ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение, в общем, относится к области онколитической виротерапии и, более конкретно, к композициям и способам для лечения, предупреждения или ингибирования пролиферативных заболеваний, особенно рака. Воплощения включают онколитический вирус, содержащий нуклеотидную(ные) последовательность(сти), кодирующую(щие) один или более чем один модулятор иммунологической контрольной точки. Воплощения также включают фармацевтическую композицию, содержащую такой онколитический вирус и, в конечном счете, фармацевтически приемлемый носитель и его применение для лечения пролиферативных заболеваний, таких как раковые заболевания.
Рак вызывается как внешними факторами (например, табак, инфекционные организмы, характер питания, химические соединения и радиация), так и внутренними факторами (например, наследственные мутации, гормоны, иммунные состояния и мутации, которые возникают из-за метаболизма). Каждый год во всем мире рак диагностируется у более чем 12 миллионов субъектов. В промышленно развитых странах приблизительно один человек из пяти умрет от рака. Несмотря на существование огромного числа химиотерапевтических средств, они часто являются неэффективными, особенно против злокачественных и метастатических опухолей, которые развиваются на очень ранней стадии заболевания. Кроме того, противоопухолевый иммунитет часто является неэффективным из-за того факта, что опухолевые клетки развили механизмы для того, чтобы избегать защиты хозяина. Одним из главных механизмов подавления иммунитета является процесс, известный как «истощение Т-клеток», который возникает из-за хронического воздействия антигенов и характеризуется повышающей регуляцией ингибирующих рецепторов. Данные ингибирующие рецепторы служат в качестве иммунологических контрольных точек для того, чтобы предотвращать неконтролируемые иммунные реакции. В литературе были описаны разные иммунологические контрольные точки, действующие на разных уровнях Т-клеточного иммунитета, включая белок 1 программируемой клеточной смерти (PD-1) и его лиганды PD-L1 и PD-L2, CTLA-4 (белок-4, ассоциированный с цитотоксическими Т-лимфоцитами), LAG3 (ген 3, активирующий лимфоциты), аттенюатор В- и Т-лимфоцитов, иммуноглобулин Т-клеток, белок 3, содержащий домен муцина (TIM-3), и иммуноглобулин-супрессор активации Т-клеток с V-доменом.
Каким бы ни был механизм действия, данные иммунологические контрольные точки могут ингибировать развитие эффективного противоопухолевого иммунного ответа. Имеется возрастающий интерес к возможной терапевтической пользе блокирования таких иммунологических контрольных точек в качестве средства ингибирования толерантности иммунной системы к опухолям и, таким образом, спасения истощенных противоопухолевых Т-клеток (Leach et al., 1996, Science 271: 1734-6). На протяжении последнего десятилетия было разработано большое число антагонистических антител (например, против Tim3, PD-L1, CTLA-4, PD1 и т.д.), и важнее всего то, что некоторые из них были ассоциированы с объективными клиническими ответами у раковых пациентов. Антитела, нацеленные на CTLA-4 (например, ипилимумаб, ервой, Bristol-Myers Squibb (BMS)), уже распространяются на рынке против метастатической меланомы. BMS сообщала о том, что из 1800 пациентов с меланомой, которых лечили ипилимумабом, 22% все еще живы через 3 года. Также продолжаются терапии антителами против PD-L1 (например, MPDL3280A, Roche), против PD-1 (например, ниволумаб, BMS).
Другим терапевтическим подходом, который возникает в области рака, являются онколитические вирусы (Hermiston, 2006, Curr. Opin. Mol. Ther. 8: 322-30). Онколитические вирусы способны к селективной репликации в делящихся клетках (например, в раковой клетке), оставляя неделящиеся клетки (например, нормальные клетки) невредимыми. Поскольку инфицированные делящиеся клетки разрущаются лизисом, они высвобождают новые инфекционные вирусные частицы для инфицирования окружающих делящихся клеток. Раковые клетки представляют собой идеальных хозяев для многих вирусов, так как они имеют инактивированный противовирусный интерфероновый путь или имеют мутировавшие гены супрессоров опухолей, что обеспечивает беспрепятственный ход вирусной репликации (Chernajovsky et al., 2006, British Med. J. 332: 170-2). В настоящее время в качестве онколитических агентов был клинически испытан целый ряд вирусов, включая аденовирус, реовирус, вирус кори, вирус простого герпеса, вирус ньюкаслской болезни и вирус осповакцины.
Некоторые вирусы являются онколитическими в природе (такие как реовирус и пикорнавирус долины Сенека), тогда так другие являются генетически модифицированными для селективности в отношении опухоли посредством модифицирования вирусного генома. Такие модификации включают функциональные делеции важных вирусных генов, применение опухоле- или тканеспецифичных промоторов для контроля экспрессии вирусных генов и модификации тропизма для перенаправления вируса к поверхности раковых клеток.
Первым онколитическим вирусом, подлежащим одобрению надзорным органом, был генетически модифицированный аденовирус, названный Н101 (Shanghai Sunway Biotech), который получил одобрение от Государственного управления Китая по контролю качества продуктов питания и лекарственных средств (SFDA) в 2005 году для лечения рака головы и шеи. Другой онколитический аденовирус, названный ONYX-015, находится на этапе продолжающихся клинических испытаний для лечения разных солидных опухолей (в фазе III для лечения рецидивирующего рака головы и шеи) (Cohen et al., 2001, Curr. Opin. Investig. Drugs 2: 1770-5). В качестве другого примера, онколитический вирус простого герпеса 1 (T-VEC) был генетически модифицирован для ослабления вирулентости вируса, увеличения селективности в отношении раковых клеток и усиления противоопухолевого иммунного ответа (посредством экспрессии GM-CSF (гранулоцитарно-макрофагальный колониестимулирующий фактор)). Клиническая эффективность при нерезецируемой меланоме была продемонстрирована в клинических испытаниях фазы II и фазы III (Senzer et al, 2009, J. Clin. Oncol. 27: 5763-71).
Вирусы осповакцины (VV) обладают многими из ключевых характеристик, необходимых для применения в онколитической виротерапии, такими как природный тропизм в отношении опухолей, сильная литическая способность, короткий жизненный цикл с быстрым распространением от клетки к клетке, высокоэффективная экспрессия генов и большая способность к клонированию. Кроме того, их доставляли миллионам индивидов во время кампании по искоренению натуральной оспы без большой озабоченности относительно безопасности. В данном отношении VV, экспрессирующий GM-CSF, с двойной делецией по ТК (тимидинкиназа) и VGF (фактор роста VV) (названный JX-963) демонстрировал значимую селективность в отношении рака у мышей, несущих опухоль (Thome et al., 2007, J Clin Invest. 117: 3350-8). На той же самой линии - JX-594 - VV с делецией ТК (штамм Wyeth), снабженный GM-CSF, были показаны многообещающие клинические данные, и скоро ожидается начало рандомизированного исследования фазы III в печеночно-клеточной карциноме.
В литературе были описаны комбинированные терапии, включающие онколитический вирус и ингибиторы иммунологических контрольных точек. В WO 2014/022138 описана комбинация облученных опухолевых клеток, онколитического аденовируса и антитела против CTLA4 для применения для лечения рака мочевого пузыря или предстательной железы. В WO 2014/047350 рассматривается рекомбинантный онколитический вирус с геном, кодирующим антитело против PD-1, вставленным в вирусный геном, без предоставления какого-либо рабочего примера, который поддерживал бы пользу такого онколитического вируса.
Техническая проблема
Можно ожидать, что рак продолжит быть серьезной глобальной угрозой для здоровья на много лет из-за большого числа причинных факторов, которые могут действовать совместно или по отдельности с инициацией или стимулированием развития рака. Кроме того, злокачественные и особенно метастатические опухоли часто являются устойчивыми к традиционным терапиям, объясняя значительный процент смертности от некоторых раковых заболеваний.
Таким образом, существует важная потребность в разработке более эффективных подходов для улучшения, предупреждения и лечения таких пролиферативных заболеваний и особенно метастатических раковых заболеваний. Согласно настоящему изобретению предложен уникальный продукт, комбинирующий онколиз для умерщвления делящихся клеток и иммунологическую контрольную точку для нарушения ассоциированной с раком иммунологической толерантности.
Данная техническая проблема решается предложением воплощений, как определено в формуле изобретения.
Другие и дополнительные аспекты, характеристики и преимущества настоящего изобретения будут очевидными из следующего описания предпочтительных в настоящее время воплощений изобретения. Данные воплощения приведены с целью раскрытия.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Настоящее изобретение касается онколитического вируса, содержащего одну или более чем одну молекулу нуклеиновой кислоты, вставленную в его геном, кодирующую один или более чем один модулятор иммунологической контрольной точки.
Онколитический вирус предпочтительно выбран из группы, состоящей из реовируса, вируса ньюкаслской болезни (NDV), вируса везикулярного стоматита (VSV), вируса кори, вируса гриппа, вируса Синдбис, аденовируса, вируса группы оспы, вируса герпеса (HSV) и тому подобных. В одном воплощении онколитический вирус представляет собой вирус осповакцины. В предпочтительном воплощении вирус осповакцины генетически модифицирован таким образом, чтобы он не имел активности тимидинкиназы (ТК) (например, геном указанного VV имеет инактивирующую мутацию в гене J2R с получением дефектного по ТК фенотипа). В качестве альтернативы или в комбинации, вирус осповакцины генетически модифицирован таким образом, чтобы он не имел активности RR (например, геном указанного VV имеет инактивирующую мутацию в гене I4L и/или F4L с получением дефектного по RR фенотипа).
В одном воплощении вирус осповакцины дополнительно экспрессирует по меньшей мере один терапевтический ген, в частности ген, кодирующий суицидный генный продукт и/или иммуностимулирующий белок.
В одном воплощении один или более чем один кодируемый модулятор иммунологической контрольной точки представляет собой антагонистическую молекулу, которая оказывает антагонистический эффект на активность PD-1, PD-L1 или CTLA4 с конкретным предпочтением в отношении антитела против PD-1 и/или антитела против CTLA4.
Согласно настоящему изобретению дополнительно предложена композиция, содержащая указанный онколитический вирус, в конечном счете, с фармацевтически приемлемым носителем. В одном воплощении данная композиция приготовлена для внутривенного или внутриопухолевого введения.
Настоящее изобретение также касается применения указанного онколитического вируса или его композиции для лечения пролиферативного заболевания, а также способа лечения, полагающегося на введение эффективного количества указанного онколитического вируса или его композиции. В одном воплощении пролиферативное заболевание, которое лечат способом по изобретению, представляет собой рак и особенно меланому, рак почки, рак предстательной железы, рак молочной железы, колоректальный рак, рак легкого и рак печени. В одном воплощении применение или способ включает дополнительную стадию, на которой указанному млекопитающему вводится фармацевтически приемлемое количество пролекарства. Введение указанного пролекартва предпочтительно происходит по меньшей мере через 3 суток после введения указанного онколитического вируса или композиции вируса.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение касается онколитического вируса, содержащего одну или более чем одну молекулу нуклеиновой кислоты, вставленную в его геном, кодирующую один или более чем один модулятор иммунологической контрольной точки.
Определения
Термины, указывающие на единственное число, в том виде, в котором они используются везде во всей данной заявке, используются в том смысле, что они означают «по меньшей мере один», «по меньшей мере первый», «один или более чем один» или «множество» упоминаемых компонентов или стадий, если контекст явно не диктует иное. Например, термин «клетка» включает множество клеток, включая их смеси.
Термин «один или более чем один» относится либо к одному, либо к числу, превышающему один (например, 2, 3, 4, 5 и т.д.).
Термин «и/или» всякий раз, когда он используется в данном документе, включает значение «и», «или» и «все или любая другая комбинация элементов, соединенных указанным термином».
Термин «примерно» или «приблизительно» в том виде, как он используется в данном документе, означает в пределах 20%, предпочтительно в пределах 10% и более предпочтительно в пределах 5% от данного значения или интервала.
Термин «содержащий» (и любая форма содержащего, такая как «содержать» и «содержит»), «имеющий» (и любая форма имеющего, такая как «иметь» и «имеет»), «включающий» (и любая форма включающего, такая как «включает» и «включать») в том виде, как они используются в данном документе, при использовании для определения продуктов, композиций и способов, являются открытыми и не исключают дополнительных, неперечисленных элементов или стадий способа. Таким образом, полипептид «содержит» аминокислотную последовательность, когда аминокислотная последовательность может быть частью конечной аминокислотной последовательности полипепида. Такой полипептид может иметь вплоть до нескольких сотен дополнительных аминокислотных остатков. Фраза «по существу состоящий из» означает исключение других компонентов или стадий, имеющих какую-либо существенную значимость. Таким образом, композиция, по существу состоящая из перечисленных компонентов, не исключала бы следовые примеси и фармацевтически приемлемые носители. Полипептид «по существу состоит из» аминокислотной последовательности, когда такая аминокислотная последовательность присутствует с фактически лишь несколькими дополнительными аминокислотными остатками. «Состоящий из» означает исключение более чем следовых элементов других компонентов или стадий. Например, полипептид «состоит из» аминокислотной последовательности, когда данный полипептид не содержит любых аминокислот, кроме перечисленной аминокислотной последовательности.
Термины «полипептид», «пептид» и «белок» относятся к полимерам из аминокислотных остатков, которые содержат по меньшей мере девять или более чем девять аминокислот, связанных посредством пептидных связей. Полимер может быть линейным, разветвленным или циклическим и может содержать встречающиеся в природе аминокислоты и/или аналоги аминокислот, и он может прерываться неаминокислотами. В качестве общего указания, если аминокислотный полимер состоит из более чем 50 аминокислотных остатков, он предпочтительно называется полипептидом или белком, тогда как, если он имеет длину 50 аминокислот или менее, он называется «пептидом».
В контексте настоящего изобретения термины «нуклеиновая кислота», «молекула нуклеиновой кислоты», «полинуклеотид» и «нуклеотидная последовательность» используются взаимозаменяемо и определяют полимер любой длины одного из полидезоксирибонуклеотидов (ДНК) (например, кДНК (комплементарная ДНК), геномная ДНК, плазмиды, векторы, вирусные геномы, выделенная ДНК, зонды, праймеры и их любая смесь) или полирибонуклеотидов (РНК) (например, мРНК (матричная РНК), антисмысловая РНК, миРНК (малая интерферирующая РНК)), или смешанных полирибо-полидезоксирибонуклеотидов. Они охватывают одно- или двухцепочечные, линейные или кольцевые, природные или синтетические, модифицированные или немодифицированные полинуклеотиды. Кроме того, полинуклеотид может содержать не встречающиеся в природе нуклеотиды и может прерываться ненуклеотидными компонентами.
Термин «аналог» или «вариант» в том виде, как он используется в данном документе, относится к молекуле (полипептиду или нуклеиновой кислоте), демонстрирующей одну или более чем одну модификацию по отношению к нативному аналогу. Может(гут) рассматриваться любая(бые) модификация(ции), включающая(щие) замену, вставку и/или делецию одного или более чем одного нуклеотида/аминокислотного остатка. Предпочтительными являются аналоги, которые сохраняют степень идентичности последовательности по меньшей мере 80%, предпочтительно по меньшей мере 85%, более предпочтительно по меньшей мере 90% и даже более предпочтительно по меньшей мере 98% идентичности с последовательностью нативного аналога.
В общем, термин «идентичность» относится к соответствию аминокислоты аминокислоте или нуклеотида нуклеотиду между двумя последовательностями полипептидов или нуклеиновых кислот. Процентная доля идентичности между двумя последовательностями представляет собой функцию числа идентичных положений, которые имеют последовательности, принимая во внимание число пробелов, которые необходимо вводить для оптимального выравнивания, и длину каждого пробела. Для определения процентной доли идентичности между аминокислотными последовательностями в данной области доступны разные компьютерные программы и математические алгоритмы, такие как, например, программа Blast, доступная в NCBI (Национальный центр биотехнологической информации) или ALIGN в Атласе последовательности и структуры белка (Dayhoffed, 1981, Suppl., 3: 482-9). В специализированных базах данных также доступны программы для определения идентичности между нуклеотидными последовательностями (например, Genbank, the Wisconsin Sequence Analysis Package, программы BESTFIT, FASTA и GAP). Для иллюстративных целей «по меньшей мере 80%-ная идентичность» означает 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%.
Термин «выделенный» в том виде, как он используется в данном документе, относится к белку, полипептиду, пептиду, полинуклеотиду, вектору и т.д., который удаляется из его природного окружения (т.е., отделяется от по меньшей мере одного другого компонента, с которым он ассоциирован в природе или обнаруживается в природе). Например, нуклеотидная последовательность является выделенной, когда она отделяется от последовательностей, обычно ассоциированных с ней в природе (например, выделяется из генома), но она может быть ассоциирована с гетерологичными последовательностями.
Термин «полученный из», «происходящий» или «происходить» используется для идентификации исходного источника компонента (например, полипептида, молекулы нуклеиновой кислоты), но подразумевается, что он не ограничивается способом, посредством которого получается компонент, который может представлять собой, например, химический синтез или способы генной инженерии.
Термин «клетка-хозяин» в том виде, как он используется в данном документе, следует понимать в широком смысле без какого-либо ограничения, касающегося конкретной организации в ткани, органе или выделенных клетках. Такие клетки могут представлять собой уникальный тип клеток или группу разных типов клеток, таких как линии культивируемых клеток, первичные клетки и делящиеся клетки. В контексте данного изобретения термин «клетки-хозяева» включает прокариотические клетки, клетки низших эукариот, таких как дрожжи, и другие эукариотические клетки, такие как клетки насекомых, растительные клетки и клетки млекопитающих (например, человека или млекопитающего, не являющегося человеком), а также клетки, способные продуцировать онколитический вирус и/или модулятор(ры) иммунологической контрольной точки для применения в данном изобретении. Данный термин также включает клетки, которые могут быть или были реципиентом векторов, описанных в данном документе, а также потомство таких клеток.
Термин «онколитический вирус» в том виде, как он используется в данном документе, относится к вирусу, способному селективно реплицироваться в делящихся клетках (например, в пролиферирующей клетке, такой как раковая клетка) с целью замедления роста и/или лизирования указанной делящейся клетки, либо in vitro, либо in vivo, при демонстрации отсутствия репликации или минимальной репликации в неделящихся клетках. Типично онколитический вирус содержит вирусный геном, упакованный в вирусную частицу (или вирион), и является инфекционным (т.е. способным инфицировать и поступать в клетку-хозяина или субъекта). Данный термин в том виде, как он используется в данном документе, охватывает вектор на основе ДНК или РНК (в зависимости от рассматриваемого вируса), а также вирусные частицы, генерированные из него.
Термин «лечение» (и любая форма лечения, такая как «осуществлять лечение», «лечить») в том виде, как он используется в данном документе, охватывает профилактику (например, предупредительные меры у субъекта, подверженного риску наличия патологического состояния, подлежащего лечению) и/или терапию (например, у субъекта, у которого поставлен диагноз наличия патологического состояния), в конечном счете, в ассоциации с традиционными терапевтическими способами воздействия. Результатом лечения является замедление, излечение, уменьшение интенсивности или контроль развития целевого патологического состояния. Например, субъекта успешно лечат против рака, если после введения онколитического вируса, как описано в данном документе, субъект демонстрирует наблюдаемое улучшение его клинического статуса.
Термин «осуществление введения» (или любая форма термина «введение», такая как «введенный») в том виде, как он используется в данном документе, относится к доставке субъекту терапевтического агента, такого как описанный в данном документе онколитический вирус.
Термин «пролиферативное заболевание» в том виде, как он используется в данном документе, охватывает любое заболевание или состояние, возникающее в результате неконтролируемого роста и распространения клеток, включая раковые заболевания, а также заболевания, ассоциированные с повышенной активностью остеокластов (например, ревматоидный артрит, остеопороз и т.д.), и сердечнососудистые заболевания (рестеноз, который возникает из-за пролиферации гладкомышечных клеток стенки кровеносного сосуда и т.д.). Термин «рак» можно использовать взаимозаменяемо с любым из терминов «опухоль», «злокачественное образование», «новообразование» и т.д. Подразумевается, что данные термины включают любой тип ткани, органа или клетки, любую стадию развития злокачественного образования (например, от предопухолевой стадии до стадии IV).
Термин «субъект» обычно относится к организму, для которого нужен или может быть полезен любой продукт и способ по изобретению. Типично организм представляет собой млекопитающее, в частности млекопитающее, выбранное из группы, состоящей из домашних животных, сельскохозяйственных животных, спортивных животных и приматов. Предпочтительно субъект представляет собой человека, которому был поставлен диагноз наличия или подверженности риску наличия пролиферативного заболевания, такого как рак. Термины «субъект» и «пациент», при отнесении к человеческому организму, могут использоваться взаимозаменяемо и охватывают субъектов мужского и женского пола. Субъект, подлежащий лечению, может быть новорожденным, ребенком в возрасте до 2 лет, молодым взрослым или взрослым.
Термин «комбинация» или «ассоциация» в том виде, как он используется в данном документе, относится к любому возможному сочетанию разных компонентов (например, онколитического вируса и одного или более чем одного вещества, эффективного в противораковой терапии). Такое сочетание включает смесь указанных компонентов, а также отдельные комбинации для сопутствующего или последовательного введений. Настоящее изобретение охватывает комбинации, содержащие эквимолярные концентрации каждого компонента, а также комбинации с очень разными концентрациями. Понятно, что оптимальная концентрация каждого компонента данной комбинации может быть определена квалифицированным специалистом в данной области.
Термин «модулятор иммунологической контрольной точки» относится к молекуле, способной позитивно или негативно модулировать функцию белка иммунологической контрольной точки (в частности, взаимодействие между антигенпрезентирующей клеткой (АРС) или раковой клеткой и эффекторной Т-клеткой). Термин «иммунологическая контрольная точка» относится к белку, прямо или опосредованно участвующему в иммунологическом пути, который при нормальных физиологических условиях является решающим для предупреждения неконтролируемых иммунных реакций и, таким образом, для поддержания аутотолерантности и/или защиты ткани. Один или более чем один модулятор иммунологической контрольной точки, используемый в данном документе, может независимо действовать на любой стадии опосредованного Т-клетками иммунитета, включая клональный отбор антигенспецифичных клеток, активацию Т-клеток, пролиферацию, транспорт к сайтам антигена и воспаления, выполнение непосредственной эффекторной функции и сигнализацию через цитокины и мембранные лиганды. Каждая из данных стадий регулируется посредством уравновешивания стимулирующих и ингибирующих сигналов, которые осуществляют точную настройку ответа. В контексте настоящего изобретения данный термин охватывает модулятор(ры) иммунологической контрольной точки, способный(ные), по меньшей мере частично, к понижающей регуляции функции ингибирующей иммунологической контрольной точки (антагонист), и/или модулятор(ры) иммунологической контрольной точки, способный(ные), по меньшей мере частично, к повышающей регуляции функции стимулирующей иммунологической контрольной точки (агонист).
Онколитический вирус
Онколитический вирус по настоящему изобретению может быть получен от любого идентифицированного в настоящее время члена семейства вируса, при условии, что он является онколитическим по его склонности к селективной репликации и умерщвлению делящихся клеток, по сравнению с неделящимися клетками. Он может представлять собой нативный вирус, который является онколитическим в природе, или может быть подвергнут генетической модификации посредством модифицирования одного или более чем одного вирусного гена таким образом, чтобы увеличить селективность в отношении опухоли и/или предпочтительную репликацию в делящихся клетках, как, например, генов, участвующих в репликации ДНК, метаболизме нуклеиновых кислот, тропизме хозяина, прикреплении к поверхности, вирулентности, лизисе и распространении (см., например, Kirn et al., 2001, Nat. Med. 7: 781; Wong et al., 2010, Viruses 2: 78-106). Также можно рассматривать помещение одного или более чем одного вирусного гена под контроль регулирующих элементов (например, промотора), специфичных в отношении события или ткани.
Типичные онколитические вирусы включают, без ограничения, реовирус, вирус долины Сенека (SW), вирус везикулярного стоматита (VSV), вирус ньюкаслской болезни (NDV), вирус простого герпеса (HSV), морбилливирус, ретровирус, вирус гриппа, вирус Синдбис, вирус группы оспы, аденовирус или тому подобные.
В одном воплощении онколитический вирус по настоящему изобретению получают из реовируса. Типичный пример включает реолизин (разрабатывается Oncolytics Biotech; NCT01166542).
В одном воплощении онколитический вирус по настоящему изобретению получают из вируса долины Сенека. Типичный пример включает NTX-010 (Rudin et al., 2011, Clin. Cancer. Res. 17(4): 888-95).
В одном воплощении онколитический вирус по настоящему изобретению получают из вируса везикулярного стоматита (VSV). Типичные примеры описаны в литературе (например, Stojdl et al., 2000, Nat. Med. 6(7): 821-5; Stojdl et al., 2003, Cancer Cell 4(4): 263-75).
В одном воплощении онколитический вирус по настоящему изобретению получают из вируса ньюкаслской болезни. Типичные примеры включают, без ограничения, штаммы 73-Т PV701 и HDV-HUJ, а также штаммы, описанные в литературе (например, Phuangsab et al., 2001, Cancer Lett. 172(1): 27-36; Lorence et al., 2007, Curr. Cancer Drug Targets 7(2): 157-67; Freeman et al., 2006, Mol. Ther. 13(1): 221-8).
В одном воплощении онколитический вирус по настоящему изобретению получают из вируса герпеса. Herpesviridae представляют собой большое семейство ДНК вирусов, которые все имеют общую структуру и состоят из относительно больших геномов на основе двухцепочечной линейной ДНК, кодирующей 100-200 генов, заключенных в икосаэдрическом капсиде, который заключен в мембрану на основе липидного бислоя. Хотя онколитический вирус герпеса и может происходить из разных типов HSV, особенно предпочтительными являются HSV1 и HSV2. Вирус герпеса может быть генетически модифицирован таким образом, чтобы ограничивать вирусную репликацию опухолями или уменьшать его цитотоксичность в неделящихся клетах. Например, может быть инактивирован любой вирусный ген, участвующий в метаболизме нуклеиновой кислоты, такой как тимидинкназа (Martuza et al., 1991, Science 252: 854-6), рибонуклеотидредуктаза (RR) (Boviatsis et al., Gene Ther. 1: 323-31; Mineta et al., 1994, Cancer Res. 54: 3363-66) или урацил-N-гликозилаза (Pyles et al., 1994, J. Virol. 68: 4963-72). Другой аспект включает мутантов вирусов с дефектами в функции генов, кодирующих факторы вирулентности, таких как ген ICP34.5 (Chambers et al., 1995, Proc. Natl. Acad. Sci. USA 92: 1411-5). Типичные примеры онколитического вируса герпеса включают NV1020 (например, Geevarghese et al., 2010, Hum. Gene Ther. 21(9): 1119-28) и T-VEC (Andtbacka et al., 2013, J. Clin. Oncol. 31, номер реферата LBA9008).
В одном воплощении онколитический вирус по настоящему изобретению получают из морбилливируса, который может быть получен из семейства парамиксовирида, с конктретным предпочтением в отношении вируса кори. Типичные примеры онколитических вирусов кори включают, без ограничения, MV-Edm (McDonald et al., 2006; Breast Cancer Treat. 99(2): 177-84) и HMWMAA (Kaufmann et al., 2013, J. Invest. Dermatol. 133(4): 1034-42).
В одном воплощении онколитический вирус по настоящему изобретению получают из аденовируса. В данной области доступны способы для генетической модификации онколитических аденовирусов. Полезная стратегия включает замену вирусных промоторов опухолеселективными промоторами или модификации продукта(тов) гена аденовируса Е1 с инактивацией его/их функции связывания с р53 или белком ретинобластомы (Rb), которые изменяются в опухолевых клетках. В природном контексте ген аденовируса E1B55kDa кооперируется с другим аденовирусным продуктом для инактивации р53 (р53 часто неправильно регулируется в раковых клетках), таким образом, предотвращая апоптоз. Типичные примеры онколитического аденовируса включают ONYX-015 (например, Khuri et al., 2000. Nat. Med 6(8): 879-85) и H101, также именуемый Oncorine (Xia et al., 2004, Ai Zheng 23(12): 1666-70).
В одном воплощении онколитический вирус по настоящему изобретению представляет собой вирус группы оспы. Термин «вирус группы оспы» в том виде, как он используется в данном документе, относится к вирусу, принадлежащему к семейству Poxviridae, с конкретным предпочтением в отношении вируса группы оспы, принадлежащего к подсемейству Chordopoxviridae, и более предпочтительно - в отношении рода Orthopoxvirus. Последовательности генома разных вирусов группы оспы, например, геномов вируса осповакцины, вируса коровьей оспы, вируса оспы птиц, вируса оспы мышей, вируса миксомы, доступны в данной области и в специализированных базах данных, таких как Genbank (номера доступа NC-006998, NC_003663, NC_005309, NC_004105, NC_001132 соответственно).
Желательно онколитический вирус группы оспы представляет собой онколитический вирус осповакцины. Вирусы осповакцины являются членами семейства вируса группы оспы, характеризуемыми геномом в 200 т.п.н. на основе двухцепочечной ДНК, который кодирует многочисленные вирусные ферменты и факторы, которые обеспечивают независимую репликацию вируса от аппарата клетки-хозяина. Большинство частиц вируса осповакцины являются внутриклеточными (IMV обозначает внутриклеточный зрелый вирион) с одиночной липидной оболочкой, и они остаются в цитозоле инфицированных клеток до лизиса. Другой инфекционной формой является частица с двойной оболочкой (EEV обозначает внеклеточный оболочечный вирион), которая отпочковывается от инфицированной клетки без ее лизирования.
Несмотря на то, что он может происходить от любого штамма вируса осповакцины, штаммы Elstree, Wyeth, Copenhagen и Western Reserve являются особенно предпочтительными. Использованная в данном документе номенклатура генов представляет собой номенклатуру штамма вируса осповакцины Copenhagen. Она также используется в данном документе для гомологичных генов других Poxviridae, если не указано иное. Однако номенклатура генов может быть другой, согласно штамму вируса группы оспы, но соответствие между штаммом Copenhagen и другими штаммами вируса осповакцины обычно доступно в литературе.
Предпочтительно онколитический вирус осповакцины по настоящему изобретению модифицирован посредством изменения одного или более чем одного вирусного гена. Указанная(ные) модификация(ции) предпочтительно приводит(дят) к синтезу (или отсутствию синтеза) дефектного белка, не способного обеспечивать активность белка, продуцированного при нормальных условиях немодифицированным геном. Модификации охватывают делецию, мутацию и/или замену одного или более чем одного нуклеотида (смежного или нет) в пределах вирусного гена или его регуляторных элементов. Модификацию(ции) можно делать целым рядом способов, известных специалистам в данной области, с использованием традиционных методик генной инженерии. Типичные модификации раскрываются в литературе с конкретным предпочтением в отношении модификаций, изменяющих вирусные гены, участвующие в метаболизме ДНК, вирулентности хозяина и пути IFN (интерферон) (см., например, Guse et al., 2011, Expert Opinion Biol. Ther. 11(5):595-608).
Более предпочтительно онколитический вирус группы оспы по настоящему изобретению модифицирован посредством изменения гена, кодирующего тимидинкиназу (локус J2R). Фермент ТК участвует в синтезе дезоксирибонуклеотидов. ТК необходима для вирусной репликации в нормальных клетках, так как данные клетки обычно имеют низкую концентрацию нуклеотидов, тогда как она является необязательной в делящихся клетках, которые содержат высокую концентрацию нуклеотидов.
Альтернативно или в комбинации онколитический вирус группы оспы по настоящему изобретению модифицируется посредством изменения по меньшей мере одного гена или обоих генов, кодирующих рибонуклеотидредуктазу (RR). В природном контексте данный фермент катализирует восстановление рибонуклеотидов до дезоксирибонуклеотидов, что представляет собой решающую стадию в биосинтезе ДНК. Вирусный фермент является аналогичным по субъединичной структуре ферменту млекопитающих, состоящему из двух гетерологичных субъединиц, обозначенных R1 и R2, кодируемых локусом I4L и F4L соответственно. Последовательности генов I4L и F4L и их положения в геноме разных вирусов группы оспы доступны в общедоступных базах данных, например, через номера доступа DQ437594, DQ437593, DQ377804, АН015635, AY313847, AY313848, NC_003391, NC_003389, NC_003310, М-35027, AY243312, DQ011157, DQ011156, DQ011155, DQ011154, DQ011153, Y16780, Х71982, AF438165, U60315, AF410153, AF380138, U86916, L22579, NC_006998, DQ121394 и NC_008291. В контексте данного изобретения могут быть инактивированы либо ген I4L (кодирующий большую субъединицу R1), либо ген F4L (кодирующий малую субъединицу R2), либо оба.
Альтернативно или в комбинации, для дальнейшего увеличения опухолеспецифичности вируса также можно следовать другим стратегиям. Типичный пример подходящей модификации включает разрушение гена, кодирующего VGF, из вирусного генома. VGF (обозначает фактор роста VV) представляет собой секретируемый белок, который экспрессируется на раннем этапе после инфекции клетки, и его функция, по-видимому, является важной для распространения вируса в нормальных клетках. Другим примером является разрушение гена A56R, кодирующего гемагглютинин, в конечном счете, в комбинации с делецией ТК (Zhang et al., 2007, Cancer Res. 67: 10038-46). Также полезным может быть разрушение гена(нов), модулирующего(щих) интерферон (например, гена B8R или B18R), или гена ингибитора каспазы-1 B13R. Другая подходящая модификация включает разрушение гена F2L, который кодирует вирусную дУТФазу, участвующую и в поддержании точности репликации ДНК, и в предоставлении предшественника для подуцирования ТМФ (тимидинмонофосфат) тимидилатсинтазой (Broyles at al., 1993, Virol. 195: 863-5). Последовательность гена F2L вируса осповакцины доступна в Genbank посредством номера доступа М25392.
В предпочтительном воплощении онколитический вирус по данному изобретению представляет собой вирус осповакцины, дефектный по ТК, в результате инактивирующих мутаций в гене J2R. В другом предпочтительном воплощении онколитический вирус по данному изобретению представляет собой вирус осповакцины, дефектный по активностям и ТК, и RR, в результате инактивирующих мутаций и в гене J2R, и в гене(нах) I4L и/или F4L, которые несет вирусный геном (например, как описано в WO 2009/065546 и Foloppe et al., 2008, Gene Ther., 15: 1361-71). В другом предпочтительном воплощении онколитический вирус по данному изобретению представляет собой вирус осповакцины, дефектный по дУТФазе в результате инактивирующих мутаций в гене F2L (например, как описано в WO 2009/065547), в конечном счете, в комбинации с нарушением по меньшей мере одной активности ТК и RR или обоих (приводя к вирусу с инактивирующими мутациями в F2L; гене F2L и J2R; F2L и I4L; или в F2L, J2R и I4L).
Терапевтические гены
В одном воплощении онколитический вирус по данному изобретению дополнительно экспрессирует по меньшей мере один терапевтический ген, вставленный в вирусный геном. «Терапевтический ген» кодирует продукт, способный обеспечивать биологическую активность, при введении субъекту подходящим образом, что, как ожидается, вызовет полезное влияние на ход или симптом патологического состояния, подлежащего лечению, либо посредством потенцирования противоопухолевой эффективности, либо усиления онколитической природы вируса. В контексте данного изобретения терапевтический ген может иметь происхождение от млекопитающего (например, человека, мыши, кролика и т.д.) или не иметь его (например, бактериальное, дрожжевое или вирусное происхождение).
В контексте данного изобретения может рассматриваться широкий спектр терапевтических генов, таких как гены, кодирующие полипептиды, которые могут компенсировать дефектные или дефицитные белки у субъекта, или гены, которые действуют через токсические эффекты, лимитируя или удаляя вредоносные клетки из организма, или гены, которые кодируют полипептиды, дающие иммунитет. Они могут представлять собой нативные гены или гены, полученные из последних посредством мутации, делеции, замены и/или добавления одного или более чем одного нуклеотида.
Преимущественно онколитический вирус по настоящему изобретению несет терапевтический ген, выбранный из группы, состоящей из генов, кодирующих суицидные генные продукты и иммуностимулирующие белки.
Суицидный ген
Термин «суицидный ген» относится к гену, кодирующему белок, способный к превращению предшественника лекарственного средства в цитотоксическое соединение. Суицидные гены включают гены, кодирующие белок, имеющий цитозиндезаминазную активность, тимидинкиназную активность, урацилфосфорибозилтрансферазную активность, активность фосфорилазы пуриновых нуклеозидов и тимидилаткиназную активность, но не ограничиваются ими. Примеры суицидных генов и соответствующих предшественников лекарственного средства, содержащих одну группировку нуклеинового основания, раскрыты в следующей таблице.
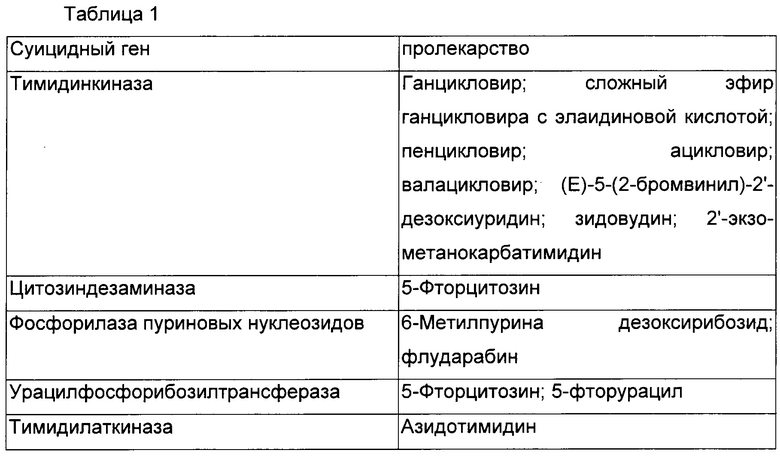
Желательно суицидный ген кодирует белок, имеющий по меньшей мере активность ЦДазы (цитозиндезаминазы). У прокариотов и низших эукариотов (она отсутствует у млекопитающих) ЦДаза участвует в метаболическом пути пиримидина, посредством чего экзогенный цитозин превращается в урацил посредством гидролитического дезаминирования. ЦДаза также дезаминирует аналог цитозина, т.е. 5-фторцитозин (5-FC), образуя, посредством этого, 5-фторурацил (5-FU), соединение, которое является высокоцитотоксическим при его превращении в 5-фтор-UMP (5-фтор-уридинмонофосфат, 5-FUMP). Молекула нуклеиновой кислоты, кодирующая ЦДазу, может быть получена из любых прокариотов и низших эукариотов, таких как Saccharomyces cerevisiae (ген FCY1), Candida albicans (ген FCA1) и Escherichia coli (ген codA). Последовательности генов и кодируемых белков ЦДаз были опубликованы и доступны в специализированных банках данных (SWISSPROT EMBL, Genbank, Medline и тому подобные). Также можно использовать функциональные аналоги данных генов. Такие аналоги предпочтительно имеют последовательность нуклеиновой кислоты, имеющую уровень идентичности по меньшей мере 70%, преимущественно по меньшей мере 80%, предпочтительно по меньшей мере 90% и наиболее предпочтительно по меньшей мере 95% с последовательностью нуклеиновой кислоты нативного гена.
Альтернативно или в комбинации, онколитический вирус по изобретению несет в его вирусном геноме суицидный ген, кодирующий полипептид, имеющий урацилфосфорибозилтрансферазную (УФРТазную) активность. У прокариотов и низших эукариотов урацил превращается в UMP посредством действия УФРТазы. Данный фермент превращает 5-FU в 5-FUMP. В качестве иллюстрации в контексте данного изобретения можно использовать последовательности нуклеиновой кислоты, кодирующие УФРТазы из Е. coli (Andersen et al., 1992, European J. Biochem. 204: 51-56), из Lactococcus lactis (Martinussen et al., 1994, J. Bacteriol. 176: 6457-63), из Mycobacterium bovis (Kim et al., 1997, Biochem. Mol. Biol. Internat. 41: 1117-24) и из Bacillus subtilis (Martinussen et al., 1995, J. Bacteriol. 177: 271-4). Однако предпочтительнее всего использовать дрожжевую УФРТазу и, в частности, УФРТазу, кодируемую S. cerevisiae (ген FUR1), последовательности которой раскрыты в Kern et al. (1990, Gene 88: 149-57). Также можно использовать функциональные аналоги УФРТазы, такие как мутант FUR1, усеченный на N-конце, описанный в ЕР 998568 (с делецией 35 первых остатков вплоть до второго остатка Met, присутствующего в положении 36 в нативном белке), который демонстрирует более высокую УФРТазную активность, чем активность нативного фермента.
Предпочтительно суицидный ген, вставленный в вирусный геном онколитического вируса по настоящему изобретению, кодирует полипептид, имеющий ЦДазную и УФРТазную активности. Такой полипептид может быть сконструирован посредством слияния двух ферментативных доменов - одного, имеющего ЦДазную активность, и второго, имеющего УФРТазную активность. Типичные полипептиды включают, без ограничения, слитые полипептиды codA::upp, FCY1::FUR1 и FCYI::FUR1[Delta] 105 (FCU1) и FCU1-8, описанные в WO 96/16183, ЕР 998568 и WO 2005/07857. Особый интерес представляет суицидный ген FCU1 (или слияние FCY1::FUR1[Delta] 105), кодирующий полипептид, содержащий аминокислотную последовательность, представленную в идентификаторе последовательности SEQ ID NO: 1 WO 2009/065546. Настоящее изобретение охватывает аналоги таких полипептидов, при условии, что они сохраняют ЦДазную и/или УФРТазную активности. Для специалиста доступно выделение молекул нуклеиновой кислоты, кодирующих ЦДазу и/или УФРТазу, из опубликованных данных, в конечном счете, конструирование их аналогов и тестирование ферментативной активности в бесклеточной или клеточной системе согласно традиционным методикам (см., например, ЕР 998568).
Иммуностимулирующие терапевтические гены
Термин «иммуностимулирующий белок» в том виде, как он используется в данном документе, относится к белку, который имеет способность стимулировать иммунную систему, специфичным или неспецифичным способом. В данной области известно огромное число белков за их способность оказывать иммуностимулирующий эффект. Примеры подходящих иммуностимулирующих белков в контексте данного изобретения включают, без ограничения, цитокины, с особым предпочтением в отношении интерлейкинов (например, IL-2, IL-6, IL-12, IL-15, IL-24), хемокинов (например, CXCL10, CXCL9, CXCL11), интерферонов (например, IFN-гамма, IFN-альфа), фактора некроза опухолей (TNF), колониестимулирующих факторов (например, GM-CSF, C-CSF, M-CSF…), белков, экспонируемых АРС (антигенпрезентирующая клетка) (например, В7.1, В7.2 и тому подобные), факторов роста (трансформирующий фактор роста - TGF, фактор роста фибробластов - FGF, фактор роста эндотелия сосудов - VEGF и тому подобные), антигенов МНС (главный комплекс гистосовместимости) класса I или II, индукторов или ингибиторов апоптоза (например, Вах, Bcl2, BcIX…), цитостатических агентов (р21, р16, Rb…), иммунотоксинов, антигенных полипептидов (антигенные полипептиды, эпитопы и тому подобное) и маркеров (бета-галактозидаза, люцифераза…). Предпочтительно иммуностимулирующий белок представляет собой интерлейкин или колониестимулирующий фактор, с особым предпочтением в отношении GM-CSF.
Модулятор(ры) иммунологических контрольных точек
Иммунологические контрольные точки и их модуляторы, а также способы применения таких соединений описаны в литературе. Согласно данному изобретению один или более чем один модулятор иммунологической контрольной точки может представлять собой, независимо, полипептид, содержащий домен, способный к связыванию с целевой иммунологической контрольной точкой и/или ингибирующий связывание лиганда с указанной целевой иммунологической контрольной точкой таким образом, чтобы осуществлять антагонистическую функцию (т.е. имеющий способность оказывать антагонистический эффект на ингибирующий сигнал, опосредованный иммунологической контрольной точкой) или агонистическую функцию (т.е. имеющий способность наращивать стимулирующий сигнал, опосредованный иммунологической контрольной точкой). Такой один или более чем один модулятор иммунологической контрольной точки может быть независимо выбран из группы, состоящей из пептидов (например, пептидные лиганды), растворимых доменов природных рецепторов, РНКи (интерферирующая РНК), антисмысловых молекул, антител и белковых каркасов.
В предпочтительном воплощении модулятор иммунологической контрольной точки представляет собой антитело. В контексте изобретения термин «антитело» («Ab») используется в самом широком смысле и охватывает встречающиеся в природе и сконструированные человеком, а также полноразмерные антитела или их функциональные фрагменты или аналоги, которые способны к связыванию с целевой иммунологической контрольной точкой или эпитопом (таким образом, сохраняющие часть, связывающуюся с мишенью). Антитело, кодируемое онколитическим вирусом по изобретению, может иметь любое происхождение, например, может быть человеческим, гуманизированным, животным (например, антитело грызуна или верблюдового) или химерным. Оно может иметь любой изотип (например, изотип IgG1, IgG2, IgG3, IgG4, IgM и т.д.). Кроме того, оно может быть гликозилированным, частично гликозилированным или негликозилированным (например, посредством мутирования одного или более чем одного остатка в пределах сайта(тов) гликозилирования). Термин антитело также включает биспецифичные или мультиспецифичные антитела, при условии, что они демонстрируют специфичность связывания, описанную в данном документе.
Для иллюстративных целей полноразмерные антитела представляют собой гликопротеины, содержащие по меньшей мере две тяжелые (Н) цепи и две легкие (L) цепи, взаимосвязанные дисульфидными связями. Каждая тяжелая цепь состоит из вариабельной области тяжелой цепи (VH) и константной области тяжелой цепи, которая составлена из трех доменов СН1, СН2 и СН3 (в конечном счете, с шарнирной областью между СН1 и СН2). Каждая легкая цепь состоит из вариабельной области легкой цепи (VL) и константной области легкой цепи, которая содержит один домен CL. Области VH и VL содержат гипервариабельные области, именуемые областями, определяющими комплементарность (CDR), с вкраплением более консервативных областей, названных каркасными областями (FR). Каждая VH и VL состоит из трех CDR и четырех FR в следующем порядке: FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4. Области CDR тяжелой и легкой цепей являются детерминантами для специфичности связывания.
Термин «гуманизированное антитело» в том виде, как он используется в данном документе, относится к антителу, не являющемуся человеческим (например, мышиному, верблюжьему, крысиному и т.д.), последовательность белка которого была модифицирована для увеличения его сходства с человеческим антителом (т.е. продуцированным в природе у человека). Способ гуманизации хорошо известен в данной области (см., например, Presta et al., 1997, Cancer Res. 57(20): 4593-9; US 5225539; US 5530101; US 6180370; WO 2012/110360). Например, антитело против иммунологической контрольной точки для применения в данном изобретении, может быть гуманизировано посредством замены одного или более чем одного остатка областей FR для того, чтобы они выглядели подобно последовательности человеческого иммуноглобулина, тогда как подавляющее большинство остатков вариабельных областей (особенно CDR) не модифицируется и соответствует остаткам иммуноглобулина, не являющегося человеческим. Относительно общего руководства, число данных замен аминокислот в областях FR типично составляет не более 20 в каждой вариабельной области VH или VL.
Термин «химерное антитело» в том виде, как он используется в данном документе, относится к антителу, содержащему один или более чем один элемент одного вида и один или более чем один элемент другого вида, например, антитело, не являющееся человеческим, содержащее по меньшей мере часть константной области (Fc) человеческого иммуноглобулина.
Онколитическим вирусом по изобетению могут экспрессироваться многие формы антитела. Показательные примеры включают, без ограничения, Fab, Fab', F(ab')2, dAb, Fd, Fv, scFv, ди-scFv, диатело и т.д. Более конкретно:
(i) фрагмент Fab, представленный одновалентным фрагментом, состоящим из доменов VL, VH, CL и СН1;
(и) фрагмент F(ab')2, представленный двухвалентным фрагментом, содержащим два фрагмента Fab, связанных по меньшей мере одним дисульфидным мостиком в шарнирной области;
(iii) фрагмент Fd, состоящий из доменов VH и СН1;
(iv) фрагмент Fv, состоящий из доменов VL и VH одного плеча антитела;
(v) фрагмент dAb, состоящий из фрагмента в виде единичного вариабельного домена (домена VH или VL);
(vi) одноцепочечный Fv (scFv), состоящий из двух доменов фрагмента Fv - VL и VH, которые слиты друг с другом, в конечном счете, с использованием линкера с получением одной белковой цепи (см., например, Bird et al., 1988, Science 242: 423-6; Huston et al., 1988, Proc. Natl. Acad. Sci. USA 85: 5879-83; US 4946778; US 5258498); и
(vii) любое другое искусственное антитело.
При необходимости такие фрагменты и аналоги могут быть подвергнуты скринингу на функциональность тем же самым способом, что и интактные антитела (например, посредством стандартного анализа ELISA (твердофазный иммуноферментный анализ).
В предпочтительном воплощении по меньшей мере один из одного или более чем одного модулятора иммунологической контрольной точки, кодируемого онколитическим вирусом по настоящему изобретению, представляет собой моноклональное антитело, с конкретным предпочтением в отношении человеческого (в котором обе каркасные области происходят из последовательностей иммуноглобулина человеческой зародышевой линии) или гуманизированного антитела согласно хорошо известному способу гуманизации.
Желательно один или более чем один модулятор иммунологической контрольной точки, кодируемый онколитическим вирусом по настоящему изобретению, оказывает по меньшей мере частичный антагонистический эффект (например, более чем на 50%) на активность ингибирующей(щих) иммунологической(ких) контрольной(ных) точки(чек), в частности, иммунологических контрольных точек, опосредованных любым из следующих: PD-1, PD-L1, PD-L2, LAG3, Tim3, BTLA и CTLA4, с конкретным предпочтением в отношении моноклонального антитела, которое специфично связывается с любым из таких белков-мишеней. Термин «специфично связывается с» относится к способности к специфичности связывания и аффинности в отношении конкретной мишени или эпитопа, даже в присутствии гетерогенной популяции других белков и биологических агентов. Таким образом, при обозначенных условиях анализа антитело, используемое в данном изобретении, предпочтительно связывается с его мишенью и не связывается в значимом количестве с другими компонентами, присутствующими в анализируемом образце или субъекте. Предпочтительно такое антитело демонстрирует высокоаффинное связывание с его мишенью с равновесной константой диссоциации, равной или меньшей 1×10-6 (например, по меньшей мере 0,5×10-6, 1×10-7, 1×10-8, 1×10-9, 1×10-10 и т.д.). В качестве альтернативы, один или более чем один кодируемый модулятор иммунологической контрольной точки имеет агонистическую функцию в том смысле, что он способен стимулировать или усиливать стимулирующие сигналы, в частности, сигналы, опосредованные CD28, с особым предпочтением в отношении любой из иммунологических контрольных точек ICOS, CD137 (4-1ВВ), ОХ40, CD27, CD40 и GITR. Стандартные анализы для оценки способности антител к связыванию в отношении иммунологических контрольных точек известны в данной области, включая, например, ELISA, вестерн-блоттинг, RIA (радиоиммуноанализ) и проточную цитометрию. Кинетику связывания (например, аффинность связывания) антител также можно оценивать стандартными анализами, известными в данной области, такими как анализ Biacore.
В предпочтительном воплощении по меньшей мере один или более чем один кодируемый модулятор контрольной точки представляет собой человеческое или гуманизированное антитело, способное оказывать антагонистический эффект на иммунологическую контрольную точку, участвующую в ответе, опосредованном Т-клетками. Предпочтительный пример модулятора иммунологической контрольной точки представлен модулятором, способным оказывать антагонистический эффект, по меньшей мере частично, на белок программируемой смерти 1 (PD-1), и особенно антителом, которое специфично связывается с человеческим PD-1. PD-1 является частью надсемейства генов иммуноглобулинов (Ig) и членом семейства CD28. Он представляет собой 55 кДа трансмембранный белок типа 1, экспрессируемый на «обученных» антигеном клетках (например, на активированных В-клетках, Т-клетках и миелоидных клетках) (Agata et al., 1996, Int. Immunol. 8: 765-72; Okazaki et al., 2002, Curr. Opin. Immunol. 14: 391779-82; Bennett et al., 2003, J. Immunol 170: 711-8). В нормальном контексте он действует посредством ограничения активности Т-клеток во время воспалительного ответа, защищая, посредством этого, нормальные ткани от разрушения (Topalian, 2012, Curr. Opin. Immunol. 24: 207-12). Для PD-1 были идентифицированы два лиганда, соответственно PD-L1 (лиганд программируемой смерти 1) и PD-L2 (лиганд программируемой смерти 2) (Freeman et al., 2000, J. Exp. Med. 192: 1027-34; Carter et al., 2002, Eur. J. Immunol. 32: 634-43). PD-L1 был идентифицирован при 20-50% человеческих раковых заболеваний (Dong et al., 2002, Nat. Med. 8: 787-9). Взаимодействие между PD-1 и PD-L1 приводило к уменьшению числа инфильтрующих опухоль лимфоцитов, уменьшению пролиферации, опосредованной рецептором Т-клеток, и ускользанию раковых клеток от иммунологического надзора (Dong et al., 2003, J. Mol. Med. 81: 281-7; Blank et al., 2005, Cancer Immunol. Immunother. 54: 307-314). Полные нуклеотидную и аминокислотную последовательности PD-1 можно найти под №доступа GenBank U64863 и NP_005009.2. В данной области доступен целый ряд антител против PD1 (см., например, антитела, описанные в WO 2004/004771; WO 2004/056875; WO 2006/121168; WO 2008/156712; WO 2009/014708; WO 2009/114335; WO 2013/043569 и WO 2014/047350). Предпочтительно онколитический вирус по настоящему изобретению кодирует и экспрессирует антитело против PD-1, которое одобрено FDA (Управление США по контролю качества пищевых продуктов и лекарственных средств) или находится на продвинутой стадии клинической разработки, как, например, антитела, распространяемые на рынке или разрабатываемые под названиями ниволумаб (также называется BMS-936558, разрабатывается Bristol Myer Squibb), пембролизумаб (также называется МК-3475; разрабатывается Merck) и пидилизумаб (также называется СТ-011; разрабатывается CureTech). Соответствующие нуклеотидные последовательности могут быть клонированы или выделены согласно стандартным методикам на основе информации, раскрытой в доступной литературе.
Другой предпочтительный пример модулятора иммунологической контрольной точки, подходящего для экспрессии онколитическим вирусом по изобретению, представлен модулятором, способным оказывать антагонистический эффект, по меньшей мере частично, на лиганд PD-1, именуемый PD-L1, и особенно антителом, которое распознает человеческий PD-L1. В данной области доступен целый ряд антител против PD-L1 (см., например, антитела, описанные в ЕР 1907000). Предпочтительные антитела против PD-L1 одобрены FDA или находятся на продвинутой стадии клинической разработки (например, MPDL3280A, разрабатывается Genentech/Roche, и BMS-936559, разрабатывается Bristol Myer Squibb, а также слияния с Fc против PD-L1 (например, АМР-224, разработанное Medimmune и AstraZeneca).
Также можно использовать антагонист недавно идентифицированного белка VISTA, который, как было показано, негативно регулирует ответы Т-клеток (Wang et al., 2011, J. Exp. Med. 208(3): 577-592). VISTA, также обозначенный PD-1H и PD-L3, имеет сходство с членами семейства PD-L1. Например, антагонисты против VISTA описываются в US 2013-0177557.
Еще один другой предпочтительный пример подходящего модулятора иммунологической контрольной точки представлен модулятором, способным оказывать антагонистический эффект, по меньшей мере частично, на белок CTLA-4, и особенно антителом, которое распознает человеческий CTLA-4. CTLA4 (антиген 4, ассоциированный с цитотоксическими Т-лимфоцитами), также известный как CD152, был идентифицирован в 1987 году (Brunet et al., 1987, Nature 328: 267-70) и кодируется геном CTLA4 (Dariavach et al., Eur. J. Immunol. 18: 1901-5). CTLA4 является членом надсемейства рецепторов иммуноглобулинов. Он экспрессируется на поверхности Т-клеток-хелперов, где он, в первую очередь, регулирует амплитуду ранних этапов активации Т-клеток. Недавняя работа свидетельствовала о том, что CTLA-4 может функционировать in vivo посредством захвата и удаления В7-1 и В7-2 из мембран антигенпрезентирующих клеток, делая их, таким образом, недоступными для запуска CD28 (Qureshi et al., Science, 2011, 332: 600-3). Полная последовательность нуклеиновой кислоты CTLA-4 может быть найдена под № доступа GenBank LI 5006. В данной области доступен целый ряд антител против CTLA-4 (см., например, антитела, описанные в US 8491895). Предпочтительные антитела против CTLA-4 в контексте данного изобретения одобрены FDA или находятся на продвинутой стадии клинической разработки. Более конкретно можно упомянуть ипилимумаб, распространяемый на рынке Bristol Myer Squibb под названием ервой (см., например, US 6984720; US 8017114), тремелимумаб, разрабатываемый Pfizer (см., например, US 7109003 и US 8143379), и одноцепочечные антитела против CTLA4 (см., например, WO 97/20574 и WO 2007/123737).
Онколитический вирус по настоящему изобретению также может экспрессировать модулятор иммунологической контрольной точки для оказания антагонистического эффекта на рецептор LAG3 (см., например, US 5773578).
Другой пример подходящего модулятора иммунологической контрольной точки представлен агонистом ОХ40, таким как агонист лиганда ОХ40 (OX40L) (см., например, US 5457035, US 7622444; WO 03/082919), или антителом, направленным на рецептор ОХ40 (см., например, US 7291331 и WO 03/106498).
Другие примеры модуляторов иммунологических контрольных точек представлены антителом против KIR или против CD96, нацеленным на ингибирующие рецепторы, которые имеют Т-клетки CD8+ или клетки NK (природный киллер).
Настоящее изобретение охватывает онколитический вирус, кодирующий более чем один модулятор иммунологической контрольной точки. Предпочтительный пример включает, без ограничения, экспрессию антитела против CTLA-4 и антитела против PD-1.
Экспрессия одной или более чем одной молекулы нуклеиновой кислоты, кодирующей модулятор(ры) иммунологической контрольной точки, если любой из терапевтических генов вставлен в вирусный геном
Молекула(лы) нуклеиновой кислоты, кодирующая(щие) модулятор иммунологической контрольной точки, и терапевтический ген могут быть легко получены стандартными методиками молекулярной биологии (например, ПЦР-амплификация, клонирование кДНК, химический синтез), используя данные о последовательностях, доступные в данной области, и предложенную в данном документе информацию. Способы клонирования антител, их фрагментов и аналогов известны в данной области (см., например, Harlow and Lane, 1988, Antibodies - A laboratory manual; Cold Spring Harbor Laboratory, Cold Spring Harbor NY). Например, молекула нуклеиновой кислоты (например, кДНК), кодирующая легкую и тяжелую цепи антитела или их CDR, может быть выделена из продуцирующей гибридомы (см., например, Kohler and Milstein, 1975, Nature 256: 495-7; Cote et al., 1983, Proc. Natl. Acad. Sci. USA 80: 2026-30; Cole et al. в Monoclonal antibodies and Cancer Therapy; Alan Liss pp77-20 96), библиотек генов иммуноглобулинов или из любого доступного источника, или нуклеотидная последовательность может быть получена химическим синтезом. Аналоги и фрагменты можно получать с использованием стандартных методик молекулярной биологии.
Молекулу(лы) нуклеиновой кислоты, кодирующую(щие) модулятор(ры) иммунологической контрольной точки, и, в конечном счете, терапевтический(кие) ген(ны) можно независимо вставлять в любом положении вирусного генома с конкретным предпочтением в отношении несущественного локуса. Вставку в онколитический вирус можно проводить посредством стандартной молекулярной биологии, например, как описано в Sambrook et al. (2001, Molecular Cloning-A Laboratory Manual, Cold Spring Harbor Laboratory). Вставку в аденовирусный вектор или вектор на основе вируса группы оспы можно проводить посредством гомологичной рекомбинации, как описано у Chartier et al. (1996, J. Virol. 70: 4805-10) и Paul et al. (2002, Cancer gene Ther. 9: 470-7) соответственно. Например, гены ТК, RR и F2L, а также межгенные области особенно подходят для вставки в онколитический вирус осповакцины, а области Е3 и Е4 - для вставки в онколитический аденовирус.
Кроме того, кодирующие нуклеотидные последовательности могут быть оптимизированы для обеспечения высокого уровня экспрессии в конкретной клетке-хозяине или субъекте. В самом деле, наблюдали то, что картины использования кодонов организмов являются в значительной степени не случайными, и использование кодонов может быть заметно отличным между разными хозяевами. Например, терапевтический ген может происходить от бактерии или низшего эукариота (например, суицидный ген), и, таким образом, он имеет неподходящую картину использования кодонов для эффективной экспрессии в высших эукариотических клетках (например, человеческих). Типично оптимизация кодонов осуществляется посредством замены одного или более чем одного «нативного» (например, бактериального или дрожжевого) кодона, соответствующего кодону, не часто используемому в интересующем организме-хозяине, одним или более чем одним кодоном, кодирующим такую же аминокислоту, который используется чаще. Нет необходимости заменять все нативные кодоны, соответствующие не часто используемым кодонам, поскольку повышенная экспрессия может достигаться даже с частичной заменой.
Кроме оптимизации использования кодонов, экспрессия в клетке-хозяине или в субъекте может дополнительно улучшаться посредством дополнительных модификаций нуклеотидной(ных) последовательности(тей). Например, могут рассматриваться разные модификации, таким образом, чтобы предотвращать кластеризацию редких, неоптимальных кодонов, присутствующих в концентрированных областях, и/или подавить, или модифицировать «негативные» элементы последвательности, которые, как ожидается, отрицательно влияют на уровни экспрессии. Такие негативные элементы последовательности включают, без ограничения, области, имеющие очень высокое (больше 80%) или очень низкое (меньше 30%) содержание GC; обогащенные AT или обогащенные GC отрезки; нестабильные прямые или инвертированные последовательности повторов; вторичные структуры RA; и/или внутренние скрытые регуляторные элементы, такие как внутренние ТАТА-боксы, chi-сайты, сайты посадки рибосомы и/или донорные/акцепторные сайты сплайсинга.
Согласно настоящему изобретению каждая из одной или более чем одной молекулы нуклеиновой кислоты, кодирующая указанный(ные) модулятор(ры) иммунологической контрольной точки, а также терапевтический(кие) ген(ны), вставленный(ные) в геном онколитического вируса по изобретению, связан(ны) функциональным образом с подходящими регуляторными элементами для его(их) экспрессии в клетке-хозяине или субъекте. Термин «регуляторные элементы» или «регуляторная последовательность» в том виде, как он используется в данном документе, относится к любому элементу, который обеспечивает, способствует или модулирует экспрессию кодирующей(щих) молекулы(кул) нуклеиновой кислоты в данной клетке-хозяине или субъекте, включая репликацию, дупликацию, транскрипцию, сплайсинг, трансляцию, стабильность и/или транспорт нуклеиновой(вых) кислоты(лот) или ее производного (т.е. мРНК). Фраза «связанный функциональным образом» в том виде, как она используется в данном документе, означает то, что связанные элементы организованы таким образом, что они согласованно функционируют в их намеченных целях. Например, промотор связан с молекулой нуклеиновой кислоты функциональным образом, если промотор осуществляет транскрипцию от сайта инициации транскрипции до терминатора указанной молекулы нуклеиновой кислоты в пермиссивной клетке-хозяине.
Специалистам в данной области будет понятно, что выбор регуляторных последовательностей может зависеть от таких факторов, как сама молекула нуклеиновой кислоты, вирус, в который она вставляется, клетка-хозяин или субъект, желательный уровень экспрессии и т.д. Промотор является особенно важным. В контексте данного изобретения он может быть конститутивным, управляющим экспрессией молекулы нуклеиновой кислоты во многих типах клеток-хозяев, или специфичным для определенных клеток-хозяев (например, регуляторные последовательности, специфичные для печени), или регулируемым в ответ на специфические события или экзогенные факторы (например, посредством температуры, питательной добавки, гормона и т.д.), или согласно фазе вирусного цикла (например, поздней или ранней). Также можно использовать промоторы, которые репрессируются во время стадии продуцирования в ответ на специфические события или экзогенные факторы для того, чтобы оптимизировать продуцирование вируса и обойти потенциальную токсичность экспрессируемого(мых) полипептида(дов).
Промоторы, подходящие для конститутивной экспрессии в клетках млекопитающих, включают немедленный ранний промотор цитомегаловируса (CMV) (US 5168062), промотор RSV, главный поздний промотор аденовируса, промотор фосфоглицерокиназы (PGK) (Adra et al., 1987, Gene 60: 65-74), промотор тимидинкиназы (ТК) вируса простого герпеса (HSV)-1 и промотор полимеразы Т7 (WO 98/10088), но не ограничиваются ими. Промоторы вируса осповакцины являются особенно адаптированными для экспрессии в онколитических вирусах группы оспы. Типичные примеры включают, без ограничения, промоторы вируса осповакцины 7.5К, H5R, 11К7.5 (Erbs et al., 2008, Cancer Gene Ther. 15(1): 18-28), TK, p28, p11, pB2R, pA35R и K1L, а также синтетические промоторы, такие как промоторы, описанные у Chakrabarti et al. (1997, Biotechniques 23: 1094-7; Hammond et al, 1997, J. Virol Methods 66: 135-8; и Kumar and Boyle, 1990, Virology 179: 151-8), a также ранние/поздние химерные промоторы. Подходящие для онколитического вируса кори промоторы включают, без ограничения, любой промотор, управляющий экспрессией транскрипционных единиц кори (Brandler and Tangy, 2008, CIMID 31: 271).
В частности, когда кодируемый(мые) модулятор(ры) иммунологической контрольной точки включает(ют) антитело и особенно mAb, с особым предпочтением в отношении антитела против PD1, экспрессия тяжелой цепи или ее фрагмента помещается под контроль промотора, который сильнее, чем промотор, используемый для экспрессии легкой цепи или ее фрагмента. В частности промотор для применения для экспрессирования тяжелого компонента дает по меньшей мере на 10%, по меньшей мере на 15%, по меньшей мере на 20%, по меньшей мере на 25% или по меньшей мере на 30% больше продукта по сравнению с экспрессией указанного легкого компонента. Подходящие промоторы для экспрессии могут быть протестированы in vitro (например, в подходящей культивируемой линии клеток) или in vivo (например, в подходящей животной модели или в субъекте). Примеры подходящих промоторов для экспрессирования тяжелого компонента указанного модулятора иммунологической контрольной точки включают промоторы CMV (вирус мозаики цветной капусты), RSV и pH5R и р11К7.5 вируса осповакцины. Примеры подходящих промоторов для экспрессирования легкого компонента указанного модулятора иммунологической контрольной точки включают промоторы PGK, бета-актина и р7.5К и pA35R вируса осповакцины.
Специалистам в данной области будет понятно, что регуляторные элементы, контролирующие экспрессию молекулы(кул) нуклеиновой кислоты, вставленной(ных) в вирусный геном, могут, кроме того, содержать дополнительные элементы для правильной инициации, регуляции и/или терминации транскрипции (например, полиА последовательности терминации транскрипции), транспорта мРНК (например, сигнальные последовательности ядерной локализации), процессинга (например, сигналы сплайсинга) и стабильности (например, интроны и некодирующие последовательности 5' и 3'), трансляции (например, инициирующий Met, трехкомпонентные лидерные последовательности, сайты связывания рибосомы IRES, сигнальные пептиды и т.д.).
Когда это целесообразно, может быть полезным включение дополнительных регуляторных элементов для облегчения экспрессии, транспорта и биологической активности по меньшей мере одного гена, вставленного в вирусный геном онколитического вируса по изобретению (т.е. терапевтического(ких) гена(нов) и/или одного или более чем одного модулятора иммунологической контрольной точки). Например, может быть включен сигнальный пептид для облегчения секреции из инфицированной клетки. Сигнальный пептид типично вставлен на N-конце белка непосредственно после Met инициатора. Выбор сигнальных пептидов является широким, и он доступен специалистам в данной области. Также можно рассматривать добавление трансмембранного домена для облегчения заякоривания кодируемого(мых) белка(ков) в подходящей мембране (например, в плазматической мембране) инфицированных клеток. Трансмембранный домен типично вставляется на С-конце белка непосредственно до или в непосредственной близости от терминатора. В данной области доступно огромное разнообразие трансмембранных доменов (см., например, WO 99/03885).
В качестве дополнительного примера также может быть добавлена пептидная метка (типично короткая пептидная последовательность, которая может быть распознана доступными антисыворотками или соединениями) для следующей экспрессии, транспора или очистки кодируемого генного продукта. В контексте данного изобретения может быть использовано огромное разнообразие пептидных меток, включая, без ограничения, метку РК, октапептид FLAG, метку MYC, метку HIS (обычно отрезок из 4-10 остатков гистидина) и е-метку (US 6686152). Пептидная(ные) метка(ки) может(гут) быть независимо расположена(ны) на N-конце белка или, в качестве альтернативы, на его С-конце, или, в качестве альтернативы, внутри или в любом из данных положений при использовании нескольких меток. Пептидные метки могут быть выявлены посредством иммунодетектирующих анализов с использованием антител против метки.
В качестве другого примера, может быть изменено гликозилирование таким образом, чтобы увеличивать биологическую активность кодируемого генного продукта (например, увеличивать). Такие модификации можно осуществлять, например, посредством мутирования одного или более чем одного остатка в пределах сайта(тов) гликозилирования. Измененные картины гликозилирования могут увеличивать способность антител стимулировать ADCC (антителозависимая клеточная цитотоксичность) и/или их аффинность в отношении их мишени.
Другим подходом, которому можно следовать в контексте настоящего изобретения, является связывание генного продукта, кодируемого онколитическим вирусом по изобретению, с внешним агентом, таким как цитотоксический агент и/или метящий агент. Термин «цитотоксический агент» в том виде, как он используется в данном документе, относится к соединению, которое является непосредственно токсичным для клеток, предотвращая их репродукцию или рост, как, например, токсины (например, ферментативно активный токсин бактериального, грибного, растительного или животного происхождения, или его фрагменты). Термин «метящий агент» в том виде, как он используется в данном документе, относится к выявляемому соединению. Метящий агент может быть выявляемым сам по себе (например, метки на основе радиоактивных изотопов или флуоресцентные метки) или, в случае ферментативной метки, может катализировать химическую модификацию соединения-субстрата, которое является выявляемым. Связывание может осуществляться посредством генетического слияния между генным продуктом (терапевтическим(кими) геном(нами) и/или модулятором(рами) иммунологической контрольной точки и внешним агентом.
В предпочтительном воплощении онколитический вирус по изобретению представляет собой вирус осповакцины (предпочтительно из штамма Copenhague), дефектный по активностям и ТК, и RR (например, в результате инактивирующих мутаций в обоих вирусных генах J2R и I4L), в геном которого вставлена молекула нуклеиновой кислоты, кодирующая антитело против PD1. Желательно элементы в виде тяжелой и легкой цепи (например, тяжелая и легкая цепи для экспрессии mAb или их вариабельные фрагменты для экспрессии Fab и scFv) помещаются под транскрипционный контроль промоторов вируса осповакцины pH5R и р7.5К соответственно. Предпочтительно молекула нуклеиновой кислоты, кодирующая антитело против PD1, вставляется в локус ТК вирусного генома. Более предпочтительно указанный вирус осповакцины снабжается суицидным геном, с особым предпочтением в отношении описанного в данном документе суицидного гена FCU1. Даже более предпочтительно суицидный ген (например, FCU1) находится под транскрипционным контролем промотора вируса осповакцины р11К7.5. Еще более предпочтительно FCU1, помещенный под контроль промотора вируса осповакцины, вставляется в локус ТК вирусного генома.
В альтернативном и также предпочтительном воплощении онколитический вирус по изобретению представляет собой вирус осповакцины (предпочтительно из штамма Wyeth), дефектный в отношении активности ТК (в результате инактивирующих мутаций в гене J2R вируса), в геном которого вставлена молекула нуклеиновой кислоты, кодирующая антитело против PD1. Более предпочтительно указанный вирус осповакцины снабжен иммуностимулирующим терапевтическим геном с особым предпочтением в отношении человеческого гена GM-CSF, описанного в данном документе. Даже более предпочтительно терапевтический ген (например, GM-CSF) находится под транскрипционным контролем синтетического раннего-позднего промотора вируса осповакцины и предпочтительно вставлен в локус ТК.
Типично онколитический вирус по настоящему изобретению продуцируется в подходящей линии клеток-хозяев с использованием традиционных методик, включающих культивирование трансфицированных или инфицированных клеток-хозяев при подходящих условиях таким образом, чтобы обеспечивать продуцирование инфекционных вирусных частиц и выделение продуцированных инфекционных вирусных частиц из культуры указанных клеток, и возможно очистку указанных выделенных инфекционных вирусных частиц. Подходящие клетки-хозяева для продуцирования онколитического вируса включают, без ограничения, человеческие линии клеток, такие как HeLa (АТСС (Американская коллекция типовых клеточных культур)), клетки 293 (Graham et al., 1997, J. Gen. Virol. 36: 59-72), HER96, PER-C6 (Fallaux et al., 1998, Human Gene Ther. 9: 1909-17), клетки птиц, такие как клетки, описанные в WO 2005/042728, WO 2006/108846, WO 2008/129058, WO 2010/130756, WO 2012/001075 и т.д., линии клеток хомяка, такие как ВНК-21 (клетки почки новорожденного хомяка-21) (АТСС CCL-10), а также первичные фибробласты эмбриона цыпленка (CEF), полученные из эмбрионов цыплят, полученных из оплодотворенных яиц. Онколитический вирус может быть по меньшей мере частично выделен до применения согласно настоящему изобретению. Можно рассматривать разные стадии очистки, включая осветление, ферментативную обработку (например, бензоназа, протеаза), стадии хроматографии и фильтрования. Подходящие способы описаны в данной области (например, WO 2007/147528; WO 2008/138533, WO 2009/100521, WO 2010/130753, WO 2013/022764).
Продуцирование модулятора иммунологической контрольной точки
В одном воплощении онколитический вирус также может использоваться в контексте данного изобретения для продуцирования методами генной инженерии одного или более чем одного модулятора иммунологической контрольной точки, который он кодирует. Он может преимущественно содержать один или более чем один дополнительный элемент, обеспечивающий поддержание, размножение или экспрессию молекулы нуклеиновой кислоты, кодирующей модулятор иммунологической контрольной точки в клетке-хозяине. Такие дополнительные элементы содержат маркерный(ные) ген(ны) для того, чтобы облегчать идентификацию и выделение клеток-хозяев-продуцентов (например, посредством комплементации ауксотрофии клетки или посредством устойчивости к антибиотику). Подходящие маркерные гены включают, без ограничения, ген дигидрофолатредуктазы (dhfr), который придает устойчивость к метотрексату (Wigler et al., 1980, Proc. Natl. Acad. Sci. USA 77: 3567; O'Hare et al., 1981, Proc. Natl. Acad. Sci. USA 78: 1527); gpt, который придает устойчивость к микофеноловой кислоте (Mulligan and Berg, 1981, Proc. Natl. Acad. Sci. USA 78: 2072); neo, который придает устойчивость к аминогликозиду G-418 (Colberre-Garapin et al., 1981, J. Mol. Biol. 150: 1); zeo, который придает устойчивость к зеомицину, kana, который придает устойчивость к канамицину; hygro, который придает устойчивость к гигромицину (Santerre et al., 1984, Gene 30: 147). Рекомбинантные вирусы, не имеющие функциональной ТК (например, в результате вставки молекулы нуклеиновой кислоты, кодирующей модулятор иммунологической контрольной точки, в локус ТК), могут быть отобраны с использованием сред, содержащих бромдезоксиуридин (BrdU). В самом деле, ТК- вирусы являются нечувствительными к средству BrdU, тогда как данное средство препятствует синтезу ДНК в вирусах ТК+. Также можно полагаться на репортерные люминисцентные или колориметрические системы, например, основанные на GFP (зеленый флуоресцентный белок), люциферазе и бета-галактозидазе.
Способы рекомбинантного продуцирования модулятора иммунологической контрольной точки являются традиционными в данной области. Типично такие способы включают (а) введение онколитического вируса, описанного в данном документе, в подходящую клетку-продуцент с получением трансфицированной или инфицированной клетки-продуцента, (б) культивирование in vitro указанной трансфицированной или инфицированной клетки-продуцента при подходящих условиях для ее роста, (в) выделение одного или более чем одного модулятора иммунологической контрольной точки из культуры клеток и (г) возможно очистку выделенного модулятора(ров) иммунологической контрольной точки. В контексте данного изобретения клетки-продуценты предпочтительно представляют собой человеческие эукариотические клетки или эукариотические клетки, не являющиеся человеческими. Предпочтительные клетки-продуценты включают, без ограничения, ВНК-21 (клетки почки новорожденного хомяка), CV-1 (линия клеток почки африканской зеленой мартышки), клетки COS (например, COS-7), клетки яичника китайского хомяка (СНО), мышиные клетки NIH/3T3, мышиные миеломные клетки NSO, клетки HeLa, клетки Vero, клетки НЕК293, клетки PERC.6 и клетки птиц (например, куриные, утиные клетки, как описано в данном документе).
Клетки-продуценты можно культивировать в традиционных ферментационных биореакторах, колбах и чашках Петри. Культивирование можно проводить при температуре, рН и содержании кислорода, подходящих для данной клетки-хозяина. В данном документе не будет сделано попыток подробного описания разных способов, известных для продуцирования белков в эукариотических клетках. Продуцирование модулятора иммунологической контрольной точки может быть внутриклеточным или, предпочтительно, он может секретироваться наружу из кетки-продуцента (например, в культуральную среду).
Модулятор иммунологической контрольной точки можно затем очищать хорошо известными способами очистки. Использованные для очистки конкретного белка условия и технология будут зависеть от таких факторов, как условия экспрессии, нетто заряд, молекулярная масса, гидрофобность, гидрофильность, и они будут очевидными для специалистов в данной области. Кроме того, уровень очистки будет зависеть от намеченного применения. Если необходимо, особенно когда модулятор иммунологической контрольной точки не секретируется наружу из клетки-продуцента, или когда он не секретируется полностью, он может быть выделен посредством стандартных методик лизиса, включающих замораживание/оттаивание, обработку ультразвуком, механическое разрушение, применение лизирующих агентов и тому подобное. В случае секреции, он может быть выделен непосредственно из культуральной среды. Подходящие методики очистки включают, без ограничения, осаждение сульфатом аммония, кислотную экстракцию, гель-электрофорез, фильтрование и хроматографические способы (например, хроматография с обращенной фазой, гель-фильтрация, ионообменная, аффинная, на фосфоцелюлозе, на основе гидрофобных взаимодействий или на гидроксиапатите и т.д.). Желательно модулятор иммунологической контрольной точки, рекомбинантно продуцируемый для онколитического вируса по изобретению, по меньшей мере частично очищают в том смысле, что он по существу не содержит других антител, имеющих отличные антигенные специфичности и/или другой клеточный материал. Кроме того, модулятор иммунологической контрольной точки может быть приготовлен согласно условиям, традиционно используемым в данной области (например, WO 2009/073569).
Терапевтическое применение
Согласно настоящему изобретению также предложена композиция, содержащая терапевтически эффективное количество онколитического вируса по изобретению, возможно с фармацевтически приемлемым носителем. Такие композиции могут вводиться один или несколько раз и посредством одинаковых или разных путей.
«Терапевтически эффективное количество» соответствует количеству онколитического вируса, которое является достаточным для получения одного или более чем одного полезного результата. Такое терапевтически эффективное количество может варьировать как функция разных параметров, в частности, способа введения; болезненного состояния; возраста и массы субъекта; способности субъекта отвечать на лечение; вида сопутствующего лечения; частоты лечения и/или потребности в профилактике или терапии. Когда рассматривается профилактическое применение, онколитический вирус или композиция по изобретению вводится в достаточной дозе для предупреждения или для задержки начала и/или установления, и/или рецидива патологического состояния (например, пролиферативного заболевания, такого как рак), особенно у субъекта, подверженного риску. Для «терапевтического» применения онколитический вирус или композиция по настоящему изобретению вводится субъекту, у которого поставлен диагноз наличия патологического состояния (например, пролиферативного заболевания, такого как рак), с целью лечения заболевания, в конечном счете, в ассоциации с одним или более чем одним традиционным терапевтическим воздействием. В частности, терапевтически эффективное количество могло бы представлять собой то количество, которое необходимо для того, чтобы вызвать наблюдаемое улучшение клинического статуса по сравнению с исходным статусом или по сравнению с ожидаемым статусом в отсутствие лечения, например, уменьшение числа опухолей; уменьшение размера опухолей, уменьшение числа или степени метастазов, увеличение продолжительности ремиссии, стабилизация (т.е. не усугубление) состояния заболевания, задержка или замедление прогрессирования заболевания или увеличения его тяжести, уменьшение интенсивности или временное облегчение болезненного состояния, пролонгированное выживание, лучший ответ на стандартное лечение, улучшение качества жизни, пониженная смертность и т.д. Терапевтически эффективное количество также могло бы быть количеством, необходимым для вызова развития эффективного неспецифичного (врожденного) и/или специфичного противоопухолевого ответа. Типично развитие иммунного ответа в конкретном ответе Т-клеток можно оценивать in vitro, в подходящих животных моделях или с использованием биологических образцов, отобранных у субъекта. Например, для осуществления наблюдения за опухолью можно использовать методики, традиционно используемые в лабораториях (например, проточная цитометрия, гистология). Также можно использовать разные доступные антитела таким образом, чтобы идентифицировать разные популяции иммунных клеток, участвующих в противоопухолевом ответе, которые присутствуют у подвергнутых лечению субъектов, такие как цитотоксические Т-клетки, активированные цитотоксические Т-клетки, клетки-природные киллеры и активированные клетки-природные киллеры. Улучшение клинического статуса можно легко оценивать посредством любого релевантного клинического измерения, типично используемого врачами или другим квалифицированным персоналом системы здравоохранения.
Подразумевается, что термин «фармацевтически приемлемый носитель» включает все и любые носители, растворители, разбавители, эксципиенты, адъюванты, диспергирующие среды, покрытия, антибактериальные и противогрибковые средства, агенты для поглощения и тому подобное, совместимые с введением млекопитающим и, в частности, человеческим субъектам.
Онколитический вирус или его композицию можно помещать в растворитель или разбавитель, подходящий для применения у человека или животного. Растворитель или разбавитель предпочтительно является изотоничным, гипотоничным или слабо гипертоничным и имеет относительно низкую ионную силу. Показательные примеры включают стерильную воду, физиологический раствор (например, хлорид натрия), раствор Рингера, растворы глюкозы, трегалозы или сахарозы, раствор Хэнка и другие водные физиологически сбалансированные солевые растворы (см., например, самое современное издание Remington: The Science and Practice of Pharmacy, A. Gennaro, Lippincott, Williams&Wilkins).
В одном воплощении композиция онколитического вируса подходящим образом буферизована для применения у человека. Подходящие буферы включают, без ограничения, фосфатный буфер (например, PBS (фосфатно-солевой буферный раствор)), бикарбонатный буфер и/или Tris буфер, способные поддерживать физиологический или слабоосновной рН (например, от приблизительно рН 7 до приблизительно рН 9).
Композиции онколитического вируса также могут содержать другие фармацевтически приемлемые эксципиенты для предоставления желательных фармацевтических или фармакодинамических свойств, включающих, например, осмолярность, вязкость, прозрачность, цвет, стерильность, стабильность, скорость растворения композиции, модифицированное или поддерживающееся высвобождение или поглощение у человеческого или животного субъекта, стимуляцию транспорта через гематоэнцефалический барьер или проникновения в конкретный орган.
Композиции онколитического вируса также могут содержать один или более чем один адъювант, способный стимулировать иммунитет (особенно иммунитет, опосредованный Т-клетками) или облегчать инфекцию опухолевых клеток при введении, например, посредством Toll-подобных рецепторов (TLR), таких как TLR-7, TLR-8 и TLR-9, включая, без ограничения, квасцы, эмульсию на основе минерального масла, такую как полный и неполный (IFA) адъювант Фрейнда, липополисахарид или его производное (Ribi et al., 1986, Immunology and Immunopharmacology of Bacterial Endotoxins, Plenum Publ. Corp., NY, p407-419), сапонины, такие как QS21 (Sumino et al., 1998, J.Virol. 72: 4931; WO 98/56415), имидазо-хинолиновые соединения, такие как имиквимод (Suader, 2000, J. Am Acad Dermatol. 43:S6), S-27609 (Smorlesi, 2005, Gene Ther. 12: 1324), и родственные соединения, такие как соединения, описанные в WO 2007/147529, цитозинфосфат-гуанозиновые олигодезоксинуклеотиды, такие как CpG (Chu et al., 1997, J. Exp. Med. 186: 1623; Tritel et al., 2003, J. Immunol. 171: 2358), и катионные пептиды, такие как IC-31 (Kritsch et al., 2005, J. Chromatogr Anal. Technol. Biomed. Life Sci. 822: 263-70).
В одном воплощении композиция онколитического вируса по настоящему изобретению может быть приготовлена с целью улучшения его стабильности, в частности, при условиях изготовления и долговременного хранения (т.е. в течение по меньшей мере 6 месяцев, предпочтительно в течение по меньшей мере двух лет) при замораживании (например, -70°С, -20°С), охлаждении (например, 4°С) или при температурах окружающей среды. В данной области доступны разные композиции вируса либо в замороженной, либо в жидкой, либо в лиофилизированной форме (например, WO 98/02522, WO 01/66137, WO 03/053463, WO 2007/056847 и WO 2008/114021, и т.д.). Твердые (например, в виде сухого порошка или лиофилизированные) композиции можно получать способом, включающим вакуумную сушку и лиофилизацию (см., например, WO 2014/053571). Для иллюстративных целей буферизованные композиции, включающие NaCl и сахар, являются особенно адаптированными для консервации вирусов (например, 10 мМ Tris, рН 8, с 5% сахарозой (масс./об.), 10 мМ глутамат натрия и 50 мМ NaCl или фосфатно-солевой буферный раствор с глицерином (10%) и NaCl).
В некоторых воплощениях композиция онколитического вируса может быть приготовлена для обеспечения правильного распределения или замедленного высвобождения in vivo. Например, она может быть приготовлена в липосомах. Можно использовать биодеградируемые, биосовместимые полимеры, такие как этиленвинилацетат, полиангидриды, полигликолевая кислота, коллаген, полиортоэфиры и полимолочная кислота. Многие способы приготовления таких композиций описаны, например, J.R. Robinson в "Sustained and Controlled Release Drug Delivery Systems", ed., Marcel Dekker, Inc., New York, 1978.
Подходящая дозировка онколитического вируса может быть адаптирована как функция разных параметров и может быть определена согласно установившейся практике практикующим специалистом в свете релевантных обстоятельств. Подходящая дозировка для онколитического вируса варьирует от приблизительно 105 до приблизительно 1013 vp (вирусные частицы), iu (инфекционная единица) или pfu (бляшкообразующие единицы), в зависимости от вируса и использованной количественной методики. В качестве общего руководства, подходящими являются дозы вируса осповакцины от приблизительно 105 до приблизительно 1013 pfu, предпочтительно от приблизительно 106 pfu до приблизительно 1011 pfu, более предпочтительно от приблизительно 107 pfu до приблизительно 5×109 pfu; причем дозы от приблизительно 108 pfu до приблизительно 109 pfu являются особенно предпочтительными, особенно для применения у человека. Количество вируса, присутствующего в образце, можно определять общепринятыми методиками титрования, например, посредством подсчета числа бляшек после инфекции пермиссивных клеток с использованием пермиссивных клеток (например, ВНК-21 или CEF), иммуноокрашивания (например, с использованием антител против вируса; Caroll et al., 1997, Virology 238: 198-211), посредством измерения поглощения при 260 нм (титры vp) или, кроме того, посредством количественной иммунофлуоресценции (титры iu).
В предпочтительном воплощении введение(ния) указанной композиции онколитического вируса обеспечивает(ют) доставку по меньшей мере 50 нг/мл в образце из организма, полученном от субъекта. В одном воплощении указанный уровень иммунологической контрольной точки получают после одного введения от приблизительно 107 pfu до приблизительно 5×109 pfu онколитического вируса, описаного в данном документе, и он может дополнительно увеличиваться с последующим(щими) введением(ниями). В другом воплощении образец из организма представляет собой кровь, сыворотку, плазму, биопсию опухоли, гомогенат опухоли, жидкость из опухоли и т.д., и его отбирают по меньшей мере от одного часа до одного месяца после указанного(ных) введения(ний). Более предпочтительно, продуцирование in situ кодируемого модулятора иммунологической контрольной точки достигало по меньшей мере 100 нг (например, по меньшей мере 200 нг/мл, по меньшей мере 400 нг/мл, по меньшей мере 500 нг/мл, по меньшей мере 700 нг/мл, по меньшей мере 800 нг/мл, по меньшей мере 900 нг/мл, по меньшей мере 1000 нг/мл, по меньшей мере 1200 нг/мл, по меньшей мере 1500 нг/мл, по меньшей мере 1800 нг/мл, по меньшей мере 2000 нг/мл, по меньшей мере 2500 нг/мл и по меньшей мере 3000 нг/мл) на мл указанного биологического образца, отобранного у субъекта через 2-8 суток (например, через 3-7 суток или более предпочтительно примерно через 5 суток) после введения указанного онколического вируса. Уровни ингибитора иммунологической контрольной точки, секретируемого in situ, можно оценивать, как описано в Примерах.
Введение
Онколитический вирус или композицию по настоящему изобретению можно вводить в одной дозе (например, болюсной инъекции) или во многих дозах. При многократных введениях их можно осуществлять одинаковым или разными путями, и они могут иметь место в том же самом или в альтернативных сайтах. Также можно следовать посредством поседовательных циклов введений, которые повторяются после периода отдыха. Интервалы между каждым введением могут составлять от нескольких часов до одного года (например, 24 ч, 48 ч, 72 ч, еженедельно, каждые две недели, ежемесячно или ежегодно). Интервалы также могут быть нерегулярными (например, после прогрессирования опухоли). Дозы могут варьировать для каждого введения в пределах описанного выше интервала.
В контексте данного изобретения применимыми являются любые традиционные пути введения, включая парентеральный, местный пути или путь введения через слизистую. Парентеральные пути предназначены для введения в виде инъекции или инфузии. Обычными типами парентеральной инъекции являются внутривенная (в вену), внутриартериальная (в артерию), внутрикожная (в дерму), подкожная (под кожу), внутримышечная (в мышцу) и внутриопухолевая (в опухоль или в непосредственной близости от нее). Инфузии типично даются посредством внутривенного пути. Введения через слизистую включают, без ограничения, пероральный/алиментарный, интраназальный, внутритрахеальный, внутрилегочный, внутривагинальный или интраректальный пути. Местное введение также может осуществляться с использованием чрескожных средств (например, пластыря и тому подобного). При введениях могут использоваться традиционные шприцы и иглы (например, инъекционные иглы Quadrafuse) или любое соединение или устройство, доступное в данной области, способное облегчать или улучшать доставку активного(ных) агента(тов) субъекту. Предпочтительные пути введения для онколитического вируса включают внутривенный и внутриопухолевый пути.
В контексте данного изобретения онколитический вирус может вводиться один или несколько раз (например, 2, 3, 4, 5, 6, 7 или 8 раз и т.д.) в дозе в пределах интервала от 107 до 5×109 pfu. Интервал времени между каждым введением может варьировать от приблизительно 1 суток до приблизительно 8 недель, преимущественно от приблизительно 2 суток до приблизительно 6 недель, предпочтительно от приблизительно 3 суток до приблизительно 4 недель и даже более предпочтительно от приблизительно 1 недели до приблизительно 3 недель (например, каждые две недели). Предпочтительная терапевтическая схема включает от 2 до 5 (например, 3) внутривенных или внутриопухолевых введения 108 или 109 pfu онколитического вируса осповакцины с приблизительно 1- или 2-недельным интервалом.
Настоящее изобретение также относится к способу лечения пролиферативного заболевания, такого как рак, включающему введение онколитического вируса, как описано в данном документе, субъекту, нуждающемуся в этом.
Настоящее изобретение также относится к способу ингибирования роста опухолевых клеток in vivo, включающему введение онколитического вируса, как описано в данном документе, субъекту, нуждающемуся в этом.
Настоящее изобретение также относится к способу усиления иммунного ответа на опухолевые клетки, включающему введение онколитического вируса, как описано в данном документе, субъекту, нуждающемуся в этом.
В одном воплощении введение онколитического вируса для применения в настоящем изобретении вызывает, стимулирует и/или переориентирует иммунный ответ. В частности введение индуцирует защитный ответ Т- или В-клеток у хозяина, которого лечат, например, против указанного онколитического вируса или, в конечном счете, против продукта, кодируемого терапевтическим(кими) геном(нами), вставленным(ными) в вирусный геом, если таковой существует. Защитный ответ Т-клеток может быть опосредован клетками CD4+ или CD8+, или как CD4+, так и CD8+клетками. Ответ В-клеток можно измерять посредством ELISA (твердофазный иммуноферментный анализ), и ответ Т-клеток можно оценивать традиционным ELISpot (метод иммуноферментных пятен), анализами ICS (внутренний контрольный образец) из любого образца (например, кровь, органы, опухоли и т.д.), отобранного у иммунизированного животного или субъекта.
В одном воплощении введение онколитического вируса обеспечивает изменение микроокружения опухоли с целью увеличения активности эффекторных клеток в опухоли, особенно эффекторных Т-лимфоцитов, и/или стимуляцию по меньшей мере частичного истощения Treg. Клетки, инфильтрующие опухоль, можно легко идентифицировать, например, традиционными анализами иммуноокрашивания.
В одном воплощении способ по изобретению обеспечивает более высокую терапевтическую эффективность, чем терапевтическая эффективность, полученная при тех же самых условиях с аналогичным онколитическим вирусом (без модулятора иммунологической контрольной точки) или с модулятором иммунологической контрольной точки, либо индивидуально, либо даже при совместном введении. В контексте данного изобретения способ по изобретению обеспечивает по меньшей мере на 5%, по меньшей мере на 10%, по меньшей мере на 15%, по меньшей мере на 20% или по меньшей мере на 25% большую терапевтическую эффективность, чем один из вируса или модулятора иммунологической контрольной точки, одиночно или при совместном введении. Более высокая терапевтическая эффективность могла бы быть подтверждена, как описано выше, в связи с термином «терапевтически эффективное количество» с конкретным предпочтением в отношении более длительного выживания.
Примеры пролиферативных заболеваний, которые можно лечить с использованием онколитического вируса, композиции или способов по изобретению, включают рак кости, рак печени, рак поджелудочной железы, рак желудка, рак толстой кишки, рак пищевода, ротоглоточный рак, рак легкого, рак головы и шеи, рак кожи, меланому, рак матки, рак шейки матки, рак яичника, рак молочной железы, рак прямой кишки, рак анальной области, рак предстательной железы, лимфому, рак эндокринной системы, рак щитовидной железы, саркому мягкой ткани, хронические и острые лейкозы, рак мочевого пузыря, рак почки, новообразование центральной нервной системы (ЦНС), глиому и т.д. Настоящее изобретение также является полезным для лечения метастатических раковых заболеваний, особенно метастатических раковых заболеваний, при которых экспрессируется PD-L1 (Iwai et al., 2005, Int. Immunol. 17: 133-44). Предпочтительные раковые заболевания, которые можно лечить с использованием онколитического вируса изобретению, включают раковые заболевания, типично отвечающие на иммунотерапию. Неограничивающие примеры предпочтительных раковых заболеваний для лечения включают меланому (например, метастатическую злокачественную меланому), рак почки (например, светлоклеточный рак), рак предстательной железы (например, аденокарцинома предстательной железы, не поддающаяся лечению гормонами), рак молочной железы, колоретальный рак, рак легкого (например, немелкоклеточный рак легкого) и рак печени (например, гепатокарцинома).
Согласно преимущественному воплощению, особенно когда онколитический вирус снабжается суицидным геном, терапия онколитическим вирусом или способы согласно настоящему изобретению могут содержать дополнительную стадию, на которой субъекту вводятся фармацевтически приемлемые количества пролекарства, преимущественно аналога цитозина, в частности 5-FC. В качестве иллюстрации можно использовать дозу от 50 до 500 мг/кг/сутки, причем предпочтительной является доза 200 мг/кг/сутки или 100 мг/кг/сутки. В контексте настоящего изобретения пролекарство вводится согласно стандартной практике (например, перорально, системно и т.д.). Предпочтительно введение имеет место после введения онколитического вируса, предпочтительно по меньшей мере через 3 суток, более предпочтительно по меньшей мере через 4 суток и даже более предпочтительно по меньшей мере через 7 суток после введения вируса. Пероральный путь является предпочтительным. Можно вводить одну дозу пролекарства или дозы, которые повторяются в течение времени, которое является достаточно длительным для обеспечения продукции токсичного метаболита в организме- или клетке-хозяине.
Онколитический вирус, композиция или способ согласно изобретению могут быть ассоциированы с одним или более чем одним веществом или терапией, эффективными в противораковой терапии, и настоящее изобретение также касается способа, который включает стадию доставки субъекту дополнительной противораковой терапии. В контексте настоящего изобретения указанная дополнительная противораковая терапия включает хирургию, лучевую терапию, химиотерапию, иммунотерапию, гормональную терапию или их комбинацию. В предпочтительном воплощении способ по изобретению включает введение одного или более чем одного вещества, эффективного в противораковой терапии. Среди фармацевтических веществ, эффективных в противораковой терапии, которые можно использовать в ассоциации или в комбинации с онколитическим вирусом, композицией или способом по изобретению, можно более конкретно упомянуть:
- алкилирующие агенты, такие как, например, митомицин С, циклофосфамид, бусульфан, ифосфамид, изосфамид, мелфалан, гексаметилмеламин, тиотепа, хлорамбуцил или дакарбазин;
- антиметаболиты, такие как, например, гемцитабин, капецитабин, 5-фторурацил, цитарабин, 2-фтордезоксицитидин, метотрексат, идатрексат, томудекс или триметрексат;
- ингибиторы топоизомеразы II, такие как, например, доксорубицин, эпирубицин, этопозид, тенипозид или митоксантрон;
- ингибиторы топоизомеразы I, такие как, например, иринотекан (СРТ-11), 7-этил-10-гидрокси-камптотецин (SN-38) или топотекан;
- антимитотические лекарственные средства, такие как, например, паклитаксел, доцетаксел, винбластин, винкристин или винорелбин;
- производные платины, такие как, например, цисплатин, оксалиплатин, спироплатин или карбоплатин;
- ингибиторы рецепторов тирозинкиназы, такие как сунитиниб (Pfizer) и сорафениб (Bayer);
- антинеопластические антитела, в частности, антитела, которые воздействуют на регуляцию рецепторов поверхности клетки, такие как трастузумаб, цетуксимаб, панитумумаб, залутумумаб, нимотузумаб, матузумаб, бевацизумаб и ранибизумаб.
- ингибиторы EGFR (рецептор эпидермального фактора роста), такие как гефитиниб, эрлотиниб и лапатиниб; и
- иммуномодулирующие агенты, такие как, например, альфа-, бета- или гамма-интерферон, интерлейкин (в частности, IL-2, IL-6, IL-10 или IL-12) или фактор некроза опухолей;
Онколитический вирус, композицию или способ согласно изобретению также можно использовать в ассоциации с лучевой терапией.
Согласно настоящему изобретению также предложены наборы, включающие другой контейнер (например, стерильный стеклянный или пластмассовый флакон) для каждой дозы вируса, подлежащей введению. Возможно данный набор может включать устройство для осуществления введения активных агентов. Набор также может включать листок-вкладыш в упаковке, включающий информацию, касающуюся композиций или индивидуального компонента и лекарственных форм в наборе.
Краткое описание графических материалов
Фиг. 1 иллюстрирует количество очищенного mAb против PD-1, Fab и scFv, продуцированных из инфицированных CEF.
Фиг. 2 иллюстрирует анализ посредством электрофореза очищенного рекомбинантного mAb (mAb вируса осповакцины) и scFv (scFv вируса осповакцины), а также имеющегося в продаже J43 (BioXCell) при восстанавливающих (R) и невосстанавливающих (NR) условиях.
Фиг. 3: эффективность репликации вирусов in vitro в LoVo, инфицированных указанными вирусами при MOI (множественность инфекции) 0,0001, 0,001 и 0,01 в сутки 5 после инфекции. Значения представляются как среднее плюс/минус SD (стандартное отклонение) трех индивидуальных определений.
Фиг. 4 иллюстрирует аминокислотные последовательности тяжелой и легкой цепей антител против PD1, экспрессируемых онколитическим вирусом осповакцины, описанным в данном документе.
Фиг. 5 иллюстрирует анализ иммуноблоттингом рекомбинантного mAb и Fab, секретируемых в супернатанты культуры клеток, полученные из CEF, инфицированных WRTG18618, WRTG18619, WRTG18620 и WRTG18621. Имеющееся в продаже J43 используется в качестве контроля; М представляет маркеры молекулярной массы, и С - негативный контроль.
Фиг. 6 иллюстрирует концентрацию рекомбинантного J43, продуцированного в сыворотке (Фиг. 6А) и в гомогенате опухоли (Фиг. 6Б) мышей, которых лечили с использованием одного введения WRTG18618 (107 pfu), инъецированного в опухоль (образующуюся в результате подкожной имплантации опухолей B16F10) или подкожно (мышам без опухолей), или с использованием внутриопухолевого введения 1 мкг или 10 мкг имеющегося в продаже J43 (Bioxcell). Концентрацию рекомбинантного J43 измеряли посредством количественной ELISA через 1, 5 и 11 суток после инъекции.
ПРИМЕРЫ
Данные примеры иллюстрируют онколитический вирус осповакцины, генетически модифицированный для экспрессии разных форм ингибиторов против иммунологических контрольных точек. Доклиническим доказательством полезных эффектов экспрессирования блокаторов иммунологических контрольных точек из вирусных векторов была демонстрация в мышиных моделях опухолей. Это подразумевает применение i) специфичных для мышей антител против иммунологических контрольных точек и ii) онколитического вируса группы оспы, способного инфицировать мышиные клетки с высокой эффективностью.
Специфичное в отношении мыши антитело хомяка J43 было выбрано для направленного воздействия на блокатор иммунологической контрольной точки - мышиный PD-1 (mPD-1). Было показано, что данное антитело блокирует взаимодействие mPD1 с PD-L1 (патент США 7858746). Антитело J43 и его изотипический контроль (IgG хомяка) доступны от BioXCell. Антитела против mPD1 и их изотипический контроль использовали для создания функциональных анализов и количественного ELISA in vitro. Кроме того, J43 и его разные формы клонировали в онколитическом вирусе и тестировали in vivo в животных моделях опухолей.
Онколитический вирус, выбранный для данных исследований, представляет собой штамм Western Reserve (WR) вируса осповакцины (VV), дефектный по тимидинкиназе (TK-) (локус J2R) и RR- (локус I4L), что делает вирус нереплицируемым в здоровых (неделящихся) клетках. В отличие от этого, предполагается то, что VV TK-RR- селективно и эффективно реплицируется в опухолевых клетках.
Векторизация молекул антитела против mPD-1 в онколитическом вирусе осповакцины ТК-RR-
Тяжелая цепь J43 демонстрировала высокую гомологию с тяжелой цепью IgG против CD79b. Таким образом, последовательность, сохраненная для клонирования тяжелой цепи, представляла собой вариабельную цепь J43 и константную цепь антитела против CD79b. Легкую цепь J43 клонировали с сигнальной последовательностью из легкой цепи антитела против CD79b. Тяжелую и легкую цепи помещали под контроль вирусных промоторов pH5R или р7.5К, которые имели немного разную силу. Помимо конструкций «целого» антитела, получали производные, соответственно, конструкции антигенсвязывающих фрагментов (Fab), меченных His, а также конструкции одноцепочечного антитела (scFv), меченного His. Для Fab также генерировали два формата конструкций, в зависимости от частей легкой или тяжелой цепи, помещенных под контроль каждого промотора. Все пять конструкций вставляли в остов VV WR с делецией TK и RR. Данные конструкции обобщены ниже:
WRTG18618 (или mAb1), соответствующая pH5R-HC - p7.5K-LC
WRTG18619 (или mAb2), соответствующая pH5R-LC - р7.5К-НС
WRTG18621 (или Fab1), соответствующая pH5R-(VH-CH1-6His) - p7.5K-LC
WRTG18620 (или Fab2), соответствующая pH5R-LC - p7.5K-(VH-CH1-6His)
WRTG18616 (scFv), соответствующая pH5R-VH-gs-VL-6His).
НС и LC представляют собой сокращения тяжелой и легкой цепей, VH и VL - вариабельных доменов тяжелой и легкой цепи, 6His - гистидиновой метки (из 6 гистидинов), и gs (полиглицин-сериновый линкер).
Конструкции 1 продемонстрировали наивысший уровень экспрессии рекомбинантного mAb или Fab с ожидаемыми профилями сборки цепи.
Маточные растворы вируса WRTG18616 (scFv), WRTG18618 (mAM) и WRTG18621 (Fab1) продуцировали в клетках ВНК-21 и очищали посредством проточного тангенциального фильтрования (TFF).
Конструирование WRTG18618 (pH5R-HC - p7.5K-LC): mAb1
Фрагмент, содержащий НС, с последующим промотором р7.5К получали синтетическим способом (Geneart: Регенсбург, Германия) и клонировали в плазмиду для переноса вируса осповакцины pTG18496, ограниченную PstI и EcoRI, с получением pTG18614. Фрагмент, содержащий LC, получали синтетическим способом и клонировали в pTG18614, ограниченную NsiI и SalI, с получением PTG18618.
Плазмида для переноса вируса осповакцины pTG18496 сконструирована для обеспечения вставки нуклеотидной последовательности, подлежащей переносу, посредством гомологичной рекомбинации в гене ТК генома VV. Она содержит фланкирующие последовательности (BRGTK и BRDTK), окружающие ген J2R, и промотор pH5R, с последующими сайтами множественного клонирования.
Аминокислотные последовательности НС и LC антитела против PD-1 приведены в SEQ ID NO: 1 и SEQ ID NO: 2 соответственно и на Фиг. 4.
Получение WRTG18618 проводили посредством гомологичной рекомбинации в первичных фибробластах куриных эмбрионов (CEF), инфицированных WR с делецией RR и трансфицированных pTG18618 посредством нуклеофекции (согласно методике Amaxa Nucleofector). Селекцию вирусов осуществляли посредством очистки бляшек после роста в клетках 143В, дефицитных по тимидинкиназе (ТК-), культивируемых в присутствии бромдезоксиуридина. Данная селекция обеспечивает то, что только rWR ТК- остаются жизнеспособными. Отсутствие загрязнения родительским WR подтверждали ПЦР (полимеразная цепная реакция).
Конструирование WRTG18619 (pH5R-LC - р7.5К-НС): mAb2
Фрагмент, содержащий LC, с последующим промотором р7.5K получали
синтетическим способом и клонировали в плазмиду для переноса вируса
осповакцины pTG18496, ограниченную PstI и EcoRI, с получением pTG18615.
Фрагмент, содержащий НС, получали синтетическим способом и клонировали в
pTG18615, ограниченную NsiI и MluI, с получением pTG18619.
Получение вируса WRTG18619 проводили в CEF посредством гомологичной
рекомбинации, как описано выше.
Конструирование WRTG18621 (pH5R-(VH-CH1-6His) - p7.5K-LC): Fab1 Фрагмент, содержащий VH-CH1-6His, с последующим промотором р7.5К
получали синтетическим способом и клонировали в плазмиду для переноса вируса
осповакцины pTG18496, ограниченную PstI и EcoRI, с получением pTG18617.
Фрагмент, содержащий LC, получали синтетическим способом и клонировали в
pTG18617, ограниченную NsiI и SalI, с получением pTG18621.
Получение вируса WRTG18621 проводили в CEF посредством гомологичной рекомбинации, как описано выше.
Конструирование WRTG18620 (pH5R-LC-p7.5K-(VH-CH1-6His): Fab2
Фрагмент, содержащий LC с последующим промотором р7.5K, получали синтетическим способом и клонировали в плазмиду для переноса вируса осповакцины pTG18496, ограниченную PstI и EcoRI, с получением pTG18615. Фрагмент, содержащий VH-CH1-6His, получали синтетическим способом и клонировали в pTG18615, ограниченную NsiI и MluI, с получением pTG18620.
Получение вируса WRTG18620 проводили в CEF посредством гомологичной рекомбинации, как описано выше.
Конструирование WRTG18616 (pH5R-VH-gs-VL-6His): scFv
Фрагмент, содержащий VH-gs-VL-6His, получали синтетическим способом и клонировали в плазмиду для переноса вируса осповакцины pTG18496, ограниченную PstI и EcoRI, с получением pTG18616.
Получение вируса WRTG18616 проводили в CEF посредством гомологичной рекомбинации, как описано выше.
Характеризация in vitro кодируемых антител против PD-1
Рекомбинантные онколитические векторы, описанные выше, тестировали для оценки продукции кодируемых молекул антител против mPD-1. Для этого инфицировали пермиссивные первичные клетки или линии клеток и отбирали супернатант.Присутствие молекул антитела против mPD-1 можно определять посредством SDS-PAGE (электрофорез в полиакриламидном геле с додецилсульфатом натрия), анализа вестерн-блоттингом, масс-спектрометрического анализа, ELISA (твердофазный иммуноферментный анализ) и функциональных анализов. Имеющиеся в продаже антитела против mPD-1 использовали в качестве контролей.
Супернатант от инфицированных CEF (moi 0,2) анализировали (через 48 и 72 ч после инфекции) посредством вестерн-блоттинга при невосстанавливающих условиях с использованием для выявления поликлонального антитела против IgG хомяка. WRTG18618 (mAb1) и WRTG18621 (Fab1) могли быть выявлены посредством ELISA. Профиль экспрессии продукта, секретированного клетками, инфицированными WRTG18618, имел сходную картину с профилем экспрессии имеющегося в продаже антитела J43, и наблюдали ожидаемую сборку продукта, секретированного клетками, инфицированными WRTG18621. Экспрессию scFv ожидаемого размера также выявляли в супернатанте культуры посредством вестерн-блоттинга.
Очистка mAb и оценка in vitro антител против PD-1
CEF культивировали в колбах F175 и инфицировали 2,7×108 pfu вирусов, экспрессирующих антитела против PD-1. Через 48-72 ч супернатанты культуры отбирали и подвергали фильтрованию на 0,2 мкм фильтрах. mAb1 очищали посредством колонки Hitrap на основе белка A (GE Healthcare), с последующей гель-фильтрацией на Superdex 200 (GE Healthcare), тогда как FaM и scFv очищали посредством HIS-Trap (GE Healthcare), с последующей гель-фильтрацией на Superdex 75 (GE Healthcare). scFv элюировали из колонки для гель-фильтрации в двух пиках: первом, соответствующем димерам, и втором - мономерам. Продуцировались значительные количества mAb, Fab и scFv, как показано на Фиг. 1. Выход составляет от 0,5 до 2 мкг/мл супернатанта согласно формату антитела.
Рекомбинантные очищенные mAb имели электрофоретический профиль, аналогичный имеющимся продаже mAb, с правильной сборкой двух легких и двух тяжелых цепей при невосстанавливающих условиях и выявлением индивидуальных легкой и тяжелой цепей при восстанавливающих условиях, как проиллюстрировано на Фиг. 2. Кроме того, очищенное scFv перемещалось на геле SDS-PAGE согласно ожидаемому размеру (см. Фиг. 2).
Рекомбинантные продукты могут быть выявлены посредством ELISA, и они являются функциональными в конкурентном ELISA (т.е. блокирование связывания мышиного PD-L1-биотин в концентрации 0,2 мкг/мл с мышиным PD-1, покрытие которым осуществлялось в концентрации 1 мкг/мл), причем mAb является более эффективным, чем Fab в данном конкурентном эксперименте. Кроме того, рекомбинантное mAb1 является более эффективным, чем его имеющийся в продаже аналог J43, а также продуцируемая VV димерная фракция scFv является более эффективной, чем мономерная фракция, с различием ЕС50 (полумаксимальная эффективная концентрация) приблизительно один log.
Способность mAb1, Fab1 и scFv, кодируемых VV, взаимодействовать с PD-1 поверхности клетки исследовали в анализах связывания на основе проточной цитометрии с использованием PD-1-позитивных Т-лимфомных линий клеток EL4 и RMA. 105 клеток EL4 или RMA инкубировали в течение 45 мин на льду со 100 мкл одного из mAb1 (5 мкг/мл), имеющегося в продаже антитела против mPD-1 J43 (5 мкг/мл, BioXCell) или негативного контроля - IgG хомяка (5 мкг/мл, BioXCell) и промывали. Связывание с PD-1 выявляли посредством инкубирования клеток в течение 45 мин на льду со 100 мкл 10 мкг/мл смеси конъюгированных с FITC моноклональных мышиных антител против антител армянского плюс сирийского хомяка (BD Pharmingen). После промывки интенсивность флуоресценции измеряли на проточном цитометре Navios™ (Beckman Coulter). Данные анализировали с использованием программы Kaluza 1.2 (Beckman Coulter). mAb1, кодируемое WRTG18618, эффективно связывалось с клетками EL4 и RMA.
Для измерения связывания Fab1 и scFv 5×105 клеток EL4 или RMA не прямо окрашивали либо 5 мкг/мл Fab1, либо 2,5 мкг/мл мономерного scFv (которые оба являются меченными His), с последующим окрашиванием моноклональным мышиным антителом против His метки, конъюгированным с РЕ (фикоэритрин) (разведенным 1/10, Miltenyi Biotec), и анализировали, как описано выше. Было показано, что Fab1 и мономерный scFv, продуцируемые из WRTG18621 и WRTG18616, эффективно связываются с поверхностью клеток EL4 и RMA. Специфичное связывание могло быть продемонстрировано посредством совместного инкубирования клеток EL4 с Fab1 или мономерным scFv и полноразмерным J43, или IgG хомяка негативного контроля от BioXCell, с последующим окрашиванием антителом против His метки, конъюгированным с РЕ. Окрашивание EL4 уменьшалось в присутствии полноразмерного J43, но не IgG хомяка негативного контроля. В целом, данные результаты продемонстрировали то, что mAb1, Fab1 и scFv, кодируемые VV, могли распознавать эндогенно экспрессируемый PD-1 в двух мышиных линиях клеток Т-лимфоцитов.
Осуществляли конкурентный анализ на основе проточной цитометрии для сравнения блокирующей активности молекул против mPD-1. Данный анализ основывается на линии клеток EL4 мышиной Т-лимфомы, которая является сильно PD-1 позитивной. Связывание mPD-L1-hFc (мышиный PD-лиганд 1, содержащий фрагмент Fc человеческого IgG) с PD1 поверхности клетки можно выявлять проточной цитометрией с использованием моноклонального антитела против hFc, меченного РЕ. Связывание mPD-L1-hFc может конкурировать с антителами против PD-1, приводя к уменьшению сигнала РЕ, связанного с клеткой.
Ингибирующую активность рекомбинантных mAb1, Fab1 и scFv на связывание PD-L1 с клетками EL4 оценивали и сравнивали с имеющимся в продаже антителом против PD-1 J43 (BioXCell). Более конкретно, 105 клеток EL4 совместно инкубировали с 2 мкг/мл mPD-L1-hFc (R&D Systems) и возрастающими концентрациями разных антител против PD-1 в 100 мкл в течение 45 мин на льду. После промывки клетки инкубировали со 100 мкл 5 мкг/мл конъюгированного с РЕ мышиного антитела против Fc человеческого IgG (BioLegend) в течение 45 мин на льду. Клетки промывали и измеряли среднюю интенсивность флуоресценции, как описано выше. При данных условиях клон J43 контрольного антитела против mPD-1 демонстрировал IC50 (полумаксимальная ингибирующая концентрация) 1,6 мкг/мл.
Как и ожидалось, контрольный IgG хомяка не показал какой-либо блокирующей активности на связывание PD-L1 с клетками EL4. Рекомбинантные mAb1, Fab1, мономерный и димерный scFv, продуцированные из WRTG18618, WRTG18621 и WRTG18616 соответственно, могли блокировать связывание лиганда с рецептором поверхности клетки, как и J43 от BioXCell. Важно то, что mAb1, кодируемое WRTG18618, демонстрировало в данном анализе повышенную блокирующую активность по сравнению с J43 от BioXCell и версиями Fab1 scFv.
Осуществляли количественный ELISA для количественного измерения концентраций антител, продуцируемых в мышиной сыворотке или рекомбинантными клетками, инфицированными WR, с использованием покрытых происходящих от кролика антител против антител хомяка для захвата J43, которые, в свою очередь, могли быть выявлены происходящим от козы антителом против антитела хомяка. Данный анализ является чувствительным к мышиной сыворотке, и стандартные кривые должны быть получены в присутствии 50%-ной мышиной сыворотки.
Анализ цитотоксичности in vitro (онколитическая активность)
Линию клеток рака толстой кишки человека LoVo трансдуцировали в суспензии при MOI 0,001 и 0,0001 вирусом осповакцины WR с делециями TK-RR без трансгена (WRTG18011) и вирусом осповакцины WR с делециями TK-RR, экспрессирующим разные молекулы против mPD-1 (WRTG18616, WRTG18618 и WRTG8621). Всего 3×105 клеток/лунку высевали в 6-луночные культуральные чашки в 2 мл среды, дополненной 10% FCS (фетальная телячья сыворотка). Клетки затем культивировали при 37°С, и выживание клеток определяли через 5 суток посредством исключения трипанового синего. Как показано на Фиг. 3, онколитическая активность разных вирусов приводила к 75-99%-ному и 99%-ному уменьшению числа жизнеспособных клеток при MOI 0,0001 и 0,001 соответственно. Вирус WR с делецией TK-RR без трансгена и вирус WR с делецией TK-RR, экспрессирующий разные молекулы против mPD-1, не показывали различий в цитотоксичности. Таким образом, экспрессия разных молекул антител против mPD-1 не влияет на онколитическую активность вируса осповакцины.
Оптимальная схема экспрессии антител
Рекомбинантные онколитические векторы тестировали иммуноблоттингом для оценки уровней экспрессии и сборки секретированных антител. Анализы иммуноблоттингом проводили на супернатантах культуры клеток, отобранных через 24 ч после инфекции CEF конструкциями mAb и Fab, описанными выше. Супернатант центрифугировали 5 мин при 16000 g, и 25 мкл готовили в буфере Лэмли без восстанавливающего агента и загружали на предварительно заполимеризованный 4-15%-ный полиакриламидный гель (Biorad). В качестве контрольной молекулы использовали имеющееся в продаже моноклональное антитело J43 (BioXcell). На гель загружали один мкг каждой молекулы. Гель-электрофорез проводили при невосстанавливающих условиях для того, чтобы сохранить сборку легких и тяжелых цепей и обеспечить оптимальное выявление (т.е. поликлональное антитело, используемое для выявления, не распознавало восстановленные цепи IgG и Fab). Белки затем переносили на PVDF (поливинилиденфторид) мембрану с использованием системы Trans-blot Turbo (набор Transblot Turbo Transfer от Biorad) с предварительно запрограммированным протоколом (высокая MW (молекулярная масса): 10 мин; постоянная сила тока 2,5 А; вплоть до 25В). Мембраны насыщали в течение ночи при 4°С в блокирующем растворе (8 мМ NaPO4, 2 мМ KPO4, 154 мМ NaCl, рН 7,2 (PBS) (фосфатно-солевой буферный раствор), дополненный 0,05% Tween 20, 5% нежирным сухим молоком, Biorad). Для иммуновыявления антител использовали конъюгированное с пероксидазой хрена (HRP) антитело козы против IgG армянского хомяка (Jackson Immunoresearch) в концентрации 80 нг/мл в буфере для разведения (PBS, 0,05% Tween 20, 0,5% нежирное сухое молоко). Разработку проводили с использованием выявляющих реактивов для вестерн-блоттинга Amersham ECL Prime, и для фиксации хемилюминисценции использовали Molecular Imager ChemiDOC™ XRP.
Как проиллюстрировано на Фиг. 5, и WRTG18618, и WRTG18619 секретировали примерно одинаковые количества продукта-антитела с кажущимся размером, соответствующим двум тяжелым и двум легким цепям, связанным вместе, сопоставимым с размером имеющегося в продаже mAb J43. Однако WRTG18619 также продуцировал неожиданный продукт, мигрирующий между маркерами 43 и 55 кДа (см. дополнительную полосу, указанную стрелкой на четвертой дорожке «18619»). Аналогичным образом, дополнительная полоса также присутствовала в супернатанте культуры, инфицированной WRTG18620 (то же самое положение и та же самая интенсивность на пятой дорожке «18620»), тогда как WRTG18621 продуцировал большое количество правильно собранного Fab без какого-либо выявляемого неправильно собранного продукта (см. кончик стрелки на шестой дорожке «18621»).
Анализы супернатантов культуры клеток, полученных после инфекции линий клеток млекопитающих ВНК21 и А549, давали такой же профиль (данные не показаны).
Глядя на схему конструкций, оказывается, что в обеих конструкциях WRTG18619 и WRTG18620 легкая цепь антитела была помещена под транскрипционный контроль промотора pH5R (сильный промотор). Одной гипотезой является то, что в данной конфигурации легкие цепи антитела могли сверхэкспрессироваться по отношению к тяжелым цепям и, таким образом, могли собираться в гомодимеры. Следовательно, данная дополнительная полоса, мигрирующая между 43 и 55 кДа, могла бы соответствовать сверхпродуцируемым легким цепям, которые собрались в гомодимер теоретической массы 47 кДа.
Следовательно, предпочтительно экспрессировать тяжелые цепи под контролем более сильного промотора, чем промотор, используемый для экспрессии легких цепей для того, чтобы уменьшать риск получения неправильной сборки (например, гомодимеров легких цепей).
В целом, данные результаты подтвердили способность рекомбинантных онколитических вирусов, описанных в данном документе, секретировать mAb и Fab на выявляемом уровне. WRTG18618 и WRTG18621 выбрали для остальных экспериментов из-за их способности продуцировать продукты-антитела, которые ближе к ожидаемым, чем продукты WRTG18619 и WRTG18620.
Экспрессия in vivo векторизированного антитела против PD1
Экспрессию векторизированного mAb J43 против PD-1 оценивали in vivo у мышей с имплантированными подкожно опухолями B16F10 или без них. WRTG18618 (конструкция mAb1) инъецировали либо внутриопухолево, либо подкожно и сравнивали с имеющимся в продаже J43 (внутриопухолевая инъекция).
Эксперименты in vivo и отбор образцов
Более конкретно, 3×105 клеток B16F10 (мышиная меланома) инъецировали подкожно (п.к.) в правые бока шестинедельных самок мышей C57BL/6. В сутки 0 (D0), когда объемы опухоли достигали 100-200 мм3, внутриопухолево (в.о.) инъецировали 100 микролитров либо WRTG18618 (107 pfu), либо имеющегося в продаже J43 (1 мкг или 10 мкг, Bioxcell).
WRTG18618 также инъецировали п. к. мышам без опухоли.
Кровь и опухоли отбирали в D1, D5 и D11. Для каждого момента времени 3 мышей анестезировали 200 мкл пентобарбитала, и отбирали кровь посредством внутрисердечной пункции. Кровь хранили при 4°С на протяжении 8 часов, и сыворотку выделяли после двух центрифугирований и выдерживали при -20°С до анализа. После отбора крови мышей умерщвляли посредством смещения шейных позвонков, и выделяли опухоль. Опухоли взвешивали, разрезали на маленькие кусочки и переносили в пробирки С-типа GentleMACS (Miltenyi), содержащие 3 мл PBS. Опухоли механически диссоциировали, применяя программу «m-imptumor01» (GentleMACS, Miltenyi). После центрифугирования при 300 g в течение 7 мин супернатанты выделяли и выдерживали при -20°С до анализа.
Концентрацию J43 измеряли количественным ELISA и в сыворотке, и в гомогенатах опухоли в разные моменты времени после инъекций вируса или имеющегося в продаже mAb J43.
Количественный ELISA
Концентрации J43 оценивали как в сыворотке, так и в гомогенатах опухоли. Более конкретно, девяносто шести-луночные планшеты (иммунопланшеты Maxisorp от Nunc) инкубировали в течение ночи при 4°С со 100 мкг/лунку 0,8 мкг/мл антитела козы против IgG хомяка (Southern Biotech) в растворе для покрытия (0,05 М карбонат Na, рН 9,6, Sigma). Планшеты промывали три раза промывочным буфером (300 мкл/лунку PBS, 0,05% Tween 20) и инкубировали 1 ч при RT (комнатная температура) с 200 мкл/лунку блокирующего раствора (PBS, 0,05% Tween 20, 5% нежирное сухое молоко). Планшеты три раза промывали промывочным буфером. Готовили стандартный интервал J43 (BioXcell) в 100%-ной мышиной сыворотке (Sigma) от 1000 до 0,488 нг/мл посредством 2-кратных последовательных разведений. Каждый стандарт затем дополнительно разводили в 2 раза в блокирующем растворе (конечная концентрация J43 от 500 до 0,244 нг/мл) для того, чтобы иметь конечную концентрацию сыворотки 50%. На планшет в двойной повторности добавляли сто мкл/лунку каждого стандарта. Образцы сыворотки разводили по меньшей мере в два раза в блокирующем буфере, при необходимости, дополнительно разводили в смеси блокирующий буфер/сыворотка (1:1, об./об.) и добавляли на планшеты 100 мкл. Планшеты инкубировали 2 ч при 37°С. После 3 промывок промывочным буфером добавляли 100 мкл/лунку конъюгированного с HRP антитела козы против IgG армянского хомяка (Jackson Immunoresearch) в концентрации 80 нг/мл, и планшеты инкубировали 1 ч при 37°С. После 3 промывок добавляли 100 мкл/лунку 3,3',5,5'-тетраметилбензидина (ТМВ, Sigma), и планшеты инкубировали при RT в течение 30 мин. Реакцию останавливали 100 мкл/лунку 2 М H2SO4, и поглощение измеряли при 450 нм с использованием планшет-ридера (TECAN Infinite М200 PRO). Полученные значения ОП переносились в программу GraphPadPrism, и осуществлялся обратный расчет концентраций в образцах с использованием стандартной кривой, аппроксимированной с использованием 5 параметров.
Как показано на Фиг. 6А, при подкожной инъекции наивным мышам (без имплантированных опухолей), продуцирование рекомбинантного J43 из WRTG18618 было очень слабым (9 нг/мл в сутки 1 и 4,5 нг/мл в сутки 5). В отличие от этого, в сыворотке мышей, несущих опухоли B16F10, секретировались большие количества рекомбинантного J43 после внутриопухолевой инъекции WRTG18618, в 9 и 1900 раз большие в D1 и D5 (83 нг/мл и 9500 нг/мл) соответственно, чем концентрация рекомбинантного J43, измеренная после подкожной инъекции вируса. Данный результат свидетельствует о том, что онколитический вирус предпочтительно размножается в опухолях, а не в подкожных здоровых тканях. По сравнению с этим, сывороточные концентрации, обнаруженные после внутриопухолевой инъекции 10 мкг имеющегося в продаже J43 достигали примерно 1000 нг/мл (1306 нг/мл в D1 и 1049 нг/мл в D5), тогда как сывороточные концентрации были значительно ниже с использованием 1 мкг имеющегося в продаже J43 (73 нг/мл в D1 и 35 нг/мл в D5).
В гомогенатах опухоли продукция рекомбинантного J43 следовала такой же тенденции, что и в сыворотке, с пиковой концентрацией в D5 (см. Фиг. 6Б). Фармакокинетика J43 после одной в.о. инъекции имеющегося в продаже mAb или WRTG18618 является очень разной. После в.о. инъекции 10 мкг имеющегося в продаже J43 максимальная концентрация антитела измерялась в сыворотке и в опухоли в D1 (1306 нг/мл и 173 нг/мл соответственно), как и ожидалось, тогда как в.о. инъекция WRTG18618 приводила к максимальным концентрациям в сыворотке и в опухоли в D5 (9500 нг/мл и 3800 нг/мл соответственно). Следует подчеркнуть то, что опухолевые концентрации рекомбинантного J43, продуцированного после в.о. инъекции вируса, были значительно выше, чем концентрации, измеренные после в.о. инъекции 10 мкг имеющегося в продаже J43 во все моменты времени (410 нг/мл по сравнению со 173 нг/мл в D1; 3800 нг/мл по сравнению с 51 нг/мл в D5 и 700 нг/мл по сравнению с концентрацией меньшей, чем порог выявления, в D11). Важно то, что продуцирование антитела все еще выявлялось в опухолях в D11 после лечения вирусом, что не происходит после инъекции имеющегося в продаже J43.
Эти данные свидетельствуют о том, что векторизация J43 в онколитических вирусах обеспечивает более сильное и более длительное накопление антитела в опухолях по сравнению с в.о. инъекцией антитела, имеющегося в продаже.
Противоопухолевая активность в модели подкожной опухоли
Противоопухолевую активность разных вирусов осповакцины, экспрессирующих антитела против PD-1, описанные выше, можно тестировать в традиционных доклинических моделях после имплантации опухолей, с последующей инъекцией конструкций. Например, мышиные раковые клетки инъецируются подкожно в бока иммунокомпетентных мышей. При достижении опухолями объема 50-70 мм3 мышей рандомизировали вслепую и лечили указанным вирусом осповакцины в дозе 1×107 PFU. Векторы или контрольный носитель (буфер, используемый для ресуспендирования вируса) непосредственно инъецируются в опухоль в сутки 10, 12 и 14 после имплантации опухоли. Размер опухоли измеряется дважды в неделю с использованием циркулей. В частности, векторизированные scFv и антитело mAb1 являются по меньшей мере такими же эффективными, как и совместное введение вектора на основе WR с имеющимся в продаже J43 для задержки роста опухоли и увеличения показателя выживания.
Специалисты в данной области узнают или смогут установить с использованием не более чем традиционного экспериментирования многочисленные эквиваленты конкретного способа и реактивов, описанных в данном документе, включая альтернативы, варианты, присоединения, делеции, модификации и замены. Такие эквиваленты считаются находящимися в пределах объема данного изобретения и покрываются следующей формулой изобретения.


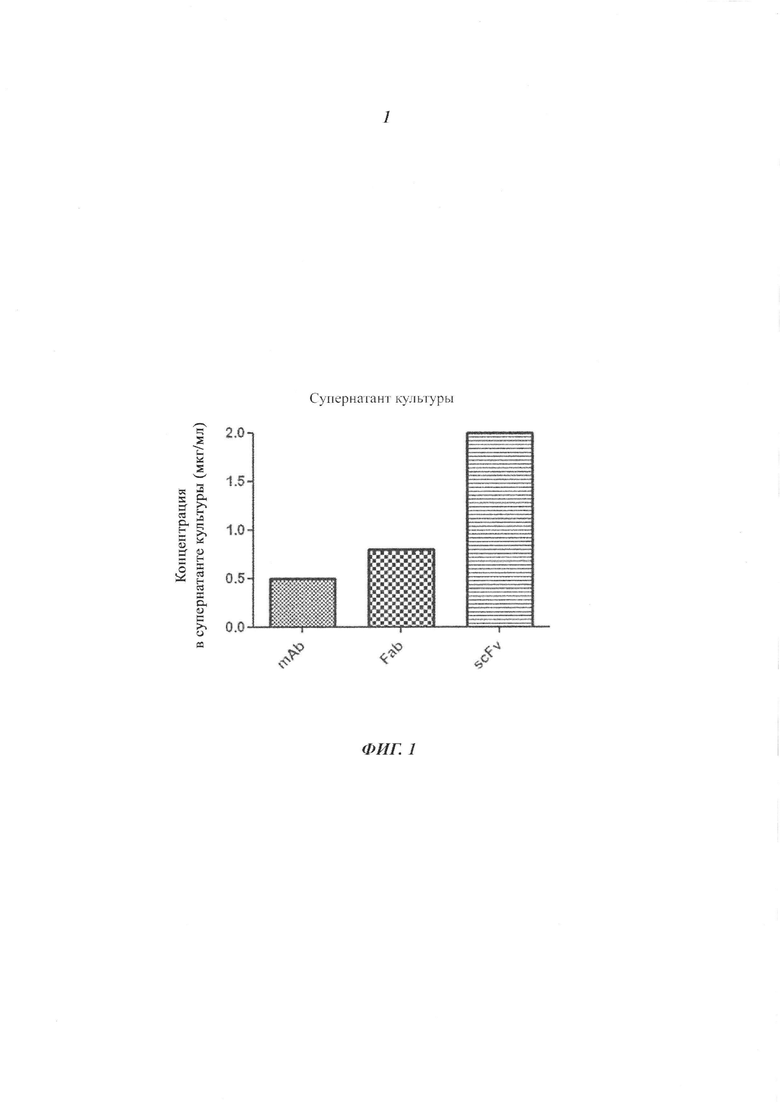

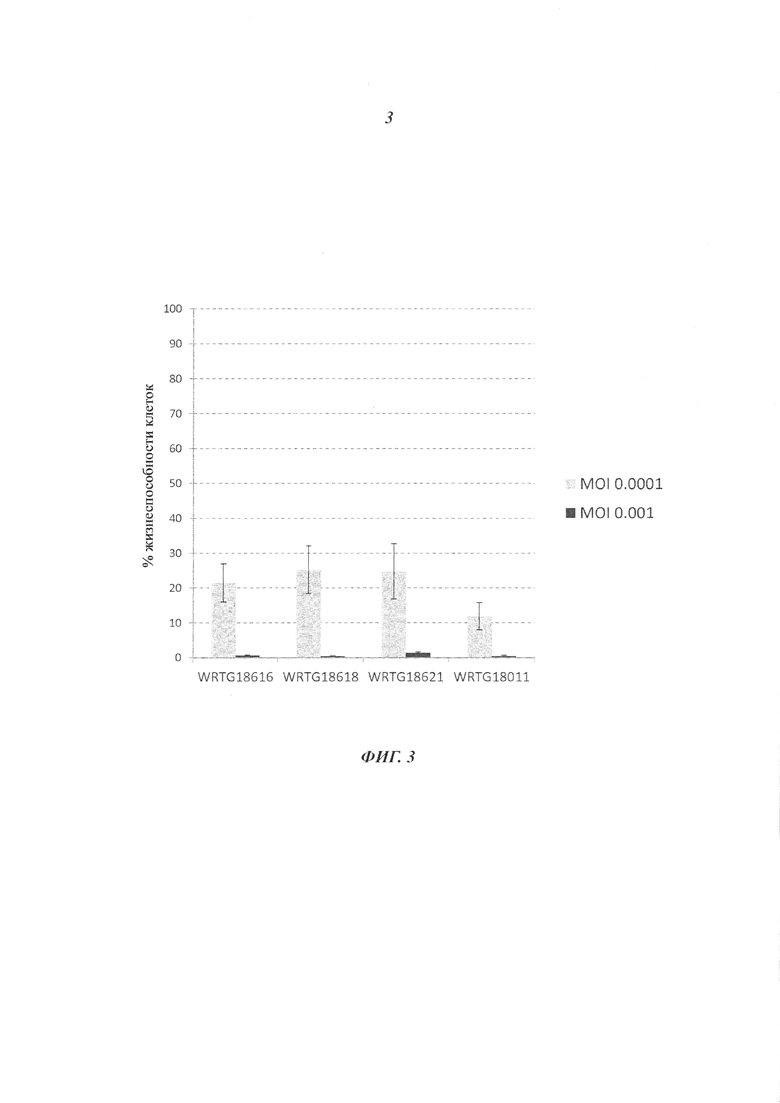
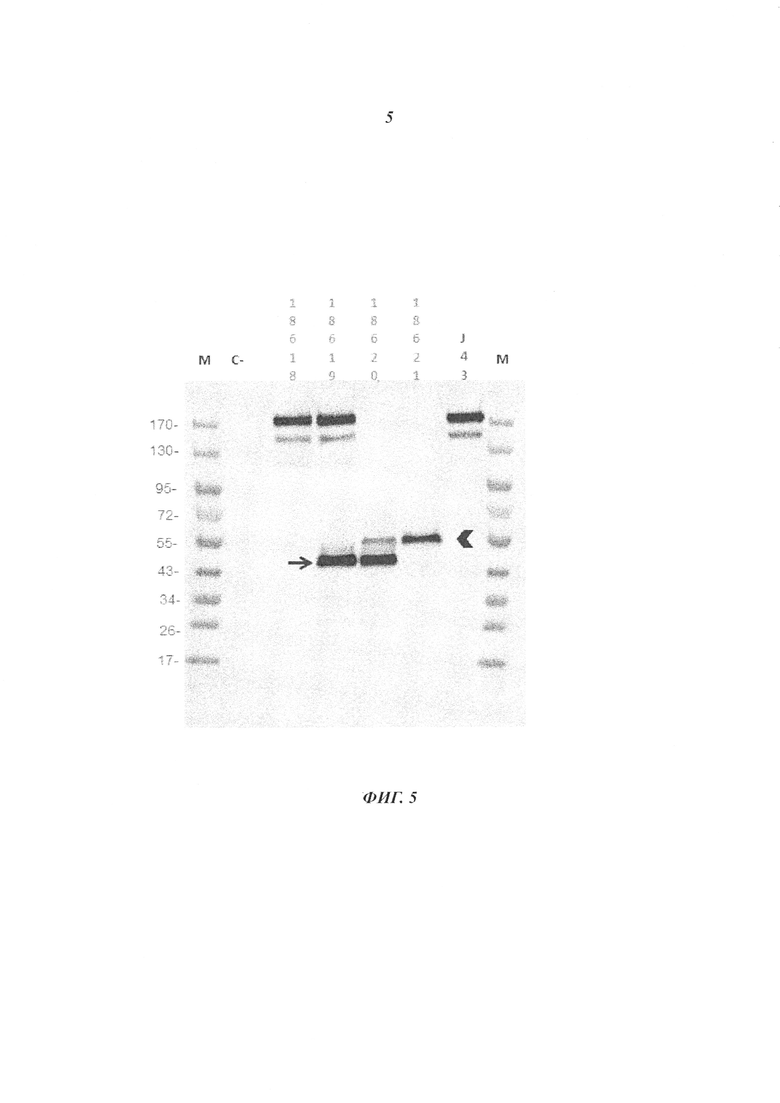
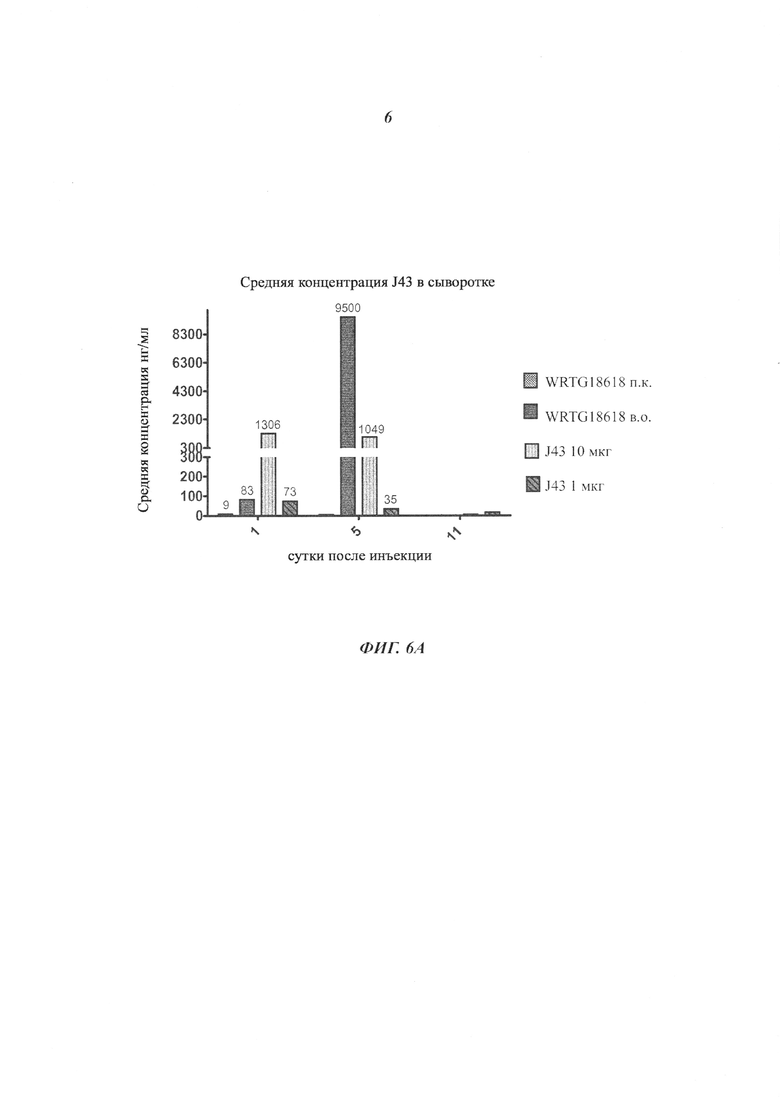
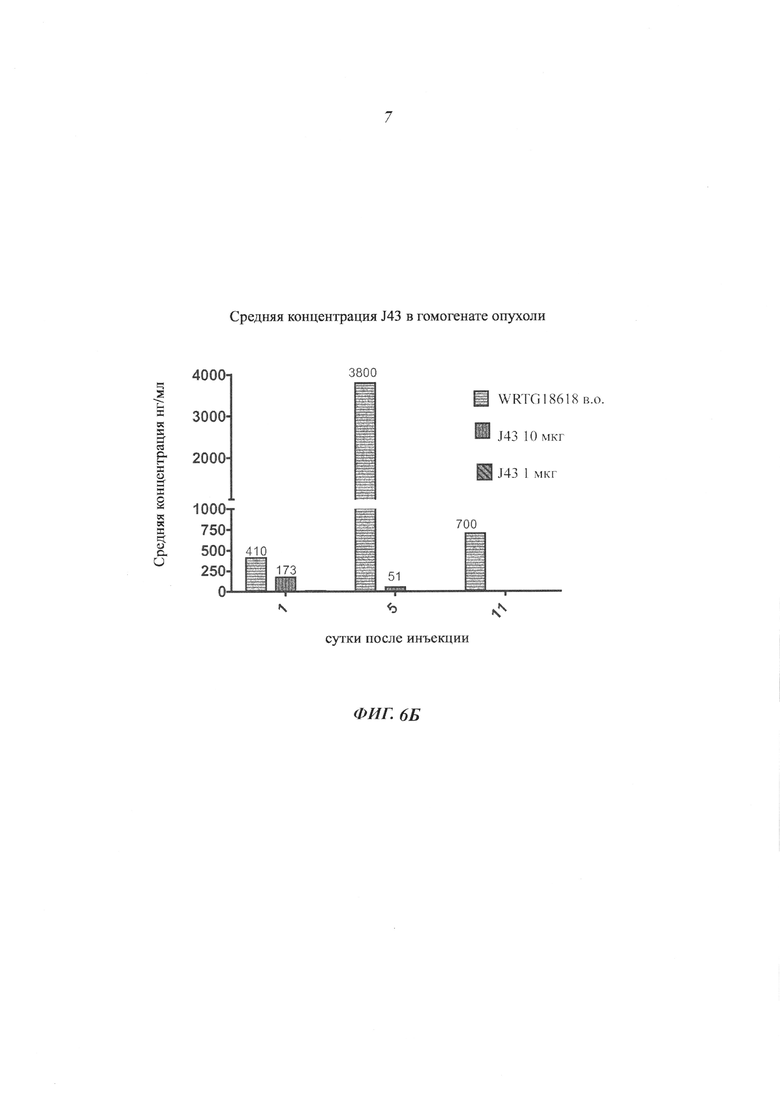
Изобретение относится к биотехнологии. Описан онколитический вирус, содержащий молекулу нуклеиновой кислоты, встроенную в его геном, кодирующую один или более чем один модулятор иммунологической контрольной точки. Указанный онколитический вирус представляет собой онколитический вирус осповакцины, дефектный по тимидинкиназе (TK) в результате инактивирующих мутаций в вирусном гене J2R и дефектный по активности рибонуклеотидредуктазы (RR) в результате инактивирующих мутаций в вирусном гене I4L. При этом указанный один или более чем один модулятор иммунологической контрольной точки включает антитело к PD-1. Описана противоопухолевая фармацевтическая композиция, содержащая данный вирус. Изобретение может быть применено для лечения пролиферативных заболеваний, таких как раковые заболевания. 4 н. и 20 з.п. ф-лы, 6 ил., 1 пр.
1. Онколитический вирус, содержащий молекулу нуклеиновой кислоты, встроенную в его геном, кодирующую один или более чем один модулятор иммунологической контрольной точки, где указанный онколитический вирус представляет собой онколитический вирус осповакцины, дефектный по тимидинкиназе (TK) в результате инактивирующих мутаций в вирусном гене J2R и дефектный по активности рибонуклеотидредуктазы (RR) в результате инактивирующих мутаций в вирусном гене I4L и где указанный один или более чем один модулятор иммунологической контрольной точки включает антитело к PD-1.
2. Онколитический вирус по п. 1, где указанный вирус осповакцины выбран из группы штаммов Elstree, Wyeth, Copenhagen и Western Reserve.
3. Онколитический вирус по п. 1, где указанный онколитический вирус дополнительно содержит по меньшей мере один терапевтический ген, встроенный в вирусный геном, указанный терапевтический ген выбран из группы, состоящей из генов, кодирующих продукты суицидных генов, и генов, кодирующих иммуностимулирующие белки.
4. Онколитический вирус по п. 3, где указанный суицидный ген выбран из группы, состоящей из генов, кодирующих белок, имеющий активность цитозиндезаминазы (ЦДаза), активность тимидинкиназы, активность урацилфосфорибозилтрансферазы (УФРТаза), активность фосфорилазы пуриновых нуклеозидов и активность тимидилаткиназы.
5. Онколитический вирус по п. 4, где указанный продукт суицидного гена имеет ЦДазную и УФРТазную активности, и предпочтительно выбран из группы, состоящей из полипептидов codA::upp, FCY1::FUR1 и FCY1::FUR1[дельта] 105 (FCU1) и FCU1-8.
6. Онколитический вирус по п. 4, где указанный онколитический вирус осповакцины содержит терапевтический суицидный ген FCU1, встроенный в его геном.
7. Онколитический вирус по п. 3, где указанный иммуностимулирующий белок представляет собой интерлейкин или колониестимулирующий фактор, с конкретным предпочтением в отношении GM-CSF (гранулоцитарно- макрофагальный колониестимулирующий фактор).
8. Онколитический вирус по п. 7, где указанный онколитический вирус осповакцины содержит встроенный в его геном терапевтический ген, кодирующий человеческий GM-CSF.
9. Онколитический вирус по п. 1, где указанный один или более чем один модулятор иммунологической контрольной точки включает антитело, которое специфично связывается с человеческим PD-1, в частности антитело против PD- 1, выбранное из группы, состоящей из ниволумаба, пембролизумаба и пидилизумаба.
10. Онколитический вирус по п. 1, где указанный один или более чем один модулятор иммунологической контрольной точки дополнительно включает антитело, которое специфично связывается с человеческим PD-L1, в частности антитело против PD-L1, выбранное из группы, состоящей из MPDL3280A и BMS- 936559.
11. Онколитический вирус по п. 1, где указанный один или более чем один модулятор иммунологической контрольной точки дополнительно включает антитело, которое специфично связывается с человеческим CTLA-4, в частности антитело против CTLA4, выбранное из группы, состоящей из ипилимумаба, тремелимумаба и одноцепочечных антител против CTLA4.
12. Противоопухолевая фармацевтическая композиция, содержащая эффективное количество онколитического вируса по любому из пп. 1-11 и фармацевтически приемлемый носитель.
13. Фармацевтическая композиция по п. 12, содержащая от 107 pfu до 5x109 pfu указанного онколитического вируса.
14. Фармацевтическая композиция по п. 12, которая приготовлена для парентерального введения.
15. Применение онколитического вируса по любому из пп. 1-11 или фармацевтической композиции по любому из пп. 12-14 для изготовления лекарства для лечения пролиферативного заболевания.
16. Применение по п. 15, где указанное пролиферативное заболевание представляет собой рак и предпочтительно рак, выбранный из группы, состоящей из меланомы, рака почки, рака предстательной железы, рака молочной железы, колоректального рака, рака легкого и рака печени.
17. Применение по п. 15, где указанный онколитический вирус вводится посредством внутривенного или внутриопухолевого пути.
18. Применение по п. 17, которое включает от 2 до 5 внутривенных или внутриопухолевых введений 108 или 109 pfu онколитического вируса осповакцины с интервалом 1 или 2 недели.
19. Применение по п. 15, которое дополнительно включает введение пролекарства и/или вещества, эффективного в противораковой терапии.
20. Применение онколитического вируса по любому из пп. 1-11 или фармацевтической композиции по любому из пп. 12-14 для лечения пролиферативного заболевания.
21. Применение по п. 20, где указанное пролиферативное заболевание представляет собой рак и предпочтительно рак, выбранный из группы, состоящей из меланомы, рака почки, рака предстательной железы, рака молочной железы, колоректального рака, рака легкого и рака печени.
22. Применение по п. 20, где указанный онколитический вирус вводится посредством внутривенного или внутриопухолевого пути.
23. Применение по п. 22, которое включает от 2 до 5 внутривенных или внутриопухолевых введений 108 или 109 pfu онколитического вируса осповакцины с интервалом 1 или 2 недели.
24. Применение по п. 20, которое дополнительно включает введение пролекарства и/или вещества, эффективного в противораковой терапии.
| US 7858746 B2, 28.12.2010 | |||
| WO 2014047350 A1, 27.03.2014 | |||
| DIAS et al | |||
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |
| Автоматическая сцепка для железнодорожного состава | 1928 |
|
SU13615A1 |
| КОЧНЕВА Г.В | |||
| и др."Онколитические поксвирусы", Журнал Молекулярная генетика, микробиология и вирусология, N1, 2012. | |||

