Данное изобретение относится к генной терапии, нано- и биотехнологии, фармацевтике, медицине и может быть использовано для доставки генетических конструкций в клетки с целью нормализации, ослабления или ликвидации патологического процесса, полного или частичного восстановления поврежденных или утраченных функций клеток, тканей и органов. Тканеспецифичность доставки генных конструкций обеспечивается благодаря использованию лигандных последовательностей к рецептору CXCR4.
Генную терапию от подходов традиционной медицины отличает то, что она ориентирована на борьбу с причиной заболевания, а не с симптомами или последствиями болезни, как в случае традиционной медицины. Первоначально генную терапию рассматривали как возможность коррекции дефекта в гене при моногенных заболеваниях путем замены мутантного фрагмента. В экспериментах на трансгенных животных было показано, что более продуктивным подходом может являться введение пациенту полноценно работающей копии гена. В настоящее время наряду с "терапевтическими" генами в клинических испытаниях применяют олигонуклеотидные последовательности ДНК и РНК. Так, перспективным направлением генной терапии является доставка антисмысловых нуклеотидов, в том числе малых интерферирующих РНК (миРНК), с целью направленной регуляции активности определенных генов.
Инъекции конструкций "голой" (naked) плазмидной ДНК были одним из первых подходов при разработке стратегии генотерапевтического лечения. В начале 90-х годов Вульфом с соавторами было показано, что инъекция плазмидной конструкции, содержащей маркерный ген lacZ, в мышцу мыши приводит к проникновению плазмиды в клетку и экспрессии гена β-галактозидазы в миофибриллах (Wolff JA, Malone RW, Williams P, Chong W, Acsadi G, Jani A, Felgner PL. Direct gene transfer into mouse muscle in vivo // Science. 1990; 247: 1465-1468). Однако основными недостатками введения "голой" плазмидной ДНК являются низкая эффективность проникновения в клетки и краткосрочность экспрессии введенных конструкций в большинстве органов и тканей. Низкая эффективность трансфекции с использованием незащищенных генетических конструкций послужила толчком к разработке методов доставки нуклеиновых кислот. В разрабатываемых подходах к доставке генетических конструкций можно выделить несколько основных направлений: использование вирусных векторов и невирусные способы доставки, включающие в себя физические способы и невирусные носители.
Вирусопосредованный перенос является высокоэффективным способом доставки нуклеиновых кислот, благодаря естественному механизму проникновения в клетки-мишени и способности защитить геном от разрушения внутриклеточными ферментами. К достоинствам данного вида векторов относится прежде всего тканеспецифичность, способность трансфецировать делящиеся и неделящиеся клетки, обеспечение долговременной экспрессии за счет интеграции в хромосому клетки-хозяина. Наиболее изученными являются векторы, созданные на основе ретровирусов, адено- и адено-ассоциировнных вирусов. Однако данный подход имеет некоторые недостатки. Ретро- и адено-ассоциированные вирусы имеют ограниченный размер клонированного фрагмента ДНК и риск инсерционного мутагенеза при встраивании вируса в геном хозяина. Серьезным недостатком аденовирусных векторов является ярко выраженный иммунный ответ при высоких дозах и повторных введениях аденовирусных конструкций (Walther W., Stein U. Viral vectors for gene transfer: a review of their use in the treatment of human disease // Drugs. - 2000. - V.60. - P.249-271, патент РФ №2252255, C12N 15/37, C12N 15/86, C12N 15/861, C12N 15/867, опубл. 2005.05.20).
На сегодняшний день разработан ряд альтернативных подходов для доставки генетических конструкций в клетки и ткани. К ним относят физические методы доставки и использование комплексов генетических конструкций с невирусными носителями. Наиболее популярными физическими методами доставки нуклеиновых кислот являются метод баллистической трансфекции и метод электропорации. При электропорации используют кратковременные электрические импульсы, приводящие к временной дестабилизации плазматической мембраны и образованию в ней пор, что увеличивает проницаемость клеток для экзогенных молекул, таких как ДНК, РНК. Метод баллистической трансфекции основан на попадании в клетки микрочастиц золота или вольфрама с адсорбированными на их поверхности генетическими конструкциями. Необходимое для попадания в клетки ускорение металлических частиц достигается с помощью электрического разряда или направленного потока гелия. Основными недостатками существующих физических методов трансфекции являются невысокая эффективность и локальность эффекта доставки нуклеиновых кислот. Они позволяют генетическим конструкциям преодолеть клеточную мембрану и избежать включения в эндосомы, предотвращая таким образом энзиматическую деградацию, но, как правило, не обеспечивают длительной персистенции введенных нуклеиновых кислот (Wells D.J. Gene therapy progress and prospects: electroporation and other physical methods // Gene Ther. - 2004. - V.11, №18. - P.1363-1369; Wang S., Joshi S., Lu S. Delivery of DNA to skin by particle bombardment // Methods Mol Biol. - 2004. - V.245. - P.185-196; Herweijer H., Wolff J.A. Progress and prospects: naked DNA gene transfer and therapy // Gene therapy. - 2003. - V.10, №6. - P.453-458).
Невирусные носители являются альтернативой вирусопосредованному переносу генетических конструкций в клетки млекопитающих. Невирусные носители легко синтезируются, легкость их модификации позволяет вносить изменения в структуру и состав молекул, тем самым, совершенствуя средства доставки. При использовании невирусных носителей отсутствуют ограничения на размер доставляемой генетической конструкции. Кроме того, они менее токсичны, в большинстве случаев не вызывают специфического иммунного ответа и более безопасны в применении in vivo по сравнению с вирусными векторами. Поэтому введение генетической конструкции, упакованной в невирусные носители, может осуществляться повторно. Исследование невирусных носителей развивается в направлении улучшения трансфецирующих свойств генетических конструкций путем образования комплексов с различными синтетическими соединениями (липидами, олиго- и полипептидами, полимерами и др.) (например, патент РФ №2336090, A61K 39/00, A61K 47/00, опубл. 2008.10.20). Совершенствование невирусных средств доставки во многом зависит от детального понимания барьеров на пути проникновения нуклеиновых кислот в клетки организма (Schmidt-Wolf G.D., Schmidt-Wolf I.G. Non-viral and hybrid vectors in human gene therapy: an update // Trends Mol Med. - 2003. - V.9, №2. - P.67-72; Gardlic R., Palffy R., Hodosy J., Turna J., Celec P. Vectors and delivery systems in gene therapy // Med Sci Monit. - 2005. - V.11, №4. - P.110-121; Wiethoff С.М., Middaugh C.R. Barriers to nonviral gene delivery // J Pharm Sci. - 2003. - V.92, №2. - P.203-217).
Пептидные носители являются перспективной группой средств доставки нуклеиновых кислот в клетки млекопитающих. Использование катионных пептидов для компактизации нуклеиновых кислот более предпочтительно, чем использование высокомолекулярных соединений, таких как гетерогенные поли-L-лизины. Основными направлениями развития пептидных носителей являются: (1) придание им эндосомолитических свойств, (2) присоединение лигандов тканеспецифичных рецепторов, (3) сигналов ядерной локализации в случае, когда необходимо доставить генетическую конструкцию в ядро.
Первым барьером на пути внутриклеточного проникновения комплексов является плазматическая мембрана. Большинство комплексов взаимодействуют с поверхностью клетки с помощью электростатических сил. Было показано, что основными неспецифическими молекулами, с которыми связываются комплексы на основе невирусных носителей, являются гликозамингликаны, которые присутствуют на поверхности всех клеток и участвуют во многих клеточных процессах, таких как дифференциация, адгезия и миграция. Для специфического взаимодействия с клеточной поверхностью в состав комплексов включают лиганды к рецепторам на поверхности клеток: трипептидный фрагмент RGD (интегрины присутствуют на поверхности многих клеток), трансферрин (его рецептор обладает повышенной экспрессией в пролиферирующих клетках), асиалоорозомукоид (асиалогликопротеиновый рецептор имеет специфическую экспрессию в гепатоцитах печени) (Schaffner Р., Dard М.М. Structure and function of RGD peptides involved in bone biology // Cell Mol Life Sci. 2003; 60: 119-132; Wu G.Y., Wu C.H. Receptor-mediated gene delivery and expression in vivo // J Biol Chem. 1988; 263: 14621-14624; Kircheis R., Wagner E. Polycation/DNA complexes for in vivo gene delivery // Gene therapy and regulation 2000; 1: 95-114).
Возможным подходом для создания системы направленного транспорта генов является модификация носителя лигандом к хемокиновому рецептору CXCR4. CXCR4 является рецептором фактора миграции стволовых клеток хемокина SDF-1 и представлен в гематопоэтических клетках, эндотелии сосудов, нейронах, мышечных сателлитных клетках. Отмечен высокий уровень экспрессии данного гена в более чем 20 видах раковых опухолей (рак груди, простаты и др.), а также в мигрирующих стволовых клетках (Juarez J, Bendall L, Bradstock К. Chemokines and their Receptors as Therapeutic Targets: The Role of the SDF-1/CXCR4 Axis. // Curr Pharm Des 2004; 10: 1245-1259). Таким образом, включение в состав молекул носителя лигандных последовательностей для связывания с рецептором CXCR4 является перспективным путем создания систем целевой доставки генов в клетки.
В результате процесса эндоцитоза комплексы носителя с нуклеиновыми кислотами оказываются заключенными внутри ранних эндосом, которые после процесса сортировки либо возвращаются на поверхность клетки, либо сливаются с лизосомами в составе так называемых поздних эндосом. Комплексы, неспособные покинуть эндосому, подвергаются деградации лизосомальными ферментами. Одним из возможных механизмов выхода из эндосомального компартмента является механизм «протонной губки» с участием молекул гистидина. Согласно гипотезе «протонной губки» свободные аминогруппы полимера при pH эндосомы ниже 6.0 способны протонироваться, т.е. присоединять вновь поступающие протоны, предотвращая закисление эндосом. Это влечет за собой поступление в эндосому дополнительных протонов, а также связанный с ним транспорт ионов хлора и воды. Вследствие этого происходит набухание эндосомы и ее последующий осмолизис.
Невирусные носители на основе катионных полимеров, такие как полилизины, ранее были исследованы в качестве средств доставки нуклеиновых кислот в клетки. Их основным недостатком является высокая токсичность. Возможной стратегией по снижению токсичности полимеров является уменьшение их молекулярного веса. Однако низкомолекулярные носители неспособны образовывать комплексы, стабильные при физиологических условиях, и эффективно защищать генетические конструкции от деградации. Для решения проблемы стабильности комплексов низкомолекулярных пептидов с нуклеиновым кислотами используют метод образования перекрестных сшивок между молекулами носителя с целью получения соединения с большим молекулярным весом. В качестве перспективных носителей на сегодняшний день рассматриваются синтетические самосшивающиеся цистеин-богатые пептиды. Наличие остатков цистеина, фланкирующих последовательность данных пептидов, обуславливает возможность образования дисульфидных связей между молекулами. Самосшивающиеся за счет остатков цистеина пептиды способны формировать плотные комплексы с плазмидной ДНК. После попадания в клетку происходит редукция дисульфидных связей и высвобождение ДНК из состава комплексов. Кроме того, редукция дисульфидных связей приводит к уменьшению токсичности соединения до уровня низкомолекулярных пептидов (McKenzie DL, Smiley Е, Kwok KY, Rice KG. Low Molecular Weight Disulfide Cross-Linking Peptides as Nonviral Gene Delivery Carriers // Bioconjugate Chem. 2000; 11: 901-909. Tanaka K, Kanazawa T, Ogawa T, Takashima Y, Fukuda T, Okada H. Disulfide crosslinked stearoyl carrier peptides containing arginine and histidine enhance siRNA uptake and gene silencing // International Journal of Pharmaceutics. 2010; 398: 219-224).
Результатом исследований пептидных носителей как средств доставки генетических конструкций должно стать создание модульного носителя, который бы содержал в себе сигналы для преодоления всех барьеров транспорта нуклеиновых кислот в клетку. Такие молекулярные коньюгаты представляют собой "искусственные вирусы", которые, однако, являются более безопасными в использовании, чем вирусные векторы. Вариант модульного носителя генетических конструкций был предложен Трентин с коллегами. Они разработали мультидоменный пептид, который содержал в своем составе цистеин-богатый пептид СНК6НС для связывания ДНК и выхода комплексов из эндосом и соединенную с ним через разделяющий спейсер последовательность сайта связывания трансглютаминазы. С помощью фактора свертываемости крови XIII комплексы внедряются в фибрильный матрикс, что обеспечивает пролонгированность генной доставки. Данный подход можно применять при тканевых повреждениях, когда существует необходимость в доставке генов ростовых факторов. Однако использование фибрильного матрикса подразумевает локальный характер доставки генных конструкций и ограничивает область применения данного модульного носителя (Trentin D, Hubbell J, Hall H. Non-viral gene delivery for local and controlled DNA release // Journal of Controlled Release. 2005; 102: 263-275).
При разработке модульных носителей особое внимание должно уделяться исследованию комбинаций лигандов-рецепторов, специфичных к клеткам-мишеням различных типов. Примером таких комбинаций может быть пара CXCR4-SDF-1α. CXCR4 является рецептором фактора миграции стволовых клеток хемокина SDF-1α и экспрессируется в гематопоэтических клетках, эндотелии сосудов, мышечных сателлитных клетках. Отмечен высокий уровень экспрессии данного гена в более чем 20 видах раковых опухолей (рак груди, простаты и др.), а также в мигрирующих стволовых клетках. Для доставки генетического материала в клетки, экспрессирующие рецептор CXCR4, Ле Бон с коллегами использовали синтетический лиганд к данному рецептору - AMD3100, который был соединен с полиэтиленимином или катионными липидами. Комплексы генетического материала с данными соединениями не приводили к достоверному повышению эффективности доставки маркерного гена в CXCR4+ клетки по сравнению с соединениями без сигналов. Носители, применяемые Ле Боном, не были эффективными, потому что специфическая доставка с их помощью возможна только при добавлении в среду трансфекции вещества, способствующего интернализации рецептора CXCR4 (форболовый эфир). (Le Bon В, Van Craynest N, Daoudi JM, Di Giorgio C, Domb AJ, Vierling P. AMD3100 Conjugates as Components of Targeted Nonviral Gene Delivery Systems: Synthesis and in Vitro Transfection Efficiency of CXCR4-Expressing Cells // Bioconjugate Chem 2004, 15: 413-423).
Таким образом, существует необходимость в создании носителя, модифицированного лигандом к рецептору CXCR4, способного обеспечить специфическую доставку нуклеиновых кислот в CXCR4+ клетки и не оказывать влияния на близлежащие ткани. Такой носитель обеспечивается настоящим изобретением.
В основу настоящего изобретения положена задача разработки носителя для направленной доставки нуклеиновых кислот в клетки, экспрессирующие рецептор CXCR4, для специфической доставки генетических конструкций в клетки, на поверхности которых представлен рецептор CXCR4, в котором за счет использования модульных носителей генетических конструкций, содержащих в своем составе последовательность-лиганд к рецептору CXCR4, достигают повышения эффективности и специфичности доставки маркерного гена.
Достижение поставленной технической задачи обеспечивается тем, что носитель для направленной доставки нуклеиновых кислот в клетки, экспрессирующие рецептор CXCR4, состоит из:
- последовательности-лиганда к рецептору CXCR4 с последовательностью аминокислот - KPVSLSYRSPSRFFESH;
- линкерного участка из двух молекул ε-аминогексановой кислоты, соединяющей последовательность-лиганд с последовательностью для компактизации нуклеиновых кислот;
- последовательности, обеспечивающей компактизацию нуклеиновых кислот и выход комплекса из эндосом - CHRRRRRRHC.
Для достижения эффективной трансфекции необходимо использование данных носителей в комплексе с генетическим материалом.
В качестве генетического материала может быть использована плазмидная ДНК, содержащая ген интереса, или олигонуклеотидные последовательности РНК, миРНК и ДНК. Изобретение поясняется с помощью фиг.1-7.
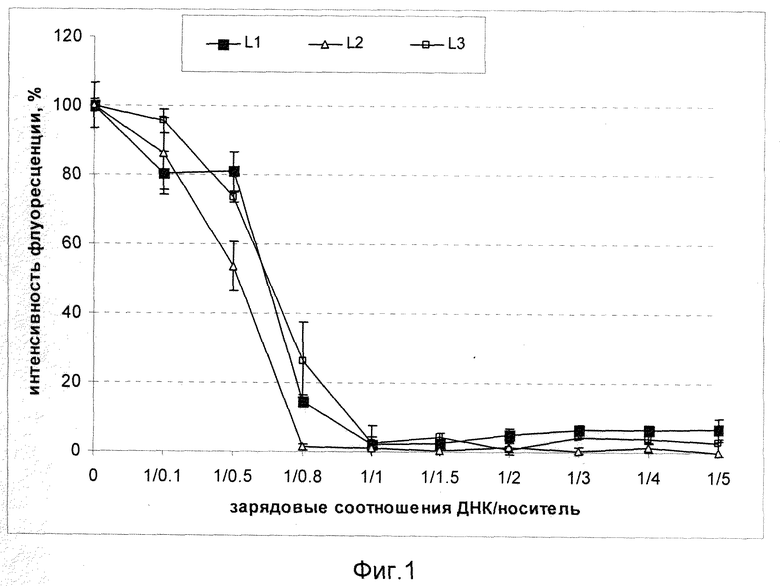
На фиг.1 показано изменение интенсивности флуоресценции бромистого этидия при увеличении зарядовых соотношений носитель/ДНК в комплексах L1/ДНК, L2/ДНК и L3/ДНК. Падение интенсивности флуоресценции свидетельствует о возрастании плотности формировавшихся комплексов. Выход кривых флюоресценции на плато указывает на то, что комплексы достигли плотности, достаточной для гашения флуоресценции бромистого этидия.
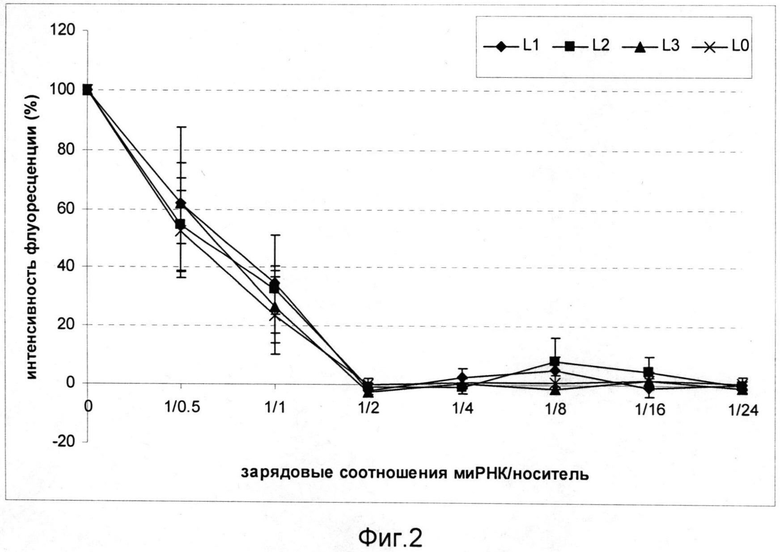
На фиг.2 продемонстрировано изменение интенсивности флуоресценции красителя SybrGreen при увеличении зарядовых соотношений носитель/миРНК в комплексах L1/миРНК, L2/миРНК и L3/миРНК. Выход кривых флюоресценции на плато указывает на то, что комплексы достигли плотности, достаточной для полного гашения флуоресценции Sybr Green.
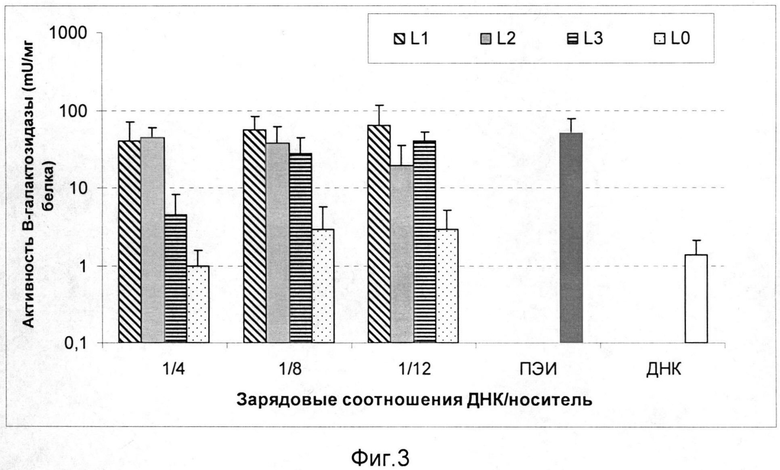
На фиг.3 приведена зависимость активности β-галактозидазы в клетках HeLa (CXCR4+) после трансфекции комплексами L1/ДНК, L2/ДНК и L3/ДНК. В этом случае использовали комплексы, сформированные при следующих зарядовых соотношениях носитель/ДНК: 4/1, 8/1, 12/1. В качестве контролей эксперимента служили интактная молекула ДНК, комплексы ПЭИ/ДНК 8/1 (положительный контроль эксперимента) и комплексы, содержащие ДНК и контрольный пептид (L0). L0 отличается от носителей в настоящем изобретении отсутствием сигнала связывания с рецептором CXCR4 и представляет собой последовательность CHRRRRRRHC. Эффективность доставки маркерного гена носителями L1, L2 и L3 была в 10 раз выше, чем контролем L0.
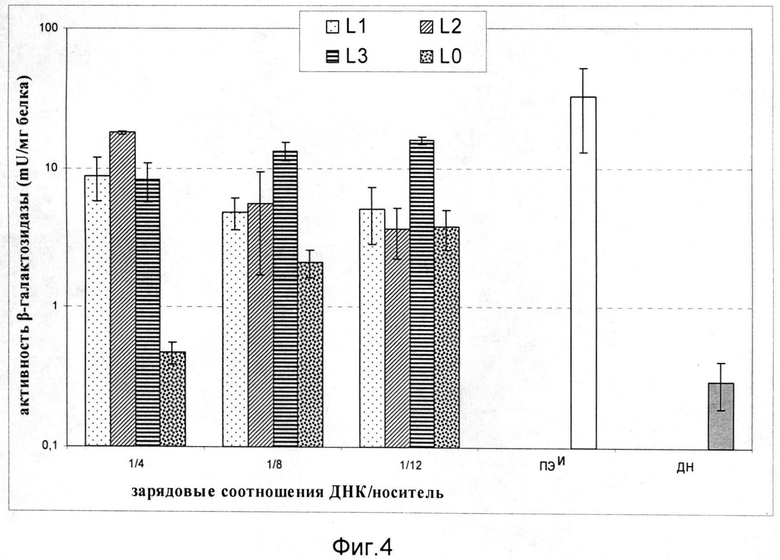
На фиг.4 приведена зависимость активности β-галактозидазы в клетках A172 (CXCR4+) после трансфекции комплексами L1/ДНК, L2/ДНК и L3/ДНК. Использованы комплексы, сформированные при следующих зарядовых соотношениях носитель/ДНК: 4/1, 8/1, 12/1. В качестве контролей эксперимента служили интактная молекула ДНК, комплексы ПЭИ/ДНК 8/1 и комплексы, содержащие ДНК и носитель L0. Эффективность доставки маркерного гена носителями L1, L2 и L3 была в 10 раз выше, чем контролем L0 при зарядовом соотношении носитель/ДНК 4/1. При увеличении зарядовых соотношений эффективность комплексов с носителем L3 была более чем в 5 раз выше, чем с носителей L0.
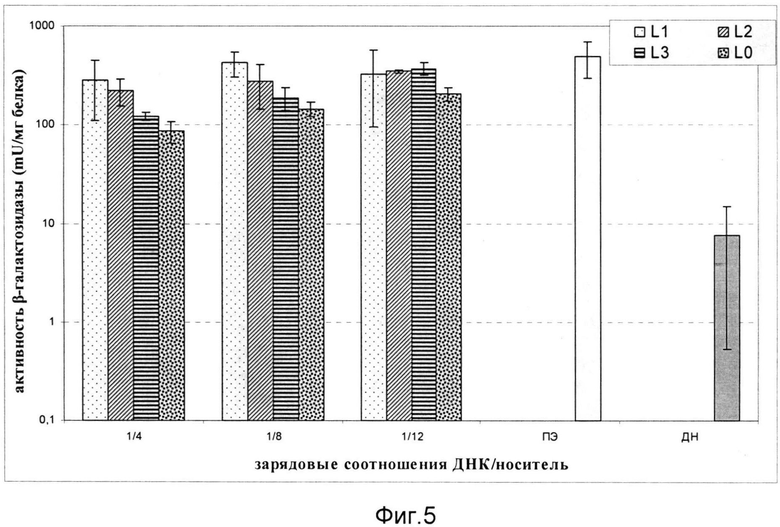
На фиг.5 показана зависимость активности β-галактозидазы в клетках СНО (CXCR4-) после трансфекции комплексами L1/ДНК, L2/ДНК и L3/ДНК. Использованы комплексы, сформированные при следующих зарядовых соотношениях носитель/ДНК: 4/1, 8/1, 12/1. В качестве контролей эксперимента служили интактная молекула ДНК, комплексы ПЭИ/ДНК 8/1 и комплексы, содержащие ДНК и носитель L0. Эффективность доставки маркерного гена носителями L1, L2 и L3 достоверно не отличалась от таковой с использованием контроля L0.
Носители без последовательности-лиганда неспособны обеспечить достоверно высокого по сравнению с контролем уровня доставки генетического материала и клетках без рецептора. В то же время эффективность комплексов ДНК с носителем L0, последовательность которого является частью молекулярного коньюгата, была достоверно выше, чем при использовании интактной плазмиды. Это свидетельствует об эффективности данной последовательности при компактизации ДНК и выходе комплексов из эндосом.
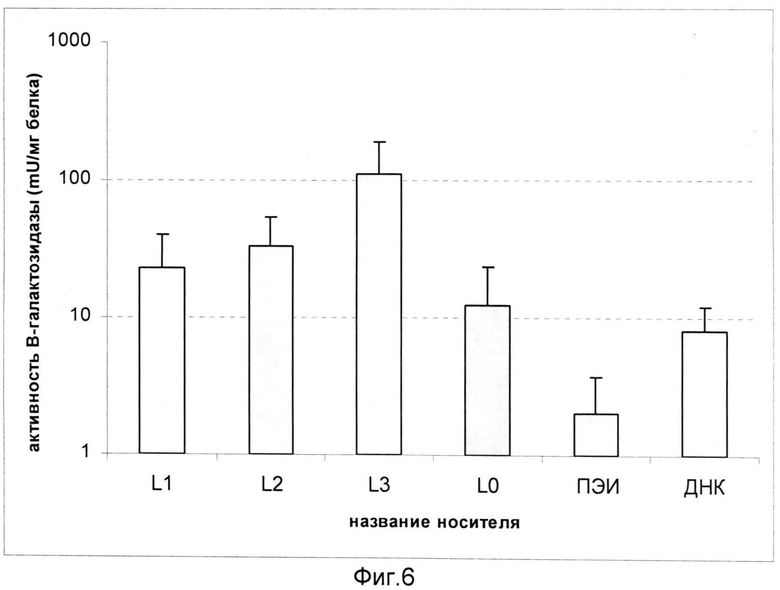
На фиг.6 показана зависимость активности β-галактозидазы в мезенхимных стволовых клетках человека, обработанных вальпроевой кислотой, после трансфекции комплексами L1/ДНК, L2/ДНК и L3/ДНК. Вальпроевая кислота увеличивает количество рецептора CXCR4 на поверхности стволовых клеток (Tsai L-K, Leng Y, Wang Z, Leeds P, Chuang D-M. The Mood Stabilizers Valproic Acid and Lithium Enhance Mesenchymal Stem Cell Migration via Distinct Mechanisms // Neuropsychopharmacology. 2010; 35: 2225-2237). Использованы комплексы, сформированные при зарядовом соотношении носитель/ДНК 4/1. В качестве контролей эксперимента служили интактная молекула ДНК, комплексы ПЭИ/ДНК 8/1 и комплексы, содержащие ДНК и носитель L0. Эффективность доставки маркерного гена носителями L2 и L3 была в 3-9 раз выше, чем контролем L0. Мезенхимные стволовые клетки человека рассматриваются в качестве перспективных агентов для клеточной и генной терапии различных заболеваний человека. В то же время они относятся к одним из наиболее трудно трансфецируемых типов клеток. Природный лиганд рецептора CXCR4 - хемокин SDF-1α, на основе которого разработана последовательность-лиганд в составе носителей, является фактором миграции стволовых клеток (Kucia М, Reca R, Miekus К, Wanzeck J, Wojakowski W, Janowska-Wieczorek A, Ratajczak J, Ratajczak MZ Trafficking of Normal Stem Cells and Metastasis of Cancer Stem Cells Involve Similar Mechanisms: Pivotal Role of the SDF-1-CXCR4 Axis // Stem Cells. 2005; 23: 879-894). Таким образом, включение в состав носителей пептида с лигандом рецептора CXCR4 позволяет повысить эффективность трансфекции мезенхимных стволовых клеток.
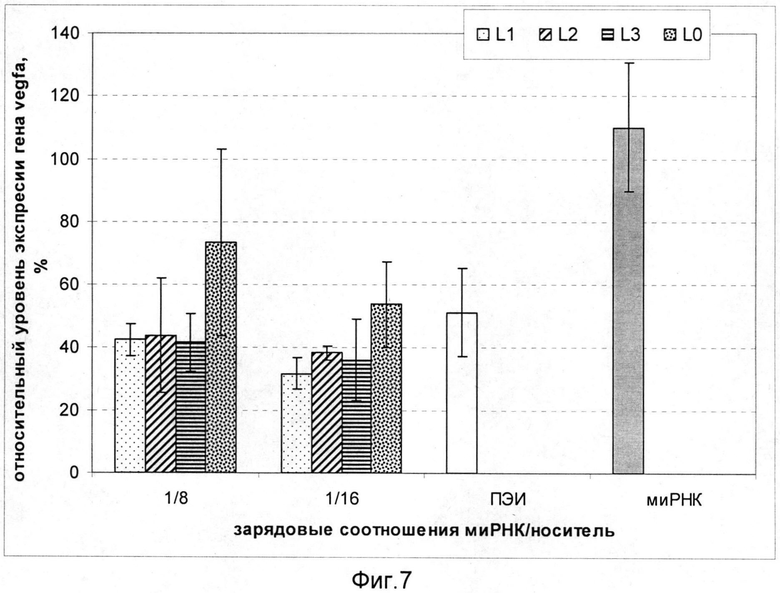
На фиг.7 показано изменение экспрессии гена VEGFA в клетках A172 (CXCR4+) после трансфекции комплексами L1/миРНК, L2/миРНК и L3/миРНК. Использованы комплексы, сформированные при следующих зарядовых соотношениях носитель/миРНК: 8/1, 16/1. В качестве контролей эксперимента служили интактная молекула миРНК, комплексы ПЭИ/миРНК 8/1 и комплексы, содержащие миРНК и носитель L0. При доставке комплексов L1/миРНК, L2/миРНК и L3/миРНК уровень экспрессии гена VEGFA был достоверно ниже по сравнению с использованием комплексов L0/миРНК при соответствующих зарядовых соотношениях. После доставки комплексов L1/миРНК уровень экспрессии составлял 43% и 32% от базального уровня при зарядовых соотношениях 8/1 и 16/1 соответственно. После доставки комплексов L2/миРНК уровень экспрессии составлял 44% и 38%, L3/миРНК - 42% и 36%. Действие «незащищенной» миРНК не приводило к подавлению экспрессии гена VEGFA.
Таким образом результаты на фиг.3, 4, 5, 6, 7 свидетельствуют в пользу специфичности доставки нуклеиновых кислот с помощью молекулярных коньюгатов из настоящего изобретения в клетки, на поверхности которых представлен рецептор CXCR4.
Осуществление изобретения можно пояснить следующим образом. Задача настоящего изобретения состоит в обеспечении эффективной и направленной доставки генетических конструкций в клетки с рецептором CXCR4 на поверхности, используя модульные носители, содержащие последовательность-лиганд к данному рецептору.
На первом этапе проводят образование комплексов одного из воплощений носителя с генетической конструкцией, содержащей ген "интереса", или с миРНК. Сформированные комплексы используют для доставки генетического материала в соответствующие клетки-мишени. Анализ эффективности проникновения в клетки оценивают с помощью ферментативных, иммуногистохимических и молекулярно-биологических методов.
Формирование комплексов проводят в изотоническом растворе. Предпочтительным является бессолевой буфер НВМ (Hepes-buffered mannitol). Размер образующихся комплексов составляет 140-250 нм.
В качестве генетических конструкций в одном из воплощений используют плазмидную ДНК.
Плазмидная ДНК содержит в своем составе маркерный (luc, lacZ) или терапевтический ген (в зависимости от заболевания) под контролем соответствующих промоторов и энхансеров (CMV, SV40 и др.) и другие элементы, необходимые, например, для репликации в клетки-хозяине или интеграции в геном. При генотерапевтичеком лечении раковых заболеваний могут быть использованы гены HLA-В7, IL-2, IL-4, TNF, IFN, Р53, тимидинкиназы и проч.
В другом воплощении в качестве генетической конструкции используют миРНК, комплементарные специфической последовательности в составе мРНК для подавления синтеза белкового продукта. Например, при генотерапевтичеком лечении опухолей могут быть использованы малые интерферирующие РНК, способные снизить экспрессию генов, кодирующих белки VEGFA, VEGFR1, VEGFR2, HGF, PDGF, FGF и др., что может приводить к снижению ангиогенеза и прекращению развития заболевания. Сформированные комплексы используют для доставки генетического материала в клетки с рецептором CXCR4 на поверхности. Проникновение комплексов с носителем из настоящего изобретения происходит преимущественно с помощью рецептор-опосредованного переноса путем связывания с внеклеточными доменами рецептора CXCR4 и последующей интернализацией рецептора.
Доза молекулярных коньюгатов и генетического материала определяется индивидуально и зависит от типа клеток, количества рецептора CXCR4 на их поверхности и сложности трансфецирования данных клеток.
При подборе условий трансфекции они создаются так, чтобы обеспечить наибольшую эффективность доставки. Предпочтительным является инкубация комплексов с клетками в течение 4 часов. Однако можно варьировать это время от 3 до 6 часов. По истечении времени инкубации производят смену среды и оставляют клетки на 24-48 часов (в зависимости от типа клеток и генетического материала) для экспрессии введенных конструкции с геном-интереса или проявления терапевтического эффекта олигонуклеотидов.
Анализ эффективности доставки проводится ферментативными, иммуногистохимическими или молекулярно-биологическими методами в зависимости от типа введенной генетической конструкции.
Пример 1. Формирование комплексов ДНК/носитель, РНК/носитель и изучение процесса комплексообразования.
В качестве генетического материала для направленной доставки генов в клетки были использованы: 1 - плазмида pCMVLacZnls, содержащая ген β-галактозидазы под контролем промотора цитомегаловируса, 2 - малая интерферирующая РНК (миРНК) против гена VEGFA. Использовали одно из воплощений носителя, содержащего последовательность-лиганд к рецептору CXCR4. Носители генетических конструкций формировали с помощью матричной полимеризации в процессе образования нуклеопептидных комплексов. Использовали следующие комбинации пептида, модифицированного лигандом рецептора CXCR4 и катионного пептида CHRRRRRRHC: носитель L1 содержит 100 мол.% пептида, модифицированного лигандом рецептора CXCR4; носитель L2 содержит 50 мол.% пептида, модифицированного лигандом рецептора CXCR4, и 50 мол.% катионного пептида CHRRRRRRHC; носитель L3 содержит 10 мол.% пептида, модифицированного лигандом рецептора CXCR4, и 90 мол.% катионного пептида CHRRRRRRHC; носитель L0 содержит 100 мол.% катионного пептида CHRRRRRRHC.
Приготавливали растворы 0,5 мкг ДНК или РНК в 10 мкл 1X буфера НВМ (5% w/v mannitol, 5 mM Hepes, pH 7.5) и растворы носителя, соответствующие различным зарядовым соотношениям ДНК/носитель или РНК/носитель, в равном объеме буфера. В пробирку-эппендорф с раствором нуклеиновой кислоты постепенно добавляли раствор носителя и интенсивно перемешивали в течение 30 секунд. Полученную смесь оставляли на 2 часа при комнатной температуре для завершения процесса формирования комплексов.
Результаты по комплексообразованию анализируют методом вытеснения бромистого этидия или SybrGreen. Измерение флуоресценции производят с помощью спектрального сканирующего мультирежимного считывающего устройства Wallac 1420D (Thermo, Finland). Вытеснение бромистого этидия наблюдается при излучении 590 нм (возбуждение при 544 нм) после добавления носителя к ДНК (50 мкг/мл), преинкубированной с интеркалирующим агентом бромистым этидием (400 нг/мл). Вытеснение красителя SybrGreen наблюдается при излучении 590 нм (возбуждение при 485 нм) после добавления носителя к РНК (50 мкг/мл), преинкубированной с однократным раствором SybrGreen. Вытеснение было посчитано по формуле (F-Ff)/(Fb-Ff), где Ff и Fb - это интенсивности флюоресценции бромистого этидия/SybrGreen в отсутствие и присутствии ДНК/РНК соответственно. Результаты представлены на фиг.1, 2.
Пример 2. Проведение трансфекции in vitro.
Клетки культуры HeLa, A172 и СНО рассевали на культуральные 48 (при использовании ДНК)/24-луночные (при использовании РНК) планшеты (Nunc) за 24 часа до трансфекции из расчета 50000/100000 клеток на лунку, содержащую 500/1000 мкл стандартной культуральной смеси, состоящей из культуральной среды DMEM (Биолот), 10% сыворотки эмбрионов коров (Биолот), 0,01% раствора гентамицина. Мезенхимные стволовые клетки человека рассевали на культуральные 48-луночные планшеты из расчета 30000 клеток на лунку, содержащую 500 мкл стандартной культуральной смеси (αМЕМ (Биолот), 15% сыворотки эмбрионов коров (Биолот), 0,01% раствора гентамицина), через 24 часа в среду трансфекции добавляли раствор вальпроевой кислоты до конечной концентрации 2,5 мМ, инкубировали при температуре 37°C в течение трех часов. Далее проводили смену среды в лунках и оставляли планшет на 48 часов.
Суспензию комплексов приготавливали согласно методике, описанной в примере 1, из расчета 2 мкг ДНК/2,7 мкг РНК на каждую лунку культурального планшета. За 10 минут до внесения суспензии комплексов НК/носитель клетки несколько раз промывали средой DMEM/αMEM и вносили в каждую лунку по 500/1000 мкл среды DMEM/αMEM. Трансфекцию проводили путем добавления суспензии комплексов НК/носитель в среду. После внесения комплексов планшеты с клетками помещали в термостат с температурой 37°C и 5% содержанием CO2 на 4 часа. По прошествии времени инкубации клетки промывали средой DMEM/αMEM и вносили в каждую лунку по 500/1000 мкл стандартной культуральной смеси. Культуральный планшет инкубировали в термостате при температуре 37°C и 5% содержанием CO2 в течение 48 часов.
Пример 3. Выявление экспрессии гена β-галактозидазы после трансфекции in vitro комплексами ДНК/носитель.
Удаляли среду из культуральный планшетов, промывали клетки в 1×PBS (pH 7.2). В каждую лунку добавляли по 80 мкл лизис буфера (25 мМ Gly-Gly, 15 мМ MgSO4, 4 мМ EGTA, 1 мМ DTT, 1 мМ PMSF; pH 7.8). По 50 мкл лизата переносили в полистироловые планшеты для измерения активности β-галактозидазы. Добавляли в каждую лунку по 50 мкл реакционного раствора 4-метилумбелиферон-β-галактопиранозид - MUG (реакционный раствор MUG: 200 мМ Na-фосфатного буфер (pH 7,3), 2 мМ MgCl2, 1 мг/мл MUG (Sigma)). Инкубировали 1,5 часа при температуре 37°C. Добавляли в каждую лунку по 100 мкл стоп - реагента (0,2 М глицин (pH 10,0)). Измерение проводили с помощью спектрального сканирующего мультирежимного считывающего устройства Wallac 1420D (Thermo, Finland) при длине волны излучения 355 нм и возбуждения 460 нм. Время экспозиции составляло 0,1 секунды. В качестве контроля использовали фермент β-галактозидазу с известной активностью (mU). Строили калибровочную кривую зависимости активности фермента от интенсивности флуоресценции. На основании калибровочной кривой получали активность β-галактозидазы в пробах. Результаты эксперимента оценивали в относительных световых единицах на 1 мг тотального белка из клеточных экстрактов в лунке культурального планшета. Общее количество белка в каждой лунке измеряли методом Бредфорд относительно калибровочной кривой по бычьему сывороточному альбумину. Результаты представлены на фиг.3, 4, 5, 6.
Пример 4. Выявление экспрессии гена VEGFA после трансфекции in vitro комплексами миРНК/носитель.
Из лунок планшета удаляли культуральную смесь, после чего 2 раза промывали раствором, содержащим трипсин и версен в соотношении 1:3. Добавляли 150 мкл раствора трипсина и версена на лунку и инкубировали при температуре 37°C в течение 10 минут. Добавляли 150 мкл 1×PBS, центрифугировали 10 мин при 2200 оборотах в минуту, сливали жидкую фракцию. Далее проводили выделение РНК с помощью раствора Trizol (Quiagen, Германия) согласно протоколу фирмы-производителя. Для синтеза кДНК смешивали 5 мкл РНК, 1 мкл гексопраймеров (15 OE/мл) и 15 мкл H2O, оставляли на 5 мин при 70°C, после чего добавляли 2,5 мкл буфера для обратной транскриптазы, 4 мкл дезоксирибонуклеотидов (2 мМ), 0,5 мкл обратной транскриптазы (200 ед. акт/мкл). Проводили амплификацию при следующих условиях: 10 мин при температуре 25°C, 60 мин при 37°C, 10 мин при 70°C. Измерение концентрации кДНК проводили на спектрофотометре NanoDrop (США). Для анализа экспрессии гена VEGFA проводили реакцию ПЦР в реальном времени использовали набор (ЗАО «Синтол», Москва), в состав которого входили 10x Eva Green, смесь дезоксирибонуклеотидов (2,5 мМ), MgCl2 (25 мМ), Taq-полимераза (5 ед./мкл), dH2O. Использовались следующие последовательности праймеров: F'-5TGCCGACAGGATGCAGAAG3', R-5'GCCGATCCACACGGAGTACT3' (к гену β-актина); F-5'CAACATCACCATGCAGATTATGC3', R-5'CCCACAGGGATTTTCTTGTCTT3' (к гену VEGFA). Амплификацию проводили в термоциклер Rotor-Gene 3000 (Corbett Research, Австралия). Полученные данные анализировали с помощью программного обеспечения Rotor-Gene version 6.1.71 (Corbett Research, Австралия). Были получены значения порогового цикла флуоресценции (Ct) для каждого образца в двух повторностях. Были построены две калибровочные кривые для исследуемого и референсного гена методом линейной регрессии. Далее с помощью метода относительного количественного анализа (Relative Quantitation), рассчитывали относительную концентрацию для каждого образца. Уровень экспрессии гена VEGFA выражали относительно уровня экспрессии гена β-актина.
Результаты представлены на фиг.7.
Настоящее изобретение может быть использовано для направленной доставки генетического материала с целью коррекции генных дефектов, придания клеткам новых функций и лечения заболеваний. Наличие экспрессии CXCR4 более чем в 20 видах раковых опухолей позволяет говорить о важности данного изобретения для генной терапии ряда онкологических заболеваний. В число таких заболеваний входят рак груди, кишечника, легких, печени, желудка, печени, лейкемии и лимфомы. Специфическая доставка суицидных генов, генов-супрессоров опухоли, интерферирующих РНК приведет к избирательной гибели клеток, в которые проникли комплексы с соответствующим генетическим материалом. Рецептор CXCR4 присутствует на поверхности большинства типов стволовых клеток, стволовых клетках гематопоэтического ряда, костного мозга, миелоидных и лимфоидных стволовых клетках. Присутствие рецептора CXCR4 на стволовых клетках, предшественниках мышечных клеток позволяет говорить о возможном использовании данного изобретения для ех vivo терапии различных заболеваний.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ НАПРАВЛЕННОЙ ДОСТАВКИ ДНК В ОПУХОЛЕВЫЕ И СТВОЛОВЫЕ КЛЕТКИ | 2008 |
|
RU2408607C2 |
| МОЛЕКУЛЯРНЫЕ КОНЪЮГАТЫ С ПОЛИКАТИОННЫМ УЧАСТКОМ И ЛИГАНДОМ ДЛЯ ДОСТАВКИ В КЛЕТКУ И ЯДРО КЛЕТКИ ДНК И РНК | 2009 |
|
RU2537262C2 |
| СРЕДСТВО ДЛЯ ВНУТРИКЛЕТОЧНОЙ ДОСТАВКИ НУКЛЕИНОВЫХ КИСЛОТ В КЛЕТКИ МЛЕКОПИТАЮЩИХ | 2014 |
|
RU2572575C1 |
| ЛЕЧЕНИЕ И ПРЕДУПРЕЖДЕНИЕ НЕЙРОДЕГЕНЕРАТИВНЫХ ЗАБОЛЕВАНИЙ С ИСПОЛЬЗОВАНИЕМ ГЕННОЙ ТЕРАПИИ | 2008 |
|
RU2471487C2 |
| Пептиды для внутриклеточной доставки нуклеиновых кислот | 2020 |
|
RU2771605C2 |
| Композиция, подавляющая экспрессию генов интерлейкина-4 и интерлейкина-13, для терапии аллергического ринита | 2018 |
|
RU2710895C1 |
| Способ получения амфифильных блок-сополимеров N,N-диметиламиноэтилметакрилата для доставки нуклеиновых кислот в живые клетки | 2014 |
|
RU2617059C2 |
| Способ получения плазмидной ДНК в составе полимерных наносфер для доставки | 2022 |
|
RU2817116C1 |
| Композиция системы доставки на основе конъюгата для доставки полинуклеотида РНК-интерференции в клетку печени и способ ее получения | 2011 |
|
RU2623160C9 |
| БИОДЕГРАДИРУЕМЫЕ КАТИОННЫЕ ПОЛИМЕРЫ | 2005 |
|
RU2440380C2 |
Изобретение относится к области генетической инженерии, молекулярной биологии и медицины. Предложен носитель для направленной доставки нуклеиновых кислот в клетки, экспрессирующие рецептор CXCR4, состоящий из последовательности-лиганда к рецептору CXCR4 с последовательностью аминокислот KPVSLSYRSPSRFFESH, линкерного участка из двух молекул ε-аминогексановой кислоты, соединяющей последовательность-лиганд с последовательностью для компактизации нуклеиновых кислот, последовательности, обеспечивающей компактизацию нуклеиновых кислот и выход комплекса из эндосом CHRRRRRRHC. Изобретение может быть использовано для направленной доставки генетических конструкций в клетки с рецептором CXCR4 на поверхности, таких как злокачественные опухолевые и стволовые клетки, с целью коррекции генных дефектов, воздействия на процессы реализации генетической информации и предотвращения заболеваний. 2 з.п. ф-лы, 7 ил., 4 пр.
1. Носитель для направленной доставки нуклеиновых кислот в клетки, экспрессирующие рецептор CXCR4, отличающийся тем, что носитель состоит из:
- последовательности-лиганда к рецептору CXCR4 с последовательностью аминокислот - KPVSLSYRSPSRFFESH;
- линкерного участка из двух молекул ε-аминогексановой кислоты, соединяющей последовательность-лиганд с последовательностью для компактизации нуклеиновых кислот;
- последовательности, обеспечивающей компактизацию нуклеиновых кислот и выход комплекса из эндосом - CHRRRRRRHC.
2. Носитель по п.1, отличающийся тем, что в качестве генетического материала для носителей используют плазмидную ДНК.
3. Носитель по п.1, отличающийся тем, что в качестве генетического материала используют олигонуклеотидные последовательности РНК и ДНК, интерферирующие РНК.
| US 20070111201 A1, 17.05.2007 | |||
| US 20040110923 A1, 10.06.2004 | |||
| СПОСОБ НАПРАВЛЕННОЙ ДОСТАВКИ ДНК В ОПУХОЛЕВЫЕ И СТВОЛОВЫЕ КЛЕТКИ | 2008 |
|
RU2408607C2 |

