1, против PD-L1, против CTLA-4, против LAG3 и т.д. (например, ипилимумаб, тремелимумаб, пембролизумаб, ниволумаб, пидилизумаб, AMP-224MEDI4736, MPDL3280A, BMS-936559 и т.д.), моноклональные антитела, блокирующие рецептор эпидермального фактора роста (в частности, цетуксимаб, панитумумаб, залутумумаб, нимотузумаб, матузумаб, трастузумаб (Herceptin™) и т.д.), и моноклональные антитела, блокирующие фактор роста эндотелия сосудов (в частности, бевацизумаб и ранибизумаб).
В дополнительных воплощениях персонализированную противораковую вакцину можно использовать в сочетании с адъювантом. Репрезентативные примеры подходящих адъювантов включают, без ограничения, лиганды TLR3 (Claudepierre et al., 2014, J. Virol. 88(10): 5242-55), лиганды TLR9 (например, CpG, такие как ODN1826 (Fend et al., 2014, Cancer Immunol. Res. 2, 1163-74) и литенимод (Li28) (Carpentier et al., 2003, Frontiers in Bioscience 8, e115-127; Carpentier et al., 2006, Neuro-Oncology 8(1): 60-6; EP 1162982; US 7700569 и US 7108844), и ингибиторы PDE5, такие как сиденафил (US 5250534, US 6469012 и ЕР 463756).
В предпочтительном воплощении способ по изобретению увеличивает время выживания у субъекта, подвергавшегося лечению, по сравнению с отсутствием лечения, например, по меньшей мере на 3 месяца. В качестве альтернативы, способ по изобретению генерирует ответ Т-клеток против указанной опухоли (ответ Т-клеток CD4+и/или CD8+).
Введения персонализированной противораковой вакцины и одной или более чем одной дополнительной противораковой терапии может осуществляться с интервалами, варьирующими от минут до недель. Например, можно предоставить субъекту рекомбинантный поксвирус и дополнительную противораковую терапию последовательно или вперемежку, но также рассматриваются сопутствующие введения обеих терапий в пределах того же самого периода времени. Ход лечения может быть определен практикующим врачом установившимся способом, и настоящим изобретением охватываются разные протоколы. Например, после хирургии и химио/лучевой терапии могут проводиться от 1 до 10 введений персонализированной противораковой вакцины.
Кроме того, после хода лечения рассматривается то, что имеется период времени, во время которого не вводится противораковое лечение перед повтором цикла(лов) лечения.
Все из процитированных выше раскрытий патентов, публикаций и записей в базах данных конкретно включаются в данный документ посредством ссылки во всей их полноте. Другие характеристики, цели и преимущества данного изобретения будут очевидными из описания и графических материалов, и из формулы изобретения. Следующие примеры включаются для демонстрации предпочтительных воплощений данного изобретения. Однако, в свете настоящего раскрытия, специалисты в данной области должны понимать то, что в конкретных воплощениях могут быть сделаны изменения, которые раскрываются без отступления от сущности и объема данного изобретения.
ПРИМЕРЫ
Материалы и методы
Конструкции, описанные ниже, получают согласно общим методикам генной инженерии и молекулярного клонирования, подробно описанным в Maniatis et al. (1989, Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor NY или последующие издания), или согласно рекомендациям изготовителя при использовании имеющегося в продаже набора. Методики ПЦР-амплификации известны специалисту в данной области (см., например, PCR protocols - A guide to methods and applications, 1990, опубликованную Innis, Gelfand, Sninsky and White, Academic Press).
Получение рекомбинантного MVA
Промоторы вируса осповакцины и синтетические гены, кодирующие разные слияния неоэпитопов, были синтезированы Geneart (Регенсбург, Германия). Последовательности оптимизировали в отношении человеческого применения кодонов, и добавляли последовательность Козака (АСС) перед кодоном инициации ATG. Кроме того, исключали некоторые мотивы: TTTTTNT, GGGGG, ССССС, которые являются вредными для экспрессии в поксвирусном векторе.
Получение многоэпитопных конструкций: MVATG 19022 и MVATG 19023
MVATG 19022 содержит экспрессионную кассету, кодирующую слияние 5 пептидов, происходящих из пяти разных антигенов: FCU1, b-gal, MUC1, Е7 HPV-16 и Е1 HPV-16. Все пептиды представляют собой 27-меры, содержащие эпитоп Т-клеток С57bI/6. Более конкретно, пентатопное слияние содержит от 5' до 3' пептид FCU1 (SEQ ID NO: 1), пептид b-Gal (SEQ ID NO: 2), пептид E1 HPV-16 (SEQ ID NO: 3), пептид E7 HPV-16 (SEQ ID NO: 4) и пептид MUC1 (SEQ ID NO: 5) с 10-мерными линкерами GS, присутствующими на N-конце данного слияния, между пептидами и после последнего пептида для оптимального процессинга данного слитого полипептида и презентации эпитопов. Для того чтобы облегчить выявление, добавляли на С-конец данного слияния последовательность метки - метки Flag (DYKDDDDK; SEQ ID NO: 6). Пентатопное слияние (SEQ ID NO: 7) помещали под контроль промотора р11К7.5 (SEQ ID NO: 8).
MVATG 19023 содержит экспрессионную кассету, кодирующую такое же пентатопное слияние, в котором на N-конец был добавлен сигнальный пептид, полученный из гликопротеина вируса бешенства (SEQ ID NO: 9). Данное модифицированное пентатопное слияние (SEQ ID NO: 10) помещали под контроль промотора р11К7.5 (SEQ ID NO: 8).
Фрагменты ДНК, соответствующие каждому пентатопному слиянию и окруженные последовательностями из примерно 30 п. о., гомологичными плазмиде для переноса вируса осповакцины, получали синтетическим путем. В плазмиду для переноса вируса осповакцины pTG18626, расщепленную NotI и Bg//II, вставляли фрагменты ДНК GeneArt®Strings™ посредством клонирования In-Fusion (набор для клонирования In-Fusion HD, Clontech), получая, соответственно, pTG19022 и pTG 19023.
Конструируется плазмида для переноса MVA - pTG 18626 - для обеспечения вставки нуклеотидной последовательности, подлежащей переносу, посредством гомологичной рекомбинации в делецию III генома MVA. Она происходит из плазмиды pUC18, в которую были клонированы фланкирующие последовательности (BRG3 и BRD3), окружающие делецию III MVA (Sutter and Moss, 1992, Proc. Natl. Acad. Sci. USA 89:10847).
Гомологичную рекомбинацию проводили с использованием гена, содержащего родительский MVA, кодирующего флуоресцентный белок mCherry в его делеции III (MVA mCherry). Преимуществом MVA mCherry является отличение клеток, которые инфицируются рекомбинантным вирусом, который успешно включил экспрессионную кассету, от клеток, которые инфицируются исходным первоначальным вирусом MVA mCherry (родительский вирус). В самом деле, ген mCherry удаляется в случае успешной рекомбинации экспрессионной кассеты в пределах делеции III, и вирусные бляшки оказываются белыми.
Хотя рекомбинация и является относительно частым событием в вирусе осповакцины, только 1-5% рекомбинантных бляшек содержат вставленную ДНК. Следовательно, для того чтобы увеличить эффективность гомологичной рекомбинации, добавили дополнительную стадию расщепления эндонуклеазой. Например, можно использовать систему CRISPR-CAS9 для специфичного получения двухнитевых разрывов в гене mCherry в MVA и, таким образом, для увеличения эффективности селекции рекомбинантного MVA (например, обычно 10-50% вирусных бляшек содержат экспрессионную кассету). При условии, что MVA реплицируется в цитоплазме, рекомендуется предоставлять фермент Cas9 без NLS (сигнал ядерной локализации) (плазмида pCas9), как описано в Yuan et al. (2015, J. Virol. doi:10.1128), наряду с пгРНК, содержащей гРНК, специфичную для последовательности mCherry.
Получение MVATG 19022 осуществляли гомологичной рекомбинацией в первичных фибробластах эмбрионов курицы (CEF). CEF выделяли из 11 или 12-суточных яиц с эмбрионами SPF (Charles River) и культивировали в культуральной среде, дополненной 4 мМ глутамином и содержащей гентамицин в конечной концентрации 40 мг/л. CEF культивировали при 37°С, 5% СO2 в течение 24 или 48 ч до инфицирования MVA. 108 клеток CEF инфицировали при MOI 0,02 родительским MVA, кодирующим mCherry, в течение 2 ч в другой культуральной среде, дополненной 2 мМ L-глутамином, и содержащей гентамицин в конечной концентрации 40 мг/л. Инфицированные клетки трипсинизировали, и 107 клеток трансфицировали нуклеофекцией 2 мкг pTG19022 (согласно технологии Аmаха Nucelofector) и, если необходимо, pCAS9 и пгРНК. Трансфицированные клетки переносили в 6-луночный планшет и инкубировали в течение 2 суток при 37°С. Инфицированные и трансфицированные клетки отбирали и использовали для выделения бляшек. Для инфицирования монослоев CEF на протяжении 30 минут при комнатной температуре использовали серийные разведения. 2×107 клеток CEF, высаженных в 60 мм культуральные чашки, инфицировали 500 мкл каждого разведения. Через 30 минут на монослой клеток наслаивали слой 5 мл культуральной среды, содержащей 1% агарозы. Культуральные чашки затем инкубировали при 37°С в атмосфере 5% СO2 на протяжении 96 ч. Выявление бляшек микроскопической проверкой облегчалось наслоением нового слоя (5 мл) культуральной среды, содержащего 1% агарозы и 0,008% нейтрального красного. Выбранные белые бляшки собирали, помещали в 500 мкл PBS (фосфатно-солевой буферный раствор) и замораживали при -20°С. Вирусную амплификацию проводили с использованием замороженных бляшек для инфицирования монослоя клеток CEF на протяжении 30 минут при комнатной температуре в 6-луночном культуральном планшете. Через 30 минут добавляли 2 мл культуральной среды, и инфицированные клетки инкубировали при 37°С в атмосфере 5% СO2. Через 72 ч собирали все содержимое чашки (амплификация 1). Возможно, вторую стадию амплификации можно осуществлять во флаконах F175. В данном случае 100 мкл амплифицированных бляшек разводили в 10 мл PBS и использовали для инфицирования свежих CEF в двух культуральных флаконах (F175, содержащий примерно 7×107 клеток) на протяжении 30 минут при комнатной температуре. Добавляли 20 мл культуральной среды, и инфицированные клетки инкубировали при 37°С в атмосфере 5% СO2 на протяжении 72 ч. Затем собирали амплификацию 2. Присутствие экспрессионной кассеты и отсутствие загрязнения родительским MVA подтверждали ПЦР. Весь данный способ мог быть осуществлен всего за три недели.
MVATG 19022 также может быть получен в большем масштабе для доклинических экспериментов с использованием нескольких флаконов F500. CEF сначала культивируются в подходящей среде, дополненной 4 мМ глутамином и содержащей гентамицин в конечной концентрации 40 мг/л, и их инкубировали на протяжении 48 ч до инфицирования при 37°С в 5% СO2. Клетки затем инфицировали объемом амплификации 2, как описано выше. Вирусную амплификацию проводили при температуре 37°С или меньше (например, 34-35°С) в 5% СO2 в течение от 72 ч до 96 ч. Инфицированные клетки и среду затем осаждали и замораживали. Неочищенный сбор разрушали с использованием высокоскоростного гомогенизатора (SILVERSON L4R) и подвергали способу очистки (например, как описано в WO 2007/147528). Вкратце, лизированный вирусный препарат может быть осветлен фильтрованием и очищен стадией фильтрования с тангенциальным потоком (TFF). Очищенный вирус ресуспендировали в подходящем буфере для приготовления вируса (например, 5% (масс./об.) сахарозы, 50 мМ NaCl, 10 мМ Tris/HCl, 10 мМ глутамат натрия, рН 8).
Получение вируса MVATG 19023 осуществляли в CEF посредством гомологичной рекомбинации, и его продуцировали, как описано выше.
Получение конструкций неопептидов СТ26: MVATG 19030 и MVATG 19038
MVATG 19030 содержит две экспрессионные кассеты, причем каждая кодирует слияние пяти неопептидов СТ26 (пентатопы), последовательности которых описываются в Kreiter et al. (2015, Nature, 520: 692-6). Для того чтобы облегчать выявление пентатопа, на С-конец каждого пентатопа добавляли последовательности метки, соответственно, метки Flag (DYKDDDDK; SEQ ID NO: 6) для пентатопа 1 и метки c-myc (EQKLISEEDL; SEQ ID NO: 11) для пентатопа 2. Меченый пентатоп 1 (SEQ ID NO: 12) помещали под контроль промотора р11К7.5 (SEQ ID NO: 8), тогда как меченый пентатоп 2 (SEQ ID NO: 13) помещали под контроль промотора pH5R (SEQ ID NO: 14).
MVATG19038 содержит две экспрессионные кассеты, кодирующие такие же пентатопные слияния, в которых на N-конец пентатопа 1 был добавлен сигнальный пептид гликопротеина вируса бешенства (SEQ ID NO: 9), а на N-конец пентатопа 2 -сигнальный пептид гликопротеина F вируса кори (SEQ ID NO: 15). Модифицированный пентатоп 1 (SEQ ID NO: 16) помещали под контроль промотора р11К7.5 (SEQ ID NO: 8), тогда как модифицированный пентатоп 2 (SEQ ID NO: 17) помещали под контроль промотора pH5R (SEQ ID NO: 14).
Фрагменты ДНК, соответствующие двум данным экспрессионным кассетам, разделенные подходящими рестрикционными сайтами и окруженные последовательностями из примерно 30 п. о., гомологичными плазмиде для переноса вируса осповакцины, получали синтетическим путем. После рестрикции SnaB1 в плазмиду для переноса вируса осповакцины pTG18626, расщепленную NotI и Bg/II, вставляли фрагменты клонирования In-Fusion (набор для клонирования In-Fusion HD, Clontech), получая, соответственно, pTG19030 и pTG19038.
Получение вирусов MVATG 19030 и MVATG 19038 осуществляли в CEF посредством гомологичной рекомбинации, как описано выше.
Получение конструкции человеческого неопептида: MVATG19111 MVATG19111 содержит три экспрессионные кассеты, которые все вместе кодируют восемнадцать человеческих неопептидов, распределенных в три слияния: первое слияние, содержащее 7 неопептидов (гептатоп), второе - 6 неопептидов (гексатоп) и третье - 5 неопептидов (пентатоп). Большинство из 18 закодированных неопептидов состояли из 29 аминокислот с мутацией в центральном положении, за исключением четырех из них: соответственно, 19-аминокислотного неопептида, имеющего мутацию, расположенную на его N-конце (NP1), неопептида, содержащего 21 аминокислоту с мутацией, расположенной на его С-конце (NP18), неопептида, содержащего 26 аминокислот, имеющего мутацию в центральном положении, как определено в данном документе, но с 3-аминокислотной делецией в С-концевой фланкирующей последовательности (NP10) и неопептида из 76 аминокислот, возникающего в результате мутации со сдвигом рамки считывания (NP15). В каждом слиянии данные неопептиды были разделены 5-аминокислотным линкером для оптимального процессинга слитого полипептида и презентации эпитопа. Использовали четыре разных линкера: GSGSG, SGSGS, GSTSG и SGTGS. Кодоны последовательности линкера были вырожденными для того, чтобы избегать повторения и ограничивать события рекомбинации во время процесса продукции поксвируса.
Для того чтобы облегчать выявление пептидных слияний, последовательности метки добавляли на С-конец каждого слияния, соответственно, метку НА (YPYDVPDYA; SEQ ID NO: 18) для гептатопа, метку HSV (QPELAPEDPED; SEQ ID NO: 19) для гексатопа и метку VSV (YTDIEMNRLGK; SEQ ID NO: 20) для пентатопа. На N-конец каждого слияния неопептидов добавляли сигнальный пептид для усиления процессинга через ЭР. Сигнальные пептиды, полученные из гликопротеина вируса бешенства (SEQ ID NO: 9), добавляли на N-конец гептатопа и пентатопа, а сигнальный пептид, полученный из гликопротеина F вируса кори (SEQ ID NO: 15) - на N-конец гексатопа.
Для контроля экспрессии слияний неопептидов использовали разные промоторы, соответственно, промотор pH5R (SEQ ID NO: 14), управляющий экспрессией гептатопа, промотор pB2R (SEQ ID NO: 21) - для экспрессии гексатопа, и промотор р11К7.5 (SEQ ID NO: 8) - для пентатопа.
Фрагмент ДНК, соответствующий трем экспрессионным кассетам, разделенным подходящими рестрикционными сайтами и окруженным последовательностями из примерно 350 п. о., гомологичными плазмиде для переноса вируса осповакцины, получали синтетическим путем, что приводило к PTG19111.
Получение вируса MVATG 19111 осуществляли в CEF посредством гомологичной рекомбинации, как описано выше.
Получение конструкции человеческого неопептида: pTG19247 Получали конструкцию плазмиды pTG19247, содержащую три экспрессионные кассеты, причем каждая кодирует десять неопептидов. Пул из 30 неопептидов отбирали на основе их прогнозируемой иммуногенности из общедоступного набора данных пациентов (исследование PRJEB3132, Секвенирование экзома образцов человеческой аденокарциномы легкого и их нормальных аналогов). Для иллюстративной цели приводится аминокислотная последовательность в SEQ ID NO: 60, 61 и 62. Каждая кассета содержит сигнальную последовательность (полученную из гликопротеина вируса бешенства и гликопротеина F вируса кори) и помещается под контроль промоторов pH5R, pB2R и p11k7.5 соответственно. Для облегчения конструирования слияний между отобранными неопептидами не включали линкеры. Получение соответствующего вируса проводили в CEF посредством гомологичной рекомбинации согласно способу, раскрытому выше. Однако получали низкое отношение белых бляшек меньше 5%, и анализы ПЦР белых бляшек выявили отсутствие образования рекомбинантного MVA, несущего слияния неопептидов (белые бляшки вероятно возникают из-за мутации в последовательности mCherry).
Экспрессия неопептидов и анализ вестерн-блоттингом
4×106 клеток CEF инфицировали при MOI 0,2 разными MVA, кодирующими слияния неопептидов, в 6-луночных планшетах. Пустой вектор MVATGN33.1 использовали в качестве негативного контроля. Через 24 часа среду отбрасывали, и клетки лизировали 300 мкл/чашку 2× буфера Тris-глицин-SDS (ссылка: LC2676; Novex), содержащего β-меркаптоэтанол (5% об./об.). Лизат затем обрабатывали ультразвуком и нагревали в течение 5 мин при 95°С. Двадцать микролитров клеточных лизатов подвергали электрофорезу в предварительно отлитом 4-15%-ном геле Criterion с использованием гелевой системы Criterion Precast (Biorad). После электрофореза белки переносили на PVDF (поливинилиденфторид) мембрану (система переноса Trans-blot® TurboTM (#170-4155, Biorad)). Иммуновыявление осуществляли с использованием антител против TAG, конъюгированных с HRP (пероксидаза хрена). На мембрану наносили разведение 1/2000 каждого из следующих антител: моноклональное антитело против FLAG М2, конъюгированное с HRP (Sigma, А8592), моноклональное антитело против метки НА, конъюгированное с HRP (Thermofisher, 26183-HRP), поликлональное антитело против метки VSV-G, конъюгированное с HRP (Thermofisher, РА1-26564), поликлональное антитело козы против метки HSV, конъюгированное с HRP (Abcam, 19392), тогда как использовали разведение 1/5000 моноклонального антитела против сМус, конъюгированного с HRP (Novex 46-0709). Иммунные комплексы выявляли с использованием реактива для вестерн-блоттинга Amersham ECL prime (GE Healthcare, RPN2236) и анализировали с использованием Molecular Imager ChemiDOC™XRS от BIO-RAD.
Анализ ELISpot (метод иммуноферментных пятен)
Подготовка чашек
Чашки (Millipore, MSIPS4W10) предобрабатывали 1 минуту 35%-ным этанолом (15 мкл/лунку) и пять раз промывали стерильной водой (200 мкл/лунку). Лунки покрывали 100 мкл 15 мкг/мл антитела против мышиного IFNγ (Mabtech, AN18, 3321-3-1000; разведенное в PBS) и инкубировали в течение ночи или в течение выходных при +4°С. В сутки эксперимента чашки пять раз промывали стерильным PBS (200 мкл/лунку) и насыщели в течение по меньшей мере 1 ч при 37°С полной средой (СМ) 200 мкл/лунку.
Получение образца
Анализы ELISpot ex vivo проводили с использованием свежих селезеночных лимфоцитов. Для каждого эксперимента селезенки от 5 животных каждой группы отбирали и объединяли. Селезенки отбирали в 4 мл полной среды (CM: RPMI 10% FCS (фетальная телячья сыворотка) или X-Vivo, если мыши несут опухоли) и затем продавливали поршнем шприца через 70 мкм клеточное сито в 6-луночном планшете. Полученную суспензию спленоцитов разводили в 2 раза в СМ, наслаивали над 4 мл среды для разделения клеток Lympholyte®-M (Cedarlane, ссылка: CL5035) и центрифугировали в течение 20 минут при 1500 g при комнатной температуре. Отбирали лимфоциты в интерфазе и дважды промывали в 10 мл среды RPMI1640 (центрифугирование 5 минут при 400 g при комнатной температуре). Лимфоциты ресуспендировали в 2 мл лизисного буфера RBC (BD PharmLyse, BD Bioscience, ссылка 555899) и инкубировали в течение 5-15 минут при комнатной температуре для лизирования эритроцитов (RBC). После одной стадии промывки в СМ (центрифугирование 5 минут при 400 g при комнатной температуре) лимфоциты ресуспендировали в 1 мл СМ и подсчитывали с использованием «счетчика частиц и анализатора размера Z2 Coulter» Beckman Coulter. Концентрацию клеток корректировали до 1×107 клеток на мл с использованием СМ.
Анализ
Во-первых, насыщающую среду удаляли посредством опустошения планшетов, и добавляли во все лунки 50 мкл СМ. Добавляли в релевантные лунки (согласно ранее определенной схеме пипетирования; каждое условие тестировали в четверной повторности) 50 мкл каждого пептида или пул пептидов, в концентрации 4 мкг/мл (т.е. для пула концентрация каждого пептида составляла 0,4 мкг/мл). В качестве позитивного контроля, добавляли 50 мкл 20 мкг/мл конкавалина А (СоnА). Во-вторых, в лунки добавляли 100 мкл каждой суспензии лимфоцитов (1×106 клеток) (за исключением лунок T8V, где добавляли только 3×105 клеток), и планшеты инкубировали в течение от 18 ч до 20 ч при 37°С в 5% СO2.
Клетки удаляли посредством опустошения планшетов, планшеты промывали 5 раз PBS (200 мкл/лунку), и распределяли во все лунки 100 мкл биотинилированного моноклонального антитела против мышиного IFNγ (Mabtech, R4-6A2, 3321-6-1000; конечная концентрация в PBS с 0,5% FCS - 1 мкг/мл). Планшеты инкубировали в течение 2 часов при комнатной температуре, затем пять раз промывали PBS, и добавляли в каждую лунку экстравидин-щелочную фосфатазу (SIGMA, Е236, 1/5000е в PBS с 0,5% FCS) (100 мкл/лунку). Планшеты инкубировали 1 ч при комнатной температуре, пять раз промывали PBS, и распределяли в каждую лунку 100 мкл раствора субстрата BClP/NBT (таблетки BClP/NBT, SIGMA, В5655; профильтрованные через 0,45 мкМ). Планшеты инкубировали при комнатной температуре, в темноте, пока в позитивных лунках не были видны различимые пятна (примерно через 5-10 минут). Развитие окраски останавливали опустошением планшетов и обильными промывками водопроводной водой. Планшеты оставляли в темноте без крышки при комнатной температуре, пока они не высыхали (по меньшей мере 1 ч).
Получение данных
Пятна подсчитывали с использованием ридера ELISpot (ридер CTL Immunospot, S5UV). Осуществляли контроль качества для каждой лунки для обеспечения того, что количества, предоставленные ридером ELISpot, соответствуют реальности картины. Результаты для каждой четверной повторности выражали как среднее число пятнообразующих единиц (sfu) на 1×106 лимфоцитов селезенки. Позитивность определяли, как описано в Moodie et al. (2006, J. Immunol Methods 315: 121-32) и Moodie et al. (2010, Cancer Immunol Immunother doi 10.1007/s00262-010-0875-4).
ПРИМЕР 1: иммунизация мультипептидной конструкцией
Конструирование и получение вектора
Как проиллюстрировано на Фиг. 1, две конструкции MVA: MVATG 19022 и MVATG 19023 получали для экспрессии слияния 5 пептидов, происходящих из разных антигенов, разделенных друг от друга 10-аминокислотными линкерами GS для оптимального процессинга слитого полипептида и презентации эпитопа. Каждый пептид представляет собой 27-мер и содержит подтвержденный Т-клеточный эпитоп С57bI/6. Последовательность, кодирующую пентатоп, помещают под контроль промотора p11k7.5 (SEQ ID NO: 8), и данную кассету вставляют в делецию III генома MVA. Более конкретно, пентатопное слияние содержит от 5' до 3' пептид, происходящий из FCU1 (SEQ ID NO: 1), пептид, происходящий из b-Gal (SEQ ID NO: 2), пептид, происходящий из Е1 HPV-16 (SEQ ID NO: 3), пептид, происходящий из Е7 HPV-16 (SEQ ID NO: 4), и пептид, происходящий из MUC1 (SEQ ID NO: 5), с 10-мерными GS линкерами, находящимися до пептида FCU1, между пептидами и после пептида MUC1. Метка Flag (SEQ ID NO: 6) присутствует на С-конце для того, чтобы облегчать оценку экспрессии. Кроме того, MVATG 19023 отличается от MVATG19022 присутствием сигнального пептида на его N-конце. Для иллюстративных целей пентатопное слияние содержит аминокислотную последовательность, как изложено в SEQ ID NO: 7 для MVATG19022 и как изложено в SEQ ID NO: 10 для MVATG19023 (с сигнальным пептидом).
Оценка экспрессии пептида
Экспрессию слияния пептидов оценивали посредством вестерн-блоттинга в CEF, инфицированных каждым вектором, с использованием антител, направленных на пептид метки Flag, включенный в экспрессионную кассету. Вкратце, CEF инфицировали или нет (имитирующий контроль) при MOI 0,2 с использованием MVATG 19022 или MVATG 19023, или пустым (т.е. нерекомбинантным) MVA (TGN33.1 в качестве негативного контроля). Клетки отбирали через 24 ч, и лизаты клеток анализировали SDS-PAGE. Иммуновыявление продуктов экспрессии осуществляли с использованием антитела против Flag. Результаты показывают то, что была выявлена полоса, соответствующая ожидаемому размеру пентатопных слияний, направляемому MVATG19022 (19,7 кДа) и MVATG19023 (22,2 кДа). Как и ожидалось, в лизатах клеток, полученных из клеток, инфицированных либо пустым MVA, либо имитирующим буфером, не были выявлены полосы.
Кроме того, наблюдали более высокие уровни экспрессии при оснащении данной кассеты пептидным сигналом, что подтверждается более интенсивной полосой, полученной с MVATG 19023 по сравнению с полосой, выявленной в клетках, инфицированных MVATG 19022.
Оценка иммуногенности
Способность этих двух векторов, экспрессирующих пентатоп, генерировать ответ Т-клеток анализировали у мышей С57bI/6 посредством ELIspot на IFNγ и сравнивали ответы, продуцированные векторами MVA, кодирующими полноразмерный антиген, соответственно, MVATG9931 (MUC-1), MVATG15637 (FCU-1), MVATG18124 (β-галактозидаза), MVATG8042 (Е7 HPV-16) и MVATG17409 (Е1 HPV-16).
Более конкретно, MVATG9931 (TG4010 с его клиническим названием) представляет собой рекомбинантный MVA, кодирующий опухолеассоциированный антиген MUC1 и человеческий интерлейкин 2 (IL-2). TG4010 в комбинации с химиотерапией стандарта помощи первой линии продемонстрировал эффективность при запущенном метастатическом немелкоклеточном раке легкого (NSCLC) в двух разных рандомизированных и контролируемых клинических испытаниях фазы 26 (Quoix et al., 2011, The Lancet Oncol 12(12): 1125-33). MVATG15637 представляет собой рекомбинантный MVA, кодирующий FCU1 дрожжевого происхождения, экспрессирующий дрожжевые ферменты ЦДазу и УФРТазу, которые превращают пролекарство фторцитозин (5-ФЦ) в цитотоксический 5-фторурацил (5-ФУ) и 5-фторуридин-5'-монофосфат (5-ФУМФ) соответственно (Husseini et al., 2017, Ann Oncol 28(1): 169-74). MVATG18124 содержит бактериальный ген LacZ, кодирующий бета-галактозидазный антиген под контролем поксвирусного промотора pH5R. MVATG8042 (TG4001 с его клиническим названием) кодирует инактивированные мутацией онкопротеины Е6 и Е7 вируса человеческой папилломы (HPV) 16 и человеческий интерлейкин-2 (WO 99/03885). MVATG17409 кодирует антиген Е1 HPV-16, модифицированный для отмены репликативной функции природного полипептида (WO 2008/092854).
Вкратце, пяти мышам С57bI/6 на группу подкожно инъецировали дважды с однонедельным интервалом (сутки 0 и 7) соответствующий MVA. Для каждой инъекции в бок животных вводили 2,5×107 БОЕ MVA (100 мкл в следующем буфере: 10 мМ Tris [трис(гидроксиметил)аминометан/НСl], 5% сахароза (масс./об.), 10 мМ глутамат натрия, 50 мМ хлорид натрия, рН 8). Через одну неделю после последней инъекции (сутки 14) всех мышей умерщвляли смещением шейных позвонков. Отбирали селезенки для выделения лимфоцитов, и Т-клетки, продуцирующие IFNγ, количественно измеряли посредством Elispot (метод иммуноферментных пятен, специфичный в отношении цитокинов), как описано в Материалах и методах. Стимуляцию проводили с использованием следующих пептидов, специфичных в отношении каждого экспрессируемого антигена/пептида:

И в качестве контролей:
T8V (специфичный в отношении MVA): TSYKFESV (позитивный контроль) (SEQ ID NO: 55)
K9i-3 (RMA, иррелевантный): KNGENAQAI (SEQ ID NO: 56)
G15H (РуМТ, иррелевантный): GICLMLFILIKRSRH (SEQ ID NO: 57)
Как проиллюстрировано на Фиг. 2А-Д, сильный клеточный ответ выявляли для 4 из 5 эпитопов, экспрессируемых MVATG 19022 и MVATG 19023 (проблемы в технической сфере затрудняют интерпретацию ответа MUC-1). За исключением ответа против Е1, MVATG 19023 имеет тенденцию давать лучший ответ, чем MVATG19022. Можно предполагать то, что адресование слияния пептидов в эндоплазматический ретикулум вероятно защищает экспрессированное слияние от деградации клеточной протеасомой и стимулирует его накопление в инфицированных клетках, которые фагоцитируются профессиональными антигенпрезентирующими клетками, такими как DC (дендритная клетка).
Важно то, что ответ IFNγ, индуцированный после иммунизации MVATG19023, находится на уровнях, сравнимых с уровнем, предоставляемым полноразмерным антигеном, и даже лучше для ответа против Е7 (Фиг. 2В). Следовательно, непосредственное соседство пептидов в остове MVA обеспечивает развитие диверсифицированного иммунного ответа без наблюдения явления иммунодоминантности, уменьшающего данные ответы.
ПРИМЕР 2: иммунизация конструкцией неопептида СТ26 Отбор неопептидов СТ26
Десять мутировавших неопептидов СТ26 отбирали на экспрессию в конструкциях MVA (аминокислотные последовательности приводятся в SEQ ID NO: 22-31) на основе разных критериев, описанных в данной области, таких как низкий балл для связывания с МНС класса I, присутствие эпитопов Т-клеток CD8+ (Kreiter et al., 2015, Nature 520 (7549): 692-6) и присутствие мутации одного остатка в геноме соответствующей линии клеток опухоли СТ26. Каждый неопептид представляет собой 27-мер, содержащий мутацию в центральном положении (положении 14) и отделенный от следующего 10-аминокислотным CS линкером для оптимального процессинга и презентации эпитопа.
Конструирование и получение вектора
Две конструкции MVA: MVATG 19030 и MVATG 19038 соответственно получали для экспрессии вышеописанных неопептидов СТ26, организованных в два слияния (пентатопы). Первое пентатопное слияние помещается под транскрипционный контроль промотора р11К7.5 (SEQ ID NO: 8), а второе - под контроль pH5R (SEQ ID NO: 14). Метка Flag (SEQ ID NO: 6) присутствует на С-конце первого пентатопного слияния, а метка c-MYC - (SEQ ID NO: 11) - на конце второго слияния. MVATG19038 отличается от MVATG19030 присутствием сигнальных пептидов, происходящих из гликопротеина вируса бешенства (SEQ ID NO: 9) на N-конце первого пентатопного слияния и из гликопротеина F вируса кори (SEQ ID NO: 15) на N-конце второго пентатопного слияния. Две данные кассеты слияния неопептидов были вставлены в делецию III генома MVA. В MVATG19030 первое и второе пентатопные слияния содержат, соответственно, аминокислотную последовательность, как изложено в SEQ ID NO: 12 и SEQ ID NO: 13 (также проиллюстрированных на Фиг. 3), тогда как MVATG19038 содержит аминокислотную последовательность, как изложено в SEQ ID NO: 16 и SEQ ID NO: 17.
Оценка экспрессии неопептида СТ26
Экспрессию слияний неопептидов СТ26 оценивали посредством вестерн-блоттинга в CEF, инфицированных MVATG19030 и MVATG19038. Вкратце, CEF инфицировали или нет (имитирующий контроль) при MOI 0,2 с использованием MVATG19030 или MVATG19038, или пустым (т.е. нерекомбинантным) MVA (TGN33.1 в качестве негативного контроля). Клетки отбирали через 24 ч, и лизаты клеток анализировали SDS-PAGE. Иммуновыявление продуктов экспрессии, включенных в экспрессионные кассеты, осуществляли с использованием антител, направленных на метки Flag и c-MYC. Антителом против MYC были выявлены неспецифичные полосы, но они также присутствовали в негативных контролях. Каким бы ни был фон, пентатопы были выявлены обоими антителами, соответствуя ожидаемому размеру слияний неопептидов, управляемых MVATG 19030 (20,6 кДа для обоих пентатопов) и MVATG 19038 (23,1 кДа для первого пентатопа и 23,5 кДа для второго пентатопа). Как и ожидалось, в лизатах клеток, полученных из клеток, инфицированных пустым MVA или имитирующим буфером, не были выявлены специфические полосы (хотя полосы фона и присутствовали в образцах, обработанных антителом против cMYC, как упомянуто выше).
Следовательно, данное исследование подтверждает экспрессию двух слияний неопептидов в инфицированных клетках. Как и в Примере 1, присутствие пептидного сигнала положительно влияло на экспрессию, и наблюдали более высокие уровни для слияний неопептидов, оснащенных пептидными сигналами, что подтверждается более интенсивными полосами, полученными с MVATG19038 по сравнению с полосами, выявленными в клетках, инфицированных MVATG19030.
Оценка иммуногенности
Способность MVATG 19030 и MVATG 19038 генерировать ответ Т-клеток против закодированных неопептидов СТ26 анализировали в опухолевой модели СТ26 мышей Balb/c посредством ELIspot на IFNγ. Обрабатывали пять животных на группу. Одной половине мышей Balb/c внутривенно инъецировали в хвостовую вену в сутки 0 опухолевые клетки СТ-26 MUC1 (2×105 клеток в 100 мкл) (группы 1-5), тогда как остающуюся половину животных (группы 6-11) оставляли необработанной. Инъекции MVA осуществляли в сутки 2 и 9 внутривенным путем с использованием 1×107 БОЕ в 100 мкл ранее описанного буфера для препарата вируса. Через одну неделю после последней иммунизации MVA (сутки 16) мышей умерщвляли. Отбирали селезенки для выделения лимфоцитов и обрабатывали, как подробно описано в Материале и методах. Стимуляцию проводили с использованием либо мутировавших неопептидов СТ26 (SEQ ID NO: 22-31), либо их немутировавших гомологов (пептиды дикого типа, описанные в SEQ ID NO: 32-41), или с использованием пептидов CD8+ СТ26 либо мутировавших (описанных в SEQ ID NO: 42-45), либо дикого типа (в SEQ ID NO: 46-49). Пептиды СТ26 дикого типа объединяли для уменьшения числа образцов для манипуляции, тогда как мутировавшие пептиды СТ26 использовали либо индивидуально, либо в пептидных пулах.
Как проиллюстрировано на Фиг. 4, после иммунизации векторами MVATG 19030 и MVATG19038 и стимуляции пулом собранных десяти мутировавших пептидов СТ26 сильный ответ IFNγ получался независимо от инъецированного вектора и животной модели (примированной клетками СТ26-MUC1 или нет). В отличие от этого, стимуляция пулом из десяти пептидов СТ26 дикого типа, а также средой негативного контроля не дает какой-либо продукции INFγ.
На Фиг. 5 проиллюстрирован ответ INFγ, генерированный против одного из мутировавших пептидов (EH27m; SEQ ID NO: 30) после иммунизации MVATG19030 и MVATG19038 как в мышиной модели, примированной, так и не примированной клетками CT26-MUC1. В обоих случаях (примированные или нет) генерируется ответ, варьирующий от 30 до 60 пятен, тогда как, в отличие от этого, стимуляция пептидом дикого типа (PE27WT SEQ ID NO: 37) не генерирует какой-либо продукции INFγ (меньше 5 пятен). Следует отметить то, что ответ, генерированный вектором MVA, оснащенным сигнальным пептидом (MVATG19038) слегка выше, чем ответ, полученный с MVATG19030, слияния которого не включают какого-либо сигнального пептида. Здесь опять не выявляется ответ (меньше 5 пятен) со средой негативного контроля.
Кроме того, ответ против мутировавшего пептида (ЕН27m) ингибируется после блокировки антителами против мышиного МНСII, как показано на Фиг. 6, что может показывать то, что данный ответ принадлежит к CD4+-типу.
В заключение, данные результаты поддерживают осуществимость получения персонализированной противораковой вакцины для человеческой противораковой терапии.
ПРИМЕР 3: конструкция человеческого неопептида
Идентификация и отбор неопептидов
Полноэкзомное секвенирование (WES) образцов опухоли и зародышевой линии пациента, страдающего от NSCLC, проводили парно-концевым способом (2×150 п. о.) для идентификации опухолеспецифичных соматических мутаций. Для информации, необработанные данные предоставляются службой секвенирования в формате стандартного файла, именуемого FASTQ. Каждый цикл секвенирования приводит к формированию файла FASTQ, который дает короткие считываемые последовательности и балл качества в расчете на основание - балл Phred. Он основывается на оценочной вероятности ошибки во время стадии распознавания оснований (Cock et al., 2010, Nucl. Acid Res. 38(6): 1767-71). При парно-концевом секвенировании формируются два файла FASTQ, причем каждый соответствует концу фрагмента библиотеки. Образующиеся файлы, таким образом, должны оставаться парными во время стадии фильтрования на качество, т.е. два считываемых фрагмента пары должны были пройти через фильтр. Критерии, оцениваемые во время данных стадий фильтрования, главным образом, основываются на баллах Phred, особенно в 3'-конце последовательности, который часто требует подрезания для сохранения точной информации для стадии распознавания варианта (Edgar and Flyvbjerg, 2015, Bioinformatics 31 (21):3476-82).
Короткие считываемые фрагменты затем выравниваются относительно эталонного генома (Li et al., 2009, Bioinformatics 25(16): 2078-79). После этой стадии картирования используются алгоритмы распознавания вариантов для выявления множества структурных вариаций, таких как однонуклеотидные варианты (SNV, включающие молчащие и немолчащие миссенс-мутаций) или маленькие вставки и делеции (инсерционно-делеционные полиморфизмы) между эталонным геномом и последовательностями пациента (Danecek P. et al., 2011, Bioinformatics 27(15): 2156-58; Nielsen et al., 2011, Nature Reviews Genetics 12(6): 443-51). Образцы из опухолевой и контрольной ткани обрабатываются в то же самое время для выявления опухолеспецифичных мутаций.
Данные мутации затем оцениваются на уровне белка для конструирования 17-мерных пептидов с мутацией в центре, когда возможно, согласно положению мутации. Данные пептиды-кандидаты затем оцениваются на уровне экспрессии с использованием данных транскриптома опухоли (Hundal et al., 2016, Genome Medecine 8(1): 11). Транскриптом опухоли измеряли посредством секвенирования РНК (RNA-Seq) в парно-концевом режиме (2×150 п. о.) для оценки экспрессии неоантигенов. Из-за чувствительности RNA-Seq, ген считался экспрессируемым при достижении глубины покрытия считываемых фрагментов выше Зх. Стадия количественного измерения начинается после картирования посредством подсчета числа считываемых фрагментов, картируемых на конкретном транскрипте, с корректировкой на размер библиотеки в общем числе картированных считываемых фрагментов на миллион (Wang et al., 2009, Nature Reviews Genetics 10(1): 57-63).
Таким образом, всего идентифицировали 20 транскриптов (соответствующих 18 уникальным генам), несущих по меньшей мере одну опухолеспецифичную мутацию. На этой основе отобрали 18 опухолеспецифичных мутаций, и сконструировали 18 неопептидов.
Данные 18 неопептидов имеют характеристики, описанные в следующей
Из 18 отобранных неопептидов:
• Все имеют размер от 19 до 76.
• 17 из 18 (94%) основываются на миссенс-мутаций, приводящей к замене одной аминокислоты пептида дикого типа, и 1 из 18 (6%) имеет мутацию на основе сдвига рамки считывания.
Среди неопептидов, несущих миссенс-мутацию:
ο они имеют размер от 19 до 29 аминокислот,
ο 15 из 17 (88%) имеют центральную мутацию, как определено в данном документе, и
ο 14 из 17 (83%) имеют размер 29 аминокислот и центральную мутацию.
Конструирование и продукция вектора
Для каждого пептида проводили поиск трансмембранного домена и сигнального пептида с использованием описания Uniprot соответствующего гена как источника информации. Ни для одного из данных неопептидов не было известно, что он является частью либо трансмембранного, либо сигнального пептида в его соответствующем белке. С использованием Uniprot искали известные функции, ассоциированные с данными пептидами. Пептиды были либо из известного домена, либо слишком короткими для образования функционального домена. Сконструировали три экспрессионные кассеты, содержащие 7 неопептидов (гептатоп), 6 неопетидов (гексатоп) и 5 неопептидов (пентатоп), с использованием следующего правила:
• 19-мерный неопептид NP1, который содержит миссенс-мутацию, расположенную на N-конце неоантигена, располагали на N-конце экспрессионной кассеты гептатопа.
• В то же время, неопептид NP18, который располагается в природе на С-конце неоантигена, располагался на С-конце экспрессионной кассеты, кодирующей пентатоп.
• Неопептид NP10 был укорочен на его последние 3 остатка из-за их потенциального участия в образовании дисульфидной связи и был помещен в середину кассеты гексатопа для отсутствия предрасположенности к какому-либо сворачиванию.
• Наконец, посредством программы Geneious спрогнозировали то, что неопептид NP13 образует потенциальную трансмембранную спираль, и, следовательно, он был помещен в конец кассеты, кодирующей гексатоп.
Данные неопептиды были разделены 5-аминокислотным спейсером для оптимального процессинга слитого полипептида и презентации эпитопа. Использовали четыре разных спейсера: GSGSG, SGSGS, GSTSG и SGTGS. Для аминокислотных последовательностей, а также последовательностей кодонов варьировали порядок для уменьшения процентной доли идентичности нуклеотидов между разными линкерами для уменьшения риска гомологичной рекомбинации между 17 линкерами, используемыми в данной конструкции. Сигнальные пептиды сливали с N-концом каждого слияния неопептидов для адресования данного пептида в ЭР. Сигнальные пептиды, полученные из гликопротеина вируса бешенства, добавляли на N-конец гептатопа и пентатопа, тогда как сигнальный пептид, происходящий из гликопротеина F вируса кори, добавляли на N-конец гексатопа.
Для того чтобы выявить экспрессию данных трех кассет, на С-конец каждой конструкции добавляли метки вирусного происхождения (т.е. метки НА (SEQ ID NO: 18), HSV (SEQ ID NO: 19) и VSV (SEQ ID NO: 20) для гептатопа, гексатопа и пентатопа соответственно).
Три данные кассеты слияния неопептидов вставляли в делецию III генома MVA, давая MVATG 19111. Более конкретно, первая кассета кодировала гептатопное слияние из 255 остатков, вторая кассета - гексатопное слияние из 239 остатков, и третья кассета - пентатопное слияние из 243 остатков. MVATG19111 продуцировался гомологичной рекомбинацией в CEF согласно способу, описанному в Материале и методах. Этот оптимизированный протокол позволил генерировать рекомбинантный MVA только за три недели и продуцировать очищенное большое количество еще за неделю.
Оценка экспрессии
Экспрессию слияний неопептидов пациента оценивали посредством вестерн-блоттинга в CEF, инфицированных MVATG19111. Вкратце, CEF инфицировали или нет (имитирующий контроль) при MOI 0,2 с использованием MVATG19111, или пустым (т.е. нерекомбинантным) MVA (TGN33.1 в качестве негативного контроля). Клетки отбирали через 24 ч, и лизаты клеток анализировали SDS-PAGE. Иммуновыявление продуктов экспрессии осуществляли с использованием антител, направленных на метки НА, HSV и VSV, включенные в экспрессионные кассеты. Результаты показывают то, что была выявлена специфическая полоса для каждого слияния неопептидов, управляемого MVATG19111. Кажущиеся размеры данных полос были слегка выше, чем ожидаемые размеры для каждого слияния: примерно 38 кДа для гептатопа (ожидаемый размер: 28,2 кДа), примерно 30 кДа для гексатопа (ожидаемый размер: 25,5 кДа) и примерно 36 кДа для пентатопа (ожидаемый размер: 25,9 кДа). Данное различие могло быть обусловлено посттрансляционными модификациями. Как и ожидалось, в лизатах клеток, полученных из клеток, инфицированных пустым MVA, либо имитирующим буфером, не были выявлены специфические полосы.
Оценка иммуногенности
По конструкции MVATG19111 нацелен на человеческие антигены, образующиеся в результате опухолеспецифичных мутаций. Для оценки иммуногенности данной вакцины, следовательно, требуется применение гуманизированной мышиной модели. Мышиная модель трансгенных мышей с человеческим лейкоцитарным антигеном (HLA)-A2.1 (HHD) предлагает возможность получения специфичных иммунных ответов на гаплотип А02*01 человеческого HLA (Firat et al. 1999, European Journal of Immunology 29(10): 3112-21). Кроме того, в данной мыши нарушена экспрессия мышиной молекулы HLA, предотвращая развитие мышиного специфичного иммунного ответа. Для оценки иммуногенности вакцины использовали мышей HHD. Мыши получали 1×107 БОЕ вакцины MVATG19111 или MVA TGN33.1 в 200 мкл фосфатно-солевого буферного раствора (PBS) посредством внутривенной инъекции в сутки 1 и сутки 7. Животных умерщвляли в С14 (сутки 14), отбирали спленоциты и замораживали до анализа. Число клеток, продуцирующих IFN-γ, определяли в ELISpot на IFN-γ.
Вкратце, суспензии одиночных клеток селезенок от мышей, вакцинированных MVATG19111 или контрольным вирусом MVATGN33.1, культивировали на планшетах ELISPOT в количестве 5×105 клеток/лунку в 200 мкл полной среды в течение 48 часов при разных условиях стимуляции. Условия стимуляции включали позитивный контроль (смесь пептидов MVA), негативный контроль (5 мкг/мл неродственного пептида), индивидуальные пептиды или один из 18 пулов пептидов, каждый из которых связан с одним из антигенов, экспрессируемых в вакцинах (5 мкг/мл перекрывающихся 20-мерных пептидов), или перекрывающиеся пептиды, соответствующие немутировавшему человеческому белку. После инкубации планшеты промывали, и клетки, продуцирующие IFN-γ, выявляли с использованием колориметрического способа, а пятна подсчитывали с использованием имеющейся в продаже автоматической программы. Иммунные ответы считали позитивными, когда число пятен было статистически отличным от аналитического фона.
У животного, получающего вакцину MVATG19111, 7 из 18 условий давали положительные иммунные ответы. Более конкретно, 4 условия были высокоиммуногенными и индуцировали выявляемые ответы Т-клеток у всех животных, тогда как 3 других были иммуногенными только у некоторых животных. Различия в иммуногенности могут быть связаны с чувствительностью анализа или вариабельностью иммунных фенотипов. Ответы были ограничены мутировавшим белком, и вакцина не вызывала иммунизацию против немутировавшего белка. У контрольного животного, получающего контрольный вирус MVATGN33.1, не наблюдался ответ.
Кроме того, ELISPOT проводили в отношении высокоиммуногенных пептидов в присутствии антитела против МНС класса II или после элиминации клеток CD8 с использованием элиминирующих шариков с антителом против CD8, как описано ранее (Puliaev et al., 2004, J. Immunol. 173(2): 910-9), для того, чтобы охарактеризовать фенотип реагирующих Т-клеток. Ответы наблюдали как после элиминации CD8, так и после блокады МСН II, демонстрируя то, что MVATG19111 могла индуцировать ответы, опосредованные и классом I (CD8), и классом II (CD4).
Продукция и терапевтическое применение
Продуцированный основной объем MVATG19111 может распределяться в индивидуальных дозах приблизительно 108 БОЕ, которые хранятся при -80°С или -20°С перед введением пациенту, от которого была получена опухоль.
ПРИМЕР 4: роль линкеров в слияниях неоэпитопов
4.1 Влияние линкеров на иммуногенность
Влияние линкеров сначала исследовали, начиная с вектора MVATG19023, описанного в Примере 1, посредством варьирования длины и последовательности линкеров. MVATG19023 кодирует слияние 5 антигенных пептидов, образующихся из FCU1, β-галактозидазы, Е1 HPV-16, Е7 HPV-16 и MUC1 (от N до С-конца данного слияния). Каждый пептид представлял собой 27-мер, содержащий подтвержденный эпитоп Т-клетки С57bI/6 (гаплотип b), расположенный в центре 27-мера, за исключением Mud, эпитоп которого соответствует 15 последним остаткам С-конца данного антигена (см. Таблицу 3). Линкеры длиной в десять остатков (5 раз GS) присутствовали на N- и С-концах каждого пептида, а также на N-конце слияния присутствовал сигнальный пептид вируса бешенства, а метка Flag - на его С-конце. Данное слияние помещали под контроль промотора р11к7.5 и вставляли в делецию III MVA.


В MVATG 19023 такой же линкер из 10 остатков присутствовал до и после каждого эпитопа. Для того чтобы обойти события гомологичной рекомбинации, которые могут происходить и приводить к частичной или полной делеции кассеты, рекомендуется вырождать последовательность ДНК каждого линкера, используя преимущество вырожденности генетического кода. Однако данное вырождение последовательности ограничивается числом, природой и длиной линкеров. В случае большого числа эпитопов, подлежащих векторизации (например, 10 или больше), генетический код не позволяет создавать достаточное разнообразие ДНК для того, чтобы избегать гомологичной рекомбинации с линкером GS пять раз. Следовательно, исследовали возможность применения меньших линкеров (GSG), остатков, отличных от серина (GAS или GTS), и, в конечном счете, полного отсутствия линкера. Вирусы продуцировали на СЕР. После осветления вирусы очищали посредством фильтрования тангенциальным потоком (TFF). Это приводит к получению MVATG190158, MVATG190159 и MVATG190157, проиллюстрированных в Таблице 4.
Иммуногенность, индуцированную данными 3 конструкциями, оценивали посредством ELISPOT и сравнивали с иммуногенностью, задаваемой MVATG 19023 в таких же условиях иммунизации, что и описанные ранее.
Результаты ELISPOT показывают то, что иммунные ответы были грубо аналогичными для MVATG19023, MVATG19158, MVATG19159 и MVATG19157 для 4 эпитопов из 5 (L15L3, I8L, I8V и R9F), тогда как ответ на EG15 имел тенденцию быть немного выше для MVATG19023, содержащего линкер из 10 остатков.
Данные результаты показывают то, что присутствие, природа и длина линкеров на N- и С-концах разных антигенных пептидов, подлежащих экспрессии, не имеет большого влияния на иммуногенность таких пептидов.
4.2 Влияние линкеров на образование рекомбинантного поксвируса
Присутствие или отсутствие линкеров также анализировали в отношении способности конструкций генерировать рекомбинантные поксвирусы в контексте вектора pTG19111 (описанного в Примере 3). Генерировали дополнительную плазмиду, содержащую такие же слияния неопептидов, что и pTG19111, но без каких-либо линкеров, получая pTG19264. Сравнивали получение рекомбинантных вирусов MVA между конструкциями без линкера (pTG 19264) и с линкером из 5 ак (аминокислота) (pTG19111) в трансфицированных клетках CEF, как описано в данном документе, причем белые бляшки подтверждают потенциальный рекомбинантный MVA, а красные бляшки - родительский (нерекомбинантный) вирус.
Как показано в Таблице 5, несмотря на то, что обе конструкции приводили в получению белых бляшек, их доля, тем не менее, была выше с вектором pTG19111, содержащим линкер.
В целом, данные результаты показывают то, что отсутствие линкеров не имеет большого влияния на иммуногенность, хотя и отрицательно изменяет долю белых бляшек, генерированных в клетках CEF, и, таким образом, получение рекомбинантного MVA. Следовательно, при необходимости, линкеры могут быть опущены для облегчения конструкции векторых кнструкций и, таким образом, уменьшения риска вредных событий гомологичной рекомбинации, особенно в конструкциях, где рассматривается экспрессия больше, чем 10 неопептидов, но с риском уменьшения доли генерированных рекомбинантных поксвирусов.
ПРИМЕР 5: анализ гидрофобности
5.1 Прогнозирование ТМ сегментов
Присутствие внутреннего или промежуточного ТМ сегмента тестировали в контексте плазмиды pTG 19247. Как упомянуто выше, pTG 19247 несет три экспрессионные кассеты, причем каждая кодирует десять неопептидов (аминокислотные последовательности которых описываются в SEQ ID NO: 60, 61 и 62), возникающих из человеческой аденокарциномы легкого (исследование PRJEB3132), с сигнальной последовательностью на N-конце каждого слияния. Для облегчения конструирования, между выбранными неопептидами не включали линкеры. Получение соответствующего MVA оценивали в CEF посредством гомологичной рекомбинации согласно способу, раскрытому выше, и подсчитывали число белых относительно красных бляшек. Было получено низкое отношение белых бляшек - меньше 5%, и анализ ПЦР белых бляшек выявил отсутствие рекомбинантного MVA, несущего слияния неопептидов.
Прогнозирование последовательностей ТМ в каждом слиянии неопептидов проводилось с использованием алгоритма прогнозирования ТМНММ. В слиянии 1 не могла быть спрогнозирована ТМ последовательность. Однако два потенциальных межпептидных ТМ сегмента были идентифицированы в слиянии 2, соответственно, в соединении неопептида 1 и неопептида 2, и в соединении неопептида 7 и неопептида 8. С другой стороны, три внутрипептидных ТМ сегмента было идентифицировано в слиянии 3, более конкретно, в пределах неопептидов 2, 4 и 5.
Затем была получена конструкция без ТМ, названная pTG19258, содержащая такое же слияние неопептидов 1, что и pTG19247, слияние 2, как проиллюстрировано в SEQ ID NO: 63, где неопептиды 1 и 2 были инвертированы так же, как и неопептиды 7 и 8, обеспечивая подавление данных межпепидных ТМ сегментов (это новое слияние 2, таким образом, содержит от N к С неопептид 2, неопептид 1, неопептид 3, неопептид 4, неопептид 6, неопептид 8, неопептид 7, неопептид 9 и неопептид 10), и слияние 3, как проиллюстрировано в SEQ ID NO: 64, где неопептиды 2, 4 и 5 были устранены. Кроме того, промотор р11К7.5, используемый для управления слиянием 3 в pTG19247, был заменен промотором р7.5К в pTG19258. Более высокая процентная доля белых бляшек была получена при трансфекции клеток CEF. Однако после выделения и амплификации 20 белых бляшек оставалось только 6 белых бляшек, и анализ ПЦР выявил, что все соответствуют родительскому вирусу, мутировавшему в mCherry (ни одна из них не соответствала рекомбинатному вирусу).
Влияние гидрофобности дополнительно исследовали посредством определения индексов гидрофобности и гидропатичности для каждого неопептида и слияния.
5.2 Анализ гидрофобности
Анализ гидрофобности проводили для определения индексов гидрофобности и гидропатичности всех неопептидов, кодируемых конструкцией MVATG 19111: pTG19247 и pTG19258, с использованием способа Kyte и Doolittle. Результаты для 3 слияний, кодируемых MVATG19111, описываются в следующей

Данные результаты показывают то, что 17 из 18 неопептидов, кодируемых MVATG 19111, демонстрируют удовлетворительные индексы гидрофобности и гидропатичности (отрицательные значения и ниже 0,1 соответственно). Только неопептид 12, содержащийся во втором слиянии, имел положительный балл, отражая гидрофобную природу данного конкретного пептида. Тем не менее, слияние 2 оставалось гидрофобным, что отражается отрицательными индексами гидрофобности (-74) и гидропатичности (-0,42).
Такое же исследование по прогнозированию проводилось для неоептидов и слияний, кодируемых pTG19247 и pTG19258, и индексы описываются в Таблицах 7 и 8 соответственно.
Результаты подчеркивают высокогидрофобную природу слияния 3, что доказывается положительными индексами гидрофобности (133,5) и гидропатичности (0,494).
Такое же исследование было сделано в контексте pTG19258, который соответствует версии pTG19247 с делецией ТМ, как описано выше. Индексы гидрофобности и гидропатичности слияний 1 и 2 являются такими же, что и индексы, описанные в Таблице 7 (такое же слияние, которое несут оба вектора и слияние 2, содержащее такие же неопептиды с инвертированным порядком неопептидов 1 и 2, и 7 и 8 в pTG19258 для устранения межпептидных ТМ-сегментов). В Таблице 8 приводятся баллы гидрофобности и гидропатичности для слияния 3 pTG19258 (слияние из 7 неопептидов, у которых были удалены неопептиды 2, 4 и 5, содержащие ТМ, по отношению к его аналогу pTG19247).
Несмотря на устранение высокогидрофобных и содержащих ТМ неопептидов 2, 4 и 5, общий индекс слияния 3 pTG19258 дает гидрофобную природу.
Следовательно, генерировали дополнительные конструкции, соответственно:
• версию pTG19258 с делецией слияния 3 (названную pTG19288)
• версию pTG19258, включающую 3-аминокислотные линкеры на N-конце первого неопептида и между каждым неопептидом, содержащимся в данных трех слияниях (pTG19267)
• устранение третьего слияния в pTG19267 (pTG19293)
• замена в pTG19258 третьего слияния другим слиянием, содержащим 10 других неопептидов, выбранных из общедоступного банка данных аденокарциномы легкого, и прогнозируемых гидрофильными (отрицательный индекс гидрофобности), без линкеров (pTG19290; SEQ ID NO: 65) или с 3-аминокислотными линкерами на N- и С-концах каждого неопептида (pTG19291).
• устранение слияния 1 в pTG19291 (pTG19298).
Получение рекомбинантных вирусов MVA оценивали в клетках CEF, трансфицированных данными новыми конструкциями плазмид, как описано в данном документе, и сравнивали с родительской плазмидой pTG19258. Красные бляшки коррелируют с родительским (нерекомбинантным) вирусом, тогда как белые бляшки являются указанием потенциального рекомбинантного MVA. Но требуются дополнительные ПЦР-анализы на панели белых и красных бляшек для различения между нерекомбинантным (т.е. родительским) и рекомбинантными вирусами (негативными в отношении mCherry или позитивными в отношении mCherry, причем последние отражают загрязнение рекомбинантных бляшек родительскими). Данные результаты обобщаются на Фиг. 8.
Как показано на Фиг. 8, pTG19258 не приводил к образованию какого-либо рекомбинантного вируса MVA. Несмотря на то, что образуются белые бляшки, вирусы, несущие слияние, не могли быть выявлены посредством анализа ПЦР, хотя в данной конструкции и не могли быть идентифицированы ТМ сегменты (тем не менее, слияние 3 дает положительный индекс, отражая его гидрофобную природу). Белые бляшки были обусловлены родительским вирусом с мутацией в маркерном гене mCherry.
Добавление линкеров между неопептидами в каждой кассете слияния позволяло уменьшать гидрофобную природу слияния 3 (индекс гидропатичности был снижен до значения меньше 0,1 - от 0,151 до 0,08). Данная модификация преобразовалась в более высокую процентную долю белых бляшек (13,3%), генерированных в клетках CEF, давая 7 рекомбинантных вирусов, подтвержденных анализами ПЦР.
Замена гидрофобного слияния 3 в pTG19258 другим слиянием, сделанным из 10 гидрофильных неопептидов (индекс гидропатичности -0,52), также была полезной для генерации рекомбинантного вируса MVA, каким бы ни было присутствие (pTG19291) или отсутствие (pTG19290) линкеров в слияниях неопептидов, давая, соответственно, 7 и 6 генерированных рекомбинантных вирусов.
Устранение слияний для сохранения только 2 слияний - каждого из 10 неопептидов, имеющих отрицательный индекс в отношении гидрофобности, увеличивало процентную долю белых бляшек до 16% и выход рекомбинантных вирусов (10 и 11 рекомбинантов, идентифицированных ПЦР).
В целом, данные результаты указывают на оптимальную конструкцию слияний неопептидов, как обобщено далее:
устранение потенциальных ТМ-сегментов (внутри или между пептидами);
выбор скорее гидрофильных неопептидов (дающих отрицательный индекс в отношении гидрофобности) или уменьшение числа гидрофобных неопептидов (дающих отрицательный индекс в отношении гидрофобности), например, до меньше, чем 40%;
организация неопептидов в слиянии(ниях);
рекомендовано присутствие сигнального пептида на N-конце каждого слияния.
присутствие 3-аминокислотных линкеров между неопептидами на N-конце первого неопептида (например, GTS, GSG или GAS, как проиллюстрировано примером на Фиг. 7)
включение в вирусный геном 1-3 слияний 6-10 неопептидов.
БИБЛИОГРАФИЧЕСКИЕ ССЫЛКИ
Acres and Bonnefoy, 2008, Expert Review of Vaccines 7, 889-93;
Altschul et al., 1990, J. Mol. Biol. 215(3):403-10;
Anderson and Schrijver, 2010, Genes 1 (1): 38-69;
Andreatta et al., 2015, Immunogenetics 67(11-12): 641-50;
Antoine et al., 1998, Virol. 244: 365-96;
Bainbridge et al., 2010, Genome Biol.11:R62;
Boegel et al., 2015, Methods Mol Biol 1310: 247-51;
Caroll et al., 1997, Virology 238: 198-211;
Carpentier et al., 2003, Frontiers in Bioscience 8, e115-127;
Carpentier et al., 2006, Neuro-Oncology 8(1): 60-6;
Chakrabarti et al. 1997, Biotechniques 23: 1094-7;
Chu and Corey, 2012, Nucleic Acid Ther. 22: 271-4;
Claudepierre et al., 2014, J. Virol. 88(10): 5242-55;
Cock ef al., 2010, Nucl. Acid Res. 38(6): 1767-71;
Danecek P. et al., 2011, Bioinformatics 27(15): 2156-58;
Edgar and Flyvbjerg, 2015, Bioinformatics 31 (21): 3476-82;
EP 1162982;
EP 463756;
Erbs et al., 2008, Cancer Gene Ther. 15(1): 18-28;
Farsaci et al., 2011, In Cancer Vaccines: From Research to Clinical Practice, Ed Bot; CRC Press, pp56-77;
Fend et al., 2014, Cancer Immunol. Res. 2, 1163-74;
Firatetal. 1999, European Journal of Immunology 29(10): 3112-21;
Gulley et al., 2008, Clin Cancer Res 14(10): 3060-9;
Guse et al., 2011, Expert Opinion Biol. Ther.11(5):595-608;
Hammond et al, 1997, J. Virol Methods 66: 135-8;
Hundal et al., 2016, Genome Medecine 8(1): 11;
Husseini et al., 2017, Ann Oncol 28(1): 169-74;
Kallol et al., 2003, J. Chromat. 1000: 637-55;
Kern et al., 1990, Gene 88: 149-57;
Kreiter et al. 2015, Nature, 520: 692-6;
Krogh et al., 2001, J. Mol. Biol. 305: 567-80;
Kumar and Boyle, 1990, Virology 179: 151-8;
Kyte and Doolittle, 1982, J. Mol. Biol. 157: 105-32;
Li et al., 2009, Bioinformatics 25(16): 2078-79;
Maniatis et al. 1989, Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor NY;
Mayr et al., 1975, Infection 3: 6-14;
Meyer et al., 1991, J. Gen. Virol. 72: 1031-8;
Moodie et al. 2006, J. Immunol Methods 315: 121-32;
Moodie et al. 2010, Cancer Immunol Immunother doi 10.1007/s00262-010-0875-4;
Ng S.B. et al, 2009, Nature 461: 272-6;
Nielsen et al., 2010, Immunology 130(3): 319-28;
Nielsen et al., 2011, Nature Reviews Genetics 12(6): 443-51;
Olivier et al., 2010, mAbs 2(4): 405-15;
Plotkin, 2008, Clin Infect Dis.47(3):401-9;
Quoixet al., 2011, The Lancet Oncology 12(12): 1125-33;
Relman, 2008, J Infect Dis. 198(1):4-5;
Rock et al., 2010, J. Immunol. 184(1): 9-15;
Rose et al., 1993, Ann. Rev. Biomol. Struc. 22: 381-415;
Sutter and Moss, 1992, Proc. Natl. Acad. Sci. USA 89: 10847-51;
Sweet et al., 1983, J. Mol. Biol. 171: 479-88;
US 5250534;
US 5972597;
US 6440422;
US 6469012;
US 6998252;
US 7108844;
US 7700569;
US 5879924;
Wang et al., 2009, Nature Reviews Genetics 10(1): 57-63;
WO 03/008533;
WO 03/053463;
WO 2005/007840;
WO 2005/042728;
WO 2006/0850082;
WO 2007/056847;
WO 2007/077256;
WO 2007/147528;
WO 2008/092854;
WO 2008/129058;
WO 2008/138533;
WO 2009/004016;
WO 2009/065546;
WO 2009/065547;
WO 2009/100521;
WO 2010/130753;
WO 2010/130756;
WO 2012/001075;
WO 2012/159754;
WO 2013/022764;
WO 2014/053571;
WO 2015/175334;
WO 2015/175340;
WO 2016/187508;
WO 2016/191545;
WO 2016/207859;
WO 97/02355;
WO 99/03885;
Yuan et al., 2015, J. Virol 89, 5176-9, doi:10.1128; Yuan et al., 2016, Viruses 8, 72, doi:10.3390
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> ТРАНСГЕН СА
<120> ПЕРСОНАЛИЗИРОВАННАЯ ВАКЦИНА
<130> B374744PCTD38479
<150> EP 17305760.5
<151> 2017-06-21
<150> EP 18305496.4
<151> 2018-04-23
<160> 70
<170> PatentIn version 3.5
<210> 1
<211> 27
<212> ПРТ
<213> искусственная последовательность
<220>
<223> FCU1 пептид
<400> 1
Cys Ser Lys Glu Gly Ile Glu Lys Tyr His Ala Ala Phe Pro Glu Val
1 5 10 15
Arg Ile Val Thr Gly Ala Leu Asp Arg Gly Leu
20 25
<210> 2
<211> 27
<212> ПРТ
<213> искусственная последовательность
<220>
<223> пептид, полученный из антигена бета-галактозидазы
<400> 2
Gly Gly Ala Asp Thr Thr Ala Thr Asp Ile Ile Cys Pro Met Tyr Ala
1 5 10 15
Arg Val Asp Glu Asp Gln Pro Phe Pro Ala Val
20 25
<210> 3
<211> 27
<212> ПРТ
<213> искусственная последовательность
<220>
<223> пептид, полученный из антигена HPV-16 E1
<400> 3
Tyr Asp Asn Asp Ile Val Asp Asp Ser Glu Ile Ala Tyr Lys Tyr Ala
1 5 10 15
Gln Leu Ala Asp Thr Asn Ser Asn Ala Ser Ala
20 25
<210> 4
<211> 27
<212> ПРТ
<213> искусственная последовательность
<220>
<223> пептид, полученный из антигена HPV-16 E7
<400> 4
Gly Pro Ala Gly Gln Ala Glu Pro Asp Arg Ala His Tyr Asn Ile Val
1 5 10 15
Thr Phe Cys Cys Lys Cys Asp Ser Thr Leu Arg
20 25
<210> 5
<211> 27
<212> ПРТ
<213> искусственная последовательность
<220>
<223> пептид, полученный из антигена MUC1
<400> 5
Tyr Glu Lys Val Ser Ala Gly Asn Gly Gly Ser Ser Leu Ser Tyr Thr
1 5 10 15
Asn Pro Ala Val Ala Ala Thr Ser Ala Asn Leu
20 25
<210> 6
<211> 8
<212> ПРТ
<213> искусственная последовательность
<220>
<223> Flag tag
<400> 6
Asp Tyr Lys Asp Asp Asp Asp Lys
1 5
<210> 7
<211> 204
<212> ПРТ
<213> искусственная последовательность
<220>
<223> гибрид из 5 пептидов, экспрессируемый MVATG19022
<400> 7
Met Gly Ser Gly Ser Gly Ser Gly Ser Gly Ser Cys Ser Lys Glu Gly
1 5 10 15
Ile Glu Lys Tyr His Ala Ala Phe Pro Glu Val Arg Ile Val Thr Gly
20 25 30
Ala Leu Asp Arg Gly Leu Gly Ser Gly Ser Gly Ser Gly Ser Gly Ser
35 40 45
Gly Gly Ala Asp Thr Thr Ala Thr Asp Ile Ile Cys Pro Met Tyr Ala
50 55 60
Arg Val Asp Glu Asp Gln Pro Phe Pro Ala Val Gly Ser Gly Ser Gly
65 70 75 80
Ser Gly Ser Gly Ser Tyr Asp Asn Asp Ile Val Asp Asp Ser Glu Ile
85 90 95
Ala Tyr Lys Tyr Ala Gln Leu Ala Asp Thr Asn Ser Asn Ala Ser Ala
100 105 110
Gly Ser Gly Ser Gly Ser Gly Ser Gly Ser Gly Pro Ala Gly Gln Ala
115 120 125
Glu Pro Asp Arg Ala His Tyr Asn Ile Val Thr Phe Cys Cys Lys Cys
130 135 140
Asp Ser Thr Leu Arg Gly Ser Gly Ser Gly Ser Gly Ser Gly Ser Tyr
145 150 155 160
Glu Lys Val Ser Ala Gly Asn Gly Gly Ser Ser Leu Ser Tyr Thr Asn
165 170 175
Pro Ala Val Ala Ala Thr Ser Ala Asn Leu Gly Ser Gly Ser Gly Ser
180 185 190
Gly Ser Gly Ser Asp Tyr Lys Asp Asp Asp Asp Lys
195 200
<210> 8
<211> 112
<212> ДНК
<213> искусственная последовательность
<220>
<223> фрагмент промотора vaccinia p11k7.5
<400> 8
ataaaaatat agtagaattt cattttgttt ttttctatgc tataaatagg atccgataaa 60
gtgaaaaata attctaattt attgcacggt aaggaagtag aatcataaag aa 112
<210> 9
<211> 23
<212> ПРТ
<213> искусственная последовательность
<220>
<223> фрагмент гликопротеина вируса бешенства (сигнальный пептид)
<400> 9
Met Val Pro Gln Ala Leu Leu Phe Val Pro Leu Leu Val Phe Pro Leu
1 5 10 15
Cys Phe Gly Lys Phe Pro Ile
20
<210> 10
<211> 226
<212> ПРТ
<213> искусственная последовательность
<220>
<223> гибрид из 5 пептидов, кодируемый MVATG19023 (+ сигнальный пептид)
<400> 10
Met Val Pro Gln Ala Leu Leu Phe Val Pro Leu Leu Val Phe Pro Leu
1 5 10 15
Cys Phe Gly Lys Phe Pro Ile Gly Ser Gly Ser Gly Ser Gly Ser Gly
20 25 30
Ser Cys Ser Lys Glu Gly Ile Glu Lys Tyr His Ala Ala Phe Pro Glu
35 40 45
Val Arg Ile Val Thr Gly Ala Leu Asp Arg Gly Leu Gly Ser Gly Ser
50 55 60
Gly Ser Gly Ser Gly Ser Gly Gly Ala Asp Thr Thr Ala Thr Asp Ile
65 70 75 80
Ile Cys Pro Met Tyr Ala Arg Val Asp Glu Asp Gln Pro Phe Pro Ala
85 90 95
Val Gly Ser Gly Ser Gly Ser Gly Ser Gly Ser Tyr Asp Asn Asp Ile
100 105 110
Val Asp Asp Ser Glu Ile Ala Tyr Lys Tyr Ala Gln Leu Ala Asp Thr
115 120 125
Asn Ser Asn Ala Ser Ala Gly Ser Gly Ser Gly Ser Gly Ser Gly Ser
130 135 140
Gly Pro Ala Gly Gln Ala Glu Pro Asp Arg Ala His Tyr Asn Ile Val
145 150 155 160
Thr Phe Cys Cys Lys Cys Asp Ser Thr Leu Arg Gly Ser Gly Ser Gly
165 170 175
Ser Gly Ser Gly Ser Tyr Glu Lys Val Ser Ala Gly Asn Gly Gly Ser
180 185 190
Ser Leu Ser Tyr Thr Asn Pro Ala Val Ala Ala Thr Ser Ala Asn Leu
195 200 205
Gly Ser Gly Ser Gly Ser Gly Ser Gly Ser Asp Tyr Lys Asp Asp Asp
210 215 220
Asp Lys
225
<210> 11
<211> 10
<212> ПРТ
<213> искусственная последовательность
<220>
<223> cMYC tag
<400> 11
Glu Gln Lys Leu Ile Ser Glu Glu Asp Leu
1 5 10
<210> 12
<211> 204
<212> ПРТ
<213> искусственная последовательность
<220>
<223> гибрид из 5 CT26 неопептидов (пентатоп 1)
<400> 12
Met Gly Ser Gly Ser Gly Ser Gly Ser Gly Ser Tyr Arg Gly Ala Asn
1 5 10 15
Leu His Leu Glu Glu Thr Leu Ala Gly Phe Trp Ala Arg Leu Leu Glu
20 25 30
Arg Leu Phe Lys Gln Leu Gly Ser Gly Ser Gly Ser Gly Ser Gly Ser
35 40 45
Gly Tyr Ile Ser Arg Val Thr Ala Gly Lys Asp Ser Tyr Ile Ala Leu
50 55 60
Val Asp Lys Asn Ile Met Gly Tyr Ile Ala Ser Gly Ser Gly Ser Gly
65 70 75 80
Ser Gly Ser Gly Ser Ala Gly Thr Gln Cys Glu Tyr Trp Ala Ser Arg
85 90 95
Ala Leu Asp Ser Glu His Ser Ile Gly Ser Met Ile Gln Leu Pro Gln
100 105 110
Gly Ser Gly Ser Gly Ser Gly Ser Gly Ser Glu Gly Asp Pro Cys Leu
115 120 125
Arg Ser Ser Asp Cys Ile Asp Glu Phe Cys Cys Ala Arg His Phe Trp
130 135 140
Thr Lys Ile Cys Lys Gly Ser Gly Ser Gly Ser Gly Ser Gly Ser Val
145 150 155 160
Thr Ser Ile Pro Ser Val Ser Asn Ala Leu Asn Trp Lys Glu Phe Ser
165 170 175
Phe Ile Gln Ser Thr Leu Gly Tyr Val Ala Gly Ser Gly Ser Gly Ser
180 185 190
Gly Ser Gly Ser Asp Tyr Lys Asp Asp Asp Asp Lys
195 200
<210> 13
<211> 206
<212> ПРТ
<213> искусственная последовательность
<220>
<223> гибрид из 5 CT26 неопептидов (пентатоп 2), кодируемый MVATG19030
<400> 13
Met Gly Ser Gly Ser Gly Ser Gly Ser Gly Ser Pro Leu Leu Pro Phe
1 5 10 15
Tyr Pro Pro Asp Glu Ala Leu Glu Ile Gly Leu Glu Leu Asn Ser Ser
20 25 30
Ala Leu Pro Pro Thr Glu Gly Ser Gly Ser Gly Ser Gly Ser Gly Ser
35 40 45
Val Ile Leu Pro Gln Ala Pro Ser Gly Pro Ser Tyr Ala Thr Tyr Leu
50 55 60
Gln Pro Ala Gln Ala Gln Met Leu Thr Pro Pro Gly Ser Gly Ser Gly
65 70 75 80
Ser Gly Ser Gly Ser Asp Lys Pro Leu Arg Arg Asn Asn Ser Tyr Thr
85 90 95
Ser Tyr Ile Met Ala Ile Cys Gly Met Pro Leu Asp Ser Phe Arg Ala
100 105 110
Gly Ser Gly Ser Gly Ser Gly Ser Gly Ser Glu Val Ile Gln Thr Ser
115 120 125
Lys Tyr Tyr Met Arg Asp Val Ile Ala Ile Glu Ser Ala Trp Leu Leu
130 135 140
Glu Leu Ala Pro His Gly Ser Gly Ser Gly Ser Gly Ser Gly Ser Glu
145 150 155 160
His Ile His Arg Ala Gly Gly Leu Phe Val Ala Asp Ala Ile Gln Val
165 170 175
Gly Phe Gly Arg Ile Gly Lys His Phe Trp Gly Ser Gly Ser Gly Ser
180 185 190
Gly Ser Gly Ser Glu Gln Lys Leu Ile Ser Glu Glu Asp Leu
195 200 205
<210> 14
<211> 114
<212> ДНК
<213> искусственная последовательность
<220>
<223> фрагмент протомора vaccinia pH5R
<400> 14
tttattctat acttaaaaaa tgaaaataaa tacaaaggtt cttgagggtt gtgttaaatt 60
gaaagcgaga aataatcata aattatttca ttatcgcgat atccgttaag tttg 114
<210> 15
<211> 28
<212> ПРТ
<213> искусственная последовательность
<220>
<223> фрагмент гликопротеина F вируса кори (сигнальный пептид)
<400> 15
Met Gly Leu Lys Val Asn Val Ser Ala Ile Phe Met Ala Val Leu Leu
1 5 10 15
Thr Leu Gln Thr Pro Thr Gly Gln Ile His Trp Gly
20 25
<210> 16
<211> 226
<212> ПРТ
<213> искусственная последовательность
<220>
<223> гибрид из 5 CT26 неопептидов, кодируемый MVATG19038 (пентатоп 1)
<400> 16
Met Val Pro Gln Ala Leu Leu Phe Val Pro Leu Leu Val Phe Pro Leu
1 5 10 15
Cys Phe Gly Lys Phe Pro Ile Gly Ser Gly Ser Gly Ser Gly Ser Gly
20 25 30
Ser Tyr Arg Gly Ala Asn Leu His Leu Glu Glu Thr Leu Ala Gly Phe
35 40 45
Trp Ala Arg Leu Leu Glu Arg Leu Phe Lys Gln Leu Gly Ser Gly Ser
50 55 60
Gly Ser Gly Ser Gly Ser Gly Tyr Ile Ser Arg Val Thr Ala Gly Lys
65 70 75 80
Asp Ser Tyr Ile Ala Leu Val Asp Lys Asn Ile Met Gly Tyr Ile Ala
85 90 95
Ser Gly Ser Gly Ser Gly Ser Gly Ser Gly Ser Ala Gly Thr Gln Cys
100 105 110
Glu Tyr Trp Ala Ser Arg Ala Leu Asp Ser Glu His Ser Ile Gly Ser
115 120 125
Met Ile Gln Leu Pro Gln Gly Ser Gly Ser Gly Ser Gly Ser Gly Ser
130 135 140
Glu Gly Asp Pro Cys Leu Arg Ser Ser Asp Cys Ile Asp Glu Phe Cys
145 150 155 160
Cys Ala Arg His Phe Trp Thr Lys Ile Cys Lys Gly Ser Gly Ser Gly
165 170 175
Ser Gly Ser Gly Ser Val Thr Ser Ile Pro Ser Val Ser Asn Ala Leu
180 185 190
Asn Trp Lys Glu Phe Ser Phe Ile Gln Ser Thr Leu Gly Tyr Val Ala
195 200 205
Gly Ser Gly Ser Gly Ser Gly Ser Gly Ser Asp Tyr Lys Asp Asp Asp
210 215 220
Asp Lys
225
<210> 17
<211> 232
<212> ПРТ
<213> искусственная последовательность
<220>
<223> гибрид из 5 CT26 неопептидов, кодируемый MVATG19038 (пентатоп 2)
<400> 17
Met Gly Leu Lys Val Asn Val Ser Ala Ile Phe Met Ala Val Leu Leu
1 5 10 15
Thr Leu Gln Thr Pro Thr Gly Gln Ile His Trp Gly Ser Gly Ser Gly
20 25 30
Ser Gly Ser Gly Ser Pro Leu Leu Pro Phe Tyr Pro Pro Asp Glu Ala
35 40 45
Leu Glu Ile Gly Leu Glu Leu Asn Ser Ser Ala Leu Pro Pro Thr Glu
50 55 60
Gly Ser Gly Ser Gly Ser Gly Ser Gly Ser Val Ile Leu Pro Gln Ala
65 70 75 80
Pro Ser Gly Pro Ser Tyr Ala Thr Tyr Leu Gln Pro Ala Gln Ala Gln
85 90 95
Met Leu Thr Pro Pro Gly Ser Gly Ser Gly Ser Gly Ser Gly Ser Asp
100 105 110
Lys Pro Leu Arg Arg Asn Asn Ser Tyr Thr Ser Tyr Ile Met Ala Ile
115 120 125
Cys Gly Met Pro Leu Asp Ser Phe Arg Ala Gly Ser Gly Ser Gly Ser
130 135 140
Gly Ser Gly Ser Glu Val Ile Gln Thr Ser Lys Tyr Tyr Met Arg Asp
145 150 155 160
Val Ile Ala Ile Glu Ser Ala Trp Leu Leu Glu Leu Ala Pro His Gly
165 170 175
Ser Gly Ser Gly Ser Gly Ser Gly Ser Glu His Ile His Arg Ala Gly
180 185 190
Gly Leu Phe Val Ala Asp Ala Ile Gln Val Gly Phe Gly Arg Ile Gly
195 200 205
Lys His Phe Trp Gly Ser Gly Ser Gly Ser Gly Ser Gly Ser Glu Gln
210 215 220
Lys Leu Ile Ser Glu Glu Asp Leu
225 230
<210> 18
<211> 9
<212> ПРТ
<213> искусственная последовательность
<220>
<223> HA tag
<400> 18
Tyr Pro Tyr Asp Val Pro Asp Tyr Ala
1 5
<210> 19
<211> 11
<212> ПРТ
<213> искусственная последовательность
<220>
<223> HSV tag
<400> 19
Gln Pro Glu Leu Ala Pro Glu Asp Pro Glu Asp
1 5 10
<210> 20
<211> 11
<212> ПРТ
<213> искусственная последовательность
<220>
<223> VSV tag
<400> 20
Tyr Thr Asp Ile Glu Met Asn Arg Leu Gly Lys
1 5 10
<210> 21
<211> 100
<212> ДНК
<213> искусственная последовательность
<220>
<223> фрагмент vaccinia pB2R promoter
<400> 21
tatattatta agtgtggtgt ttggtcgatg taaaattttt gtcgataaaa attaaaaaat 60
aacttaattt attattgatc tcgtgtgtac aaccgaaatc 100
<210> 22
<211> 27
<212> ПРТ
<213> искусственная последовательность
<220>
<223> CT26 пептид 1
<400> 22
Tyr Arg Gly Ala Asn Leu His Leu Glu Glu Thr Leu Ala Gly Phe Trp
1 5 10 15
Ala Arg Leu Leu Glu Arg Leu Phe Lys Gln Leu
20 25
<210> 23
<211> 27
<212> ПРТ
<213> искусственная последовательность
<220>
<223> CT26 пептид 2
<400> 23
Gly Tyr Ile Ser Arg Val Thr Ala Gly Lys Asp Ser Tyr Ile Ala Leu
1 5 10 15
Val Asp Lys Asn Ile Met Gly Tyr Ile Ala Ser
20 25
<210> 24
<211> 27
<212> ПРТ
<213> искусственная последовательность
<220>
<223> CT26 пептид 3
<400> 24
Ala Gly Thr Gln Cys Glu Tyr Trp Ala Ser Arg Ala Leu Asp Ser Glu
1 5 10 15
His Ser Ile Gly Ser Met Ile Gln Leu Pro Gln
20 25
<210> 25
<211> 27
<212> ПРТ
<213> искусственная последовательность
<220>
<223> CT26 неопептид 4
<400> 25
Glu Gly Asp Pro Cys Leu Arg Ser Ser Asp Cys Ile Asp Glu Phe Cys
1 5 10 15
Cys Ala Arg His Phe Trp Thr Lys Ile Cys Lys
20 25
<210> 26
<211> 27
<212> ПРТ
<213> искусственная последовательность
<220>
<223> CT26 неопептид 5
<400> 26
Val Thr Ser Ile Pro Ser Val Ser Asn Ala Leu Asn Trp Lys Glu Phe
1 5 10 15
Ser Phe Ile Gln Ser Thr Leu Gly Tyr Val Ala
20 25
<210> 27
<211> 27
<212> ПРТ
<213> искусственная последовательность
<220>
<223> CT26 неопептид 6
<400> 27
Pro Leu Leu Pro Phe Tyr Pro Pro Asp Glu Ala Leu Glu Ile Gly Leu
1 5 10 15
Glu Leu Asn Ser Ser Ala Leu Pro Pro Thr Glu
20 25
<210> 28
<211> 27
<212> ПРТ
<213> искусственная последовательность
<220>
<223> CT26 неопептид 7
<400> 28
Val Ile Leu Pro Gln Ala Pro Ser Gly Pro Ser Tyr Ala Thr Tyr Leu
1 5 10 15
Gln Pro Ala Gln Ala Gln Met Leu Thr Pro Pro
20 25
<210> 29
<211> 27
<212> ПРТ
<213> искусственная последовательность
<220>
<223> CT26 неопептид 8
<400> 29
Asp Lys Pro Leu Arg Arg Asn Asn Ser Tyr Thr Ser Tyr Ile Met Ala
1 5 10 15
Ile Cys Gly Met Pro Leu Asp Ser Phe Arg Ala
20 25
<210> 30
<211> 27
<212> ПРТ
<213> искусственная последовательность
<220>
<223> CT26 неопептид 9
<400> 30
Glu Val Ile Gln Thr Ser Lys Tyr Tyr Met Arg Asp Val Ile Ala Ile
1 5 10 15
Glu Ser Ala Trp Leu Leu Glu Leu Ala Pro His
20 25
<210> 31
<211> 27
<212> ПРТ
<213> искусственная последовательность
<220>
<223> CT26 неопептид 10
<400> 31
Glu His Ile His Arg Ala Gly Gly Leu Phe Val Ala Asp Ala Ile Gln
1 5 10 15
Val Gly Phe Gly Arg Ile Gly Lys His Phe Trp
20 25
<210> 32
<211> 27
<212> ПРТ
<213> искусственная последовательность
<220>
<223> немутированный CT26 пептид 1
<400> 32
Tyr Arg Gly Ala Asn Leu His Leu Glu Glu Thr Leu Ala Glu Phe Trp
1 5 10 15
Ala Arg Leu Leu Glu Arg Leu Phe Lys Gln Leu
20 25
<210> 33
<211> 27
<212> ПРТ
<213> искусственная последовательность
<220>
<223> немутированный CT26 пептид 2
<400> 33
Gly Tyr Ile Ser Arg Val Thr Ala Gly Lys Asp Ser Tyr Leu Ala Leu
1 5 10 15
Val Asp Lys Asn Ile Met Gly Tyr Ile Ala Ser
20 25
<210> 34
<211> 27
<212> ПРТ
<213> искусственная последовательность
<220>
<223> немутированный CT26 пептид 3
<400> 34
Ala Gly Thr Gln Cys Glu Tyr Trp Ala Ser Arg Ala Leu Gly Ser Glu
1 5 10 15
His Ser Ile Gly Ser Met Ile Gln Leu Pro Gln
20 25
<210> 35
<211> 27
<212> ПРТ
<213> искусственная последовательность
<220>
<223> немутированный CT26 пептид 4
<400> 35
Glu Gly Asp Pro Cys Leu Arg Ser Ser Asp Cys Ile Asp Gly Phe Cys
1 5 10 15
Cys Ala Arg His Phe Trp Thr Lys Ile Cys Lys
20 25
<210> 36
<211> 27
<212> ПРТ
<213> искусственная последовательность
<220>
<223> немутированный CT26 пептид 5
<400> 36
Val Thr Ser Ile Pro Ser Val Ser Asn Ala Leu Asn Trp Arg Glu Phe
1 5 10 15
Ser Phe Ile Gln Ser Thr Leu Gly Tyr Val Ala
20 25
<210> 37
<211> 27
<212> ПРТ
<213> искусственная последовательность
<220>
<223> немутированный CT26 пептид 6
<400> 37
Pro Leu Leu Pro Phe Tyr Pro Pro Asp Glu Ala Leu Glu Thr Gly Leu
1 5 10 15
Glu Leu Asn Ser Ser Ala Leu Pro Pro Thr Glu
20 25
<210> 38
<211> 27
<212> ПРТ
<213> искусственная последовательность
<220>
<223> немутированный CT26 пептид 7
<400> 38
Val Ile Leu Pro Gln Ala Pro Ser Gly Pro Ser Tyr Ala Ile Tyr Leu
1 5 10 15
Gln Pro Ala Gln Ala Gln Met Leu Thr Pro Pro
20 25
<210> 39
<211> 27
<212> ПРТ
<213> искусственная последовательность
<220>
<223> немутированный CT26 пептид 8
<400> 39
Asp Lys Pro Leu Arg Arg Asn Asn Ser Tyr Thr Ser Tyr Thr Met Ala
1 5 10 15
Ile Cys Gly Met Pro Leu Asp Ser Phe Arg Ala
20 25
<210> 40
<211> 27
<212> ПРТ
<213> искусственная последовательность
<220>
<223> немутированный CT26 пептид 9
<400> 40
Glu Val Ile Gln Thr Ser Lys Tyr Tyr Met Arg Asp Val Thr Ala Ile
1 5 10 15
Glu Ser Ala Trp Leu Leu Glu Leu Ala Pro His
20 25
<210> 41
<211> 27
<212> ПРТ
<213> искусственная последовательность
<220>
<223> немутированный CT26 пептид 10
<400> 41
Glu His Ile His Arg Ala Gly Gly Leu Phe Val Ala Asp Glu Ile Gln
1 5 10 15
Val Gly Phe Gly Arg Ile Gly Lys His Phe Trp
20 25
<210> 42
<211> 9
<212> ПРТ
<213> искусственная последовательность
<220>
<223> CD8 CT26 эпитоп 7M1
<400> 42
Pro Ser Tyr Ala Thr Tyr Leu Gln Pro
1 5
<210> 43
<211> 9
<212> ПРТ
<213> искусственная последовательность
<220>
<223> CD8 CT26 эпитоп 7M2
<400> 43
Tyr Ala Thr Tyr Leu Gln Pro Ala Gln
1 5
<210> 44
<211> 9
<212> ПРТ
<213> искусственная последовательность
<220>
<223> CD8 CT26 эпитоп 7M3
<400> 44
Thr Tyr Leu Gln Pro Ala Gln Ala Gln
1 5
<210> 45
<211> 9
<212> ПРТ
<213> искусственная последовательность
<220>
<223> CD8 CT26 эпитоп 2M
<400> 45
Ile Ala Leu Val Asp Lys Asn Ile Met
1 5
<210> 46
<211> 9
<212> ПРТ
<213> искусственная последовательность
<220>
<223> CD8 CT26 эпитоп 7WT1
<400> 46
Pro Ser Tyr Ala Ile Tyr Leu Gln Pro
1 5
<210> 47
<211> 9
<212> ПРТ
<213> искусственная последовательность
<220>
<223> CD8 CT26 эпитоп 7WT2
<400> 47
Tyr Ala Ile Tyr Leu Gln Pro Ala Gln
1 5
<210> 48
<211> 9
<212> ПРТ
<213> искусственная последовательность
<220>
<223> CD8 CT26 эпитоп 7WT3
<400> 48
Ile Tyr Leu Gln Pro Ala Gln Ala Gln
1 5
<210> 49
<211> 9
<212> ПРТ
<213> искусственная последовательность
<220>
<223> CD8 CTй- эпитоп 2 WT
<400> 49
Leu Ala Leu Val Asp Lys Asn Ile Met
1 5
<210> 50
<211> 8
<212> ПРТ
<213> искусственная последовательность
<220>
<223> стимулирующий пептид
<400> 50
Ile Ala Tyr Lys Tyr Ala Gln Leu
1 5
<210> 51
<211> 8
<212> ПРТ
<213> искусственная последовательность
<220>
<223> стимулирующий пептид
<400> 51
Ile Cys Pro Met Tyr Ala Arg Val
1 5
<210> 52
<211> 9
<212> ПРТ
<213> искусственная последовательность
<220>
<223> стимулирующий пептид
<400> 52
Arg Ala His Tyr Asn Ile Val Thr Phe
1 5
<210> 53
<211> 15
<212> ПРТ
<213> искусственная последовательность
<220>
<223> стимулирующий пептид
<400> 53
Leu Ser Tyr Thr Asn Pro Ala Val Ala Ala Thr Ser Ala Asn Leu
1 5 10 15
<210> 54
<211> 15
<212> ПРТ
<213> искусственная последовательность
<220>
<223> стимулирующий пептид
<400> 54
Glu Lys Tyr His Ala Ala Phe Pro Glu Val Arg Ile Val Thr Gly
1 5 10 15
<210> 55
<211> 8
<212> ПРТ
<213> искусственная последовательность
<220>
<223> стимулирующий пептид
<400> 55
Thr Ser Tyr Lys Phe Glu Ser Val
1 5
<210> 56
<211> 9
<212> ПРТ
<213> искусственная последовательность
<220>
<223> нерелевантный пептид
<400> 56
Lys Asn Gly Glu Asn Ala Gln Ala Ile
1 5
<210> 57
<211> 15
<212> ПРТ
<213> искусственная последовательность
<220>
<223> нерелевантный пептид
<400> 57
Gly Ile Cys Leu Met Leu Phe Ile Leu Ile Lys Arg Ser Arg His
1 5 10 15
<210> 58
<211> 27
<212> ПРТ
<213> искусственная последовательность
<220>
<223> пример неопептида аденокарциномы легких, который дает
отрицательные баллы по шкале гидрофобности и гидропатии
<400> 58
Gly Gln Ser Leu Pro Met Thr His Ser Leu Lys Leu Ser Lys Thr Asn
1 5 10 15
Arg Thr Leu Phe Leu Leu Gly Val Thr Lys Tyr
20 25
<210> 59
<211> 27
<212> ПРТ
<213> искусственная последовательность
<220>
<223> пример неопептида аденокарциномы легких, который дает
положительные баллы по шкале гидрофобности и гидропатии
<400> 59
Gly Leu Met Gly Ile Val Val Gly Thr Val Phe Ile Ile Arg Gly Leu
1 5 10 15
Arg Ser Val Gly Ala Ser Arg His Gln Gly Leu
20 25
<210> 60
<211> 270
<212> ПРТ
<213> искусственная последовательность
<220>
<223> гибрид 1 из десяти неопептидов, кодируемый pTG19247
<400> 60
Lys Glu His Gly Thr His Gln Gln Leu Leu Ala Gln Lys Val Ile Tyr
1 5 10 15
Phe Ser Met Val Ser Val Gln Ala Gly Thr Lys Ala Trp Glu Ser Trp
20 25 30
Arg Ser Glu Val Gly Lys Gln Leu Met Pro Leu Tyr Glu Glu Tyr Val
35 40 45
Val Leu Lys Asn Glu Met Asp Val Ser Lys Glu Asn Glu Asn Ser Met
50 55 60
Val Pro Gln Arg Ala Pro Val Gly Ser Leu Ser Val Lys Asn Lys Ala
65 70 75 80
His Gln Met Lys Ala Arg Leu Thr Gln Glu Leu Gln Gln Ala Asn Asn
85 90 95
Met His Asn Val Leu Gln Ala Glu Leu Asp Lys Leu His Leu His Tyr
100 105 110
Pro Ile Glu Arg Gly Leu Val Thr Gly Cys Asp Asp Met Glu Lys Leu
115 120 125
Trp Lys His Leu Phe Glu Arg Ala Val Pro Tyr Met Ala Lys Phe Val
130 135 140
Ile Phe Ala Lys Ile Asn Asp Pro Arg Glu Gly Arg Leu Arg Cys Tyr
145 150 155 160
Cys Met Lys Thr Val Val Val Met Leu Gln Asn Val Val Pro Ala Lys
165 170 175
Val Ser Leu Val Cys Val Val Lys Pro Asp Glu Phe Trp Ser Gly Val
180 185 190
Lys Val Ser Val Tyr Ala Val Pro Asp Lys Met Asn Gln Ala Asp Tyr
195 200 205
Ala Leu Asp Ala Ala Val Thr Leu Trp Asn Asn Gly Arg Asn His Leu
210 215 220
Ile Phe Asn Leu Tyr Phe Gly Thr Trp Pro Asp Tyr Thr Glu Asp Val
225 230 235 240
Gly Phe Asp Ile Arg Gln Pro Pro Gly Lys Ala Leu Glu Trp Leu Ala
245 250 255
His Ile Tyr Trp Asp Asp Asp Lys Arg Tyr Ser Pro Ser Leu
260 265 270
<210> 61
<211> 267
<212> ПРТ
<213> искусственная последовательность
<220>
<223> гибрид 2 из 10 неопептидов, кодируемый pTG19247
<400> 61
Gly Gln Ser Leu Pro Met Thr His Ser Leu Lys Leu Ser Lys Thr Asn
1 5 10 15
Arg Thr Leu Phe Leu Leu Gly Val Thr Lys Tyr Gly Leu Met Gly Ile
20 25 30
Val Val Gly Thr Val Phe Ile Ile Arg Gly Leu Arg Ser Val Gly Ala
35 40 45
Ser Arg His Gln Gly Leu Trp Tyr Val Arg Met Asn Arg Arg Arg Leu
50 55 60
Lys Gly Glu Thr Gly Met Glu Asp Cys Val Met Ala Leu Glu Thr Leu
65 70 75 80
Phe Thr Ala Ile Pro Leu Glu Ile Phe Ile Val Thr Met Glu Asn Asp
85 90 95
Phe Pro Gly Gly Val Ile Gly Lys Ile His Ala Thr Gly Arg Gly Arg
100 105 110
Ser Lys Leu Lys Ser Gly Ile Gly Ala Leu Val Leu Pro Gly Val Ser
115 120 125
Thr Ala Asp Ile Ser Ser Asn His Gln Arg Glu Asn Ser Gly His Arg
130 135 140
Arg Asp Gln Ile Thr Glu Lys Asp Ala Ala Leu Cys Val Leu Ile Asp
145 150 155 160
Glu Met His Asn Tyr Met Tyr Trp Tyr Arg Gln Asp Pro Gly Met Val
165 170 175
Leu Lys Leu Ile Tyr Tyr Ser Val Gly Ala Gly Ile Thr Gly Trp Met
180 185 190
Tyr Gly Thr Val Gln Arg Thr Gly Arg Thr Arg Met Leu Pro Ala Asn
195 200 205
Tyr Val Glu Ala Ile Glu Glu Trp Glu Cys Leu Asp Ser Ala Gln Gln
210 215 220
Arg Leu His Arg Asp Val Met Leu Glu Asn Tyr Gly Asn Leu Phe Ser
225 230 235 240
Leu Phe Thr Val Ser Pro Thr Leu Tyr Phe Leu His Ser Val Ser Leu
245 250 255
Pro Ala Leu Asp Leu Lys Pro Gly Glu Gly Ala
260 265
<210> 62
<211> 270
<212> ПРТ
<213> искусственная последовательность
<220>
<223> гибрид 3 из 10 неопептидов, кодируемый pTG19247
<400> 62
Val Val Gly Trp Arg Ser Gly Val Glu Lys Asp Leu Asp Lys Val Leu
1 5 10 15
Gln Thr His Ser Val Phe Val Asn Val Ser Lys Lys Gly Ile Arg Thr
20 25 30
Leu Leu Phe Ala Leu Met Met Ser Phe Pro Ala Leu Phe Asn Ile Gly
35 40 45
Leu Leu Leu Phe Leu Val Glu Ala Thr Asn Ala Ser Val Phe Lys Tyr
50 55 60
Tyr Ile His Gly Leu Ser Asp Leu Ile Asp Cys Cys Asp Leu Gly Tyr
65 70 75 80
His Ser Ile Phe Tyr Leu Leu Lys Ile Ala Asn Phe Ser Asn Leu Ile
85 90 95
Phe Leu His Leu Lys Arg Arg Val Lys Ser Val Ile Val His His Arg
100 105 110
Ile Val Gly Cys Ser Leu Ala Val Ile Cys Gly Val Leu Tyr Gly Ser
115 120 125
Thr Phe Val Pro Ile Ile Tyr Phe Leu Val Ile Ala Leu Lys Asn Ala
130 135 140
Val Glu Ile Tyr Val Trp Ala Pro Lys Pro Tyr His Lys Phe Met Ala
145 150 155 160
Phe Lys Pro Gly Leu Asp Arg Glu Gln Leu Tyr Trp Glu Leu Ser Lys
165 170 175
Leu Thr His Ser Ile Thr Glu Leu Gly Pro Tyr Thr Leu Leu Leu Gln
180 185 190
Ile Ser Asn Trp Phe Ile Asn Ala Arg Arg Cys Ile Leu Pro Asp Met
195 200 205
Leu Gln Gln Arg Arg Asn Asp Pro Thr Ile Ser Leu Ser Gly Cys Ala
210 215 220
Val Gln Met Phe Leu Ser Leu Ala Met Gly Thr Thr Glu Cys Val Leu
225 230 235 240
Leu Gly Met Gln Ile Leu Ser Lys Leu Leu Leu Asp Leu Ser Cys Glu
245 250 255
Phe Ser Leu Leu Lys Ser Glu Cys His Arg Val Lys Met Gln
260 265 270
<210> 63
<211> 267
<212> ПРТ
<213> искусственная последовательность
<220>
<223> гибрид 2 без ТМ с инверсией неопептидов 1 и 2 и 7 и 8
<400> 63
Gly Leu Met Gly Ile Val Val Gly Thr Val Phe Ile Ile Arg Gly Leu
1 5 10 15
Arg Ser Val Gly Ala Ser Arg His Gln Gly Leu Gly Gln Ser Leu Pro
20 25 30
Met Thr His Ser Leu Lys Leu Ser Lys Thr Asn Arg Thr Leu Phe Leu
35 40 45
Leu Gly Val Thr Lys Tyr Trp Tyr Val Arg Met Asn Arg Arg Arg Leu
50 55 60
Lys Gly Glu Thr Gly Met Glu Asp Cys Val Met Ala Leu Glu Thr Leu
65 70 75 80
Phe Thr Ala Ile Pro Leu Glu Ile Phe Ile Val Thr Met Glu Asn Asp
85 90 95
Phe Pro Gly Gly Val Ile Gly Lys Ile His Ala Thr Gly Arg Gly Arg
100 105 110
Ser Lys Leu Lys Ser Gly Ile Gly Ala Leu Val Leu Pro Gly Val Ser
115 120 125
Thr Ala Asp Ile Ser Ser Asn His Gln Arg Glu Asn Ser Gly His Arg
130 135 140
Arg Asp Gln Ile Thr Glu Lys Asp Ala Ala Leu Cys Val Leu Ile Asp
145 150 155 160
Glu Met Gly Trp Met Tyr Gly Thr Val Gln Arg Thr Gly Arg Thr Arg
165 170 175
Met Leu Pro Ala Asn Tyr Val Glu Ala Ile His Asn Tyr Met Tyr Trp
180 185 190
Tyr Arg Gln Asp Pro Gly Met Val Leu Lys Leu Ile Tyr Tyr Ser Val
195 200 205
Gly Ala Gly Ile Thr Glu Glu Trp Glu Cys Leu Asp Ser Ala Gln Gln
210 215 220
Arg Leu His Arg Asp Val Met Leu Glu Asn Tyr Gly Asn Leu Phe Ser
225 230 235 240
Leu Phe Thr Val Ser Pro Thr Leu Tyr Phe Leu His Ser Val Ser Leu
245 250 255
Pro Ala Leu Asp Leu Lys Pro Gly Glu Gly Ala
260 265
<210> 64
<211> 189
<212> ПРТ
<213> искусственная последовательность
<220>
<223> гибрид 3 без TM с супрессией неопептидов 2, 4 и 5
<400> 64
Val Val Gly Trp Arg Ser Gly Val Glu Lys Asp Leu Asp Lys Val Leu
1 5 10 15
Gln Thr His Ser Val Phe Val Asn Val Ser Lys Glu Ala Thr Asn Ala
20 25 30
Ser Val Phe Lys Tyr Tyr Ile His Gly Leu Ser Asp Leu Ile Asp Cys
35 40 45
Cys Asp Leu Gly Tyr His Phe Leu Val Ile Ala Leu Lys Asn Ala Val
50 55 60
Glu Ile Tyr Val Trp Ala Pro Lys Pro Tyr His Lys Phe Met Ala Phe
65 70 75 80
Lys Pro Gly Leu Asp Arg Glu Gln Leu Tyr Trp Glu Leu Ser Lys Leu
85 90 95
Thr His Ser Ile Thr Glu Leu Gly Pro Tyr Thr Leu Leu Leu Gln Ile
100 105 110
Ser Asn Trp Phe Ile Asn Ala Arg Arg Cys Ile Leu Pro Asp Met Leu
115 120 125
Gln Gln Arg Arg Asn Asp Pro Thr Ile Ser Leu Ser Gly Cys Ala Val
130 135 140
Gln Met Phe Leu Ser Leu Ala Met Gly Thr Thr Glu Cys Val Leu Leu
145 150 155 160
Gly Met Gln Ile Leu Ser Lys Leu Leu Leu Asp Leu Ser Cys Glu Phe
165 170 175
Ser Leu Leu Lys Ser Glu Cys His Arg Val Lys Met Gln
180 185
<210> 65
<211> 270
<212> ПРТ
<213> искусственная последовательность
<220>
<223> гибрид 3, кодируемый pTG19290
<400> 65
Ser Ser Thr Ile Met Asp Val Asp Ser Thr Ile Ser Ser Trp Arg Ser
1 5 10 15
Thr Pro Ala Met Met Asn Gly Gln Gly Ser Thr Lys Ser Lys Lys Val
20 25 30
Ser Asp Ile Lys Glu Glu Val Leu Ser Ala Ala Cys His Pro Val Gln
35 40 45
His Ser Lys Val Glu Tyr Tyr His Ile Glu Val Met Lys Lys Lys Met
50 55 60
Tyr Ile Phe Trp Gly Lys Met Ser Arg Arg Thr Leu Leu Pro Ile Pro
65 70 75 80
Thr Ser Trp Tyr Leu Glu Asn Ile Tyr Pro Asp Ser Gln Ile Thr Arg
85 90 95
Arg Tyr Tyr Ser Leu Gly Glu Ile Arg Asn Val Glu Ser Pro Phe Asn
100 105 110
Val Val Ala Trp His Gly Asn Tyr Thr Leu Tyr Lys Tyr Asn Leu Lys
115 120 125
Asn Phe Met Val Ile Asn Ser Asp Tyr Glu Leu Ala Glu Val His Lys
130 135 140
Ala Leu Leu Asp Asn Lys Asn Lys Val Leu Glu Val Lys Lys Pro Pro
145 150 155 160
Arg Phe Glu Lys Gly Pro Leu Pro Gly Glu Asp Trp Thr Ile Phe Lys
165 170 175
Tyr Asn His Ser Thr Tyr Gln Pro Val Leu Leu Thr Glu Asp Arg Trp
180 185 190
Val Arg Asn Gln Ala Asn Leu Asp Lys Glu His Val Pro Leu Leu Lys
195 200 205
Ile Glu Glu Pro Pro Ser Thr Ala Leu Asp Thr Glu Glu Glu Glu Asp
210 215 220
Asp Thr Ile His Met Trp Asn Ala Ile Met Thr Phe Tyr Ser Ala Leu
225 230 235 240
Ile Asp Leu Ser Thr Val Glu Lys His Cys Gly Lys Val Pro Tyr Pro
245 250 255
Leu Ser Tyr Val Pro Ala Val Leu Ser Glu Leu Thr Asp Gln
260 265 270
<210> 66
<211> 5
<212> ПРТ
<213> искусственная последовательность
<220>
<223> линкер из 5ак
<400> 66
Gly Ser Gly Ser Gly
1 5
<210> 67
<211> 5
<212> ПРТ
<213> искусственная последовательность
<220>
<223> линкер из 5ак
<400> 67
Ser Gly Ser Gly Ser
1 5
<210> 68
<211> 10
<212> ПРТ
<213> искусственная последовательность
<220>
<223> линкер из 10ак
<400> 68
Gly Ser Gly Ser Gly Ser Gly Ser Gly Ser
1 5 10
<210> 69
<211> 5
<212> ПРТ
<213> искусственная последовательность
<220>
<223> линкер из 5ак
<400> 69
Gly Ser Thr Ser Gly
1 5
<210> 70
<211> 5
<212> ПРТ
<213> искусственная последовательность
<220>
<223> линкер из 5 ак
<400> 70
Ser Gly Thr Gly Ser
1 5
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ КОНСТРУИРОВАНИЯ РЕКОМБИНАНТНОГО ПОКСВИРУСА ДЛЯ ТЕРАПЕВТИЧЕСКОЙ ВАКЦИНЫ | 2020 |
|
RU2827444C1 |
| КОМБИНИРОВАННАЯ ТЕРАПИЯ ДЛЯ ЛЕЧЕНИЯ РАКА ПУТЕМ ВНУТРИВЕННОГО ВВЕДЕНИЯ РЕКОМБИНАНТНОГО MVA И АНТИТЕЛА | 2018 |
|
RU2795103C2 |
| ПРОМОТОРЫ ДЛЯ УСИЛЕНИЯ ЭКСПРЕССИИ В ПОКСВИРУСАХ | 2016 |
|
RU2753884C2 |
| ДИСПЛЕЙ ИНТЕГРАЛЬНОГО МЕМБРАННОГО БЕЛКА НА ВНЕКЛЕТОЧНЫХ ОБОЛОЧЕЧНЫХ ВИРИОНАХ ПОКСВИРУСА | 2017 |
|
RU2759846C2 |
| ВАКЦИННАЯ КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ РАКА | 2018 |
|
RU2782261C2 |
| СУБЪЕДИНИЧНАЯ ВАКЦИНА ДЛЯ ЛЕЧЕНИЯ ИЛИ ПРЕДОТВРАЩЕНИЯ ИНФЕКЦИИ ДЫХАТЕЛЬНЫХ ПУТЕЙ | 2020 |
|
RU2811991C2 |
| ВАКЦИНА ПРОТИВ АФРИКАНСКОЙ ЧУМЫ СВИНЕЙ | 2020 |
|
RU2819672C2 |
| СТАБИЛИЗИРОВАННЫЕ БЕЛКИ F RSV И ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2795459C2 |
| Вакцина на основе AAV5 для индукции специфического иммунитета к вирусу SARS-CoV-2 и/или профилактики коронавирусной инфекции, вызванной SARS-CoV-2 | 2020 |
|
RU2783313C1 |
| Вакцина на основе AAV5 для индукции специфического иммунитета к вирусу SARS-CoV-2 и/или профилактики коронавирусной инфекции, вызванной SARS-CoV-2 | 2020 |
|
RU2760301C1 |

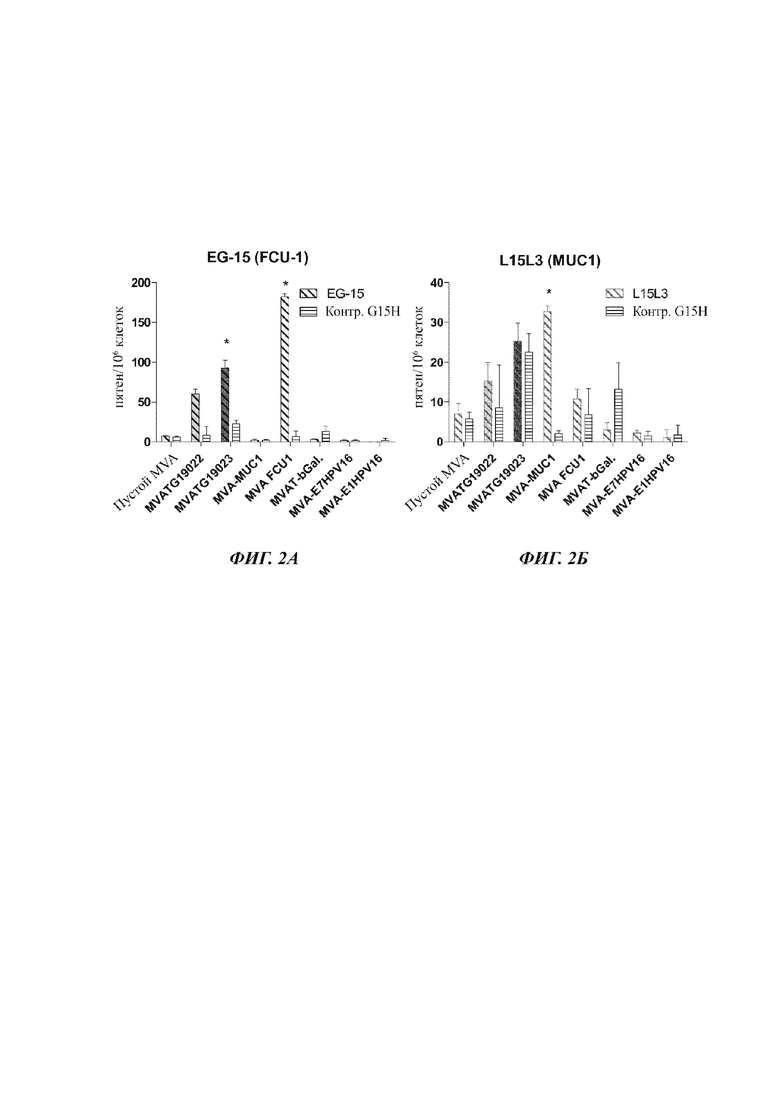
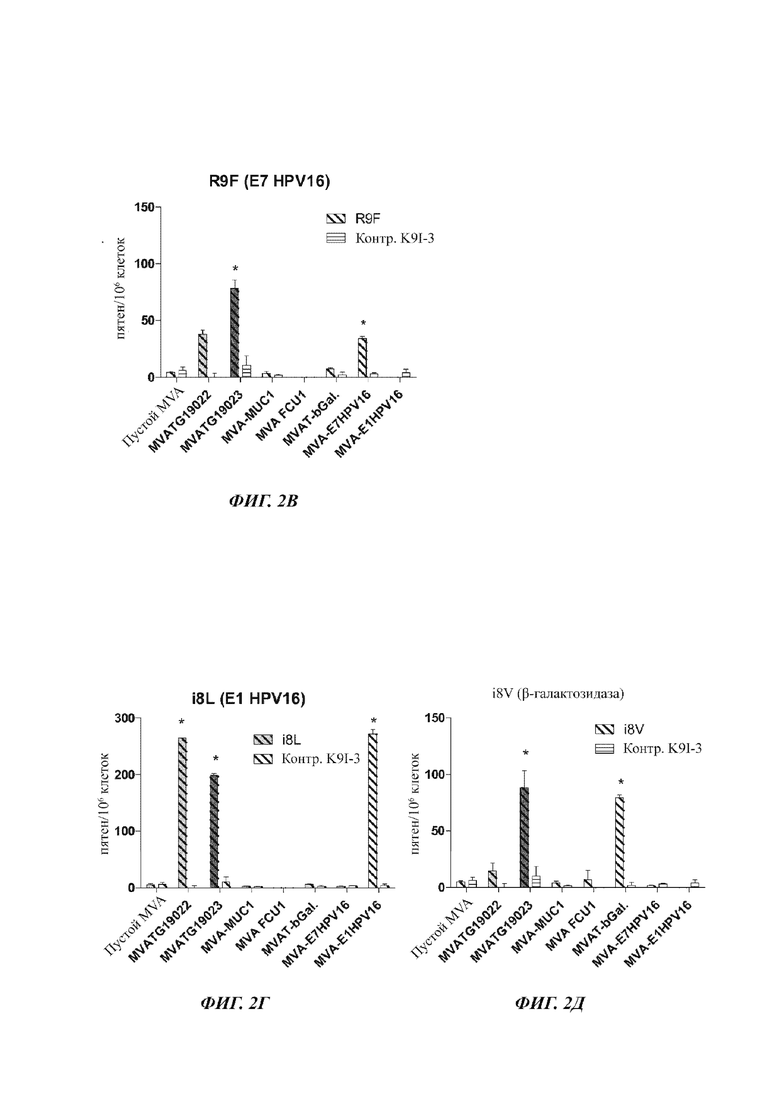

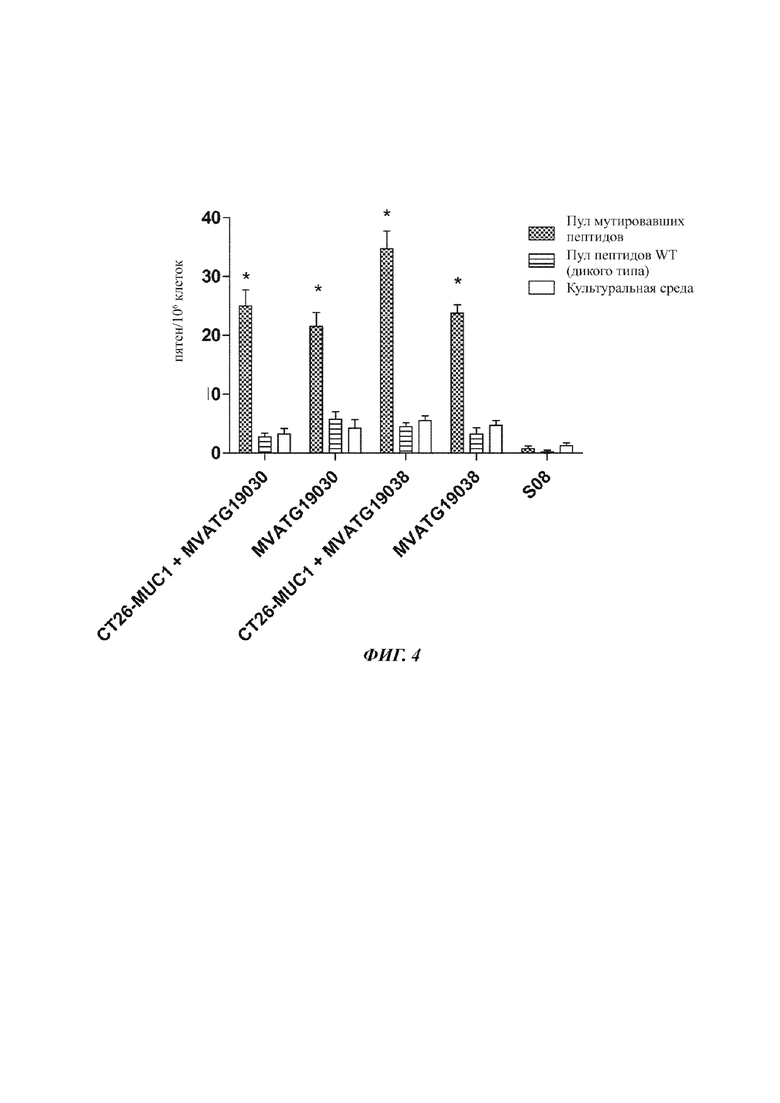
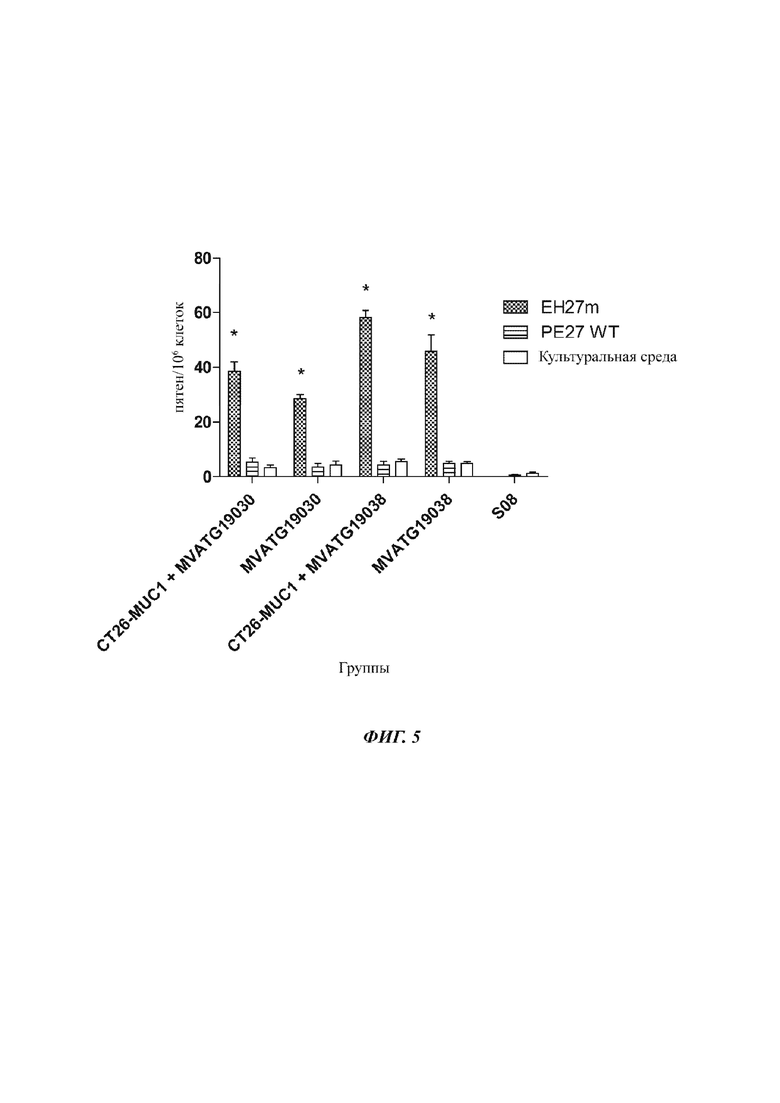
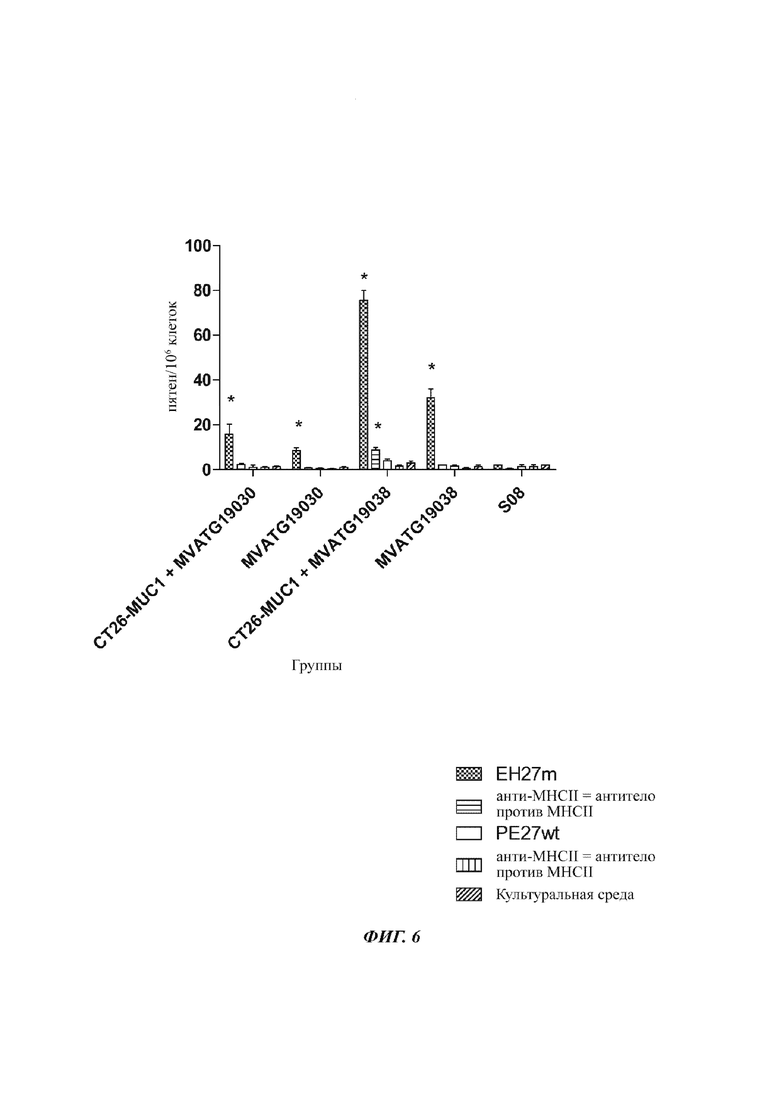

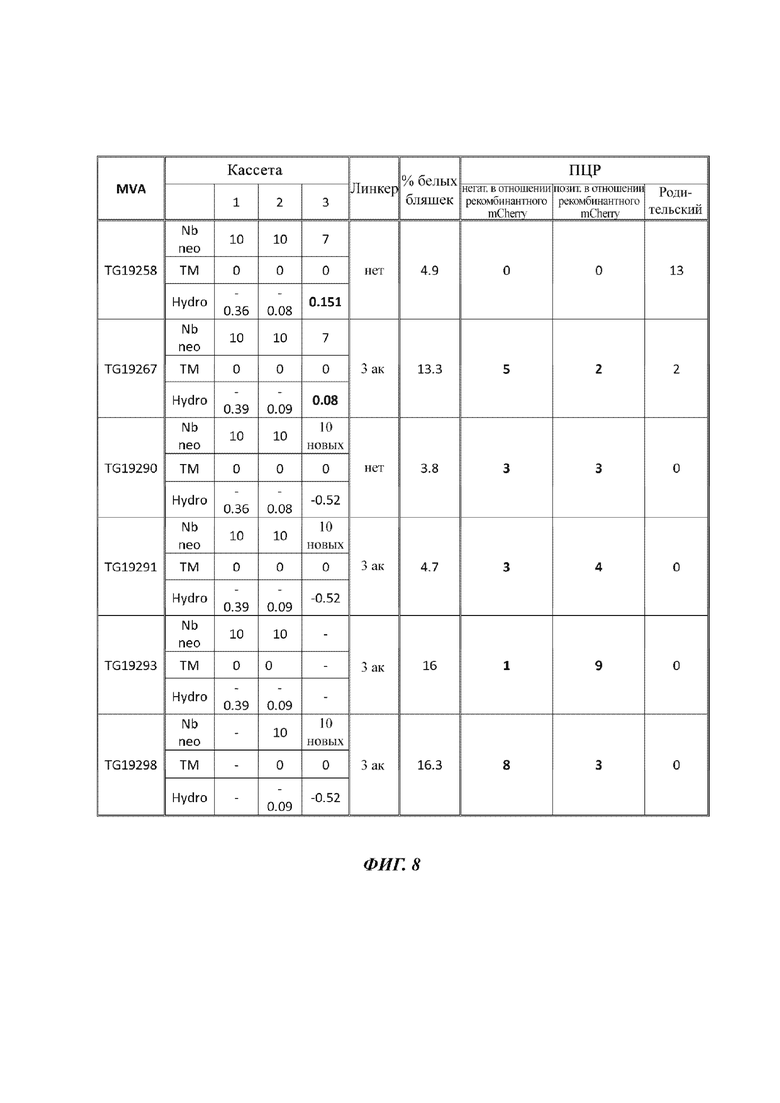
Группа изобретений относится к персонализированной противораковой вакцине, способу её получения и применению. Вакцина содержит рекомбинантный вирус осповакцины, кодирующий от 6 до 35 пептидов, где каждый пептид является фрагментом неоантигена, содержащего неоэпитоп, а также фланкирующую(-ие) последовательность(-ти) на одной или обеих сторонах этого неоэпитопа, и содержит по меньшей мере одну опухолеспецифичную мутацию, появившуюся вместе с неопластическим превращением и/или прогрессированием раковых клеток. Способ получения включает (1) стадию идентификации от 6 до 35 пептидов, подходящих для кодирования рекомбинантным вирусом осповакцины, включающую: а) экстрагирование ДНК из образца опухоли и неопухолевого образца, б) выбор областей-мишеней, в) их секвенирование, г) идентификацию одной или более чем одной опухолеспецифичной мутации посредством сравнения последовательностей ДНК, полученных из указанных образцов опухоли и неопухолевых образцов; и (2) стадию получения рекомбинантного вируса осповакцины, включающую получение синтетической нуклеиновой кислоты, кодирующей отобранные пептиды в виде одного или более чем одного слияния из 2-15 пептидов, и получение рекомбинантного пептид-экспрессирующего вируса осповакцины. Применение вакцины позволяет лечить рак или предупреждать его рецидивы, обеспечивая стимуляцию иммунного ответа Т-клеток. 4 н. и 34 з.п. ф-лы, 12 ил., 8 табл., 5 пр.
1. Персонализированная противораковая вакцина, содержащая рекомбинантный вирус осповакцины, кодирующий от 6 до 35 пептидов, где каждый пептид является фрагментом неоантигена, содержащего неоэпитоп, а также фланкирующую(-ие) последовательность(-ти) на одной или обеих сторонах этого неоэпитопа, и содержит по меньшей мере одну опухолеспецифичную мутацию, появившуюся вместе с неопластическим превращением и/или прогрессированием раковых клеток и выбранную из миссенс-мутаций, делеций, вставок, мутаций со сдвигом рамки считывания и мутаций в сайте сплайсинга; по меньшей мере 60% указанных пептидов содержит миссенс-мутацию или мутацию со сдвигом рамки считывания; и несколько пептидов экспрессируется рекомбинантным вирусом осповакцины в виде одного или более слияний из 2-15 пептидов, при этом указанное слияние не содержит какого-либо трансмембранного (ТМ) сегмента.
2. Персонализированная противораковая вакцина по п. 1, в которой указанный рекомбинантный вирус осповакцины представляет собой дефектный по репликации вирус осповакцины.
3. Противораковая вакцина по п. 2, где указанный дефектный по репликации вирус осповакцины представляет собой модифицированный вирус осповакцины Анкара (MVA).
4. Персонализированная противораковая вакцина по любому из пп. 1-3, в которой по меньшей мере 80% указанных пептидов содержат миссенс-мутацию или мутацию со сдвигом рамки считывания.
5. Персонализированная противораковая вакцина по п. 4, в которой по меньшей мере 60% указанных пептидов содержат миссенс-мутацию.
6. Персонализированная противораковая вакцина по любому из пп. 1-5, в которой указанные пептиды имеют длину от 16 до 90 аминокислотных остатков, предпочтительно от 17 до 85 аминокислотных остатков и более предпочтительно от 18 до 80 аминокислотных остатков.
7. Персонализированная противораковая вакцина по любому из пп. 4-6, в которой указанные пептиды, содержащие миссенс-мутацию, имеют длину от 18 до 29 остатков и указанные пептиды, содержащие мутацию со сдвигом рамки считывания, имеют длину от 30 до 80 аминокислотных остатков.
8. Персонализированная противораковая вакцина по любому из пп. 4-7, в которой по меньшей мере 80%, предпочтительно по меньшей мере 85%, более предпочтительно по меньшей мере 90% пептидов, имеющих миссенс-мутацию, несут замененную аминокислоту в центральном положении.
9. Персонализированная противораковая вакцина по любому из пп. 1-8, в которой все пептиды экспрессируются рекомбинантным вирусом осповакцины в виде одного или более чем одного слияния из 2-15 пептидов.
10. Персонализированная противораковая вакцина по любому из пп. 1-9, в которой указанное слияние дает отрицательный индекс в отношении гидрофобности.
11. Персонализированная противораковая вакцина по любому из пп. 1-9, в которой указанное слияние демонстрирует индекс гидропатичности, равный или меньший 0,1.
12. Персонализированная противораковая вакцина по любому из пп. 1-11, в которой рекомбинантный вирус осповакцины кодирует 2 или 3 слияния, где:
каждое слияние содержит от 5 до 10 пептидов;
каждое слияние содержит линкеры, предпочтительно длиной 3 аминокислоты, на N-конце первого пептида и между каждым пептидом;
каждое слияние содержит сигнальный пептид на N-конце указанного слияния;
каждое слияние возможно содержит метку на C-конце указанного слияния; и
каждое слияние:
демонстрирует общий отрицательный индекс гидрофобности или общий индекс гидропатичности, равный или меньший 0,1, и
не содержит какого-либо потенциального ТМ сегмента.
13. Персонализированная противораковая вакцина по любому из пп. 1-12, в который указанный рекомбинантный вирус осповакцины представляет собой рекомбинантный MVA, содержащий от одной до трех кассет – каждая для экспрессии слияния из 5-10 пептидов – под транскрипционным контролем промотора вируса осповакцины, выбранного из группы, состоящей из p11k7.5, pH5R, p7.5K и pB2R.
14. Персонализированная противораковая вакцина по любому из пп. 1-13, в который указанный рекомбинантный вирус осповакцины дополнительно кодирует один или более чем один терапевтический ген, предпочтительно выбранный из группы, состоящей из суицидных и иммуностимулирующих генов.
15. Способ получения персонализированной противораковой вакцины по любому из пп. 1-14, включающий:
(1) стадию идентификации от 6 до 35 пептидов, подходящих для кодирования рекомбинантным вирусом осповакцины, где указанные пептиды содержат по меньшей мере одну опухолеспецифичную мутацию, появившуюся вместе с неопластическим превращением и/или прогрессированием раковых клеток и выбранную из миссенс-мутаций, делеций, вставок, мутаций со сдвигом рамки считывания и мутаций в сайте сплайсинга, при этом по меньшей мере 60% указанных пептидов содержат миссенс-мутацию или мутацию со сдвигом рамки считывания, где указанная стадия идентификации включает в себя следующие подстадии а)-г):
а) экстрагирование ДНК из указанного образца опухоли и указанного неопухолевого образца,
б) выбор областей-мишеней, предпочтительно полных кодирующих областей генома (экзом),
в) секвенирование указанных областей-мишеней (например, экзома) из указанной экстрагированной ДНК и
г) идентификация одной или более чем одной опухолеспецифичной мутации посредством сравнения последовательностей ДНК, полученных из указанных образцов опухоли и неопухолевых образцов; и
(2) стадию получения указанного рекомбинантного вируса осповакцины, включающую получение синтетической нуклеиновой кислоты, кодирующей отобранные пептиды в виде одного или более чем одного слияния из 2-15 пептидов, где указанное слияние не содержит трансмембранного (ТМ) сегмента, и получение рекомбинантного пептид-экспрессирующего вируса осповакцины.
16. Способ по п. 15, в котором указанный образец опухоли представляет собой биопсию опухоли, предпочтительно содержащую больше 20% опухолевых клеток, и где указанный неопухолевый образец представляет собой биологическую жидкость, цитологический материал или биопсию.
17. Способ по любому из пп. 15, 16, в котором указанная стадия идентификации дополнительно включает по меньшей мере одну из следующих дополнительных подстадий д) – и):
д) ранжирование потенциального(ных) пептида(дов) по их уровням экспрессии в опухоли либо на уровне транскрипции мРНК, либо на уровне трансляции белка;
е) отбор неаутоэкспрессируемой(мых) опухолеспецифичной(ных) мутации(ций);
ж) прогнозирование иммунного потенциала неоэпитопа(пов), содержащегося(щихся) в указанном(ных) пептиде(дах);
з) прогнозирование присутствия потенциальных(ного) ТМ сегмента(тов) в самих неоэпитопах и/или слияниях пептидов, содержащих несколько пептидов;
и) ранжирование потенциального(ных) пептида(дов) и/или слияния(ний) пептидов по их степени гидрофобности и, в частности, отбор пептида(дов) и/или слияния(ний) пептидов таким образом, что указанный(ные) пептид(ды) и/или слияние(ния) пептидов демонстрирует(ют) отрицательный общий индекс гидрофобности и/или общий индекс гидропатичности, равный или меньший 0,1.
18. Способ по любому из пп. 15-17, в котором молекула(лы) нуклеиновой кислоты, кодирующая(щие) указанный один или более чем один пептид, подлежащая(щие) вставке в геном рекомбинантного вируса осповакцины, организуется(ются) в одной или более чем одной экспрессионной кассете под контролем подходящих регуляторных элементов, обеспечивающих экспрессию у субъекта.
19. Способ по п. 17 или 18, в котором указанная(ные) молекула(лы) нуклеиновой кислоты или экспрессионная(ные) кассета(ты), кодирующая(щие) указанные пептиды, вставляется(ются) в геном родительского вируса осповакцины с образованием указанного рекомбинантного вируса осповакцины, и предпочтительно где указанный родительский вирус осповакцины содержит ген флуоресцентного репортера, клонированный в месте вставки, который отбирается на молекулу(лы) нуклеиновой кислоты или экспрессионную(ные) кассету(ты), кодирующую(щие) пептид.
20. Способ по п. 19, в котором указанный флуоресцентный репортер выбран из группы, состоящей из GFP (зеленый флуоресцентный белок), eGFP (усиленный зеленый флуоресцентный белок), флуоресцентного белка AmСyan 1 и mCherry.
21. Способ по п. 20, в котором указанная стадия получения указанного рекомбинантного вируса осповакцины включает дополнительную стадию расщепления эндонуклеазой, способной генерировать по меньшей мере один двухнитевой разрыв в указанной нуклеотидной последовательности флуоресцентного репортера, но в которой указанная эндонуклеаза не расщепляет геном вируса осповакцины.
22. Способ по п. 21, в котором указанная эндонуклеаза выбрана из группы, состоящей из нуклеаз с цинковыми пальцами (ZFN), эффекторных нуклеаз, подобных активатору транскрипции (TALEN), нуклеаз кластерных коротких палиндромных повторов, разделенных регулярными промежутками (CRISPR)/Cas9, и рестрикционных ферментов с уникальным сайтом расщепления в пределах гена флуоресцентного репортера.
23. Способ по любому из пп. 15-22, в котором указанная стадия получения рекомбинантного вируса осповакцины обеспечивает получение указанного рекомбинантного вируса осповакцины самое большее за 4 недели, предпочтительно самое большее за 3 недели с отношением рекомбинантного к родительскому, составляющим от 2 до 0,05.
24. Способ по любому из пп. 15-23, где указанный способ дополнительно включает стадию изготовления, где указанная стадия изготовления включает стадию амплификации в подходящей клетке-продуценте в подходящем масштабе, стадию выделения продуцированного рекомбинантного вируса осповакцины из культуры клеток и возможную стадию очистки выделенного рекомбинантного вируса осповакцины.
25. Способ по п. 24, в котором указанная клетка-продуцент представляет собой фибробласты эмбриона курицы (CEF).
26. Способ по п. 24 или 25, в котором указанная стадия выделения продуцированного рекомбинантного вируса осповакцины включает стадию лизиса, в которой мембрана клетки-продуцента разрушается, предпочтительно с использованием высокоскоростного гомогенизатора.
27. Способ по любому из пп. 24-26, в котором указанная стадия очистки включает стадию фильтрования тангенциальным потоком (TFF).
28. Способ по любому из пп. 24-27, в котором в указанной стадии изготовления достигается продукция по меньшей мере 109 БОЕ (бляшкообразующая единица), желательно по меньшей мере 5х109 БОЕ и предпочтительно приблизительно 1010 БОЕ или более для персонализированной противораковой вакцины.
29. Персонализированная противораковая вакцина, полученная способом по любому из пп. 15-28 и содержащая рекомбинантный вирус осповакцины, кодирующий от 6 до 35 пептидов, где каждый пептид является фрагментом неоантигена, содержащего неоэпитоп, а также фланкирующую(-ие) последовательность(-ти) на одной или обеих сторонах этого неоэпитопа, и содержит по меньшей мере одну опухолеспецифичную мутацию, появившуюся вместе с неопластическим превращением и/или прогрессированием раковых клеток и выбранную из миссенс-мутаций, делеций, вставок, мутаций со сдвигом рамки считывания и мутаций в сайте сплайсинга; по меньшей мере 60% указанных пептидов содержит миссенс-мутацию или мутацию со сдвигом рамки считывания; и несколько пептидов экспрессируется рекомбинантным вирусом осповакцины в виде одного или более слияний из 2-15 пептидов, при этом указанное слияние не содержит какого-либо трансмембранного (ТМ) сегмента.
30. Персонализированная противораковая вакцина по любому из пп. 1-14 и 29, где указанная персонализированная противораковая вакцина представляет собой композицию, содержащую терапевтически эффективное количество указанного рекомбинантного вируса осповакцины и фармацевтически приемлемый носитель.
31. Персонализированная противораковая вакцина по п. 30, в которой указанное терапевтически эффективное количество для индивидуальных доз составляет от приблизительно 5х107 БОЕ до приблизительно 109 БОЕ.
32. Применение персонализированной противораковой вакцины по любому из пп. 30 и 31 для изготовления лекарственного средства для применения у субъекта, нуждающегося в этом, для лечения рака или предупреждения его рецидива у субъекта.
33. Применение по п. 32, где указанный рак представляет собой солидную опухоль.
34. Применение по п. 33, где указанный рак представляет собой рак мозга, такой как, например, астроцитомы, эмбриональные опухоли, герминомы, атипическая тератоидная/рабдоидная опухоль центральной нервной системы, краниофарингиома, эпендимома, глиома и глиобластома, а также рак головы и шеи.
35. Применение по п. 33, где указанный рак представляет собой рак легкого и, в частности, NSCLC (немелкоклеточный рак легкого) с особым предпочтением в отношении аденокарциномы, плоскоклеточной карциномы и крупноклеточной карциномы.
36. Применение по п. 33, где указанный рак представляет собой рак яичника.
37. Применение по любому из пп. 32-36, где указанную персонализированную противораковую вакцину вводят пациенту посредством внутривенных, подкожных, внутримышечных или внутриопухолевых инъекций.
38. Применение по любому из пп. 32-37, где указанную персонализированную противораковую вакцину вводят в сочетании с одной или более чем одной дополнительной противораковой терапией/ями, которые имеют пользу в лечении рака.
| WO 2015085233 A1, 11.06.2015 | |||
| WO 2014168874, 16.10.2014 | |||
| SEBASTIAN KREITER et al | |||
| "Mutant MHC class II epitopes drive therapeutic immune responses to cancer", NATURE, vol | |||
| Способ обогащения руд флотацией (отпениванием) с прибавлением масла | 1917 |
|
SU520A1 |
| УСТАНОВКА ДЛЯ НАНЕСЕНИЯ ПОКРЫТИЯ | 1997 |
|
RU2199400C2 |
| WO 2007147528 A1, 27.12.2007 | |||
| ВАКЦИНА НА ОСНОВЕ В-КЛЕТОК, НАГРУЖЕННЫХ ЛИГАНДОМ Т-КЛЕТОК-ПРИРОДНЫХ КИЛЛЕРОВ И АНТИГЕНОМ | 2006 |
|
RU2420311C2 |

