Настоящее изобретение относится к нуклеиновым кислотам, связывающимся с MCP-1, и к их применению для получения лекарственного препарата и диагностического средства соответственно.
Человеческий MCP-1 (моноцитарный хемоаттрактант, белок-1; альтернативные названия: MCAF [моноцитарный хемоаттрактант и активирующий фактор]; CCL2; SMC-CF [колониестимулирующий фактор гладких мышц]; HC-11; LDCF; GDCF; TSG-8; SCYA2; A2; код доступа SwissProt, P13500) независимо подразделяется на три группы (Matsushima 1988; Rollins 1989; Yoshimura 1989). Этот белок состоит из 76 аминокислот и, подобно всем хемокинам, характеризуется тем, что он имеет гепарин-связывающий сайт. Две внутримолекулярные дисульфидные связи придают данной молекуле стабильную жесткую структуру. Кроме того, MCP-1 у своего амино-конца содержит пироглутамат. В положении Thr 71 находится потенциальный сайт O-связанного гликозилирования. Другие члены семейства MCP были также обнаружены у человека (MCP-2, -3, -4) и у мышей (MCP-2, -3, -5). Человеческие белки имеют приблизительно 70% гомологию с человеческим MCP-1.
Структура MCP-1 была выявлена с помощью ЯМР (Handel 1996) и рентгеноструктурного анализа (Lubkowski 1997). Мономер MCP-1 имеет укладку, типичную для хемокинов, у которых за амино-концевыми цистеинами расположена длинная петля, переходящая в три антипараллельных β-складчатых слоя в мотиве “греческий ключ” (Greek key). На своем конце этот белок имеет α-спираль, которая перекрывается тремя β-слоями (код доступа к базе данных PDB - 1DOK).
Хотя трехмерная структура MCP-1 у млекопитающих различных видов, в основном, сохраняется, однако эволюционно аминокислотная последовательность не является высококонсервативной. Результаты сопоставления последовательностей путем выравнивания продемонстрировали 55% общее сходство человеческой и мышиной последовательностей MCP-1 (также называемого JE) в первых 76 аминокислотах. Помимо аминокислотной последовательности, мышиный MCP-1 отличается от человеческого MCP-1 размером молекулы (125 аминокислот) и степенью гликозилирования. Мышиный MCP-1 содержит карбокси-концевой домен, состоящий из 49 аминокислот, который отсутствует в человеческом MCP-1 и который не является необходимым для биологической активности in vitro. Ниже представлены проценты идентичности аминокислотных последовательностей человеческого MCP-1 с последовательностями MCP-1 различных видов:
Принимая во внимание высокую степень дивергенции, следует отметить, что для успешного проведения фармакологических исследований на грызунах-моделях может оказаться необходимым получение антагонистов MCP-1 грызунов.
MCP-1 представляет собой сильный аттрактант моноцитов/макрофагов, базофилов, активированных Т-клеток и NK-клеток. Клетки различных типов широкого ряда, такие как эндотелиальные клетки, эпителиальные клетки, фибробласты, кератиноциты, синовиальные клетки, мезангиальные клетки, остеобласты, клетки гладких мышц, а также множество опухолевых клеток экспрессируют MCP-1 (Baggiolini 1994). Экспрессия этого белка стимулируется провоспалительными агентами нескольких типов, такими как IL-1β, TNF-α, IFN-γ, ЛПС (липополисахарид) и GM-CSF.
Свойством MCP-1, которое является довольно необычным для всей системы различных хемокинов, является его высокая степень специфичности к своему рецептору, а именно его высокая аффинность связывания только с хемокиновым рецептором CCR2. Подобно всем хемокиновым рецепторами, CCR2 представляет собой GPCR (рецептор, сопряженный с G-белком) (Dawson 2003). CCR2, очевидно, экспрессируется в двух несколько отличающихся формах, образующихся в результате альтернативного сплайсинга мРНК, кодирующей карбокси-концевую область, а именно: CCR2a и CCR2b (Charo 1994). Эти рецепторы экспрессируются в моноцитах, в миелоидных клетках-предшественниках и в активированных T-клетках (Myers 1995; Qin 1996). Константа диссоциации MCP-1, связанного с рецептором, трансфицированным в клетки HEK-293, составляет 260 пM, что соответствует величинам, измеренным на моноцитах (Myers 1995; Van Riper 1993). Активация рецептора CCR2b, присутствующего на трансфицированных клетках HEK-293, белком MCP-1 приводит к ингибированию аденилилциклазы при концентрации 90 пМ и к мобилизации внутриклеточного кальция при несколько более высоких концентрациях и, вероятно, не зависит от гидролиза фосфатидилинозита. Такое влияние на высвобождение аденилилциклазы и внутриклеточного кальция в значительной степени ингибируется коклюшным токсином, что указывает на участие гетеротримерных G-белков типа Gi в передаче сигнала (Myers 1995).
MCP-1 участвует в рекрутинге моноцитов в воспаленной ткани. Присутствующие в этой ткани макрофаги высвобождают хемокины, такие как MCP-1 и т.п., и цитокины, такие как TNF, IL-1β и т.п., которые активируют эндотелиальные клетки, экспрессирующие серию адгезивных молекул. Образовавшийся “липкий” эндотелий опосредует эффект “катящихся” моноцитов, перемещающихся вдоль поверхности сосудистого русла. При этом моноциты сталкиваются с присутствующим на эндотелиальной поверхности MCP-1, который связывается с CCR2 на моноцитах и активирует их. В конечном счете, это приводит к полному прекращению распространения моноцитов вдоль эндотелия и их транспорта в окружающие ткани, где указанные моноциты дифференцируются в макрофаги и мигрируют в область максимальной концентрации MCP-1.
MCP-1 является членом семейства хемокинов, которое представляет собой семейство небольших (примерно 8-14 кДа) гепарин-связывающих, главным образом, основных и структурно родственных молекул. Они образуются преимущественно в воспаленных тканях и регулируют рекрутинг, активацию и пролиферацию лейкоцитов (Baggiolini 1994; Springer 1995; Schall 1994). Хемокины селективно индуцируют хемотаксис нейтрофилов, эозинофилов, базофилов, моноцитов, макрофагов, тучных клеток и Т- и В-клеток. Хемокины, помимо их хемотаксического действия, могут селективно вызывать и другие эффекты в отвечающих клетках, такие как изменение формы клеток, временное увеличение концентрации свободных внутриклеточных ионов кальция, дегрануляция, активация интегринов, образование биологических активных липидов, таких как лейкотриены, простагландины, тромбоксаны, или “дыхательный взрыв” (высвобождение молекул активного кислорода для деструкции патогенных микроорганизмов или опухолевых клеток). Таким образом, посредством индуцирования высвобождения других провоспалительных медиаторов, хемотаксиса и экстравазации лейкоцитов на участке инфицирования или воспаления, хемокины стимулируют усиление воспалительного ответа.
По расположению первых двух из четырех консервативных цистеиновых остатков хемокины подразделяются на четыре класса: CC или β-хемокины, в которых цистеины присутствуют в тандеме; CXC или α-хемокины, которые разделены одним дополнительным аминокислотным остатком; XC или γ-хемокины и лимфотактин, которые были обнаружены лишь недавно и которые имеют только один дисульфидный мостик; и CX3C-хемокины, которые отличаются тем, что они имеют три аминокислотных остатка между цистеинами, причем лишь одним известным до настоящего времени членом этого класса является мембраносвязанный фракталкин (Bazan 1997).
Хемокины CXC, в частности те хемокины CXC, которые имеют у своего амино-конца аминокислотную последовательность ELR, действуют, главным образом, на нейтрофилы. Примерами хемокинов CXC, активных по отношению к нейтрофилам, являются IL-8, GROα, -β и -γ, NAP-2, ENA-78 и GCP-2. Хемокины CC действуют на лейкоциты широкого ряда, такие как моноциты, макрофаги, эозинофилы, базофилы, а также Т- и В-лимфоциты (Oppenheim 1991; Baggiolini 1994; Miller 1992; Jose 1994; Ponath 1996). Примерами таких хемокинов являются I-309, MCP-1, -2, -3, -4, MIP-1α и -β, RANTES и эотаксин.
Хемокины действуют посредством рецепторов, которые принадлежат к суперсемейству из семи трансмембранных рецепторов, связанных с G-белком (GPCR; Murphy 2000). Обычно считается, что взаимодействие хемокина и хемокинового рецептора является неспецифическим, то есть один хемокин может связываться со многими хемокиновыми рецепторами, и наоборот, один хемокиновый рецептор может взаимодействовать с несколькми хемокинами. Некоторыми известными рецепторами для хемокинов CC являются: CCR1, который связывается с MIP-1α и RANTES (Neote 1993; Gao 1993); CCR2, который связывается с хемокинами, включая MCP-1, -2, -3 и -4 (Charo 1994; Myers 1995; Gong 1997; Garcia-Zepeda 1996); CCR3, который связывается с хемокинами, включая эотаксин, RANTES и MCP-3 (Ponath 1996b); CCR4, который, как было обнаружено, передает сигнал в ответ на действие MCP-1, MIP-1α и RANTES (Power 1995); и CCR5, который, как было показано, передает сигнал в ответ на действие MIP-1α и -β и RANTES (Boring 1996; Raport 1996; Samson 1996).
Как указывалось выше, все четыре члена семейства MCP (1-4) связываются с CCR2, тогда как MCP-2, MCP-3 и MCP-4 могут также взаимодействовать с CCR1 и CCR3 (Gong 1997; Heath 1997; Uguccioni 1997), а MCP-2 может взаимодействовать с CCR5 (Ruffing 1998). Другим хемокином СС, обнаруживающим высокую степень гомологии с хемокинами семейства MCP, является эотаксин, который был впервые выделен из жидкости бронхоальвеолярного лаважа, взятого у сенсибилизированных аллергеном морских свинок (Jose 1994). Было показано, что эотаксин также обладает способностью активировать CCR2 (Martinelli 2001).
Проблема, рассматриваемая в настоящем изобретении, заключается в необходимости разработки способов, в основе которых лежит специфическое взаимодействие с МСР-1. Более конкретно, проблема, рассматриваемая в настоящем изобретении, может быть решена с применением способов, основанных на использовании нуклеиновых кислот, специфически взаимодействующих с MCP-1.
Другая проблема, рассматриваемая в настоящем изобретении, заключается в необходимости разработки способа приготовления лекарственного средства для лечения заболеваний человека или животных, где указанное заболевание характеризуется прямым или опосредованным участием MCP-1 в механизме патогенеза этого заболевания.
Еще одна проблема, рассматриваемая в настоящем изобретении, заключается в необходимости разработки способа получения диагностического средства для лечения заболевания, которое характеризуется прямым или опосредованным участием MCP-1 в механизме патогенеза этого заболевания.
Эти и другие проблемы, рассматриваемые в настоящем изобретении, могут быть решены с применением объекта настоящего изобретения, заявленного в независимых пунктах прилагаемой формулы изобретения. Предпочтительные варианты могут быть взяты из зависимых пунктов прилагаемой формулы изобретения.
Проблема, рассматриваемая в настоящем изобретении, может быть также решена в соответствии с первым аспектом изобретения с использованием нуклеиновой кислоты, предпочтительно связывающейся с MCP-1 и выбранной из группы, включающей нуклеиновую кислоту типа 1A, нуклеиновую кислоту типа 1B, нуклеиновую кислоту типа 2, нуклеиновую кислоту типа 3, нуклеиновую кислоту типа 4 и нуклеиновую кислоту, имеющую любую нуклеотидную последовательность, выбранную из последовательностей SEQ ID NО:87-115.
В первом подаспекте первого аспекта изобретения нуклеиновая кислота типа 1A содержит, в направлении 5'→3', первый фрагмент “бокс В1А”, второй фрагмент “бокс В2”, третий фрагмент “бокс В3”, четвертый фрагмент “бокс В4”, пятый фрагмент “бокс В5”, шестой фрагмент “бокс В6” и седьмой фрагмент “бокс В1В”, где
первый фрагмент “бокс B1A” и седьмой фрагмент “B1B бокс” необязательно гибридизуются друг с другом, и в результате такой гибридизации образуется двухцепочечная структура,
первый фрагмент “бокс B1A” содержит нуклеотидную последовательность AGCRUG,
второй фрагмент “бокс B2” содержит нуклеотидную последовательность CCCGGW,
третий фрагмент “бокс В3” содержит нуклеотидную последовательность GUR,
четвертый фрагмент “бокс В4” содержит нуклеотидную последовательность RYA,
пятый фрагмент “бокс В5” содержит нуклеотидную последовательность GGGGGRCGCGAYC,
шестой фрагмент “бокс В6” содержит нуклеотидную последовательность UGCAAUAAUG или URYAWUUG, а
седьмой фрагмент “бокс В1В” содержит нуклеотидную последовательность CRYGCU.
В предпочтительном варианте первого подаспекта изобретения
первый фрагмент “бокс В1А” содержит нуклеотидную последовательность AGCGUG.
В одном из вариантов первого подаспекта изобретения
второй фрагмент “бокс В2” содержит нуклеотидную последовательность CCCGGU.
В одном из вариантов первого подаспекта изобретения
третий фрагмент “бокс В3” содержит нуклеотидную последовательность GUG.
В одном из вариантов первого подаспекта изобретения
четвертый фрагмент “бокс В4” содержит нуклеотидную последовательность GUA.
В одном из вариантов первого подаспекта изобретения
пятый фрагмент “бокс В5” содержит нуклеотидную последовательность GGGGGGCGCGACC.
В одном из вариантов первого подаспекта изобретения
шестой фрагмент “бокс В6” содержит нуклеотидную последовательность UACAUUUG.
В одном из вариантов первого подаспекта изобретения
седьмой фрагмент “бокс В1В” содержит нуклеотидную последовательность CACGCU.
В одном из вариантов первого подаспекта изобретения нуклеиновая кислота содержит нуклеотидную последовательность SEQ ID NО:21.
Во втором подаспекте первого аспекта изобретения нуклеиновая кислота типа 1В содержит, в направлении 5'→3', первый фрагмент “бокс В1А”, второй фрагмент “бокс В2”, третий фрагмент “бокс В3”, четвертый фрагмент “бокс В4”, пятый фрагмент “бокс В5”, шестой фрагмент “бокс В6” и седьмой фрагмент “бокс B1B”, где:
первый фрагмент “бокс В1А” и седьмой фрагмент “бокс B1B” необязательно гибридизуются друг с другом, и в результате такой гибридизации образуется двухцепочечная структура,
первый фрагмент “бокс В1А” содержит нуклеотидную последовательность AGYRUG,
второй фрагмент “бокс В2” содержит нуклеотидную последовательность CCAGCU или CCAGY,
третий фрагмент “бокс В3” содержит нуклеотидную последовательность GUG,
четвертый фрагмент “бокс В4” содержит нуклеотидную последовательность AUG,
пятый фрагмент “бокс В5” содержит нуклеотидную последовательность GGGGGGCGCGACC,
шестой фрагмент “бокс В6” содержит нуклеотидную последовательность CAUUUUA или CAUUUA и
седьмой фрагмент “бокс В1В” содержит нуклеотидную последовательность CAYRCU.
В одном из вариантов второго подаспекта изобретения
первый фрагмент “бокс В1А” содержит нуклеотидную последовательность AGCGUG.
В одном из вариантов второго подаспекта изобретения
второй фрагмент “бокс В2” содержит нуклеотидную последовательность CCAGU.
В одном из вариантов второго подаспекта изобретения
шестой фрагмент “бокс В6” содержит нуклеотидную последовательность CAUUUUA.
В одном из вариантов второго подаспекта изобретения
седьмой фрагмент “бокс В1В” содержит нуклеотидную последовательность CACGCU.
В одном из вариантов второго подаспекта изобретения нуклеиновая кислота содержит нуклеотидные последовательности SEQ ID NO:28 и SEQ ID NO:27.
В третьем подаспекте первого аспекта изобретения нуклеиновая кислота типа 2 содержит, в направлении 5'→3', первый фрагмент “бокс B1A”, второй фрагмент “бокс В2” и третий фрагмент “бокс В1В”, где
первый фрагмент “бокс В1А” и третий фрагмент “бокс В1В” необязательно гибридизуются друг с другом, и в результате такой гибридизации образуется двухцепочечная структура,
первый фрагмент “бокс В1А” содержит нуклеотидную последовательность, выбранную из группы, содержащей ACGCA, CGCA и GCA,
второй фрагмент “бокс В2” содержит нуклеотидную последовательность CSUCCCUCACCGGUGCAAGUGAAGCCGYGGCUC, а
третий фрагмент “бокс В1В” содержит нуклеотидную последовательность, выбранную из группы, содержащей UGCGU, UGCG и UGC.
В одном из вариантов третьего подаспекта изобретения
второй фрагмент “бокс В2” содержит нуклеотидную последовательность CGUCCCUCACCGGUGCAAGUGAAGCCGUGGCUC.
В одном из вариантов третьего подаспекта изобретения
a) первый фрагмент “бокс В1А” содержит нуклеотидную последовательность ACGCA, а
третий фрагмент “бокс В1В” содержит нуклеотидную последовательность UGCGU; или
b) первый фрагмент “бокс В1А” содержит нуклеотидную последовательность CGCA, а
третий фрагмент “бокс В1В” содержит нуклеотидную последовательность UGCG; или
c) первый фрагмент “бокс В1А” содержит нуклеотидную последовательность GCA, а
третий фрагмент “бокс В1В” содержит нуклеотидную последовательность UGC или UGCG.
В одном из вариантов третьего подаспекта изобретения
первый фрагмент “бокс В1А” содержит нуклеотидную последовательность GCA.
В предпочтительном варианте третьего подаспекта изобретения
третий фрагмент “бокс В1В” содержит нуклеотидную последовательность UGCG.
В одном из вариантов третьего подаспекта изобретения нуклеиновая кислота включает в себя нуклеотидные последовательности SEQ ID NO:37, SEQ ID NО:116, SEQ ID NО:117 и SEQ ID NO:278.
В четвертом подаспекте первого аспекта изобретения нуклеиновая кислота типа 3 содержит, в направлении 5'→3', первый фрагмент “бокс В1А”, второй фрагмент “бокс В2А”, третий фрагмент “бокс В3”, четвертый фрагмент “бокс В2В”, пятый фрагмент “бокс В4”, шестой фрагмент “бокс В5A”, седьмой фрагмент “бокс В6”, восьмой фрагмент “бокс В5B” и девятый фрагмент “бокс В1В”, где
первый фрагмент “бокс В1А” и девятый фрагмент “бокс В1В” необязательно гибридизуются друг с другом, и в результате такой гибридизации образуется двухцепочечная структура,
второй фрагмент “бокс В2A” и четвертый фрагмент “бокс В2B” необязательно гибридизуются друг с другом, и в результате такой гибридизации образуется двухцепочечная структура,
шестой фрагмент “бокс В5A” и восьмой фрагмент “бокс В5B” необязательно гибридизуются друг с другом, и в результате такой гибридизации образуется двухцепочечная структура,
первый фрагмент “бокс В1А” содержит нуклеотидную последовательность, выбранную из группы, содержащей GURCUGC, GKSYGC, KBBSC и BNGC,
второй фрагмент “бокс В2А” содержит нуклеотидную последовательность GKMGU,
третий фрагмент “бокс В3” содержит нуклеотидную последовательность KRRAR,
четвертый фрагмент “бокс В2В” содержит нуклеотидную последовательность ACKMC,
пятый фрагмент “бокс В4” содержит нуклеотидную последовательность, выбранную из группы, содержащей CURYGA, CUWAUGA, CWRMGACW и UGCCAGUG,
шестой фрагмент “бокс В5А” содержит нуклеотидную последовательность, выбранную из группы, содержащей GGY и CWGC,
седьмой фрагмент “бокс В6” содержит нуклеотидную последовательность, выбранную из группы, содержащей YAGA, CKAAU и CCUUUAU,
восьмой фрагмент “бокс В5В” содержит нуклеотидную последовательность, выбранную из группы, содержащей GCYR и GCWG, и
девятый фрагмент “бокс В1В” содержит нуклеотидную последовательность, выбранную из группы, содержащей GCAGCAC, GCRSMC, GSVVM и GCNV.
В одном из вариантов четвертого подаспекта изобретения
третий фрагмент “бокс В3” содержит нуклеотидную последовательность GAGAA или UAAAA.
В одном из вариантов четвертого подаспекта изобретения
пятый фрагмент “бокс В4” содержит нуклеотидную последовательность CAGCGACU или CAACGACU.
В одном из вариантов четвертого подаспекта изобретения
пятый фрагмент “бокс В4” содержит нуклеотидную последовательность CAGCGACU, а бокс B3 содержит нуклеотидную последовательность UAAAA.
В одном из вариантов четвертого подаспекта изобретения
пятый фрагмент “бокс В4” содержит нуклеотидную последовательность CAACGACU, а бокс B3 содержит нуклеотидную последовательность GAGAA.
В одном из вариантов четвертого подаспекта изобретения
седьмой фрагмент “бокс В6” содержит нуклеотидную последовательность UAGA.
В одном из вариантов четвертого подаспекта изобретения
a) первый фрагмент “бокс В1А” содержит нуклеотидную последовательность GURCUGC, а
девятый фрагмент “бокс В1В” содержит нуклеотидную последовательность GCAGCAC; или
b) первый фрагмент “бокс В1А” содержит нуклеотидную последовательность GKSYGC, а
девятый фрагмент “бокс В1В” содержит нуклеотидную последовательность GCRSMC; или
c) первый фрагмент “бокс В1А” содержит нуклеотидную последовательность KBBSC, а
девятый фрагмент “бокс В1В” содержит нуклеотидную последовательность GSVVM; или
d) первый фрагмент “бокс В1А” содержит нуклеотидную последовательность BNGC, а
девятый фрагмент “бокс В1В” содержит нуклеотидную последовательность GCNV.
В предпочтительном варианте четвертого подаспекта изобретения
a) первый фрагмент “бокс В1А” содержит нуклеотидную последовательность GUGCUGC, а
девятый фрагмент “бокс В1В” содержит нуклеотидную последовательность GCAGCAC; или
b) первый фрагмент “бокс В1А” содержит нуклеотидную последовательность GUGCGC, а
девятый фрагмент “бокс В1В” содержит нуклеотидную последовательность GCGCAC; или
c) первый фрагмент “бокс В1А” содержит нуклеотидную последовательность KKSSC, а
девятый фрагмент “бокс В1В” содержит нуклеотидную последовательность GSSMM; или
d) первый фрагмент “бокс В1А” содержит нуклеотидную последовательность SNGC, а
девятый фрагмент “бокс В1В” содержит нуклеотидную последовательность GCNS.
В другом предпочтительном варианте четвертого подаспекта изобретения
первый фрагмент “бокс В1А” содержит нуклеотидную последовательность GGGC, а
девятый фрагмент “бокс В1В” содержит нуклеотидную последовательность GCCC.
В одном из вариантов четвертого подаспекта изобретения второй фрагмент “бокс В2A” содержит нуклеотидную последовательность GKMGU, а четвертый фрагмент “бокс В2В” включает нуклеотидную последовательность ACKMC.
В предпочтительном варианте четвертого подаспекта изобретения второй фрагмент “бокс В2A” содержит нуклеотидную последовательность GUAGU, а четвертый фрагмент “бокс В2B” содержит нуклеотидную последовательность ACUAC.
В одном из вариантов четвертого подаспекта изобретения
a) шестой фрагмент “бокс В5A” содержит нуклеотидную последовательность GGY, а
восьмой фрагмент “бокс В5B” содержит нуклеотидную последовательность GCYR; или
b) шестой фрагмент “бокс В5A” содержит нуклеотидную последовательность CWGC, а
восьмой фрагмент “бокс В5B” содержит нуклеотидную последовательность GCWG.
В предпочтительном варианте четвертого подаспекта изобретения
шестой фрагмент “бокс В5A” содержит нуклеотидную последовательность GGC, а
восьмой фрагмент “бокс В5B” содержит нуклеотидную последовательность GCCG.
В более предпочтительном варианте четвертого подаспекта изобретения шестой фрагмент “бокс В5A” гибридизуется с нуклеотидами GCY восьмого фрагмента “бокс B5B”.
В одном из вариантов четвертого подаспекта изобретения нуклеиновая кислота содержит нуклеотидную последовательность SEQ ID NO:56.
В одном из вариантов четвертого подаспекта изобретения нуклеиновая кислота содержит нуклеотидную последовательность, выбранную из группы, содержащей нуклеотидные последовательности SEQ ID NO:57-61, SEQ ID NO:67-71 и SEQ ID NO:73.
В пятом подаспекте первого аспекта изобретения нуклеиновая кислота типа 4 содержит, в направлении 5'→3', первый фрагмент “бокс В1А”, второй фрагмент “бокс В2” и третий фрагмент “бокс В1В”, где
первый фрагмент “бокс В1А” и третий фрагмент “бокс В1В” необязательно гибридизуются друг с другом, и в результате такой гибридизации образуется двухцепочечная структура,
первый фрагмент “бокс В1А” содержит нуклеотидную последовательность, выбранную из группы, содержащей AGCGUGDU, GCGCGAG, CSKSUU, GUGUU и UGUU;
второй фрагмент “бокс В2” содержит нуклеотидную последовательность, выбранную из группы, содержащей AGNDRDGBKGGURGYARGUAAAG, AGGUGGGUGGUAGUAAGUAAAG и CAGGUGGGUGGUAGAAUGUAAAGA, а
третий фрагмент “бокс В1В” содержит нуклеотидную последовательность, выбранную из группы, содержащей GNCASGCU, CUCGCGUC, GRSMSG, GRCAC и GGCA.
В одном из вариантов пятого подаспекта изобретения
a) первый фрагмент “бокс В1А” содержит нуклеотидную последовательность GUGUU, а
третий фрагмент “бокс В1В” содержит нуклеотидную последовательность GRCAC;
b) первый фрагмент “бокс В1А” содержит нуклеотидную последовательность GCGCGAG, а
третий фрагмент “бокс В1В” содержит нуклеотидную последовательность CUCGCGUC; или
c) первый фрагмент “бокс В1А” содержит нуклеотидную последовательность CSKSUU, а
третий фрагмент “бокс В1В” содержит нуклеотидную последовательность GRSMSG; или
d) первый фрагмент “бокс В1А” содержит нуклеотидную последовательность UGUU, а
третий фрагмент “бокс В1В” содержит нуклеотидную последовательность GGCA; или
e) первый фрагмент “бокс В1А” содержит нуклеотидную последовательность AGCGUGDU, а
третий фрагмент “бокс В1В” содержит нуклеотидную последовательность GNCASGCU.
В предпочтительном варианте пятого подаспекта изобретения первый фрагмент “бокс В1А” содержит нуклеотидную последовательность CSKSUU, а третий фрагмент “бокс В1В” содержит нуклеотидную последовательность GRSMSG.
В более предпочтительном варианте пятого подаспекта изобретения первый фрагмент “бокс В1А” содержит нуклеотидную последовательность CCGCUU, а третий фрагмент “бокс В1В” содержит нуклеотидную последовательность GGGCGG.
В одном из вариантов пятого подаспекта изобретения
второй фрагмент “бокс В2” содержит нуклеотидную последовательность AGGUGGGUGGUAGUAAGUAAAG.
В одном из вариантов пятого подаспекта изобретения нуклеиновая кислота содержит нуклеотидную последовательность SEQ ID NO:80.
В одном из вариантов 1-5 подаспектов изобретения нуклеиновая кислота обладает способностью связываться с MCP-1, предпочтительно с человеческим MCP-1.
В одном из вариантов 1-5 подаспектов изобретения нуклеиновая кислота обладает способностью связываться с хемокином, где указанный хемокин выбран из группы, включающей эотаксин, MCP-1, MCP-2 и MCP-3.
В одном из вариантов 1-5 подаспектов изобретения нуклеиновая кислота обладает способностью связываться с хемокином, где указанный хемокин выбран из группы, включающей человеческий эотаксин, человеческий MCP-1, человеческий MCP-2 и человеческий MCP-3.
В одном из вариантов 1-5 подаспектов изобретения нуклеиновая кислота обладает способностью связываться с MCP-1, где указанный MCP-1 предпочтительно выбран из группы, включающей обезьяний MCP-1, лошадиный MCP-1, кроличий MCP-1, коровий MCP-1, собачий MCP-1, свиной MCP-1 и человеческий MCP-1.
В одном из вариантов 1-5 подаспектов изобретения нуклеиновая кислота обладает способностью связываться с человеческим MCP-1.
В предпочтительном варианте 1-5 подаспектов изобретения MCP-1 имеет аминокислотную последовательность SEQ ID NO:1.
Проблема, рассматриваемая в настоящем изобретении, может быть решена в соответствии со вторым аспектом настоящего изобретения с использованием нуклеиновой кислоты, предпочтительно связывающейся с мышиным MCP-1, где указанная нуклеиновая кислота содержит нуклеотидную последовательность SEQ ID NO:122, SEQ ID NO:253 и SEQ ID NO:254.
Проблема, рассматриваемая в настоящем изобретении, может быть решена в соответствии с третьим аспектом настоящеего изобретения с использованием нуклеиновой кислоты, предпочтительно связывающейся с мышиным MCP-1, где указанная нуклеиновая кислота содержит нуклеотидную последовательность SEQ ID NO:127.
В одном из вариантов второго и третьего аспектов изобретения мышиный MCP-1 содержит аминокислотную последовательность SEQ ID NO:2.
В одном из вариантов 1-3 аспектов изобретения нуклеиновая кислота имеет модификацию, где указанная модификация предпочтительно представляет собой высокомолекулярную группу, и/или указанная модификация предпочтительно позволяет изменять свойства нуклеиновой кислоты согласно первому, второму или третьему аспектам изобретения, а именно время ее присутствия в организме животного или человека, предпочтительно в организме человека.
В предпочтительном варианте 1-3 аспектов изобретения указанная модификация выбрана из группы, включающей группу ГЭК и группу ПЭГ.
В более предпочтительном варианте 1-3 аспектов изобретения указанной модификацией является молекула ПЭГ, состоящая из прямого или разветвленного ПЭГ, где молекулярная масса указанной молекулы ПЭГ предпочтительно составляет примерно от 20 до 120 кД, более предпочтительно примерно от 30 до 80 кД, а наиболее предпочтительно примерно 40 кД.
В альтернативном более предпочтительном варианте 1-3 аспектов изобретения указанной модификацией является молекула ГЭК, где молекулярная масса указанной молекулы ГЭК предпочтительно составляет примерно от 10 до 130 кД, более предпочтительно примерно от 30 до 130 кД, а наиболее предпочтительно примерно 100 кД.
В одном из вариантов 1-3 аспектов изобретения указанная модифицирующая молекула присоединена к нуклеиновой кислоте посредством линкера.
В одном из вариантов 1-3 аспектов изобретения указанная модифицирующая молекула присоединена к нуклеиновой кислоте у ее 5'-концевого нуклеотида и/или у ее 3'-концевого нуклеотида, и/или к нуклеотиду указанной нуклеиновой кислоты между 5'-концевым нуклеотидом и 3'-концевым нуклеотидом.
В одном из вариантов 1-3 аспектов изобретения нуклеотидами указанной нуклеиновой кислоты или нуклеотидами, образующими указанную нуклеиновую кислоту, являются L-нуклеотиды.
В одном из вариантов 1-3 аспектов изобретения указанной нуклеиновой кислотой является L-нуклеиновая кислота.
В одном из вариантов 1-3 аспектов изобретения указанная молекула нуклеиновой кислоты, обладающая способностью связываться с MCP-1, состоит из L-нуклеотидов.
Проблема, рассматриваемая в настоящем изобретении, может быть решена в соответствии с четвертым аспектом изобретения с использованием фармацевтической композиции, содержащей нуклеиновую кислоту согласно первому, второму и третьему аспектам изобретения и, необязательно, дополнительный компонент, где указанный дополнительный компонент выбран из группы, включающей себя фармацевтически приемлемые наполнители, фармацевтически приемлемые носители и фармацевтически активные средства.
В одном из вариантов четвертого аспекта изобретения указанная фармацевтическая композиция включает в себя нуклеиновую кислоту согласно 1-3 аспектам изобретения и фармацевтически приемлемый носитель.
Проблема, рассматриваемая в настоящем изобретении, может быть решена в соответствии с пятым аспектом с применением нуклеиновой кислоты согласно первому, второму и третьему аспектам изобретения, используемой для получения лекарственного средства.
В одном из вариантов пятого аспекта изобретения указанное лекарственное средство может быть использовано в медицине или в ветеринарии.
Проблема, рассматриваемая в настоящем изобретении, может быть решена в соответствии с шестым аспектом изобретения с применением нуклеиновой кислоты согласно первому, второму и третьему аспектам изобретения, используемой для получения диагностических средств.
В одном из вариантов пятого аспекта изобретения и в одном из вариантов шестого аспекта изобретения лекарственное средство и диагностическое средство соответственно предназначены для лечения и/или предупреждения и диагностики соответственно заболеваний или расстройств, выбранных из группы, включающей в себя воспалительные заболевания, аутоиммунные заболевания, аутоиммунный энцефаломиелит, инсульт, острый и хронический рассеянный склероз, хроническое воспалительное заболевание, ревматоидный артрит, болезни почек, рестеноз, рестеноз после ангиопластики, острые и хронические аллергические реакции, первичные и вторичные иммунологические или аллергические реакции, астму, конъюнктивит, бронхит, рак, атеросклероз, атеросклеротическую сердечно-сосудистую недостаточность или инсульт, псориаз, псориатический артрит, воспаление нервной системы, атопический дерматит, колит, эндометриоз, увеит, заболевания сетчатки, включая дегенерацию желтого пятна, отслоение сетчатки, диабетическую ретинопатию, ретролентальную фиброплазию, пигментный ретинит, пролиферирующую витреоретинопатию и центральную серозную хориоретинопатию; идеопатический фиброз легких, саркоидоз, полимиозит и дерматомиозит; для профилактики иммуносупрессии и снижения риска инфицирования; а также для лечения сепсиса, воспаления почек, гломерулонефрита, быстропрогрессирующего гломерулонефрита, пролиферирующего гломерулонефрита, диабетической нефропатии, обструктивной нефропатии, острого тубулярного некроза и диффузного гломерулосклероза, системной красной волчанки, хронического бронхита, болезни Бехчета, амиотрофического бокового склероза (АБС), раннего атеросклероза, ассоциированного с болезнью Кавазаки, инфаркта миокарда, ожирения, хронической болезни печени, болезни Пейрони, острого поражения хорды спинного мозга, заболеваний, ассоциированных с трансплантацией легких или почек, миокардита, болезни Альцгеймера и невропатии, карциномы молочной железы, карциномы желудка, рака мочевого пузыря, рака яичника, гамартомы, карциномы прямой и ободочной кишки, аденомы толстой кишки, панкреатита, хронической обструктивной болезни легких (ХОБЛ) и воспалительных заболеваний кишечника, таких как болезнь Крона или язвенный колит.
Не ограничиваясь какой-либо конкретной теорией, следует отметить, что применение нуклеиновых кислот согласно изобретению в диагностических целях основано, главным образом, на повышенном или пониженном уровне хемокинов, где указанный хемокин выбран из группы, включающей эотаксин, MCP-1, MCP-2 и MCP-3, причем более предпочтительным является MCP-1. Для специалиста в данной области очевидно, что большинство из вышеупомянутых заболеваний характеризуется таким повышенным или пониженным уровнем хемокинов.
Проблема, рассматриваемая в настоящем изобретении, может быть решена в соответствии с седьмым аспектом настоящего изобретения с использованием комплекса, включающего в себя хемокин и нуклеиновую кислоту согласно первому, второму и третьему аспектам изобретения, где указанный хемокин выбран из группы, включающей в себя эотаксин, MCP-1, MCP-2 и MCP-3, при этом предпочтительным комплексом является кристаллический комплекс.
В одном из вариантов седьмого аспекта изобретения указанный хемокин выбран из группы, включающей в себя человеческий эотаксин, человеческий MCP-1, человеческий MCP-2 и человеческий MCP-3.
В одном из вариантов седьмого аспекта изобретения указанным хемокином является MCP-1, предпочтительно выбранный из группы, включающей в себя человеческий МСР-1, обезьяний MCP-1, лошадиный MCP-1, кроличий MCP-1, коровий MCP-1, собачий MCP-1 и свиной MCP-1, при этом более предпочтительным является человеческий MCP-1.
Проблема, рассматриваемая в настоящем изобретении, может быть решена в соответствии с восьмым аспектом настоящего изобретения с использованием нуклеиновой кислоты согласно первому, второму и третьему аспектам изобретения для детектирования хемокина, где указанный хемокин выбран из группы, включающей в себя эотаксин, MCP-1, MCP-2 и MCP-3.
В одном из вариантов восьмого аспекта изобретения указанный хемокин выбран из группы, включающей в себя человеческий эотаксин, человеческий MCP-1, человеческий MCP-2 и человеческий MCP-3.
В одном из вариантов восьмого аспекта изобретения указанным хемокином является MCP-1, где указанный МСР-1 предпочтительно выбран из группы, включающей в себя человеческий МСР-1, обезьяний MCP-1, лошадиный MCP-1, кроличий MCP-1, коровий MCP-1, собачий MCP-1 и свиной MCP-1, при этом более предпочтительным является человеческий MCP-1.
Проблема, рассматриваемая в настоящем изобретении, может быть решена в соответствии с девятым аспектом настоящего изобретения с применением способа скрининга антагониста хемокина или агониста хемокина, где указанный способ включает следующие стадии:
- получение кандидата на антагонист и/или кандидата на агонист хемокина,
- получение нуклеиновой кислоты согласно первому, второму или третьему аспектам изобретения,
- получение тест-системы, которая обеспечивает детекцию сигнала в присутствии антагониста хемокина и/или агониста хемокина, и
- установление того факта, является ли данный кандидат антагонистом хемокина и/или агонистом хемокина,
где указанный хемокин выбран из группы, включающей в себя эотаксин, MCP-1, MCP-2 и MCP-3.
В одном из вариантов девятого аспекта изобретения указанный хемокин выбран из группы, включающей в себя человеческий эотаксин, человеческий MCP-1, человеческий MCP-2 и человеческий MCP-3.
В одном из вариантов девятого аспекта изобретения указанным хемокином является MCP-1, где указанный МСР-1 предпочтительно выбран из группы, включающей в себя человеческий МСР-1, обезьяний MCP-1, лошадиный MCP-1, кроличий MCP-1, коровий MCP-1, собачий MCP-1 и свиной MCP-1, при этом более предпочтительным является человеческий MCP-1.
Проблема, рассматриваемя в настоящем изобретении, может быть решена в соответствии с десятым аспектом настоящего изобретения с применением способа скрининга агониста хемокинов или антагониста хемокина, где указанный способ включает следующие стадии:
- получение хемокина, иммобилизованного на фазе, предпочтительно на твердой фазе,
- получение нуклеиновой кислоты согласно первому, второму или третьему аспектам изобретения, где указанная нуклеиновая кислота согласно первому аспекту изобретения предпочтительно является меченой,
- добавление кандидата на агонист хемокина и/или кандидата на антагонист хемокина, и
- определение того, является ли данный кандидат агонистом хемокина и/или антагонистом хемокина,
где указанный хемокин выбран из группы, включающей в себя эотаксин, MCP-1, MCP-2 и MCP-3.
В одном из вариантов десятого аспекта изобретения указанную стадию определения осуществляют для того, чтобы установить, заменена ли указанная нуклеиновая кислота кандидатом на агонист хемокина или кандидатом на антагонист хемокина.
В одном из вариантов десятого аспекта изобретения указанный хемокин выбран из группы, включающей в себя человеческий эотаксин, человеческий MCP-1, человеческий MCP-2 и человеческий MCP-3.
В одном из вариантов десятого аспекта изобретения указанным хемокином является MCP-1, где указанный МСР-1 предпочтительно выбран из группы, включающей в себя человеческий МСР-1, обезьяний MCP-1, лошадиный MCP-1, кроличий MCP-1, коровий MCP-1, собачий MCP-1 и свиной MCP-1, при этом более предпочтительным является человеческий MCP-1.
Проблема, рассматриваемая в настоящем изобретении, может быть решена в соответствии с одиннадцатым аспектом настоящего изобретения с применением набора для детектирования хемокина, где указанный набор включает нуклеиновую кислоту согласно первому, второму и третьему аспектам изобретения и где указанный хемокин выбран из группы, включающей в себя эотаксин, MCP-1, MCP-2 и MCP-3.
В одном из вариантов одиннадцатого аспекта изобретения указанный хемокин выбран из группы, включающей в себя человеческий эотаксин, человеческий MCP-1, человеческий MCP-2 и человеческий MCP-3.
В одном из вариантов одиннадцатого аспекта изобретения указанным хемокином является MCP-1, где указанный МСР-1 предпочтительно выбран из группы, включающей в себя человеческий МСР-1, обезьяний MCP-1, лошадиный MCP-1, кроличий MCP-1, коровий MCP-1, собачий MCP-1 и свиной MCP-1, при этом более предпочтительным является человеческий MCP-1.
Проблема, рассматриваемая в настоящем изобретении, может быть решена в соответствии с двенадцатым аспектом настоящего изобретения с применением антагониста хемокина, полученного способом согласно десятому или девятому аспектам изобретения, где указанный хемокин выбран из группы, включающей в себя эотаксин, MCP-1, MCP-2 и MCP-3.
В одном из вариантов двенадцатого аспекта изобретения указанный хемокин выбран из группы, включающей в себя человеческий эотаксин, человеческий MCP-1, человеческий MCP-2 и человеческий MCP-3.
В одном из вариантов двенадцатого аспекта изобретения указанным хемокином является MCP-1, где указанный МСР-1 предпочтительно выбран из группы, включающей в себя человеческий МСР-1, обезьяний MCP-1, лошадиный MCP-1, кроличий MCP-1, коровий MCP-1, собачий MCP-1 и свиной MCP-1, при этом более предпочтительным является человеческий MCP-1.
Проблема, рассматриваемая в настоящем изобретении, может быть решена в соответствии с тринадцатым аспектом настоящего изобретения с применением агониста хемокина, полученного способом согласно десятому или девятому аспектам изобретения, где указанный хемокин выбран из группы, включающей в себя эотаксин, MCP-1, MCP-2 и MCP-3.
В одном из вариантов тринадцатого аспекта изобретения указанный хемокин выбран из группы, включающей в себя человеческий эотаксин, человеческий MCP-1, человеческий MCP-2 и человеческий MCP-3.
В одном из вариантов тринадцатого аспекта изобретения указанным хемокином является MCP-1, где указанный МСР-1 предпочтительно выбран из группы, включающей в себя человеческий МСР-1, обезьяний MCP-1, лошадиный MCP-1, кроличий MCP-1, коровий MCP-1, собачий MCP-1 и свиной MCP-1, при этом более предпочтительным является человеческий MCP-1.
Для специалиста в данной области очевидно, что агонистом хемокина и/или антагонистом хемокина предпочтительно является агонист и антагонист соответственно, стимулирующий и ингибирующий соответствующий описанный здесь хемокин. В соответствии с этим указанным агонистом и антагонистом хемокина является, например, агонист MCP-1 и антагонист MCP-1 соответственно.
Проблема, рассматриваемая в настоящем изобретении, может быть решена в соответствии с четырнадцатым аспектом настоящего изобретения с применением способа детектирования нуклеиновой кислоты согласно первому, второму или третьему аспектам изобретения в образце, где указанный способ включает стадии:
а) получение образца, содержащего нуклеиновую кислоту согласно изобретению;
b) получение зонда для захвата, где указанный зонд для захвата по меньшей мере частично комплементарен первой части нуклеиновой кислоты согласно первому, второму или третьему аспекту изобретения, и детектирующего зонда, где указанный детектирующий зонд по меньшей мере частично комплементарен второй части нуклеиновой кислоты согласно первому, второму или третьему аспекту изобретения, или, альтернативно, указанный зонд для захвата по меньшей мере частично комплементарен второй части нуклеиновой кислоты согласно первому, второму или третьему аспекту изобретения, а указанный детектирующий зонд по меньшей мере частично комплементарен первой части нуклеиновой кислоты согласно первому, второму или третьему аспекту изобретения;
c) осуществление реакции указанного зонда для захвата и детектирующего зонда либо одновременно, либо последовательно в любом порядке с нуклеиновой кислотой согласно первому, второму или третьему аспекту изобретения или с ее частью;
d) необязательное детектирование гибридизации указанного зонда для захвата с нуклеиновой кислотой согласно первому, второму или третьему аспекту изобретения, описанной на стадии а); и
e) детектирование комплекса, полученного, как описано на стадии (с), и состоящего из нуклеиновой кислоты согласно первому, второму или третьему аспекту изобретения и зонда для захвата и детектирующего зонда.
В одном из вариантов четырнадцатого аспекта изобретения указанный детектирующий зонд включает детектируемое средство, и/или указанный зонд для захвата может быть иммобилизован на носителе, предпочтительно на твердом носителе.
В одном из вариантов четырнадцатого аспекта изобретения любой детектирующий зонд, который не является частью данного комплекса, удаляют из реакционной смеси, в результате чего на стадии (е) детектируется только детектирующий зонд, который является частью указанного комплекса.
В одном из вариантов четырнадцатого аспекта изобретения стадия (е) включает в себя стадию сравнения сигнала, вырабатываемого детектируемым средством в том случае, если зонд для захвата и детектирующий зонд гибридизуются в присутствии нуклеиновой кислоты согласно первому, второму или третьему аспекту изобретения или ее части и в отсутствие указанной нуклеиновой кислоты или ее части.
В одном из вариантов четырнадцатого аспекта изобретения детектируемой нуклеиновой кислотой является нуклеиновая кислота, имеющая нуклеотидную последовательность SEQ ID NО:37, 116, 117 или 278, а зонд для захвата или детектирующий зонд содержит нуклеотидную последовательность SEQ ID NO:255 или SEQ ID NO:256.
В одном из вариантов четырнадцатого аспекта изобретения детектируемой нуклеиновой кислотой является нуклеиновая кислота, имеющая нуклеотидную последовательность SEQ ID NО:122, 253 или 254, а зонд для захвата или детектирующий зонд содержит нуклеотидную последовательность SEQ ID NO:281 и SEQ ID NO:282.
Проблема, рассматриваемая в настоящем изобретении, может быть решена с применением объекта настоящего изобретения, заявленного в независимых пунктах прилагаемой формулы изобретения. Предпочтительный вариант может быть взят из зависимых пунктов прилагаемой формулы изобретения.
Отличительные свойства нуклеиновой кислоты согласно изобретению, описанной в настоящей заявке, могут быть реализованы специалистом в любом аспекте настоящего изобретения, где указанная нуклеиновая кислота используется отдельно или в любой комбинации.
Человеческий MCP-1, а также мышиный MCP-1 представляют собой основные белки, имеющие аминокислотную последовательность SEQ ID NО:1 и 2 соответственно.
Обнаружение того факта, что могут быть идентифицированы короткие нуклеиновые кислоты, обладающие высокой аффинностью связывания с MCP-1, было полной неожиданностью, поскольку согласно наблюдениям Eaton et al. (1997) считалось, что генерирование аптамеров, то есть D-нуклеиновых кислот, связывающихся с молекулой-мишенью и направленных на основный белок, вообще говоря, представляет значительные трудности, поскольку мишень такого типа продуцирует высокое и неспецифическое отношение “сигнал-шум”. Такое высокое отношение “сигнал-шум” является результатом высокой неспецифической аффинности, которой обладают нуклеиновые кислоты по отношению к основным молекулам-мишеням, таким как MCP-1.
Как более подробно описано в формуле изобретения и в примере 1, еще более неожиданным для авторов настоящего изобретения было обнаружение ряда различных MCP-1-связывающих молекул нуклеиновой кислоты, в результате чего большинство нуклеиновых кислот могло быть охарактеризовано по их нуклеотидным фрагментам, также называемым здесь “боксами”. Различные MCP-1-связывающие молекулы нуклеиновой кислоты могут быть подразделены на категории, исходя из указанных боксов и некоторых структурных признаков и элементов соответственно. Различные категории нуклеиновых кислот, определенные таким образом, также называются здесь типами нуклеиновых кислот, более конкретно типом 1А, типом 1В, типом 2, типом 3 и типом 4.
Нуклеиновые кислоты согласно изобретению также включают нуклеиновые кислоты, которые по существу гомологичны описанным здесь конкретным последовательностям. Термин “по существу гомологичный” означает, что такая гомология составляет по меньшей мере 75%, предпочтительно 85%, более предпочтительно 90%, а наиболее предпочтительно более чем 95%, 96%, 97%, 98% или 99%.
Фактический процент гомологичных нуклеотидов, присутствующих в нуклеиновой кислоте согласно изобретению, зависит от общего числа нуклеотидов, присутствующих в нуклеиновой кислоте. Процент модификации может быть вычислен, исходя из общего числа нуклеотидов, присутствующих в нуклеиновой кислоте.
Гомология может быть определена методом, известным специалистам в данной области. Более конкретно алгоритм сравнения последовательностей позволяет вычислять процент идентичности тестируемой(ых) последовательности(ей) со сравниваемой последовательностью, исходя из расчетных параметров программы. Тестируемой последовательностью предпочтительно является последовательность или молекула нуклеиновой кислоты, которая, как считается или как может быть проанализировано, является гомологичной или до некоторой степени гомологичной другой молекуле нуклеиновой кислоты, где указанная другая молекула нуклеиновой кислоты также называется эталонной последовательностью. В одном из вариантов изобретения эталонной последовательностью является молекула нуклеиновой кислоты, описанная в настоящей заявке, более предпочтительно молекула нуклеиновой кислоты, имеющая любую из последовательностей SEQ ID NО:10-129, 132-256 и 278-282. Оптимальное выравнивание последовательностей для сравнения может быть осуществлено, например, с использованием алгоритма локальной гомологии Смита и Уотермана (Smith & Waterman, 1981) или алгоритма выравнивания для выявления гомологии Ниделмана и Вюнша (Needleman & Wunsch, 1970), с использованием метода поиска сходства Пирсона и Липмана (Pearson & Lipman, 1988) и с использованием компьютерных устройств для реализации указанных алгоритмов (GAP, BESTFIT, FASTA и TFASTA в пакете программ для генетических исследований Wisconsin Genetics Software Package, Genetics Computer Group, 575 Science Dr., Madison, Wis.) или путем визуального наблюдения.
Одним из примеров алгоритма, подходящего для определения процента идентичности последовательности, является алгоритм, используемый в качестве основного инструментального средства поиска путем локального выравнивания последовательностей (далее называемый "BLAST") (см., например, Altschul et al. 1990 и Altschul et al. 1997). Программа для осуществления BLAST-анализов является общедоступной и имеется на сайте Национального центра биотехнологической информации (далее обозначаемого “NCBI"). Параметры по умолчанию, используемые для определения идентичности последовательностей с помощью компьютерной программы, имеющейся в NCBI, например программы BLASTN (для нуклеотидных последовательностей) и программы BLASTP (для аминокислотных последовательностей), описаны McGinnis et al. (McGinnis et al., 2004).
Термин “заявленная нуклеиновая кислота” или “нуклеиновая кислота согласно изобретению” также включает нуклеиновые кислоты, содержащие нуклеотидные последовательности, описанные в настоящей заявке или их части, предпочтительно такие нуклеиновые кислоты или их части, которые участвуют в связывании с MCP-1. В одном из вариантов изобретения предпочтительно используемый здесь термин “заявленная нуклеиновая кислота” также включает нуклеиновую кислоту, связывающуюся с любой молекулой, выбранной из группы, включающей в себя MCP-2, MCP-3, MCP-4 и эотаксин. Специалистам в данной области очевидно, что отдельные нуклеиновые кислоты согласно изобретению связываются с одной или несколькими такими молекулами. В одном из вариантов изобретения такой нуклеиновой кислотой является одна из описанных здесь молекул нуклеиновой кислоты или ее производное и/или метаболит, где, в отличие от описанных здесь молекул нуклеиновой кислоты, указанное производное и/или метаболит предпочтительно представляют собой усеченную нуклеиновую кислоту. Такое усечение может присутствовать на любом конце или на обоих концах нуклеиновых кислот, описанных в настоящей заявке. Кроме того, такое усечение может присутствовать во внутренней последовательности нуклеиновой кислоты, то есть оно может присутствовать в положении(ях) нуклеотида(ов), находящемся(ихся) между 5'- и 3'-концевыми нуклеотидами соответственно. Более того, такое усечение может включать делецию по меньшей мере одного нуклеотида в описанной здесь последовательности нуклеиновых кислот. Такое усечение может также относится к более чем одному фрагменту заявленной(ых) нуклеиновой(ых) кислоты (кислот), где указанный фрагмент может иметь длину по меньшей мере в один нуклеотид. Связывание нуклеиновой кислоты согласно изобретению предпочтительно с молекулой, выбранной из группы, включающей в себя MCP-1, MCP-2, MCP-3, MCP-4 и эотаксин, может быть определено специалистами с помощью рутинных экспериментов или с применением либо описанного здесь способа, либо известного способа, а предпочтительно способа, описанного в экспериментальной части настоящего изобретения. В одном из вариантов изобретения, относящегося к связыванию нуклеиновых кислот согласно изобретению с МСР-1, если это не оговорено особо, такое связывание также означает связывание нуклеиновых кислот согласно изобретению с любой молекулой, выбранной из группы, включающей в себя MCP-2, MCP-3, MCP-4 и эотаксин.
Нуклеиновые кислоты согласно изобретению могут представлять собой D-нуклеиновые кислоты или L-нуклеиновые кислоты. Предпочтительными заявленными нуклеиновыми кислотами являются L-нуклеиновые кислоты. Кроме того, одна или несколько частей нуклеиновой кислоты могут присутствовать в виде D-нуклеиновых кислот, либо по меньшей мере одна или несколько частей нуклеиновых кислот представляют собой L-нуклеиновые кислоты. Термин “часть” нуклеиновых кислот означает по меньшей мере один нуклеотид. Такие нуклеиновые кислоты обычно называются здесь D- и L-нуклеиновыми кислотами соответственно. Следовательно, в особенно предпочтительном варианте изобретения нуклеиновые кислоты согласно изобретению состоят из L-нуклеотидов и включают по меньшей мере один D-нуклеотид. Такой D-нуклеотид предпочтительно присоединен к части, отличающейся от фрагментов, определяющих нуклеиновые кислоты согласно изобретению, а предпочтительно к тем частям, где происходит взаимодействие с другими частями нуклеиновой кислоты. Предпочтительно такой D-нуклеотид присоединен к концу любого из фрагментов и любой нуклеиновой кислоты согласно изобретению соответственно. В другом предпочтительном варианте изобретения такие D-нуклеотиды могут действовать в качестве спейсера или линкера, предпочтительно связывающего модифицирующие молекулы, такие как ПЭГ и ГЭК, с нуклеиновыми кислотами согласно изобретению.
Кроме того, в любом из вариантов настоящего изобретения каждая и любая из молекул нуклеиновой кислоты, описанных здесь во всей своей полноте и определяемых термином “последовательность(и) нуклеиновой(ых) кислоты (кислот)”, ограничивается конкретной(ыми) нуклеотидной(ыми) последовательностью(ями). Другими словами, в таком варианте изобретения термины “включающий или “включает(ют)” должны быть интерпретированы как “содержащий” или “состоящий из”.
Кроме того, в соответствии с настоящим изобретением заявленные нуклеиновые кислоты являются частью более длинной нуклеиновой кислоты, где указанная более длинная нуклеиновая кислота включает несколько частей, и по меньшей мере одной из таких частей является нуклеиновая кислота согласно изобретению или ее часть. Другой(ими) частью(ями) указанных более длинных нуклеиновых кислот может быть любая одна или несколько D- нуклеиновых кислот, либо одна или несколько L-нуклеиновых кислот. В соответствии с настоящим изобретением может быть использована любая их комбинация. Указанная(ые) другая(ие) часть(и) более длинной нуклеиновой кислоты, взятая(ые) отдельно или вместе, либо во всей своей полноте, либо в конкретной комбинации, может иметь функцию, отличающуюся от связывания, предпочтительно от связывания с MCP-1. Одной из возможных функций является взаимодействие с другими молекулами, которые предпочтительно отличаются от MCP-1, такое как, например, иммобилизация, перекрестное связывание, детектирование или амплификация. В другом варианте изобретения нуклеиновые кислоты согласно изобретению включают, в виде отдельных или комбинированных молекул, несколько нуклеиновых кислот согласно изобретению. Такие нуклеиновые кислоты, включающие несколько нуклеиновых кислот согласно изобретению, также охватываются термином “более длинная нуклеиновая кислота”.
Используемыми здесь L-нуклеиновыми кислотами являются нуклеиновые кислоты, состоящие из L-нуклеотидов, предпочтительно полностью состоящие из L-нуклеотидов.
Используемыми здесь D-нуклеиновыми кислотами являются нуклеиновые кислоты, состоящие из D-нуклеотидов, предпочтительно полностью состоящие из D-нуклеотидов.
Используемые здесь термины “нуклеиновая кислота” и “молекула нуклеиновой кислоты” являются взаимозаменяемыми, если это не оговорено особо.
Кроме того, если это не оговорено особо, то любая описанная здесь нуклеотидная последовательность расположена в направлении 5'→3'.
Независимо от того, состоит ли заявленная нуклеиновая кислота из D-нуклеотидов, L-нуклеотидов или их комбинаций, где указанной комбинацией может быть, например, произвольная комбинация или определенная последовательность фрагментов, состоящих по меньшей мере из одного L-нуклеотида и по меньшей мере одной D-нуклеиновой кислоты, указанная нуклеиновая кислота может состоять из дезоксирибонуклеотида(ов), рибонуклеотида(ов) или их комбинаций.
Конструирование заявленных нуклеиновых кислот в виде L-нуклеиновой кислоты является предпочтительными по ряду причин. L-нуклеиновые кислоты представляют собой энантиомеры природных нуклеиновых кислот. Тогда как D-нуклеиновые кислоты являются не очень стабильными в водных растворах, особенно в биологических системах или в биологических образцах, что обусловлено присутствием в них нуклеаз широкого ряда. Природные нуклеазы, в частности нуклеазы клеток животных, не способны разлагать L- нуклеиновые кислоты. Поэтому в такой системе, включая организм животного и человека, биологическое время полужизни L- нуклеиновой кислоты является более продолжительным. Поскольку L-нуклеиновая кислота не разлагается, то образования продуктов разложения нуклеазы не происходит, а следовательно, и не наблюдается каких-либо побочных эффектов. Этот аспект определяет отличие L-нуклеиновой кислоты, фактически, от всех других соединений, которые используются для лечения заболеваний и/или расстройств, ассоциированных с присутствием MCP-1. L-нуклеиновые кислоты, которые специфически связываются с молекулой-мишенью по механизму, отличающемуся от механизма спаривания оснований Уотсона-Крика, либо аптамеры, которые частично или полностью состоят из L-нуклеотидов, в частности те части аптамера, которые участвуют в его связывании с молекулой-мишенью, также называются оптическими изомерами.
Кроме того, в соответствии с настоящим изобретением, заявленные нуклеиновые кислоты, также называемые здесь нуклеиновыми кислотами согласно изобретению, независимо от того, присутствуют ли они в виде D-нуклеиновых кислот, L-нуклеиновых кислот или D,L-нуклеиновых кислот, или в виде ДНК или РНК, могут существовать в форме одноцепочечных или двухцепочечных нуклеиновых кислот. Обычно заявленными нуклеиновыми кислотами являются одноцепочечные нуклеиновые кислоты, которые имеют определенные вторичные структуры, образованные первичной последовательностью, а поэтому могут также образовывать третичные структуры. Однако заявленные нуклеиновые кислоты могут быть также двухцепочечными, а это означает, что две цепи, которые являются комплементарными или частично комплементарными другу другу, гибридизуются друг с другом. Это придает стабильность нуклеиновой кислоте, что может, в частности, оказаться предпочтительным в случае, если нуклеиновая кислота присутствует в природной D-форме, а не в L-форме.
Заявленные нуклеиновые кислоты могут быть модифицированы. Такие модификации могут быть внесены в одно положение нуклеотида нуклеиновой кислоты и хорошо известны специалистам. Примеры таких модификаций описаны, среди прочих, в работах Venkatesan (2003), Kusser (2000), Aurup (1994), Cummins (1995), Eaton (1995), Green (1995), Kawasaki (1993), Lesnik (1993) и Miller (1993). Такими модификациями могут быть атом H, атом F, группа O-CH3 или группа NH2 во 2'-положении отдельных нуклеотидов, из которых состоит нуклеиновая кислота. Кроме того, нуклеиновая кислота согласно изобретению может включать по меньшей мере один нуклеотид LNA. В одном из вариантов изобретения указанная нуклеиновая кислота согласно изобретению состоит из нуклеотидов LNA.
В одном из вариантов изобретения нуклеиновыми кислотами согласно изобретению могут быть многокомпонентные нуклеиновые кислоты. Используемая здесь многокомпонентная нуклеиновая кислота состоит по меньшей мере из двух нуклеотидных цепей. Эти по меньшей мере две цепи нуклеиновой кислоты образуют функциональное звено, где указанное функциональное звено представляет собой лиганд, присоединенный к молекуле-мишени. Указанные по меньшей мере две цепи нуклеиновой кислоты могут происходить от любой из заявленных нуклеиновых кислот в результате расщепления нуклеиновой кислоты с образованием двух цепей, либо в результате синтеза одной нуклеиновой кислоты, соответствующей первой части заявленной нуклеиновой кислоты, то есть полноразмерной нуклеиновой кислоты, и другой нуклеиновой кислоты, соответствующей второй части полноразмерной нуклеиновой кислоты. Следует отметить, что для получения многокомпонентной нуклеиновой кислоты, которая имеет более двух цепей, описанных выше, может быть применен как метод расщепления, так и метод синтеза. Другими словами, по меньшей мере две цепи нуклеиновой кислоты обычно отличаются от двух цепей, которые являются комплементарными и гибридизуются друг с другом, хотя между различными нуклеиновыми кислотами может иметь место определенная степень комплементарности.
И, наконец, в соответствии с настоящим изобретением может быть получена полностью замкнутая, то есть кольцевая структура нуклеиновых кислот согласно изобретению, а именно нуклеиновые кислоты согласно изобретению могут быть получены в виде замкнутых колец предпочтительно посредством ковалентной связи, причем более предпочтительно, чтобы такая ковалентная связь присутствовала между 5'- и 3'-концом описанных здесь последовательностей нуклеиновой кислоты.
Авторами настоящего изобретения было установлено, что нуклеиновые кислоты согласно изобретению имеют наиболее подходящий интервал величин КD.
Определение константы связывания может быть осуществлено с использованием так называемого устройства Biacore, которое также известно специалистам в данной области. Рассматриваемая здесь аффинность может быть также измерена с помощью “анализа на ингибирование”, описанного в примерах. Подходящей мерой для выражения интенсивности связывания нуклеиновой кислоты согласно изобретению с мишенью, а в данном случае с MCP-1, является так называемая величина KD, которая, как и метод ее определения, известна специалистам.
Нуклеиновые кислоты согласно изобретению характеризуются определенной величиной KD. Предпочтительная величина KD для нуклеиновых кислот согласно изобретению составляет ниже 1 мкM. Считается, что величина KD, равная примерно 1 мкM, является характерной для неспецифического связывания нуклеиновой кислоты с мишенью. Специалистам в данной области очевидно, что величины KD для группы соединений, таких как нуклеиновые кислоты согласно изобретению, варьируются в пределах определенного интервала значений. Вышеупомянутая величина KD, равная примерно 1 мкМ, является предпочтительно верхним пределом величин KD. Предпочтительный нижний предел величин KD для связывания нуклеиновых кислот с мишенью может составлять приблизительно 10 пикомоль или более. В соответствии с настоящим изобретением величины KD для отдельных нуклеиновых кислот, связывающихся с MCP-1, предпочтительно входят в этот интервал. Предпочтительные интервалы величин могут быть определены путем выбора любого первого числа, входящего в этот интервал, и любого второго числа, входящего в этот интервал. Предпочтительными верхними пределами этих значений являются 250 нM и 100 нM, а предпочтительными нижними пределами этих значений являются 50 нM, 10 нM, 1 нM, 100 пМ и 10 пМ.
Молекулы нуклеиновой кислоты согласно изобретению могут иметь любую длину при условии, что они будут обладать способностью связываться с молекулой-мишенью. Следует отметить, что нуклеиновые кислоты согласно изобретению имеют предпочтительные длины. Обычно такая длина составляет от 15 до 120 нуклеотидов. При этом следует отметить, что нуклеиновые кислоты согласно изобретению могут иметь длину, составляющую примерно от 15 до 120 нуклеотидов. Более предпочтительно длина нуклеиновых кислот согласно изобретению составляет примерно от 20 до 100 нуклеотидов, примерно от 20 до 80 нуклеотидов, примерно от 20 до 60 нуклеотидов, примерно от 20 до 50 нуклеотидов и примерно от 30 до 50 нуклеотидов.
В соответствии с настоящим изобретением описанные здесь нуклеиновые кислоты содержат группу, которая предпочтительно представляет собой высокомолекулярную группу и/или которая предпочтительно изменяет свойства нуклеиновой кислоты, такие как, среди прочих, время ее присутствия в организме животного, предпочтительно в организме человека. Особенно предпочтительным вариантом такой модификации является ПЭГилирование и ГЭКилирование нуклеиновых кислот согласно изобретению. Используемый здесь термин “ПЭГ” означает полиэтиленгликоль, а термин “ГЭК” означает гидроксиэтилированный крахмал. Используемый здесь термин “ПЭГилирование” предпочтительно означает модификацию нуклеиновой кислоты согласно изобретению, где указанная модификация состоит из молекулы ПЭГ, присоединенной к нуклеиновой кислоте согласно изобретению. Используемый здесь термин “ГЭКилирование” предпочтительно означает модификацию нуклеиновой кислоты согласно изобретению, где указанная модификация состоит из молекулы ГЭК, присоединенной к нуклеиновой кислоте согласно изобретению. Эти модификации, а также метод модифицирования нуклеиновой кислоты с использованием таких модификаций описаны в заявке на Европейский патент EP 1306382, содержание которой во всей своей полноте вводится в настоящее описание посредством ссылки.
Молекулярная масса модифицирующей молекулы, состоящей из высокомолекулярной группы или содержащей такую группу, предпочтительно составляет примерно от 2000 дo 200000 Да, более предпочтительно от 20000 дo 120000 Дa, в частности, если такой высокомолекулярной группой является ПЭГ, и предпочтительно примерно от 3000 дo 180000 Да, а еще более предпочтительно от 5000 дo 130000 Да, в частности, если такой высокомолекулярной группой является ГЭК. Способ модификации молекулой ГЭК описан, например, в заявке на патент Германии DE 12004006249.8, содержание которой во всей своей полноте вводится в настоящее описание посредством ссылки.
В соответствии с настоящим изобретением ПЭГ и ГЭК могут быть использованы в виде прямой или разветвленной цепи, как описано в патентных заявках WO2005074993 и PCT/EP02/11950. Такая модификация может быть в принципе введена в молекулы нуклеиновой кислоты согласно изобретению в любом их положении. Такую модификацию предпочтительно вводят у 5'-концевого нуклеотида, у 3'-концевого нуклеотида и/или в любом положении нуклеотидов, расположенных между 5'-нуклеотидом и 3'-нуклеотидом молекулы нуклеиновой кислоты.
Модифицирующая молекула, предпочтительно молекула ПЭГ и/или ГЭК, может быть присоединена к молекуле нуклеиновой кислоты согласно изобретению либо непосредственно, либо посредством линкера. Кроме того, в соответствии с настоящим изобретением молекула нуклеиновой кислоты согласно изобретению содержит одну или несколько модифицирующих молекул, предпочтительно одну или несколько молекул ПЭГ и/или ГЭК. В одном из вариантов изобретения к молекуле нуклеиновой кислоты согласно изобретению может быть присоединена, посредством отдельной линкерной молекулы, более чем одна молекула ПЭГ или ГЭК. Линкер, используемый в соответствии с настоящим изобретением, сам может быть прямым или разветвленным. Линкеры такого типа известны специалистам, а также описаны в патентных заявках WO2005074993 и PCT/EP02/11950.
Не ограничиваясь какой-либо конкретной теорией, следует лишь отметить, что в результате модификации нуклеиновых кислот согласно изобретению, содержащих высокомолекулярную группу, такую как полимер, а более конкретно, описанные здесь полимеры, которые предпочтительно являются физиологически приемлемыми, может быть изменена кинетика экскреции. Более конкретно, следует отметить, что из-за увеличения молекулярной массы таких модифицированных заявленных нуклеиновых кислот и из-за отсутствия метаболизма нуклеиновых кислот, в частности, если они присутствуют в L-форме, их экскреция из организма животного, предпочтительно из организма млекопитающего, а еще более предпочтительно из организма человека уменьшается. Поскольку экскреция обычно происходит через почки, то авторы настоящего изобретения выдвинули предположение, что скорость клубочковой фильтрации модифицированной таким образом нуклеиновой кислоты значительно снижается по сравнению со скоростью фильтрации нуклеиновых кислот, в которых отсутствует высокомолекулярная модифицирующая группа указанного типа, что приводит к увеличению времени присутствия модифицированных нуклеиновых кислот в организме. В соответствии с этим, особенно примечательным является то, что, несмотря на такую высокомолекулярную модификацию, специфичность нуклеиновой кислоты согласно изобретению не нарушается. Таким образом, нуклеиновые кислоты согласно изобретению имеют неожиданные свойства, которыми обычно не обладают фармацевтически активные соединения, а именно такие свойства, при которых для замедленного высвобождения таких нуклеиновых кислот не требуется изготовления фармацевтического препарата с пролонгированным высвобождением. Вместо этого нуклеиновые кислоты согласно изобретению в своей модифицированной форме, включающей высокомолекулярную часть, сами могут быть использованы в качестве препарата пролонгированного высвобождения. Таким образом, модификация(и) молекул нуклеиновой кислоты, описанных в настоящей заявке, и модифицированные таким образом молекулы нуклеиновой кислоты и любая их композиция могут обеспечивать конкретные предпочтительно регулируемые фармакокинетические свойства и биологическое распределение этих молекул. Такими свойствами также являются время присутствия в кровотоке и распределение в тканях. Указанные модификации также описаны в патентной заявке PCT/EP02/11950.
Однако в соответствии с настоящим изобретением описанные здесь нуклеиновые кислоты не содержат каких-либо модификаций, в частности, каких-либо высокомолекулярных модифицирующих групп, таких как ПЭГ или ГЭК. Такой вариант является особенно предпочтительным в том случае, если нуклеиновая кислота обнаруживает предпочтительное распределение в любом органе или ткани-мишени организма. Средства, содержащие такую нуклеиновую кислоту и имеющие указанный профиль распределения, позволяют достигать эффективных локальных концентраций в ткани-мишени при сохранении низкой системной концентрации. Такое средство может быть использовано в низких дозах, что является не только экономически выгодным, но также позволяет снижать уровень нежелательного воздействия содержащейся в нем нуклеиновой кислоты на другие ткани и тем самым снижать потенциальный риск возникновения побочных эффектов.
Заявленные нуклеиновые кислоты, которые также называются здесь нуклеиновыми кислотами согласно изобретению и/или антагонистами согласно изобретению, могут быть использованы для получения или приготовления лекарственного средства. Такой лекарственный препарат или фармацевтическая композиция согласно изобретению содержат по меньшей мере одну из заявленных нуклеиновых кислот, необязательно в комбинации с другими фармацевтически активными соединениями, где сама заявленная нуклеиновая кислота предпочтительно действует как фармацевтически активное соединение. В предпочтительных вариантах изобретения такие лекарственные средства содержат по меньшей мере фармацевтически приемлемый носитель. Такими носителями могут быть, например, вода, буфер, PBS, раствор глюкозы, предпочтительно 5% сбалансированный солевой раствор глюкозы, крахмал, сахар, желатин или любое другое фармацевтически приемлемое вещество-носитель. Такие носители по существу известны специалистам в данной области. Следует отметить, что любые варианты, применения и аспекты, относящиеся к лекарственному средству согласно изобретению, могут также относиться и к фармацевтической композиции согласно изобретению и наоборот.
Патологии, заболевания и расстройства, для лечения и/или предупреждения которых используются нуклеиновые кислоты, фармацевтические композиции и лекарственные средства, заявленные в настоящем изобретении или приготовленные в соответствии с настоящим изобретением, ассоциируются с прямым или непрямым участием MCP-1 в соответствующем механизме патогенеза. Однако лечению и предупреждению могут быть также подвергнуты такие патологии, заболевания и расстройства, в патогенезе которых прямо или опосредованно участвуют MCP-2, MCP-3, MCP-4 и/или эотаксин. Для специалиста в данной области очевидно, что такие нуклеиновые кислоты согласно изобретению, в частности, могут быть использованы, например, для лечения заболеваний, в патогенезе которых участвуют, в широком смысле этого слова, MCP-2, MCP-3, MCP-4 и эотаксин, где указанные нуклеиновые кислоты взаимодействуют и связываются с указанными MCP-2, MCP-3, MCP-4 и эотаксином соответственно.
Более конкретно, такое применение обусловлено, среди прочих, профилем экспрессии MCP-1, который, как предполагается, играет важную роль в развитии заболеваний человека, ассоциированных с инфильтрацией мононуклеарных клеток. Такая инфильтрация клеток характерна для многих воспалительных и аутоиммунных заболеваний.
Было показано, что у животных-моделей MCP-1 экспрессируется в головном мозге после фокальной ишемии (Kim 1995; Wang 1995) и во время экспериментального аутоиммунного энцефаломиелита (Hulkower 1993; Ransohoff 1993; Banisor 2005). MCP-1 является хемокином, который может играть важную роль в нацеливании на мононуклеарные клетки в патологическом процессе, проиллюстрированном на этих животных-моделях, таком как инсульт и рассеянный склероз.
Об уникальной роли системы MCP-1/CCR2 в хемоаттракции моноцитов, а следовательно, и в развитии хронического воспаления, свидетельствует большое количество данных, указывающих на то, что: (i) MCP-1- или CCR2-дефицитные мыши обнаруживают заметное снижение уровня макрофагального хемотаксического ответа, при этом другие функции остаются нормальными (Kuziel 1997, Kurihara 1997, Boring 1997, Lu 1998); (ii) несмотря на функциональную избыточность других хемокинов in vitro, потеря всего лишь одной эффекторной функции MCP-1 является достаточной для нарушения транспорта моноцитов у нескольких животных с моделью воспаления (Lloyd 1997, Furuichi 2003, Egashira 2002, Galasso 2000, Ogata 1997, Kennedy 1998, Gonzalo 1998, Kitamoto 2003); (iii) при многих воспалительных заболеваниях уровни MCP-1 увеличиваются. Действительно, считается, что MCP-1 играет определенную роль в развитии многих заболеваний, имеющих или не имеющих воспалительный компонент, таких как ревматоидный артрит (Koch 1992, Hosaka 1994, Akahoshi 1993, Harigai 1993, Rollins 1996), заболевание почек (Wada 1996, Viedt 2002), рестеноз после ангиопластики (Economou 2001), аллергия и астма (Alam 1996; Holgate 1997, Gonzalo 1998), рак (Salcedo 2000, Gordillo 2004), атеросклероз (Nelken 1991, Yla-Herttuala 1991, Schwartz 1993; Takeya 1993, Boring 1998), псориаз (Vestergaard 2004), воспаление нервной системы (Huang 2001), атопический дерматит (Kaburagi 2001), колит (Okuno 2002), эндометриоз (Jolicoeur 2001), увеит (Tuaillon 2002), заболевания сетчатки (Nakazawa 2007), идиопатический фиброз легких и саркоидоз (Iyonaga 1994) и полимиозит/дерматомиозит (De Bleecker 2002).
Терапевтическое лечение средствами, направленными против MCP-1, или антагонистами CCR2 должно воздействовать на избыточный транспорт воспалительных моноцитов, но при этом может сохраняться базальный уровень транспорта фагоцитов, что позволяет избежать развития общей иммуносупрессии и увеличения риска возникновения инфекций (Dawson 2003).
Кроме того, исходя из все возрастающего количества данных о молекулярных механизмах воспалительного процесса и взаимосвязи локально секретируемых медиаторов воспаления, могут быть идентифицированы новые мишени для терапии почечных заболеваний (Holdsworth 2000, Segerer 2000). Одной из таких мишеней, для которой имеются надежные данные об ее экспрессии, а также имеются результаты интервенционного исследования с использованием специфических антагонистов на соответствующих животных моделях, является MCP-1. Этот белок играет далеко не последнюю роль в рекрутинге иммунных клеток в области пораженных воспалением почек. Инфильтрация иммунных клеток является, по всей вероятности, главным механизмом структурного поражения почек и снижения функции почек при развитии различных форм почечного заболевания.
Все типы почечных клеток могут экспрессировать хемокины, включая MCP-1, после стимуляции in vitro (Segerer 2000); при этом существует большое число стимуляторов, запускающих экспрессию MCP-1 in vitro, включая цитокины, кислородные радикалы, иммунные комплексы и липидные медиаторы.
В здоровых почках крыс и мышей MCP-1 не экспрессируется, но его экспрессия легко индуцируется у грызунов с моделью острого и хронического воспаления почек, включая гломерулонефрит, опосредуемый иммунными комплексами, быстро прогрессирующий гломерулонефрит, пролиферирующий гломерулонефрит, диабетическую нефропатию, обструктивную нефропатию или острый тубулярный некроз (Segerer 2000, Anders 2003). Данные об экспрессии MCP-1 у грызунов четко коррелируют с данными о соответствующей экспрессии, обнаруженной в биоптате почек человека (Rovin 1994, Cockwell 1998, Wada 1999). Кроме того, экспрессия MCP-1 в почках человека ассоциируется с активностью заболевания, и снижение уровня такой экспрессии при соответствующей терапии приводит к ремиссии заболевания (Amann 2003).
Инфильтрация гломерулярных мононуклеарных клеток ассоциируется с развитием диффузного гломерулосклероза у пациентов с диабетической нефропатией. MCP-1 играет важную роль в рекрутинге и аккумуляции моноцитов и лимфоцитов в почечных клубочках (Banba 2000, Morii 2003).
Очевидно, что локально продуцируемый MCP-1, в частности, участвует в инициации и прогрессировании поражения тубулярной и интерстициальной структуры почек, как было зарегистрировано в экспериментах с использованием трансгенных мышей с нефритом, индуцированным нефротоксичной сывороткой (NSN). MCP-1 обнаруживался, главным образом, в эндотелиальных клетках сосудов, в тубулярных эпителиальных клетках и в инфильтрированных мононуклеарных клетках при интерстициальных поражениях. MCP-1-опосредуемая активация тубулярных эпителиальных клеток подтверждает мнение, что MCP-1 играет определенную роль в воспалении тубулярной и интерстициальной системы, то есть отличительного признака прогрессирующего заболевания почек (Wada 2001, Viedt 2002).
Благодаря гомологии с MCP-1, с одной стороны, и с MCP-2, MCP-3, MCP-4 и эотаксином, с другой стороны, нуклеиновые кислоты согласно изобретению или по меньшей мере те из них, которые взаимодействуют или связываются с MCP-2, MCP-3, MCP-4 и эотаксином соответственно, могут быть использованы как таковые для лечения, предупреждения и/или диагностики любого заболевания, в патогенезе которого прямо или опосредованно участвуют MCP-2, MCP-3, MCP-4 и эотаксин соответственно. Используемый здесь термин “участвует” предпочтительно означает, что если соответствующая молекула, которая участвует в патогенезе данного заболевания, не обладает одной, несколькими или всеми функциями, ассоциированными с механизмом патогенеза, лежащим в основе развития данного заболевания, то либо данное заболевание может быть вылечено или до определенной степени ослаблено, либо может быть предотвращена вспышка данного заболевания, при этом по меньшей мере один из симптомов или любые признаки данного заболевания могут быть ослаблены или устранены соответственно, в результате чего состояние данного индивидуума соответственно становится идентичным или почти идентичным состоянию, наблюдаемому у индивидуума, не страдающего указанным заболеванием или не подверженного риску развития такого заболевания.
Действительно, поскольку MCP-1-связывающие нуклеиновые кислоты согласно изобретению взаимодействуют или связываются с человеческим или мышиным MCP-1, то для среднего специалиста в данной области очевидно, что MCP-1-связывающие нуклеиновые кислоты согласно изобретению могут быть с успехом использованы для лечения, предупреждения и/или диагностики любого из описанных здесь заболеваний человека или животных.
Указанные члены семейства белков-хемоаттрактантов моноцитов (MCP), то есть MCP-2, MCP-3, MCP-4 и эотаксин, имеют высокую степень сходства с MCP-1. При этом эотаксин, MCP-2, -3 и -4 взаимодействуют, но не исключительно, посредством рецептора CCR3, то есть характерного хемокинового рецептора, присутствующего на человеческих эозинофилах (Heath 1997). Рецептор CCR3 активируется в опухолях, таких как Т-клеточная лимфома кожи (Kleinhans 2003), глиобластома (Kouno 2004), или почечно-клеточная карцинома (Johrer 2005).
Более конкретно, повышенные уровни эотаксина непосредственно ассоциируются с диагнозом астмы и с нарушением легочной функции (Nakamura 1999). Повышенные уровни экспрессии эотаксина в области аллергического воспаления наблюдались у пациентов с атопической и неатопической астмой (Ying 1997, Ying 1999). Кроме того, мРНК, кодирующие MCP-2 и -4, конститутивно экспрессируются в различных тканях; однако их физиологические функции в этих тканях остаются неизвестными. При сепсисе уровни MCP-2, а также уровни MCP-1 в плазме повышаются (Bossink 1995), а экспрессия MCP-3 наблюдается у астматиков (Humbert 1997). И, наконец, MCP-4 может обнаруживаться на поверхности просвета сосудов, пораженных атеросклерозом (Berkhout 1997).
В соответствии с этим, заболеваниями, и/или расстройствами, и/или патологическими состояниями, которые могут быть подвергнуты лечению и/или предупреждению с применением лекарственных средств согласно изобретению, являются, но не ограничиваются ими, воспалительные заболевания, аутоиммунные заболевания, аутоиммунный энцефаломиелит, инсульт, острый и хронический рассеянный склероз, хроническое воспалительное заболевание, ревматоидный артрит, болезни почек, рестеноз, рестеноз после ангиопластики, острые и хронические аллергические реакции, первичные и вторичные иммунологические или аллергические реакции, астма, конъюнктивит, бронхит, рак, атеросклероз, атеросклеротическая сердечно-сосудистая недостаточность или инсульт, псориаз, псориатический артрит, воспаление нервной системы, атопический дерматит, колит, эндометриоз, увеит, заболевание сетчатки, включая дегенерацию желтого пятна, отслоение сетчатки, диабетическую ретинопатию, ретролентальную фиброплазию, пигментный ретинит, пролиферирующий витреоретинит и центральную серозную хориоретинопатию; идеопатический фиброз легких, саркоидоз, полимиозит, дерматомиозит; иммуносупрессия и риск инфицирования; сепсис, воспаление почек, гломерулонефрит, быстро прогрессирующий гломерулонефрит, пролиферирующий гломерулонефрит, диабетическая нефропатия, обструктивная нефропатия, острый тубулярный некроз и диффузный гломерулосклероз, системная красная волчанка, хронический бронхит, болезнь Бехчета, амиотрофический боковой склероз (АБС), ранний атеросклероз, ассоциированный с болезнью Кавазаки, инфаркт миокарда, ожирение, хроническая болезнь печени, болезнь Пейрони, острое поражение хорды спинного мозга, заболевание, ассоциированное с трансплантацией легких или почек, миокардит, болезнь Альцгеймера и невропатия, карцинома молочной железы, карцинома желудка, рак мочевого пузыря, рак яичника, гамартома, карцинома прямой и ободочной кишки, аденома толстой кишки, панкреатит, хроническая обструктивная болезнь легких (ХОБЛ) и воспалительные заболевания кишечника, такие как болезнь Крона или язвенный колит.
В другом варианте изобретения указанное лекарственное средство включает другое фармацевтически активное средство. Такими другими фармацевтически активными соединениями являются, среди прочих, но не ограничиваются ими, соединения, которые, как известно, регулируют кровяное давление и могут быть использованы для лечения диабета, такие как ингибиторы ангиотензин-конвертирующего фермента (ACE) и блокаторы рецептора ангиотензина. В другом варианте изобретения дополнительным фармацевтически активным соединением может быть также одно из соединений, которые снижают уровень инфильтрации иммунных клеток на участке хронического воспаления или по существу подавляют резко выраженный иммунный ответ, который вырабатывается при хронических воспалительных состояниях и который приводит к поражению ткани. Такими соединениями могут быть, но не ограничиваются ими, стероиды или иммунодепрессанты, и эти соединения предпочтительно выбирают из группы, включающей кортикостероиды, такие как преднизон, метилпреднизолон, гидрокортизон, дексаметазон, и иммунодепрессанты общего действия, такие как циклофосфамид, циклоспорин, хлорамбуцил, азатиоприн, такролимус или микофенолят-мофетил. Кроме того, в других вариантах изобретения могут быть использованы более специфические блокаторы костимуляции Т-клеток, например блокаторы CD154, CD40, CD28, CD86 или CD80, или агенты, истощающие T- и/или B-клетки, например анти-CD20 антитело. И наконец, другим фармацевтически активным средством может быть модулятор активности любого другого хемокина, который может быть агонистом или антагонистом хемокина или агонистом или антагонистом хемокинового рецептора. Альтернативно или дополнительно таким другим фармацевтически активным средством является другая нуклеиновая кислота согласно изобретению. Альтернативно указанный лекарственный препарат содержит по меньшей мере одну дополнительную нуклеиновую кислоту, которая связывается с молекулой-мишенью, отличающейся от MCP-1, или обладает функцией, отличающейся от функции одной из нуклеиновых кислот согласно изобретению.
В соответствии с настоящим изобретением указанное лекарственное средство альтернативно или дополнительно может быть использовано для предупреждения, в принципе, любого из заболеваний, описанных в связи с применением указанного лекарственного средства для лечения таких заболеваний. Следовательно, соответствующие маркеры, то есть маркеры соответствующих заболеваний, известны специалистам. Соответствующим маркером предпочтительно является MCP-1. Альтернативно и/или дополнительно соответствующий маркер выбран из группы, включающей MCP-2, MCP-3, MCP-4 и эотаксин. Другую группу маркеров выбирают из группы, включающей аутореактивные антитела, присутствующие в плазме, такие как, например, антитела против дцДНК или ревматоидный фактор.
В одном из вариантов лекарственных средств согласно изобретению такое лекарственное средство может быть использовано в комбинации с другими курсами терапии любых из описанных здесь заболеваний, в частности заболеваний, которые могут быть подвергнуты лечению лекарственным препаратом согласно изобретению.
"Комбинированная терапия" (или "совместная терапия") включает введение лекарственного средства согласно изобретению и по меньшей мере второго средства как части конкретного курса лечения, которое, как предполагается, дает благоприятный эффект при совместном действии этих терапевтических средств, то есть лекарственного средства согласно изобретению и указанного второго средства. Благоприятный эффект такой комбинированной терапии включает, но не ограничивается им, совместное фармакокинетическое или фармакодинамическое действие, ассоциированное с введением комбинации терапевтических средств. Введение указанной комбинации терапевтических средств обычно осуществляют в течение определенного периода времени (обычно в течение нескольких минут, часов, дней или недель, в зависимости от выбранного курса комбинированной терапии).
"Комбинированная терапия" может включать, но не только, введение двух или более из указанных терапевтических средств как части отдельных курсов монотерапии, которые иногда и случайно приводят к продуцированию комбинированных эффектов согласно изобретению. "Комбинированная терапия" включает последовательное введение этих терапевтических средств, при котором каждое терапевтическое средство вводят в различные периоды времени, а также по существу одновременное введение этих терапевтических средств или по меньшей мере двух из указанных терапевтических средств. Такое по существу одновременное введение может быть осуществлено, например, путем введения индивидууму одной капсулы, содержащей фиксированное отношение каждого терапевтического средства, или множества отдельных капсул, содержащих каждое из указанных терапевтических средств.
Последовательное или по существу одновременное введение каждого терапевтического средства может быть осуществлено любым подходящим способом, включая, но не ограничиваясь ими, местное введение, пероральное введение, внутривенное введение, внутримышечное введение и прямую абсорбцию через ткани слизистой оболочки. Указанные терапевтические средства могут быть введены одним и тем же способом или различными способами. Так, например, первое терапевтическое средство данной комбинации может быть введено путем инъекции, а другие терапевтические средства данной комбинации могут быть введены местно.
Альтернативно, например, все терапевтические средства могут быть введены местно, либо они могут быть введены путем инъекции. Последовательность введения этих терапевтических средств не имеет решающего значения, если это не оговорено особо. "Комбинированная терапия" может также включать введение описанных выше терапевтических средств в дополнительной комбинации с другими биологически активными ингредиентами. Если комбинированная терапия дополнительно включает немедикаментозное лечение, то такое лечение может быть осуществлено в любое подходящее время при условии, что при этом будет достигаться благоприятный эффект от совместного действия комбинированной терапии терапевтическими средствами и немедикаментозной терапии. Так, например, в конкретных случаях благоприятный эффект может быть достигнут даже в том случае, если немедикаментозное лечение проводят через большой интервал времени после введения терапевтических средств, возможно, через несколько дней или даже недель.
Как указывалось выше в описании общих терминов, лекарственное средство согласно изобретению может быть введено, в принципе, в любой форме, известной специалистам. Предпочтительным способом введения является системное введение, а более предпочтительным является парентеральное введение предпочтительно путем инъекции. Альтернативно данное лекарственное средство может быть введено местно. Другие способы введения включают внутримышечное введение, внутрибрюшинное введение и подкожное введение, а также пероральное введение, интраназальное введение, интратрахеальное введение или внутрилегочное введение, при этом предпочтительным способом введения является наименее инвазивное и наиболее эффективное введение.
Парентеральное введение обычно включает подкожное, внутримышечное или внутривенное введение и вливание. Кроме того, один из способов парентерального введения предусматривает имплантацию систем медленного или пролонгированного высвобождения, обеспечивающих поддержание постоянного уровня лекарственного средства и хорошо известных среднему специалисту в данной области.
Кроме того, предпочтительные лекарственные средства согласно изобретению могут быть введены в виде лекарственных форм для интраназального введения с помощью подходящих носителей, используемых для интраназального местного введения, ингаляторов или лекарственных форм для чрескожного введения с использованием чрескожных пластырей, хорошо известных среднему специалисту в данной области. Само собой разумеется, что введение лекарственного средства с помощью системы для чрескожной доставки, осуществляемое в процессе данного курса лечения, обычно проводят непрерывно, а не периодически. Другими предпочтительными препаратами для местного введения являются кремы, мази, лосьоны, аэрозольные спреи и гели, в которых концентрация активного ингредиента обычно составляет в пределах от 0,01% до 15% масс./масс. или масс./об.
Лекарственное средство согласно изобретению обычно содержит эффективное количество активного(ых) компонента(ов), используемого(ых) в данной терапии, включая, но не ограничиваясь ими, молекулу нуклеиновой кислоты согласно изобретению, растворенную или диспергированную в фармацевтически приемлемой среде. Фармацевтически приемлемыми средами или носителями являются любые и все растворители, дисперсионные среды, материалы для покрытий, антибактериальные и противогрибковые средства, агенты, придающие изотоничность, и агенты, замедляющие абсорбцию, и т.п. Применение таких сред и агентов для получения фармацевтически активных веществ хорошо известно специалистам. В лекарственное средство согласно изобретению могут быть также включены дополнительные активные ингредиенты.
В другом своем аспекте настоящее изобретение относится к фармацевтической композиции. Такая фармацевтическая композиция содержит по меньшей мере одну из нуклеиновых кислот согласно изобретению и предпочтительно фармацевтически приемлемый носитель. Таким носителем может быть любой носитель или любое связующее вещество, которые обычно используются специалистами и/или являются известными. Более конкретно, таким связующим веществом или носителем является любое связующее вещество или носитель, обсуждаемые в связи с получением описанного здесь лекарственного препарата. В другом варианте изобретения указанная фармацевтическая композиция содержит дополнительное фармацевтически активное средство.
Получение лекарственного препарата и фармацевтической композиции будет очевидно для специалиста исходя из описания настоящего изобретения. Обычно такие композиции могут быть получены в виде инъекций жидких растворов или суспензий, а также в виде твердых форм, подходящих для их растворения или суспендирования в жидкости перед инъекцией, в виде таблеток или других твердых форм для перорального введения, в виде капсул с пролонгированным высвобождением или в виде других часто используемых форм, включая глазные капли, кремы, лосьоны, мази, ингаляторы и т.п. Особенно эффективным может быть также применение стерильных препаратов, таких как промывки на основе физиологического раствора, которые используются хирургами, практическими врачами или медицинскими работниками для обработки конкретных участков операционного поля. Композиции могут быть также доставлены с помощью микроскопических устройств, микрочастиц или тампонов.
После получения лекарственное средство может быть введено способом, совместимым с данной лекарственной композицией, и в фармакологически эффективном количестве. Такие композиции могут быть легко введены в виде различных лекарственных форм, таких как растворы для инъекций, описанные выше, но могут быть также использованы и капсулы для высвобождения лекарственных средств и т.п.
В этой связи следует отметить, что количество активного ингредиента и объем вводимой композиции зависят от конкретного индивидуума или пациента, подвергаемого лечению. Выбор конкретных количеств активного соединения, необходимого для введения, зависит от заключения врача и является индивидуальным для каждого конкретного индивидуума.
Обычно используется минимальный объем лекарственного средства, необходимый для диспергирования активных соединений. Подходящие схемы введения также могут варьироваться, но обычно их подбирают путем предварительного введения данного соединения и мониторинга наблюдаемых результатов, а затем вводят дополнительные регулируемые дозы через дополнительные интервалы времени.
Так, например, для перорального введения в форме таблетки или капсулы (например, желатиновой капсулы), активный компонент лекарственного средства, то есть молекулы нуклеиновой кислоты согласно изобретению и/или другое фармацевтически активное средство, также называемое здесь терапевтическим(ими) средством(ами) или активным(ми) соединением(ями), могут быть объединены с пероральным нетоксическим фармацевтически приемлемым инертным носителем, таким как этанол, глицерин, вода и т.п. Кроме того, при желании или необходимости, в данную смесь могут быть включены подходящие связующие вещества, замасливатели, дезинтегрирующие вещества и красители. Подходящими связующими веществами являются крахмал, алюмосиликат магния, крахмальная паста, желатин, метилцеллюлоза, натрий-содержащая карбоксиметилцеллюлоза и/или поливинилпирролидон, природные сахара, такие как глюкоза или бета-лактоза, кукурузные подсластители, природные и синтетические смолы, такие как аравийская камедь, трагакантовая камедь или альгинат натрия, полиэтиленгликоль, воски и т.п. Замасливателами, используемыми в этих лекарственных формах, являются олеат натрия, стеарат натрия, стеарат магния, бензоат натрия, ацетат натрия, хлорид натрия, двуокись кремния, тальк, стеариновая кислота, ее магниевая или кальциевая соль и/или полиэтиленгликоль и т.п. Дезинтеграторами являются, но не ограничиваются ими, крахмал, метилцеллюлоза, агар, бентонит, крахмалы, содержащие ксантановую камедь, агар, альгиновая кислота или их натриевые соли, или вспенивающие смеси и т.п. Разбавителями являются, например, лактоза, декстроза, сахароза, маннит, сорбит, целлюлоза и/или глицин.
Лекарственное средство согласно изобретению может быть также введено в виде пероральных лекарственных форм, таких как таблетки или капсулы с длительным и пролонгированным высвобождением, драже, порошки, гранулы, эликсиры, настойки, суспензии, сиропы и эмульсии. Суппозитории преимущественно получают из жирных эмульсий или суспензий.
Фармацевтическая композиция или лекарственное средство могут быть стерилизованы и/или они могут содержать адъюванты, такие как консерванты, стабилизаторы, смачивающие или эмульгирующие агенты, стимуляторы растворения, соли для регуляции осмотического давления и/или буферы. Кроме того, они могут также содержать другие терапевтически ценные вещества. Композиции получают стандартными методами смешивания, гранулирования или нанесения покрытий, и такие композиции обычно содержат примерно от 0,1% до 75%, предпочтительно примерно от 1% до 50% активного ингредиента.
Жидкие, в частности инъецируемые, композиции могут быть получены, например, путем растворения, диспергирования и т.п. Активное соединение растворяют в фармацевтически чистом растворителе или смешивают с указанным растворителем, таким как, например, вода, физиологический раствор, водный раствор декстрозы, глицерин, этанол и т.п., в результате чего получают раствор или суспензию для инъекций. Кроме того, могут быть приготовлены твердые формы, подходящие для растворения в жидкости перед инъекцией.
Наполнителями для твердых композиций являются фармацевтически чистые маннит, лактоза, крахмал, стеарат магния, натрий-содержащий сахарин, тальк, целлюлоза, глюкоза, сахароза, карбонат магния и т.п. Активное соединение, определенное выше, может быть также приготовлено в виде суппозитория с использованием, например, полиалкиленгликоля, например пропиленгликоля в качестве носителя. В некоторых вариантах изобретения суппозитории преимущественно получают из жирных эмульсий или суспензий.
Лекарственные средства и молекулы нуклеиновой кислоты согласно изобретению соответственно могут быть также введены в форме липосомных систем для доставки, таких как небольшие однослойные везикулы, крупные однослойные везикулы и многослойные везикулы. Липосомы могут быть образованы из различных холестерин-содержащих фосфолипидов, стеариламина или фосфатидилхолинов. В некоторых вариантах изобретения пленку, состоящую из липидных компонентов, гидратируют водным раствором лекарственного средства с получением липидного слоя, в который инкапсулируется лекарственное средство, и такой процесс хорошо известен среднему специалисту в данной области. Так, например, описанные здесь молекулы нуклеиновой кислоты могут быть получены в виде комплекса с липофильным соединением или неиммуногенным высокомолекулярным соединением, сконструированным методами, известными специалистам. Кроме того, липосомы могут содержать такие молекулы нуклеиновой кислоты на своей поверхности для доставки и внедрения цитотоксических агентов вовнутрь клетки, опосредуя таким образом цитолиз клеток. Репрезентативные комплексы, ассоциированные с нуклеиновыми кислотами, приводятся в патенте США № 6011020.
Лекарственные средства и молекулы нуклеиновой кислоты согласно изобретению соответственно могут быть также объединены с растворимыми полимерами, используемыми в качестве носителей для доставки лекарственного средства. Такими полимерами могут быть поливинилпирролидон, сополимер пирана, полигидроксипропил-метакриламид-фенол, полигидроксиэтил-аспанамидфенол или полиэтиленоксидополилизин, замещенный пальмитоильными остатками. Кроме того, лекарственные средства и молекулы нуклеиновой кислоты согласно изобретению соответственно могут быть объединены с биологически разлагаемыми полимерами определенного класса, которые могут быть использованы для достижения регулируемого высвобождения лекарственного средства, и такими полимерами являются, например, полимолочная кислота, полиэпсилон-капролактон, полигидроксимасляная кислота, полиортоэфиры, полиацетали, полидигидропираны, полицианоакрилаты и перекрестно связанные или амфипатические блоксополимеры гидрогелей.
Если это необходимо, то вводимые фармацевтические композиции и лекарственные средства соответственно могут также содержать небольшие количества нетоксичных вспомогательных веществ, таких как смачивающие или эмульгирующие агенты, рН-забуферивающие агенты, и другие вещества, такие как, например, ацетат натрия и олеат триэтаноламина.
Схему введения доз молекул нуклеиновой кислоты и лекарственных средств согласно изобретению соответственно выбирают в зависимости от ряда факторов, включая тип, вид, возраст, массу, пол и состояние здоровья пациента; тяжесть состояния, подвергаемого лечению; способ введения; почечную и печеночную функцию пациента; и конкретно используемый аптамер или его соль. Любой врач или ветеринар может легко определить и прописать эффективное количество лекарственного средства, необходимого для предупреждения, подавления или прекращения прогрессирования данного патологического состояния.
Для лечения любых из описанных здесь заболеваний эффективные уровни нуклеиновой кислоты согласно изобретению в плазме предпочтительно должны составлять от 500 фM дo 500 мкM.
Молекулы нуклеиновой кислоты и лекарственные средства согласно изобретению соответственно могут быть введены в виде разовой дозы ежедневно, через два или три дня, раз в неделю, раз в две недели, один раз в месяц или один раз в три месяца.
В соответствии с настоящим изобретением описанное здесь лекарственное средство включает описанную здесь фармацевтическую композицию.
В другом своем аспекте настоящее изобретение относится к способу лечения индивидуума, нуждающегося в таком лечении, где указанный способ включает введение фармацевтически активного количества по меньшей мере одной из нуклеиновых кислот согласно изобретению. В одном из вариантов изобретения указанный индивидуум страдает заболеванием или подвержен риску развития у него такого заболевания, где указанным заболеванием является любое из описанных здесь заболеваний, в частности любое из заболеваний, рассматриваемых в связи с применением любой из нуклеиновых кислот согласно изобретению для приготовления лекарственного средства.
Следует отметить, что нуклеиновая кислота, а также антагонисты согласно изобретению могут быть использованы не только в качестве лекарственного средства или для его приготовления, но также и в косметических целях, в частности, для устранения воспалительных региональных поражений кожи, ассоциированных с MCP-1. Поэтому другим состоянием или заболеванием, которое может быть подвергнуто лечению или может быть предотвращено с использованием нуклеиновой кислоты, лекарственного средства и/или фармацевтической композиции согласно изобретению, являются воспалительные региональные поражения кожи.
Предпочтительно используемые здесь диагностическое вещество, диагностический реагент или диагностическое средство являются подходящими для прямого или опосредованного детектирования MCP-1, предпочтительно описанного здесь MCP-1, а более предпочтительно описанного здесь MCP-1, ассоциированного с различными рассматриваемыми здесь расстройствами и заболеваниями. Однако молекулы нуклеиновой кислоты согласно изобретению до определенной степени также связываются с любыми, некоторыми или со всеми MCP-2, MCP-3, MCP-4 и эотаксином, поэтому такие молекулы нуклеиновой кислоты могут быть также использованы для диагностики заболеваний и расстройств соответственно, механизм патогенеза которых прямо или опосредованно связан или ассоциирован со сверхэкспрессией или сверхактивностью MCP-2, MCP-3, MCP-4 и/или эотаксина. Такое диагностическое средство является подходящим для детектирования и/или мониторинга любых из описанных здесь расстройств и заболеваний соответственно. Указанный метод детекции может быть осуществлен посредством связывания нуклеиновых кислот согласно изобретению с MCP-1. Такое связывание может быть детектировано прямым или непрямым методом. Соответствующие методы и средства известны специалистам. Нуклеиновые кислоты согласно изобретению, среди прочих, могут содержать метку, которая позволяет детектировать нуклеиновые кислоты согласно изобретению, предпочтительно нуклеиновую кислоту, связанную с MCP-1. Такую метку предпочтительно выбирают из группы, включающей радиоактивные, ферментные и флуоресцентные метки. В принципе, все известные анализы, разработанные для антител, могут быть адаптированы для нуклеиновых кислот согласно изобретению, то есть антитело, связывающееся с мишенью, заменяют нуклеиновой кислотой, связывающейся с мишенью. В анализах с использованием антител, а именно немеченых связывающихся с мишенью антител, детекцию предпочтительно проводят с помощью “второго” антитела, которое модифицировано радиоактивными, ферментными и флуоресцентными метками и связывается с антителом, связывающимся с мишенью у своего Fc-фрагмента. В случае нуклеиновой кислоты, предпочтительно нуклеиновой кислоты согласно изобретению, указанную нуклеиновую кислоту модифицируют указанной меткой, где данная метка предпочтительно выбрана из группы, включающей биотин, Cy-3 и Cy-5, и детектируется с помощью антитела против такой метки, например антитела против биотина, анти-Cy3 антитела или анти-Cy5 антитела, или в случае, если такой меткой является биотин, то ее детектируют с помощью стрептавидина или авидина, которые по своей природе связываются с биотином. Указанное антитело, стрептавидин или авидин, в свою очередь, предпочтительно модифицируют соответствующей меткой, например, радиоактивной, ферментной или флуоресцентной меткой (такой как “второе антитело”).
В другом варианте изобретения молекулы нуклеиновой кислоты согласно изобретению детектируют или анализируют с помощью второго средства для детекции, где указанным средством для детекции является молекулярный “маяк”. Методика на основе молекулярного “маяка” известна специалистам в данной области. Вкратце, нуклеиновокислотные зонды, которые также называются молекулярными “маяками”, представляют собой обратный комплемент по отношению к образцу детектируемой нуклеиновой кислоты, и поэтому они гибридизуются с частью образца детектируемой нуклеиновой кислоты. После связывания с образцом нуклеиновой кислоты флуорофорные группы молекулярного “маяка” отделяются, что приводит к изменению флуоресцентного сигнала, предпочтительно к изменению его интенсивности. Такое изменение коррелирует с количеством имеющегося образца нуклеиновой кислоты.
Следует отметить, что детекция MCP-1 с использованием нуклеиновых кислот согласно изобретению, в частности, позволяет детектировать MCP-1, определенный в настоящей заявке.
Что касается детекции MCP-1, то предпочтительный способ такой детекции включает следующие стадии:
(а) получение образца, тестируемого на присутствие MCP-1,
(b) получение нуклеиновой кислоты согласно изобретению,
(c) проведение реакции образца с нуклеиновой кислотой предпочтительно в реакционном сосуде,
где стадия (a) может быть проведена перед стадией (b), либо стадия (b) может быть проведена перед стадией (a).
В предпочтительном варианте изобретения осуществляют дополнительную стадию (d), которая включает анализ на прохождение реакции образца с нуклеиновой кислотой. Предпочтительно нуклеиновую кислоту в стадии (b) иммобилизуют на поверхности. Такой поверхностью может быть поверхность реакционного сосуда, такого как реакционная пробирка, лунка планшета или поверхность предмета, содержащегося в таком реакционном сосуде, например сферы. Иммобилизация нуклеиновой кислоты на указанной поверхности может быть осуществлена любыми средствами, известными специалистам, включая, но не ограничиваясь ими, нековалентное или ковалентное связывание. Предпочтительно такое связывание осуществляется посредством ковалентной химической связи между поверхностью и нуклеиновой кислотой. Однако в соответствии с настоящим изобретением нуклеиновая кислота может быть опосредованно иммобилизована на поверхности, где такая опосредованная иммобилизация включает использование дополнительного компонента или пары партнеров по взаимодействию. Таким дополнительным компонентом предпочтительно является соединение, которое специфически взаимодействует с иммобилизуемой нуклеиновой кислотой, также называемой партнером по взаимодействию, и тем самым опосредует присоединение нуклеиновой кислоты к поверхности. Указанного партнера по взаимодействию предпочтительно выбирают из группы, включающей в себя нуклеиновые кислоты, полипептиды, белки и антитела. Предпочтительным партнером по взаимодействию является антитело, более предпочтительно моноклональное антитело. Альтернативно указанным партнером по взаимодействию является нуклеиновая кислота, предпочтительно функциональная нуклеиновая кислота. Более предпочтительно такую функциональную нуклеиновую кислоту выбирают из группы, включающей аптамеры, оптические изомеры и нуклеиновые кислоты, которые по меньшей мере частично комплементарны другой нуклеиновой кислоте. В другом альтернативном варианте изобретения связывание нуклеиновой кислоты с поверхностью опосредуется многокомпонентным партнером по взаимодействию. Такой многокомпонентный партнер по взаимодействию предпочтительно представляет собой пару партнеров по взаимодействию, либо такой партнер по взаимодействию состоит из первого члена и второго члена, где указанный первый член состоит из нуклеиновой кислоты или присоединен к нуклеиновой кислоте, а указанный второй член присоединен к поверхности или входит в состав данной поверхности. Многокомпонентный партнер по взаимодействию предпочтительно может быть выбран из группы, состоящей из пар партнеров по взаимодействию, включающих биотин и авидин, биотин и стрептавидин и биотин и нейтравидин. Предпочтительным первым членом такой пары партнеров по взаимодействию является биотин.
Предпочтительным результатом осуществления указанного способа является образование иммобилизованного комплекса MCP-1 и нуклеиновой кислоты, где более предпочтительным объектом детектирования является указанный комплекс. В соответствии с одним из вариантов изобретения осуществляют детекцию MCP-1, присутствующего в данном комплексе.
Соответствующим средством для детекции, удовлетворяющим данному требованию, является, например, любое детектируемое средство, специфичное для этой (этих) части (частей) MCP-1. Особенно предпочтительным средством для детекции является детектируемое средство, выбранное из группы, включающей нуклеиновые кислоты, полипептиды, белки и антитела, способы получения которых известны специалистам.
Способ детектирования MCP-1 также включает в себя удаление образца из реакционного сосуда, предпочтительно используемого для осуществления стадии (c).
В другом варианте изобретения указанный способ также включает в себя стадию иммобилизации партнера по взаимодействию с MCP-1 на поверхности, предпочтительно на поверхности, определяемой выше, где указанный партнер по взаимодействию определен в настоящей заявке, предпочтительно, как указано выше в описании соответствующего способа, более предпочтительно в различных вариантах изобретения, и такими партнерами являются нуклеиновые кислоты, полипептиды, белки и антитела. В этом варианте изобретения особенно предпочтительным детектируемым средством является нуклеиновая кислота согласно изобретению, где указанная нуклеиновая кислота может быть предпочтительно меченой или немеченой. В случае, если указанная нуклеиновая кислота является меченой, то она может быть детектирована прямо или опосредованно. Такой способ детекции может также включать использование второго средства для детекции, которое также предпочтительно выбрано из группы, включающей нуклеиновые кислоты, полипептиды, белки и варианты, указанные в различных описанных здесь вариантах изобретения. Такое детектируемое средство предпочтительно является специфичным в отношении нуклеиновой кислоты согласно изобретению. В более предпочтительном варианте изобретения вторым детектируемым средством является молекулярный “маяк”. В предпочтительном варианте изобретения нуклеиновая кислота или второе детектируемое средство либо оба этих средства могут включать детектируемую метку. Детектируемую метку предпочтительно выбирают из группы, включающей биотин, бромдезоксиуридиновую метку, дигоксигениновую метку, флуоресцентную метку, УФ-метку, радиоактивную метку и хелатообразующую молекулу. Альтернативно второе детектируемое средство взаимодействует с детектируемой меткой, которая предпочтительно содержится в нуклеиновой кислоте, состоит из нуклеиновой кислоты или присоединена к нуклеиновой кислоте. Особенно предпочтительные комбинации приводятся ниже, где
детектируемой меткой является биотин, а вторым средством для детекции является антитело против биотина, или
детектируемой меткой является биотин, а вторым средством для детекции является авидин или авидин-несущая молекула, или
детектируемой меткой является биотин, а вторым детектируемым средством является стрептавидин или стрептавидин-несущая молекула, или
детектируемой меткой является биотин, а вторым детектируемым средством является нейтравидин или нейтравидин-несущая молекула, или где
указанной детектируемой меткой является бромдезоксиуридин, а указанным вторым детектируемым средством является антитело против бромдезоксиуридина, или
указанной детектируемой меткой является дигоксигенин, а указанным вторым детектируемым средством является антитело против дигоксигенина, или
указанной детектируемой меткой является хелатообразующая молекула, а указанным вторым средством для детекции является радионуклид,
при этом предпочтительно, чтобы указанная детектируемая метка была присоединена к нуклеиновой кислоте. Следует отметить, что комбинация такого типа может быть также использована в варианте, где нуклеиновая кислота присоединена к поверхности. В указанном варианте изобретения предпочтительно, чтобы детектируемая метка была присоединена к партнеру по взаимодействию.
И, наконец, в соответствии с настоящим изобретением второе детектируемое средство детектируют с использованием третьего детектируемого средства, где указанным третьим детектируемым средством предпочтительно является фермент, а более предпочтительно, прохождение ферментативной реакции после детектирования второго детектируемого средства, либо указанным третьим детектируемым средством является средство для обнаружения радиоактивного излучения, а более предпочтительно радиоактивного излучения, испускаемого радионуклидом. При этом предпочтительно, чтобы указанное третье детектируемое средство специфически детектировалось вторым детектируемым средством и/или взаимодействовало с этим средством.
Кроме того, в одном из вариантов изобретения, в котором партнер по взаимодействию с MCP-1 иммобилизован на поверхности, а нуклеиновая кислота согласно изобретению предпочтительно присоединена к комплексу, образованному партнером по взаимодействию и MCP-1, образец может быть удален из реакционной смеси, более предпочтительно из реакционного сосуда после проведения стадии c) и/или стадии d).
В одном из вариантов изобретения нуклеиновая кислота согласно изобретению включает флуоресентную молекулу, где флуоресцентное излучение указанной флуоресцентной молекулы изменяется после образования комплекса между нуклеиновой кислотой и MCP-1 и отличается от излучения свободного MCP-1.
В другом варианте изобретения нуклеиновая кислота представляет собой производное нуклеиновой кислоты согласно изобретению, где указанное производное включает вместо аденозина по меньшей мере одно флуоресцентное производное аденозина. В предпочтительном варианте изобретения указанным флуоресцентным производным аденозина является этеноаденозин.
В другом варианте изобретения комплекс, состоящий из производного нуклеиновой кислоты согласно изобретению и MCP-1, детектируют с использованием флуоресценции.
В одном из вариантов указанного способа сигнал формируется на стадии (c) или на стадии (d), предпочтительно такой сигнал коррелирует с концентрацией MCP-1 в образце.
В предпочтительном аспекте изобретения указанные анализы могут быть осуществлены в 96-луночных планшетах, где компоненты иммобилизованы в реакционных сосудах, как описано выше, и в лунках, служащих в качестве реакционных сосудов.
Следует отметить, что все вышесказанное также относится к MCP-2, MCP-3, MCP-4 и/или эотаксину и по меньшей мере, до определенной степени, к нуклеиновым кислотам согласно изобретению, которые также связываются или взаимодействуют с MCP-2, MCP-3, MCP-4 и/или эотаксином.
Нуклеиновая кислота согласно изобретению может быть также использована в качестве исходного материала для разработки лекарственных средств. В принципе существует два возможных подхода. Одним из таких подходов является скрининг библиотек соединений, где указанными библиотеками соединений предпочтительно являются библиотеки низкомолекулярных соединений. В одном из вариантов изобретения указанным скринингом является широкомасштабный скрининг. Предпочтительно широкомасштабный скрининг соединений в анализе с использованием мишеней осуществляют быстрым и эффективным методом проб и ошибок. В наилучшем случае этот анализ осуществляют путем колориметрических измерений. Библиотеки, используемые в этих целях, известны специалистам.
Альтернативно нуклеиновая кислота согласно изобретению может быть использована для рациональной разработки лекарственных средств. Предпочтительно рациональная разработка лекарственных средств представляет собой конструирование основной структуры терапевтических средств. Исходя из трехмерной структуры мишени, которую обычно идентифицируют такими методами как рентгеновская кристаллография или спектроскопия ядерного магнитного резонанса, для поиска в базах данных, содержащих структуры многих различных химических соединений, используются компьютерные программы. Такой выбор осуществляют на компьютере, а идентифицированные соединения могут быть затем протестированы в лаборатории.
Рациональное конструирование лекарственных средств может быть осуществлено на основе любойнуклеиновой кислоты согласно изобретению и включает создание структуры, предпочтительно трехмерной структуры, аналогичной структуре заявленных нуклеиновых кислот или идентичной частям структуры заявленных нуклеиновых кислот, опосредующим связывание. В любом случае такая структура обладает такими же связывающими свойствами, что и заявленные нуклеиновые кислоты, или аналогичными свойствами. В любой дополнительной или альтернативной стадии рационального конструирования лекарственных средств предпочтительную трехмерную структуру частей нуклеиновых кислот, связывающихся с нейромедиатором, имитируют химическими группами, которые отличаются от нуклеотидов и нуклеиновых кислот. Благодаря такой имитации может быть сконструировано соединение, отличающееся от нуклеиновых кислот. Таким соединением предпочтительно являются небольшие молекулы или пептид.
В случае скрининга библиотек соединений, например, с помощью анализа на конкурентное связывание, известного специалистам, могут быть обнаружены соответствующие аналоги MCP-1, агонисты MCP-1 или антагонисты MCP-1. Такие анализы на конкурентное связывание могут быть проведены следующим образом. Заявленную нуклеиновую кислоту, предпочтительно оптический изомер, который представляет собой связывающуюся с мишенью L-нуклеиновую кислоту, присоединяют к твердой фазе. Для идентификации аналогов MCP-1 в данный анализ может быть включен меченый MCP-1. Потенциальный аналог может конкурировать с молекулами MCP-1 за связывание с оптическим изомером, что приводит к снижению интенсивности сигнала, продуцируемого соответствующей меткой. Скрининг для поиска агонистов или антагонистов может включать приведение анализа в клеточной культуре, известного специалистам.
Набор согласно изобретению может включать в себя по меньшей мере одну или несколько заявленных нуклеиновых кислот. Кроме того, этот набор может включать в себя по меньшей мере один или несколько позитивных или негативных контролей. В качестве позитивного контроля может служить, например, MCP-1, в частности MCP-1, к которому заявленная нуклеиновая кислота является специфичной или с которым она связывается, предпочтительно в жидкой форме. В качестве негативного контроля может служить, например, пептид, который обладает биофизическими свойствами, аналогичными свойствам MCP-1, но не распознается заявленными нуклеиновыми кислотами. Кроме того, указанный набор может включать один или несколько буферов. В данном наборе могут содержаться различные ингредиенты в осушенной или в лиофилизованной форме или в виде жидкого раствора. Набор может включать один или несколько контейнеров, которые, в свою очередь, могут содержать один или несколько ингредиентов данного набора. В другом варианте изобретения указанный набор включает инструкцию или вкладыш с инструкцией, в которой имеется информация для потребителя по применению данного набора и его различных ингредиентов.
Для оценки фармакокинетического и биодинамического профиля нуклеиновой кислоты согласно изобретению в нескольких физиологических жидкостях, тканях и органах человека и животных обычно проводят фармацевтическую и биоаналитическую оценку указанной нуклеиновой кислоты. Для этих целей могут быть применены любые методы детекции, описанные в настоящей заявке или известные специалистам. В другом аспекте настоящего изобретения для детекции нуклеиновой кислоты согласно изобретению проводят анализ методом “сэндвич”-гибридизации. В этом детектирующем анализе используют зонд для захвата и детектирующий зонд. Зонд для захвата комплементарен первой части нуклеиновой кислоты согласно изобретению, а детектирующий зонд комплементарен второй части нуклеиновой кислоты согласно изобретению. Оба зонда, то есть зонд для захвата и детектирующий зонд, могут быть образованы нуклеотидами ДНК, модифицированными нуклеотидами ДНК, модифицированными нуклеотидами РНК, нуклеотидами РНК, нуклеотидами LNA и/или нуклеотидами PNA.
Следовательно, зонд для захвата включает фрагмент последовательности, комплементарный 5'-концу нуклеиновой кислоты согласно изобретению, а детектирующий зонд включает фрагмент последовательности, комплементарный 3'-концу нуклеиновой кислоты согласно изобретению. В этом случае зонд для захвата иммобилизуют на поверхности или матрице у его 5'-конца, причем этот зонд для захвата может быть иммобилизован непосредственно у своего 5'-конца или посредством линкера, расположенного между 5'-концом и поверхностью, или матрицей. Однако, в принципе, линкер может быть связан с каждым нуклеотидом зонда для захвата. Линкер может быть образован гидрофильными линкерами, известными специалистам, или нуклеотидами D-ДНК, модифицированными нуклеотидами D-ДНК, нуклеотидами D-РНК, модифицированными нуклеотидами D-РНК, нуклеотидами D-LNA, нуклеотидами PNA, нуклеотидами L-РНК, нуклеотидами L-ДНК, модифицированными нуклеотидами L-РНК, модифицированными нуклеотидами L-ДНК и/или нуклеотидами L-LNA.
Альтернативно зонд для захвата включает фрагмент последовательности, комплементарный 3'-концу нуклеиновой кислоты согласно изобретению, а детектирующий зонд включает фрагмент последовательности, комплементарный 5'-концу нуклеиновой кислоты согласно изобретению. В этом случае зонд для захвата иммобилизуют на поверхности или матрице у его 3'-конца, причем этот зонд для захвата может быть иммобилизован непосредственно у своего 3'-конца или посредством линкера, расположенного между 3'-концом и поверхностью, или матрицей. Однако, в принципе, линкер может быть связан с каждым нуклеотидом фрагмента последовательности, комплементарного нуклеиновой кислоте согласно изобретению. Линкер может быть образован гидрофильными линкерами, известными специалистам, или нуклеотидами D-ДНК, модифицированными нуклеотидами D-ДНК, нуклеотидами D-РНК, модифицированными нуклеотидами D-РНК, нуклеотидами D-LNA, нуклеотидами PNA, нуклеотидами L-РНК, нуклеотидами L-ДНК, модифицированными нуклеотидами L-РНК, модифицированными нуклеотидами L-ДНК и/или нуклеотидами L-LNA.
Число нуклеотидов зонда для захвата и детектирующего зонда, которое может быть гибридизовано с нуклеиновой кислотой согласно изобретению, варьируется и может зависеть от числа нуклеотидов зонда для захвата, и/или детектирующего зонда, и/или самой нуклеиновой кислоты согласно изобретению. Общее число нуклеотидов зонда для захвата и детектирующего зонда, которое может гибридизоваться с нуклеиновой кислотой согласно изобретению, должно представлять собой максимальное число нуклеотидов, составляющих нуклеиновую кислоту согласно изобретению. Минимальное число нуклеотидов (2-10 нуклеотидов) детектирующего зонда и зонда для захвата должно гибридизоваться с 5'-концом или 3'-концом нуклеиновой кислоты согласно изобретению соответственно. Для достижения высокой специфичности и селективности между нуклеиновой кислотой согласно изобретению и другими нуклеиновыми кислотами, присутствующими в анализируемых образцах, общее число нуклеотидов зонда для захвата и детектирующего зонда должно быть равным числу нуклеотидов, составляющих нуклеиновую кислоту согласно изобретению, или должно быть равным максимальному числу нуклеотидов, составляющих нуклеиновую кислоту согласно изобретению.
Кроме того, детектируемый зонд предпочтительно содержит маркерную молекулу или метку, которая может быть детектирована, как описано выше. Такая метка или маркерная молекула может быть, в принципе, связана с каждым нуклеотидом указанного детектирующего зонда. Метка или маркер предпочтительно расположены у 5'-конца или у 3'-конца детектирующего зонда, где между нуклеотидами детектирующего зонда, комплементарными нуклеиновой кислоте согласно изобретению, могут быть встроены метка или линкер. Этот линкер может быть образован гидрофильными линкерами, известными специалистам, или нуклеотидами D-ДНК, модифицированными нуклеотидами D-ДНК, нуклеотидами D-РНК, модифицированными нуклеотидами D-РНК, нуклеотидами D-LNA, нуклеотидами PNA, нуклеотидами L-РНК, нуклеотидами L-ДНК, модифицированными нуклеотидами L-РНК, модифицированными нуклеотидами L-ДНК и/или нуклеотидами L-LNA.
Детектирование нуклеиновой кислоты согласно изобретению может быть осуществлено следующим образом. У одного своего конца нуклеиновая кислота согласно изобретению гибридизуется с зондом для захвата, а у другого своего конца - с детектируемым зондом. После этого не связанный детектирующий зонд удаляют, например, в одну или несколько стадий промывки. Затем может быть определено количество связанного детектирующего зонда, который предпочтительно содержит метку или маркерную молекулу.
В предпочтительном варианте изобретения используемый здесь термин “лечение” предпочтительно включает, дополнительно или альтернативно, предупреждение заболевания и/или мониторинг его течения.
Используемые здесь термины “заболевание” и “расстройство” предпочтительно являются взаимозаменяемыми, если это не оговорено особо.
Используемый здесь термин “включает” предпочтительно не ограничивается компонентами, перечисленными или описываемыми данным термином. Однако в альтернативном варианте термин “включает” может означать “содержит”, а поэтому ограничивается компонентами, перечисленными или описываемыми данным термином.
Различные идентификационные номера последовательностей SEQ ID NO:, химическая природа используемых здесь молекул нуклеиновой кислоты согласно изобретению и молекул-мишеней MCP-1, а также их фактические последовательности и их природные источники систематизированы в нижеследующей таблице.

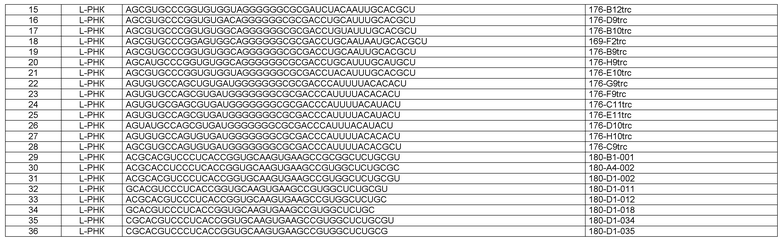
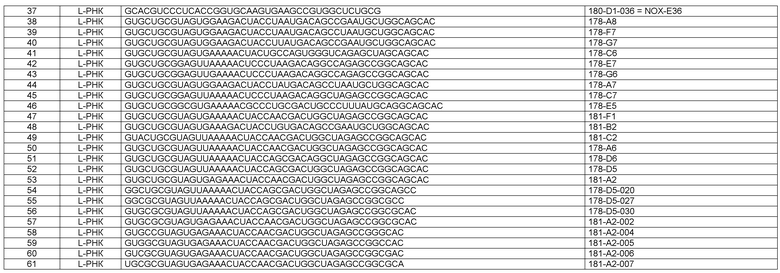











Для иллюстрации настоящего изобретения ниже представлены графические материалы, примеры и список последовательностей, исходя из которых будут более очевидны другие отличительные признаки, варианты и преимущества настоящего изобретения.
На фиг.1 проиллюстровано сопоставление последовательностей родственных лигандов РНК, связывающихся с человеческим MCP-1, с указанием мотива последовательности (“тип 1A”), который в предпочтительном варианте изобретения играет по существу главную роль в связывании с человеческим MCP-1.
На фиг.2 проиллюстрировано сопоставление последовательностей родственных лигандов РНК, связывающихся с человеческим MCP-1, с указанием мотива последовательности (“тип 1В”), который в предпочтительном варианте изобретения играет по существу главную роль в связывании с человеческим MCP-1 и с производными лигандов РНК 180-D1-002.
На фиг.3 проиллюстрировано сопоставление последовательностей родственных лигандов РНК, связывающихся с человеческим MCP-1, с указанием мотива последовательности (“тип 2”), который в предпочтительном варианте изобретения играет по существу главную роль в связывании с человеческим MCP-1.
На фиг.4 проиллюстрировано сопоставление последовательностей родственных лигандов РНК, связывающихся с человеческим MCP-1, с указанием мотива последовательности (“тип 3”), который в предпочтительном варианте изобретения играет по существу главную роль в связывании с человеческим MCP-1.
На фиг.5 проиллюстрированы производные лигандов РНК 178-D5 и 181-A2 (лигандов РНК человеческого MCP-1 с мотивом последовательности “типа 3”).
На фиг.6 проиллюстрировано сопоставление последовательностей родственных лигандов РНК, связывающихся с человеческим MCP-1, с указанием мотива последовательности (“тип 4”), который в предпочтительном варианте изобретения играет по существу главную роль в связывании с человеческим MCP-1 (другие последовательности).
На фиг.7 представлена таблица последовательностей нескольких различных лигандов РНК, связывающимся с человеческим MCP-1, которые не являются родственными мотивам MCP-1-связывающих последовательностей “типа 1A”, “типа 1B”; “типа 2”, “типа 3” или “типа 4”.
На фиг.8 проиллюстрировано сопоставление последовательностей производных лиганда РНК 188-A3-001 и 189-G7-001, которые связываются с мышиным MCP-1;
На фиг.9 представлен результат анализа на связывание аптамера D-NOX-E36 с биотинилированным человеческим D-MCP-1, проводимого при комнатной температуре и при 37°C, где указанный результат представлен как связывание аптамера при всех концентрациях биотинилированного человеческого D-MCP-1.
На фиг.10 представлен результат анализа на связывание аптамера D-mNOX-E36 с биотинилированным мышиным D-MCP-1, проводимого при комнатной температуре и при 37°C, где указанный результат представлен как связывание аптамера при всех концентрациях биотинилированного мышиного D-MCP-1.
На фиг.11 проиллюстрировано MCP-1-индуцированное высвобождение Ca++ в клетках THP-1 и была построена кривая “доза-ответ” для человеческого MCP-1, которая указывает на то, что 50% эффективная концентрация (EC50) составляет приблизительно 3 нМ и представляет собой интенсивности флуоресценции минус контроль при всех концентрациях человеческого MCP-1.
На фиг.12 проиллюстрирована эффективность оптического изомера NOX-E36 в анализе на высвобождение кальция, где клетки стимулировали 3 нM человеческого MCP-1, предварительно инкубированного при 37°C с различными количествами оптического изомера NOX-E36, и где указанные данные представлены как процент от контроля при всех концентрациях NOX-E36.
На фиг.13 проиллюстрирована эффективность оптического изомера mNOX-E36 в анализе на высвобождение кальция, где клетки стимулировали 5 нM мышиного MCP-1, предварительно инкубированного при 37°C с различными количествами оптического изомера mNOX-E36, и где указанные данные представлены как процент от контроля при всех концентрациях mNOX-E36.
На фиг.14 проиллюстрирован хемотаксис клеток ТНР-1, индуцированный человеческим MCP-1, где после 3-часовой миграции клеток THP-1 в направлении различных концентраций MCP-1 была построена кривая “доза-ответ” для МСР-1 и где полученные данные представлены как Х-кратное увеличение по сравнению с контролем при всех концентрациях человеческого MCP-1.
На фиг.15 проиллюстрирована эффективность оптического изомера NOX-E36 в анализе на хемотаксис, где указанные клетки мигрировали в направлении к 0,5 нМ человеческого МСР-1, предварительно инкубированного при 37°C с различными количествами оптического изомера NOX-E36, и где полученные данные представлены как процент от контроля при всех концентрациях оптического изомера NOX-E36.
На фиг.16 проиллюстрирована эффективность оптического изомера mNOX-E36 в анализе на хемотаксис, где указанные клетки мигрировали в направлении к 0,5 нМ мышиного МСР-1, предварительно инкубированного при 37°C с различными количествами оптического изомера mNOX-E36, и где полученные данные представлены как процент от контроля при всех концентрациях оптического изомера mNOX-E36.
На фиг.17 представлена сенсорограмма Biacore 2000 с указанием значения КD связывания оптического изомера NOX-E36 с человеческим MCP-1, который был иммобилизован на сенсорном чипе PioneerF1 путем проведения процедуры связывания с амином, где указанная сенсорограмма представлена как зависимость ответа (RU) от времени.
На фиг.18 представлена сенсорограмма Biacore 2000, иллюстрирующая связывание оптического изомера NOX-E36 с белками, принадлежащими к семейству человеческих MCP (huMCP-1, huMCP-2, huMCP-3), и с человеческим эотаксином, которые были иммобилизованы на сенсорном чипе PioneerF1 и СМ4 соответственно путем проведения процедуры связывания с амином, где указанная сенсорограмма представлена как зависимость ответа (RU) от времени.
На фиг.19 представлена сенсорограмма Biacore 2000, иллюстрирующая связывание оптического изомера NOX-E36 с MCP-1 различных видов (собачьим MCP-1, обезьяньим MCP-1, человеческим MCP-1, свиным MCP-1, кроличьим MCP-1, мышиным MCP-1, крысиным MCP-1), где различные формы MCP-1 были иммобилизованы на сенсорном чипе PioneerF1 и СМ4 соответственно путем проведения процедуры связывания с амином и где указанная сенсорограмма представлена как зависимость ответа (RU) от времени.
На фиг.20 представлена сенсорограмма Biacore 2000 с указанием значения КD связывания оптического изомера 181-A2-018 с человеческим MCP-1, который был иммобилизован на сенсорном чипе CM4 путем проведения процедуры связывания с амином, где указанная сенсорограмма представлена как зависимость ответа (RU) от времени.
На фиг.21 представлена сенсорограмма Biacore 2000, иллюстрирующая связывание оптического изомера 181-A2-018 с белками, принадлежащими к семейству человеческих MCP (huMCP-1, huMCP-2, huMCP-3), и с человеческим эотаксином, которые были иммобилизованы на сенсорном чипе PioneerF1 и СМ4 соответственно путем проведения процедуры связывания с амином, где указанная сенсорограмма представлена как зависимость ответа (RU) от времени.
На фиг.22 представлена сенсорограмма Biacore 2000, иллюстрирующая связывание оптического изомера 181-A2-018 с MCP-1 различных видов (собачьим MCP-1, обезьяньим MCP-1, человеческим MCP-1, свиным MCP-1, кроличьим MCP-1, мышиным MCP-1, крысиным MCP-1), где различные формы MCP-1 были иммобилизованы на сенсорном чипе PioneerF1 и СМ4 соответственно путем проведения процедуры связывания с амином и где указанная сенсорограмма представлена как зависимость ответа (RU) от времени.
На фиг.23 проиллюстрировано проводимое с помощью программы Clustal W сопоставление последовательностей MCP-1 от млекопитающих различных видов, а также человеческого MCP-2, MCP-3 и эотаксина (только в положениях 1-76).
На фиг.24A представлена таблица, в которой систематизирована специфичность связывания NOX-E36 и 181-A2-018 с MCP-1 от млекопитающих различных видов, а также человеческого MCP-2, MCP-3 и эотаксина.
На фиг.24B представлена таблица, в которой систематизирована селективность NOX-E36, определенная в анализе Biacore, где биотинилированный NOX-E36 был иммобилизован на поверхности сенсорного чипа, и было проанализировано связывание панели различных хемокинов CC и CXC с NOX-E36.
На фиг.24C проиллюстрирован кинетический анализ взаимодействия NOX-E36 с хемокинами, определенного в анализе Biacore, где указанные хемокины были ковалентно иммобилизованы на поверхности сенсорного чипа СМ5, и были введены различные концентрации NOX-E36, а характер связывания с NOX-E36s был проанализирован с помощью компьютерной программы BiaEvaluation.
На фиг.24D проиллюстрирована кривая “доза-ответ” для хемотаксиса клеток THP-1, стимулированных МIP-1α, где эффективная концентрация составляла приблизительно 0,2 нM.
На фиг.24E проиллюстрировано ингибирование MIP-1α-индуцированного хемотаксиса с использованием NOX-E36. NOX-E36 не оказывал влияния на MIP-1α-индуцированный хемотаксис клеток THP-1.
На фиг.25 проиллюстрирована эффективность оптического изомера NOX-E36-3'-ПЭГ в анализе на высвобождение кальция, где клетки были стимулированы 3 нМ человеческого МСР-1, предварительно инкубированного при 37°C с различными количествами оптического изомера NOX-E36-3'-ПЭГ, и где полученные данные представлены как процент от контроля при всех концентрациях оптического изомера NOX-E36-3'-ПЭГ.
На фиг.26 проиллюстрирована эффективность оптического изомера NOX-E36-3'-ПЭГ в анализе на хемотаксис, где клетки мигрировали в направлении 0,5 нМ человеческого МСР-1, предварительно инкубированного при 37°C с различными количествами оптического изомера NOX-E36-3'-ПЭГ, и где полученные данные представлены как процент от контроля при всех концентрациях оптического изомера NOX-E36-3'-ПЭГ.
На фиг.27А проиллюстрирована эффективность оптического изомера NOX-E36-5'-ПЭГ в анализе на высвобождение кальция, где клетки были стимулированы 3 нМ человеческого МСР-1, предварительно инкубированного при 37°С с различными количествами оптического изомера NOX-E36-5'-ПЭГ, и где полученные данные представлены как процент от контроля при всех концентрациях оптического изомера NOX-E36-5'-ПЭГ.
На фиг.27B проиллюстрирована эффективность оптического изомера NOX-E36-5'-ПЭГ в анализе на хемотаксис, где клетки мигрировали в направлении 0,5 нМ человеческого MCP-1, предварительно инкубированного при 37°C с различными количествами оптического изомера NOX-E36-5'-ПЭГ, где полученные данные представлены как процент от контроля при всех концентрациях оптического изомера NOX-E36-5'-ПЭГ.
На фиг.28 проиллюстрировано высвобождение Ca++ в клетках ТНР-1, индуцированное мышиным МСР-1, и была построена кривая “доза-ответ” для мышиного МСР-1, которая указывала на 50% эффективную концентрацию (EC50), составляющую приблизительно 5 нМ, где полученные данные представлены как интенсивность флуоресценции минус контроль при всех концентрациях мышиного MCP-1.
На фиг.29 проиллюстрирована эффективность оптического изомера mNOX-E36-3'-ПЭГ против мышиного MCP-1 в анализе на высвобождение кальция, где клетки были стимулированы 3 нM мышиного MCP-1, предварительно инкубированного при 37°C с различными количествами оптического изомера mNOX-E36-3'-ПЭГ, и где полученные данные представлены как процент от контроля при всех концентрациях оптического изомера mNOX-E36-3'-ПЭГ.
На фиг.30 проиллюстрирован хемотаксис клеток ТHP-1, индуцированный мышиным МСР-1, и после 3-часовой миграции клеток THP-1 по направлению к различным концентрациям mMCP-1 строили кривую “доза-ответ” для mMCP-1, где полученные данные представлены как Х-кратное увеличение по сравнению с контролем при всех концентрациях мышиного МСР-1.
На фиг.31 проиллюстрирована эффективность оптического изомера mNOX-E36-3'-ПЭГ против мышиного МСР-1 в анализе на хемотаксис, где клетки мигрировали по направлению к 0,5 нМ мышиного МСР-1, предварительно инкубированного при 37°С с различными количествами оптического изомера mNOX-E36-3'-ПЭГ, и где полученные данные представлены как процент от контроля при всех концентрациях антимышиного оптического изомера mNOX-E36-3'-ПЭГ.
На фиг.32 проиллюстрирована сенсорограмма Biacore 2000 с указанием величины КD связывания аптамера D-mNOX-E36 с мышиным D-MCP-1, который был иммобилизован на сенсорном чипе PioneerF1 путем проведения процедуры связывания с амином, где полученные данные представлены как зависимость ответа (RU) от времени.
На фиг.33 представлена сенсорограмма Biacore 2000, иллюстрирующая связывание аптамера D-mNOX-E36 с человеческим D-MCP-1 и мышиным D-MCP-1, где указанные две различные формы D-MCP-1 были иммобилизованы на сенсорных чипах PioneerF1 и CM4 соответственно путем проведения процедуры связывания с амином и где полученные данные представлены как зависимость ответа (RU) от времени.
На фиг.34 проиллюстрированы почечные срезы, взятые у 24-недельных MRLlpr/lpr-мышей и окрашенные периодной шиффовой кислотой (PAS) и антителами против Mac-2 (макрофаги) и CD3 (T-клетки), как указано на фигуре, где представленные изображения являются репрезентативными для 7-12 мышей в каждой группе (исходное увеличение PAS: x 100, PAS-вставки: x 400, Mac2: x 400, CD3: x 100.
На фиг.35 представлена таблица, в которой приводятся параметры почечных функций и гистологические данные для различных групп 24-недельных MRLlpr/lpr-мышей.
На фиг.36 представлена количественная оценка гистологических изменений, проводимая с помощью морфометрии и осуществляемая на окрашенных серебром срезах, полученных от мышей всех групп; где А - индекс объема интерстициальной ткани; В - индекс увеличения объема трубочек и С - индекс поражения тубулярных клеток вычисляли как процент в поле зрения микроскопа при большом увеличении и выражали как среднее ± ср.кв.от.
На фиг.37 показан коэффициент выживаемости MRLlpr/lpr -мышей в различных группах обработки, вычисленный с помощью анализа Каплана-Майера.
На фиг.38 проиллюстрирована экспрессия мРНК для СС-хемокинов CCL2 и CCL5 в почках, определенная с помощью ОТ-ПЦР в реальном времени с использованием полноразмерной почечной РНК, собранной у 5 мышей в каждой группе, где уровни мышиной РНК для каждой группы выражены в единицах на соответствующую экспрессию рРНК 18S.
На фиг.39 проиллюстрировано уменьшение патологических изменений в легких после обработки оптическим изомером mNOX-E36-3'ПЭГ; ткани легких были взяты у мышей всех групп в возрасте 24 недель и была проведена полуколичественная оценка этих тканей; обработка оптическими изомерами mNOX-E36 и mNOX-E36-3'ПЭГ приводила к уменьшению перибронхиолярного воспаления у MRLlpr/lpr-мышей; изображения являются репрезентативными для 7-11 мышей в каждой группе и даны с исходным увеличением x 100.
На фиг.40 проиллюстрированы манифестации волчанки на коже MRLlpr/lpr-мышей в возрасте 24 недель, которые обычно наблюдаются на морде или на шее животного (левая мышь), но которые реже встречались у мышей, обработанных оптическим изомером, направленным против mCCL2 (правая мышь).
На фиг.41 представлены данные анализа сыворотки и данные гистологического анализа у MRLlpr/lpr-мышей в возрасте 24 недель.
На фиг.42 проиллюстрирована фармакокинетика ПЭГилированных и неПЭГилированных оптических изомеров против mCCL2 в плазме во время исследований и показан график зависимости концентрации оптического изомера mNOX-E36 в плазме от времени.
На фиг.43 проиллюстрирован проточный цитометрический анализ на CCR2 в костном мозге и в периферической крови у 24-недельных MRLlpr/lpr-мышей, обработанных носителем или оптическим изомером mNOX-E36-3'-ПЭГ; полученные данные представлены как средний процент CCR2-позитивных клеток ± ср.кв.от. в костном мозге или в периферической крови у 5 мышей в каждой группе.
На фиг.44 представлены уровни CCL2 в сыворотке PoC-ПЭГ- (белые столбцы) и уровни mNOX-E36-3'-ПЭГ (mNOX-E36-P)-обработанных (черные столбцы) db/db-мышей 1К, где указанные уровни были определены с помощью ELISA в различные периоды времени, как показано на данной фигуре; полученные данные выражены как среднее ± ср.кв.от.; *, p<0,05 mNOX-E36-3'ПЭГ (mNOX-E36-P) и PoC-ПЭГ.
На фиг.45 показано число Mac-2- и Ki-67-позитивных клеток, инфильтрованных в клубочках и в интерстициальной ткани необработанных db/db-мышей, РОС-ПЭГ-обработанных db/db-мышей или mNOX-E36-3'-ПЭГ-обработанных db/db-мышей.
На фиг.46 проиллюстрирован диабетический гломерулосклероз у 6-месячных db/db-мышей, где почечные срезы, взятые у мышей различных групп, были окрашены периодной шиффовой кислотой, а 15 клубочков от каждого почечного среза были оценены на степень поражения гломерулосклерозом; представленные здесь изображения, с исходным увеличением 400x, иллюстрируют степень поражения клубочков по шкале соответствующих оценок, как показано на данной фигуре; указанный график иллюстрирует средний процент каждой оценки ± ср.кв.от. для всех мышей каждой группы (n=7-10); *, p<0,05 для mNOX-E36-3'-ПЭГ (mNOX-E36-P)- и PoC-ПЭГ (PoC-P)-обработанных db/db-мышей 1К.
На фиг.47 показан уровень клубочковой фильтрации (GFR) у 6-месячных mNOX-E36-3'-ПЭГ (mNOX-E36-P)- и PoC-ПЭГ(PoC-P)-обработанных db/db-мышей 1К, где GFR определяли по кинетике клиренса ФИТЦ-инулина у групп PoC-ПЭГ- и mNOX-E36-3'-ПЭГ-обработанных db/db-мышей 1К по окончании исследования.
На фиг.48 показан объем атрофии канальцев и интерстициальной ткани у 6-месячных db/db-мышей, где изображения окрашенных серебром почечных срезов иллюстрируют репрезентативные почки, взятые у мышей соответствующих групп (исходное увеличение 100х); величины выражены как среднее ± ср.кв.от. соответствующего индекса морфометрического анализа, проводимого у 7-10 мышей в каждой группе; *, p<0,05 для db/db-мышей 2К по сравнению с мышами BKS дикого типа; #, p<0,05 для db/db-мышей 1К по сравнению с db/db-мышами 2К; †, p<0,05 для mNOX-E36-3'-ПЭГ (mNOX-E36-ПЭГ-обработанных мышей) по сравнению с PoC-ПЭГ-обработанными db/db-мышами 1К.
На фиг.49 проиллюстрирована экспрессия мРНК CCL2 в почках db/db-мышей, определенная с помощью ОТ-ПЦР в реальном времени с использованием полноразмерной РНК почек, взятых у 6-10 мышей в каждой группе, где указанные уровни мРНК у мышей в каждой группе выражены по отношению к соответствующей экспрессии рРНК 18S.
На фиг.50 проиллюстрирована пространственная экспрессия CCL2 в почках db/db-мышей, определенная методом иммунологического окрашивания, а изображения иллюстрируют репрезентативные срезы почек, взятых у 6-месячных мышей соответствующих групп, как показано на данной фигуре (исходное увеличение - 200x).
Пример 1
Нуклеиновые кислоты, связывающиеся с человеческим МСР-1
С использованием биотинилированного человеческого D-MCP-1, служащего в качестве мишени, может быть получено несколько нуклеиновых кислот, которые связываются с человеческим МСР-1 и нуклеотидные последовательности которых представлены на фигурах 1-7. Нуклеиновые кислоты были охарактеризованы на уровне аптамеров, т.е. на уровне D-нуклеиновых кислот с помощью анализов на конкурентное ингибирование или на прямое ингибирование с использованием биотинилированного человеческого D-MCP-1 (пример 4), либо на уровне оптических изомеров, т.е. на уровне L-нуклеиновой кислоты с использованием природной конфигурации MCP-1 (L-MCP) путем измерения методом поверхностного плазмонного резонанса на оборудовании Biacore 2000 (пример 7) или с помощью анализа на высвобождение Ca++ в клеточной культуре in vitro (пример 5), или анализа на хемотаксис in vitro (пример 6).
Полученные таким образом молекулы нуклеиновой кислоты имеют различные мотивы последовательностей, и основные четыре типа этих последовательностей определены на фиг.1 и 2 (типа 1А/1В), на фиг.3 (типа 2), на фиг.4 и 5 (типа 3) и на фиг.6 (типа 4). Дополнительные MCP-1-связывающие нуклеиновые кислоты, которые не являются родственными друг другу и другим мотивам последовательностей, описанных в настоящей заявке, перечислены на фиг.7. В целях определения мотивов нуклеотидных последовательностей, для обозначения нетипичных нуклеотидов используются сокращения согласно ИЮПАК:
Любые последовательности нуклеиновых кислот или последовательности фрагментов и боксов соответственно, если это не оговорено особо, расположены в направлении 5'→3'.
MCP-1-связывающие нуклеиновые кислоты типа 1А (фиг.1)
Как показано на фиг.1, все последовательности MCP-1-связывающих нуклеиновых кислот типа 1A включают несколько последовательностей фрагментов или боксов, где боксы B1A и B1B представляют собой 5'- и 3'-концевые фрагменты, которые могут гибридизоваться друг с другом. Однако такая гибридизация необязательно должна происходить в молекуле, присутствующей фактически в физиологических условиях. Боксы B2, B3, B4, B5 и бокс B6 фланкируются боксом B1A и боксом B1B.
Нуклеиновые кислоты были охарактеризованы на уровне аптамеров с помощью анализа на прямое и конкурентное ингибирование с использованием биотинилированного человеческого D-MCP-1 и классифицированы по характеру связывания (пример 4). Отобранные последовательности синтезировали в виде оптического изомера (пример 3) и тестировали с использованием природной конфигурации MCP-1 (L-MCP) в анализе in vitro на высвобождение Ca++ в клеточной культуре (пример 5).
Последовательности MCP-1-связывающих нуклеиновых кислот типа 1A могут отличаться по определенным боксам, которые влияют на аффинность связывания с MCP-1. Исходя из анализа на связывание различных MCP-1-связывающих нуклеиновых кислот, систематизированных как МСР-1-связывающие нуклеиновые кислоты типа 1A , ниже отдельно описаны боксы B1A, B2, B3, B4, B5, B6 и B1B и их нуклеотидные последовательности, а указанные более предпочтительные боксы определены по их роли, которую они играют в связывании с MCP-1, а именно:
- боксы B1A и B1B, представляющие собой 5'- и 3'-концевые фрагменты, которые могут гибридизоваться друг с другом, где B1A представляет собой AGCRUG, предпочтительно AGCGUG; и где B1B представляет собой CRYGCU, предпочтительно CACGCU;
- бокс B2, который представляет собой CCCGGW, предпочтительно CCCGGU;
- бокс B3, который представляет собой GUR, предпочтительно GUG;
- бокс B4, который представляет собой RYA, предпочтительно GUA;
- бокс B5, который представляет собой GGGGGRCGCGAYC, предпочтительно GGGGGGCGCGACC;
- бокс B6, который представляет собой UGCAAUAAUG или URYAWUUG, предпочтительно UACAUUUG.
Как описано на фиг.1, молекула нуклеиновой кислоты, обозначаемая 176-E10trc, обладает наилучшей аффинностью связывания с MCP-1 (как аптамер в анализе на ингибирование при КD=5 нМ, а также как оптический изомер с IC50=4-5 нM в анализе in vitro на высвобождение Ca++ в клеточной культуре), И поэтому она может иметь оптимальную последовательность и оптимальную комбинацию элементов последовательностей B1A, B2, B3, B4, B5, B6 и B1B.
МСР-1-связывающие нуклеиновые кислоты типа 1B (фиг.2)
Как показано на фиг.2, все последовательности типа 1B содержат несколько последовательностей фрагментов или боксов, где указанные боксы B1A и B1B представляют собой 5'- и 3'-концевые фрагменты, которые могут гибридизоваться друг с другом, а боксы B2, B3, B4, B5 и бокс B6 фланкированы боксом B1A и боксом B1B. Однако такая гибридизация необязательно должна происходить в молекуле, присутствующей фактически в физиологических условиях.
Нуклеиновые кислоты были охарактеризованы на уровне аптамеров с помощью анализа на прямое и конкурентное ингибирование с использованием биотинилированного человеческого D-MCP-1 и классифицированы по характеру связывания (пример 4). Отобранные последовательности синтезировали в виде оптического изомера (пример 3) и тестировали с использованием природной конфигурации MCP-1 (L-MCP) в анализе in vitro на высвобождение Ca++ в клеточной культуре (пример 5).
Последовательности MCP-1-связывающих нуклеиновых кислот типа 1В могут отличаться по определенным боксам, которые влияют на аффинность связывания с MCP-1. Исходя из анализа на связывание различных MCP-1-связывающих нуклеиновых кислот, систематизированных как МСР-1-связывающие нуклеиновые кислоты типа 1В , ниже отдельно описаны боксы B1A, B2, B3, B4, B5, B6 и B1B и их нуклеотидные последовательности, а указанные более предпочтительные боксы определены по их роли, которую они играют в связывании с MCP-1, а именно:
- боксы B1A и B1B, которые могут гибридизоваться друг с другом, где B1A представляет собой AGYRUG, предпочтительно AGCGUG; и где B1B представляет собой CAYRCU, предпочтительно CACGCU;
- бокс B2, который представляет собой CCAGCU или CCAGY, предпочтительно CCAGU;
- бокс B3, который представляет собой GUG;
- бокс B4, который представляет собой AUG;
- бокс B5, который представляет собой GGGGGGCGCGACC;
- бокс B6, который представляет собой CAUUUUA или CAUUUA, предпочтительно CAUUUUA.
Как описано на фиг.2, молекула нуклеиновой кислоты, обозначаемая 176-С9trc, обладает наилучшей аффинностью связывания с MCP-1 (как аптамер в анализе на ингибирование при КD=5 нМ, а также как оптический изомер с IC50=4-5 нM в анализе in vitro на высвобождение Ca++ в клеточной культуре), и поэтому она может иметь оптимальную последовательность и оптимальную комбинацию элементов последовательностей B1A, B2, B3, B4, B5, B6 и B1B.
МСР-1-связывающие нуклеиновые кислоты типа 2 (фиг.3)
Как показано на фиг.3, все последовательности типа 2 содержат несколько последовательностей фрагментов или боксов, где указанные боксы B1A и B1B представляют собой 5'- и 3'-концевые фрагменты, которые могут гибридизоваться друг с другом, а бокс B2 представляет собой центральный элемент последовательности. Однако такая гибридизация необязательно должна происходить в молекуле, присутствующей фактически в физиологических условиях.
Нуклеиновые кислоты были охарактеризованы на уровне аптамеров с помощью анализа на прямое и конкурентное ингибирование с использованием биотинилированного человеческого D-MCP-1 и классифицированы по характеру связывания (пример 4). Отобранные последовательности синтезировали в виде оптического изомера (пример 3) и тестировали с использованием природной конфигурации MCP-1 (L-MCP) в анализе in vitro на высвобождение Ca++ в клеточной культуре (пример 5) или в анализах in vitro на хемотаксис (пример 6).
Последовательности MCP-1-связывающих нуклеиновых кислот типа 3 могут отличаться по определенным боксам, которые влияют на аффинность связывания с MCP-1. Исходя из анализа на связывание различных MCP-1-связывающих нуклеиновых кислот, систематизированных как МСР-1-связывающие нуклеиновые кислоты типа 2, ниже отдельно описаны боксы B1A, B2 и B1B и их нуклеотидные последовательности, а указанные более предпочтительные боксы определены по их роли, которую они играют в связывании с MCP-1, а именно:
- боксы B1A и B1B, 5'- и 3'-концевые фрагменты, которые могут гибридизоваться друг с другом, где B1A представляет собой ACGCA, а B1B представляет собой UGCGU, или B1A представляет собой CGCA, а B1B представляет собой UGCG, или B1A представляет собой GCA, а B1B представляет собой UGCG или UGC; предпочтительно B1A представляет собой GCA, а B1B представляет собой UGCG;
- бокс B2, CSUCCCUCACCGGUGCAAGUGAAGCCGYGGCUC, предпочтительно CGUCCCUCACCGGUGCAAGUGAAGCCGUGGCUC.
Как показано на фиг.3, нуклеиновая кислота, обозначаемая 180-D1-002, а также производные 180-D1-002, такие как 180-D1-011, 180-D1-012, 180-D1-035 и 180-D1-036 (= NOX-E36), обладают наилучшей аффинностью связывания с MCP-1, как аптамеры в анализе на ингибирование или в анализе на конкурентное ингибирование при КD<1 нМ, а поэтому они могут иметь оптимальную последовательность и оптимальную комбинацию элементов последовательностей B1A, B2, и B1B.
Для молекулы нуклеиновой кислоты D-NOX-E36 (D-180-D1-036; SEQ ID NО:159) была определена константа диссоциации (KD), равная 890±65 пМ при комнатной температуре (RT) и 146±13 пМ при 37°C (пример 4; фиг.9). Соответствующий оптический изомер NOX-E36 (180-D1-036; SEQ ID NO:37) обнаруживает ингибирующую концентрацию (IC50) 3-4 нM в анализе in vitro на высвобождение Ca++ (пример 5; фиг.12) и приблизительно 0,5 нM в анализе in vitro на хемотаксис (пример 6; фиг.15). Для ПЭГилированных производных NOX-E36, NOX-E36-3'ПЭГ и NOX-E36-5'ПЭГ IC50, определенная в анализе на высвобождение Ca++, составляет приблизительно 3 нМ (пример 5, фиг.25 и фиг.27A), а в анализе на хемотаксис она составляет <1 нM (пример 6; фиг.26 и фиг.27B).
МСР-1-связывающие нуклеиновые кислоты типа 3 (фиг.4+5)
Как показано на фиг.4 и 5, все последовательности типа 3 включают несколько последовательностей фрагментов или боксов, где три пары боксов являются характерными для МСР-1-связывающих нуклеиновых кислот типа 3. Оба бокса B1A и B1B, а также боксы B2A и B2B и боксы B5A и B5B обладают способностью гибридизоваться друг с другом. Однако такая гибридизация необязательно должна происходить в молекуле, присутствующей фактически в физиологических условиях. Между этими потенциально гибридизованными элементами последовательности находятся негибридизующиеся нуклеотиды, определяемые как бокс В3, бокс В4 и бокс В6.
Нуклеиновые кислоты были охарактеризованы на уровне аптамеров с помощью анализа на прямое и конкурентное ингибирование с использованием биотинилированного человеческого D-MCP-1 и классифицированы по характеру связывания (пример 4). Отобранные последовательности синтезировали в виде оптического изомера (пример 3) и тестировали с использованием природной конфигурации MCP-1 (L-MCP) в анализах in vitro на хемотаксис (пример 6) или в анализах с применением методов измерения Biacore (пример 7).
Последовательности MCP-1-связывающих нуклеиновых кислот типа 3 могут отличаться определенными боксами, которые влияют на аффинность связывания с MCP-1. Исходя из анализа на связывание различных MCP-1-связывающих нуклеиновых кислот, систематизированных как МСР-1-связывающие нуклеиновые кислоты типа 3, ниже отдельно описаны боксы B1A, B2A, B3, B2B, B4, B5A, B6, B5B, B1B и их нуклеотидные последовательности, а указанные более предпочтительные боксы определены по их роли, которую они играют в связывании с MCP-1, а именно:
- боксы B1A и B1B, 5'- и 3'-концевые фрагменты, которые могут гибридизоваться друг с другом, где B1A представляет собой GURCUGC, а B1B представляет собой GCAGCAC; при этом предпочтительно B1A представляет собой GUGCUGC, а B1B представляет собой GCAGCAC; или B1A представляет собой GKSYGC, а B1B представляет собой GCRSMC; при этом предпочтительно B1A представляет собой GUGCGC, а B1B представляет собой GCGCAC; или B1A представляет собой KBBSC, а B1B представляет собой GSVVM; при этом предпочтительно B1A представляет собой KKSSC, а B1B представляет собой GSSMM; или B1A представляет собой BNGC, а B1B представляет собой GCNV; при этом предпочтительно B1A представляет собой SNGC, а B1B представляет собой GCNS, а наиболее предпочтительно B1A представляет собой GGGC, а B1B представляет собой GCCC;
- боксы B2A и B2B, фрагменты, которые могут гибридизоваться друг с другом, где B2A представляет собой GKMGU, а B2B представляет собой ACKMC; при этом предпочтительно B2A представляет собой GUAGU, а B2B представляет собой ACUAC;
- бокс B3, который представляет собой KRRAR, предпочтительно UAAAA или GAGAA;
- бокс B4, который представляет собой CURYGA, или CUWAUGA, или CWRMGACW, или UGCCAGUG, предпочтительно CAGCGACU или CAACGACU;
- боксы B5A и B5B, фрагменты, которые могут гибридизоваться друг с другом, где B5A представляет собой GGY, а B5B представляет собой GCYR, причем GCY может гибридизоваться с нуклеотидами B5A; или B5A представляет собой CWGC, а B5B представляет собой GCWG; при этом предпочтительно B5A представляет собой GGC, а B5B представляет собой GCCG;
- бокс B6, который представляет собой: YAGA или CKAAU или CCUUUAU, а предпочтительно UAGA.
Как показано на фиг.4 и 5, нуклеиновая кислота, обозначенная 178-D5, и ее производное 178-D5-030, а также нуклеиновая кислота 181-A2 и ее производные 181-A2-002, 181-A2-004, 181-A2-005, 181-A2-006, 181-A2-007, 181-A2-017, 181-A2-018, 181-A2-019, 181-A2-020, 181-A2-021 и 181-A2-023 обладают наилучшей аффинностью связывания с MCP-1. 178-D5 и 178-D5-030 были определены как аптамеры в анализе или на прямое или конкурентное ингибирование (пример 4) с КD приблизительно 500 пМ. В том же самом эксперименте было определено, что КD нуклеиновой кислоты 181-A2 составляет приблизительно 100 пМ. В анализе Biacore (пример 7) было определено, что КD нуклеиновой кислоты 181-A2 и ее производных по отношению к MCP-1 составляет 200-300 пМ. В анализе на высвобождение Ca++ и в анализе на хемотаксис с культивированными клетками (примеры 5 и 6 соответственно) было определено, что IC50 для обеих нуклеиновых кислот 178-D5 и 181-A2 составляет приблизительно 500 пМ. Поэтому нуклеиновые кислоты 178-D5 и 181-A2, а также их производные могут иметь оптимальную последовательность и оптимальную комбинацию элементов последовательностей B1A, B2A, B3, B2B, B4, B5A, B6, B5B и B1B.
МСР-1-связывающие нуклеиновые кислоты типа 4 (фиг.6)
Как показано на фиг.6, все последовательности типа 4 содержат несколько последовательностей, фрагментов или боксов, где указанные боксы B1A и B1B представляют собой 5'- и 3'-концевые фрагменты, которые могут гибридизоваться друг с другом, а бокс B2 представляет собой центральный элемент последовательности.
Нуклеиновые кислоты были охарактеризованы на уровне аптамеров с помощью прямых анализов на ингибирование с использованием биотинилированного человеческого D-MCP-1 и классифицированы по характеру связывания (пример 4). Отобранные последовательности синтезировали в виде оптического изомера (пример 3) и тестировали с использованием природной конфигурации MCP-1 (L-MCP) в анализе in vitro на высвобождение Ca++ в клеточной культуре (пример 5) и/или в анализе на хемотаксис (пример 6).
Последовательности MCP-1-связывающих нуклеиновых кислот типа 4 могут отличаться определенными боксами, которые влияют на аффинность связывания с MCP-1. Исходя из анализа на связывание различных MCP-1-связывающих нуклеиновых кислот, систематизированных как МСР-1-связывающие нуклеиновые кислоты типа 2 , ниже отдельно описаны боксы B1A, B2 и B1B и их нуклеотидные последовательности, а указанные более предпочтительные боксы определены по их роли, которую они играют в связывании с MCP-1, а именно:
- боксы B1A и B1B, 5'- и 3'-концевые фрагменты, которые могут гибридизоваться друг с другом, где B1A представляет собой AGCGUGDU, а B1B представляет собой GNCASGCU; или B1A представляет собой GCGCGAG, а B1B представляет собой CUCGCGUC; или B1A представляет собой CSKSUU, а B1B представляет собой GRSMSG; или B1A представляет собой GUGUU, а B1B представляет собой GRCAC; или B1A представляет собой UGUU, а B1B представляет собой GGCA; предпочтительно B1A представляет собой CSKSUU, а B1B представляет собой GRSMSG; а наиболее предпочтительно B1A представляет собой CCGCUU, а B1B представляет собой GGGCGG; и
- бокс B2, который представляет собой AGNDRDGBKGGURGYARGUAAAG, или AGGUGGGUGGUAGUAAGUAAAG, или CAGGUGGGUGGUAGAAUGUAAAGA, а предпочтительно AGGUGGGUGGUAGUAAGUAAAG.
Как показано на фиг.6, нуклеиновые кислоты, обозначенные 174-D4-004 и 166-A4-002, имеют наилучшую аффинность связывания с MCP-1 (как оптические изомеры с IC50=2-5 нM, как было определено in vitro в анализе на выcвобождение Ca++ в клеточной культуре), и поэтому они могут иметь оптимальную последовательность и оптимальную комбинацию элементов последовательности B1A, B2 и B1B.
Кроме того, было идентифицировано 29 других МСР-1-связывающих нуклеиновых кислот, которые не могут быть описаны как комбинация элементов нуклеотидной последовательности, представленная МСР-1-связывающими нуклеиновыми кислотами типов 1-4. Эти последовательности перечислены на фиг.7.
Следует отметить, что любая из последовательностей, проиллюстрированных на фиг.1-7, представляет собой нуклеиновые кислоты согласно изобретению, включая их усеченные формы, а также их удлиненные формы, при условии, что такие усеченные и удлиненные молекулы нуклеиновой кислоты соответственно еще обладают способностью связываться с мишенью.
Пример 2
Нуклеиновые кислоты, которые связываются с мышиным MCP-1
С использованием биотинилированного мышиного D-MCP-1 в качестве мишени может быть получено несколько молекул нуклеиновой кислоты, связывающихся с D-MCP-1. Результат анализа последовательностей этих молекул нуклеиновой кислоты представлен на фиг.8.
Нуклеиновые кислоты были охарактеризованы на уровне аптамеров с помощью анализа на ингибирование с использованием биотинилированного мышиного D-MCP-1 и классифицированы по характеру связывания (пример 4). Отобранные последовательности синтезировали в виде оптического изомера (пример 3) и тестировали с использованием природной конфигурации MCP-1 (L-MCP) в анализе in vitro на высвобождение Ca++ в клеточной культуре (пример 5) или в анализе на хемотаксис (пример 6).
Как показано на фиг.8, D-188-A3-001 и D-189-G7-001 и их производные связываются с D-MCP-1 с субнаномолярным КD, как было определено в анализе на ингибирование (фиг.8).
Для молекулы D-mNOX-E36 (= D-188-A3-007; SEQ ID NО:244) была определена константа диссоциации (KD), составляющая 0,1-0,2 нM при 37°C (пример 4; фиг.10). Соответствующий оптический изомер mNOX-E36 (188-A3-007; SEQ ID NО:122) обнаруживает ингибирующую концентрацию (IC50), составляющую приблизительно 12 нM в анализе in vitro на высвобождение Ca++ (пример 5; фиг.13) и приблизительно 7 нM в анализе in vitro на хемотаксис (пример 6; фиг.16). Для ПЭГилированных производных mNOX-E36, mNOX-E36-3'ПЭГ (SEQ ID NО:254) IC50, определенная в анализе на высвобождение Ca++, составляет приблизительно 8 нM (пример 5, фиг.29), а в анализе на хемотаксис она составляет приблизительно 3 нМ (пример 6; фиг.31).
Следует отметить, что любая из последовательностей, проиллюстрированных на фиг.1-7, представляет собой нуклеиновые кислоты согласно изобретению, включая их усеченные формы, а также их удлиненные формы, при условии, что такие усеченные и удлиненные молекулы нуклеиновой кислоты соответственно еще обладают способностью связываться с мишенью.
Пример 3
Синтез и дериватизация аптамеров и оптических изомеров
Лабораторный синтез
Аптамеры и оптические изомеры получали путем твердофазного синтеза на синтезаторе ABI 394 (Applied Biosystems, Foster City, CA, USA) с применением метода фосфорамидитного химического синтеза 2'TBDMS-РНК (M.J. Damha, K.K. Ogilvie, Methods in Molecular Biology, Vol.20 Protocols for oligonucleotides and analogs, ed. S. Agrawal, p.81-114, Humana Press Inc. 1993). rA(N-Bz)-, rC(Ac)-, rG(N-ibu)- и rU-фосфорамидиты в D- и L-конфигурации закупали у компании ChemGenes, Wilmington, MA. Аптамеры и оптические изомеры очищали с помощью гель-электрофореза.
Крупномасштабный синтез с модификацией
Оптический изомер NOX-E36 получали путем твердофазного синтеза на синтезаторе ÄktaPilot100 (Amersham Biosciences; General Electric Healthcare, Freiburg) с применением фосфорамидитного способа химического синтеза 2'TBDMS-РНК (M.J. Damha, K.K. Ogilvie, Methods in Molecular Biology, Vol.20 Protocols for oligonucleotides and analogs, ed. S. Agrawal, p.81-114, Humana Press Inc. 1993). L-rA(N-Bz)-, L-rC(Ac)-, L-rG(N-ibu)- и L-rU-фосфорамидиты закупали у компании ChemGenes, Wilmington, MA. 5'-амино-модификатор был закуплен у American International Chemicals Inc. (Framingham, MA, USA). Синтез немодифицированного оптического изомера начинали на L-riboG-модифицированном стекле с регулируемым размером пор (CPG) 1000 Å (Link Technology, Glasgow, UK); при этом для получения 3'-NH2-модифицированного оптического изомера использовали 3'-амино-модификатор-CPG, 1000 Å (ChemGenes, Wilmington, MA). Для связывания (15 минут на цикл) использовали 0,3 M бензилтиотетразола (CMS-Chemicals, Abingdon, UK) в ацетонитриле и 3,5 эквивалентов соответствующего 0,1 M фосфорамидитного раствора в ацетонитриле. Был проведен цикл окисления-кэпирования. Дополнительные стандартные растворители и реагенты для олигонуклеотидного синтеза закупали у компании Biosolve (Valkenswaard, NL). Оптический изомер представлял собой синтезированный DMT-ON, и после снятия защиты он был очищен с помощью препаративной ОФ-ВЭЖХ (Wincott F. et al. (1995) Nucleic Acids Res 23:2677) с использованием среды Source 15RPC (Amersham). 5'DMT-группу удаляли путем добавления 80% уксусной кислоты (30 мин при комнатной температуре). Затем добавляли водный 2 M раствор NaOAc и оптический изомер обессоливали путем фильтрации в тангенциальном потоке с использованием регенерированной целлюлозной мембраны 5 K (Millipore, Bedford, MA).
ПЭГилирование NOX-E36
Для увеличения времени присутствия оптического изомера в плазме in vivo оптический изомер NOX-E36 ковалентно связывали с молекулой полиэтиленгликоля (ПЭГ) в 40 кДa у 3'-конца или у 5'-конца.
3'-ПЭГилирование NOX-E36
Для ПЭГилирования (подробное техническое описание метода ПЭГилирования можно найти в заявке на Европейский патент ЕP 1306382) очищенный 3'амино-модифицированный оптический изомер растворяли в смеси H2O (2,5 мл), ДМФ (5 мл) и буфера A (5 мл; полученного путем смешивания лимонной кислоты • H2O [7 г], борной кислоты [3,54 г], фосфорной кислоты [2,26 мл] и 1 M NaOH [343 мл] и добавления H2O до конечного объема 1 л; pH был доведен до значения 8,4 путем добавления 1 M HCl).
pH раствора оптического изомера доводили до значения 8,4 путем добавления 1 M NaOH. Затем через каждые 30 минут добавляли четыре порции 0,6 эквивалентов сложного ПЭГ-NHS-эфира в 40 кДа (Nektar Therapeutics, Huntsville, AL) при 37°C до достижения максимального выхода 75-85%. В процессе добавления сложного ПЭГ-NHS-эфира pH реакционной смеси поддерживали при 8-8,5 с использованием 1 M NaOH.
Реакционную смесь смешивали с 4 мл раствора мочевины (8 М), 4 мл буфера А и 4 мл буфера В (0,1 M ацетата триэтиламмония в H2O) и нагревали до 95°C в течение 15 минут. Затем ПЭГилированный оптический изомер очищали с помощью ОФ-ВЭЖХ с использованием среды Source 15RPC (Amersham) в градиенте ацетонитрила (буфер В; буфер С: 0,1 M ацетат триэтиламмония в ацетонитриле). Избыток ПЭГ элюировался при 5% буфера C, а ПЭГилированный оптический изомер элюировался при 10-15% буфера С. Фракции продукта с чистотой >95% (как было оценено с помощью ВЭЖХ) объединяли и смешивали с 40 мл 3М NaOAC. ПЭГилированный оптический изомер обессоливали путем фильтрации в тангенциальном потоке (на регенерированной целлюлозной мембране 5К, Millipore, Bedford MA).
5'-ПЭГилирование NOX-E36
Для ПЭГилирования (подробное техническое описание метода ПЭГилирования можно найти в заявке на Европейский патент ЕP 1306382) очищенный 5'амино-модифицированный оптический изомер растворяли в смеси H2O (2,5 мл), ДМФ (5 мл) и буфера A (5 мл; полученного путем смешивания лимонной кислоты • H2O [7 г], борной кислоты [3,54 г], фосфорной кислоты [2,26 мл] и 1 M NaOH [343 мл] и добавления воды до конечного объема 1 л; pH был доведен до значения 8,4 путем добавления 1 M HCl).
pH раствора оптического изомера доводили до значения 8,4 путем добавления 1 M NaOH. Затем через каждые 30 минут добавляли шесть порций 0,25 эквивалентов сложного ПЭГ-NHS-эфира в 40 кДа (Nektar Therapeutics, Huntsville, AL) при 37°C до достижения максимального выхода 75-85%. В процессе добавления сложного ПЭГ-NHS-эфира pH реакционной смеси поддерживали при 8-8,5 с использованием 1 M NaOH.
Реакционную смесь смешивали с 4 мл раствора мочевины (8 М), 4 мл буфера А и 4 мл буфера В (0,1 M ацетата триэтиламмония в H2O) и нагревали до 95°C в течение 15 минут. Затем ПЭГилированный оптический изомер очищали с помощью ОФ-ВЭЖХ с использованием среды Source 15RPC (Amersham) в градиенте ацетонитрила (буфер В; буфер С: 0,1 M ацетат триэтиламмония в ацетонитриле). Избыток ПЭГ элюировался при 5% буфера C, а ПЭГилированный оптический изомер элюировался при 10-15% буфера С. Фракции продукта с чистотой >95% (как было оценено с помощью ВЭЖХ) объединяли и смешивали с 40 мл 3 М NaOAC. ПЭГилированный оптический изомер обессоливали путем фильтрации в тангенциальном потоке (на регенерированной целлюлозной мембране 5К, Millipore, Bedford MA).
Пример 4
Определение констант связывания (Анализ на ингибирование)
Прямой анализ на ингибирование
Аффинность связывания аптамеров с D-MCP-1 измеряли в формате анализа на ингибирование при 20 или 37°C соответственно. Аптамеры представляют собой 5'-фосфат, меченный полинуклеотид-киназой T4 (Invitrogen, Karlsruhe, Germany) в присутствии [γ-32P]-меченой АТФ (Hartmann Analytic, Braunschweig, Germany). Удельная радиоактивность меченых аптамеров составляла 200000-800000 имп.мин/пмоль. Аптамеры инкубировали после денатурации и ренатурации при концентрации 20 пМ при 37°C в буфере для отбора (20 мM трис-HCl, pH 7,4; 137 мM NaCl; 5 мM KCl; 1 мM MgCl2; 1 мM CaCl2; 0,1% [масс./об.] твина-20) вместе с различными количествами биотинилированного D-MCP-1 в течение 4-12 часов для достижения равновесия при низких концентрациях. Для предотвращения адсорбции партнеров по связыванию на поверхностях используемой пластиковой лабораторной посуды или иммобилизующей матрицы к буферу для отбора добавляли 10 мкг/мл альбумина человеческой сыворотки (Sigma-Aldrich, Steinheim, Germany) и 10 мкг/мл дрожжевой РНК (Ambion, Austin, USA). Интервал концентраций биотинилированного D-MCP-1 составлял от 8 пМ до 100 нM, а общий объем реакционной смеси составлял 1 мл. Пептид и комплекс “пептид-аптамер” иммобилизовали на 1,5-мкл частицах стрептавидина Ultralink Plus (Pierce Biotechnology, Rockford, USA), которые были предварительно уравновешены буфером для отбора и ресуспендированы в общем объеме 6 мкл. Частицы выдерживали в суспензии в течение 30 минут при соответствующей температуре в термомиксере. Излучение иммобилизованной радиоактивной метки количественно оценивали в сцинтилляционном счетчике после удаления супернатанта и соответствующей промывки. Затем строили кривую зависимости процента связывания от концентрации биотинилированного D-MCP-1, а константы диссоциации вычисляли с помощью компьютерных алгоритмов (GRAFIT; Erithacus Software; Surrey U.K.) при допустимом стехиометрическом отношении 1:1.
Анализ на конкурентное ингибирование
Для сравнения различных D-MCP-1-связывающих аптамеров осуществляли ранговый анализ на конкурентное связывание аптамеров. Для этой цели аптамер с наилучшей аффинностью подвергали радиоактивному мечению (см. выше) и использовали в качестве эталона. После денатурации и ренатурации его инкубировали при 37°C с биотинилированным D-MCP-1 в 1 мл селективного буфера в условиях, способствующих приблизительно 5-10% неконкурентному связыванию с пептидом после иммобилизации и промывки на агарозе NeutrAvidin или на стрептавидине Ultralink Plus (оба реагента были закуплены у Pierce). Избыток денатурированных и ренатурированных вариантов немеченого D-РНК-аптамера добавляли к различным концентрациям (например, 2, 10 и 50 нМ) меченого эталонного аптамера для проведения параллельных реакций связывания. Тестируемые аптамеры конкурировали с эталонным аптамером за связывание с мишенью, что приводило к уменьшению сигнала связывания в зависимости от их параметров связывания. Затем аптамер, который обнаруживал наибольшую активность в данном анализе, служил в качестве нового эталона для сравнительного анализа других аптамерных вариантов.
Пример 5
Определение ингибирующей концентрации в анализе на высвобождение Ca ++
Клетки THP-1 (DSMZ, Braunschweig) культивировали в течение ночи при плотности 0,3×106 клеток/мл при 37°C и 5% CO2 в среде RPMI 1640 с реагентом GlutaMAX (Invitrogen), которая дополнительно содержала 10% фетальную телячью сыворотку, 50 единиц/мл пенициллина, 50 мкг/мл стрептомицина и 50 мкM β-меркаптоэтанола.
Оптические изомеры инкубировали с рекомбинантным человеческим MCP-1 (Bachem) в сбалансированном солевом растворе Хэнкса (HBSS), содержащем 1 мг/мл альбумина бычьей сыворотки, 5 мM пробенецида и 20 мM HEPES (HBSS+), в течение 15-60 минут при 37°C в планшете с 96 глубокими лунками малого профиля емкостью 0,2 мл («стимулирующий раствор»).
Для загрузки красителя-индикатора кальция клетки центрифугировали при 300×g в течение 5 минут, ресуспендировали в 4 мл раствора красителя-индикатора (10 мкM fluo-4 [Molecular Probes], 0,08% плюроника 127 [Molecular Probes] в HBSS+) и инкубировали в течение 60 минут при 37°C. Затем добавляли 11 мл HBSS+ и клетки центрифугировали, как описано выше, один раз промывали 15 мл HBSS+, а затем ресуспендировали в HBSS+ до достижения клеточной плотности 1,1×106/мл. 90 мкл полученной клеточной суспензии добавляли в каждую лунку черного 96-луночного планшета.
Измерение флуоресцентных сигналов проводили при возбуждении на длине волны 485 нм и излучении на длине волны 520 нм в ридере для считывания для многолуночных планшетов Fluostar Optima (BMG). Для параллельного измерения нескольких образцов лунки одного (перпендикулярного) ряда 96-луночного планшета регистрировали одновременно. Для определения базисной линии первые три считывания проводили со временем запаздывания 4 секунды. Затем считывание прерывали и планшет вынимали из аппарата. С использованием многоканальной пипетки 10 мкл стимулирующего раствора добавляли в лунки, а затем планшет снова помещали в ридер и продолжали измерение. В целом было проведено 20 измерений с интервалами времени в 4 секунды.
Для каждой лунки определяли разность между максимальной флуоресценцией и базовым значением, после чего строили график зависимости от концентрации MCP-1, или в экспериментах по ингибированию высвобождения кальция оптическими изомерами строили график зависимости от концентрации оптического изомера.
Определение 50% эффективной концентрации (EC 50 ) для человеческого MCP-1
После стимуляции клеток THP-1 различными концентрациями hMCP-1 и построения графика разности между максимальными и фоновыми сигналами получали кривую “доза-ответ” для человеческого MCP-1, которая указывала на то, что 50% эффективная концентрация (EC50) составляла примерно 2-4 нМ (фиг.11). Данную концентрацию использовали в последующих экспериментах по ингибированию высвобождения Ca++ под действием оптических изомеров.
Определение 50% эффективной концентрация (EC 50 ) для мышиного MCP-1
После стимуляции клеток THP-1 различными концентрациями mMCP-1 и построения графика разности между максимальными и фоновыми сигналами получали кривую “доза-ответ” для мышиного MCP-1, которая указывала на то, что 50% эффективная концентрация (EC50) составляла примерно 5 нМ (фиг.28). Данную концентрацию использовали в последующих экспериментах по ингибированию высвобождения Ca++ под действием оптических изомеров.
Пример 6
Определение ингибирующей концентрации в анализе на хемотаксис
Клетки THP-1, культивированные, как описано выше, центрифугировали, один раз промывали в HBH (HBSS, содержащем 1 мг/мл альбумина бычьей сыворотки и 20 мM HEPES) и ресуспендировали при плотности 3×106 клеток/мл. 100 мкл этой суспензии наносили на вставочную пластину Transwell с размером пор 5 мкм (Corning, #3421). В нижних отсеках MCP-1 предварительно инкубировали вместе с оптическими изомерами в различных концентрациях в 600 мкл HBH при 37°C в течение 20-30 минут, а затем добавляли клетки. Клетки оставляли на 3 часа при 37°C для миграции. Затем вставочные пластины удаляли и в нижние отсеки добавляли 60 мкл 440 мкM резазурина (Sigma) в забуференном фосфатом физиологическом растворе. После инкубирования при 37°C в течение 2,5 часов интенсивность флуоресценции измеряли при возбуждении на длине волны 544 нм и излучении на длине волны 590 нм в планшет-ридере для многолуночных планшетов Fluostar Optima (BMG).
Определение 50% эффективной концентрации (EC 50 ) для человеческого MCP-1
После 3-часовой миграции клеток THP-1 в направлении различных концентраций человеческого MCP-1 строили кривую “доза-ответ” для человеческого MCP-1, по которой было определено, что максимальная эффективная концентрация составляет примерно 1 нМ, и которая указывала на снижение активации при более высоких концентрациях (фиг.14). Для дальнейших экспериментов по ингибированию хемотаксиса оптическими изомерами использовали MCP-1 при концентрации 0,5 нМ.
Определение 50% эффективной концентрация (EC 50 ) для мышиного MCP-1
После 3-часовой миграции клеток THP-1 в направлении различных концентраций мышиного MCP-1 строили кривую “доза-ответ” для мышиного MCP-1, по которой было определено, что максимальная эффективная концентрация составляет примерно 1-3 нМ, и которая указывала на снижение активации при более высоких концентрациях (фиг.30). Для дальнейших экспериментов по ингибированию хемотаксиса оптическими изомерами использовали мышиный MCP-1 при концентрации 0,5 нМ.
Пример 7
Анализ на связывание, проводимый с помощью поверхностного плазмонного резонанса
7.1 Оценка специфичности NOX-E36, 181-A2-018 и mNOX-E36
Для анализа на связывание нуклеиновых кислот с человеческим MCP-1 и родственными белками использовали устройство Biacore 2000 (Biacore AB, Uppsala, Sweden). После связывания посредством амино-групп белки диализовали против воды в течение 1-2 час (с использованием смешанных сложных эфиров целлюлозы Millipore VSWP с размером пор 0,025 мкм) для удаления нежелательных аминов. Перед связыванием с белком сенсорные чипы PioneerF1 или CM4 (Biacore AB) активировали путем введения 35 мкл 1:1-разведения 0,4 M NHS и 0,1 M EDC при скорости потока 5 мкл/мин. Затем вводили хемокин в концентрациях 0,1-1,5 мкг/мл при скорости потока 2 мкл/мин до тех пор, пока чувствительность прибора не достигала значений в пределах 1000-2000 RU (относительных единиц). Непрореагировавшие сложные NHS-эфиры дезактивировали путем введения 35 мкл раствора гидрохлорида этаноламина (pH 8,5) при скорости потока 5 мкл/мин. Сенсорный чип премировали два раза связывающим буфером и уравновешивали при 10 мкл/мин в течение 1-2 часов до тех пор, пока не были достигнуты стабильные базовые значения. Для всех белков кинетические параметры и константы диссоциации оценивали путем введения серии инъекций оптических изомеров в концентрациях 1000, 500, 250, 125, 62,5, 31,25 и 0 нМ в селективном буфере (Трис-HCl, 20 мМ; NaCl, 137 мМ; KCl, 5 мМ; CaCl2, 1 мМ; MgCl2, 1 мМ; Твин 20, 0,1% [масс./об.]; pH 7,4). Во всех экспериментах данный анализ осуществляли при 37°C с использованием команды Kinject, с помощью которой было определено время ассоциации 180 секунд и время диссоциации 360 секунд при скорости потока 10 мкл/мин. Анализ данных и вычисление констант диссоциации (КD) проводили с помощью компьютерной программы BIAevaluation 3.0 (BIACORE AB, Uppsala, Sweden) с использованием алгоритма построения стехиометрической лангмюровской модели 1:1.
7.1.1 NOX-E36 и 181-A2-018 (нуклеиновые кислоты, специфичные к человеческому MCP-1)
Все сенсорограммы проиллюстрированы только для человеческого MCP-1 (фиг.17 и 20 соответственно); а для других белков сенсорограммы, построенные с использованием оптического изомера в концентрации 125 нМ, приводятся лишь в целях иллюстрации (фиг.18/19 и 21/22).
Анализ на взаимодействие NOX-E36·hMCP-1: рекомбинантный человеческий MCP-1 иммобилизовали на сенсорном чипе PioneerF1 в соответствии с рекомендациями производителя (путем проведения процедуры связывания с амином) до тех пор, пока чувствительность прибора не достигала значения 1381 RU (относительных единиц). Полученная константа диссоциации (КD) для связывания NOX-E36 с человеческим MCP-1 составляла приблизительно 890 пМ (фиг.17).
Анализ на взаимодействие 181-A2-018·hMCP-1: рекомбинантный человеческий MCP-1 иммобилизовали на сенсорном чипе СМ4 в соответствии с рекомендациями производителя (путем проведения процедуры связывания с амином) до тех пор, пока чувствительность прибора не достигала значения 3111 RU (относительных единиц). Полученная константа диссоциации (КD) для связывания 181-A2-018 с человеческим MCP-1 составляла приблизительно 370 пМ (фиг.20).
Для определения специфичности NOX-E36 и 181-A2-018 различные человеческие белки семейства MCP-1, а также человеческий эотаксин иммобилизовали на сенсорном чипе PioneerF1 и CM4 (hMCP-1, 1754 RU; hMCP-2, 1558 RU; hMCP-3, 1290 RU; эотаксин, 1523 RU). Кинетический анализ показал, что NOX-E36 связывался с эотаксином и с hMCP-2 с константами диссоциации (KD) 5-10 нМ, но не распознавал hMCP-3 (фиг.18 и 24A). В противоположность этому, 181-A2-018 связывался с эотаксином, hMCP-2 и hMCP-3, но с несколько более низкой аффинностью (10-20 нМ; фиг.21 и 24A).
Межвидовую перекрестную реактивность NOX-E36 и 181-A2-018 оценивали на сенсорном чипе PioneerF1 и СМ4 с использованием человеческого МСР-1 (1460 RU), обезьяньего МСР-1 (1218 RU), свиного МСР-1 (1428 RU), собачьего МСР-1 (1224 RU), кроличьего МСР-1 (1244 RU), крысиного МСР-1 (1267 RU) и мышиного МСР-1 (1361 RU), иммобилизованных в соответствии с процедурой связывания с амином. Кинетический анализ показал, что NOX-E36 связывался с человеческим, обезьяньим, свиным и собачьим MCP-1 со сравнимыми константами диссоциации (КD), составляющими 0,89-1,2 нМ, но не распознавал мышиный, крысиный и кроличий MCP-1 (фиг.19 и 24A). 181-A2-018 связывался с человеческим и обезьяньим MCP-1 со сравнимыми константами диссоциации (КD) 0,5-0,6 нМ, а со свиным, кроличьим и собачьим MCP-1 он связывался с гораздо более низкой аффинностью. Крысиный и мышиный MCP-1 не распознавались оптическим изомером NOX-A2-018 (фиг.22 и 24A).
Последовательности, а также степень гомологии (в процентах) идентичных аминокислот белков MCP1 различных видов и близкородственных человеческих белков представлены на фиг.23, а вычисленные величины КD для NOX-E36 и 181-A2-018 представлены в виде таблицы на фиг.24A.
7.1.2 mNOX-E36 (нуклеиновая кислота, специфичная к мышиному MCP-1)
Для анализа характера связывания mNOX-E36 3759 RU синтетического биотинилированного мышиного D-MCP-1 (проточная кювета 3) и 3326 RU биотинилированного человеческого D-MCP-1 (проточная кювета 4) иммобилизовали на сенсорном чипе, конъюгированном со стрептавидином (Biacore AB, Freiburg, Germany) соответственно. Растворы аптамера mNOX-E36 (D-РНК) в концентрациях 500, 250, 125, 62,5, 31,25 и 0 нМ были введены с использованием команды Kinject, с помощью которой было определено время ассоциации 180 секунд и время диссоциации 360 секунд. Проточную кювету 1 использовали в качестве контроля (буфер и декстрановая матрица) (поверхность SA-чипа Biacore), а на проточной кювете 2 был иммобилизован неспецифический D-пептид для определения неспецифического связывания аптамера. На фиг.32 представлена сенсорограмма кинетики связывания D-NOX-E36 с мышиным D-MCP-1 с вычисленной константой диссоциации (КD), составляющей 200-300 пМ. mNOX-E36 не связывался с человеческим D-MCP-1 (фиг.33), но для ясности на этой фигуре представлена только сенсорограмма, полученная с использованием 125 нМ оптического изомера.
7.2 Оценка селективности NOX-E36
Селективность NOX-E36 оценивали методом поверхностного плазмонного резонанса путем иммобилизации 5'-биотинилированного NOX-E36 на стрептавидине (на SA-чипе). 352 RU оптического изомера NOX-E36 на проточной кювете (FC) 1 и такое же количество 5'-концевого биотинилированного нефункционального контрольного оптического изомера (POC) на FC 2 иммобилизовали посредством связывания со стрептавидином/биотином. FC3 использовали в качестве контроля для определения неспецифического связывания с декстрановой поверхностью сенсорного SA-чипа.
100 нМ панели человеческих хемокинов всех четырех подгрупп (CC, CXC, CX3C и XC) вводили в течение 360 секунд, и комплексы оставляли на 360 секунд для диссоциации при скорости потока 10 мкл/мин и при 37°C. После ассоциации (ответ 1; степень взаимодействия) и после диссоциации (ответ 2, аффинность взаимодействия) строили график единиц отклика. После каждого введения поверхность чипа регенерировали 1 М хлоридом натрия с 0,1% твином в течение 240 сек, а затем иммобилизованные оптические изомеры подвергали рефолдингу в течение 2 минут при физиологических условиях (рабочий буфер). Введение каждого хемокина повторяли 3 раза. CXCL1, CXCL2, CXCL6 и CXCL9 обнаруживали неспецифическое связывание с рибонуклеиновыми кислотами и с декстрановой поверхностью чипа. Специфическое высокоаффинное связывание с иммобилизованным NOX-E36 можно было детектировать только для CCL2/MCP1, CCL8/MCP2, CCL11/эотаксина, CCL3/MIP1α и CXCL7/NAP-2 (фиг.24B). Обнаружение того, что MCP-2 и эотаксин связываются с NOX-E36, не является неожиданным и объясняется относительно высокой гомологией между этими хемокинами, а именно между неожиданно позитивными CCL3/MIP-1α и CXCL7/NAP-2 и MCP-1, которая составляет 62 и 70%, как показали тесты in vitro на функциональное ингибирование, проводимые в соответствии с настоящим изобретением или известные специалистам соответственно.
И, наконец, кинетические параметры взаимодействия между NOX-E36 и CCL2/MCP1, CCL8/MCP2, CCL11/эотаксином, CCL3/MIP1α, CXCL7/NAP-2, CCL7/MCP-3 и CCL13/MCP-4 были определены в «инвертированной» системе. В данном случае хемокины были иммобилизованы и был введен свободный NOX-E36 (подробное описание протокола см. 7.1). Кинетические данные систематизированы на фиг.24C.
7.3 Оценка функциональности антагониста MIP-1α in vitro
Измерения методом Biacore указывали на перекрестную реактивность NOX-E36 и MIP-1α. С помощью функционального анализа in vitro, проводимого в клеточной культуре, необходимо проверить, может ли Biacore-связывание NOX-E36 с MIP-1α также коррелировать с функциональными свойствами, например с антагонизмом.
Для этих целей проводили эксперименты по хемотаксису на клетках THP-1, которые могут быть стимулированы MIP-1α. Клетки THP-1, культивированные, как описано выше, центрифугировали, один раз промывали в HBH (в HBSS, содержащем 1 мг/мл альбумина бычьей сыворотки и 20 мМ HEPES) и ресуспендировали при 3×106 клеток/мл. 100 мкл этой суспензии наносили на вставочные пластины Transwell с размером пор 5 мкм (Corning, #3421). В нижних отсеках MCP-1 предварительно инкубировали вместе с оптическими изомерами в различных концентрациях в 600 мкл HBH при 37°C в течение 20-30 минут, а затем добавляли клетки. Клетки оставляли на 3 часа при 37°C для миграции. Затем вставочные пластины удаляли и в нижние отсеки добавляли 60 мкл 440 мкM резазурина (Sigma) в забуференном фосфатом физиологическом растворе. После инкубирования при 37°C в течение 2,5 часов интенсивность флуоресценции измеряли при возбуждении на длине волны 544 нм и излучении на длине волны 590 нм в ридере для считывания многолуночных планшетов Fluostar Optima (BMG).
После 3-часовой миграции клеток THP-1 по направлению к различным концентрациям человеческого MIP-1α строили кривую «доза-ответ» для человеческого MIP-1α, которая указывала на то, что 50% эффективная концентрация (EC50) составляла примерно 1 нМ и на снижение активации при более высоких концентрациях (фиг.24D). В последующих экспериментах по ингибированию хемотаксиса оптическими изомерами использовали концентрацию MIP-1α=0,5 нМ.
Эксперименты по определению ингибирования хемотаксиса под действием NOX-E36 осуществляли с использованием 0,5 нМ MIP-1α в качестве стимулятора. Совершенно очевидно, что NOX-E36 не ингибировал MIP-1α-индуцированный хемотаксис при наивысшей тестируемой концентрации MIP-1α, равной 1 мкМ. В качестве позитивного контроля параллельно осуществляли соответствующий эксперимент с использованием MCP-1 в качестве стимулятора (фиг.24E).
Пример 8
Лечение заболевания типа волчанки у MRL lpr/lpr -мышей с использованием оптического изомера-антагониста mMCP-1
Блокирование провоспалительных медиаторов представляет собой эффективный способ лечения хронического воспаления (Steinman 2004). CC-хемокины, помимо TNF и интерлейкинов, являются важными кандидатами на специфические антагонисты, поскольку CC-хемокины опосредуют рекрутинг лейкоцитов из внутрисосудистого пространства на участки воспаления (Baggiolini 1998, Luster 2005). Этот факт является очень важным аргументом в пользу того, что MCP-1 (=CCL2) и его соответствующий хемокиновый рецептор CCR2 играют решающую роль в аутоиммунном поражении ткани, таком как клинические манифестации системной красной волчанки (Gerard & Rollins 2001). Так, например, MRLlpr/lpr-мыши, дефицитные по гену Ccl2 или по гену Ccr2, были резистентными к аутоиммунному ответу волчаночного типа (Perez de Lema 2005, Tesch 1999). Следовательно, система CCL2/CCR2 может представлять собой перспективную терапевтическую мишень, например, для лечения волчаночного нефрита. Действительно, пролонгированная генотерапия или перенос трансфецированных клеток, которые приводят к in situ продуцированию NH2-усеченной MCP-1, позволяют значительно снизить аутоиммунное повреждение тканей у MRLlpr/lpr-мышей. Однако такие экспериментальные подходы не могут быть применены для лечения человека из-за неконтролируемого продуцирования данного антагониста и образования опухолей (Hasegawa 2003, Shimizu 2004). Поэтому необходимо получить новые антагонисты CCL2 с благоприятным фармакокинетическим профилем in vivo. В этом примере показано, что блокирование мышиного CCL2 оптическими изомерами mNOX-E36 или mNOX-E36-3'ПЭГ, направленными против mCCL2, может быть использовано для лечения волчаночного нефрита и устранения манифестаций другого заболевания, например системной красной волчанки. Терапия с использованием оптического изомера против mCCL2 замедленного действия является эффективной для лечения волчаночного нефрита, аутоиммунного перибронхита и кожной болезни волчаночного типа у MRLlpr/lpr-мышей и не связана с какими-либо вышеуказанными проблемами, возникающими при терапевтической блокаде CCL2/CCR2.
Животные и экспериментальный протокол
10-недельные самки MRLlpr/lpr-мышей были закуплены у компании Harlan Winkelmann (Borchen, Germany) и содержались в нормальных условиях с 12-часовым циклом смены дня и ночи. Воду и стандартный корм (Ssniff, Soest, Germany) давали ad libitum. В возрасте 14 недель группам из 12 мышей три раза в неделю подкожно вводили инъекции следующих оптических изомеров в 5% глюкозе (объем инъекции 4 мл/кг): mNOX-E36, 1,5 мкмоль/кг; mNOX-E36-3'ПЭГ, 0,9 мкмоль/кг; нефункционального контрольного оптического изомера PoC (5'-UAAGGAAACUCGGUCUGAUGCGGU AGCGCUGUGCAGAGCU-3'), 1,9 мкмоль/кг; PoC-ПЭГ, 0,9 мкмоль/кг и носителя (5% глюкозы). Уровни mNOX-E36 и mNOX-E36-3'ПЭГ в плазме определяли в пробах крови, которые брали еженедельно из ретроорбитального синуса через 3 или 24 часа после инъекции соответственно. Уровни оптического изомера в пробах плазмы определяли модифицированным методом «сэндвич»-гибридизации, как описано в примере 8. Мышей умерщвляли путем смещения шейных позвонков по достижении ими возраста 24 недель.
Оценка системной красной волчанки
Кожные поражения регистрировали методом полуколичественной оценки (Schwarting 2005). Массовое отношение селезенки и массы мезентериальных лимфоузлов к общей массе тела вычисляли как показатели ассоциированного с волчанкой лимфопролиферативного синдрома. По окончании исследований у каждого животного брали пробы крови и мочи, причем кровь брали из ретроорбитального венозного сплетения под общей анестезией эфиром. По окончании исследования у каждого животного брали пробы крови и мочи, вычисляли отношение альбумина/креатинина в моче и определяли титры сывороточных анти-дцДНК аутоантител изотипа IgG, как описано ранее (Pawar 2006). Скорость клубочковой фильтрации (GFR) определяли в течение 24 недель по кинетике клиренса ФИТЦ-инулина из плазмы (Sigma-Aldrich, Steinheim, Germany) через 5, 10, 15, 20, 35, 60 и 90 минут после одной инъекции ударной дозы (Qi 2004). Интенсивность флуоресценции определяли при возбуждении на длине волны 485 нм и считывали при излучении на длине волны 535 нм. GFR вычисляли, исходя из двухкомпонентной модели, с использованием компьютерной программы построения нелинейной кривой регрессии (GraphPad Prism, GraphPad Software Inc., San Diego, CA). Уровни цитокинов в сыворотке определяли с использованием коммерчески доступных наборов для ELISA-анализа на IL-6, IL-12p40 (OptEiA, BD Pharmingen) и IFN-α (PBL Biomedical Labs, USA). Почки и легкие от всех мышей фиксировали в 10%-ном забуференном формалине, обрабатывали и заливали в парафин. 5 мкм-срезы для окрашивания серебром и периодной шиффовой кислотой получали в соответствии со стандартными протоколами (Anders 2002). Степень тяжести поражения почек оценивали по индексам активности и по показателям, характерным для хронического состояния, как описано в литературе для человеческого волчаночного нефрита (Austin 1984), и проводили морфометрию интерстициального поражения почек, как было описано ранее (Anders 2002). Тяжесть перибронхиального воспаления оценивали полуколичественным методом по 0-4-балльной системе. Для осуществления иммунного окрашивания фиксированные в формалине и залитые в парафин срезы подвергали депарафинизации и регидратации. Эндогенную пероксидазу блокировали 3% гидроксидом водорода и разведение антигена осуществляли в растворе для разведения антигена (Vector, Burlingame, CA) в автоклаве. Биотин блокировали с использованием набора для блокирования авидина/биотина (Vector). Предметные стекла инкубировали с «первыми» антителами в течение одного часа, а затем с биотинилированными «вторыми» антителами (против крысиных IgG, Vector) и с реагентом ABC (Vector). В перерывах между стадиями инкубирования предметные стекла промывали в забуференном фосфатом физиологическом растворе. В качестве детектируемой системы использовали 3'3'-диаминобензидин (DAB, Sigma, Taufkirchen, Germany) с усиливающим металлическим реактивом, в результате чего получали продукт черного цвета. В качестве контрастного красителя использовали метиловый зеленый, и предметные стекла дегидратировали и помещали в аппарат Histomount (Zymed Laboratories, San Francisco, CA). В качестве «первых» антител использовали: крысиное анти-Mac2 антитело (макрофаги, Cederlane, Ontario, Canada, 1:50), антитело против мышиного CD3 (1:100, клон 500A2, BD), антитело против мышиного IgG1 (1:100, M32015, Caltag Laboratories, Burlingame, CA, USA), антитело против мышиного IgG2a (1:100, M32215, Caltag) и антитело против мышиного C3 (1:200, GAM/C3c/ФИТЦ, Nordic Immunological Laboratories, Tilburg, Netherlands). В качестве негативного контроля проводили инкубирование с антителом соответствующего изотипа. Для проведения количественного анализа гломерулярные клетки подсчитывали в 15 корковых клубочках на один срез. Гломерулярные Ig- и C3c-отложения оценивали на 15 срезах корковых клубочков по 0-3-балльной шкале.
Получение РНК и проведение количественной ОТ-ПЦР (TaqMan) в реальном времени
Почечную ткань каждой мыши подвергали быстрой заморозке в жидком азоте и хранили при -80°C. От каждого животного получали полноразмерную почечную РНК и осуществляли обратную транскрипцию, как описано в литературе (Anders 2002). Праймеры и зонды поставлялись компанией PE Biosystems, Weiterstadt, Germany. Указанные праймеры (300 нМ), используемые для детектирования рРНК Ccl2, Ccl5 и 18S, и предварительно полученный реагент для анализа TaqMan поставлялись фирмой PE Biosystems.
Проточная цитометрия
По окончании исследования от мышей всех групп получали образцы цельной крови и костного мозга. Проточную цитометрию осуществляли с применением аппарата FACScalibur и с использованием предварительно охарактеризованного анти-mCCR2 антитела MC21 (Mack 2001). Для детекции использовали биотинилированное антитело против крысиного IgG (BD Biosciences). В качестве антитела контрольного изотипа использовали крысиный IgG2b (BD Biosciences).
Статистический анализ
Данные выражали как среднее ± среднеквадратическая ошибка (SEM). Сравнение между группами осуществляли с использованием одномерного ANOVA. Для проведения множества сравнений использовали поправку Бонферрони рosthoc. Величина p<0,05 рассматривалась как показатель статистической значимости.
«Сэндвич»-анализ на гибридизацию
Количество оптического изомера в образцах оценивали с помощью «сэндвич»-анализа на гибридизацию, проводимого на основе анализа, описанного Drolet et al. 2000 (Pharm Res 17:1503). Пробы крови брали одновременно для мониторинга клиренса NOX-E36 из плазмы. Выбранные ткани приготавливали для определения концентраций оптического изомера.
Приготовление планшетов для гибридизации
Оптический изомер mNOX-E36 количественно оценивали с помощью невалидизированного «сэндвич»-анализа на гибридизацию. Вкратце, зонд для захвата mNOX-E36 (SEQ ID:281) иммобилизовали на белых 96-луночных планшетах для связывания с ДНК (Corning Costar, Wiesbaden, Germany) при концентрации 0,75 мМ в 0,5 M фосфате натрия, 1 мМ EDTA, pH 8,5, в течение ночи при 4°C. Лунки два раза промывали и блокировали 0,5% масс./об. BSA в 0,25 M фосфате натрия, 1 мМ EDTA, pH 8,5, в течение 3 часов при 37°C, а затем снова промывали и хранили при 4°C вплоть до их использования. Перед гибридизацией лунки предварительно нагревали до 37°C и два раза промывали предварительно нагретым промывочным буфером (3×SSC, 0,5% [масс./об.] додецилсаркозинат натрия, pH 7,0, причем сначала приготавливали 20x маточный раствор [3 M NaCl, 0,3 M Na3-цитрат] без лауроилсаркозината натрия и соответствующим образом разводили).
Получение образцов
Анализ всех образцов проводили в дубликатах. Пробы плазмы оттаивали на льду, интенсивно перемешивали и быстро центрифугировали в охлажденной настольной центрифуге. Тканевые гомогенаты оттаивали при комнатной температуре и центрифугировали в течение 5 минут при максимальной скорости и при комнатной температуре. Для проведения анализа брали только 5 мкл каждого образца, а затем их снова помещали в холодильник на хранение. Образцы разводили буфером для гибридизации (8 нМ зонда для детекции mNOX-E36 [SEQ ID NO:282] в промывочном буфере) при комнатной температуре в соответствии со следующей схемой:
Были проанализированы все разведения образцов. mNOX-E36-стандарт серийно разводили для построения по 8 точкам калибровочной кривой, охватывающей диапазон 0-4 нМ. Получение и анализ контрольных образцов не проводили. Калибровочный стандарт был идентичен значениям, полученным для тестируемых образцов.
Гибридизация и детекция
Образцы нагревали в течение 10 минут при 95°C и охлаждали до 37°C. Комплексы «оптический изомер/детектирующий зонд» подвергали гибридизации с иммобилизованными зондами для захвата в течение 30 минут при 37°C. Несвязанные оптические изомеры удаляли путем двухкратной промывки промывочным буфером и 1×TBST (20 мМ трис-Cl, 137 мМ NaCl, 0,1% твин 20, pH 7,5) соответственно. Гибридизованные комплексы детектировали с использованием конъюгата стрептавидина-щелочной фосфатазы, разведенного 1:5000 в 1×TBST, в течение 1 часа при комнатной температуре. Для удаления несвязанного конъюгата лунки снова промывали 1×TBST и 20 мМ трис-Cl, 1 мМ MgCl2, pH 9,8 (два раза каждым). И, наконец, лунки заполняли 100 мл субстрата CSDP (Applied Biosystems, Darmstadt, Germany) и инкубировали в течение 45 минут при комнатной температуре. Хемилюминесценцию измеряли на микропланшет-ридере FLUOstar Optima (BMG Labtechnologies, Offenburg, Germany).
Анализ данных
Для количественного анализа данных было использовано следующее разведение анализируемого образца:
крысиная плазма в EDTA 1:2000.
Данные, полученные от группы мышей, обработанных носителем (которым не вводили оптический изомер), вычитали как фоновый сигнал.
Описанный здесь «сэндвич»-анализ на гибридизацию аналогичным образом может быть применим для оптических изомеров NOX-36, NOX-E36-5'-ПЭГ и NOX-E36-3'-ПЭГ, при этом должны быть использованы соответствующий зонд для захвата NOX-E36 (SEQ ID:255) и соответствующий зонд для детекции NOX-E36 (SEQ ID:256) (данные не приводятся).
Результаты
mNOX-E36-3'ПЭГ повышают степень выживаемости и ослабляют тяжесть почечного заболевания у MRL lpr/lpr -мышей
У самок MRLlpr/lpr-мышей развивался пролиферативный и ассоциированный с иммунным комплексом гломерулонефрит, который имел очень высокую степень сходства с диффузным пролиферативным волчаночным нефритом у человека и приводил к гибели этих мышей. В этом протоколе терапевтического исследования обработанных MRLlpr/lpr-мышей в возрасте 14-24 недель обрабатывали ПЭГилированным и неПЭГилированным оптическим изомером, направленным против mCCL2, а также ПЭГилированным и неПЭГилированным контрольным ("PoC")-оптическим изомером или носителем. На этой стадии у MRLlpr/lpr-мышей, обработанных носителем, PoC или PoC-ПЭГ, наблюдался диффузный пролиферативный гломерулонефрит, характеризующийся инфильтрацией клубочковых макрофагов и инфильтрацией смешанных воспалительных клеток перигломерулярной и интерстициальной тканей, состоящих из гломерулярных и интерстициальных Mac2-позитивных макрофагов и интерстициальных CD3-позитивных лимфоцитов (фиг.34 и 35). mNOX-E36-3'ПЭГ снижал показатели активности и хронического характера течения волчаночного нефрита, а также вышеупомянутых маркеров воспаления почек (фиг.35). Не-ПЭГилированная молекула mNOX-E36 была менее эффективна в отношении хронического характера заболевания и количества интерстициальных макрофагов и Т-клеток (фиг.35). Прогрессирующее хроническое заболевание почек было также идентифицировано по атрофии канальцев и по конфлюэнтным участкам интерстициального фиброза у мышей, обработанных носителем, PoC- и PoC-ПЭГ (фиг.34). При применении морфометрии для количественной оценки этих изменений было обнаружено, что ПЭГилированный и неПЭГилированный mNOX-E36 снижал объем повреждения интерстициальной ткани, степень поражения клеток канальцев и уровень дилатации канальцев, то есть показатели тяжести и прогноза хронического заболевания почек (фиг.36). mNOX-E36-3'ПЭГ, но не неПЭГилированный mNOX-E36, снижал показатели 50%-ной смертности (фиг.37). Таким образом, mNOX-E36-3'ПЭГ способен снижать число инфильтратов почечных макрофагов и Т-клеток, улучшать динамику течения волчаночного нефрита и повышать выживаемость (при болезни почек) MRLlpr/lpr-мышей. Для того чтобы определить, оказывает ли лечение оптическими изомерами mNOX-E36 и mNOX-E36-3'ПЭГ благоприятное воздействие на течение внутрипочечного воспаления у MRLlpr/lpr-мышей, была проведена ОТ-ПЦР в реальном времени в целях оценки уровней экспрессии провоспалительных хемокинов CCL2 и CCL5, которые, как было показано ранее, постоянно стимулировались в почках MRLlpr/lpr-мышей в процессе прогрессирования почечного заболевания (Perez de Lema 2001). Обработка мышей в возрасте от 14 до 24 недель оптическими изомерами mNOX-E36 и mNOX-E36-3'ПЭГ приводила к снижению уровней экспрессии мРНК CCL2 и CCL5 в почках по сравнению с уровнем экспрессии, наблюдаемым у контрольных мышей, обработанных носителем (фиг.38).
Оптические изомеры, направленные против CCL2, снижают степень внепочечного аутоиммунного поражения ткани у MRL lpr/lpr -мышей
Обычно ткани кожи и легких у MRLlpr/lpr-мышей также подвержены аутоиммунному повреждению. У мышей, обработанных носителем, аутоиммунное заболевание легких характеризуется умеренной инфильтрацией перибронхиолярных и периваскулярных воспалительных клеток, и у 60% таких мышей наблюдается поражение кожи (фиг.39, 40 и 35). У мышей, обработанных обоими mNOX-E36 и mNOX-E36-3'ПЭГ, наблюдалось снижение степени перибронхиального воспаления и поражения кожи по сравнению с MRLlpr/lpr-мышами, обработанными носителем, PoC- и PoC-ПЭГ соответственно (фиг.39, 40 и 35). Следовательно, CCL2-специфические оптические изомеры не ограничиваются воздействием на волчаночный нефрит и также воздействуют на другие манифестации аутоиммунного поражения ткани у MRLlpr/lpr-мышей.
mNOX-E36 и лимфопролиферативный синдром, аутоантитела против дцДНК и уровни цитокинов в сыворотке у MRL lpr/lpr -мышей
У самок MRLlpr/lpr-мышей развивался лимфопролиферативный синдром, характеризующийся обширной спленомегалией и увеличением цервикальных, подмышечных, паховых и мезентериальных лимфоузлов. Ни mNOX-E36, ни mNOX-E36-3'ПЭГ не оказывали какого-либо воздействия на массу селезенки и лимфоузлов у MRLlpr/lpr-мышей (фиг.41). Аутоиммунный ответ у MRLlpr/lpr-мышей характеризуется продуцированием аутоантител против множества ядерных антигенов, включая дцДНК. В сыворотке 24-недельных MRLlpr/lpr-мышей присутствовали высокие уровни аутоантител IgG, IgG1, IgG2a, IgG2b против дцДНК. Ни один из mNOX-E36 и mNOX-E36-3'ПЭГ не оказывал какого-либо воздействия на эти аутоантитела против ДНК (фиг.41). Заболевание волчаночного типа у обработанных носителем MRLlpr/lpr-мышей характеризуется повышенными уровнями IFN-α, IL-12p40 и IL-6 в сыворотке. Ни один из mNOX-E36 и mNOX-E36-3'ПЭГ не оказывал какого-либо влияния на эти медиаторы воспаления (фиг.41). Таким образом, оба варианта mNOX-E36 не оказывали влияния на лимфопролиферацию, продуцирование анти-дцДНК IgG и на уровни цитокинов в сыворотке MRLlpr/lpr-мышей.
Уровни mNOX-E36 и mNOX-E36-3'ПЭГ в плазме у MRL lpr/lpr -мышей
Уровни mNOX-E36 и mNOX-E36-3'ПЭГ в плазме определяли еженедельно для мониторинга эффекта лекарственного средства в процессе развития прогрессирующего почечного заболевания у MRLlpr/lpr-мышей. Во время исследования медианные уровни mNOX-E36 в плазме через 3 часа после инъекции и mNOX-E36-3'ПЭГ через 24 часа после инъекции составляли приблизительно 300 нМ и 1 мкМ соответственно (фиг.42). Таким образом, ПЭГилирование приводило к увеличению уровней mNOX-E36 в плазме при прогрессирующем почечном заболевании у MRLlpr/lpr-мышей и не модулировало фармакокинетику обоих оптических изомеров.
mNOX-E36-3'ПЭГ блокирует миграцию моноцитов из костного мозга
Было обнаружено, что миграция моноцитов из костного мозга во время бактериальной инфекции приводит к стимуляции хемокинового рецептора CCR2 (Serbina 2006), однако роль CCL2 в вырабатывании аутоиммунного ответа остается гипотетической. Поэтому была проведена оценка популяции CCR2-позитивных моноцитов в периферической крови и в костном мозге 24-недельных MRLlpr/lpr-мышей, обработанных mNOX-E36-3'ПЭГ и носителем. Обработка оптическим изомером mNOX-E36-3'ПЭГ приводила к увеличению содержания CCR2-позитивных клеток в костном мозге от 13% до 26%, а в периферической крови эта популяция снижалась с 26% до 11% (фиг.43). Полученные данные подтверждают роль CCL2 в оттоке CCR2-позитивных клеток из костного мозга во время развития аутоиммунного заболевания у MRLlpr/lpr-мышей.
Выводы
С применением технологии оптических изомеров был получен новый и специфический антагонист mCCL2, который эффективно блокирует mCCL2 in vitro и in vivo. Действительно, лечение оптическим изомером замедленного действия, а именно изомером, направленным против CCL2, значительно снижает степень повреждения ткани при прогрессирующем аутоиммунном заболевании волчаночного типа у MRLlpr/lpr-мышей. Эти данные подтверждают центральную роль CCL2 в развитии хронического воспалительного поражения ткани и позволяют идентифицировать CCL2-специфические оптические изомеры как новое терапевтическое средство против аутоиммунного повреждения ткани.
Пример 9
Лечение диабетической нефропатии у мышей с диабетической односторонней нефроэктомией с использованием оптического изомера против mMCP-1
Диабетическая нефропатия остается главной причиной развития абсолютной почечной недостаточности, поскольку воздействие на ангиотензин-зависимые патологические механизмы не всегда приводит к предотвращению прогрессирования заболевания (Zimmet 2001; Ritz 1999; United States Renal Data System 2004; Svensson 2003). Следовательно, для лечения диабетической нефропатии помимо существующих терапевтических методов необходимо разработать другие стратегии лечения.
Данные, полученные в недавно проводимых экспериментальных исследованиях, позволяют установить связь прогрессирования диабетической нефропатии с внутрипочечным воспалением (Galkina 2006; Mora 2005; Meyer 2003; Tuttle 2005). Так, например, микофенолят-мофетил, метотрексат или облучение снижают уровень экскреции альбумина мочи и степень гломерулосклероза у крыс с диабетической нефропатией, индуцированной стрептозотоцином (Yozai 2005; Utimura 2003). Тем не менее, молекулярные и клеточные механизмы внутрипочечного воспаления при диабетической нефропатии пока недостаточно охарактеризованы. У пациентов с диабетической нефропатией наблюдаются повышенные уровни маркеров острой фазы воспаления в сыворотке, но это не является показателем внутрипочечного воспаления (Dalla Vestra 2005; Navarro 2003). У пациентов с диабетической нефропатией наблюдается экскреция высоких уровней моноцитарного белка-хемоаттрактанта 1, а именно CC-хемокина (MCP-1/CCL2) в моче, который может быть более специфичным к внутрипочечному воспалению (Morii 2003; Tashiro 2002; Takebayashi 2006). Действительно, MCP-1/CCL2 экспрессируется человеческими мезангиальными клетками, подвергаемыми воздействию либо высокими концентрациями глюкозы, либо конечными продуктами глубокого гликозилирования (Ihm 1998; Yamagishi 2002). CCL2 участвует в сложном многостадийном процессе рекрутинга лейкоцитов из интраваскулярных компартментов в экстраваскулярные компартменты, то есть в клубочках и в почечной интерстициальной ткани (Baggiolini 1998). Действительно, инфильтраты макрофагов обычно присутствуют у человека и у экспериментального животного с диабетическим гломерулосклерозом и тубулярноинтерстициальным поражением (Bohle 1991; Furuta 1993; Chow 2007). У Ccl2-дефицитных мышей с диабетом типа 1 или типа 2 наблюдаются более низкие уровни клубочковых макрофагов, что обусловлено меньшим поражением клубочков (Chow 2004; Chow 2006). В этих исследованиях была также продемонстрирована функциональная роль CCL2 в развитии гломерулярной патологии при диабетической нефропатии типа 1 и типа 2. Следовательно, CCL2 может представлять собой потенциальную терапевтическую мишень для диабетической нефропатии, и для лечения таких патологий целесообразно использовать антагонисты CCL2 с нужными фармакокинетическими профилями. В этом примере авторы настоящего изобретения сообщают об эффектах ПЭГилированного оптического изомера mNOX-E36-3'ПЭГ против CCL2 у db/db-мышей с диабетом типа 2, у которых наблюдается прогрессирующая диабетическая нефропатия. Авторами настоящего изобретения было показано, что оптический изомер, направленный против CCL2, должен быть эффективным для лечения диабетической нефропатии.
Животные и протокол эксперимента
5-недельные самцы db/db мышей C57BLKS или мышей C57BLKS дикого типа было получены от Taconic (Ry, Denmark), и эти мыши содержались в вентиляционных клетках с верхним фильтром в условиях 12-часового цикла “день/ночь” и с неограниченным доступом к корму и воде на протяжении всего исследования. Перед началом исследования клетки, подстилки, норы, корм и вода были стерилизованы в автоклаве. При достижении мышами возраста 6 недель этих мышей подвергали односторонней нефроэктомии (мыши “1K") или ложной хирургической операции (мыши “2K"), которую проводили путем бокового разреза в 1 см, как описано в литературе для db/db-мышей и мышей дикого типа (Bower 1980). У мышей ложно прооперированной группы почки оставались на месте. Через 10 недель, при достижении мышами возраста 4 месяца, db/db-мышей 1K разделяли на две группы, которым три раза в неделю подкожно вводили инъекцию либо mNOX-E36-3'ПЭГ, либо PoC-ПЭГ в 5% глюкозы (доза 0,9 мкмоль/кг; объем инъекции 1 мл/кг). Лечение продолжали в течение 8 недель (до достижения мышами возраста 6 месяцев), а затем животных умерщвляли и получали ткани для гистологического анализа. На все экспериментальные процедуры было получено разрешение Федеральных регуляторных органов.
Оценка диабетической нефропатии
Все иммуногистологические исследования осуществляли на залитых в парафин срезах, как описано в литературе (Anders 2002). В качестве “первых” антител использовали следующие антитела: крысиное анти-Mac2 антитело (гломерулярные макрофаги, Cederlane, Ontario, Canada, 1:50) и анти-Ki-67 антитело (пролиферация клеток, Dianova, Hamburg, Germany, 1:25). Для проведения гистопатологического анализа часть почек, взятых у каждой мыши, фиксировали 10% формалином в забуференном фосфатом физиологическом растворе и заливали в парафин. 3 мкм-срезы окрашивали реагентом Шиффа, а именно периодной кислотой или серебром в соответствии с инструкциями поставщиков (Bio-Optica, Milano, Italy). Клубочковые склеротические поражения оценивались наблюдателем полуколичественным слепым методом по следующей шкале оценок: 0 = поражение отсутствует, 1 = склеротическое поражение <25%, 2 = склеротическое поражение 25-49%, 3 = склеротическое поражение 50-74%, 4 = склеротическое поражение 75-100% соответственно. Было проанализировано 15 клубочков на срез. Показатели интерстициального объема и дилатации клубочков определяли путем наложения сетки из 100 точек на 10 неперекрывающихся кортикальных полей, как описано в литературе (Anders 2002). Количество интерстициальных клеток было определено наблюдателем слепым методом в 15 полях зрения при большом увеличении микроскопа (hpf, 400x). Из депарафинизированных клубочков получали РНК-препарат и проводили количественную ОТ-ПЦР в реальном времени (TaqMan). После инкубирования в буфере для лизиса (10 мМ трис-HCl, 0,1 мМ EDTA, 2% ДСН и 20 мкг/мл протеиназы K) в течение 16 часов при 60°C проводили экстракцию РНК фенолом-хлороформом. Гломерулярную РНК растворяли в 10 мкл воды, не содержащей РНКазы. РНК всех органов и гломерулярную РНК подвергали обратной транскрипции и ОТ-ПЦР в реальном времени, как описано в литературе (Anders 2002, Cohen 2002). Контроль, состоящий из ddH2O, не содержал генов-мишеней и генов “домашнего хозяйства”. Олигонуклеотидный праймер (300 нМ) и зонды (100 нМ) для рРНК mCcl2, Gapdh и 18S представляют собой ранее разработанные аналитические реагенты TaqMan, поставляемые от PE. Праймеры и зонды поставлялись от ABI Biosystems, Weiterstadt, Germany. Скорость клубочковой фильтрации (GFR) определяли по кинетике клиренса ФИТЦ-инулина из плазмы (Sigma-Aldrich, Steinheim, Germany) через 5, 10, 15, 20, 35, 60 и 90 минут после одной инъекции ударной дозы (Qi 2004). Интенсивность флуоресценции определяли при возбуждении на длине волны 485 нм и считывали при излучении на длине волны 535 нм. GFR вычисляли, исходя из двухкомпонентной модели, с использованием компьютерной программы построения нелинейной кривой регрессии (GraphPad Prism, GraphPad Software Inc., San Diego, CA). Все данные выражали как среднее ± среднеквадратическая ошибка среднего (SEM). Сравнение между группами осуществляли с использованием ANOVA, а для проведения множества сравнений использовали поправку Бонферрони рost-hoc. Величина p<0,05 рассматривалась как показатель статистической значимости.
Результаты
mNOX-E36-3'ПЭГ снижает число гломерулярных макрофагов и степень генерализованного гломерулосклероза у db/db-мышей с односторонней нефроэктомией
Если отсутствие функционального CCL2 ассоциируется со снижением уровня рекрутинга гломерулярных макрофагов у db/db-мышей (Chow 2007), а mNOX-E36-3'ПЭГ обладает способностью блокировать CCL2-опосредуемый рекрутинг макрофагов in vitro и in vivo, то mNOX-E36-3'ПЭГ должен нарушать рекрутинг почечных макрофагов у db/db-мышей с прогрессирующей диабетической нефропатией типа 2. Для проверки этой гипотезы авторами настоящего изобретения были введены подкожные инъекции mNOX-E36-3'ПЭГ или PoC-ПЭГ 4-месячным db/db-мышам с односторонней нефроэктомией ("1K"). Обработку проводили в течение 8 недель, после чего брали ткани для оценки тяжести диабетической нефропатии. В течение этого периода времени mNOX-E36-3'ПЭГ-обработка не оказывала значительного влияния на количество лейкоцитов или тромбоцитов, при этом уровни глюкозы в крови или масса тела заметно повышались у db/db-мышей во всех группах по сравнению с мышами BLKS, у которых отсутствовал диабет (данные не приводятся). Интересно отметить, что mNOX-E36-3'ПЭГ повышал уровни CCL2 в сыворотке у db/db-мышей 1K, что указывало на то, что антагонист CCL2 сохранял уровни CCL2 в кровотоке (фиг.44). В подтверждение гипотезы, выдвинутой авторами настоящего изобретения, следует отметить, что mNOX-E36-3'ПЭГ значительно снижал количество гломерулярных макрофагов, т.е. на 40% по сравнению с числом гломерулярных макрофагов у db/db-мышей, обработанных PoC-ПЭГ или носителем, что ассоциируется со снижением числа Ki-67-позитивных пролиферирующих клеток в клубочках у mNOX-E36-3'ПЭГ-обработанных db/db-мышей (фиг.45). Полученные результаты ассоциируются с заметной положительной динамикой генерализованного диабетического гломерулосклероза у db/db-мышей 1К (фиг.46). Действительно, mNOX-E36-3'ПЭГ-обработка приводила к снижению тяжести диабетического гломерулосклероза у db/db-мышей 1К до уровня, наблюдаемого у db/db-мышей того же возраста, которые не были подвергнуты нефроэктомии (“2К”) (фиг.46). Полученные данные показали, что замедленная блокада CCL2-зависимого рекрутинга гломерулярных макрофагов с использованием mNOX-E36-3'ПЭГ приводит к предупреждению генерализованного диабетического гломерулосклероза у db/db-мышей с диабетом типа 2.
mNOX-E36-3'ПЭГ увеличивает скорость GFR у db/db-мышей 1К
Благоприятные эффекты mNOX-E36-3'ПЭГ-обработки в отношении диабетического гломерулосклероза у db/db-мышей 1К должны ассоциироваться с увеличением GFR. Авторами настоящего изобретения была проанализирована кинетика клиренса ФИТЦ-инулина как маркера GFR у db/db-мышей (Qi 2004). Авторами настоящего изобретения было обнаружено, что по сравнению с нормальной GFR, составляющей примерно 250 мл/мин у db/db-мышей (Qi 2004), у 6-месячных db/db-мышей 1К, которым был введен PoC-ПЭГ, GFR снижалась и составляла 112±23 мл/мин (фиг.47). mNOX-E36-3'ПЭГ-обработка приводила к значительному увеличению GFR до 231±30 мл/мин у db/db-мышей 1K (p<0,001), что позволяет предположить, что блокирование CCL2-зависимого рекрутинга гломерулярных макрофагов может также улучшать почечную функцию у мышей с диабетом типа 2.
mNOX-E36-3'ПЭГ снижает число интерстициальных макрофагов и степень тубулярноинтерстициального поражения у db/db-мышей 1K
Прогрессирующая диабетическая нефропатия у человека ассоциируется с увеличением числа интерстициальных макрофагов и повышением тяжести тубулярноинтерстициального поражения (Bohle 1991). У db/db-мышей 2К до достижения ими 8-месячного возраста инфильтрации интерстициальных макрофагов и значительного тубулярноинтерстициального поражения не наблюдалось (Chow 2007). Ранняя односторонняя нефроэктомия ускоряла развитие тубулярноинтерстициальной патологии у db/db-мышей (Ninichuk 2005), и поэтому авторами настоящего изобретения была проведена количественная оценка уровней интерстициальных макрофагов, степени дилатации канальцев и объема интерстициальной ткани, которые являются показателями тубулярноинтерстициального поражения у 6-месячных мышей всех групп. В то же время у db/db-мышей 1K, по сравнению с db/db-мышами 2К, наблюдалось увеличение числа интерстициальных макрофагов и значительное увеличение степени дилатации канальцев и увеличение объема интерстициальной ткани (фиг.45, фиг.48). mNOX-E36-3'ПЭГ-обработка приводила к снижению числа интерстициальных макрофагов на 53%, а также степени дилатации канальцев и объема интерстициальной ткани у db/db-мышей 1K (фиг.45, фиг.48). Таким образом, блокирование CCL2-зависимого рекрутинга почечных макрофагов также приводит к предотвращению тубулярноинтерстициального поражения у db/db-мышей с диабетом типа 2.
mNOX-E36-3'ПЭГ снижает уровень почечной экспрессии Ccl2 у db/db-мышей 1K
Инфильтраты макрофагов усиливают воспалительные ответы на участках поражения ткани, например, локальную экспрессию CCL2. Авторами настоящего изобретения была выдвинута гипотеза, что mNOX-E36-3'ПЭГ-зависимое снижение числа почечных макрофагов должно ассоциироваться со снижением уровня экспрессии почечного CCL2. Для количественной оценки экспрессии мРНК CCL2 у db/db-мышей авторами настоящего изобретения была проведена ОТ-ПЦР в реальном времени. mNOX-E36-3'ПЭГ снижал уровни мРНК CCL2 в почках 6-месячных db/db-мышей 1K по сравнению с уровнями, наблюдаемыми у PoC-ПЭГ-обработанных мышей того же возраста (фиг.49). Для дополнительной оценки пространственной экспрессии CCL2 авторами настоящего изобретения было осуществлено иммунологическое окрашивание почечных срезов на белок CCL2. У db/db-мышей 1K, по сравнению с db/db-мышами 2К или мышами дикого типа, экспрессия CCL2 заметно усиливалась в клетках клубочков и канальцев, а также в интерстициальных клетках (фиг.50). У этих мышей, по сравнению с db/db-мышами 1К, обработанными носителем или PoC-ПЭГ, mNOX-E36-3'ПЭГ заметно снижал интенсивность окрашивания на CCL2 во всех компартментах. Полученные данные показали, что блокирование CCL2-зависимого рекрутинга почечных макрофагов оптическим изомером mNOX-E36-3'ПЭГ приводит к снижению уровня локальной экспрессии CCL2 у db/db-мышей 1К.
Выводы
Существующее мнение о том, что воспаление играет важную роль в прогрессировании диабетической нефропатии у человека, вызывает все больший интерес (Tuttle 2005), и поэтому для лечения этого заболевания особое внимание следует обратить на MCP-1/CCL2 как на потенциальную мишень. В этом примере авторами настоящего изобретения было показано, что обработка односторонне нефроэктомизированных 6-месячных мышей с диабетом оптическим изомером mNOX-E36-3'ПЭГ приводит к снижению числа гломерулярных (и интерстициальных) макрофагов, что связано с меньшей степенью пролиферации гломерулярных клеток. Кроме того, mNOX-E36-3'ПЭГ-обработка приводит к заметному снижению уровня экспрессии мРНК CCL2 в почках/клубочках. Более того, при проведении терапии данной группы снижение числа гломерулярных макрофагов и уровня пролиферации гломерулярных клеток ассоциируется с защитой ткани от гломерулосклероза и со значительным увеличением скорости клубочковой фильтрации. Благоприятное воздействие mNOX-E36-3'ПЭГ на гломерулярную патологию и почечную функцию у мышей с диабетом соответствует данным исследований, в которых были использованы другие антагонисты CCL2 в других моделях гломерулярного поражения (Lloyd 1997, Hasegawa 2003, Tang 1996, Wenzel 1997, Fujinaka 1997, Schneider 1999). Интересно отметить, что замедленное начало блокады CCL2 также приводит к снижению числа интерстициальных макрофагов, ассоциированному с меньшей степенью тубулярноинтерстициальной патологии у db/db-мышей 1K.
В целом, полученные данные позволяют признать, что CCL2 является перспективной терапевтической мишенью при лечении диабетической нефропатии, а также подтверждают предположение, что индуцирование блокады CCL2 оптическим изомером - даже в запущенной стадии заболевания - может служить защитным механизмом.
Библиография
Ниже приводятся полные библиографические данные цитируемых здесь документов, содержание которых, если это не оговорено особо, вводится в настоящее описание посредством ссылки.
Akahoshi T, Wada C, Endo H, Hirota K, Hosaka S, Takagishi K, Kondo H, Kashiwazaki S, Matsushima K, (1993). Expression of monocyte chemotactic and activating factor in rheumatoid arthritis. Regulation of its production in synovial cells by interleukin-1 and tumor necrosis factor. Arthritis Rheum. 36:762.
Alam R, York J, Moyars M, Stafford S, Grant J.A, Lee J, Forsythe P, Sim T, Ida N, (1996). Increased MCP-1, RANTES, and MIP-1α in bronchoalveolar lavage fluid of allergic asthmatic patients. Am. J. Respir. Crit. Care Med. 153:1398.
Altschul S.F, Gish W, Miller W, Myers E.W, Lipman D.J. (1990), Basic local alignment search tool. J Mol. Biol. 215(3):403-10.
Altschul S.F, Madden T.L, Schaffer A.A, Zhang J, Zhang Z, Miller W, Lipman D.J, (1997). Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res. Sep 1; 25(17):3389-402.
Amann B, Tinzmann R, Angelkort B, (2003). ACE inhibitors improve diabetic nephropathy through suppression of renal MCP-1. Diabetes Care 26:2421.
Anders H.J, Vielhauer V, Frink M, Linde Y, Cohen C.D, Blattner S.M, Kretzler M, Strutz F, Mack M, Grone H.J, Onuffer J, Horuk R, Nelson P.J, Schlöndorff D, (2002). A chemokine receptor CCR-1 antagonist reduces renal fibrosis after unilateral ureter ligation. J. Clin. Invest. 109:251.
Anders H.J, Vielhauer V, Schlöndorff D (2003). Chemokines and chemokine receptors are involved in the resolution or progression of renal disease. Kidney Int. 63:401.
Aurup H et al. (1994). Nucleic Acids Res 22:20.
Austin H.A. 3rd, Muenz L.R, Joyce K.M, Antonovych T.T, Balow J.E, (1984). Diffuse proliferative lupus nephritis: identification of specific pathologic features affecting renal outcome. Kidney Int. 25:689.
Baggiolini M, Dewald B, Moser B (1994). Interleukin-8 and related chemotactic cytokines - CXC and CC chemokines. Adv. Immunol. 55:97.
Baggiolini M, (1998). Chemokines and leukocyte traffic. Nature 392:565.
Banba N, Nakamura T, Matsumura M, Kuroda H, Hattori Y, Kasai K, (2000). Possible relationship of monocyte chemoattractant protein-1 with diabetic nephropathy. Kidney Int. 58:684.
Banisor I, Leist T.P, Kalman B, (2005). Involvement of β-chemokines in the development of inflammatory demyelination. J. Neuroinflammation 2:7.
Bazan J.F, Bacon K.B, Hardiman G, Wang W, Soo K, Rossi D, Greaves D.R, Zlotnik A, Schall T.J, (1997). A new class of membrane-bound chemokine with a CX3C motif. Nature 385:640.
Berkhout T.A. (1997). J Biol. Chem. 272:16404.
Bohle A, Wehrmann M, Bogenschutz O, Batz C, Muller C.A, Muller G.A, (1991). The pathogenesis of chronic renal failure in diabetic nephropathy. Investigation of 488 cases of diabetic glomerulosclerosis. Pathol. Res. Pract. 187:251.
Boring L, Gosling J, Chensue S.W, Kunkel SL, Farese R.V. Jr, Broxmeyer H.E, Charo I.F, (1997). Impaired monocyte migration and reduced type 1 (Th1) cytokine responses in C-C chemokine receptor 2 knockout mice. J. Clin. Invest. 100:2552.
Boring L, Gosling J, Cleary M, Charo I.F, (1998). Decreased lesion formation in CCR2-/- mice reveals a role for chemokines in the initiation of atherosclerosis. Nature 394:894.
Boring L, Gosling J, Monteclaro F.S, Lusis A.J, Tsou C.L, Charo I.F, (1996). Molecular cloning and functional expression of murine JE (monocyte chemoattractant protein 1) and murine macrophage inflammatory protein 1alpha receptors: evidence for two closely linked C-C chemokine receptors on chromosome 9. J. Biol. Chem. 271:7551.
Bossink A.W, Paemen L, Jansen P.M, Hack C.E, Thijs L.G, Van Damme J, (1995). Plasma levels of the chemokines monocyte chemotactic proteins-1 and -2 are elevated in human sepsis. Blood 86:3841.
Bower G, Brown D.M, Steffes M.W, Vernier R.L, Mauer S.M, (1980). Studies of the glomerular mesangium and the juxtaglomerular apparatus in the genetically diabetic mouse. Lab. Invest. 43:333.
Charo I.F, Myers S.J, Herman A, Franci C, Connolly A.J, Coughlin S.R, (1994). Molecular cloning and functional expression of two monocyte chemoattractant protein 1 receptors reveals alternative splicing of the carboxyl-terminal tails. Proc. Natl Acad. Sci. USA 91:2752.
Chow F.Y, Nikolic-Paterson D.J, Ma F.Y, Ozols E, Rollins B.J, Tesch G.H, (2007). Monocyte chemoattractant protein-1-induced tissue inflammation is critical for the development of renal injury but not type 2 diabetes in obese db/db mice. Diabetologica 50:471.
Chow F.Y, Nikolic-Paterson D.J, Ozols E, Atkins R.C, Rollin B.J, Tesch G.H, (2006). Monocyte chemoattractant protein-1 promotes the development of diabetic renal injury in streptozotocin-treated mice. Kidney Int. 69:73.
Chow F, Ozols E, Nikolic-Paterson D.J, Atkins R.C, Tesch G.H, (2004). Macrophages in mouse type 2 diabetic nephropathy: Correlation with diabetic state and progressive renal injury. Kidney Int. 65:116.
Cockwell P, Howie A.J, Adu D, Savage C.O. (1998). In situ analysis of C-C chemokine mRNA in human glomerulonephritis. Kidney Int. 54:827.
Cohen C.D, Gröne H.J, Gröne E.F, Nelson P.J, Schlöndorff D, Kretzler M, (2002). Laser microdissection and gene expression analysis on formaldehyde-fixed archival tissue. Kidney Int. 61:125.
Cummins L.L. et al. (1995). Nucleic Acids Res 23:2019.
Dalla Vestra M, Mussap M, Gallina P, Bruseghin M, Cernigoi A.M, Saller A, Plebani M, Fioretto P, (2005). Acute-phase markers of inflammation and glomerular structure in patients with type 2 diabetes. J. Am. Soc. Nephrol. 16 Suppl 1:S78.
Dawson J, Miltz W, Mir A.K, Wiessner C, (2003). Targeting monocyte chemoattractant protein-1 signalling in disease. Expert Opin. Ther. Targets 7:35.
De Bleecker J.L, De Paepe B, Vanwalleghem I.E, Schroder J.M, (2002). Differential expression of chemokines in inflammatory myopathies. Neurology 58:1779.
Drolet D.W, Nelson J, Tucker C.E, Zack P.M, Nixon K, Bolin R, Judkins M.B, Farmer J.A, Wolf J.L, Gill S.C, Bendele R.A, (2000). Pharmacokinetics and safety of an anti-vascular endothelial growth factor aptamer (NX1838) following injection into the vitreous humor of rhesus monkeys. Pharm. Res. 17:1503.
Eaton B.E. et al. (1995). Chem Biol 2:633.
Eaton B.E, Gold L, Hicke B.J, Janjic N, Jucker F.M, Sebosta D.P, Tarasow T.M, Willis M.C, Zichi D.A, (1997). Bioorg Med. Chem 5:1087.
Economou E, Tousoulis D, Katinioti A, Stefanadis C, Trikas A, Pitsavos C, Tentolouris C, Toutouza M.G, Toutouzas P, (2001). Chemokines in patients with ischaemic heart disease and the effect of coronary angioplasty. Int. J. Cardiol. 80:55.
Egashira K, Zhao Q, Kataoka C, Ohtani K, Usui M, Charo I.F, Nishida K, Inoue S, Katoh M, Ichiki T, Takeshita A, (2002). Importance of monocyte chemoattractant protein-1 pathway in neointimal hyperplasia after periarterial injury in mice and monkeys. Circ. Res. 90:1167.
Fujinaka H, Yamamoto T, Takeya M, Feng L, Kawasaki K, Yaoita E, Kondo D, Wilson C.B, Uchiyama M, Kihara I, (1997). Suppression of anti-glomerular basement membrane nephritis by administration of anti-monocyte chemoattractant protein-1 antibody in WKY rats. J. Am. Soc. Nephrol. 8:1174.
Furuichi K, Wada T, Iwata Y, Kitagawa K, Kobayashi K-I, Hashimoto H, Ishiwata Y, Tomosugi N, Mukaida N, Matsushima K, Egashira K, Yokoyama H, (2003). Gene therapy expressing amino-terminal truncated monocyte chemoattractant protein-1 prevents renal ischemia-reperfusion injury. J. Am. Soc. Nephrol. 14:1066.
Furuta T, Saito T, Ootaka T, Soma J, Obara K, Abe K, Yoshinaga K, (1993). The role of macrophages in diabetic glomerulosclerosis. Am. J. Kidney Dis. 21:480.
Galasso J.M, Liu Y, Szaflarski J, Warren J.S, Silverstein F.S, (2000). Monocyte chemoattractant protein-1 is a mediator of acute excitotoxic injury in neonatal rat brain. Neuroscience 101:737.
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=pubmed&dopt=Abstract&list_uids=16394109&query_hl=55&itool=pubmed_docsum. Leukocyte recruitment and vascular injury in diabetic nephropathy. J. Am. Soc. Nephrol. 17:368-377.
Gao J.L, Kuhns D.B, Tiffany H.L, McDermott D, Li X, Francke U, Murphy P.M, (1993). Structure and functional expression of the human macrophage inflammatory protein 1 alpha/RANTES receptor. J. Exp. Med. 177:1421.
Garcia-Zepeda E.A, Combadiere C, Rothenberg M.E, Sarafi M.N, Lavigne F, Hamid Q, Murphy P.M, Luster A.D, (1996). Human monocyte chemoattractant protein (MCP)-4 is a novel CC chemokine with activities on monocytes, eosinophils, and basophils induced in allergic and nonallergic inflammation that signals through the CC chemokine receptors (CCR)-2 and -3. J. Immunol. 157:5613.
Gerard C, Rollins, B.J. Chemokines and disease. Nat. Immunol. 6:1182.
Gong X, Gong W, Kuhns D.B, Ben-Baruch A, Howard O.M, Wang J.M. (1997). Monocyte chemotactic protein-2 (MCP-2) uses CCR1 and CCR2B as its functional receptors. J. Biol. Chem. 272:11682.
Gonzalo J.A, Lloyd C.M, Wen D, Albar J.P, Wells T.N.C, Proudfoot A, Martinez-A C, Dorf M, Bjerke T, Coyle A.J, Gutierrez-Ramos J.C, (1998). The coordinated action of CC chemokines in the lung orchestrates allergic inflammation and airway hyperresponsiveness. J. Exp. Med. 188:157.
Gordillo G.M, Onat D, Stockinger M, Roy S, Atalay M, Beck F.M, Sen C.K, (2004). A key angiogenic role of moncyte chemoattractant protein-1 in hemangioendothelioma proliferation. Am. J. Physiol. Cell Physiol. 287:C866.
Green L.S, et al. (1995). Chem Biol 2:683.
Handel T.M, Domaille P.J. (1996). Heteronuclear (1H, 13C, 15N) NMR assignments and solution structure of the monocyte chemoattractant protein-1 (MCP-1) dimer. Biochemistry 35:6569.
Harigai M, Hara M, Yoshimura T, Leonard E.J, Inoue K, Kashiwazaki S (1993). Monocyte chemoattractant protein-1 (MCP-1) in inflammatory joint diseases and its involvement in the cytokine network of rheumatoid synovium. Clin. Immunol. Immunopathol. 69:83.
Hasegawa H, Kohno M, Sasaki M, Inoue A, Ito M.R, Terada M, Hieshima K, Maruyama H, Miyazaki J, Yoshie O, Nose M, Fujita S (2003). Antagonist of monocyte chemoattractant protein 1 ameliorates the initiation and progression of lupus nephritis and renal vasculitis in MRL/lpr mice. Arthritis Rheum. 48:2555.
Heath H, Qin S et al. (1997). Chemokine receptor usage by human eosinophils. The importance of CCR3 demonstrated using an antagonistic monoclonal antibody. J Clin Invest 99:178.
Holdsworth S.R, Kitching A.R, Tipping P.G, (2000). Chemokines as therapeutic targets in renal disease. Curr. Opin. Nephrol. Hypertens. 9:505.
Holgate S.T, Bodey K.S, Janezic A, Frew A.J, Kaplan A.P, Teran L.M, (1997). Release of RANTES, MIP-1α, and MCP-1 into asthmatic airways following endobronchial allergen challenge. Am. J. Respir. Crit. Care Med. 156:1377.
Hosaka S. et al. (1994). Clin. Exp. Immunol. 97:451.
Huang D.R, Wang J, Kivisakk P, Rollins B.J, Ransohoff R.M. (2001). Absence of monocyte chemoattractant protein 1 in mice leads to decreased local macrophage recruitment and antigen-specific T helper cell type 1 immune response in experimental autoimmune encephalomyelitis. J. Exp. Med. 193:713.
Hulkower K, Brosnan C.F, Aquino D.A, Cammer W, Kulshrestha S, Guida M.P, Rapoport D.A, Berman J.W, (1993). Expression of CSF-1, c-fms, and MCP-1 in the central nervous system of rats with experimental allergic encephalomyelitis. J. Immunol. 150:2525.
Humbert M, Ying S, Corrigan C, Menz G, Barkans J, Pfister R, Meng Q, Van Damme J, Opdenakker G, Durham S.R, Kay A.B, (1997). Bronchial mucosal expression of the genes encoding chemokines RANTES and MCP-3 in symptomatic atopic and nonatopic asthmatics: relationship to the eosinophil-active cytokines interleukin (IL)-5, granulocyte macrophage-colony-stimulating factor, and IL-3. Am J Respir Cell Mol. Biol. 16:1.
Ihm C.G, Park J.K, Hong S.P, Lee T.W, Cho B.S, Kim M.J, Cha D.R, Ha H, (1998). A high glucose concentration stimulates the expression of monocyte chemotactic peptide 1 in human mesangial cells. Nephron 79:33.
Iyonaga K, Takeya M, Saita N, Sakamoto O, Yoshimura T, Ando M, Takahashi K, (1994). Monocyte chemoattractant protein-1 in idiopathic pulmonary fibrosis and other interstitial lung diseases. Hum. Pathol. 25:455.
Johrer K, Zelle-Rieser C, Perathoner A, Moser P, Hager M, Ramoner R, Gander H, Holtl L, Bartsch G, Greil R, Thurnher M, (2005). Up-regulation of functional chemokine receptor CCR3 in human renal cell carcinoma. Clin Cancer Res 11:2459.
Jolicoeur C, Lemay A, Akoum A (2001). Comparative effect of danazol and a GnRH agonist on monocyte chemotactic protein-1 expression by endometriotic cells. Am. J. Reprod. Immunol. 45:86.
Jose P.J, Griffiths-Johnson D.A, Collins P.D, Walsh D.T, Moqbel R, Totty N.F, Truong O, Hsuan J.J, Williams T.J. Eotaxin: a potent eosinophil chemoattractant cytokine detected in a guinea pig model of allergic airways inflammation. J. Exp. Med. 179:881.
Kaburagi Y, Shimada Y, Nagaoka T, Hasegawa M, Takehara K, Sato S, (2001). Enhanced production of CC-chemokines (RANTES, MCP-1, MIP-1α, MIP-1β, and eotaxin) in patients with atopic dermatitis. Arch. Dermatol. Res. 293:350.
Kawasaki A.M. et al. (1993). J Med Chem 36:831.
Kennedy K.J, Strieter R.M, Kunkel S.L, Lukacs N.W, Karpus W.J. (1998). Acute and relapsing experimental autoimmune encephalomyelitis are regulated by differential expression of the CC chemokines macrophage inflammatory protein-1α and monocyte chemotactic protein-1. J. Neuroimmunol. 91:98.
Kim J.S, Gautam S.C, Chopp M, Zaloga C, Jones M.L, Ward P.A, Welch K.M, (1995). Expression of monocyte chemoattractant protein-1 and macrophage inflammatory protein-1 after focal cerebral ischemia in the rat. J. Neuroimmunol. 56:127.
Kitamoto S, Egashira K. (2003). Anti-monocyte chemoattractant protein-1 gene therapy for cardiovascular diseases. Expert Rev. Cardiovasc. Ther. 1:393.
Kleinhans M, Tun-Kyi A, Gilliet M, Kadin M.E, Dummer R, Burg G, and Nestle F.O, (2003). Functional expression of the eotaxin receptor CCR3 in CD30+ cutaneous T-cell lymphoma. Blood 101:1487.
Koch A.E, Kunkel S.L, Harlow L.A, Johnson B, Evanoff H.L, Haines G.K, Burdick M.D, Pope R.M, Strieter R.M. (1992). Enhanced production of monocyte chemoattractant protein-1 in rheumatoid arthritis. J. Clin. Invest. 90:772.
Kouno J, Nagai H, Nagahata T, Onda M, Yamaguchi H, Adachi K, Takahashi H, Teramoto A, and Emi M, (2004). Up-regulation of CC chemokine, CCL3L1, and receptors, CCR3, CCR5 in human glioblastoma that promotes cell growth. J Neurooncol 70:301.
Kurihara T, Warr G, Loy J, Bravo R (1997). Defects in macrophage recruitment and host defense in mice lacking the CCR2 chemokine receptor. J. Exp. Med. 186:1757.
Kusser W (2000). J Biotechnol. 74:27-38.
Kuziel W.A, Morgan S.J, Dawson T.C, Griffin S, Smithies O, Ley K, Maeda N, (1997). Severe reduction in leukocyte adhesion and monocyte extravasation in mice deficient in CC chemokine receptor 2. Proc. Natl Acad. Sci. U S A 94:12053.
Lesnik E.A. et al. (1993). Biochemistry 32:7832.
Lloyd C.M, Minto A.W, Dorf M.E, Proudfoot A, Wells T.N.C, Salant D.J, Gutierrez-Ramos J.C, (1997). RANTES and monocyte chemoattractant protein-1 (MCP-1) play an important role in the inflammatory phase of crescentic nephritis, but only MCP-1 is involved in crescent formation and interstitial fibrosis. J. Exp. Med. 185:1371.
Lu B.B, Rutledge B.J, Gu L, Fiorillo J, Lukacs N.W, Kunkel S.L, North R, Gerard C, Rollins B.J. (1998). Abnormalities in monocyte recruitment and cytokine expression in monocyte chemoattractant protein-1 deficient mice. J. Exp. Med. 187:601.
Lubkowski J, Bujacz G, Boque L, Domaille P.J, Handel T.M, Wlodawer A, (1997). The structure of MCP-1 in two crystal forms provides a rare example of variable quaternary interactions. Nat Struct. Biol 4:64.
Mack M, Cihak J, Simonis C, Luckow B, Proudfoot A.E, Plachy J, Bruhl H, Frink M, Anders H.J, Vielhauer V, Pfirstinger J, Stangassinger M, Schlöndorff D, (2001). Expression and characterization of the chemokine receptors CCR2 and CCR5 in mice. J. Immunol. 166:4697.
Martinelli R, Sabroe I, LaRosa G, Williams T.J, Pease J.E. The CC chemokine eotaxin (CCL11) is a partial agonist of CC chemokine receptor 2b. J Biol Chem 276:42957.
Matsushima K, Morishita K, Yoshimura T, Lavu S, Kobayashi Y, Lew W, Appella E, Kung H.F, Leonard E.J, Oppenheim J.J. (1989). Molecular cloning of a human monocyte-derived neutrophil chemotactic factor (MDNCF) and the induction of MDNCF mRNA by interleukin 1 and tumor necrosis factor. J. Exp. Med. 167:1883.
McGinnis S, Madden T.L. (2004). BLAST: at the core of a powerful and diverse set of sequence analysis tools. Nucleic Acids Res. 32(Web Server issue):W20-5.
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=pubmed&dopt=Abstract&list_uids=12472806&query_hl=62&itool=pubmed_DocSum. (2003). Immunosuppression for diabetic glomerular disease? Kidney Int. 63:377.
Miller M.D, Krangel M.S, (1992). Biology and biochemistry of the chemokines: a family of chemotactic and inflammatory cytokines. Crit. Rev. Immunol. 12:17.
Miller L.E. et al. (1993). J Physiol 469:213.
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=pubmed&dopt=Abstract&list_uids=16316588&query_hl=14&itool=pubmed_docsum. The role of inflammation as a pathogenic factor in the development of renal disease in diabetes. Curr. Diab. Rep. 5:399.
Morii T, Fujita H, Narita T, Shimotomai T, Fujishima H, Yoshioka N, Imai H, Kakei M, Ito S. (2003). Association of monocyte chemoattractant protein-1 with renal tubular damage in diabetic nephropathy. J. Diabetes Complications 17:11.
Murphy P.M, Baggiolini M, Charo I.F, Hebert C.A, Horuk R, Matsushima K, Miller L.H, Oppenheim J.J, Power C.A. (2000). International union of pharmacology. XXII. Nomenclature for chemokine receptors. Pharmacol. Rev. 52:145.
Nakamura H, Weiss S.T, Israel E, Luster A.D, Drazen J.M, Lilly C.M. (1999). Eotaxin and impaired lung function in asthma. Am J Respir Crit Care Med. 160:1952.
Nakazawa T, Hisatomi T, Nakazawa C, Noda K, Maruyama K, She H, Matsubara A, Miyahara S, Nakao S, Yin Y, Benowitz L, Hafezi-Moghadam A, Miller J.W. (2007). Monocyte chemoattractant protein 1 mediated retinal detachment-induced photoreceptor apoptosis. Proc. Natl. Acad. Sci. U S A 104:2425.
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=pubmed&dopt=Abstract&list_uids=12830456&query_hl=65&itool=pubmed_docsum (2003). Inflammatory parameters are independently associated with urinary albumin in type 2 diabetes mellitus. Am. J. Kidney Dis. 42:53.
Myers S.J, Wong L.M, Charo I.F. (1995). Signal transduction and ligand specificity of the human monocyte chemoattractant protein-1 receptor in transfected embryonic kidney cells. J. Biol. Chem. 270:5786.
Needleman & Wunsch (1970), A general method applicable to the search for similarities in the amino acid sequence of two proteins. J Mol Biol. 48(3):443-53.
Nelken N.A, Coughlin S.R, Gordon D, Wilcox J.N. (1991). Monocyte chemoattractant protein-1 in human atheromatous plaques. J. Clin. Invest. 88:1121.
Neote K, DiGregorio D, Mak J.Y, Horuk R, Schall T.J, (1993). Molecular cloning, functional expression, and signaling characteristics of a C-C chemokine receptor. Cell 72:415.
Ninichuk V, Gross O, Reichel C, Khandoga A, Pawar R.D, Ciubar R, Segerer S, Belemezova E, Radomska E, Luckow B, de Lema GP, Murphy P.M, Gao J.L, Henger A, Kretzler M, Horuk R, Weber M, Krombach F, Schlondorff D, Anders H.J, (2005). Delayed chemokine receptor 1 blockade prolongs survival in collagen 4A3-deficient mice with Alport disease. J. Am. Soc. Nephrol. 16:977.
Ogata H, Takeya M, Yoshimura T, Takagi K, Takahashi K, (1997). The role of monocyte chemoattractant protein-1 (MCP-1) in the pathogenesis of collagen-induced arthritis in rats. J. Pathol. 182:106.
Okuno T, Andoh A, Bamba S, Araki Y, Fujiyama Y, Fujiyama M, Bamba T, (2002). Interleukin-1β and tumor necrosis factor-α induce chemokine and matrix metalloproteinase gene expression in human colonic subepithelial myofibroblasts. Scand. J. Gastroenterol. 37:317.
Oppenheim J.J, Zachariae C.O, Mukaida N, Matsushima K (1991). Properties of the novel proinflammatory supergene "intercrine" cytokine family. Annu. Rev. Immunol. 9:617.
Pawar R.D, Patole P.S, Zecher D, Segerer S, Kretzler M, Schlöndorff D, Anders H.J. (2006). Toll-like receptor-7 modulates immune complex glomerulonephritis. J. Am. Soc. Nephrol. 17:141.
Pearson & Lipman (1988), Improved tools for biological sequence comparison. Proc. Nat'l. Acad. Sci. USA 85: 2444.
Perez de Lema G, Maier H, Franz T.J, Escribese M, Chilla M.S, Segerer S, Camarasa N, Schmid H, Banas B, Kalaydjiev S, Busch D.H, Pfeffer K, Mampaso F, Schlöndorff D, Luckow B (2005). Chemokine receptor CCR2 deficiency reduces renal disease and prolongs survival in MRL/lpr lupus-prone mice. J. Am. Soc. Nephrol. 16:3592.
Perez de Lema G, Maier H, Nieto E, Vielhauer V, Luckow B, Mampaso F, Schlöndorff D. Chemokine expression precedes inflammatory cell infiltration and chemokine receptor and cytokine expression during the initiation of murine lupus nephritis. J. Am. Soc. Nephrol. 12:1369.
Ponath P.D, Qin S, Ringler D.J, Clark-Lewis I, Wang J, Kassam N, Smith H, Shi X, Gonzalo J.A, Newman W, Gutierrez-Ramos J.C, Mackay CR (1996a). Cloning of the human eosinophil chemoattractant, eotaxin. Expression, receptor binding, and functional properties suggest a mechanism for the selective recruitment of eosinophils. J. Clin. Invest. 97:604.
Ponath P.D, Qin S, Post T.W, Wang J, Wu L, Gerard N.P, Newman W, Gerard C, Mackay C.R, (1996b). Molecular cloning and characterization of a human eotaxin receptor expressed selectively on eosinophils. J. Exp. Med. 183:2437.
Power C.A, Meyer A, Nemeth K, Bacon K.B, Hoogewerf A.J, Proudfoot A.E, Wells T.N, (1995). Molecular cloning and functional expression of a novel CC chemokine receptor cDNA from a human basophilic cell line. J. Biol. Chem. 270:19495.
Qi Z, Whitt I, Mehta A, Jin J, Zhao M, Harris R.C, Fogo A.B, Breyer M.D, (2004). Serial determination of glomerular filtration rate in conscious mice using FITC-inulin clearance. Am. J. Physiol. Renal Physiol. 286:F590.
Qin S, LaRosa G, Campbell J.J, Smith-Heath H, Kassam N, Shi X, Zeng L, Buthcher E.C, Mackay CR (1996). Expression of monocyte chemoattractant protein-1 and interleukin-8 receptors on subsets of T-cells: correlation with transendothelial chemotactic potential. Eur. J. Immunol. 26:640.
Ransohoff R.M. et al. (1993). FASEB J 7:592.
Raport C.J, Gosling J, Schweickart V.L, Gray P.W, Charo I.F. (1996). Molecular cloning and functional characterization of a novel human CC chemokine receptor (CCR5) for RANTES, MIP-1β, and MIP-1α. J. Biol. Chem. 271:17161.
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=pubmed&dopt=Abstract&list_uids=10561134&query_hl=6 End-stage renal failure in type 2 diabetes: A medical catastrophe of worldwide dimensions. Am. J. Kidney Dis. 34:795-808.
Rollins B.J, Stier P, Ernst T, Wong G.G. (1989). The human homolog of the JE gene encodes a monocyte secretory protein. Mol. Cell Biol. 9:4687.
Rollins B.J. (1996). Monocyte chemoattractant protein 1: a potential regulator of monocyte recruitment in inflammatory disease. Mol. Med. Today 2:198.
Rovin B.H, Rumancik M, Tan L, Dickerson J. (1994). Glomerular expression of monocyte chemoattractant protein-1 in experimental and human glomerulonephritis. Lab. Invest. 71:536.
Ruffing N, Sullivan N. et al. (1998). CCR5 has an expanded ligand-binding repertoire and is the primary receptor used by MCP-2 on activated T cells. Cell Immunol. 189:160.
Salcedo R, Ponce M.L, Young H.A, Wasserman K, Ward J.M, Keinman H.K, Oppenheim J.J, Murphy W.J (2000). Human endothelial cells express CCR2 and respond to MCP-1: direct role of MCP-1 in angiogenesis and tumor progression. Blood 96:34.
Samson M, Labbe O, Mollereau C, Vassart G, Parmentier M. (1996). Molecular cloning and functional expression of a new human CC-chemokine receptor gene. Biochemistry 35:3362.
Schall T.J, Bacon K.B (1994). Chemokines, leukocyte trafficking, and inflammation. Curr. Opin. Immunol. 6:865.
Schneider A, Panzer U, Zahner G, Wenzel U, Wolf G, Thaiss F, Helmchen U, Stahl R.A. (1999). Monocyte chemoattractant protein-1 mediates collagen deposition in experimental glomerulonephritis by transforming growth factor-beta. Kidney Int. 56:135.
Schwarting A, Paul K, Tschirner S, Menke J, Hansen T, Brenner W, Kelly V.R, Relle M, Galle P.R. (2005). Interferon-beta: a therapeutic for autoimmune lupus in MRL-Faslpr mice. J. Am. Soc. Nephrol. 16:3264.
Schwartz C.J, Valente A.J, Sprague E.A. (1993). A modern view of atherogenesis. Am. J. Cardiol. 71:9B.
Segerer S, Nelson P.J, Schlöndorff D. (2000). Chemokines, chemokine receptors, and renal disease: from basic science to pathophysiologic and therapeutic studies. J. Am. Soc. Nephrol. 11:152.
Shimizu S, Nakashima H, Masutani K, Inoue Y, Miyake K, Akahoshi M, Tanaka Y, Egashira K, Hirakata H, Otsuka T, Harada M. (2004). Anti-monocyte chemoattractant protein-1 gene therapy attenuates nephritis in MRL/lpr mice. Rheumatology (Oxford) 43:1121.
Smith & Waterman (1981), Adv. Appl. Math. 2: 482.
Springer T.A. (1995). Traffic signals on endothelium for lymphocyte recirculation and leukocyte emigration. Annu. Rev.Physiol. 57:827.
Steinman L. (2004). Immune therapy for autoimmune diseases. Science 305:212.
Svensson M, Sundkvist G, Arnqvist HJ, Bjork E, Blohme G, Bolinder J, Henricsson M, Nystrom L, Torffvit O, Waernbaum I, Ostman J, Eriksson J.W. (2003). Signs of nephropathy may occur early in young adults with diabetes despite modern diabetes management: Results from the nationwide population-based Diabetes Incidence Study in Sweden (DISS). Diabetes Care 26:2903.
Takebayashi K, Matsumoto S, Aso Y, Inukai T. (2006). Association between circulating monocyte chemoattractant protein-1 and urinary albumin excretion in nonobese Type 2 diabetic patients. J. Diabetes Complications 20:98.
Takeya M, Yoshimura T, Leonard E.J, Takahashi K. (1993). Detection of monocyte chemoattractant protein-1 in human atherosclerotic lesions by an anti-monocyte chemoattractant protein-1 monoclonal antibody. Hum. Pathol. 24:534.
Tang W.W, Qi M, Warren J.S. (1996). Monocyte chemoattractant protein 1 mediates glomerular macrophage infiltration in anti-GBM Ab GN. Kidney Int. 50:665.
Tashiro K, Koyanagi I, Saitoh A, Shimizu A, Shike T, Ishiguro C, Koizumi M, Funabiki K, Horikoshi S, Shirato I, Tomino Y. (2002). Urinary levels of monocyte chemoattractant protein-1 (MCP-1) and interleukin-8 (IL-8), and renal injuries in patients with type 2 diabetic nephropathy. J. Clin. Lab. Anal. 16:1.
Tesch G.H, Maifert S, Schwarting A, Rollins B.J, Kelley V.R. (1999). Monocyte chemoattractant protein 1-dependent leukocytic infiltrates are responsible for autoimmune disease in MRL-Fas(lpr) mice. J. Exp. Med. 190:1813.
Tuaillon N, Shen de F, Berger R.B, Lu B, Rollins B.J, Chan C.C. (2002). MCP-1 expression in endotoxin-induced uveitis. Invest. Ophthalmol. Vis. Sci. 43:1493.
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=pubmed&dopt=Abstract&list_uids=15872083&query_hl=4. (2005). Linking metabolism and immunology: diabetic nephropathy is an inflammatory disease. J. Am. Soc. Nephrol. 16:1537.
Uguccioni M, Mackay C.R. et al. (1997). High expression of the chemokine receptor CCR3 in human blood basophils. Role in activation by eotaxin, MCP-4, and other chemokines. J Clin Invest 100:1137.
United States Renal Data System (2004). Annual data report: Incidence and prevalence 2004. Am. J. Kidney Dis. 45:S77.
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=pubmed&dopt=Abstract&list_uids=12472785&query_hl=87&itool=pubmed_docsum. (2003). Mycophenolate mofetil prevents the development of glomerular injury in experimental diabetes. Kidney Int. 63:209.
Van Riper G, Siciliano S, Fischer P.A, Meurer R, Springer M.S, Rosen H. (1993). Characterization and species distribution of high affinity GTP-coupled receptors for human rantes and monocyte chemoattractant protein 1. J. Exp. Med. 177:851.
Venkatesan N. et al. (2003). Curr Med. Chem. 10:1973.
Vestergaard C, Just H, Baumgartner Nielsen J, Thestrup-Pedersen K, Deleuran M. (2004). Expression of CCR2 on monocytes and macrophages in chronically inflamed skin in atopic dermatitis and psoriasis. Acta Derm. Venereol. 84:353.
Viedt C, Orth S.R. (2002). Monocyte chemoattractant protein-1 (MCP-1) in the kidney: does it more than simply attract monocytes? Nephrol. Dial. Transplant. 17:2043.
Wada T, Furuichi K, Segada-Takaeda C, Ahimizu M, Sakai N, Takeda S.I, Takasawa K, Kida H, Kobayashi K.I, Mukaida N, Ohmoto Y, Matsushima K, Yokoyama H. (1999). MIP-1α and MCP-1 contribute to crescents and interstitial lesions in human crescentic glomerulonephritis. Kidney Int. 56:995.
Wada T, Yokoyama H, Matsushima K, Kobayashi K.I. (2001). Chemokines in renal diseases. Int. Immunopharmacol. 1:637.
Wada T, Yokoyama H, Furuichi K, Kobayashi K.I, Harada K, Naruto M, Su S.B, Akiyama M, Mukaida N, Matsushima K. (1996). Intervention of crescentic glomerulonephritis by antibodies to monocyte chemotactic and activating factor (MCAF/MCP-1). FASEB J. 10:1418.
Wang X, Yue T.L, Barone F.C, Feuerstein G.Z. (1995). Monocyte chemoattractant protein-1 messenger RNA expression in rat ischemic cortex. Stroke 26:661.
Wenzel U, Schneider A, Valente A.J, Abboud H.E, Thaiss F, Helmchen U.M, Stahl R.A. (1997). Monocyte chemoattractant protein-1 mediates monocyte/macrophage influx in anti-thymocyte antibody-induced glomerulonephritis. Kidney Int. 51:770.
Yamagishi S, Inagaki Y, Okamoto T, Amano S, Koga K, Takeuchi M, Makita Z. (2002). Advanced glycation end product-induced apoptosis and overexpression of vascular endothelial growth factor and monocyte chemoattractant protein-1 in human-cultured mesangial cells. J. Biol. Chem. 277:20309.
Ying S, Robinson D.S, Meng Q, Rottman J, Kennedy R, Ringler D.J, Mackay C.R, Daugherty B.L, Springer M.S, Durham S.R, Williams T.J, Kay A.B. (1997). Enhanced expression of eotaxin and CCR3 mRNA and protein in atopic asthma. Association with airway hyperresponsiveness and predominant co-localization of eotaxin mRNA to bronchial epithelial and endothelial cells. Eur J Immunol 27:3507.
Ying S, Meng Q, Zeibecoglou K, Robinson D.S, Macfarlane A, Humbert M, Kay A.B. (1999). Eosinophil chemotactic chemokines (eotaxin, eotaxin-2, RANTES, monocyte chemoattractant protein-3 (MCP-3), and MCP-4), and C-C chemokine receptor 3 expression in bronchial biopsies from atopic and nonatopic (Intrinsic) asthmatics. J Immunol 163:6321.
Yla-Herttuala S, Lipton B.A, Rosenfeld M.E, Sarkioja T, Yoshimura T, Leonard E.J, Witztum J.L, Steinberg D. (1991). Expression of monocyte chemoattractant protein 1 in macrophage-rich areas of human and rabbit atherosclerotic lesions. Proc. Natl Acad. Sci. U S A 88:5252.
Yoshimura T, Robinson E.A, Tanaka S, Appella E, Leonard E.J. (1989). Purification and amino acid analysis of two human monocyte chemoattractants produced by phytohemagglutinin-stimulated human blood mononuclear leukocytes. J. Immunol. 142:1956.
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=pubmed&dopt=Abstract&list_uids=16177002&query_hl=61&itool=pubmed_DocSumMethotrexate prevents renal injury in experimental diabetic rats via anti-inflammatory actions. J. Am. Soc. Nephrol. 16:3326.
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=pubmed&dopt=Abstract&list_uids=11742409&query_hl=3&itool=pubmed_DocSum. (2001). Global and societal implications of the diabetes epidemic. Nature 414:782.
Отличительные признаки настоящего изобретения, формула изобретения и/или графический материал, представленные в настоящей заявке, могут быть использованы как отдельно, так и в любой их комбинации в целях осуществления настоящего изобретения в его различных формах.
| название | год | авторы | номер документа |
|---|---|---|---|
| СВЯЗЫВАЮЩАЯ МСР-1 НУКЛЕИНОВАЯ КИСЛОТА И ЕЕ ПРИМЕНЕНИЕ | 2008 |
|
RU2542973C2 |
| CXCL8-СВЯЗЫВАЮЩИЕ НУКЛЕИНОВЫЕ КИСЛОТЫ | 2019 |
|
RU2815444C2 |
| СВЯЗЫВАЮЩИЕ SDF-1 НУКЛЕИНОВЫЕ КИСЛОТЫ И ИХ ПРИМЕНЕНИЕ | 2008 |
|
RU2612388C2 |
| SDF-1-СВЯЗЫВАЮЩИЕ НУКЛЕИНОВЫЕ КИСЛОТЫ | 2007 |
|
RU2590709C2 |
| АНТИТЕЛА ПРОТИВ CCR2 | 2009 |
|
RU2547595C2 |
| АНТИКИНОВЫЕ АНТИТЕЛА, КОТОРЫЕ СВЯЗЫВАЮТСЯ С НЕСКОЛЬКИМИ СС-ХЕМОКИНАМИ | 2010 |
|
RU2574786C2 |
| СВЯЗЫВАЮЩИЕСЯ С SDF1 НУКЛЕИНОВЫЕ КИСЛОТЫ И ИХ ПРИМЕНЕНИЕ ПРИ ЛЕЧЕНИИ РАКА | 2011 |
|
RU2679495C2 |
| СРЕДСТВА ИНГИБИРОВАНИЯ ЭКСПРЕССИИ ПРОТЕИНКИНАЗЫ 3 | 2007 |
|
RU2553561C2 |
| СПОСОБ ПОЛИАЛКОКСИЛИРОВАНИЯ НУКЛЕИНОВЫХ КИСЛОТ, КОТОРЫЙ ПОЗВОЛЯЕТ ИЗВЛЕКАТЬ И ПОВТОРНО ИСПОЛЬЗОВАТЬ ИЗБЫТОК ПОЛИАЛКОКСИЛИРУЮЩЕГО РЕАГЕНТА | 2017 |
|
RU2765027C2 |
| КОМПОЗИЦИЯ ДНК ДЛЯ ВЫЗОВА ИММУННОГО ОТВЕТА ПРОТИВ ОПУХОЛЕАССОЦИИРОВАННЫХ МАКРОФАГОВ | 2007 |
|
RU2459631C2 |


















































































































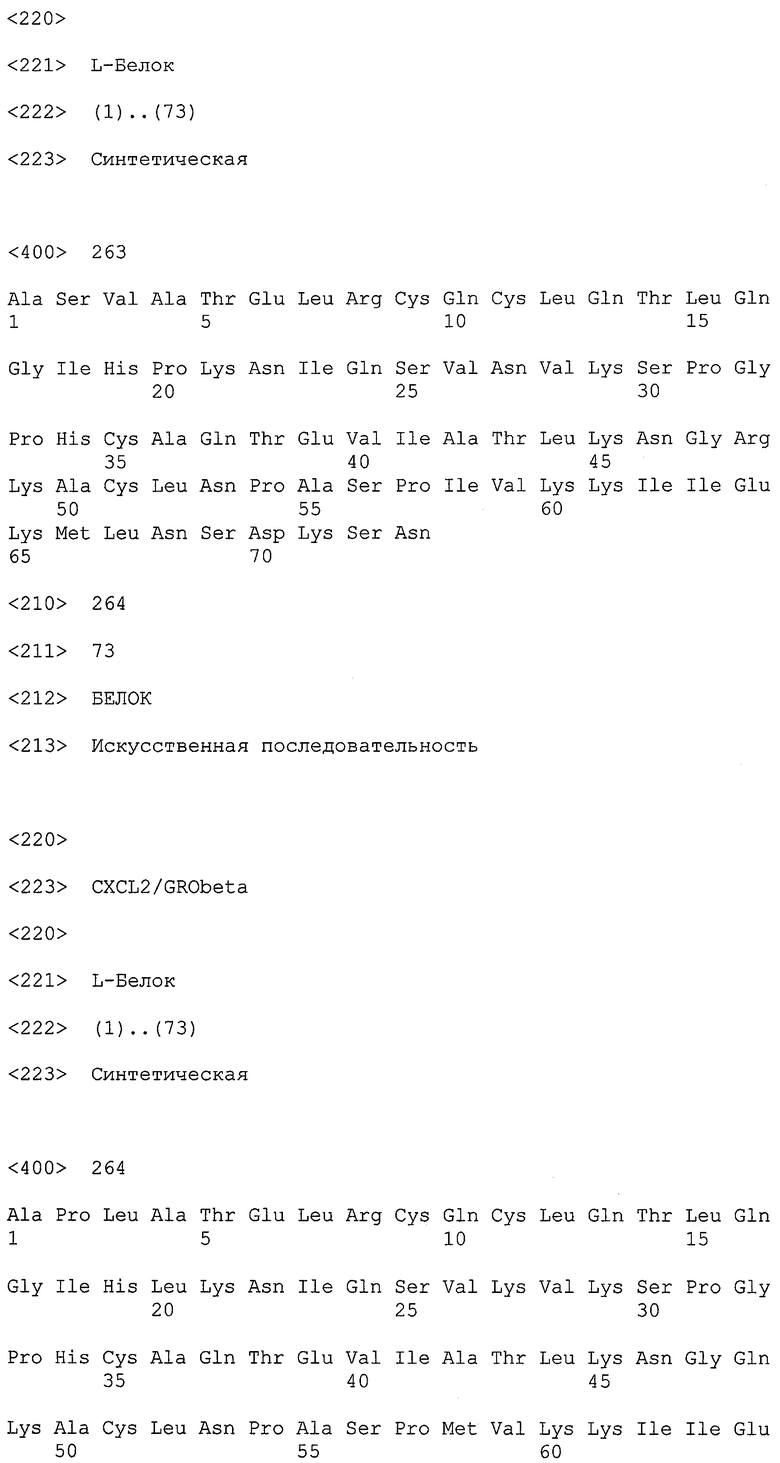













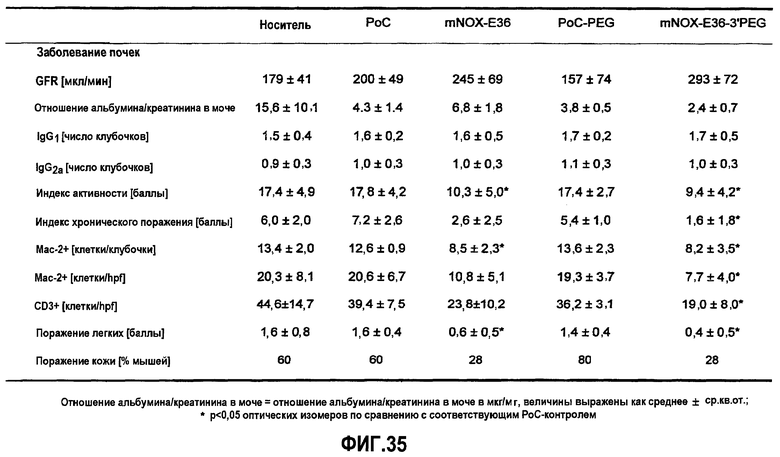
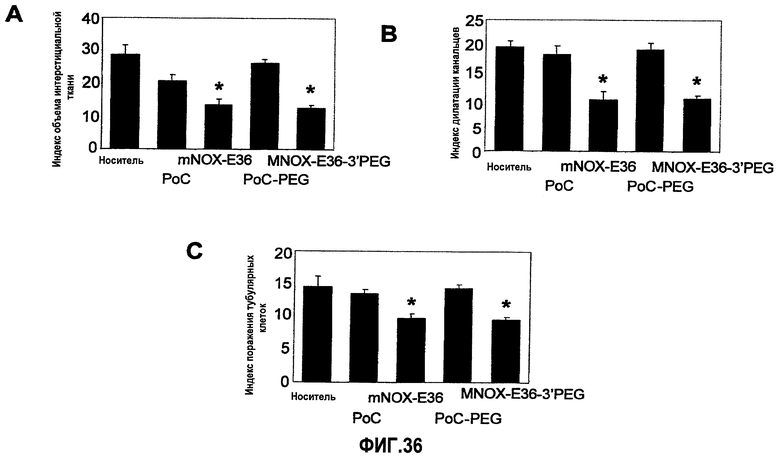
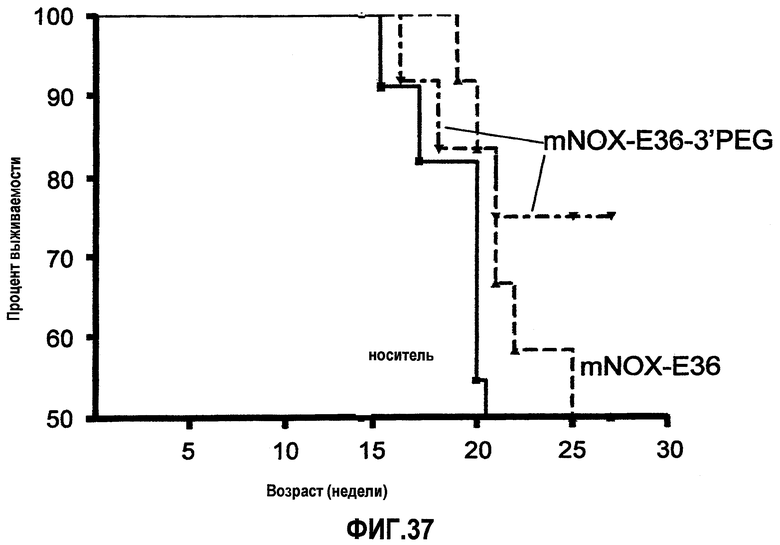
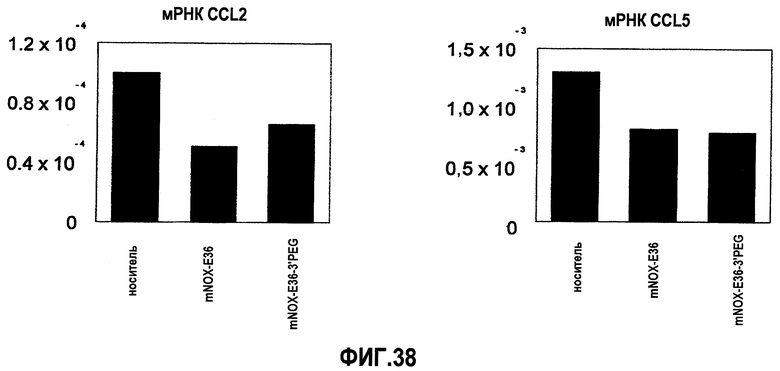
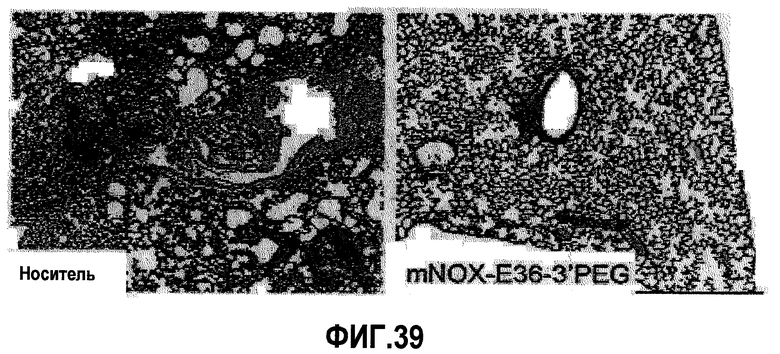

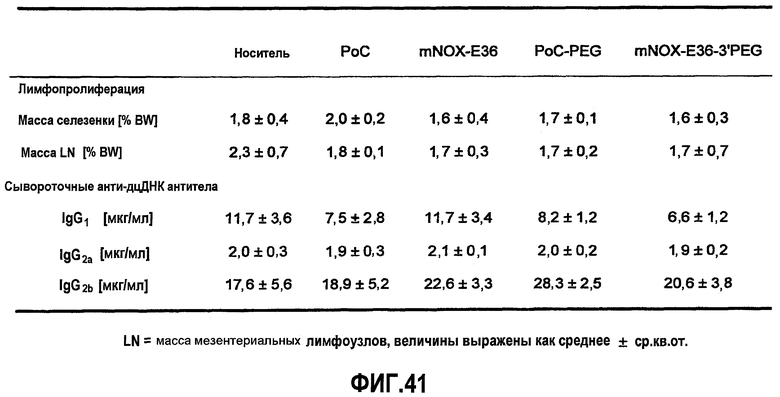
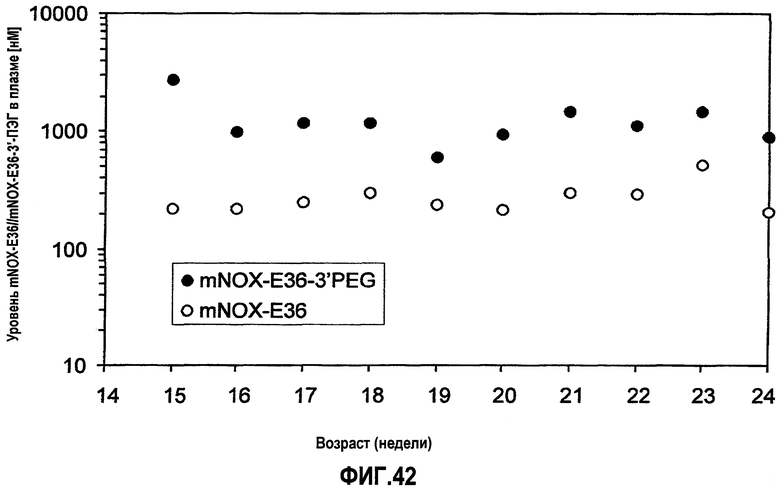
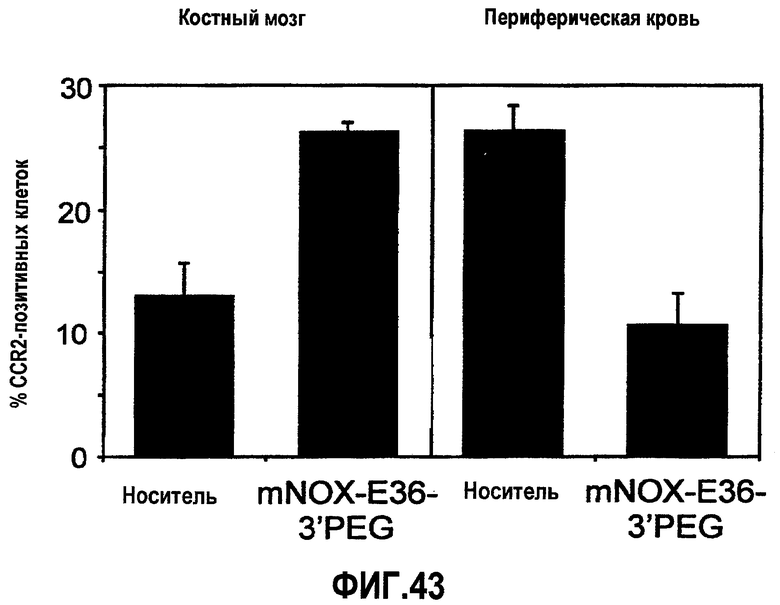
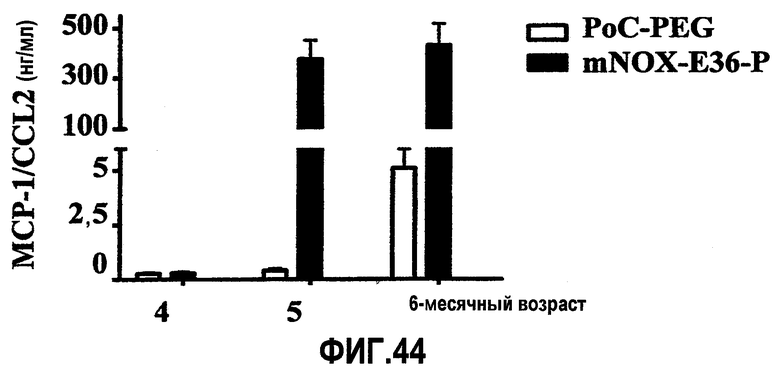
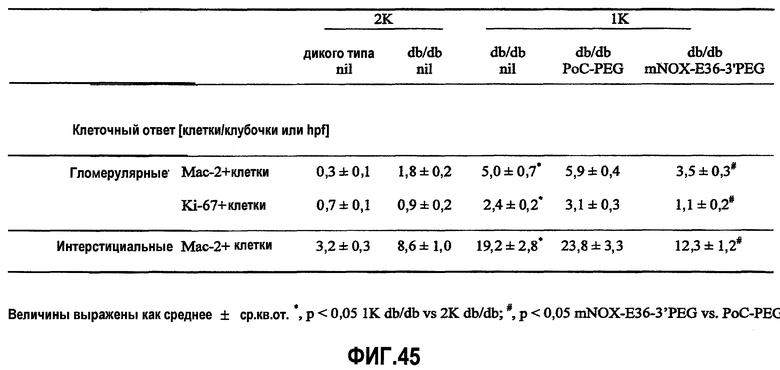
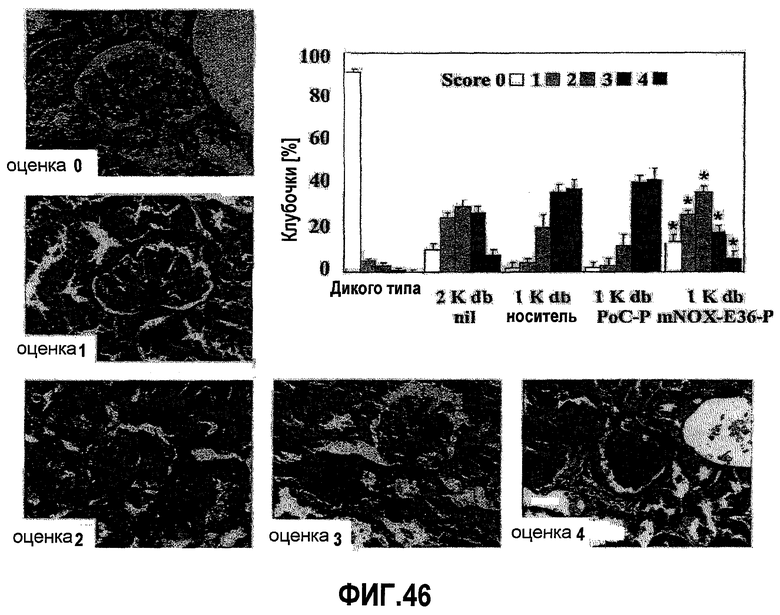
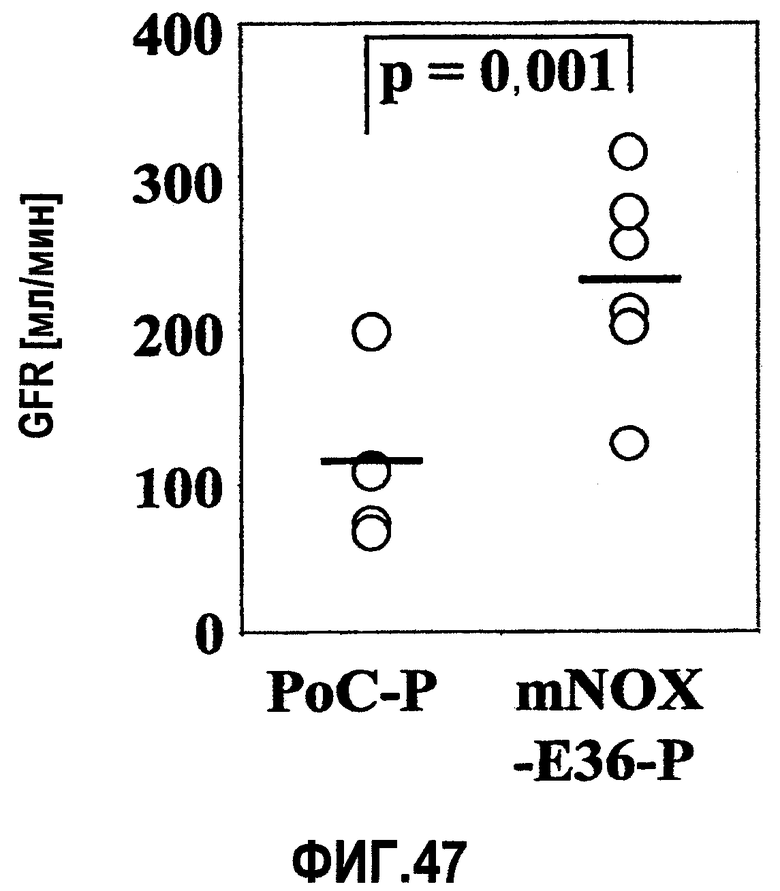
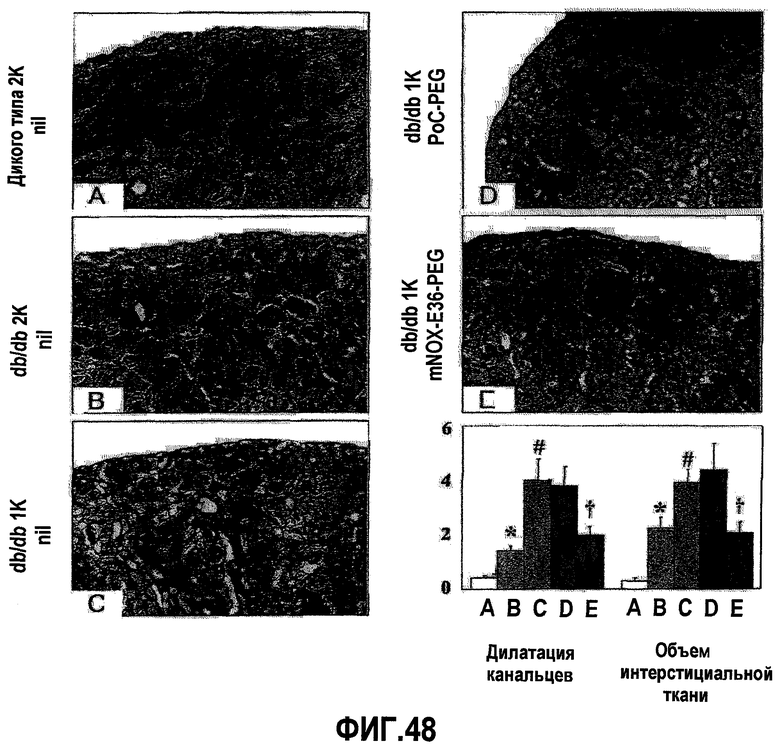
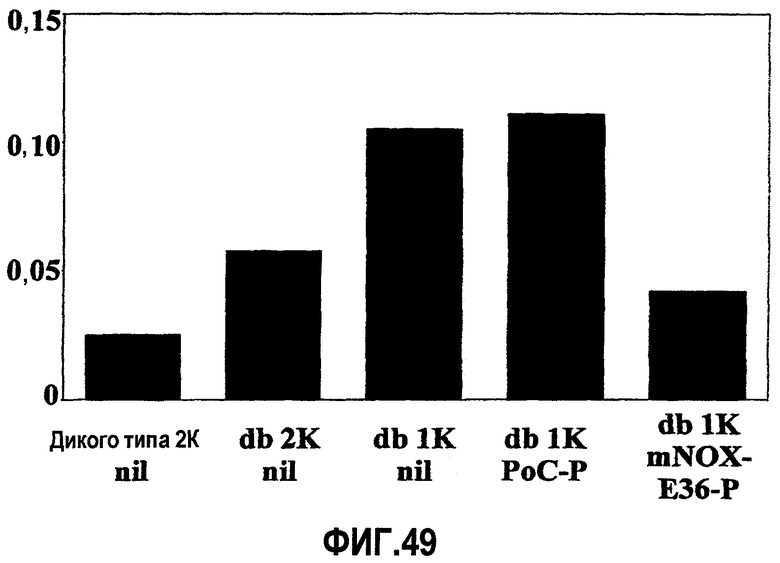
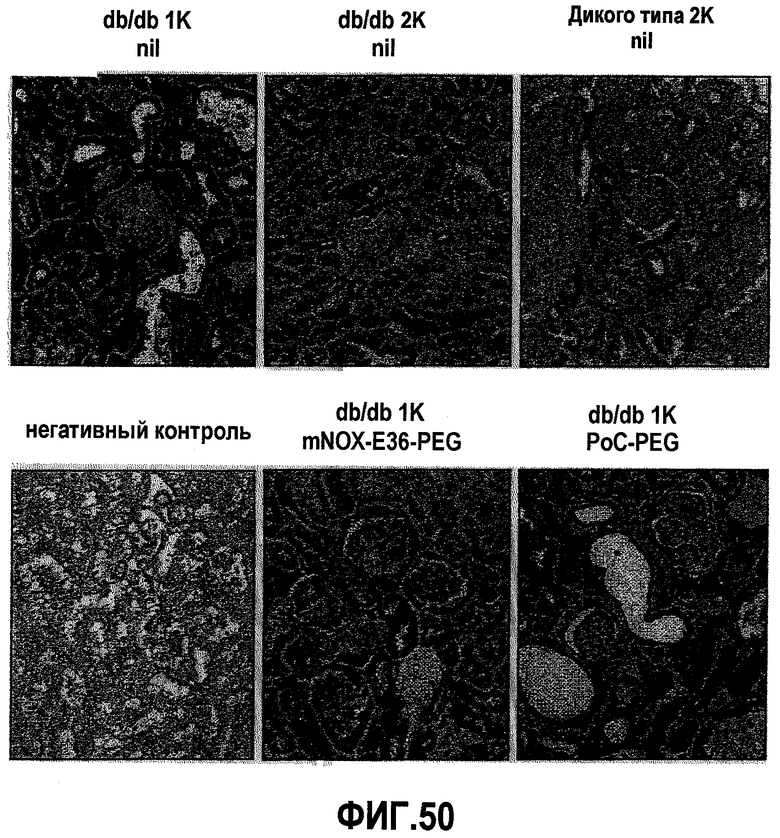
Изобретение относится к области молекулярной биологии, биохимии и медицины. Предложена L-нуклеиновая кислота - антагонист MCP-1 и способ её детектирования. Изобретение может быть использовано в лечении заболеваний, обусловленных MCP-1-механизмом патогенеза. 2 н. и 14 з.п. ф-лы, 50 ил., 1 табл., 9 пр.
1. L-нуклеиновая кислота, представляющая собой антагонист МСР-1 человека, связывающаяся с МСР-1, где нуклеиновая кислота, в направлении 5'→3', состоит из первого фрагмента "бокс В1А", второго фрагмента "бокс В2" и третьего фрагмента "бокс B1B", где
первый фрагмент "бокс В1А" состоит из нуклеотидной последовательности, выбранной из группы, состоящей из ACGCA, CGCA и GCA,
второй фрагмент "бокс В2" состоит из нуклеотидной последовательности CSUCCCUCACCGGUGCAAGUGAAGCCGYGGCUC, а
третий фрагмент "бокс В1В" состоит из нуклеотидной последовательности, выбранной из группы, состоящей из UGCGU, UGCG и UGC.
2. L-нуклеиновая кислота по п.1, гдевторой фрагмент "бокс В2" состоит из нуклеотидной последовательности CGUCCCUCACCGGUGCAAGUGAAGCCGUGGCUC.
3. L-нуклеиновая кислота по п.1 и 2, где
a) первый фрагмент "бокс В1А" состоит из нуклеотидной последовательности ACGCA, a
третий фрагмент "бокс В1В" состоит из нуклеотидной последовательности UGCGU; или
b) первый фрагмент "бокс В1А" состоит из нуклеотидной последовательности CGCA, a
третий фрагмент "бокс В1В" состоит из нуклеотидной последовательности UGCG; или
с) первый фрагмент "бокс В1А" состоит из нуклеотидной последовательности GCA, а
третий фрагмент "бокс В1В" состоит из нуклеотидной последовательности UGC или UGCG.
4. L-нуклеиновая кислота по п.1, гдепервый фрагмент "бокс В1А" состоит из нуклеотиднойпоследовательности GCA.
5. L-нуклеиновая кислота по.1, где третий фрагмент "бокс В1В" состоит из нуклеотиднойпоследовательности UGCG.
6. L-нуклеиновая кислота по п.1, где L-нуклеиновая кислота состоит из нуклеотидной последовательности SEQ ID NO:37, SEQ ID NO0:116, SEQ ID NO:117 или SEQ ID NO:278.
7. L-нуклеиновая кислота по п.1, где первый фрагмент "бокс В1А" и третий фрагмент "бокс В1В" гибридизуются друг с другом, и в результате такой гибридизации образуется двухцепочечная структура.
8. L-нуклеиновая кислота по п.1, содержащая модификацию, где модифицирован либо 5'-концевой нуклеотид, либо 3'-концевой нуклеотид указанной нуклеиновой кислоты и где указанная модификация представляет собой высокомолекулярную группу и/или указанная модификация позволяет изменять время присутствия L-нуклеиновой кислоты по любому из пп.1-7 в организме животного или человека.
9. L-нуклеиновая кислота по п.8, где указанная модификация выбрана из группы, включающей группу ГЭК и группу ПЭГ.
10. L-нуклеиновая кислота по п.9, где указанной модификацией является группа ПЭГ, состоящая из прямого или разветвленного ПЭГ, где молекулярная масса указанной молекулы ПЭГ предпочтительно составляет от 20 до 120 кДа, более предпочтительно примерно от 30 до 80 кДа, а наиболее предпочтительно примерно 40 кДа.
11. L-нуклеиновая кислота по п.9, где указанной модификацией является группа ГЭК, где молекулярная масса указанной молекулы ГЭК предпочтительно составляет от 10 до 130 кДа, более предпочтительно от 30 до 130 кДа, а наиболее предпочтительно 100 кДа.
12. Способ детектирования L-нуклеиновой кислоты по любому из пп.1-11 в образце, включающий стадии
a) получения образца, содержащего L-нуклеиновую кислоту согласно изобретению;
b) получения зонда для захвата, где указанный зонд для захвата комплементарен первой части L-нуклеиновой кислоты по любому из пп.1-11, и детектирующего зонда, где указанный детектирующий зонд комплементарен второй части L-нуклеиновой кислоты по любому из пп.1-11, или, альтернативно, указанный зонд для захвата комплементарен второй части L-нуклеиновой кислоты по любому из пп.1-11, а детектирующий зонд комплементарен первой части L-нуклеиновой кислоты по любому из пп.1-11;
c) осуществления реакции указанного зонда для захвата и детектирующего зонда либо одновременно, либо последовательно в любом порядке с L-нуклеиновой кислотой по любому из пп.1-11 или ее частью;
d) необязательного детектирования гибридизации указанного зонда для захвата с L-нуклеиновой кислотой по любому из пп.1-11, описанной на стадии (а); и
е) детектирования комплекса, полученного, как описано на стадии (с), и состоящего из L-нуклеиновой кислоты по любому из пп.1-11, зонда для захвата и детектирующего зонда, где указанный детектирующий зонд содержит детектирующее средство.
13. Способ по п.12, где указанный детектирующий зонд содержит детектирующее средство; и/или где указанный зонд для захвата может быть иммобилизован на носителе, предпочтительно на твердом носителе.
14. Способ по п.12 или 13, где любой детектирующий зонд, который не является частью данного комплекса, удаляют из реакционной смеси, в результате чего на стадии (е) детектируется только детектирующий зонд, который является частью указанного комплекса.
15. Способ по п.12 или 13, где стадия (е) включает стадию сравнения сигнала, вырабатываемого детектирующим средством в том случае, если зонд для захвата и детектирующий зонд гибридизуются в присутствии L-нуклеиновой кислоты по любому из пп.1-14 или ее части и в отсутствие указанной L-нуклеиновой кислоты или ее части.
16. Способ по п.12 или 13, где указанной детектируемой L-нуклеиновой кислотой является L-нуклеиновая кислота, имеющая нуклеотидную последовательность SEQ ID NO:37, 116, 117 или 278, а указанный зонд для захвата или детектирующий зонд содержит нуклеотидную последовательность SEQ ID NO:255 или SEQ ID NO:256.
| RHODES A | |||
| et al | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Способ получения бумажной массы из стеблей хлопчатника | 1912 |
|
SU506A1 |
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| WO 2005108431, 17.11.2005 | |||
| WO 200038714, 06.07.2000 | |||
| RU 2010108222, 20.09.2011 | |||

