Изобретение относится к области сельского хозяйства, в частности к способам генетико-селекционных исследований.
Известен способ оценки вклада двойного диплоидного элементарного генома, включающий выращивание полиплоидных растений пшеницы разного количественного и качественного геномного состава и их диплоидных сородичей - источников исходных геномов, определение площади флагового листа и использование в качестве оценки фактического значения признака в полиплоиде к теоретически рассчитанному исходя из суммарного признака диплоидных сородичей - источников исходных геномов (С.Е.Дунаев, Р.Л.Богуславский, М.И.Зеленский. Сравнительная характеристика видов пшеницы по морфологии листа и содержанию хлорофилла в условиях Дагестана и Ленинградской области // Труды по прикладной ботанике, генетике и селекции. - 1989. - т.127. - С.115-122).
Недостатком данного способа является необходимость выращивания наряду с полиплоидными видами их диплоидных сородичей - источников исходных геномов.
Известен способ оценки вклада генома в сложный количественный признак пшеницы, включающий выделение из мягкой пшеницы T.aestivum L. ее тетраформы и использование в качестве оценки сравнительного анализа массы 1000 семян T.aestivum AuBD и ее тетраформы АuB (В.Г.Конарев, Н.К.Губарева, И.П.Гаврилюк, В.Бушук. Идентификация генома Д у пшениц по глиадину // Вестник с/х науки. - 1972. - N 7. - С.108-114).
Недостатком этого способа является трудность получения тетраформы, которая сопряжена со скрещиванием и бэкроссированием до 7-8 поколения, кроме того, в результате плохой всхожести и слабой жизнеспособности очень трудно получить семена тетраформ.
Известен способ оценки вклада геномов в количественный признак, когда для сравнения используются представители первой филогенетической ветви и их гомологи по второй филогенетической ветви, в частности T.aestivum AuBD, T.persicum Vav. AuB и их гомологи T.miguschovae Zhir. AbGD, T.militinae AbG соответственно (Патент на изобретение №2101931, 20.01.1998) - ближайший аналог. Суть способа в выявлении наличия или отсутствия вклада элементарных диплоидных геномов при сравнении видов пшениц с известными уровнями количественных признаков с соответствующими гомологами. Схематично это выглядит следующим образом:

Благодаря такому подходу было установлено, что T.miguschovae AbAbGDgd неполный гомолог T.aestivum AuAuBBDD, поскольку у него не проявляется вклад одного из трех элементарных диплоидных геномов. То есть согласно феномогеномным исследованиям у T.aestivum- трехфеномный, а у T.miguschovae - двухфеномный уровень количественных признаков, что и отражено на схеме.
Недостатком является выращивание наряду с основными видообразцами их гомологов.
Цель изобретения - упростить и снизить трудоемкость оценки реализации элементарных двойных диплоидных геномов в полиплоидных растениях. При заявляемом способе исключается возделывание видов - гомологов, т.к. используют естественные полиплоидные видообразцы, а также полученные из T.aestivum L. AABBDD (2n=42) мелкогабитусные диплоидные макромутанты (2n=14).
Суть изобретения в использовании этих мелкогабитусных диплоидных форм в качестве феномогеномных маркеров (двойной диплоидный геном-феном), при помощи которых определяют вклады элементарных геномов в сложный количественный признак полиплоидного полигеномного растения.
Так, из данных табл.1 видно, что такой важный и сложный количественный признак, как масса зерна с колоса, характеризующая продуктивность растения, у мелкогабитусного мутанта и диплоидного T.monococcum L. очень близки. В то же время разница между образцами с диплоидным набором хромосом (мелкогабитусный мутант и T.monococcum) с озимой пшеницей Рассвет-1 по данному признаку примерно троекратная, а у яровой мягкой - двукратная. Это можно показать по средним показателям, используя при этом, как данные мелкогабитусного мутанта, так и диплоидного T.monococcum. К примеру: 2,11: 0,73=2,89≈3 или 2,11:0,69=3,06≈3.
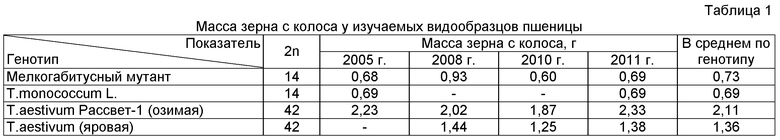
Относительно двукратного преимущества яровой мягкой - 1,36: 0,73=1,86≈2 или 1,36:0,69=1,97≈2. То есть у озимой мягкой пшеницы проявляются вклады всех трех его элементарных диплоидных геномов, а у яровой только двух. Следовательно признак мелкогабитусного мутанта (2n=14) можно использовать как феномогеномный маркер для выявления вклада элементарных диплоидных геномов в признаках полигеномного полиплоидного растения, в частности мягкой пшеницы (2n=42). Таким образом, используя в эксперименте всего лишь два образца пшеницы, мы существенно упрощаем способ, снижаем трудоемкость и затраты при оценке вкладов элементарных двойных диплоидных геномов в признак полиплоидной пшеницы.
Пример 1. Для того чтобы определить реализуются ли вклады всех элементарных геномов в признаке озимой мягкой пшеницы T.aestivum L. AABBDD (2n=42), достаточно вырастить одновременнно в одинаковых условиях ее и мелкогабитусный мутант (2n=14), а затем, используя признак мелкогабитусного диплоидного мутанта как феномогеномный маркер, расчетным путем выявить вклады скольких элементарных геномов формируют сложный признак данного растения. То есть 2,11: 0,73=2,89≈3 (см.табл.1). Чтобы подтвердить, факт реализации всех трех элементарных диплоидных геномов, проверим через χ2 (табл.2).
2n=14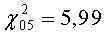 .
.
В качестве наблюдаемых частот (f) принимаем вычисленный вклад элементарного диплоидного генома (для чего делим признак озимой мягкой пшеницы на 3, зная ее трехгеномную природу и 42-хромосомный генотип: 2,11: 3=0,703; в качестве ожидаемых частот признаки феномогеномного маркера (признак мелкогабитусного мутанта (2n=14) 0,730 см. табл.1). Как видно из данных табл.2, теоретически рассчитанная масса зерна с колоса (2,190) очень близка к реальной (2,110) и нулевая гипотеза не отвергается. Таким образом, доказано, что в данном признаке проявляются вклады всех трех элементарных двойных диплоидных геномов (трехфеномный уровень), входящих в генотип полиплоидной мягкой пшеницы. Значит, используя признак мелкогабитусного мутанта, мы существенно упрощаем способ оценки реализации вкладов элементарных двойных диплоидных геномов в признак полиплоидной пшеницы.
Пример 2. Применяя показатели мелкогабитусного мутанта можно показать реализацию вкладов только двух элементарных геномов в признаке яровой мягкой пшеницы T.aestivum L. AABBDD, так как эти растения также выращивались одновременно с мелкогабитусным мутантом: 1,36: 0,73=1,86≈2 (см.табл.1). Таким образом, данный количественный признак у мягкой яровой пшеницы определяется вкладами двух элементарных геномов, то есть, несмотря на гексаплоидный трехгеномный статус, яровая мягкая пшеница обладает двухфеномным уровнем данного показателя.
Пример 3. Более того, используя признак мелкогабитусного мутанта как феномогеномный маркер, показан «двухфеномный» тетраплоидный уровень сложного количественного признака у гексаплоидной пшеницы T.compactum Host. AABBDD и Tsphaerococcum Perciv. AABBDD (табл.3).
Так, T.sphaerococcum AABBDD (Шарада исходная), обладает двухфеномным уровнем массы зерна с колоса: 1,55: 0,73=2,12≈2, тогда как мягкая пшеница T.aestivum AABBDD, выделенная из нее и улучшенная форма первой, трехфеномным уровнем данного признака: 2,19: 0,73=3 и 2,17:0,73=2,97≈3 соответственно. T.compactum AABBDD, использовавшийся в этом же опыте обладает двухфеномным уровнем: 1,46:0,73=2. Следовательно, используя показатели диплоидного мелкогабитусного мутанта, как феномо-геномный маркер, показано, что в продукционных признаках T.sphaerococcum и T.compactum реализуются вклады только двух элементарных двойных диплоидных геномов, тогда как у T.aestivum и Шарады улучшенной всех трех элементарных двойных диплоидных геномов. Следует подчеркнуть, что все эти видообразцы возделывались одновременно, мягкими пшеницами, о которых речь шла выше.
Таким образом, используя показатели мелкогабитусного диплоидного мутанта, мы существенно упростили и снизили трудоемкость и затраты на определение вкладов элементарных двойных диплоидных геномов в признаки полиплоидной пшеницы.
Изобретение относится к области сельского хозяйства, в частности к способам генетико-селекционных исследований. Изобретение представляет собой способ оценки реализации элементарных двойных диплоидных геномов в полиплоидных полигеномных пшеницах, включающий выращивание в одинаковых условиях полиплоидных пшениц и мелкогабитусного диплоидного мутанта, анализ их количественных признаков и последующее сравнение результатов анализа, по которым оценивают реализацию диплоидного генома. При этом в качестве феномогеномного маркера для определения вклада элементарного диплоидного генома в признак полиплоидной пшеницы используют полученную из T.aestivum L. AABBDD фенотипически идентичную ей мелкогабитусную диплоидную форму. Изобретение позволяет упростить оценку реализации элементарных двойных диплоидных геномов в полиплоидных полигеномных пшеницах. 3 табл., 3 пр.
Способ оценки реализации элементарных двойных диплоидных геномов в полиплоидных полигеномных пшеницах, включающий выращивание в одинаковых условиях полиплоидных пшениц и мелкогабитусного диплоидного мутанта, анализ их количественных признаков и последующее сравнение результатов анализа, по которым оценивают реализацию диплоидного генома, отличающийся тем, что в качестве феномогеномного маркера для определения вклада элементарного диплоидного генома в признак полиплоидной пшеницы используют полученную из T.aestivum L. AABBDD фенотипически идентичную ей мелкогабитусную диплоидную форму.
| РОМАНОВ Б.В, Улучшение продукционных характеристик шарозерной пшеницы, Вестник Российской академии сельскохозяйственных наук, N5, 2010, с | |||
| Устройство для выпрямления многофазного тока | 1923 |
|
SU50A1 |
| РОМАНОВ Б.В., Закономерность дозированного характера изменений количественных признаков пшеницы при полиплоидизации, Сельскохозяйственная биология, N5, 2001, с 105-112 | |||
| СПОСОБ ОЦЕНКИ РЕАЛИЗАЦИИ ГЕНОМОВ В ПОЛИПЛОИДНЫХ ПШЕНИЦАХ | 1993 |
|
RU2101931C1 |

