УРОВЕНЬ ТЕНИКИ
[0001] Настоящее описание относится к изолированным полинуклеотидам, кодирующим дельта-9-элонгазу, дельта-9-элонгазам, которые кодируют указанные изолированные полинуклеотиды, векторам экспрессии, содержащим указанные изолированные полинуклеотиды, клеткам-хозяевам, содержащим указанные векторы экспрессии, и способам получения дельта-9-элонгаз и полиненасыщенных жирных кислот.
[0002] Полиненасыщенные жирные кислоты (ПНЖК) играют большую роль в правильном функционировании живых организмов. Например, ПНЖК представляют собой важные компоненты плазматической мембраны клетки, в которой они присутствуют в виде фосфолипидов. Они также являются предшественниками простациклинов, эйкозаноидов, лейкотриенов и простагландинов млекопитающих. Кроме того, ПНЖК необходимы для правильного формирования развивающегося головного мозга младенца, а также для образования и восстановления тканей. В связи с биологической значимостью ПНЖК, предпринимаются попытки эффективного получения ПНЖК, а также промежуточных продуктов, позволяющих их получить.
[0003] В биосинтез ПНЖК вовлечено множество ферментов, в основном десатураз и элонгаз (см. Фигуру 1). Десатуразы катализируют введение элементов ненасыщенности (например, двойных связей) между атомами углерода в субстрате-алкильной цепи жирной кислоты. Элонгазы катализируют добавление 2-углеродной единицы к жирной кислоте-субстрату. Например, линолевую кислоту (ЛК, 18:2n-6) получают из олеиновой кислоты (ОК, 18:1n-9) с помощью дельта-12-десатуразы. Эйкозандиеновую кислоту (ЭДК, 20:2n-6) получают из линолевой кислоты (ЛК, 18:2n-6) с помощью дельта-9-элонгазы. Дигомо-гамма-линоленовую кислоту (ДГЛК, 20:3n-6) получают из эйкозандиеновой кислоты (ЭДК, 20:2n-6) с помощью дельта-8-десатуразы. Арахидоновую кислоту (АРК, 20:4n-6) получают из дигомо-гамма-линоленовой кислоты (ДГЛК, 20:3n-6) с помощью дельта-5-десатуразы (см. Фигуру 1).
[0004] Элонгазы катализируют превращение гамма-линоленовой кислоты (ГЛК, 18:3n-6) в дигомо-гамма-линоленовую кислоту (ДГЛК, 20:3n-6) и превращение стеаридоновой кислоты (СДК, 18:4n-3) в эйкозатетраеновую кислоту (ЭТК, 20:4n-3). Элонгаза также катализирует превращение арахидоновой кислоты (АРК, 20:4n-6) в адреновую кислоту (АДК, 22:4n-6) и превращение эйкозапентаеновой кислоты (ЭПК, 20:5n-3) в омега-3-докозапентаеновую кислоту (22:5n-3). Дельта-9-элонгаза удлиняет полиненасыщенные жирные кислоты, содержащие элемент ненасыщенности в положении 9 атома углерода. Например, дельта-9-элонгаза катализирует превращение линолевой кислоты (ЛК, 18:2n-6) в эйкозандиеновую кислоту (ЭДК, 20:2n-6) и превращение альфа-линоленовой кислоты (АЛК, 18:3n-3) в эйкозатриеновую кислоту (ЭТрК, 20:3n-3). Омега-3-ЭТрК затем можно превратить в омега-3-ЭТК с помощью дельта-8-десатуразы. Омега-3-ЭТК затем можно использовать для получения других полиненасыщенных жирных кислот, таких как омега-3-ЭПК, которые можно добавлять в фармацевтические композиции, пищевые композиции, корма для животных, а также другие продукты, такие как косметические средства.
[0005] Обнаруженные ранее элонгазы отличаются по субстратам, на которые они действуют. Более того, они присутствуют как у животных, так и в растениях. Элонгазы, обнаруженные у млекопитающих, обладают способностью использовать в качестве субстрата насыщенные, мононенасыщенные и полиненасыщенные жирные кислоты. Напротив, обнаруженные в растениях элонгазы специфичны к насыщенным или мононенасыщенным жирным кислотам. Таким образом, существует потребность в элонгазе, специфичной к ПНЖК, для получения полиненасыщенных жирных кислот в растениях.
[0006] Полагают, что как у растений, так и у животных процесс элонгации состоит из четырех этапов (Lassner и др., The Plant Cell 8:281-292 (1996)). КоА представляет собой переносчик ацильной группы. Первый этап включает конденсацию малонил-КоА с длинноцепочечным ацил-КоА с получением диоксида углерода и β-кетоацил-КоА, в котором ацильная молекула удлинилась на два атома углерода. Последующие реакции включают восстановление до β-гидроксиацил-КоА, дегидрирование с получением еноил-КоА и повторное восстановление с получением удлиненного ацил-КоА. Исходная реакция конденсации представляет собой не только субстрат-специфичный этап, но и стадию, лимитирующую скорость реакции.
[0007] Следует отметить, что животные не могут уменьшать насыщенность ниже положения дельта-9 и, следовательно, не могут превращать олеиновую кислоту (ОК, 18:1n-9) в линолевую кислоту (ЛК, 18:2n-6). Аналогичным образом, альфа-линоленовая кислота (АЛК, 18:3n-3) не может быть синтезирована млекопитающими, так как у них отсутствует дельта-15-десатуразная активность. Тем не менее, у млекопитающих и водорослей альфа-линоленовая кислота может быть превращена в стеаридоновую кислоту (СДК, 18:4n-3) с помощью дельта-6-десатуразы (см. WO 96/13591; см. также патент США номер 5552306), а затем подвергнута элонгации с получением эйкозатетраеновой кислоты (ЭТК, 20:4n-3). Эта полиненасыщенная жирная кислота (т.е. ЭТК, 20:4n-3) затем может быть превращена в эйкозапентаеновую кислоту (ЭПК, 20:5-3) с помощью дельта-5-десатуразы. Другие эукариоты, включая грибы и растения, содержат ферменты, которые уменьшают насыщенность в положениях атомов углерода 12 (см. WO 94/11516 и патент США номер 5443974) и 15 (см. WO 93/11245). Следовательно, основные полиненасыщенные жирные кислоты животные получают из пищи и/или в результате десатурации и элонгации линолевой кислоты или альфа-линоленовой кислоты. В связи с неспособностью млекопитающих образовывать указанные незаменимые длинноцепочечные жирные кислоты, большой интерес представляет выделение генов, вовлеченных в биосинтез ПНЖК, из видов, которые в природе продуцируют данные жирные кислоты, и экспрессирование данных генов в системе микроорганизма, растения или животного, которую можно изменить, чтобы обеспечить получение промышленных количеств одной или нескольких ПНЖК. Следовательно, существует выраженная потребность в ферментах элонгазах, генах, кодирующих данные ферменты, а также рекомбинантных способах получения указанных ферментов.
[0008] В связи с приведенным обсуждением, также существует определенная потребность в маслах, содержащих ПНЖК на более высоком, чем присутствующие в природе, уровне, а также в маслах, обогащенных новыми ПНЖК. Такие масла можно получить путем выделения и экспрессии генов элонгазы.
[0009] Одной из наиболее важных длинноцепочечных ПНЖК является эйкозапентаеновая кислота (ЭПК). ЭПК обнаруживают в грибах, а также в маслах из морепродуктов. Докозагексаеновая кислота (ДГК) представляет собой другую важную длинноцепочечную ПНЖК. ДГК чаще всего обнаруживают в рыбьем жире, а также ее можно выделить из ткани головного мозга млекопитающего. Арахидоновая кислота (АРК) представляет собой третью важную длинноцепочечную ПНЖК. АРК обнаруживают в мицелиальных грибах, и ее также можно выделить из тканей млекопитающего, включая печень и надпочечники.
[0010] АРК, ЭПК и/или ДГК, например, можно получить либо с помощью альтернативного пути с участием дельта-8-десатуразы/дельта-9-элонгазы, либо с помощью обычного пути дельта-6 (см. Фигуру 1). Ранее были идентифицированы элонгазы, которые активны в отношении жирных кислот - субстратов в обычном пути дельта-6 получения длинноцепочечных ПНЖК, особенно АРК, ЭПК и ДГК. В обычном пути дельта-6 превращения ЛК в ДГЛК и АЛК в омега-3-ЭТК используют фермент дельта-6-десатуразу для превращения ЛК в ГЛК и АЛК в стеаридоновую кислоту (СДК) и фермент дельта-6-элонгазу для превращения ГЛК в ДГЛК и СДК в омега-3-ЭТК. Тем не менее, в некоторых случаях, альтернативный путь с участием дельта-8-десатуразы/дельта-9-элонгазы может оказаться предпочтительным по сравнению с обычным путем дельта-6. Например, если определенные остаточные промежуточные продукты омега-6 или омега-3 жирных кислот, такие как ГЛК или СДК, не желательны в процессе получения ДГЛК, омега-3-ЭТК, АРК, ЭПК, омега-3-докозапентаеновой кислоты, омега-6-докозапентаеновой кислоты, АДК и/или ДГК, в качестве альтернативы обычному пути дельта-6 можно применять другой путь с участием дельта-8-десатуразы/дельта-9-элонгазы, чтобы избежать образования ГЛК и СДК.
[0011] В настоящем описании идентифицировали новый источник дельта-9-элонгазы для получения длинноцепочечных ПНЖК, в частности ДГЛК, ЭТК, АРК, ЭПК, омега-3-докозапентаеновой кислоты, омега-6-докозапентаеновой кислоты, АДК и/или ДГК. Фермент дельта-9-элонгаза согласно настоящему описанию превращает, например, ЛК в омега-6-ЭДК и АЛК в омега-3-ЭТрК. Получение ДГЛК из омега-6-ЭДК и АРК из ДГЛК затем катализируется ферментом дельта-8-десатуразой и дельта-5-десатуразой, соответственно.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0012] В одном своем аспекте, настоящее описание относится к изолированной молекуле нуклеиновой кислоты, или фрагменту указанной нуклеиновой кислоты, которая включает изолированную последовательность нуклеотидов, кодирующую полипептид, обладающий элонгазной активностью, или которая является комплементарной указанной последовательности, при этом последовательность аминокислот указанного полипептида по меньшей мере на 68% идентична последовательности аминокислот, выбранной из группы, состоящей из SEQ ID NO:18 и SEQ ID NO:20.
[0013] В другом своем аспекте, настоящее описание относится к изолированной последовательности нуклеотидов, или фрагменту указанной последовательности, которая включает или комплементарна по меньшей мере 68% последовательности нуклеотидов, выбранной из группы, состоящей из SEQ ID NO:17 и SEQ ID NO:19,. Указанная изолированная последовательность нуклеотидов или фрагмент указанной последовательности кодирует функционально активную элонгазу, которая использует полиненасыщенную жирную кислоту в качестве субстрата, в частности, функционально активную дельта-9-элонгазу.
[0014] Указанную последовательность нуклеотидов можно получить, например, из Euglenoid sp., в частности, можно выделить, например, из Euglena deses Ehr. CCMP 2916.
[0015] В другом аспекте, настоящее описание относится к очищенному полипептиду, который кодирует описанная выше изолированная последовательность нуклеотидов, а также к очищенному полипептиду, который удлиняет полиненасыщенные жирные кислоты, содержащие элемент ненасыщенности в положении атома углерода 9, и обладает по меньшей мере 68% идентичности аминокислотной последовательности с последовательностью аминокислот, выбранной из группы, состоящей из SEQ ID NO:18 и SEQ ID NO:20.
[0016] В еще одном аспекте, настоящее описание относится к вектору экспрессии. Указанный вектор экспрессии содержит последовательность нуклеотидов, функционально связанную с регуляторной последовательностью, при этом указанная последовательность нуклеотидов включает или комплементарна по меньшей мере 68% последовательности нуклеотидов, выбранной из группы, состоящей из SEQ ID NO:17 и SEQ ID NO:19. Настоящее описание также относится к клетке-хозяину, содержащему указанный вектор экспрессии. Клетка-хозяин может представлять собой, например, эукариотическую клетку или прокариотическую клетку. Подходящие эукариотические клетки и прокариотические клетки описаны в данной заявке. Настоящее описание также относится к трансгенному семени, содержащему указанный вектор экспрессии.
[0017] В другом аспекте, настоящее описание относится к клетке растения, семени растения, растению или ткани растения, содержащим описанный выше вектор экспрессии, при этом экспрессия последовательности нуклеотидов указанного вектора экспрессии приводит к продуцированию по меньшей мере одной полиненасыщенной жирной кислоты указанными клеткой растения, растением или тканью растения. Полиненасыщенная жирная кислота может быть выбрана, например, из группы, состоящей из омега-6-ЭДК и омега-3-ЭТрК и комбинации перечисленных жирных кислот. Настоящее описание также охватывает одно или несколько растительных масел или жирных кислот, экспрессированных описанной выше клеткой растения, семенем растения, растением или тканью растения.
[0018] Кроме того, настоящее описание относится к способу получения дельта-9-элонгазы. Указанный способ включает следующие этапы: а) выделение последовательности нуклеотидов, которая включает или комплементарна по меньшей мере 68% последовательности нуклеотидов, выбранной из группы, состоящей из SEQ ID NO:17 и SEQ ID NO:19; b) конструирование вектора экспрессии, содержащего: i) указанную выделенную последовательность нуклеотидов, функционально связанную с и) регуляторной последовательностью; и с) введение указанного вектора экспрессии в клетку-хозяина на время и при условиях, которые достаточны для экспрессирования дельта-9-элонгазы, в соответствующих случаях. Указанная клетка-хозяин может, например, представлять собой эукариотическую клетку или прокариотическую клетку. В частности, эукариотическая клетка может представлять собой, например, клетку млекопитающего, клетку насекомого, клетку растения или клетку гриба. Клетка растения может быть получена из масличного растения, выбранного из группы, состоящей из сои, видов Brassica, сафлора, подсолнечника, кукурузы, хлопчатника и льна.
[0019] Дополнительно, настоящее описание относится к способу получения полиненасыщенной жирной кислоты, включающему следующие этапы: а) выделение последовательности нуклеотидов, которая включает или комплементарна по меньшей мере 68% последовательности нуклеотидов, выбранной из группы, состоящей из SEQ ID NO:17 и SEQ ID NO:19; b) конструирование вектора экспрессии, содержащего указанную выделенную последовательность нуклеотидов, функционально связанную с регуляторной последовательностью; с) введение указанного вектора экспрессии в клетку-хозяина на время и при условиях, которые достаточны для экспрессирования дельта-9-элонгазы; и d) обеспечение взаимодействия указанной экспрессированной дельта-9-элонгазы и полиненасыщенной жирной кислоты-субстрата с тем, чтобы превратить указанную полиненасыщенную жирную кислоту-субстрат в первую полиненасыщенную жирную кислоту-продукт. Указанная «полиненасыщенная жирная кислота-субстрат» представляет собой, например, ЛК или АЛК, и «первая полиненасыщенная жирная кислота-продукт» представляет собой, например, омега-6-ЭДК или омега-3-ЭТрК, соответственно. Данный способ может дополнительно включать этап воздействия на первую полиненасыщенную жирную кислоту-продукт по меньшей мере одной десатуразы, по меньшей мере одной дополнительной элонгазы или комбинации перечисленных ферментов с тем, чтобы превратить указанную первую полиненасыщенную жирную кислоту-продукт во вторую или последующую полиненасыщенную жирную кислоту-продукт. Указанная вторая или последующая полиненасыщенная жирная кислота-продукт может представлять собой, например, ДГЛК или омега-3-ЭТК, АРК, ЭПК, ДПК, ДГК или комбинации перечисленных жирных кислот.
[0020] В другом аспекте, настоящее описание относится к способу получения полиненасыщенной жирной кислоты в клетке-хозяине, включающему следующие этапы: а) выделение последовательности нуклеотидов, которая включает или комплементарна по меньшей мере 68% последовательности нуклеотидов, выбранной из группы, состоящей из SEQ ID NO:17 и SEQ ID NO:19; b) конструирование вектора экспрессии, содержащего указанную выделенную последовательность нуклеотидов, функционально связанную с регуляторной последовательностью; с) введение i) указанного вектора экспрессии и ii) по меньшей мере одной дополнительной конструкции рекомбинантной ДНК, содержащей указанную выделенную последовательность нуклеотидов, кодирующую дельта-8-десатуразу и функционально связанную с по меньшей мере одной регуляторной последовательностью, в клетку-хозяина на время и при условиях, которые достаточны для экспрессирования дельта-9-элонгазы и дельта-8-десатуразы; и d) обеспечения взаимодействия указанных экспрессированной дельта-9-элонгазы и дельта-8-десатуразы на полиненасыщенную жирную кислоту-субстрат, выбранную из группы, состоящей из ЛК, АЛК и комбинации перечисленных жирных кислоте тем, чтобы превратить указанную полиненасыщенную жирную кислоту-субстрат в первую полиненасыщенную жирную кислоту-продукт. Указанная первая полиненасыщенная жирная кислота-продукт может представлять собой, например, ДГЛК, омега-3-ЭТК или комбинации перечисленных жирных кислот. Данный способ может дополнительно включать этап воздействия на указанную первую полиненасыщенную жирную кислоту-продукт по меньшей мере одной десатуразы, по меньшей мере одной дополнительной элонгазы или комбинации перечисленных ферментов с тем, чтобы превратить указанную первую полиненасыщенную жирную кислоту-продукт во вторую или последующую полиненасыщенную жирную кислоту-продукт. Указанная вторая или последующая полиненасыщенная жирная кислота-продукт может представлять собой, например, АРК, ЭПК, ДПК, ДГК или комбинацию перечисленных жирных кислот. В одном аспекте, указанный способ может дополнительно включать введение в клетку-хозяина конструкции рекомбинантной ДНК, содержащей i) выделенную последовательность нуклеотидов, кодирующую дельта-5-десатуразу, функционально связанную с ii) регуляторной последовательностью. Указанная клетка-хозяин может представлять собой описанную выше клетку-хозяина.
[0021] В другом своем аспекте, настоящее описание относится к способу получения трансгенного растения, включающему трансформирование клетки растения по меньшей мере одной выделенной последовательностью нуклеотидов, или фрагментом указанной последовательности, которая включает или комплементарна по меньшей мере 68% последовательности нуклеотидов, выбранной из группы, состоящей из SEQ ID NO:17 и SEQ ID NO:19, и регенерирование трансгенного растения из трансформированной клетки растения. Указанную клетку растения можно получить из масличного растения, выбранного из группы, состоящей из сои, видов Brassica, сафлора, подсолнечника, кукурузы, хлопчатника и льна. В другом аспекте, настоящее описание относится к семени, полученному из такого трансгенного растения, полученного с помощью указанного способа.
[0022] Также следует отметить, что каждой последовательности нуклеотидов и аминокислот, упомянутой в настоящем описании, был присвоен конкретный идентификационный номер последовательности. В Перечне последовательностей (который приложен к настоящему описанию), включенном в данную заявку посредством ссылки, перечислены все такие последовательности и соответствующие им номера.
КРАТКОЕ ОПИСАНИЕ ФИГУР
[0023] На Фигуре 1 показана путь биосинтеза жирных кислот и роль дельта-9-элонгазы в этом пути.
[0024] На Фигурах 2А и 2В показано выравнивание последовательностей нуклеотидов SEQ ID NO:26 и SEQ ID NO:27, которые представляют собой последовательности нуклеотидов вариантов Eug-MO7-ELO#10 и Eug-MO7-ELO#14, соответственно, клонированных в сайты Ват HI/Hind III вектора pYX242, что обсуждается в Примере 3.
Варианты обведены прямоугольниками.
[0025] На Фигурах 3А и 3В показано выравнивание последовательностей аминокислот дельта-9-элонгазы из Euglena deses Ehr. CCMP 2916 (Eug-MO7-ELO-10) (SEQ ID NO:18) с известными дельта-9-элонгазами из Euglena gracialis (SEQ ID NO:4), Isochrysis galbana (SEQ ID NO:2); элонгазой Elov 14 мыши (номер доступа AAG47667; SEQ ID NO:21), элонгазой ELOVL2 человека (номер доступа NP_060240; SEQ ID NO:22) и элонгазой С. elegans (номер доступа AF244356; SEQ ID NO:23). Неизменяющиеся остатки заштрихованы.
[0026] На Фигуре 4А показана последовательность аминокислот дельта-9-элонгазы из Pavlova salma (номер доступа AAY15135; SEQ ID NO:1).
[0027] На Фигуре 4В показана последовательность аминокислот дельта-9-элонгазы из Isochrysis galbana (номер доступа AF390174; SEQ ID NO:2).
[0028] На Фигуре 4С показана последовательность аминокислот дельта-9-элонгазы из Eutreptiella sp. (SEQ ID NO:3).
[0029] На Фигуре 4D показана последовательность аминокислот дельта-9-элонгазы из Euglena gracialis (номер доступа САТ16687; SEQ ID NO:4).
[0030] На Фигуре 4Е показана последовательность аминокислот дельта-9-элонгазы из Euglena anabena (SEQ ID NO:5).
[0031] На Фигуре 5А показана последовательность нуклеотидов (SEQ ID NO:6) клона plate2_MO7, получение которого описано в Примере 1.
[0032] На Фигуре 5В показана предсказанная последовательность аминокислот (SEQ ID NO:7) клона plate2_MO7, получение которого описано в Примере 1.
[0033] На Фигуре 6А показана последовательность нуклеотидов (SEQ ID NO:13) предполагаемого 3′-конца фрагмента гена plate2_MO7, получение которого описано в Примере 2.
[0034] На Фигуре 6В показана предсказанная последовательность аминокислот (последовательности SEQ ID NO:14 и 30-32) предполагаемого 3′-конца фрагмента гена plate2_MO7, получение которого описано в Примере 2. Последовательности SEQ ID NO:14 и 30-32 отделены значком "*", который представляет собой стоп-кодон.
[0035] На Фигуре 6С показана последовательность SEQ ID NO:14.
[0036] На Фигуре 6D показана последовательность SEQ ID NO:30.
[0037] На Фигуре 6Е показана последовательность SEQ ID NO:31.
[0038] На Фигуре 6F показана последовательность SEQ ID NO:32.
[0039] На Фигуре 7А показана последовательность нуклеотидов (SEQ ID NO:17) предполагаемой дельта-9-элонгазы из Euglena deses Ehr. CCMP 2916 (Eug-MO7-ELO#10), получение которого описано в Примере 3.
[0040] На Фигуре 7В показана предсказанная последовательность аминокислот (SEQ ID NO:18), кодируемая последовательностью нуклеотидов (SEQ ID NO:17) предполагаемой дельта-9-элонгазы из Euglena deses Ehr. CCMP 2916 (Eug-MO7-ELO#10), получение которого описано в Примере 3.
[0041] На Фигуре 8А показана последовательность нуклеотидов (SEQ ID NO:19) варианта дельта-9-элонгазы из Euglena deses Ehr. CCMP 2916 (Eug-MO7-ELO#14), получение которого описано в Примере 3.
[0042] На Фигуре 8В показана предсказанная последовательность аминокислот (SEQ ID NO:20), кодируемая последовательностью нуклеотидов (SEQ ID NO:19) варианта дельта-9-элонгазы из Euglena deses Ehr. CCMP 2916 (Eug-MO7-ELO#14), получение которого описано в Примере 3.
[0043] На Фигуре 9А показана последовательность аминокислот элонгазы Elovl4 мыши (номер доступа AAG47667; SEQ ID NO:21).
[0044] На Фигуре 9В показана последовательность аминокислот элонгазы ELOVL2 человека (номер доступа NP_060240; SEQ ID NO:22).
[0045] На Фигуре 9С показана последовательность аминокислот элонгазы С. elegans (номер доступа AF244356; SEQ ID NO:23).
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0046] Настоящее описание относится к последовательностям нуклеотидов (например, гену) и транслированным последовательностям аминокислот гена дельта-9-элонгазы из Euglenoid sp., например, Euglena deses Ehr., в частности Euglena deses Ehr. CCMP 2916. Более того, настоящее описание также охватывает варианты применения указанного гена и фермента, кодируемого указанным геном. Например, ген и соответствующий фермент можно применять для получения полиненасыщенных жирных кислот, таких как, например, омега-6-ЭДК, омега-3-ЭТрК, ДГЛК, омега-3-ЭТК, АРК, ЭПК, омега-3-докозапентаеновая кислота, омега-6-докозапентаеновая кислота, АДК, ДГК или любая комбинация перечисленных жирных кислот, которые можно добавить в фармацевтические композиции, пищевые композиции и в другие полезные продукты.
Определения
[0047] В данной заявке, формы единственного числа включают ссылку на множественное число, если в контексте явно не указано иное. При перечислении в данной заявке числовых диапазонов, предполагается, что каждое число, находящееся в рамках данных диапазонов, указано в явной форме с одинаковой степенью точности. Например, предполагается, что в диапазоне 6-9, в дополнение к 6 и 9, указаны числа 7 и 8, а в диапазоне 6,0-7,0 в явной форме указаны числа 6,0, 6,1, 6,2, 6,3, 6,4, 6,5, 6,6, 6,7, 6,8, 6,9 и 7,0.
[0048] Химерная конструкция: В данной заявке, формулировка "химерная конструкция" относится к комбинации молекул нуклеиновых кислот, которые обычно не встречаются вместе в природе. Соответственно, химерная конструкция может включать регуляторные последовательности и кодирующие последовательности, которые получают из различных источников, или регуляторные последовательности и кодирующие последовательности, полученные из одного и того же источника, но расположенные в порядке, отличном от обычно встречающегося в природе.
[0049] Кодирующая последовательность: В данной заявке, термин "кодирующая последовательность" относится к последовательности ДНК, которая кодирует определенную последовательность аминокислот. "Регуляторные последовательности" относятся к последовательностям нуклеотидов, расположенным против хода транскрипции (5′-некодирующие последовательности), внутри или по ходу транскрипции (3′-некодирующие последовательности) от кодирующей последовательности, при этом указанные последовательности нуклеотидов влияют на транскрипцию, процессинг или стабильность РНК или трансляцию связанной с ними кодирующей последовательности. Регуляторные последовательности могут включать, но не ограничены перечисленными, промоторы, последовательности, направляющие трансляцию, интроны и регуляторные последовательности полиаденилирования.
[0050] Комплементарность: В данной заявке, термин "комплементарность" относится к степени схожести между двумя фрагментами ДНК. Ее определяют путем измерения способности смысловой нити одного фрагмента ДНК гибридизоваться с антисмысловой нитью другого фрагмента ДНК при условиях, подходящих для образования двойной спирали. В двойной спирали, аденин присутствует на одной нити, тимин присутствует на другой нити. Аналогично, там, где гуанин присутствует на одной нити, цитозин обнаруживают на другой. Чем больше схожесть между последовательностями нуклеотидов двух фрагментов ДНК, тем больше способность образовывать гибридные дуплексы между нитями двух указанных фрагментов ДНК.
[0051] Кодируется, гибридизация и строгие условия: В настоящем описании формулировка "кодируется" относится к последовательности нуклеиновых кислот, которая кодирует полипептидную последовательность, при этом указанная полипептидная последовательность или ее часть содержит аминокислотную последовательность, состоящую из по меньшей мере трех последовательных аминокислот, более предпочтительно из по меньшей мере восьми последовательных аминокислот и еще более предпочтительно из по меньшей мере 15 последовательных аминокислот полипептида, кодируемого указанной последовательностью нуклеиновых кислот.
[0052] В объем настоящего описания также входит изолированная последовательность нуклеотидов, которая кодирует фермент, обладающий активностью элонгазы ПНЖК, и которая способна гибридизоваться при умеренно строгих условиях с нуклеиновой кислотой, обладающей последовательностью нуклеотидов, которая включает или комплементарна последовательности нуклеотидов, включающей SEQ ID NO:17 или SEQ ID NO:19 (как показано на Фигурах 7А и 8А, соответственно). Молекула нуклеиновой кислоты "способна гибридизоваться" с другой молекулой нуклеиновой кислоты, если одноцепочечная форма указанной молекулы нуклеиновой кислоты может отжигаться с другой молекулой нуклеиновой кислоты при подходящих условиях, то есть при подходящей температуре и ионной силе (см., Sambrook и др., Molecular Cloning: A Laboratory Manual, второе издание (1989), Cold Spring Harbor Laboratory Press, Колд Спринг Харбор, Нью-Йорк)). Условия (температура и ионная сила) определяют "строгость" гибридизации. Для "гибридизации" необходимо, чтобы две нуклеиновые кислоты содержали комплементарные последовательности. Тем не менее, в зависимости от строгости гибридизации, могут встречаться несовпадения между основаниями. Подходящая строгость гибридизации нуклеиновых кислот зависит от длин нуклеиновых кислот и степени их комплементарности. Такие параметры хорошо известны специалистам в данной области. В частности, чем больше степень подобия или гомологии между двумя последовательностями нуклеотидов, тем больше значение Tm для гибридов нуклеиновых кислот с данными последовательностями. Для гибридов длиной больше, чем 100 нуклеотидов, вывели уравнения для вычисления Tm (см., Sambrook и др., выше). Для гибридизации с более короткими нуклеиновыми кислотами, положение несовпадений становится более существенным, и длина олигонуклеотида определяет его специфичность (см., Sambrook и др., выше).
[0053] Обычно строгие условия представляют собой такие условия, при которых концентрация соли меньше, чем приблизительно 1,5 М ионов Na, обычно концентрация ионов Na (или других солей) приблизительно составляет от 0,01 до 1,0 М при рН от 7,0 до 8,3 и температура составляет по меньшей мере приблизительно 30°С для коротких зондов (например, размером от 10 до 50 нуклеотидов) и по меньшей мере приблизительно 60°С для длинных зондов (например, размером больше, чем 50 нуклеотидов). Строгих условий также можно добиться путем добавления дестабилизирующих агентов, таких как формамид. Пример условий низкой строгости включает гибридизацию в буферном растворе с содержанием формамида от 30 до 35%, 1 М NaCl, 1% ДСН (додецилсульфата натрия) при 37°С и промывку от 1 Х до 2 Х SSC (20 Х SSC=3,0 M NaCl/0,3 М тринатриевого цитрата) при температуре от 50 до 55°С. Пример умеренно строгих условий включает гибридизацию в буферном растворе с содержанием формамида от 40 до 45%, 1 М NaCl, 1% ДСН при 37°С и промывку от 0,5 Х до 1 Х SSC при температуре от 55 до 60°С. Пример высоко строгих условий включает гибридизацию в буферном растворе с 50% формамидом, 1 М NaCl, 1% ДСН при 37°С и промывку 0,1 Х SSC при температуре от 60 до 65°С.
[0054] Экзон: В данной заявке, термин "экзон" относится к части последовательности гена, которая транскрибируется и обнаруживается в зрелой информационной РНК, полученной из указанного гена, но не обязательно является частью последовательности, которая кодирует конечный продукт гена.
[0055] Экспрессия, антисмысловое ингибирование и ко-супрессия: В данной заявке, термин "экспрессия" относится к получению функционального конечного продукта. Экспрессия гена включает транскрипцию гена и трансляцию мРНК с образованием белка-предшественника или зрелого белка.
[0056] В данной заявке, формулировка "антисмысловое ингибирование" относится к получению транскриптов антисмысловых РНК, способных подавлять экспрессию целевого белка.
[0057] В данной заявке, термин "ко-супрессия" относится к продукции смысловых транскриптов РНК, способных подавлять экспрессию идентичных или по существу сходных чужеродных или эндогенных генов (см. патент США номер 5231020).
[0058] Фрагмент или часть фрагмента, который функционально эквивалентен: Термины "фрагмент или часть фрагмента, который функционально эквивалентен" и "функционально эквивалентенный фрагмент или часть фрагмента", которые используют взаимозаменяемо в настоящем описании, относятся к части или подпоследовательности изолированной молекулы нуклеиновой кислоты, у которой сохраняется способность изменять экспрессию гена или вызывать некоторый фенотип независимо от того, кодирует или нет указанный фрагмент или часть фрагмента активный фермент. Например, фрагмент или часть фрагмента можно применять при разработке химерных конструкций для получения желательного фенотипа у трансформированного растения. Химерные конструкции можно разработать специально, чтобы применять для ко-супрессии или антисмыслового ингибирования, путем присоединения фрагмента или части фрагмента нуклеиновой кислоты, независимо от того, кодирует он или нет активный фермент, в подходящей ориентации по отношению к промоторной последовательности растения.
[0059] Ген, нативный ген, чужеродный ген и трансген: В данной заявке, термин "ген" относится к молекуле нуклеиновой кислоты, которая экспрессирует определенный белок, включая регуляторные последовательности, предшествующие (5′-некодирующие последовательности) и следующие (3′-некодирующие последовательности) за кодирующей последовательностью.
[0060] В данной заявке, формулировка "нативный ген" относится к гену в таком виде, в котором он встречается в природе, с его собственными регуляторными последовательностями.
[0061] "Чужеродный" ген относится к гену, обычно не встречающемуся в организме хозяина, но который был введен в организм хозяина путем переноса генов. Чужеродные гены могут включать нативные гены, вставленные в ненативный организм, или химерные конструкции.
[0062] В данной заявке, термин "трансген" относится к гену, который был введен в геном с помощью процедуры трансформации.
[0063] Виды Gossvpium: В данной заявке, формулировка "виды Gossypium" относится к любым из перечисленных растений: Gossypium arboreum, Gossypium barbadense, Gossypium herbaceum, Gossypium hirsutum, Gossypium hirsutum var hirsutum, Gossypium hirsutum var marie-galante, Gossypium lapideum, Gossypium sturtianum, Gossypium thuberi, Gossypium thurberi, Gossypium tomentosum или Gossypium tormentosum.
[0064] Гомология: Термины "гомология", "гомологичный", "по существу сходный" и "по существу соответствующий" используются взаимозаменяемо в настоящем описании и относятся к молекулам нуклеиновых кислот, в которых изменение одного или нескольких нуклеотидных оснований не влияет на способность указанной молекулы нуклеиновой кислоты опосредовать экспрессию гена или определять некоторый фенотип. Данные термины также относятся к модификациям молекул нуклеиновых кислот согласно настоящему описанию, таким как делеция или вставка одного или нескольких нуклеотидов, которые по существу не изменяют функциональных свойств полученной в результате этого молекулы нуклеиновой кислоты по сравнению с исходной, немодифицированной молекулой. Следовательно, для специалистов в данной области должно быть очевидно и понятно, что в объем настоящего описания входят не только конкретные типичные последовательности.
[0065] Клетка-хозяин: В данной заявке, под формулировкой "клетка-хозяин" подразумевают клетку, которая содержит изолированную последовательность нуклеиновых кислот или ее фрагмент согласно настоящему описанию. Клетка-хозяин могут представлять собой прокариотическую клетку (например, такую как Escherichia coli, цианобактерии и Bacillus subtilis) или эукариотическую клетку (например, такую как клетку гриба, насекомого, растения или млекопитающего).
[0066] Примеры клеток гриба, которые можно использовать, представляют собой Saccharomyces spp., Candida spp., Lipomyces spp., Yarrowia spp., Kluyveromyces spp., Hansenula spp., Aspergillus spp., Pemcillium spp., Neurospora spp., Trichoderma spp.и Pichia spp.Особенно предпочтительной клеткой гриба является Saccharomyces cerevisiae.
[0067] Клетки растения могут быть клетками однодольного или двудольного растения. Особенно предпочтительные клетки растения получают из масличных растений, таких как Glycine max (например, соя), виды Brassica, Carthamus tinctorius L. (например, сафлор), Helianthus annuus (например, подсолнечник), Zea mays (например, кукуруза), виды Gossypium (хлопчатник) и Linum usitatissimum (например, лен).
[0068] Идентичность, идентичность последовательностей и процент идентичности последовательностей (% идентичности): Термины "идентичность" или "идентичность последовательностей", которые используют взаимозаменяемо в настоящем описании, когда они используются в контексте последовательностей нуклеотидов или последовательностей полипептидов, относятся к основаниям нуклеиновых кислот или к аминокислотным остаткам в двух последовательностях, которые оказываются одинаковыми, когда их выравнивают на максимальное соответствие в рамках определенного окна сравнения. Таким образом, идентичность определяют как степень одинаковости, соответствия или эквивалентности между одинаковыми нитями (либо смысловыми, либо антисмысловыми) двух фрагментов ДНК или полипептида.
[0069] "Процент идентичности последовательностей" или "% идентичности" рассчитывают путем сравнения двух оптимально выровненных последовательностей на протяжении определенного участка, определения количества положений, в которых встречаются идентичные основания в обеих последовательностях, для получения количества совпадающих положений, деления количества таких положений на общее количество положений в сравниваемом фрагменте и умножения результата на 100. Оптимальное выравнивание последовательностей можно осуществить с помощью алгоритма Смита-Ватермана, Appl. Math. 2:482 (1981), с помощью алгоритма Нидлмана-Вунша, J. Mol. Biol. 48:443 (1970), с помощью способа по Пирсону и Липману, Proc. Natl. Acad. Sci. (USA) 85:2444 (1988), и с помощью компьютерных программ, которые осуществляют соответствующие алгоритмы (например, Higgins и др., CABIOS. 5L151-153 (1989)), FASTDB (Intelligenetics), BLAST (National Center for Biomedical Information; Altschul и др., Nucleic Acids Research 25:3389-3402 (1997)), PILEUP (Genetics Computer Group, Мэдисон, Висконсин) или GAP, BESTFIT, FASTA и TFASTA (пакет программ Wisconsin Genetics, версия 7.0, Genetics Computer Group, Мэдисон, Висконсин) (см. патент США номер 5912120). Подходящие примеры процента идентичности последовательностей включают, но не ограничены перечисленными, 68%, 69%, 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%. Идентичность можно определить с помощью любых из компьютерных программ, описанных в данной заявке.
[0070] Опосредованно или непосредственно: В данной заявке, в объем термина "опосредованно", когда он используется применительно к использованию гена и соответствующего фермента для получения полиненасыщенных жирных кислот, входит случай, когда первая кислота превращается во вторую кислоту (т.е. интермедиат пути) под воздействием первого фермента (например, ЛК в омега-6-ЭДК, например, посредством дельта-9-элонгазы), а затем вторая кислота превращается в третью кислоту под воздействием второго фермента (например, омега-6-ЭДК в ДГЛК, например, посредством дельта-8-десатуразы).
[0071] В данной заявке, в объем термина "непосредственно", когда он используется применительно к использованию гена и соответствующего фермента для получения полиненасыщенных жирных кислот, входит случай, когда фермент непосредственно превращает первую кислоту во вторую кислоту, при этом вторая кислота затем используется в композиции (например, превращение ЛК в омега-6-ЭДК, например, посредством дельта-9-элонгазы, или омега-3-ЭТрК в омега-3-ЭТК, например, посредством дельта-8-десатуразы).
[0072] Интрон: В данной заявке, термин "интрон" относится к последовательности, находящейся внутри гена, которая не кодирует часть последовательности белка. Следовательно, такие последовательности транскрибируются в РНК, но затем вырезаются и не транслируются. Данный термин также используют для вырезанных последовательностей РНК.
[0073] Изолированный: В данной заявке, термин "изолированный" или "выделенный" относится к молекуле нуклеиновой кислоты (ДНК или РНК), или к белку, или к биологически активной его части, которые удалены от окружения, в котором они встречаются в природе, или от их источника с помощью обычных методик, известных в данной области (например, от бактерий, водорослей, грибов, растений, позвоночных, млекопитающих и т.д.). Изолированные молекулы нуклеиновых кислот или белки практически или по существу свободны от компонентов, которые обычно сопутствуют или взаимодействуют с молекулами нуклеиновых кислот или белками во встречающемся в природе окружении.
[0074] Изолированный фрагмент нуклеиновой кислоты или изолированная последовательность нуклеиновых кислот: В данной заявке, формулировка "изолированный фрагмент нуклеиновой кислоты" или "изолированная последовательность нуклеиновых кислот" относится к полимеру РНК или ДНК, который состоит из одной или двух цепочек и возможно содержит синтетические, искусственные или измененные нуклеотидные основания. Изолированный фрагмент нуклеиновой кислоты в виде полимера ДНК может состоять из одного или нескольких фрагментов кДНК, геномной ДНК или синтетической ДНК ("фрагмент" определенного полинуклеотида относится к полинуклеотидной последовательности, которая включает непрерывную последовательность, состоящую по меньшей мере из приблизительно 6 последовательных нуклеотидов, предпочтительно по меньшей мере из приблизительно 8 последовательных нуклеотидов, более предпочтительно по меньшей мере из приблизительно 10 последовательных нуклеотидов, по меньшей мере из приблизительно 15 последовательных нуклеотидов, по меньшей мере из приблизительно 20 последовательных нуклеотидов, по меньшей мере из приблизительно 25 последовательных нуклеотидов и т.д., идентичную или комплементарную участку данной последовательности нуклеотидов). Для описания нуклеотидов (обычно находятся в 5′-монофосфатной форме) пользуются однобуквенными обозначениями, описанными далее: "А" для аденилата или дезоксиаденилата (для РНК или ДНК, соответственно), "С" для цитидилата или дезоксицитидилата, "G" для гуанилата или дезоксигуанилата, "U" для уридилата, "Т" для дезокситимидилата, "R" для пуринов (А или G), "Y" для пиримидинов (С или Т), "К" для G или Т, "Н" для А, или С, или Т, "I" для инозина и "N" для любого нуклеотида.
[0075] Зрелый и предшественник: В данной заявке, термин "зрелый", когда он используется применительно к термину "белок", относится к полипептиду, который подвергся посттрансляционному процессингу; т.е. к полипептиду, из которого были удалены все пре- или пропоследовательности, присутствующие в первичном продукте трансляции. В данной заявке, термин "предшественник", когда он используется применительно к термину "белок", относится к первичному продукту трансляции мРНК; т.е. то есть к полипептиду, в котором все еще присутствуют пре- или пропоследовательности. Пре- или пропоследовательности могут представлять собой, но не ограничены сигналами внутриклеточной локализации.
[0076] 3′-некодируюшие последовательности: В данной заявке, формулировка "3′-некодирующие последовательности" относится к последовательностям ДНК, расположенным по ходу транскрипции от кодирующей последовательности, которые содержат регуляторные последовательности полиаденилирования и другие последовательности, кодирующие регуляторные сигналы, способные влиять на процессинг мРНК или экспрессию гена. Сигнал полиаденилирования обычно описывают как сигнал, влияющий на добавление участков полиадениловой кислоты к 3′-концу предшественника мРНК. Примеры применения различных 3′-некодирующих последовательностей описаны у Ingelbrecht и др., (1989) Plant Cell 1:671 680.
[0077] Не встречающийся в природе: В настоящем описании формулировка "не встречающийся в природе" относится к чему-либо искусственному, не соответствующему обычно встречающемуся в природе.
[0078] Функционально связанный: В данной заявке, формулировка "функционально связанный" относится к такой связи последовательностей нуклеиновых кислот в одной молекуле нуклеиновой кислоты, что функция одной из последовательностей регулируется другой из них. Например, промотор называют функционально связанным с кодирующей последовательностью, если он способен регулировать экспрессию данной кодирующей последовательности (т.е. указанная кодирующая последовательность находится под транскрипционным контролем данного промотора). Кодирующие последовательности могут быть функционально связаны с регуляторными последовательностями в смысловой или антисмысловой ориентации. В другом примере, комплементарные участки РНК согласно настоящему описанию могут быть функционально связаны либо непосредственно, либо опосредованно, с 5′-стороны от целевой мРНК или с 3′-стороны от целевой мРНК, или внутри целевой мРНК, или первый комплементарный участок находится с 5′-стороны и комплементарный ему участок находится с 3′-стороны от целевой мРНК.
[0079] Растение: В данной заявке, термин "растение" относится к целым растениям, органам растения, тканям растения, семенам, клеткам растения, зернам и потомству растения. Клетки растения включают, без ограничения, клетки семян, суспензионных культур, зародышей, меристематических участков, каллусной ткани, листьев, корней, побегов, гаметофитов, спорофитов, пыльцы и микроспор.
[0080] Полимеразная цепная реакция или ПЦР: В данной заявке, формулировка "полимеразная цепная реакция" или "ПЦР" относится к способу синтезирования больших количеств определенных фрагментов ДНК, состоящему из ряда повторяющихся циклов (Perkin Elmer Cetus Instruments, Норуолк, Коннектикут). Обычно двунитевую ДНК денатурируют нагреванием, отжигают при низкой температуре два праймера, комплементарные к 3′-границам целевого фрагмента, а затем удлиняют при промежуточной температуре. Один набор этих трех последовательных этапов называют циклом.
[0081] ПЦР представляет собой эффективный способ для амплификации ДНК в миллионы раз путем повторных репликаций матрицы за короткий период времени (Mullis и др., Cold Spring Harbor Symp.Quant. Biol. 51:263 273 (1986); Eriich и др., заявка на европейский патент 50424; заявка на европейский патент 84796; заявка на европейский патент 258017, заявка на европейский патент 237362; Mullis, заявка на европейский патент 201184, Mullis и др., патент США номер 4683202; Eriich, патент США номер 4582788; и Saiki и др., патент США номер 4683194). В данном процессе в качестве затравки для синтеза ДНК используют наборы специфичных синтезированных in vitro олигонуклеотидов. Разработка праймеров зависит от последовательностей ДНК, которые нужно проанализировать. Указанный способ осуществляют с помощью большого количества циклов (обычно 20-50) плавления матрицы при высокой температуре, позволяя праймерам отжигаться на комплементарные последовательности матрицы, а затем реплицирования матрицы под воздействием ДНК полимеразы. Продукты ПЦР анализируют путем разделения в агарозных гелях, а затем окрашивания бромистым этидием и визуализации путем УФ-трансиллюминации. В качестве альтернативы, можно добавить в ПЦР радиоактивные дНТФ, чтобы ввести метку в продукты. В этом случае продукты ПЦР визуализируют путем экспонирования геля с рентгеновской пленкой. Дополнительное преимущество радиоактивного мечения продуктов ПЦР состоит в том, что можно произвести количественный анализ уровней отдельных продуктов амплификации.
[0082] Промотор и энхансер: В данной заявке, термин "промотор" относится к последовательностям ДНК, способным контролировать экспрессию кодирующей последовательности или функциональной РНК. Промоторная последовательность состоит из близких и более отдаленных расположенных против хода транскрипции элементов, последние элементы часто называют энхансерами.
[0083] В данной заявке, термин "энхансер" относится к последовательностям ДНК, которые могут стимулировать активность промотора и могут представлять собой природный элемент промотора или гетерологичный элемент, вставленный для повышения уровня активности или тканеспецифичности промотора. Промоторные последовательности также могут располагаться внутри транскрибируемых участков генов, и/или по ходу транскрипции от транскрибируемых последовательностей. Промоторы могут быть получены полностью из нативного гена или могут состоять из различных элементов, полученных из различных встречающихся в природе промоторов, или даже могут включать синтетические фрагменты ДНК. Для специалиста в данной области должно быть очевидно, что различные промоторы могут направлять экспрессию гена в различных тканях или типах клеток, или на различных стадиях развития, или в ответ на различные условия окружающей среды. Промоторы, которые вызывают экспрессирование гена в большинстве типов клеток в большинство моментов времени, обычно называют "конститутивными промоторами". Постоянно обнаруживают новые промоторы различных типов, пригодные для клеток растения; многочисленные их примеры можно найти в сборнике Okamuro и Goldberg, (1989) Biochemistry of Plants 15:1 82. Также должно быть очевидно, что поскольку в большинстве случаев точные границы регуляторных последовательностей полностью не определены, молекулы ДНК с некоторыми вариациями могут обладать идентичной промоторной активностью.
[0084] Рекомбинантный: В данной заявке, термин "рекомбинантный" относится к искусственному соединению двух в противном случае отдельных фрагментов последовательности, например, путем химического синтеза или путем манипулирования над изолированными фрагментами нуклеиновых кислот с помощью методик генной инженерии.
[0085] Рекомбинантная конструкция, экспрессионная конструкция и рекомбинантная экспрессионная конструкция: Формулировки "рекомбинантная конструкция", "экспрессионная конструкция" и "рекомбинантная экспрессионная конструкция" используют взаимозаменяемо в настоящем описании, и они относятся к функциональным единицам генетического материала, которые можно вставить в геном клетки, применяя стандартную методику, хорошо известную специалисту в данной области. Такую конструкцию можно применять саму по себе или совместно с вектором. Если используют вектор, тогда выбор вектора зависит от способа, который будет использоваться для трансформирования растений-хозяев, что хорошо известно специалистам в данной области. Например, можно применять плазмидный вектор. Квалифицированный специалист хорошо осведомлен о генетических элементах, которые должны присутствовать в векторе для успешной трансформации, селекции и размножения клеток-хозяев, содержащих любую из изолированных молекул нуклеиновых кислот согласно настоящему описанию. Для квалифицированного специалиста также должно быть очевидно, что различные независимые события трансформации приведут к различным уровням и паттернам экспрессии (Jones и др., (1985) EMBO J. 4:2411 2418; De Almeida и др., (1989) Mol. Gen. Genetics 218:78 86), и, следовательно, что нужно осуществить скрининг множества событий для получения линий, обладающих желательным уровнем и паттерном экспрессии. Такой скрининг можно осуществить путем анализа ДНК по Саузерну, Нозерн-анализа экспрессии мРНК, анализа методом вестерн-блот экспрессии белка или фенотипического анализа.
[0086] РНК-транскрипт, информационная РНК. кДНК, функциональная РНК и эндогенная РНК: В настоящем описании формулировка "РНК-транскрипт" относится к продукту, возникшему в результате транскрипции последовательности ДНК, катализируемой РНК-полимеразой. Если РНК-транскрипт представляет собой идеально комплементарную копию последовательности ДНК, его называют первичным транскриптом, или РНК-транскрипт может представлять собой последовательность РНК, полученную в результате посттранскрипционного процессинга первичного транскрипта, тогда его называют зрелой РНК.
[0087] В данной заявке, формулировка "информационная РНК (мРНК)" относится к РНК, которая не содержит интронов и которая может транслироваться в белок в клетке.
[0088] В данной заявке, термин "кДНК" относится к ДНК, которая синтезирована по матрице мРНК с помощью фермента обратной траскриптазы и комплементарна ей. КДНК может быть одноцепочечной, или ее можно превратить в двунитевую форму, применяя фрагмент Кленова ДНК-полимеразы I. "Смысловая" РНК относится к РНК-транскрипту, который включает мРНК и может транслироваться в белок внутри клетки или in vitro. "Антисмысловая РНК" относится к РНК-транскрипту, который комплементарен всему целевому первичному транскрипту, или мРНК, или ее части, который блокирует экспрессию целевого гена (патент США номер 5107065). Комплементарность антисмысловой РНК может наблюдаться с любым участком транскрипта определенного гена, т.е. с 5′-некодирующей последовательностью, 3′-некодирующей последовательностью, с интронами или с кодирующей последовательностью.
[0089] В настоящем описании, формулировка "функциональная РНК" относится к антисмысловой РНК, рибозиму РНК или другой РНК, которая может не транслироваться, но несмотря на это, влияет на клеточные процессы.
[0090] Термины "комплемент" и "обратный комплемент" используют взаимозаменяемо в настоящем описании применительно к мРНК-транскриптам, и они предназначены для определения антисмысловой РНК по отношению к информационной.
[0091] В данной заявке, формулировка "эндогенная РНК" относится к любой РНК, которая кодируется любой последовательностью нуклеиновых кислот, присутствующей в геноме хозяина, перед трансформированием рекомбинантной конструкцией согласно настоящему описанию, либо встречающейся в природе, либо не встречающейся в природе, т.е. введенной с помощью рекомбинантных средств, мутагенеза и т.д.
[0092] Подобие: Термин "подобие", если он используется для описания "подобия" между двумя последовательностями аминокислот, белками или полипептидами, относится к присутствию некоторого количества идентичных, а также консервативных аминокислотных остатков в обеих последовательностях. Чем выше степень подобия между двумя последовательностями аминокислот, тем больше соответствие, подобие или эквивалентность двух последовательностей.
[0093] Стабильная трансформация, временная трансформация и трансформация: В данной заявке, формулировка "стабильная трансформация" относится к переносу молекулы нуклеиновой кислоты в геном организма хозяина, включая как геном ядра, так и геном органелл, что приводит к генетически стабильному наследованию.
[0094] Наоборот, в данной заявке формулировка "временная трансформация" относится к переносу молекулы нуклеиновой кислоты в ядро или в содержащую ДНК органеллу организма хозяина, что приводит к экспрессии гена без встраивания в геном или стабильного наследования. Организмы хозяина, трансформированные молекулами нуклеиновых кислот, называют "трансгенными" организмами. Предпочтительный способ трансформирования клеток риса, кукурузы и других однодольных представляет собой способ трансформирования с помощью ускорения частиц или "генной пушки" (Klein и др., (1987) Nature (Лондон) 327:70 73; патент США номер 4945050), или способ, опосредованный Agrobacterium, с применением подходящей Ti-плазмиды, содержащей трансген (Ishida Y. и др., (1996) Nature Biotech. 14:745 750).
[0095] В данной заявке, термин "трансформация" относится как к стабильной трансформации, так и к временной трансформации.
[0096] Последовательность, направляющая трансляцию: В данной заявке, формулировка "последовательность, направляющая трансляцию" относится к последовательностям ДНК, расположенным между промоторной последовательностью гена и кодирующей последовательностью. Последовательность, направляющая трансляцию, присутствует в полностью процессированной мРНК и расположена против хода транскрипции от последовательности, инициирующей трансляцию. Последовательность, направляющая трансляцию, может влиять на процессинг первичного транскрипта до мРНК, на стабильность мРНК или на эффективность трансляции. Примеры направляющих трансляцию последовательностей описаны у Turner, R. и Foster, G. D. (1995) Molecular Biotechnology 3:225.
[0097] Все патенты, публикации патентов и приоритетные документы, цитированные в данном описании, настоящим полностью включены в данную заявку посредством ссылки. Ген дельта-9-элонгазы и кодируемый им фермент
[0098] Фермент, кодируемый геном дельта-9-элонгазы согласно настоящему описанию, незаменим для получения (с помощью альтернативного пути с участием дельта-8-десатуразы/дельта-9-элонгазы) длинноцепочечных полиненасыщенных жирных кислот (ДЦ-ПНЖК), имеющих длину 20 или более атомов углерода. Последовательность нуклеотидов изолированного гена дельта-9-элонгазы ССМР 2916 Euglena deses Ehr. показана на Фигуре 7А, и предсказанная последовательность аминокислот соответствующего белка показана на Фигуре 7 В.
[0099] Превращение ЛК в ДГЛК и АЛК в омега-3-ЭТК с помощью фермента дельта-9-элонгазы и фермента дельта-8-десатуразы называют альтернативным путем с участием дельта-8-десатуразы/дельта-9-элонгазы. В обычном дельта-6-пути превращения ЛК в ДГЛК и АЛК в омега-3-ЭТК, фермент дельта-6-десатуразу применяют для превращения ЛК в ГЛК и АЛК в СДК, а дельта-6-элонгазу применяют для превращения ГЛК в ДГЛК, и СДК в омега-3-ЭТК, соответственно. В каждом пути, получение АРК или ЭПК затем катализируется, например, дельта-5-десатуразой. ДГК, например, можно получить путем превращения ЭПК в омега-3-докозапентаеновую кислоту (ДПК), а затем омега-3-докозапентаеновой кислоты в ДГК, применяя, например, дельта-5-элонгазу и дельта-4-десатуразу, соответственно.
[00100] Хотя, например, ДГЛК, омега-3-ЭТК, АРК, ЭПК, омега-3-докозапентаеновую кислоту, омега-6-докозапентаеновую кислоту, АДК и/или ДГК можно получить с помощью любого из альтернативных путей с участием дельта-8-десатуразы/дельта-9-элонгазы или с помощью обычного дельта-6-пути, в некоторых случаях, альтернативный путь с участием дельта-8-десатуразы/дельта-9-элонгазы может оказаться предпочтительным по сравнению с обычным дельта-6-путем. Например, если нежелательно образование определенных остаточных интермедиатов омега-6 или омега-3 жирных кислот, таких как ГЛК или СДК, при получении ДГЛК, омега-3-ЭТК, АРК, ЭПК, омега-3-докозапентаеновой кислоты, омега-6-докозапентаеновой кислоты, АДК и/или ДГК, можно использовать альтернативный путь с участием дельта-8-десатуразы/дельта-9-элонгазы в качестве альтернативы обычному дельта-6-пути, чтобы избежать образования ГЛК и СДК.
[00101] Выше обсуждалось, что дельта-9-элонгаза представляет собой фермент, необходимый для альтернативного пути синтеза с участием дельта-8-десатуразы/дельта-9-элонгазы. ЭПК, например, не может быть синтезирована с помощью альтернативного пути с участием дельта-8-десатуразы/дельта-9-элонгазы в отсутствии гена дельта-9-элонгазы и фермента, кодируемого им. На Фигуре 1 показано, что изолированный фермент дельта-9-элонгаза согласно настоящему описанию превращает, например, АЛК в омега-3-ЭТрК и ЛК в омега-6-ЭДК. Получение омега-3-ЭТК из омега-3-ЭТрК и ЭПК из омега-3-ЭТК затем катализируется, например, дельта-8-десатуразой и дельта-5-десатуразой, соответственно. Благодаря использованию альтернативного пути с участием дельта-8-десатуразы/дельта-9-элонгазы, можно исключить образование интермедиатов жирных кислот ГЛК и СДК.
[00102] Следует отметить, что в объем настоящего описания также входят последовательности нуклеотидов (и соответствующие кодируемые ими белки), представляющие собой последовательности, включающие, состоящие или комплементарные по меньшей мере 68%, 69%, 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% нуклеотидов в последовательности (т.е. обладающие идентичностью последовательности) с SEQ ID NO:17 (т.е. изолированной последовательности нуклеотидов гена дельта-9-элонгазы ССМР 2916 Euglena deses Ehr.) или с SEQ ID NO:19 (т.е. вариантом гена дельта-9-элонгазы ССМР 2916 Euglena deses Ehr.). Такие последовательности можно получить от человека, а также из других, не относящихся к человеку источников (например, из С. elegans или мыши).
[00103] Более того, в объем настоящего описания также входят фрагменты и производные, включающие или состоящие из последовательности нуклеотидов SEQ ID NO:17 (показанной на Фигуре 7А) или SEQ ID NO:19 (показанной на Фигуре 8А)), а также из последовательностей, полученных из других источников и обладающих описанными выше свойствами комплементарности или соответствия. Функциональные эквиваленты описанных последовательностей (т.е. последовательностей, обладающих дельта-9-элонгазной активностью) также входят в объем настоящего описания.
[00104] Фрагменты, полученные из SEQ ID NO:17 или SEQ ID NO:19, могут включать или состоять из от 10 до приблизительно 780 нуклеотидов, от 10 до приблизительно 700 нуклеотидов, от 10 до приблизительно 650 нуклеотидов, от 10 до приблизительно 500 нуклеотидов, от 10 до приблизительно 250 нуклеотидов, от 10 до приблизительно 100 нуклеотидов, от 10 до приблизительно 50 нуклеотидов или от 15 до 40 нуклеотидов. В одном аспекте, фрагменты последовательностей SEQ ID NO:17 и SEQ ID NO:19 кодируют полипептид, обладающий дельта-9-элонгазной активностью. В другом аспекте, фрагменты последовательностей SEQ ID NO:17 и SEQ ID NO:19 можно применять в качестве праймеров и зондов. Способы получения праймеров и зондов хорошо известны специалистам в данной области. Длина таких праймеров и зондов может достигать от 10 до 50 нуклеотидов, предпочтительно от 15 до 40 нуклеотидов.
[00105] Варианты последовательностей нуклеотидов SEQ ID NO:17 или SEQ ID NO:19 также предусмотрены в данной заявке. Такие варианты могут включать вставки, замены или делеции одной или более пар оснований. Неограничивающие примеры вариантов последовательности нуклеотидов SEQ ID NO:17, входящих в объем настоящего описания, показаны в Таблице А ниже. Один конкретный пример варианта последовательности SEQ ID NO:17 представляет собой последовательность SEQ ID NO:19 (см. Фигуру 8А).
[00106] В объем настоящего описания также входят последовательности нуклеотидов из других источников, которые обладают описанными выше свойствами комплементарности или соответствия последовательности SEQ ID NO:17 или SEQ ID NO:19. Функциональные эквиваленты SEQ ID NO:17 или SEQ ID NO:19 (т.е. последовательности, обладающие дельта-9-элонгазной активностью) также входят в объем настоящего описания.
[00107] В объем настоящего описания также входят последовательности нуклеотидов или фрагменты указанных последовательностей, кодирующие полипептид, обладающий дельта-9-элонгазной активностью, при этом последовательность аминокислот указанного полипептида идентична на по меньшей мере 68%, 69%, 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% последовательности аминокислот, включающей SEQ ID NO:18 или SEQ ID NO:20. Такие последовательности можно получить из человека, а также из других не относящихся к человеку источников (например, из С.elegans или мыши).
[00108] В объем настоящего описания также входит изолированный и/или очищенный полипептид, который удлиняет полиненасыщенные жирные кислоты, содержащие элемент ненасыщенности в положении атома углерода 9 (т.е. обладает дельта-9-элонгазной активностью) и последовательность которого подобна или идентична по меньшей мере на 68%, 69%, 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% последовательности аминокислот SEQ ID NO:18 (показанной на Фигуре 7 В) или SEQ ID NO:20 (показанной на Фигуре 8 В)). В частности, в объем настоящего описания входит очищенный полипептид, обладающий последовательностью аминокислот SEQ ID NO:18 или SEQ ID NO:20.
[00109] Фрагменты полипептида, имеющего последовательность SEQ ID NO:18 или SEQ ID NO:20, также предусмотрены в данной заявке. Такие фрагменты могут включать или состоять из от 10 до приблизительно 260 последовательных аминокислот, от 10 до приблизительно 200 последовательных аминокислот, от 10 до приблизительно 100 последовательных аминокислот, от 10 до приблизительно 50 последовательных аминокислот, от 10 до приблизительно 40 последовательных аминокислот, от 10 до приблизительно 30 последовательных аминокислот или от 10 до приблизительно 20 последовательных аминокислот.
[00110] Варианты полипептида, имеющего последовательность SEQ ID NO:18 или SEQ ID NO:20, также предусмотрены в данной заявке. Такие варианты могут включать одну или более вставок, замен или делеций аминокислот. Неограничивающие примеры вариантов последовательности аминокислот SEQ ID NO:18, входящие в объем настоящего описания, показаны в Таблице В ниже. Один конкретный пример варианта последовательности SEQ ID NO:18 представляет собой SEQ ID NO:20 (см. Фигуру 8 В).
Получение фермента дельта-9-элонгазы
[00111] После того, как нуклеиновая кислота (например, ген), кодирующая фермент дельта-9-элонгазу была выделена и/или очищена, ее можно внедрить либо в прокариотическую, либо в эукариотическую клетку-хозяин с помощью вектора или конструкции. Указанный вектор, например, бактериофаг, космида или плазмида, может содержать последовательность нуклеотидов, кодирующую фермент дельта-9-элонгазу, а также любую регуляторную последовательность (например, промотор), которая способна функционировать в клетке-хозяине и способна вызывать экспрессию дельта-9-элонгазы, кодируемой указанной последовательностью нуклеотидов. Регуляторная последовательность находится в функциональной связи или функционально связана с последовательностью нуклеотидов (выше отмечено, что "регуляторный" означает "функционально связанный" с кодирующей последовательностью, если регуляторная последовательность влияет на транскрипцию или экспрессию кодирующей последовательности). Подходящие промоторы включают, например, промоторы генов, кодирующих алкогольдегидрогеназу, глицеральдегид-3-фосфатдегидрогеназу, фосфоглюкоизомеразу, фосфоглицераткиназу, кислую фосфатазу, промотор Т7, промотор TPI, промоторы генов, кодирующих лактазу, металлотионеин, предранний промотор цитомегаловируса, промоторы генов, кодирующих кислый белок молочной сыворотки, глюкоамилазу, и промоторы, активируемые в присутствии галактозы, например, GAL1 и GAL10. Кроме того, в вектор также можно ввести последовательности нуклеотидов, которые кодируют другие белки, олигосахариды, липиды и т.п., а также другие регуляторные последовательности, такие как сигнал полиаденилирования (например, сигнал полиаденилирования (поли-А) SV-40T-антигена, овальбумина или гормона роста крупного рогатого скота). Выбор последовательностей, присутствующих в конструкции, зависит от продуктов, которые необходимо экспрессировать, а также от природы клетки-хозяина.
[00112] Выше отмечено, что как только вектор был сконструирован, его можно внедрить в выбранную клетку-хозяин с помощью способов, известных средним специалистам в данной области, включая, например, трансфекцию, трансформацию и электропорацию (см. Molecular Cloning: A Laboratory Manual, 2oe изд., том 1-3, ред. Sambrook и др., Cold Spring Harbor Laboratory Press (1989)). Клетку-хозяина затем культивируют при подходящих условиях, позволяющих экспрессию генов, что приводит к продукции клеткой желательных ПНЖК, которые затем извлекают и очищают с помощью обычных методик, известных в данной области.
[00113] Примеры подходящих прокариотических клеток-хозяев включают, но не ограничены перечисленными, бактерии, такие как Escherichia coli. Bacillus subtilis, а также цианобактерии, такие как Spirulina spp. (т.е. сине-зеленые водоросли). Эукариотическая клетка, например, может представлять собой клетку млекопитающего, клетку насекомого, клетку растения или клетку гриба. Клетка гриба может представлять собой, например, Saccharomyces spp., Candida spp., Lipomyces spp., Yarrowia spp., Aspergillus spp., Penicillium spp., Neurospora spp., Kluyveromyces spp., Hansenula spp., Trichoderma spp.или Pichia spp. В частности, клетка гриба может представлять собой клетку дрожжей, например, такую как Saccharomyces spp., Candida spp., Hansenula spp. и Pichia spp. Клетка дрожжей также может представлять собой клетку Saccharomyces cerevisiae. Клетка растения включает, но не ограничена клетками масличных растений, таких как Glycine max (например, сои), видов Brassica, Carthamus tinctorius L. (например, сафлора), Helianthus annus (например, подсолнечника), Zea mays (например, кукурузы), видов Gossypium (например, хлопчатника) и Linum usitatissimum (например, льна).
[00114] Можно добиться временной или стабильной экспрессии в клетке-хозяине. Можно осуществить временную экспрессию с введенных конструкций, которые содержат сигналы экспрессии, функционирующие в клетке-хозяине, при этом указанные конструкции не реплицируются и редко встраиваются в клетку-хозяина, или если клетка-хозяин не пролиферирует. Временную экспрессию также можно осуществить путем стимуляции активности регулируемого промотора, функционально связанного с интересующим геном, хотя у таких индуцируемых систем часто обнаруживают низкий основной уровень экспрессии. Стабильной экспрессии можно добиться путем внедрения конструкции, которая может встраиваться в геном хозяина или которая автономно реплицируется в клетке-хозяине. Можно осуществить селекцию клеток, стабильно экспрессирующих интересующий ген, путем применения селектируемого маркера, расположенного на экспрессионной конструкции или трансфицированного вместе с ней, а затем селекции клеток, экспрессирующих указанный маркер. Если стабильная экспрессия происходит в результате встраивания, место встраивания конструкции в геном хозяина может быть произвольным, или его можно выбрать заранее путем применения конструкций, содержащих участки гомологии с геномом хозяина, подходящие для направленной рекомбинации с соответствующим локусом хозяина. Если конструкции предназначены для встраивания в эндогенный локус, все или некоторые из регуляторных областей транскрипции и трансляции могут обеспечиваться указанным эндогенным локусом.
[00115] Для экспрессии фермента дельта-9-элонгазы и, в конечном итоге, интересующей(их) ПНЖК, также можно использовать трансгенное млекопитающее. В частности, как только была создана описанная выше конструкция, ее можно внедрить в пронуклеус зародыша. Зародыша затем можно имплантировать в реципиентную самку. В качестве альтернативы, также можно применять способ передачи ядра (Schnieke и др., Science 278:2130-2133 (1997)). Затем позволяют беременности и родам идти естественным путем (см., например, патент США номер 5750176 и патент США номер 5,700,671). Образцы молока, тканей или других жидкостей организма потомства должны содержать измененные уровни ПНЖК, по сравнению с уровнями, обычно обнаруживаемыми у нетрансгенного животного. Затем можно контролировать продукцию измененных или повышенных уровней ПНЖК у последующих поколений и, таким образом, контролировать внедрение в их геномы гена, кодирующего желательный фермент десатуразу. Млекопитающее, используемое в качестве хозяина, можно выбрать из группы, состоящей, например, из мыши, крысы, кролика, свиньи, козы, овцы, лошади и коровы. Тем не менее, можно использовать любое млекопитающее, при условии, что оно обладает способностью принимать в свой геном ДНК, кодирующую интересующий фермент.
[00116] Для экспрессии полипептида дельта-9-элонгазы, функциональные участки инициации и терминации транскрипции и трансляции функционально связывают с ДНК, кодирующей полипептид элонгазы. Участки инициации и терминации транскрипции и трансляции получают из множества неисключительных источников, включая ДНК, которую нужно экспрессировать, гены, про которые известно или предполагается, что они способны экспрессироваться в желательной системе, векторы экспрессии, участки, полученные путем химического синтеза; или из эндогенного локуса клетки-хозяина. Экспрессия в ткани растения и/или части растения происходит с некоторой эффективностью, особенно если ткань или часть растения представляет собой такую ткань или часть, которую собирают рано, например, семя, листья, фрукты, цветы, корни и т.д. Экспрессию в определенной части растения можно обеспечить путем применения специфической регуляторной последовательности, такой как регуляторные последовательности, описанные в патентах США с номерами 5463174, 4943674, 5106739, 5175095, 5420034, 5188958 и 5589379.
[00117] В качестве альтернативы, экспрессированный белок может представлять собой фермент, который вызывает образование продукта, который может внедряться, либо непосредственно, либо после дополнительных модификаций, во фракцию сока растения-хозяина. Экспрессия гена дельта-9-элонгазы, или антисмысловых транскриптов дельта-9-элонгазы, может изменять уровни определенных ПНЖК, или их производных, обнаруживаемых в частях растения и/или в тканях растения.
[00118] Участок, кодирующий полипептид дельта-9-элонгазы, можно экспрессировать либо отдельно, либо совместно с другими генами (например, с геном, кодирующим дельта-8-десатуразу, геном, кодирующим дельта-5-десатуразу, геном, кодирующим дельта-17-десатуразу, геном, кодирующим дельта-5-элонгазу и/или геном, кодирующим дельта-4-десатуразу), для получения таких тканей и/или частей растения, которые содержат большие количества желательных ПНЖК, или в которых состав ПНЖК наиболее близко напоминает таковой в грудном молоке человека (см. WO 95/24494). Терминатор можно получить из 3′-области гена, из которого был получен участок инициации, или из отличного гена. Известно большое число терминаторов, которые оказались достаточно подходящими для множества хозяев того же и отличного рода и вида. Терминатор обычно выбирают с точки зрения удобства, а не благодаря какому-либо конкретному свойству.
[00119] Выше отмечено, что растение (например, Glycine max (сою) или Brassica napus (канолу)) или ткань растения также можно использовать в качестве хозяина или клетки-хозяина, соответственно, для экспрессии фермента дельта-9-элонгазы, который, в свою очередь, можно применять для получения полиненасыщенных жирных кислот. В частности, желательные ПНЖК можно экспрессировать в семенах. Способы получения масел из семян известны в данной области. Таким образом, в дополнение к обеспечению источника ПНЖК, можно влиять на компоненты масел из семян путем экспрессии гена дельта-9-элонгазы, а также, возможно, генов десатураз (например, дельта-8-десатуразы, дельта-17-десатуразы, дельта-5-десатуразы, дельта-4-десатуразы и т.д.) и генов других элонгаз (например, дельта-5-элонгазы и т.д.), чтобы получить масла из семян, которые можно добавить в пищевые композиции, фармацевтические композиции, корма для животных и косметические средства. Опять же, вектор, который содержит последовательность ДНК, кодирующую дельта-9-элонгазу, функционально связанную с промотором, будут вводить в ткань растения или растение на некоторое время и при условиях, достаточных для экспрессирования гена дельта-9-элонгазы. Указанный вектор также может содержать один или более генов, которые кодируют другие ферменты, например, дельта-4-десатуразу, дельта-5-десатуразу, дельта-6-десатуразу, дельта-10-десатуразу, дельта-12-десатуразу, дельта-15-десатуразу, дельта-17-десатуразу, дельта-19-десатуразу, дельта-6-элонгазу и/или дельта-5-элонгазу. Ткань растения или растение могут продуцировать подходящий субстрат, на который действуют ферменты, или можно ввести в ткань растения, клетку растения или растение вектор, кодирующий ферменты, который продуцирует такие субстраты. В дополнение, субстрат можно распылить на ткани растения, экспрессирующие подходящие ферменты. Путем применения этих различных методик, можно получить ПНЖК, используя клетки растения, ткань растения или растение. Также следует отметить, что в объем настоящего описания также входит трансгенное растение, содержащее описанный выше вектор, в котором экспрессия последовательности нуклеотидов указанного вектора приводит к получению полиненасыщенной жирной кислоты, например, в семенах трансгенного растения.
[00120] Регенерация, развитие и культивирование растений из отдельных трансформированных протопластов растения или из различных трансформированных эксплантатов хорошо известно в данной области (Weissbach и Weissbach, Methods/or Plant Molecular Biology, (ред.). Academic Press, Inc., Сан-Диего, Калифорния, (1988)). Данный процесс регенерации и роста обычно включает этапы селекции трансформированных клеток, культивирования этих отдельных клеток путем прохождения ими обычных стадий эмбрионального развития, в частности, стадии коренящегося ростка. Трансгенные зародыши и семена регенерируют аналогичным способом. Полученные в результате этого трансгенные коренящиеся побеги впоследствии высаживают в подходящую среду для выращивания растений, такую как почва.
[00121] Развитие или регенерация растений, содержащих чужеродный, экзогенный ген, который кодирует интересующий белок, хорошо известна в данной области. Предпочтительно, регенерированные растения подвергают самоопылению для получения гомозиготных трансгенных растений. В противном случае, пыльцу, полученную из регенерированных растений, скрещивают с выращиваемыми из семян растениями агрономически важных линий. В качестве альтернативы, пыльцу из растений данных важных линий используют для опыления регенерированных растений. Трансгенное растение согласно настоящему описанию, содержащее желательный полипептид, культивируют, применяя способы, хорошо известные специалисту в данной области.
[00122] Существует множество способов регенерации растений из ткани растения. Конкретный способ регенерации будет зависеть от исходной ткани растения и конкретного вида растения, которое нужно регенерировать.
[00123] Способы трансформирования двудольных растений, главным образом, путем применения Agrobacterium tumefaciens, и получения трансгенных растений были опубликованы для хлопчатника (патент США номер 5004863, патент США номер 5159135, патент США номер 5518908); сои (патент США номер 5569834, патент США номер 5416011, McCabe и др., BiolTechnology 6:923 (1988), Christou и др., Plant Physiol. 87:671 674 (1988)); Brassica (патент США номер 5463174); земляного ореха (Cheng и др., Plant Cell Rep.15:653 657 (1996), McKently, и др., Plant Cell Rep. 14:699 703 (1995)); папайи и гороха (Grant и др., Plant Cell Rep.15:254 258, (1995)).
[00124] Также было опубликовано трансформирование однодольных растений с применением электропорации, бомбардировки частицами и Agrobacterium. Трансформирования и регенерации растений удалось добиться для спаржи (Bytebier и др., Proc. Natl. Acad. Sci. (США) 84:5354, (1987)); ячменя (Wan и Lemaux, Plant Physiol 104:37 (1994)); Zea mays (Rhodes и др., Science 240:204 (1988), Gordon-Kamm и др., Plant Cell, 2:603 618 (1990), Fromm и др., BiolTechnology 8:833 (1990), Koziel и др., BiolTechnology 11:194, (1993), Armstrong и др., Crop Science 35:550 557 (1995)); овса (Somers и др., BiolTechnology 10: 15 89 (1992)); ежи сборной (Нот и др., Plant Cell Rep.7:469 (1988)); риса (Toriyama и др., TheorAppl. Genet. 205:34, (1986); Part и др.. Plant Mol. Biol. 32:1135 1148, (1996); Abedinia и др., Aust. J. Plant Physiol. 24:133 141 (1997); Zhang и Wu, Theor. Appl. Genet. 76:835 (1988); Zhang и др. Plant Cell Rep. 7:379, (1988); Battraw и Hall, Plant Sci. 86:191 202 (1992); Christou и др., Bio/Technology 9:957 (1991)); ржи (De la Репа и др., Nature 325:274 (1987)); сахарного тростника (Bower и Birch, Plant J. 2:409 (1992)); овсяницы гигантской (Wang и др., BiolTechnology 10:691 (1992)) и пшеницы (Vasil и др., Bio/Technology 10:667 (1992); патент США номер 5631152).
[00125] Были разработаны анализы экспрессии генов, исходя из временной экспрессии клонированных конструкций нуклеиновых кислот, путем введения молекул нуклеиновых кислот в клетки растения посредством обработки полиэтиленгликолем, электропорации или бомбардировки частицами (Marcotte и др.. Nature 335:454 457 (1988); Marcotte и др., Plant Cell 1:523 532 (1989); McCarty и др., Cell 66:895 905 (1991); Hattori и др., Genes Dev. 6:609 618 (1992); Goff и др., EMBOJ. 9:2517 2522 (1990)).
[00126] Можно применять системы временной экспрессии для делеционного анализа генных конструкций (см., в основном, Maliga и др., Methods in Plant Molecular Biology, Cold Spring Harbor Press (1995)). Должно быть очевидно, что любые из молекул нуклеиновых кислот согласно настоящему описанию можно ввести в клетку растения для постоянной или временной экспрессии в комбинации с другими генетическими элементами, такими как векторы, промоторы, энхансеры и т.д.
[00127] В дополнение к описанным выше процедурам, практикующие специалисты также знакомы со стандартными источниками, в которых описаны конкретные условия и процедуры для конструирования, манипулирования и выделения макромолекул (например, молекул ДНК, плазмид и т.д.), получения рекомбинантных организмов и скрининга и выделения клонов (см., например, Sambrook и др., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Press (1989); Maliga и др., Methods in Plant Molecular Biology, Cold Spring Harbor Press (1995); Birren и др., Genome Analysis: Detecting Genes, 1, Колд Спринг Харбор, Нью-Йорк (1998); Birren и др., Genome Analysis: Analyzing DNA, 2, Колд Спринг Харбор, Нью-Йорк (1998); Plant Molecular Biology: A Laboratory Manual, ред. dark, Springer, Нью-Йорк (1997)).
[00128] Субстраты, которые может продуцировать клетка-хозяин либо по природе, либо в результате введения трансгена, а также ферменты, которые могут кодироваться последовательностями ДНК, присутствующими в векторе, который впоследствии вводят в клетку-хозяина, показаны на Фигуре 1.
[00129] Принимая во внимание вышесказанное, в объем настоящего описания входит способ получения фермента дельта-9-элонгазы, включающий следующие этапы: 1) выделение последовательности нуклеотидов, которая включает или комплементарна по меньшей мере 68% последовательности нуклеотидов, кодирующей фермент дельта-9-элонгазу (например, последовательности нуклеотидов, выбранной из группы, состоящей из SEQ ID NO:17 и SEQ ID NO:19); 2) конструирование вектора экспрессии, содержащего указанную последовательность нуклеотидов, функционально связанную с регуляторной последовательностью; и 3) введение указанного вектора в клетку-хозяина на время и при условиях, которые достаточны для получения фермента дельта-9-элонгазы.
[00130] В объем настоящего описания также входит способ получения полиненасыщенных жирных кислот. В одном аспекте, указанный способ включает: 1) выделение последовательности нуклеотидов, которая включает или комплементарна по меньшей мере 68% последовательности нуклеотидов, кодирующей фермент дельта-9-элонгазу (например, последовательности нуклеотидов, выбранной из группы, состоящей из SEQ ID NO:17 и SEQ ID NO:19); 2) конструирование вектора экспрессии, содержащего указанную последовательность нуклеотидов, функционально связанную с регуляторной последовательностью; 3) введение указанного вектора экспрессии в клетку-хозяина на время и при условиях, достаточных для получения фермента дельта-9-элонгазы; и 4) обеспечение взаимодействия указанной экспрессированной дельта-9-элонгазы с полиненасыщенной жирной кислотой субстратом с тем, чтобы превратить указанную полиненасыщенную жирную кислоту-субстрат в первую полиненасыщенную жирную кислоту-продукт. Примеры ПНЖК субстрата включают ЛК, АЛК и комбинации перечисленных жирных кислот. Примеры первой полиненасыщенной жирной кислоты-продукта, которую можно получить с помощью данного способа, представляют собой омега-6-ЭДК, омега-3-ЭТрК или обе жирные кислоты омега-6-ЭДК и омега-3-ЭТрК. Например, если ЛК подвергают воздействию фермента дельта-9-элонгазы, она превращается в омега-6-ЭДК. В другом примере, если АЛК подвергают воздействию фермента дельта-9-элонгазы, она превращается в омега-3-ЭТрК.
[00131] Указанный способ может дополнительно включать этап(ы) воздействия на указанную первую полиненасыщенную жирную кислоту-продукт по меньшей мере одной десатуразы, по меньшей мере одной дополнительной элонгазы или комбинациями перечисленных ферментов, и возможно повторения данного этапа (т.е. воздействие на второй или последующие полиненасыщенные жирные кислоты-продукты десатуразы или элонгазы (которая может быть такой же или отличной от любой использованной ранее десатуразы или элонгазы) для превращения указанной первой полиненасыщенной жирной кислоты-продукта во вторую или последующую (например, третью, четвертую, пятую, шестую и т.д.) полиненасыщенную жирную кислоту-продукт). Данный этап можно повторить столько раз, сколько необходимо, до тех пор, пока не получат желательную полиненасыщенную жирную кислоту-продукт. Например, если первая полиненасыщенная жирная кислота-продукт представляет собой омега-6-ЭДК, указанный способ может дополнительно включать воздействие на омега-6-ЭДК, например, дельта-8-десатуразой, которая превращает омега-6-ЭДК в ДГЛК (вторая полиненасыщенная жирная кислота-продукт). ДГЛК затем при необходимости можно превратить в АРК (третью полиненасыщенную жирную кислоту-продукт) путем воздействия на ДГЛК, например, дельта-5-десатуразой. АРК затем можно подвергнуть воздействию дельта-17-десатуразы для получения ЭПК (четвертой полиненасыщенной жирной кислоты-продукта). Более того, при необходимости ЭПК можно подвергнуть воздействию дельта-5-элонгазы для получения ДПК (пятой полиненасыщенной жирной кислоты- продукта). ДПК затем при необходимости можно подвергнуть воздействию дельта-4-десатуразы для получения ДГК (шестой полиненасыщенной жирной кислоты-продукта). В другом примере, если первая полиненасыщенная жирная кислота- продукт представляет собой омега-3-ЭТрК, указанный способ может дополнительно включать воздействие на омега-3-ЭТрК, например, дельта-8-десатуразой, которая превращает омега-3-ЭТрК в ЭТК (вторую полиненасыщенную жирную кислоту-продукт). ЭТК затем можно превратить в ЭПК (третью полиненасыщенную жирную кислоту-продукт) путем воздействия на ЭТК, например, дельта-5-десатуразой. ЭПК можно дополнительно превратить в другие полиненасыщенные жирные кислоты, описанные выше.
[00132] В другом аспекте, указанный способ включает: 1) выделение последовательности нуклеотидов, которая включает или комплементарна по меньшей мере 68% последовательности нуклеотидов, кодирующей фермент дельта-9-элонгазу (например, последовательности нуклеотидов, выбранной из группы, состоящей из SEQ ID NO:17 и SEQ ID NO:19); 2) конструирование вектора экспрессии, содержащего указанную выделенную последовательность нуклеотидов, функционально связанную с регуляторной последовательностью; 3) введение указанного вектора экспрессии и по меньшей мере одной дополнительной конструкции рекомбинантной ДНК, содержащей выделенную последовательность нуклеотидов, кодирующую дельта-8-десатуразу и функционально связанную с по меньшей мере одной регуляторной последовательностью, в клетку-хозяина на время и при условиях, которые достаточны для экспрессии дельта-9-элонгазы и дельта-8-десатуразы; и 4) обеспечение взаимодействия указанной экспрессированной дельта-9-элонгазы и дельта-8-десатуразы с полиненасыщенной жирной кислотой-субстратом, выбранной из группы, состоящей из ЛК, АЛК и комбинации перечисленных жирных кислот с тем, чтобы превратить указанную полиненасыщенную жирную кислоту-субстрат в первую полиненасыщенную жирную кислоту-продукт. Примеры первой полиненасыщенной жирной кислоты-продукта включают ДГЛК, омега-3-ЭТК и комбинации перечисленных жирных кислот. Более того, указанный способ может дополнительно включать этап(ы) воздействия на первую полиненасыщенную жирную кислоту-продукт по меньшей мере одной дополнительной десатуразы или по меньшей мере одной дополнительной элонгазы и, возможно, повторения данного этапа (то есть, осуществления воздействия на указанные вторую или последующую полиненасыщенную жирную кислоту-продукт десатуразы или элонгазы (которая может быть такой же или отличной от любой десатуразы или элонгазы, использованной ранее)) для превращения первой полиненасыщенной жирной кислоты-продукта (например, ДГЛК и/или омега-3-ЭТК) во вторую или последующую (например, третью, четвертую, пятую, шестую и т.д.) полиненасыщенную жирную кислоту-продукт. Данный этап можно повторить столько раз, сколько необходимо, до тех пор, пока не получат желательную полиненасыщенную жирную кислоту-продукт. В одном аспекте, указанный способ дополнительно включает введение в указанную клетку-хозяина конструкции рекомбинантной ДНК, содержащей выделенную последовательность нуклеотидов, кодирующую дельта-5-десатуразу, функционально связанную с регуляторной последовательностью.
[00133] Таким образом, дельта-9-элонгазу можно применять для получения полиненасыщенных жирных кислот, которые, в свою очередь, можно применять для определенных полезных целей или можно применять для получения других ПНЖК. Применения гена дельта-9-элонгазы
[00134] Выше отмечено, что выделенный ген дельта-9-элонгазы и фермент дельта-9-элонгаза, кодируемый данным геном, могут иметь множество применений. Например, указанный ген и соответствующий фермент можно применять непосредственно или опосредованно получения полиненасыщенных жирных кислот, например, дельта-9-элонгазу можно применять для получения омега-6-ЭДК, омега-3-ЭТрК, ДГЛК, омега-3-ЭТК, АРК, ЭПК, омега-3-докозапентаеновой кислоты, омега-6-докозапентаеновой кислоты, АДК и/или ДГК. Подразумевают, что в объем термина "непосредственно" входит случай, когда фермент непосредственно превращает указанную кислоту в другую кислоту, последнюю из которых используют в композиции (например, превращение ЛК в омега-6-ЭДК). Подразумевают, что в объем термина "опосредованно" входит случай, когда кислота превращается в другую кислоту (т.е. в промежуточный продукт пути синтеза) под воздействием фермента (например, ЛК в омега-6-ЭДК), а затем последнюю кислоту превращают в другую кислоту с помощью фермента, не являющегося элонгазой (например, омега-6-ЭДК в ДГЛК, например, с помощью дельта-8-десатуразы). Данные полиненасыщенные жирные кислоты (т.е. жирные кислоты, полученные либо непосредственно, либо опосредованно, с помощью активности фермента дельта-9-элонгазы) можно добавить, например, в пищевые композиции, фармацевтические композиции, косметические средства и корма для животных, при этом все из перечисленных входят в объем настоящего описания. Данные применения подробно описаны ниже.
Пищевые композиции
[00135] Настоящее описание включает пищевые композиции. Такие композиции, для целей настоящего описания, включают любой продукт питания или препарат для потребления человеком, включая энтеральное или парентеральное потребление, который при приеме внутрь (а) служит для питания или формирования тканей или в качестве источника энергии и/или (b) для сохранения, восстановления или поддержания достаточного пищевого статуса или метаболической функции.
[00136] Питательная композиция согласно настоящему описанию содержит по меньшей мере одно масло или кислоту, полученную непосредственно или опосредованно путем применения гена дельта-9-элонгазы, описанного в данной заявке, и может находиться либо в твердой, либо в жидкой форме. Кроме того, указанная композиция может включать съедобные питательные макроэлементы, витамины и минералы в количествах, необходимых для конкретного применения. Количество таких ингредиентов будет изменяться в зависимости от того, предполагается ли применение указанной композиции нормальными, здоровыми младенцами, детьми или взрослыми с определенными потребностями, например которые сопутствуют некоторым метаболическим состояниям (например, метаболическим расстройствам).
[00137] Примеры питательных макроэлементов, которые можно добавить в указанную композицию, включают, но не ограничены перечисленными: съедобные жиры, углеводы и белки. Примеры таких съедобных жиров включают, но не ограничены перечисленными: кокосовое масло, соевое масло и моно- и диглицериды. Примеры таких углеводов включают, но не ограничены перечисленными: глюкозу, съедобную лактозу и гидролизованный крахмал. Кроме того, примеры белков, которые можно использовать в питательной композиции согласно настоящему описанию, включают, но не ограничены перечисленными: соевые белки, электродиализированную молочную сыворотку, электродиализированное обезжиренное молоко, молочную сыворотку или гидролизаты данных белков.
[00138] Что касается витаминов и минералов, в пищевые композиции согласно настоящему описанию можно добавить следующие из них: кальций, фосфор, калий, натрий, хлорид, магний, марганец, железо, медь, цинк, селен, йод и витамины А, Е, D, С и комплекс витаминов группы В. Также можно добавить другие подобные витамины и минералы.
[00139] Компоненты, применяемые в пищевых композициях согласно настоящему описанию, должны быть полуочищенными или очищенными. Под полуочищенным или очищенным подразумевают материал, который получили путем очистки природного материала или с помощью синтеза.
[00140] Примеры пищевых композиций согласно настоящему описанию включают, но не ограничены перечисленными: составы для младенцев, пищевые добавки, пищевые заменители и композиции для регидратации. Пищевые композиции, представляющие особый интерес, включают, но не ограничены перечисленными: композиции, используемые для энтеральной и парентеральной подкормки младенцев, специальные составы для младенцев, добавки для пожилых и добавки для субъектов с затрудненным пищеварением и/или пониженным всасыванием.
[00141] Питательную композицию согласно настоящему описанию также можно добавить в продукт питания, даже если дополнение к диете не требуется. Например, композицию можно добавить в продукт питания любого типа, включая, но не ограничиваясь перечисленными: маргарины, модифицированные масла, сыры, молоко, йогурт, шоколад, конфету, закуски, салатные масла, кулинарные масла, кулинарные жиры, мясо, рыбу и напитки.
[00142] В предпочтительном варианте реализации согласно настоящему описанию, питательная композиция представляет собой энтеральный питательный продукт, более предпочтительно энтеральный питательный продукт для применения взрослыми или детьми. Данную композицию можно вводить взрослым или детям, подверженным стрессу или обладающим определенными потребностями вследствие хронических или острых болезненных состояний. Указанная композиция может включать, в дополнение к полиненасыщенным жирным кислотам, полученным в соответствии с настоящим описанием, питательные макроэлементы, витамины и минералы, описанные выше. Питательные макроэлементы могут присутствовать в количествах, эквивалентных присутствующим в молоке человека или эквивалентных в пересчете на энергию, т.е. в пересчете на калории.
[00143] Способы составления жидких или твердых энтеральных и парентеральных питательных формул хорошо известны в данной области.
[00144] Энтеральную формулу, например, можно стерилизовать и впоследствии использовать как готовую к употреблению (RTF) или хранить в виде концентрированной жидкости или порошка. Порошок можно получить путем распылительной сушки формулы, полученной, как указано выше, а затем ресуспендировать путем регидратации концентрата. Питательные формулы для применения взрослыми или детьми хорошо известны в данной области и доступны для приобретения (например, в Similac®, Ensure®, Jevity® и Alimentum® из подразделения Ross Products Division, Abbott Laboratories, Колумбус, Огайо). Масло или кислоту, полученную в соответствии с настоящим описанием, можно добавить в любую из данных формул.
[00145] Энергетическая плотность пищевых композиций согласно настоящему описанию, находящихся в жидкой форме, может находиться в диапазоне от приблизительно 0,6 ккал до приблизительно 3 ккал на мл. В твердой или порошкообразной форме, питательные добавки могут включать от приблизительно 1,2 до более чем 9 ккал на грамм, предпочтительно приблизительно от 3 до 7 ккал на грамм. Обычно осмоляльность жидкого продукта должна быть меньше, чем 700 миллиосмоль и, более предпочтительно, меньше, чем 660 миллиосмоль.
[00146] Питательная формула может включать описанные выше питательные макроэлементы, витамины и минералы, в дополнение к ПНЖК, полученным в соответствии с настоящим описанием. Присутствие данных дополнительных компонентов помогает индивиду удовлетворять минимальную ежедневную потребность в данных элементах. В дополнение к обеспечению ПНЖК, также может быть желательно добавление в композицию цинка, меди, фолиевой кислоты и антиоксид антов. Полагают, что данные вещества стимулируют подверженную стрессам иммунную систему и, следовательно, обеспечат дополнительную пользу для индивида, принимающего указанную композицию. В фармацевтическую композицию также можно добавить данные элементы.
[00147] В более предпочтительном варианте реализации, питательная композиция включает, в дополнение к антиоксидантам и по меньшей мере одной ПНЖК, источник углевода, при этом по меньшей мере 5 процентов массы указанного углевода составляет неусвояемый олигосахарид. В более предпочтительном варианте реализации, питательная композиция дополнительно включает белок, таурин и карнитин.
[00148] Выше отмечено, что ПНЖК, полученные в соответствии с настоящим описанием, или производные указанных ПНЖК, можно добавить в пищевые заменители или добавки, особенно, в формулу для младенцев, для пациентов, получающих питание внутривенно, или для предупреждения или лечения истощения или других состояний или болезненных состояний. В качестве теоретической основы, следует отметить, что грудное молоко человека обладает профилем жирных кислот, включающим от приблизительно 0,15% до приблизительно 0,36% ДГК, от приблизительно 0,03% до приблизительно 0,13% ЭПК, от приблизительно 0,30% до приблизительно 0,88% АРК, от приблизительно 0,22% до приблизительно 0,67% ДГЛК и от приблизительно 0,27% до приблизительно 1,04% ГЛК. Таким образом, жирные кислоты, такие как АРК, ЭПК и/или ДГК, полученные в соответствии с настоящим описанием, можно применять для изменения, например, композиции в составах для младенцев, чтобы лучше воспроизвести содержание ПНЖК в грудном молоке человека, или для изменения присутствующих ПНЖК, обычно обнаруживаемых в молоке не относящегося к человеку млекопитающего. В частности композиция для применения в фармакологической или пищевой добавке, особенно в заменителе грудного молока или добавке к нему, будет предпочтительно включать одну или более из АРК, ЭПК, ДГЛК и ДГК. Более предпочтительно, масло будет включать от приблизительно 0,3 до 30% АРК и от приблизительно 0,2 до 30% ДГЛК.
[00149] Парентеральные пищевые композиции, содержащие от приблизительно 2 до приблизительно 30 процентов массы жирных кислот, в пересчете на триглицериды, входят в объем настоящего описания. Возможно могут быть включены другие витамины, особенно жирорастворимые витамины, такие как витамин A, D, Е и L-карнитин. При необходимости, можно добавить консервант, такой как альфа-токоферол, в количестве, составляющем приблизительно 0,1% по массе. [00150] В дополнение, соотношения АРК и ДГЛК могут быть пригодными для данного конкретного конечного применения. При составлении лекарственной формы в виде заменителя грудного молока или добавки к нему, будет предусмотрена композиция, которая содержит одну или более из АРК, ДГЛК и ГЛК, в соотношении от приблизительно 1:19:30 до приблизительно 6:1:0,2, соответственно. Например, грудное молоко животных может различаться по соотношению АРК:ДГЛК:ГЛК в диапазоне от 1:19:30 до 6:1:0,2, который включает промежуточные соотношения, которые предпочтительно составляют приблизительно 1:1:1, 1:2:1, 1:1:4. При совместном получении в клетке-хозяине, можно изменять скорость и процент превращения субстрата-предшественника, такого как ЭДК и ДГЛК, в АРК, чтобы точно контролировать соотношения ПНЖК. Например, скорость превращения ДГЛК в АРК от 5% до 10% можно применять для получения соотношения АРК к ДГЛК, равного приблизительно 1:19, тогда как скорость превращения, равную приблизительно от 75% до 80%, можно применять для получения соотношения АРК к ДГЛК, равного приблизительно 6:1. Следовательно, как в системе культуры клеток, так и в животном- хозяине можно регулировать время, степень и специфичность экспрессии элонгазы, а также экспрессии десатуразы (такой как, но не ограничиваясь дельта-8-десатуразой) и других элонгаз, для модулирования уровней и соотношений ПНЖК. ПНЖК/кислоты, полученные в соответствии с настоящим описанием (например, АРК и ЭПК), затем можно объединить с другими ПНЖК/кислотами (например, ДГЛК) в желательных концентрациях и соотношениях.
[00151] Дополнительно, ПНЖК, полученные в соответствии с настоящим описанием, или клетки-хозяева, содержащие ПНЖК, также можно применять в качестве пищевых добавок для животных, чтобы изменить состав жирных кислот в ткани или молоке животного на более желательный для потребления человеком или животным. [00152] Примеры некоторых питательных добавок, составов для младенцев, питательных заменителей и других питательных растворов, в которых используются полиненасыщенные жирные кислоты, полученные согласно настоящему описанию, приведены ниже.
[0153] Составы для младенцев: Примеры составов для младенцев включают, но не ограничены перечисленными: состав с железом Isomil® Soy Formula with Iron, состав от диареи Isomil® DF Soy Formula For Diarrhea, состав с железом Isomil® Advance® Soy Formula with Iron, готовый к употреблению состав с железом Isomil® Advance® 20 Soy Formula With Iron Ready To Feed, состав для младенцев Similac® Infant Formula, состав для младенцев с железом Similac® Advance® Infant Formula with Iron, состав для младенцев с железом Similac® NeoSure® Advance® Infant Formula With Iron, готовый к употреблению, обогащающей грудное молоко человека с низким содержанием железа, состав Similac Natural Care Advance Low-Iron Human Milk Fortifier Ready To Use, каждый из которых доступен для приобретения в Abbott Nutrition (Колумбус, Огайо). Различные ПНЖК согласно настоящему описанию можно заменить и/или добавить в формулы для младенцев, описанные в данной заявке, и в другие формулы для младенцев, известные специалистам в данной области.
[00154] Питательные составы: Примеры питательных составов включают, но не ограничены перечисленными: ENSURE®, ENSURE® HIGH PROTEIN, ENSURE PLUS®, ENSURE® POWDER, ENSURE® PUDDING, ENSURE® WITH FIBER, Oxepa™ Nutritional Product, каждый из которых доступен для приобретения в Abbott Nutrition (Колумбус, Огайо). Различные питательные добавки, описанные выше и известные специалистам в данной области, можно заменить и/или дополнить ПНЖК, полученными в соответствии с настоящим описанием.
Фармацевтические композиции
[00155] В объем настоящего описания также входит фармацевтическая композиция, содержащая одну или более кислот и/или масел, полученных с применением генов дельта-9-элонгазы, описанных в данной заявке, в соответствии со способами, описанными в данной заявке. В частности, такая фармацевтическая композиция может включать одну или более кислот и/или масел, а также стандартный, хорошо известный, нетоксичный фармацевтически приемлемый носитель, адъювант или среду, такую как, например, фосфатно-солевой буферный раствор, вода, этанол, полиолы, растительные масла, смачивающий агент или эмульсия, такая как эмульсия вода/масло. Композиция может находиться либо в жидкой, либо в твердой форме. Например, композиция может быть в виде таблетки, капсулы, проглатываемой жидкости или порошка, инъецируемого состава или топической мази или крема. Подходящую текучесть можно поддерживать, например, путем обеспечения необходимого размера частиц в случае дисперсии и путем применения поверхностно-активных веществ. Также может оказаться желательным включение изотонических агентов, например, сахаров, хлорида натрия и тому подобных. Помимо таких инертных разбавителей, композиция также может содержать адъюванты, такие как смачивающие агенты, эмульгирующие и суспендирующие агенты, подслащивающие агенты, ароматизирующие агенты и отдушки.
[00156] Суспензии, в дополнение к активным соединениям, могут включать суспендирующие агенты, такие как, например, этоксилированные изостеариловые спирты, полиоксиэтиленсорбит и сложные эфиры сорбитана, микрокристаллическая целлюлоза, метагидроксид алюминия, бентонит, агар-агар и трагакант или смеси указанных веществ.
[00157] Твердые лекарственные формы, такие как таблетки и капсулы, можно получить, применяя методики, хорошо известные в данной области. Например, ПНЖК, полученные в соответствии с настоящим описанием, можно таблетировать вместе с обычной основой таблетки, такой как лактоза, сахароза и кукурузный крахмал, в комбинации со связующими веществами, такими как гуммиарабик, кукурузный крахмал или желатин, дезинтегрирующими агентами, такими как картофельный крахмал или альгиновая кислота, и лубрикантом, таким как стеариновая кислота или стеарат магния. Капсулы можно получить путем включения данных вспомогательных веществ в желатиновую капсулу вместе с антиоксидантами и соответствующей(ими) ПНЖК. Компоненты антиоксиданты и ПНЖК должны соответствовать описанным выше нормам.
[00158] Для внутривенного введения, ПНЖК, полученные в соответствии с настоящим описанием, или производные ПНЖК, можно включить в коммерческие составы, такие как Intralipids™. Типичный профиль жирных кислот в нормальной плазме взрослого человека включает от 6,64 до 9,46% АРК, от 1,45 до 3,11% ДГЛК и от 0,02 до 0,08% ГЛК. Данные ПНЖК или их метаболические предшественники можно вводить отдельно или в комбинации с другими ПНЖК, чтобы добиться нормального профиля жирных кислот у пациента. При необходимости, отдельные компоненты составов могут предоставляться отдельно, в виде набора, для однократного или многократного применения. Типичная дозировка конкретной жирной кислоты находится в диапазоне от 0,1 мг до 20 г (до 100 г) ежедневно и предпочтительно составляет от 10 мг до 1,2, 5 или 10 г ежедневно.
[00159] Возможные пути введения фармацевтических композиций согласно настоящему описанию включают, например, энтеральный (например, пероральный и ректальный) и парентеральный путь. Например, жидкую композицию можно вводить, например, перорально или ректально. Кроме того, гомогенную смесь можно полностью диспергировать в воде, смешать при стерильных условиях с физиологически приемлемыми разбавителями, консервантами, буферами или пропеллентами с получением спрея или ингалятора.
[00160] Путь введения, конечно, будет зависеть от требуемого эффекта. Например, если указанную композицию используют для лечения грубой, сухой или постаревшей кожи, для лечения поврежденной или обожженной кожи или для лечения кожи или волос, пораженных болезнью или патологическим состоянием, возможно, ее можно применять топически.
[00161] Дозировку композиции для введения пациенту может определить средний специалист в данной области, и она будет зависеть от различных факторов, таких как вес пациента, возраст пациента, иммунный статус пациента и т.д.
[00162] Что касается формы, указанная композиция может представлять собой, например, раствор, дисперсию, суспензию, эмульсию или стерильный порошок, который затем растворяют.
[00163] В настоящее описание также входит лечение различных расстройств путем применения фармацевтических и/или пищевых композиций, описанных в данной заявке. В частности, композиции согласно настоящему описанию можно применять для лечения рестеноза после ангиопластики. Более того, симптомы воспаления, ревматоидного артрита, астмы и псориаза также можно лечить с помощью композиций согласно настоящему описанию. Также существуют доказательства того, что ПНЖК могут участвовать в метаболизме кальция; таким образом, композиции согласно настоящему описанию, возможно, можно применять для лечения или предупреждения остеопороза и камней в почках или мочевыводящих путях.
[00164] Дополнительно, композиции согласно настоящему описанию также можно применять для лечения рака. Было показано, что злокачественные клетки имеют измененный состав жирных кислот. Показали, что добавление жирных кислот замедляет их рост, вызывает гибель клеток и повышает их чувствительность к химиотерапевтическим агентам. Более того, композиции согласно настоящему описанию также можно применять для лечения кахексии, связанной с раком.
[00165] Композиции согласно настоящему описанию также можно применять для лечения диабета (см. патент США номер 4826877 и Horrobin и др., Am. J. din. Nutr. 1993) том 57 (прил.) 732S-737S). Измененный метаболизм и состав жирных кислот продемонстрировали у страдающих диабетом животных.
[00166] Более того, композиции согласно настоящему описанию, содержащие ПНЖК, полученные либо непосредственно, либо опосредованно путем применения фермента дельта-9-элонгазы, также можно применять для лечения экземы, для снижения кровяного давления и для улучшения экзаменационных оценок по математике. Кроме того, композиции согласно настоящему описанию можно применять для ингибирования агрегации тромбоцитов, расширения кровеносных сосудов, снижения уровней холестерина, ингибирования пролиферации гладкой мускулатуры стенки сосуда и фиброзной ткани (Brenner и др.. Adv. Exp.Med. Biol. (1976) том 83, стр.85-101), уменьшения или предотвращения кровотечения в желудочно-кишечном тракте и других побочных эффектов нестероидных противовоспалительных лекарственных средств (см. патент США номер 4666701), предупреждения или лечения эндометриоза и предменструального синдрома (см. патент США номер 4758592) и лечения миалгического энцефаломиелита и хронической усталости после вирусных инфекций (см. патент США номер 5116871).
[00167] Дополнительные применения композиций согласно настоящему описанию включают применение для лечения СПИДа, множественного склероза и воспалительных заболеваний кожи, а также для поддержания общего здорового состояния. [00168] Дополнительно, композицию согласно настоящему описанию можно применять для косметических целей. Ее можно добавить в уже известные косметические композиции, таким образом, чтобы получить смесь, или можно применять в качестве самостоятельной композиции. Применения в ветеринарии
[00169] Следует отметить, что описанные выше фармацевтические и пищевые композиции можно применять у животных (т.е. у домашних или не домашних), также, как и у людей, поскольку животные испытывают многие из таких же потребностей и находятся во многих из таких же состояний, что и люди. Например, масло или кислоты согласно настоящему описанию можно применять в пищевых добавках для животных или аквакультур, пищевых заменителях для животных, витаминах для животных или в топических мазях для животных.
[00170] Настоящее описание можно проиллюстрировать с помощью следующих неограничивающих примеров.
Пример 1: Конструирование библиотеки кДНК из ССМР 2916 Euglena deses Ehr. и анализ последовательности для выделения предполагаемых кандидатов дельта-9-элонгазы
[00171] Анализ состава жирных кислот некоторых морских водорослей выявил присутствие значительного количества докозагексаеновой кислоты (ДГК, 22:6n-3) (15% общей массы липидов) в ССМР 2916 Euglena deses Ehr. (см. таблицу 1). В дополнение, в данном организме присутствовали интермедиаты альтернативного пути с участием дельта-8-десатуразы/дельта-9-элонгазы (см. Фигуру 1), что свидетельствует о том, что данный путь активен в этом организме. Таким образом, предположили, что в данном организме присутствует активная дельта-9-элонгаза, способная превращать линолевую кислоту (ЛК, 18:2n-6) в омега-6-эйкозандиеновую кислоту (омега-6-ЭДК, 20:2n-6), или альфа-линоленовую кислоту (АЛК, 18:3, n-3) в омега-3-эйкозатриеновую кислоту (омега-3-ЭТрК, 20:3n-3), а также активная дельта-8-десатураза, способная превращать омега-6-эйкозандиеновую кислоту (омега-6-ЭДК, 20:2 n-6) в дигомо-гамма-линоленовую кислоту (ДГЛК, 20:3 n-6), или омега-3-эйкозатриеновую кислоту (омега-3-ЭТрК, 20:3n-3) в омега-3-эйкозатетраеновую кислоту (омега-3-ЭТК, 20:4n-3) (см. Фигуру 1).
[00172] Целью данного исследования было выделение полноразмерного гена дельта-9-элонгазы из ССМР 2916 Euglena deses Ehr. и описание ферментативной активности его продукта путем экспрессии в гетерологичном хозяине, Saccharomyces cerevisiae. [00173] Для выделения полноразмерных генов из ССМР 2916 Euglena deses Ehr., сконструировали библиотеку микро-кДНК, применяя суммарную РНК, выделенную из данного организма. Для этого получили клеточную массу ССМР 2916 Euglena deses Ehr. из Provasoli-Guillard-National Center for Marine Phytoplankton (CCMP-Bigelow Laboratories, Вест-Бутбэй, Мэн) и выделили из нее суммарную РНК, применяя набор Qiagen RNeasy Maxi kit (Qiagen, Валенсия, Калифорния), следуя протоколу производителя. Вкратце, замороженную клеточную массу размельчали в жидком азоте, применяя ступку и пестик, суспендировали в буфере RLT (Qiagen RNeasy Plant Mini kit) и пропускали через QiaShredder. РНК очищали с помощью колонок RNeasy maxi, следуя инструкциям производителя.
[00174] Конструирование библиотеки микро-кДНК заказали в Agencourt Biosciences (Уолтем, Массачусетс), которую там сконструировали, применяя 50 мкг РНК из ССМР 2916 Euglena deses Ehr., следуя запатентованной технологии. В Agencourt используют несколько уникальных и запатентованных этапов в процессе синтеза первой нити, которые, в конечном итоге, позволяют добиться на 25 - 30% большей эффективности, чем при обычно применяемых методиках. Во время запатентованного процесса, РНК подвергают обратной транскрипции в онДНК, используя условия, разработанные для уменьшения или исключения явлений внутреннего праймирования. Комбинация данного процесса и специализированной программы проведения циклов повышает количество полноразмерных клонов. После синтеза второй нити, клоны кДНК затем отбирают по размеру, чтобы они были больше, чем 1,2 т.п.о., для уменьшения преимущественного клонирования маленьких, укороченных кДНК. Для библиотеки с большими размерами вставок, размер вставки выбирают >4 т.п.о., чтобы повысить количество клонов с большими вставками. После отбора по размеру, концы кДНК защищали и кДНК расщепляли с помощью редкощепящего фермента. "Редкощепящий" фермент рестрикции, сайт для которого вводят в клоны во время этапа праймирования кДНК, затем используют для получения клонов для направленного клонирования в вектор pAGEN. Вероятность того, что "редкощепящий" фермент рестрикции расщепит сами клоны кДНК, в 20 раз ниже, следовательно, данный фермент позволяет получить гораздо больше полноразмерных клонов, чем при процессах конструирования библиотек кДНК, в которых используют обычные ферменты рестрикции, которые расщепляют клон на произвольных участках. В результате получают вставку с 5′-тупым концом и 3′-выступом, который образовался в результате расщепления редкощепящим ферментом рестрикции. Благодаря данному процессу, не требуется дополнительное легирование адаптера для обеспечения направленного клонирования. Это в целом улучшает эффективность процесса клонирования. Указанный вектор специально сконструирован для направленного клонирования без применения 5′-адаптеров, что дополнительно повышает эффективность трансформации благодаря уменьшению количества манипуляций над кДНК в процессе клонирования. После того, как закончили составлять первичную библиотеку кДНК, в ней проверяют количество независимых клонов, процент рекомбинантных клонов и средний размер вставки.
[00175] Клонами затем трансформировали DH10B Е. coli (бактериальные клетки, устойчивые к фагу Т1). Титр полученной библиотеки составлял 3,2×106 КОЕ/мл, с количеством независимых колоний 3,52×107 и со средним размером вставки 1,3 т.п.о.
[00176] Секвенировали 4224 клона из данной библиотеки кДНК и анализировали последовательности без вектора, применяя BLAST, чтобы идентифицировать последовательности, содержащие гомологию с известными последовательностями дельта-9-элонгазы. Анализ с помощью BLAST выявил пять предполагаемых совпадений в библиотеке кДНК ССМР 2916 Euglena deses Ehr. с гомологией с известными последовательностями дельта-9-элонгазы из Pavlova saUna (номер доступа AAY 15135; SEQ ID NO:1; Фигура 4А), Isochrysis galbana (номер доступа AF390174; SEQ ID NO:2; Фигура 4B), Eutreptiella sp.(см. WO 2007/061845 A2; SEQ ID NO:3; Фигура 4С), Euglena gracialis (номер доступа САТ16687; SEQ ID NO:4; Фигура 4D) и Euglena anabena (см. WO 2008/0194685 Al; SEQ ID NO:5; Фигура 4Е).
[00177] У одного клона EST, обозначенного ′plate2_MO7′ (SEQ ID NO:6; Фигура 5А), полученного в результате секвенирования клонов из библиотеки кДНК ССМР 2916 Euglena deses Ehr., обнаружили высокую гомологию последовательности с ранее идентифицированными дельта-9-элонгазами. Длина данного фрагмента ДНК составляла 744 п.о., и предсказанная последовательность аминокислот (SEQ ID NO:7; Фигура 5 В) обладала идентичностью (идентичность последовательности аминокислот составляла 66%) с последовательностью дельта-9-элонгазы из Euglena gracialis (SEQ ID NO:4). Фрагмент гена plate2_MO7, похоже, содержал сайт инициации транскрипции ′ATG′ гена, что выяснили исходя из выравнивания с другими дельта-9-элонгазами, но не содержал 3′-конец предполагаемой дельта-9-элонгазы ССМР 2916 Euglena deses Ehr.
Пример 2: Выделение 3′-конца элонгазы plate2 MO7 из ССМР 2916 Euslena deses Ehr.
[00178] Последовательность клона plate2_MO7 из Примера 1 использовали в качестве матрицы для получения 3′-конца.
[00179] Первую нить кДНК синтезировали с помощью набора для быстрой амплификации концов кДНК SMART™ RACE kit (BD Biosciences), следуя инструкциям производителя. Для синтеза готовой к 3′-RACE кДНК, 1,5 мкг суммарной РНК из ССМР 2916 Euglena deses Ehr. и 1 мкл праймера к 3′-кодирующей последовательности (5′-AAGCAGTGGTATCAACGCAGAGTAC(T)30VN-3′, в котором N=А, С, G или Т; и V=А G или С (SEQ ID NO:8)) (12 мкМ) смешивали в суммарном объеме 5 мкл в безнуклеазной пробирке для ПЦР, инкубировали при 70°С в течение 2 минут и быстро охлаждали на льду. После непродолжительного центрифугирования в пробирки добавляли 2 мкл 5Х буфера для первой нити [250 мМ Tris-HCl (рН-8,3), 375 мМ КСl и 30 мМ MgCl2], 1 мкл 0,1 М дитиотреитола (ДТТ) и 1 мкл 10 мМ смеси дНТФ. После инкубирования при 42°С в течение 2 минут, в пробирку добавляли 1 мкл обратной траскриптазы (PowerScript™ RT, BD Biosciences) и инкубировали при 42°С в течение 90 минут. Первую нить кДНК разбавляли в 100 мкл буфера трицин- этилендиаминтетрауксусной кислоты (ЭДТК) [10 мМ трицин-КОН (рН 8,5), 1,0 мМ ЭДТК] и ферменты инактивировали прогревом на 72°С в течение 7 минут.
[00180] Для выделения 3′-конца фрагмента гена элонгазы Euglenoid sp. (т.е. последовательности клона plate2_MO7), разработали праймеры, исходя из сведений о последовательности части гена элонгазы plate2_MO7. Первичную ПЦР-амплификацию осуществляли, применяя готовую к 3′-RACE кДНК в качестве матрицы и следующие праймеры: Eug Elo МО-7 FP1 (геноспецифичный праймер) (5′ - AGG CGC TGT GGA TCT TCG TCT TCC - 3′) (SEQ ID NO:9), в комбинации со смесью праймеров для RACE Universal Primer Mix A (UPM, BD Biosciences):
[00181] Длинный праймер (0,4 мкМ): 5′ - СТА АТА CGA CTC ACT ATA GCA AGC AGT GGT АТС AAC GCA GAG T - 3′ (SEQ ID NO:10); и
[00182] Короткий праймер (2 мкМ): 5′ - СТА АТА CGA CTC ACT ATA GGG С - 3′ (SEQ IDNO:11).
[00183] Амплификацию осуществляли, используя 0,25 мкл (100 мМ) геноспецифичного праймера, 0,25 мкл (100 мМ) смеси праймеров UPM, 2,5 мкл матрицы кДНК, 2,5 мкл 2,5 мМ дНТФ, 5 мкл 5Х буфера для ПЦР (Advantage® GC II polymerase buffer (Clontech), 200 мМ трицин-КОН (рН 9,2), 75 мМ ацетат калия, 17,5 мМ ацетат магния, 25% ДМСО, 18,75 мкг/мл БСА, 0,005% Tween 20, 0,005% Nonidet-P40), 2,5 мкл реагента GC Melt Reagent (Clontech), 0,5 мкл полимеразы 50Х Advantage® GC I polymerase (Clontech) и 11,5 мкл воды Milli-Q® (Millipore), в конечном объеме реакции, равном 25 мкл. Образцы сначала денатурировали при 94°С в течение 3 минут, а затем осуществляли 2 цикла: 94°С в течение 30 секунд, 64°С в течение 30 секунд и 68°С в течение 1,3 минут; 3 цикла: 94°С в течение 30 секунд, 62°С в течение 30 секунд и 68°С в течение 1,30 минут; 4 цикла: 94°С в течение 30 секунд, 60°С в течение 30 секунд и 68°С в течение 1,30 минут; и 26 циклов: 94°С в течение 30 секунд, 58°С в течение 30 секунд и 68°С в течение 1,30 минут. Конечный цикл элонгации при 68°С в течение 10 минут осуществляли перед остановкой реакции при 4°С.
[00184] Анализ ПЦР-продуктов выявил очень слабые полосы, что, вероятно, было следствием низких уровней транскриптов гена элонгазы в клетке. Следовательно, осуществляли гнездовую ПЦР-реакцию, применяя в качестве матрицы 1 мкл продукта описанной выше первичной ПЦР-реакции. Праймеры, использованные для гнездовой ПЦР, представляли собой Eug Elo МО-7 FP2 (геноспецифичный праймер): 5′ - ТСС CCG TGC CGA AGT CGT ТСА ТСА СС - 3′ (SEQ ID NO:12) и смесь праймеров Universal Primer Mix A (UPM) (последовательности SEQ ID NO:10 и 11). Условия ПЦР-реакции и параметры циклов были такими же, как и используемые для первичной ПЦР-реакции.
[00185] Ампликон размером 548 п.о. (SEQ ID NO:13; Фигура 6А), полученный с помощью гнездовой ПЦР, очищали в геле, применяя набор для очистки Qiagen Gel Purification kit (Qiagen), клонировали в вектор pTZ57R/T (Т/А cloning vector, MBI Fermentas) и секвенировали. Секвенирование позволило выявить, что данный фрагмент (SEQ ID NO:13) содержал целый 3′-конец фрагмента элонгазы plate2_MO7, наряду со стоп-кодоном ′TAG′ и находящимся по ходу транскрипции от него участком, содержащим полиА-хвост. Предсказанная последовательность аминокислот данного фрагмента (последовательности SEQ ID NO:14 и 30-32) показана на Фигуре 6 В. Первая звездочка обозначает стоп-сайт кодируемого plate 2_MO7 белка.
Пример 3: Выделение полноразмерного гена элонгазы plate2 MO7 из ССМР 2916 Euglena deses Ehr.
[00186] Полноразмерную последовательность гена элонгазы plate2_MO7 получали путем ПЦР-амплификации, применяя библиотеку кДНК Euglena deses Ehr. в качестве матрицы и праймеры, разработанные таким образом, чтобы они включали 5′- и 3′-концы гена plate2_MO7, исходя из сведений о последовательности, полученных в Примере 1 и Примере 2. В дополнение, в праймеры были включены сайты BamHI/HindIII (подчеркнуты) для облегчения клонирования гена в сайты BamHI/HindIII дрожжевого вектора экспрессии pYX242. Использовали следующие последовательности праймеров:
[00187] Прямой праймер МO7-Еlо: 5′ - САС CAT GGA ТСС ATG GAC GTC GCG ACT ACG CTG G - 3′ (SEQ ID NO:15), и
[00188] Обратный праймер МO7-Еlо: 5′ - ACG CGT AAG CTT СТА GTC САС TTT CTT CTC АТС CTT C - 3′ (SEQ ID NO:16).
[00189] Амплификацию осуществляли, применяя 0,5 мкл (100 мкМ) каждого праймера, 1 мкл (~110 нг) пула плазмид библиотеки кДНК Euglena deses Ehr. в качестве матрицы, 5 мкл 2,5 мМ дНТФ, 10 мкл 5Х буфера Phusion GC Buffer (Finnzymes), 5 мкл ДМСО, 0,5 мкл (1 ед.) полимеразы Phusion polymerase (Finnzymes) и 27,5 мкл воды Milli-Q® (Millipore). Образцы сначала денатурировали при 98°С в течение 3 минут, а затем осуществляли 2 цикла: 98°С в течение 8 секунд, 60°С в течение 12 секунд и 72°С в течение 45 секунд; и 28 циклов: 98°С в течение 8 секунд, 58°С в течение 12 секунд и 72°С в течение 45 секунд. Осуществляли конечный цикл элонгации при 72°С в течение 3 минут перед остановкой реакции при 4°С.
[00190] В результате ПЦР получили продукт размером -789 п.о., который клонировали в сайты Ват Hi/Hind III вектора pYX242 и трансформировали им E.coli DH5a (Invitrogen). Полученную таким образом плазмидную ДНК секвенировали с получением полноразмерной последовательности гена размером 789 п.о., обозначенного ′Eug-MO7-ELO#10′ (SEQ ID NO:17; Фигура 7А). SEQ ID NO:17 депонировали в Американскую коллекцию типовых культур (АТСС), 10801 University Boulevard, Манассас, Вирджиния 20110-2209 10 июля, 2009 г., в соответствии с Будапештским договором, и ей был присвоен номер депозита АТСС ___. Считали, что данный ген кодирует предполагаемую дельта-9-элонгазу из ССМР 2916 Euglena deses Ehr., и имеет предсказанную длину 262 аминокислоты (SEQ ID NO:18; Фигура 7 В). Данный ген использовали для исследования экспрессии, чтобы описать ферментативную активность его продукта.
[00191] В дополнение к клону Eug-MO7-ELO#10, в процессе секвенирования были идентифицированы дополнительные варианты клона, последовательности которых несколько отличались на некоторых участках на протяжении полноразмерного гена. Данные варианты последовательности, вероятно, появились в процессе ПЦР-амплификации. Анализ последовательности одного такого варианта, Eug-MO7-ELO #14, выявил несколько нуклеотидных и соответствующих аминокислотных замен по сравнению с исходным клоном Eug-MO7-ELO#10 (см. таблицу 2 и Фигуры 2А и 2В). Нуклеотидная (SEQ ID NO:19) и предсказанная аминокислотная последовательность (SEQ ID NO:20) Eug-MO7-ELO #14 показана на Фигурах 8А и 8В, соответственно. Для анализа экспрессии использовали как исходный клон Eug-MO7-ELO#10, так и вариант Eug-MO7-ELO #14.
[00192] Запрос последовательности Eug-MO7-ELO #10 в BLAST с поиском подобия с последовательностями, находящимися в базе данных BLAST ′nr′, выявил, что предсказанная последовательность аминокислот, кодируемая Eug-MO7-ELO#10 (SEQ ID NO:18), обладает наибольшей идентичностью последовательности аминокислот (36% идентичности последовательностей) с дельта-9-элонгазой Isochrysis galbana (SEQ ID NO:2). Попарное выравнивание SEQ ID NO:18 с известной дельта-9-элонгазой из Euglena gracialis (номер доступа CAT 16687; SEQ ID NO:4) выявило гораздо большую идентичность последовательностей аминокислот (66% идентичности). При этом для попарного выравнивания использовали параметры по умолчанию программы Vector NTI®AlignX. Попарное выравнивание с последовательностью дельта-9-элонгазы Pavlova salina (SEQ ID NO:1) выявило лишь -15% идентичности последовательностей.
[00193] В отличие от десатураз, ферменты элонгазы содержат очень мало высококонсервативных фрагментов. Данные ферменты представляют собой высокогидрофобные белки, содержащие от четырех до пяти гидрофобных участков, которые предположительно пронизывают мембрану. В дополнение, в них был обнаружен высококонсервативный гистидиновый блок (НХХНН) (SEQ ID NO:28), включенный в четвертый пронизывающий мембрану участок, который необходим для осуществления ферментативной активности (см. Leonard и др., "Elongation of long-chain fatty acids", Prog Lipid Res. (2004) том 43, стр.36-54). В некоторых элонгазах, первый остаток гистидина во фрагменте ′НХХНН′ (SEQ ID NO:28) заменен на глутамин (Q), что приводит к образованию консервативного фрагмента ′QXXHH′ (SEQ ID NO:29). Данный фрагмент QXXHH (SEQ ID NO:29) обнаруживают в большинстве дельта-9-элонгаз, включая Eug-MO7-ELO#10. В дополнение, элонгаза Eug-MO7-ELO#10 содержит другие неизменяющиеся остатки, которые присутствуют в большинстве элонгаз, известных на данный момент и описанных в Leonard и др., "Elongation of long-chain fatty acids". Prog LipidRes. (2004) том 43, стр.36-54. [00194] На Фигурах 3А и 3В изображено выравнивание последовательности аминокислот элонгазы Eug-MO7-ELO#10 с другими известными элонгазами, которые обладают различной субстратной специфичностью. Данные элонгазы включают элонгазу Elov14 мыши (номер доступа AAG47667; SEQ ID NO:21; Фигура 9А), элонгазу ELOVL2 человека (номер доступа NP_060240; SEQ ID NO:22; Фигура 9 В) и элонгазу С. elegans (номер доступа AF244356; SEQ ID NO:23), в дополнение к дельта-9-элонгазам из Euglena gracialis (SEQ ID NO:4) и Isochrysis galbana (SEQ ID NO:2). Неизменяющиеся аминокислоты заштрихованы в выровненных последовательностях. Принято считать, что данные неизменяющиеся остатки являются существенными для функционирования данных элонгирующих ферментов, так как они высоко консервативны между разными видами. Выравнивание осуществляли, применяя программное обеспечение Vector NTI, в котором реализуется измененный алгоритм ClustalW. Пример 4: Определение ферментативной активности предполагаемой дельта-9-элонгазы, кодируемой геном Eug-MO7-ELO#10
[00195] Eug-MO7-ELO#10 и вариант Eug-MO7-ELO#14, кодирующие предполагаемую дельта-9-элонгазу, клонировали в сайты BamHI/HindIII дрожжевого вектора экспрессии, pYX242 (Novagen), соответственно. Данными конструкциями трансформировали клетки компетентного штамма SC334 Saccharomyces cerevisiae. Трансформацию дрожжей осуществляли с помощью набора Alkali-Cation Yeast Transformation Kit (QBioGene), придерживаясь условий, рекомендованных производителем. Трансформанты отбирали по ауксотрофии по лейцину в средах, в которых отсутствовал лейцин (DOB [-Leu]).
[00196] Для определения элонгирующей активности ферментов, кодируемых Eug-MO7-ELO#10 и Eug-MO7-ELO#14, трансформанты растили в присутствии 50 мкМ специфичных жирных кислот - субстратов (перечислены ниже), и субстратную специфичность определяли по превращению специфичного продукта:
[00197] Для дельта-9-элонгазной активности:
[00198] Линолевая кислота (18:2 n-6) ⇒ Эйкозандиеновая кислота (ЭДК, 20:2 n-6)
[00199] Альфа-линоленовая кислота (18:3 n-3) ⇒ Эйкозатриеновая кислота (ЭТрК, 20:3 n-3)
[00200] Для C18-элонгазной активности:
[00201] Гамма-линоленовая кислота (ГЛК, 18:3 n-6) ⇒ Дигомо-гамма-линоленовая кислота (ДГЛК, 20:3 n-б)
[00202] Стеаридоновая кислота (СДК, 18:4 n-3) ⇒ Омега-3-эйкозатетраеновая кислота (омега-3-ЭТК, 20:4 n-3)
[00203] Для С20-элонгазной активности:
[00204] Арахидоновая кислота (АРК, 20:4 n-6) ⇒ Адреновая кислота (омега-6-АДК, 22:4 n-6)
[00205] Эйкозапентаеновая кислота (ЭПК, 20:5 n-3) ⇒ Омега-3-докозапентаеновая кислота (омега-3-ДПК, 22:5 n-3)
[00206] В качестве отрицательного контроля использовали штамм 334 S. Cerevisiae, в котором экспрессировался вектор pYX242.
[00207] Трансформированные колонии, выделенные из селективной среды DOB [-Leu], растили в течение ночи в 10 мл жидкого бульона YPD при 30°С, при интенсивном встряхивании. 5 мл данной культуры, культивированной в течение одной ночи, затем добавляли в 45 мл селективной среды (DOB [-Leu]), содержащей 50 мкМ (конечная концентрация) различных жирных кислот- субстратов (как описано), и интенсивно встряхивали (250 об/мин) в течение от 48 до 72 часов (как указано) при 24°С.
[00208] Для экстрагирования всех липиов, дрожжевые клетки откручивали при 2000 об/мин в течение 15 минут и добавляли 0,5 мл воды, образцы встряхивали, а затем добавляли 10 мл метанола при аккуратном перемешивании. Затем добавляли 20 мл хлороформа, образцы встряхивали в течение 1 минуты при высокой скорости встряхивания и оставляли на 2 часа при комнатной температуре. Потом в образец добавляли 6 мл солевого раствора, а затем центрифугировали при 2200 об/мин в течение 10 минут. Верхнюю фракцию хлороформа переносили в чистую/сухую 30 мл виалу и выпаривали хлороформ досуха при 40°С в потоке азота. Как только растворители полностью выпарились, в каждую виалу добавляли 2 мл хлороформа и образцы дериватизировали.
[00209] Для дериватизации липидов в метиловые эфиры жирных кислот (МЭЖК), в каждую пробирку вносили 100 мкл внутреннего стандарта (17,216 мкг/100 мкл) тригептадеканоина. Хлороформ выпаривали досуха в атмосфере азота при 40°С, добавляли 2 мл трифторида бора в 14% метаноле, а затем добавляли 2 капли (-50 мкл) толуола. Через каждую виалу пропускали азот и грели в течение 15 минут при 95°С.После того, как виалы охладились, добавляли 2 мл солевого раствора и экстрагировали липиды 4 мл гексана при интенсивном встряхивании в течение 1 минуты. Гексановый экстракт затем переносили в 20 мл чистые/сухие пробирки с завинчивающейся крышкой, добавляли 5 мл деионизованной воды (ди-Н2O), встряхивали образец и центрифугировали при 1500 об/мин в течение 4 минут. Промытый гексан затем переносили в 20 мл пробирку с реагентом. Гексан выпаривали досуха, и каждый образец вновь разбавляли 0,5 мл свежего гексана. Конечное разбавление гексаном встряхивали, чтобы диспергировать липиды. Весь образец затем погружали в виалы для автоматического отбора проб GC auto sampler vials и инжектировали 4 мкл для анализа. GC были откалиброваны с помощью стандарта NuChek Std. 461.
[00210] Процент превращения субстрата в продукт рассчитывали, применяя формулу:
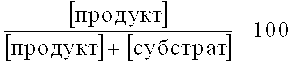
[00211] В Таблице 3 представлена ферментативная активность белков, кодируемых Eug-MO7-ELO#10 и Eug-MO7-ELO#14, на основании процента превращения добавленного субстрата. Белок, кодируемый Eug-MO7-ELO#10, превращал 10,5% ЛК (18:2n-6) в ЭДК (20:2 n-6) и 23,2% АЛК (18:3п-3) в ЭТрК (20:3n-3). Это свидетельствовало о том, что ген Eug-MO7-ELO#10 кодирует дельта-9-элонгазу, которая может узнавать как n-6, так и n-3 жирные кислоты-субстраты. Белок, кодируемый измененным клоном Eug-MO7-ELO#14, также проявлял дельта-9-элонгирующую активность, превращая 7,84% ЛК (18:2n-6) в ЭДК (20:2 n-6) и 17,15% АЛК (18:3n-3) в ЭТрК (20:3n-3). Тем не менее, его активность была ниже, чем у белка, кодируемого исходной последовательностью Eug-MO7-ELO#10. Это свидетельствует о том, что остатки, которые отличаются между Eug-MO7-ELO#14 и Eug-MO7-ELO#10, являются существенными для дельта-9-элонгирующей активности данного фермента.
[00212] Очень низкая фоновая (неспецифичное превращение субстрата) активность была обнаружена для контроля, содержащего только вектор (см. таблицу 3). Ферменты, кодируемые как Eug-MO7-ELO#10, так и Eug-MO7-ELO#14, не проявляли активность на каких-либо других тестируемых субстратах ПНЖК (см. таблицу 4), что свидетельствует о том, что данный фермент специфичен к субстратам, вовлеченным в альтернативный путь с участием дельта-8-десатуразы/дельта-9-элонгазы (см. Фигуру 1).
b Количество образовавшегося продукта
с % превращения = ([продукт]/{[продукт]+[субстрат]})×100
MO7-ELO#10 и Eug-MO7-ELO#14, экспрессированных в штамме SC334
Saccharomyces cerevisiae.
b Количество образовавшегося продукта
с % превращения = ([продукт]/{[продукт]+[субстрат]})×100
Пример 5: Экспрессия дельта-9-элонгазы ′Eug-МO7-ELO#10′ в семенах растения
[00213] Кодирующую последовательность элонгазы Eug-MO7-ELO#10 амплифицировали с помощью ПЦР плазмиды, содержащей соответствующий ген, со следующими смысловым и антисмысловым олигонуклеотидными праймерами (введенные сайты для ферментов рестрикции подчеркнуты):
[00214] 5′ - TATAGAATTCAAATGGACGTCGCGACTACGCTG - 3′ (SEQ ID NO:24), и
[00215] 5′ - TATTCTCGAGTTCTAGTCCACTTTCTTCTCATCCTTC - 3′ (SEQ ID NO:25).
[00216] ПЦР-реакцию осуществляли с использованием высокоточной полимеразы Phusion polymerase (New England Biolabs). Амплифицированный с помощью ПЦР ген расщепляли ферментами рестрикции EcoRI и XhoI, и полученный в результате этого продукт соединяли по 5′-концу со специфичным для семян промотором глицинина-1 из сои и по 3′-концу с 3′-нетранслируемой областью глицинина-1 в бинарном векторе р0308-DsRed с получением плазмиды ′pEugELO′. Регуляторные элементы глицинина-1 были ранее описаны в Nielsen и др., "Characterization of the glycinin gene family in soybean" Plant Cell (1989) том 1, стр.313-328. Данный вектор также содержит трансген Ds-Red под контролем промотора вируса мозаики маниока для селекции трансформированных семян с помощью флуоресценции и маркера устойчивости к канамицину для селекции бактерий. В качестве контроля для данных экспериментов, ген дельта-9-элонгазы Isochrysis galbana (SEQ ID NO:2) также клонировали в виде фрагмента EcoRI/XhoI под контроль промотора глицинина-1 в p0308-Ds-Red с получением плазмиды ′pIsoD9′.
[00217] pEugELO и pIsoD9 внедряли в штамм С58 МР90 Agrobacterium tumefaciens путем электропорации. Устойчивые к канамицину агробактерии затем использовали для трансформирования экотипа Co1-0 Arabidopsis thaliana с помощью способа погружения цветочных почек ("floral dip") (Clough и др., "Floral dip: a simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana". Plant J (1998) том 16, стр.735-743). После погружения цветочных почек в агробактерии, растения оставляли при 22°С с длиной дня 16 часов до достижения зрелости, а затем высушивали. Для данных экспериментов использовали мутанта fad3/fae1 Arabidopsis, который содержит низкие уровни альфа-линоленовой кислоты и жирные кислоты с очень длинной цепью (≥С20), но при этом в масле из его семян содержатся повышенные уровни линолевой кислоты (Cahoon и др., "Conjugated fatty acids accumulate to high levels in phospholipids of metabolically engineered soybean and Arabidopsis seeds" Phytochemistry (2006) том 67, стр.1166-1176). Данное генетическое окружение дает профиль жирных кислот, приблизительно соответствующий их профилю в маслах из семян таких культур, как сафлор и соя с низким содержанием линоленовой кислоты. Трансгенные семена, полученные из растений Arabidopsis после погружения в агробактерии, идентифицировали по флуоресценции маркерного белка DsRed, применяя методику, описанную в Pidkowich и др., "Modulating seed beta-ketoacyl-acyl carrier protein synthase II level converts the composition of a temperate seed oil to that of a palm-like tropical oil", Proc Natl Acad Sci USA (2007) том 104, стр.4742-4747. Липиды, включая триацилглицерины, входящие в состав трансгенных по одному гену и нетрансгенных контрольных семян, подвергали непосредственной переэтерификации путем применения реагента гидроксида триметилсульфония (TMSH), как описано в Cahoon и Shanklin "Substrate-dependent mutant complementation to select fatty acid desaturase variants for metabolic engineering of plant seed oils", Proc Natl Acad Sci USA (2000) том 97, стр.12350-12355. Метиловые эфиры жирных кислот, полученные из отдельных семян, анализировали с помощью газовой хроматографии с пламенно-ионизационным детектированием, применяя газовый хроматограф Agilent 6890, оборудованный колонкой INNOWax (длина 30 м × внутренний диаметр 0,25 мм) и с возможностью программно изменять температуру печи от 185°С (выдерживание 1 минуту) до 230°С (выдерживание 2 минуты) со скоростью 7°С/минуту. Метиловые эфиры входящих в состав семян жирных кислот идентифицировали на основании времени их удержания по сравнению с метиловыми эфирами известных жирных кислот из семян дикого типа Co1-0 Arabidopsis thaliana, и путем сравнения времени удержания с таковым для стандартных метиловых эфиров жирных кислот.
[00218] В Таблице 5 показаны составы жирных кислот семян, трансформированных только Т1, полученных в результате шести независимых событий трансформации из растений, трансформированных конструкцией pEugELO. Также показаны составы жирных кислот семян, трансформированных только Т1, представляющих независимые события трансформации, из растений, трансформированных конструкцией pIsoD9, контрольной дельта-9-элонгазой (см. таблицу 6). Основным изменением в составе жирных кислот трансгенных семян, полученных в результате трансформации pEugELO, по сравнению с нетрансформированными семенами fad3/fae1 (см. таблицу 7), оказалось присутствие высоких уровней ЭДК (20:2n-6, дельта-11,14). В данных семенах, относительные количества 20:2 находились в диапазоне от 40% до 49% (масс./масс.) общего количества жирных кислот. Для сравнения, 20:2 составляли >0,5% общего количества жирных кислот в семенах нетрансгенного fad3/fae1 (см. Таблицу 7). Это сопровождалось одновременным уменьшением относительных количеств ЛК (18:2n-6, дельта-9, 12) от приблизительно 50% в нетрансгенных семенах fad3/fae1 (см. Таблицу 7) до всего 14% в pEugELO. Этот результат согласуется с тем фактом, что 18:2 служит первичным субстратом для синтеза 20:2, осуществляемого элонгазой Eug-MO7-ELO#10. Количества эйкозеновой кислоты (20:1 дельта-11) и эйкозановой кислоты (20:0) также были повышены в трансформированных pEugELO семенах, по сравнению с нетрансгенными семенами fad3/fae1, но каждая из указанных жирных кислот составляла <3% общего количества жирных кислот в трансгенных семенах. Данные открытия указывают на то, что элонгаза Eug-MO7-ELO#10 в растениях обладает субстратным предпочтением С is ПНЖК, таких как ЛК (18:2n-6), и представляет собой эффективный фермент для получения 20:2 в семенах, которые обогащены ЛК (18:2n-6). Для сравнения, семена, сконструированные для экспрессии дельта-9-ELO Isochrysis galbana (pIsoD9), накапливали 20:2 в количествах от 30 до 40% общего количества жирных кислот и каждую из 20:0 и 20:1 в количествах <3% общего количества жирных кислот (см. Таблицу 6).
Пример 6: Коэкспрессия дельта-9-элонгазы ′Eug-МO7-ELO#10 с дельта-8-десатуразой
[00219] Можно совместно экспрессировать Eug-MO7-ELO#10 с дельта-8-десатуразой, чтобы воссоздать альтернативный путь с участием дельта-8-десатуразы/дельта-9-элонгазы, приводящий к продукции АРК. В дополнение, также возможно ко-экспрессировать три гена, дельта-9-элонгазу ′Eug-MO7-ELO#10′ вместе с дельта-8-десатуразой и дельта-5-десатуразой, в гетерологичных хозяевах, таких как масличные растения или маслянистые дрожжи, чтобы воссоздать путь биосинтеза АРК, который позволит получить АРК в данных гетерологичных хозяевах.
[00220] В связи с приведенным выше описанием, должно быть очевидно, что было достигнуто несколько объектов согласно настоящему описанию и были получены другие полезные результаты.
[00221] Так как в описанный выше объект можно внести различные изменения, без отклонения от объема настоящего описания, предполагается, что все объекты, входящие в приведенное выше описание, должны рассматриваться как иллюстрирующие, но не в ограничивающем смысле.
| название | год | авторы | номер документа |
|---|---|---|---|
| МУЛЬТИЗИМЫ И ИХ ИСПОЛЬЗОВАНИЕ В ПОЛУЧЕНИИ ПОЛИНЕНАСЫЩЕННЫХ ЖИРНЫХ КИСЛОТ | 2008 |
|
RU2517608C2 |
| ДЕЛЬТА-5-ДЕСАТУРАЗА И ЕЕ ПРИМЕНЕНИЕ ДЛЯ ПОЛУЧЕНИЯ ПОЛИНЕНАСЫЩЕННЫХ ЖИРНЫХ КИСЛОТ | 2007 |
|
RU2469092C2 |
| ГЕНЫ ДЕЛЬТА-8-ДЕСАТУРАЗЫ, ФЕРМЕНТЫ, КОДИРУЕМЫЕ ИМИ, И ИХ ПРИМЕНЕНИЕ | 2009 |
|
RU2517615C2 |
| КОНСТРУКЦИИ НУКЛЕИНОВОЙ КИСЛОТЫ И СПОСОБЫ ПОЛУЧЕНИЯ МАСЛА С ИЗМЕНЕННЫМ СОСТАВОМ ИЗ СЕМЯН | 2007 |
|
RU2392795C2 |
| ПОЛУЧЕНИЕ ДГК И ДРУГИХ ДЦ-ПНЖК В РАСТЕНИЯХ | 2012 |
|
RU2644241C2 |
| ДЕЛЬТА 6-АЦЕТИЛЕНАЗА/ДЕЛЬТА-6-ДЕСАТУРАЗА И ДЕЛЬТА-6-ДЕСАТУРАЗА ИЗ Ceratodon purpureus | 2000 |
|
RU2268939C2 |
| ЭНХАНСЕР ПАЛОЧКОВИДНОГО ВИРУСА САХАРНОГО ТРОСТНИКА (SCBV) И ЕГО ПРИМЕНЕНИЕ В ФУНКЦИОНАЛЬНОЙ ГЕНОМИКЕ РАСТЕНИЙ | 2013 |
|
RU2639517C2 |
| НОВАЯ ЛИЗОФОСФОЛИПИД-АЦИЛТРАНСФЕРАЗА | 2010 |
|
RU2534559C2 |
| Brassica juncea КАЧЕСТВА ОМЕГА-9 | 2009 |
|
RU2557316C2 |
| ФРАГМЕНТ ДНК (ВАРИАНТЫ), ВЕКТОР, СПОСОБ ПОЛУЧЕНИЯ РАСТЕНИЯ, СПОСОБ ИНДУЦИРОВАНИЯ ПРОДУКЦИИ ГАММА-ЛИНОЛЕНОВОЙ КИСЛОТЫ (ВАРИАНТЫ), СПОСОБ ИНДУЦИРОВАНИЯ ПРОДУКЦИИ ОКТАДЕКАТЕТРАЕНОВОЙ КИСЛОТЫ И ДЕЛЬТА-6-ДЕСАТУРАЗА ЦИАНОБАКТЕРИЙ | 1992 |
|
RU2152996C2 |




















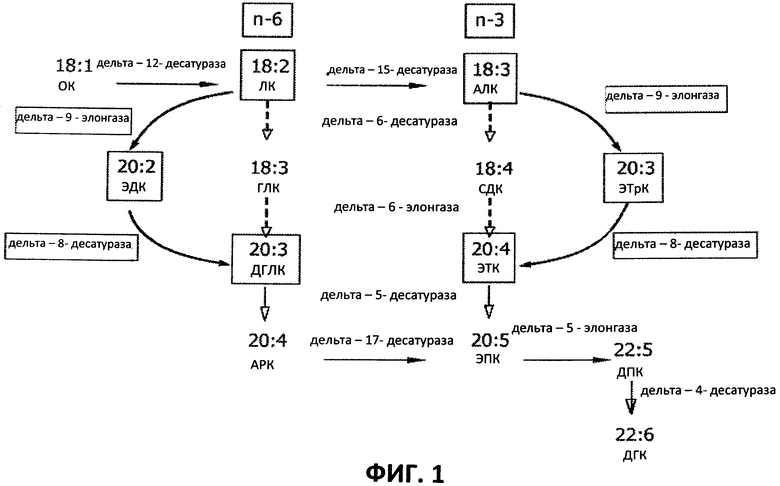
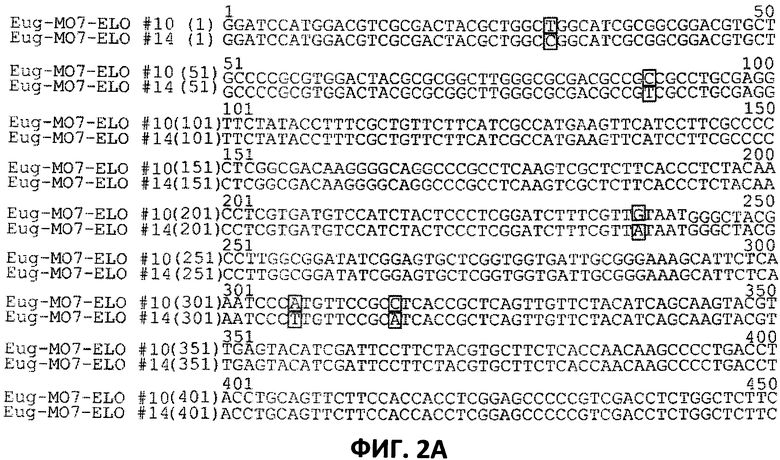
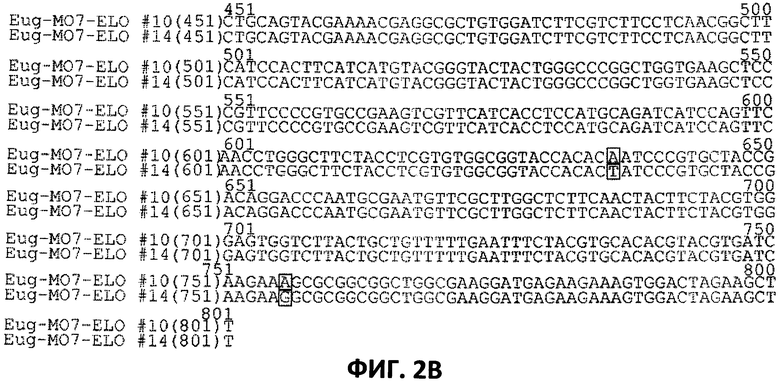
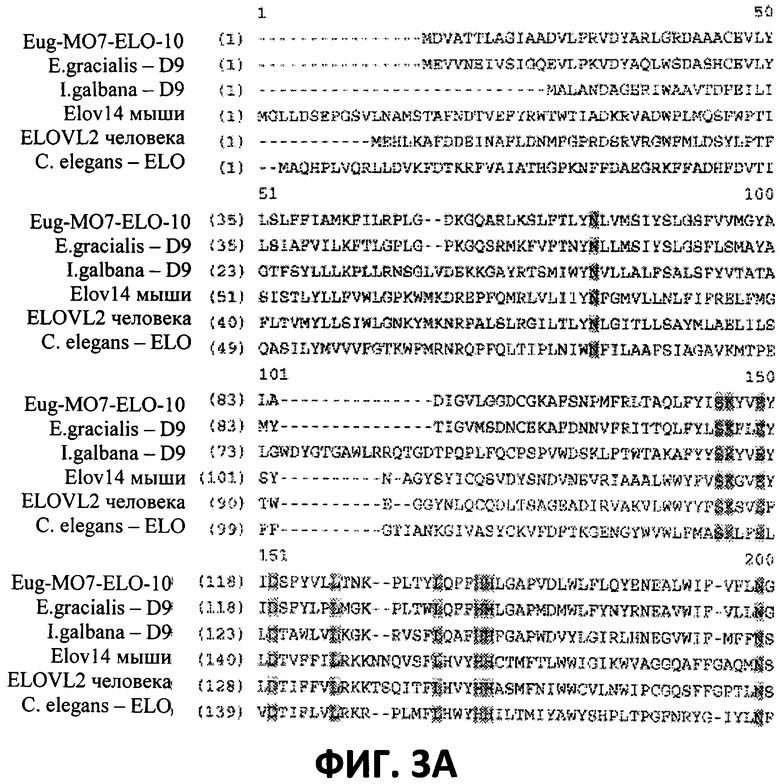








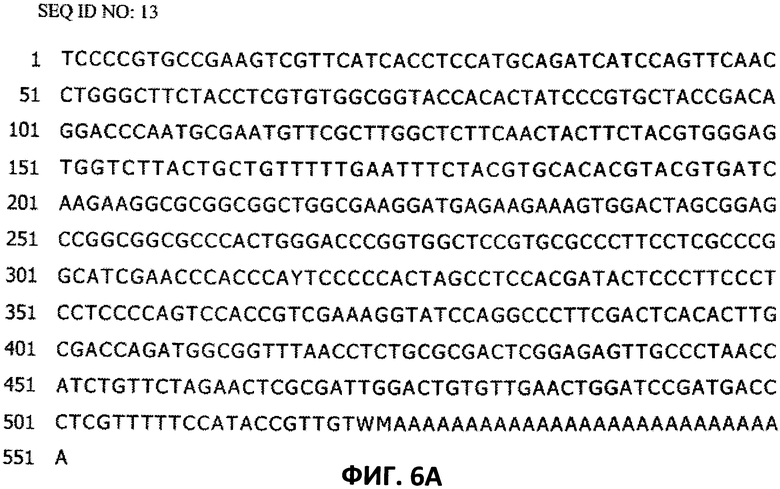












Изобретение относится к области биохимии, в частности к изолированной молекуле нуклеиновой кислоты, кодирующей полипептид, обладающий дельта-9-элонгазной активностью, а также к очищенному полипептиду, обладающему дельта-9-элонгазной активностью, кодируемому вышеуказанной изолированной молекулой нуклеиновой кислоты. Раскрыты вектор экспрессии, содержащий вышеуказанную изолированную молекулу нуклеиновой кислоты, а также клетка-хозяин, трансгенное растение и трансгенное семя, его содержащие. Изобретение позволяет эффективно получать масла, обогащенные ненасыщенными полимерными жирными кислотами. 8 н. и 10 з.п. ф-лы, 9 ил., 9 табл., 6 пр.
1. Изолированная молекула нуклеиновой кислоты, кодирующая полипептид, обладающий дельта-9-элонгазной активностью, причем указанная молекула нуклеиновой кислоты характеризуется изолированной последовательностью нуклеотидов, кодирующей полипептид, характеризующийся последовательностью аминокислот, выбранной из группы, состоящей из SEQ ID NO: 18 и SEQ ID NO: 20, или комплементарна указанной последовательности нуклеотидов.
2. Изолированная молекула нуклеиновой кислоты, кодирующая полипептид, обладающий дельта-9-элонгазной активностью, причем указанная молекула нуклеиновой кислоты характеризуется последовательностью нуклеотидов, выбранной из группы, состоящей из SEQ ID NO: 17 и SEQ ID NO: 19, или комплементарна указанной последовательности нуклеотидов.
3. Изолированная молекула нуклеиновой кислоты по п. 1, отличающаяся тем, что указанная изолированная молекула нуклеиновой кислоты кодирует функционально активную дельта-9-элонгазу, которая использует полиненасыщенную жирную кислоту в качестве субстрата.
4. Изолированная молекула нуклеиновой кислоты по п. 2, отличающаяся тем, что указанная изолированная молекула нуклеиновой кислоты кодирует функционально активную дельта-9-элонгазу, которая использует полиненасыщенную жирную кислоту в качестве субстрата.
5. Изолированная молекула нуклеиновой кислоты по п. 1, отличающаяся тем, что указанная изолированная молекула нуклеиновой кислоты получена из Euglenoid sp.
6. Изолированная молекула нуклеиновой кислоты по п. 5, отличающаяся тем, что указанная изолированная молекула нуклеиновой кислоты получена из Euglena deses Ehr. ССМР 2916.
7. Изолированная молекула нуклеиновой кислоты по п. 2, отличающаяся тем, что указанная изолированная молекула нуклеиновой кислоты получена из Euglenoid sp.
8. Изолированная молекула нуклеиновой кислоты по п. 7, отличающаяся тем, что указанная изолированная молекула нуклеиновой кислоты получена из Euglena deses Ehr. ССМР 2916.
9. Очищенный полипептид, обладающий дельта-9-элонгазной активностью, кодируемый изолированной молекулой нуклеиновой кислоты по п. 1 или 2.
10. Очищенный полипептид, обладающий дельта-9-элонгазной активностью, который удлиняет полиненасыщенные жирные кислоты, содержащие ненасыщенную связь в положении атома углерода 9, причем указанный полипептид характеризуется аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO: 18 и SEQ ID NO: 20.
11. Вектор экспрессии, содержащий изолированную молекулу нуклеиновой кислоты, функционально связанную с регуляторной последовательностью, при этом указанная молекула нуклеиновой кислоты характеризуется последовательностью нуклеотидов, выбранной из группы, состоящей из SEQ ID NO: 17 и SEQ ID NO: 19, или комплементарна указанной последовательности нуклеотидов.
12. Клетка-хозяин для продуцирования полипептида, обладающего дельта-9-элонгазной активностью, причем указанная клетка-хозяин содержит вектор экспрессии по п. 11.
13. Клетка-хозяин по п. 12, отличающаяся тем, что указанная клетка-хозяин выбрана из группы, состоящей из эукариотической клетки и прокариотической клетки.
14. Клетка-хозяин по п. 13, отличающаяся тем, что указанная эукариотическая клетка выбрана из группы, состоящей из: клетки млекопитающего, не являющегося человеком, клетки насекомого, клетки растения и клетки гриба.
15. Клетка-хозяин по п. 14, отличающаяся тем, что указанная клетка растения получена из масличного растения, выбранного из группы, состоящей из: сои, видов Brassica, сафлора, подсолнечника, кукурузы, хлопчатника и льна.
16. Трансгенное растение для продуцирования полипептида, обладающего дельта-9-элонгазной активностью, причем указанное трансгенное растение содержит вектор экспрессии по п. 11, где экспрессия изолированной молекулы нуклеиновой кислоты в указанном векторе экспрессии приводит к продуцированию по меньшей мере одной полиненасыщенной жирной кислоты указанным растением.
17. Трансгенное растение по п. 16, отличающееся тем, что полиненасыщенная жирная кислота выбрана из группы, состоящей из омега-6-эйкозандиеновой кислоты (омега-6-ЭДК), омега-3-эйкозатриеновой кислоты (омега-3-ЭТрК) и комбинации перечисленных жирных кислот.
18. Трансгенное семя для продуцирования полипептида, обладающего дельта-9-элонгазной активностью, которое содержит вектор экспрессии по п. 11.
| WO2008124048 А2, 16.10.2008. WO2008128240 A1, 16.10.2008 | |||
| WO2007136877 А2, 29.11.2007 | |||
| SINGH S.P | |||
| et al., Metabolic engineering of new fatty acids in plants, Curr Opin Plant Biol, 2005, Vol | |||
| Топка с несколькими решетками для твердого топлива | 1918 |
|
SU8A1 |
| ЛОСЬ Д | |||
| А., Структура, регуляция экспрессии и функционирование десатураз жирных кислот, Успехи биологической химии, т | |||
| Механический грохот | 1922 |
|
SU41A1 |
| Деревянное стыковое устройство | 1920 |
|
SU163A1 |

