Изобретение относится к биотехнологии, к генной инженерии, микробиологической промышленности и ветеринарии и представляет собой дрожжевой штамм - продуцент секреторного рекомбинантного иммунного интерферона гамма собаки, содержащий сконструированную in vitro плазмиду, которая обеспечивает синтез секреторного рекомбинантного иммунного интерферона гамма собаки.
В настоящее время количество домашних собак в России и мире стремительно растет в связи с чем проблема профилактики и лечения вирусных болезней домашних животных приобретает особую остроту. Наиболее опасными высоколетальными вирусными заболеваниями собак являются чума собак, парвовирус собак, вирус парагриппа собак, инфекционный гепатит собак. Действенным способом лечения может выступать терапия на основе интерферонов, в частности интерферона гамма. Иммунный интерферон, гамма-интерферон, относится к обширной группе эволюционно родственных белков, получивших название интерферонов. Образование гамма-интерферона происходит в Т-лимфоцитах и ЕК-клетках (естественных киллерах) под воздействием, соответственно, вирусов и опухолевых клеток. Гамма-интерферон активирует специфический клеточный иммунитет, стимулирует цитотоксичность макрофагов, повышает устойчивость организма к различным инфекциям. Использование аутентичного гамма-интерферона собаки потенциально обладает гораздо меньшим побочным эффектом со стороны иммунной системы собак и близких видов по сравнению с применением аналогичных белков человека. Однако, систематические исследования спектра биологического действия аутентичного гамма-интерферона собаки Canis familiaris, как и его применение в качестве лекарственного препарата затруднены, так как в настоящее время в России не существует препаратов гамма-интерферона собаки.
Перспективным подходом для получения гамма-интерферона собаки Canis familiaris в значительных количествах является использование микроорганизмов в качестве продуцентов этого препарата. Использование бактерии Escherichia coli для этих целей рискованно, так как указанная бактерия является условно патогенным микроорганизмом, и препараты гамма-интерферона, полученные из бактериальных клеток, могут содержать эндотоксины. Вышеизложенное свидетельствует о перспективности создания штаммов-продуцентов гамма-интерферона собаки на основе других микроорганизмов, в частности, дрожжей [1]. Применение непатогенных микроорганизмов (дрожжей), не содержащих токсических и пирогенных факторов, в качестве продуцентов белков животных, позволяет использовать рекомбинантные белки в ветеринарной практике. Дрожжи Komagataella pastoris (Pichia pastoris) выглядят в данном случае особенно перспективными, так как сочетают такие важные параметры как непатогенность, простота культивирования и низкая себестоимость конечного продукта, а также отсутствие иммуногенных альфа-1,3-маннозных связей в составе гликопротеинов.
В качестве прототипа принят патент JP 11266868 [2]. В указанном случае для продукции гамма-интерферона собаки в качестве организма-продуцента применяют дрожжи Saccharomyces cerevisiae. Для культивирования дрожжей используют среды относительно простого состава, для них разработаны способы повышения уровня продукции и секреции рекомбинантных белков. Недостатком этой системы экспрессии является гипергликозилирование секретируемых гликопротеинов [3]. Кроме того, известно, что рекомбинантные гликопротеины содержат маннозные остатки, связанные альфа-1-3 связями, которые являются сильными антигенными детерминантами [4]. Это обстоятельство ограничивает использование рекомбинантных гликопротеинов, синтезированных дрожжами S. cerevisiae, в качестве лекарственных препаратов. Гамма-интерферон собаки относится к гликопротеинам, поэтому для его продукции система экспрессии на основе дрожжей S. cerevisiae не применима.
Заявленное изобретение свободно от указанного недостатка за счет того, что в качестве продуцента используются дрожжи Komagataella pastoris. Поэтому получение гамма-интерферона собаки с помощью дрожжей Komagataella pastoris, которые не обладают указанным ограничивающим фактором, является перспективным.
Целью предлагаемого объекта изобретения является создание штамма дрожжей Komagataella pastoris, который может быть использован для получения ветеринарного препарата на основе интерферона гамма собаки (Canis familiaris) для профилактики и лечения инфекционных вирусных заболеваний домашних животных, таких как чума собак, парвовирус собак, вирус парагриппа собак, инфекционный гепатит собак.
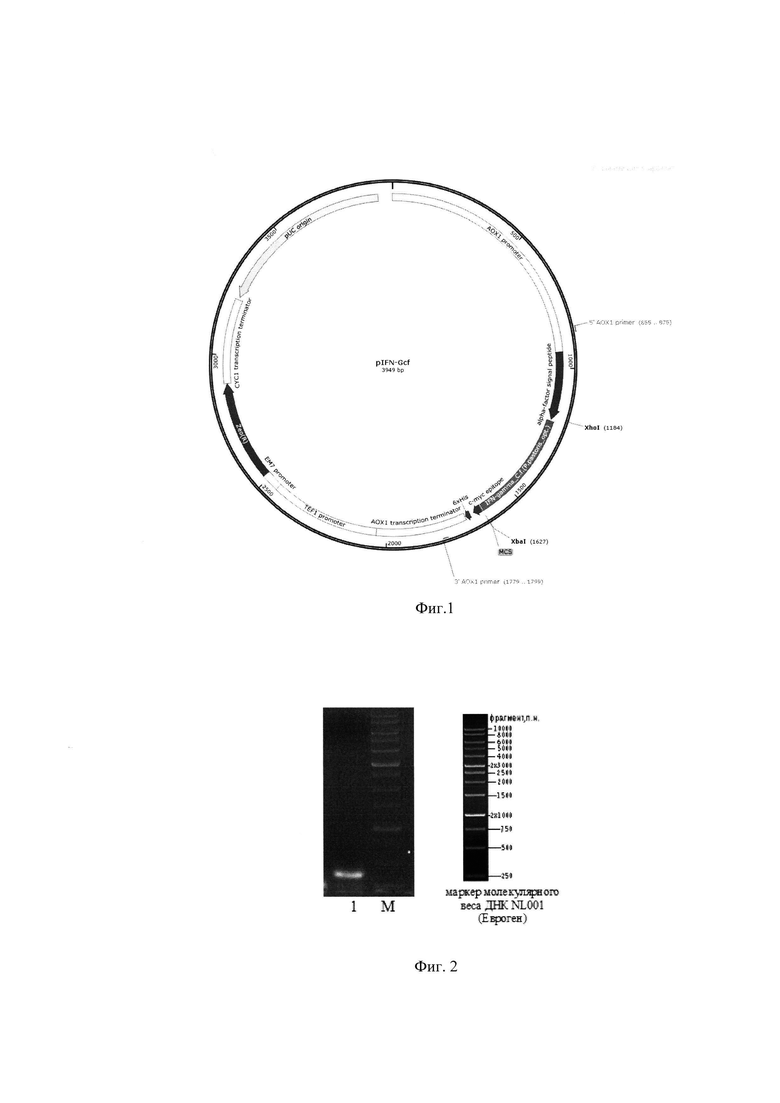
Плазмида pIFN-Gcf, обеспечивающая синтез гамма-интерферона собаки трансформированными ею клетками дрожжей, состоит из следующих элементов:
- XhoI-XbaI - фрагмент плазмидной ДНК дрожжевого вектора pPICZαA размером 3 568 п.о. (из которого удалена область сайта множественного клонирования размером 25 п.о. между сайтами распознавания рестриктаз XhoI и XbaI), включающий ген устойчивости к зеоцину, бактериальную область инициации репликации; фрагмент 5'-некодирующей области дрожжевого гена АОХ1 размером 940 п.о., содержащий область, обеспечивающую активацию транскрипции этого гена в присутствии метанола в качестве источника углерода в культуральной среде; фрагмент гена АОХ1 размером 342 п.о., содержащий область терминации транскрипции этого гена;
- XhoI-XbaI - фрагмент размером 449 п.о., содержащий кодирующую часть гена гамма-интерферона собаки;
Схема плазмидной ДНК pIFN-Gcf изображена на Фиг. 1, где XhoI и XbaI - сайты распознавания соответствующих рестриктаз. Общий размер плазмиды pIFN-Gcf - 3949 п.о.
Для достижения цели используют следующий способ конструирования плазмиды, обеспечивающей синтез гамма-интерферона собаки в клетках дрожжей. Из плазмиды pPICZαA (Invitrogen) с помощью ферментов рестрикции XbaI и XhoI вырезают фрагмент сайта множественного клонирования размером 25 п.о., после чего вектор дефосфорилируют.Вставку - кодирующую часть гена гамма-интерферона собаки размером 449 п.о. - получают путем обработки ферментами рестрикции XbaI и XhoI промежуточного вектора pAL2T-IFN-Gcf, сконструированного ранее путем лигирования ПЦР продукта фрагмента гена гамма-интерферона собаки с линеаризованным вектором pAL2T (Евроген). Нуклеотидная последовательность гена гамма-интерферона собаки (SEQ ID NO: 1) была оптимизирована для экспрессии в дрожжах Komagataella pastoris путем замены нативных для млекопитающих кодонов на наиболее часто встречающиеся у данных дрожжей.
Полученной лигазной смесью трансформируют клетки штамма XL1Blue (Евроген), с последующим высеванием на низкосолевую среду LB с добавлением зеоцина. С помощью рестрикционного и ПЦР анализа доказывают наличие плазмиды pIFN-Gcf у трансформантов.
После интеграции указанной последовательности в вектор pPICZαA открытая рамка считывания итогового рекомбинантного белка гамма-интерферона собаки содержит: сигнал секреции αMF-фактор дрожжей Saccharomyces cerevisiae, фрагмент открытой рамки считывания гена гамма-интерферона собаки, оптимизированного для экспрессии в дрожжах Komagataella pastoris путем замены нативных для млекопитающих кодонов на наиболее часто встречающиеся у данного вида дрожжей, а также последовательностей эпитопа сМус и метки Hisx6 для очистки итогового рекомбинантного белка методом аффинной хроматографии (SEQ ID NO: 2).
Выбор конструкции плазмиды для продукции гамма-интерферона собаки обусловлен следующими причинами. Плазмида pIFN-Gcf получена на основе дрожжевого интегративного вектора pPICZαA. В результате трансформации линеаризованной плазмидой и последующей гомологичной рекомбинации происходит встраивание экспрессионной кассеты в хромосому дрожжей Komagataella pastoris, что обеспечивает стабильное поддержание клонированного гена гамма-интерферона. В состав плазмиды входит ген устойчивости к зеоцину, что позволяет селективно отбирать бактериальных и дрожжевых трансформантов на селективной среде с добавлением зеоцина.
Экспрессия гена гамма-интерферона собаки в составе плазмиды pIFN-Gcf находится под контролем промотора гена АОХ1, содержащим области, обеспечивающие активацию транскрипции в присутствии метанола в культуральной среде, а также область инициации транскрипции. Промотор гена АОХ1 относится к числу наиболее сильных дрожжевых промоторов. Уровень экспрессии генов, находящихся под контролем АОХ1 промотора, эффективно регулируется источниками углерода. Транскрипция гена АОХ1 полностью блокирована при выращивании дрожжей на среде с глюкозой, на среде с глицерином наблюдается только базальный уровень экспрессии гена. Использование метанола в качестве единственного источника углерода значительно усиливает экспрессию гена АОХ1 и, следовательно, генов, находящихся под контролем АОХ1 промотора. Это позволяет регулировать синтез гамма-интерферона собаки в клетках дрожжей. Регулируемая экспрессия клонированного гена позволяет существенно снизить метаболическую нагрузку на клетку дрожжей.
В качестве продуцента гамма-интерферона используют штамм Y-3486-IFN-Gcf. Штамм Y-3486-IFN-Gcf получен при трансформации штамма дрожжей Komagataella pastoris Y-3486 (wt) (ВКПМ) плазмидой pIFN-Gcf. Штамм Y-3486 не несет гена устойчивости к зеоцину, что позволяет селективно отбирать трансформантов, несущих плазмиду pIFN-Gcf.
Штамм дрожжей Komagataella pastoris Y-3486-IFN-Gcf характеризуется следующими признаками.
Культурально-морфологические признаки.
На полной дрожжевой среде образует кремоватые колонии, гладкие, блестящие, пастообразной консистенции, клетки от круглых до овальных (2,0-4,0)×(2,2-5,8) мкм. На жидкой среде образует осадок и пленку. Культура имеет характерный запах метилотрофных дрожжей.
Клетки хорошо растут на полной дрожжевой среде YEPD: 2% пептона, 1% дрожжевого экстракта, 2% глюкозы или 1% глицерина.
Кроме того, клетки хорошо растут на минеральной среде SC: 1,34% Yeast Nitrogen Base («Difco», США), 2% глюкозы (1% глицерина, 0,5% метанола), а также на других синтетических средах для дрожжей.
Физиолого-биохимические признаки.
Клетки растут в пределах от 4 до 35°С. Оптимальной температурой выращивания является 30°С. При росте в аэробных условиях клетки незначительно защелачивают среду. Оптимум рН для роста составляет 4,5-6,5.
В качестве источника углерода клетки могут использовать многие простые соединения, такие как: глюкоза, глицерин, метанол.
В качестве источника азота клетки могут использовать минеральные соли в аммонийной форме, аминокислоты, мочевину.
Клетки способны к аэробному росту.
Существенными признаками штамма является устойчивость к зеоцину.
Способ получения плазмиды pIFN-Gcf проиллюстрирован следующим примером.
ПРИМЕР 1.
Нуклеотидная последовательность рекомбинантного гена гамма-интерферона собаки (SEQ ID NO: 1) получена методом химического синтеза и включает фрагмент открытой рамки считывания гена гамма-интерферона собаки, оптимизированный для экспрессии в дрожжах Komagataella pastoris путем замены нативных для млекопитающих кодонов на наиболее часто встречающиеся у данных дрожжей, а также последовательностей эпитопа сМус и метки Hisx6 для очистки итогового рекомбинантного белка методом аффинной хроматографии. Оптимизированная последовательность была лигирована в промежуточный вектор pUC57 и предоставлена в данном виде компанией, осуществлявшей услуги по синтезу гена (Евроген).
Для амплификации указанного фрагмента гена гамма-интерферона собаки использовали следующие праймеры ПЦР (5'->3'): прямой праймер CTCGAGAAAAGACAAGCTATGTTCTT, содержащий сайт рестрикции XhoI, и обратный праймер TCTAGACCCTTAGAAGCTCTTCTACCTCT, содержащий сайт рестрикции XbaI.
Режим реакции ПЦР:
1 цикл: 95°С - 3 мин.
30 циклов:
95°С - 30 сек.
55°С - 30 сек.
72°С - 60 сек.
1 цикл: 72°С - 3 мин.
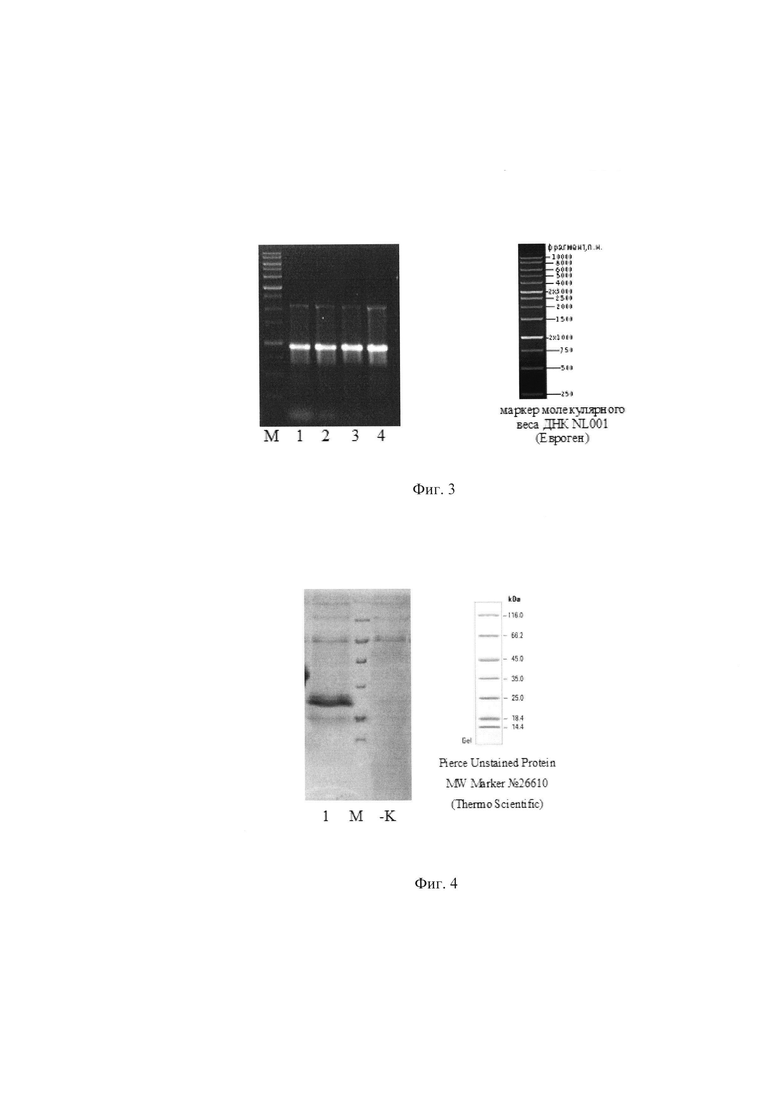
Далее проводили электрофорез продукта ПЦР реакции в 0,7% агарозном геле в буфере ТВЕ (0,1 М трис-боратный буфер, рН 8,3, содержащий 1 мМ ЭДТА). Для контроля размера амплифицированного фрагмента ДНК при электрофорезе использован молекулярный маркер 1 kb NL001 (Евроген). Результат проверки - электрофореграмма продуктов ПЦР при амплификации фрагмента гена гамма-интерферона собаки - представлен на Фиг. 2, где 1 - проба ПЦР продукта, М - маркер молекулярного веса ДНК. Размер ПЦР продукта соответствует расчетному в 449 п.о.
Выделение ПЦР фрагмент гена гамма-интерферона собаки из агарозного геля проводят по методике, разработанной фирмой QIAGEN. Полоску геля с фрагментом ДНК помещают в пробирку и добавляют раствор QX1 (300 мкл на 100 мг геля). Пробу нагревают до 50°С, добавляют реактив QIAEX (10 мкл на 5 мкг ДНК) и инкубируют при температуре 50°С в течение 10 минут, периодически перемешивая. Далее центрифугируют 30 секунд при 15000 об/мин, супернатант отбрасывают, осадок дважды экстрагируют растворами QX2 и QX3, удаляют супернатант центрифугированием при 15000 об/мин в течение 30 секунд. Осадок высушивают на воздухе, растворяют в 20 мкл буфера ТЕ, центрифугируют 30 секунд при 15000 об/мин., супернатант переносят в новую пробирку.
Выделенный ПЦР фрагмент гена гамма-интерферона собаки лигируют с линеаризованным вектором pAL2T (Евроген). Для этого 10 мкл реакционной смеси (2 мкл 5Х Quick ligation буфер, 1 мкл pAL2-T вектор (50 нг/мкл), 1 мкл Quick-TA Т4 ДНК лигаза, 3 мкл ПЦР продукт (50 нг/мкл), 3 мкл вода) инкубировали в течение 30 минут при 22°С. Продуктом лигирования выступает плазмида pAL2T-IFN-Gcf.
Полученную лигазную смесь используют для трансформации компетентных бактерий Escherichia coli. 200 мкл компетентных клеток с добавлением 1-2 мкл лигазной смеси инкубируют 15-20 минут во льду. Далее проводят тепловой шок в течение 60-90 секунд с возвращением клеток на 2 минуты в лед. После добавления среды LB (1% пептона, 0,5% дрожжевого экстракта, 1% хлористого натрия) проводят инкубацию в течение одного часа. Аликвоту в 100 мкл высевают на твердую среду LB с ампициллином. Колонии трансформантов отбирают после инкубирования в течение 12 часов при температуре 37°С.
Клетки бактерий Escherichia coli, содержащие плазмиду pAL2T-IFN-Gcf выращивают при 37°С в течение ночи в 1 л питательной среды LB (1% пептона, 0,5% дрожжевого экстракта, 1% хлористого натрия), содержащей ампициллин в концентрации 50 мг/л. Клетки собирают центрифугированием при 5000об/мин в течение 10 минут при 4°С, суспендируют в 20 мл 25 мМ трис-хлоридного буфера (рН 8,0), содержащего 10 мМ ЭДТА и 50 мМ глюкозы, добавляют 30 мг лизоцима и инкубируют 10 минут при комнатной температуре. Далее добавляют 40 мл 0,2М гидроокиси натрия, содержащей 1% додецилсульфата натрия, осторожно перемешивают и инкубируют в течение 10 минут при 4°С. Раствор нейтрализуют добавлением 30 мл 3 М ацетата натрия (рН 5,0) и выдерживают в течение 10 минут при 4°С. После этого центрифугируют при 14000 об/мин в течение 40 минут при 4°. К супернатанту добавляют 0,6 объема изо-пропилового спирта, выдерживают 20 минут при комнатной температуре и центрифугируют при 14000 об/мин в течение 20 минут при 20°С. Полученный осадок промывают 70% этиловым спиртом, высушивают в вакууме и растворяют в 4 мл дистиллированной воды. Далее добавляют 4,2 г хлористого цезия и 0,36 мл раствора бромистого этидия (10 мг/мл). Полученный раствор выдерживают в течение 1 часа при 4°С, затем центрифугируют при 15000 об/мин в течение 15 минут. Супернатант центрифугируют при 70000 об/мин в течение 16 часов в центрифуге TL 100 («Beckman»). После центрифугирования отбирают полосу плазмидной ДНК (нижнюю из двух флюоресцирующих в ультрафиолетовом свете полос), дважды экстрагируют бромистый этидий равным объемом изо-амилового спирта, разбавляют в два раза дистиллированной водой и осаждают плазмидную ДНК двумя объемами этилового спирта и 1/15 объема 3 М ацетата натрия (рН 5,0). Осадок собирают центрифугированием при 10000 об/мин в течение 10 минут, промывают 70% этиловым спиртом и растворяют в 0,5-1 мл буфера ТЕ (10 мМ трис-хлоридный буфер, рН 8,0, содержащий 1 мМ ЭДТА).
Концентрацию плазмидной ДНК определяют по поглощению раствора при длине волны 260 нм. Чистоту препарата контролируют при помощи электрофореза в 0,7% агарозном геле в буфере ТВЕ (0,1 М трис-боратный буфер, рН 8,3, содержащий 1 мМ ЭДТА).
Гидролиз плазмиды pAL2T-IFN-Gcf рестриктазами XbaI и XhoI проводят в 10 мМ трис-хлоридном буфере (рН 8,0), содержащем 100 мМ хлористого калия, 5 мМ хлористого магния, 0,02% тритона Х-100 и 0,1 мг/мл бычьего сывороточного альбумина. К 5 мкг плазмидной ДНК в объеме 20 мкл добавляют по 5 ед. каждой рестриктазы, после чего пробу инкубируют в течение 2 часов при 37°С. Далее проводят разделение продуктов рестрикции методом электрофореза в агарозном геле и проводят выделение фрагмента, соответствующего вставке синтетического гена гамма-интерферона собаки. Выделение фрагмента вставки ДНК из агарозного геля проводят по указанной выше методике QIAGEN.
Аналогично проводят обработку плазмиды pPICZαA рестриктазами XbaI и XhoI. Дополнительно продукт рестрикции дефосфорилируют с помощью щелочной фосфатазы AnP (NEB). Для этого на 20 мкл реакционной смеси добавляют: 2 мкл десятикратного буфера, 1-3 мкл плазмидной ДНК (50 нг/мкл), деионизованная вода - до 20 мкл, 1 мкл (5 ед.) щелочной фосфатазы. Инкубируют в течение 30 минут при 37°С. Производят остановку реакции нагревом до 80°С в течение 2 минут. Очищают ДНК гель-фильтрацией, на спин-колонках с последующим высаждением 96%-ным этанолом для удаления фосфатазы.
Для получения плазмиды pIFN-Gcf проводят лигирование 1 мкг дефосфорилированного линеаризованного вектора pPICZαA, гидролизованного рестриктазами XbaI и XhoI, и 3 мкг вставки целевого гена, гидролизованного рестриктазами XbaI и XhoI, в 10 мкл 70 мМ трис-хлоридного буфера (рН 7,6), содержащего 5 мМ дитиотреитола, 5 мМ хлористого магния, 1 мМ АТФ, добавляя 10 ед. ДНК-лигазы фага Т4 и инкубируя при 14°С в течение ночи.
Полученной лигазной смесью трансформируют клетки штамма Escherichia coli XL1-Blue recA1 endA1 gyrA96 thi-1 hsdR17 supE44 relA1 lac [F'proAB lacIqZΔM15 Tn10 (Tetr)] (Евроген). Для этого клетки Escherichia coli выращивают в 100 мл среды LB при 37°С до достижения культурой густоты клеточной суспензии, соответствующей 0,4-0,6 ед. оптической плотности при длине волны 550 нм. Клеточную суспензию охлаждают в ледяной бане, центрифугируют при 5000 об/мин в течение 10 минут при 4°С. Клетки суспендируют в 100 мл 10 мМ хлористого натрия, собирают центрифугированием в тех же условиях. Далее клетки суспендируют в 50 мл 75 мМ хлористого кальция, выдерживают в ледяной бане в течение 40 минут, осаждают центрифугированием в тех же условиях и суспендируют в 1 мл 75 мМ хлористого кальция. К суспензии компетентных клеток добавляют глицерин до конечной концентрации 15%, разделяют на аликвоты и хранят при -70°С. Перед трансформацией суспензию компетентных клеток размораживают в ледяной бане, добавляют лигазную смесь и инкубируют в ледяной бане в течение 40 минут. Далее клетки подвергают действию теплового шока при 42°С в течение 2 минут, после чего инкубируют в 1,5 мл среды LB при 37°С в течение 1 часа. Клетки собирают центрифугированием при 5000 об/мин в течение 10 минут и высевают на чашки Петри с низкосолевой средой LB, содержащей, 0,5% NaCl, 2% агара и 25 мкг/мл зеоцина. Чашки инкубируют при 37°С в течение 12-16 часов.
Из выросших отдельных клонов трансформантов выделяют плазмидную ДНК при помощи методики, использованной для получения плазмиды pAL2T-IFN-Gcf, за исключением того, что клетки Escherichia coli выращивают в 10 мл LB, и, соответственно, объемы всех растворов уменьшают в 100 раз, и проводят проверку методом ПЦР наличия вставки целевого гена. Для этого используются праймеры к 5'-АОХ1 промотору и 3'-АОХ1 в составе плазмиды: (5'->3'): прямой праймер GACTGGTTCCAATTGACAAGC, соответствующий 5'-АОХ1 промотору, и обратный праймер GCAAATGGCATTCTGACATCC, соответствующий 3'-AOX1 терминатору. Результат проверки - электрофореграмма ПЦР для 4-х проб на матрице плазмиды pIFN-Gcf -представлен на Фиг. 3, где 1, 2, 3, 4 - пробы ПЦР продуктов, М - маркер молекулярного веса ДНК. Размер ПЦР продукта соответствует расчетному в 945 п.о.
Режим реакции ПЦР:
1 цикл: 95°С - 3 мин.
30 циклов:
95°С - 30 сек.
55°С - 30 сек.
72°С - 90 сек.
1 цикл: 72°С - 2 мин.
Из выявленного таким образом клона препаративно выделяют плазмиду pIFN-Gcf, так же, как описано для плазмиды pAL2T-IFN-Gcf, и гидролизуют рестриктазой PmeI в однократном буфере (50 мМ ацетат натрия, 20 мМ Трис-ацетат, 10 мМ ацетат магния, 100 мкг/мл бычий сывороточный альбумин, рН 7.9 при 25°С) (NEB). PmeI-фрагмент плазмиды pIFN-Gcf, размером 3949 п.о, выделяют по описанной выше методике фирмы QIAGEN, исключая стадию нагревания и используют его для трансформации клеток дрожжей, как описано в примере 2.
ПРИМЕР 2.
Для получения штамма дрожжей Komagataella pastoris Y-3486-IFN-Gcf -продуцента секреторного рекомбинантного иммунного интерферона гамма собаки, клетки дрожжей штамма Komagataella pastoris Y-3486 (wt) трансформируют плазмидой pIFN-Gcf.
Клетки дрожжей выращивают в 100 мл среды YEPD при 30°С до достижения культурой оптической плотности, соответствующей 2-4 ед поглощения при длине волны 600 нм. Клетки дважды промывают стерильной водой, после чего суспендируют в 0,3 мл 100 мМ раствора ацетата лития и инкубируют при 30°С в течение 30 минут. К 50 мкл полученной суспензии клеток добавляют 0,1-1 мкг плазмидной ДНК, 50 мкг ДНК спермы лосося, предварительно денатурированной нагреванием (10 минут при 100°С) и 0,3 мл раствора 100 мМ ацетата лития, содержащего 40% полиэтиленгликоля 4000. Далее пробу инкубируют 30 минут при 30°С и 20 минут при 42°С, помещают на 15 секунд в ледяную баню и центрифугируют 10 секунд при 10000 об/мин. Клетки суспендируют в 1 мл стерильной воды и высевают на твердую полную дрожжевую среду YPD с содержанием 100 мкг/ мл зеоцина. Клоны трансформантов вырастают через 2-3 суток.
Для анализа продукции интерферона гамма собаки клетками трансформантов их выращивают при 30°С в 100 мл жидкой среды BMGY (2% пептона, 1% дрожжевого экстракта, 1% глицерина, 10 мл 1 М калий-фосфатного буфера, рН 6,0, 1,34% Yeast Nitrogen Base («Difco», США) до стационарной фазы роста в течение 2 суток. Клетки собирают центрифугированием при 5000 об/мин в течение 10 минут, супернатант сливают, и переносят всю биомассу в 100 мл жидкой среды BMMY (2% пептона, 1% дрожжевого экстракта, 0,5% метанола, 10 мл 1 М калий-фосфатного буфера, рН 6,0, 1,34% Yeast Nitrogen Base («Difco», США) для индукции экспрессии гена гамма-интерферона. Индукцию проводят при 30° в течение 4 суток. По окончании индукции отбирают пробы культуральной жидкости, из которых белок концентрируют осаждением 80% раствором сульфата аммония. Разделение белков проводят в 15% полиакриламидном геле в денаиурирующих условиях (электродный буфер: 25 мМ трис, 192 мМ глицин, 0,1% додецилсульфат натрия, рН 8,3; буфер для геля: 375 мМ трис-хлоридный буфер, рН 8,8). Параллельно проводят разделение белков контрольного штамма (штамм дикого типа Y-3489), выращенного в идентичных условиях. В качестве стандартов молекулярной массы используют лактатдегидрогеназу (35,0 кДа), эндонуклеазу рестрикции Bsp98I (25,0 кДа), (3-лактоглобулин (18,4 кДа), лизоцим (14,4 кДа). Среди секретируемых белков штамма-продуцента Y-3486-IFN-Gcf появляется дополнительная белковая полоса с молекулярной массой около 19,4 кДа, что соответствует расчетной массе рекомбинантного интерферона гамма собаки (SEQ ID NO: 2), рассчитанной сервисом https://sciencegateway.org/tools/proteinmw.htm. Результаты электрофореза белков культуральной жидкости показаны на Фиг. 4, где 1 - белки из культуральной жидкости штамма Y-3486-IFN-Gcf белка, М - маркер молекулярного веса белков, -К - белки из культуральной жидкости штамма дикого типа Y-3486 белка.
Уровень синтеза рекомбинантного интерферона гамма собаки определяют, сравнивая интенсивность окрашивания полосы рекомбинантного белка с полосой маркерного белка соответствующего веса. В качестве маркера концентрации был принят β-лактобулин (молекулярный вес - 18,4 кДа, концентрация 0,1 мг/мл). Согласно полученным данным, штамм Y-3486-IFN-Gcf синтезируют около 527 мг/л культуры дрожжей.
Предложенный штамм дрожжей Komagataella pastoris Y-3486-IFN-Gcf, оказался эффективным продуцентом рекомбинантного интерферона гамма собаки и может использоваться для получения ветеринарного препарата для профилактики и лечения инфекционных вирусных заболеваний домашних животных. Использованные источники информации:
1. Celik Е., Calik P. Production of recombinant proteins by yeast cells // Biotechnol. Adv. 2012. V. 30(5). P. 1108-1118.
2. Патент JP 11266868 (прототип).
3. Kukuruzinska M.A., Lennon K. Diminished activity of the first N-glycosylation enzyme, dolichol-P-dependent N-acetylglucosamine-l-P transferase (GPT), gives rise to mutant phenotypes in yeast // Biochim. Biophys. Acta. 1995. V. 1247. P. 51-59
4. Ballou C.E. A study of the immunogenicity of three yeast mannans // J. Biol. Chem. 1970. V. 245. P. 1197-1203.
Изобретение относится к биотехнологии, к генной инженерии, микробиологической промышленности и ветеринарии и представляет собой дрожжевой штамм - продуцент секреторного рекомбинантного иммунного интерферона гамма собаки, содержащий сконструированную in vitro плазмиду, которая обеспечивает синтез секреторного рекомбинантного иммунного интерферона гамма собаки. Из плазмиды pPICZαA с помощью гидролиза рестриктазами XhoI и XbaI удаляют область сайта множественного клонирования размером 25 п.о. и далее лигируют по данным сайтам рестрикции последовательность рекомбинантного гена гамма-интерферона собаки. Рекомбинантный ген включает фрагмент открытой рамки считывания гена гамма-интерферона, оптимизированного для экспрессии в дрожжах Komagataella pastoris. Проводят очистку итогового рекомбинантного белка методом аффинной хроматографии, затем лигазной смесью трансформируют клетки бактерий Escherichia coli и отбирают клоны, содержащие рекомбинантную плазмидную ДНК pIFN-Gcf. Изобретение позволяет создать штамм дрожжей Komagataella pastoris, который может быть использован для получения ветеринарного препарата на основе интерферона гамма собаки (Canis familiaris) для профилактики и лечения инфекционных вирусных заболеваний домашних животных, таких как чума собак, парвовирус собак, вирус парагриппа собак, инфекционный гепатит собак. 3 н.п. ф-лы, 4 ил., 2 пр.
1. Рекомбинантная плазмидная ДНК pIFN-Gcf, обеспечивающая биосинтез секреторного рекомбинантного интерферона гамма собаки, имеющая размер 3949 п.о. и состоящая из следующих элементов:
- XhoI-XbaI - фрагмент плазмидной ДНК дрожжевого вектора pPICZαA размером 3568 п.о. (из которого удалена область сайта множественного клонирования размером 25 п.о. между сайтами распознавания рестриктаз XhoI и XbaI), включающий ген устойчивости к зеоцину, бактериальную область инициации репликации; фрагмент 5'-некодирующей области дрожжевого гена АОХ1 размером 940 п.о., содержащий область, обеспечивающую активацию транскрипции этого гена в присутствии метанола в качестве источника углерода в культуральной среде; фрагмент гена АОХ1 размером 342 п.о., содержащий область терминации транскрипции этого гена;
- XhoI-XbaI - фрагмент размером 449 п.о., содержащий кодирующую часть гена гамма-интерферона собаки;
- уникальные сайты распознавания следующих рестриктаз: XhoI - 1185 п.о.; HindIII - 873 п.о.; XbaI - 628 п.о.; HpaI - 1496 п.о.; BsrGI - 3125 п.о; PmeI - 414 п.о.
2. Способ конструирования рекомбинантной плазмидной ДНК pIFN-Gcf по п. 1, обеспечивающей синтез секреторного рекомбинантного интерферона гамма собаки в клетках дрожжей Komagataella pastoris, заключающийся в том, что из плазмиды pPICZαA с помощью гидролиза рестриктазами XhoI и XbaI удаляют область сайта множественного клонирования размером 25 п.о. и далее лигируют по данным сайтам рестрикции последовательность рекомбинантного гена гамма-интерферона собаки, включающего фрагмент открытой рамки считывания гена гамма-интерферона, оптимизированного для экспрессии в дрожжах Komagataella pastoris путем замены нативных для млекопитающих кодонов на наиболее часто встречающиеся у данных дрожжей, а также последовательностей эпитопа сМус и метки Hisx6 для очистки итогового рекомбинантного белка методом аффинной хроматографии, затем лигазной смесью трансформируют клетки бактерий Escherichia coli и отбирают клоны, содержащие рекомбинантную плазмидную ДНК pIFN-Gcf.
3. Рекомбинантный штамм дрожжей Komagataella pastoris Y-3489-IFN-Gcf - продуцент рекомбинантного интерферона гамма собаки, полученный трансформацией штамма дрожжей Komagataella pastoris Y-3489 (ВКПМ) плазмидой pIFN-Gcf по п. 1.
| JP 11266868 A, 05.10.1999 | |||
| KUKURUZNSKA M.A., et al, Diminished activity of the first N-glycosylation enzyme, dolichol-P-dependent N-acetylglucosamine-l-P transferase (GPT), gives rise to mutant phenotypes in yeast, Biochim | |||
| Biophys | |||
| Acta, 1995, v | |||
| Устройство для автоматического регулирования магнитного потока проходящего через якорь динамо-машины | 1923 |
|
SU1247A1 |
| P | |||
| Способ запрессовки не выдержавших гидравлической пробы отливок | 1923 |
|
SU51A1 |
| ТЮРИН О.В., Разработка системы экспрессии генов на основе метилотрофных дрожжей | |||

