Изобретение относится к области биотехнологии, в частности к генной инженерии, и может быть использовано в медицине для лечения патологий, связанных с избыточным ангиогенезом, включая рост злокачественных опухолей, ревматоидный артрит, псориаз и диабетическую ретинопатию. Предлагается способ, позволяющий локально подавлять рост кровеносных сосудов путем введения в области избыточного ангиогенеза эффективного количества протеолитически неактивных рекомбинантных форм активатора плазминогена урокиназного типа (uPA), конкурентно подавляющих протеолитическое действие нативного активатора плазминогена урокиназного типа и пространственно разобщающих рецепторные белки, участвующие в реализации его сигнальных эффектов.
Введение
В процессе инициации ангиогенеза необходимым моментом является дестабилизация существующих сосудов, включающая в себя разрушение межклеточных контактов эндотелиальных клеток, деградацию белков базальной мембраны и внеклеточного матрикса, последующую миграцию эндотелиальных клеток и формирование новых сосудов. Урокиназа играет ключевую роль в начальных этапах ангиогенеза, обеспечивая как околоклеточный протеолиз, так и стимуляцию миграции клеток сосудистой стенки. Урокиназа и активированный ею плазмин непосредственно участвуют в деградации таких белков базальной мембраны, как фибронектин и ламинин, а также вызывают повышение экспрессии и последующую активацию матриксных металлопротеиназ. Урокиназа стимулирует миграцию клеток сосудистой стенки, а также протеолитическую активацию таких факторов роста, как VEGF и HGF, которые стимулируют пролиферацию и миграцию эндотелиальных клеток [1-5]. Так, повышение экспрессии урокиназы в эндотелиальных клетках увеличивает их инвазивный потенциал. Урокиназа участвует в индукции ангиогенеза в сетчатке глаза под действием HGF [6], а также в стимуляции роста кровеносных сосудов в ишемизированном миокарде, вызванной действием VEGF [7].
Напротив, блокирование взаимодействия урокиназы с ее рецептором подавляет ее ангиогенные эффекты. В природе это может происходить за счет формирования так называемого «связывающего пептида», который образуется в результате протеолитического расщепления урокиназы плазмином или металлопротеиназами в положениях Lys135-Lys136 или Glu143-Leu144. «Связывающий пептид» блокирует взаимодействие урокиназы с ее рецептором, что вызывает подавление миграции эндотелиальных клеток и ангиогенеза in vivo, стимулированных ангиогенными факторами роста [8]. Этот же пептид способен блокировать инвазивные свойства опухолей и замедлять их рост и метастазирование за счет подавления ангиогенеза [9].
In vitro было показано, что добавление антител, блокирующих урокиназный рецептор, в среду культивирования ингибирует формирование капилляроподобных структур [10, 11]. Разрушение комплекса uPA-uPAR ингибирует миграцию и пролиферацию эндотелиальных клеток сетчатки, которые были индуцированы HGF. Более того, нокаут гена урокиназы проявляется в снижении роста кровеносных сосудов в ишемизированных задних конечностях мышей [12], значительно худшем восстановлении миокарда после инфаркта, а также снижении эффекта в ответ на лечение VEGF.
Таким образом, уже на основе теоретического анализа вопроса можно сделать заключение, что процесс ангиогенеза можно подавлять, воздействуя на протеолитическую активность урокиназы, а также на ее взаимодействие с рецептором.
Уровень техники
По данным анализа литературных источников первое свидетельство, подтвердившее возможность подавления прогрессирования опухоли ингибированием активности урокиназы, было получено с использованием антител к урокиназе [13, 14]. Впоследствии усилия в этой области были направлены на разработку низкомолекулярных синтетических ингибиторов uPA, обладающих высокой эффективностью, селективностью и фармакокинетическими свойствами, позволяющими их использовать как лекарственные препараты для противоопухолевой терапии. Дизайн ингибиторов трипсиноподобных ферментов базировался на структурном анализе и отталкивался от активного центра трипсина, специфика связывания которого в значительной степени определяется остатком Asp 189, расположенным в основе первичного якорного участка для субстратов, названного специфичным карманом S1 [15]. Кристаллическая структура каталитического домена uPA обладает подобной трипсину топологией, в которой Asp189 сохранен, придавая участку S1 афинность к положительно заряженным остаткам Arg и Lys [16]. Поэтому большинство синтетических ингибиторов uPA, созданных к настоящему времени, обладают общей структурной особенностью, заключенной в моно- или биароматической части, обладающей функцией амидина или гуанидина и действующей как миметик аргинина. Однако в выборе компонентов существует строгое ограничение, обусловленное необходимостью ингибирования урокиназы, не затрагивая активность других подобных трипсину сериновых протеаз и особенно tPA и плазмина - ключевых ферментов фибринолиза.
В самых ранних исследованиях селективными ингибиторами урокиназы считались паразамещенные производные бензамидина, а также 4-хлоро- и 4-трифлюорометилфенилгуанидины и амилорид, но ни один из них не обладал высокой эффективностью, поскольку у всех у них Ki находился в пределах микромолярного диапазона [17]. Когда структура амилорида была взята за отправную точку, поиск более мощных ингибиторов привел к синтезу двух бензо[b]тиофен-2-карбоксамидинов с 4 замещениями, B428 и B623 с Ki 0.53 и 0.16 микроМ соответственно [18]. Противоопухолевые свойства B428 и B623 были проверены в нескольких работах, которые продемонстрировали способность данных ингибиторов подавлять в культуре клеток процессы, опосредованные действием урокиназы. In vivo же упомянутые ингибиторы проявляли антипролиферативный и противоинвазивные эффекты при их испытании на моделях опухолевых ксеноплантов [19-22]. Для ингибитора B428 было проведено исследование с целью определения структурных детерминант его афинности и селективности к молекуле урокиназы и другим трипсиноподобным сериновым протеазам [23, 24]. Интересно, что, несмотря на кристаллографические данные, свидетельствующие о высоком подобии S1 участков uPA и tPA, значительная специфичность к молекуле uPA может быть достигнута через взаимодействие ингибиторов уже на одном только участке S1. Фактически позиция 190 в tPA тромбине и факторе Ха занята аланином, тогда как в uPA и трипсине - серином; именно последний способен создавать дополнительную водородную связь с амидиновыми ингибиторами, придавая им более высокую степень афинности к молекуле uPA по сравнению с молекулой tPA. Другие элементы аминокислотной последовательности в участке S1 и во фланговых областях вносят дополнительные структурные различия, которые все более дифференцируют S1 участки трипсиноподобных сериновых протеаз, придавая уникальность структуры каждому из ферментов, и тем самым предоставляют возможность для создания новых специфических ингибиторов. Кроме того, точность взаимодействия uPA с ингибиторами зависит от малых карманов и/или субсайтов фермента, которые могут быть использованы как дополнительные мишени для оптимизации структуры проектируемых ингибиторов. Дело обстоит именно так для S1 бета- и S1′ субсайтов, которые были использованы для конструирования 8- и 6-замещенных 2-нафтамидинов благодаря доступности 8-й и 6-й позиции 2-нафтамидина соответственно [25-27]. В последние несколько лет было предложено много новых низкомолекулярных ингибиторов урокиназы, включая производные 4-аминоарилгуанидина и 4-аминобензамидина, 2-пиридинилгуанидины, 1-изохинолинилгуанидины и 1-(7-сульфонамидоизохинолинил)-гуанидины, производные мексилетина, содержащие терминальный бром 4-хлоро-3-алкоксизокумарины, 4-оксазолидиноновый аналон UK122, производные 5-тиометилтиофенеамидина, N-1-адамантил)-N′-(4-гуанидинобензил) мочевина (WX-293T) и WX-UK1 (производное 3-амидинофенилаланина) [27-35]. Однако среди этих ингибиторов только WX-UK1 (WILEX, Мюнхен, Германия) дошел до стадии клинических испытаний. WX-UK1, обладая Ki к человеческой урокиназе в 0.6 микроМ, значительно уменьшал инвазию и распространение раковых клеток, а также рост первичной опухоли на моделях туморогенеза in vivo [36-38]. После этих доклинических результатов в I фазе исследований были определены максимально допустимые дозы при назначении WX-UK1: одного или в комбинации с капецитабином (XelodaTM) - пропрепаратом цитостатика 5-флюорурацила для перорального приема. Результаты исследования показали, что WX-UK1 (сам по себе) безопасен и нетоксичен в диапазоне протестированных доз. Кроме того, когда WX-UK1 назначался в комбинации с капецитабином, никаких достоверных изменений в частоте или интенсивности побочных явлений, типичных для капецитабина, не наблюдалось. В это испытание I фазы вошли 25 пациентов с прогрессирующими солидными опухолями. В результате лечения у нескольких из них течение заболевания стабилизировалось, а у трех пациентов (у двух из которых был метастазирующий рак молочной железы) наблюдали частичный ответ. Эти результаты особенно примечательны, поскольку у этих трех пациентов на момент включения в исследование болезнь имела прогрессирующее течение. Так как WX-UK1 не всасывался в желудочно-кишечном тракте, Wilex разработал пропрепарат для перорального приема - WX-671 - для системной доставки активного WX-UK1. После перорального приема WX-671 расщепляется до активного ингибитора WX-UK1 конститутивными внутриклеточными редуктазами. Эффективность этого пропрепарата в сочетании с классическими цитостатиками в настоящее время оценивается во II фазе двух клинических испытаний. В одно из исследований вошли пациенты с локально прогрессирующим неметастазирующим неоперабельным раком поджелудочной железы. Время появления первого метастаза и выживание без прогрессирования болезни являлись главными конечными точками. Пациенты проходят лечение цитостатиком Гемцитабином: только им или в комбинации с WX-671. Второе исследование представляет собой двойное слепое мультицентровое клиническое испытание II фазы, включившее в себя пациентов с метастазирующим раком молочной железы. Это исследование позволит оценить эффективность комбинированной терапии WX-671 и капецитабином по сравнению с монотерапией капецитабином.
Другой класс селективных ингибиторов урокиназы представлен пептидными молекулами, идентифицированными с использованием фаговых библиотек. Экотин, димерный ингибитор сериновых протеиназ широкой специфичности, обнаруженный в периплазме Е.coli, использовался для построения фаговых библиотек, экспрессирующих варианты экотина, и те, которые обладали высокой аффинностью к урокиназе, были отобраны; затем, используя смешанный комбинированный дизайн структуры экотина и скрининг фаговых библиотек экотина, были получены новые пептидные производные экотина с более высокой активностью и селективностью в отношении uPA [39-41]. В похожем исследовании с использованием случайных фаговых пептидных последовательностей и урокиназы в качестве мишени была изолирована ограниченная дисульфидным мостиком последовательность CSWRGLENHRMC. Этот пептид, названный упаин-1 (upain-1), связывается как с активным участком, так и с поверхностными петлями uPA и обладает высокой специфичностью ингибирования молекулы урокиназы [42]. Более того, на основе минимальных (от ди- до тетрапептидных) последовательностей были созданы ингибиторы-пептидомиметики. На участке связывания они содержат группы аргинина, бензамидина или фенилгуанидина и формируют короткую антипараллельную бета-структуру с сегментом фермента от Ser214 до Gly216, которая воспроизводит способ связывания ингибиторов PAIs [43-46]. У одного из этих ингибиторов - бензилсульфонил-D-Ser-Ser-амидинобензиаламида (с Ki 20 нм) была выявлена значительная антиметастатическая активность и увеличение выживания на модели ксенопланта фибросаркомы [46].
Также известны различные природные и синтетические соединения, обладающие способностью подавлять процесс образования новых сосудов. Их действие обычно направлено либо на ингибирование ангиогенных факторов роста (антитела против VEGF, растворимые формы рецепторов VEGF и FGF), либо на подавление чувствительности эндотелиальных клеток к факторам роста (сурамин, пентозан), либо на блокирование проведения внутриклеточных сигналов или на подавление пролиферации и миграции эндотелиальных клеток. Однако круг этих соединений сегодня весьма ограничен.
До настоящего времени большинство созданных ингибиторов uPA действуют по конкурентному и обратимому механизму; однако недавно были созданы необратимые ингибиторы на основе дифенилфосфоната. Группа дифенилфосфоната реагирует с сериновым остатком активного центра и образует устойчивый фосфонилированный и необратимо ингибированный фермент. Такая группа была включена в структуру аргинилтрипептида или в ароматические аналоги аргинина, кроме того, были внесены дополнительные структурные модификации для оптимизации ингибирующих свойств [47; 48].
Несмотря на многочисленность лекарственных кандидатов, нацеленных на ингибирование урокиназы и ее рецепторов, до сих пор ни одно из соединений с успехом не преодолело доклинических и клинических испытаний. Упомянутые кандидаты проявляют либо недостаточную селективность, либо неприемлемую токсичность.
В связи с этим нами была предпринята попытка разработать способ подавления ангиогенеза, лишенный недостатков, характерных для использования известных соединений-ингибиторов проангиогенного действия системы активатора плазминогена урокиназного типа.
Раскрытие изобретения
При разработке настоящего изобретения во внимание были приняты следующие факты.
Первое. Для подавления роста сосудов необходимо подавление клеточной миграции и пролиферации, а также формирования капилляров. Урокиназа стимулирует клеточную миграцию, пролиферацию и формирование капилляров.
Второе. Урокиназа стимулирует рост сосудов эффективнее общепризнанного стимулятора ангиогенеза - фактора роста сосудистого эндотелия, который обладает более широким, чем предполагалось ранее, спектром действия и, соответственно, нежелательными побочными эффектами при использовании для стимуляции роста кровеносных сосудов.
Третье. Помимо стимуляции эндотелиальных клеток, для формирования кровеносных сосудов требуется ремоделирование внеклеточного матрикса. Урокиназа, активируя плазмин и матриксные металлопротеиназы, также обеспечивает необходимое ремоделирование матрикса.
Четвертое. Разные домены урокиназы опосредуют ее разные проангиогенные эффекты. Условием реализации проангиогенных эффектов урокиназы является совместно локализованная активность доменов урокиназы.
С учетом этого решение поставленной задачи предполагало:
а) создание генно-инженерных конструкций, включающих модифицированные последовательности гена урокиназы (uPA); наработку и очистку рекомбинантных белков на основе структуры урокиназы;
б) разработку in vitro и in vivo моделей для оценки влияния рекомбинантных белков на основе урокиназы на процессы миграции, формирования капилляров и роста сосудов;
в) подбор оптимальных концентраций и времени регистрации эффектов рекомбинантных белков на основе структуры урокиназы для выявления их влияния на процесс ангиогенеза;
г) оценку эффективности подавления рекомбинантными белками на основе структуры урокиназы миграции эндотелиальных клеток, формирования капилляров и роста сосудов на моделях in vitro и in vivo.
При осуществлении изобретения впервые были использованы рекомбинантные плазмиды, содержащие последовательности кДНК рекомбинантных белков на основе структуры урокиназы (аминокислотная последовательность протеолитически неактивной урокиназы и аминокислотная последовательность рекомбинантного белка на основе крингл-домена урокиназы.
Были получены рекомбинантные формы урокиназы: полноразмерная протеолитически неактивная урокиназа и крингл-домен урокиназы. С помощью использования антител к нативным фрагментам структуры урокиназы и рекомбинантного рецептора урокиназы (uPAR) подтвердили, что структура рекомбинантных форм урокиназы и их связывание с рецепторами идентично структуре и связыванию с рецептором нативной урокиназы (пример 1).
На основании полученных данных в дальнейших экспериментах по изучению способности рекомбинантных форм урокиназы подавлять избыточный ангиогенез нами были использованы рекомбинантные формы урокиназы, сходные по своей структуре с нативной молекулой урокиназы.
На адекватных in vitro и in vivo животных моделях было получено экспериментальное подтверждение того, что рекомбинантные молекулы урокиназы способны в значительной степени подавлять миграцию эндотелиоцитов, формирование микротрубочек (аналог капилляров in vitro), а также рост капилляров и сосудов среднего калибра в имплантате Матригеля™. Миграцию эндотелиоцитов в большей степени подавлял крингл-домен урокиназы (пример 3), в то время как полноразмерная протеолитически неактивная урокиназа в большей степени подавляла формирование микротрубочек in vitro и рост капилляров и сосудов среднего калибра на модели опухолевого ангиогенеза in vivo (примеры 4 и 5).
Впервые для подавления избыточного ангиогенеза были применены рекомбинантные формы урокиназы: полноразмерная протеолитически неактивная урокиназа и крингл-домен урокиназы. При этом было установлено, что сочетанное использование крингл-домена и протеолитически неактивной урокиназы более эффективно подавляет рост капилляров и сосудов среднего калибра в имплантате Матригеля™, чем использование только одного из них.
Таким образом, в результате создания настоящего изобретения разработан способ подавления нежелательного (избыточного) ангиогенеза, сущность которого состоит в локальном использовании терапевтически эффективных количеств:
(а) от 50 нМ до 0,5 мкМ рекомбинантной молекулы полноразмерной протеолитически неактивной урокиназы с аминокислотной последовательностью SEQ ID NO: 1;
(б) от 50 нМ до 0,5 мкМ рекомбинантной молекулы крингл-домена урокиназы с аминокислотной последовательностью SEQ ID NO: 2;
(в) эквимолярной (1:1) комбинации рекомбинантных форм урокиназы, охарактеризованных в (а) и (б).
При этом основным техническим результатом является повышение эффективности способа: при использовании моделей in vitro и in vivo удалось добиться значительного снижения степени миграции эндотелиоцитов, снижения формирования микротрубочек in vitro, а также уменьшения числа формирующихся капилляров и сосудов среднего калибра в сравнении с контрольными опытами, в которых использовалось эквивалентное количество полноразмерной урокиназы дикого типа. Дополнительным результатом данного способа следует считать отказ от применения для подавления избыточного ангиогенеза небезопасных синтетических ингибиторов сериновых протеаз.
Краткое описание чертежей
Фигура 1. Электрофорез препаратов белков в ДСН - 14% ПААГ. Гель окрашивали раствором Кумасси R-250. 1-3 - в неденатурирующих условиях: 1. крингл-домен; 2. неактивная урокиназа; 3. полноразмерная урокиназа; 4-6 - в денатурирующих условиях: 4. полноразмерная урокиназа; 5. неактивная урокиназа; 6. крингл-домен.
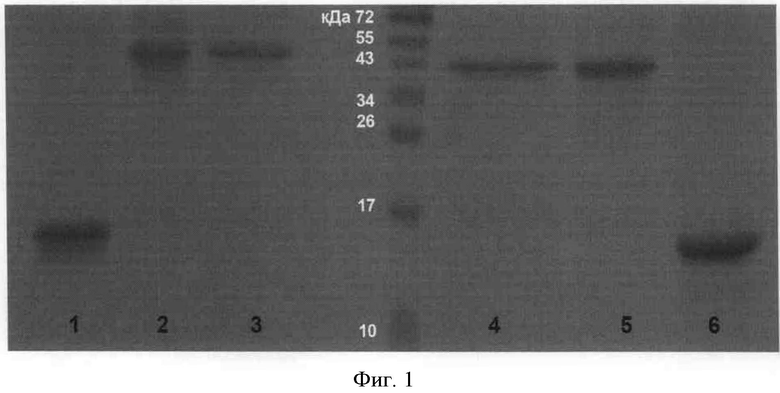
Фигура 2. Иммунохемилюминисцентное окрашивание полученных препаратов рекомбинантных форм урокиназы, разделенных электрофорезом в ПААГ в неденатурирующих условиях. 1 - полноразмерная урокиназа; 2 - протеолитически неактивная урокиназа; 3 - крингл-домен; 4 - полноразмерная урокиназа; 5 - протеолитически неактивная урокиназа; 6 - крингл-домен.
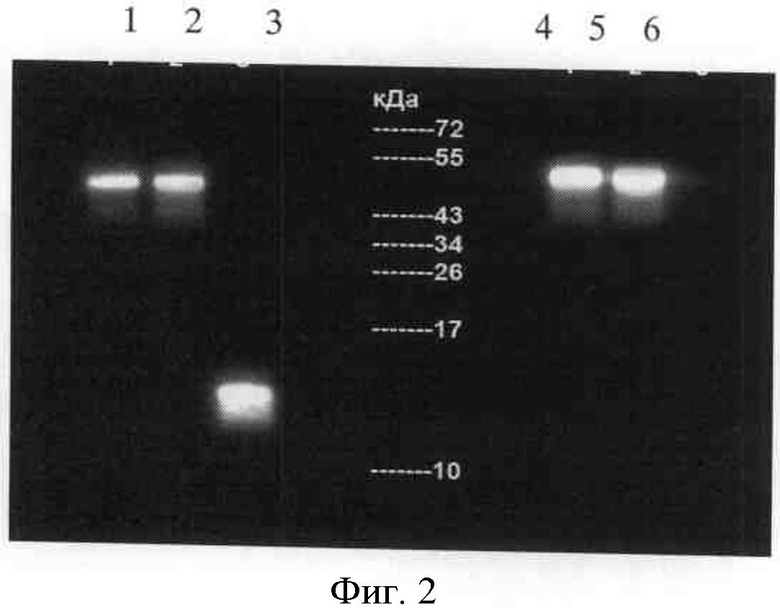
Фигура 3. Протеолитическая активность рекомбинантных форм урокиназы. Протеолитическую активность оценивали с использованием в качестве субстрата синтетического пептида пиро-Glu-Gly-Arg-pNA (S-2444, Sigma).
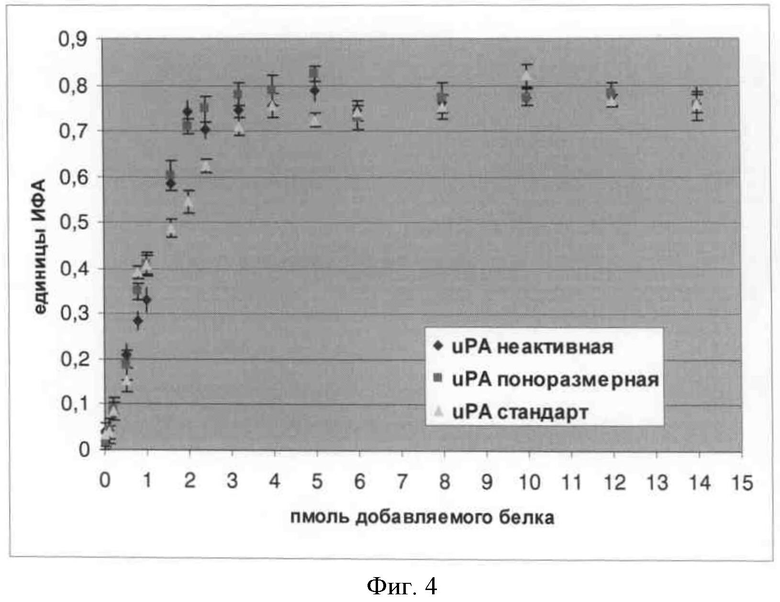
Фигура 4. Связывание рекомбинантных форм урокиназы со специфичным рецептором урокиназы.
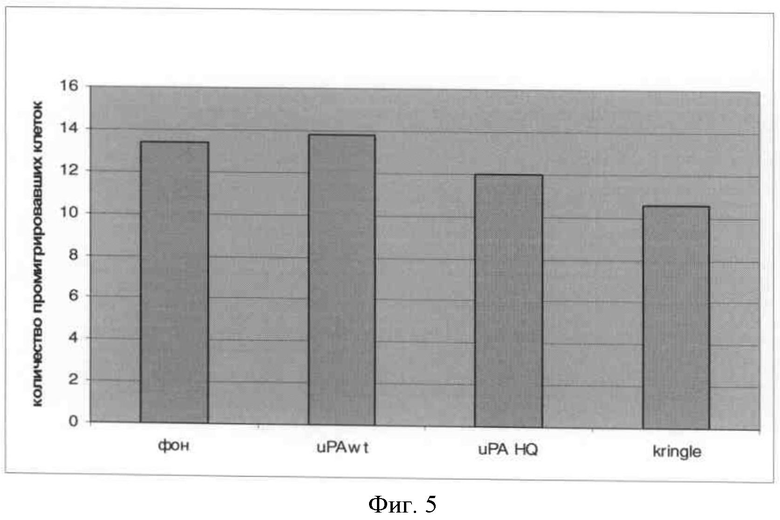
Фигура 5. Миграция эндотелиальных клеток в присутствии рекомбинантных белков на основе урокиназы.
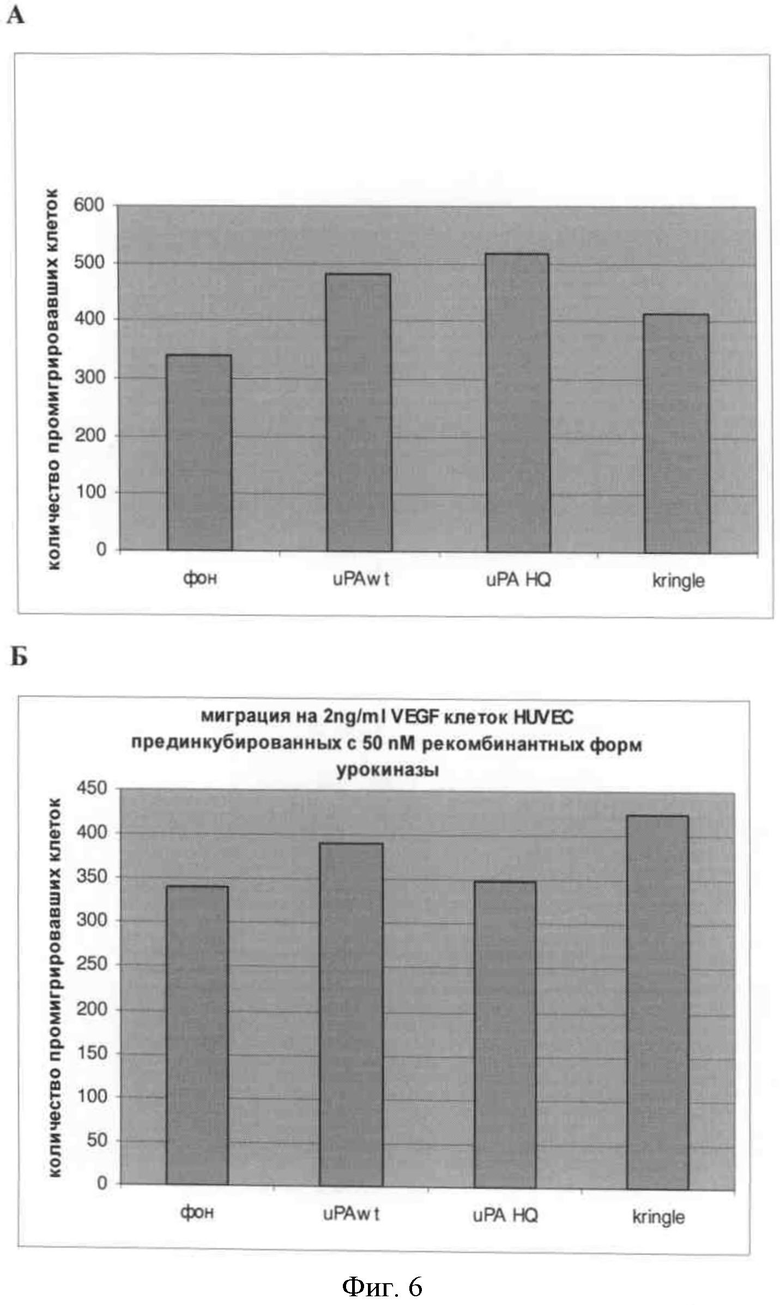
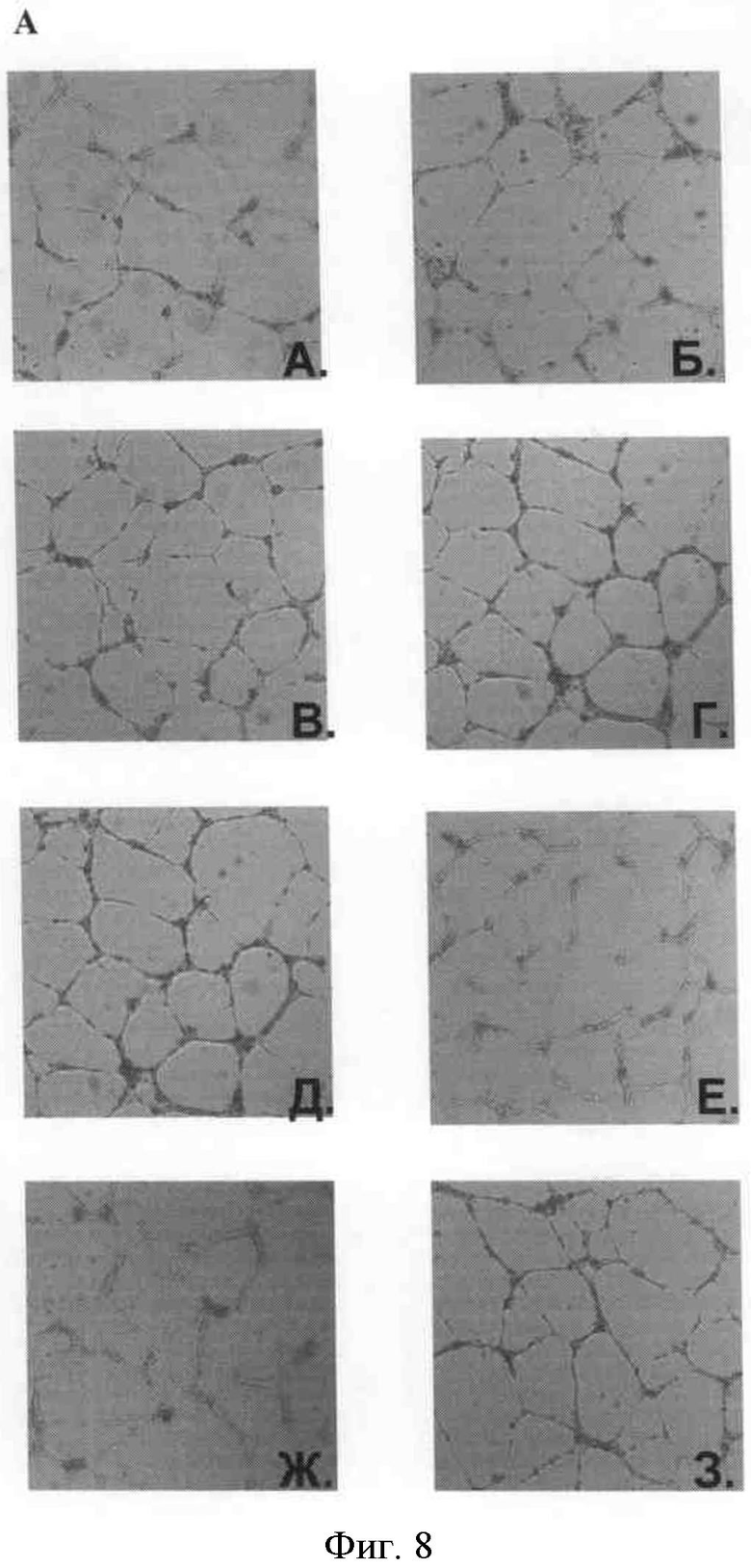
Фигура 6. Формирование капилляроподобных структур в присутствии рекомбинантных форм урокиназы. A) На рисунке слева показаны сформированные капилляроподобные структуры, стимулированные: A. 5% фетальной сывороткой быка; Б. 15% фетальной сывороткой быка; В. VEGF 1 нг/мл; Г. VEGF 2 нг/мл; Д. VEGF 5 нг/мл; Е. VEGF 2 нг/мл в присутствии 150 нМ крингл-домена; Ж. VEGF 2 нг/мл в присутствии 150 нМ протеолитически неактивной урокиназы; З. VEGF 2 нг/мл в присутствии 150 нМ нативной формы урокиназы.
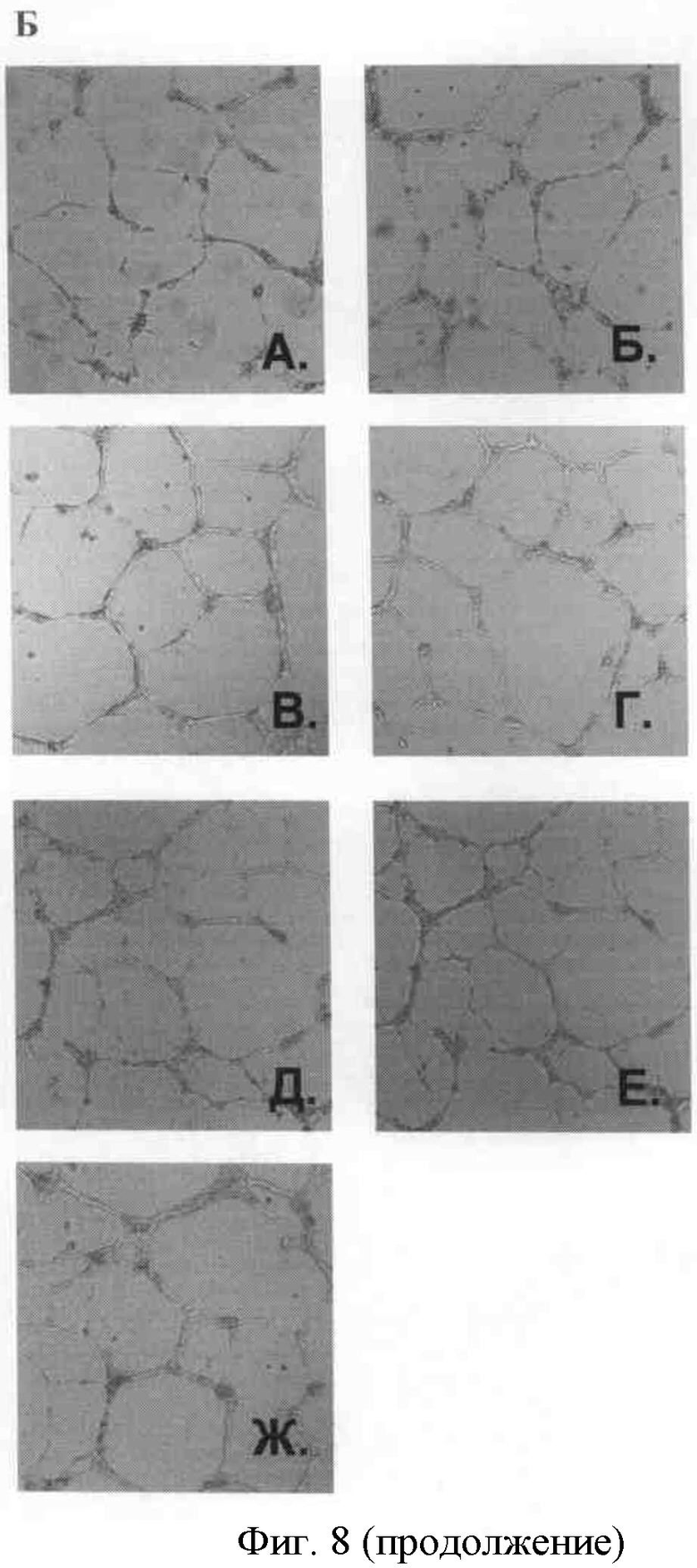
Б) На рисунке показаны сформированные капилляроподобные структуры, стимулированные: А. 5% фетальной сывороткой быка; Б. 15% фетальной сывороткой быка; В. bFGF 10 нг/мл в присутствии 150 нМ крингл-домена; Г. bFGF 10 нг/мл в присутствии 150 нМ протеолитически неактивной урокиназы; Д-Е. bFGF 10 нг/мл в присутствии 150 нМ полноразмерной протеолитически активной урокиназы.
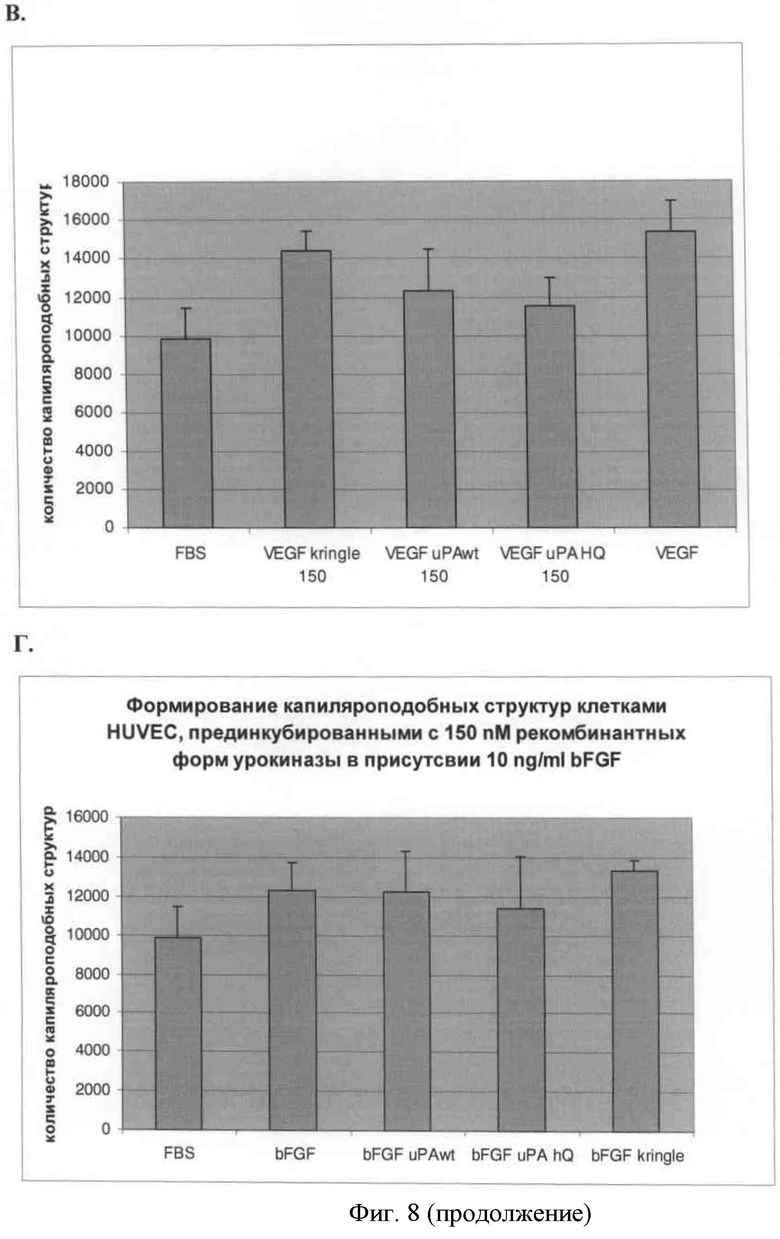
В) Формирование капилляроподобных структур эндотелиальными клетками в присутствии 2 нг/мл фактора роста сосудистого эндотелия и 150 нМ урокиназы и рекомбинантных белков на основе урокиназы.
Г) Формирование капилляроподобных структур эндотелиальными клетками в присутствии 10 нг/мл основного фактора роста фибробластов и 150 нМ урокиназы и рекомбинантных белков на основе урокиназы.
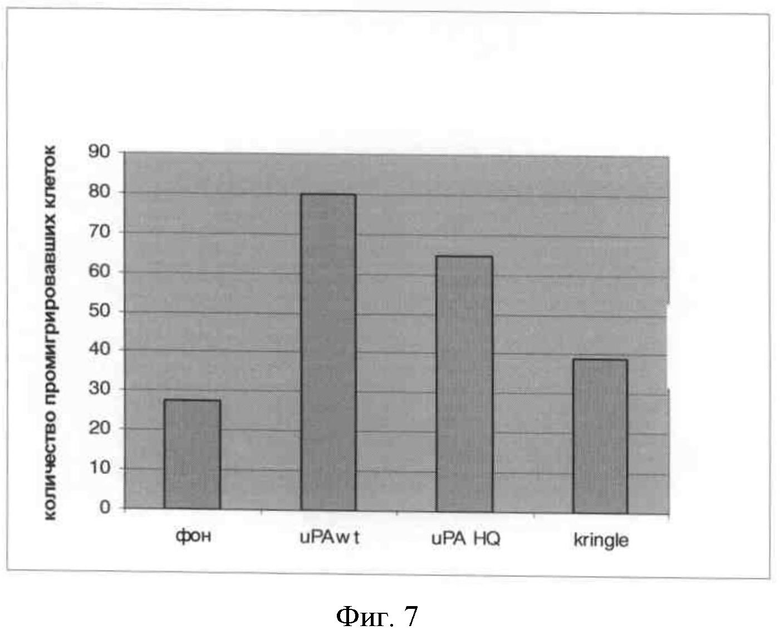
Фигура 7
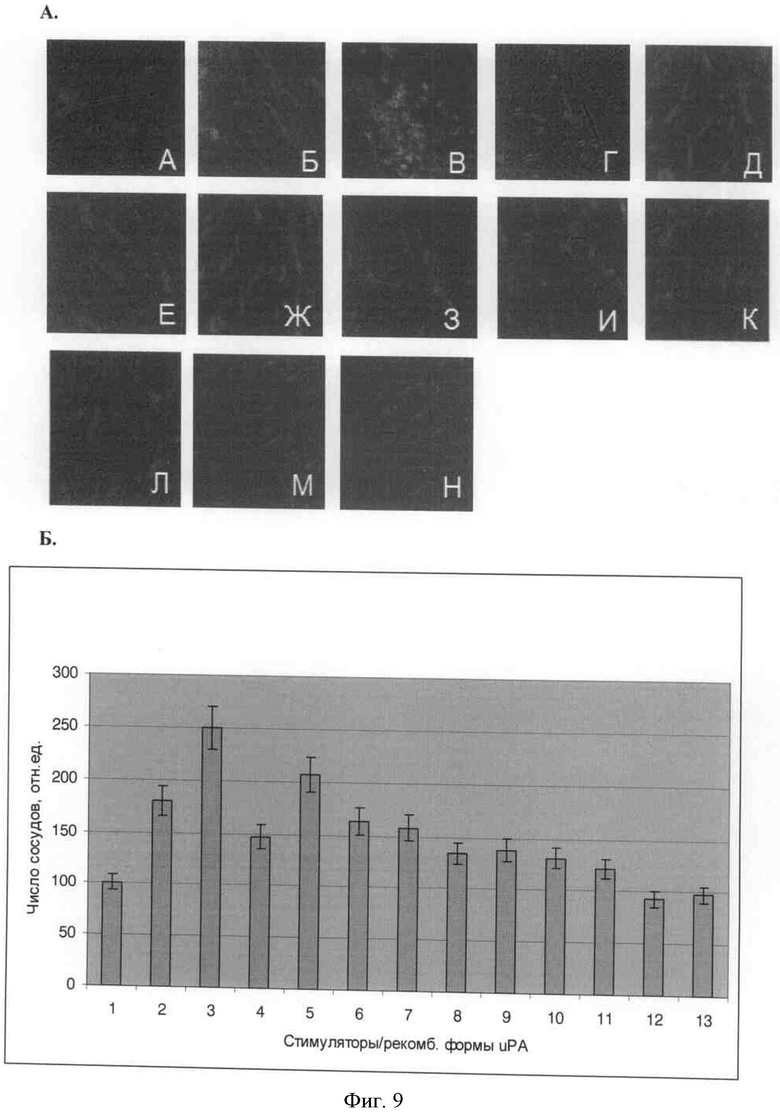
А) Оценка формирования сосудов в имплантах Матригеля™, модель in vivo. Синим цветом окрашены ядра, красным - формирующиеся сосуды: А - без дополнительных стимулов; Б - фактор роста сосудистого эндотелия, 20 нг/мл; В - фактор роста сосудистого эндотелия, 50 нг/мл; Г - основный фактор роста фибробластов, 10 нг/мл; Д - основный фактор роста фибробластов, 20 нг/мл; Е - фактор роста сосудистого эндотелия, 20 нг/мл + протеолит. неакт. урокиназа, 50 нМ; Ж - фактор роста сосудистого эндотелия, 20 нг/мл + крингл-домен урокиназы, 50 нМ; З - основный фактор роста фибробластов, 10 нг/мл + протеолит. неакт. урокиназа, 50 нМ; И - основный фактор роста фибробластов, 10 нг/мл + крингл-домен урокиназы, 50 нМ; К - фактор роста сосудистого эндотелия, 20 нг/мл + по 50 нМ протеолит. неакт. урокиназы и крингл-домена; Л - основный фактор роста фибробластов, 10 нг/мл + по 50 нМ протеолит. неакт. урокиназы и крингл-домена; М - 50 нМ протеолит. неакт. урокиназа; Н - 50 нМ крингл-домен урокиназы.
Б) Количественная обработка данных иммуногистохимического анализа. Число образовавшихся сосудов нормировано на количество сосудов, сформированных в Матригеле™ без использования дополнительных стимулов (столбец 1).
В подкожно вводимый Матригель™ вносили стимулы и рекомбинантные формы урокиназы: 1 - без дополнительных стимулов; 2 - фактор роста сосудистого эндотелия, 20 нг/мл; 3 - фактор роста сосудистого эндотелия, 50 нг/мл; 4 - основный фактор роста фибробластов, 10 нг/мл; 5 - основный фактор роста фибробластов, 20 нг/мл; 6 - фактор роста сосудистого эндотелия, 20 нг/мл + протеолит. неакт. урокиназа, 50 нМ; 7 - фактор роста сосудистого эндотелия, 20 нг/мл + крингл-домен урокиназы, 50 нМ; 8 - основный фактор роста фибробластов, 10 нг/мл + протеолит. неакт. урокиназа, 50 нМ; 9 - основный фактор роста фибробластов, 10 нг/мл + крингл-домен урокиназы, 50 нМ; 10 - фактор роста сосудистого эндотелия, 20 нг/мл + по 50 нМ протеолит. неакт. урокиназы и крингл-домена; 11 - основный фактор роста фибробластов, 10 нг/мл + по 50 нМ протеолит. неакт. урокиназы и крингл-домена; 12-50 нМ протеолит. неакт. урокиназа; 13-50 нМ крингл-домен урокиназы.
SEQ ID NO: 1

Выделены шрифтами: полужирным прописным (II) - начало каталитического домена; полужирным прописным курсивом (PRF) - плазминовый сайт; полужирным прописным с подчеркиванием (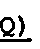 - триада аминокислот активного центра; жирным строчным - аминокислоты, отличные от опубликованной последовательности изоформы 1 (Swissprot Р00749).
- триада аминокислот активного центра; жирным строчным - аминокислоты, отличные от опубликованной последовательности изоформы 1 (Swissprot Р00749).
SEQ ID NO: 2

Выделены шрифтами: полужирным прописным курсивом (PRF) - плазминовый сайт; жирным строчным - аминокислоты, отличные от опубликованной последовательности изоформы 1 (Swissprot Р00749).
Осуществление изобретения
При осуществлении изобретения, помимо методов, подробно раскрытых в нижеследующих примерах, были использованы стандартные технологии генной инженерии, а также опубликованные нами ранее методики культивирования эндотелиальных клеток и модели изучения ангиогенеза in vitro и in vivo.
Пример 1. Получение рекомбинантных форм урокиназы
Рекомбинантные формы урокиназы были получены с использованием технологии рекомбинантных ДНК. Для производства рекомбинантных форм урокиназы были использованы штаммы-продуценты, созданные лабораторией генной инженерии НИИ экспериментальной кардиологии РК НПК МСЗ РФ (для получения протеолитически неактивной урокиназы штамм SC2404; для получения крингл-домена - штамм SC1923).
Наращивание штаммов-продуцентов
Экспрессирующие рекомбинантные формы урокиназы клетки культивировали, используя среду LB [Bertani, G., 1952], содержащую: 1% пептон, 0.5% дрожжевой экстракт и 1% NaCl. Среду стерилизовали автоклавированием в паровом автоклаве при температуре 121°C в течение 60 минут. Использовали стандартную коммерчески доступную среду LB, поставляемую в виде сухого порошка, и готовили ее согласно рекомендациям производителя [Sigma Aldrich L-3022].
Для наработки рекомбинантных белков использовали бактериальные штаммы-продуценты на основе клеток E.coli линии BL21(DE3) (для получения протеолитически неактивной урокиназы штамм SC2404; для получения крингл-домена - штамм SC1923). После размораживания образцов указанных штаммов-продуцентов получали малые культуры, которые растили в течение ночи при 37°C при интенсивной аэрации (260 об/мин). Основная биомасса выращивалась в объеме не менее 2 л в течение 12 часов при 37°C и интенсивной аэрации (260 об/мин).
Разрушение клеточной стенки
Для выделения белков, экспрессированных в указанных штаммах-продуцентах, использовали ферментативную обработку клеточных стенок бактерий лизоцимом, а также обработку клеток ультразвуком. Лизоцимом обрабатывали бактериальные клетки в течение 30 минут при +4°C, а ультразвуком воздействовали 3-4 раза (по 40 секунд) при мощности 80-100 Вт и одновременном охлаждении на ледяной бане. Полученный раствор центрифугировали 10 мин при 20000 g при температуре +4°C.
Выделение и очистка протеолитически неактивной урокиназы и крингл-домена из рефолдинговой смеси
Рекомбинантные формы урокиназы нарабатывали в клетках штамма-продуцента в виде нерастворимой фракции - телец включения, требующей трехкратной отмывки буфером 50 мМ трис/HCl, pH=7.5, содержащим 1 М NaCl, и последующей трехкратной отмывки буфером 50 мМ трис/HCl, pH=7.5. Промытый осадок растворяли в 6 М гуанидин/HCl pH 9.0, содержащем 10 мМ β-меркаптоэтанола (50 мл раствора на 0.5 л клеточной культуры), при необходимости раствор подогревали до 50-60°C. К полученному раствору медленно при перемешивании добавляли 10-кратный объем 1 М гуанидин/HCl pH 9.0, содержащего Red/Ox пару 1 мМ глутатион восстановленный / 0.1 мМ глутатион окисленный. Полученный раствор белков оставляли для рефолдинга на 14 часов при температуре +4°C. Рефолдинговую смесь центрифугировали 10 мин при 20000 g. pH белкового раствора доводили 1 М Трис-HCl pH=2,5 до значения pH 7.4.
Для очистки протеолитически неактивой урокиназы и крингл-домена использовали двухстадийную хроматографическую очистку. Первой стадией была очистка с помощью металл-хелатной аффинной хроматографии. Второй стадией очистки была иммуноафинная хроматография с иммобилизованными на сорбенте антителами.
Образцы полученных рекомбинантных белков разделяли с помощью ДСН-электрофореза в 14% ПААГ по методу Лэммли. Гель окрашивали раствором Кумасси R-250. Гель фотографировали на трансиллюминаторе с помощью цифровой камеры. В результате было установлено, что молекулярный вес полученных белков соответствует протеолитически неактивой урокиназе и крингл-домену урокиназы (фиг.1).
Пример 2. Свойства рекомбинантных форм урокиназы
Анализ функциональных характеристик протеолитически неактивой урокиназы и крингл-домена, полученных в примере 1, проводили с помощью иммуноблоттинга и иммуноферментного анализа.
Нативностъ конформации очищенных белков
Для определения нативности конформации рекомбинантных белков использовали антитела, распознающие антигенные детерминанты, формируемые аминокислотными остатками белка.
Белки разделяли с помощью ДСН-электрофореза в ПААГ, а затем переносили их из геля на ПВДФ-мембрану, где окрашивали с использованием специфических моноклональных антител против значимых детерминант нативной урокиназы человека, в концентрации 0,2 мкг/мл (UNG - к крингл-домену, UIG - к протеолитическому домену). В качестве вторичных антител использовали антитела козы к иммуноглобулинам мыши, конъюгированные с пероксидазой хрена AffiniPure (Н+L), в разведении 1:2000. Для детекции белков инкубировали мембрану с хемилюминесцентным субстратом. Люминесцентные сигналы регистрировали с помощью чувствительной цифровой видеокамеры. Белки разделяли с помощью ДСН-электрофореза в 14% ПААГ и осуществляли электроперенос на мембрану.
Нативная урокиназа и полученные рекомбинантные белки связывали антитела UNG (дорожки 1-3 на фиг.2). Нативная урокиназа и полученная протеолитически неактивная урокиназа также связывали антитела UIG в отличие от крингл-домена (сравни дорожки 5 и 6 на фиг.2). Связывание специфичных антител свидетельствует о сохранности значимых детерминант в структуре белков, а значит и о нативной конформации полученных рекомбинантных белков.
Протеолитическая активность рекомбинантных белков на основе структуры урокиназы
Активность урокиназы определяли по протеолитической активности с использованием в качестве субстрата синтетического пептида пиро-Glu-Gly-Arg-pNA (S-2444, Sigma). Ферментативная активность урокиназы при данном подходе определяется путем сравнения активности исследуемого препарата урокиназы и активности прилагаемого к набору стандарта.
Для этого препарат рекомбинантного белка урокиназы в концентрации 2 мкМ преинкубировали с плазмином (конечная концентрация 0,005 мг/мл) в течение 1 часа при 37°C в буфере PBS, содержащем 0,1% BSA и 0,05% Tween-20. Для определения доли двухцепочечной формы урокиназы, содержащейся в препарате, часть образца не активировали плазмином. Образец, активированный плазмином, разбавляли до конечной концентрации урокиназы 5-30 нМ буфером PBS / 0,1% BSA / 0,05% Tween-20, содержащим ингибитор плазмина апротинин в концентрации 100 калликреиновых ед./мл (940 мкл буфера + 10 мкл образца). Неактивированный образец разбавляли таким же буфером до конечной концентрации урокиназы 50-300 нМ. К пробам, объем которых составлял по 950 мкл, добавляли по 50 мкл раствора специфичного субстрата урокиназы S-2444 в буфере PBS / 0,1% BSA / 0,05% Tween-20 (1,92 мг/мл). Каталитическую активность определяли по высвобождению паранитроанилида путем измерения поглощения при длине волны 405 нм каждую минуту. Рассчитывали каталитическую активность по формуле МкКат/л= =1,74*Объем пробы (1 мл)*A (среднее изменение поглощения за 1 мин на линейном участке)/Объем образца (10 мкл).
Мы обнаружили, что полученные рекомбинантные белки (протеолитически неактивная урокиназа и крингл-домен) не обладают протеолитической активностью в отличие от контрольной урокиназы (сравни графики на фиг.3).
Связывание рекомбинантных белков на основе структуры урокиназы с рецептором урокиназы
Для проверки способности рекомбинантных белков на основе структуры урокиназы связываться с рецептором урокиназы использовали свойство очищенного урокиназного рецептора сохранять способность связывать лиганды.
Для этого рекомбинантный рецептор иммобилизовали на дне лунок 96-луночного пластикового планшета для ИФА инкубированием в течение 1 часа с урокиназным рецептором (0,11 нмоль в буфере 0.1 М NaHCO3 pH 9.5) при 37°C и постоянном перемешивании. Параллельно с лунками с рецептором белки-лиганды наносили в чистые лунки для контроля качества титровки системы. Промывку и блокирование неспецифических участков связывания здесь и далее проводили блокирующим буфером 50 мМ трис/HCl pH 7.5, содержащим 10 мг/мл БСА, 20 мг/мл желатина, 0,02% Tween-20. Добавляли рекомбинантные формы урокиназы в количестве 0-0,014 нмоль и инкубировали в течение 1 часа. Промывали 4 раза по 5 минут при 37°C при перемешивании в блокирующем буфере. В лунки вносили биотинилированные поликлональные антитела козы к урокиназе (GAHUK) в концентрации 2 мкг/мл в блокирующем буфере. Промывали 4 раза по 5 минут при 37°C при перемешивании в блокирующем буфере. Вносили нейтравидин, конъюгированный с полипероксидазой хрена, специфически узнающий остатки биотина. Промывали 4 раза по 5 минут при 37°C при перемешивании в блокирующем буфере. Инкубировали с субстратом орто-фенилендиамином (0,1 мг/мл) до развития окраски. Реакцию останавливали добавлением 0,5 объема 10% H2SO4. Интенсивность окраски измеряли на планшетном спектрофлуориметре при длине волны 492 нм.
Было установлено, что характер связывания рекомбинантной протеолитически неактивной формы урокиназы, содержащей ростовой домен, не отличался от связывания нативной стандартной полноразмерной урокиназы и связывания рекомбинантной полноразмерной урокиназы (сравни кривые на фиг.4).
Таким образом, функциональная активность полученных рекомбинантных форм урокиназы соответствует протеолитически неактивой урокиназе и крингл-домену урокиназы.
Пример 3. Подавление миграции эндотелиальных клеток под действием рекомбинантных форм урокиназы
Влияние полученных протеолитически неактивной урокиназы и крингл-домена на миграцию эндотелиальных клеток изучали с помощью камеры Бойдена производства «Neuroprobe Inc.», США. Для этого эндотелиальные клетки вены пуповины человека, достигшие конфлюэнтного состояния, перед экспериментом культивировали в бессывороточной среде DMEM/0,1% БСА в течение 24 ч.
В качестве хемоаттрактантов использовали различные формы урокиназы человека, которые помещали в нижние ячейки камеры Бойдена. Хемоаттрактант и эндотелиальные клетки были разделены поликарбонатным пористым фильтром с диаметром пор 8 мкм («Nucleopore Согр.», США), покрытым коллагеном I типа (100 нг/мл) (Vitrogen 100, «Celtrix Pharmaceuticals Inc.», США).
В верхние ячейки камеры в бессывороточной среде DMEM/0,1% БСА вносили по 50 мкл суспензии эндотелиальных клеток, полученной при обработке монослоя клеток раствором трипсина и ЭДТА (0,05/0,02%). Число эндотелиальных клеток, вносимых в лунку, составляло 75000 клеток. Камеру с эндотелиальными клетками и добавленными контрольными стимулами и исследуемыми веществами инкубировали в CO2-инкубаторе в течение 3 часов. Эмигрировавшие эндотелиальные клетки снимали специальным скребком с верхней поверхности фильтра, а эндотелиальные клетки на нижней поверхности фиксировали в метаноле и окрашивали красителем DiffQuick («Baxter», США). Мембрану с окрашенными эндотелиальными клетками сканировали на сканере ScanJet II СХ с использованием программ Deskscan и NIH Image. Интенсивность миграции оценивали по площади пиков, полученных сканированием поля окрашенных эндотелиальных клеток. Результаты представлены в виде отношения полученных площадей к контрольной площади, полученной для клеток, прошедших через фильтр в отсутствие хемоаттрактанта.
При анализе способности рекомбинантных форм урокиназы подавлять миграцию эндотелиальных клеток было обнаружено, что урокиназа дикого типа не влияет на миграцию эндотелиальных клеток в ответ на 10 нг/мл bFGF, а рекомбинантная протеолитически неактивная урокиназа и ее рекомбинантный крингл-домен вызывают снижение клеточной миграции ниже начального уровня (сравни столбец uPAwt со столбцами uPA HQ и kringle на фиг.5). Крингл-домен проявляет в этом отношении большую активность (сравни столбцы uPA HQ и kringle на фиг.5).
Таким образом, рекомбинантная протеолитически неактивная урокиназа и рекомбинантный крингл-домен урокиназы подавляют миграцию эндотелиальных клеток, в то время как полноразмерная протеолитически активная урокиназа не влияет на миграцию эндотелиальных клеток. На основании полученных результатов о негативном влиянии протеолитически неактивной урокиназы и рекомбинантного крингл-домена урокиназы на миграцию эндотелиальных клеток можно сделать вывод о потенциальной возможности применения этих рекомбинантных белков для подавления ангиогенеза.
Пример 4. Подавление формирования капилляроподобных структур in vitro под действием рекомбинантных форм урокиназы
Для оценки формирования капилляроподобных структур использовали 24-луночный планшет, дно которого покрывали Матригелем™ (BD Biosciences). В каждую лунку высеивали 5×104 HUVEC (эндотелиальных клеткок из пуповины человека). Для определения влияния иммобилизованных форм урокиназы на формирование капилляров в Матригель™ вносили исследуемые стимуляторы или ингибиторы. Клетки инкубировали в среде культивирования для эндотелиальных клеток в CO2-инкубаторе при 37°C в течение 24 ч. Формируемые капилляроподобные структуры фотографировали. Суммарную длину формирующихся капилляроподобных структур оценивали в 5 случайных полях зрения в каждой лунке с помощью программы "MetaMorph 5.0" ("Universal Imaging"). Для обеспечения достоверности результатов эксперименты повторяли трижды в 3 параллелях, используя по 9 лунок на экспериментальную точку.
На представленных рисунках видно (фиг.6), что рекомбинантный крингл-домен и протеолитически неактивная урокиназа влияют на формирование эндотелиальных микротрубочек, стимулированное основным фактором роста фибробластов. При этом протеолитически неактивная урокиназа подавляет формирование эндотелиальных микротрубочек.
На графиках (фиг.6) представлены результаты математической обработки полученных изображений. Математическая обработка позволяет учитывать длину образующихся эндотелиальных микротрубочек на единицу площади.
Аналогичный эффект был продемонстрирован при использовании в качестве стимулятора образования микротрубочек основного фактора роста фибробластов. На графике видно незначительное уменьшение длины эндотелиальных микротрубочек при использовании протеолитически неактивной урокиназы в качестве ингибитора.
Результаты математической обработки демонстрируют незначительное, но достоверное уменьшение длины эндотелиальных микротрубочек при использовании протеолитически неактивной урокиназы в качестве ингибитора и фактора роста сосудистого эндотелия в качестве стимулятора.
Тем не менее, при визуальной оценке процесса образования эндотелиальных микротрубочек видно, что на этот процесс влияет не только протеолитически неактивная урокиназа, но и рекомбинантный крингл-домен урокиназы.
В результате исследования способности рекомбинантной протеолитически неактивной урокиназы и рекомбинантного крингл-домена урокиназы ингибировать формирование микротрубочек было продемонстрировано, что протеолитически неактивная урокиназа незначительно, но достоверно уменьшает длину образующихся эндотелиальных микротрубочек. Продемонстрировано влияние рекомбинантной протеолитически неактивной урокиназы и ее рекомбинантного крингл-домена на морфологию образующихся микротрубочек.
На основании результатов исследования способности рекомбинантной протеолитически неактивной урокиназы и рекомбинантного крингл-домена урокиназы влиять на формирование микротрубочек можно сделать вывод о потенциальной возможности применения этих рекомбинантных конструкций для подавления ангиогенеза.
Пример 5. Подавление ангиогенеза in vivo под действием рекомбинантных форм урокиназы
Для проверки гипотезы о том, что рекомбинантная протеолитически неактивная урокиназа и ее рекомбинантный крингл-домен могут подавлять формирование кровеносных сосудов в организме, было проведено исследование влияния рекомбинантных форм урокиназы на ангиогенез на модели in vivo.
Для оценки влияния рекомбинантных форм урокиназы на рост кровеносных сосудов in vivo была выбрана модель ангиогенеза в подкожном имплантате Матригеля™.
Матригель™ является продуктом секреции клеточной линии опухоли (мышиной саркомы Энгельбрета-Хольм-Сварма) и поэтому может рассматриваться как модель опухолевого ангиогенеза. Матригель™, содержащий в своем составе комплекс факторов роста и матриксных белков, стимулирует прорастание сосудов. Добавляя в его состав исследуемые факторы, можно оценить их влияние на ангиогенез.
Мышь наркотизировали внутрибрюшинным введением 2,5% раствора авертина;
Матригель™ размораживали на льду, не допуская нагревания выше +4°C (при +8°C гель полимеризуется). В Матригель™ вводили исследуемые добавки, не допуская повышения температуры. Каждой мыши вводили подкожно в каждый бок по 500 мкл Матригеля™. Инъекции проводили инсулиновым шприцем с иглой толщиной 23 G таким образом, чтобы Матригель™ успевал полимеризоваться под кожей и образовывал желеобразный имплантат неправильной формы. Экспериментальные группы включали по 5 мышей. Через 10 дней после подкожного введения Матригеля™ с исследуемыми добавками мышей умерщвляли путем цервикальной дислокации. Затем изымали имплантаты и замораживали их в жидком азоте в среде для заморозки тканей Tissue-Tek (Sakura, Япония). Оценку плотности кровеносных сосудов на срезах Матригеля™ проводили с помощью иммунофлуоресцентного окрашивания. Для этого с помощью криомикротома приготовляли срезы толщиной 6 мкм; которые фиксировали в 4% растворе формалина (Sigma, США) при комнатной температуре в течение 10 мин; отмывали зафиксированные срезы фосфатно-солевым буфером (PBS); инкубировали в 0,05М растворе ацетата аммония на PBS (10 минут). Затем отмывали срезы PBS и в течение 30 минут инкубировали срезы в 1% растворе БСА (бычий сывороточный альбумин) на PBS, содержащем 10% сыворотки. Затем срезы инкубировали в течение 1 часа при комнатной температуре с моноклональными антителами к CD31; трижды отмывали PBS и помещали в раствор вторичных антител на 1 час. В качестве отрицательного контроля часть срезов инкубировали с неспецифическими изотипическими иммуноглобулинами вместо первых антител. Ядра клеток докрашивали флуоресцентным красителем DAPI. После трехкратной отмывки в PBS срезы заключали в среду Mounting Medium Vectashield™ (Vector Laboratories Inc.). Для предотвращения «выгорания» флуоресцентных меток готовые образцы хранили при 4°C в темноте. Полученные препараты анализировали при помощи флуоресцентного микроскопа ZeissAxiovert200M. Документирование изображений производили с помощью цифровой видеокамеры Axiocam HRc (Zeiss, Германия). Изображения обрабатывали в программе Axiovision. Размер сосудов и их плотность оценивали с помощью программного обеспечения MetaMorph 5.0 (Universal Imaging) и ClickCounter. Подсчет сосудов проводили в 5 полях зрения на срезе (по 12 срезов на каждый имплантат) при 20-кратном увеличении, отдельно выделяя капилляры, сосуды среднего диаметра и крупные сосуды. Полученное число сосудов в поле зрения нормировали на единицу площади среза.
Мы обнаружили, что рекомбинантная протеолитически неактивная форма урокиназы и рекомбинантный крингл-домен урокиназы подавляют формирование сосудов в имплантате Матригеля™ как на фоне стимуляции ангиогенеза фактором роста эндотелия сосудов и основным фактором роста фибробластов, так и базальный уровень формирования сосудов в Матригеле™ без введения дополнительных стимуляторов ангиогенеза (фиг.7). По результатам математической обработки наблюдаемые эффекты рекомбинантных форм урокиназы на ангиогенез имели достоверный характер.
Список литературы
1. Naldini L, Tamagnone L, Vigna E, Sachs M, Hartmann G, Birchmeier W, Daikuhara Y, Tsubouchi H, Blasi F, Comoglio PM. EMBO J. 1992 Dec; 11(13):4825-33.
2. Tkachuk V, Stepanova V, Little PJ, Bobik A. Clin Exp Pharmacol Physiol. 1996 Sep; 23(9):759-65.
3. Carmeliet P, Moons L, Herbert JM, Crawley J, Lupu F, Lijnen R, Collen D.
Circ Res. 1997 Nov; 81(5):829-39.
4. Mazar AP, Henkin J, Goldfarb RH. Angiogenesis. 1999; 3(l):15-32.
5. van Hinsbergh VW, Engelse MA, Quax PH.
Arterioscler Thromb Vasc Biol. 2006 Apr; 26(4):716-28. Epub 2006 Feb 9. Review.
6. Colombo ES, Menicucci G, McGuire PG, Das AInvest Ophthalmol Vis Sci. 2007 Apr; 48(4): 1793-800.
7. Heymans S, Luttun A, Nuyens D, Theilmeier G, Creemers E, Moons L, Dyspersin GD, Cleutjens JP, Shipley M, Angellilo A, Levi M, Nübe O, Baker A, Keshet E, Lupu F, Herbert JM, Smits JF, Shapiro SD, Baes M, Borgers M, Collen D, Daemen MJ, Carmeliet P. Nat Med. 1999 Oct; 5(10):1135-42.
8. Ugwu F, Van Hoef B, Bini A, Collen D, Lijnen HR Biochemistry. 1998 May 19; 37(20):7231-7236.
9. Guo Y, Higazi AA, Arakelian A, Sachais BS, Cines D, Goldfarb RH, Jones TR, Kwaan H, Mazar AP, Rabbani SA. FASEB J. 2000 Jul; 14(10):1400-10.
10. Koolwijk P, van Erck MG, de Vree WJ, Vermeer MA, Weich HA, Hanemaaijer R, van Hinsbergh VW. J Cell Biol. 1996 Mar; 132(6): 1177-88.
11. Quax PH, Grimbergen JM, Lansink M, Bakker AH, Blatter MC, Belin D, van Hinsbergh VW, Verheijen JH. Arterioscler Thromb Vasc Biol. 1998 May; 18(5):693-701.
12. Deindl E, Ziegelhöffer T, Kanse SM, Fernandez B, Neubauer E, Carmeliet P, Preissner KT, Schaper W. FASEB J. 2003 Jun; 17(9):1174-6. Epub 2003 Apr 8.
13. Kirchheimer JC, Wojta J, Christ G, Binder BR. Proc Natl Acad Sci USA. 1989 Jul; 86(14):5424-8.
14. Ossowski L, Reich E Cell. 1983 Dec; 35(3 Pt 2):611-9.
15. Graf L, Craik CS, Patthy A, Roczniak S, Fletterick RJ, Rutter WJ. Biochemistry. 1987 May 5; 26(9):2616-23.
16. Spraggon G, Phillips C, Nowak UK, Ponting CP, Saunders D, Dobson CM, Stuart DI, Jones EY. Structure. 1995 Jul 15; 3(7):681-91.
17. Vassalli JD, Belin D. FEBS Lett. 1987 Apr 6; 214(1):187-91.
18. Towle MJ, Lee A, Maduakor EC, Schwartz CE, Bridges AJ, Littlefield BA. Cancer Res. 1993 Jun 1; 53(11):2553-9.
19. Alonso, D.F.; Farias, E.F.; Ladeda, V.; Davel, L.; Puricelli, L.; Bal de Kier Joffe, E Tumor Model. Breast Cancer Res. Treat. 1996, 40, 209-223.
20. Alonso, D.F.; Tejera, A.M.; Farias, E.F.; Bal de Kier Joffe, E.; Gomez, D.E. Anticancer Res. 1998, 18, 4499-4504.
21. Rabbani, S.A.; Harakidas, P.; Davidson, D.J.; Henkin, J.; Mazar, A.P. Int. J. Cancer 1995, 63, 840-845.
22. Xing, R.H.; Mazar, A.; Henkin, J.; Rabbani, S.A. Cancer Res. 1997, 57, 3585-3593.
23. Katz, В.A.; Mackman, R.; Luong, C.; Radika, K.; Martelli, A.; Sprengeler, P.A.; Wang, J.; Chan, H.; Wong, L. Chem. Biol. 2000, 7, 299-312.
24. Nienaber, V.L.; Davidson, D.; Edalji, R.; Giranda, V.L.; Klinghofer, V.; Henkin, J.; Magdalinos, P.; Mantei, R.; Merrick, S.; Severin, J.M.; Smith, R.A.; Stewart, K.; Walter, K.; Wang, J.; Wendt, M.; Weitzberg, M.; Zhao, X.; Rockway, T. Structure 2000, 8, 553-563.
25. Nienaber, V.; Wang, J.; Davidson, D.; Henkin, J. J. Biol. Chem. 2000, 275, 7239-7248.
26. Wendt, M.D.; Rockway, T.W.; Geyer, A.; McClellan, W.; Weitzberg, M.; Zhao, X.; Mantei, R.; Nienaber, V.L.; Stewart, K.; Klinghofer, V.; Giranda, V.L. J. Med. Chem. 2004, 47, 303-324.
27 Bruncko, M.; McClellan, W.J.; Wendt, M.D.; Sauer, D.R.; Geyer, A.; Dalton, C.R.; Kaminski, M.A.; Weitzberg, M.; Gong, J.; Dellaria, J.F.; Mantei, R.; Zhao, X.; Nienaber, V.L.; Stewart, K.; Klinghofer, V.; Bouska, J.; Rockway, T.W.; Giranda, V.L. Bioorg. Med. Chem. Lett. 2005, 15, 93-98.
28. Spencer, J.R.; McGee, D.; Allen, D.; Katz, B.A.; Luong, C.; Sendzik, M.; Squires, N.; Mackman, R.L. Bioorg. Med. Chem. Lett. 2002, 12, 2023-2026.
29. Barber, C.G.; Dickinson, R.P.; Fish, P.V. Bioorg. Med. Chem. Lett. 2004, 14, 32273230.
30. Barber, C.G.; Dickinson, R.P.; Horne, V.A. Bioorg. Med. Chem. Lett. 2002, 12, 181-184.
31. Fish, P.V.; Barber, C.G.; Brown, D.G.; Butt, R.; Collis, M.G.; Dickinson, R.P.; Henry, B.Т.; Horne, V.A.; Huggins, J.P.; King, E.; O′Gara, M.; McCleverty, D.; McIntosh, F.; Phillips, C.; Webster, R. J. Med. Chem. 2007, 50, 2341-2351.
32. Frederickson, M.; Callaghan, O.; Chessari, G.; Congreve, M.; Cowan, S.R.; Matthews, J.E.; McMenamin, R.; Smith, D.M.; Vinkovi.., M.; Wallis, N.G. J. Med. Chem. 2008, 51, 183-186.
33. Heynekamp, J.J.; Hunsaker, L.A.; Vander Jagt, T.A.; Deck, L.M.; Vander Jagt, D.L. BMC Chem. Biol. 2006, 6, 1.
34. Zhu, M.; Gokhale, V.M.; Szabo, L.; Munoz, R.M.; Baek, H.; Bashyam, S.; Hurley, L.H.; Von Hoff, D.D.; Han, H. Mol. Cancer Ther. 2007, 6, 1348-1356.
35. Subasinghe, N.L.; Illig, C.; Hoffman, J.; Rudolph, M.J.; Wilson, K.J.; Soil, R.; Randle, Т.; Green, D.; Lewandowski, F.; Zhang, M.; Bone, R.; Spurlino, J.; DesJarlais, R.; Deckman, I.; Molloy, C.J.; Manthey, C; Zhou, Z.; Sharp, C.; Maguire, D.; Crysler, C.; Grasberger, B. Structure-Based Design, Bioorg. Med. Chem. Lett. 2001, 11, 1379-1382.
36. Sperl, S.; Jacob, U.; Arroyo de Prada, N.; Sturzebecher, J.; Wilhelm, O.G.; Bode, W.; Magdolen, V.; Huber, R.; Moroder, L. Proc. Natl. Acad. Sci. USA 2000, 97, 5113-5118.
37. Ertongur, S.; Lang, S.; Mack, В.; Wosikowski, K.; Muehlenweg, В.; Gires, O. Int. J. Cancer 2004, 110, 815-824.
38. Setyono-Han, В.; Stuerzebecher, J.; Schmalix, W.A.; Muehlenweg, В.; Sieuwerts, A.; Timmermans, M.; Magdolen, V.; Schmitt, M.; Klijn, J.G. M.; Foekens, J.A. Thromb. Haemost. 2005, 93, 779-786.
39. Wang, С.I.; Yang, Q.; Craik, C.S. J. Biol. Chem. 1995, 270, 12250-12256.
40. Yang, S.Q.; Craik, C.S. J. Mol. Biol. 1998, 279, 1001-1011.
41. Laboissiere, M.C.; Young, M.M.; Pinho, R.G.; Todd, S.; Fletterick, R.J.; Kuntz, I.; Craik, C.S. J. Biol. Chem. 2002, 277, 26623-26631.
42. Hansen, M.; Wind, Т.; Blouse, G.E.; Christensen, A.; Petersen, H.H.; Kjelgaard, S.; Mathiasen, L.; Holtet, T.L.; Andreasen, P.A.A J. Biol. Chem. 2005, 280, 38424-38437.
43. Tamura, S.Y.; Weinhouse, M.I.; Roberts, C.A.; Goldman, E.A.; Masukawa, K.; Anderson, S.M; Cohen, C.R.; Bradbury, A.E.; Bernardino, V.Т.; Dixon, S.A.; Ma, M.G.; Nolan, T.G.; Brunck, Т.K. Bioorg. Med. Chem. Lett. 2000, 10, 983-987.
44. Kunzel, S.; Schweinitz, A.; Reissmann, S.; Sturzebecher, J.; Steinmetzer, T. Bioorg. Med. Chem. Lett. 2002, 12, 645-648.
45. Zeslawska, E.; Jacob, U.; Schweinitz, A.; Coombs, G.; Bode, W.; Madison, E. J. Mol. Biol. 2003, 328, 109-118.
46. Schweinitz, A.; Steinmetzer, Т.; Banke, I.J.; Arlt, M.J.; Sturzebecher, A.; Schuster, O.; Geissler, A.; Giersiefen, H.; Zeslawska, E.; Jacob, U.; Kruger, A.; Sturzebecher, J. J. Biol. Chem. 2004, 279, 33613-33622.
47. Joossens, J.; Van der Veken, P.; Lambeir, A.M.; Augustyns, K.; Haemers, A. J. Med. Chem. 2004, 47, 2411-2413.
48. Sieczyk, M.; Oleksyszyn, J. Bioorg. Med. Chem. Lett. 2006, 16, 2886-28890.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО АНТИАНГИОГЕННОГО ПОЛИПЕПТИДА | 2012 |
|
RU2488635C1 |
| ДВУХСЛОЙНАЯ, ПЛОСКАЯ, ПРОЗРАЧНАЯ ПОДЛОЖКА ГИДРОГЕЛЯ ДЛЯ ДЛИТЕЛЬНОГО КУЛЬТИВИРОВАНИЯ КЛЕТОК | 2016 |
|
RU2614260C1 |
| МНОГОФУНКЦИОНАЛЬНЫЕ И ПОЛИВАЛЕНТНЫЕ ИНГИБИТОРЫ АНГИОГЕНЕЗА | 2005 |
|
RU2398878C2 |
| ПРИМЕНЕНИЕ FVIIa ИЛИ АНТАГОНИСТА ТКАНЕВОГО ФАКТОРА ДЛЯ РЕГУЛЯЦИИ ГЕННОЙ ЭКСПРЕССИИ И КЛЕТОЧНОЙ МИГРАЦИИ ИЛИ ХЕМОТАКСИСА | 2000 |
|
RU2268744C2 |
| РЕКОМБИНАНТНЫЙ ГИБРИДНЫЙ ПОЛИПЕПТИД, СПОСОБНЫЙ ИНГИБИРОВАТЬ ПРОЛИФЕРАЦИЮ ЭНДОТЕЛИАЛЬНЫХ КЛЕТОК ЧЕЛОВЕКА IN VITRO, И СПОСОБ ЕГО ПОЛУЧЕНИЯ | 2011 |
|
RU2465283C1 |
| СПОСОБ ПОВЫШЕНИЯ АНГИОГЕННОЙ АКТИВНОСТИ СТРОМАЛЬНЫХ КЛЕТОК ЖИРОВОЙ ТКАНИ | 2011 |
|
RU2531502C2 |
| ПЕПТИДЫ НАПРАВЛЕННОГО ДЕЙСТВИЯ НА VEGFR-1/NRP-1 | 2008 |
|
RU2488592C2 |
| СПОСОБ ОЦЕНКИ АНГИОГЕННОГО ПОТЕНЦИАЛА ПРОГЕНИТОРНЫХ КЛЕТОК У БОЛЬНЫХ СЕРДЕЧНО-СОСУДИСТЫМИ ЗАБОЛЕВАНИЯМИ | 2011 |
|
RU2548801C2 |
| ШТАММ Е. coli BL21(DE3)/pTEV-TMS - ПРОДУЦЕНТ ГИБРИДНОГО БЕЛКА TrxTEVrs-TMS, ПРЕДНАЗНАЧЕННОГО ДЛЯ ПРОТЕОЛИТИЧЕСКОГО РАСЩЕПЛЕНИЯ С ОБРАЗОВАНИЕМ АНТИАНГИОГЕННОГО ПЕПТИДА ТУМАСТИНА, ПРОИЗВОДНОГО ФРАГМЕНТА [L69K-95] ТУМСТАТИНА ЧЕЛОВЕКА, И СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО АНТИАНГИОГЕННОГО ПЕПТИДА | 2015 |
|
RU2625008C2 |
| СПОСОБ СТИМУЛЯЦИИ РЕГЕНЕРАТИВНЫХ ПРОЦЕССОВ В ИШЕМИЗИРОВАННЫХ ТКАНЯХ | 2011 |
|
RU2497529C2 |

Изобретение относится к области биотехнологии, в частности к молекулярной медицине, и может быть использовано в медицинской практике для лечения патологий, связанных с избыточным ангиогенезом, включая рост злокачественных опухолей, ревматоидный артрит, псориаз и диабетическую ретинопатию. Способ включает введение в область избыточного ангиогенеза эффективного количества протеолитически неактивных рекомбинантных форм активатора плазминогена урокиназного типа (урокиназы) в сочетании с рекомбинантным крингл-доменом активатора плазминогена урокиназного типа, конкурентно подавляющих протеолитическое действие нативного активатора плазминогена урокиназного типа и пространственно разобщающих рецепторные белки, опосредующие регуляторные эффекты системы урокиназы. Способ позволяет локально подавлять рост кровеносных сосудов. 3 н. и 1 з.п. ф-лы, 9 ил., 5 пр.
1. Способ локального подавления роста кровеносных сосудов путем совместного введения в участок избыточного ангиогенеза эффективного количества протеолитически неактивной рекомбинантной формы активатора плазминогена урокиназного типа и рекомбинантного крингл-домена активатора плазминогена урокиназного типа, причем в качестве рекомбинантной формы активатора плазминогена используют рекомбинантную молекулу полноразмерной протеолитически неактивной урокиназы с аминокислотной последовательностью SEQ ID NO:1., препятствующую протеолитической активности эндогенной урокиназы, а в качестве рекомбинантного крингл-домена используют рекомбинантный крингл-домен урокиназы с аминокислотной последовательностью SEQ ID NO:2, пространственно разобщающий рецепторные белки, которые опосредуют эффекты урокиназы, и также вытесняющий протеолитическую активность эндогенной урокиназы из участков связывания крингл-домена урокиназы.
2. Способ по п.1, отличающийся тем, что используют комбинацию (1:1) полноразмерной протеолитически неактивной урокиназы с аминокислотной последовательностью SEQ ID NO:1 и крингл-домена урокиназы с аминокислотной последовательностью SEQ ID NO:2.
3. Рекомбинантная полноразмерная протеолитически неактивная урокиназа, имеющая аминокислотную последовательность SEQ ID NO:1, для использования в способе по п.1.
4. Рекомбинантный протеолитически неактивный крингл-домен урокиназы, имеющий аминокислотную последовательность SEQ ID NO:2, для использования в способе по п.1.
| КОМПОЗИЦИИ, ПОДАВЛЯЮЩИЕ РАЗВИТИЕ КРОВЕНОСНЫХ СОСУДОВ, И СПОСОБЫ ИХ ИСПОЛЬЗОВАНИЯ | 2001 |
|
RU2304433C2 |
| KOOLWIJK P et.al | |||
| Cooperative effect of TNFalpha, bFGF, and VEGF on the formation of tubular structures of human microvascular endothelial cells in a fibrin matrix | |||
| Role of urokinase activity, J Cell Biol | |||
| Предохранительное устройство для паровых котлов, работающих на нефти | 1922 |
|
SU1996A1 |
| RU 2001120927 A, 27.07.2003 | |||

