Настоящее изобретение относится к молекулам нуклеиновых кислот, связывающихся с CXC-хемокиновым происходящим из стромальных клеток фактором-1 (SDF-1), способам лечения заболеваний и их применению в производстве лекарственного средства.
Хемокины. Хемокины представляют собой семейство структурно родственных, связывающих гепарин основных низкомолекулярных белков с молекулярной массой 8-14 кДа. С функциональной точки зрения их можно классифицировать на провоспалительные, гомеостатические, либо двойного действия (Moser, Wolf et al. 2004). Воспалительные хемокины индуцируются патогенными факторами, цитокинами или факторами роста и рекрутируют эффекторные лейкоциты к очагам инфекции, воспаления, тканевым повреждениям и опухолям. Такие хемокины регулируют рекрутинг, активацию и пролиферацию белых кровяных клеток, также называемых лейкоцитами (Schall and Bacon 1994; Springer 1995; Baggiolini 1998). Хемокины селективно индуцируют хемотаксис нейтрофилов, эозинофилов, базофилов, моноцитов, макрофагов, тучных клеток, T- и B-клеток. В дополнение к своему хемотаксическому эффекту они могут селективно вызывать другие эффекты у реактивных клеток, такие как изменения клеточной формы, временное увеличение концентрации свободных внутриклеточных ионов кальция, дегрануляция, активация интегринов, образование биологически активных липидов, таких как, помимо прочего, лейкотриены, простагландины, тромбоксаны, либо окислительный всплеск, то есть высвобождение реактивных форм кислорода для разрушения патогенных организмов или опухолевых клеток. Таким образом, провоцируя высвобождение дополнительных провоспалительных медиаторов, хемотаксис и проникновение лейкоцитов к очагам инфекции или воспаления, хемокины запускают усиление воспалительного ответа. Гомеостатические хемокины, с другой стороны, экспрессируются преимущественно в костном мозге и лимфоидных тканях, и вовлечены в гемопоэз, иммунологический надзор и адаптивные иммунные ответы (Godessart 2005).
Класс хемокинов. По взаимному расположению первых двух из четырех консервативных остатков цистеина хемокины подразделяют на четыре класса: CC или β-хемокины, у которых цистеины расположены один за другим; CXC или α-хемокины, где они разделены одним дополнительным аминокислотным остатком; XC или γ-хемокины, у которых есть лишь одна дисульфидная связь, причем лимфотактин, также именуемый XCL1, до настоящего времени является единственным представителем этого класса; и CX3C-хемокины, для которых характерны три аминокислотных остатка между цистеинами, при этом мембраносвязанный фракталкин является единственным представителем данного класса (Bazan, Bacon et al. 1997). Обычно хемокины имеют два названия: одно, которое присвоено на основании их функции, и другое, которое является систематическим названием согласно особенностям последовательности.
Хемокины класса CXC. Хемокины класса CXC действуют преимущественно на нейтрофилы, конкретнее, те хемокины класса CXC, которые несут аминокислотную последовательность ELR на своем аминоконце. Примерами хемокинов класса CXC, действующих на нейтрофилы, являются IL-8/CXCL8, GROα/CXCL1, GROβ/CXCL2, и GROγ/CXCL3, NAP-2/CXCL7, ENA-78/CXCL5, SDF-1/CXCL12 и GCP-2/CXCL6. Хемокины класса CXC действуют на более широкий ряд лейкоцитов, таких как моноциты, макрофаги, эозинофилы, базофилы, также как и T- и B-лимфоциты (Oppenheim, Zachariae et al. 1991; Miller and Krangel 1992; Baggiolini, Dewald et al. 1994; Jose, Griffiths-Johnson et al. 1994; Ponath, Qin et al. 1996). Их примерами являются I-309/CCL1; MCP-1/CCL2, MCP-2/CCL8, MCP-3/CCL7, MCP-4/CCL13, MIP-1α/CCL3 и MIP-1β/CCL4, RANTES/CCL5, а также эотаксин/CCL11.
Рецепторы хемокинов класса CXC. Хемокины действуют непосредственно через рецепторы, принадлежащие суперсемейству G-белок сопряженных рецепторов, семь раз пронизывающих мембрану, также обозначаемых GPCR (Murphy, Baggiolini et al. 2000). Вообще говоря, взаимодействия хемокина и хемокинового рецептора обычно бывают случайными в том смысле, что один хемокин может связывать несколько хемокиновых рецепторов, и, наоборот, один хемокиновый рецептор может взаимодействовать с несколькими различными хемокинами. Некоторые известные рецепторы хемокинов класса CXC включают CXCR1, который связывает GROα, GCP-2 и IL-8; CXCR2, который связывает хемокины, включающие GROα, GROβ, GROγ, ENA-78 и IL-8; CXCR3, который связывает хемокины, включающие PF4, MIG, IP-10 и I-TAC; CXCR4, который, как считается до сих пор, лишь генерирует сигнал в ответ на SDF-1, а также CXCR5, который, как показано, генерирует сигнал в ответ на BCA-1 (Godessart 2005). Кроме CXCR4, был идентифицирован новый рецептор SDF-1, названный RDC1/CXCR7 (Balabanian, Lagane et al. 2005, Burns, Summers et al. 2006).
SDF-1. Происходящий из стромальных клеток фактор-1 (сокр.: SDF-1; синонимы - CXCL12; PBSF [фактор, стимулирующий рост пре-В-клеток]; TPAR-1 [TPA-репрессированный ген 1]; SCYB12; TLSF [стимулирующий фактор клеток лимфомы тимуса]; hIRH [человеческий интеркрин, пониженный при гепатомах]) представляет собой ангиогенный хемокин класса CXC, который не содержит ELR-мотив, типичный для хемокинов, подобных IL-8 (Salcedo, Wasserman et al. 1999; Salcedo and Oppenheim 2003), но связывает и активирует G-белок сопряженный рецептор CXCR4. Данный хемокин независимо был открыт тремя группами, либо путем клонирования кДНК, несущих N-концевые сигнальные последовательности (Tashiro, Tada et al. 1993), благодаря своей способности стимулировать ранние предшественники B-клеток при экспрессии стромальными клетками линии PA6 (Nagasawa, Kikutani et al. 1994), либо путем выделения из библиотеки кДНК, созданной из фибробластов эмбриона мыши, обработанных тетрадодеканоилфорболацетатом (сокр.: TPA) – C-активатором протеинкиназы (Jiang, Zhou et al. 1994). В результате альтернативного сплайсинга существуют две формы SDF-1, - SDF-1α (68 аминокислотных остатков) и SDF-1β, который по сравнению с SDF-1α несет пять дополнительных остатков на C-конце (Shirozu, Nakano et al. 1995). Биологическое значение сплайсинга этих двух вариантов изучено не полностью.
Последовательности SDF-1. Весьма значительной является консервативность последовательностей SDF-1 из различных видов: SDF-1α человека (SEQ ID NO:1) и SDF-1α мыши (SEQ ID NO:2) фактически идентичны. Существует единственная консервативная замена V (валина) на I (изолейцин) в положении 18 (Shirozu, Nakano et al. 1995).
ЯМР-структура SDF-1. Существует модель ЯМР-структуры (выборка из базы данных PDB, 1SDF) для SDF-1 [8-68]. Было обнаружено, что SDF-1 представляет собой мономер с неупорядоченной N-концевой областью. Различия с другими хемокинами найдены главным образом в способе упаковки гидрофобного ядра (кора) и поверхностном распределении зарядов (Crump, Gong et al. 1997).
Физиологические активности SDF-1. Физиологические активности SDF-1: Поскольку CXCR4, рецептор SDF-1, широко экспрессируется на лейкоцитах, зрелых дендритных клетках, клетках эндотелия, клетках мозга и мегакариоцитах, активности SDF-1 являются плейотропными. Данный хемокин, к настоящему моменту идентифицированный лучше любого другого, проявляет самый широкий диапазон биологических функций, особенно вне иммунной системы. Самые существенные функциональные эффекты SDF-1 представляют собой:
«Хоминг» и прикрепление эпителиальных клеток к неоваскулярным участкам в сосудистой части оболочки сетчатки. Было показано, что SDF-1 вовлечен в «хоминг» эпителиальных клеток к сосудистой оболочке в процессе неоваскуляризации в ткани глаза. Точная роль этих клеток все еще исследуется, но опубликованная гипотеза такова, что эпителиальные клетки вовлечены в формирование аберрантных кровеносных сосудов (Sengupta, Caballero et al. 2005).
Стволовые клетки. SDF-1 необходим, чтобы поддерживать стволовые клетки и клетки-предшественники, например, гемопоэтические клетки-предшественники (обычно CD34+) в костном мозге взрослых людей. Для мобилизации клеток CD34+ при трансплантации гемопоэтических стволовых клеток можно применять AMD3100, селективный антагонист CXCR4. Клетки CD34+ мигрируют in vitro и in vivo вдоль градиента SDF-1, продуцируемого стромальными клетками (Aiuti, Webb et al. 1997).
Развитие и хемотаксис B-клеток. SDF-1 поддерживает пролиферацию пре-В-клеток и увеличивает рост предшественников В-клеток костного мозга (Nagasawa, Kikutani et al. 1994); это индуцирует специфическую миграцию пре- и про-B-клеток, хотя и не действует как существенный хемоаттрактант для зрелых B-клеток (D'Apuzzo, Rolink et al. 1997; Bleul, Schultze et al. 1998). По-видимому, SDF-1 важен для позиционирования B-клеток во вторичной лимфоидной ткани.
Хемотаксис T-клеток. SDF-1 представляет собой один из наиболее эффективных хемоаттрактантов для T-клеток; CXCR4 представлен у многих субпопуляций T-клеток (Bleul, Farzan et al. 1996).
Эмбриональное развитие. SDF-1 и его рецептор, CXCR4, являются существенными для эмбрионального развития. Нокаутные по SDF-1 и CXCR4 мыши не выживают в перинатальный период; они имеют дефекты межжелудочковой перегородки сердца или неправильное развитие мозжечка в дополнение к сниженному количеству B-клеток и миелоидных предшественников (Nagasawa, Hirota et al. 1996; Ma, Jones et al. 1998; Zou, Kottmann et al. 1998). SDF-1 также необходим для нормальной онтогении гемопоэза в процессе эмбрионального развития (Juarez and Bendall 2004).
ВИЧ инфекция. SDF-1 способен ингибировать Т-лимфотропное попадание ВИЧ-1 в линии клеток, несущие CXCR4, а экспрессия SDF-1 может оказывать сильное влияние на патогенез СПИД, поскольку полиморфизм гена SDF-1 человека влияет на появление СПИД (Bleul, Farzan et al. 1996).
Другие заболевания. Считается, что измененные уровни экспрессии SDF-1 или его рецептора, CXCR4, либо измененные ответы на эти молекулы связаны со многими заболеваниями людей, такими как ретинопатия (Brooks, Caballero et al. 2004; Butler, Guthrie et al. 2005; Meleth, Agron et al. 2005); рак молочной железы (Muller, Homey et al. 2001; Cabioglu, Sahin et al. 2005), яичников (Scotton, Wilson et al. 2002), поджелудочной железы (Koshiba, Hosotani et al. 2000), щитовидной железы (Hwang, Chung et al. 2003) и носоглотки (Wang, Wu et al. 2005); глиома (Zhou, Larsen et al. 2002); нейробластома (Geminder, Sagi-Assif et al. 2001); В-клеточный хронический лимфолейкоз (Burger, Tsukada et al. 2000); гипогаммаглобулинемия (Gulino, Moratto et al. 2004; Balabanian, Lagane et al. 2005; Kawai, Choi et al. 2005); синдромы иммунологической недостаточности (Arya, Ginsberg et al. 1999; Marechal, Arenzana-Seisdedos et al. 1999; Soriano, Martinez et al. 2002); патологическая неоваскуляризация (Salvucci, Yao et al. 2002; Yamaguchi, Kusano et al. 2003; Grunewald, Avraham et al. 2006); воспаление (Murdoch 2000; Fedyk, Jones et al. 2001; Wang, Guan et al. 2001); рассеянный склероз (Krumbholz, Theil et al. 2006); ревматоидный артрит/остеоартрит (Buckley, Amft et al. 2000; Kanbe, Takagishi et al. 2002; Grassi, Cristino et al. 2004).
Антагонизм SDF-1 и его рецептора. В моделях на экспериментальных животных антагонисты SDF-1 или его рецептора доказали свою эффективность для блокировки роста и/или метастатического распространения человеческих раковых клеток различного происхождения, такого как поджелудочная железа (Guleng, Tateishi et al. 2005; Saur, Seidler et al. 2005), толстая кишка (Zeelenberg, Ruuls-Van Stalle et al. 2003; Guleng, Tateishi et al. 2005), молочная железа (Muller, Homey et al. 2001; Lapteva, Yang et al. 2005), легкие (Phillips, Burdick et al. 2003), глиобластома и медуллобластома (Rubin, Kung et al. 2003), предстательная железа (Sun, Schneider et al. 2005), остеосаркома (Perissinotto, Cavalloni et al. 2005), меланома (Takenaga, Tamamura et al. 2004), желудок (Yasumoto, Koizumi et al. 2006) и множественная миелома (Menu, Asosingh et al. 2006). Помимо этого, анти-SDF-1 терапия на животных моделях оказалась полезной в профилактике неоваскуляризации сетчатки глаза (Butler, Guthrie et al. 2005, Mames, Mattheus et al. 2006), нефрита (Balabanian, Couderc et al. 2003) и артрита (Matthys, Hatse et al. 2001; Tamamura, Fujisawa et al. 2004; De Klerck, Geboes et al. 2005). Кроме того, с целью мобилизации клеток CD34+ для трансплантации гемопоэтических стволовых клеток применяли AMD3100, селективный антагонист CXCR4. Клетки CD34+ мигрировали in vitro и in vivo вдоль градиента SDF-1, создаваемого стромальными клетками (Aiuti, Webb et al. 1997).
SDF-1 и глазные заболевания. SDF-1 является участником в патологии заболеваний задней части глаза, таких как диабетическая ретинопатия (сокр. DR) (Fong, Aiello et al. 2004) и возрастная макулярная дегенерация (сокр. AMD) (Ambati, Anand et al. 2003). Оба эти заболевания повреждают глаз и приводят к постепенной потере зрения, завершаясь слепотой. Повреждение происходит из-за несоответствующего роста кровеносных сосудов в задней части глаза - процесс, известный как хороидальная неоваскуляризация (сокр. CNV). Во время CNV новые кровеносные сосуды, происходящие из сосудистой оболочки, мигрируют через разрыв в оболочке Бруха в пигментный эпителий под сетчаткой (сокр. sub-RPE) или в пространство под сетчаткой. Аномальные сосуды могут кровоточить, что называют также интраретинальным кровоизлиянием, или приводить к подтеканию жидкости под сетчатку. Это может привести к шрамам и приподнять макулу, что искажает зрение.
Диабетическая ретинопатия. DR - главное следствие диабета, часто встречающееся у пациентов с диабетом как типа 1, так и типа 2. В США насчитывается приблизительно 16 миллионов диабетиков, и почти 8 миллионов из них имеют некоторую форму DR. Если пролиферативную диабетическую ретинопатию (сокр. PDR) оставить без лечения, приблизительно 60% пациентов становятся слепыми на один или оба глаза в течение 5 лет. С тревожным повышением распространенности диабета в Северной Америке, Европе и многих развивающихся странах быстро растет и популяция пациентов. Например, у пациентов с диабетом случаи слепоты встречаются в 25 раз чаще, чем среди всего населения. Кроме того, DR - наиболее частая причина слепоты у граждан средних лет, составляя, по меньшей мере, 12 процентов всех новых случаев в Соединенных Штатах ежегодно. Действуют программы массовых обследований для того, чтобы проверить зрение пациентов с диабетом, и своевременно предоставить необходимое доступное лечение.
Непосредственные причины DR плохо изучены, но предполагается, что заболевание возникает вследствие комбинации исходных причин: ослабленная саморегуляция притока крови к сетчатке глаза; накопление сорбита в клетках сетчатки и накопление конечных продуктов повышенного гликозилирования во внеклеточной жидкости. Все эти факторы прямо или косвенно связаны с гипергликемией - избытком сахара в кровотоке.
Симптомы DR и AMD похожи. Пациенты теряют клетки сетчатки, и происходят микроаневризмы, то есть кровоизлияния, в базальной мембране сетчатки. Кроме того, эндотелиальный фактор роста сосудов (сокр. VEGF), инсулиноподобный фактор роста 1 (IGF-1) и другие факторы крови, возможно включающие SDF-1, привлекают новые васкулярные клетки и способствуют образованию повреждающих кровеносных сосудов.
Возрастная макулярная дегенерация. AMD разрушает центральное зрение человека. Ранние стадии заболевания могут быть даже незаметны, потому что симптомы среди пациентов различны. Иногда у пациента затронут только один глаз. Или зрение может быть ослаблено в обоих глазах, но незначительно. Данное заболевание вызывает искаженное или дефектное цветовосприятие. Часто отмечается наличие темного пятна в центре поля зрения.
Этиология (это означает «течение») заболевания плохо изучена. Часто полагают, что AMD представляет собой старение самого внешнего слоя сетчатки. Физические изменения происходят в центре сетчатки, также известном как макула, представляющем собой часть сетчатки, от которой зависит самое острое зрение.
Влажная форма AMD начинается как продолжение сухой формы заболевания. Приблизительно 90% пациентов страдают сухой формой AMD, которая приводит к истончению макулярных тканей и патологическим отклонениям в их пигментации. Для остальных характерна влажная форма, которая включает хороидальную неоваскуляризацию и часто - образование макулярного отека и кровоизлияние в сетчатке глаза или под сетчаткой глаза. Все это может привести к быстрому ухудшению остроты зрения.
Обычно самая распространенная причина слепоты у людей в возрасте старше 55 лет, влажная форма AMD, поражает приблизительно от 4% до 5% населения Соединенных Штатов в возрасте 65-74 лет, и почти 10% из тех, кто старше 75 лет. В одних только Соединенных Штатах уже насчитывается 5 миллионов человек старше 80 лет, пораженных этим заболеванием, и ожидается, что еще 5 миллионов человек заболеют до 2020 г.
Опухоли. Опухоли (включая солидные и гематологические новообразования и злокачественные опухоли) – это не только массы раковых клеток: инфильтрация опухолей иммунными клетками является особенностью рака. У многих человеческих раковых образований есть сложная сеть хемокинов, которая влияет на глубину и фенотип этого инфильтрата, так же как на рост опухоли, выживаемость, миграцию и ангиогенез. Большинство солидных опухолей содержит много доброкачественных стромальных клеток. Действительно, стромальные клетки иногда превосходят численностью раковые клетки. Преобладающие стромальные клетки, находящиеся в раковых образованиях, являются макрофагами, лимфоцитами, эндотелиальными клетками и фибробластами.
SDF-1 в опухолях. Клетки различных типов рака имеют различные профили экспрессии рецептора хемокина, но рецептор SDF-1, CXCR4, обычно встречается в опухолевых клетках мыши и человека: опухолевые клетки, по меньшей мере, из 23 различных типов человеческих раковых образований эпителиального, мезенхимного и гемопоэтического происхождения, экспрессируют CXCR4 (Balkwill 2004), при этом SDF-1 является единственным известным лигандом для CXCR4. Кроме костного мозга и вторичной лимфоидной ткани, где он экспрессируется конститутивно, SDF-1 обнаружен в участках локализации первичной опухоли в лимфоме (Corcione, Ottonello et al. 2000) и в опухолях головного мозга как нейронного, так и астроцитного происхождения. Кроме того, он присутствует в высоких уровнях при раке яичников (Scotton, Wilson et al. 2002) и раке поджелудочной железы (Koshiba, Hosotani et al. 2000), так же как и в местах метастазов в молочной железе (Muller, Homey et al. 2001), а также при раке щитовидной железы (Hwang, Chung et al. 2003), нейробластоме и гематологической злокачественной опухоли (Geminder, Sagi-Assif et al. 2001). Напротив, экспрессия CXCR4 является низкой или отсутствует в нормальной молочной железе (Muller, Homey et al. 2001), яичниках (Scotton, Wilson et al. 2002) и эпителии простаты (Sun, Schneider et al. 2005).
Кроме CXCR4, был идентифицирован новый рецептор SDF-1: RDC1/CXCR7 (Balabanian, Lagane et al. 2005, Burns, Summers et al. 2006). In vitro и in vivo исследования клеточных линий рака простаты показывают, что нарушения экспрессии CXCR7/RDC1 связаны с повышенными адгезивными и инвазивными активностями, вдобавок к выигрышу в выживаемости. Кроме того, было замечено, что уровни CXCR7/RDC1 регулируются посредством CXCR4 (Wang et al., 2008). In vitro и in vivo исследования показали, что оба рецептора SDF-1, а именно - CXCR4 и CXCR7, способствуют росту опухоли, метастатическому потенциалу и устойчивости к (вызванному химиотерапией) апоптозу в случаях многих опухолей, например, при раке молочной железы, глиобластомах, раке яичников, нейробластоме, раке легкого, раке ободочной и прямой кишки и раке простаты (Burns et al., 2006; Li et al., 2008; Scotton et al., 2002; Yang et al., 2008; Zagzag et al., 2008).
Таким образом, вероятно, экспрессия CXCR4 и CXCR7 является основной характеристикой некоторых опухолей.
Ингибирование хемокин-рецепторной передачи сигнала как терапевтическая возможность лечения рака. Ингибирование хемокин-рецепторной передачи сигнала в опухолевых клетках имеет потенциальные возможности вызывать задержку роста или апоптоз, и предотвращать инвазию и метастазирование in vivo, как показано следующими данными: нокдаун CXCR4 посредством миРНК уничтожал рост опухоли молочной железы (Lapteva, Yang et al. 2005); T-гибридомные клетки, трансфицированные конструкцией, предотвращающей экспрессию CXCR4 на поверхности, не могли далее метастазировать к отдаленным органам, будучи внутривенно введены мышам (Zeelenberg, Ruuls-Van Stalle et al. 2001); в похожих экспериментах с клетками рака ободочной и прямой кишки, метастазы в легких и печени значительно уменьшались (Zeelenberg, Ruuls-Van Stalle et al. 2003); анти-CXCR4 антитела ингибировали распространение ксенотрансплантатов рака молочной железы в лимфоузлы (Muller, Homey et al. 2001); обработка лимфобластных клеток анти-CXCR4 или анти-SDF-1 антителами задерживала рост опухоли у (NOD)/SCID мышей (Bertolini, Dell'Agnola et al. 2002); анти-SDF-1 антитела ингибировали развитие в органе метастазов клеток немелкоклеточного рака легких (сокр. NSCLC) (Phillips, Burdick et al. 2003); системное введение антагониста CXCR4, AMD3100 (при помощи AnorMED Inc.), ингибировало рост ксенотрансплантатов внутричерепной глиобластомы и медуллобластомы, а также повышенный апоптоз опухолевых клеток в течение 24 часов (Rubin, Kung et al. 2003); анти-SDF-1 антитела ингибировали рост MCF-7 клеток рака молочной железы с примесями связанных с карциномой фибробластов (Orimo, Gupta et al. 2005); нейтрализация CXCR4 антителами блокировала метастазирование рака предстательной железы и рост костных опухолей (Sun, Schneider et al. 2005); развитие метастазов в легких после инъекции клеток остеосаркомы было предупреждено введением T134 – пептидного антагониста CXCR4 (Perissinotto, Cavalloni et al. 2005).
Различные авторы приходят к выводу, что адресное воздействие на систему SDF-1/CXCR4 предоставляет раковым больным новые терапевтические возможности:
Человеческие опухоли яичников в значительной степени экспрессируют SDF-1 плюс, - на более низком уровне, - VEGF. Появление обоих белков вызвано гипоксией в опухоли. Патологические концентрации любого из этих белков в отдельности не были достаточны, чтобы вызвать ангиогенез in vivo, но SDF-1 и VEGF вместе, в патологических концентрациях, эффективно и синергистически вызывали неоваскуляризацию. Таким образом, нарушение синергии в этой системе, а не один лишь VEGF, является новой эффективной антиангиогенной стратегией лечения рака (Kryczek, Lange et al. 2005);
Клеточные линии рака молочной железы, наделенные аутокринным SDF-1/CXCR4-путем передачи сигнала, показывают агрессивное поведение. Это включает увеличение инвазивности и миграции, наряду с более быстрым ростом. Таким образом, система SDF-1/CXCR4 предоставляет важную информацию для прогнозирования агрессивности и устанавливает важные цели для терапии человеческого рака молочной железы (Kang, Watkins et al. 2005);
Миграция и метастазирование клеток мелкоклеточного рака легких (сокр. SCLC), – которые экспрессируют высокие уровни CXCR4, – регулируются с помощью SDF-1. Активация CXCR4 способствует адгезии к дополнительным клеткам (таким как стромальные клетки) и внеклеточным матричным молекулам в пределах микроокружения опухоли. Эти адгезионные взаимодействия приводят к повышенной резистентности клеток SCLC к химиотерапии. По этой причине ингибиторы системы SDF-1/CXCR4 также увеличивают чувствительность к химиотерапии клеток SCLC и приводят к новым направлениям в терапии пациентов с SCLC (Hartmann, Burger et al. 2004) и другими опухолями.
Хемокин-рецепторная передача сигнала и направленная миграция стволовых клеток. Система SDF-1/CXCR4 возникает как центральный регулятор направленной миграции различных типов стволовых клеток в организме. Поскольку большая часть, если не все, злокачественные новообразования берут начало в клеточных компартментах стволовых клеток/клеток-предшественников, раковые стволовые клетки также экспрессируют CXCR4 на своей поверхности и, в результате, заставляют систему SDF-1/CXCR4 участвовать в наведении их направленной миграции/метастазирования на органы, экспрессирующие SDF-1, например, такие как лимфоузлы, легкие, печень и кости. Вследствие этого у стратегий, нацеленных на модуляцию системы SDF-1/CXCR4, есть важные клинические применения как в восстановительной медицине, чтобы доставлять нормальные стволовые клетки к тканям, так и в клинической онкологии, чтобы ингибировать метастазирование раковых стволовых клеток (Kucia, Reca et al. 2005).
Мобилизация стволовых клеток. Лейкоциты, также известные как белые кровяные клетки, включают нейтрофилы, макрофаги, эозинофилы, базофилы/тучные клетки, B-клетки и T-клетки. Белые кровяные клетки непрерывно замещаются через систему гемопоэза, под действием колониестимулирующих факторов (CSF) и различных цитокинов на стволовые клетки и клетки-предшественники в гемопоэтических тканях. Наиболее широко известным из этих факторов является гранулоцитарный колониестимулирующий фактор (сокр. G-CSF), который был одобрен для использования как средство нейтрализации отрицательного воздействия химиотерапии, стимулируя производство лейкоцитов и клеток-предшественников (мобилизацию стволовых клеток периферической крови). Существует целый ряд антигенов клеточной поверхности, которые используются как маркеры для характеристики популяций стволовых клеток и клеток-предшественников. Эти маркеры также подлежат уточнению всякий раз, когда обнаруживаются новые, более специфичные маркеры. Гемопоэтические стволовые клетки в настоящее время характеризуются как являющиеся CD34+, c-kit+, Sca-1+, CD45+, lin- и CD38- (CD38 является также маркером клеточной линии, поэтому он не имеет отношения к lin-). Костный мозг также является носителем для некоторых других типов стволовых клеток, которые не являются гемопоэтическими, но могут дать начало другим типам клеток и тканей. Мезенхимные стволовые клетки характеризуются как CD34+, Sca-1+, lin-, BMPR+ и/или STRO-1+, тканеспецифичные стволовые клетки костного мозга: в настоящее время определяются как являющиеся CXCR4+, CD34+, CD45–. Субпопуляции тканеспецифичных стволовых клеток костного мозга включают (Majka et al. 2005):
скелетные стволовые клетки: Myf5+, MyoD+
кардиальные стволовые клетки: NKx2.5+, GATA4+
печеночные стволовые клетки: CK19+, α-фетопротеин+
нейронные стволовые клетки: нестин+, GATA4+
Сообщалось, что некоторые другие факторы повышают содержание белых кровяных клеток и клеток-предшественников как у людей, так и у животных. Эти средства включают гранулоцит-макрофаговый колониестимулирующий фактор (сокр. GM-CSF), интерлейкин-1 (сокр. IL-1), интерлейкин-3 (сокр. IL-3), интерлейкин-8 (сокр. IL-8), PIXY-321 (сокр. GM-CSF/IL-3 химерный белок), воспалительный белок макрофага (сокр. MIP), GROβ (CXCL2) и GROβT (CXCL2Δ4), фактор стволовых клеток, тромбопоэтин и ростовой онкоген, в виде индивидуальных средств, либо в комбинации (Broxmeyer, Benninger et al. 1995; Glaspy, Даvis et al. 1996; Rosenfeld, Bolwell et al. 1996; Glaspy, Shpall et al. 1997; Vadhan-Raj, Murray et al. 1997; Broxmeyer, Orazi et al. 1998; Dаle, Liles et al. 1998; Pruijt, Willemze et al. 1999; King, Horowitz et al. 2001).
Хотя эндогенные факторы роста фармакологически эффективны, хорошо известные недостатки белков и пептидов, применяемых в качестве фармацевтических препаратов, подчеркивают необходимость добавления к набору таких факторов роста дополнительных средств, которые эффективны до той же степени, то есть которые повышают содержание, соответственно, клеток-предшественников лейкоцитов и стволовых клеток, предпочтительно - повышают их уровень в периферической крови субъекта. Соответственно, одна проблема, приоритетная для настоящей заявки, состоит в том, чтобы предоставить средства и способы, которые повышают содержание клеток-предшественников лейкоцитов и стволовых клеток, соответственно, более конкретно - повышают их уровень в периферической крови субъекта. Дальнейшая проблема, приоритетная для настоящей заявки, состоит в том, чтобы предоставить средства и способы лечения заболеваний, которые вызваны или связаны с низким уровнем клеток-предшественников лейкоцитов и стволовых клеток, соответственно.
Стволовые клетки либо мобилизировали, чтобы напрямую запустить репарацию поврежденных тканей у того же самого пациента, у которого они были мобилизированы, либо их мобилизировали и брали у донора, совместимого по человеческому лейкоцитарному антигену (HLA), и вводили пациенту либо внутривенно, либо непосредственно в пораженную ткань. Последнее может быть также проделано со стволовыми клетками, которые были мобилизированы у самого пациента. Перед введением стволовых клеток они могут быть размножены и/или дифференцированы in vitro.
Аллергические заболевания дыхательных путей и контактные аллергии. Было обнаружено, что SDF-1 действует как хемотаксическое средство на зрелые тучные клетки и их предшественников, – особенно когда гистамин высвобождается из зрелых тучных клеток, например, с помощью опосредованной IgE передачи сигнала посредством связывания с Fcε-рецептором на поверхности тучных клеток (Godot, Arock et al. 2007). При моделировании аллергии дыхательных путей на мышах опосредованная антителами нейтрализация CXCR4, экспрессируемого лейкоцитами, как указывалось выше, снижала гиперреактивность дыхательных путей. Антитела также наполовину снижали эозинофилию легких, особенно в бронхоальвеолярной промывной жидкости и интерстициальной ткани, указывая, что CXCR4-опосредованные сигналы способствуют воспалению легкого. Нейтрализация SDF-1α приводит к похожему ослаблению как аллергического воспаления легких, так и гиперреактивности дыхательных путей (Gonzalo, Lloyd et al. 2000). Есть также свидетельство, что SDF-1 способствует ангиогенезу. Это было явно показано Хошино и сотрудниками при астме, путем анализа бронхиальных биопсий на предмет ангиогенеза и экспрессии SDF-1. Иммуногистохимический анализ на срезах этих биопсий показал, что у субъектов больных астмой была более высокая степень васкуляризации и большее число SDF-1-положительных клеток по сравнению с контрольными субъектами (Hoshino, Aoike et al. 2003).
Кроме того, клинические и экспериментальные данные указывают, что инфильтрирующие кожу лейкоциты играют важную роль в инициировании и лечении атопического дерматита, и было показано, что SDF-1 представляет собой важный фактор рекрутинга T-лимфоцитов и дендритных клеток, особенно – клеток типа дендритных клеток Лангерганса (Gombert, Dieu-Nosjean et al. 2005).
Псориаз. Псориаз представляет собой воспалительное кожное заболевание с преимущественно аутоиммунным компонентом. Псориаз характеризуется сильной лейкоцитарной инфильтрацией пораженной кожи, при этом T-клетки играют заметную роль. Чжоу и сотрудники нашли среди прочего повышенную экспрессию мРНК SDF-1 в псориазных повреждениях кожи (Zhou, Krueger et al. 2003).
Воспаление суставов. В литературе есть свидетельство причастности системы SDF-1-CXCL4 к воспалению суставов. Маттис и сотр. показали, что AMD3100, сильный и специфичный антагонист CXCR4, ингибировал аутоиммунное воспаление суставов у мышей с врожденным отсутствием IFNγ-рецепторов (Matthys, Hatse et al. 2001). Экспрессию SDF-1 также наблюдали в синовиальных биопсиях и с помощью RT-PCR (полимеразной цепной реакции в реальном масштабе времени) у субъектов, страдающих спондилоартропатией, ревматоидным артритом, псориатическим артритом и дегенеративным заболеванием суставов (остеоартритом). Однако во всех случаях не наблюдали суперэкспрессии (Gu, Marker-Hermann et al. 2002). Сходные результаты были получены для CXCR4 с помощью RT-PCR.
Ревматоидный артрит. Совсем недавно было обнаружено, что уровни SDF-1 в синовиальной жидкости у пациентов с ревматоидным артритом были повышены по сравнению с пациентами с остеоартритом (Kim, Cho et al. 2007). Авторы также описали, что в клеточной культуре экспрессия SDF-1 у подобных фибробластам синовиоцитов была активирована совместным культивированием этих клеток с T-клетками. Этот эффект также наблюдался при добавлении IL-17, - T-клеточного цитокина, - к культуральной среде.
Иммуногистохимический анализ на срезах биопсий суставов показал, что SDF-1 экспрессируется в синовиальной оболочке суставов, затронутых псориатическим артритом. Быстрое и существенное клиническое улучшение наблюдалось после лечения инфликсимабом у всех 9 пациентов. Это сопровождалось снижением синовиальных уровней SDF-1 среди других факторов роста (биопсии были взяты после 8 недель терапии) (Gu, Marker-Hermann et al. 2002).
Хотя существуют описания того, что лейкоцитарная инфильтрация является характерной особенностью аллергических заболеваний, аллергических реакций и воспалительных процессов при аутоиммунных заболеваниях, пока не разработано никакого эффективного лечения таких заболеваний. На настоящий момент необходимы новейшие средства, влияющие на течение таких болезней и, предпочтительно, влияющие на лейкоцитарную инфильтрацию. Соответственно, дальнейшая проблема, приоритетная для настоящей заявки, состоит в том, чтобы предоставить средства и способы для ингибирования или сокращения лейкоцитарной инфильтрации в ткани. Дальнейшая проблема, приоритетная для настоящей заявки, состоит в том, чтобы предоставить средства и способы лечения заболеваний, которые вызваны или связаны с лейкоцитарной инфильтрацией в ткани или с повышенным уровнем инфильтрации таких лейкоцитов в ткани.
Передача сигнала в системе SDF-1 и его рецептора влияет на миграцию клеток внутри организма, предпочтительно, из одной ткани - в другую ткань, из ткани - в периферическую кровь и/или из периферической крови - в ткань, приводя к некоторым заболеваниям и нарушениям. Специфическое вмешательство, предпочтительно - ингибирование взаимодействия между SDF-1 и рецептором или рецепторами SDF-1, может вызвать улучшение течения некоторых заболеваний и нарушений. Ввиду вышеизложенного, еще одна дальнейшая, более общая проблема, приоритетная для настоящей заявки, состоит в том, чтобы предоставить средства и способы, влияющие на миграцию клеток внутри организма, предпочтительно из одной ткани в другую ткань, из ткани - в периферическую кровь и/или из периферической крови - в ткань, посредством чего такая миграция приводит к некоторым заболеваниям и нарушениям или связана с ними. На настоящий момент дальнейшая проблема, приоритетная для настоящей заявки, состоит в том, чтобы предоставить средства и способы, которые вызваны миграцией клеток внутри организма или связаны с ней, предпочтительно из одной ткани в другую ткань, из ткани - в периферическую кровь и/или из периферической крови - в ткань, приводя к некоторым заболеваниям и нарушениям.
Решение проблемы, приоритетной для настоящего изобретения, отражено в независимых пунктах формулы изобретения. Предпочтительные варианты осуществления могут быть взяты из зависимых пунктов формулы изобретения.
Конкретнее, проблема, приоритетная для настоящей заявки, решена в первом аспекте с помощью связывания молекулы нуклеиновой кислоты с SDF-1, где молекула нуклеиновой кислоты оказывает влияние на миграцию клеток.
В первом варианте осуществления первого аспекта клетки экспрессируют рецептор SDF, где рецептор SDF-1 является одним, предпочтительно выбранным из CXCR4 и CXCR7.
Во втором варианте осуществления первого аспекта, который также является вариантом осуществления первого варианта осуществления первого аспекта, миграция клеток включает мобилизацию клеток-предшественников, стволовых клеток, раковых клеток, долгоживущих плазмоцитов, B-клеток и/или T-клеток в периферическую кровь субъекта, где, предпочтительно, данные B-клетки и/или T-клетки являются В-клетками памяти и/или T-клетками памяти.
В третьем варианте осуществления первого аспекта, который также является вариантом осуществления второго варианта осуществления первого аспекта, клетки-предшественники и/или стволовые клетки включают CD34+ клетки-предшественники.
В четвертом варианте осуществления первого аспекта, который также является вариантом осуществления второго и третьего вариантов осуществления первого аспекта, мобилизация клеток-предшественников и/или стволовых клеток происходит в гемопоэтической ткани.
В пятом варианте осуществления первого аспекта, который также является вариантом осуществления четвертого варианта осуществления первого аспекта, данной гемопоэтической тканью является, по меньшей мере, одна либо миелоидная ткань, либо лимфоидная ткань, где, предпочтительно, данная миелоидная ткань расположена в костном мозге, и, предпочтительно, данная лимфоидная ткань расположена в слизистой оболочке пищеварительного тракта, дыхательном пути, лимфоузлах, селезенке, тимусе и/или лимфоидных фолликулах в воспаленной ткани.
В шестом варианте осуществления первого аспекта молекула нуклеиновой кислоты ингибирует миграцию лейкоцитов.
В седьмом варианте осуществления первого аспекта, который также является вариантом осуществления шестого варианта осуществления первого аспекта, данные лейкоциты представляют собой T-лимфоциты, B-лимфоциты, моноциты, макрофаги, эозинофилы, нейтрофилы, базофилы, дендритные клетки и/или тучные клетки.
В восьмом варианте осуществления первого аспекта, который также является вариантом осуществления шестого и седьмого вариантов осуществления первого аспекта, в процессе миграции лейкоцитов данные лейкоциты аккумулируются в ткани, где, предпочтительно, данная аккумуляция лейкоцитов приводит к воспалению указанной ткани.
В девятом варианте осуществления первого аспекта, который также является вариантом осуществления восьмого варианта осуществления первого аспекта, данная ткань включает кожу, слизистую оболочку, органы, выбираемые, не ограничиваясь только ими, из глаз, мозга, легких, почек, сердца, печени, желудочно-кишечного тракта, селезенки, костей и/или лимфатической системы, предпочтительно кожи и/или слизистой оболочки дыхательных путей.
В десятом варианте осуществления первого аспекта, который также является вариантом осуществления первого аспекта и любого из вариантов, от первого до девятого, осуществления первого аспекта, молекулу нуклеиновой кислоты выбирают из группы, включающей молекулы нуклеиновых кислот типа A, молекулы нуклеиновых кислот типа B, молекулы нуклеиновых кислот типа C и молекулы нуклеиновых кислот, имеющих последовательности нуклеиновых кислот согласно любой из SEQ ID NO:142, SEQ ID NO:143 и SEQ ID NO:144.
В одиннадцатом варианте осуществления первого аспекта, который также является вариантом осуществления десятого варианта осуществления первого аспекта, молекулы нуклеиновых кислот типа A содержат следующую коровую нуклеотидную последовательность:
5’ AAAGYRACAHGUMAAXAUGAAAGGUARC 3’ (SEQ ID NO:19)
где XA либо отсутствует, либо представляет собой A.
В двенадцатом варианте осуществления первого аспекта, который также является вариантом осуществления одиннадцатого варианта осуществления первого аспекта, молекулы нуклеиновых кислот типа A содержат коровую нуклеотидную последовательность, выбранную из группы, включающей
5’ AAAGYRACAHGUMAAUGAAAGGUARC 3’ (SEQ ID NO:20),
5’ AAAGYRACAHGUMAAAUGAAAGGUARC 3’ (SEQ ID NO:21) и
5’ AAAGYAACAHGUCAAUGAAAGGUARC 3’ (SEQ ID NO:22),
предпочтительно, коровая нуклеотидная последовательность включает 5’ AAAGYAACAHGUCAAUGAAAGGUARC 3’ (SEQ ID NO:22).
В 13м варианте осуществления первого аспекта, который также является вариантом осуществления одиннадцатого и двенадцатого вариантов осуществления первого аспекта, молекула нуклеиновой кислоты содержит в 5’->3’ направлении первый участок нуклеотидов, коровую нуклеотидную последовательность и второй участок нуклеотидов.
В 14м варианте осуществления первого аспекта, который также является вариантом осуществления одиннадцатого и двенадцатого вариантов осуществления первого аспекта, молекула нуклеиновой кислоты включает в 5’->3’ направлении второй участок нуклеотидов, коровую нуклеотидную последовательность и первый участок нуклеотидов.
В 15м варианте осуществления первого аспекта, который также является вариантом осуществления 13го и 14го вариантов осуществления первого аспекта, молекула нуклеиновой кислоты включает первый и второй участок нуклеотидов, а указанный первый и указанный второй участок нуклеотидов дополнительно гибридизуют друг с другом, где в процессе гибридизации формируется двухцепочечная структура.
В 16м варианте осуществления первого аспекта, который также является вариантом осуществления любого из вариантов, от 13го до 15го, осуществления первого аспекта, данная структура двойной цепи содержит от четырех до шести пар оснований, предпочтительно, - пять пар оснований.
В 17м варианте осуществления первого аспекта, который также является вариантом осуществления любого из вариантов, от 13го до 16го, осуществления первого аспекта, первый участок нуклеотидов включает нуклеотидную последовательность 5’ X1X2NNBV 3’ (SEQ ID NO:44), а второй участок нуклеотидов включает нуклеотидную последовательность 5’ BNBNX3X4 3’ (SEQ ID NO:45),
где X1 либо отсутствует, либо представляет собой R, X2 представляет собой S, X3 представляет собой S и X4 либо отсутствует, либо представляет собой Y;
или
X1 отсутствует, X2 либо отсутствует, либо представляет собой S, X3 либо отсутствует, либо представляет собой S и X4 отсутствует.
В 18м варианте осуществления первого аспекта, который также является вариантом осуществления любого из вариантов, от 13го до 17го, осуществления первого аспекта, первый участок нуклеотидов включает нуклеотидную последовательность 5’ RSHRYR 3’ (SEQ ID NO:23), а второй участок нуклеотидов включает нуклеотидную последовательность 5’ YRYDSY 3’ (SEQ ID NO:24),
предпочтительно, первый участок нуклеотидов включает нуклеотидную последовательность 5’ GCUGUG 3’, а второй участок нуклеотидов включает нуклеотидную последовательность 5’ CGCAGC 3’.
В 19м варианте осуществления первого аспекта, который также является вариантом осуществления любого из вариантов, от 13го до 17го, осуществления первого аспекта, первый участок нуклеотидов включает нуклеотидную последовательность 5’ X2BBBS 3’ (SEQ ID NO:42), а второй участок нуклеотидов включает нуклеотидную последовательность 5’ SBBVX3 3’ (SEQ ID NO:43),
где X2 либо отсутствует, либо представляет собой S и X3 либо отсутствует, либо представляет собой S;
предпочтительно, первый участок нуклеотидов включает нуклеотидную последовательность 5’ CUGUG 3’, а второй участок нуклеотидов включает нуклеотидную последовательность 5’ CGCAG 3’;
или первый участок нуклеотидов включает нуклеотидную последовательность 5’ GCGUG 3’, а второй участок нуклеотидов включает нуклеотидную последовательность 5’ CGCGC 3’.
В 20м варианте осуществления первого аспекта, который также является вариантом осуществления любого из вариантов, от 11го до 19го, осуществления первого аспекта, молекула нуклеиновой кислоты имеет последовательность нуклеиновой кислоты согласно любой из SEQ ID NO:5-18, 25-41, 133, 137, 139-141.
В 21м варианте осуществления первого аспекта, который также является вариантом осуществления десятого варианта осуществления первого аспекта, молекулы нуклеиновых кислот типа B содержат следующую коровую нуклеотидную последовательность:
5’ GUGUGAUCUAGAUGUADWGGCUGWUCCUAGUYAGG 3’ (SEQ ID NO:57).
В 22м варианте осуществления первого аспекта, который также является вариантом осуществления 21го варианта осуществления первого аспекта, молекулы нуклеиновых кислот типа B включают коровую нуклеотидную последовательность:
GUGUGAUCUAGAUGUADUGGCUGAUCCUAGUCAGG (SEQ ID NO:58).
В 23м варианте осуществления первого аспекта, который также является вариантом осуществления любого из вариантов, от 21го до 22го, осуществления первого аспекта, молекула нуклеиновой кислоты включает в 5’->3’ направлении первый участок нуклеотидов, коровую нуклеотидную последовательность и второй участок нуклеотидов.
В 24м варианте осуществления первого аспекта, который также является вариантом осуществления любого из вариантов, от 21го до 22го, осуществления первого аспекта, молекула нуклеиновой кислоты включает в 5’->3’ направлении второй участок нуклеотидов, коровую нуклеотидную последовательность и первый участок нуклеотидов.
В 25м варианте осуществления первого аспекта, который также является вариантом осуществления любого из вариантов, от 23го до 24го, осуществления первого аспекта, молекула нуклеиновой кислоты включает первый и второй участок нуклеотидов, а указанный первый и указанный второй участок нуклеотидов дополнительно гибридизуют друг с другом, где в процессе гибридизации формируется двухцепочечная структура.
В 26м варианте осуществления первого аспекта, который также является вариантом осуществления любого из вариантов, от 21го до 25го, осуществления первого аспекта, данная структура двойной цепи содержит от четырех до шести пар оснований, предпочтительно, - пять пар оснований.
В 27м варианте осуществления первого аспекта, который также является вариантом осуществления любого из вариантов, от 23го до 26го, осуществления первого аспекта, первый участок нуклеотидов включает нуклеотидную последовательность 5’ X1X2SVNS 3’ (SEQ ID NO:77), а второй участок нуклеотидов включает нуклеотидную последовательность 5’ BVBSX3X4 3’ (SEQ ID NO:78), где
X1 либо отсутствует, либо представляет собой A, X2 представляет собой G, X3 представляет собой C и X4 либо отсутствует, либо представляет собой U;
или
X1 отсутствует, X2 либо отсутствует, либо представляет собой G, X3 либо отсутствует, либо представляет собой C и X4 отсутствует.
В 28м варианте осуществления первого аспекта, который также является вариантом осуществления любого из вариантов, от 23го до 27го, осуществления первого аспекта, первый участок нуклеотидов включает нуклеотидную последовательность 5’ X1GCRWG 3’ (SEQ ID NO:59), а второй участок нуклеотидов включает нуклеотидную последовательность 5’ KRYSCX4 3’(SEQ ID NO:60),
где X1 либо отсутствует, либо представляет собой A и X4 либо отсутствует, либо представляет собой U.
В 29м варианте осуществления первого аспекта, который также является вариантом осуществления любого из вариантов, от 23го до 28го, осуществления первого аспекта, первый участок нуклеотидов включает нуклеотидную последовательность 5’ X1GCGUG 3’ (SEQ ID NO:75), а второй участок нуклеотидов включает нуклеотидную последовательность 5’ UACGCX4 3’ (SEQ ID NO:76),
где X1 либо отсутствует, либо представляет собой A и X4 либо отсутствует, либо представляет собой U,
предпочтительно, первый участок нуклеотидов включает нуклеотидную последовательность 5’ AGCGUG 3’, а второй участок нуклеотидов включает нуклеотидную последовательность 5’ UACGCU 3’.
В 30м варианте осуществления первого аспекта, который также является вариантом осуществления любого из вариантов, от 23го до 27го, осуществления первого аспекта, первый участок нуклеотидов включает нуклеотидную последовательность 5’ X2SSBS 3’ (SEQ ID NO:73), а второй участок нуклеотидов включает нуклеотидную последовательность 5’ BVSSX3 3’ (SEQ ID NO:74),
где X2 либо отсутствует, либо представляет собой G и X3 либо отсутствует, либо представляет собой C,
предпочтительно, первый участок нуклеотидов включает нуклеотидную последовательность 5’ GCGUG 3’, а второй участок нуклеотидов включает нуклеотидную последовательность 5’ UACGC 3’.
В 31м варианте осуществления первого аспекта, который также является вариантом осуществления любого из вариантов, от 21го до 30го, осуществления первого аспекта, молекула нуклеиновой кислоты имеет последовательность нуклеиновой кислоты согласно любой из SEQ ID NO:46-56, 61-72 и 132.
В 32м варианте осуществления первого аспекта, который также является вариантом осуществления десятого варианта осуществления первого аспекта, молекулы нуклеиновых кислот типа C включают коровую нуклеотидную последовательность GGUYAGGGCUHRXAAGUCGG (SEQ ID NO:90),
где XA либо отсутствует, либо представляет собой A.
В 33м варианте осуществления первого аспекта, который также является вариантом осуществления 32го варианта осуществления первого аспекта, молекулы нуклеиновых кислот типа C включают коровую нуклеотидную последовательность, выбранную из группы, включающей
5’ GGUYAGGGCUHRAAGUCGG 3’ (SEQ ID NO:91),
5’ GGUYAGGGCUHRAGUCGG 3’ (SEQ ID NO:92), и
5’ GGUUAGGGCUHGAAGUCGG 3’ (SEQ ID NO:93), предпочтительно, коровая нуклеотидная последовательность включает 5’ GGUUAGGGCUHGAAGUCGG 3’ (SEQ ID NO:93).
В 34м варианте осуществления первого аспекта, который также является вариантом осуществления любого из вариантов, от 32го до 33го, осуществления первого аспекта, молекула нуклеиновой кислоты включает в 5’->3’ направлении первый участок нуклеотидов, коровую нуклеотидную последовательность и второй участок нуклеотидов.
В 35м варианте осуществления первого аспекта, который также является вариантом осуществления любого из вариантов, от 32го до 33го, осуществления первого аспекта, молекула нуклеиновой кислоты включает в 5’->3’ направлении второй участок нуклеотидов, коровую нуклеотидную последовательность и первый участок нуклеотидов.
В 36м варианте осуществления первого аспекта, который также является вариантом осуществления любого из вариантов, от 34го до 35го, осуществления первого аспекта, молекула нуклеиновой кислоты включает первый и второй участок нуклеотидов и где по меньшей мере часть указанного первого участок нуклеотидов и по меньшей мере часть указанного второго участок нуклеотидов дополнительно гибридизуют друг с другом, где в процессе гибридизации формируется двухцепочечная структура.
В 37м варианте осуществления первого аспекта, который также является вариантом осуществления любого из вариантов, от 34го до 36го, осуществления первого аспекта, длина первого участка нуклеотидов и длина второго участка нуклеотидов по отдельности и независимо составляет от 0 до 17 нуклеотидов, предпочтительно - от 4 до 10 нуклеотидов и более предпочтительно – от 4 до 6 нуклеотидов.
В 38м варианте осуществления первого аспекта, который также является вариантом осуществления любого из вариантов, от 36го до 37го, осуществления первого аспекта, структура двойной цепи включает от 4 до 10 пар оснований, предпочтительно - от 4 до 6 пар оснований, более предпочтительно - 5 пар оснований.
В 39м варианте осуществления первого аспекта, который также является вариантом осуществления 38го варианта осуществления первого аспекта, структура двойной цепи включает от 4 до 10 идущих подряд пар оснований, предпочтительно - от 4 до 6 идущих подряд пар оснований, более предпочтительно - 5 идущих подряд пар оснований.
В 40м варианте осуществления первого аспекта, который также является вариантом осуществления любого из вариантов, от 34го до 39го, осуществления первого аспекта, первый участок нуклеотидов включает нуклеотидную последовательность 5’ RKSBUSNVGR 3’ (SEQ ID NO:120), а второй участок нуклеотидов включает нуклеотидную последовательность 5’ YYNRCASSMY 3’ (SEQ ID NO:121),
предпочтительно, первый участок нуклеотидов включает нуклеотидную последовательность 5’ RKSBUGSVGR 3’ (SEQ ID NO:122), а второй участок нуклеотидов включает нуклеотидную последовательность 5’ YCNRCASSMY 3’ (SEQ ID NO:123).
В 41м варианте осуществления первого аспекта, который также является вариантом осуществления любого из вариантов, от 34го до 39го, осуществления первого аспекта, первый участок нуклеотидов включает нуклеотидную последовательность 5’ XSSSSV 3’ (SEQ ID NO:124), а второй участок нуклеотидов включает нуклеотидную последовательность 5’ BSSSXS 3’ (SEQ ID NO:125), где Xs либо отсутствует, либо представляет собой S.
В 42м варианте осуществления первого аспекта, который также является вариантом осуществления любого из вариантов, от 34го до 39го и 41го, осуществления первого аспекта, первый участок нуклеотидов включает нуклеотидную последовательность 5’ SSSSR 3’ (SEQ ID NO:130), а второй участок нуклеотидов включает нуклеотидную последовательность 5’ YSBSS 3’ (SEQ ID NO:131),
предпочтительно, первый участок нуклеотидов включает нуклеотидную последовательность 5’ SGGSR 3’ (SEQ ID NO:126), а второй участок нуклеотидов включает нуклеотидную последовательность 5’ YSCCS 3’ (SEQ ID NO:127).
В 43м варианте осуществления первого аспекта, который также является вариантом осуществления любого из вариантов, от 34го до 39го, 41го и 42го, осуществления первого аспекта, первый участок нуклеотидов включает нуклеотидную последовательность 5’ GCSGG 3’ (SEQ ID NO:128), а второй участок нуклеотидов включает нуклеотидную последовательность 5’ CCKGC 3’ (SEQ ID NO:129),
предпочтительно, первый участок нуклеотидов включает нуклеотидную последовательность 5’ GCCGG 3’, а второй участок нуклеотидов включает нуклеотидную последовательность 5’ CCGGC 3’.
В 44м варианте осуществления первого аспекта, который также является вариантом осуществления любого из вариантов, от 34го до 39го, осуществления первого аспекта, первый участок нуклеотидов включает нуклеотидную последовательность 5’ CGUGCGCUUGAGAUAGG 3’, а второй участок нуклеотидов включает нуклеотидную последовательность 5’ CUGAUUCUCACG 3’.
В 45м варианте осуществления первого аспекта, который также является вариантом осуществления любого из вариантов, от 34го до 39го, осуществления первого аспекта, первый участок нуклеотидов включает нуклеотидную последовательность 5’ UGAGAUAGG 3’, а второй участок нуклеотидов включает нуклеотидную последовательность 5’ CUGAUUCUCA 3’.
В 46м варианте осуществления первого аспекта, который также является вариантом осуществления любого из вариантов, от 34го до 39го, осуществления первого аспекта, первый участок нуклеотидов включает нуклеотидную последовательность 5’ GAGAUAGG 3’, а второй участок нуклеотидов включает нуклеотидную последовательность 5’ CUGAUUCUC 3’.
В 47м варианте осуществления первого аспекта, который также является вариантом осуществления любого из вариантов, от 32го до 46го, осуществления первого аспекта, молекула нуклеиновой кислоты имеет последовательность нуклеиновой кислоты согласно любой из SEQ ID NO:79-89, 94-119 и 134-136.
В 48м варианте осуществления первого аспекта, который также является вариантом осуществления десятого варианта осуществления первого аспекта, молекула нуклеиновой кислоты имеет последовательность нуклеиновой кислоты согласно любой из SEQ ID NO:142-144.
В 49м варианте осуществления первого аспекта, который также является вариантом осуществления первого аспекта и любого из вариантов, от первого до 48го, осуществления первого аспекта, молекула нуклеиновой кислоты является антагонистом SDF-1.
В 50м варианте осуществления первого аспекта, который также является вариантом осуществления первого аспекта и любого из вариантов, от первого до 48го, осуществления первого аспекта, молекула нуклеиновой кислоты является антагонистом в рецепторной системе SDF-1, где рецептором SDF-1 в рецепторной системе SDF-1 является какой-либо один, предпочтительно выбранный из CXCR4 и CXCR7.
В 51м варианте осуществления первого аспекта, который также является вариантом осуществления первого аспекта и любого из вариантов, от первого до 50го, осуществления первого аспекта, SDF-1 представляет собой SDF-1 человека и/или рецептор SDF-1 рецепторной системы SDF-1 представляет собой рецептор SDF-1 человека.
В 52м варианте осуществления первого аспекта, который также является вариантом осуществления первого аспекта и любого из вариантов, от первого до 51го, осуществления первого аспекта, SDF-1 включает аминокислотную последовательность согласно SEQ ID NO:1.
В 53м варианте осуществления первого аспекта, который также является вариантом осуществления первого аспекта и любого из вариантов, от первого до 52го, осуществления первого аспекта, нуклеиновая кислота включает модификацию.
В 54м варианте осуществления первого аспекта, который также является вариантом осуществления 53го варианта осуществления первого аспекта, данную модификацию выбирают из группы, включающей остаток HES и остаток PEG.
В 55м варианте осуществления первого аспекта, который также является вариантом осуществления 54го варианта осуществления первого аспекта, данная модификация представляет собой остаток PEG, содержащий линейный или разветвленный PEG, где молекулярная масса остатка PEG предпочтительно составила от примерно 2 до 180 кДа, более предпочтительно - от примерно 60 до 140 кДа и наиболее предпочтительно – примерно 40 кДа.
В 56м варианте осуществления первого аспекта, который также является вариантом осуществления 54го варианта осуществления первого аспекта, данная модификация представляет собой остаток HES, где молекулярная масса остатка HES предпочтительно составила от примерно 10 до 130 кДа, более предпочтительно - от примерно 30 до 130 кДа и наиболее предпочтительно – примерно 100 кДа.
В 57м варианте осуществления первого аспекта, который также является вариантом осуществления первого аспекта и любого из вариантов, от первого до 56го, осуществления первого аспекта, нуклеотиды данной нуклеиновой кислоты представляют собой L-нуклеотиды, предпочтительно, - нуклеотиды последовательностей, согласно любой из SEQ ID NO:19, 20, 21, 22, 57, 58, 90, 91, 92 и 93.
Проблема, приоритетная для настоящей заявки, решена во втором аспекте с помощью фармацевтической композиции, содержащей нуклеиновую кислоту согласно первому аспекту и любым вариантам их осуществления, и необязательно - по меньшей мере один дополнительный компонент, где данный дополнительный компонент выбирают из группы, включающей фармацевтически приемлемые вспомогательные средства и фармацевтически активные средства.
Проблема, приоритетная для настоящей заявки, решена в третьем аспекте с помощью использования нуклеиновой кислоты согласно первому аспекту и любым вариантам их осуществления для производства лекарственного средства.
В первом варианте осуществления третьего аспекта данное лекарственное средство использовали для мобилизации клеток-предшественников и/или стволовых клеток в периферическую кровь, и/или для лечения заболеваний и/или нарушений, предпочтительно выбранных из группы, включающей заживление ран; ожоги; нарушения, вызванные или связанные с поврежденной тканью органа, и/или поврежденной сосудистой системой, где такие нарушения отобраны из повреждений сетчатки и сосудистой оболочки глаза, инсульта, поражения миокарда, инфаркта миокарда, ишемии после трансплантации органа и травматического повреждения; и гемопоэтические нарушения, где такие нарушения отобраны из апластической анемии, лейкоза, лекарственной анемии и лейкопении, а также бактериальной инфекции при лейкопении.
Во втором варианте осуществления третьего аспекта данное лекарственное средство предназначено для мобилизации раковых клеток в периферическую кровь субъекта.
В третьем варианте осуществления третьего аспекта, который также является вариантом осуществления второго варианта осуществления третьего аспекта, данные раковые клетки отобраны из лейкозных клеток, клеток лимфомы, раковых стволовых клеток, раковых клеток с метастатическим потенциалом и раковых метастазов.
В четвертом варианте осуществления третьего аспекта, который также является вариантом осуществления любого из вариантов, от второго до третьего, осуществления третьего аспекта, данное лекарственное средство применяется в сочетании со вторым фармацевтически активным средством, где второе фармацевтически активное средство пригодно для мобилизации раковых клеток в периферическую кровь субъекта, где второе фармацевтически активное средство предпочтительно выбрано из мобилизирующих раковые клетки средств.
В пятом варианте осуществления третьего аспекта, который также является вариантом осуществления любого из вариантов, от второго до четвертого, осуществления третьего аспекта, данное лекарственное средство применяют в сочетании с третьим фармацевтически активным средством, где третье фармацевтически активное средство повреждает, разрушает и/или метит раковые клетки в периферической крови, где данная метка приводит к активации защитных сил организма.
В шестом варианте осуществления третьего аспекта, который также является вариантом осуществления любого из вариантов, от второго до пятого, осуществления третьего аспекта, субъект последовательно или одновременно подвергается химиотерапии и/или лучевой терапии.
В седьмом варианте осуществления третьего аспекта, который также является вариантом осуществления любого из вариантов, от пятого до шестого, осуществления третьего аспекта, данное лекарственное средство применяют для лечения и/или профилактики рака, предпочтительно – солидных опухолей и гематологического рака, более предпочтительно - лейкоза, лимфомы и миеломы.
В восьмом варианте осуществления третьего аспекта данное лекарственное средство предназначено для мобилизации долгоживущих плазмоцитов, B-клеток и/или T-клеток в периферическую кровь субъекта, где предпочтительно B-клетки и/или T-клетки являются В-клетками памяти и/или T-клетками памяти.
В девятом варианте осуществления третьего аспекта, который также является вариантом осуществления восьмого варианта осуществления третьего аспекта, данное лекарственное средство применяют в сочетании со вторым фармацевтически активным средством, где второе фармацевтически активное средство применяют для мобилизации долгоживущих плазмоцитов, B-клеток и/или T-клеток памяти в периферическую кровь субъекта, где второе фармацевтически активное средство предпочтительно выбрано из мобилизирующих клетки средств.
В десятом варианте осуществления третьего аспекта, который также является вариантом осуществления любого из вариантов, от восьмого до девятого, осуществления третьего аспекта, данное лекарственное средство применяют в сочетании с третьим фармацевтически активным средством, и третье фармацевтически активное средство повреждает, разрушает и/или метит долгоживущие плазмоциты, B-клетки и/или T-клетки в периферической крови, где данная метка приводит к активации защитных сил организма.
В одиннадцатом варианте осуществления третьего аспекта, который также является вариантом осуществления любого из вариантов, от восьмого до десятого, осуществления третьего аспекта, субъект последовательно или одновременно подвергается химиотерапии и/или лучевой терапии.
В двенадцатом варианте осуществления третьего аспекта, который также является вариантом осуществления любого из вариантов, от восьмого до одиннадцатого, осуществления третьего аспекта, лекарственное средство применяют для лечения и/или профилактики:
системных аутоиммунных заболеваний , где такое системное аутоиммунное заболевание предпочтительно выбрано из аллергии, аутоиммунной гемолитической анемии с комбинированным синдромом холодовых и тепловых агглютининов, синдрома системной воспалительной реакции, геморрагического шока, сахарного диабета 1го типа, диффузной склеродермии, полихондрии, аутоиммунного плюригландулярного синдрома, системной эритематозной волчанки и ее проявлений, ревматоидного артрита, ревматических заболеваний глаза, мозга, легких, почек, сердца, печени, желудочно-кишечного тракта, селезенки, кожи, костей, лимфатической системы, крови или других органов;
аутоиммунных заболеваний желудочно-кишечного тракта, где такое аутоиммунное заболевание желудочно-кишечного тракта предпочтительно выбрано из болезни Крона, язвенного колита, заболевания брюшной полости, глютеновой энтеропатии, воспалительного заболевания кишечника, панкреатита, эозинофильного эзофагита;
аутоиммунных заболеваний кожи , где такое аутоиммунное заболевание кожи предпочтительно выбрано из псориаза, крапивницы, дерматомиозита, обыкновенной пузырчатки, листовидной пузырчатки, буллезного пемфигоида, кольцевидной/лентовидной склеродермии, витилиго, герпетиформного дерматита или болезни Дюринга, склерозирующего лишая;
аутоиммунных заболеваний сосудистой системы , где такое аутоиммунное заболевание сосудистой системы предпочтительно выбрано из васкулитов, предпочтительно – височного артериита, васкулита, сосудистого истечения, ревматической полимиалгии, атеросклероза, синдрома Черджа-Стросса, синдрома Такаясу, синдрома Гудпасчера, предпочтительно, главным образом затрагивающего почки, конкретнее – почечные клубочки, и/или также главным образом затрагивающего легкие, гломерулонефрита, узелкового полиартериита, болезни Бехчета;
аутоиммунных заболеваний нервной системы , где такое аутоиммунное заболевание нервной системы предпочтительно выбрано из рассеянного склероза, хронической воспалительной демиелинизирующей полинейропатии, нейрокогнитивной дисфункции, синдрома мышечной скованности, синдрома Гийена-Барре, тяжелой псевдопаралитической миастении, синдрома Ламберта-Итона;
аутоиммунных заболеваний опорно-двигательного аппарата , где такое аутоиммунное заболевание опорно-двигательного аппарата предпочтительно выбрано из анкилозирующего спондилита, саркоидоза, ревматической полимиалгии, полимиозита, псориатического артрита, ревматического полиартрита, полихондрии, фибромиалгии, ювенильного ревматоидного артрита, болезни Лайма, реактивного артрита, спондилоартропатии, дегенеративного заболевания сустава;
других аутоиммунных заболеваний , где такие другие аутоиммунные заболевания предпочтительно выбраны из синдрома Когана, аутоиммунного адреналита, болезни Меньера, местных воспалений, гнездной алопеции, острых воспалительных заболеваний, первичного биллиарного цирроза, синдрома Шегрена, склеродермий, таких как диффузная склеродермия, CREST-синдром и/или кольцевидная/лентовидная склеродермия, аутоиммунного увеита, тиреоидита Хашимото, базедовой болезни, аутоиммунного гепатита, гломерулонефрита, антифосфолипидного синдрома, идиопатического фиброза легких, аутоиммунного бесплодия, заболеваний иммунного комплекса и перитонита;
отторжения трансплантата пересаженного органа , где такой орган выбран из печени, почки, кишечника, легких, сердца, кожи, конечностей, роговой оболочки глаза, островков Лангерганса, костного мозга, кровеносных сосудов, поджелудочной железы;
и/или реакции «трансплантат против хозяина» после пересадки костного мозга.
В 13м варианте осуществления третьего аспекта данное лекарственное средство предназначено для ингибирования миграции лейкоцитов.
В 14м варианте осуществления третьего аспекта, который также является вариантом осуществления 13го варианта осуществления третьего аспекта, данное лекарственное средство предназначено для профилактики и/или лечения отторжения трансплантата пересаженными органами, такими как печень, почки, кишечник, легкие, сердце, кожа, конечности, роговая оболочка глаза, островки Лангерганса, костный мозг, кровеносные сосуды и поджелудочная железа.
В 15м варианте осуществления третьего аспекта, который также является вариантом осуществления 13го варианта осуществления третьего аспекта, данное лекарственное средство применяют для лечения и/или профилактики воспаления, возникающего во время течения или имеющего отношение к:
системным аутоиммунным заболеваниям , где такое системное аутоиммунное заболевание предпочтительно выбрано из аллергии, аутоиммунной гемолитической анемии с комбинированным синдромом холодовых и тепловых агглютининов, синдрома системной воспалительной реакции, геморрагического шока, сахарного диабета 1го типа, диффузной склеродермии, полихондрии, аутоиммунного плюригландулярного синдрома, системной эритематозной волчанки и ее проявлений, ревматоидного артрита, ревматических заболеваний глаза, мозга, легких, почек, сердца, печени, желудочно-кишечного тракта, селезенки, кожи, костей, лимфатической системы, крови или других органов;
аутоиммунным заболеваниям желудочно-кишечного тракта , где такое аутоиммунное заболевание желудочно-кишечного тракта предпочтительно выбрано из болезни Крона, язвенного колита, заболевания брюшной полости, глютеновой энтеропатии, воспалительного заболевания кишечника, панкреатита, эозинофильного эзофагита;
аутоиммунным заболеваниям кожи , где такое аутоиммунное заболевание кожи предпочтительно выбрано из псориаза, крапивницы, дерматомиозита, обыкновенной пузырчатки, листовидной пузырчатки, буллезного пемфигоида, кольцевидной/лентовидной склеродермии, витилиго, герпетиформного дерматита или болезни Дюринга, склерозирующего лишая;
аутоиммунным заболеваниям сосудистой системы , где такое аутоиммунное заболевание сосудистой системы предпочтительно выбрано из васкулитов, предпочтительно – височного артериита, васкулита, сосудистого истечения, ревматической полимиалгии, атеросклероза, синдрома Черджа-Стросса, синдрома Такаясу, синдрома Гудпасчера, предпочтительно, главным образом затрагивающего почки, конкретнее – почечные клубочки, и/или также главным образом затрагивающего легкие, гломерулонефрита, узелкового полиартериита, болезни Бехчета;
аутоиммунным заболеваниям нервной системы , где такое аутоиммунное заболевание нервной системы предпочтительно выбрано из рассеянного склероза, хронической воспалительной демиелинизирующей полинейропатии, нейрокогнитивной дисфункции, синдрома мышечной скованности, синдрома Гийена-Барре, тяжелой псевдопаралитической миастении, синдрома Ламберта-Итона;
аутоиммунным заболеваниям опорно-двигательного аппарата , где такое аутоиммунное заболевание опорно-двигательного аппарата предпочтительно выбрано из анкилозирующего спондилита, саркоидоза, ревматической полимиалгии, полимиозита, псориатического артрита, ревматического полиартрита, полихондрии, фибромиалгии, ювенильного ревматоидного артрита, болезни Лайма, реактивного артрита, спондилоартропатии, дегенеративного заболевания сустава;
и/или другим аутоиммунным заболеваниям, где такие другие аутоиммунные заболевания предпочтительно выбраны из синдрома Когана, аутоиммунного адреналита, болезни Меньера, местных воспалений, гнездной алопеции, острых воспалительных заболеваний, первичного биллиарного цирроза, синдрома Шегрена, склеродермий, таких как диффузная склеродермия, CREST-синдром и/или кольцевидная/лентовидная склеродермия, аутоиммунного увеита, тиреоидита Хашимото, базедовой болезни, аутоиммунного гепатита, гломерулонефрита, антифосфолипидного синдрома, идиопатического фиброза легких, аутоиммунного бесплодия, заболеваний иммунного комплекса и перитонита.
В 16м варианте осуществления третьего аспекта, который также является вариантом осуществления 13го варианта осуществления третьего аспекта, данное лекарственное средство предназначено для лечения и/или профилактики аллергических реакций кожи и/или слизистой оболочки дыхательных путей, предпочтительно - сенной лихорадки, астмы, гиперреактивности дыхательных путей и/или дерматита.
В 17м варианте осуществления третьего аспекта, который также является вариантом осуществления 16го варианта осуществления третьего аспекта, данный дерматит представляет собой контактный дерматит и/или атопический дерматит.
Проблема, приоритетная для настоящей заявки, решена в четвертом аспекте с помощью способа получения клеток-предшественников и/или стволовых клеток от первого субъекта, каковой способ включает:
а) введение данному субъекту нуклеиновой кислоты, согласно первому аспекту и любым вариантам их осуществления, в количестве, эффективном для мобилизации указанных клеток-предшественников и/или стволовых клеток в периферическую кровь указанного субъекта;
b) последующий сбор указанных клеток-предшественников и/или стволовых клеток от указанного субъекта.
В первом варианте осуществления четвертого аспекта сбор клеток-предшественников и/или стволовых клеток осуществляли посредством афереза, лейкафереза, клеточного сортинга и/или проточной цитометрии.
Во втором варианте осуществления четвертого аспекта, который также является вариантом осуществления четвертого аспекта и первым вариантом осуществления четвертого аспекта, первый субъект или второй субъект были последовательно или одновременно подвержены химиотерапии и/или лучевой терапии.
В третьем варианте осуществления четвертого аспекта, который также является вариантом осуществления второго варианта осуществления четвертого аспекта, после химиотерапии и/или лучевой терапии первого субъекта или второго субъекта собранные клетки-предшественники и/или стволовые клетки первого субъекта или второго субъекта вводили в периферическую кровь первого субъекта или второго субъекта.
В четвертом варианте осуществления четвертого аспекта, который также является вариантом осуществления четвертого аспекта и любого из вариантов, от первого до третьего, осуществления четвертого аспекта, собранные клетки-предшественники и/или стволовые клетки выращивали, и подращенные клетки-предшественники и/или стволовые клетки вводили первому субъекту или второму субъекту, где, предпочтительно, подращенные клетки-предшественники и/или стволовые клетки вводили посредством внутривенной или местной инъекции.
В пятом варианте осуществления четвертого аспекта, который также является вариантом осуществления четвертого аспекта и любого из вариантов, от первого до четвертого, осуществления четвертого аспекта, данный способ применяли для лечения раковых, предпочтительно, солидных опухолей и гематологических злокачественных новообразований.
Проблема, приоритетная для настоящей заявки, решена в пятом аспекте с помощью молекулы нуклеиновой кислоты согласно первому аспекту и любым вариантам их осуществления, для применения в способе согласно четвертому аспекту и любым вариантам их осуществления.
Проблема, приоритетная для настоящей заявки, решена в шестом аспекте с помощью способа истощения долгоживущих плазмоцитов, B-клеток и/или T-клеток самого субъекта, каковой способ включает:
а) введение данному субъекту нуклеиновой кислоты, согласно первому аспекту и любым вариантам их осуществления, в количестве, эффективном для мобилизации указанных долгоживущих плазмоцитов, B-клеток и/или T-клеток в периферическую кровь указанного субъекта;
b) последующий сбор указанных долгоживущих плазмоцитов, B-клеток и/или T-клеток от указанного субъекта;
где истощенные и собранные T-клетки являются, предпочтительно, T-клетками памяти.
В первом варианте осуществления шестого аспекта сбор долгоживущих плазмоцитов, B-клеток и/или T-клеток осуществляли посредством афереза, клеточного сортинга и/или проточной цитометрии, предпочтительно - посредством проточной цитометрии с поверхностными маркерами, подходящими для указанных клеток.
В шестом варианте осуществления четвертого аспекта, который также является вариантом осуществления четвертого аспекта и любого из вариантов, от первого до пятого, осуществления четвертого аспекта; и который также представляет собой второй вариант осуществления шестого аспекта, который также является вариантом осуществления шестого аспекта и первым вариантом осуществления шестого аспекта, данный способ применяют для лечения и/или профилактики:
системных аутоиммунных заболеваний , где такое системное аутоиммунное заболевание предпочтительно выбрано из аллергии, аутоиммунной гемолитической анемии с комбинированным синдромом холодовых и тепловых агглютининов, синдрома системной воспалительной реакции, геморрагического шока, сахарного диабета 1го типа, диффузной склеродермии, полихондрии, аутоиммунного плюригландулярного синдрома, системной эритематозной волчанки и ее проявлений, ревматоидного артрита, ревматических заболеваний глаза, мозга, легких, почек, сердца, печени, желудочно-кишечного тракта, селезенки, кожи, костей, лимфатической системы, крови или других органов;
аутоиммунных заболеваний желудочно-кишечного тракта, где такое аутоиммунное заболевание желудочно-кишечного тракта предпочтительно выбрано из болезни Крона, язвенного колита, заболевания брюшной полости, глютеновой энтеропатии, воспалительного заболевания кишечника, панкреатита, эозинофильного эзофагита;
аутоиммунных заболеваний кожи , где такое аутоиммунное заболевание кожи предпочтительно выбрано из псориаза, крапивницы, дерматомиозита, обыкновенной пузырчатки, листовидной пузырчатки, буллезного пемфигоида, кольцевидной/лентовидной склеродермии, витилиго, герпетиформного дерматита или болезни Дюринга, склерозирующего лишая;
аутоиммунных заболеваний сосудистой системы , где такое аутоиммунное заболевание сосудистой системы предпочтительно выбрано из васкулитов, предпочтительно – височного артериита, васкулита, сосудистого истечения, ревматической полимиалгии, атеросклероза, синдрома Черджа-Стросса, синдрома Такаясу, синдрома Гудпасчера, предпочтительно, главным образом затрагивающего почки, конкретнее – почечные клубочки, и/или также главным образом затрагивающего легкие, гломерулонефрита, узелкового полиартериита, болезни Бехчета;
аутоиммунных заболеваний нервной системы , где такое аутоиммунное заболевание нервной системы предпочтительно выбрано из рассеянного склероза, хронической воспалительной демиелинизирующей полинейропатии, нейрокогнитивной дисфункции, синдрома мышечной скованности, синдрома Гийена-Барре, тяжелой псевдопаралитической миастении, синдрома Ламберта-Итона;
аутоиммунных заболеваний опорно-двигательного аппарата , где такое аутоиммунное заболевание опорно-двигательного аппарата предпочтительно выбрано из анкилозирующего спондилита, саркоидоза, ревматической полимиалгии, полимиозита, псориатического артрита, ревматического полиартрита, полихондрии, фибромиалгии, ювенильного ревматоидного артрита, болезни Лайма, реактивного артрита, спондилоартропатии, дегенеративного заболевания сустава;
других аутоиммунных заболеваний, где такие другие аутоиммунные заболевания предпочтительно выбраны из синдрома Когана, аутоиммунного адреналита, болезни Меньера, местных воспалений, гнездной алопеции, острых воспалительных заболеваний, первичного биллиарного цирроза, синдрома Шегрена, склеродермий, таких как диффузная склеродермия, CREST-синдром и/или кольцевидная/лентовидная склеродермия, аутоиммунного увеита, тиреоидита Хашимото, базедовой болезни, аутоиммунного гепатита, гломерулонефрита, антифосфолипидного синдрома, идиопатического фиброза легких, аутоиммунного бесплодия, заболеваний иммунного комплекса и перитонита;
отторжения трансплантата пересаженного органа, где такой орган выбран из печени, почки, кишечника, легких, сердца, кожи, конечностей, роговой оболочки глаза, островков Лангерганса, костного мозга, кровеносных сосудов, поджелудочной железы;
и/или реакции «трансплантат против хозяина » после пересадки костного мозга.
Проблема, приоритетная для настоящей заявки, решена в седьмом аспекте с помощью молекулы нуклеиновой кислоты, согласно первому аспекту и любым вариантам их осуществления, для применения в способе согласно шестому аспекту и любым вариантам их осуществления.
Проблема, приоритетная для настоящей заявки, решена в восьмом аспекте с помощью использования нуклеиновой кислоты, как определено в пункте формулы изобретения по любому из вариантов, от 10го до 57го, осуществления первого аспекта, для производства лекарственного средства, где данное лекарственное средство предназначено для лечения и/или профилактики нефропатии, предпочтительно - диабетической нефропатии.
Проблема, приоритетная для настоящей заявки, решена в девятом аспекте с помощью использования нуклеиновой кислоты, как определено в пункте формулы изобретения по любому из вариантов, от 10го до 57го, осуществления первого аспекта, для производства лекарственного средства, где данное лекарственное средство предназначено для лечения и/или профилактики гипертензии, предпочтительно - легочной гипертензии.
Проблема, приоритетная для настоящей заявки, решена в десятом аспекте с помощью использования нуклеиновой кислоты, как определено в пункте формулы изобретения по любому из вариантов, от 10го до 57го, осуществления первого аспекта, для производства лекарственного средства, где данное лекарственное средство предназначено для лечения и/или профилактики фиброза, предпочтительно - идиопатического фиброза легких.
В первом варианте осуществления десятого аспекта данное лекарственное средство предназначено для лечения и/или профилактики фиброза в рамках процесса заживления ран.
Проблема, приоритетная для настоящей заявки, решена в одиннадцатом аспекте с помощью использования нуклеиновой кислоты, как определено в пункте формулы изобретения по любому из вариантов осуществления, от 10го до 57го, для производства лекарственного средства, где данное лекарственное средство предназначено для лечения заболевания и/или нарушения, включающего ангиогенез и/или неоваскуляризацию.
В первом варианте осуществления одиннадцатого аспекта данное лекарственное средство применяют для комбинированной терапии со средством, ингибирующим VEGF.
Во втором варианте осуществления одиннадцатого аспекта, который также является вариантом осуществления первого варианта осуществления одиннадцатого аспекта, данное лекарственное средство применяют у субъектов, которые слабо реагируют на терапию со средством, ингибирующим VEGF.
В третьем варианте осуществления одиннадцатого аспекта данное лекарственное средство применяют у субъектов, которые не реагируют на терапию со средством, ингибирующим VEGF.
В четвертом варианте осуществления одиннадцатого аспекта данное заболевание и/или нарушение включает и/или связано с хороидальной неоваскуляризацией.
В пятом варианте осуществления одиннадцатого аспекта, который также является вариантом осуществления одиннадцатого аспекта и любого из вариантов, от первого до четвертого, осуществления одиннадцатого аспекта, данное заболевание и/или нарушение выбирают из группы, включающей заболевания сетчатки, предпочтительно - возрастной макулярной дегенерации, диабетической ретинопатии, окклюзии вен в сетчатке, макулярного отека и отека сосудов сетчатки глаза.
В шестом варианте осуществления одиннадцатого аспекта, который также является вариантом осуществления одиннадцатого аспекта и любого из вариантов, от первого до третьего, осуществления одиннадцатого аспекта, данное заболевание выбирают из группы, включающей раковые, предпочтительно - солидные опухоли и метастазы.
В 18м варианте осуществления третьего аспекта данное лекарственное средство предназначено для введения субъекту, который подвержен, или должен быть подвержен, курсу лечения, истощающему клетки-предшественники и/или стволовые клетки у субъекта, предпочтительно – истощает в периферической крови.
В 19м варианте осуществления третьего аспекта, который также является вариантом осуществления третьего аспекта, а также 18го варианта осуществления третьего аспекта, данное лекарственное средство должно быть введено субъекту, который подвержен, или должен быть подвержен, химиотерапии и/или лучевой терапии.
В 20м варианте осуществления третьего аспекта данное лекарственное средство предназначено для восстановления или укрепления иммунной системы субъекта.
Проблема, приоритетная для настоящей заявки, решена в двенадцатом аспекте с помощью молекулы, ингибирующей передачу сигнала между SDF-1 и рецептором SDF-1, для производства лекарственного средства, где данное лекарственное средство предназначено для лечения и/или профилактики нефропатии, предпочтительно - диабетической нефропатии.
В первом варианте осуществления двенадцатого аспекта молекула представляет собой молекулу, связывающую SDF-1, либо молекулу, связывающую рецептор SDF-1, и содержит целенаправленные нуклеиновые кислоты, подобные аптамерам и шпигельмерам, антитела и низкомолекулярные синтетические препараты.
Во втором варианте осуществления двенадцатого аспекта, который также является вариантом осуществления двенадцатого аспекта и первого варианта осуществления двенадцатого аспекта, молекула представляет собой молекулу нуклеиновой кислоты, как определено в вариантах, от десятого до 57го, осуществления первого аспекта.
В третьем варианте осуществления двенадцатого аспекта молекула представляет собой молекулу, ингибирующую экспрессию SDF-1 или рецептора SDF-1, и содержит молекулы миРНК, рибозимы, антисмысловые молекулы и ингибиторы факторов транскрипции.
Проблема, приоритетная для настоящей заявки, решена в 13м аспекте с помощью молекулы нуклеиновой кислоты, согласно любому из вариантов, от пятого до 57го, осуществления первого аспекта, для применения в способе ингибирования миграции лейкоцитов.
Проблема, приоритетная для настоящей заявки, решена в 14м аспекте с помощью молекулы, связывающей SDF-1, не проникающей через гематоэнцефалический барьер, с целью применения для мобилизации стволовых клеток, полученных из костного мозга, или с целью применения в производстве лекарственного средства, предпочтительно - лекарственного средства для мобилизации стволовых клеток, полученных из костного мозга.
В первом варианте осуществления 14го аспекта данное лекарственное средство применяли для облегчения течения поражений центральной нервной системы и/или для стимуляции восстановления тканей после инсульта, предпочтительно - ишемического инсульта.
Во втором варианте осуществления 14го аспекта, который также является вариантом осуществления первого варианта осуществления 14го аспекта, молекула, связывающая SDF-1, содержит целенаправленные нуклеиновые кислоты, выбранные из группы, включающей аптамеры, шпигельмеры, антитела и низкомолекулярные синтетические препараты.
В третьем варианте осуществления 14го аспекта, который также является вариантом осуществления 14го аспекта и любого из вариантов, от первого до второго, осуществления 14го аспекта, молекула является молекулой нуклеиновой кислоты, как определено в любом из вариантов, от девятого до 57го, осуществления первого аспекта.
Проблема, приоритетная для настоящей заявки, решена в 15м аспекте с помощью молекулы нуклеиновой кислоты, согласно первому аспекту и любым из вариантов, от первого до 57го, осуществления первого аспекта, для применения в лечении заболевания, как определено в любом из предыдущих пунктов формулы изобретения.
Проблема, приоритетная для настоящей заявки, решена в 16м аспекте с помощью использования нуклеиновой кислоты, как определено в вариантах, от 10го до 57го, осуществления первого аспекта, для производства лекарственного средства, где данное лекарственное средство предназначено для лечения и/или профилактики WHIM синдрома (хронической нециклической нейтропении).
Проблема, приоритетная для настоящей заявки, решена в 17м аспекте с помощью использования нуклеиновой кислоты, как определено в вариантах, от 10го до 57го, осуществления первого аспекта, для производства лекарственного средства, где данное лекарственное средство предназначено для лечения и/или профилактики роста и метастазирования рака, а также роста неоплазии.
Проблема, приоритетная для настоящей заявки, решена в 18м аспекте с помощью использования нуклеиновой кислоты, как определено в вариантах, от 10го до 57го, осуществления первого аспекта, для производства лекарственного средства, где данное лекарственное средство вводят субъекту до химиотерапии, предпочтительно – химиотерапии, назначенной для лечения рака.
Не стремясь быть связанными рамками какого-либо одного толкования, авторы настоящего изобретения предполагают, что молекулы нуклеиновых кислот согласно настоящему изобретению ингибируют связывание SDF-1 со своим рецептором SDF-1 и, таким образом, прямо или косвенно, влияют на миграцию клеток, предпочтительно - миграцию клеток из периферической крови в какую-либо одну или более ткань(и), а также из тканей - в периферическую кровь.
Однако, не стремясь быть связанными рамками какого-либо одного толкования, авторы настоящего изобретения предполагают далее, что молекулы нуклеиновых кислот согласно настоящему изобретению, ингибирующие связывание SDF-1 с рецептором SDF-1, приводят к миграции клеток-предшественников, стволовых клеток, раковых клеток, долгоживущих плазмоцитов, B-клеток и/или T-клеток, предпочтительно - путем мобилизации, из тканей в периферическую кровь, посредством ингибирования взаимодействия между SDF-1 и рецептором SDF-1.
Более того и вновь не стремясь быть связанными рамками какого-либо одного толкования, авторы настоящего изобретения предполагают, что молекулы нуклеиновых кислот согласно настоящему изобретению, ингибирующие связывание SDF-1 со своими рецепторами SDF-1, предотвращают миграцию лейкоцитов, таких как T-лимфоциты, B-лимфоциты, моноциты, макрофаги, эозинофилы, нейтрофилы, базофилы и/или дендритные и тучные клетки, предпочтительно, в тканях, таких как кожа или слизистая оболочка, и, предпочтительно, помогают смягчать аутоиммунные заболевания и аллергические реакции кожи и слизистой оболочки.
К тому же и по-прежнему не стремясь быть связанными рамками какого-либо одного толкования, авторы настоящего изобретения смогли показать, что применение средств, ингибирующих SDF-1, таких как молекулы нуклеиновых кислот согласно настоящему изобретению, может быть полезным при лечении нефропатии, предпочтительно - диабетической нефропатии; гипертензии, предпочтительно - легочной гипертензии; фиброза, предпочтительно - фиброза легких, и при лечении заболеваний и/или нарушений, включающих неоваскуляризацию, предпочтительно хороидальную неоваскуляризацию.
Что касается различных заболеваний, патологических состояний и нарушений, которые можно излечить или предотвратить с помощью молекул нуклеиновых кислот согласно настоящему изобретению, или композиций, предпочтительно - фармацевтических композиций, содержащих таковые, следует подтвердить, что такие заболевания, патологические состояния и нарушения являются теми же, что описаны в данном документе, включая конкретно и те, что описаны и предложены во вводной части настоящей заявки. На настоящий момент, соответствующие отрывки являются неотъемлемой частью настоящего раскрытия, указывая на возможность применения данных молекул нуклеиновых кислот для профилактики и лечения, соответственно, указанных заболеваний, патологических состояний и нарушений.
После внутривенного введения связывающей SDF-1 молекулы нуклеиновой кислоты, такой как NOX-A12-JE40, клетки находятся в крови, где эффект связывающей SDF-1 нуклеиновой кислоты NOX-A12-JE40 был отмечен после проведения определения количества клеток цельной крови с применением гемоцитометра (см. пример 10). В то время как гемопоэтические стволовые клетки/гемопоэтические клетки-предшественники представлены только в виде небольшой доли мобилизированных белых кровяных клеток, существуют и другие типы клеток, выходящих в кровь. К ним относятся моноциты и нейтрофильные гранулоциты, также как и тканесвязанные стволовые клетки/клетки-предшественники, мезенхимные стволовые клетки, долгоживущие B-клетки плазмы, не ограничиваясь этими клетками.
На основании этих результатов авторы настоящего изобретения сделали вывод, что ингибирование передачи сигнала от SDF-1 к рецептору SDF-1 с помощью связывающей SDF-1 молекулы нуклеиновой кислоты согласно настоящему изобретению приводит к влиянию на миграцию клеток. Предпочтительно, данные клетки экспрессируют рецептор SDF-1.
В соответствии с этим, термин «миграция», предпочтительно используемый в данном документе, относится к миграции и/или перемещению из одной ткани в другую ткань, из ткани в периферическую кровь и/или из периферической крови в ткань. Миграцию клеток можно проверить в TAX-тесте (миграция in vitro), как показано в примере 5 и/или применяя гемоцитометр и FACS анализ, как показано в примере 10 (эксперимент in vivo). Кроме того, можно проделать иммуногистохимию ткани, где мигрировавшие клетки обнаруживаются антителами, направленными на специфичные для данных клеток поверхностные маркеры. Используемый в данном документе термин «рецептор SDF-1», независимо от того, используется ли он в единственном или во множественном числе, относится к любому рецептору, с которым связывается SDF-1. На сегодняшний день известны два рецептора, CXCR 4 (Godessart 2005) и CXCR7 (Burns, Summers, et al. 2006), которые являются предпочтительными рецепторами, связывающими SDF-1.
Развитие клеток крови. Развитие и созревание клеток крови является комплексным процессом. Зрелые кровяные клетки происходят из гемопоэтических исходных клеток, также называемых клетками-предшественниками, и стволовых клеток, присутствующих в особых гемопоэтических тканях, включая костный мозг. В пределах этих сред обитания гемопоэтические клетки пролиферируют и дифференцируются, прежде чем вступить в процесс кровообращения. Хемокиновый рецептор CXCR4 и его природный лиганд SDF-1, вероятно, являются существенными в этом процессе (Maekawa and Ishii 2000; Nagasawa 2000). Это было показано в соответствии с сообщениями, что нокаутные по CXCR4 или SDF-1 мыши обнаруживают нарушения гемопоэза (Ma, Jones et al. 1998; Tachibana, Hirota et al. 1998; Zou, Kottmann et al. 1998). Также известно, что CD34+ клетки-предшественники экспрессируют CXCR4 и нуждаются в SDF-1, производимом стромальными клетками костного мозга для хемоатракции и приживления трансплантата (Peled, Petit et al. 1999), и что in vitro SDF-1 является хемотаксическим как для клеток CD34+ (Aiuti, Webb et al. 1997; Viardot, Kronenwett et al. 1998), так и для клеток-предшественников/стволовых клеток (Jo, Rafii et al. 2000). SDF-1 также является важным хемоаттрактантом, передающим сигнал через рецептор CXCR4, для некоторых других более дифференцированных предшественников и зрелых кровяных клеток, включая T-лимфоциты и моноциты (Bleul, Fuhlbrigge et al. 1996), про- и пре-B-лимфоциты (Fedyk, Ryyan et al. 1999), а также мегакариоциты (Riviere, Subra et al. 1999; Abi-Younes, Sauty et al. 2000; Hodohara, Fujii et al. 2000; Majka, Janowska-Wieczorek et al. 2000; Gear, Suttitanamongkol et al. 2001). Из-за этих взаимоотношений между различными типами клеток и зависимости от SDF-1 и рецептора SDF-1, молекулы нуклеиновой кислоты фактически могут быть направлены к различным типам клеток согласно настоящему изобретению.
Таким образом, вкратце, представляется, что SDF-1 способен управлять местонахождением и дифференцировкой клеток, содержащих рецепторы SDF-1, предпочтительно - рецепторы CXCR4, являются ли эти клетки стволовыми клетками, то есть клетками, которые являются CD34+, и/или клетками-предшественниками, которые приводят к образованию указанных типов колоний в ответ на специфические стимулы; они могут быть CD34+, либо CD34-, либо несколько более дифференцированными клетками.
В последнее время значительное внимание было сосредоточено на многих CD34+ клетках, мобилизированных в банке клеток-предшественников периферической крови, используемых для аутологичной и аллографтной трансплантации стволовых клеток. Полагается, что популяция CD34+ клеток является компонентом, ответственным, прежде всего, за сокращение времени восстановления после химиотерапии, и вероятнее всего, что данные клетки ответственны за приживление трансплантата на долгий срок и восстановление гемопоэза (Croop, Cooper et al. 2000). Механизм, с помощью которого CD34+ клетки повторно приживляются, может быть обусловлен хемотаксическими эффектами SDF-1 на экспрессирующие CXCR4 клетки (Ponomaryov, Peled et al. 2000; Voermans, Kooi et al. 2001). Совсем недавно было показано, что зрелые гемопоэтические стволовые клетки ответственны за восстановление поврежденной сердечной ткани у мышей (Jackson, Majka et al. 2001; Kocher, Schuster et al. 2001).
В данном документе термин «клетки-предшественники» относится к клеткам, которые в ответ на некоторые стимулы могут образовывать дифференцированные гемопоэтические или миелоидные клетки. Присутствие клеток-предшественников может быть оценено по способности клеток в образце формировать колониеобразующие единицы различных типов, включая, например, CFU-GM (что означает колониеобразующие единицы гранулоцитов-макрофагов); CFU-GEMM (что означает колониеобразующие единицы поливалентных клеток); BFU-E (колониеобразующая эритроидная «взрывообразующая» единица); HPP-CFC (что означает высоко пролиферативные потенциально колониеобразующие клетки); или другие типы дифференцированных колоний, которые могут быть получены в культуре с помощью известных протоколов.
В данном документе стволовые клетки являются менее дифференцированными формами клеток-предшественников. Как правило, такие клетки часто положительны по CD34. Однако некоторые стволовые клетки не содержат этот маркер. Клетки CD34+ могут быть проанализированы с помощью сортинга флуоресцентно-активированных клеток (сокр. FACS), и, таким образом, их присутствие в образце может быть оценено посредством этого метода.
Как правило, CD34+ клетки присутствуют в крови только на низких уровнях, но в костном мозге присутствуют в больших количествах. Хотя другие типы клеток, такие как эндотелиальные клетки и тучные клетки, также могут проявить этот маркер, CD34 считают признаком присутствия стволовых клеток.
Не стремясь быть связанными рамками какого-либо одного толкования, авторы настоящего изобретения пришли к заключению, что в дополнение к стволовым клеткам и/или клеткам-предшественникам, на раковые клетки, долгоживущие плазмоциты, B-клетки и/или T-клетки оказывает влияние ингибирующее действие нуклеиновой кислоты, согласно настоящему изобретению, на передачу сигнала между SDF-1 и его рецептором SDF-1. B-клетки и T-клетки предпочтительно являются B-клетками памяти и T-клетками памяти. Ингибирование передачи сигнала между SDF-1 и рецептором SDF-1 приводит к миграции, включающей мобилизацию, в периферическую кровь.
Мобилизация стволовых клеток, клеток-предшественников, раковых клеток, долгоживущих плазмоцитов, B-клеток и/или T-клеток предпочтительно происходит в гемопоэтических тканях, где данные гемопоэтические ткани выбирают из группы, включающей миелоидную ткань и лимфоидную ткань. Миелоидная ткань локализована в костном мозге. Лимфоидная ткань локализована в слизистой оболочке желудочно-кишечного тракта и дыхательного пути, лимфоузлах, селезенке и/или тимусе.
В данном документе раковые клетки представляют собой клетки, относящиеся к новообразованию, и являются предпочтительно выбранными из лейкозных клеток, миеломных клеток, клеток лимфомы, раковых стволовых клеток, раковых клеток с метастатическим потенциалом и метастазами рака. Клетки, относящиеся к новообразованию, обычно имеют общие генетические или эпигенетические отклонения, свидетельство клональности. Для некоторых типов неоплазмы, например, лимфомы и лейкоза, подтверждение клональности, как теперь полагают, необходимо (хотя и не достаточно) для характеризации клеточной пролиферации как относящейся к новообразованию.
После сильного иммунного ответа с продукцией активных антител окончательно дифференцированными B-клетками (плазмоцитами) большинство этих клеток умирает, чему соответствует прекращение заболевания и/или устранение чужеродного антигена (например, вирусного белка). Однако существует меньшинство В-клеток памяти, способных превратиться в плазмобласты с помощью T-клетки. У этих плазмобластов есть способность мигрировать в ответ на хемотаксические стимулы в тканевые ниши, где они выживают как долгоживущие плазмоциты «памяти» в течение многих лет. Такие ниши могут быть как в костном мозге, так и на периферии, например, в воспаленных тканях. Эти долгоживущие плазмоциты вероятнее всего ответственны за сохранение титров защитных антител в плазме, которые часто сохраняются в течение многих лет после инфекции или прививки (Tarlinton et al., 2008).
Мобилизация клеток-предшественников и/или стволовых клеток предпочтительно происходит из гемопоэтических тканей, где данную гемопоэтическую ткань выбирают из группы, включающей миелоидную ткань и лимфоидную ткань. Миелоидная ткань локализована в костном мозге. Лимфоидная ткань локализована в слизистой оболочке желудочно-кишечного тракта и дыхательного пути, лимфоузлах, селезенке, тимусе и лимфоидных фолликулах в воспаленной ткани. Предпочтительно, данная мобилизация клеток, как сообщалось ранее, включает миграцию клеток, как сообщалось ранее, в периферическую кровь.
Как показано ранее, было обнаружено, что SDF-1 действует как хемотаксическое средство на зрелые и исходные тучные клетки; особенно, когда зрелые тучные клетки высвобождают гистамин, например, путем передачи IgE-опосредованного сигнала посредством связывания с Fcε-рецептором на поверхности тучной клетки (Godot, Arock et al. 2007). Заболевания, подобные аллергическим реакциям кожи и слизистой оболочки дыхательных путей, такие как сенная лихорадка и астма, дерматит, особенно контактный дерматит и атопический дерматит, часто включают миграцию лейкоцитов к пораженным тканям и их аккумуляцию там. Экспрессию SDF-1 также наблюдали в синовиальных биопсиях и посредством RT-PCR субъектов, пораженных такими воспалениями суставов, как спондилоартропатия, ревматоидный артрит, псориатический артрит и дегенеративное заболевание суставов (остеоартрит). Взаимодействие связывающих SDF-1 нуклеиновых кислот согласно настоящему изобретению с SDF-1, поэтому, может оказывать положительное действие на пациентов, страдающих от воспаления суставов.
С помощью этого, и принимая во внимание отмеченное участие SDF-1 и рецепторов SDF-1, связывание SDF-1, а также взаимодействие между SDF-1 и ингибирующими рецептор SDF-1 молекулами нуклеиновых кислот согласно настоящему изобретению может помочь смягчить течение такого заболевания, где ингибирование SDF-1 молекулами нуклеиновых кислот согласно настоящему изобретению приводит к сокращению и/или ингибированию миграции лейкоцитов, где лейкоциты предпочтительно выбраны из группы, включающей T-лимфоциты, B-лимфоциты, моноциты, макрофаги, мегакариоциты, эозинофилы, нейтрофилы, базофилы, дендритные клетки и/или тучные клетки.
Миграция лейкоцитов приводит к их аккумуляции в ткани, где, предпочтительно, данная аккумуляция лейкоцитов приводит к воспалению указанной ткани, где данная ткань включает кожу и/или слизистую оболочку, предпочтительно слизистую оболочку дыхательных путей, и некоторые органы, выбранные без каких-либо ограничений из глаз, мозга, легких, почек, сердца, печени, желудочно-кишечного тракта, селезенки, кожи, костей и/или лимфатической системы.
Антагонистом SDF-1 является такая молекула, которая связывается с SDF-1 и ингибирует функцию SDF-1, предпочтительно в клеточном тесте или в модели in vivo, как описано в примерах.
Кроме того, настоящее изобретение основано на неожиданном факте, что существует возможность создать нуклеиновые кислоты, специфически и с высокой аффинностью связывающиеся с SDF-1. Такие нуклеиновые кислоты также предпочтительно упоминаются в данном документе как молекулы нуклеиновых кислот согласно настоящему изобретению, нуклеиновые кислоты согласно настоящему изобретению, патентоспособные нуклеиновые кислоты или патентоспособные молекулы нуклеиновых кислот.
SDF-1 представляет собой щелочной пептид, имеющий аминокислотную последовательность в соответствии с SEQ ID NO:1. Расчетное pI SDF-1 составляет 9,70. В данном документе термин «SDF-1» относится к любому SDF-1, без каких-либо ограничений включающему SDF-1 млекопитающих. Предпочтительно, SDF-1 млекопитающих выбирают из группы, включающей SDF-1 мышей, крыс, кроликов, хомяков, обезьян и человека. Более предпочтительно SDF-1 является SDF-1 человека, также называемым SDF-1α (SEQ ID NO:1) и/или SDF-1β человека (SEQ ID NO:2), наиболее предпочтительно SDF-1 человека, также называемым SDF-1α (SEQ ID NO:1).
Тот факт, что можно обнаружить высокоаффинное связывание нуклеиновых кислот с SDF-1, на настоящий момент является неожиданным, с тех пор как Итон и Голд с сотрудниками (Eaton, Gold et al. 1997) заметили, что создание аптамеров, то есть D-нуклеиновых кислот, связывающихся с молекулой-мишенью, направленных на щелочной белок, в большинстве случаев очень затруднено, поскольку этот тип мишеней производит высокое, но неспецифическое отношение «сигнал-шум». Это высокое отношение «сигнал-шум» происходит из-за высокой неспецифической аффинности, показываемой нуклеиновыми кислотами для щелочных мишеней, таких как SDF-1.
Особенности нуклеиновой кислоты согласно настоящему изобретению, как описано в данном документе, могут быть реализованы в любом аспекте настоящего изобретения, где используется данная нуклеиновая кислота, либо отдельно, либо в любой комбинации.
Не стремясь быть связанными рамками какого-либо одного толкования, авторы настоящего изобретения предполагают, что наблюдаемая специфичность связывающих SDF-1 нуклеиновых кислот согласно настоящему изобретению вносит свой вклад в некоторые структурные особенности и, в частности, в одну из нуклеотидных последовательностей, также упоминаемых здесь как коровые последовательности, которые будут обсуждены далее более детально, в соответствии с чем приводятся ссылки на фигуры с 1 по 8 и на пример 1. Однако следует понимать, что указанные фигуры и пример 1 включают лишь некоторые из указанных структурных особенностей, которые не должны быть обязательно реализованы во всех без исключения нуклеиновых кислотах согласно настоящему изобретению.
Как изложено более подробно в формуле изобретения и примере 1, различные связывающие SDF-1 человека молекулы нуклеиновой кислоты можно классифицировать на основании указанных боксов и некоторых структурных особенностей и элементов, соответственно. Различные категории, определенные таким образом, также упоминаются в данном документе как типы, а конкретнее, как тип A, тип B и тип C.
В предпочтительном варианте осуществления данная нуклеиновая кислота согласно настоящему изобретению является единичной молекулой нуклеиновой кислоты. В следующем варианте осуществления единичная молекула нуклеиновой кислоты представлена как совокупность единичных молекул нуклеиновой кислоты. Предпочтительно, термины «нуклеиновая кислота» и «молекула нуклеиновой кислоты» в данном документе применяются взаимозаменяемым образом, если не указано противоположное.
Специалист в данной области подтвердит, что молекула нуклеиновой кислоты в соответствии с данным изобретением предпочтительно состоит из нуклеотидов, ковалентно связанных друг с другом, предпочтительно через фосфодиэфирные связи или мостики.
Нуклеиновые кислоты согласно настоящему изобретению должны также включать нуклеиновые кислоты, которые существенным образом гомологичны конкретным последовательностям, раскрытым в данном документе. Термин «существенным образом гомологичны» следует понимать так, что гомология составляет, по меньшей мере, 75%, предпочтительно - 85%, более предпочтительно - 90% и наиболее предпочтительно - больше чем 95%, 96%, 97%, 98% или 99%.
Фактическое процентное содержание гомологичных нуклеотидов в нуклеиновой кислоте согласно настоящему изобретению будет зависеть от общего количества нуклеотидов в нуклеиновой кислоте. Частичное изменение процентов может быть основано на общем количестве нуклеотидов в нуклеиновой кислоте.
Определение гомологии известно любому специалисту, квалифицированному в данной области. Конкретнее, алгоритм сравнения последовательностей рассчитывает процент совпадения последовательностей для тестируемой(ых) последовательности(ей) относительно последовательности сравнения, исходя из предварительно заданных программных параметров. Тестируемая последовательность предпочтительно представляет собой последовательность или молекулу нуклеиновой кислоты, которая уже была проверена, или должна быть проверена, является ли она гомологичной, а если так, то до какой степени, другой молекуле нуклеиновой кислоты, на основании чего такая другая молекула нуклеиновой кислоты также называется последовательностью сравнения. В некотором варианте осуществления, последовательность сравнения представляет собой молекулу нуклеиновой кислоты, как описано в данном документе, более предпочтительно - молекулу нуклеиновой кислоты, имеющей последовательность нуклеиновой кислоты согласно любой из SEQ ID NO:5-144. Оптимальное выравнивание последовательностей для сравнения может быть проведено, например, алгоритмом поиска локальных гомологий Смита и Вотермана (Smith & Waterman, 1981), алгоритмом поиска гомологий Нидельмана и Вунча (Needleman & Wunsch, 1970), способом поиска подобия Пирсона и Липмана (Pearson & Lipman, 1988), с помощью компьютеризированных реализаций этих алгоритмов (GAP, BESTFIT, FASTA, и TFASTA в пакете программ Wisconsin Genetics Software Package, Genetics Computer Group, 575 Science Dr., Madison, Wis.), или путем визуальной проверки.
Один пример алгоритма, который является подходящим для определения процента идентичности последовательностей, является алгоритмом, используемым в основном инструменте поиска локального выравнивания (в дальнейшем - «BLAST»), см., например, работы Альтшуль и др. (Altschul et al. 1990 и Altschul et al., 1997). Программное обеспечение для выполнения BLAST-анализов общедоступно через Национальный центр биотехнологической информации (в дальнейшем – «NCBI»). Параметры по умолчанию, использованные при определении идентичности последовательностей с помощью программного обеспечения, поставляемого NCBI, например, BLASTN (для нуклеотидных последовательностей) и BLASTP (для аминокислотных последовательностей), описаны у Макгинниса и др. (McGinnis et al., 2004).
Термин «патентоспособная нуклеиновая кислота» или «нуклеиновая кислота согласно настоящему изобретению» будет также включать те нуклеиновые кислоты, которые содержат последовательности нуклеиновых кислот, раскрытых в данном документе, или их части, предпочтительно - в тех случаях, когда данные нуклеиновые кислоты или указанные части участвуют в связывании SDF-1. Такая нуклеиновая кислота может происходить из тех, которые раскрыты в данном документе, например, путем процессинга (концевого усечения). Усечение может относиться как к любому, так и к обоим концам нуклеиновой кислоты, как раскрыто в данном документе. К тому же, усечение может иметь отношение к внутренней последовательности нуклеотидов, то есть оно может иметь отношение к нуклеотиду(ам) между 5’ и 3’ концевыми нуклеотидами, соответственно. Кроме того, усечение должно включать делецию хотя бы одного нуклеотида из последовательности нуклеиновых кислот, раскрытых в данном документе. Усечение может также иметь отношение к более чем одному участку нуклеотидов патентоспособной(ых) нуклеиновой(ых) кислот(ы), где участок нуклеотидов может быть длиной всего один нуклеотид. Связывание нуклеиновой кислоты согласно настоящему изобретению может быть обнаружено любым специалистом в данной области с помощью общепринятых лабораторных исследований, либо с помощью применения или воспроизведения способа, описанного в данном документе, предпочтительно - описанного в данном документе в разделе «Примеры».
Нуклеиновыми кислотами согласно настоящему изобретению могут быть либо D-нуклеиновые кислоты, либо L-нуклеиновые кислоты. Предпочтительно, патентоспособные нуклеиновые кислоты представляют собой L-нуклеиновые кислоты. Кроме того, возможно, что одна или несколько частей нуклеиновой кислоты присутствуют в форме D-нуклеиновых кислот, либо по меньшей мере одна или несколько частей нуклеиновых кислот представляют собой L-нуклеиновые кислоты. Термин «часть» нуклеиновых кислот должен означать по меньшей мере один нуклеотид. Такие нуклеиновые кислоты обычно упоминаются в данном документе как D-и L-нуклеиновые кислоты, соответственно. Поэтому в особенно предпочтительном варианте осуществления нуклеиновые кислоты, согласно настоящему изобретению, состоят из L-нуклеотидов и включают по меньшей мере один D-нуклеотид. Такой D-нуклеотид предпочтительно присоединен к части, отличающейся от участков нуклеотидов, определяющих нуклеиновые кислоты согласно настоящему изобретению, предпочтительно к тем их частям, где происходит взаимодействие с другими частями нуклеиновой кислоты. Предпочтительно, такой D-нуклеотид присоединен к концевой части любого из участков нуклеотидов и любой нуклеиновой кислоты согласно настоящему изобретению, соответственно. В следующем предпочтительном варианте осуществления такие D-нуклеотиды могут действовать как спейсеры или линкеры, предпочтительно присоединяя модификации, такие как PEG и HES, к нуклеиновым кислотам согласно настоящему изобретению.
Также в пределах настоящего изобретения находится то, что каждая и любая из молекул нуклеиновых кислот, описанных здесь в их полноте с точки зрения их последовательности(ей) нуклеиновой кислоты, ограничена конкретной нуклеотидной последовательностью(ями). Другими словами, термины «включающий(е)» или «включает(ют)» будут интерпретироваться в таком варианте осуществления в значении «содержащий(е)» или «состоящий(е) из».
Также в пределах настоящего изобретения находится то, что нуклеиновые кислоты согласно настоящему изобретению являются частью более длинной нуклеиновой кислоты, где эта более длинная нуклеиновая кислота включает несколько частей, где по меньшей мере одной такой частью является нуклеиновая кислота или ее часть согласно настоящему изобретению. Другая(ие) часть(и) этих более длинных нуклеиновых кислот может(гут) быть одной или несколькими либо D-нуклеиновой(ыми) кислотой(ами), либо L-нуклеиновой(ыми) кислотой(ами). Любая комбинация может использоваться применительно к настоящему изобретению. Эта(и) другая(ие) часть(и) более длинной нуклеиновой кислоты может(гут) показать функцию, отличную от связывания, предпочтительно от связывания с SDF-1. Одной возможной функцией является предоставление возможности взаимодействия с другими молекулами, причем такие другие молекулы предпочтительно отличаются от SDF-1, что необходимо, например, для иммобилизации, перекрестной сшивки, детекции или амплификации. В следующем варианте осуществления настоящего изобретения нуклеиновые кислоты согласно изобретению включают в виде отдельных или объединенных фрагментов некоторые нуклеиновые кислоты по настоящему изобретению. Такая нуклеиновая кислота, включающая некоторые из нуклеиновых кислот по настоящему изобретению, также охватывается термином «более длинная нуклеиновая кислота».
L-нуклеиновые кислоты в данном документе представляют собой нуклеиновые кислоты, состоящие из L-нуклеотидов, предпочтительно полностью состоящие из L-нуклеотидов.
D-нуклеиновые кислоты в данном документе представляют собой нуклеиновые кислоты, состоящие из D-нуклеотидов, предпочтительно полностью состоящие из D-нуклеотидов.
Термины «нуклеиновая кислота» и «молекула нуклеиновой кислоты» используются в данном документе взаимозаменяемым образом, если только явно не указано обратное.
Кроме того, если только явно не указано обратное, любая нуклеотидная последовательность представлена в данном документе в направлении 5’ → 3’.
Независимо от того, состоит ли патентоспособная нуклеиновая кислота из D-нуклеотидов, L-нуклеотидов или комбинации обоих, причем данная комбинация представляет собой, например, случайную комбинацию или определенную последовательность участков нуклеотидов, состоящих из по меньшей мере одного L-нуклеотида и по меньшей мере одной D-нуклеиновой кислоты, данная нуклеиновая кислота может состоять из дезоксирибонуклеотида(ов), рибонуклеотида(ов) или их комбинаций.
Разработка патентоспособных нуклеиновых кислот в виде L-нуклеиновой кислоты выгодна по нескольким причинам. L-нуклеиновые кислоты представляют собой энантиомеры нуклеиновых кислот природного происхождения. D-нуклеиновые кислоты, однако, не очень устойчивы в водных растворах и, в частности, в биологических системах или биологических образцах из-за широко распространенного присутствия нуклеаз. Нуклеазы природного происхождения, в частности, нуклеазы из клеток животных, не способны деградировать L-нуклеиновые кислоты. Из-за этого биологическое время полужизни L-нуклеиновой кислоты в такой системе, включая организмы животного и человека, значительно возрастает. Вследствие отсутствия способности деградировать L-нуклеиновую кислоту не образуется никаких продуктов нуклеазной деградации и, таким образом, не наблюдается никаких побочных эффектов, возникающих по этой причине. Этот аспект выделяет L-нуклеиновую кислоту от фактически всех остальных соединений, которые используются в терапии заболеваний и/или нарушений, включающих присутствие SDF-1. L-нуклеиновые кислоты, которые специфически связываются с целевой молекулой посредством механизма, отличного от спаривания оснований по Уотсону и Крику, или аптамеры, которые состоят частично или полностью из L-нуклеотидов, в частности, те части аптамера, которые участвуют в связывании аптамера с целевой молекулой, также называют шпигельмерами.
В пределах настоящего изобретения находится то, что первый и второй участок нуклеотидов, фланкирующие коровую нуклеотидную последовательность, в принципе могут гибридизоваться друг с другом. В процессе такой гибридизации образуется структура двойной цепи. Любой специалист в данной области подтвердит, что такая гибридизация может как происходить, так и не происходить, в частности, в условиях in vitro и/или in vivo. Кроме того, в случае такой гибридизации не обязательной является ситуация, когда гибридизация происходит по всей длине двух участков нуклеотидов, где, по меньшей мере, исходя из правил спаривания оснований, может произойти такая гибридизация и, следовательно, образование структуры двойной спирали. В данном документе структура двойной спирали является частью молекулы или структурой, образованной двумя или более отдельными цепями, где присутствуют по меньшей мере одна, предпочтительно, две или более пары оснований, спаренных, предпочтительно, в соответствии с правилами спаривания оснований по Уотсону и Крику. Также любой специалист в данной области подтвердит, что иное спаривание оснований, такое как спаривание оснований по Хугстену (Hoogsten), может иметь место или образовывать такую структуру двойной спирали.
Также в пределах настоящего изобретения находится то, что патентоспособные нуклеиновые кислоты, независимо от того, присутствуют ли они в виде D-нуклеиновых кислот, L-нуклеиновых кислот, или D,L-нуклеиновых кислот, либо являются ли они ДНК или РНК, могут присутствовать в виде одноцепочечных или двухцепочечных нуклеиновых кислот. Как правило, патентоспособные нуклеиновые кислоты представляют собой одноцепочечные нуклеиновые кислоты, которые характеризуются определенными вторичными структурами, обусловленными первичными последовательностями, и могут, следовательно, также формировать третичные структуры. Патентоспособные нуклеиновые кислоты, однако, могут также быть двухцепочечными в том смысле, что две цепи, которые комплиментарны или частично комплиментарны друг к другу, гибридизованы друг с другом. Это придает стабильность нуклеиновой кислоте, которая, в частности, будет предпочтительной, если данная нуклеиновая кислота будет присутствовать в природной D-форме, а не L-форме.
Патентоспособные нуклеиновые кислоты могут быть модифицированы. Такие модификации могут иметь отношение к единственному нуклеотиду нуклеиновой кислоты и хорошо известны в данной области. Примеры таких модификаций описаны, среди других, Венкатсаном и др. (Venkatesan, Kim et al. 2003), а также Куссером (Kusser 2000). Такой модификацией может служить атом H, атом F или O-CH3 группа, или NH2 группа по 2’ положению индивидуального нуклеотида, из которых состоит данная нуклеиновая кислота. Кроме того, нуклеиновая кислота согласно настоящему изобретению может включать по меньшей мере один LNA нуклеотид. В некотором варианте осуществления данная нуклеиновая кислота согласно настоящему изобретению состоит из LNA нуклеотидов.
В некотором варианте осуществления нуклеиновые кислоты согласно настоящему изобретению могут быть многокомпонентной нуклеиновой кислотой. Многокомпонентная нуклеиновая кислота, как используется в данном документе, представляет собой нуклеиновую кислоту, которая состоит из по меньшей мере двух цепей нуклеиновой кислоты. Эти по меньшей мере две цепи нуклеиновой кислоты формируют функциональную единицу, где функциональная единица представляет собой лиганд для целевой молекулы. По меньшей мере две цепи нуклеиновой кислоты могут быть получены из любой патентоспособной нуклеиновой кислоты либо путем расщепления нуклеиновой кислоты, получая две цепи, либо путем синтеза одной нуклеиновой кислоты, соответствующей первой части патентоспособной, то есть полноразмерной нуклеиновой кислоты, и другой нуклеиновой кислоты, соответствующей второй части полноразмерной нуклеиновой кислоты. Следует подтвердить, что можно применять как расщепление, так и синтез для производства многокомпонентной нуклеиновой кислоты, где насчитывается более двух цепей, пример чего приводился выше. Другими словами, по меньшей мере две цепи нуклеиновой кислоты, как правило, отличаются от двух цепей, являющихся комплиментарными и гибридизованными друг с другом, хотя некоторая степень комплиментарности между различными частями нуклеиновой кислоты может существовать.
В конечном итоге также в пределах настоящего изобретения находится то, что реализована полностью закрытая, то есть кольцевая, структура нуклеиновых кислот согласно настоящему изобретению, то есть то, что нуклеиновые кислоты согласно настоящему изобретению закрыты, предпочтительно посредством ковалентного химического связывания, где более предпочтительно такое ковалентное химическое связывание происходит между 5’ концом и 3’ концом последовательностей нуклеиновой кислоты, как раскрыто в данном документе.
Авторы настоящего изобретения обнаружили, что для нуклеиновых кислот согласно настоящему изобретению характерен очень удобный диапазон значений Kd.
Возможностью определить константу связывания является исследование методом поверхностного плазмонного резонанса при помощи так называемого прибора Biacore (Biacore AB, Упсала, Швеция), который также известен специалисту в данной области. Аффинность, в данном документе, также измеряли при помощи «теста пошагового связывания», как описано в примерах. Подходящим количественным показателем для выражения интенсивности связывания между нуклеиновой кислотой и мишенью, которая в данном случае представляет собой SDF-1, является так называемая величина Kd, которая сама по себе, равно как и метод ее определения, известны специалисту в данной области.
Для нуклеиновых кислот согласно настоящему изобретению характерны определенные значения Kd. Предпочтительно, значение Kd, показанное нуклеиновыми кислотами согласно настоящему изобретению, меньше 1 мкМ. Значение Kd около 1 мкМ указано, чтобы служить характеристикой для неспецифического связывания нуклеиновой кислоты с мишенью. Как подтвердит любой специалист в данной области, значение Kd группы соединений, таких как нуклеиновые кислоты согласно настоящему изобретению, находится в пределах определенного диапазона. Вышеупомянутая Kd, величиной около 1 мкМ, представляет собой предпочтительный верхний предел значения Kd. Предпочтительный нижний предел для Kd связывающих цель нуклеиновых кислот может составлять около 10 пМ или выше. В пределах настоящего изобретения находится то, что значения Kd связывания отдельных нуклеиновых кислот с грелином находятся предпочтительно внутри этого диапазона. Предпочтительные диапазоны могут быть определены путем выбора любого первого числа внутри этого диапазона и любого второго числа внутри этого диапазона. Предпочтительные верхние значения составляют 250 нМ и 100 нМ, предпочтительные нижние значения - 50 нМ, 10 нМ, 1 нМ, 100 пМ и 10 пМ.
Молекулы нуклеиновых кислот согласно настоящему изобретению могут иметь любую длину при условии, что они все еще в состоянии связываться с целевой молекулой. В данной области технологий допускается, что существуют предпочтительные длины нуклеиновых кислот согласно настоящему изобретению. Как правило, данная длина находится между 15 и 120 нуклеотидами. Специалисты в данной области подтвердят, что любое целое число между 15 и 120 является возможной длиной для нуклеиновых кислот согласно настоящему изобретению. Более предпочтительные диапазоны длины нуклеиновых кислот согласно настоящему изобретению являются длинами от примерно 20 до 100 нуклеотидов, от примерно 20 до 80 нуклеотидов, от примерно 20 до 60 нуклеотидов, от примерно 20 до 50 нуклеотидов и от примерно 20 до 40 нуклеотидов.
В пределах настоящего изобретения находится то, что нуклеиновые кислоты, раскрытые в данном документе, включают остаток, который предпочтительно является высокомолекулярным остатком, и/или который предпочтительно позволяет изменять характеристики нуклеиновой кислоты с точки зрения, помимо прочего, времени удержания в организме животных, предпочтительно в организме человека. Особенно предпочтительным вариантом осуществления такой модификации являются модификации нуклеиновых кислот согласно настоящему изобретению с помощью PEG и HES. В данном документе PEG обозначает поли(этиленгликоль), а HES - гидроксиэтилкрахмал. Модификация с помощью PEG в данном документе является модификацией нуклеиновой кислоты согласно настоящему изобретению, где такая модификация состоит из остатка PEG, который присоединен к нуклеиновой кислоте согласно настоящему изобретению. Модификация с помощью HES, в данном документе, является модификацией нуклеиновой кислоты согласно настоящему изобретению, где такая модификация состоит из остатка HES, который присоединен к нуклеиновой кислоте согласно настоящему изобретению. Эти модификаторы, так же как процесс модификации нуклеиновой кислоты с помощью таких модификаторов, описаны в европейской заявке на патент EP 1306382, раскрытие которого включено во всей полноте в виде ссылки в данном документе.
Предпочтительно, молекулярная масса модификатора, состоящего из высокомолекулярного остатка или включающего его, составляет от примерно 2000 до 200000 Да, предпочтительно от 40000 до 120000 Да, в частности, когда таким высокомолекулярным остатком является PEG, и предпочтительно от примерно 3000 до 180000 Да, более предпочтительно от 60000 до 140000 Да, в частности, когда таким высокомолекулярным остатком является HES. Процесс модификации с помощью HES, например, описан в заявке на патент Германии DE 12004006249.8, раскрытие которого включено во всей полноте в виде ссылки в данном документе.
В пределах настоящего изобретения находится то, что как PEG, так и HES могут использоваться как в линейной, так и разветвленной формах, как далее описано в заявках на патент WO2005074993 и PCT/EP02/11950. Такая модификация может, в принципе, быть проведена с молекулами нуклеиновых кислот по настоящему изобретению по любому положению в них. Предпочтительно, такую модификацию проводят либо с 5’-концевым нуклеотидом, либо с 3'-концевым нуклеотидом и/или с любым нуклеотидом между 5’ нуклеотидом и 3’ нуклеотидом молекулы нуклеиновой кислоты.
Модификатор и, предпочтительно, остаток PEG и/или HES может быть присоединен к молекуле нуклеиновой кислоты по настоящему изобретению либо непосредственно, либо через линкер. Также в пределах настоящего изобретения находится то, что молекула нуклеиновой кислоты согласно настоящему изобретению включает один или более модификатор, предпочтительно один или более остаток PEG и/или HES. В некотором варианте осуществления отдельная молекула линкера присоединяет более одного остатка PEG или остатка HES к молекуле нуклеиновой кислоты согласно настоящему изобретению. Линкер, используемый в связи с настоящим изобретением, сам по себе может быть либо линейным, либо разветвленным. Линкеры этого типа известны специалистам в данной области и дополнительно описаны в заявках на патент WO2005074993 и PCT/EP02/11950.
Не стремясь быть связанными рамками какого-либо одного толкования, по-видимому, модифицируя нуклеиновые кислоты согласно настоящему изобретению высокомолекулярным остатком, таким как полимер, а более конкретно, полимеры, раскрытые в данном документе, которые предпочтительно являются физиологически приемлемыми, - изменяется кинетика экскреции. Более конкретно: очевидно, что из-за увеличенной молекулярной массы таких модифицированных патентоспособных нуклеиновых кислот и из-за нуклеиновых кислот, не участвующих в метаболизме, особенно, когда они находятся в L-форме, снижается экскреция из организма животных, предпочтительно из организма млекопитающих, и более предпочтительно из организма человека. Поскольку экскреция обычно происходит через почки, авторы настоящего изобретения предполагают, что скорость клубочковой фильтрации таким образом модифицированной нуклеиновой кислоты значительно снижается по сравнению с нуклеиновыми кислотами, у которых не проводилось подобной высокомолекулярной модификации, приводящей к увеличению времени пребывания в организме. В связи с вышеизложенным, особенно примечательно, что несмотря на такую высокомолекулярную модификацию специфичность нуклеиновой кислоты согласно настоящему изобретению не затронута неблагоприятным образом. На настоящий момент, у нуклеиновых кислот согласно настоящему изобретению есть такие неожиданные особенности, обычно не ожидаемые от фармацевтически активных соединений, что фармацевтический состав, обеспечивающий замедленное высвобождение, не обязательно нужен, чтобы обеспечить замедленное высвобождение. Точнее нуклеиновые кислоты согласно настоящему изобретению в их модифицированной форме, включающей высокомолекулярный остаток, уже сами по себе могут использоваться в качестве состава с замедленным высвобождением. На настоящий момент, модификатор(ы) молекул нуклеиновых кислот, как раскрыто в данном документе, и модифицированные с их участием молекулы нуклеиновых кислот, и любая композиция, включающая их же, могут обеспечивать особые, предпочтительно, контролируемые фармакокинетики и биораспределения таковых. Это также включает время удержания в кровотоке и распределение по тканям. Такие модификаторы далее описаны в заявке на патент PCT/EP02/11950.
Однако в пределах настоящего изобретения также находится то, что нуклеиновые кислоты, раскрытые в данном документе, не включают никаких модификаций и, в частности, никакую высокомолекулярную модификацию, такую как модификация с помощью PEG или HES. Такой вариант осуществления особенно предпочтителен, когда желательно быстрое выведение нуклеиновых кислот из организма после введения. Такое быстрое выведение могло бы быть желательно в случае in vivo диагностической визуализации или особых требований к терапевтической дозировке, при использовании нуклеиновых кислот или лекарственных средств, включающих то же самое, согласно настоящему изобретению.
Патентоспособные нуклеиновые кислоты, которые также упоминаются в данном документе как нуклеиновые кислоты согласно настоящему изобретению, и/или антагонисты согласно настоящему изобретению, могут использоваться для создания или изготовления лекарственного средства. Такое лекарственное средство содержит по меньшей мере одну из патентоспособных нуклеиновых кислот, необязательно вместе с дополнительными фармацевтически активными соединениями, где патентоспособная нуклеиновая кислота предпочтительно действует непосредственно как фармацевтически активное соединение. Такие лекарственные средства включают в предпочтительных вариантах осуществления по меньшей мере фармацевтически приемлемую лекарственную основу. Такая основа может представлять собой, например, воду, буфер, PBS, раствор глюкозы, раствор сахарозы, раствор маннозы, предпочтительно 5%-ый уравновешенный раствор сахарозы, крахмал, сахар, желатин или любое другое приемлемое вещество основы. Такие основы в большинстве случаев известны специалисту в данной области. Специалист в данной области подтвердит, что любые варианты осуществления, способы применения и аспекты лекарственного средства по настоящему изобретению, либо имеющие отношение к таковому, также применимы для фармацевтической композиции по настоящему изобретению и наоборот.
Показания, заболевания и нарушений, для лечения и/или профилактики которых применяют нуклеиновые кислоты, фармацевтические композиции и лекарственные средства в соответствии с настоящим изобретением, или приготовленные в соответствии с настоящим изобретением, являются результатом вовлечения, прямого или косвенного, SDF-1 в соответствующий патогенетический механизм.
Разумеется, поскольку связывающие SDF-1 нуклеиновые кислоты согласно настоящему изобретению взаимодействуют или связываются с SDF-1 человека или мыши, специалист в данной области в общих чертах поймет, что связывающие SDF-1 нуклеиновые кислоты согласно настоящему изобретению легко могут использоваться для лечения, профилактики и/или диагностики любого заболевания людей и животных, описанного в данном документе. В связи с вышеизложенным необходимо констатировать, что молекулы нуклеиновых кислот согласно настоящему изобретению могут использоваться для лечения и профилактики каких-либо заболеваний, нарушений или патологических состояний, описанных в данном документе, независимо от механизма действия, лежащего в основе такого заболевания, нарушения и патологического состояния.
Далее приводится обоснование использования молекул нуклеиновых кислот согласно настоящему изобретению в связи с различными заболеваниями, нарушениями и патологическими состояниями, тем самым делая возможной заявленную терапевтическую, профилактическую и диагностическую применимость молекул нуклеиновых кислот согласно настоящему изобретению. Чтобы избежать любого ненужного повторения, необходимо констатировать, что из-за вовлечения оси SDF-1/рецептор SDF-1, как представлено в общих чертах в связи с вышеизложенным, указанная ось может быть обращена с помощью молекул нуклеиновых кислот согласно настоящему изобретению таким образом, чтобы был достигнут требуемый терапевтический, профилактический и диагностический эффект. Кроме того, необходимо констатировать, что специфика заболеваний, нарушений и патологических состояний пациентов и любые детали режима лечения, описанные в связи с вышеизложенным, могут быть предметом предпочтительных вариантов осуществления текущей заявки.
Стимуляция стволовых клеток и/или клеток-предшественников в крови, а конкретнее, в периферической крови, является, кроме прочего, полезной при процедурах для облегчения последствий лечебных протоколов, которые оказывают отрицательное влияние на костный мозг, такие как те, что приводят к лейкопении. Таковыми являются известные побочные эффекты химиотерапии и лучевой терапии. Нуклеиновые кислоты по настоящему изобретению также увеличивают успех пересадки костного мозга, стимулируют заживление ран и лечение ожогов, и помогают при восстановлении поврежденной ткани органа. Они также оказывают противодействие бактериальным инфекциям, которые распространены при лейкозе. На настоящий момент молекулы нуклеиновых кислот согласно настоящему изобретению могут использоваться для любой такой цели, а также лечения и профилактики, соответственно, любого такого заболевания и патологического состояния.
Мобилизация стволовых клеток для регенерации тканей. В случаях поражения органов, например, из-за неправильной работы сосудистой сети или травмы, регенерация тканей желательна, но часто не достижима. Показано, что стволовые клетки костного мозга, предпочтительно - аутологические стволовые клетки, оказывают благоприятное воздействие при моделировании множества заболеваний на животных, и в некоторых случаях оказываются полезными для людей.
Репарация сетчатки и пигментного эпителия сетчатки. Происходящие из костного мозга стволовые клетки были исследованы при моделировании на мышах относящегося к сетчатке глаза сосудистого и дегенеративного заболевания. Было показано, что эти стволовые клетки присоединяются к поврежденным участкам, стабилизируют патологическую сосудистую сеть и ускоряют неоваскуляризацию гипоксических областей, будучи введенными путем инъекции в стекловидное тело или под сетчатку глаза (Friedlander et al., 2007; Otani et al., 2002). Другие авторы обнаружили, что подгруппа полученных из костного мозга стволовых клеток, которые были мобилизированы с помощью G-CSF, нацелена на участки повреждения пигментного эпителия сетчатки глаза. Было обнаружено, что помимо системного введения препарата ни сбор, ни внутриглазная инъекция стволовых клеток не были необходимы. Напротив, привлечение клеток к участку ранения/повреждения было достигнуто путем кратковременной ограниченной суперэкспрессии хемокинов, например, SDF-1 (Li et al., 2007; Li et al., 2006).
Сердечная репарация. Накопленные данные указывают на то, что миокард отвечает на рост или ранение рекрутингом стволовых клеток и/или клеток-предшественников, которые участвуют в процессах репарации и регенерации. Моделированием на мышах инфаркта миокарда Франсиоли и др. показали, что клетки c-kit+, которые являются стволовыми клетками, аккумулируются в области инфаркта через 1-2 недели после инфаркта, вероятнее всего для того, чтобы способствовать процессам репарации (Fransioli et al., 2008). Даун и др. сообщили о потенциальной терапевтической эффективности происходящих из костного мозга Sca-1+/Lin-/CD45-очень маленьких, подобных эмбриональным, стволовых клеток (сокр. VSEL). При моделировании на мышах инфаркта миокарда (сокр. MI) трансплантации относительно небольшого количества CD45-VSEL было достаточно, чтобы улучшить функцию левого желудочка, и облегчило гипертрофию мышечных клеток после MI (Даwn et al., 2008).
Репарация и снижение интенсивности воспаления после ишемического инсульта. Швартинг и др. исследовали действие инъекции меченых зеленым флуоресцентным пептидом Lin(-)-гемопоэтических стволовых клеток на размер инфаркта, апоптозный некроз клеток, постишемическое воспаление и транскрипцию гена цитокина у моделей на мышах ишемической травмы головного мозга. Спустя 24 часа после инъекции клетки были найдены в селезенке, а позже - в ишемической паренхиме мозга, экспрессирующими микроглиальные, но не нейронные маркерные белки. Оценка повреждения ткани показала значительно меньшие зоны инфаркта, и меньший апоптозный некроз нейронов в периинфарктных участках у животных, обработанных Lin(-)-гемопоэтическими стволовыми клетками. Анализ инфильтрации иммунных клеток в ишемические полушария обнаружил сокращение проникновения T-клеток и макрофагов у обработанных мышей (Schwarting et al., 2008). В работе Imitola et al. ранее сообщалось, что нервные стволовые клетки привлекались к местам поражения центральной нервной системы (сокр. CNS) с помощью SDF-1α/CXCR4 метаболического пути.
В свете этих результатов мобилизация достаточного числа стволовых клеток костного мозга, вероятно, будет выгодна, чтобы вызвать процессы репарации (Tang et al., 2007). Блокировка SDF-1 или его рецептора CXCR4 является многообещающим подходом в этой области, поскольку это мобилизирует множество стволовых клеток костного мозга. Правильные дозировки, схемы применения и потенциальная локализация мобилизированного средства могут быть определены квалифицированным специалистом в данной области, выполняющим стандартные экспериментальные работы, где предпочтительно предполагается, что стволовые клетки являются мобилизированными, но все еще способны отвечать на хемотаксические сигналы так, что они могут быть рекрутированы к местам повреждения ткани.
Поскольку связывающие SDF-1 молекулы нуклеиновой кислоты согласно настоящему изобретению ингибируют передачу сигнала между SDF-1 и рецептором SDF-1, такие связывающие SDF-1 молекулы нуклеиновой кислоты можно использовать для создания лекарственного средства, которое может быть применено для мобилизации клеток-предшественников и/или стволовых клеток в периферическую кровь, и/или для лечения заболеваний и/или нарушений, предпочтительно выбранных из группы, включающей: заживление ран; ожоги; нарушения, вызванные или связанные с поврежденной тканью органа и/или поврежденной сосудистой системой, где такие нарушения отобраны из повреждений сетчатки и сосудистой оболочки глаза, инсульта, поражения миокарда, инфаркта миокарда, ишемии после трансплантации органа и травматического повреждения; и гемопоэтические нарушения, где такие нарушения отобраны из апластической анемии, лейкоза, лекарственной анемии и лейкопении, а также бактериальной инфекции при лейкопении.
Для мобилизации клеток-предшественников и/или стволовых клеток данное лекарственное средство может применяться в сочетании со вторым фармацевтически активным средством, где функцией указанного второго фармацевтически активного средства является мобилизация клеток-предшественников и/или стволовых клеток. Мобилизирующие клетки средства без ограничений выбирают из гранулоцитарно-моноцитарного колониестимулирующего фактора (сокр. GM-CSF), интерлейкина-8 (сокр. IL-8), воспалительного белка макрофагов (сокр. MIP), ростового онкогена, блокирующих CXCR4 средств, таких как AMD3001 (Aiuti, Webb et al. 1997) и гранулоцитарного колониестимулирующего фактора (сокр. G-CSF).
В случаях поражения CNS использование макромолекулярного ингибитора SDF-1 для мобилизации происходящих из костного мозга стволовых клеток может быть особенно выгодным. Хотя клетки и остаются чувствительными для градиентов SDF-1, они маскируются макромолекулярным ингибитором SDF-1 в периферийных тканях, пока в организме поддерживается достаточно высокая концентрация. Однако так как макромолекулярный ингибитор SDF-1 не может пройти гематоэнцефалический барьер, градиенты SDF-1, возникшие вследствие суперэкспрессии SDF-1 на участках гипоксии или поражений CNS, сохраняются в CNS, и могут привлекать некоторые из клеток, мобилизированные к участку повреждения, где они могут выполнять свое назначение, как описано выше. В предпочтительном варианте осуществления такой макромолекулярный ингибитор SDF-1 представляет собой молекулу нуклеиновой кислоты согласно настоящему изобретению.
Сочетание блокировки SDF-1 и химиотерапии. Приблизительно 20% клеток B-линии дифференцировки острых лимфообластных лейкемий неизлечимы традиционной химиотерапией. Преклинические эксперименты, использующие ex-vivo лейкозные клетки в совместной культуре со стромальными клетками, предоставили свидетельства, что взаимодействие лейкозных бластных клеток с компонентами костного мозга защищает лейкозные клетки от химиотерапии (Mudry, Fortney et al. 2000; Garrido, Appelbaum et al. 2001; Tabe, Jin et al. 2007). Есть также сообщения о моделях на мышах, у которых ингибирование молекул на клеточных поверхностях, которые важны для адгезии клеток и, тем самым, для хоминга (злокачественных) гемопоэтических клеток, улучшает эффективность химиотерапии и приводит к уничтожению лейкозных стволовых клеток (Matsunaga, Takemoto et al. 2003; Jin, Hope et al. 2006). SDF-1, как известно, играет важную роль при хоминге и удержании стволовых клеток в нишах костного мозга. Существуют накопленные преклинические, и одно сообщение о клиническом доказательстве, что блокирование CXCR4, рецептора SDF-1 у гемопоэтических клеток, также приводит к высвобождению бластных клеток острого миелолейкоза из костного мозга в периферическую кровь, где они могут служить мишенями для химиотерапии (например, с помощью средства цитарабин) (Fierro, Brenner et al. 2008) или других средств, приводящих к гибели опухолевых клеток (например, биологические препараты сами по себе или в сочетании с антителозависимой клеточноопосредованной цитотоксичностью или комплементзависимой цитотоксичностью). Дополнительно, Чжин и др. недавно заметили, что лечение хронического миелолейкоза с помощью ингибиторов тирозинкиназы (например, иматиниба) приводит к активации CXCR4 на лейкозных клетках. В результате это приводит к возросшему хомингу в костном мозге и вызывает блок G0-G1 цикла, что приводит клетки в состояние неподвижности и нечувствительности к дальнейшим попыткам химиотерапии (Jin, Tabe et al. 2008). В свете вышесказанного, настоящее изобретение предлагает комбинированную терапию химиотерапевтическим средством вместе с CXCR4 или с ингибитором SDF-1, таким как связывающие SDF-1 молекулы нуклеиновой кислоты согласно настоящему изобретению, включая, помимо прочего, NOX-A12-JE40, чтобы уменьшить хоминг лейкозных клеток в костном мозге и мобилизировать неподвижные лейкозные клетки, которые вернулись назад в костный мозг до терапии. Из-за потери сигналов ниши клетки могут развиваться в направлении клеточного цикла и поэтому становятся более чувствительными к химиотерапии. Химиотерапия и соответствующие средства, применяемые в связи с вышеизложенным, известны в данной области и, например, представляют собой антитела, такие как ритуксимаб, ибритумомаб тиуксетан, тозитумомаб; алкилирующие средства, такие как цисплатин и карбоплатин, также как и оксалиплатин, мехлорэтамин; циклофосфамид, хлорамбуцил; антиметаболиты, такие как пурин, азатиоприн, меркаптопурин; растительные алкалоиды и терпеноиды, такие как алкалоиды барвинка и таксаны; подофиллотоксин; эпотилон; и ингибиторы топоизомеразы, такие как камптотецины.
Поэтому лекарственное средство, как раскрыто в данном документе, может использоваться для мобилизации раковых клеток в периферическую кровь субъекта, где раковые клетки отобраны без каких-либо ограничений из группы лейкозных клеток, миеломных клеток, лимфомных клеток, раковых стволовых клеток, раковых клеток с метастатическим потенциалом и метастазов рака.
Лекарственное средство согласно настоящему изобретению можно применять в сочетании со вторым лекарственным средством или вторым фармацевтически активным средством, которое можно применять для мобилизации раковых клеток в периферическую кровь субъекта. Второе фармацевтически активное средство включает мобилизующие клетки средства, как было ранее раскрыто в данном документе.
Кроме того, лекарственные средства согласно настоящему изобретению можно применять в сочетании с третьим лекарственным средством или третьим фармацевтически активным средством, где данное третье фармацевтически активное средство повреждает, уничтожает и/или метит раковые клетки. Такие уничтожающие раковые клетки лекарственные средства являются предпочтительно выбранными без каких-либо ограничений из группы, включающей антитела, такие как ритуксимаб, ибритумомаб тиуксетан, тозитумомаб; алкилирующие средства, такие как цисплатин и карбоплатин, также как и оксалиплатин, мехлорэтамин; циклофосфамид, хлорамбуцил; антиметаболиты, такие как пурин, азатиоприн, меркаптопурин; растительные алкалоиды и терпеноиды, такие как алкалоиды барвинка и таксаны; подофиллотоксин; эпотилон; ингибиторы топоизомеразы, такие как камптотецины.
Третье лекарственное средство или фармацевтически активное средство обладает химиотерапевтическим действием или может его оказать.
Лекарственные средства, метящие раковые клетки, приводят к активации защитных сил организма, направленных к меченой таким образом раковой клетке, где лекарственные средства, метящие раковые клетки, без каких-либо ограничений отобраны из группы моноклональных антител. Они работают, нацеливаясь на опухолеспецифичные антигены, тем самым усиливая иммунный ответ клетки-хозяина на опухолевые клетки, с которыми соединяется само средство. Примерами являются трастузумаб (торговое наименование герцептин), цетуксимаб и ритуксимаб (торговые наименования: ритуксан или мабтера).
Комбинированная терапия лекарственным средством согласно настоящему изобретению может проводиться со вторым и/или третьим лекарственным средством.
Субъект, которого лечили лекарственным средством для мобилизации раковых клеток, может быть последовательно или одновременно подвергнут лучевой терапии. В некотором варианте осуществления лучевая терапия может использоваться как альтернативное лечение вместо третьего лекарственного средства или третьего фармацевтически активного средства.
Данное лекарственное средство согласно настоящему изобретению, в сочетании со вторым лекарственным средством или вторым фармацевтически активным средством, либо без него, с третьим лекарственным средством или третьим фармацевтически активным средством, либо без него, и с лучевой терапией, либо без нее, может применяться для лечения и/или профилактики рака, предпочтительно - солидных опухолей и гематологических злокачественных новообразований, более предпочтительно - лейкоза, лимфомы, и миеломы, где предпочтительно данное лекарственное средство согласно настоящему изобретению применяют в сочетании с третьим лекарственным средством или лучевой терапией.
Долгоживущие плазмоциты, мобилизация B-клеток и T-клеток памяти при аутоиммунном заболевании
B-клетки и/или T-клетки памяти возвращаются из организма в костный мозг и, возможно, другие места, например, лимфоузлы, и/или удерживаются там градиентом SDF-1, который образован благодаря экспрессии SDF-1 стромальными клетками (Parretta, Cassese et al. 2005; Zhang, Nakajima et al. 2005; Radbruch, Muehlinghaus et al. 2006). В нишах эти клетки находятся в состоянии покоя, где они не чувствительны к модифицирующим заболевание лекарственным средствам (например, цитостатические лекарственные средства или метотрексат), которые обычно используются для лечения аутоиммунного заболевания. Как только они покидают нишу, они могут быстро делиться и начать повышать восстановленный иммунный ответ, если они встречаются с родственным им антигеном, что может привести к бессрочному аутоиммунному заболеванию. Вмешательство связывающих SDF-1 нуклеиновых кислот в градиент SDF-1 в костном мозге и других нишах или блокирование CXCR4 может привести к мобилизации B-клеток и/или T-клеток и делает возможным их элиминацию из крови посредством афереза или их таргетирование с соответствующим лекарственным средством.
После острого иммунного ответа с активным производством антител окончательно дифференцированными B-клетками (плазмоциты) большинство этих клеток умирают с разрешением воспалительного процесса или с устранением чужеродного антигена (например, вирусного белка). Однако существует меньшинство B-клеток памяти, которые могут превратиться в лимфобласты с помощью T-клетки. У этих лимфобластов есть способность в ответ на хемотаксические стимулы мигрировать в ниши, где они выживают в виде долгоживущих плазмоцитов «памяти» в течение нескольких лет. Во время перемещения клетки теряют экспрессию CXCR5 и CCR7 и больше не мигрируют к соответствующим лигандам. Экспрессия CXCR4, однако, поддерживается. Таким образом, данные клетки будут двигаться в сторону участков экспрессии SDF-1, где они могут встретиться с долгосрочными сигналами выживания, которые могут быть отличными от SDF-1 или дополнительными к нему (Minges Wols et al., 2007). Эти ниши могут находиться как в костном мозге, так и в периферийных органах, например, в воспаленных тканях. Долгоживущие плазмоциты вероятнее всего ответственны за сохранение защитных титров антител плазмы, которые часто сохраняются в течение многих лет после инфекции или вакцинации (Tarlinton et al., 2008). Плазмоциты не столь же восприимчивы к изменяющим течение заболевания лекарственным средствам, как другие B- и T-клетки, вероятнее всего потому, что они не делятся. Кроме того, они не могут быть предназначены для лечения антителом против CD20, так как плазмоциты не несут CD20. При аутоиммунных заболеваниях, которые поддерживаются аутоантителами, секретируемыми плазмоцитами и долгоживущими плазмоцитами, было бы выгодно элиминировать эти клетки так, чтобы иммунная система прекратила повреждать собственную ткань. Процедуру очистки крови пациента от плазмоцитов можно осуществить аферезом, например, с помощью антитела против CD138 (синдекан-1) на аффинной матрице (например, колонка или гранулы) (Minges Wols & Witte, 2008; Wijdenes et al., 1996). Для того чтобы устранить не только плазмоциты в кровотоке, но также и резидентные долгоживущие плазмоциты, было бы предпочтительно мобилизировать последние из их ниш. Это можно проделать системным введением молекул связывающей SDF-1 нуклеиновой кислоты согласно настоящему изобретению, что подавляет градиенты SDF-1 и мобилизирует эти клетки, которые двигаются в сторону SDF-1 в периферическую кровь.
Следовательно, данное лекарственное средство согласно настоящему изобретению может использоваться для мобилизации долгоживущих плазмоцитов, B-клеток и/или T-клеток в периферическую кровь субъекта.
Лекарственное средство согласно настоящему изобретению можно применять в сочетании со вторым лекарственным средством или вторым фармацевтически активным средством, которое можно применять для мобилизации долгоживущих плазмоцитов, B-клеток и/или T-клеток в периферическую кровь субъекта. Второе лекарственное средство или второе фармацевтически активное средство содержит мобилизующие клетки средства, как было ранее раскрыто в данном документе, но не ограничивается только ими.
Хотя данные клетки мобилизированы с помощью только лишь лекарственного средства согласно настоящему изобретению или в сочетании со вторым лекарственным средством или фармацевтически активным средством, как описано выше, они могут быть очищены из крови с помощью афереза, клеточного сортинга и/или проточной цитометрии (например, с помощью клеточного сортинга с активацией флуоресценции [FACS] и/или клеточного сортинга с магнитной активацией [MACS] с подходящими маркерами поверхности долгоживущих плазмоцитов, B-клеток или T-клеток).
Кроме того, лекарственное средство согласно настоящему изобретению можно применять в сочетании с третьим лекарственным средством или третьим фармацевтически активным средством, где данное третье лекарственное средство или третье фармацевтически активное средство повреждает, уничтожает и/или метит долгоживущие плазмоциты, B-клетки и/или T-клетки в периферической крови. Такие лекарственные средства, повреждающие долгоживущие плазмоциты, B-клетки и/или T-клетки, выбирают без ограничений из каких-либо изменяющих течение заболевания лекарственных средств, например, метотрексат или цитотоксические средства. Примеры лекарственных средств, метящих долгоживущие плазмоциты, B-клетки и/или T-клетки, приводящих к активации защитных сил организма, направленных на меченные, как показано выше, долгоживущие плазмоциты, B-клетки и/или T-клетки, без каких-либо ограничений включают выбранные из группы, состоящей из ритуксимаба, антител, связывающих рецептор IL-6, или антител, связывающих синдекан-1.
Комбинированную терапию с помощью лекарственного средства согласно настоящему изобретению можно проводить со вторым и/или с третьим лекарственным средством.
Субъект, обработанный в соответствии с настоящим изобретением лекарственным средством согласно настоящему изобретению для мобилизации долгоживущих плазмоцитов, B-клеток и/или Т-клеток, может одновременно или последовательно проходить лучевую терапию, которая повреждает или уничтожает делящиеся клетки иммунной системы пациента, которая в конечном счете поражает саму себя. Лучевая терапия может использоваться как альтернатива лечению с помощью третьего лекарственного средства. Употребление лекарственного средства согласно настоящему изобретению в сочетании со вторым лекарственным средством или без него, либо второго фармацевтически активного средства с третьим лекарственным средством или без него, либо третьего фармацевтически активного средства с лучевой терапией или без нее, может использоваться для профилактики и/или лечения аутоиммунных заболеваний, мобилизация долгоживущих плазмоцитов, B-клеток и/или Т-клеток по отдельности или в рамках более комплексной терапевтической концепции может быть полезной для лечения различных заболеваний, включая помимо прочего,
системные аутоиммунные заболевания , где такое системное аутоиммунное заболевание предпочтительно выбрано из аллергии, аутоиммунной гемолитической анемии с комбинированным синдромом холодовых и тепловых агглютининов, синдрома системной воспалительной реакции, геморрагического шока, сахарного диабета 1го типа, диффузной склеродермии, полихондрии, аутоиммунного плюригландулярного синдрома, системной эритематозной волчанки и ее проявлений, ревматоидного артрита, ревматических заболеваний глаза, мозга, легких, почек, сердца, печени, желудочно-кишечного тракта, селезенки, кожи, костей, лимфатической системы, крови или других органов;
аутоиммунные заболевания желудочно-кишечного тракта , где такое аутоиммунное заболевание желудочно-кишечного тракта предпочтительно выбрано из болезни Крона, язвенного колита, заболевания брюшной полости, глютеновой энтеропатии, воспалительного заболевания кишечника, панкреатита, эозинофильного эзофагита;
аутоиммунные заболевания кожи , где такое аутоиммунное заболевание кожи предпочтительно выбрано из псориаза, крапивницы, дерматомиозита, обыкновенной пузырчатки, листовидной пузырчатки, буллезного пемфигоида, кольцевидной/лентовидной склеродермии, витилиго, герпетиформного дерматита или болезни Дюринга, склерозирующего лишая;
аутоиммунные заболевания сосудистой системы , где такое аутоиммунное заболевание сосудистой системы предпочтительно выбрано из васкулитов, предпочтительно височного артериита, васкулита, сосудистого истечения, ревматической полимиалгии, атеросклероза, синдрома Черджа-Стросса, синдрома Такаясу, синдрома Гудпасчера, предпочтительно, главным образом затрагивающего почки, конкретнее – почечные клубочки, и/или также главным образом затрагивающего легкие, гломерулонефрита, узелкового полиартериита, болезни Бехчета;
аутоиммунные заболевания нервной системы , где такое аутоиммунное заболевание нервной системы предпочтительно выбрано из рассеянного склероза, хронической воспалительной демиелинизирующей полинейропатии, нейрокогнитивной дисфункции, синдрома мышечной скованности, синдрома Гийена-Барре, тяжелой псевдопаралитической миастении, синдрома Ламберта-Итона;
аутоиммунные заболевания опорно-двигательного аппарата , где такое аутоиммунное заболевание опорно-двигательного аппарата предпочтительно выбрано из анкилозирующего спондилита, саркоидоза, ревматической полимиалгии, полимиозита, псориатического артрита, ревматического полиартрита, полихондрии, фибромиалгии, ювенильного ревматоидного артрита, болезни Лайма, реактивного артрита, спондилоартропатии, дегенеративного заболевания сустава;
и другие аутоиммунные заболевания, где такие другие аутоиммунные заболевания предпочтительно выбраны из синдрома Когана, аутоиммунного адреналита, болезни Меньера, местных воспалений, гнездной алопеции, острых воспалительных заболеваний, первичного биллиарного цирроза, синдрома Шегрена, склеродермий, таких как диффузная склеродермия, CREST-синдром и/или кольцевидная/лентовидная склеродермия, аутоиммунного увеита, тиреоидита Хашимото, базедовой болезни, аутоиммунного гепатита, гломерулонефрита, антифосфолипидного синдрома, идиопатического фиброза легких, аутоиммунного бесплодия, заболеваний иммунного комплекса и перитонита.
Кроме того, употребление лекарственного средства согласно настоящему изобретению в сочетании со вторым лекарственным средством или вторым фармацевтически активным средством, либо без них, может использоваться для лечения и/или профилактики реакции «трансплантат против хозяина» после пересадки костного мозга и отторжения трансплантата пересаженным органом, где такой орган выбран из печени, почки, кишечника, легких, сердца, кожи, конечностей, роговой оболочки глаза, островков Лангерганса, костного мозга, кровеносных сосудов и поджелудочной железы.
При моделировании на мышах аллергического заболевания дыхательных путей, антитело, имеющее целью CXCR4 на лейкоцитах, уменьшало гиперреактивность дыхательных путей и эозинофилию легких, указывая, что CXCR4-опосредованные сигналы способствуют воспалению легких (Gonzalo, Lloyd et al. 2000). Есть также свидетельство, что SDF-1 способствует ангиогенезу. Клинические и экспериментальные данные указывают, что инфильтрирующие кожу лейкоциты играют важную роль в инициировании и поддержании атопического дерматита, и было показано, что SDF-1 представляет собой важный фактор рекрутинга T-лимфоцитов и дендритных клеток (Gombert, Dieu-Nosjean et al. 2005). Так как лейкоциты и, конкретно, T-клетки экспрессируют рецептор SDF-1, CXCR4, и демонстрируют хемотаксический ответ на градиенты SDF-1, нарушение этих градиентов путем связывания SDF-1 и нейтрализации нуклеиновой кислоты согласно настоящему изобретению годится для помощи при воспалительных заболеваниях с чрезмерным воспалением бактериального или вирусного происхождения, либо без такового, при воспалении легких и/или кожи, предпочтительно – при псориазе.
Ревматоидный артрит вероятно является системным аутоиммунным заболеванием, которое обычно начинается в малых суставах кистей рук и стоп. Его признаком является воспаление синовиальной мембраны, для которого характерна инфильтрация мембраны и окружающей ткани лейкоцитами, особенно макрофагами, T-клетками и B-клетками. Этот процесс, а также секреция протеаз и провоспалительных цитокинов приводят к уплотнению синовиальной мембраны и росту рыхлой ткани, называемой паннус. Он пролиферирует вокруг сустава и инвазивно - в кость и хрящ, приводя к их необратимому разрушению. Паннус вызывает формирование новообразованных сосудов для своего собственного кровоснабжения. Ивамото и др. недавно обнаружили, что в синовиальной ткани и синовиальной жидкости пациентов с RA повышено содержание целого ряда хемокинов, среди которых находится и SDF-1 (Iwamoto et al., 2008). Активация SDF-1 также была показана ранее in vitro, путем добавления синовиальной жидкости от пациентов с RA к культивируемым фибробласт-подобным синовиоцитам. Эта жидкость или добавление IL-17 индуцировали экспрессию в синовиоцитах мРНК SDF-1. Индукцию экспрессии SDF-1 можно отменить добавлением антител против IL17 (Kim et al., 2007). В случае воспаленного сустава SDF-1 может действовать тройственным образом. Во-первых, как хемоаттрактант для лейкоцитов, во-вторых, как аттрактант для эндотелиальных клеток-предшественников, которые необходимы для ангиогенеза, или в-третьих, как пусковой сигнал экспрессии расположенного в 5'-3' направлении фактора роста, такого как VEGF, который затем приводит к росту новообразованных сосудов.
На основании этих экспериментов авторы настоящего изобретения предполагают, что лекарственное средство, используемое для ингибирования миграции лейкоцитов, предпочтительно из периферической крови к ткани, может обеспечить лечение и/или профилактику заболевания и/или нарушения, как изложено ниже.
Миграция лейкоцитов может быть инициирована чужеродными антигенами пересаженных органов, где пересаженные органы выбраны из печени, почки, кишечника, легких, сердца, кожи, конечностей, роговой оболочки глаза, островков Лангерганса, костного мозга, кровеносных сосудов и поджелудочной железы. По этой причине лекарственное средство, которое приводит к ингибированию миграции лейкоцитов, может использоваться для профилактики и/или лечения отторжения трансплантатов пересаженных органов, как раскрыто в данном документе.
Кроме того, миграция лейкоцитов может быть инициирована воспалением, которое происходит при или связано с:
системными аутоиммунными заболеваниями , где такое системное аутоиммунное заболевание предпочтительно выбрано из аллергии, аутоиммунной гемолитической анемии с комбинированным синдромом холодовых и тепловых агглютининов, синдрома системной воспалительной реакции, геморрагического шока, сахарного диабета 1го типа, диффузной склеродермии, полихондрии, аутоиммунного плюригландулярного синдрома, системной эритематозной волчанки и ее проявлений, ревматоидного артрита, ревматических заболеваний глаза, мозга, легких, почек, сердца, печени, желудочно-кишечного тракта, селезенки, кожи, костей, лимфатической системы, крови или других органов;
аутоиммунными заболеваниями желудочно-кишечного тракта , где такое аутоиммунное заболевание желудочно-кишечного тракта предпочтительно выбрано из болезни Крона, язвенного колита, заболевания брюшной полости, глютеновой энтеропатии, воспалительного заболевания кишечника, панкреатита, эозинофильного эзофагита;
аутоиммунными заболеваниями кожи , где такое аутоиммунное заболевание кожи предпочтительно выбрано из псориаза, крапивницы, дерматомиозита, обыкновенной пузырчатки, листовидной пузырчатки, буллезного пемфигоида, кольцевидной/лентовидной склеродермии, витилиго, герпетиформного дерматита или болезни Дюринга, склерозирующего лишая;
аутоиммунными заболеваниями сосудистой системы , где такое аутоиммунное заболевание сосудистой системы предпочтительно выбрано из васкулитов, предпочтительно – височного артериита, васкулита, сосудистого истечения, ревматической полимиалгии, атеросклероза, синдрома Черджа-Стросса, синдрома Такаясу, синдрома Гудпасчера, предпочтительно, главным образом затрагивающего почки, конкретнее, почечные клубочки, и/или также главным образом затрагивающего легкие, гломерулонефрита, узелкового полиартериита, болезни Бехчета;
аутоиммунными заболеваниями нервной системы, где такое аутоиммунное заболевание нервной системы предпочтительно выбрано из рассеянного склероза, хронической воспалительной демиелинизирующей полинейропатии, нейрокогнитивной дисфункции, синдрома мышечной скованности, синдрома Гийена-Барре, тяжелой псевдопаралитической миастении, синдрома Ламберта-Итона;
аутоиммунными заболеваниями опорно-двигательного аппарата, где такое аутоиммунное заболевание опорно-двигательного аппарата предпочтительно выбрано из анкилозирующего спондилита, саркоидоза, ревматической полимиалгии, полимиозита, псориатического артрита, ревматического полиартрита, полихондрии, фибромиалгии, ювенильного ревматоидного артрита, болезни Лайма, реактивного артрита, спондилоартропатии, дегенеративного заболевания сустава;
другими аутоиммунными заболеваниями , где такие другие аутоиммунные заболевания предпочтительно выбраны из синдрома Когана, аутоиммунного адреналита, болезни Меньера, местных воспалений, гнездной алопеции, острых воспалительных заболеваний, первичного биллиарного цирроза, синдрома Шегрена, склеродермий, таких как диффузная склеродермия, CREST-синдром и/или кольцевидная/лентовидная склеродермия, аутоиммунного увеита, тиреоидита Хашимото, базедовой болезни, аутоиммунного гепатита, гломерулонефрита, антифосфолипидного синдрома, идиопатического фиброза легких, аутоиммунного бесплодия, заболеваний иммунного комплекса и перитонита.
По этой причине лекарственное средство, которое приводит к ингибированию или снижению миграции лейкоцитов, может использоваться для профилактики и/или лечения воспаления, которое происходит при аутоиммунных заболеваниях или связано с ними, как раскрыто в данном документе.
Как было показано при моделировании на мышах аллергического заболевания дыхательных путей, лекарственное средство, приводящее к ингибированию миграции лейкоцитов, может быть эффективным при лечении и/или профилактике аллергических реакций кожи и/или слизистой оболочки дыхательных путей, предпочтительно - поллиноза, астмы, гиперреактивности дыхательных путей и/или дерматита, предпочтительно контактного дерматита и/или атопического дерматита.
Мобилизация клеток-предшественников и/или стволовых клеток нуклеиновой кислотой согласно настоящему изобретению предусматривает способ получения этих клеток от субъекта. При этом эффективное количество молекул нуклеиновых кислот согласно изобретению вводят субъекту, что приводит к мобилизации клеток-предшественников и/или стволовых клеток в периферическую кровь субъекта. Клетки субъекта могут быть собраны из периферической крови субъекта, соответственно, аферезом, лейкаферезом, клеточным сортингом и/или проточной цитометрией, как раскрыто в данном документе.
После сбора клеток-предшественников и/или стволовых клеток у субъекта, данный субъект, предпочтительно первый субъект, последовательно или одновременно может быть подвергнут химиотерапии или лучевой терапии.
Химиотерапия и лучевая терапия обычно затрагивают быстро делящиеся клетки. Их используют для лечения рака, потому что раковые клетки делятся чаще, чем большинство здоровых клеток. Однако, поскольку клетки костного мозга также часто делятся, обработки высокими дозами могут серьезно повредить или разрушить костный мозг пациента. Без здорового костного мозга пациент больше не в состоянии производить клетки крови, необходимые для переноски кислорода, борьбы с инфекцией и предотвращения кровотечения. Трансплантация стволовых клеток периферической крови заменяет стволовые клетки, которые были разрушены в процессе лечения. Здоровые, пересаженные стволовые клетки могут восстановить способность костного мозга производить клетки крови, необходимые пациенту.
Трансплантация стволовых клеток периферической крови чаще всего используется при лечении лейкоза и лимфомы. Она наиболее эффективна, когда лейкемия или лимфома находятся в стадии ремиссии (признаки и симптомы рака исчезли). Трансплантация стволовых клеток периферической крови также используется при лечении других раковых новообразований, таких как нейробластома (рак, который возникает в незрелых нервных клетках и поражает, главным образом, младенцев и детей), и множественная миелома. Исследователи апробируют трансплантацию стволовых клеток периферической крови в клинических испытаниях (исследованиях) для лечения различных типов рака (NCI, 2001).
Как показано на фиг.41 и 43, некоторые типы рака, такие как лейкемия и солидные опухоли, лимфопролиферативные нарушения и доброкачественные нарушения, такие как аутоиммунные заболевания, гемопоэтические нарушения можно лечить трансплантацией аллогенных и/или аутологических гемопоэтических стволовых клеток (сокр. HSCT) (Gratwohl, Baldomero et al. 2002).
Если химиотерапию или лучевую терапию проводили следующим шагом предпочтительно у первого субъекта, при этом химиотерапию или лучевую терапию предпочтительно проводили или назначали для того, чтобы повредить или разрушить опухолевые клетки, то собранные клетки-предшественники и/или стволовые клетки предпочтительно первого субъекта можно ввести обратно в периферическую кровь предпочтительно первого субъекта. С другой стороны, второй субъект может предоставить собранные у него клетки-предшественники и/или стволовые клетки первому субъекту, у которого низкий уровень клеток-предшественников и/или стволовых клеток, либо те клетки-предшественники и/или стволовые клетки были разрушены ранее, например, при химиотерапии или лучевой терапии.
Данный способ может использоваться для лечения рака, предпочтительно солидных опухолей, гематологических опухолей или злокачественных новообразований.
При аутоиммунном заболевании, включая помимо прочего, например, красную волчанку, ревматоидный артрит, особенно в тяжелых случаях, один из вариантов терапии состоит в том, чтобы провести курс лечения с отделением миелоцитов или более селективный - с отделением лимфоцитов, и после этого возвратить пациенту гемопоэтическую систему (Burt, Marmont et al. 2006). Гемопоэтические стволовые клетки и гемопоэтические клетки-предшественники, необходимые для пересадки гемопоэтической системы, эффективно иммобилизовали молекулами нуклеиновых кислот согласно настоящему изобретению, включая помимо прочего, SDF-1 связывающий шпигельмер NOX-A12-JE40, или с помощью другой стратегии блокирования SDF-1 или его соответствующего рецептора CXCR4.
Как вариант, гемопоэтические стволовые клетки/гемопоэтические клетки-предшественники собирают из крови и используют для реконструкции иммунной системы пациента после удаления гемопоэтической системы, включая аутореактивные лимфоциты, с помощью химиотерапии и/или лучевой терапии. По существу, на этом основано радикальное лечение аутоиммунного заболевания.
Мобилизация долгоживущих плазмоцитов, B-клеток и/или T-клеток нуклеиновой кислотой согласно настоящему изобретению предусматривает способ извлечения этих клеток от субъекта. С помощью этого эффективное количество молекул нуклеиновых кислот согласно изобретению вводят субъекту, что приводит к мобилизации долгоживущих плазмоцитов, B-клеток и/или T-клеток в периферическую кровь субъекта. Предпочтительно, собранные T-клетки являются T–клетками памяти. Клетки могут быть удалены у субъекта из периферической крови субъекта, соответственно, аферезом, лейкаферезом, клеточным сортингом и/или проточной цитометрией, как раскрыто в данном документе. Предпочтительно, такое удаление проводят проточной цитометрией с поверхностными маркерами, подходящими для указанных клеток.
Данный способ сбора клеток-предшественников и/или стволовых клеток и долгоживущих плазмоцитов, B-клеток и/или T-клеток, соответственно, может быть использован для лечения и/или профилактики системных аутоиммунных заболеваний, аутоиммунных заболеваний желудочно-кишечного тракта, аутоиммунных заболеваний кожи, аутоиммунных заболеваний сосудистой системы, аутоиммунных заболеваний нервной системы, аутоиммунных заболеваний опорно-двигательного аппарата и других аутоиммунных заболеваний, как раскрыто в данном документе.
Диабетическая нефропатия (сокр. DN) является главной причиной конечной стадии почечного заболевания. Хотя ингибиторы ангиотензина могут предотвратить прогрессирование заболевания во многих случаях, не существует никакой приемлемой терапии при отсутствии реакции. При DN клубочковый пучок подвергается медленной, но прогрессирующей структурной модификации, характеризуемой клубочковой гипертрофией, нодозной и диффузной аккумуляцией внеклеточного мезангиального матрикса и повреждением подоцитов. Последнее, как думают, является причиной прогрессирования микроальбуминурии на ранних стадиях до выраженной протеинурии и гломерулосклероза на последних стадиях DN. Начало и прогрессирование DN вызывает многочисленные патологические механизмы, включая отложение конечных продуктов глубокого гликозилирования, эндотелиальную дисфункцию и повышенную локальную экспрессию факторов роста и провоспалительных медиаторов. Как правило, хемокины принадлежат последней группе факторов, потому что определенные хемокины провоцируют воспаление путем рекрутинга и активации иммунных клеток при DN, как и при других типах заболеваний почек. Например, направленная делеция или терапевтическое блокирование моноцитарного хемоаттрактантного белка MCP-1/CCL2 может предотвратить гломерулосклероз, блокируя рекрутинг макрофагов в клубочках мышей с диабетом типа 1 или типа 2 (T1D/T2D) (Chow et al., 2007; Chow et al., 2006; Kulkarni et al., 2007).
Ничего не известно о SDF-1 при DN. DN лишена либо пролиферации подоцитов, либо аутоиммунитета, следовательно, вышеупомянутые исследования вряд ли способны оценить преобладающую функциональную роль SDF-1 при DN. Однако у авторов настоящего изобретения есть причины предполагать, что прогрессирующее реструктурирование клубочковой структуры до гломерулосклероза, морфологического варианта заживления ран, могло бы включать передачу сигналов SDF-1. На основании доступных данных по другим стадиям заболевания остается неясным, защищает ли SDF-1 преимущественно от DN, например, сохраняя целостность ткани и поддерживая регенерацию, либо SDF-1 преимущественно провоцирует DN, например, усугубляя гломерулосклероз. Данные авторов настоящего изобретения, использовавших связывающую SDF-1 нуклеиновую кислоту NOX-A12-JE40 (SEQ ID NO:132) в качестве молекулы, представляющей молекулы нуклеиновых кислот согласно настоящему изобретению, при моделировании на мышах диабетической нефропатии, подтверждают последнее, что указывает на ранее неизвестный патологический механизм гломерулосклероза, и на SDF-1 как потенциальную терапевтическую мишень при DN в примере 12. NOX-A12-JE40 (SEQ ID NO:132) поэтому может быть полезным как терапевтическое средство для лечения или профилактики диабетической нефропaтии. Механизм действия также может быть потенциально опосредован мобилизацией полученных из костного мозга клеток-предшественников и/или стволовых клеток (Ito et al., 2001). Поэтому нуклеиновые кислоты SDF-1, раскрытые в данном документе, могут использоваться для изготовления лекарственного средства, где данное лекарственное средство предназначено для лечения и/или профилактики нефропaтии и предпочтительно - диабетической нефропaтии.
Легочная артериальная гипертензия.
Легочная гипертензия (сокр. PH) представляет собой серьезное заболевание малопонятной этиологии, для которого характерно повышенное давление в легочной артерии, приводящее к прогрессирующей правосторонней сердечной недостаточности и, в конечном итоге, к смерти. PH происходит в результате утолщения интимы мелких легочных резистивных артерий, которое в свою очередь происходит, по крайней мере, частично, в результате дисфункции и пролиферации эндотелиальных и гладкомышечных клеток. Повышенная пролиферация эндотелиальных клеток в сосудах и мускуляризация сосудистой системы являются патологическими особенностями ремоделирования легочных сосудов, и было продемонстрировано, что этот процесс связан с индуцированным гипоксией производством ангиогенных факторов, воспалительных медиаторов и вазоконстрикторов. Ямадзи-Кеган и др. обнаружили в ex vivo исследовании органной культуры легкого мыши, что CXCL12/SDF-1 был активирован и может быть вовлечен во внутрилегочный рекрутинг клеток кровотока суперэкспрессированным индуцированным гипоксией митогенетическим фактором (сокр. HIMF) (Yamaji-Kegan, Su et al. 2006). Также было описано влияние SDF-1 на ремоделирование сосудов вообще и при легочной артериальной гипертензии в частности (Schober and Zernecke 2007). Поэтому блокирование SDF-1 SDF-связывающими нуклеиновыми кислотами, как раскрыто в данном документе, может использоваться для лечения и/или профилактики гипертензии, предпочтительно - легочной гипертензии, конкретнее, легочной артериальной гипертензии.
Идиопатический фиброз легких
Легочная ткань пациентов с идиопатическим фиброзом легких по описаниям имеет более высокие численности клеток, экспрессирующих как SDF-1, так и CXCR4, чем нормальные легкие. В модели на мышах блеомицин-индуцированного фиброза легких Ксу и др. приводят данные, показывающие, что введение антагониста CXCR4 (TN14003) значительно уменьшало фиброз легкого (Xu, Mora et al. 2007). Поэтому блокирование SDF-1 SDF-связывающими нуклеиновыми кислотами раскрыто настолько, что может использоваться для лечения и/или профилактики идиопатического фиброза легких.
Фиброз при заживлении ран
После ожоговой травмы в коже крысы, свиньи и человека, как было показано, происходит суперэкспрессия SDF-1. Хотя это и имеет определенные преимущества в течение короткого периода времени после ранения, как полагают, это скорее стимулирует притяжение лейкоцитов (например, эозинофилов) и фиброз, а не способствует эпителизации (Avniel, Arik et al. 2006). Посредством ингибирования CXCR4 или SDF-1 связывающими SDF-1 нуклеиновыми кислотами, как раскрыто в данном документе, можно получить заживление раны с меньшим фиброзом.
Как описано ранее, было показано, что SDF-1 участвует в хоминге эндотелиальных клеток к сосудистой оболочке глаза во время неоваскуляризации в глазной ткани, где точная роль этих клеток все еще исследуется (Sengupta, Caballero et al. 2005). Однако авторы настоящего изобретения смогли показать с помощью двух независимых животных моделей, что блокирование SDF-1 SDF-1-связывающими нуклеиновыми кислотами согласно настоящему изобретению, как раскрыто в данном документе, приводит к сокращению неоваскуляризации.
Модель на животных «вызванной лазером хороидальной неоваскуляризации» применяли, чтобы оценить действие экспериментальных лекарственных препаратов на новообразованные сосуды сетчатки и сосудистой оболочки глаза. Это происходит при таких заболеваниях, как влажная или «пролиферативная» форма возрастной макулярной дегенерации (сокр. AMD), диабетическая ретинопатия и окклюзия вен сетчатки глаза. Как было показано, CXCR4 экспрессируется при индуцированной лазером CNV (Lima e Silva et al., FASEB J. 21: 2007). Он был локализован совместно с клетками, экспрессирующими CD45 и F4/80, означая то, что эти клетки представляют собой полученные из BM макрофаги. Ингибиторы CXCR4 уменьшали индуцированную лазером CNV. Но не был исследован случай, если клетки CXCR4 также экспрессируют SDF1. Как успешно показано в примере 11 в данном документе, связывающая SDF-1 нуклеиновая кислота NOX-A12-JE40 (SEQ ID NO:132), как представитель молекул нуклеиновых кислот согласно настоящему изобретению, блокирует неоваскуляризацию у моделей CNV на животных.
Модель индуцированной кислородом ретинопатии на мышах представляет собой модель для имитации индуцированной гипоксией неоваскуляризации сетчатки, наблюдаемой при DR, особенно при пролиферативной DR, и при AMD (Smith, Wesolowski et al. 1994). Модель также называют моделью ретинопатии недоношенности, поскольку недоношенные дети, помещенные в инкубаторы в больницах, теряют зрение из-за слишком длительного пребывания в инкубаторах в условиях повышенного содержания кислорода, что приводит к аномальному росту сосудов в сетчатке глаза во время пребывания в инкубаторе и после их возвращения к условиям с нормальным содержанием кислорода. Как описано в примере 14, при моделировании на мышах связывающая SDF-1 нуклеиновая кислота NOX-A12-JE40 (SEQ ID NO:132), как представитель молекул нуклеиновых кислот согласно настоящему изобретению, значительно ингибирует формирование сосудистого пучка и тем самым улучшает общую оценку ретинопатии на день P17 наблюдений, как раскрыто в данном документе.
Кроме того, как в показано в примере 9, в стандартном тесте на ангиогенез в органной культуре, - тесте аортального кольца на прорастание сосудов, - связывающая SDF-1 нуклеиновая кислота 193-G2-012-5'-PEG (NOX-A12-JE40 (SEQ ID NO:132), как представитель молекул нуклеиновых кислот согласно настоящему изобретению, блокирует индуцированное с помощью SDF-1 прорастание.
По этой причине связывающие SDF-1 нуклеиновые кислоты, как раскрыто в данном документе, могут использоваться для производства лекарственного средства, где данное лекарственное средство предназначено для лечения заболеваний и/или нарушений, которые вызывают ангиогенез и/или неоваскуляризацию, предпочтительно хороидальную неоваскуляризацию. Моделирование неоваскуляризации на животных показало, что связывающие SDF-1 нуклеиновые кислоты, раскрытые в данном документе, могут использоваться как лекарственное средство для лечения заболеваний и/или нарушений, выбранных из группы, включающей заболевания сетчатки, предпочтительно возрастной макулярной дегенерации, диабетической ретинопатии, окклюзии вен в сетчатке, макулярного отека и отека сосудов сетчатки глаза.
Неоваскуляризация предпочтительно определяется в данном документе как образование функциональных капиллярных сетей с перфузией красных кровяных телец. Неоваскуляризация отличается от ангиогенеза тем, что для ангиогенеза, главным образом, характерны протрузия и разрастание капиллярных зачатков и отростков от ранее существовавших кровеносных сосудов.
Ингибирование отека сосудов сетчатки глаза
В процессе возрастной макулярной дегенерации (AMD), диабетической ретинопатии и окклюзии сосудов в сетчатке часто наблюдается формирование макулярного отека. Обычно усиленное подтекание местной сосудистой системы является причиной образования отека. Часто это представляет собой последствие воспалительных процессов, которое изменяет архитектуру сосудистой системы или структуру недоразвитых, подтекающих новообразованных сосудов.
Формирование макулярного отека может привести к быстрому ухудшению остроты зрения из-за того, что нарушена поставка питательных веществ и других сигнальных молекул к сетчатке глаза.
SDF-1 вносит вклад в несколько факторов, которые могут привести к формированию отека. Посредством ингибирования экспрессии белка непроницаемых перегородок, окклюдина (Butler et al., 2005), стенки сосудов могут расслабляться. Это, кроме того, вызывает инвазию лейкоцитов, что может создать провоспалительные окружающие условия, и это может стимулировать экспрессию VEGF (Liang et al., 2007; Salcedo et al., 1999), который сначала был известен как «фактор подтекания сосудов», поскольку его присутствие приводит к формированию подтекающих сосудов.
Было неясно, приведет ли ингибирование SDF-1, например, SDF-1-связывающими нуклеиновыми кислотами, как раскрыто в этом изобретении, к какому-нибудь физиологическому эффекту, поскольку SDF-1 в здоровых глазах экспрессируется только на низких уровнях (Lima e Silva et al., 2007).
Одна связывающая SDF-1 нуклеиновая кислота была проверена на модели индуцированного с помощью VEGF подтекания сосудов сетчатки глаза кролика. Подтекание, которое наблюдали с помощью флуорометрии, возникало не вследствие прямого кратковременного эффекта VEGF-инъекции в стекловидное тело, а скорее за счет каскадов реакций, которые активируются и реагируют более медленно (Edelman et al., 2005). Связывающая SDF-1 нуклеиновая кислота значительно уменьшала подтекание сосудов сетчатки глаза дозозависимым образом.
Ингибирование васкулогенеза в дополнение к неблагоприятному исходу VEGF-ингибирования или после него
Недавно Рэдди и др. показали, что SDF-1 может способствовать росту опухолевых сосудов даже при небольшом присутствии VEGF. Вероятно, это является вторым, VEGF-независимым каскадом реакций для стимуляции новообразованных сосудов (Reddy, Zhou et al. 2008). Вмешиваясь в передачу сигналов SDF-1, связывающие SDF-1 нуклеиновые кислоты, такие как те, что соответствуют настоящему изобретению, могут по этой причине быть полезны в качестве лечения, направленного против ангиогенеза. Это может быть особенно эффективно для не ответивших на анти-VEGF терапию пациентов, невосприимчивых к анти-VEGF терапии пациентов, или при комбинированной терапии с лекарственными средствами против VEGF для всех показаний, которые включают ангиогенез, конкретнее, при пролиферативном заболевании сетчатки глаза, где пролиферативные заболевания сетчатки глаза отобраны из AMD, DR и окклюзии вен в сетчатке, а также при раке, предпочтительно солидных опухолях и метастазах.
Лекарственные средства, ингибирующие функцию VEGF, без каких-либо ограничений включают бевацизумаб (авастин), пегаптаниб (макуген) и ранибизумаб (луцентис).
На настоящий момент связывающие SDF-1 молекулы нуклеиновых кислот, раскрытые в данном документе, могут использоваться для производства лекарственного средства, где данное лекарственное средство предназначено для комбинированной терапии с ингибирующим VEGF лекарственным средством и/или для применения к субъектам, которые отвечают слабо или не отвечают совсем на терапию с ингибирующим VEGF лекарственным средством. Термин «слабо», используемый в данном документе в связи с ответом на любую терапию в связи с любым аспектом и вариантом осуществления настоящей заявки, в этом контексте означает, что в течении заболевания не достигнуто никакой ремиссии.
WHIM синдром
Синдром WHIM (хроническая нециклическая нейтропения) представляет собой иммунодефицитное состояние, которое часто характеризуется укороченной формой рецептора CXCR4. Это влечет за собой повышенную сенсибилизацию к рецепторному лиганду SDF-1 (CXCL-12) и, таким образом, к более сильному хемотаксису. По этой причине, чтобы достигнуть нормальной направленной миграции стволовых клеток, является выгодным понизить биологически активную концентрацию SDF-1 в организме с помощью блокирующего SDF-1 средства, такого как связывающая SDF-1 молекула нуклеиновой кислоты согласно настоящему изобретению, или использовать блокатор рецептора CXCR4 (Lagane, Chow et al. 2008).
Разделение и/или истощение таких клеток, как клетки-предшественники и/или стволовые клетки, долгоживущие плазмоциты, B-клетки памяти и/или T-клетки памяти из организма, предпочтительно из крови, может быть выполнено с помощью афереза, клеточного сортинга и проточной цитометрии.
Аферез представляет собой технологию, в которой кровь субъекта проводят через аппарат, который выделяет один конкретный компонент или группу компонентов, и возвращает остаток в кровоток. В зависимости от веществ и/или клеток, которые требуется удалить, при аферезе использовали различные процессы, включая сбор стволовых клеток, процедуры абсорбции и аффинной хроматографии.
Лейкаферез представляет собой лабораторную процедуру, при которой белые кровяные клетки отделяют от образца крови. Это может быть сделано, чтобы уменьшить очень высокий счет белых кровяных клеток у больных раком (лейкемия), или удалить белые кровяные клетки для переливания.
Клеточный сортинг представляет собой процесс, где смешанные популяции клеток разделяют на две или более популяции, как показано на примере для проточной цитометрии, предпочтительно с помощью клеточного сортинга с активацией флуоресценции (FACS) и клеточного сортинга с магнитной активацией (MACS).
Проточная цитометрия представляет собой метод подсчета, контроля и сортировки микроскопических частиц наподобие различных популяций клеток, суспендированных в потоке жидкости. Она позволяет одновременный мультипараметрический анализ физических и/или химических особенностей одиночных клеток, протекающих через оптическое и/или электронное детектирующее устройство. Клеточный сортинг с активацией флуоресценции является специализированным типом проточной цитометрии. Он обеспечивает способ сортировки гетерогенной смеси биологических клеток в два или более контейнера, по одной клетке за один прием, основанный на специфическом рассеянии света и особенностях флуоресценции каждой клетки. Флуоресцентное окрашивание частицы/клетки может быть проведено посредством инкубации частиц/клеток с флуоресцентным красителем, который связывается с частицей/клеткой. Он представляет собой полезный научный прибор, поскольку он обеспечивает быструю, объективную и количественную регистрацию флуоресцентных сигналов от отдельных клеток, так же как и физическое разделение клеток, представляющих особый интерес. Акроним FACS зарегистрирован как торговая марка и принадлежит Becton Dickinson. Клеточный сортинг с магнитной активацией (MACS) является названием торговой марки, зарегистрированным Miltenyi Biotec, для способа разделения различных популяций клеток в зависимости от их поверхностных антигенов (CD молекулы). В связи с этим смесь клеток, которые необходимо разделить, инкубируют с магнитными гранулами, покрытыми антителами против специфического поверхностного антигена. Это заставляет клетки, экспрессирующие этот антиген, присоединяться к магнитным гранулам. Впоследствии клеточный раствор наносят на колонку, помещенную в сильное магнитное поле. На этой стадии клетки, прикрепленные к гранулам (экспрессирующие антиген), остаются на колонке, хотя другие клетки (не экспрессирующие антиген) элюируются. Посредством этого способа клетки могут быть разделены по положительному или отрицательному сродству к специфическому(им) антигену(ам).
В следующем варианте осуществления лекарственное средство включает дополнительное фармацевтически активное средство. Такими дополнительными фармацевтически активными соединениями могут быть те, что известны специалистам в данной области и являются предпочтительно выбранными из группы, включающей антагонисты хемокинов или цитокинов, кортикостероиды, и т.п. Любому специалисту в данной области понятно, что приведенные различные показания в соответствии с настоящим изобретением могут быть реализованы с помощью нуклеиновых кислот согласно настоящему изобретению, указанное(ые) дополнительное(ые) фармацевтически активное(ые) средство(а) может(гут) быть любым(и) из тех, что в принципе являются подходящими для лечения и/или профилактики такого заболевания. Молекулы нуклеиновых кислот согласно настоящему изобретению, особенно если они присутствуют или используются в качестве лекарственного средства, предпочтительно комбинируют с гранулоцит-макрофаговым колониестимулирующим фактором (GM-CSF), интерлейкином-1 (IL-1), интерлейкином-3 (IL-3), интерлейкином-8 (IL-8), PIXY-321 (GM-CSF/IL-3 химерный белок), воспалительным белком макрофага (MIP), фактором стволовых клеток, тромбопоэтином и ростовым онкогеном, в виде индивидуальных средств, либо в комбинации.
В качестве альтернативы или в дополнение к этому, такое дополнительное фармацевтически активное средство представляет собой дополнительную нуклеиновую кислоту согласно настоящему изобретению. В качестве альтернативы, данное лекарственное средство включает по меньшей мере еще одну нуклеиновую кислоту, которая связывается с целевой молекулой, отличной от SDF-1, или характеризуется функцией, отличной от таковой для нуклеиновых кислот согласно настоящему изобретению.
Любой специалист в данной области подтвердит, что патентоспособные нуклеиновые кислоты могут использоваться фактически при любом заболевании, где антагонист SDF-1 может быть введен пациенту, которому такой антагонист необходим, и такой антагонист подходит для устранения причины заболевания или нарушения, либо, по крайней мере, сводит к минимуму последствия заболевания или нарушения. Такое последствие включает без каких-либо ограничений патологическую неоваскуляризацию, воспаление и метастазирование. Применимость нуклеиновых кислот согласно настоящему изобретению в связи с этими и другими заболеваниями или нарушениями вытекает, среди прочего, из вовлеченности SDF-1, как обрисовано в общих чертах во вводной части настоящего патентного описания, которое приводится здесь в виде ссылки, чтобы избежать любого ненужного повторения.
В одном варианте осуществления лекарственного средства по настоящему изобретению такое лекарственное средство предназначено для использования в комбинации с другими способами лечения для любого из заболеваний, раскрытых в данном документе, особенно тех, для которых данное лекарственное средство по настоящему изобретению должно использоваться.
«Комбинированная терапия» (или «совместная терапия») включает введение лекарственного средства по данному изобретению и по меньшей мере второго средства в качестве части соответствующего курса лечения, предназначенного для обеспечения положительного эффекта совместного действия этих терапевтических средств, то есть лекарственного средства по настоящему изобретению и указанного второго средства. Положительный эффект комбинации включает без каких-либо ограничений фармакокинетическое или фармакодинамическое совместное действие, следующее из комбинации терапевтических средств. Комбинированное введение этих терапевтических средств, как правило, выполняется в течение определенного периода времени (обычно минуты, часы, дни или недели, в зависимости от выбранной комбинации).
«Комбинированная терапия» может быть, но обычно не может быть, предназначена для осуществления введения двух или более этих терапевтических средств как части отдельных курсов монотерапии, что случайно и произвольно приводит к комбинациям по настоящему изобретению. «Комбинированная терапия» имеет в виду включение в себя введения этих терапевтических средств последовательным образом, а именно, когда каждое терапевтическое средство вводят в различное время, так же как и введения этих терапевтических средств, или по меньшей мере двух из терапевтических средств, фактически одновременным образом. Фактически одновременного введения можно добиться, например, вводя субъекту единственную капсулу, имеющую постоянную пропорцию каждого терапевтического средства, или единственную капсулу, имеющую много отделов для каждого терапевтического средства.
Последовательное или фактически одновременное введение каждого терапевтического средства может быть произведено любым соответствующим способом введения, включая, помимо прочего, местные способы введения, пероральные способы введения, внутривенные способы введения, внутримышечные способы введения и прямое поглощение через ткани мембран слизистой оболочки. Терапевтические средства можно вводить одним и тем же способом введения или различными способами введения. Например, первое терапевтическое средство данной комбинации можно вводить посредством инъекции, хотя другие терапевтические средства комбинации можно вводить наружно.
Альтернативно, например, все терапевтические средства можно вводить наружно или все терапевтические средства можно вводить посредством инъекции. Последовательность, в которой вводят терапевтические средства, не строго критична, если не отмечено иначе. «Комбинированная терапия» также может включать введение терапевтических средств описанным выше способом в дополнительной комбинации с другими биологически активными компонентами. Там, где комбинированная терапия дополнительно включает немедикаментозное лечение, данное немедикаментозное лечение может проводиться в любое подходящее время столь долго, сколько потребуется для достижения положительного эффекта от совместного действия комбинации терапевтических средств и немедикаментозного лечения. Например, в соответствующих случаях положительный эффект тем не менее был достигнут, когда немедикаментозное лечение было временно передвинуто от момента введения терапевтических средств, возможно на дни или даже недели.
Как изложено в общих чертах выше, лекарственное средство согласно настоящему изобретению может быть введено, в принципе, в любой форме, известной специалистам в данной области. Предпочтительный способ введения представляет собой систематическое введение, более предпочтительно - парентеральное введение, предпочтительно - посредством инъекции. Альтернативно, лекарственное средство может быть введено местно. Другие способы введения включают внутримышечный, внутрибрюшинный и подкожный, пероральный, интраназальный, интратрахеальный или легочный, при этом предпочтение отдается такому способу введения, который наименее агрессивен, хотя и гарантирует эффективность.
Парентеральное введение обычно используется для подкожных, внутримышечных или внутривенных инъекций и инфузий. Кроме того, один подход для парентерального введения предусматривает имплантацию систем замедленного высвобождения или непрерывного высвобождения, гарантирующих поддержание постоянного уровня дозировки, которые известны специалисту в данной области.
Кроме того, предпочтительные лекарственные средства по настоящему изобретению могут быть введены в интраназальной форме посредством наружного применения подходящих интраназальных основ, лекарственных форм для ингаляции, или посредством трансдермальных способов введения, применяя те формы трансдермальных пластырей, которые известны специалистам в данной области. Конечно, для того чтобы быть введенной в форме трансдермальной системы доставки, доза введения скорее должна быть непрерывна, чем прерывиста, на протяжении всей схемы приема лекарственного средства. Другие предпочтительные наружные препараты включают кремы, мази, лосьоны, аэрозольные спреи и гели, в которых концентрация активного компонента обычно колеблется от 0,01% до 15%, масс./масс. или масс./об.
В дополнение к прямому введению субъекту, предпочтительные лекарственные средства по настоящему изобретению могут использоваться в протоколах лечения ex vivo, чтобы приготовить клеточные культуры, применяемые затем для пополнения запасов клеток крови субъекта. Лечение ex vivo может быть проведено на аутологических клетках, собранных из периферической крови, или костного мозга, или аллотрансплантатов от подобранных доноров. Концентрация предпочтительных лекарственных средств по настоящему изобретению или в комбинации с другими средствами, такими как воспалительный белок макрофагов, находится в пределах компетенции специалиста в данной области.
Субъекты, реагирующие на способ по данному изобретению, включают как медицинских, так и ветеринарных субъектов вообще, включая здоровых и больных людей. Среди других субъектов, для которых полезны способы и средства по данному изобретению, находятся кошки, собаки, крупные млекопитающие, птицы, такие как цыплята, и т.п. Вообще говоря, любому субъекту, которому полезно повышение количества клеток-предшественников и/или стволовых клеток, или чьи клетки-предшественники и/или стволовые клетки желательны для трансплантации стволовых клеток, подходит введение по способу изобретения.
Типичные патологические состояния, для которых можно добиться улучшения или иным образом принести пользу с помощью способа и средств по данному изобретению, включают гемопоэтические нарушения, такие как апластическая анемия, лейкозы, индуцированные лекарственными средствами анемии и гемопоэтические недостаточности вследствие лучевой терапии или химиотерапии. Способ и средства по данному изобретению также полезны для более успешных трансплантаций во время и после иммуносупрессивных терапий, так же как и в проведении более эффективного заживления ран и лечения бактериального воспаления. Способ по настоящему изобретению дополнительно полезен для лечения субъектов с иммунной недостаточностью или с иным образом ослабленной иммунной системой. Типичные патологические состояния, для которых можно добиться улучшения или иным образом принести пользу с помощью способа по настоящему изобретению, включают тех субъектов, кто заражен ретровирусами, а конкретнее, зараженных вирусом иммунодефицита человека (ВИЧ). Способ по данному изобретению, таким образом, предназначается для широкого спектра патологических состояний, для которых подходящим является повышение количества клеток-предшественников и/или стволовых клеток у субъекта, либо где будет полезен сбор клеток-предшественников и/или стволовых клеток для последующей трансплантации стволовых клеток.
Нуклеиновую кислоту по настоящему изобретению, в некотором варианте осуществления, также вводят, чтобы восстановить миокард с помощью мобилизированных стволовых клеток костного мозга.
Лекарственное средство по настоящему изобретению обычно содержит эффективное количество активного(ых) компонента(ов) терапии, включая, помимо прочего, молекулу нуклеиновой кислоты по настоящему изобретению, растворенную или диспергированную в фармацевтически приемлемой среде. Фармацевтически приемлемые среды или лекарственные основы включают любые и все растворители, дисперсионные среды, покрытия, противомикробные и противогрибковые средства, изотонические и сдерживающие абсорбцию средства и т.п. Употребление таких сред и средств для фармацевтически активных веществ известно в данной области. Дополнительные активные компоненты могут также быть включены в лекарственное средство по настоящему изобретению.
В дополнительном аспекте настоящее изобретение имеет отношение к фармацевтической композиции. Такая фармацевтическая композиция включает, по меньшей мере, одну из нуклеиновых кислот согласно настоящему изобретению и предпочтительно - фармацевтически приемлемый связующий компонент. Таким связующим компонентом может служить любой связующий компонент, используемый и/или известный в данной области. Более подробно, такой связующий компонент представляет собой любой из связующих компонентов, обсуждавшихся в связи с изготовлением лекарственного средства, раскрытых в данном документе. В дополнительном варианте осуществления фармацевтическая композиция включает дополнительное фармацевтически активное средство.
Изготовление лекарственного средства и фармацевтической композиции с учетом настоящего раскрытия будет известно специалистам в данной области. Как правило, такие составы могут быть изготовлены в виде средств для инъекций, либо в виде жидких растворов, либо в виде суспензий; сухих форм, подходящих для растворения, или суспендирования в жидкости перед инъекцией; в виде таблеток или других твердых частиц для перорального введения; в виде капсул пролонгированного действия; или в любой другой форме, используемой в настоящее время, включая глазные капли, кремы, лосьоны, бальзамы, средства для ингаляции и т.п. Употребление стерильных составов, таких как составы для мытья на основе физиологического раствора, хирургами, врачами или патронажными медицинскими работниками для обработки конкретных поверхностей в операционном поле, может также быть особенно полезным. Композиции могут также быть доставлены посредством микрозонда, микрочастицы или губки.
Если исходить из технологии изготовления состава, лекарственное средство может быть введено способом, соответствующим дозируемым составам, и в том количестве, при котором это будет фармакологически эффективно. Составы свободно вводят во множестве лекарственных форм, таких как растворы для инъекций такого рода, как описано выше, но также могут применяться капсулы с высвобождением лекарственного средства и им подобные.
В этом контексте количество активного компонента и объем композиции, которые должны быть введены, зависят от подлежащего лечению человека или субъекта. Конкретное количество активных соединений, требуемое для введения, зависит от назначения практикующего врача и является индивидуальным для каждого человека.
Обычно используется минимальный объем лекарственного средства, необходимого для диспергирования активных соединений. Подходящие схемы введения также различны, но их необходимо индивидуализировать, первоначально вводя соединения и контролируя результаты, а затем давая дополнительно контролируемые дозы через новые интервалы.
Например, для перорального введения в форме таблетки или капсулы (например, желатиновой капсулы), активный лекарственный компонент, то есть молекула нуклеиновой кислоты по настоящему изобретению и/или любое дополнительное фармацевтически активное средство, также называемое в данном документе как терапевтическое(ие) средство(а) или активное(ые) соединение(я), может(гут) быть объединен(ы) с пероральной, нетоксичной, фармацевтически приемлемой инертной лекарственной основой, такой как этанол, глицерин, вода и т.п. Кроме того, по желанию или необходимости, подходящие связующие компоненты, смазывающие вещества, дезинтегрирующие средства и красящие средства могут также быть включены в смесь. Подходящие связующие компоненты включают крахмал, алюмосиликат магния, крахмальную пасту, желатин, метилцеллюлозу, натрий карбоксиметилцеллюлозу и/или поливинилпирролидон, сахара природного происхождения, такие как глюкоза или бета-лактоза, сахаристые вещества из кукурузы, натуральные и синтетические смолы, такие как гуммиарабик, трагакантовая камедь или альгинат натрия, полиэтиленгликоль, воски и т.п. Смазывающие вещества, используемые в этих лекарственных формах, включают олеат натрия, стеарат натрия, стеарат магния, бензоат натрия, ацетат натрия, хлорид натрия, диоксид кремния, тальк, стеариновую кислоту, ее магниевые или кальциевые соли и/или полиэтиленгликоль и т.п. Дезинтегрирующие средства включают, без ограничения, крахмал, метилцеллюлозу, агар, бентонит, крахмалы ксантановой камеди, агар, альгиновую кислоту или ее натриевую соль, или шипучие смеси, и т.п. Разбавители включают, например, лактозу, декстрозу, сахарозу, маннит, сорбит, целлюлозу и/или глицин.
Лекарственное средство по данному изобретению может быть также введено в таких пероральных лекарственных формах, как таблетки или капсулы, пилюли, порошки, гранулы, эликсиры, настойки, суспензии, сиропы и эмульсии пролонгированного действия и замедленного высвобождения. Суппозитории изготавливают преимущественно из жирных эмульсий или суспензий.
Фармацевтическая композиция или лекарственное средство может быть стерилизована и/или содержать вспомогательные лекарственные вещества, такие как консервирующие, стабилизирующие, увлажняющие или эмульгирующие средства, способствующие растворению факторы, соли для регулировки осмотического давления и/или буферы. Кроме того, они могут также содержать другие терапевтически ценные вещества. Данные композиции изготавливают согласно обычным способам смешивания, гранулирования или нанесения покрытий, и они обычно содержат от примерно 0,1% до 75%, предпочтительно от примерно 1% до 50%, активного компонента.
Жидкие, в частности, вводимые путем инъекции, композиции могут быть, например, изготовлены посредством растворения, диспергирования и т.д. Активное соединение растворяют или смешивают с фармацевтически чистым растворителем, таким как, например, вода, физиологический раствор, водный раствор декстрозы, глицерин, этанол и т.п., чтобы с их помощью образовать раствор или суспензию для инъекций. Дополнительно могут быть составлены сухие формы, подходящие для растворения в жидкости перед инъекцией.
Для сухих композиций наполнители включают фармацевтические сорта маннита, лактозы, крахмала, стеарата магния, сахарина натрия, талька, целлюлозы, глюкозы, сахарозы, карбоната магния, и т.п. Активное соединение, определенное выше, может быть также составлено в форме суппозиториев, используя, например, полиалкиленгликоли, например, пропиленгликоль, в качестве лекарственной основы. В некоторых вариантах осуществления суппозитории изготавливают преимущественно из жирных эмульсий или суспензий.
Лекарственные средства и молекулы нуклеиновых кислот, соответственно, по настоящему изобретению могут быть также введены в форме липосомных систем доставки, таких как малые моноламеллярные везикулы, большие моноламеллярные везикулы и мультиламеллярные везикулы. Липосомы могут быть сформированы из множества фосфолипидов, содержащих холестерин, октадециламин или фосфатидилхолины. В некоторых вариантах осуществления пленка липидных компонентов гидратируется водным раствором лекарственного средства до формы липидного слоя, заключающего лекарственное средство в капсулу, что хорошо известно специалисту в данной области. Например, молекулы нуклеиновых кислот, описанные в данном документе, могут быть изготовлены в виде комплекса с липофильным соединением или с неиммуногенным, высокомолекулярным соединением, сконструированного с помощью способов, известных в данной области. Дополнительно, липосомы могут переносить такие молекулы нуклеиновых кислот на своей поверхности для нацеливания и переноса цитотоксического средства внутри себя до уничтожения клетки-посредника. Пример комплексов, ассоциированных с нуклеиновой кислотой, представлен в патенте США № 6011020.
Лекарственные средства и молекулы нуклеиновых кислот, соответственно, по настоящему изобретению могут быть также присоединены к растворимым полимерам в виде нацеливаемых лекарственных основ. Такие полимеры могут включать поливинилпирролидон, сополимер пирана, полигидроксипропил-метакриламидфенол, полигидроксиэтил-аспанамидфенол или полиэтиленоксид-полилизин, замещенный пальмитоильными остатками. Кроме того, лекарственные средства и молекулы нуклеиновых кислот, соответственно, по настоящему изобретению могут быть присоединены к классу биодеградируемых полимеров, полезных для достижения управляемого высвобождения лекарственного средства, например, полимолочная кислота, полиэпсилонкапролактон, полигидроксимасляная кислота, сложные полиортоэфиры, полиацетали, полидигидропираны, полицианакрилаты, и перекрестно-сшитые или амфифильные блоксополимеры гидрогелей.
Фармацевтическая композиция и лекарственное средство, соответственно, для успешного введения необязательно могут также содержать незначительное количество нетоксичных вспомогательных веществ, таких как увлажняющие или эмульгирующие средства, поддерживающие pH средства, и другие вещества, такие как, например, ацетат натрия и олеат триэтаноламина.
Схема приема лекарственного средства, использующая молекулы нуклеиновых кислот и лекарственные средства, соответственно, по настоящему изобретению, отобрана в соответствии с множеством факторов включая тип, биологический вид, возраст, вес, пол и медицинское состояние пациента; серьезность патологического состояния, подлежащего лечению; способ введения; почечная и печеночная функция пациента; и какой конкретный аптамер или его соль используется. Врач или ветеринар обычной квалификации могут без труда определить и предписать эффективное количество лекарственного средства, необходимого для предотвращения, противодействия или угнетения прогрессирования патологических состояний.
Эффективные уровни нуклеиновых кислот в плазме, согласно настоящему изобретению, предпочтительно находятся в пределах от 500 фМ до 500 мкМ при лечении любых заболеваний, раскрытых в данном документе.
Молекулы нуклеиновых кислот и лекарственные средства, соответственно, по настоящему изобретению предпочтительно могут быть введены в единственной дозе ежедневно, каждый второй или третий день, еженедельно, каждую вторую неделю, в единственной дозе ежемесячно или каждый третий месяц.
В пределах настоящего изобретения находится то, что лекарственное средство, описанное в данном документе, составляет фармацевтическую композицию, раскрытую в данном документе.
В дополнительном аспекте настоящее изобретение имеет отношение к способу лечения субъекта, которому такое лечение необходимо, где данный способ включает введение фармацевтически активного количества, по крайней мере, одной из нуклеиновых кислот согласно настоящему изобретению. В некотором варианте осуществления субъект страдает от заболевания или подвержен риску развития такого заболевания, где данное заболевание является любым из раскрытых в данном документе, в частности, любым из заболеваний, раскрытых в связи с употреблением любой из нуклеиновых кислот согласно настоящему изобретению для производства лекарственного средства.
В данном документе термин «лечение» включает в предпочтительном варианте осуществления дополнительную или альтернативную профилактику и/или последующее наблюдение.
В данном документе термины «заболевание» и «нарушение» должны использоваться взаимозаменяемым образом, если не указано обратное.
В данном документе термин «включает» предпочтительно не предназначен для ограничения содержания предметной области, следующего вслед за таким термином или описываемого им. Однако в альтернативном варианте осуществления термин «включает» должен быть понят в значении «содержит» и, таким образом, как ограничение содержания предметной области, следующего вслед за таким термином или описываемого им.
Различные SEQ ID NO, химическая природа молекул нуклеиновых кислот согласно настоящему изобретению и целевых SDF-1 молекул, используемых в данном документе, их фактическая последовательность и внутренний номер ссылки суммированы в следующей таблице.
Нужно заметить, что нуклеиновые кислоты были характеризованы на уровне аптамера, то есть на уровне D-нуклеиновой кислоты (D-РНК), с помощью биотинилированного человеческого D-SDF-1 (SEQ ID NO:4), или на уровне шпигельмера, то есть L-нуклеиновой кислоты (L-РНК) с помощью SDF-1 природной конфигурациии, L-SDF-1 (человеческий SDF-1α, SEQ-ID NO:1). Различные нуклеиновые кислоты делят между собой одно внутреннее ссылочное название, но существуют один SEQ.ID NO: для молекулы D-РНК (аптамер) и один SEQ ID NO: для молекулы L-РНК (шпигельмер), соответственно.
SDF-1 человека/обезьяны/кошки
SDF-1 мышей
Таблица 1(C)
192-A10-001-5’-PEG40
Настоящее изобретение далее проиллюстрировано фигурами, примерами и списком последовательностей, из которых могут быть восприняты дополнительные свойства, варианты осуществления и преимущества, где
На фиг.1 показан сравнительный анализ первичной структуры последовательностей соответствующих лигандов РНК, связывающихся с SDF-1 человека, указан мотив последовательности («тип A»), который в предпочтительном варианте осуществления во всей полноте является существенным для связывания с SDF-1 человека;
На фиг.2A показаны производные лиганда РНК 192-A10-001 (лиганд РНК SDF-1 человека мотива последовательности «типа A»);
На фиг.2B показаны производные лиганда РНК 192-A10-001 (лиганд РНК SDF-1 человека мотива последовательности «типа A»);
На фиг.3 показан сравнительный анализ первичной структуры последовательностей соответствующих лигандов РНК, связывающихся с SDF-1 человека, указан мотив последовательности («тип B»), который в предпочтительном варианте осуществления во всей полноте является существенным для связывания с SDF-1 человека;
На фиг.4A показаны производные лигандов РНК 193-C2-001 и 193-G2-001 (лиганды РНК SDF-1 человека мотива последовательности «типа B»);
На фиг.4B показаны производные лигандов РНК 193-C2-001 и 193-G2-001 (лиганды РНК SDF-1 человека мотива последовательности «типа B»);
На фиг.5 показан сравнительный анализ первичной структуры последовательностей соответствующих лигандов РНК, связывающихся с SDF-1 человека, указан мотив последовательности («тип C»), который в предпочтительном варианте осуществления во всей полноте является существенным для связывания с SDF-1 человека;
На фиг.6 показаны производные лиганда РНК 190-A3-001 (лиганд РНК SDF-1 человека мотива последовательности «типа C»);
На фиг.7A показаны производные лиганда РНК 190-D5-001 (лиганд РНК SDF-1 человека мотива последовательности «типа C»);
На фиг.7B показаны производные лиганда РНК 190-D5-001 (лиганд РНК SDF-1 человека мотива последовательности «типа C»);
На фиг.8 показаны производные лиганда РНК 197-B2 (лиганд РНК SDF-1 человека мотива последовательности «типа C»);
На фиг.9 показаны дополнительные лиганды РНК, связывающиеся с SDF-1 человека;
На фиг.10 показан индуцированный с помощью SDF-1 человека хемотаксис клеток линии Jurkat человеческого Т-клеточного лейкоза, при этом после 3 часов миграции клеток линии Jurkat человеческого Т-клеточного лейкоза по направлению к различным концентрациям SDF-1 человека была получена кривая дозовой зависимости для SDF-1 человека, представлено в виде зависимости сигнала флуоресценции от концентрации SDF-1 человека;
На фиг.11 показан результат анализа связывания связывающего SDF-1 человека аптамера 192-A10-001 с биотинилированным человеческим D-SDF-1 при 37°C, представлено в виде зависимости связывания аптамера от концентрации биотинилированного человеческого D-SDF-1;
На фиг.12 показана эффективность связывающего SDF-1 человека шпигельмера 192-A10-001 в хемотаксическом тесте; клетки получили возможность мигрировать в сторону 0,3 нМ SDF-1 человека, преинкубированного при 37°C с различными количествами шпигельмера 192-A10-001, представлено в виде зависимости доли контроля в процентах от концентрации шпигельмера 192-A10-001;
На фиг.13 показан результат анализа конкурентного связывания связывающих SDF-1 человека аптамеров 192-A10-001, 192-F10-001, 192-C9-001, 192-E10-001, 192-C10-001, 192-D11-001, 192-G11-001, 192-H11-001, 192-D10-001, 192-E9-001 и 192-H9-001 с биотинилированным человеческим D-SDF-1 при 37°C, представлено в виде связывания меченого аптамера 192-A10-001 (использован в качестве контроля сравнения, замещаемого немечеными аптамерами) при 1 нМ и 5 нМ немеченых аптамерах 192-A10-001, 192-F10-001, 192-C9-001, 192-E10-001, 192-C10-001, 192-D11-001, 192-G11-001, 192-H11-001, 192-D10-001, 192-E9-001 и 192-H9-001;
На фиг.14 показан результат анализа связывания связывающего SDF-1 человека аптамера 192-A10-008 с биотинилированным человеческим D-SDF-1 при 37°C, представлено в виде зависимости связывания аптамера от концентрации биотинилированного человеческого D-SDF-1;
На фиг.15 показана сенсограмма, полученная на приборе Biacore 2000, отображающая значение KD связывающего SDF-1 человека шпигельмера 192-A10-008, связавшегося с SDF-1 человека, который был иммобилизован на сенсорном чипе PioneerF1 с помощью процедуры сочетания аминов, представлено в виде зависимости ответа (отн. ед.) от времени, дополнительно приведены скорости ассоциации и диссоциации и значения KD шпигельмеров 192-A10-008 и 192-A10-001;
На фиг.16 показана эффективность связывающего SDF-1 шпигельмера 192-A10-008 в хемотаксическом тесте; клетки получили возможность мигрировать в сторону 0,3 нМ SDF-1 человека, преинкубированного при 37°C с различными количествами шпигельмера 192-A10-008, представлено в виде зависимости доли контроля в процентах от концентрации шпигельмера 192-A10-008;
На фиг.17 показана сенсограмма, полученная на приборе Biacore 2000, отображающая значение KD шпигельмера 193-G2-01, связавшегося с SDF-1 человека, который был иммобилизован на сенсорном чипе PioneerF1 с помощью процедуры сочетания аминов, представлено в виде зависимости ответа (отн. ед.) от времени, дополнительно приведены скорости ассоциации и диссоциации и значения KD шпигельмеров 193-G2-001 и 193-C2-001;
На фиг.18 показан результат анализа связывания человеческого анти-SDF-1 аптамера 193-G2-012 с биотинилированным человеческим D-SDF-1 при 37°C, представлено в виде зависимости связывания аптамера от концентрации биотинилированного человеческого D-SDF-1;
На фиг.19 показан результат анализа конкурентного связывания связывающих SDF-1 человека аптамеров 190-A3-001, 190-A3-003, 190-A3-004, 190-A3-007, 191-D5-001, 191-D5-002, 191-D5-003, 191-D5-004, 191-D5-005, 191-D5-006 и 191-D5-007 с биотинилированным человеческим D-SDF-1 при 37°C, представлено в виде связывания меченого аптамера 190-A3-001 или 191-D5-001 (использованы в качестве контроля сравнения, замещаемого немечеными аптамерами) при 500 нМ, 50 нМ и 10 нМ немеченых аптамерах 190-A3-001, 190-A3-003, 190-A3-004, 190-A3-007, 191-D5-001, 191-D5-002, 191-D5-003, 191-D5-004, 191-D5-005, 191-D5-006 и 191-D5-007;
На фиг.20 показан результат анализа связывания связывающих SDF-1 человека аптамеров 190-A3-004 и 191-D5-007 с биотинилированным человеческим D-SDF-1 при 37°C, представлено в виде зависимости связывания аптамера от концентрации биотинилированного человеческого D-SDF-1;
На фиг.21 показана сенсограмма, полученная на приборе Biacore 2000, отображающая значение KD шпигельмера 191-D5-007, связавшегося с SDF-1 человека, который был иммобилизован на сенсорном чипе PioneerF1 с помощью процедуры сочетания аминов, представлено в виде зависимости ответа (отн. ед.) от времени, дополнительно приведены скорости ассоциации и диссоциации и значения KD шпигельмеров 191-D5-001, 191-D5-007, 190-A3-003 и 197-B2;
На фиг.22 показана эффективность связывающего SDF-1 шпигельмера 190-A3-004 в хемотаксическом тесте; клетки получили возможность мигрировать в сторону 0,3 нМ SDF-1 человека, преинкубированного при 37°C с различными количествами шпигельмера 190-A3-004, представлено в виде зависимости доли контроля в процентах от концентрации шпигельмера 190-A3-004;
На фиг.23A показана эффективность связывающих SDF-1 шпигельмеров 193-G2-012-5’-PEG, 197-B2-006-5’-PEG, 191-D5-007-5’-PEG и 191-A10-008-5’-PEG в хемотаксическом тесте; клетки получили возможность мигрировать в сторону 0,3 нМ SDF-1 человека, преинкубированного при 37°C с различными количествами шпигельмеров 193-G2-012-5’-PEG, 197-B2-006-5’-PEG, 191-D5-007-5’-PEG и 191-A10-008-5’-PEG, представлено в виде зависимости доли контроля в процентах от концентрации шпигельмеров 193-G2-012-5’-PEG, 197-B2-006-5’-PEG, 191-D5-007-5’-PEG и 191-A10-008-5’-PEG;
На фиг.23B показана эффективность связывающих SDF-1 шпигельмеров 197-B2-006-5’PEG и 197-B2-006-31b-5’-PEG в хемотаксическом тесте; клетки получили возможность мигрировать в сторону 0,3 нМ SDF-1 человека, преинкубированного при 37°C с различными количествами шпигельмеров 197-B2-006-5’PEG и 197-B2-006-31b-5’-PEG, представлено в виде зависимости доли контроля в процентах от концентрации шпигельмеров 197-B2-006-5’PEG и 197-B2-006-31b-5’-PEG;
На фиг.24A показана сенсограмма, полученная на приборе Biacore 2000, отображающая значения KD шпигельмеров 193-G2-012-5’-PEG, 191-A10-008-5’-PEG и 191-A10-001-5’-PEG, связавшихся с SDF-1 человека, который был иммобилизован на сенсорном чипе PioneerF1 с помощью процедуры сочетания аминов, представлено в виде зависимости ответа (отн. ед.) от времени;
На фиг.24B показана сенсограмма, полученная на приборе Biacore 2000, отображающая значения KD шпигельмеров 197-B2-006-5’PEG, 197-B2-006-31b-5’-PEG и 191-D5-007-5’-PEG, связавшихся с SDF-1 человека, который был иммобилизован на сенсорном чипе PioneerF1 с помощью процедуры сочетания аминов, представлено в виде зависимости ответа (отн. ед.) от времени;
На фиг.25A показана эффективность связывающих SDF-1 шпигельмеров 192-A10-001, 192-A10-001-5’-HES130 и 192-A10-001-5’-HES100 в хемотаксическом тесте; клетки получили возможность мигрировать в сторону 0,3 нМ SDF-1 человека, преинкубированного при 37°C с различными количествами шпигельмеров 192-A10-001, 192-A10-001-5’-HES130 и 192-A10-001-5’-HES100, представлено в виде зависимости доли контроля в процентах от концентрации шпигельмеров 192-A10-001, 192-A10-001-5’-HES130 и 192-A10-001-5’-HES100;
На фиг.25B показана эффективность связывающих SDF-1 шпигельмеров 192-A10-001, 192-A10-001-5’-PEG30 и 192-A10-001-5’-PEG40 в хемотаксическом тесте; клетки получили возможность мигрировать в сторону 0,3 нМ SDF-1 человека, преинкубированного при 37°C с различными количествами шпигельмеров 192-A10-001, 192-A10-001-5’-PEG30 и 192-A10-001-5’-PEG40, представлено в виде зависимости доли контроля в процентах от концентрации шпигельмеров 192-A10-001, 192-A10-001-5’-PEG30 и 192-A10-001-5’-PEG40;
На фиг.26 показана неэффективность контрольного шпигельмера в хемотаксическом тесте; клетки получили возможность мигрировать в сторону 0,3 нМ SDF-1 человека или мыши, преинкубированного при 37°C с различными количествами контрольного шпигельмера, представлено в виде зависимости доли контроля в процентах от концентрации контрольного шпигельмера;
На фиг.27 показан индуцированный с помощью SDF-1 мыши хемотаксис клеток линии Jurkat человеческого Т-клеточного лейкоза, при этом после 3 часов миграции клеток линии Jurkat человеческого Т-клеточного лейкоза по направлению к различным концентрациям SDF-1 была получена кривая дозовой зависимости для SDF-1, представлено в виде сигнала флуоресценции;
На фиг.28 показана эффективность связывающих SDF-1 шпигельмеров 192-A10-001 и 191-D5-007-5’PEG в хемотаксическом тесте; клетки получили возможность мигрировать в сторону 0,3 нМ SDF-1 мыши, преинкубированного при 37°C с различными количествами шпигельмеров 192-A10-001 и 191-D5-007-5’PEG, представлено в виде зависимости доли контроля в процентах от концентрации шпигельмеров 192-A10-001 и 191-D5-007-5’PEG;
На фиг.29 показана эффективность связывающего SDF-1 шпигельмера 192-A10-001 в тесте на связывание CXCR4-рецептора с помощью [125J]-SDF-1α человека, преинкубированного при 37°C с различными количествами шпигельмера 192-A10-001, специфично связанный [125J]-SDF-1α наносили на график в зависимости от концентрации шпигельмера 192-A10-001; и
На фиг.30 показано ингибирование MAP-киназного стимулирования экспрессирующих CXCR4 клеток посредством 1 нМ SDF-1α человека связывающим SDF-1 человека шпигельмером 192-A10-001;
На фиг.31 показано ингибирование индуцированного с помощью SDF-1 прорастания сосудов связывающим SDF-1 человека шпигельмером 193-G2-012-5’-PEG и PEG-модифицированным контрольным шпигельмером в тесте аортального кольца на прорастание сосудов, где кольца крысиной аорты погружали в коллагеновый матрикс и инкубировали в течение 6 дней с SDF-1 в присутствии или отсутствие шпигельмеров (a: контроль; b: 10 нМ SDF-1; c: 10 нМ SDF-1 + 1 мкМ связывающий SDF-1 человека шпигельмер 193-G2-012-5’-PEG; d: 10 нМ SDF-1 + 1 мкМ PEG-модифицированный контрольный шпигельмер);
На фиг.32 показано ингибирование индуцированного с помощью SDF-1 прорастания сосудов связывающим SDF-1 человека шпигельмером 193-G2-012-5’-PEG и PEG-модифицированным контрольным шпигельмером в тесте аортального кольца на прорастание сосудов, где показатели прорастания сосудов показаны в виде среднего значения +/- SD (стандартное отклонение) для 5 колец на одно патологическое состояние (*: значение для SDF-1 существенно отличается от контроля (критерий Манна-Уитни; p=0,009); **: значение для SDF-1 + связывающего SDF-1 человека шпигельмера 193-G2-012-5’-PEG существенно отличается от такового для SDF-1 (критерий Манна-Уитни; p=0,028);
На фиг.33 показано схематическое изображение лечения животных и способы, примененные для получения данных о высвобождении стволовых клеток, согласно примеру 10;
На фиг.34 показано арифметическое число CD117+ и Ly-6 A/E+ клеток (гемопоэтические стволовые клетки/ гемопоэтические клетки-предшественники) в расчете на микролитр плазмы, высвобождаемые за время от 1 до 48 часов после внутривенной инъекции NOX-A12-JE40, NOX-A12-NO30, revNOX-A12-JE40 (контрольный шпигельмер), AMD3100, G-CSF (нейпоген) или лекарственной основы (5% глюкоза); на графике показаны средние значения и стандартные отклонения;
На фиг.35 показаны колониеобразующие единицы в расчете на микролитр крови для мышей C57BL/6 по истечении 6 час для инъекций NOX-A12-производных и AMD3100, или 48 час для инъекций G-CSF (нейпоген) и лекарственной основы, как указано в подписи к фигуре; на диаграмме показаны средние значения и стандартные отклонения для 5 мышей (тройной повтор для каждой группы);
На фиг.36 показаны результаты исследований на мышах индуцированной лазером хороидальной неоваскуляризации, где NOX-A12-JE40 уменьшал площадь новообразованных сосудов в обработанном NOX-A12-JE40 глазу, при прямом сравнении с площадью в обработанном лекарственной основой глазу (раствор Рингера) у того же самого животного (левая диаграмма); средние значения неоваскулярных площадей отдельных повреждений в обработанных NOX-A12-JE40 и лекарственной основой глазах мышей после поражения лазером показаны на диаграмме справа;
На фиг.37 показаны результаты повторных обработок здоровых и больных диабетом мышей, с удалением одной почки или без этого, лекарственной основой, NOX-A12-JE40, revNOX-A12-JE40 (контрольным шпигельмером), где NOX-A12-JE40 повышает число баллов по шкале оценки гломерулосклероза;
На фиг.38 показаны результаты моделирования на животных подтекания сосудов сетчатки глаза после индукции посредством введения в стекловидное тело VEGF и обработки животных различными концентрациями связывающего SDF-1 шпигельмера NOX-A12-JE40, лекарственной основы или кенакорт-ретарта; у этих моделей подтекание сосудистой системы сетчатки измеряли посредством фотометрии флуоресцеина через 48 час после инъекции в стекловидное тело VEGF;
На фиг.39 показано число баллов по шкале оценки ретинопатии (глаз, обработанный лекарственной основой [ось x] против глаза, обработанного шпигельмером NOX-A12-JE40 [ось y]) отдельных мышей при моделировании на мышах индуцированной кислородом ретинопатии, которая является моделью, имитирующей индуцированную гипоксией неоваскуляризацию сетчатки глаза, наблюдаемую при диабетической ретинопатии или AMD;
На фиг.40 показаны значения p для статистического расхождения между обработанными шпигельмером NOX-A12-JE40 и обработанными лекарственной основой глазами для отдельно измеренных параметров ретинопатии и числа баллов по шкале оценки ретинопатии, измеренных при моделировании на мышах индуцированной кислородом ретинопатии, которая является моделью, имитирующей индуцированную гипоксией неоваскуляризацию сетчатки глаза, наблюдаемую при диабетической ретинопатии или AMD; значения p определяли с помощью знаковых ранговых критериев Уилкоксона;
На фиг.41 показан уровень лейкоцитов в крови после внутривенного введения 13,4 мг (рассчитано относительно олигонуклеотидной составляющей) связывающей SDF-1 нуклеиновой кислоты NOX-A12-JE40, где на графике регистрировался только один момент времени после введения лекарственной основы (5% глюкоза), поскольку это значение было решено считать постоянным; однако NOX-A12-JE40 мобилизирует значительное количество лейкоцитов обратимым образом;
На фиг.42 показано краткое описание показаний для аллогенных трансплантаций гемопоэтических стволовых клеток (сокр. HSCT) и число пациентов в Европе за период 1990–2000 гг. (Gratwohl, Baldomero et al. 2002);
На фиг.43 показано краткое описание показаний для аутогенных трансплантаций гемопоэтических стволовых клеток (сокр. HSCT) и число пациентов в Европе за период 1990–2000 гг. (Gratwohl, Baldomero et al. 2002).
Пример 1
Нуклеиновые кислоты, связывающие SDF-1 человека
Используя биотинилированный человеческий D-SDF-1 в качестве мишени, могут быть созданы некоторые связывающие SDF-1 человека нуклеиновые кислоты, нуклеотидные последовательности которых представлены на фиг.1-9. Нуклеиновые кислоты были характеризованы на уровне аптамера, то есть на уровне D-нуклеиновой кислоты с помощью биотинилированного человеческого D-SDF-1, или на уровне шпигельмера, то есть L-нуклеиновой кислоты с помощью SDF-1 природной конфигурациии (L-SDF-1).
Аптамеры анализировали с помощью биотинилированного человеческого D-SDF-1, посредством конкурентного или прямого замещающих тестов на связывание с биотинилированным человеческим D-SDF-1 (пример 4). Шпигельмеры анализировали с помощью SDF-1 природной конфигурациии (L-SDF-1) с помощью измерений методом поверхностного плазмонного резонанса, применяя прибор Biacore 2000 (пример 6) и in vitro хемотаксического теста на культуре клеток (пример 5).
Произведенные таким образом молекулы нуклеиновых кислот демонстрируют различные мотивы последовательностей, три главных типа определены на фиг.1, 2A и 2B (тип A), фиг.3, 4A и 4B (тип B), фиг.5, 6, 7A, 7B и 8 (тип C). Для определения мотивов нуклеотидных последовательностей используются сокращения ИЮПАК для неопределенных нуклеотидов:
Если не указано противоположное, любая последовательность нуклеиновых кислот или последовательность участков нуклеотидов и боксы, соответственно, указаны в 5’ → 3’ направлении.
1.1 Связывающие SDF-1 нуклеиновые кислоты типа A
Как представлено на фиг.1, все последовательности связывающих SDF-1 нуклеиновых кислот типа A содержат одну коровую нуклеотидную последовательность, фланкированную 5’- и 3’-концевыми участками нуклеотидов, способными гибридизоваться друг с другом. Однако такая гибридизация не обязательно приводится в представлении молекулы.
Нуклеиновые кислоты были характеризованы на уровне аптамеров посредством прямого и конкурентного замещающих тестов на связывание с биотинилированным человеческим D-SDF-1, чтобы отнести их к какой-либо категории, принимая во внимание их динамику связывания (пример 4). Отобранные последовательности синтезировали в виде шпигельмеров (пример 3) и анализировали с помощью SDF-1 природной конфигурациии (L-SDF) с помощью in vitro хемотаксического теста на культуре клеток (пример 5) и измерений методом поверхностного плазмонного резонанса, применяя прибор Biacore 2000 (пример 6).
Последовательности установленных боксов или участков нуклеотидов могут различаться между связывающими SDF-1 нуклеиновыми кислотами типа A, что влияет на аффинность связывания SDF-1. На основании анализа связывания различных связывающих SDF-1 нуклеиновых кислот, охарактеризованных как связывающие SDF-1 нуклеиновые кислоты типа A, данная коровая нуклеотидная последовательность и ее нуклеотидные последовательности, как описано ниже, по отдельности и, более предпочтительно, - все вместе, являются существенными для связывания SDF-1:
Коровая нуклеотидная последовательность всех идентифицированных последовательностей связывающих SDF-1 нуклеиновых кислот типа A содержит последовательность AAAGYRACAHGUMAAXAUGAAAGGUARC (тип A формула-1), где XA либо отсутствует, либо представляет собой «A». Если «A» отсутствует, последовательность коровой нуклеотидной последовательности может быть охарактеризована как тип A формула-2 (AAAGYRACAHGUMAA-UGAAAGGUARC). Связывающая SDF-1 нуклеиновая кислота 191-A6 типа A (коровая нуклеотидная последовательность AAAGUAACACGUAAAAUGAAAGGUAAC), несущая дополнительный нуклеотид «A» в составе коровой нуклеотидной последовательности и все еще способная связывать SDF-1, позволяет вывести другую коровую нуклеотидную последовательность (AAAGYRACAHGUMAAAUGAAAGGUARC, тип A формула-3). Как пример для всех остальных нуклеиновых кислот из связывающих SDF-1 нуклеиновых кислот типа A, связывающая SDF-1 нуклеиновая кислота 192-A10-001 типа A была охарактеризована по свой аффинности связывания SDF-1 человека. Равновесную константу связывания KD определяли с помощью замещающего теста на связывание (KD=1,5 нМ, фиг.11) и измерений методом поверхностного плазмонного резонанса (KD=1,0 нМ, фиг.15). Значение IC50 (полумаксимальная концентрация ингибирования), равное 0,12 нМ для 192-A10-001, было получено с помощью in vitro хемотаксического теста в культуре клеток (фиг.12). Соответственно, все связывающие SDF-1 нуклеиновые кислоты типа A, представленные на фиг.1, анализировали в конкурентном замещающем тесте на связывание в сравнении с 192-A10-001 (фиг.13; не все протестированные связывающие SDF-1 нуклеиновые кислоты типа A показаны на фиг.13). Связывающие SDF-1 нуклеиновые кислоты 192-B11 и 192-C10 типа A показали те же аффинности связывания, что и 192-A10-001 в этих конкурентных лабораторных исследованиях. Более слабая аффинность связывания была определена для связывающих SDF-1 нуклеиновых кислот 192-G10, 192-F10, 192-C9, 192-E10, 192-D11, 192-G11, 192-H11 и 191-A6 типа A. Связывающие SDF-1 нуклеиновые кислоты 192-D10, 192-E9 и 192-H9 типа A имели значительно более слабую аффинность связывания, чем 192-A10-001 (фиг.13).
Как указывалось выше, связывающие SDF-1 нуклеиновые кислоты 192-B11 и 192-C10 типа A показали ту же аффинность связывания с SDF-1, что и 192-A10-001. Однако они показали небольшие различия нуклеотидных последовательностей в коровой нуклеотидной последовательности. Исходя из вышеизложенного, консенсусная последовательность трех молекул, связывающихся с SDF-1 почти с той же высокой аффинностью, может быть охарактеризована нуклеотидной последовательностью AAAGYAACAHGUCAAUGAAAGGUARC (тип A формула-4), в силу чего нуклеотидная последовательность коровой нуклеотидной последовательности 192-A10-001 (нуклеотидная последовательность: AAAGCAACAUGUCAAUGAAAGGUAGC) представляет собой нуклеотидную последовательность с лучшей аффинностью связывания среди связывающих SDF-1 нуклеиновых кислот типа A.
Пять или шесть из шести нуклеотидов 5’-концевого участка нуклеотидов связывающих SDF-1 нуклеиновых кислот типа A могут гибридизоваться с соответствующими пятью или шестью нуклеотидами из шести нуклеотидов 3’-концевого участка нуклеотидов связывающих SDF-1 нуклеиновых кислот типа A, образуя концевую спираль. Хотя эти нуклеотиды вариабельны по некоторым положениям, различные нуклеотиды допускают гибридизацию пяти или шести из шести нуклеотидов 5’- и 3’-концевых участков нуклеотидов в каждом. 5’-концевый и 3’-концевый участки нуклеотидов связывающих SDF-1 нуклеиновых кислот типа A, как показано на фиг.1, могут быть охарактеризованы общей формулой для 5’-концевого участка нуклеотидов («RSHRYR», тип A формула-5-5’) и для 3’-концевого участка нуклеотидов («YRYDSY», тип A формула-5-3’). Усеченные производные связывающей SDF-1 нуклеиновой кислоты 192-A10-001 типа A анализировали в конкурентном замещающем тесте на связывание в сравнении с исходной молекулой 192-A10-001 и 192-A10-008 (фиг.2A и 2B). Эти эксперименты показали, что усечение шести концевых нуклеотидов (5’конец: GCUGUG; 3’конец: CGCAGC) у 192-A10-001 и до пяти нуклеотидов (5’конец: CUGUG; 3’конец: CGCAG) у производного 192-A10-002 может быть проведено без снижения аффинности связывания. Однако усечение до четырех концевых нуклеотидов (5’конец: UGUG; 3’конец: CGCA; 192-A10-003) или менее (192-A10-004/ -005/ -006/ -007) приводит к пониженной аффинности связывания с SDF-1 (фиг.2A). Определенные 5’-концевые и 3’-концевые участки нуклеотидов длиною в пять нуклеотидов и четыре нуклеотида производных связывающей SDF-1 нуклеиновой кислоты 192-A10-001 типа A, как показано на фиг.2A и 2B, могут быть описаны общей формулой для 5’-концевого участка нуклеотидов («X2BBBS», тип A формула-6-5’) и для 3’-концевого участка нуклеотидов («SBBVX3»; тип A формула-6-3’), где X2 либо отсутствует, либо представляет собой «S», а X3 либо отсутствует, либо представляет собой «S».
Нуклеотидная последовательность 5’- и 3’-концевых участков нуклеотидов влияет на аффинность связывания связывающих SDF-1 нуклеиновых кислот типа A. Это показано не только с помощью нуклеиновых кислот 192-F10 и 192-E10, но также и с помощью производных 192-A10-001 (фиг.2B). Коровые нуклеотидные последовательности 192-F10 и 192-E10 идентичны 192-B11 и 192-C10, но включают небольшие отличия по 3’-концу 5’-концевого участка нуклеотидов и по 5’-концу 3’-концевого участка нуклеотидов, приводя к пониженной аффинности связывания.
Замещения 5’- и 3’-концевых нуклеотидов «CUGUG» и «CGCAG» связывающей SDF-1 нуклеиновой кислоты 192-A10-002 типа A на «GCGCG» и «CGCGC» (192-A10-015) приводит к пониженной аффинности связывания, тогда как замещения на «GCGUG» и «CGCGC» (192-A10-008) приводят к той же самой аффинности связывания, что показана для 192-A10-002 (фиг.2B, фиг.15, фиг.12, фиг.16). Кроме того, девять производных связывающей SDF-1 нуклеиновой кислоты 192-A10-001 типа A (192-A10-014/ -015/ -016/ -017/ -018/ -019/ -020/ -021/ -022/ -023), несущие четыре 5’- и 3’-концевых нуклеотида, соответственно, были испытаны как аптамеры на свои аффинности связывания в сравнении с 192-A10-001 или ее производной 192-A10-008 (обе имеют идентичные аффинности связывания с SDF-1). Все клоны показали более слабые, значительно более слабые или намного более слабые аффинности связывания с SDF-1, такие как 192-A10-001 (шесть нуклеотидов, образующих концевую спираль), или как 192-A10-008 с пятью концевыми нуклеотидами, соответственно (фиг.2B). Соответственно, последовательность и число нуклеотидов 5’- и 3’-концевых участках нуклеотидов являются существенными для эффективного связывания с SDF-1. Как показано для связывающих SDF-1 нуклеиновых кислот 192-A10-002 и 192-A10-08 типа A, предпочтительными комбинациями 5’- и 3’-концевых участков нуклеотидов являются «CUGUG» и «CGCAG» (5’- и 3’-концевые участки нуклеотидов связывающей SDF-1 нуклеиновой кислоты 192-A10-002 типа A), а также «GCGUG» и «CGCGC» (5’- и 3’-концевые участки нуклеотидов связывающей SDF-1 нуклеиновой кислоты 192-A10-008 типа A).
Однако, комбинируя 5’- и 3’-концевые участки нуклеотидов всех испытанных связывающих SDF-1 нуклеиновых кислот типа A, общая формула для 5’-концевого участка нуклеотидов связывающих SDF-1 нуклеиновых кислот типа A представляет собой «X1X2NNBV» (тип A формула-7-5’), а общая формула для 3’-концевого участка нуклеотидов связывающих SDF-1 нуклеиновых кислот типа A представляет собой «BNBNX3X4» (тип A формула-7-3’), где
X1 представляет собой «R» или отсутствует, X2 представляет собой «S», X3 представляет собой «S» и X4 представляет собой «Y» или отсутствует;
либо
X1 отсутствует, X2 представляет собой «S» или отсутствует, X3 представляет собой «S» или отсутствует и X4 отсутствует.
1.2 Cвязывающие SDF-1 нуклеиновые кислоты типа B
Как представлено на фиг.3, все последовательности связывающих SDF-1 нуклеиновых кислот типа B содержат одну коровую нуклеотидную последовательность, фланкированную 5’- и 3’-концевыми участками нуклеотидов, способными гибридизоваться друг с другом. Однако такая гибридизация не обязательно приводится в представлении молекулы.
Нуклеиновые кислоты были характеризованы на уровне аптамеров посредством прямого и конкурентного замещающих тестов на связывание с биотинилированным человеческим D-SDF-1, чтобы отнести их к какой-либо категории, принимая во внимание их динамику связывания (пример 4). Отобранные последовательности синтезировали в виде шпигельмеров (пример 3) и анализировали с помощью SDF-1 природной конфигурациии (L-SDF) с помощью in vitro хемотаксического теста на культуре клеток (пример 5) и измерений методом поверхностного плазмонного резонанса, применяя прибор Biacore 2000 (пример 6).
Последовательности установленных боксов или участков нуклеотидов могут различаться между связывающими SDF-1 нуклеиновыми кислотами типа B, что влияет на аффинность связывания SDF-1. На основании анализа связывания различных связывающих SDF-1 нуклеиновых кислот, охарактеризованных как связывающие SDF-1 нуклеиновые кислоты типа B, данная коровая нуклеотидная последовательность и ее нуклеотидные последовательности, как описано ниже, по отдельности и, более предпочтительно, - все вместе, являются существенными для связывания SDF-1:
Коровая нуклеотидная последовательность всех идентифицированных последовательностей связывающих SDF-1 нуклеиновых кислот типа B содержит последовательность GUGUGAUCUAGAUGUADWGGCUGWUCCUAGUYAGG (тип B формула-1). Связывающие SDF-1 нуклеиновые кислоты 193-G2-001, 193-C2-001 и 193-F2-001 типа B, отличающиеся лишь по одному положению от коровой нуклеотидной последовательности, анализировали с помощью замещающего теста на связывание в сравнении со связывающей SDF-1 нуклеиновой кислотой 192-A10-001 типа A (KD=1,5 нМ определяли в замещающем тесте на связывание [фиг.11], KD=1,0 нМ определяли посредством измерений методом поверхностного плазмонного резонанса [фиг.15], IC50=0,12 нМ; [фиг.12]). Каждая из трех протестированных связывающих SDF-1 нуклеиновых кислот типа B показала превосходящее связывание с SDF-1 человека по сравнению со связывающей SDF-1 нуклеиновой кислотой 192-A10-001 типа A, в соответствии с чем аффинность связывания у 193-G2-001 фактически та же, что и у 193-C2-001, и у 193-F2-001 (фиг.3). Данные свидетельствуют, что разница в нуклеотидной последовательности коровой нуклеотидной последовательности связывающих SDF-1 нуклеиновых кислот 193-G2-001, 193-C2-001 и 193-F2-001 типа B не влияла на аффинность связывания с SDF-1. Как пример, связывающая SDF-1 нуклеиновая кислота 193-G2-001 типа B была охарактеризована по свой аффинности связывания SDF-1 человека. Равновесную константу связывания KD определяли с помощью замещающего теста на связывание (KD=0,3 нМ) и измерений методом поверхностного плазмонного резонанса (KD=0,5 нМ, фиг.17). Значение IC50 (полумаксимальная концентрация ингибирования), равное 0,08 нМ для 193-G2-001, было получено с помощью in vitro хемотаксического теста в культуре клеток. Напротив, связывающие SDF-1 нуклеиновые кислоты 193-B3-002, 193-H3-002, 193-E3-002 и 193-D1-002 типа B, отличающиеся от коровой нуклеотидной последовательности, имели худшие связывающие свойства (фиг.3). В результате, связывающие SDF-1 нуклеиновые кислоты типа B с улучшенной аффинностью связывания с SDF-1 разделяют между собой коровую нуклеотидную последовательность с последовательностью GUGUGAUCUAGAUGUADUGGCUGAUCCUAGUCAGG (тип B формула-2).
Четыре, пять или шесть из шести нуклеотидов 5’-концевого участка нуклеотидов связывающих SDF-1 нуклеиновых кислот типа B могут гибридизоваться с соответствующими четырьмя, пятью или шестью нуклеотидами из шести нуклеотидов 3’-концевого участка нуклеотидов связывающих SDF-1 нуклеиновых кислот типа B, образуя концевую спираль. Хотя эти нуклеотиды вариабельны по некоторым положениям, различные нуклеотиды допускают гибридизацию четырех, пяти или шести из шести нуклеотидов 5’- и 3’-концевых участков нуклеотидов в каждом. 5’-концевый и 3’-концевый участки нуклеотидов связывающих SDF-1 нуклеиновых кислот типа B, как показано на фиг.3, могут быть охарактеризованы общей формулой для 5’-концевого участка нуклеотидов (тип B формула-3-5’; «X1GCRWG», где X1 представляет собой «A» или отсутствует) и для 3’-концевого участка нуклеотидов (тип B формула-3-3’; «KRYSCX4», где X4 представляет собой «U» или отсутствует). Связывающие SDF-1 нуклеиновые кислоты 193-G1-002, 193-D2-002, 193-A1-002 и 193-D3-002 типа B имеют более слабые аффинности связывания с SDF-1, хотя они разделяют идентичную коровую нуклеотидную последовательность (тип B формула-2) с 193-C2-001, 193-G2-001 и 193-F2-001 (фиг.3). Неудовлетворительные связывающие свойства связывающих SDF-1 нуклеиновых кислот 193-G1-002, 193-D2-002, 193-A1-002 и 193-D3-002 типа B могут объясняться числом нуклеотидов и последовательностью 5’- и 3’-концевых участков нуклеотидов.
Усеченные производные связывающих SDF-1 нуклеиновых кислот 193-G2-001 и 193-C2-001 типа B анализировали в конкурентном замещающем тесте на связывание в сравнении с 193-G2-001 и 193-G2-012, соответственно (фиг.4A и 4B). Эти эксперименты показали, что усечение шести концевых нуклеотидов (5’конец: AGCGUG; 3’конец: UACGCU) у связывающих SDF-1 нуклеиновых кислот 193-G2-001 и 193-C2-001 типа B до пяти нуклеотидов (5’конец: GCGUG; 3’конец: UACGC) приводит к молекулам со сходной аффинностью связывания (193-C2-002 и 193-G2-012). Равновесную константу диссоциации KD определяли с помощью замещающего теста на связывание (KD=0,3 нМ, фиг.18). Усечение до четырех (5’конец: CGUG; 3’конец: UACG; 193-C2-003) или менеее нуклеотидов (193-C2-004, 193-C2-005, 193-C2-006, 193-C2-007) приводит к пониженной аффинности связывания с SDF-1, что было определено с помощью конкурентного замещающего теста на связывание (фиг.4A). Нуклеотидная последовательность пяти концевых нуклеотидов на 5’- и 3’-конце, соответственно, влияла на аффинность связывания связывающих SDF-1 нуклеиновых кислот типа B. Замещения 5’- и 3’-концевых нуклеотидов «GCGUG» и «UACGC» (193-C2-002, 193-G2-12) на «GCGCG» и «CGCGC» (193-G2-013) привели к пониженной аффинности связывания. Кроме того, были протестированы четыре различные производные связывающей SDF-1 нуклеиновой кислоты 193-G2-001 типа B с концевой спиралью длиной в четыре спаренные нуклеотидные основания (193-G2-014/ -015/ -016/ -017). Все они показали пониженные аффинности связывания с SDF-1 (фиг.4B). По этой причине последовательность и длина 5’- и 3’-концевых нуклеотидов имеют существенное значение для эффективного связывания с SDF-1. Как показано на фиг.4A и 4B, 5’-концевые и 3’-концевые участки нуклеотидов производных связывающих SDF-1 нуклеиновых кислот 193-C2-003 и 193-G2-012 типа B длиною в пять и четыре нуклеотида могут быть описаны общей формулой для 5’-концевого участка нуклеотидов («X2SSBS», тип B формула-4-5’), где X2 либо отсутствует, либо представляет собой «G», и для 3’-концевого участка нуклеотидов («BVSSX3», тип B формула-4-3’), где также X3 либо отсутствует, либо представляет собой «C». Как показано для связывающих SDF-1 нуклеиновых кислот 193-G2-001 и 193-C2-01 типа B и их производных 193-G2-012 и 193-C2-002, предпочтительной комбинацией 5’- и 3’-концевых участков нуклеотидов является «X1GCGUG» (5’-концевый участок нуклеотидов; тип B формула 5-5’) и «UACGCX4» (3’-концевый участок нуклеотидов; тип B формула 5-3’), где X1 либо представляет собой «A», либо отсутствует, а X4 представляет собой «U» или отсутствует.
Однако, комбинируя 5’- и 3’-концевые участки нуклеотидов всех испытанных связывающих SDF-1 нуклеиновых кислот типа B, общая формула для 5’-концевого участка нуклеотидов связывающих SDF-1 нуклеиновых кислот типа B представляет собой «X1X2SVNS» (тип B формула-6-5’), а общая формула для 3’-концевого участка нуклеотидов связывающих SDF-1 нуклеиновых кислот типа B представляет собой «BVBSX3X4» (тип B формула-6-3’), где
X1 представляет собой «A» или отсутствует, X2 представляет собой «G», X3 представляет собой «C» и X4 представляет собой «U» или отсутствует;
либо X1 отсутствует, X2 представляет собой «G» или отсутствует, X3 представляет собой «C» или отсутствует и X4 отсутствует;
1.3 Связывающие SDF-1 нуклеиновые кислоты типа C
Как представлено на фиг.5, все последовательности связывающих SDF-1 нуклеиновых кислот типа C содержат одну коровую нуклеотидную последовательность, фланкированную 5’- и 3’-концевыми участками нуклеотидов, способными гибридизоваться друг с другом. Однако такая гибридизация не обязательно приводится в представлении молекулы.
Нуклеиновые кислоты были характеризованы на уровне аптамеров посредством прямого и конкурентного замещающих тестов на связывание с биотинилированным человеческим D-SDF-1, чтобы отнести их к какой-либо категории, принимая во внимание их динамику связывания (пример 4). Отобранные последовательности синтезировали в виде шпигельмеров (пример 3) и анализировали с помощью SDF-1 природной конфигурациии (L-SDF) посредством in vitro хемотаксического теста на культуре клеток (пример 5) и измерений методом поверхностного плазмонного резонанса, применяя прибор Biacore 2000 (пример 6).
Последовательности установленных боксов или участков нуклеотидов могут различаться между связывающими SDF-1 нуклеиновыми кислотами типа C, что влияет на аффинность связывания SDF-1. На основании анализа связывания различных связывающих SDF-1 нуклеиновых кислот, охарактеризованных как связывающие SDF-1 нуклеиновые кислоты типа C, данная коровая нуклеотидная последовательность и ее нуклеотидные последовательности, как описано ниже, по отдельности и, более предпочтительно, - все вместе, являются существенными для связывания SDF-1:
Коровая нуклеотидная последовательность всех идентифицированных последовательностей связывающих SDF-1 нуклеиновых кислот типа C содержит последовательность GGUYAGGGCUHRXAAGUCGG (тип C формула-1), где XA либо отсутствует, либо представляет собой «A». За исключением связывающей SDF-1 нуклеиновой кислоты 197-D1 типа C коровая нуклеотидная последовательность всех идентифицированных последовательностей связывающих SDF-1 нуклеиновых кислот типа C содержит нуклеотидную последовательность GGUYAGGGCUHRAAGUCGG (тип C формула-2). Связывающая SDF-1 нуклеиновая кислота 197-D1 типа C (коровая нуклеотидная последовательность: GGUUAGGGCUAA-AGUCGG), утратившая один нуклеотид «A» в коровой нуклеотидной последовательности и все еще связывающая SDF-1, позволяет вывести другую коровую нуклеотидную последовательность (GGUYAGGGCUHR-AGUCGG, тип C формула-3). Исходно, все связывающие SDF-1 нуклеиновые кислоты типа C, как представлено на фиг.5, анализировали конкурентным замещающим тестом на связывание в сравнении со связывающей SDF-1 нуклеиновой кислотой 192-A10-001 типа A (KD=1,5 нМ определяли замещающим тестом и путем измерений методом поверхностного плазмонного резонанса; IC50=0,12 нМ). Связывающие SDF-1 нуклеиновые кислоты 191-D5-001, 197-B2, 190-A3-001, 197-H1, 197-H3 и 197-E3 типа C показали более слабые аффинности связывания, чем 192-A10-001, в конкурентных лабораторных исследованиях. Значительно более слабую аффинность связывания определили для 191-A5, 197-B1, 197-D1, 197-H2 и 197-D2 (фиг.5). Молекулы и их производные были дополнительно охарактеризованы повторными конкурентными замещающими тестами на связывание, измерениями методом плазмонного резонанса и in vitro хемотаксическим тестом. Связывающую SDF-1 нуклеиновую кислоту 191-D5-001 типа C охарактеризовали по ее аффинности связывания с SDF-1 человека, где равновесную константу связывания KD определяли методом поверхностного плазмонного резонанса (KD=0,8 нМ, фиг.21). Значение IC50 (полумаксимальная концентрация ингибирования), равное 0,2 нМ для 191-D5-001, было получено с помощью in vitro хемотаксического теста в культуре клеток. Аффинность связывания связывающей SDF-1 нуклеиновой кислоты 197-B2 типа C для SDF-1 человека определяли методом поверхностного плазмонного резонанса (KD=0,9 нМ), значение ее IC50 (полумаксимальная концентрация ингибирования), равное 0,2 нМ, было измерено с помощью in vitro хемотаксического теста в культуре клеток. Эти данные показывают, что связывающие SDF-1 нуклеиновые кислоты 191-D5-001 и 197-B2 типа C имеют схожую аффинность связывания с SDF-1 (фиг.5 и 8).
Связывающая SDF-1 нуклеиновая кислота 190-A3-001 типа C (48 нуклеотидов) включает 5’-концевый участок нуклеотидов длиной 17 нуклеотидов и 3’-концевый участок нуклеотидов длиной 12 нуклеотидов, где с одной стороны, четыре нуклеотида на 5’-конце 5’-концевого участка нуклеотидов и четыре нуклеотида на 3’-конце 3’-концевого участка нуклеотидов могут гибридизоваться друг с другом, образуя концевую спираль. С другой стороны, нуклеотиды «UGAGA» в 5’-концевом участке нуклеотидов могут гибридизоваться с нуклеотидами «UCUCA» в 3’-концевом участке нуклеотидов, образуя концевую спираль. Усечение до восьми нуклеотидов 5’-концевого участка нуклеотидов («GAGAUAGG») и до девяти нуклеотидов 3’-концевого участка нуклеотидов («CUGAUUCUC») молекулы 190-A3-001 (где шесть из восьми/девяти нуклеотидов 5’- и 3’-концевых участков нуклеотидов могут гибридизоваться друг с другом) не влияют на аффинность связывания с SDF-1 (190-A3-004; фиг.6 и фиг.19). Равновесную константу связывания KD 190-A3-004 определяли с помощью замещающего теста на связывание (KD=4,6 нМ, фиг.20) и измерений методом поверхностного плазмонного резонанса (KD=4,7 нМ). Значение IC50 (полумаксимальная концентрация ингибирования), равное 0,1 нМ для 190-A3-004, было получено с помощью in vitro хемотаксического теста в культуре клеток (фиг.22). Однако отсечение двух нуклеотидов у 5’-концевого участка нуклеотидов приводит к очень сильному снижению аффинности связывания (190-A3-007; фиг.6 и фиг.19).
Связывающие SDF-1 нуклеиновые кислоты 191-D5-001, 197-B2 и 197-H1 типа C (коровая нуклеотидная последовательность: GGUUAGGGCUAGAAGUCGG), 197-H3/191-A5 типа C (коровая нуклеотидная последовательность: GGUUAGGGCUCGAAGUCGG) и 197-E3/197-B1 типа C (коровая нуклеотидная последовательность: GGUUAGGGCUUGAAGUCGG) содержат фактически идентичную коровую нуклеотидную последовательность (тип C формула-4; нуклеотидная последовательность: GGUUAGGGCUHGAAGUCGG). 191-D5-001, 197-B2 и 197-H1 не содержат 5’- и 3’-концевые участки нуклеотидов (у 197-H3 и у 197-E3 5’- и 3’-концевые участки нуклеотидов идентичны таковым у 197-B2). Однако соответствующие десять (197-B2, 197-E3, 197-H3) или девять из десяти (191-D5-001, 197-H1) нуклеотидов 5’-концевого участка нуклеотидов могут гибридизоваться с соответствующими десятью (197-B2, 197-E3, 197-H3) или девятью из десяти (191-D5-001, 197-H1) нуклеотидов 3’-концевого участка нуклеотидов (фиг.5). Таким образом, 5’-концевый участок нуклеотидов связывающих SDF-1 нуклеиновых кислот 197-B2, 191-D5-001, 197-H1, 197-E3 и 197-H3 типа C, как отмечалось ранее, плюс 191-A5, 197-B1, 197-H2, 197-D1 и 197-D2 включают типовую общую нуклеотидную последовательность «RKSBUSNVGR» (тип C формула-5-5’). 3’-концевый участок нуклеотидов связывающих SDF-1 нуклеиновых кислот 197-B2, 191-D5-001, 197-H1, 197-E3 и 197-H3 типа C, как отмечалось ранее, плюс 191-A5, 197-B1, 197-H2, 197-D1 и 197-D2 включают типовую общую нуклеотидную последовательность «YYNRCASSMY» (тип C формула-5-3’), где 5’- и 3’-концевые участки нуклеотидов связывающих SDF-1 нуклеиновых кислот 197-B2, 191-D5-001, 197-H1, 197-E3 и 197-H3 типа C являются предпочтительными. Эти предпочтительные 5’- и 3’-концевые участки нуклеотидов связывающих SDF-1 нуклеиновых кислот 197-B2, 191-D5-001, 197-H1, 197-E3 и 197-H3 типа C могут быть охарактеризованы общими формулами «RKSBUGSVGR» (тип C формула-6-5’; 5’-концевый участок нуклеотидов) и «YCNRCASSMY» (тип C формула-6-3’; 3’-концевый участок нуклеотидов).
Были сконструированы усеченные производные связывающей SDF-1 нуклеиновой кислоты 191-D5-001 типа C и испытаны в конкурентном замещающем тесте на связывание в сравнении с исходной молекулой 191-D5-001 (фиг.7A, фиг.7B и фиг.19). Сперва длину 5’- и 3’-концевых участков нуклеотидов сокращали от десяти нуклеотидов (191-D5-001) каждый до семи нуклеотидов каждый (191-D5-004), как представлено на фиг.7A, где девять из десяти (191-D5-001), или шесть из семи нуклеотидов (191-D5-004) 5’-концевого участка нуклеотидов и 3’-концевого участка нуклеотидов, соответственно, могли гибридизоваться друг с другом. Усечение семи нуклеотидов, соответственно, у 5’- и у 3’-концевого участка нуклеотидов (где шесть из семи нуклеотидов могли гибридизоваться друг с другом) приводило к пониженной аффинности связывания с SDF-1 (191-D5-004). Концевые участки нуклеотидов связывающей SDF-1 нуклеиновой кислоты 191-D5-004 типа C модифицировали, где не образующий пары нуклеотид «A» в составе 3’-концевого участка нуклеотидов 191-D5-004 замещали на «C» (191-D5-005). Эта модификация приводила к усилению связывания. Это производное связывающей SDF-1 нуклеиновой кислоты 191-D5-005 типа C показало связывание с SDF-1, сходное с 191-D5-001. Последующее усечение 5’- и 3’-концевого участка нуклеотидов до пяти нуклеотидов, соответственно, приводило к молекуле общей протяженностью в 29 нуклеотидов (191-D5-007). Вследствие сходных черт 191-D5-001 и связывающих SDF-1 нуклеиновых кислот 197-B2, 191-D5-001, 197-H1, 191-A5, 197-H3, 197-B1, 197-E3, 197-D1, 197-H2 и 197-D2 типа C, а также вследствие характеристик, показанных 191-D5-007, можно предположить, что 5’- и 3’-концевые участки нуклеотидов в принципе могут быть усечены до пяти нуклеотидов, где нуклеотидная последовательность «CGGGA» для 5’-концевого участка нуклеотидов и «UCCCG» для 3’-концевого участка нуклеотидов были успешно испытаны (связывающая SDF-1 нуклеиновая кислота 191-D5-007 010 типа C). Связывающая SDF-1 нуклеиновая кислота 191-D5-007 типа C неожиданно некоторым образом лучше связывается с SDF-1, чем 191-D5-001 (определено на уровне аптамера с помощью конкурентного теста связывания). Равновесную константу связывания KD 191-D5-007 определяли с помощью замещающего теста на связывание (KD=2,2 нМ, фиг.20) и измерений методом поверхностного плазмонного резонанса (KD=0,8 нМ, фиг.21). Значение IC50 (полумаксимальная концентрация ингибирования), равное 0,1 нМ для 191-D5-007, было получено с помощью in vitro хемотаксического теста в культуре клеток. Далее следовало усечение обоих концевых участков нуклеотидов до четырех нуклеотидов (191-D5-010, фиг.7A).
Последующие производные типа C связывающей SDF-1 нуклеиновой кислоты 191-D5-001 (191-D5-017/ -024/ -029), несущие 5’- и 3’-концевые участки нуклеотидов соответствующих четырех нуклеотидов также показали пониженные аффинности связывания с SDF-1 в конкурентном замещающем тесте на связывание в сравнении с 191-D5-007 (фиг.7B). Также были дополнительно протестированы альтернативные 5’- и 3’-концевые участки нуклеотидов длиной, соответствующей пяти нуклеотидам (191-D5-017-29a, 191-D5-017-29b, 191-D5-019-29a, 191-D5-024-29a, 191-D5-024-29b). Общая формула этих производных для 5’-концевого участка нуклеотидов представляет собой «XSSSSV» (тип C формула-7-5’), а для 3’-участка нуклеотидов представляет собой «BSSSXS» (тип C формула-7-3’), где XS отсутствует или представляет собой «S». Два из пяти протестированных вариантов показали аффинности связывания с SDF-1, идентичные 191-D5-007 (191-D5-024-29a, 191-D5-024-29b; фиг.7B). Последовательности 5’-концевых и 3’-концевых участков нуклеотидов производных 191-D5-001, которые показывают наилучшие аффинности связывания с SDF-1 и включают 5’-концевый и 3’-концевый участок нуклеотидов из пяти нуклеотидов, соответственно (191-D5-007, 191-D5-024-29a, 191-D5-024-29b), могут быть охарактеризованы общей формулой (5’-концевый участок нуклеотидов: «SGGSR», тип C формула-8-5’; 3’-концевый участок нуклеотидов: «YSCCS», тип C формула-8-3’).
Усеченные производные связывающей SDF-1 нуклеиновой кислоты 197-B2 типа C анализировали в конкурентном замещающем тесте на связывание в сравнении с исходной молекулой 197-B2 и 191-D5-007 (фиг.8). С помощью конкурентного замещающего теста на связывание в сравнении с 191-D5-007 было показано, что 197-B2 имеет ту же аффинность связывания с SDF-1, что и 191-D5-007. 5’- и 3’-концевые участки нуклеотидов укорачивали без потери аффинности связывания от десяти нуклеотидов (197-B2) каждый до пяти нуклеотидов каждый (197-B2-005), где нуклеотиды 5’-концевого участка нуклеотидов и 3’-концевого участка нуклеотидов полностью могли гибридизоваться друг с другом. Если 5’-концевый («GCGGG») и 3’-концевый («CCUGC») участок нуклеотидов 197-B2-005 замещали на «GCCGG» (5’-концевый участок нуклеотидов) и на «CCGGC» (3’-концевый участок нуклеотидов) 197-B2-006, аффинность связывания с SDF-1 присутствовала в полной мере. Поскольку 197-B2 и 191-D5-001 (и их производные) содержат коровую нуклеотидную последовательность (GGUUAGGGCUAGAAGUCGG) и были протестированы некоторые производные 191-D5 с 5’- и 3’-концевыми участками нуклеотидов длиной в соответствующие четыре нуклеотида, дополнительное усечение 5’- и 3’-концевых участков нуклеотидов было опущено. Были сконструированы две дополнительные производные, содержащие шесть нуклеотидов на 5’- и 3’-конце (5’- и 3’-концевые участки нуклеотидов), соответственно. Аффинность связывания с SDF-1 обеих молекул (197-B2-006-31a и 197-B2-006-31b) является той же самой, что и для 191-D5-007 и 197-B2-006 (фиг.8). Последовательности 5’-концевых и 3’-концевых участков нуклеотидов производных 197-B2, которые показывают наилучшую аффинность связывания с SDF-1 и включают 5’-концевые и 3’-концевые участки нуклеотидов пяти нуклеотидов, соответственно, могут быть охарактеризованы общей формулой (5’-концевый участок нуклеотидов: «GCSGG», тип C формула-9-5’; 3’-концевый участок нуклеотидов: «CCKGC», тип C формула-9-3’).
Комбинируя предпочтительные 5’- и 3’-участки нуклеотидов усеченных производных связывающих SDF-1 нуклеиновых кислот 191-D5-001 (5’-концевый участок нуклеотидов: «SGGSR», тип C формула-8-5’; 3’-концевый участок нуклеотидов: «YSCCS», тип C формула-8-3’) и 197-B2 типа C (5’-концевый участок нуклеотидов: «GCSGG», тип C формула-9-5’; 3’-концевый участок нуклеотидов: «CCKGC», тип C формула-9-3’), совместная предпочтительная общая формула для 5’-концевого и 3’-концевого участка нуклеотидов представляет собой «SSSSR» (5’-концевый участок нуклеотидов, тип C формула-10-5’) и «YSBSS» (3’-концевый участок нуклеотидов: тип C формула-10-3’).
1.4 Дополнительные связывающие SDF-1 нуклеиновые кислоты
Кроме того, были идентифицированы три дополнительные связывающие SDF-1 нуклеиновые кислоты, не содержащие связывающих SDF-1 мотивов «типа A», «типа B» и «типа C». Их анализировали как аптамеры замещающим тестом на связывание (фиг.9).
Следует понимать, что любая из последовательностей, показанная на фиг.1-9, является нуклеиновой кислотой согласно настоящему изобретению, включая эти их усеченные формы, но также включая и эти их удлиненные формы, однако при условии, что таким образом усеченные и удлиненные, соответственно, молекулы нуклеиновых кислот все еще способны связываться с мишенью.
Пример 2
Модификация остатком PEG массой 40 кДа и иная модификация связывающих SDF шпигельмеров
Для того, чтобы продлить продолжительность пребывания шпигельмера в плазме in vivo, шпигельмеры 193-G2-012, 192-A10-008, 191-D5-007, 197-B2-006 и 197-B2-006-31b ковалентно присоединяли к остатку полиэтиленгликоля (PEG) массой 40 кДа на 5'-конце, как описано в главе 3: (PEG-модифицированные клоны: 193-G2-012-5’-PEG, 192-A10-008-5’PEG, 191-D5-007-5’PEG, 197-B2-006-5’PEG и 197-B2-006-31b-5’PEG).
PEG-модифицированные молекулы шпигельмеров анализировали с помощью in vitro TAX-теста в культуре клеток (глава 5) и с помощью измерений методом плазмонного резонанса на приборе Biacore (глава 6). Все модифицированные PEG массой 40 кДа шпигельмеры все еще были способны ингибировать индуцированный с помощью SDF-1 хемотаксис и связываться с SDF-1 в низком наномолярном диапазоне концентраций (фиг.23A, 23B, 24A и фиг.24B).
Кроме того, cвязывающий SDF шпигельмер 192-A10-001 модифицировали 40 кДа-PEG, 30 кДа-PEG, 100 кДа-HES или 130 кДа-HES (PEG- и HES-модифицированные клоны: 192-A10-001-5’PEG40, 192-A10-001-5’PEG30, 192-A10-001-5’HES100, 192-A10-001-5’HES130; методика присоединения в главе 3). Как представлено на фиг.25A и фиг.25B, ни остаток PEG, ни остаток HES не влияли на способность шпигельмеров эффективно ингибировать индуцированный с помощью SDF-1 хемотаксис.
Пример 3
Синтез и получение производных аптамеров и шпигельмеров
3.1 Мелкосерийный синтез
Аптамеры и шпигельмеры получали твердофазным синтезом на синтезаторе ABI 394 (Applied Biosystems, Foster City, CA, USA), с помощью 2’TBDMS-РНК-фосфорамидитной химической технологии (Damha and Ogilvie, 1993). rA(N-Bz)-, rC(Ac)-, rG(N-ibu)- и rU- фосфорамидиты в D- и L-конфигурации были приобретены у ChemGenes, Wilmington, MA. Аптамеры и шпигельмеры очищали гель-электрофорезом.
3.2 Крупносерийный синтез плюс модификация
Шпигельмеры получали твердофазным синтезом на синтезаторе ÄktaPilot100 (Amersham Biosciences; General Electric Healthcare, Freiburg) с помощью 2’TBDMS-РНК-фосфорамидитной химической технологии (Damha and Ogilvie, 1993). L-rA(N-Bz)-, L-rC(Ac)-, L-rG(N-ibu)- и L-rU- фосфорамидиты были приобретены у ChemGenes (Wilmington, MA, USA). 5’-амино-модификатор был приобретен у American International Chemicals Inc. (Framingham, MA, USA). Синтез шпигельмеров начинали на модифицированных L-riboG, L-riboC, L-riboA, L-riboU соответственным образом стеклянных бусинах CPG с размерами пор 1000 Å (Link Technology, Glasgow, UK). Для присоединия следующего нуклеотида (15 мин на цикл) применяли 0,3M бензилтиотетразол (American International Chemicals Inc., Framingham, MA, USA) в ацетонитриле, и 3,5 эквивалентов соответственного 0,2M раствора фосфорамидита в ацетонитриле. Применяли цикл окислительного кэппинга. Дальнейшие стандартные растворители и реагенты для олигонуклеотидного синтеза были приобретены у Biosolve (Valkenswaard, NL). Шпигельмеры синтезировали DMT-ON; после снятия защит их очищали посредством препаративной обращенно-фазовой ВЭЖХ (Wincott F. et al., 1995), применяя носитель для обращенно-фазовой хроматографии Source15RPC medium (Amersham). 5’DMT-группу удаляли 80% уксусной кислотой (90 мин при комнатной температуре). Следующим шагом добавляли водный раствор 2M NaOAc и шпигельмер обессоливали фильтрованием в тангенциальном потоке при помощи регенерированной целлюлозной мембраны с пределом исключения 5 кДа (Millipore, Bedford, MA).
3.3 PEG-модификация
Для того чтобы продлить продолжительность пребывания шпигельмера в плазме in vivo, шпигельмеры ковалентно присоединяли к остатку полиэтиленгликоля (PEG) массой 40 кДа на 5'-конце.
Для PEG-модификации (технические детали данного способа PEG-модификации можно посмотреть в заявке на европейский патент EP 1306382) очищенные 5’-аминомодифицированные шпигельмеры растворяли в смеси H2O (2,5 мл), DMF (5 мл) и буфера A (5 мл; получен смешиванием лимонной кислоты • H2O [7 г], борной кислоты [3,54 г], фосфорной кислоты [2,26 мл] и 1M NaOH [343 мл] и доведением водой до конечного объема 1 л; pH=8,4 подводили с помощью 1M HCl).
Значение pH раствора шпигельмера доводили до 8,4 с помощью 1M NaOH. Затем PEG-NHS сложный эфир (Nektar Therapeutics, Huntsville, AL) массой 40 кДа добавляли при 37°C каждые 30 мин шестью порциями по 0,25 экв. до достижения максимального выхода продуктов реакции от 75 до 85%. Значение pH реакционной смеси во время добавления сложного эфира PEG-NHS поддерживали при 8–8,5 с помощью 1M NaOH.
Реакционную смесь гомогенизировали с 4 мл раствора мочевины (8M) и 4 мл буфера B (0,1 M ацетат триэтиламмония в H2O) и нагревали до 95°C в течение 15 мин. Затем PEG-модифицированный шпигельмер очищали посредством препаративной обращенно-фазовой ВЭЖХ, применяя носитель для обращенно-фазовой хроматографии Source 15RPC medium (Amersham), в градиенте концентрации ацетонитрила (буфер B; буфер C: 0,1 M ацетат триэтиламмония в ацетонитриле). Избыток PEG элюировали 5% буфером C, PEG-модифицированный шпигельмер - при 10–15% буфера C. Фракции с продуктом чистотой >95% (как оценивали с помощью ВЭЖХ) объединяли и смешивали с 40 мл 3M NaOAC. PEG-модифицированный шпигельмер обессоливали фильтрованием в тангенциальном потоке (регенерированная целлюлозная мембрана с пределом исключения 5 кДа, Millipore, Bedford MA).
3.4 HES-модификация
Для того чтобы продлить продолжительность пребывания шпигельмера в плазме in vivo, шпигельмеры ковалентно присоединяли к гидроксиэтилкрахмалу (HES) различной молекулярной массы >130 кДа и степенью замещения >0,5. Предпочтительным местом для конъюгации является 5’-конец шпигельмера.
Для HES-модификации (технические детали данного способа HES-модификации нуклеиновых кислот см. в выложенном описании к неакцептованной заявке на патент ФРГ DE 101 12 825 A1, а для D/L-нуклеиновых кислот - PCT WO 02/080979 A2), очищенный 5’-аминомодифицированный шпигельмер растворяли в бикарбонате натрия (0,3M, 1 мл) и значение pH доводили до 8,5.
Что касается данного шпигельмера, 5-кратный избыток свободной кислоты HES (3,3 ммоль, Supramol, Rosbach, Germany) и ди(N-сукцинимидил)карбоната (3,3 ммоль) добавляли к N,N-диметилформамиду (1 мл), чтобы получить раствор активированного N-гидроксисукцинимидного сложного эфира HES. Чтобы растворить все вещества, участвующее в реакции, смесь недолго перемешивали при 60°C, охлаждали до 25°C и затем перемешивали в течение 1,5 час при 25°C. Раствор шпигельмера добавляли в раствор активированного HES, и полученную смесь перемешивали при 25°C и pH 8,5. Реакцию контролировали аналитической ионообменной ВЭЖХ. Обычно конъюгацию продолжали вплоть до >75% в пределах 1 час.
Для очистки с помощью ионообменной ВЭЖХ на носителе Source 15Q medium (GE, Freiburg, Germany) реакционную смесь гомогенизировали с 10-кратным количеством буфера A (1 мМ EDTA, 25 мМ Tris, 10 мМ NaClO4 в системе вода/ацетонитрил 9:1, pH 4). Избыток HES элюировали при 5% буфере B (1 мМ EDTA, 25 мМ Tris, 500 мМ NaClO4 в системе вода/ацетонитрил 9:1, pH 4), где конъюгат «HES-шпигельмер» элюировали при 20-30% буфера B. Фракции с продуктом чистотой >95% (как оценивали с помощью ВЭЖХ) объединяли и обессоливали фильтрованием в тангенциальном потоке (регенерированная целлюлозная мембрана с пределом исключения 5 кДа, Millipore, Bedford MA).
Пример 4
Определение констант связывания (замещающий тест на связывание)
4.1 Прямой замещающий тест на связывание
Аффинности аптамеров к биотинилированному D-SDF-1 человека измеряли в формате замещающего теста на связывание при 37°C. Аптамеры метили фосфатом по 5’-концу T4 полинуклеотидкиназой (Invitrogen, Karlsruhe, Germany), применяя [γ-32P]-меченый АТФ (Hartmann Analytic, Braunschweig, Germany). Удельная радиоактивность меченых аптамеров составила 200000–800000 импульсов в минуту/пмоль. Аптамеры инкубировали после денатурации и ренатурации при 10, 20, 30 или 40 пМ концентрации при 37°C в буфере селекции (20 мМ Tris-HCl pH 7,4; 137 мМ NaCl; 5 мМ KCl; 1 мМ MgCl2; 1 мМ CaCl2; 0,1% [вес/объем] Tween-20) вместе с переменными количествами биотинилированного D-SDF-1 человека в течение 4-12 часов, чтобы достигнуть равновесия при малых концентрациях. Буфер селекции дополняли 10 мкг/мл сывороточного альбумина человека (Sigma-Aldrich, Steinheim, Germany) и 10 мкг/мл дрожжевой РНК (Ambion, Austin, USA), чтобы предотвратить адсорбцию партнеров по связыванию на поверхностях используемой лабораторной посуды из пластика или иммобилизационной матрице. Диапазон концентраций биотинилированного D-SDF-1 человека находился в пределах от 8 пМ до 100 нМ; полный объем реакции составил 1 мл. Пептид и комплексы «пептид-аптамер» были иммобилизованы на 1,5 мкл частиц Streptavidin Ultralink Plus (Pierce Biotechnology, Rockford, USA), которые были предварительно уравновешены буфером селекции и ресуспендированы в общем объеме 6 мкл. Частицы выдерживали в суспензии в течение 30 мин при соответствующей температуре в термомиксере. Иммобилизованную радиоактивность просчитывали на сцинцилляционном счетчике после отделения супернатанта и подходящей промывки. На график наносили процент связывания против концентрации биотинилированного D-SDF-1 человека, а значения констант диссоциации получали при использовании алгоритмов программного обеспечения (GRAFIT; Erithacus Software; Surrey U.K.), считая стехиометрическое отношение равным 1:1.
4.2 Конкурентный замещающий тест на связывание
Чтобы сравнить различные связывающие D-SDF-1 аптамеры, проводили конкурентный ранжирующий тест. С этой целью самый аффинный из доступных аптамер радиоактивно метили (см. выше), и он служил образцом сравнения. После денатурации и ренатурации его инкубировали при 37°C с биотинилированным D-SDF-1 человека в 1 мл буфера селекции при условиях, приводящих приблизительно к 5–10% связыванию с пептидом, после иммобилизации и промывки на носителе NeutrAvidin агарозе или Streptavidin Ultralink Plus (оба от компании Pierce) в отсутствие конкуренции. Избыток денатурированных и ренатурированных немеченых вариантов D-РНК-аптамеров добавляли до различных концентраций (например, 2, 10 и 50 нМ) вместе с меченым аптамером сравнения к параллельным реакциям связывания. Тестируемые аптамеры конкурировали с аптамером сравнения за связывание мишени, тем самым снижая сигнал связывания в зависимости от своих связывающих свойств. Аптамер, оказавшийся в этом тесте самым активным, может затем служить новым образцом для сравнительного анализа дальнейших вариантов аптамеров.
Пример 5
Анализ ингибирования индуцированного с помощью SDF-1 хемотаксиса связывающими SDF-1 шпигельмерами
Клетки линии Jurkat человеческого Т-клеточного лейкоза (полученные от DSMZ, Braunschweig) культивировали при 37°C и 5% CO2 в среде RPMI 1640 с добавкой к питательной среде Glutamax (Invitrogen, Karlsruhe, Germany), содержащей 10% фетальную бычью сыворотку, 100 Ед/мл пенициллина и 100 мкг/мл стрептомицина (Invitrogen, Karlsruhe, Germany). За один день до эксперимента клетки с плотностью 0,3×106/мл (9×106/30 мл) пересевали в новую колбу в стандартной среде (Invitrogen, Karlsruhe, Germany).
Для эксперимента клетки центрифугировали (5 мин при 300 g), ресуспендировали, подсчитывали и однократно промывали в 15 мл HBH (сбалансированный солевой раствор Хэнкса, содержащий 1 мг/мл бычьего сывороточного альбумина и 20 мМ HEPES; Invitrogen, Karlsruhe, Germany). Затем клетки ресуспендировали до плотности 3×106/мл или 1,33×106/мл в зависимости от типа используемого фильтровального планшета. Затем в течение нескольких часов клеткам давали возможность мигрировать через пористые мембраны фильтровальных планшетов в сторону раствора, содержащего SDF-1 и различные количества шпигельмера. Применяли либо планшеты и вставки Transwell с пористой поликарбонатной мембраной, размер пор 5 мкм (Corning; 3421), либо планшеты MultiScreen MIC (Millipore, MAMIC5S10).
5.1 Протокол для планшет Transwell
Стимулирующие растворы (SDF-1 + различные концентрации шпигельмера) изготавливали в 600 мкл HBH в нижних отделах планшетов Transwell и инкубировали в течение 20–30 мин. Все условия воспроизводили по меньшей мере дважды. Вставки переносили в лунки, содержащие стимулирующие растворы, и по 100 мкл суспензии клеток с плотностью 3×106/мл добавляли во вставки (3×105 клеток/лунку). Затем клеткам давали возможность мигрировать в течение 3 час при 37°C.
После этого вставки удаляли и добавляли 60 мкл рабочего раствора резазурина (Sigma, Deisenhofen, Germany; 440 мкМ в PBS; Biochrom, Berlin, Germany) в лунки (также и в калибровочные лунки). Затем планшеты инкубировали при 37°C в течение от 2,5 до 3 час. После инкубации по 200 мкл из каждой лунки переносили в черный 96–луночный планшет. Измерение сигналов флуоресценции проводили при 544 нм (возбуждение) и 590 нм (эмиссия) на многоканальном планшетном ридере Fluostar Optima (BMG, Offenburg, Germany).
5.2 Протокол для планшетов Millipore MultiScreen
Стимулирующие растворы (SDF-1 + различные концентрации шпигельмера) изготавливали в виде 10-кратных растворов в 0,2 мл малогабаритной 96-трубочном планшете. С помощью пипетки раскапывали по 135 мкл HBH в нижние отделы планшетов MultiScreen и добавляли по 15 мкл стимулирующих растворов. Все условия воспроизводили трижды. По прошествии от 20 до 30 мин данную фильтровальную планшету вставляли в планшет, содержащий стимулирующие растворы, и добавляли по 75 мкл суспензии клеток с плотностью 1,33×106/мл в лунки фильтровального планшета (1×105 клеток/лунку). Затем клеткам давали возможность мигрировать в течение 3 час при 37°C.
После этого вставной планшет удаляли и добавляли 20 мкл рабочего раствора резазурина (440 мкМ в PBS) в нижние лунки. Затем планшеты инкубировали при 37°C в течение от 2,5 до 3 час. После инкубации по 100 мкл из каждой лунки переносили в черный 96–луночный планшет. Измерение сигналов флуоресценции проводили, как описано выше.
5.3 Количественное определение
Для количественного определения значения флуоресценции корректировали на величину фоновой флуоресценции (лунки без клеток). Затем вычисляли разницу между экспериментальными параметрами в присутствии и в отсутствие SDF-1. Значение для образца без шпигельмера (только SDF-1) было принято за 100%, а значения для образцов со шпигельмером были рассчитаны в виде доли в процентах от него. Для построения зависимости «доза-ответ» значения в процентах отмечали на графике против концентрации шпигельмера, а величину IC50 (концентрация шпигельмера, при которой присутствует 50% активности без шпигельмера) определяли графически из полученной кривой.
5.4 Результаты
5.4.1 Дозозависимая стимуляция клеток линии Jurkat с помощью SDF-1 человека
Было обнаружено, что SDF-1 человека стимулирует миграцию клеток линии Jurkat дозозависимым образом с полумаксимальной стимуляцией приблизительно при 0,3 нМ (фиг.11).
5.4.2 Дозозависимое ингибирование индуцированного с помощью SDF-1 человека хемотаксиса связывающими SDF-1 шпигельмерами
Когда клеткам давали возможность мигрировать в сторону раствора, содержащего SDF-1 человека плюс повышающиеся концентрации связывающих SDF-1 шпигельмеров, наблюдали дозозависимое ингибирование. Соответствующие значения IC50 протестированных шпигельмеров определены в примере 1. Когда неспецифичный контрольный шпигельмер использовали вместо связывающих SDF-1 шпигельмеров, не наблюдали никакого эффекта ингибирования вплоть до 1 мкМ (фиг.26).
5.4.3 Дозозависимое ингибирование индуцированного с помощью SDF-1 мышей хемотаксиса связывающими SDF-1 шпигельмерами
SDF-1 обладает высокой межвидовой консервативностью: SDF-1 мышей отличается от SDF-1α человека на одну аминокислоту (изолейцин в положении 18 вместо валина). SDF-1 мышей может стимулировать хемотаксис клеток линии Jurkat (фиг.27), и было обнаружено, что это действие может ингибироваться шпигельмерами 192-A10-001 и 191-D5-007-5’-PEG с той же эффективностью, что и в случае SDF-1 человека (фиг.28).
Пример 6
Анализ связывания методом поверхностного плазмонного резонанса
Прибор Biacore 2000 (Biacore AB, Uppsala, Sweden) применяли, чтобы проанализировать связывание шпигельмеров с SDF-1α человека. Когда требовалось достичь присоединения SDF-1α через аминогруппы, SDF-1α диализовали против воды в течение 1–2 час (Millipore VSWP сложные эфиры смешанной целлюлозы; размер пор - 0,025 мкм), чтобы удалить препятствующие процессу амины. Сенсорные чипы CM4 (BIACORE AB, Uppsala, Sweden) активировали перед присоединением белка 35 мкл инъекцией разведения 1:1 0,4М NHS и 0,1 М EDC при скорости потока 5 мкл/мин. Затем вводили хемокин в концентрациях 0,1–1,5 мкг/мл при скорости потока 2 мкл/мин, пока выходной сигнал прибора находился в диапазоне 1000–2000 RU (относительные единицы). Непрореагировавшие сложные эфиры NHS дезактивировали инъекцией 35 мкл раствора гидрохлорида этаноламина (pH 8,5) при скорости потока 5 мкл/мин. Сенсорный чип дважды подвергали первичной обработке буфером связывания и уравновешивали при 10 мкл/мин в течение 1–2 час, пока не стабилизировалась базовая линия. Для всех белков кинетические параметры и константы диссоциации оценены с помощью ряда инъекций шпигельмера при 1000, 500, 250, 125, 62,5, 31,25 и 0 нМ концентрациях в буфере селекции (Tris-HCl, 20 мМ; NaCl, 137 мМ; KCl, 5 мМ; CaCl2, 1 мМ; MgCl2, 1 мМ; Tween 20, 0,1% [вес/объем]; pH 7,4). Во всех экспериментах анализ выполняли при 37°C, применяя приборную команду «Kinject», определяющую значение времени ассоциации 180 сек и времени диссоциации 360 сек, при скорости потока 10 мкл/мин. Анализ данных и вычисление констант диссоциации (KD) проводили с помощью пакета программ BIAevaluation 3.0 (BIACORE AB, Uppsala, Sweden), применяя стехиометрический 1:1 аппроксимирующий алгоритм Ленгмюра.
Пример 7
Ингибирование связывающими SDF-1 шпигельмерами связывания [125J]-SDF-1 с экспрессирующими CXCR4 клетками
7.1 Способ
Клон кДНК, кодирующий CXCR4-рецептор человека (NM_003467.2) был приобретен у OriGene Technologies (Rockville, MD) и клонирован в pCR3.1-вектор (Invitrogen, Karlsruhe, Germany). Полученный вектор трансфицировали в клетки CHO-K1 (DSMZ, Braunschweig, Germany), применяя Lipofectamin 2000 (Invitrogen), и стабильно экспрессирующие линии клеток отбирали обработкой генетицином. Экспрессию рецепторов подтверждали посредством RT-PCR.
Для тестов на связывание экспрессирующие CXCR4 клетки рассевали на покрытые полилизином 24-луночные планшеты при плотности клеток 1×105 клеток/лунку и культивировали в течение ночи при 37°C и 5% CO2 в среде CHO-Ultra medium (Cambrex, Verviers, Belgium), содержащей 50 Ед/мл пенициллина, 50 мкг/мл стрептомицина и 0,5 мг/мл генетицина.
Для тестов на связывание удаляли среду, и клетки однократно промывали сбалансированным солевым раствором Хэнкса, дополнительно содержащим 20 мМ HEPES, 1 мг/мл бычьего сывороточного альбумина, 0,1 мг/мл бацитрацина (HBB). Затем клетки инкубировали в 0,2 мл HBB в течение 1 час при комнатной температуре вместе с 50 пМ [125J]-SDF-1 (PerkinElmer, Rodgau, Germany) и различными концентрациями шпигельмера.
Неспецифическое связывание определяли добавлением немеченой SDF-1 человека (R&D Systems, Wiesbaden, Germany) до конечной концентрации 0,5 мкМ к нескольким лункам.
После инкубационного периода супернатант удаляли и лунки промывали 3 раза ледяным HBB. После этого клетки лизировали 0,1 мл 0,1M NaOH. Лизаты переносили в сцинтилляционные флаконы и после добавления 4 мл сцинтилляционной смеси Unisafe 1 Liquid Szintillation cocktail (Zinsser, Frankfurt, Germany) просчитывали на сцинтилляционном счетчике Beckman LS6500.
Поскольку значения для неспецифического связывания (связывания в присутствии высокого количества немеченого SDF-1) были несколько выше значений общего связывания при высокой концентрации (500 пМ) шпигельмера, разницу между максимальным связыванием («max») и связыванием в присутствии 500 пМ шпигельмера использовали для вычисления значений IC50.
7.2 Результаты
Нанесение на график связанного [125J]-SDF-1 в зависимости от концентрации шпигельмера показало, что связывание SDF-1 может быть блокировано шпигельмером 192-A10-001 с IC50 около 60 пМ (фиг.29).
Пример 8
Ингибирование связывающими SDF-1 шпигельмерами индуцированной с помощью SDF-1 активации MAP-киназы
8.1 Способ
Экспрессирующие CXCR4 клетки CHO рассевали на 6-луночные планшеты при плотности 0,5×106 клеток/лунку и культивировали около трех часов при 37°C и 5% CO2 в среде CHO-Ultra medium (Cambrex, Verviers, Belgium), содержащей 50 Ед/мл пенициллина, 50 мкг/мл стрептомицина и 0,5 мг/мл генетицина. После присоединения клеток среду удаляли и замещали на среду Хэма F12, содержащую 50 Ед/мл пенициллина, 50 мкг/мл стрептомицина. Клетки затем инкубировали в течение ночи при 37°C и 5% CO2. За три часа до стимуляции среду еще раз замещали на среду Хэма F12. Клетки стимулировали посредством 1 нМ SDF-1 человека и различными количествами шпигельмера в течение 5 или 10 минут. После этого среду удаляли, и клетки однократно недолго промывали 1 мл ледяного фосфатно-солевого буферного раствора (PBS), за чем следовал лизис с помощью SDS-буфера для образца (Tris/HCl, pH 6,8, 62,5 мМ; глицерин, 10%; SDS, 2%; бромфеноловый синий, 0,01%; бета-меркаптоэтанол, 5%). В каждую лунку добавляли 1 мкл 0,5 Ед/мкл бензоназы (Merck, Darmstadt, Germany), и после инкубации в течение от 5 до 10 мин при комнатной температуре лизаты переносили в пробирки Эппендорфа, инкубировали при 95°C в течение 5 мин и хранили при –20°C до дальнейшего анализа.
По 25 мкл лизатов разделяли на 10% денатурирующих SDS-полиакриламидных гелях. Затем белки переносили с помощью электроблоттинга на нитроцеллюлозные мембраны HybondECL (Amersham/GE Healthcare, Munich, Germany). После блоттинга мембраны окрашивали с помощью пурпурно-красного (0,2% в 3% трихлоруксусной кислоте) для контроля нагрузки и переноса белка, а затем блокировали путем инкубации в TBS-T (трис-солевой буфер (20 мМ Tris/HCl, pH 7,6, 137 мМ NaCl) с 0,1% Tween 20), содержащем 10% обезжиренного сухого молока, при 2–8°C в течение ночи.
Затем мембраны инкубировали с антителом кролика против фосфо-MAP-киназы (1:1000 в 10% молоке в TBS-T) в течение 2 час при комнатной температуре. После трехкратной промывки в течение 5 мин посредством TBS-T мембрану инкубировали с конъюгатом HRP с антителами против IgG кролика (1:2000 в 10% молоке в TBS-T) в течение 1 час при комнатной температуре. Затем мембрану опять промывали трижды по 5 мин в TBS-T, за чем следовала инкубация в течение 1 мин в хемилюминесцентном реагенте LumiGloR. Люминесценцию детектировали посредством экспозиции с хемилюминесцентными пленками HyperfilmTMECL (Amersham/GE Healthcare) в течение от 30 секунд до 2 минут. Антитела и реагент для детекции люминесценции были компонентами набора антител для технологии клеточной сигнализации PhosphoPlus p44/42 MAP Kinase (Thr202/Tyr204) Antibody kit from Cell Signaling Technology (New England Biolabs, Frankfurt a. M., Germany).
8.2 Результаты
Стимуляция экспрессирующих CXCR4 клеток с помощью 1 нМ SDF-1 человека в течение 5 мин приводит к выраженной стимуляции MAP-киназы, на что указывает увеличение интенсивности электрофоретической полосы, отображающей активированную MAP-киназу. Эта активация MAP-киназы может быть дозозависимым образом ингибирована шпигельмером 191-A10-001 (фиг.30).
Пример 9
Функциональный анализ связывающего SDF-1 человека шпигельмера 193-G2-012-5’-PEG в тесте аортального кольца на прорастание сосудов
Чтобы проверить, функционален ли связывающий SDF-1 человека шпигельмер 193-G2-012-5’-PEG также и в стандартном тесте на ангиогенез в культуре органов, были выполнены тесты аортального кольца на прорастание сосудов. Этот тест, в котором оценивали длину и обилие подобных сосудам прорастаний от эксплантатов, стал наиболее широко используемой моделью ангиогенеза в культурах органов (Auerbach et al. 2003). Было уже показано, что SDF-1 индуцирует прорастание сосудов в этом типе тестов (Salcedo et al. 1999).
Аорты крыс нарезали на кольца, погружали в коллагеновый матрикс и инкубировали с SDF-1 и SDF-1 плюс связывающий SDF-1 человека шпигельмер 193-G2-012-5’-PEG или SDF плюс нефункциональный PEG-модифицированный контрольный шпигельмер, не связывающий SDF-1. По прошествии от 6 до 7 дней анализировали прорастание сосудов (то есть отростков эндотелиальных клеток), фотографируя и определяя индекс прорастания сосудов.
9.1 Способ
Аорты самцов крыс получали от компании Bagheri Life sciences (Berlin, Germany). Аорты были свежепрепарированными и их транспортировали на льду в среде MCDB 131 (Invitrogen, Karlsruhe, Germany), содержащей 50 Ед/мл пенициллина, 50 мкг/мл стрептомицина (оба от Invitrogen, Karlsruhe, Germany) и 2,5 мкг/мл фунгизона (Cambrex, USA).
Для эксперимента изолированную аорту переносили в чашку для культуры клеток вместе со средой, и удаляли остаточную соединительную ткань. Затем аорту нарезали скальпелем на кольца длиной приблизительно от 1 до 2 мм. Кольца интенсивно промывали (по меньшей мере пять раз) в среде Medium199 (Invitrogen, Карлсруэ, Германия), а затем помещали в лунки 24-луночных планшетов, содержащих 450 мкл раствора коллагена на лунку. Этот раствор коллагена изготавливали, смешивая 9 мл коллагена хвоста крысы (3 мг/мл в 0,1% уксусной кислоте; Sigma, Deisenhofen, Germany) с 1,12 мл 10X среды Medium199 (Invitrogen, Karlsruhe, Germany), 1,12 мл 10X буфера для коллагена (0,05н NaOH, 200 мМ HEPES, 260 мМ NaHCO3) и 0,6 мл 200 мМ глутамина. Кольца ориентировали таким образом, чтобы обрезанные края были перпендикулярны основанию лунки. Коллагену давали затвердеть, инкубируя планшеты в течение по меньшей мере одного часа при 37°C. После этого добавляли 1 мл на лунку среды MCDB131 с добавками (SDF-1 и шпигельмеры). Затем кольца инкубировали при 37°C в течение шести-семи дней. В качестве контроля прорастания сосудов дополнительно проводили эксперименты в присутствии VEGF (эндотелиальный фактор роста сосудов).
Прорастание сосудов документировали, фотографируя цифровой камерой. В некоторых случаях кольца фиксировали добавлением 1 мл 10% параформальдегида и хранили при 2-8°C для последующей документации. Фотографии анализировали с помощью пакета программ для обработки изображения Scion Image. После калибровки с помощью снимка, сделанного со ступенчатого микрометра столика микроскопа, на расстоянии 0,33 мм от одного края кольца прочерчивали линию. Вдоль этой линии программным обеспечением вычерчивалась графическая гистограмма, гистограммы распечатывали и подсчитывали пики (соответствовавшие проросткам, пересекающим данную линию). Это число было принято как индекс прорастания сосудов. Для каждого патологического состояния оценивали по 4-5 колец. Статистический анализ был выполнен с помощью программы WinSTAT для Excel.
9.2 Результаты
Можно показать, что SDF-1 вызывает прорастание сосудов, и что этот эффект может быть блокирован связывающим SDF-1 человека шпигельмером 193-G2-012-5’-PEG. Не наблюдалось никакой блокировки индуцированного с помощью SDF-1 прорастания сосудов нефункциональным PEG-модифицированным контрольным шпигельмером (фиг.31 и 32).
Пример 10
Мобилизация гемопоэтических стволовых клеток/гемопоэтических клеток-предшественников (HSC/HPC) у мышей разовой внутривенной инъекцией NOX-A12, модифицированного остатками PEG с молекулярной массой 30 или 40 кДа
10.1 Пробные вещества и схема введения
С помощью инъекций мышам внутривенно вводили по 13,4 мг/кг связывающего SDF-1 шпигельмера NOX-A12-JE40 (SEQ ID NO:132), связывающего SDF-1 шпигельмера NOX-A12-NO30 (SEQ ID NO:242) или контрольного шпигельмера revNOX-A12 (SEQ ID NO:243), у которого отсутствует активность связывания с SDF-1. Через 1, 6, 24 или 48 час после инъекции [по 5 мышей на каждое вещество и каждый момент времени] животных забивали, и собирали кровь. Контрольным группам с помощью инъекции вводили лекарственную основу (5% глюкозу), AMD-3100 (Sigma, France, 5 мг/кг подкожно) или гранулоцитарный колониестимулирующий фактор (G-CSF, нейпоген) (2,5 мкг/мышь/инъекцию, каждые 12 час). Забой животных проводили в те же моменты времени, что указаны выше (см. фиг.33). Для группы G-CSF (нейпоген): Из-за схемы инъекции по одной инъекции каждые 12 час, животные, которых забивали через один и шесть часов, по этой причине получили только одну инъекцию, группа, забитая через 24 час, получила 2 инъекции (спустя 0 и 12 час), группа, забитая через 48 час, получила четыре инъекции (спустя 0, 12, 24 и 36 час) (см. фиг.33). Число кровяных клеток определяли в гемоцитометре.
10.2 Детекция HSC/HPC
Пятьдесят (50) мкл цельной крови сперва инкубировали с блокирующим FcR реагентом (кат. № 130-092-575, Miltenyi Biotec, Paris, France) в буфере для окрашивания (PBS [кат. № 17-516F, Lonza], 0,2% BSA [кат. № A7030, Sigma], 0,02% NaN3 [кат. № S2002, Sigma]) и затем инкубировали при комнатной температуре в темноте в течение 30 мин с конъюгированными с FITC антителами против CD117 и конъюгированными с PE антителами против Ly-6A/E, либо с соответствующими изотипами (как описано в таблице ниже).
Красные кровяные клетки лизировали с помощью процедуры «фиксация-лизис». Вкратце, буфер для процедуры «фиксация-лизис» можно приготовить добавлением 25 мкл раствора 3 из набора IOTest (10X фиксирующий раствор [кат. № A07800, Beckman Coulter, Villepinte, France]) к 1 мл лизирующего раствора VersaLyse [кат. № A09777, Beckman Coulter], и 1 мл смеси добавляли к окрашенным клеткам. После встряхивания на вортексе и инкубации в течение 10 мин в темноте при комнатной температуре клетки центрифугировали и однократно промывали 3 мл буфера для окрашивания, а также ресуспендировали в 1 мл сравнительного раствора микрогранул (PKH26, кат. № P7458, 220000 гранул/мл, Sigma, разбавленный 1/2 в буфере для окрашивания). Образцы хранили на льду защищенными от света до проведения FACS-анализа.
Флуоресценцию поверхности клеток анализировали с помощью проточного цитометра (FACS, CyFlow® space), применяя возбуждение лазером с длиной волны 488 нм. Для каждого образца накапливали в общей сложности по 10000 элементарных актов.
10.3 Вычисление абсолютного числа клеток на мкл
Объем образцов крови был 50 мкл. В конце процедуры подготовки белые клетки крови (которые содержались в 50 мкл образцов крови) ресуспендировали в 0,75 мл раствора микрогранул (полученный разбавлением 1/2 концентрированного раствора, содержащего 220000 гранул в 1 мл). Число клеток на мл этого последнего раствора составило: (CN/BN)×(220000/2), а общее количество клеток: (CN/BN)×(220000/2)×(0,75/1), где CN - число одиночных импульсов счета для клетки, а BN - число одиночных импульсов счета для микрогранул.
В связи с этим, это общее число клеток содержалось в 50 мкл крови, а абсолютное число клеток (ACN) на мкл крови составило: (CN/BN)×(220000/2)×(0,75/1)×(1/50).
10.4 Определение числа колониеобразующих единиц (КОЕ)
Только кровь, взятую в те моменты времени, когда число гемопоэтических клеток-предшественников (HPC)/гемопоэтических стволовых клеток (HSC) было максимальным, отбирали для испытаний в КОЕ-тестах (один и тот же момент времени для всех мышей на каждое вещество). Истощение красных кровяных клеток (RBC) в образцах нормальной периферической крови проводили посредством добавления 10 объемов 10x лизирующего буфера (0,8% NH4Cl с 0,1 мМ EDTA) (кат. № 07800, StemCell Technologies) в периферическую кровь, перемешивания путем опрокидывания 3-4 раза пробирки и инкубации на льду в течение 5-15 минут. После центрифугирования в течение 7 мин при 1200 об/мин образец дважды промывали от белых кровяных клеток (WBC) средой Дульбекко, модифицированной по способу Исков (IMDM), содержащей 2% термоинактивированную фетальную бычью сыворотку (FBS, кат. № DE14-802F, Lonza). Содержащие ядра WBC подсчитывали с помощью гемоцитометра (MS9-5 counter (Melet Schloesing, Osny, France)) после разбавления 1/20 в 3% уксусной кислоте с метиленовым синим (кат. № 07060, StemCell Technologies).
Клетки наносили на планшеты по три точки в 0,9% метилцеллюлозе, содержащей IMDM (кат. № MethoCult® M03434, StemCell Technologies), уже содержащей 2% термоинактивированную FBS, рекомбинантный мышиный (rm) фактор стволовых клеток (SCF, рост тучных клеток, а также миелоидных и лимфоидных предшественников), rm IL-3 и rh IL-6 (рост ранних миелоидных предшественников всех клеточных линий) и рекомбинантный человеческий эритропоэтин (rh EPO, рост эритроидных предшественников). Вкратце, 10-кратно сконцентрированные WBC (0,4 мл) разводили в метилцеллюлозной полной IMDM (конечную концентрацию предстоит определить во время сертификационного эксперимента), тщательно встряхивали на вортексе и оставляли на 2-5 минут, чтобы дать пузырькам исчезнуть перед диспергированием. 1,1 мл метилцеллюлозной среды, содержащей смесь клеток, диспергировали, используя шприц с люэровским наконечником и иглу с тупым концом 16G (кат. № 28110, StemCell Technologies), в каждую из трех 35 мм чашек для культивирования (кат. № 27150, StemCell Technologies). Чашки для культивирования осторожно наклоняли и поворачивали, чтобы распределить метилцеллюлозу равномерно. Большие чашки для культивирования с добавленной открытой стерильной водой с вложенной 35 мм чашкой для культивирования использовали, чтобы поддерживать 35 мм чашки для культивирования во время инкубации при 37°C, 5%-ом CO2, при 95%-ой влажности в течение 7-12 дней.
После завершения инкубации колонии подсчитывали вручную как BFU-E (колониеобразующие «взрывообразующие» единицы эритроидов), CFU-GM (колониеобразующие единицы гранулоцитов и/или макрофагов) и CFU-GEMM (колониеобразующие единицы гранулоцитов-эритроцитов-макрофагов-мегакариоцитов) с помощью инвертированного микроскопа и 60 мм чашки для подсчета колоний с координатной сеткой (кат. № 27500, StemCell Technologies). Число колоний нормировали в виде КОЕ/мл крови.
10.5 Результаты проточной цитометрии
Связывающие SDF-1 шпигельмеры NOX-A12-JE40 и NOX-A12-NO30 приводят к значительному увеличению HPC/HSC, что определено двойным окрашиванием CD117 и Ly-6 A/E в FACS-анализе (см. фиг.34). Пик наблюдали через 6 час после введения шпигельмера. NOX-A12-JE40 приводит к высвобождению 700 CD117+Ly-6 A/E+ клеток на мкл. Связывающий SDF-1 шпигельмер NOX-A12-NO30 высвобождает 300 CD117+Ly-6 A/E+ клеток на мкл крови. В контрольной (лекарственная основа) группе насчитывали 100 CD117+Ly-6 A/E+ клеток на мкл крови. Эффект G-CSF на мобилизацию HSC/HPC был заметен у мышей, получивших 4 инъекции G-CSF и забитых через 48 час после первой инъекции (250 CD117+Ly-6 A/E+ клеток на мкл). AMD-3100 и связывающий SDF-1 шпигельмер revNOX-A12-JE40 не показали никакого эффекта (см. фиг.34 для графического представления всех моментов времени и всех групп).
В результате обработки связывающим SDF-1 шпигельмером NOX-A12-JE40 временное увеличение HSC/HPC сопровождалось временным увеличением общего содержания белых кровяных клеток, к чему приводят главным образом повышенные количества макрофагов, гранулоцитов и нейтрофилов, а также эозинофилов. Было обнаружено, что нейпоген вызвал увеличение количества белых кровяных клеток (счет WBC), сопряженное со снижением количества эозинофилов [%] (см. фиг.41).
10.6 Результаты КОЕ-тестов
По сравнению с лекарственной основой (0,8 КОЕ/мл), все вещества приводили к повышенным средним значениям общих КОЕ. Однако revNOX-A12-JE40 (6 час) и AMD3100 (6 час) показали только незначительные эффекты (1,1 и 1,5 КОЕ/мл, соответственно). Одна инъекция связывающего SDF-1 шпигельмера NOX-A12-JE40 приводила приблизительно к таким же значениям КОЕ/мл крови через 6 час, что и четыре инъекции G-CSF (определяли через 48 час после первой инъекции и через 12 час после 4й инъекции) (3,7 и 3,2 КОЕ/мл, соответственно). Это в четыре раза выше значения, определенного для лекарственной основы. Связывающий SDF-1 шпигельмер NOX-A12-NO30 удваивал счет КОЕ по сравнению с лекарственной основой (1,9 КОЕ/мл). См. фиг.35 для общего осмотра.
Пример 11
Ингибирование хороидальной неоваскуляризации (CNV)
Моделирование на животных «индуцированной лазером хороидальной неоваскуляризации» применяли, чтобы оценить действие экспериментальных лекарственных препаратов на новообразованные сосуды сетчатки и сосудистой оболочки глаза. Это происходит при таких заболеваниях, как влажная или «пролиферативная» форма возрастной макулярной дегенерации (AMD), диабетическая ретинопатия и окклюзия вен сетчатки глаза. Как было показано, CXCR4 экспрессируется при индуцированной лазером CNV (Lima e Silva et al., FASEB J. 21:2007). Он был локализован совместно с CD45 и F4/80 экспрессирующими клетками, означая то, что эти клетки представляют собой полученные из BM макрофаги. Ингибиторы CXCR4 уменьшали индуцированную лазером CNV. Но не был исследован случай, если эти клетки также экспрессируют SDF1. В этом исследовании авторами оценивалось, блокирует ли неоваскуляризацию связывающий SDF-1 шпигельмер NOX-A12-JE40 (SEQ ID NO:132).
11.1 Способы
Двадцати двум мышам C57/BL6J, не моложе 12 недель, проводили анестезию и наносили по 3 лазерных ожога на глаз. Животные обнаруживают симптомы хороидальной неоваскуляризации на местах лазерных ожогов. Один день спустя 2 мкл 440 мкМ раствора связывающего SDF-1 шпигельмера NOX-A12-JE40, растворенного в растворе Рингера, вводили путем инъекции в стекловидное тело в один глаз (доза: 0,88 нмоль = 12,9 мкг [только олигонуклеотидная часть молекулы]=48 мкг [молекула целиком, включая PEG]), в то время как другой глаз получил раствор Рингера в качестве контроля. Спустя 14 дней после обработки лазером животным проводили перфузию декстран-флуоресцеина, и изготавливали хороидальные тотальные препараты. Тотальные препараты оценивали по изменениям сосудов сосудистой оболочки и площади CNV перепонки.
Поскольку в один глаз был введен шпигельмер, а в другой глаз каждого животного был введен только буфер, применяли знаковые ранговые критерии Уилкоксона, которые имеют отношение к разнице между обработанным и контрольным глазом каждого животного. Знаковый ранговый критерий Уилкоксона анализирует разницы между двумя связанными измерениями, в данном случае - между обработанным и контрольным глазом каждого животного. Он выявляет существенные различия, даже если серия обработанных глаз и серия контрольных глаз не являются статистически различными. Использовали следующую R-команду: wilcoxsign_test(V1~V2, data=d0, distribution=«exact»). p<0,05 является значимым на уровне 95%.
11.2 Результаты
Оказалось возможным количественно оценить тринадцать из 22 мышей. Среднее значение площади неоваскуляризации обработанных NOX-A12-JE40 глаз было меньше, чем среднее для глаз, обработанных раствором Рингера, указывая на то, что шпигельмер снижает формирование CNV. Значение p, рассчитанное с помощью теста согласованных пар знаковых рангов Уилкоксона, составило 0,021. Поэтому было сделано заключение, что связывающий SDF-1 шпигельмер NOX-A12-JE40 значительно снижает индуцированную лазером хороидальную неоваскуляризацию у моделей CNV на мышах, обещая перспективный терапевтический эффект (см. фиг.36).
Пример 12
Эффективность при диабетической нефропатии
Гломерулосклероз, например, при диабете, остается основной причиной конечной стадии заболевания почек, поскольку направленность лечения на ангиотензин-зависимые патологические механизмы не всегда предотвращает прогрессирование заболевания. Следовательно, необходимы другие стратегии лечения, добавляющие что-либо новое в терапевтический арсенал для лечения гломерулосклероза. Данные недавних экспериментальных исследований устанавливают взаимосвязь между прогрессированием гломерулосклероза у мышей и людей, страдающих диабетом, с внутрипочечным воспалением. Например, мофетила микофенолат, метотрексат или облучение понижают выделение альбумина с мочой и гломерулосклероз у крыс с диабетом, индуцированным стрептозотоцином. Несмотря на это, молекулярные и клеточные механизмы внутрипочечного воспаления при диабетической нефропатии остаются недостаточно охарактеризованными. Пациенты с диабетической нефропатией имеют повышенный титр маркеров острой фазы воспаления в сыворотке, но это может и не означать внутрипочечного воспаления.
В этом исследовании позднее блокирование SDF-1 связывающим SDF-1 шпигельмером NOX-A12-JE40 (SEQ ID NO:132) анализировали на db/db мышах, перенесших удаление одной почки в возрасте 6 недель. Введение NOX-A12-JE40 начинали в возрасте 4 месяцев и проводили 3 раза в неделю при дозе 50 мг/кг подкожно (соответствует 13,4 мг/кг олигонуклеотидной части). Спустя два месяца животных забивали и определяли число баллов по шкале оценки гломерулосклероза.
Исходя из этого числа баллов, каждый конкретный клубочек оценивали в баллах по лейкоцитарной инфильтрации и заживлению ткани. Число баллов 0 описывает здоровые клубочки, в то время как число баллов 4 описывает полностью фиброзную форму (Ninichuk, Clauss et al. 2008).
12.2 Результаты
Хотя мыши «дикого типа» в основном не имеют повреждений почек в возрасте шести месяцев, db/db мыши того же возраста демонстрируют значительное поражение почек. Поражение почки у db/db мышей, лишенных одной почки в возрасте шести недель, является даже еще более сильным.
Связывающий SDF-1 шпигельмер NOX-A12-JE40 (SEQ ID NO:132), но не неспецифичный контрольный шпигельмер revNOX-A12-JE40 (SEQ ID NO:243) обратной последовательности, значительно облегчает поражение почки, наблюдаемое у лишенных одной почки db/db мышей через шесть месяцев: наблюдалось значительно меньше клубочков с наивысшим числом баллов поражения (3 и 4). Вместо этого, у большего числа клубочков наблюдали менее заметные перемены (инфильтрацию лейкоцитов) (см. фиг.37).
Пример 13
Cвязывающий SDF-1 шпигельмер NOX-A12-JE40 ингибирует индуцированное с помощью VEGF подтекание сосудов сетчатки глаза у пигментированных кроликов
Подтекание сосудов сетчатки случается при некоторых заболеваниях глаз, таких как возрастная макулярная дегенерация и окклюзия вен в сетчатке. Это приводит к макулярному отеку, ухудшающему зрение.
13.1 Способы
У модели на животном подтекание сосудов сетчатки глаза можно вызвать инъекцией VEGF в стекловидное тело кроликов. У этой модели подтекание сосудистой системы сетчатки глаза измеряли фотометрией флуоресцеина через 48 час после инъекции VEGF в стекловидное тело. Инъекцию испытательного образца проводили за 5 дней до VEGF-стимуляции. Подтекание, наблюдаемое через два дня после введения VEGF, решено считать не имеющим непосредственного отношения к усилению подтекания, которое кратковременно наблюдается после VEGF-стимуляции, а скорее эффектом более продолжительных последующих процессов, запускаемых инъекцией VEGF (Edelman, Lutz et al. 2005).
В этом исследовании использовали по восемь пигментированных кроликов (Fauve de Bourgogne, возраст - 2-3 месяца) на группу.
Были выделены следующие группы:
- 4 NOX-A12-JE40-дозовые группы (105 нмоль, 40 нмоль, 8 нмоль, 1,6 нмоль)
- эталонная группа (кенакорт ретард (R) (4% ацетонид триамцинолона), 2 мг в стекловидное тело) и
- лекарственная основа (5% глюкоза для инфузий).
Объем инъекции составлял 50 мкл для всех групп. Спустя пять дней после введения вещества в правые глаза, также только в правые глаза вводили VEGF: по 500 нг рекомбинантного VEGF165 человека в 50 мкл PBS. Через 48 час измеряли подтекание сетчатки в глазах обезболенных животных с помощью окулярной флуорометрии (1 час после внутривенной инъекции флуоресцеина натрия (10% в 0,9% хлориде натрия, 50 мг/кг)). Вкратце, интенсивность флуоресценции сканировали in vivo вдоль оптической оси от роговой оболочки до сетчатки в обоих глазах, используя устройство флуоротрон (FM-2 Fluorotron Master). Полученные кривые распределения интенсивности интегрировали, и вычисляли отношение площадей под кривой (AUC) у обработанного глаза к необработанному глазу. Затем вычисляли групповые средние значения и стандартные отклонения и представляли в виде графика.
13.2 Результаты
Инъекции в стекловидное тело 105 и 40 нмоль связывающего SDF-1 шпигельмера NOX-A12-JE40 в 50 мкл глюкозы за 5 дней перед инъекцией рекомбинантного VEGF165 человека (500 нг в 50 мкл) существенно ингибировали индуцированное с помощью VEGF подтекание сосудистой системы сетчатки через 48 час после VEGF-стимуляции. Эталонный образец, триамцинолон, также показал сильное снижение подтекания сосудов. Эксперименты на приборе BiaCore, проделанные раньше, подтвердили, что при используемых концентрациях не происходит никакого VEGF-связывания связывающим SDF-1 шпигельмером NOX-A12-JE40. По этой причине есть основания полагать, что NOX-A12-JE40 блокирует нижеследующий каскад реакций с участием VEGF (например, рекрутинг лейкоцитов, ослабление тесных эндотелиальных соединений в результате суперэкспрессии SDF-1), что приводит к продолжительному подтеканию сосудов (см. фиг.38). По этой же причине NOX-A12-JE40 может быть полезен для лечения макулярного отека или отека сетчатки как таковых, либо вторичных по отношению к возрастной макулярной дегенерации или окклюзии вен в сетчатке.
Пример 14
Связывающий SDF-1 шпигельмер NOX-A12-JE40 ингибирует ангиогенез при индуцированной кислородом ретинопатии
Модель индуцированной кислородом ретинопатии на мышах представляет собой модель для имитации индуцированной гипоксией неоваскуляризации сетчатки, наблюдаемой при диабетической ретинопатии, особенно при пролиферативной диабетической ретинопатии и при возрастной макулярной дегенерации (Smith, Wesolowski et al. 1994). Модель также называют моделью ретинопатии недоношенных, поскольку недоношенные дети, помещенные в инкубаторы в больницах, теряют зрение из-за слишком длительного пребывания в инкубаторах в условиях повышенного содержания кислорода, что приводит к аномальному росту сосудов в сетчатке глаза во время пребывания в инкубаторе и после их возвращения к условиям с нормальным содержанием кислорода.
14.1 Способы
При моделировании на мышах, новорожденных C57BL/6J мышей инкубировали при 75% содержании кислорода в течение дней P5-P12 после рождения. После возращения к условиям с нормальным содержанием кислорода животные обнаруживают симптомы неоваскуляризации сетчатки из-за относительной гипоксии. Связывающий SDF-1 шпигельмер NOX-A12-JE40 (880 пмоль в 2 мкл раствора Рингера) вводили путем инъекции на день P12. На день P17 мышам проводили перфузию декстран-флуоресцеина для визуализации сосудистой системы сетчатки. Тотальные препараты сетчатки использовали для оценки изменений сосудов сосудистой системы сетчатки в закодированной форме с помощью системы балльных оценок. Перфузия FITC-декстрана позволяет количественно оценить только перфузированные сосуды. Для того чтобы визуализовать несформированные, еще не перфузированные сосуды, тотальные препараты окрашивали на изолектин-B4 и анализировали под флуоресцентным микроскопом соответствующим образом. Глаза, обработанные связывающим SDF-1 шпигельмером, сравнивали с глазами, обработанными лекарственной основой, внутри-индивидуальным образом. Статическую значимость вычисляли с помощью знаковых ранговых критериев Уилкоксона. Значение p=0,05 соответствует 95% достоверности.
Следующие параметры оценивали в баллах:
1) число сосудистых пучков
2) абсолютный размер области пучка
3) относительный размер области пучка
4) число прорастаний сосудов (клубочки)
5) абсолютный размер лишенной сосудов зоны
6) относительный размер лишенной сосудов зоны
Число баллов по шкале оценки ретинопатии вычисляли, исходя из этих параметров по FITC-декстрановым фотоснимкам (Higgins, Yu et al. 1999). Доверительные уровни для данных параметров и показатель по шкале оценки ретинопатии показаны на фиг.40.
14.2 Результаты
Из 34 проверенных мышей 24 тотальных препарата можно было оценить после перфузии FITC-декстрана, и 15 можно было оценить после окрашивания на изолектин. Единственная инъекция 880 пмоль связывающего SDF-1 шпигельмера NOX-A12-JE40 на день P12 значительно ингибировала формирование клубочков и тем самым улучшила общее число баллов по шкале оценки ретинопатии, как наблюдали на день P17 (см. фиг.39 и 40). По этой причине NOX-A12-JE40 может иметь благоприятное воздействие на заболевания с индуцированной гипоксией неоваскуляризацией, особенно, заболеваний глаза (например, диабетическая ретинопатия, AMD).
Ссылки
Полные библиографические данные документов, перечисленных в данном документе, если не указано обратное, представляют собой нижеследующее, где раскрытие указанных ссылок включено в данный документ в виде ссылки.
Abi-Younes, S., A. Sauty, et al. (2000). "The stromal cell-derived factor-1 chemokine is a potent platelet agonist highly expressed in atherosclerotic plaques." Circ Res 86(2):131-8.
Aiuti, A., I. J. Webb, et al. (1997). "The chemokine SDF-1 is a chemoattractant for human CD34+ hematopoietic progenitor cells and provides a new mechanism to explain the mobilization of CD34+ progenitors to peripheral blood." J Exp Med 185(1):111-20.
Altschul S.F, Gish W, Miller W, Myers E.W, Lipman D.J. (1990), Basic local alignment search tool. J Mol. Biol. 215(3):403-10.
Altschul S.F, Madden T.L, Schaffer A.A, Zhang J, Zhang Z, Miller W, Lipman D.J. (1997). Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res. 25(17):3389-402.
Ambati, J., A. Anand, et al. (2003). An animal model of age-related macular degeneration in senescent Ccl-2- or Ccr-2-deficient mice. Nat Med. 9:1390-7.
Arya, S. K., C. C. Ginsberg, et al. (1999). "In vitro phenotype of SDF1 gene mutant that delays the onset of human immunodeficiency virus disease in vivo." J Hum Virol 2(3):133-8.
Auerbach et al. (2003) Angiogenesis assays: a critical overview. Clin. Chem. 49:32-40.
Avniel, S., Z. Arik, et al. (2006). "Involvement of the CXCL12/CXCR4 pathway in the recovery of skin following burns." J Invest Dermatol. 126(2):468-76.
Baggiolini, M. (1998). "Chemokines and leukocyte traffic." Nature 392(6676):565-8.
Baggiolini, M., B. Dewald, et al. (1994). "Interleukin-8 and related chemotactic cytokines--CXC and CC chemokines." Adv Immunol 55: 97-179.
Balabanian K, Lagane B, Infantino S, Chow KY, Harriague J, Moepps B, Arenzana-Seisdedos F, Thelen M, Bachelerie F. (2005) The chemokine SDF-1/CXCL12 binds to and signals through the orphan receptor RDC1 in T lymphocytes. J Biol. Chem. 280(42):35760-35766.
Balabanian, K., B. Lagane, et al. (2005). "WHIM syndromes with different genetic anomalies are accounted for by impaired CXCR4 desensitization to CXCL12." Blood 105(6):2449-57.
Balabanian, K., J. Couderc, et al. (2003). "Role of the chemokine stromal cell-derived factor 1 in autoantibody production and nephritis in murine lupus." J Immunol. 170(6):3392-400.
Balkwill, F. (2004). "Cancer and the chemokine network." Nat Rev Cancer 4(7):540-50.
Bazan, J. F., K. B. Bacon, et al. (1997). "A new class of membrane-bound chemokine with a CX3C motif." Nature 385(6617):640-4.
Bertolini, F., C. Dell'Agnola, et al. (2002). "CXCR4 neutralization, a novel therapeutic approach for non-Hodgkin's lymphoma." Cancer Res 62(11):3106-12.
Bleul, C. C., J. L. Schultze, et al. (1998). "B lymphocyte chemotaxis regulated in association with microanatomic localization, differentiation state, and B cell receptor engagement." J Exp Med 187(5):753-62.
Bleul, C. C., M. Farzan, et al. (1996). "The lymphocyte chemoattractant SDF-1 is a ligand for LESTR/fusin and blocks HIV-1 entry." Nature 382(6594):829-33.
Bleul, C. C., R. C. Fuhlbrigge, et al. (1996). "A highly efficacious lymphocyte chemoattractant, stromal cell-derived factor 1 (SDF-1)." J Exp. Med. 184(3):1101-9.
Brooks, H. L., Jr., S. Caballero, Jr., et al. (2004). "Vitreous levels of vascular endothelial growth factor and stromal-derived factor 1 in patients with diabetic retinopathy and cystoid macular edema before and after intraocular injection of triamcinolone." Arch Ophthalmol. 122(12):1801-7.
Broxmeyer, H. E., A. Orazi, et al. (1998). "Myeloid progenitor cell proliferation and mobilization effects of BB10010, a genetically engineered variant of human macrophage inflammatory protein-1alpha, in a phase I clinical trial in patients with relapsed/refractory breast cancer." Blood Cells Mol. Dis 24(1):14-30.
Broxmeyer, H. E., L. Benninger, et al. (1995). "Effects of in vivo treatment with PIXY321 (GM-CSF/IL-3 fusion protein) on proliferation kinetics of bone marrow and blood myeloid progenitor cells in patients with sarcoma." Exp. Hematol. 23(4):335-40.
Buckley, C. D., N. Amft, et al. (2000). "Persistent induction of the chemokine receptor CXCR4 by TGF-beta 1 on synovial T cells contributes to their accumulation within the rheumatoid synovium." J Immunol. 165(6):3423-9.
Burger, J. A., N. Tsukada, et al. (2000). "Blood-derived nurse-like cells protect chronic lymphocytic leukemia B cells from spontaneous apoptosis through stromal cell-derived factor-1." Blood 96(8):2655-63.
Burns J.M, Summers B.C, Wang Y, Melikian A, Berahovich R, Miao Z, Penfold M.E, Sunshine M.J, Littman D.R, Kuo C.J, Wei K, McMaster B.E, Wright K, Howard M.C, Schall T.J. (2006) A novel chemokine receptor for SDF-1 and I-TAC involved in cell survival, cell adhesion, and tumor development. J Exp. Med. 203(9):2201-2213.
Burt, R. K., A. Marmont, et al. (2006). "Randomized controlled trials of autologous hematopoietic stem cell transplantation for autoimmune diseases: the evolution from myeloablative to lymphoablative transplant regimens." Arthritis Rheum 54(12):3750-60.
Butler J.M, Guthrie S.M, Koc M, Afzal A, Caballero S, Brooks H.L, Mames R.N, Segal M.S, Grant M.B, Scott E.W. (2005) SDF-1 is both necessary and sufficient to promote proliferative retinopathy. J Clin. Invest 115(1):86-93.
Butler, J. M., S. M. Guthrie, et al. (2005). "SDF-1 is both necessary and sufficient to promote proliferative retinopathy." J Clin. Invest 115(1):86-93.
Cabioglu, N., A. Sahin, et al. (2005). "Chemokine receptor CXCR4 expression in breast cancer as a potential predictive marker of isolated tumor cells in bone marrow." Clin. Exp. Metastasis 22(1):39-46.
Chow F.Y, Nikolic-Paterson D.J, Ma F.Y, Ozols E, Rollins B.J, Tesch G.H. (2007) Monocyte chemoattractant protein-1-induced tissue inflammation is critical for the development of renal injury but not type 2 diabetes in obese db/db mice. Diabetologia 50(2):471-480.
Chow F.Y, Nikolic-Paterson D.J, Ozols E, Atkins R.C, Rollin B.J, Tesch G.H. (2006) Monocyte chemoattractant protein-1 promotes the development of diabetic renal injury in streptozotocin-treated mice. Kidney Int 69(1):73-80
Corcione, A., L. Ottonello, et al. (2000). "Stromal cell-derived factor-1 as a chemoattractant for follicular center lymphoma B cells." J Natl. Cancer Inst 92(8):628-35.
Croop, J. M., R. Cooper, et al. (2000). "Large-scale mobilization and isolation of CD34+ cells from normal donors." Bone Marrow Transplant 26(12):1271-9.
Crump, M. P., J. H. Gong, et al. (1997). "Solution structure and basis for functional activity of stromal cell-derived factor-1; dissociation of CXCR4 activation from binding and inhibition of HIV-1." Embo J 16(23):6996-7007.
Dale, D. C., W. C. Liles, et al. (1998). "Effects of granulocyte-macrophage colony-stimulating factor (GM-CSF) on neutrophil kinetics and function in normal human volunteers." Am J Hematol. 57(1):7-15.
Damha, M.J., Ogilvie, K.K.(1993) Methods in Molecular Biology, Vol.20 Protocols for oligonucleotides and analogs, ed. S. Agrawal, pp.81-114, Humana Press Inc. 1993.
D'Apuzzo, M., A. Rolink, et al. (1997). "The chemokine SDF-1, stromal cell-derived factor 1, attracts early stage B cell precursors via the chemokine receptor CXCR4." Eur J Immunol. 27(7):1788-93.
Dawn B, Tiwari S, Kucia M.J, Zuba-Surma E.K, Guo Y, Sanganalmath S.K, Abdel-Latif A, Hunt G, Vincent R.J, Taher H, Reed N.J, Ratajczak M.Z, Bolli R (2008) Transplantation of bone marrow-derived very small embryonic-like stem cells attenuates left ventricular dysfunction and remodeling after myocardial infarction. Stem Cells 26(6):1646-1655.
De Klerck, B., L. Geboes, et al. (2005). "Pro-inflammatory properties of stromal cell-derived factor-1 (CXCL12) in collagen-induced arthritis." Arthritis Res Ther 7(6):R1208-20.
Eaton, B. E., L. Gold, et al. (1997). "Post-SELEX combinatorial optimization of aptamers." Bioorg Med. Chem. 5(6):1087-96.
Edelman J.L, Lutz D, Castro M.R (2005). Corticosteroids inhibit VEGF-induced vascular leakage in a rabbit model of blood-retinal and blood-aqueous barrier breakdown. Exp. Eye Res 80(2):249-258.
Edelman, J. L., D. Lutz, et al. (2005). "Corticosteroids inhibit VEGF-induced vascular leakage in a rabbit model of blood-retinal and blood-aqueous barrier breakdown." Exp Eye Res 80(2):249-58.
Fedyk, E. R., D. H. Ryyan, et al. (1999). "Maturation decreases responsiveness of human bone marrow B lineage cells to stromal-derived factor 1 (SDF-1)." J Leukoc Biol. 66(4):667-73.
Fedyk, E. R., D. Jones, et al. (2001). "Expression of stromal-derived factor-1 is decreased by IL-1 and TNF and in dermal wound healing." J Immunol. 166(9):5749-54.
Fierro, F. A., S. Brenner, et al. (2008). "Combining SDF-1/CXCR4 antagonism and chemotherapy in relapsed acute myeloid leukemia." Leukemia.
Fong, D. S., L. P. Aiello, et al. (2004). "Diabetic retinopathy." Diabetes Care 27(10):2540-53.
Fransioli J, Bailey B, Gude N.A, Cottage C.T, Muraski J.A, Emmanuel G, Wu W, Alvarez R, Rubio M, Ottolenghi S, Schaefer E, Sussman M.A (2008) Evolution of the c-kit-positive cell response to pathological challenge in the myocardium. Stem Cells 26(5):1315-1324.
Friedlander M, Dorrell M.I, Ritter M.R, Marchetti V, Moreno S.K, El-Kalay M, Bird A.C, Banin E, Aguilar E (2007). Progenitor cells and retinal angiogenesis. Angiogenesis 10(2):89-101.
Garrido, S. M., F. R. Appelbaum, et al. (2001). "Acute myeloid leukemia cells are protected from spontaneous and drug-induced apoptosis by direct contact with a human bone marrow stromal cell line (HS-5)." Exp. Hematol. 29(4):448-57.
Gear, A. R., S. Suttitanamongkol, et al. (2001). "Adenosine diphosphate strongly potentiates the ability of the chemokines MDC, TARC, and SDF-1 to stimulate platelet function." Blood 97(4):937-45.
Geminder, H., O. Sagi-Assif, et al. (2001). "A possible role for CXCR4 and its ligand, the CXC chemokine stromal cell-derived factor-1, in the development of bone marrow metastases in neuroblastoma." J Immunol. 167(8):4747-57.
Glaspy, J. A., E. J. Shpall, et al. (1997). "Peripheral blood progenitor cell mobilization using stem cell factor in combination with filgrastim in breast cancer patients." Blood 90(8):2939-51.
Glaspy, J., M. W. Davis, et al. (1996). "Biology and clinical potential of stem-cell factor." Cancer Chemother Pharmacol 38 Suppl: S53-7.
Godessart, N. (2005). "Chemokine receptors: attractive targets for drug discovery." Ann N Y Acad Sci 1051:647-57.
Godot, V., M. Arock, et al. (2007). "H4 histamine receptor mediates optimal migration of mast cell precursors to CXCL12." J Allergy Clin. Immunol. 120(4):827-34.
Gombert, M., M. C. Dieu-Nosjean, et al. (2005). "CCL1-CCR8 interactions: an axis mediating the recruitment of T cells and Langerhans-type dendritic cells to sites of atopic skin inflammation." J Immunol. 174(8):5082-91.
Gonzalo, J. A., C. M. Lloyd, et al. (2000). "Critical involvement of the chemotactic axis CXCR4/stromal cell-derived factor-1 alpha in the inflammatory component of allergic airway disease." J Immunol. 165(1):499-508.
Grassi, F., S. Cristino, et al. (2004). "CXCL12 chemokine up-regulates bone resorption and MMP-9 release by human osteoclasts: CXCL12 levels are increased in synovial and bone tissue of rheumatoid arthritis patients." J Cell Physiol. 199(2):244-51.
Gratwohl A, Baldomero H, Horisberger B, Schmid C, Passweg J, Urbano-Ispizua A (2002). Current trends in hematopoietic stem cell transplantation in Europe. Blood 100(7):2374-2386.
Grunewald, M., I. Avraham, et al. (2006). "VEGF-induced adult neovascularization: recruitment, retention, and role of accessory cells." Cell 124(1):175-89.
Gu, J., E. Marker-Hermann, et al. (2002). "A 588-gene microarray analysis of the peripheral blood mononuclear cells of spondyloarthropathy patients." Rheumatology (Oxford) 41(7):759-66.
Guleng, B., K. Tateishi, et al. (2005). "Blockade of the stromal cell-derived factor-1/CXCR4 axis attenuates in vivo tumor growth by inhibiting angiogenesis in a vascular endothelial growth factor-independent manner." Cancer Res 65(13):5864-71.
Gulino, A. V., D. Moratto, et al. (2004). "Altered leukocyte response to CXCL12 in patients with warts hypogammaglobulinemia, infections, myelokathexis (WHIM) syndrome." Blood 104(2):444-52.
Hartmann, T. N., M. Burger, et al. (2004). "The role of adhesion molecules and chemokine receptor CXCR4 (CD184) in small cell lung cancer." J Biol Regul Homeost Agents 18(2):126-30.
Higgins, R. D., K. Yu, et al. (1999). "Diltiazem reduces retinal neovascularization in a mouse model of oxygen induced retinopathy." Curr Eye Res 18(1):20-7.
Hodohara, K., N. Fujii, et al. (2000). "Stromal cell-derived factor-1 (SDF-1) acts together with thrombopoietin to enhance the development of megakaryocytic progenitor cells (CFU-MK)." Blood 95(3):769-75.
Hoshino, M., N. Aoike, et al. (2003). "Increased immunoreactivity of stromal cell-derived factor-1 and angiogenesis in asthma." Eur Respir J 21(5):804-9.
Hwang, J. H., H. K. Chung, et al. (2003). "CXC chemokine receptor 4 expression and function in human anaplastic thyroid cancer cells." J Clin. Endocrinol. Metab 88(1):408-16.
Ito T, Suzuki A, Imai E, Okabe M, Hori M (2001). Bone marrow is a reservoir of repopulating mesangial cells during glomerular remodeling. J Am Soc Nephrol. 12(12):2625-2635.
Iwamoto T, Okamoto H, Toyama Y, Momohara S (2008) Molecular aspects of rheumatoid arthritis: chemokines in the joints of patients. FEBS J.
Jackson, K. A., S. M. Majka, et al. (2001). "Regeneration of ischemic cardiac muscle and vascular endothelium by adult stem cells." J Clin. Invest 107(11):1395-402.
Jiang, W., P. Zhou, et al. (1994). "Molecular cloning of TPAR1, a gene whose expression is repressed by the tumor promoter 12-O-tetradecanoylphorbol 13-acetate (TPA)." Exp Cell Res 215(2):284-93.
Jin, L., K. J. Hope, et al. (2006). "Targeting of CD44 eradicates human acute myeloid leukemic stem cells." Nat Med 12(10):1167-74.
Jin, L., Y. Tabe, et al. (2008). "CXCR4 up-regulation by imatinib induces chronic myelogenous leukemia (CML) cell migration to bone marrow stroma and promotes survival of quiescent CML cells." Mol. Cancer Ther 7(1):48-58.
Jo, D. Y., S. Rafii, et al. (2000). "Chemotaxis of primitive hematopoietic cells in response to stromal cell-derived factor-1." J Clin. Invest 105(1):101-11.
Jose, P. J., D. A. Griffiths-Johnson, et al. (1994). "Eotaxin: a potent eosinophil chemoattractant cytokine detected in a guinea pig model of allergic airways inflammation." J Exp. Med 179(3):881-7.
Juarez, J. and L. Bendall (2004). "SDF-1 and CXCR4 in normal and malignant hematopoiesis." Histol Histopathol 19(1):299-309.
Kanbe, K., K. Takagishi, et al. (2002). "Stimulation of matrix metalloprotease 3 release from human chondrocytes by the interaction of stromal cell-derived factor 1 and CXC chemokine receptor 4." Arthritis Rheum 46(1):130-7.
Kang, H., G. Watkins, et al. (2005). "Stromal cell derived factor-1: its influence on invasiveness and migration of breast cancer cells in vitro, and its association with prognosis and survival in human breast cancer." Breast Cancer Res 7(4):R402-10.
Kawai, T., U. Choi, et al. (2005). "Enhanced function with decreased internalization of carboxy-terminus truncated CXCR4 responsible for WHIM syndrome." Exp. Hematol. 33(4):460-8.
Kim K.W, Cho M.L, Kim H.R, Ju J.H, Park M.K, Oh H.J, Kim J.S, Park S.H, Lee S.H, Kim H.Y (2007). Up-regulation of stromal cell-derived factor 1 (CXCL12) production in rheumatoid synovial fibroblasts through interactions with T lymphocytes: role of interleukin-17 and CD40L-CD40 interaction. Arthritis Rheum 56(4):1076-1086.
King, A. G., D. Horowitz, et al. (2001). "Rapid mobilization of murine hematopoietic stem cells with enhanced engraftment properties and evaluation of hematopoietic progenitor cell mobilization in rhesus monkeys by a single injection of SB-251353, a specific truncated form of the human CXC chemokine GRObeta." Blood 97(6):1534-42.
Kocher, A. A., M. D. Schuster, et al. (2001). "Neovascularization of ischemic myocardium by human bone-marrow-derived angioblasts prevents cardiomyocyte apoptosis, reduces remodeling and improves cardiac function." Nat Med. 7(4):430-6.
Koshiba, T., R. Hosotani, et al. (2000). "Expression of stromal cell-derived factor 1 and CXCR4 ligand receptor system in pancreatic cancer: a possible role for tumor progression." Clin Cancer Res 6(9):3530-5.
Krumbholz, M., D. Theil, et al. (2006). "Chemokines in multiple sclerosis: CXCL12 and CXCL13 up-regulation is differentially linked to CNS immune cell recruitment." Brain 129:200-211.
Kryczek, I., A. Lange, et al. (2005). "CXCL12 and vascular endothelial growth factor synergistically induce neoangiogenesis in human ovarian cancers." Cancer Res 65(2):465-72.
Kucia, M., R. Reca, et al. (2005). "Trafficking of normal stem cells and metastasis of cancer stem cells involve similar mechanisms: pivotal role of the SDF-1-CXCR4 axis." Stem Cells 23(7):879-94.
Kulkarni O, Pawar R.D, Purschke W, Eulberg D, Selve N, Buchner K, Ninichuk V, Segerer S, Vielhauer V, Klussmann S, Anders H.J. (2007) Spiegelmer inhibition of CCL2/MCP-1 ameliorates lupus nephritis in MRL-(Fas)lpr mice. J Am Soc Nephrol. 18(8):2350-2358.
Kusser, W. (2000). "Chemically modified nucleic acid aptamers for in vitro selections: evolving evolution." J Biotechnol 74(1):27-38.
Lagane, B., K. Y. Chow, et al. (2008). "CXCR4 dimerization and {beta}-arrestin-mediated signaling account for the enhanced chemotaxis to CXCL12 in WHIM syndrome." Blood.
Lapteva, N., A. G. Yang, et al. (2005). "CXCR4 knockdown by small interfering RNA abrogates breast tumor growth in vivo." Cancer Gene Ther 12(1):84-9.
Li J.K, Yu L, Shen Y, Zhou L.S, Wang Y.C, Zhang J.H (2008). Inhibition of CXCR4 activity with AMD3100 decreases invasion of human colorectal cancer cells in vitro. World J Gastroenterol. 14(15):2308-2313
Li Y, Atmaca-Sonmez P, Schanie C.L, Ildstad S.T, Kaplan H.J, Enzmann V (2007) Endogenous bone marrow derived cells express retinal pigment epithelium cell markers and migrate to focal areas of RPE damage. Invest Ophthalmol. Vis Sci 48(9):4321-4327.
Li Y, Reca R.G, Atmaca-Sonmez P, Ratajczak M.Z, Ildstad S.T, Kaplan H.J, Enzmann V (2006). Retinal pigment epithelium damage enhances expression of chemoattractants and migration of bone marrow-derived stem cells. Invest Ophthalmol Vis Sci 47(4):1646-1652.
Liang Z, Brooks J, Willard M, Liang K, Yoon Y, Kang S, Shim H (2007) CXCR4/CXCL12 axis promotes VEGF-mediated tumor angiogenesis through Akt signaling pathway. Biochem Biophys Res Commun 359(3):716-722.
Lima e Silva R, Shen J, Hackett S.F, Kachi S, Akiyama H, Kiuchi K, Yokoi K, Hatara M.C, Lauer T, Aslam S, Gong Y.Y, Xiao W.H, Khu N.H, Thut C, Campochiaro P.A (2007) The SDF-1/CXCR4 ligand/receptor pair is an important contributor to several types of ocular neovascularization. FASEB J 21(12):3219-3230.
Ma, Q., D. Jones, et al. (1998). "Impaired B-lymphopoiesis, myelopoiesis, and derailed cerebellar neuron migration in CXCR4- and SDF-1-deficient mice." Proc. Natl. Acad Sci U S A 95(16):9448-53.
Maekawa, T. and T. Ishii (2000). "Chemokine/receptor dynamics in the regulation of hematopoiesis." Intern Med 39(2):90-100.
Majka M, Kucia M, Ratajczak M.Z. (2005) Stem cell biology - a never ending quest for understanding. Acta Biochim Pol. 52(2):353-358.
Majka, M., A. Janowska-Wieczorek, et al. (2000). "Stromal-derived factor 1 and thrombopoietin regulate distinct aspects of human megakaryopoiesis." Blood 96(13):4142-51.
Mames R.N, Mattheus A, Butler J, Brown G, Jorgensen M, Scott E (2006). New anti SDF-1 antinody prevents retinal neovascularization in primates. ARVO; May 1, 2006; Fort Lauderdale.
Marechal, V., F. Arenzana-Seisdedos, et al. (1999). "Opposite effects of SDF-1 on human immunodeficiency virus type 1 replication." J Virol. 73(5):3608-15.
Matsunaga, T., N. Takemoto, et al. (2003). "Interaction between leukemic-cell VLA-4 and stromal fibronectin is a decisive factor for minimal residual disease of acute myelogenous leukemia." Nat Med 9(9):1158-65.
Matthys, P., S. Hatse, et al. (2001). "AMD3100, a potent and specific antagonist of the stromal cell-derived factor-1 chemokine receptor CXCR4, inhibits autoimmune joint inflammation in IFN-gamma receptor-deficient mice." J Immunol. 167(8):4686-92.
McGinnis S, Madden T.L (2004). BLAST: at the core of a powerful and diverse set of sequence analysis tools. Nucleic Acids Res. 32(Web Server issue):W20-5.
Meleth, A. D., E. Agron, et al. (2005). "Serum inflammatory markers in diabetic retinopathy." Invest Ophthalmol. Vis Sci 46(11):4295-301.
Menu, E., K. Asosingh, et al. (2006). "The involvement of stromal derived factor 1alpha in homing and progression of multiple myeloma in the 5TMM model." Haematologica.
Miller, M. D. and M. S. Krangel (1992). "Biology and biochemistry of the chemokines: a family of chemotactic and inflammatory cytokines." Crit Rev Immunol. 12(1-2):17-46.
Minges Wols H.A, Ippolito J.A, Yu Z, Palmer J.L, White.A, Le P.T, Witte P.L. (2007). The effects of microenvironment and internal programming on plasma cell survival. Int Immunol. 19(7):837-846.
Minges Wols H.A, Witte P.L. (2008). Plasma cell purification from murine bone marrow using a two-step isolation approach. J Immunol Methods 329(1-2):219-224
Moser, B., M. Wolf, et al. (2004). "Chemokines: multiple levels of leukocyte migration control." Trends Immunol. 25(2):75-84.
Mudry, R. E., J. E. Fortney, et al. (2000). "Stromal cells regulate survival of B-lineage leukemic cells during chemotherapy." Blood 96(5):1926-32.
Muller, A., B. Homey, et al. (2001). "Involvement of chemokine receptors in breast cancer metastasis." Nature 410(6824):50-6.
Murdoch, C. (2000). "CXCR4: chemokine receptor extraordinaire." Immunol Rev 177:175-84.
Murphy, P. M., M. Baggiolini, et al. (2000). "International union of pharmacology. XXII. Nomenclature for chemokine receptors." Pharmacol. Rev 52(1):145-76.
Nagasawa, T. (2000). "A chemokine, SDF-1/PBSF, and its receptor, CXC chemokine receptor 4, as mediators of hematopoiesis." Int J Hematol. 72(4):408-11.
Nagasawa, T., H. Kikutani, et al. (1994). "Molecular cloning and structure of a pre-B-cell growth-stimulating factor." Proc. Natl. Acad Sci U S A 91(6):2305-9.
Nagasawa, T., S. Hirota, et al. (1996). "Defects of B-cell lymphopoiesis and bone-marrow myelopoiesis in mice lacking the CXC chemokine PBSF/SDF-1." Nature 382(6592):635-8.
NCI NCI. (2001) Bone Marrow Transplantation and Peripheral Blood Stem Cell Transplantation: Questions and Answers. Fact Sheet, Vol.2008.
Needleman & Wunsch (1970), A general method applicable to the search for similarities in the amino acid sequence of two proteins. J Mol. Biol. 48(3):443-53.
Ninichuk, V., S. Clauss, et al. (2008). "Late onset of Ccl2 blockade with the Spiegelmer mNOX-E36-3'PEG prevents glomerulosclerosis and improves glomerular filtration rate in db/db mice." Am J Pathol. 172(3):628-37.
Oppenheim, J. J., C. O. Zachariae, et al. (1991). "Properties of the novel proinflammatory supergene "intercrine" cytokine family." Annu Rev Immunol. 9:617-48.
Orimo, A., P. B. Gupta, et al. (2005). "Stromal fibroblasts present in invasive human breast carcinomas promote tumor growth and angiogenesis through elevated SDF-1/CXCL12 secretion." Cell 121(3):335-48.
Otani A, Kinder K, Ewalt K, Otero FJ, Schimmel P, Friedlander M (2002) Bone marrow-derived stem cells target retinal astrocytes and can promote or inhibit retinal angiogenesis. Nat Med 8(9):1004-1010.
Parretta, E., G. Cassese, et al. (2005). "CD8 cell division maintaining cytotoxic memory occurs predominantly in the bone marrow." J Immunol. 174(12):7654-64.
Pearson & Lipman (1988), Improved tools for biological sequence comparison. Proc. Nat'l. Acad. Sci. USA 85:2444.
Peled, A., I. Petit, et al. (1999). "Dependence of human stem cell engraftment and repopulation of NOD/SCID mice on CXCR4." Science 283(5403):845-8.
Perissinotto, E., G. Cavalloni, et al. (2005). "Involvement of chemokine receptor 4/stromal cell-derived factor 1 system during osteosarcoma tumor progression." Clin Cancer Res 11(2 Pt 1):490-7.
Phillips, R. J., M. D. Burdick, et al. (2003). "The stromal derived factor-1/CXCL12-CXC chemokine receptor 4 biological axis in non-small cell lung cancer metastases." Am J Respir Crit Care Med 167(12):1676-86.
Ponath, P. D., S. Qin, et al. (1996). "Cloning of the human eosinophil chemoattractant, eotaxin. Expression, receptor binding, and functional properties suggest a mechanism for the selective recruitment of eosinophils." J Clin. Invest 97(3):604-12.
Ponomaryov, T., A. Peled, et al. (2000). "Induction of the chemokine stromal-derived factor-1 following DNA damage improves human stem cell function." J Clin. Invest 106(11):1331-9.
Pruijt, J. F., R. Willemze, et al. (1999). "Mechanisms underlying hematopoietic stem cell mobilization induced by the CXC chemokine interleukin-8." Curr Opin. Hematol. 6(3):152-8.
Radbruch, A., G. Muehlinghaus, et al. (2006). "Competence and competition: the challenge of becoming a long-lived plasma cell." Nat Rev Immunol 6(10):741-50.
Reddy, K., Z. Zhou, et al. (2008). "Stromal cell-derived factor-1 stimulates vasculogenesis and enhances Ewing's sarcoma tumor growth in the absence of vascular endothelial growth factor." Int J Cancer.
Riviere, C., F. Subra, et al. (1999). "Phenotypic and functional evidence for the expression of CXCR4 receptor during megakaryocytopoiesis." Blood 93(5):1511-23.
Rosenfeld, C. S., B. Bolwell, et al. (1996). "Comparison of four cytokine regimens for mobilization of peripheral blood stem cells: IL-3 alone and combined with GM-CSF or G-CSF." Bone Marrow Transplant 17(2):179-83.
Rubin, J. B., A. L. Kung, et al. (2003). "A small-molecule antagonist of CXCR4 inhibits intracranial growth of primary brain tumors." Proc Natl Acad Sci U S A 100(23):13513-8.
Salcedo R, Wasserman K, Young H.A, Grimm M.C, Howard O.M, Anver M.R, Kleinman H.K, Murphy W.J, Oppenheim J.J. (1999) Vascular endothelial growth factor and basic fibroblast growth factor induce expression of CXCR4 on human endothelial cells: In vivo neovascularization induced by stromal-derived factor-1alpha. Am J Pathol 154(4):1125-1135.
Salcedo, R. and J. J. Oppenheim (2003). "Role of chemokines in angiogenesis: CXCL12/SDF-1 and CXCR4 interaction, a key regulator of endothelial cell responses." Microcirculation 10(3-4):359-70.
Salcedo, R., K. Wasserman, et al. (1999). "Vascular endothelial growth factor and basic fibroblast growth factor induce expression of CXCR4 on human endothelial cells: In vivo neovascularization induced by stromal-derived factor-1alpha." Am J Pathol 154(4):1125-35.
Salvucci, O., L. Yao, et al. (2002). "Regulation of endothelial cell branching morphogenesis by endogenous chemokine stromal-derived factor-1." Blood 99(8):2703-11.
Saur, D., B. Seidler, et al. (2005). "CXCR4 expression increases liver and lung metastasis in a mouse model of pancreatic cancer." Gastroenterology 129(4):1237-50.
Schall, T. J. and K. B. Bacon (1994). "Chemokines, leukocyte trafficking, and inflammation." Curr Opin. Immunol. 6(6):865-73.
Schober, A. and A. Zernecke (2007). "Chemokines in vascular remodeling." Thromb Haemost 97(5):730-7.
Schwarting S, Litwak S, Hao W, Bahr M, Weise J, Neumann H (2008) Hematopoietic Stem Cells Reduce Postischemic Inflammation and Ameliorate Ischemic Brain Injury. Stroke.
Scotton C.J, Wilson J.L, Scott K, Stamp G, Wilbanks G.D, Fricker S, Bridger G, Balkwill F.R. (2002) Multiple actions of the chemokine CXCL12 on epithelial tumor cells in human ovarian cancer. Cancer Res 62(20):5930-5938.
Scotton, C. J., J. L. Wilson, et al. (2002). "Multiple actions of the chemokine CXCL12 on epithelial tumor cells in human ovarian cancer." Cancer Res 62(20):5930-8.
Sengupta, N., S. Caballero, et al. (2005). "Preventing stem cell incorporation into choroidal neovascularization by targeting homing and attachment factors." Invest Ophthalmol Vis Sci 46(1):343-8.
Shirozu, M., T. Nakano, et al. (1995). "Structure and chromosomal localization of the human stromal cell-derived factor 1 (SDF1) gene." Genomics 28(3):495-500.
Smith & Waterman (1981), Adv. Appl. Math. 2:482.
Smith, L. E., E. Wesolowski, et al. (1994). "Oxygen-induced retinopathy in the mouse." Invest Ophthalmol Vis Sci 35(1):101-11.
Soriano, A., C. Martinez, et al. (2002). "Plasma stromal cell-derived factor (SDF)-1 levels, SDF1-3'A genotype, and expression of CXCR4 on T lymphocytes: their impact on resistance to human immunodeficiency virus type 1 infection and its progression." J Infect Dis 186(7):922-31.
Springer, T. A. (1995). "Traffic signals on endothelium for lymphocyte recirculation and leukocyte emigration." Annu Rev Physiol 57:827-72.
Stem Cells: Scientific Progress and Future Research Directions. Department of Health and Human Services. June 2001. <http://stemcells.nih.gov/info/scireport/2001report>.
Sun, Y. X., A. Schneider, et al. (2005). "Skeletal localization and neutralization of the SDF-1(CXCL12)/CXCR4 axis blocks prostate cancer metastasis and growth in osseous sites in vivo." J Bone Miner Res 20(2):318-29.
Tabe, Y., L. Jin, et al. (2007). "Activation of integrin-linked kinase is a critical prosurvival pathway induced in leukemic cells by bone marrow-derived stromal cells." Cancer Res 67(2):684-94.
Tachibana, K., S. Hirota, et al. (1998). "The chemokine receptor CXCR4 is essential for vascularization of the gastrointestinal tract." Nature 393(6685):591-4.
Takenaga, M., H. Tamamura, et al. (2004). "A single treatment with microcapsules containing a CXCR4 antagonist suppresses pulmonary metastasis of murine melanoma." Biochem Biophys Res Commun 320(1):226-32.
Tamamura, H., M. Fujisawa, et al. (2004). "Identification of a CXCR4 antagonist, a T140 analog, as an anti-rheumatoid arthritis agent." FEBS Lett 569(1-3):99-104.
Tang Y, Yasuhara T, Hara K, Matsukawa N, Maki M, Yu G, Xu L, Hess DC, Borlongan CV (2007) Transplantation of bone marrow-derived stem cells: a promising therapy for stroke. Cell Transplant 16(2):159-169.
Tarlinton D, Radbruch A, Hiepe F, Dorner T (2008) Plasma cell differentiation and survival. Curr Opin Immunol 20(2):162-169.
Tashiro, K., H. Tada, et al. (1993). "Signal sequence trap: a cloning strategy for secreted proteins and type I membrane proteins." Science 261(5121):600-3.
Vadhan-Raj, S., L. J. Murray, et al. (1997). "Stimulation of megakaryocyte and platelet production by a single dose of recombinant human thrombopoietin in patients with cancer." Ann Intern Med 126(9):673-81.
Venkatesan, N., S. J. Kim, et al. (2003). "Novel phosphoramidite building blocks in synthesis and applications toward modified oligonucleotides." Curr Med Chem 10(19):1973-91.
Viardot, A., R. Kronenwett, et al. (1998). "The human immunodeficiency virus (HIV)-type 1 coreceptor CXCR-4 (fusin) is preferentially expressed on the more immature CD34+ hematopoietic stem cells." Ann Hematol. 77(5):193-7.
Voermans, C., M. L. Kooi, et al. (2001). "In vitro migratory capacity of CD34+ cells is related to hematopoietic recovery after autologous stem cell transplantation." Blood 97(3):799-804.
Wang J, Shiozawa Y, Wang Y, Jung Y, Pienta K.J, Mehra R, Loberg R, Taichman R.S (2008) The role of CXCR7/RDC1 as a chemokine receptor for CXCL12/SDF-1 in prostate cancer. J Biol. Chem. 283(7):4283-4294.
Wang, J., E. Guan, et al. (2001). "Role of tyrosine phosphorylation in ligand-independent sequestration of CXCR4 in human primary monocytes-macrophages." J Biol. Chem. 276(52):49236-43.
Wang, N., Q. L. Wu, et al. (2005). "Expression of chemokine receptor CXCR4 in nasopharyngeal carcinoma: pattern of expression and correlation with clinical outcome." J Transl. Med 3:26.
Wijdenes J, Vooijs W.C, Clement C, Post J, Morard F, Vita N, Laurent P, Sun R.X, Klein B, Dore J.M. (1996) A plasmocyte selective monoclonal antibody (B-B4) recognizes syndecan-1. Br J Haematol. 94(2):318-323
Wincott F, DiRenzo A, Shaffer C, Grimm S, Tracz D, Workman C, Sweedler D, Gonzalez C, Scaringe S, Usman N. (1995). Synthesis, deprotection, analysis and purification of RNA and ribozymes. Nucleic Acids Res. 23(14):2677-84.
Xu, J., A. Mora, et al. (2007). "Role of the SDF-1/CXCR4 axis in the pathogenesis of lung injury and fibrosis." Am J Respir Cell Mol. Biol. 37(3):291-9.
Yamaguchi, J., K. F. Kusano, et al. (2003). "Stromal cell-derived factor-1 effects on ex vivo expanded endothelial progenitor cell recruitment for ischemic neovascularization." Circulation 107(9):1322-8.
Yamaji-Kegan, K., Q. Su, et al. (2006). "Hypoxia-induced mitogenic factor has proangiogenic and proinflammatory effects in the lung via VEGF and VEGF receptor-2." Am J Physiol Lung Cell Mol. Physiol. 291(6):L1159-68.
Yang J, Zhang B, Lin Y, Yang Y, Liu X, Lu F. (2008) Breast cancer metastasis suppressor 1 inhibits SDF-1alpha-induced migration of non-small cell lung cancer by decreasing CXCR4 expression. Cancer Lett.
Yasumoto, K., K. Koizumi, et al. (2006). "Role of the CXCL12/CXCR4 axis in peritoneal carcinomatosis of gastric cancer." Cancer Res 66(4):2181-7.
Zagzag D, Esencay M, Mendez O, Yee H, Smirnova I, Huang Y, Chiriboga L, Lukyanov E, Liu M, Newcomb E.W. (2008) Hypoxia- and vascular endothelial growth factor-induced stromal cell-derived factor-1alpha/CXCR4 expression in glioblastomas: one plausible explanation of Scherer's structures. Am J Pathol 173(2):545-560.
Zeelenberg, I. S., L. Ruuls-Van Stalle, et al. (2001). "Retention of CXCR4 in the endoplasmic reticulum blocks dissemination of a T cell hybridoma." J Clin. Invest 108(2):269-77.
Zeelenberg, I. S., L. Ruuls-Van Stalle, et al. (2003). "The chemokine receptor CXCR4 is required for outgrowth of colon carcinoma micrometastases." Cancer Res 63(13):3833-9.
Zhang, X., T. Nakajima, et al. (2005). "Tissue trafficking patterns of effector memory CD4+ T cells in rheumatoid arthritis." Arthritis Rheum 52(12):3839-49.
Zhou, Y., P. H. Larsen, et al. (2002). "CXCR4 is a major chemokine receptor on glioma cells and mediates their survival." J Biol. Chem. 277(51):49481-7.
Zou, Y. R., A. H. Kottmann, et al. (1998). "Function of the chemokine receptor CXCR4 in haematopoiesis and in cerebellar development." Nature 393(6685):595-9.
Отличительные признаки настоящего изобретения, раскрытого в данном описании, пункты формулы изобретения и/или рисунки, как отдельно, так и в любом их сочетании, могут быть материалами для реализации данного изобретения в его различных формах.
| название | год | авторы | номер документа |
|---|---|---|---|
| SDF-1-СВЯЗЫВАЮЩИЕ НУКЛЕИНОВЫЕ КИСЛОТЫ | 2007 |
|
RU2590709C2 |
| СВЯЗЫВАЮЩИЕСЯ С SDF1 НУКЛЕИНОВЫЕ КИСЛОТЫ И ИХ ПРИМЕНЕНИЕ ПРИ ЛЕЧЕНИИ РАКА | 2011 |
|
RU2679495C2 |
| СВЯЗЫВАЮЩАЯ МСР-1 НУКЛЕИНОВАЯ КИСЛОТА И ЕЕ ПРИМЕНЕНИЕ | 2008 |
|
RU2542973C2 |
| НОВЫЕ СВЯЗЫВАЮЩИЕ C5a НУКЛЕИНОВЫЕ КИСЛОТЫ | 2013 |
|
RU2645261C2 |
| CXCL8-СВЯЗЫВАЮЩИЕ НУКЛЕИНОВЫЕ КИСЛОТЫ | 2019 |
|
RU2815444C2 |
| ГУМАНИЗИРОВАННЫЕ АНТИТЕЛА К CXCR4 ДЛЯ ЛЕЧЕНИЯ РАКА | 2011 |
|
RU2595394C2 |
| ПРИМЕНЕНИЕ АНТИТЕЛА I-3859 ДЛЯ ВЫЯВЛЕНИЯ И ДИАГНОСТИКИ РАКА | 2012 |
|
RU2636032C2 |
| АНТИТЕЛА К CXCR4 И ИХ ПРИМЕНЕНИЕ ДЛЯ ЛЕЧЕНИЯ РАКА | 2009 |
|
RU2573897C2 |
| АНТИТЕЛА ПРОТИВ CXCR4 ДЛЯ ЛЕЧЕНИЯ ИНФЕКЦИИ ВИЧ | 2010 |
|
RU2573902C2 |
| АПТАМЕР ПРОТИВ МИДКИНА И ЕГО ПРИМЕНЕНИЕ | 2007 |
|
RU2460794C2 |



































































































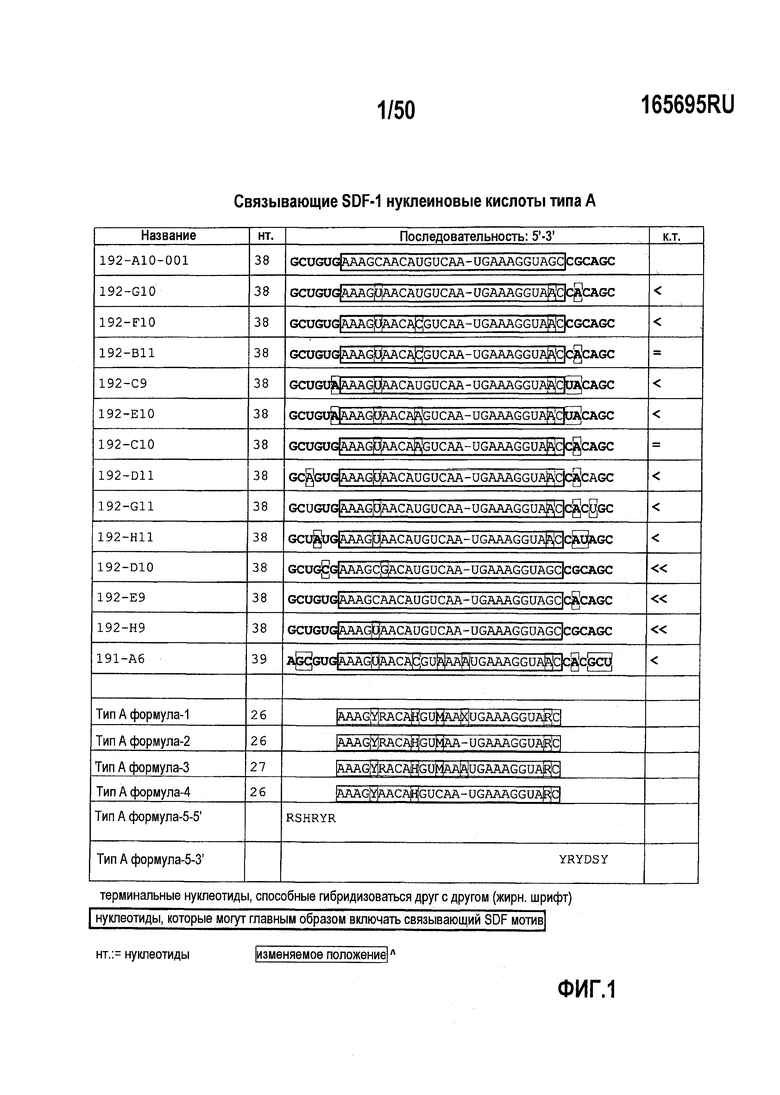




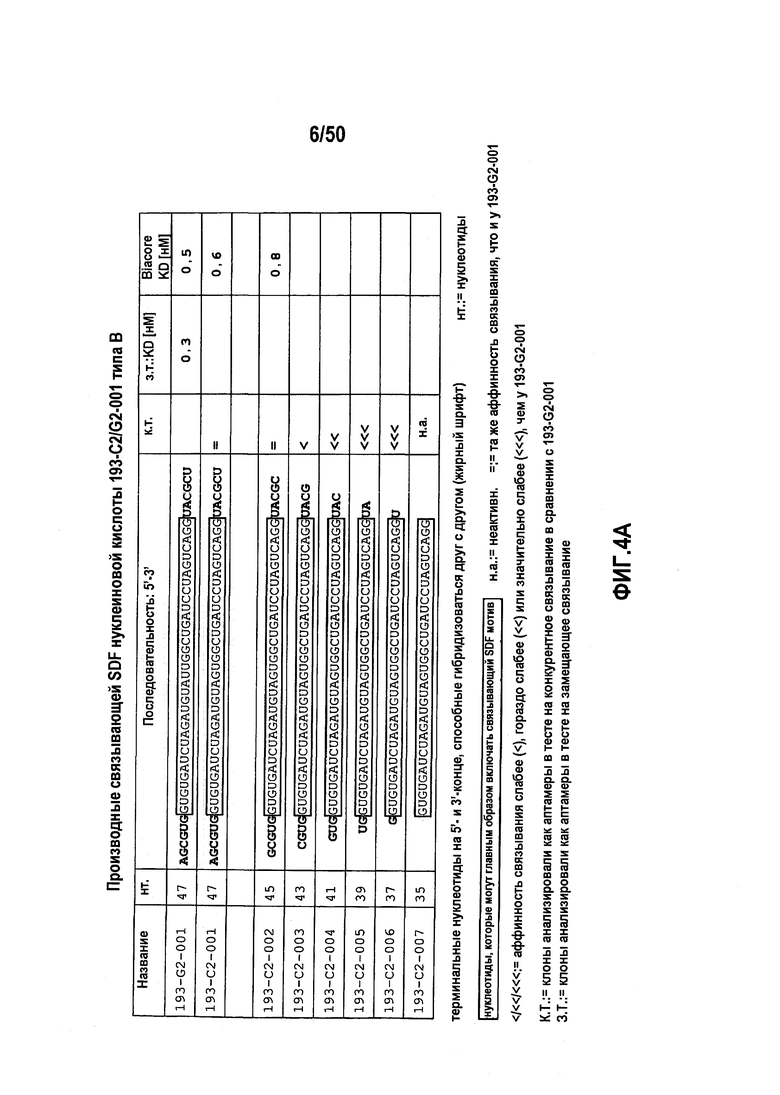





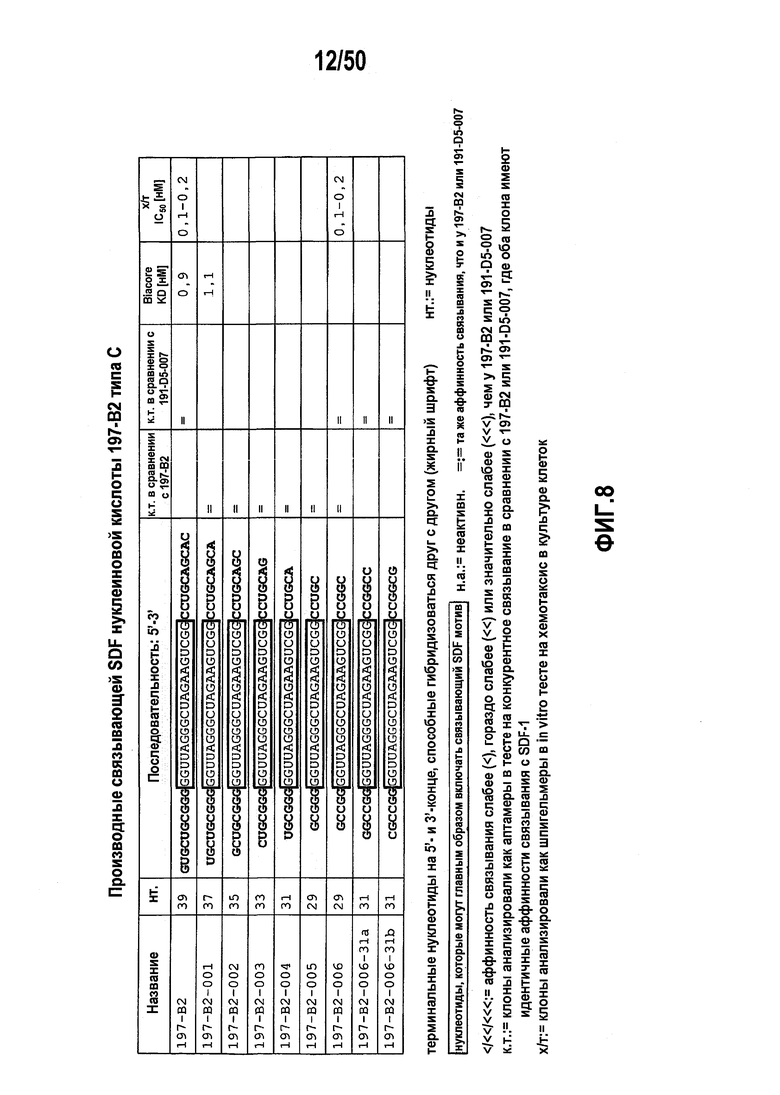


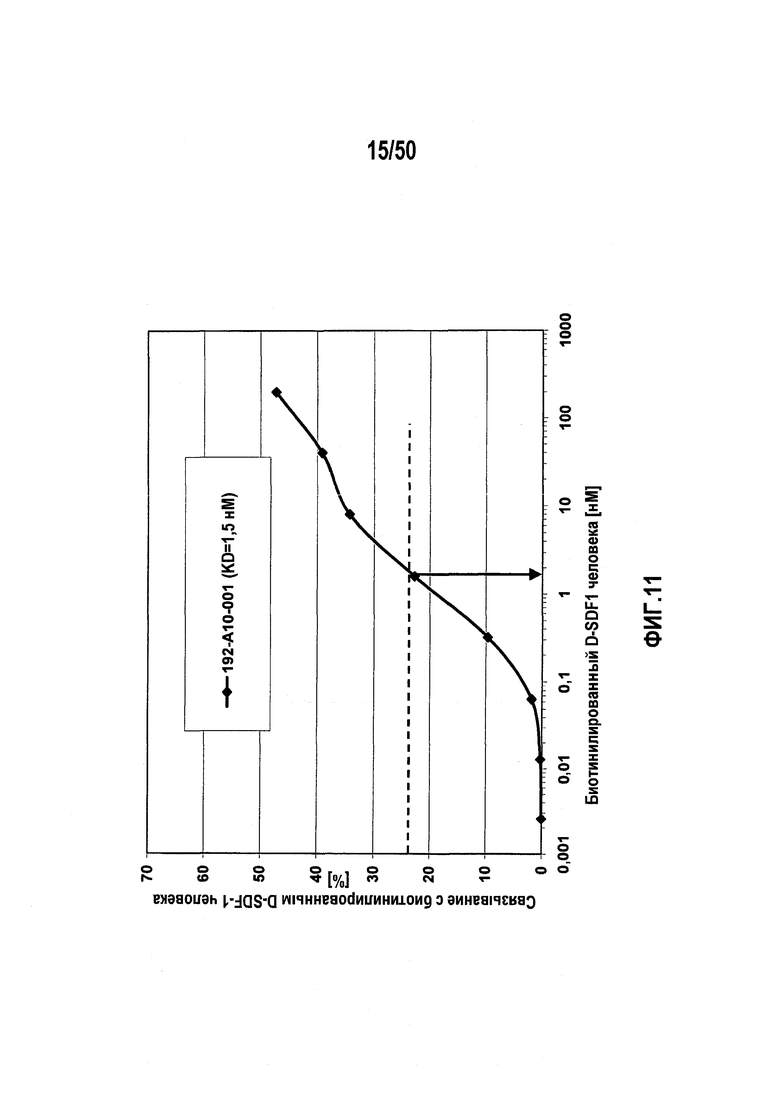

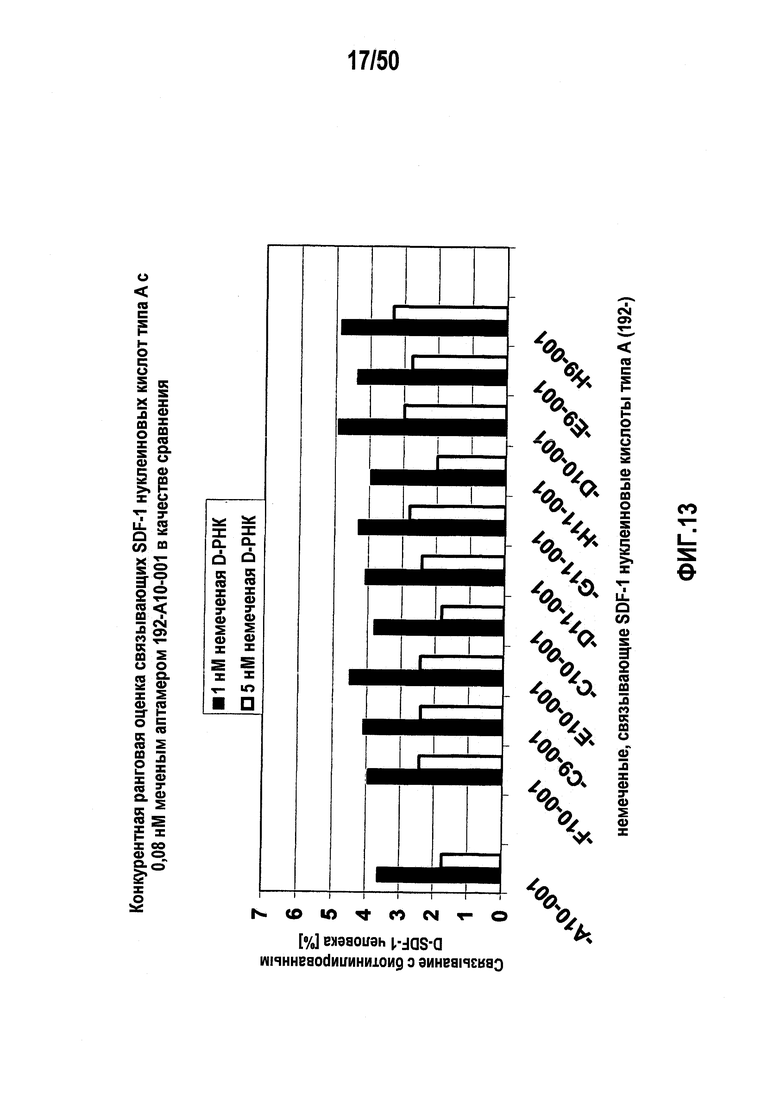
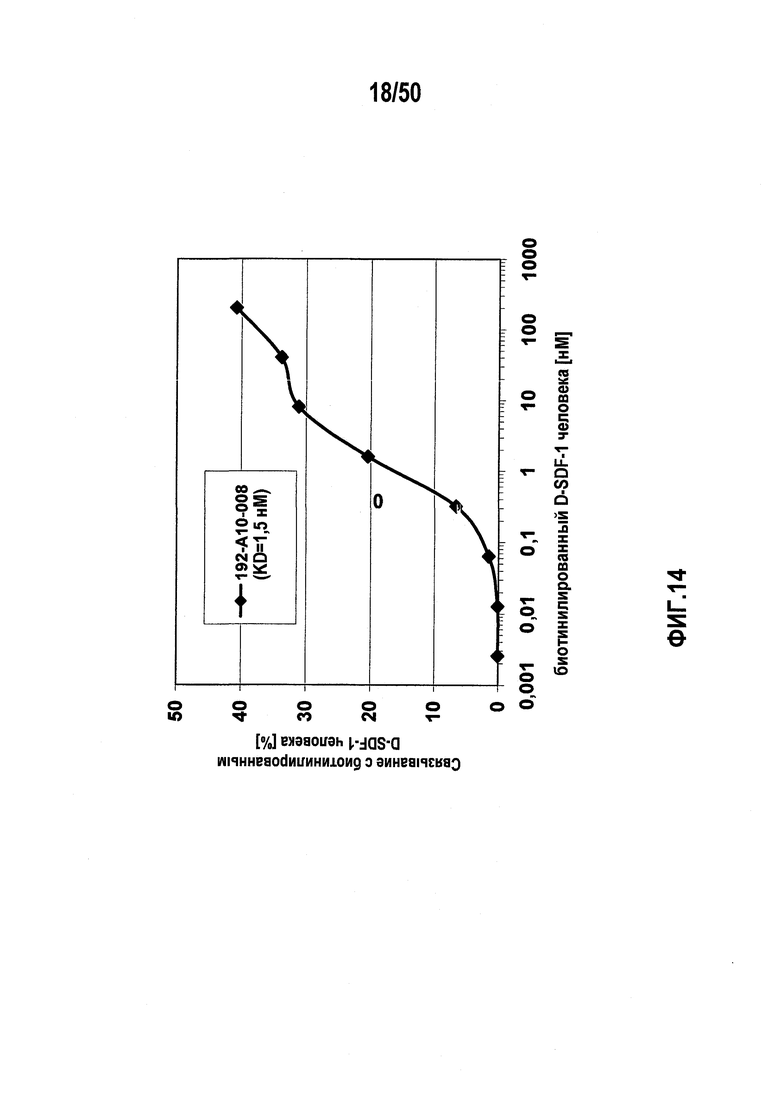
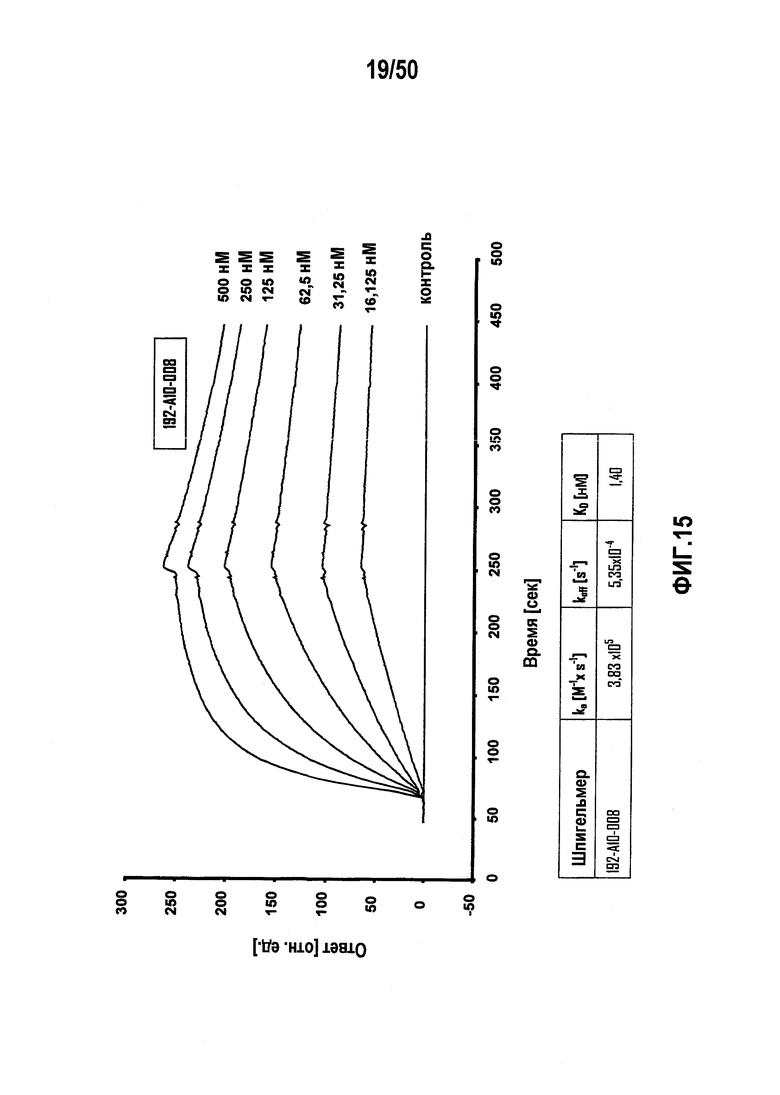

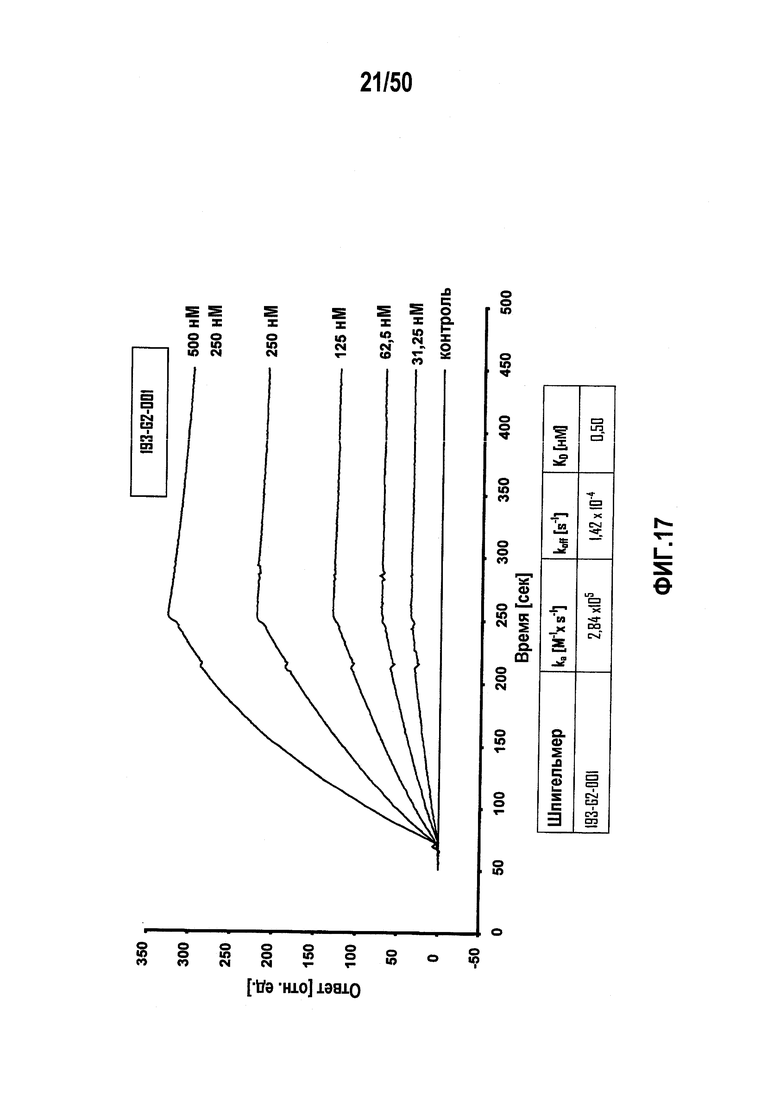
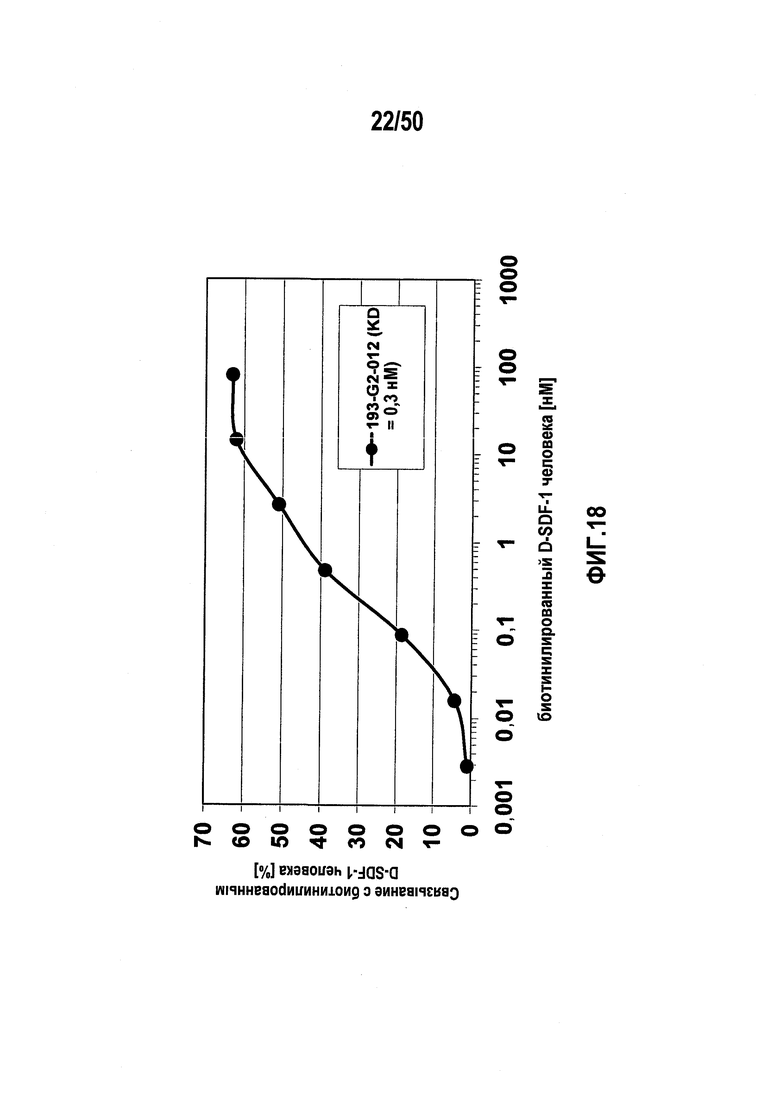


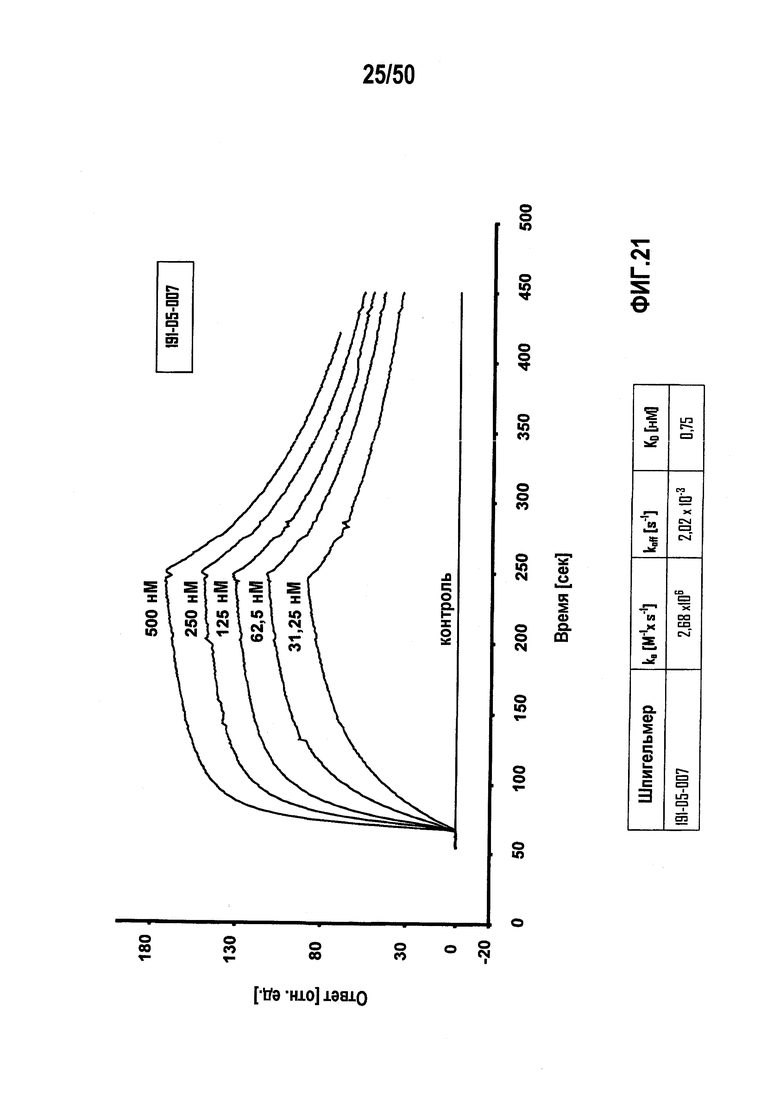


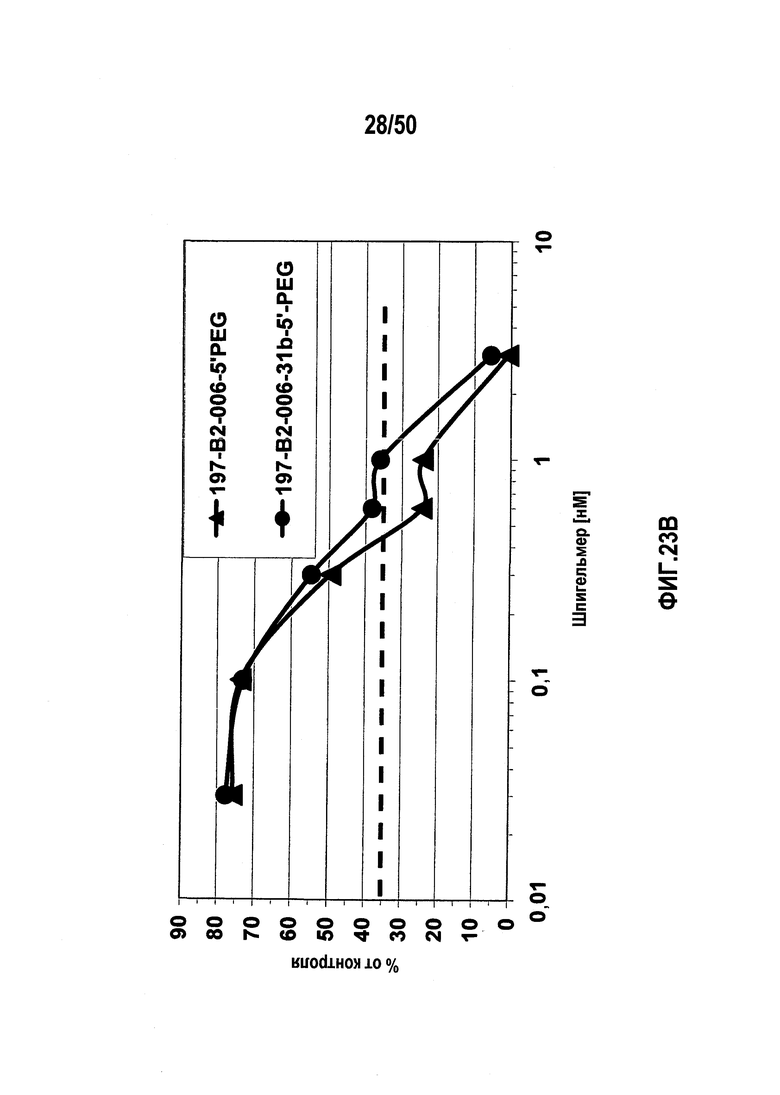


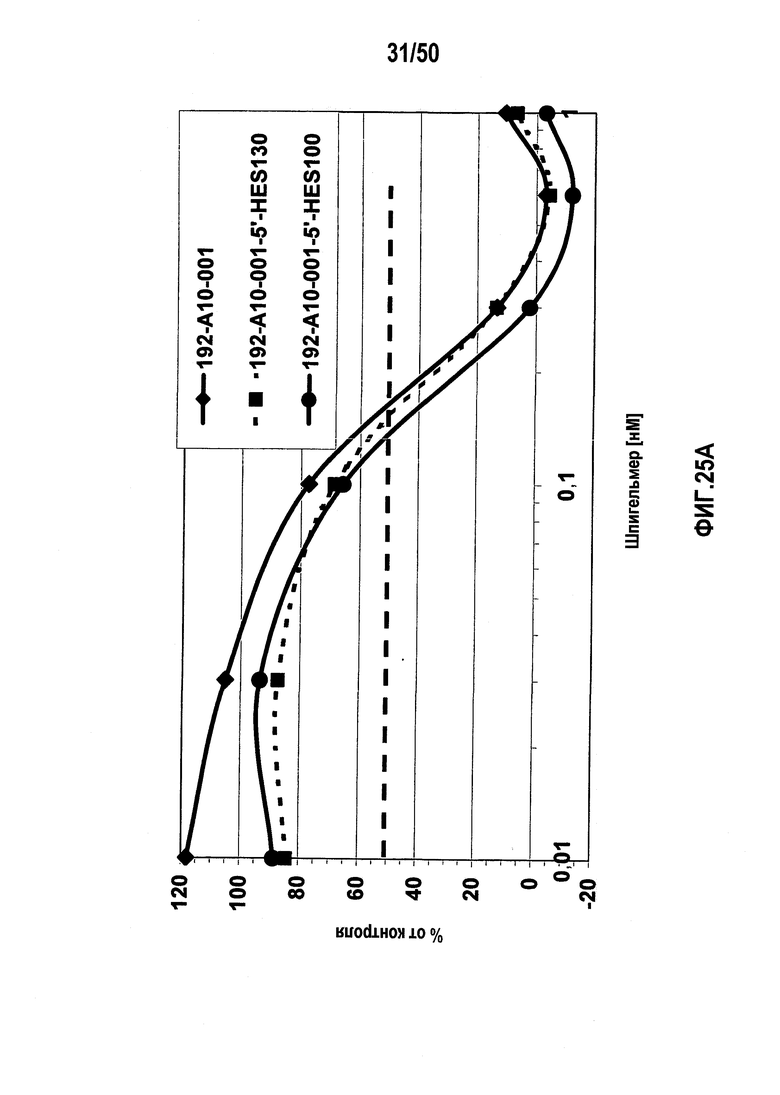

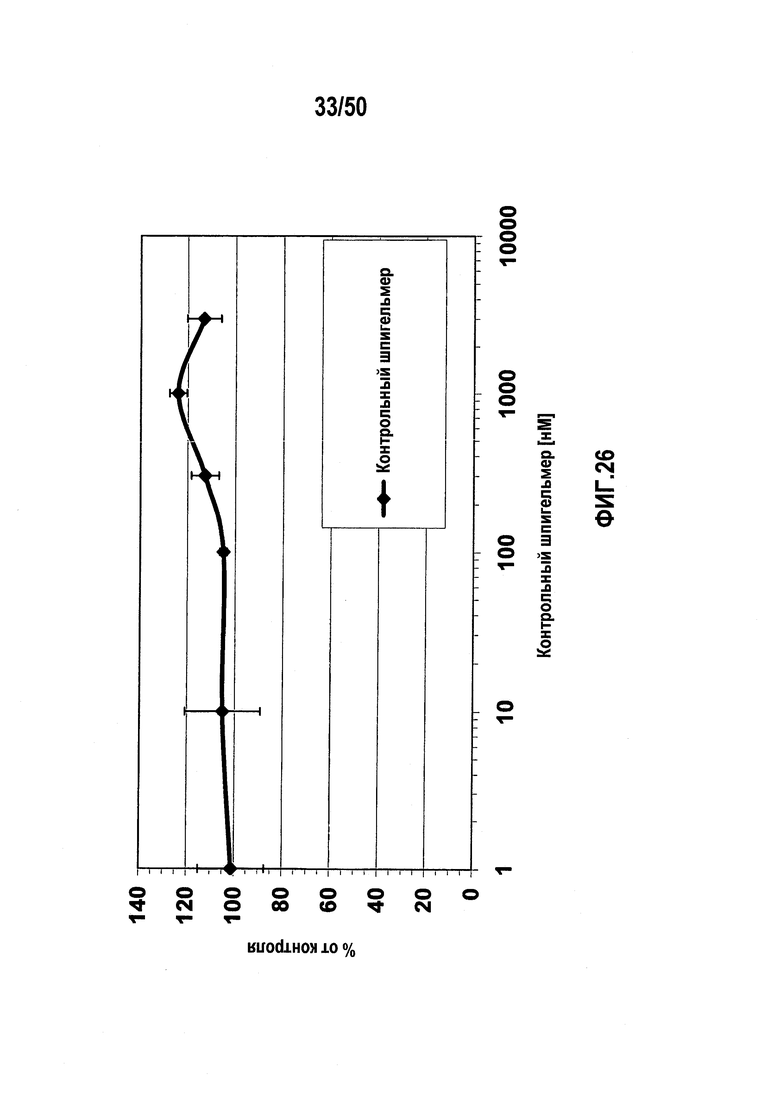
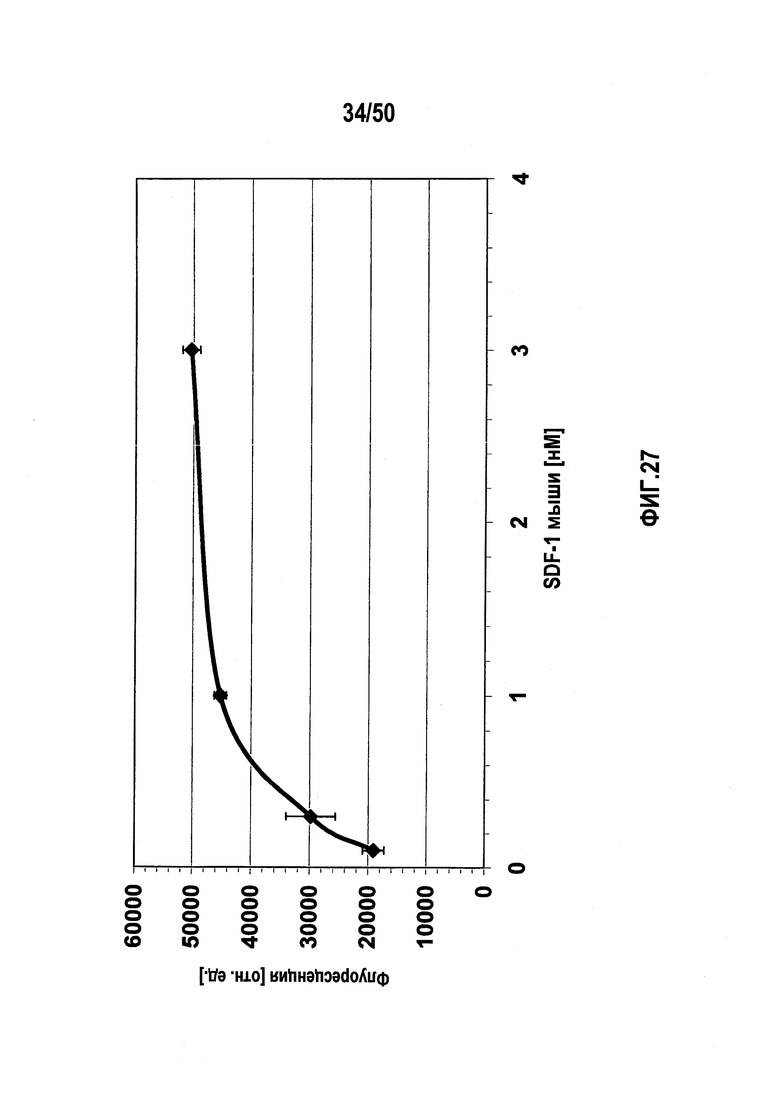
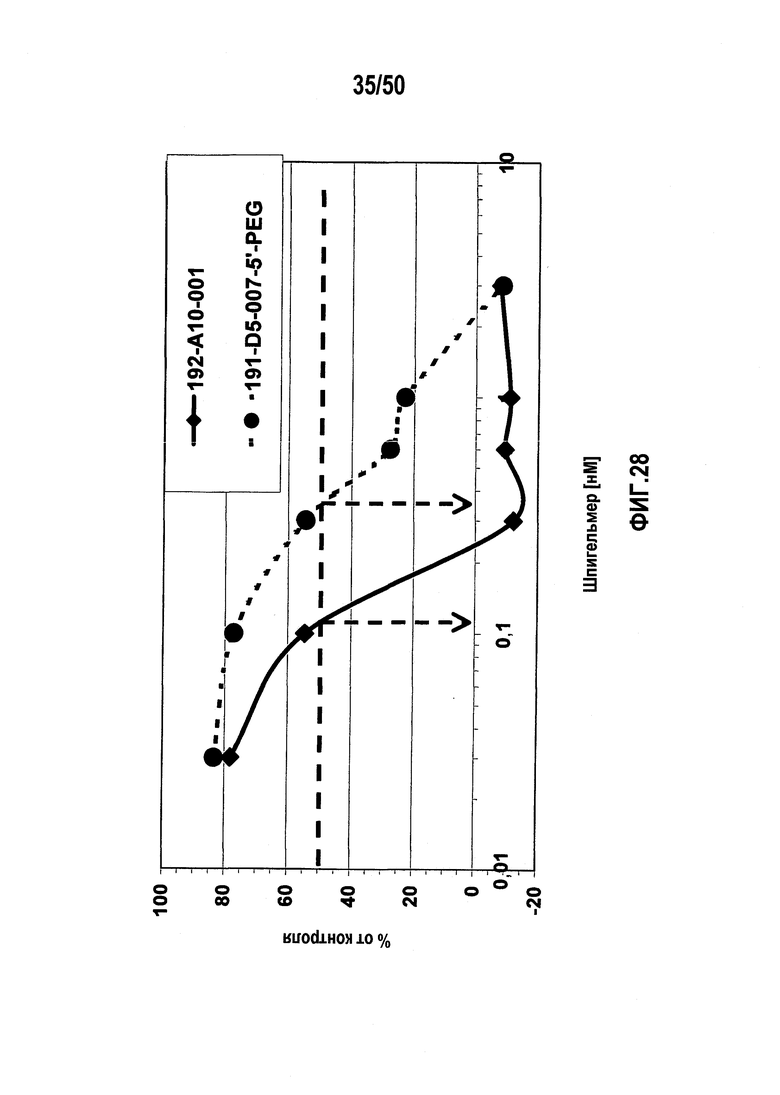
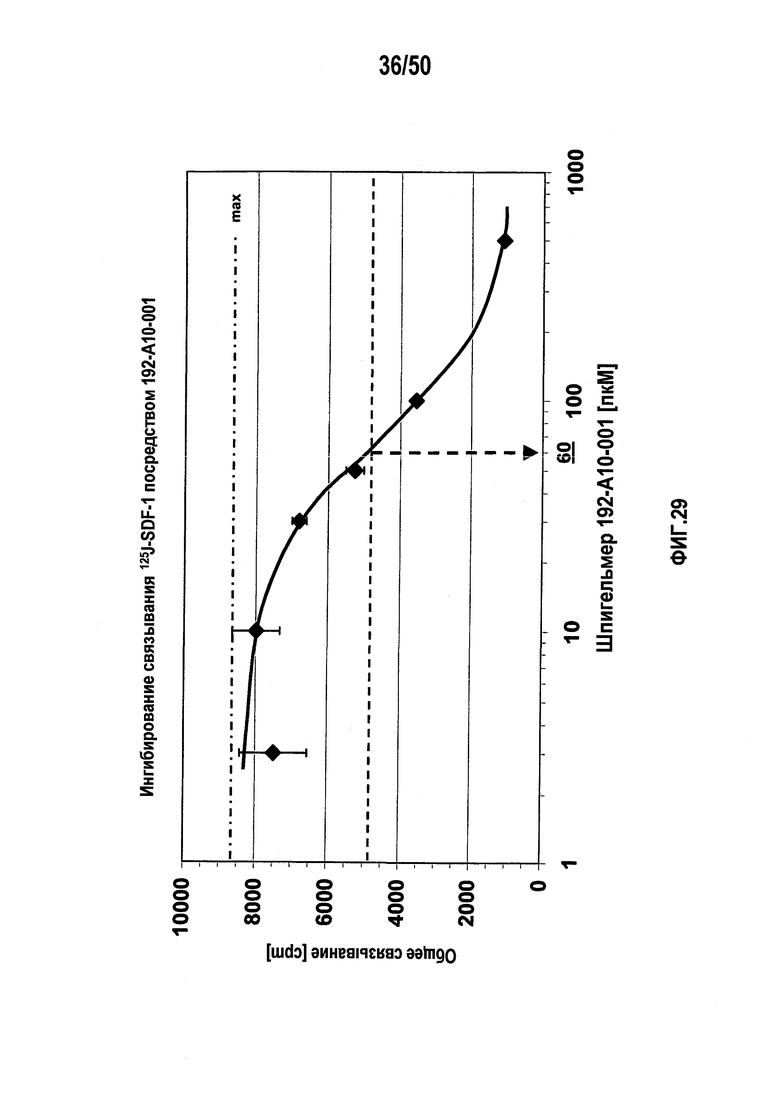

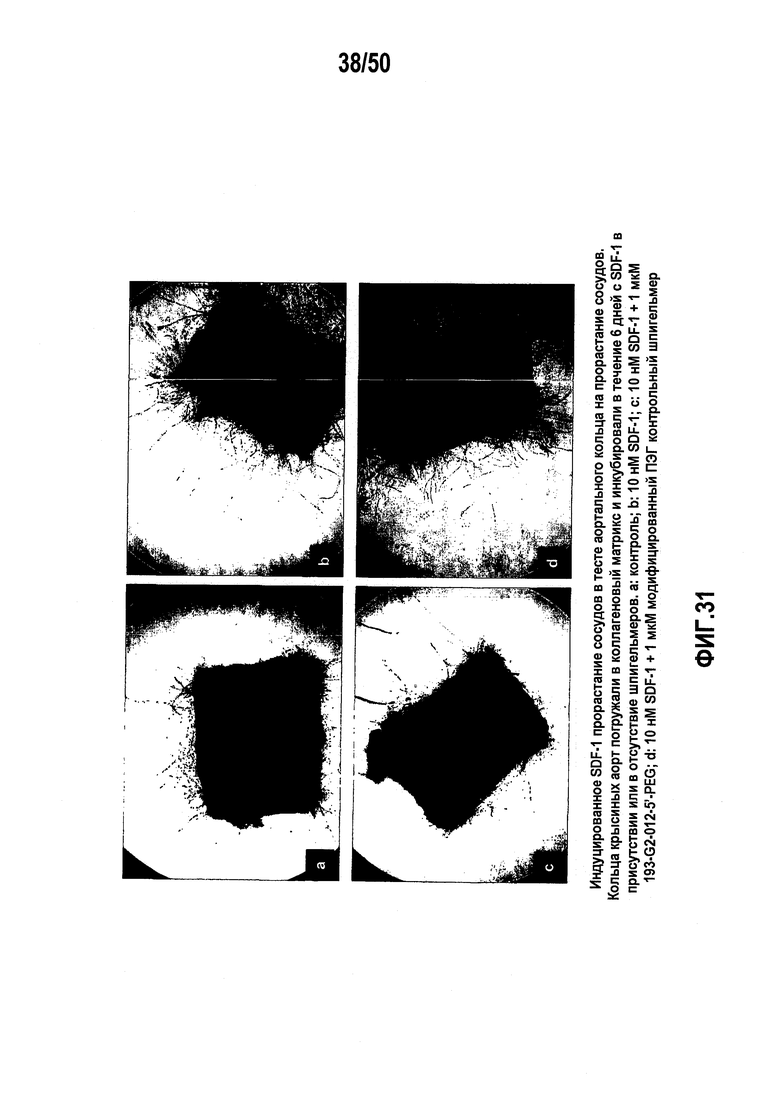



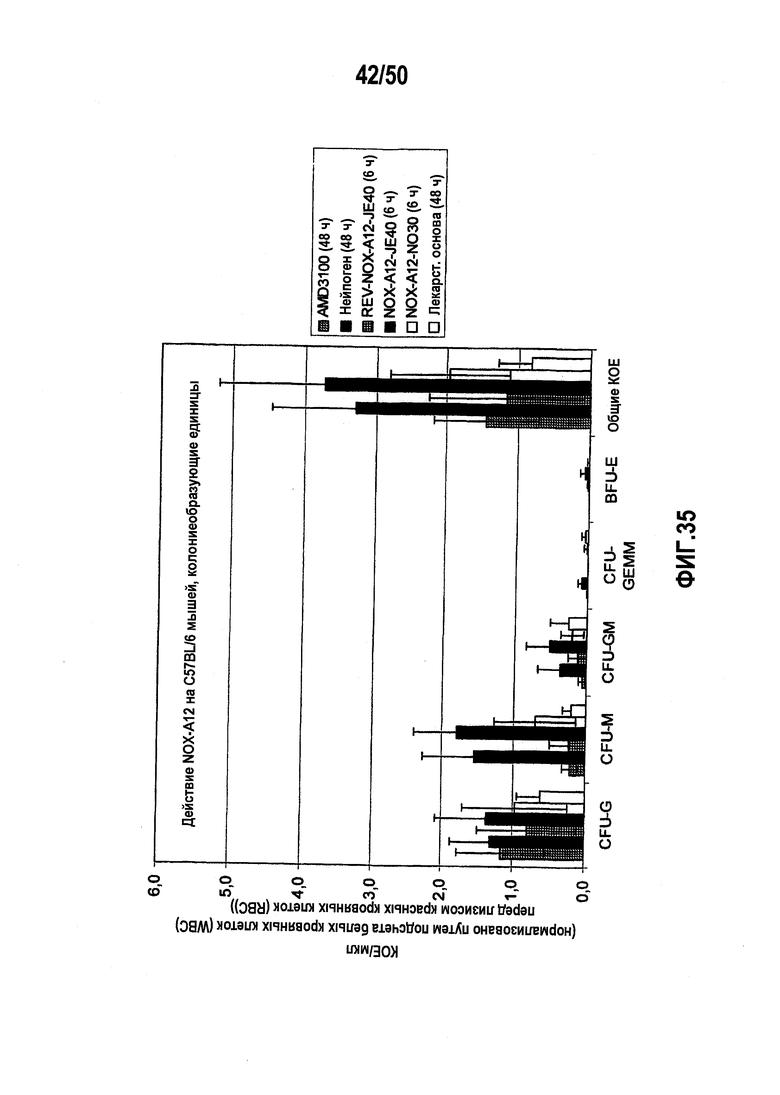

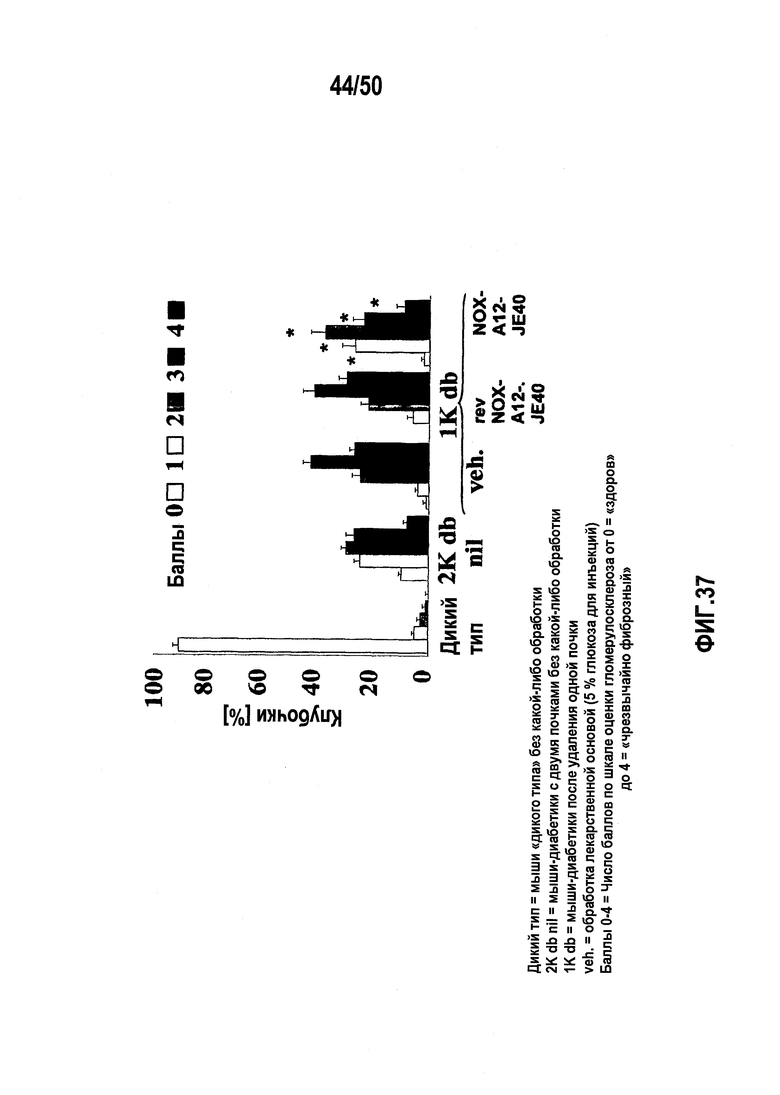


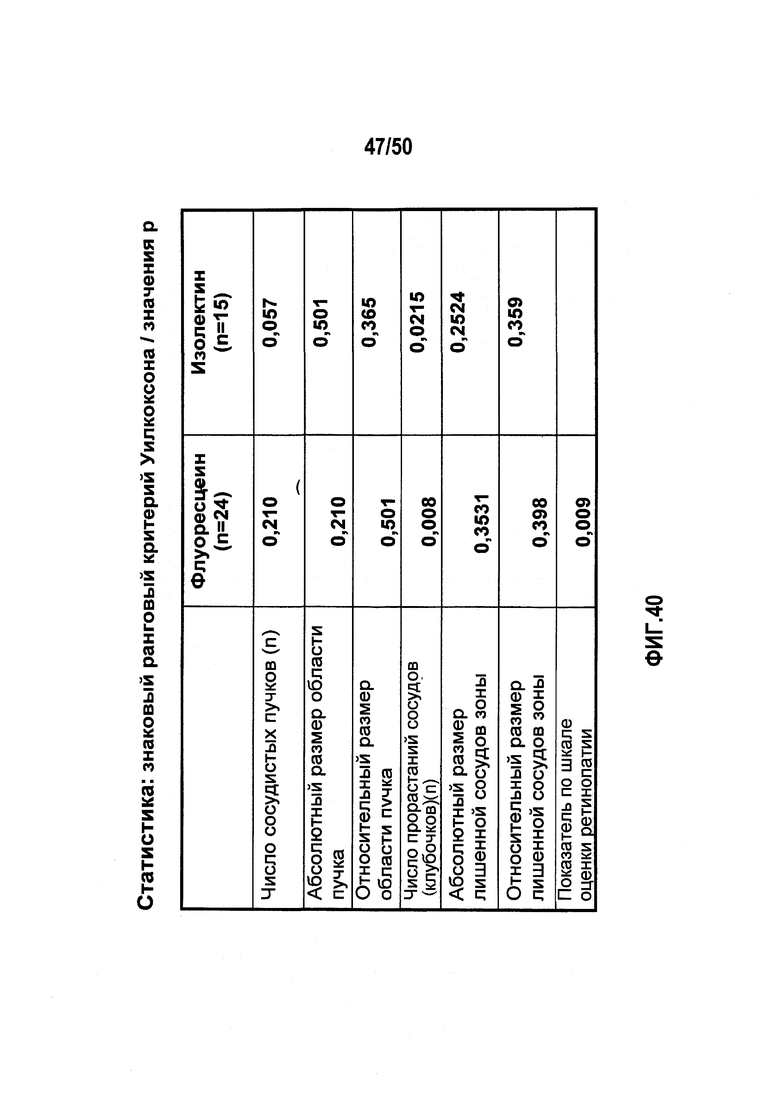
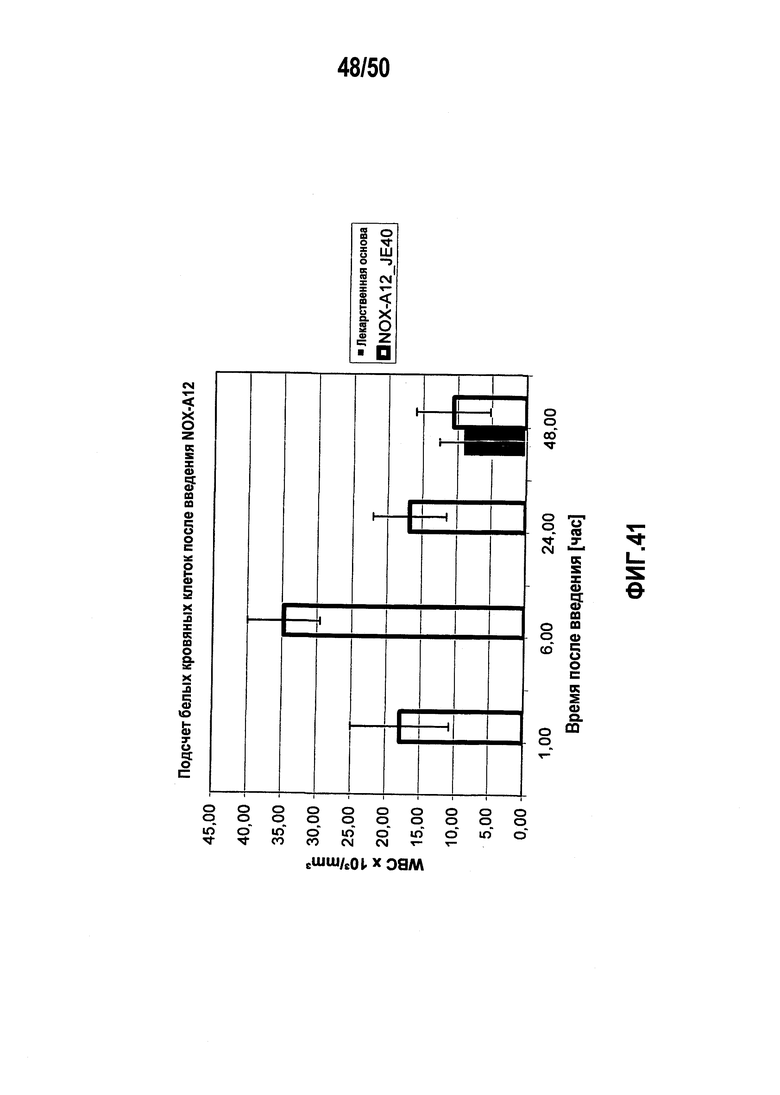


Изобретение относится к области биотехнологии, конкретно к клеточным технологиям, и может быть использовано в медицине для получения лекарственного средства для лечения рака. Получают молекулу L-нуклеиновой кислоты, связывающуюся с SDF-1, которая содержит в 5'→3' направлении первый участок нуклеотидов, коровую нуклеотидную последовательность и второй участок нуклеотидов или в 5'→3' направлении второй участок нуклеотидов, коровую нуклеотидную последовательность и первый участок нуклеотидов. При этом коровая нуклеотидная последовательность содержит 5'GUGUGAUCUAGAUGUADWGGCUGWUCCUAGUYAGG3' (SEQ ID NO: 57), где указанный первый и второй участок нуклеотидов имеет достаточную степень обратной комплиментарности для гибридизации друг с другом, где при гибридизации формируется двухнитевая структура и где двухнитевая структура состоит из четырех-шести пар оснований. Указанная нуклеиновая кислота может быть модифицирована HES или PEG. Изобретение позволяет способствовать мобилизации клеток, экспрессирующих рецепторы SDF-1, в периферическую кровь пациента. 5 н. и 4 з.п. ф-лы, 43 ил., 3 табл., 14 пр.
1. Молекула L-нуклеиновой кислоты, связывающаяся с SDF-1, для применения в способе влияния на миграцию клеток, экспрессирующих рецепторы SDF-1, где миграция клеток включает мобилизацию указанных клеток в периферическую кровь пациента, где молекула нуклеиновой кислоты содержит в 5'→3' направлении первый участок нуклеотидов, коровую нуклеотидную последовательность и второй участок нуклеотидов или в 5'→3' направлении второй участок нуклеотидов, коровую нуклеотидную последовательность и первый участок нуклеотидов,
где коровая нуклеотидная последовательность содержит следующую нуклеотидную последовательность:
5'GUGUGAUCUAGAUGUADWGGCUGWUCCUAGUYAGG3' (SEQ ID NO: 57), где указанный первый и второй участок нуклеотидов имеет достаточную степень обратной комплиментарности для гибридизации друг с другом, где при гибридизации формируется двухнитевая структура и где двухнитевая структура состоит из четырех-шести пар оснований.
2. Молекула L-нуклеиновой кислоты, связывающаяся с SDF-1, для применения в способе влияния на миграцию клеток, экспрессирующих рецепторы SDF-1, где миграция клеток включает мобилизацию указанных клеток в периферическую кровь пациента, где молекула нуклеиновой кислоты содержит в 5'→3' направлении первый участок нуклеотидов, коровую нуклеотидную последовательность и второй участок нуклеотидов или в 5'→3' направлении второй участок нуклеотидов, коровую нуклеотидную последовательность и первый участок нуклеотидов, где коровая нуклеотидная последовательность содержит следующую нуклеотидную последовательность:
5'GUGUGAUCUAGAUGUADWGGCUGWUCCUAGUYAGG3' (SEQ ID NO: 57), где указанный первый и второй участок нуклеотидов имеет достаточную степень обратной комплиментарности для гибридизации друг с другом, где при гибридизации формируется двухнитевая структура, и где двухнитевая структура состоит из четырех-шести пар оснований,
и где нуклеиновая кислота содержит модификацию, где модификация выбрана из группы, включающей молекулу HES и молекулу PEG.
3. Молекула L-нуклеиновой кислоты по п. 1 или 2, где молекула нуклеиновой кислоты содержит коровую нуклеотидную последовательность
GUGUGAUCUAGAUGUADUGGCUGAUCCUAGUCAGG (SEQ ID NO: 58).
4. Молекула L-нуклеиновой кислоты по п. 1 или 2, где первый участок нуклеотидов содержит нуклеотидную последовательность 5'X1X2SVNS3' (SEQ ID NO: 77) и второй участок нуклеотидов содержит нуклеотидную последовательность 5'BVBSX3X43' (SEQ ID NO: 78), где
X1 либо отсутствует, либо представляет собой А, X2 представляет собой G, X3 представляет собой С и X4 либо отсутствует, либо представляет собой U; или
X1 отсутствует, X2 либо отсутствует, либо представляет собой G, X3 либо отсутствует, либо представляет собой С и X4 отсутствует.
5. Молекула L-нуклеиновой кислоты по п. 1 или 2, где первый участок нуклеотидов содержит нуклеотидную последовательность 5'X2SSBS3' (SEQ ID NO: 73) и второй участок нуклеотидов содержит нуклеотидную последовательность 5'BVSSX33' (SEQ ID NO: 74),
где X2 либо отсутствует, либо представляет собой G и X3 либо отсутствует, либо представляет собой С,
предпочтительно первый участок нуклеотидов содержит нуклеотидную последовательность 5'GCGUG3' и второй участок нуклеотидов содержит нуклеотидную последовательность 5'UACGC3'.
6. Молекула L-нуклеиновой кислоты по п. 1 или 2, где молекула нуклеиновой кислоты имеет нуклеотидную последовательность, представленную любой из SEQ ID NOs: 46-56, 61-65, 67-72 и 132.
7. Фармацевтическая композиция, содержащая молекулу нуклеиновой кислоты по п. 1 или 2 в количестве, эффективном для мобилизации клеток, экспрессирующих рецепторы SDF-1, в периферическую кровь субъекта, и по меньшей мере дополнительное фармацевтически активное средство, приемлемое для применения при лечении рака, и где фармацевтическая композиция используется для лечения рака и фармацевтически активное средство повреждает, разрушает и/или метит раковые клетки, где метка приводит к активации защитных сил организма.
8. Способ получения гемопоэтических или миелоидных клеток-предшественников и/или стволовых клеток от субъекта, включающий:
а) введение субъекту нуклеиновой кислоты по п. 1 или 2 в количестве, эффективном для мобилизации указанных гемопоэтических или миелоидных клеток-предшественников и/или стволовых клеток в периферическую кровь указанного субъекта;
b) последующий сбор указанных клеток-предшественников и/или стволовых клеток от указанного субъекта.
9. Применение молекулы нуклеиновой кислоты по п. 1 или 2 для получения лекарственного средства для лечения рака, где лекарственное средство вводят до химиотерапии или радиотерапии и где лекарственное средство применяется при химиотерапии или радиотерапии.
| WO 2006032144 A1, 30.03.2006 | |||
| US 2003017485 A1, 23.01.2003 | |||
| US 7141363 B2, 28.11.2006 | |||
| US 2004131585 A1, 08.07.2004 | |||
| NETELENBOS T | |||
| et al., Proteoglycans on bone marrow endothelial cells bind and present SDF-1 towards hematopoietic progenitor cells, Leukemia, 2003, v.17, n.1, p.175-184 | |||
| АДГЕЗИВНЫЕ МОЛЕКУЛЫ СОСУДОВ И МОДУЛЯЦИЯ ИХ ФУНКЦИЙ | 2000 |
|
RU2244748C2 |
| УОЛЛИС Д | |||
| и др., Клиническая иммунология и аллергология, 2000, Москва, "Практика", Глава 15. | |||

