Изобретение относится к медицине, паразитологии и молекулярной генетике, в частности к диагностике гельминтозов человека и животных, к генотипированию и видовой идентификации на разных стадиях жизненного цикла возбудителей шистосомного или церкариального дерматита человека - четырех европейских видов кровяных сосальщиков из рода Trichobilharzia (класс Tremanoda, отряд Strigeata, семейство Schistosomatidae), и диагностическим тестам и к наборам для них.
Трематоды семейства Schistosomatidae имеют важное практическое значение в медицине и ветеринарии. Во взрослом состоянии они паразитируют в кровеносной системе человека, некоторых млекопитающих и птиц, а на личиночных стадиях - в пресноводных моллюсках разных таксономических групп. Церкарии шистосом, выходя из зараженных моллюсков, короткое время живут в воде в свободном состоянии и для продолжения развития и размножения обязательно должны проникнуть в окончательного хозяина, в кровеносной системе которого формируются раздельнополые мариты.
Наибольшую опасность для человека представляют виды, вызывающие шистосомозы и шистосомные дерматиты. Возбудителями шистосомозов человека считают пять из девяти известных видов рода Schistosoma - S. mansoni, S. haematobium, S. japonicum, S. intercalatum, S. mekongi. Они распространены более чем в 70 странах мира, преимущественно в поясе с жарким климатом, и вызывают различные формы шистосомозов: кишечный, мочеполовой, японский, интеркалатный и меконговый (Яроцкий Л.С. Шистосомозы. М.: Медицина, 1982. С.279).
Представители другого рода шистосом, а именно рода Trichobilharzia, являются возбудителями шистосомозов домашних и диких водоплавающих птиц, в первую очередь уток и гусей. Известно, что паразитирование шистосом у птиц приводит к функциональным нарушениям и их гибели (Беэр С.А., Воронин М.В. Церкариозы в урбанизированных экосистемах. М.: Наука. 2007. С.240).
Шистосомы птиц, личинки которых проникают в кожу человека при купании в естественных водоемах, являются причиной церкариозов. Согласно принятой в Российской федерации «Международной статистической классификации болезней (МКБ-10)» заболевание называется «церкариальный дерматит или чесотка купальщиков». Синонимами церкариального дерматита служат употребляемые в просторечье другие названия: «зуд купальщиков», «зуд пловцов», «водный зуд», «церкариоз», а в зарубежных источниках чаще встречается термин «шистосоматидный дерматит». Церкариальные дерматиты у людей в настоящее время зарегистрированы почти во всех странах Европы, в Америке, Азии и Африке. По самым общим оценкам российских экспертов, более 120 очагов церкариозов насчитывается на европейской и азиатской территории России (Беэр С.А., Воронин М.В. Церкариозы в урбанизированных экосистемах. М.: Наука. 2007. С.240).
Циклы развития всех сосальщиков семейства Schistosomatidae весьма сложные. Развитие шистосом происходит со сменой партеногенетического и раздельнополого поколений, а также сменой окончательного и промежуточного хозяев. Под оболочкой яйца находится вполне развитая личинка - мирацидий. Появившись из яйца в воде, после непродолжительного плавания мирацидий проникает в промежуточного хозяина - моллюска. На этой стадии формируется мешковидная неподвижная спороциста, которая производит дочерние спороцисты, из которых развиваются личиночные формы с вилообразным хвостом - церкарии, или фуркоцеркарии. Для этих личинок характерна пара темно-окрашенных глазков, расположенных в передней части тела, по которым их сразу можно идентифицировать. Церкарии - подвижные личинки, обладающие рядом черт, свойственных взрослым гельминтам. Для продолжения развития они должны проникнуть в организм позвоночного животного, который получил название окончательного или дефинитивного хозяина. Химическим стимулом для прикрепления и внедрения личинок птичьих шистосом в хозяина служат свободные жирные кислоты, находящиеся в верхних слоях кожного покрова позвоночного животного. Именно сходство водорастворимых фракций липидов - церамидов и холестерола, образующихся на коже человека и водоплавающих птиц приводят к ошибочному проникновению личинок в неспецифических хозяев (Гинецинская Т.А. Трематоды - их жизненные циклы, биология и эволюция. Л.: Наука. 1968. С.411).
При описании комплекса клинических проявлений церкариальных дерматитов у людей показано, что интервал от момента внедрения личинок в кожу до появления первых признаков заболевания, по некоторым оценкам, составляет от 30 минут до 18 дней, а средняя продолжительность клинических проявлений заболевания составляет в среднем 5 дней. Например, церкарии вида Т. szidati вызывали кожный зуд, который сохранялся на протяжении 12 дней. При интенсивном зуде и присоединении вторичной инфекции кожные проявления церкариального дерматита были аналогичны клинической картине чесотки (Березанцев Ю. Ф., Курочкин Ю.В. Изучение шистосоматидных церкариозов в СССР. 2. Клиника, патогенез, профилактика церкариозов. Медицинская паразитология, 1966. № 4. С.434-438).
В настоящее время внутри рода Trichobilharzia выделяют более 40 видов паразитов птиц. Из них в Европе известно четыре вида, вызывающих церкариальный дерматит человека: Т. szidati Neuhaus, 1952 (синоним Т. ocellata Brumpt,1931); T.franki Muller and Kimming, 1994; Т. regenti Horák, Kolářová and Dvořák 1998; и Т. salmaticencis Simon-Martin and Simon-Vicente, 1999 (Horak P., Kolarova L. and Adema С.М. Biology of the schistosome genus Trichobilharzia. Adv Parasitol. 2002. V.52. P.155-233). Из них три вида (Т. szidati, Т. franki, Т. salmaticensis) относят к висцеральным формам, т.е. паразитирующим в кровеносных сосудах внутренних органов птицы, и один вид - Т. regenti паразитирует в носовой полости и центральной нервной системе птиц (назальная форма). Наряду с видами Trichobilharzia spp. повсеместно распространен другой вид птичьих шистосом - Bilharziella polonica [Kowalewsky, 1895] Looss, 1899. Как синоним, ранее использовалось и другое название - Pseudobilharziella kowalewskii (Ejsmont, 1929) или Ornithobilharzia polonica (Kowalewsky, 1895) Tanabe, 1925. Схема жизненного цикла этого представителя рода Bilharziella почти ничем не отличается от такового для рода Trichobilharzia. В отличие от трихобильгарций, паразитирующих на прудовиках (сем. Lymnaeidae), партениты В. polonica инфицируют исключительно пресноводных катушек сем. Planorbidae. Считается, что в отдельных случаях церкарии этого полигостального вида наряду с трихобильгарциями могут вызывать "лихорадку купальщика" (Беэр С.А., Воронин М.В. Церкариозы в урбанизированных экосистемах. М.: Наука. 2007. С.240). Так, сформированные шистосомулы наблюдали в легких при экспериментальном заражении мышей церкариями В. polonica (Horak P., Kolarova L. Survival of bird schistosomes in mammalian lungs. Int. J. Parasitol. 2000. V.30. P.65-68).
При экспериментальном заражении показан более высокий патогенный эффект церкарии для млекопитающих в случае назальной формы Trichobilharzia regenti (Horak Р., Kolarova L., Dvorak J. Trichobilharzia regenti n.sp.(Schistosomatidae, Bilharziellinae), a new nasal schistosome from Europe. Parasite 1998: №5. P.349-357. Horak P., Kovar L., Kolarova L. and Nebesarova J. 1998. Horak P., Dvorak J., Kolarova L., Trefil L. Trichobilharzia regenti, a pathogen of the avian and mammalian central nervous system. Parasitology 1999. V.119. P.577-581).
Зрелые самки Т. regenti откладывают яйца в венозных пазухах носа птиц, и через некоторое время яйца с носовой слизью попадают в воду. В отличие от назальных видов (таких как Т. regenti, Т. arcuata, Т. aureliani, Т. nasicola, Т. rodhaini) висцеральные виды птичьих шистосом рода Trichobilharzia (Г. szidati, Т. salmanticensis, Т. franki, Т. alaskensis и другие) откладывают яйца в кровеносных капиллярах внутренних органов окончательного хозяина. Затем яйца попадают в просвет кишечника и с пометом птиц выбрасываются наружу. Horak с соавторами в экспериментах по заражению утят церкариями T.regenti наблюдали симптомы частичного или полного нервного паралича конечностей птиц, потерю ими равновесия и нарушение ориентации в пространстве.
Известные проведенные гистологические исследования выявили дистрофические и некротические изменения в нейронах спинного мозга и в белом веществе головного мозга при последующей миграции шистосомул Т. regenti. На этом основании центральная нервная система млекопитающих и птиц рассматривается как основной путь перемещения шистосомул Т. regenti к месту окончательной локализации - носовым пазухам (Horak P., Dvorak J., Kolarova L., Trefil L. Trichobilharzia regenti, a pathogen of the avian and mammalian central nervous system. Parasitology 1999. V.119. P.577-581).
Недостатком известных способов является отсутствие четких морфологических критериев и гостальной специфичности, поэтому видовая идентификация сосальщиков рода Trichobilharzia стандартными способами затруднительна и в настоящее время возможна только с помощью молекулярно-биологических методов.
Известно маркирование ядерного и митохондриального генома для представителей популяций трех европейских видов, а именно для Т. szidati, Т. franki и Т. regenti, а также для недавно обнаруженного на территории Белоруссии нового вида T.sp.var.narochanica (Хрисанфова Г.Г., Лопаткин А.А., Мищенков В.А. и др. Генетическая изменчивость птичьих шистосом (Класс Trematoda, сем. Schistosomatidae) озера Нарочь: идентификация нового вида в группе Trichobilharzia ocellata. Доклады Академии Наук, 2009. Т.428. №5. С.698-702). Церкарии этого вида птичьих шистосом выявлены с помощью генотипирования возбудителей церкариального дерматита в водоемах Белоруссии.
Известна геномная вариабельность птичьих шистосом рода Trichobilharzia, выявленная при изучении ITS1 рДНК для церкариальных изолятов из Женевского озера (Picard D., Jousson О. Genetic variability among cercariae of the Schistosomatidae (Trematoda: Digenea) causing swimmer's itch in Europe. 2001. Parasite V.8. P.237-242). В этом способе продемонстрирован высокий уровень внутривидовой гетерогенности ITS1 рДНК церкариальных изолятов вида Т. franki.
В известном способе диагностики полиморфные рДНК применяли для дифференциации церкарий трех видов Т. szidati, Т. franki и Т. regenti, обнаруженных на пресноводных моллюсках в Чехии (Dvorak J., Vanacova S., Hampi V., Flegr J., Horak P. Comparison of European Trichobilharzia species based on ITS1 and ITS2 sequences. Parasitology. 2002. V.124. P.307-313). В качестве молекулярного маркера использовали последовательности ITS1 рДНК и участок 5.8S-ITS2 рДНК. Показано, что все три вида отличаются между собой как по длине, так и по структуре ITS1, ITS2 рДНК.
Недостатком известного способа видовой диагностики является то, что в случае контаминации чужеродной ДНК предлагаемые в указанной работе праймеры могут давать дополнительные фрагменты амплификации, затрудняя, таким образом, четкую видовую идентификацию трихобильгарций.
Известен способ видовой диагностики на разных стадиях жизненного цикла гельминтов - возбудителей церкариального дерматита человека - молекулярная тест-система, разработанная для обнаружения в биологических образцах ДНК представителей птичьих шистосом рода Trichobilharzia (Korsunenko, А.V. Chrisanfova G.G., Ryskov A.P., Movsessian S.O., Vasilyev V.A., Semyenova S.K. Detection of European Trichobilharzia Schistosomes (Т. franki, T. szidati, and T. regenti) Based on Novel Genome Sequences. Journal of Parasitology. 2010. V.96, №4, P.802-806). Недостатком известного способа является невозможность видовой идентификации европейских шистосом рода Trichobilharzia.
Следует выбрать прототипом один из наиболее близких прототипов. Наиболее близким к предлагаемому способу видовой диагностики на разных стадиях жизненного цикла гельминтов - возбудителей церкариального дерматита человека являются молекулярные тест-системы, разработанные в настоящее время для обнаружения в биологических образцах только ДНК различных видов возбудителей шистосомоза человека из рода Schistosoma (патент США 6818402, опубл. 16 ноября 2004; или патент США W09705158 (A1), опубл. 13 февраля 1997).
Недостатком известного способа видовой диагностики является невозможность идентификации видов шистосом из рода Trichobilharzia, тем более на разных стадиях жизненного цикла.
Задачей предлагаемого изобретения является создание быстрого, простого и надежного способа для одновременной возможности детекции и видовой идентификации четырех европейских видов птичьих шистосом рода Trichobilharzia на разных стадиях жизненного цикла, включая стадию моллюска, с использованием в качестве генетического маркера консервативной последовательности участка ядерной рибосомной ДНК.
В результате использования предлагаемого способа видовой ДНК-диагностики появляется возможность быстрой детекции и видовой идентификации возбудителей церкариального дерматита на разных стадиях жизненного цикла паразита, что значительно повышает эффективность санитарно-эпидемиологических мероприятий, направленных на выявление природных очагов церкариоза, их мониторинг и разработку методов борьбы с ними.
Вышеуказанный технический результат достигается тем, что в предлагаемом способе видовой ДНК-дифференциации на разных стадиях жизненного цикла гельминтов - возбудителей церкариального дерматита человека, включающем выделение ДНК возбудителя, амплификацию мишени с использованием специфических праймеров, электрофоретическое разделение, генотипирование и видовую дифференциацию возбудителей церкариального дерматита - четырех видов птичьих шистосом из рода Trichobilharzia - кровяных сосальщиков, T. szidati, T. regenti, T.franki и T.sp. var narochanica проводят путем амплификации участков последовательности ядерной рибосомальной ДНК в образцах половозрелых гельминтов или их личиночных стадий, принадлежащих к четырем видам, и/или на рДНК инфицированных шистосомами пресноводных моллюсков из семейства Lymnaeidae с помощью полимеразной цепной реакции и использованием четырех олигонуклеотидных праймеров, затем разделяют указанные амплифицированные последовательности рДНК с помощью электрофореза с получением образцов видоспецифичного электрофоретического паттерна гельминта, при этом на каждой из выделенных ДНК осуществляют в двух реакциях ПЦР с набором из четырех предлагаемых праймеров, а именно F, R1, R2, R3, где в первой реакции используют праймеры F, R1, R2, во второй реакции - праймеры F, R3, при этом используют биологический маркер, содержащий полиморфную ДНК и характеризующий видовую принадлежность четырех европейских видов птичьих шистосом рода Trichobilharzia, и представляющий собой нуклеотидную последовательность из группы 4 праймеров следующего состава
F: 5'-CTTTCCATCTATCACGATGCACT-3'
R1: 5'-ATGATAATGTGCATAACACACC-3'
R2: 5'-GCCGTTTATTTATATGTATGTG-3'
R3: 5'-CAAGCCGTTTATTWATATATAACGG-3',
причем используют набор для диагностики и видовой идентификации возбудителей церкариального дерматита, содержащий биологический маркер, реакционную смесь, состоящую из 60 мМ трис-HCl, 10 мМ сульфата аммония, 0,1% TWEEN 20, по 100 мкМ каждого dNTP, 0,5 мкМ MgCl2, 0,9 единиц Taq-полимеразы и эталонные ДНК для каждого вида трихобильгарций - положительный контроль, а полученные амплификационные продукты визуализируют и дифференцируют путем электрофореза в агарозном геле и после окрашивания гелей бромистым этидием идентифицируют по размеру (длине) выявляемые полиморфные маркеры, где в первой реакции амплификации один фрагмент размером 255 п.н. характерен для вида Т. regenti, один фрагмент длиной 316 п.н. характерен для вида Т. szidati, два фрагмента размером 255 и 316 п.н. характерны для вида Т. sp. var narochanica, а во второй реакции амлификационный фрагмент размером 258 п.н. детектируется только у вида Т. franki.
В предлагаемом способе детекции и видовой дифференциации на разных стадиях жизненного цикла возбудителей церкариального дерматита, включающем сбор образцов марит или образцов моллюсков, предположительно инфицированных шистосомами, выделение ДНК из них, исследование полученной ДНК с помощью двух реакций ПЦР с двумя разными наборами праймеров, визуализацию этих маркерных ДНК с помощью электрофореза в агарозном геле, а затем по результатам двух реакций амплификации определение видовой принадлежности шистосом из рода Trichobilharzia в исследуемых образцах по размеру и числу маркерных ДНК, где на основании состава ПНР-паттерна в реакции I с праймерной системой I идентифицируют три из четырех видов (T. szidati, Т. regenti и Т. sp.var narochanica), а в реакции II с праймерной системой II идентифицируют четвертый вид Т. franki. Размер амплифицированных продуктов определяют с помощью маркера молекулярных масс (100 bp Ladder).
Способ видовой ДНК-дифференциации на разных стадиях жизненного цикла возбудителей церкариального дерматита, в том числе четырех видов рода Trichobilharzia, Т. franki, Т. szidati, Т. regenti, Т. sp.var narochanica Trichobilharzia, Т. franki, Т. szidati, Т. regenti, Т. sp. var narochanica, с применением предлагаемых биологических маркеров и набора осуществляют следующим образом.
Осуществляют геномное типирование гельминтов с использованием метода полимеразной цепной реакции (ПНР). В предлагаемом способе получают молекулярно-генетические характеристики представителей четырех видов рода Trichobilharzia, Т. franki, Т. szidati, Т. regenti, Т. narochanica, находящихся на разных стадиях жизненного цикла, в том числе у взрослых половозрелых особей (марит) и личинок (церкарий и спороцист), паразитирующих на пресноводных моллюсках из семейства Lymnaeidae, являющихся промежуточными хозяевами птичьих шистосом на территории России, Белорусси, Западной и Центральной Европы.
Для этого собирают образцы взрослых шистосом, паразитирующих в кровеносной системе домашних или диких утиных птиц, а также образцы церкарий и/или спороцист шистосом, инфицирующих пресноводных моллюсков из семества Lymnaeidae, выделяют из них ДНК, проводят две реакции амплификации с разными наборами праймеров, визуализируют полученные ПЦР-продукты с помощью электрофоретического разделения амплификатов. На заключительном этапе на основании полученных ПЦР-паттернов определяют видовую принадлежность паразита по наличию в амплифицируемом спектре одного или двух маркерных фрагментов, эталонных для каждого из четырех видов рода Trichobilharzia, Т. franki, Т. szidati, Т. regenti, Т. sp. var narochanica.
Сбор материала и консервация проб.
Взрослых половозрелых гельминтов выделяют в основном из кровеносных сосудов воротной системы печени у утиных птиц. Для выделения назального вида шистосом у зараженной птицы проводят смывы из носовой полости, где могут быть локализованы яйца трематод. Кроме того, при вскрытии птицы анализируют поверхность головного мозга и спинную хорду на наличие паразитов. Для сбора личиночных стадий, церкарий, используют два подхода. Подвижных церкарий собирают пипеткой из воды, в которой инкубировали инфицированного моллюска.
Второй способ состоит в отсечении у моллюска части тела с гепатопанкреасом и последующем отделении препаровальной иглой под микроскопом визуально детектируемых спороцист с содержащимися в них оцеллятными церкариями. При нарушении целостности спороцист церкарии шистосом активно выходят в воду и могут быть собраны в достаточном количестве вышеуказанным способом. Для последующего выделения ДНК могут быть использованы не только отдельные церкарии или несколько личинок, но и кусочки спороцист. Образцы фиксируют в 70% спирте и хранят при +4°С. Для более продолжительного хранения пригодны кельвинаторы, поддерживающие температуру ниже -50°С, а также жидкоазотные сосуды Дьюара.
Выделение ДНК
Для выделения геномной ДНК из церкарий, спороцист или ткани инфицированного моллюска используют набор реагентов QIAamp® DNA Micro Kit (QIAGEN GmbH, Germany). С помощью скальпеля или препаровальных игл под бинокуляром отделяют небольшую часть инфицированного гепатопанкреаса или 1-10 фрагментов спороцист либо с помощью пипетки отбирают 1-100 церкарий и переносят в 1.5 мл пробирки "Эппендорф". Вес образцов не должен превышать 10 мг. К биологическому материалу добавляют по 180 мкл буфера ATL, 20 мкл протеиназы К и перемешивают на вортексе в течение 15 сек. Пробирки инкубируют в термостате при температуре 56°С до полного лизиса биологического материала (1-3 ч). Далее добавляют 200 мкл буфера AL, перемешивают на вортексе в течение 15 с до получения гомогенной раствора. Затем добавляют 200 мкл этанола (96%) и инкубируют 5 мин при комнатной температуре (15-25°С).
Аккуратно переносят каждый лизат на колонки QIAamp MinElute Column, закрывают крышки и центрифугируют 1 мин при 8000 об/мин (6000x g). Колонки дважды перемещают в чистые 2 мл пробирки, промывая каждый раз буфером (500 мкл буфера AW1), и центрифугируют в течение 1 мин при 8000 об/мин (6000x g). Для полного освобождения фильтра колонки от жидкости центрифугируют пробирки в течение 3 мин при 14000 об/мин (20000x g). Колонки с фильтрами помещают в чистые 1.5 мл пробирки и вносят по 20-50 мкл буфера АЕ или дистиллированной воды в центр мембраны, а затем инкубируют при комнатной температуре 5 мин и центрифугируют 1 мин при 14000 об/мин (20000x g). Растворенную в буфере ДНК хранят при -20 °С.
°С.
Для генотипирования взрослых гельминтов используют стандартные наборы для выделения ДНК из тканей, например, набора реагентов DIAtom™ Prep 200 (Москва). Набор реагентов Diatom™ DNA Prep200 основан на использовании лизирующего реагента с гуанидинтиоцианатом, который предназначен для лизиса клеток, солюбилизации клеточного дебриса, а также для денатурации клеточных нуклеаз. В присутствии лизирующего реагента ДНК эффективно сорбируется на NucleoS™-сорбенте, затем легко отмывается от белков и солей спиртовым раствором. ДНК, элюированная из сорбента раствором ЭкстраГен Е™ или деионизованной водой, может напрямую использоваться по назначению. Продолжительность выделения ДНК из тканей составляет ~3 часа. Концентрацию полученных препаратов ДНК определяют на спектрофотометре ThermoSpectronic (USA).
Аналитический электрофорез
Для определения концентрации и качества выделенных образцов используют метод электрофореза в агарозном геле. Гель-электрофорез проводится в горизонтальной камере в 1x трис-боратным буфере (ТВЕ), рН 7,5-7,8 (0,089 М трис-бората, 0,089 М борной кислоты и 0,002 М ЭДТА). При внесении ДНК в карманы геля используется буфер для нанесения (ксиленцианол, бромфеноловый синий и глицерин). После электрофореза гель окрашивают в растворе бромистого этидия в течение 5-10 минут или добавляют бромистый этидий непосредственно в гель. Вхождение ДНК в гель осуществляют при 20V в течение 15-20 минут, далее электрофорез ведут при 60V в течение 2-3 часов. В качестве маркера концентрации используется ДНК фага λ, внесенная в разных количествах, например, 0,5; 1; 2 мкг.
Полимеразная цепная реакция (ПЦР).
1. Проведение ПЦР.
Выделенную ДНК используют в качестве матрицы в двух последовательных или параллельных реакциях амплификации (реакция I и II) со специфичными праймерами, которую проводят в термоциклере «MJ Research» (USA) при температурных и временных режимах, представленных в таблице 1. Реакционная смесь объемом 25 мкл содержит ПЦР-буфер (62 мМ Трис-HCl, 15,4 мМ (NH4)2SO4, 0,01% Tween-20), 5 мМ MgCl2, 0,35 мМ каждого dNTP, 0,5 pmol каждого праймера, 0,6 единиц Taq-полимеразы («Fermentas», Vilnius, Lithuania) и 10-100 нг геномной ДНК. Для предотвращения испарения с поверхности раствора во время реакции в каждую пробирку добавляют 20-25 мкл минерального масла. В таблице 1 приведены температурные и временные режимы ПЦР.
Для проведения ПЦР используют четыре праймера, подобранных к последовательностям ITS2 рДНК T. franki, T. szidati, T. regenti, T. narochanica.
В таблице 2 приведен нуклеотидный состав праймеров. Для каждой из двух реакций используют один и тот же прямой праймер F. Два обратных праймера R1 и R2 используют в реакции I, а в реакцию II входит один обратный праймер R3. В реакции I по присутствию единичных фрагментов длиной 255 п.н. и 316 п.н. дифференцируют два из четырех видов птичьих шистосом из рода Trichobilharzia (Т. szidati, Т. regenti), а по наличию в спектрах одновременно двух фрагментов длиной 255 п.н. и 316 п.н. идентифицируют третий вид - Т. narochanica. В реакции II фрагмент размером 258 п.н. амплифицируется только на ДНК Т. franki, и не амплифицируется на ДНК трех остальных видов.
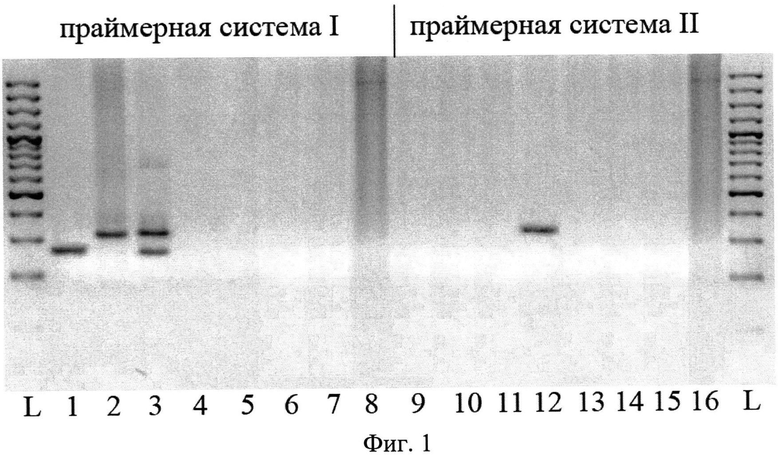
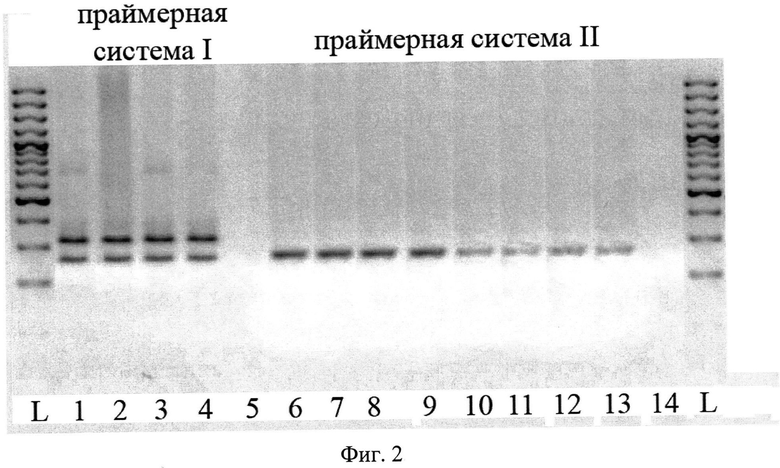
Сущность предлагаемого способа поясняется на фиг.1 и фиг.2.
На фиг.1 представлена электрофореграмма в агарозном геле способа определения видовой принадлежности птичьих шистосом рода Trichobilharzia. Дорожки 1, 9 - T. regenti, дор. 2, 10 - Т. szidati; дор. 3, 11 - T. sp. var. narochanica; дор. 4, 12 - Т. franki. На остальных дорожках представлены амплификационные спектры других видов трематод. На дор. 5, 13 - Bilharziella polonica; дор. 6, 14 - Cotylurus sp. (сем. Strigeidae); дор. 7, 15 - Diplostomum sp. (сем. Diplostomidae); дор. 8. 16 - Bolbophorus sp. (сем. Postodiplostomidae).
На фиг.2 представлена электрофореграмма в агарозном геле способа определения видовой принадлежности птичьих шистосом рода Trichobilharzia на разных стадиях жизненного цикла. Дор. 1 - марита T. sp. var narochanica; дор. 2 - церкарии T. sp. var narochanica; дор. 3 - спороцисты T. sp. var narochanica; дор. 4 - печень моллюска Radix sp, зараженного Т. sp. var narochanica; дор. 5 - печень незараженного моллюска Radix sp.; дор. 6 - марита T. franki; дор. 7 - церкарии Т. franki; дор. 8 - спороцисты Т. franki; дор. 9 - печень моллюска Radix sp, зараженного Т. franki (5γ /мкл); дор. 10, 11, 12, 13 - ДНК Т. franki в разведении 1γ/мкл: 0.5γ/мкл: 0.25γ/мкл: 0.05γ/мкл; дор. 14 - печень незараженного моллюска Radix sp.
Из образцов птичьих шистосом четырех видов рода Trichobilharzia выделяют ДНК способами, указанными выше. На каждой из выделенных ДНК осуществляют ПЦР в два этапа с набором из четырех предлагаемых праймеров, а именно F, R1, R2 (реакция I) и F, R3 (реакция II). Полученные амплификационные продукты разделяют путем электрофореза в агарозном геле и после окрашивания геля бромистым этидием визуализируют и идентифицируют по числу и размеру (длине) выявляемые полиморфные маркеры.
Примеры выполнения предлагаемого способа.
Пример 1.
На фиг.1 представлен набор полиморфных ДНК, выявляемых в реакции I и II с помощью четырех предлагаемых праймеров на ДНК птичьих шистосом четырех видов: Т. regenti (дор. 1, 9), Т. szidati (дор. 2, 10), T. sp var. narochanica (дор. 3, 11), Т. franki (дор. 4, 12), а также птичьей шистосомы В. polonica (дор. 5, 13) и фуркоцеркарии трематод Cotylurus sp.(сем. Strigeidae) (дор. 6, 14); Diplostomum sp.(сем. Diplostomidae) (дор. 7, 15); Bolbophorus sp.(сем. Postodiplostomidae) (дор. 8. 16).
ДНК выделяют указанным выше способом и осуществляют идентификацию видов трихобильгарций путем сравнения амплификационных паттернов, причем сочетание праймера F и двух обратных праймеров R1 и R2 (праймерная система I, реакция I) позволяет детектировать у Т. szidati и Т. regenti по одному фрагменту разного размера - 316 п.н. и 255 п.н., соответственно. У третьего вида T.sp.var. narochanica эта праймерная система выявляет паттерны сразу из двух фрагментов размером 255 и 316 п.н., а для четвертого вида Т. franki продукты амплификации полностью отсутствуют. Использование праймерной системы II (F, R3) на ДНК четырех видов позволяет визуализировать в реакции II один фрагмент размером 258 п.н. и только в случае Т. franki строгая видоспецифичность обнаруженной изменчивости подтверждается полным отсутствием продуктов амплификации при использовании в качестве мишеней ДНК нескольких близкородственных видов трематод из семейств Schistosomatidae (В. polonica, дорожки 5, 13), сем. Strigeidae (Cotylurus sp., дор. 6, 14), Diplostomidae (дор. 7, 15) и сем. Postodiplostomidae (Bolbophorus sp., дор. 8, 16).
Пример 2.
ДНК выделяют указанным выше способом из марит, церкарий, спороцист T. sp. var narochanica (дор. 1-3), печени моллюска Radix sp, зараженного T. sp. var narochanica (дор. 4), из марит, церкарий, спороцист T. franki (дор. 6-8), печени моллюска Radix sp, зараженного T. franki (дор. 9), а также из печени незараженного моллюска Radix sp.(дор. 5, 14) (Фиг.2). На каждой из выделенных ДНК осуществляют ПНР с первой (дор. 1-5) или второй (дор. 6-14) праймерной системой. Полученные амплификационные продукты визуализируют путем электрофореза в агарозном геле и после окрашивания гелей бромистым этидием идентифицируют по размеру (длине) выявляемые полиморфные маркеры. Отсутствие амплификатов в спектрах неинфицированных моллюсков, принадлежащих к роду Radix из семейства Lymnaeidae, и наличие одного (для T. franki) или двух (для T. sp. var narochanica) диагностических фрагментов в спектрах марит, церкарий, спороцист и печени моллюска, инфицированной личинками двух исследованных видов трихобильгарций, подтверждает диагностическую ценность предлагаемых примеров для детектирования возбудителей церкариального дерматита на разных стадиях жизненного цикла, а именно диагностики на стадии моллюска. Разведение ДНК инфицированного моллюска (с исходной концентрацией ДНК 5γ/мкл, дорожка 9) в 5 (дорожка 10), 10 (дорожка 11), 20 (дорожка 12) и 100 раз (дорожка 13) не снижает чувствительности предлагаемого способа детекции личинок птичьих шистосом. По наличию в спектре инфицированного моллюска двух фрагментов длиной 255 п.н. и 316 п.н. определяется принадлежность личинок паразита к виду T. sp. var narochanica., а одного фрагмента длиной 258 п.н. - к виду T. franki.
Изобретение относится к паразитологии и касается способа видовой ДНК-дифференциации гельминтов - возбудителей церкариального дерматита человека. Дифференциацию четырех видов Trichobilharzia: Т. szidati, T.regenti, T.franki и T.sp.var.narochanica осуществляют путем амплификации участков последовательности ядерной рибосомальной ДНК (рДНК) в образцах половозрелых гельминтов, их личиночной стадии и/или на рДНК инфицированных указанными гельминтами пресноводных моллюсков из семейства Lymnaeidae с помощью ПЦР и четырех олигонуклотидных праймеров следующего состава:
F: 5'-CTTTCCATCTATCACGATGCACT-3'
R1: 5'-ATGATAATGTGCATAACACACC-3'
R2: 5'-GCCGTTTATTTATATGTATGTG-3'
R3: 5'-CAAGCCGTTTATTWATATATAACGG-3'.
Полученные амплификационные продукты визуализируют и дифференцируют и идентифицируют по размеру (длине), причем один амплификационный фрагмент размером 255 п.н. характерен для вида T.regenti, один фрагмент длиной 316 п.н. характерен для вида Т.szidati, два фрагмента размером 255 и 316 п.н. характерны для вида T.sp.var.narochanica, а амплификационный фрагмент размером 258 п.н. детектируется только у вида T.franki. Представленный способ позволяет одновременно детектировать и проводить видовую идентификацию четырех видов птичьих шистосом рода Trichobilharzia на разных стадиях жизни. 2 ил., 2 табл., 2 пр.
Способ видовой ДНК-дифференциации птичьих шистосом из рода Trichobilharzia - возбудителей церкариального дерматита человека на разных стадиях жизненного цикла, включающий выделение ДНК- возбудителя, амплификацию мишени с использованием специфических праймеров, ПЦР и электрофоретическое разделение, отличающийся тем, что проводят дифференциацию четырех видов Trichobilharzia: Т. szidati, T.regenti, T.franki и T.sp.var.narochanica путем амплификации участков последовательности ядерной рибосомальной ДНК (рДНК) в образцах половозрелых гельминтов, их личиночной стадии и/или на рДНК инфицированных указанными гельминтами пресноводных моллюсков из семейства Lymnaeidae с помощью ПЦР и четырех олигонуклотидных праймеров следующего состава:
F: 5'-CTTTCCATCTATCACGATGCACT-3'
R1: 5'-ATGATAATGTGCATAACACACC-3'
R2: 5'-GCCGTTTATTTATATGTATGTG-3'
R3: 5'-CAAGCCGTTTATTWATATATAACGG-3',
а также при использовании набора, содержащего биологический маркер, представляющий собой полиморфную ДНК и характеризующий видовую принадлежность шистосом рода Trichobilharzia и реакционную смесь, состоящую из 60 мМ трис-HCl, 10 мМ сульфата аммония, 0,1% TWEEN 20, по 100 мкМ каждого dNTP, 0,5 мкМ MgCl2, 0,9 единиц Taq-полимеразы, разделяют указанные амплифицированные последовательности рДНК с помощью электрофореза и эталонных ДНК для каждого вида Trichobilharzia - положительный контроль, с получением образцов видоспецифического электрофоретического паттерна гельминта, при этом на каждой из выделенных ДНК осуществляют две реакции ПЦР с набором из четырех предлагаемых праймеров, где в первой реакции используют праймеры - F, R1, R2, а во второй реакции - F, R3, полученные амплификационные продукты визуализируют и дифференцируют путем электрофореза в агарозном геле и после окрашивания гелей бромистым этидием идентифицируют по размеру (длине), причем в первой реакции один амплификационный фрагмент размером 255 п.н. характерен для вида T.regenti, один фрагмент длиной 316 п.н. характерен для вида Т.szidati, два фрагмента размером 255 и 316 п.н. характерны для вида T.sp.var.narochanica, а во второй реакции амплификационный фрагмент размером 258 п.н. детектируется только у вида T.franki.
| ХРИСАНФОВА Г.Г | |||
| и др | |||
| Генетическая изменчивость птичьих шистосом (класс Trematoda, сем | |||
| Schistosomatidae) озера Нарочь: идентификация нового вида в группе Trichobilharzia ocellata // Докдады Академии Наук | |||
| Колосоуборка | 1923 |
|
SU2009A1 |
| US 6818402 B2, 16.11.2004 | |||
| J | |||
| DVORAK et al., Comparison of European Trichobilharzia species based on ITS1 and ITS2 sequences, Parasitology, 2002, Vol.124, No.3, abstr. | |||

