Изобретение относится к области молекулярной биологии и может быть использовано для выявления в генах человека мутаций сдвига рамки трансляции и нонсенс-мутаций, ответственных за развитие ряда онкологических заболеваний.
На сегодняшний день хорошо известно, что частой причиной возникновения семейных, то есть наследуемых, форм онкозаболеваний являются мутации в генах, кодирующих онкосупрессоры. В гене BRCA1 описано более 800, а в гене BRCA2 более 700 мутаций, связанных с формированием наследственных форм рака молочной железы и рака яичников [BIC Database, http://research.nhgri.nih.gov/projects/bic/index.shtml].
Кроме того, в этих генах описано значительное количество полиморфных вариантов аллелей, не связанных с онкологией. Этот факт затрудняет выявление значимых мутаций при использовании различных вариантов гетеродуплексного анализа, так как эти методы выявляют любые изменения в последовательности ДНК, что требует дополнительной идентификации методом прямого секвенирования.
Однако, большая часть (около 90%) онкологически значимых мутаций в генах BRCA11 BRCA2 явлются мутациями сдвига рамки трансляции и нонсенс-мутациями, которые приводят к преждевременной терминации трансляции. В результате образуются более короткие продукты, не способные выполнять функцию онкосупрессоров. Для выявления таких мутаций используют метод анализа укороченных белков (Protein Truncation Test, PTT), основанный на анализе продуктов трансляции в бесклеточной системе in vitro. Наличие коротких продуктов оценивается визуально после электрофоретического разделения белков с последующим проведением авторадиографии или Вестерн блота [Den Dunnen J., Van Ommen G. The protein truncation test: A review. Hum Mutat (1999) 14: 95-102]. В целом, метод анализа укороченных белков - дорогая, трудоемкая и недостаточно хорошо воспроизводимая процедура, результаты ее часто интерпретируются неоднозначно.
Известен способ выявления нонсенс-мутаций и мутаций сдвига рамки, который заключается в клонировании амплифицированных фрагментов гена в составе плазмиды в единой трансляционной рамке с геном-репортером с последующей трансформацией дрожжевых клеток, так называемый "yeast based stop-codon assay" [Патент США 6682885, МКИ C12Q 1/68, 2004]. Целостность рамки слитого белка оценивается по активности белка, кодируемого геном-репортером. Отсутствие окраски колоний клеток-трансформантов на индикаторных чашках свидетельствует о наличии мутации в исследуемом фрагменте ДНК. Недостатком этого подхода является сложность работы с линиями дрожжевых клеток: клетки долго растут, метод требует специальных сред для культивирования клеток, нередко возникают проблемы с трансформацией.
Работа с бактериальной системой вектор-хозяин существенно проще. Наиболее близкий к заявленному метод детекции мутаций, нарушающих целостность рамки считывания, заключается в конструировании бактериальных плазмид с последующей трансформацией клеток Escherichia coli [Заявка США 20030064365, МКИ C12Q 1/68, С12/Р 19/34]. Амплифицированный фрагмент гена встраивают в плазмидный вектор в единой трансляционной рамке с геном-репортером, кодирующим α-пептид β-галактозидазы E.coli (LacZα). Однако, использование LacZa-содержащих плазмид для детекции нонсенс-мутаций и мутаций сдвига рамки обладает существенным недостатком: эта система часто дает ложно-отрицательные результаты: колонии клеток-трансформантов дают Lac+-фенотип, несмотря
на наличие стоп-кодонов в тестируемой ДНК. Проблема заключается в том, что бактериальная клетка может использовать в качестве сайтов инициации трансляции кодоны эукариотической ДИК, расположенные после кодонов-терминаторов трансляции и находящиеся в единой трансляционной рамке с LacZα, так называемая реинициация трансляции [Gutkina N.I. et al., Limitations on the recombinant plasmid selection by Lac+/Lac- colony phenotype detection. Biopchem. Biophys. Res. Commun. (2002) 298: 37-40; Real S.M. et al.. Development of a premature stop codon-detection method based on a bacterial two-hybrid system. BMC Biotechnology (2006) 6: 38]. Таким образом, детекция мутаций с использованием LacZα-содержащих плазмид возможна только в участках гена, не содержащих сайты реинициации трансляции, что является серьезным ограничением при исследовании таких сложных генов, как BRCA1.
Задачей, на решение которой направлено настоящее изобретение, является разработка способа детекции нонсенс-мутаций и мутаций сдвига рамки, в котором используется бактериальная система вектор-хозяин, но в то же время результаты анализа не зависят от реинициации трансляции с кодонов внутри эукариотической ДНК.
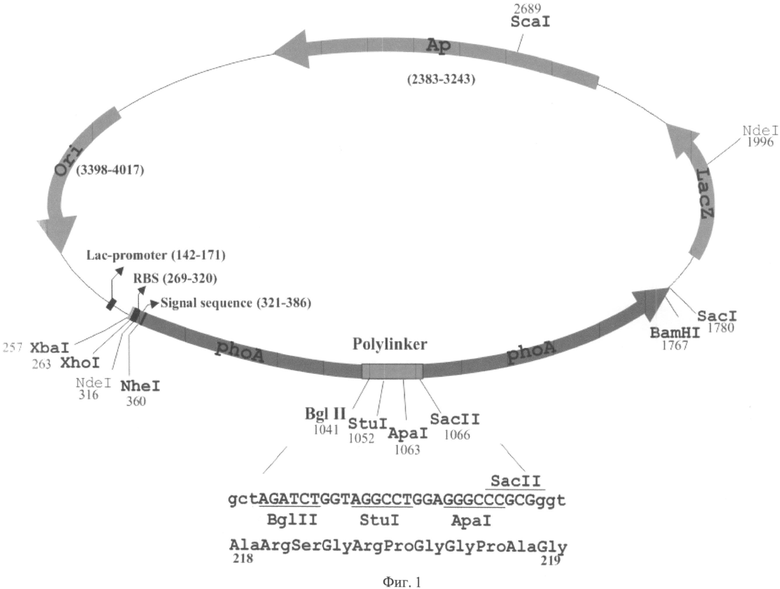
Для решения этой задачи мы сконструировали новый плазмидный вектор размером 4184bp (Фиг.1), предназначенный для клонирования фрагментов гена BRCA1 в единой трансляционной рамке с геном щелочной фосфатазы E.coli (phoA), активность которой определяется при использовании хромогенного субстрата 5-бром-4-хлор-3-индолил-β-D-фосфата (Х-фосфата). Принципиальное отличие заявляемой плазмиды от LacZα-содержащих плазмид заключается в том, что полилинкер встроен между 218 и 219 кодонами гена. phoA, в то время как полилинкер в составе LacZα-содержащих плазмид встроен в ген LacZα после 6-15 кодонов, не имеющих существенного влияния на β-галактозидазную активность репортерного белка. В то же время, при использовании заявляемой плазмиды, белок, синтезированный в случае реинициации трансляции внутри фрагмента BRCA1, не может обладать ферментативной активностью из-за отсутствия N-концевой части, составляющей половину всей последовательности фермента.
Схема заявляемой плазмиды представлена на Фиг.1. Для создания этого вектора resphoA был переклонирован из плазмиды pCGVI [Guzman C.A. et aL, A novel Escherichia coli expression-export vector containing alkaline phosphatase as an insertional inactivadon screening system. Gene (1994) 148: 171-172] в плазмиду pUC19 в два этапа: XhoI-BamHI фрагмент плазмиды pCGVI клонировали в плазмиду pGEM7Zf(+) по сайгам XhoI-BamHI, затем XbaI-SacI фрагмент полученнной плазмиды клонировали в pUC19. Затем внутрь последовательности phoA между 218 и 219 кодонами была встроена последовательность размером 27 п.о., содержащая сайты узнавания эндонуклеаз рестрикции (Фиг.1) BglII (1041), StuI (1052), ApaI (1063), SacII (1066) и предназначенная для клонирования фрагментов гена BRCA1 (полилинкер). В скобках после названия ферментов указана позиция, по которой происходит разрезание.
Район, который был выбран для встройки полилинкера, кодирует участок полипептидной цепи, расположенный на поверхности белка вдали от активного центра и не участвующий в формировании димеров щелочной фосфатазы, необходимых для проявления ферментативной активности.
Таким образом, XbaI-SacI фрагмент плазмиды (Фиг.1) размером 1523 п.о. содержит:
- фрагмент, кодирующий щелочную фосфатазу E.coli (387-1766) (Фиг.1),
- фрагмент гена atpE E.coli, содержащий сайт инициации трансляции (269-320) (Фиг.1),
- встроенную в ген phoA последовательность, содержащую сайты узнавания эндонуклеаз рестрикции BglII(1041), StuI(1052), ApaI(1063), SacII (1066) и предназначенную для клонирования фрагментов renaBRCA1 (полилинкер) (Фиг.1),
фрагмент, кодирующий N-концевую часть сигнальной последовательности гена бета-лактамазы (bla) и С-концевую часть сигнальной последовательности гена phoA, который обеспечивает секрецию щелочной фосфатазы в периплазму бактериальной клетки (321-386 н.) (Фиг.1).
Часть плазмиды, соответствующая последовательности pUC19, содержит следующие функционально значимые элементы: Lac-промотор (142-171 н.), участок начала репликации ori (3398-4017 н.), ген, определяющий устойчивость к ампициллину Ар (2383-3243 н.)
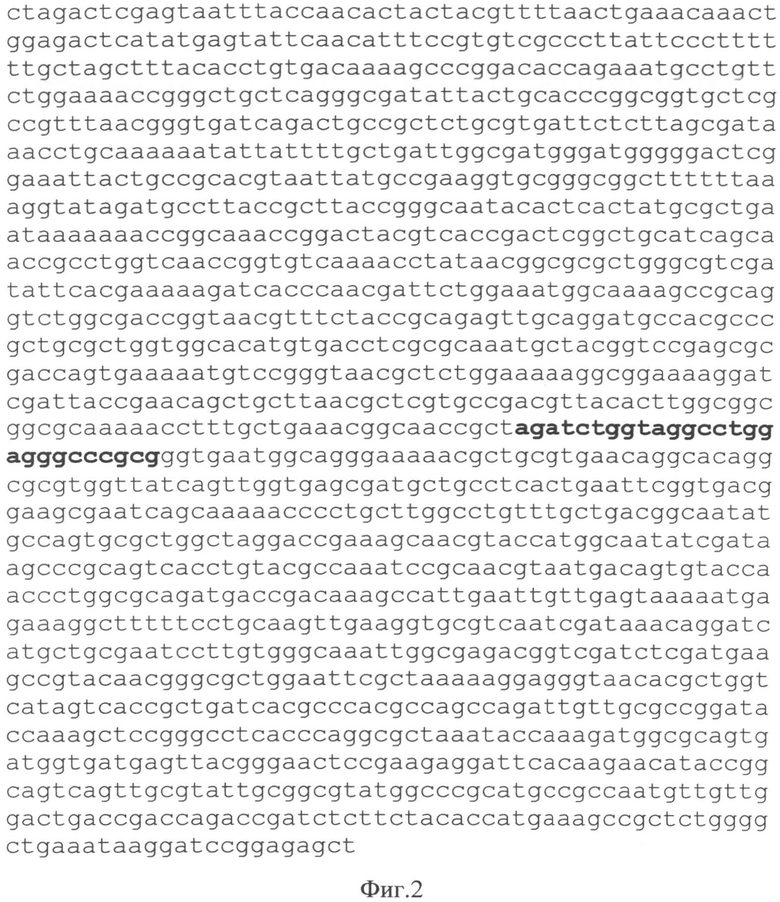
Последовательность XbaI-SacI фрагмента плазмиды представлена на Фиг.2.
Задача изобретения была решена также способом, в котором используют рекомбинантные плазмиды, созданные с применением вышеописанного плазмидного вектора, при этом исследуемый фрагмент гена находится в единой трансляционной рамке с геном щелочной фосфатазы Е.coli, наличие в исследуемом фрагменте гена мутаций, нарушающих целостность рамки считывания, оценивают визуально по отсутствию окраски колоний клеток Е.coli, трансформированных полученной рекомбинантной плазмидой, в присутствии субстрата щелочной фосфатазы.
Предлагаемый способ детекции мутаций состоит из следующих этапов:
1. Амплификация фрагмента гена с использованием в качестве матрицы геномной ДНК пациента.
2. Гидролиз амплификата эндонуклеазами рестрикции. Возможны следующие варианты:
а) использовать уже имеющиеся в амплификате сайты, как это сделано в примере 1.
б)необходимые сайты узнавания рестриктаз вводятся в последовательность праймеров, как это было сделано в примере 2.
3. Гидролиз эндонуклеазами рестрикции плазмидной ДНК.
4. Очистка вектора и клонируемых фрагментов ДНК: продукты гидролиза эндонуклеазами рестрикции подвергаются электрофоретическому разделению в агарозном геле с последующей элюпией из кусочков геля нужных фрагментов.
5. Лигирование исследуемого фрагмента ДНК и плазмидной ДНК.
6. Трансформация лигазной смесью вектор-фрагмент компетентных клеток E.coli (штамм XL-1 Blue) и посев клеточной суспензии на чашки с агаром, содержащие ампициллин, тетрациклин и субстрат для щелочной фосфатазы Х-фосфат. Использование phoA- штаммов не является абсолютно необходимым, так как природный PhoA-промотор клеток-хозяев репрессируется высокой концентрацией неорганического фосфата, в то время как в плазмидном векторе природный PhoA-промотор отсутствует.
7. Визуальная оценка окраски колоний клеток-трансформантов. Синяя окраска клонов свидетельствует об отсутствии в исследуемом фрагменте гена какой-либо мутации, нарушающей целостность рамки считывания. Отсутствие окраски (белые клоны) свидетельствует о наличии в исследуемом фрагменте либо нонсенс-мутации, либо мутации сдвига рамки считывания.
Предлагаемый способ обладает следующими преимуществами:
1. Способ позволяет выявлять любые, в том числе неизвестные, нонсенс-мутации и мутации сдвига рамки считывания.
2. Способ позволяет выявлять только значимые для развития патологии мутации, так как исключается детекция полиморфных вариантов, не приводящих к возникновению стоп-кодонов.
3. Использование для клонирования фрагментов ДНК заявляемой плазмиды позволяет избежать ложноотрицательных результатов, вызванных возможностью реинициации трансляции на участках, расположенных после стоп-кодонов, возникших в результате мутации.
4. Способ хорошо различает разные аллели гена - мутантный и дикого типа, так как каждый клон клеток содержит только один вариант плазмиды, что позволяет однозначно интерпретировать результаты.
5. Способ не требует использования дефицитных по щелочной фосфатазе штаммов клеток, так как фосфатазная активность клетки-хозяина подавляется добавлением в питательную среду неорганического фосфата. При этом, щелочная фосфатаза, кодируемая заявляемой плазмидой, сохраняет активность, что позволяет использовать для трансформации широко-доступные бактериальные штаммы
6. Способ относительно прост и не требует больших материальных затрат.
Изобретение иллюстрируют следующие рисунки:
Фиг.1 Схема заявляемой плазмиды.
Начало нумерации плазмиды соответствует началу нумерации pUC19 (GenBank Accession # M77789). Полужирным шрифтом выделены уникальные сайты узнавания эндонуклеаз рестрикции Фиг.2 Последовательность XbaI-SacI фрагмента, встроенного в pUC19.
Полужирным шрифтом выделена последовательность полилинкера.
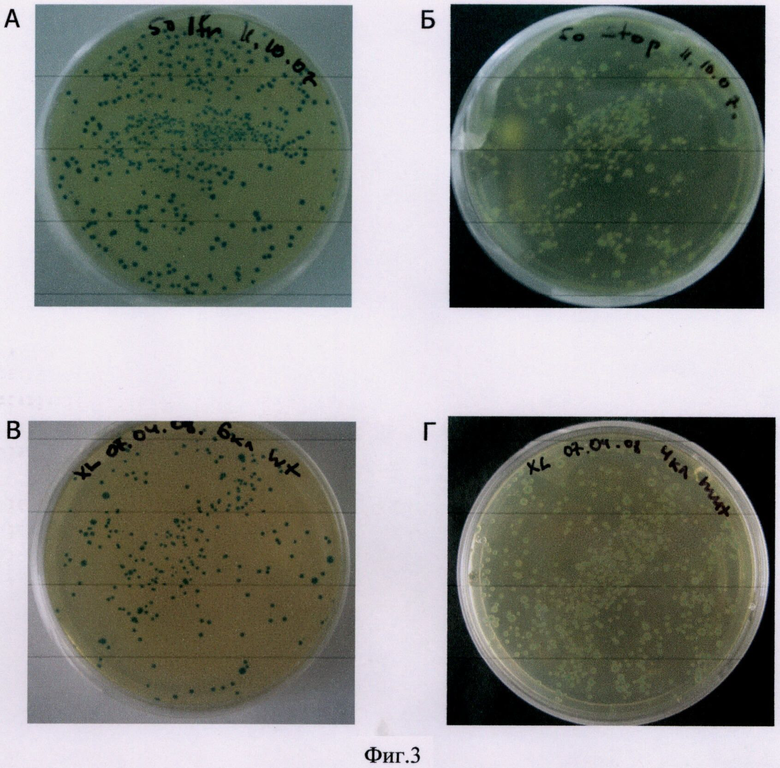
Фиг.3 Окраска колоний клеток E.coli (XL I-Blue), трансформированных различными рекомбинантными плазмидами.
А - плазмида со встройкой фрагмента экзона 11 гена BRCA1 без мутаций.
Б - плазмида со встройкой фрагмента экзона 11 с двумя стоп-кодонами.
В - плазмида со встройкой экзона 20 гена BRCA1 без мутации.
Г - плазмида со встройкой экзона 20 гена BRCA1 с мутацией 5382insC
Изобретение иллюстрируют нижеприведенные примеры.
Пример 1.
Выявление нонсенс-мутации в фрагменте экзона 11 гена BRCA1. Следующие праймеры использовались для амплификации экзона 11 гена BRCA1:
F-праймер: 5'-CTCATGGAAGGTAAAGAACCTGC-3'
R-праймер: 5'-TTTCTTTAAGGACCCAGAGTGG-3'
Реакционная смесь содержала 16 мМ (NH4)2SO4, 67 мМ Tris-HCl (pH 8.8), 0.01% Twin 20, 4 мМ MgCl2, 300 мкМ каждого dNTP, 0.5 мкМ каждого праймера, 50 нг геномной ДНК и 1 ед. Taq ДНК полимеразы. Условия термоциклирования: денатурация 94°С - 2 мин, затем 35 циклов (94°С - 20 сек, 60°С - 30 сек, 72°С - 60 сек). В качестве матрицы использовали геномную ДНК, не содержащую мутаций в экзоне 11 гена BRCA1. Продукт амплификации гидролизовали эндонуклеазами рестрикции BglII и HpaI. После электрофоретического разделения продуктов гидролиза BglII-HpaI фрагмент экзона 11 размером 227 п.н. элюировали из геля и лигировали с плазмидной ДНК, гидролизованной BglII и PceI (изошизомер StuI). Полученная рекомбинантная плазмида содержит фрагмент экзона 11 в единой трансляционной рамке с геном щелочной фосфатазы. Компетентные клетки (Е.coli, штамм XL1-Blue) готовили стандартным методом с использованием CaCl2. Компетентные клетки инкубировали с лигазной смесью (вектор+фрагмент) и высевали на чашки с 1.5% агаром, содержащим 50 мкг/мл ампициллина, 12,5 мкг/мл тетрациклина, 30 мМ Na-фосфатный буфер (рН 7,5) и 160 мкг/мл Х-фосфата. Колонии клеток, трансформированных полученной рекомбинантной плазмидой, были интенсивно окрашены, что свидетельствует об отсутствии мутаций, нарушающих целостность рамки считывания (фиг.3А).
Для того, чтобы продемонстрировать возможность детекции нонсенс-мутаций, BglII-KpnI фрагмент экзона 11 был заменен на химически синтезированный фрагмент с однонуклеотидными заменами, которые приводят к появлению стоп-кодонов внутри этого фрагмента. Нарушение целостности рамки трансляции привело к полному отсутствию окраски колоний клеток, трансформированных полученной плазмидой (фиг.3Б).
Пример 2.
Выявление мутации 5382insC (мутации сдвига рамки трансляции) в экзоне 20 гена BRCA1. В этом примере фрагмент ДНК, содержащий экзон 20, получали, используя в качестве матрицы геномную ДНК пациента с диагнозом рак молочной железы, для которого ранее было доказано, что он является носителем очень распространенной у таких больных мутации 5382insC. Следующие праймеры использовались для амплификации экзона 20 гека BRCA1:
F-праймер: 5'-AATCGCCTAGATCTCAGCATGATTTTGAAGTCAGAGG-3'
R-праймер: 5'-TTAATTAAGGGCCCAGGAGGGAGCTTTACCTTTCTG-3'
Полужирным шрифтом выделена комплементарная часть праймеров. На 5'-конце добавлена последовательность, которая содержит сайты узнавания эндонуклеаз рестрикции (подчеркнуты) BglII (F-праймер) и Apal (R-праймер). Реакционная смесь содержала 16 мМ (NH4)2SO4, 67 мМ Tris-HCl (рН 8.8), 0.01% Twin 20, 4 мМ MgCl2, 300 мкМ каждого dNTP, 0.5 мкМ каждого праймера, 50 нг геномной ДИК и 1 ед. Taq ДНК полимеразы. Условия термоциклирования: денатурация 94°С - 2 мин, затем 35 циклов (94°С - 20 сек, 60°С - 30 сек, 72°С - 60 сек). Продукт амплификации после электрофоретической очистки гидролизовали BglII и ApaI и лигировали с плазмидой, линеаризованной BglII и ApaI. Полученная рекомбинантная плазмида содержит экзон 20 в единой трансляционной рамке с геном щелочной фосфатазы. Были получены 2 варианта рекомбинантных плазмид: а) встройка содержит экзон 20 без мутации, б) встройка содержит экзон 20 с мутацией 5382insC. В первом случае, окраска колоний клеток-трансформантов была ярко-синей, во втором случае, клоны были белые (фиг.3В и 3Г, соответственно).
Таким образом, наличие в клонированных фрагментах гена BRCA1 мутации сдвига рамки или нонсенс-мутации, нарушающих целостность рамки считывания, приводит к строгому подавлению фосфатазной активности, что позволяет однозначно различить аллель дикого типа и мутантный аллель.
Данный способ может быть применен для детекции мутаций и в других сложных генах, содержащих большое количество мутаций, приводящих к преждевременной терминации трансляции.
Настоящее изобретение относится к области молекулярной биологии. Предложен способ выявления в гене BRCA1 мутаций сдвига рамки считывания и нонсенс-мутаций, заключающийся в создании рекомбинантных плазмид, в которых амплифицированный фрагмент гена находится в единой трансляционной рамке с геном щелочной фосфатазы Е.соli (phoA). Сконструирован плазмидный вектор pPhoA-frame, который содержит последовательность ДНК, кодирующую щелочную фосфатазу E.coli. Внутрь этой последовательности встроен фрагмент ДНК, содержащий сайты узнавания эндонуклеаз рестрикции BglII, StuI, ApaI, SacII и предназначенный для клонирования фрагментов гена BRCA1 (полилинкер). Амплифицированный фрагмент гена BRCA1 встраивается в плазмидный вектор pPhoA-frame в единой трансляционной рамке с phoA. Наличие в исследуемом фрагменте гена мутаций, нарушающих целостность рамки считывания, оценивается визуально по отсутствию окраски колоний клеток E.coli, трансформированных полученной рекомбинантной плазмидой, на индикаторной чашке, содержащей субстрат для щелочной фосфатазы. Предлагаемый способ позволяет выявлять только значимые для развития патологии мутации, так как исключается детекция полиморфных вариантов, не приводящих к появлению стоп-кодонов и в большинстве случаев не оказывающих существенного влияния на функцию белка. Метод позволяет выявить наличие любых, в том числе и неизвестных, мутаций, нарушающих целостность рамки. Способ может быть использован для выявления в генах человека мутаций сдвига рамки трансляции и нонсенс-мутаций, ответственных за развитие ряда онкологических заболеваний. 2 н.п. ф-лы, 3 ил., 2 пр.
1. Плазмидный вектор размером 4184 п.н., предназначенный для клонирования фрагментов кодирующей части гена BRCA1 с целью выявления в них нонсенс-мутаций и мутаций сдвига рамки считывания, включающий фрагмент размером 1523 п.н., встроенный в плазмиду pUC19 по сайтам XbaI и SacI, и содержащий следующие функционально значимые элементы:
фрагмент, кодирующий щелочную фосфатазу E.coli;
фрагмент гена atpE E.coli, содержащий сайт инициации трансляции; и
встроенную в ген phoA (между 218 и 219 кодонами) последовательность, содержащую сайты узнавания эндонуклеаз рестрикции BglII (1041), StuI (1052), ApaI (1063), SacII (1066) и предназначенную для клонирования фрагментов гена BRCA1 (полилинкер);
где остальные структурные элементы и их взаиморасположение указаны на фиг.1.
2. Способ выявления в гене BRCA1 человека нонсенс-мутаций и мутаций сдвига рамки трансляции, включающий создание рекомбинантных плазмид, отличающийся тем, что используют плазмиды, созданные с использованием плазмидного вектора по 1, при этом исследуемый фрагмент гена, размер которого не превышает 227 п.н., встраивают в полилинкер, расположены между 218 и 219 кодонами гена phoA, в единой трансляционной рамке с геном щелочной фосфатазы E.coli, а фрагменты кодирующей части гена представляют собой амплификаты экзонов, большая часть которых (21 из 22) не превышает 200 п.н., либо фрагментов большого экзона, и способ детекции мутаций состоит из следующих этапов:
1) амплификация фрагмента гена с использованием в качестве матрицы геномной ДНК пациента, при этом необходимые сайты узнавания рестриктаз вводят в последовательность праймеров,
2) гидролиз амплификата и плазмидной ДНК эндонуклеазами рестрикции,
3)лигирование исследуемого фрагмента ДНК и плазмидной ДНК,
4) трансформация лигазной смесью вектор-фрагмент компетентных клеток E.coli (штамм XL-1 Blue) и посев клеточной суспензии на индикаторные чашки, при этом фосфатазная активность клетки-хозяина подавляется добавлением в питательную среду неорганического фосфата, в то время как щелочная фосфатаза, кодируемая плазмидой, сохраняет активность,
5) визуальная оценка окраски колоний клеток-трансформантов: синяя окраска клонов свидетельствует об отсутствии в исследуемом фрагменте гена какой-либо мутации, нарушающей целостность рамки считывания, а отсутствие окраски (белые клоны) свидетельствует о наличии в исследуемом фрагменте либо нонсенс-мутации, либо мутации сдвига рамки считывания.
| WO 200206527 А2, 24.01.2002 | |||
| GUZMAN C.A | |||
| et al | |||
| A novel Escherichia coli expression-export vector containing alkaline phosphatase as an insertional inactivation screening system | |||
| Gene | |||
| Прибор для охлаждения жидкостей в зимнее время | 1921 |
|
SU1994A1 |
| ВЕКТОР РМТ440 С ПОЗИТИВНОЙ СЕЛЕКЦИЕЙ ДЛЯ КЛОНИРОВАНИЯ ФРАГМЕНТОВ ЧУЖЕРОДНОЙ ДНК | 1996 |
|
RU2105064C1 |

