Область техники, к которой относится изобретение
Описана новая регулирующая клетки активность фактора коагуляции VII (FVII) или антагониста тканевого фактора, такого как, например, инактивированный фактор коагуляции VIIa (FVIIai) клеток, экспрессирующих тканевой фактор (TF). Настоящее изобретение относится к способу регуляции клеточной миграции или хемотаксиса с помощью контактирования клетки с FVIIa или с другим агонистом TF, или с FVIIai или с другим антагонистом TF и определения миграции указанной клетки. Изобретение также относится к применению FVIIa или другого агониста TF, или FVIIai, или другого антагониста TF для получения лекарства для регуляции клеточной миграции у больного. Более того, настоящее изобретение относится к способу лечения и к способу определения активности соединений, в частности предполагаемых лекарств, которые связаны с клеточной миграцией.
Известный уровень техники
Внешний путь свертывания крови инициируется при связывании циркулирующего в крови FVIIa с интегральным белком мембран, тканевым фактором (TF). Роль TF в свертывании крови интенсивно изучается. Считается, что вовлечение FVIIa в качестве протеолитического фермента в каскад свертывания крови ограничивается внеклеточной частью клеток, экспрессирующих TF. Внутриклеточная активность FVIIa впервые была предположена, когда была выявлена гомология последовательности TF и суперсемейства цитокиновых/интерфероновых или гемопоэтических рецепторов. Подкласс I семейства гемопоэтических рецепторов включает рецепторы гормона роста, пролактина, интерлейкинов с 1 по 7, факторов, стимулирующих колонии гранулоцитов и макрофагов, эритропоэтина и тромбопоэтина. Подкласс II включает TF и рецепторы интерферона а и b.
Сходство TF с данным классом рецепторов дополнительно подтвердилось с выявлением кристаллической структуры. Характеристикой данного класса рецепторов цитокинов, который включает рецепторы интерферона b и g, а также ИЛ-10, является то, что их активация ведет к быстрому фосфорилированию по тирозину самих рецепторов, а также группы внутриклеточных белков. В течение минут после первоначального фосфорилирования по тирозину активируется набор митоген-активируемых (Ser/Thr) киназ (МАРК). Данные киназы связаны с несколькими параллельными сигнальными путями. При исследовании предполагаемой внутриклеточной сигнальной активности FVIIa показано, что он индуцирует мобилизацию свободного внутриклеточного кальция (Ca2+) в клеточной линии J82 карциномы мочевого пузыря человека, которая конститутивно экспрессирует TF, и в клетках эндотелия пупочной вены, которые были предварительно обработаны интерлейкином-1 для экспрессии TF, но не способен вызывать какую-либо активацию внутриклеточных тирозинкиназ подобно цитокинам. В заключение предполагается, что FVIIa зависимым от TF способом индуцирует мобилизацию внутриклеточного Ca+2 путем активации фосфолипазы С. Механизм, с помощью которого FVIIa активирует фосфолипазу С не известен, но активация тирозинкиназы определенно исключена.
Последние данные ряда лабораторий указывают на те, что TF может влиять на спектр важных биологических функций, отличных от коагуляции, таких как ангиогенез, васкуляризация эмбрионов и метастазирование опухолей. В настоящее время, однако, неясно каким образом TF вносит вклад в данные биологические процессы. Внеклеточный домен TF состоит из двух модулей, подобных фибронектину типа III, что типично для внеклеточного домена рецепторов цитокинов класса II, это дает возможность предположить, что TF может играть роль в передаче сигнала, главной функции рецептора цитокинов. Однако TF имеет очень короткий цитоплазматический домен (длиной только из 21 аминокислотного остатка) и лишен прилегающих к мембране мотивов, которые опосредуют связывание нерецепторных киназ Януса (Jaks), существенных для передачи сигнала рецепторов цитокинов. Несмотря на это, некоторые биохимические данные предполагают наличие у TF функции трансдукции сигнала. Анализ последовательности белка TF человека выявил предполагаемый сайт фосфорилирования в цитоплазматическом домене, который консервативен для TF мыши, крысы и кролика. Специфические остатки серина в цитоплазматическом хвосте TF фосфорилируются в клетках после стимуляции с помощью активатора протеинкиназы С. Цитоплазматический хвост TF человека фосфорилируется in vitro по множеству сайтов при инкубации с лизатами клеток U87-MG. На потенциальную роль цитоплазматического домена TF в передаче сигнала указывается также в исследованиях, которые показали, что прометастатическое действие TF принципиально зависит от цитоплазматического домена TF. Кроме того, обнаружено, что цитоплазматический домен TF взаимодействует с актинсвязывающим белком 280 (АВР-280) и поддерживает клеточную адгезию и миграцию путем привлечения АВР-280 к опосредуемым TF адгезионным контактам.
Однако показано также, что TF участвует в определенных типах клеточной сигнализации, действуя в качестве кофактора для своего физиологического лиганда FVIIa во внеклеточной сигнализации посредством протеолитического механизма. Например, показано, что связывание FVIIa с TF клеточной поверхности индуцирует осцилляции внутриклеточного Са2+ в ряде клеток, экспрессирующих TF, временное фосфорилирование тирозина в моноцитах, активацию MAP киназы, изменение экспрессии генов в фибробластах и повышенную экспрессию рецептора урокиназы в опухолевых клетках. Каталитически неактивный FVIIa (FVIIai) не способен индуцировать многие из указанных сигнальных ответов, начиная с осцилляции Са2+ до активации MAP киназы и снижения активности генов, и, похоже, что каталитическая активность FVIIa может требоваться для, по меньшей мере, некоторых путей проведения сигнала, опосредуемого TF-FVIIa. В настоящее время мало известно о пути(ях) проведения сигнала, которые индуцируются протеолитически активным FVIIa, и о том, как возникающие под действием FVIIa сигналы могли бы участвовать в ангиогенезе и в метастазировании опухоли.
Для изучения временной программы транскрипции, которая лежит в основе индуцированного FVIIa ответа, в настоящей работе заявители исследовали ответ фибробластов человека на действие FVIIa с помощью микрочипов (микроматриц) для кДНК. Результаты показали, что клеточная экспрессия некоторых генов значительно изменяется в фибробластах после экспозиции с FVIIa. Одним из таких генов является Суr61, индуцируемый ростовым фактором промежуточный ранний ген, продукт которого, как показано, способствует клеточной адгезии, повышает индуцированный ростовым фактором синтез ДНК и стимулирует клеточную миграцию фибробластов и эндотелиальных клеток.
Краткое изложение существа изобретения
Настоящее изобретение относится к применению FVII и/или FVIIa, и/или другого агониста TF, и/или FVIIai, и/или другого антагониста TF в терапевтическом лечении патологических состояний, которые могут иметь отношение к миграции клеток или которые лечатся с помощью специфической регуляции клеточной миграции или хемотаксиса.
В другом варианте изобретение относится к применению FVII и/или FVII, и/или другого агониста TF, и/или FVIIai, и/или другого антагониста TF в терапевтическом лечении патологических состояний, которые могут иметь отношение к регуляции экспрессии, по меньшей мере, одного гена в клетке, например гена Суr61.
В другом варианте изобретение относится к способу индукции или усиления клеточной миграции, включающему стадию контактирования указанной клетки с агонистом тканевого фактора.
В одном осуществлении агонистом тканевого фактора является FVII или FVIIa.
В другом варианте изобретение относится к способу снижения или торможения клеточной миграции, включающему стадию контактирования клетки с антагонистом тканевого фактора.
В одном осуществлении антагонистом тканевого фактора является модифицированный FVII.
В одном осуществлении клетка представляет собой клетку человека, экспрессирующую тканевой фактор, включая фибробласты, гладкомышечные клетки, опухолевые клетки, гематопоэтические клетки и эпителиальные клетки.
В одном осуществлении модифицированный фактор FVII выбран из фактора VII, модифицированного с помощью Dansyl-Phe-Pro-Arg хлорметилкетона, Dansyl-Glu-Gly-Arg хлорметилкетона, Dansyl-Phe-Phe-Arg хлорметилкетона, Phe-Phe-Arg хлорметилкетона, Dansyl-D-Phe-Pro-Arg хлорметилкетона, Dansyl-D-Glu-Gly-Arg хлорметилкетона, Dansyl-D-Phe-Phe-Arg хлорметилкетона и D-Phe-Phe-Arg хлорметилкетона.
В другом варианте изобретение относится к способу индукции или усиления заживления ран у больного, включающему введение указанному больному эффективного количества фармацевтической композиции, включающей фактор VIIa или фактор VII, или другой агонист тканевого фактора, или их сочетание.
В другом варианте изобретение относится к способу торможения или снижения клеточной миграции, инвазии, индуцированной миграцией клеточной пролиферации или ангиогенеза у больного, страдающего заболеванием или состоянием, связанным с нежелательной клеточной миграцией, инвазией, индуцированной миграцией клеточной пролиферацией или ангиогенезом, включающему введение указанному больному эффективного количества фармацевтической композиции, включающей антагонист тканевого фактора.
В одном осуществлении заболевание или состояние представляет собой первичный опухолевый рост, опухолевую инвазию или метастаз.
В другом варианте изобретение относится к применению агониста тканевого фактора для производства лекарства для индукции или усиления клеточной миграции.
В другом варианте изобретение относится к применению антагониста тканевого фактора для производства лекарства для снижения или торможения клеточной миграции.
В другом варианте изобретение относится к способу регуляции экспрессии, по меньшей мере, одного гена в клетке, включающему стадию либо контактирования указанной клетки с агонистом тканевого фактора, либо контактирования указанной клетки с антагонистом тканевого фактора.
В одном осуществлении ген представляет собой ген, принадлежащий к семейству генов CCN.
В другом осуществлении ген выбран из группы, состоящей из Cyr61, CTFG, рецептора D2 дофамина, EST Incyte PD 395116 или рецептора P2U нуклеотидов.
В одном осуществлении ген представляет собой ген Cyr61.
В одном осуществлении регуляция представляет собой индукцию или усиление экспрессии. В другом осуществлении регуляция представляет собой снижение или торможение экспрессии.
В одном осуществлении FVII или FVIIa или другой агонист тканевого фактора индуцирует или усиливает экспрессию генов, а модифицированный FVII или другой антагонист тканевого фактора снижает или тормозит экспрессию генов, например, когда ген представляет собой ген, принадлежащий к семейству генов CCN или ген, выбранный из группы, состоящей из Cyr61, CTFG, рецептора D2 дофамина, EST Incyte PD 395116 или рецептора P2U нуклеотида.
В другом осуществлении FVII или FVIIa или другой агонист тканевого фактора снижает или тормозит экспрессию генов, а модифицированный FVII или другой антагонист тканевого фактора индуцирует или усиливает экспрессию генов, например, когда ген представляет собой EST PD674714.
Патологические состояния, которые можно лечить, представляют собой такие патологические состояния, как, например, атеросклероз, закрепление опухоли, опухолевый рост, опухолевая инвазия, метастаз или ангиогенез. Другие состояния, которые можно лечить, представляют собой, например, заживление ран, включая регенерацию стенок сосудов и лечение ожогов, или воспаление, или регуляцию клеточной миграции in vitro, как, например, при росте ткани.
Перечень фигур
Фиг.1А и 1В: Анализ с помощью проточной цитометрии экспрессии TF в фибробластах (1А). Клетки красили либо с помощью мышиного моноклонального антитела против IgG, конъюгированного с флуоресцеинизотиоцианатом (ФИТЦ) (незакрашенная область), что использовалось в качестве негативного контроля, либо с помощью моноклонального антитела против тканевого фактора (TF), конъюгированного с ФИТЦ (закрашенная область). На фиг.1В показана прокоагулянтная активность фибробластов. В фибробластах с экспрессией TF вызывалось 10-кратное увеличение РСА по сравнению с моноцитами, не экспрессирующими TF.
Фиг.2А: Действие FVIIa и FFR-FVIIa на индуцированный PDGF-BB хемотаксис фибробластов человека. ▪ показывает хемотаксический ответ фибробластов на различные концентрации PDGF-BB.
Фибробласты, инкубированные с 100 нМ FVIIa (•) или 100 нМ FFR-VIIa (о), мигрировали в направлении различных концентраций PDGF-BB. Результаты представляют собой среднее и SEM (стандартная ошибка средней) из трех отдельных экспериментов. Величины Р менее 0,05, * считались статистически достоверными (t-критерий Стьюдента).
Фиг.3 А-D: Влияние различных концентраций FVIIa или FFR-FVIIa на индуцированный PDGF-BB хемотаксис фибробластов. ▪ показывает миграцию фибробластов по направлению к различным концентрациям PDGF-BB. Клетки инкубировали с 12,5 (А), 25 (В), 50(С) и 100 (D) нМ FVIIa (•) или FFR-FVIIa (о) и анализировали в камере Boyden по отношению к различным концентрациям PDGF-BB. Результаты представляют собой среднее и SEM из трех отдельных экспериментов. *=р<0,05, **=р<0,01 и ***=р<0,001 по t-критерию Стьюдента.
Фиг.4А: смесь трех моноклональных антител против TF блокирует действие FVIIa и FFR-FVIIa на индуцированный PDGF-BB хемотаксис фибробластов. ▪ показывает миграцию фибробластов по направлению к PDGF-BB в отсутствие антител против TF, • фибробласты, преинкубированные с антителами против TF и 100 нМ FVIIa, и о фибробласты, преинкубированные с антителами против TF и 100 нМ FFR-FVIIa. Результаты представляют собой среднее и SEM из трех отдельных экспериментов.
Фиг.5А и 5В: Влияние FXa на хемотаксический ответ на PDGF-BB, индуцированный FVIIa. Фибробласты проинкубировали с 200 нМ ТАР (фиг.5А) (▪) или с 0,2-2 мкМ ТАР (фиг.5В) (▪) и затем с 100 нМ FVIIa (•). ТАР присутствовал во всех экспериментах. Хемотаксис индуцировали различными концентрациями PDGF-BB (5А) или 0,1 нг/мл PDGF-BB (5В). Результаты представляют собой среднее и SD (стандартное отклонение) двух отдельных экспериментов.
Фиг.6А: влияние тромбина на хемотаксический ответ на PDGF-BB, индуцированный FVIIa. Фибробласты проинкубировали с 5 Ед/мл (конечная концентрация) гирудина и затем с 100 нМ FVIIa. Гирудин присутствовал во всех экспериментах. Хемотаксис индуцировали различными концентрациями PDGF-B. ▪ показывает клетки, инкубированные с одним гирудином, а • - клетки с гирудином и FVIIa. Результаты представляют собой среднее и SD двух отдельных экспериментов.
Фиг.7А: влияние ингибирования Р13'-киназы на хемотаксис фибробластов, инкубированных с FVIIa. Клетки проинкубировали с различными концентрациями LY294002 в течение 30 мин при 37°С, а затем 100 нМ FVIIa (•) или без FVIIa (▪). Ингибитор присутствовал во время анализа хемотаксиса. Хемотаксис индуцировали 0,1 нг/мл PDGF-BB. Результаты представляют собой среднее и SD двух отдельных экспериментов.
Фиг.8А и 8В: влияние ингибирования PLC на хемотаксис фибробластов, инкубированных с FVIIa. Клетки инкубировали с различными концентрациями U73122 (активным ингибитором PLC) (8A) или U73343 (неактивным контролем) (8В) в течение 30 мин при 37°С, перед инкубацией в присутствии или в отсутствие 100 нМ FVIIa, и затем анализировали в камере Boyden по отношению к градиенту концентрации 0,1 нг/мл PDGF-BB. Агенты присутствовали во время всего эксперимента. ▪ показывает клетки с одним U73122 или U73343, • - клетки с U73122 или U73343 и FVIIa. Результаты представляют собой среднее и SD двух отдельных экспериментов.
Фиг.9: выделение инозитолтрифосфата (IP3) из фибробластов, стимулированных только FVIIa, FFR-FVIIa или в сочетании PDGF-BB. Клетки метили в течение ночи с помощью мио [3H] инозитола, инкубировали в присутствии или в отсутствие 100 нМ FVIIa или FFR-FVIIa в отсутствие или в присутствии 10 нг/мл или 100 нг/мл PDGF-BB. После этого клетки анализировали на предмет выделения 1Рз. Незакрашенные столбики показывают клетки без FVIIa или FFR-FVIIa (контроль), заштрихованные столбики показывают клетки с FFR-FVIIa, а черные столбики показывают клетки, инкубированные с FVIIa.
Фиг.10: фосфорилирование по тирозину PLC-γ1 в ответ на один PDGF-BB (контроль), FVIIa или FFR-FVIIa в сочетании с PDGF-BB. Клетки инкубировали с 100 нМ FVIIa или FFR-FVIIa в течение одного часа, а затем в присутствии или в отсутствие PDGF-BB в указанных концентрациях. Клетки лизировали, и фосфорилирование по тирозину PLC-γ1 определяли, как описано в методах.
Фиг.1. Нозерн-блот анализ, подтверждающий данные, полученные с помощью теста с применением микрочипов для кДНК. Десять г суммарной РНК (из тех же образцов РНК, которые применяли для выделения поли(А)РНК для получения зондов для гибридизации кДНК в микрочипах) были применены для Нозерн-блот анализа с применением в качестве зонда 32Р-меченным Суr61 (кДНК неполной длины, полученная от Genomic Systems). Вставка В. Сигналы гибридизации, количественно оцененные с помощью Phosphorimager (Molecular Dynamics).
Фиг.2 и 2В. Зависимая от времени индуцированная фактором VIIa экспрессия Суr61. Находящиеся в покое монослои клеток WI-38 обрабатывали фактором VIIa (5 г/мл) (2А) или PDGF-BB (10 нг/мл) (2В) в течение различных периодов времени. Суммарную РНК (10 г) подвергали Нозерн-блот анализу с применением в качестве зонда радиоактивно-меченного Суr61. Окрашенная бромистым этидием 28S рибосомальная РНК соответствующего блота показана на нижней вставке в качестве контроля внесения РНК.
Фиг.3. Дозозависимая индуцированная фактором VIIa экспрессия Суr61. Находящиеся в покое монослои клеток WI-38 обрабатывали различными дозами фактора VIIa, 0, 0,1, 0,5, 2,0 и 5,0 г/мл в течение 45 мин. Суммарную РНК (10 г) подвергали Нозерн-блот анализу с применением в качестве зонда радиоактивно-меченного Суr61. Окрашенная бромистым этидием 28S рибосомальная РНК соответствующего блота показана на нижней вставке в качестве контроля внесения РНК.
Фиг.4. Каталитическая активность фактора VIIa необходима для индуцированной экспрессии Суr61. Находящиеся в покое монослои клеток WI-38 обрабатывали контрольной бессывороточной средой или бессывороточной средой, содержащей фактор VIIa (5 г/мл) или инактивированным по активному сайту фактором VIIa (VIIai, 5 г/мл) в течение 45 мин. Суммарную РНК (10 г) подвергали Нозерн-блот анализу с применением в качестве зонда радиоактивно-меченного Суr61. Окрашенная бромистым этидием 28S рибосомальная РНК соответствующего блота показана на нижней вставке в качестве контроля внесения РНК.
Фиг.5. Индуцированная фактором VIIa экспрессия Суг61 не снимается специфическими ингибиторами фактора Ха и тромбина. Находящиеся в покое монослои клеток WI-38 обрабатывали контрольной средой и средой, содержащей фактор VIIa (5 г/мл; 100 нМ) в течение 45 мин. Клетки проинкубировали с 200 нМ рекомбинантного ТАР (дорожка 3) или гирудином (дорожка 4) в течение 30 мин перед экспозицией с фактором VIIa в течение 45 мин. Суммарную РНК (10 г) подвергали Нозерн-блот анализу с применением в качестве зонда радиоактивно-меченного Суr61. Окрашенная бромистым этидием 28S рибосомальная РНК соответствующего блота показана на нижней вставке в качестве контроля внесения РНК.
Фиг.6. Действие актиномицина D и циклогексимида на индуцированный фактором VIIa стационарный уровень мРНК Суr61. Находящиеся в покое монослои клеток WI-38 проинкубировали с контрольным растворителем, актиномицином D (10 г/мл) или циклогек-симидом (10 г/мл) в течение 30 мин перед тем как клетки экспонировали с фактором VIIa (5 г/мл) в течение 45 мин. Суммарную РНК (10 г) подвергали Нозерн-блот анализу с применением в качестве зонда радиоактивно-меченного Суr61. Окрашенная бромистым этидием 28S рибосомальная РНК соответствующего блота показана на нижней вставке в качестве контроля внесения РНК.
Фиг.7. Фактор VIIa индуцирует экспрессию CTGF.
Находящиеся в покое монослои клеток WI-38 обрабатывали фактором VIIa (5 г/мл) в течение различных периодов времени. Суммарную РНК (10 г) подвергали Нозерн-блот анализу с применением в качестве зонда радиоактивно-меченного CTGF. Окрашенная бромистым этидием 28S рибосомальная РНК соответствующего блота показана в качестве контроля внесения РНК.
Подробное описание изобретения
Настоящее изобретение относится к применению FVII или FVIIa или другого агониста TF для производства фармацевтической композиции для индукции или усиления клеточной миграции.
В другом варианте настоящее изобретение относится к применению FVII, FVIIa или другого агониста TF для производства фармацевтической композиции для индукции или усиления заживления ран или ангиогенеза.
В еще одном варианте настоящее изобретение относится к применению FVIIai или другого антагониста TF для производства фармацевтической композиции для ингибирования или предотвращения клеточной миграции.
В одном осуществлении предметом является клеточная миграция.
В другом варианте настоящее изобретение относится к применению FVIIai или другого антагониста TF для производства фармацевтической композиции для ингибирования или предотвращения ангиогенеза, метастазирования, опухолевого роста или опухолевой инвазии.
В другом варианте настоящее изобретение касается способа индукции или усиления клеточной миграции у больного, включающего введение указанному больному эффективного количества FVII или FVIIa или другого агониста TF.
В еще одном варианте настоящее изобретение касается способа торможения или предотвращения клеточной миграции у больного, включающего введение указанному больному эффективного количества FVIIai или другого антагониста TF.
В отдельном осуществлении эффективное количество представляет собой дневную дозу от приблизительно 5 мкг/кг/день до приблизительно 500 мкг/кг/день.
В другом осуществлении антагонист TF включает модифицированный FVIIa, например FFR-FVIIa.
В настоящем изобретении представляется механизм осуществления активности FVII и/или FVIIa, который касается стимуляции клеточной миграции. Такой механизм создает основу для выявления участия FVII и/или FVIIa в патологических состояниях, в которых принимают участие экспрессирующие TF клетки, подобные эндотелиальным клеткам, эпителиальным клеткам, фибробластам, гладкомышечным клеткам и моноцитам/макрофагам. Более того, в изобретении предлагается основа для идентификации конкретных фармакологических мишеней, которые уместны для терапевтического вмешательства.
Таким образом, настоящее изобретение относится к применению FVII и/или FVIIa и/или FVIIai для терапевтического лечения патологических состояний, которые могут быть связаны с клеточной миграцией или которые могут быть подвергнуты лечению с помощью специфической регуляции клеточной миграции.
В другом варианте настоящее изобретение относится к способу выявления потенциальных лекарств, которые регулируют клеточную миграцию, при этом данный способ включает
a) культивирование экспрессирующей TF клетки;
b) измерение миграции клетки;
c) инкубацию клетки с потенциальным лекарством и
d) измерение миграции инкубированной клетки и определение какого-либо изменения уровня миграции по сравнению с миграцией, измеренной на стадии b, причем такое изменение является показателем биологической активности потенциального лекарства в указанной клетке.
В целом компоненты крови, участвующие в том, что было обозначено как коагуляторный "каскад", представляют собой проферменты или зимогены, ферментативно неактивные белки, которые превращаются в протеолитические ферменты под действием активатора, который сам является активированным фактором свертывания. Факторы коагуляции, которые уже подверглись такому превращению, и обычно обозначаемые как "активные факторы" обозначаются добавлением буквы "а" к наименованию коагуляторного фактора (например, фактор VIIa).
Термин "цинковый хелатор" предназначен для включения соединения, которое связывает фактор VIIa и индуцирует замещение ионов кальция на ионы цинка в факторе VIIa, тем самым ингибируя активность фактора VIIa или комплекса тканевой фактор-фактор VIIa (TF-FVIIa).
Подходящим антагонистом TF согласно изобретению может быть хелатирующее цинк соединение, например, дигидроксамат или дигидразид с группами гидроксамата или гидразида, расположенными друг относительно друга таким образом, что они способны хелатировать ион цинка. Хелатирующее цинк соединение действует в сочетании с FVIIa. Ионы Zn2+ проявляют свое тормозное действие за счет конкуренции со стимулирующим действием ионов Са2+. Предполагается, что ионы Zn2+ вытесняют ионы Са2+ из одного или более сайта(сайтов) связывания кальция в FVIIa. Хелатирующие цинк соединения, например, гидроксаматы и гидразиды, способны действовать как сильные агенты, способствующие связыванию ионов цинка при конкуренции с ионами кальция. Тем самым, специфические соединения потенцируют ингибирование цинком активности комплекса фактор VIIa/тканевой фактор. Активность фактора VIIa в комплексе с тканевым фактором может быть ингибирована с помощью механизма, в котором хелатор цинка связывается с фактором VIIa и облегчает замещение Са2+ на Zn2+. За счет данного действия хелатор оказывает модулирующее влияние на TF при нормальной концентрации свободных ионов Са2+ и Zn2+ в крови.
Термин "FVII" или "фактор VII" обозначает "одноцепочечный" (зимогенный) коагуляторный фактор VII. Термин "фактор VIIa" или "FVIIa" обозначает "двухцепочечный" активированный коагуляторный фактор VII, расщепленный за счет специфического расщепления по пептидной связи Arg152-Ile153. FVII и FVIIa могут быть выделены из крови или получены рекомбинантными способами. Очевидно, что применение способов, описанных здесь, не зависит от происхождения очищенного фактора VIIa и, следовательно, настоящее изобретение рассматривается как охватывающее применение любых препаратов фактора VII или FVIIa, пригодных для указанного здесь применения. Предпочтительным является FVIIa человека. FVII или FVIIa также предназначены для включения вариантов FVII, в которых один или более аминокислотных остатков был(были) заменен(ы).
Термин "модифицированный фактор VII", "инактивированный FVII" или "FVIIai" предназначен для обозначения FVIIa, содержащего, по меньшей мере, одну модификацию в каталитическом центре, которая существенно снижает способность модифицированного FVIIa активировать FX и FIX. Термины можно применять взаимозаменяемо. Такая модификация включает аминокислотное замещение (или замену) одного или более остатков из каталитической триады Ser344, Asp142 и His193, а также включает модификацию остатков каталитической триады ингибиторами сериновой протеазы, такими как фосфорорганические соединения, сульфанилфторид, пептид галометилкетон или азапептид. FFR-FVIIa служит одним из примеров производного FVIIai, полученного с помощью блокады активного центра FVIIa необратимым ингибитором, D-фенилаланин-L-фенилаланин-L-аргининхлорметилкетон (FFR cmk). Другими подходящими производными FVIlai являются инактивированный FVIIa, полученный или который может быть получен путем блокирования активного центра с помощью L-фенилаланин-L-фенилаланин-L-аргининхлорметилкетона, дансил-L-фенилаланин-L-фенилаланин-L-аргининхлорметилкетона или дансил-D-фенилаланин-L-фенилаланин-L-аргининхлорметилкетона. Предпочтительным является FFR-FVIIa (FVIIa, инактивированный с помощью FFR cmk).
Термин "протеинкиназа" предназначен для обозначения фермента, способного фосфорилировать серии и/или треонин, и/или тирозин в пептидах и/или белках.
Термин "потенциальное лекарство" предназначен для обозначения любого образца, имеющего биологическую функцию или оказывающего биологическое действие в клеточной системе. Образец может быть образцом биологического материала, таким как микробиологический или растительный экстракт, или он может быть образцом, содержащим соединение или смесь соединений, полученных с помощью органического синтеза или генетическими методами.
Термин "агонист TF" включает соединения, индуцирующие
a) передачу сигнала путем прямого связывания с TF (например, FVIIa),
b) стимуляцию каскада МАРК,
c) блокирование ингибирования МАРК (например, ингибиторы РТР-азы),
в которых агонисты являются потенциальными лекарствами, как определено выше.
Термин "антагонист FT" включает
a) агенты, которые конкурируют с FVIIa за связывание с TF без проведения сигнала, например FVIIai,
b) агенты, которые связываются с FVIIa и предотвращают связывание с TF, например Zn-гидроксамат,
с) агенты, которые тормозят проведение сигнала, препятствуя членам каскада МАРК,
d) агенты, которые связываются с FVIIa/TF и предотвращают проведение сигнала,
e) агенты, которые связываются с FVIIa/TF/FX и предотвращают проведение сигнала,
f) агенты, которые блокируют активацию фактора Х человека, катализируемую комплексом тканевой фактор/фактор VIIa человека,
причем данные антагонисты являются потенциальными лекарствами, как определено выше.
Термин "фармакологические мишени" предназначен для обозначения белка, который может менять миграцию экспрессирующих TF клеток.
Термин "репортерный ген" предназначен для обозначения конструкции ДНК, которая после транскрипции дает белок, который может быть выявлен.
Термин "промоторный элемент SRE" обозначает последовательность ДНК, которая связывает транскрипционные факторы, индуцируемые компонентами, присутствующими в сыворотке.
Термин "экспрессирующая TF клетка" обозначает любую клетку млекопитающего, которая экспрессирует TF.
Термин "фосфорилирование белка" предназначен для обозначения фосфорилирования серина и/или треонина, и/или тирозина в пептидах и/или белках.
Модуляция или регуляция клеточной миграции определяется как способность FVIIa или другого агониста TF, или FVIIai или другого антагониста TF 1) повышать или снижать характер, нормальный или ненормальный, протекающей клеточной миграции, 2) инициировать нормальную клеточную миграцию и 3) инициировать ненормальную клеточную миграцию.
В данном контексте термин "лечение" включает как предотвращение неблагоприятного состояния, так и регуляцию уже существующего состояния с целью торможения или сведения к минимуму этого состояния. Таким образом, профилактическое введение FVIIa или другого агониста TF, или FVIIai или другого антагониста TF включается в термин "лечение".
В данном контексте термин "одна единица" определяется как количество фактора VII, присутствующее в 1 мл нормальной плазмы, соответствующее приблизительно 0,5 мкг белка. После активации 50 единиц соответствуют приблизительно 1 мкг белка.
В данном контексте термин "больной" определяется как любое животное, в частности млекопитающее, такое как человек. Термин "субъект" применяют взаимозаменяемо с термином "больной".
Сокращения
Тканевой фактор (TF) является клеточным рецептором фактора VIIa (FVIIa), и комплекс служит главным инициатором свертывания крови. Заявители изучали влияние связывания FVIIa с TF на клеточную миграцию и проведение сигнала фибробластов человека, которые экспрессируют TF на высоком уровне. Фибробласты, инкубированные с FVIIa, мигрировали в направлении концентрационного градиента PDGF-BB при концентрации приблизительно в сто раз меньшей, чем в случае фибробластов, не связанных с FVIIa. Антитела против TF ингибировали повышение хемотаксиса, вызванное FVIIa/TF. Более того, при ингибированном по активному сайту FVIIa (FFR-FVIIa) наблюдалось выраженное подавление хемотаксиса, индуцированного PDGF-BB. Была исключена возможность того, что гиперхемотаксис индуцируется предполагаемым образованием FXa и активностью тромбина.
FVIIa индуцировал образование инозитол-1,4,5-трифосфата в той же степени, что и PDGF-BB; эффекты FVIIa и PDGF-BB были аддитивными. FFR-FVIIa не вызывал какого-либо выделения инозитол-1,4,5-трифосфата. Ответ на PDGF-BB и FVIIa в виде клеточной миграции полностью блокировался ингибитором PLC, что позволяет предположить, что для ответа важна активация PLC. Таким образом, связывание FVIIa с TF может быть независимым от свертывания, может модулировать клеточные ответы, такие как хемотаксис и для этого необходима каталитическая активность FVIIa.
Полагают, что TF играет роль в метастазировании опухолевой клетки, но механизм пока не известен. Однако, совсем недавно Ott et al. идентифицировали связывающийся с актином белок 280 (АВР-280) в качестве лиганда цитоплазматического домена TF, что служит молекулярным путем, посредством которого TF может поддерживать метастазирование опухолевой клетки. Молекулярные сигналы и биологические функции, трансдуцируемые FVIIa/TF, являются, однако, еще малопонятными.
В фибробластах человека имеет место конститутивная экспрессия TF. Данные клетки экспрессируют также рецепторы фактора роста тромбоцитов (PDGF). PDGF индуцирует в своих клетках-мишенях митогенность, реорганизацию актина и направленную клеточную миграцию (хемотаксис). Ранее заявители показали, что PDGF-BB является эффективным хемотаксическим фактором для фибробластов человека и что хемотаксический ответ опосредуется рецептором β-класса. Таким образом, данные клетки были выбраны для изучения предполагаемой трансдукции сигнала и клеточной миграции, индуцированной связыванием FVIIa с TF.
Ниже заявители впервые показывают четкую связь между передачей сигнала, индуцированного связыванием FVIIa с TF, и клеточным ответом на ростовой фактор. Заявители представляют результаты, свидетельствующие о том, что в фибробластах человека комплекс FVIIa/TF ведет к гиперхемотаксическому ответу на PDGF-BB. Более того, ингибированный по активному сайту FVIIa (FFR-FVIIa) дозозависимо подавлял направленную миграцию к PDGF-BB. С помощью применения специфических ингибиторов PLC и фосфатидили-нозитол-3'-киназы (PI3'-киназы) заявители также показывают, что гиперхемотаксический ответ по направлению к PDGF-BB, индуцированный сигналом FVIIa/TF, зависит от активности фосфолипазы С (PLC), но не зависит от PI3'-киназы. FVIIa и PDGF-BB аддитивным образом индуцировали образование инозитол-1,4,5-трифосфата (IP3), одного из вторичных посредников, выделяющихся после активации PLC.
TF конститутивно экспрессируется на плазматической мембране многих экстраваскулярных клеток, таких как фибробласты стромы в сосудистой адвентициальной оболочке и в фиброзных капсулах печени, селезенки и почек. Таким образом, экспрессия TF обнаруживается в местах, физически отделенных от циркулирующей крови и обеспечивающих гемостатическую оболочку. После травмы данный барьер, как полагают, защищает организм от потери крови. TF может, однако, индуцироваться в моноцитах/макрофагах, гладкомышечных клетках сосудов, эндотелиальных клетках и ряде опухолевых клеток под действием множества агентов, включая цитокины и ростовые факторы. Индукция на транскрипционном уровне происходит вскоре после стимуляции, что указывает на TF как относящийся к росту немедленно-ранний ген.
В данной работе заявители исследовали роль TF в качестве рецептора с сигнальной функцией. Заявители показывают, что фибробласты человека с конститутивной экспрессией TF после связывания лиганда FVIIa мигрируют по направлению к крайне низким концентрациям PDGF-BB. Один TF/FVIIa не индуцировал повышенную спонтанную миграцию, т.е. случайную миграцию. Таким образом, сочетание внутриклеточного проведения сигнала FVIIa/TF и ростового фактора PDGF-BB было необходимо для достижения ответа в виде подвижности. Обязательным было не только связывание с TF, но и каталитическая активность TF/FVIIa, поскольку ингибированный по активному сайту FVIIa не вызывал повышенного миграционного ответа. Более того, ингибирующие моноклональные антитела предотвращали увеличение хемотаксического ответа под действием FVIIa. Заявители также исключили возможность того, что происходит непрямая передача сигнала за счет FXa или тромбина, поскольку ТАР и гирудин не оказывали влияния на индуцированный FVIIa/TF хемотаксис. Вместо этого заявители обнаружили, что возрастающие концентрации FFR-FVIIa активно ингибируют индуцированный PDGF-BB хемотаксис. Фибробласты, инкубированные с FFR-FVIIa, проявляли совершенно нормальную случайную миграцию. Ингибирующее действие FFR-FVIIa на индуцированный PDGF-BB хемотаксис не наблюдалось в присутствии сочетания антител против TF, что тем самым исключает возможность того, что FFR-FVIIa является токсичным. Результаты скорее позволяют предположить, что в клетках, экспрессирующих β-рецепторы PDGF и TF, комплекс FVIIa/TF важен для хемотаксического ответа на PDGF-BB.
Данные заявителей о том, что FVIIa повышает образование IP3, и ранее сообщавшиеся данные о том, что FVIIa/TF индуцирует осцилляции Са2+, особенно в клетках MDCK, убедительно поддерживают мнение о том, что в ряде клеток PLC активируется в результате передачи сигнала FVIIa/TF. Кроме того, гиперхемотаксический ответ фибробластов на PDGF-BB, индуцированный FVIIa/TF, блокировался дозозависимо ингибитором PLC. Заявители ранее обнаружили сходный гиперхемотаксический ответ на PDGF-BB в клетках Y934F с мутантным β-рецептором PDGF, в которых наблюдалось повышенное фосфорилирование и активация PLC-γ1. В данных клетках повышенное фосфорилирование PLC-γ1 коррелировало с троекратно повышенным образованием IP3 по сравнению с клетками, экспрессирующими β-рецептор PDGF дикого типа. Сочетание FVIIa/TF и PDGF-BB вызывало приблизительно двукратное увеличение образования IP3 в фибробластах человека. Индуцированное FVIIa/TF образование IP3, однако, не коррелировало с фосфорилированием PLC-γ1. Нельзя исключить индуцированное FVIIa/TF фосфорилирование по тирозину PLC-γ2, но это кажется маловероятным, поскольку экспрессия PLC-γ2 в фибробластах человека очень низка. Более того, внутриклеточная часть TF не наделена внутренней протеин-тирозинкиназной активностью. Данные результаты позволяют предположить, что FVIIa/TF индуцирует активацию β и/или δ изоферментов PLC. При анализе выделения IP3 в культуральную среду клеток добавляли 0,1% FBS, содержащую лишь приблизительно 0,1 нМ FXa. Заявители обнаружили, что для индукции образования IP3 требуется концентрация FXa, превышающая 20 нМ. Остается неизвестным механизм, посредством которого активируется изоферменты β или δ PLC. Полагают, что активация включает кооперацию между TF и ассоциированным с мембраной белком.
Позднее была обнаружена связь TF с цитоскелетом. Было показано молекулярное взаимодействие между цитоплазматическим доменом TF и белком, связывающим актиновый филамент АВР 280. Более того, было найдено, что TF находится в тесном контакте с актином и белками, связывающими актиновый филамент, такими как α-актин и АВР280 в ламеллиподиях и в областях складчатой мембраны распространяющихся эпителиальных клеток. АВР 280, член филаминового подсемейства, необходим для нормального функционирования ламеллиподий и, следовательно, очень важен для клеточной подвижности. Изоферменты РI3'-киназы и PLC участвуют в хемотаксических ответах, таких как мобилизация связывающих актин белков. В предшествующих исследованиях заявители наблюдали, что PI3'-киназный путь в индуцированном β-рецептором PDGF хемотаксисе, по-видимому, менее важен в клетках с гиперэкспрессией и повышенной активностью PLC-γ1. To же самое наблюдалось в случае клеток с FVIIa, связанным с TF. Это указывает на то, что величина активации изоферментов РI3'-киназы и PLC должна определять, какой из данных путей будет доминирующим. Взятые вместе указанные данные показывают, что клеточная миграция является важной морфогенетической функцией, индуцируемой проведением сигнала FVIIa/TF.
Хемотаксис играет ведущую роль в заживлении ран, ангиогенезе и метастазировании. Хемотаксис также представляет собой важный компонент развития атеросклеротических бляшек. В данных процессах множество клеток экспрессирует TF, а также PDGF и рецепторы PDGF. Рестеноз является главным осложнением после процедуры вмешательства в обструктивные артерии. Было показано, что PDGF вовлечен в ответ стенок сосудов (образование неоинтимы) на механическое повреждение путем опосредования миграции и пролиферации гладкомышечных клеток и фибробластов. Заявители впервые показали, что связывание FVIIa с экспрессирующими TF клетками вызывает повышенный хемотаксический ответ на PDGF, который не зависит от свертывания.
В настоящее время немного известно о пути(путях) проведения сигнала, индуцированного протеолитически активным VIIa, и о том, как генерируемые VIIa сигналы могли бы участвовать в клеточных процессах. Одна из возможностей заключается в том, что FVIIa мог бы индуцировать экспрессию ростовых регуляторов, действующих на следующих этапах в индукции клеточных процессов. Для исследования данной возможности в настоящей работе заявители определяли изменения в транскрипционной программе фибробластов человека в ответ на экспозицию с VIIa с применением микрочипов (микроматриц) кДНК, которые содержат более 8000 отдельных генов человека. Заявители выбрали фибробласты, поскольку данные клетки в норме контактируют с сывороткой, которая содержит ростовые факторы и активированные факторы свертывания в контексте сосудистой травмы из-за физических (например, хирургических) и патофизиологических состояний. Временная программа генной экспрессии, наблюдаемая в ответ на сыворотку, позволяет предположить, что фибробласты запрограммированы интерпретировать внезапную экспозицию с сывороткой не только как общий митогенный стимул, но и как специфический физиологический сигнал. Характеристика транскрипционной активации в ответ на сыворотку и ростовые факторы предполагает также, что фибробласты служат активными участниками в коммуникации между различными клетками, которые совместно контролируют воспаление, ангиогенез и заживление ран.
Анализ с помощью кДНК-микрочипов мРНК, выделенной из фибробластов, экспонированных с VIIa в течение 90 мин, показывает повышение экспрессии Суr61. Нозерн-блот анализ подтвердил индуцированную VIIa экспрессию Суr61 в фибробластах. VIIa повышает также экспрессию Суr61 в гладкомышечных клетках сосудов, хотя и не столь выражение, как в фибробластах. Индукция экспрессии Суr61 зависит от каталитической активности FVIIa, поскольку FVIIai не способен индуцировать экспрессию Суr61. Хотя фактор Ха и тромбин также могли индуцировать экспрессию Суr61 (данные не показаны), данные соединения не участвуют в индуцированной FVIIa экспрессии Суr61. Заявители не обнаружили доказательств образования следов фактора Ха и тромбина в их экспериментальной системе. Более того, специфический ингибитор фактора Ха и тромбина не оказывал существенного влияния на индуцированную FVIIa экспрессию Суr61.
Cyr61 является немедленно-ранним геном, транскрипционно активируемым в фибробластах ростовыми факторами сыворотки. Он кодирует 40-кДа, обогащенный цистеином гепаринсвязывающий белок, который ассоциирует с внеклеточным матриксом и клеточными поверхностями. Cyr61 является членом растущего семейства генов, кодирующих консервативные модульные белки, для которых характерно наличие N-концевого секреторного сигнала, последующих четырех модульных структурных доменов и 38 цистеиновых остатков, большей частью консервативных среди членов семейства. В настоящее время семейство белков состоит из шести отдельных членов, включая Cyr61, фактор роста соединительной ткани (CTGF) и онко-белок птиц Nov (что, таким образом, дало наименование семейству CCN) (Семейство CCN описывается дополнительно в Lau et al., Exp. Cell Res 248: 44-57, 1999). Показано, что белок Cyr61 (i) стимулирует прикрепление и распространение эндотелиальных клеток подобно действию фибронектина, (ii) усиливает действие bFGF и PDGF на скорость синтеза ДНК в фибробластах и эндотелиальных клетках сосудов, (iii) способствует клеточной миграции как фибробластов, так и эндотелиальных клеток. Недавние исследования показывают, что Cyr61 действует в качестве лиганда интегрина αγβ3 рецептора адгезии, известный как вовлеченный в сигнальные пути, которые регулируют ряд клеточных процессов, включая ангиогенез и метастазирование опухолей. Очищенный белок Cyr61, как показано, стимулирует непосредственную миграцию микроваскулярной эндотелиальной клетки человека в культуре через αγβ3 зависимый путь и индуцирует неоваскуляризацию роговицы крыс. Более того, экспрессия Cyr61 в опухолевых клетках стимулирует опухолевый рост и васкуляризацию.
Основываясь на настоящих данных, которые показывают, что FVIIa индуцирует экспрессию Суr61 в фибробластах, полагают, что Суr61, индуцированный FVIIa, ответственен за опосредованную FVIIa клеточную миграцию и метастазирование опухоли за счет действия через интегрин αγβ3. Таким образом, Суr61 связывает протеолитический сигнал FVIIa-TF с интегриновым сигнальным путем. Наблюдения о том, что каталитическая активность VIIa необходима для миграции гладкомышечных клеток и опухолевых клеток, и для метастазирования опухоли, находятся в соответствии с другим наблюдением, свидетельствующим о том, что каталитическая активность FVIIa необходима для индукции Суr61.
В дополнение к Cyr61 VIIa мог бы также индуцировать другие регуляторы, которые могли бы опосредовать индуцированные FVIIa биологические ответы. Было показано, что связывание FVIIa с TF поверхности опухолевых клеток поджелудочной железы вызывает избирательную гиперэкспрессию гена uPAR. Ранее с помощью метода дифференциального дисплея заявители показали стимуляцию транскрипции гена поли(А)полимеразы в фибробластах, экспонированных с FVIIa. Хотя было бы интересно выяснить, покажет ли также способ с кДНК микрочипами дифференциальную экспрессию РАР, фильтр не содержал кДНК РАР. Кроме Cyr61, микрочипы с кДНК заявителей также показали дифференциальную экспрессию четырех других генов (смотри результаты), но соотношение дифференциальной экспрессии было очень близким к границе статистической значимости. Поскольку в предварительных экспериментах заявители не смогли подтвердить их дифференциальную экспрессию с помощью Нозерн-блот анализа, а также в результате отсутствия каких-либо предположительно имеющих отношение данных о способности продуктов данных генов опосредовать индуцированные FVIIa биологические ответы, заявители далее не проводили анализа их экспрессии. Однако, поскольку CTGF является структурно сходной с Суr61 молекулой и вызывает те же биологические ответы, что и Суr61, заявители исследовали экспрессию CTGF, несмотря на то, что относительное соотношение экспрессии CTGF в обработанном FVIIa образце по сравнению с контрольным образцом в анализе с помощью кДНК микрочипов равно 1,8 (реальной величиной в анализе обычно считается 2). Результаты показали, что FVIIa также индуцирует экспрессию CTGF и что кинетика индуцированной VIIa экспрессии CTGF сходна с таковой Суr61.
Хотя CTGF ведет себя весьма сходно с Суr61, между ними существуют незначительные различия. Например, (а) CTGF, как показано, митогенен сам по себе, в то время как Суr61 не имеет присущей ему митогенной активности, но усиливает индуцируемый ростовыми факторами синтез ДНК, (b) Суr61 стимулирует хемотаксис, в то время как CTGF стимулирует как хемотаксис, так и хемокинез, (с) хотя как Суr61, так и CTGF являются ассоциированными с ЕСМ сигнальными молекулами, CTGF, как показано, секретируется в культуральную среду. Таким образом, возможно, что FVIIa местно регулирует клеточные функции через Суr61, поскольку оказывает дистантное действие, влияя на секрецию CTGF.
Схема лечения любого больного, подвергаемого лечению FVIIa или другим агонистом TF, или FVIIai или другим антагонистом TF, как здесь отмечается, должна быть определена специалистом в данной области. Суточная доза, которая вводится при лечении, может быть определена врачом и должна зависеть от конкретного применяемого соединения, от способа введения и от веса и состояния больного. Эффективное количество, которое подходит как суточная доза, составляет от приблизительно 5 мкг/кг/день до приблизительно 500 мкг/кг/день, предпочтительно от приблизительно 10 мкг/кг/день до 300 мкг/кг/день, более предпочтительно от приблизительно 15 мкг/кг/день до 200 мкг/кг/день, наиболее предпочтительно от приблизительно 20 мкг/кг/день до 100 мкг/кг/день.
FVIIa или другой агонист TF, или FVIIai или другой антагонист TF должны вводиться в виде однократной дозы, но их можно также давать в виде множественных доз, предпочтительно с интервалами в 4-6-12 часов в зависимости от получаемой дозы и состояния больного.
FVIIa или другой агонист TF, или FVIIai или другой антагонист TF могут вводиться внутривенно или они могут быть введены с помощью непрерывной или периодической инфузии, или они могут быть введены прямо в подходящее место, так, например, введены непосредственно в опухоль. FVIIa или другой агонист TF, или FVIIai или другой антагонист TF предпочтительно вводят с помощью внутривенных инъекций и в количестве приблизительно 100-100000 единиц на кг массы тела и предпочтительно в количестве приблизительно 250-25000 единиц на кг массы тела, что соответствует приблизительно 5-500 мкг/кг, при этом доза может быть повторена 2-4 раза в течение 24 часов.
Традиционные способы для получения фармацевтических композиций, которые могут быть применены в соответствии с настоящим изобретением, описаны, например, в Remington's PharmaceuticalSciences, 1985.
Композиции, применяемые в соответствии с настоящим изобретением, получают с помощью способов, известных per se специалистам в данной области.
Вкратце, фармацевтические препараты, которые подходят для применения в соответствии с настоящим изобретением, готовят путем смешивания FVII, FVIIa или другого агониста TF, или FVIIai или другого антагониста TF, предпочтительно в очищенной форме, с подходящими адъювантами и подходящим носителем или разбавителем. Подходящие физиологически приемлемые носители или разбавители включают стерильную воду или физиологический раствор. Подходящие в данном отношении адъюванты включают кальций, белки (например, альбумины) или другие инертные пептиды (например, глицилглицин), или аминокислоты (например, глицин или гистидин) для стабилизации очищенного фактора VIIa. Другие физиологически приемлемые адъюванты представляют собой невосстановливаемые сахара, многоатомные спирты (например, сорбит, маннит или глицерин), полисахариды, такие как декстрины низкого молекулярного веса, детергенты (например, полисорбат) и антиоксиданты (например, бисульфит и аскорбат). Адъюванты обычно присутствуют в концентрации от 0,001 до 4% вес/об. Фармацевтический препарат может также содержать ингибиторы протеаз, например апротинин, и консервирующие агенты.
Препараты могут быть стерилизованы, например, с помощью фильтрации через задерживающий бактерии фильтр, путем включения в композиции стерилизующих агентов, с помощью облучения композиций или путем нагревания композиций. Они могут быть также получены в форме стерильных твердых композиций, которые можно растворять в стерильной воде или в какой-либо другой стерильной среде, подходящей для введения, заблаговременно или немедленно перед применением.
В различных вариантах настоящее изобретение касается:
Способа регуляции экспрессии, по меньшей мере, одного гена в клетке, включающего стадии:
a) контактирования указанной клетки с фактором VII (а) или антагонистом тканевого фактора
b) определения экспрессии указанного гена в указанной клетке.
Указанного выше способа, в котором указанная клетка представляет собой клетку сосудов человека, экспрессирующую тканевой фактор, включая фибробласты и гладкомышечные клетки.
Способа, в котором указанный ген выбран из группы, состоящей из Cyr61, CTFG, D2 рецептора дофамина, EST Incyte PD 395116 или P2U рецептора нуклеотидов.
Способа, в котором указанный антагонист тканевого фактора представляет собой модифицированный фактор VII (а), известный как фактор VIIai.
Способа, в котором экспрессия указанного гена увеличена.
Способа, в котором экспрессия указанного гена ингибирована или сведена к минимуму.
Способа увеличения экспрессии указанного гена, включающего контактирование клетки с фактором VIIa.
Способа торможения экспрессии указанного гена, включающего контактирование клетки с модифицированным фактором VII, известным как FVIIai.
Способа, в котором указанный ген представляет собой EST PD674714.
Способа регуляции клеточной миграции, включающего стадии:
a) контактирования указанной клетки с фактором VIIa или антагонистом тканевого фактора;
b) определения миграции указанной клетки.
Способа, в котором указанная клетка представляет собой клетку человека, экспрессирующую тканевой фактор, включая фибробласты, гладкомышечные клетки, опухолевые клетки, гематопоэтические клетки и эпителиальные клетки.
Способа, в котором антагонист тканевого фактора представляет собой модифицированный фактор VIIa, известный как фактор VIIai.
Способа, в котором модифицированный фактор VII выбран из Dansyl-Phe-Pro-Arg хлорметилкетона, Dansyl-Glu-Gly-Arg хлорметилкетона, Dansyl-Phe-Phe-Arg хлорметилкетона и Phe-Phe-Arg хлорметилкетона.
Способа увеличения миграции клеток, включающего контактирование клетки с FVIIa или с агонистом тканевого фактора.
Способа прекращения или торможения миграции клеток, включающего контактирование клетки с антагонистом тканевого фактора.
Способа индукции или увеличения заживления ран у больного, включающего введение указанному больному эффективного количества фармацевтической композиции, включающей фактор VIIa или агонист тканевого фактора.
Способа торможения инвазивности опухолевых клеток, включающего контактирование указанных клеток с эффективным количеством антагониста тканевого фактора.
Способа торможения клеточной миграции, инвазии, клеточной пролиферации, индуцированной миграцией, или ангиогенеза у больного с заболеванием или состоянием, связанным с нежелательной клеточной миграцией, инвазией, клеточной пролиферацией, индуцированной миграцией, или ангиогенезом, включающего введение указанному больному эффективного количества фармацевтической композиции, включающей антагонист тканевого фактора.
Способа, в котором заболевание или состояние представляет собой первичный опухолевый рост, инвазию опухоли или метастазирование.
Способа, в котором антагонист тканевого фактора представляет собой модифицированный фактор VII, известный как FVIIai.
Применения фактора VIIa или антагониста тканевого фактора для получения лекарства для регуляции клеточной миграции.
Применения, в котором фактор VIIa применяют для получения лекарства для увеличения клеточной миграции.
Применения, в котором антагонист тканевого фактора применяют для получения лекарства для прекращения или торможения клеточной миграции.
Способа, в котором антагонист тканевого фактора представляет собой модифицированный фактор VIIa, известный как фактор VIIai.
Применения, в котором модифицированный фактор VII выбран из Dansyl-Phe-Pro-Arg хлорметилкетона, Dansyl-Glu-Gly-Arg хлорметилкетона, Dansyl-Phe-Phe-Arg хлорметилкетона и Phe-Phe-Arg хлорметилкетона.
Настоящее изобретение дополнительно иллюстрируется следующими примерами, которые, однако, не созданы как ограничивающие объем защищаемого изобретения. Детали, раскрытые в последующем описании и в последующих примерах, могут как отдельно, так и в любом их сочетании быть материалом для осуществления изобретения в его различных формах.
Примеры
Пример 1
Получение FVII
Очищенный фактор VIIa человека, пригодный в настоящем изобретении, предпочтительно получают методом рекомбинантных ДНК, например, как описано Hagen et al., Proc. Natl. Acad. Sci. USA 83: 2412-2416, 1986 или как описано в Европейском патенте No. 200421 (ZymoGenetics). Фактор VIIa, полученный с помощью рекомбинантной технологии, может быть аутентичным фактором VIIa или в той или иной степени модифицированным фактором VIIa, представленным таким образом, что такой фактор VIIa по существу имеет такую же биологическую активность в отношении свертывания крови, как и аутентичный фактор VIIa. Такой модифицированный фактор VIIa может быть получен путем модификации последовательности нуклеиновой кислоты, кодирующей фактор VIIa, либо путем изменения кодонов аминокислот, либо путем удаления некоторых из кодонов аминокислот в нуклеиновой кислоте, кодирующей природный FVII, с помощью известных способов, например, с помощью сайт-направленного мутагенеза.
Фактор VIIa может быть также получен с помощью способов, описанных Broze and Majerus, J. Biol. Chem. 255(4): 1242-1247, 1980 и Hedner and Kisiel, J. Clin. Invest. 71: 1836-1841, 1983. Данные способы дают фактор VII без детектируемых количеств других факторов свертывания крови. Еще более очищенный препарат фактора VII может быть получен путем включения дополнительной гельфильграции в качестве завершающей стадии очистки. После этого фактор VII превращают в активированный VIIa с помощью известных способов, например, с помощью нескольких различных белков плазмы, таких как фактор XIIa, IXa или Ха. В другом варианте, как описано Bjoem et al. (Research Disclosure, 269 September 1986, pp. 564-565), фактор VII может быть активирован пропусканием через колонку для ионообменной хроматографии, такую как Mono Q® (Pharmacia fine Chemicals) или ей подобную.
Пример 2
Получение FVIIai
Модифицированный фактор VII, пригодный в настоящем изобретении, получают, например, как описано в Международных публикациях Nos. 92/15686, 94/27631, 96/12800 и 97/47651 ZymoGenetics/Novo Nordisk).
Пример 3
Влияние FVIIa и FFR-FVIIa на хемотаксический ответ фибробластов на PDGF-BB
Фибробласты, экспрессирующие активный TF (фиг.1А и фиг.1В), инкубировали с 100 нМ FVIIa и высевали в верхней части модифицированной камеры Boyden, тогда как среду, содержащую 10% FBS и PDGF-BB в различных концентрациях, вносили ниже 150 мкм микропорового фильтра. Миграцию клеток в условиях, когда среду, содержащую 10% FBS без PDGF-BB, вносили ниже фильтра, использовали в качестве показателя случайной миграции и принимали за 100% миграцию. Значительный миграционный ответ был отмечен при концентрации PDGF-BB 0,01 нг/мл в клетках, стимулированных FVIIa, при сравнении с 1 нг/мл PDGF-BB в случае клеток, не связанных с FVIIa, т.е. при 100-кратной разнице в концентрации (фиг.2А). При концентрациях PDGF-BB 0,01-0,1 нг/мл миграционный ответ на FVIIa увеличивался дозо-зависимо, начиная с 25 нМ, с максимальным эффектом при 50-100 нМ FVIIa (фиг.3A-D). Увеличение случайной миграции не наблюдалось после активации с помощью VIla. Для того чтобы проверить, является ли протеолитически активный FVIIa обязательным для гиперхемотаксического ответа на PDGF-BB, фибробласты также инкубировали с 100 нМ FFR-FVIIa и анализировали в камере Boyden сходным образом (фиг.2А). При низких концентрациях PDGF-BB, 0,01-1 нг/мл, повышенного хемотаксиса с FFR-FVIIa не наблюдалось. Напротив, при 100 нМ FFR-FVIIa достигалось выраженное подавление хемотаксиса, индуцированного 10-50 нг/мл PDGF-BB (фиг.2А и 3A-D). Когда фибробласты предварительно инкубировали со смесью трех различных антител против TF, а затем с FVIIa или FFR-FVIIa, миграционный ответ на PDGF-BB был равным ответу фибробластов в отсутствие лиганда, связанного с TF (фиг.4А). Неспецифичное моноклональное IgG антитело не предотвращало гиперхемотаксис, индуцированный FVIIa, не тормозило миграционный ответ, вызванный FFR-FVIIa (данные не показаны). Наличие IgG антител или трех антител против TF не изменяло случайную миграцию фибробластов (данные не показаны).
Пример 4
Гиперхемотаксический ответ не опосредуется FXa или тромбином
Поскольку трансдукция индуцированного FVIIa сигнала, ведущего к гиперхемотаксическому ответу на PDGF-BB, зависела от каталитической активности FVIIa, было важно определить, осуществляется ли проведение сигнала прямо, или через FXa или тромбин, образующиеся посредством комплекса FVIIa/TF. Ответ в виде повышенной миграции, опосредуемый FVIIa/TF, не блокировался 0,2-10 мкМ антикоагулянтного пептида клеща (ТАР), специфически блокирующего активный центр FXa и предотвращающего последующую активацию каскада коагуляции, ведущего к образованию тромбина (фиг.5А,5В). Добавление 5 Ед/мл гирудина, специфического ингибитора тромбина, не оказывало никакого влияния на индуцированный FVIIa гиперхемотаксис (фиг.6А). ТАР и гирудин не влияли на миграцию фибробластов в ответ на PDGF в отсутствие лиганда FVIIa (фиг.6А, 5В, 6А). Таким образом, маловероятно, чтобы влияние FVIIa на хемотаксис опосредовалось через активацию FX или тромбина.
Пример 5
На гиперхемотаксический ответ на PDGF-BB влияют зависимые от PLC пути, но не влияет РI3'-киназа.
Недавно было показано, что РI3'-киназа важна для индуцируемого через β-рецептор PDGF хемотаксиса. В связи с этим, заявители исследовали способность LY294002, специфического ингибитора РI3'-киназы, блокировать хемотаксический ответ, индуцированный за счет сигнала FVIIa/TF. Перед добавлением 100 нМ FVIIa фибробласты предварительно обрабатывали LY294002 в указанной концентрации в течение 30 минут при 37°С и анализировали в камере Boyden как описано. На протяжении эксперимента концентрацию PDGF-BB поддерживали на постоянном уровне 0,1 нг/мл, т.е. при очень низкой концентрации, при которой FVIIa/TF индуцировал значительный хемотаксический ответ. LY294002 присутствовал на всем протяжении экспериментов. На фиг.7А показано, что миграционный ответ на PDGF-BB, опосредуемый проведением сигнала FVIIa/TF, не изменялся при ингибировании РI3'-киназы.
Для изучения вопроса о том, участвует ли активация специфичной в отношении фосфатидилинозитолов фосфолипазы С (PLC) в индуцируемом FVIIa/TF хемотаксическом ответе, заявители предварительно инкубировали фибробласты с различными концентрациями U73122, специфического ингибитора PLC, в течение 30 минут при 37°С перед добавлением 100 нМ FVIIa; после этого клетки подвергали анализу на хемотаксис в присутствии ингибитора. Близкий аналог, U73343, не оказывающий влияния на PLC, применялся в качестве отрицательного контроля. В этих экспериментах также поддерживали концентрацию PDGF-BB на постоянном уровне, равном 0,1 нМ. Предварительная обработка клеток активным ингибитором PLC, U73122, дозо-зависимо тормозила гиперхемотаксический ответ на 0,1 нг/мл PDGF-BB с полным подавлением при 1 мкМ (фиг.8А и 8В). При применении неактивного аналога, U73343, действие на хемотаксис не наблюдалось.
Пример 6
FVIIa/TF индуцирует активацию PLC
Для дальнейшей оценки важности активности PLC для гиперхемотаксического ответа заявители проанализировали также прямое действие FVIIa/TF на активность PLC в фибробластах. Активация PLC ведет к образованию двух вторичных посредников, инозитол-1,4,5-трифосфата (IP3) и диацилглицерина. Фибробласты инкубировали с мио [3H]инозитолом в течение ночи и затем с 100 нМ FVIIa или FFR-FVIIa в течение 60 минут с последующей инкубацией в отсутствие или в присутствии PDGF-BB в указанных концентрациях. Обработка одним FVIIa в концентрации 100 нМ в течение 60 минут вызывала выделение IP3 в фибробластах в той же степени, что и обработка одним PDGF-BB в концентрации 10 нг/мл и 100 нг/мл (фиг.9). Более того, сочетание 100 нМ FVIIa и 10 нг/мл или 100 нг/мл PDGF-BB вызывало двукратное повышение выделения 1Рз. Ингибированный по активному центру FVIIa не вызывал выделение 1Рз. Данные результаты ясно показывают, что PLC активируется после связывания FVIIa с TF.
Пример 7
Фосфорилирование PLC-γ1 не увеличивается при проведении сигнала TF/FVIIa в фибробластах
Для того чтобы определить, ответственна ли изоформа PLC-γ1, активируемая некоторыми рецепторами-тирозинкиназами, за повышенную активность PLC, индуцируемую FVIIa/TF, исследовали фосфорилирование по тирозину PLC-γ1. Фибробласты инкубировали в отсутствие или в присутствии 100 нМ FVIIa или FFR-FVIIa в течение одного часа с последующей стимуляцией с 0, 2, 10 или 100 нг/мл PDGF-BB. Через 5 минут инкубации клетки лизировали, PLC-γ1 иммунопреципитировали, выделяли с помощью электрофореза в ПАГ с Na-ДДС и подвергали иммуноблоттингу с помощью антител против фосфотирозина. Хотя было зафиксировано значительное повышение фосфорилирования по тирозину PLC-γ1 при возрастающих концентрациях PDGF-BB, добавление одного FVIIa к фибробластам не вызывало какого-либо фосфорилирования по тирозину PLC-γ1 (фиг.10). Более того, сочетание FVIIa и PDGF-BB в различных концентрациях не индуцировало дополнительного фосфорилирования по сравнению со стимуляцией одним PDGF-BB (фиг.10). FFR-FVIIa не влиял на Фосфорилирование по тирозину PLC-γ1 (фиг.10). Таким образом, за повышение активности PLC после стимуляции FVIIa ответственны изоформы PLC, отличные от PLC-γ1.
Пример 8
Способы
Клеточные культуры. Фибробласты кожи человека, AG1518 и AG1523 выращивали до состояния конфлуентности в среде Игла MEM с добавкой 10% фетальной телячьей сыворотки (FBS). Перед применением клетки разделяли обработкой трипсином (2,5 мг/мл в течение 10 мин при 37°С), промывали уравновешенным солевым раствором Хенка и ресуспендировали в среде Игла MEM с 10% FBS или среде Хема с добавкой 0,1% FBS.
Белки. FVIIa человека (Novo Nordisk A/S, Gentofte, Дания) экспрессировали и очищали. FFR-FVIIa (Novo Nordisk) получали путем блокирования FVIIa по активному центру с помощью D-Phe-L-Phe-L-Arg-хлорметилкетона. Рекомбинантный противосвертывающий пептид клеща (ТАР) был любезно предоставлен д-ром Р.Vlasuk, Corvas (San Diego, CA). Гирудин получен от Sigma. LY294002, U73122.и U73343 были получены от Biomol (Plymouth Meeting, PA). Моноклональные антитела против TF, TF8-5G9, TF9-5B7 и MTFH-1 (Morrissey, J.H., Fair, D.S., Edgington, T.S. Monoclonal antibody analysis of purified and cell-associated tissue factor. Thromb. Res. 52, 247-261 (1988)) были любезно предоставлены д-ром James H. Morrissey, Oklahoma Medical Research Foundation. Антитело против фосфотирозина, PY99 получено от Santa Cruz, Калифорния.
Проточная цитометрия. Поверхностную экспрессию TF анализировали с помощью иммунофлуоресценции с применением проточного цитометра (Coulter Epics XL-MCL, Beckman Coulter, Fullerton, CA, Coulter Electronics, США). Аппарат ежедневно калибровали калибровочными шариками Immuno-Check™ или Flow-Check™ Coulter. Для экспериментов с непрямой иммунофлуоресценцией фибробласты AG1518 или AG1523 промывали дважды ЗФР, содержащим 0,1% бычьего сывороточного альбумина (БСА), инкубировали в течение 30 минут на льду с меченым флуоресцеинизотиоцианатом (FITC) моноклональным антителом против TF человека (4508CJ, American Diagnostica, Greenwich, Ct, США). В качестве отрицательного контроля применяли моноклональное IgGI против глюкозоксидазы Aspergillus niger (Dakopatts). В каждом образце определяли среднюю интенсивность флуоресценции канала (MFI) и процент позитивных клеток.
Определение активности TF. Прокоагулянтную активность TF определяли, как описано Lindmark et al. (Lindmark E., Tenno Т., Chen, J., Siegbahn, A. IL-10 inhibits LPS-induced human monocyte tissue factor expression in whole blood. Br. J. Haematol. 102, 597-604 (1998)). Вкратце, аликвоты, содержащие 0,2×105 фибробластов AG1518 или AG1523, дважды промывали ЗФР и помещали в лунки 96-луночного планшета для микротитрования (Nunc, Roskilde, Дания). Прокоагулянтную активность измеряли двустадийным амидолитическим способом, в котором хромогенный субстрат, S-2222 (Chromogenix, Mölndal, Швеция), расщепляют за счет FXa, который, в свою очередь, образуется в результате активации FX под действием комплекса TF/FVIIa. В лунки добавляли реакционную смесь, содержащую в конечной концентрации 0,6 мМ S-2222, 0,2 мМ CaCl2 и факторы свертывания из концентрата факторов Prothromplex-T™ TIM4 (Baxter, Vienna, Австрия) в конечной концентрации 1 Ед/мл FVII и 1,2 Ед/мл FX, и измеряли изменение в поглощении при 405 нм через 30 минут инкубации при 37°С. Измерения проводили в трех параллелях.
Тестирование хемотаксиса. Миграционный ответ фибробластов определяли методом лидирующего фронта в модифицированной камере Boyden, как описано ранее (Siegbahn A., Hanimacher A., Westermark В., Heldin C-H. Differential effects of the various isoforms of platelet-derived growth factor on chemotaxis of fibroblasts, monocytes, and granulocytes. J. Clin. Invest. 85, 916-920 (1990) и Nister M., Hammacher A., Mellström К., Siegbahn A., Ronnstrand L., Westermark В., Heldin C-H. A glioma-derived PDGF A chain homodimer has different functionsi activities from a PDGF AB heterodimer purified from human platelets. Cell 52, 791-799 (1988)). Микропоровые фильтры (с размером пор 8 мкм) покрывали раствором коллагена типа 1 при комнатной температуре в течение ночи. Фильтры сушили на воздухе в течение 30 минут непосредственно перед применением. Фибробласты кожи человека AG1523 выращивали до состояния конфлуентности в среде Игла MEM с добавкой 10% FBS. Клетки отделяли обработкой трипсином (2,5 мг/мл в течение 10 мин при 37°С) и суспендировали в среде Игла MEM с 10% FBS. Перед анализом фибробласты инкубировали в течение 10 минут в присутствии или в отсутствие FVIIa или FFR-VIIa. Поверх фильтра камеры Boyden наносили сто микролитров суспензии клеток (2×105 клеток/мл). PDGF-BB разводили средой для анализа (средой Игла MEM с 10% FBS) и вносили под фильтр камеры. Клетки инкубировали в течение 6 часов при 37°С во влажной камере, содержащей 95% воздуха/5% CO2. FVIIa или FFR-VIIa присутствовали на протяжении всего эксперимента. После этого фильтры отделяли, фиксировали в этаноле, окрашивали Mayer's Hemalun и монтировали на стекле. Миграцию измеряли как расстояние, на котором оказались ядра двух наиболее далеко мигрировавших фибробластов в фокусе одного поля с большим увеличением (12,5×24). Расстояние миграции для каждого фильтра рассчитывали как среднее показаний, по меньшей мере, для трех различных частей фильтра. В экспериментах применяли от двух до четырех отдельных фильтров для каждой концентрации хемоаттрактанта. В каждой серии экспериментов в качестве контроля служила миграция фибробластов в направлении среды тестирования.
В случаях, когда применяли моноклональные антитела против TF или ингибиторы факторов свертывания, ТАР и гирудин, клетки проинкубировали в течение 10 минут с этими агентами, а затем с или без FVIIa или FFR-VIIa перед проведением тестирования хемотаксиса. Антитела, ТАР или гирудин присутствовали также на протяжении всего эксперимента по хемотаксису. В экспериментах по тестированию влияния различных ингибиторов, LY294002, U73122 или U73343, на миграционный ответ, клетки проинкубировали в течение 30 минут с ингибиторами в указанной концентрации, и ингибиторы присутствовали также на протяжении всего эксперимента.
Анализ выделения инозитолтрифосфата (IРз). Шестилуночные планшеты с полуконфлуентными культурами фибробластов человека AG1518 инкубировали в течение ночи (приблиз. 20 часов) с 2 мкКи мио (3H)инозита (Amersham) в 2 мл среды Хема F12 с 0,1% FBS. Среду заменяли средой Хема F12 с 0,1% FBS (содержащую 2 мМ CaCl2) и 20 мМ LiCl, и клетки инкубировали в течение 15 минут при 37°С. После этого клетки инкубировали в отсутствие или в присутствии 100 нМ FVIIa или 100 нМ FFR-VIIa в течение одного часа. Добавляли PDGF-BB (0, 10 или 100 нг/мл), и инкубацию продолжали в течение 10 минут при 37°С. Анализ IP3 проводили, как описано ранее Eriksson et al. (Eriksson A., Nanberg E., Rönnstrand L., Engström U., Rupp E., Carpenter G., Heldin C-H., Claesson-Welsh L. Demonstration of functionally different interactions between phospholipase C-γ and the two types of platelet-derived growth factor receptors. J. Biol. Chem. 270, 7773-7781 (1995)).
Анализ индуцируемого агонистом фосфорилирования под действием PLC-γ1. Полуконфлуентные культуры AG1518 истощали в отношении сыворотки в течение ночи (приблизительно 20 часов) в среде, содержащей 0,1% FBS, и затем инкубировали в отсутствие или в присутствии 100 нМ FVIIa или FFR-FVIIa в течение одного часа с последующей инкубацией с 0, 2, 10 или 100 нг/мл PDGF-BB в течение 5 минут при 37°С. Клетки лизировали и PLC-γ1 осаждали в высокой степени, как описано ранее (Hansen К., Johnell М., Siegbahn A., Rorsman С., Engstrom U., Wernstedt С., Heldin С-Н., Ronnstrand L. Mutation of a Src phosphorylation site in the PDGF β-receptor leads to increased PDGF-stimulated chemotaxis but decreased mitogenesis. EMBO J. 15, 5299-5313 (1996)), с помощью анти-PLC-γ1 антисыворотки, выработанной путем иммунизации кроликов пептидом, соответствующим карбоксильному концу PLC-γ1 быка (Artega C.L., Johnson M.D., Todderud G., Coffey R.J., Carpenter G., Page D.L. Elevated content of the tyrosine kinase substrate phospholipase C-γ1 in primary human breast carcinomas. Proc. Natl. Acad. Sci. USA 88, 10435-10439 (1991). Образцы разделяли с помощью ДСН-ПААГ электрофореза и проводили иммуноблоттинг с антителом к фосфотирозину PY99.
Статистический анализ. Данные анализировали с применением пакета программ Statistica ТМ для Windows (StatSoft, Tulsa, Okla. USA). t-критерий Стьюдента для зависимых образцов применяли для определения статистической достоверности различий между разными рядами данных. Величины р<0,05 рассматривали как статистически достоверные.
Белки. Рекомбинантный VIIa человека, предоставленный Novo Nordisk (Gentofte, Дания), восстанавливали в стерильной воде до концентрации от 1 до 1,3 мг/мл. Маточные растворы VIIa проверяли на загрязнение следовыми количествами эндотоксина с применением амебоцитного лизата из limulus (Bio Whittaker) и ничего обнаружено не было (уровень определения - 30 пг). Рекомбинантный противосвертывающий белок клеща (ТАР) был любезно предоставлен George Vlasuk, (Corvas, San Diego, CA) и рекомбинантный гирудин получен либо от Sigma (St. Louis, МО), либо от Calbiochem (San Diego, CA). Очищенный фактор Ха человека и тромбин были получены от Enzyme Research Laboratories (Southbend, IN).
Анализ с помощью кДНК-микрочипов. Клетки WI-38 культивировали до состояния 80% конфлуентности, и на 24 часа удаляли сыворотку для введения в состояние покоя, как описано выше. Культуральную среду заменяли свежей бессывороточной DMEM (с добавкой 5 мМ CaCl2), и давали клеткам стабилизироваться в течение 2 час в культуральном инкубаторе. После этого клетки обрабатывали очищенным рекомбинантным VIIa (5 мкг/мл) в течение 90 мин. По завершении 90 мин обработки выделяли суммарную РНК из необработанных (контрольных) и обработанных VIIa клеток с помощью Trizol (GIBCO BRL). Поли(А)РНК очищали двукратным пропусканием через колонки Oligo Тех для выделения мРНК, как описано в технической инструкции производителя (Qiagen). Восемьсот нг (800 нг) высокоочищенной поли(А)РНК из контрольных и обработанных VIla клеток отправляли в службу анализа с помощью кДНК-микрочипов (Human UniGEM V microarray, Genome Systems Inc, St. Louis, МО).
Нозерн-блот анализ. Суммарную РНК получали с помощью реагента TRIZOL из покоящегося монослоя клеток WI-38, экспонированных с VIla и другими материалами, как описано ранее. Нозерн-блот анализ проводили стандартным способом. Вкратце, 10 мкг суммарной РНК фракционировали по размеру с помощью гель-электрофореза в гелях 1% агароза/6% формальдегид и переносили на нитроцеллюлозную мембрану с помощью способа капиллярного блоттинга. Нозерн-реплики прегибридизовали при 42°С в растворе, содержащем 50% формамид, 5×SSC, 50 мМ Трис. HCl, рН 7,5, 0,1% пирофосфат натрия, 1% ДДС-Na, 1% поливинилпирролидон, 1% Ficoll, 25 мМ ЭДТА, 100 мкг/мл денатурированной ДНК из молок лососевых и 1% БСА, и гибридизовали с 32Р-меченым зондом кДНК Суr61 (106 имп/мин/мл). Гибридизованные мембраны экспонировали либо с Dupont NEF, либо с Fuji RX рентгеновской пленкой. Для целей количественной оценки мембраны экспонировали с фосфорным экраном в течение от 1 до 4 час, и экспонированные экраны анализировали в Phosphorimager (Molecular Dynamics) с помощью программы "image-quant". Для получения средних величин единицы (импульсы), полученные в различных экспериментах, нормализовали по внутреннему контролю (импульсы, присутствующие в контрольном-обработанном образце).
Хромогенный анализ. Клетки WI-38 культивировали в 96-луночном культуральном планшете и вводили их в состояние покоя, как описано выше. После промывки клеток в культуральные лунки, содержащие клетки, или лунки, заполненные буфером (без клеток), добавляли FVIIa (5 мкг/мл) в 1,00 мкг содержащего кальций буфера. Через 30 мин инкубации в лунки добавляли 25 мкг хромогенных субстратов для фактора Ха и тромбина, т.е. Chromozym X и Chromozym TH. Через 3 ч развития окраски снимали показания с планшета на микропланшетном ридере. В качестве контролей клетки инкубировали со следовыми концентрациями фактора Ха (от 50 до 0,1 нг/мл) или тромбина (от 0,1 до 0,002 Ед/мл). Не обнаружено различий в поглощении при 450 нм между лунками с VIIa, добавленным к клеткам, или VIIa, добавленным в лунки, не содержащие клетки. Показания были ниже, когда показания снимали при наименьшей концентрации фактора Ха или тромбина, и они представляют собой хромогенную активность VIIa.
Пример 9
Анализ с помощью микрочипов кДНК. Покоящиеся фибробласты экспонировали с контрольной бессывороточной средой или с бессывороточной средой с добавленным VIIa (5 мкг/мл) в течение 90 мин (3 сосуда Т-75 для каждой обработки). После обработки собирали суммарную РНК и выделяли поли(А)РНК. Шестьсот нг мРНК метили либо Су3, либо Су5 флуоресцентной меткой и затем гибридизовали с чипом UniGem Human V, содержащим 8000 ESTs с подтвержденными последовательностями, представляющими до 5000 известных генов человека (платная услуга, предоставляемая Genome System Inc). Применение контрольного планшета, в котором известные концентрации кДНК сравнения вносили в реакцию образования зонда для измерения чувствительности и отслеживания реакции обратной транскрипции, определения очистки, эффективности гибридизации и общей оценки качества и прохождения анализа, показало успешность процесса гибридизации. Общий анализ экспериментальных данных показал минимальные различия в сигналах гибридизации между контрольными и обработанными VII образцами. Лишь небольшое количество генов показало умеренно дифференцированную экспрессию. Заявители обнаружили повышение экспрессии 5 генов (от 3,5- до 2-кратного повышения при обработке VIIa), в то время как экспрессия одного гена снижалась после обработки VIIa (2,4-кратное снижение) (+/-2 является принятой оценкой определения минимальной величины действительных соотношений). Индивидуальность гена с повышенной в 3,5 раза экспрессией не была обнаружена из-за его особой природы. Другие гены, стимулируемые VIIa, представляют собой Суr61 (2,5 раз), рецептор D2 дофамина (2,2 раза), EST Incyte PD 395116 (2 раза) и рецептор P2U нук-леотидов (2 раза). Интересно отметить, что CTGF, ген, принадлежащий к семейству Суr61, экспрессировался в обработанных VIIa клетках в 1,8 раза больше, чем в контрольных клетках. Транскрипт со сниженной экспрессией в обработанных VIIa клетках представлял собой EST PD674714. Заявители выбрали для дальнейшего анализа Суr61.
Пример 10
Подтверждение дифференциальной экспрессии Суr61. Для подтверждения данных, полученных с микрочипом, заявители использовали образцы РНК из контрольных и обработанных VIIa клеток (те же самые образцы РНК, что и применявшиеся для получения поли (А) РНК для получения зонда в микрочиповом анализе) для Нозерн-блот анализа с применением в качестве зонда радиоактивно-меченной кДНК Суr61. Данные показывают, что зонд кДНК Суr61 гибридизуется с единственным транскриптом (приблизительно 2,0 т.п.о.) РНК, выделенной из контрольных и обработанных VIIa клеток. Однако интенсивность гибридизационного сигнала была значительно выше в случае РНК, выделенной из клеток, обработанных VIIa (фиг.1). Количественная оценка сигнала гибридизации показала, что экспрессия Суr61 была в 2,8 раза выше в клетках, экспонированных с VIIa, по сравнению с контрольными обработанными клетками.
Пример 11
Кинетика индуцированной VIIa экспрессии Суr61. Для определения кинетики экспрессии Суr61 покоящиеся фибробласты обрабатывали в течение различных периодов времени 5 мкг/мл VIIa. Суммарную РНК экстрагировали и подвергали Нозерн-блот анализу. Как показано на фиг.2, экспрессия Суr61 возрастала в зависимости от времени в обработанных VIIa клетках. Экспрессия достигала максимума через приблизительно 45 мин, после чего снижалась до базального уровня в пределах от 2 до 3 ч. Поскольку сообщалось, что экспрессия Суr61 в мышиных фибробластах после стимуляции сывороткой и ростовым фактором поддерживается в течение нескольких часов (до от 8 до 10 ч) до наступления репрессии, заявители исследовали действие сыворотки и PDGF на кинетику экспрессии Суr61 в покоящихся фибробластах человека, WI-38. Как показано на фиг.2В, Суr61 экспрессируется лишь временно после стимуляции PDGF и становится полностью репрессированным через 2 ч после стимуляции. Сходные результаты были получены с индуцированной сывороткой экспрессией Суr61 (данные не показаны).
Пример 12
Зависимая от дозы фактора VIIa индукция экспрессии Суr61. Для определения дозозависимости от VIIa покоящиеся фибробласты обрабатывали различными дозами FVIIa (от 0,1 до 5 мкг/мл) в течение 45 мин, после чего образцы суммарной РНК клеток подвергали Нозерн-блот анализу. Как показано на фиг.3, обработка фибробластов уже такой низкой концентрацией FVIIa, как 0,1 мкг/мл, была достаточна для индукции экспрессии Суr61, а концентрация FVII(a) в плазме (0,5 мкг/мл, 10 нМ приводила к выраженному ответу, близкому к максимальному.
Пример 13
Каталитическая активность фактора VIIa необходима для индукции Суr61. Для проверки того, что необходима ли каталитическая активность VIIa для индукции Суr61, клетки WI-38 обрабатывали VIIa и инактивированным по активному центру FVIIa (FVIIai) в течение 45 мин, и экспрессию Суr61 оценивали с помощью Нозерн-блот анализа. Как показано на фиг.4, FVIlai был не способен индуцировать экспрессию Суr61, что свидетельствует о необходимости протеолитической активности FVIIa. В данном контексте важно отметить, что было показано, что FVIIai связывается с TF клеточной поверхности с тем же или более высоким сродством по сравнению с FVIIa. Кажется маловероятным, что индуцированная VIIa экспрессия Суr61 в экспериментах заявителей была обусловлена образованием нижележащих факторов свертывания, FXa и тромбина. С помощью чувствительного хромогенного анализа заявители не обнаружили доказательств образования фактора Ха и тромбина в их экспериментальной системе (чувствительность определения - 10 пг). Более того, специфические ингибиторы фактора Ха и тромбина, т.е. противосвертывающий белок клеща и гирудин соответственно, были не способны предотвращать индуцированную VIIa экспрессию Суr61 (фиг.5).
Пример 14
Участие транскрипционного механизма в индукции стационарного уровня мРНК Суr61 под действием VIIa. Для исследования того, участвует ли транскрипция в опосредуемом VIIa повышении стационарного уровня мРНК Суr61, покоящиеся клетки WI-38 инкубировали с актиномицином D (10 мкг/мл) в течение 30 мин перед добавлением VIIa на 45 мин. Как показано на фиг.6, актиномицин D ингибировал стимулирующее действие VIIa. Данный результат указывает на транскрипционный механизм индукции Суr61.
Для исследования того, необходим ли синтез белка de novo для индукции мРНК Суr61 под действием VIIa, клетки WI-38 предварительно обрабатывали ингибитором синтеза белка циклогексимидом перед экспозицией клеток с VIIa в течение 45 мин. Как показано на фиг.6, стимулирующий эффект VIIa не блокировался циклогексимидом. Напротив, циклогексимид значительно повышал индуцированный VIIa стационарный уровень мРНК Суr61.
| название | год | авторы | номер документа |
|---|---|---|---|
| МОДИФИЦИРОВАННЫЙ ФАКТОР VII | 1995 |
|
RU2214833C2 |
| НОВЫЕ АНТИТЕЛА К ТКАНЕВОМУ ФАКТОРУ В КАЧЕСТВЕ АНТИКОАГУЛЯНТОВ | 2003 |
|
RU2345789C2 |
| СПОСОБ МОДУЛИРОВАНИЯ ВАСКУЛЯРИЗАЦИИ | 2005 |
|
RU2378006C2 |
| ПРОИЗВОДНЫЕ ФАКТОРА VII СВЕРТЫВАНИЯ КРОВИ | 2002 |
|
RU2338752C2 |
| НОВЫЕ СЛИТЫЕ БЕЛКИ ТРОМБОМОДУЛИНА, ОБЕСПЕЧИВАЮЩИЕ НАПРАВЛЕННЫЙ ПЕРЕНОС К ТКАНЕВОМУ ФАКТОРУ, В КАЧЕСТВЕ АНТИКОАГУЛЯНТОВ | 2003 |
|
RU2320366C2 |
| ПОЛИПЕПТИД ФАКТОРА VII СВЕРТЫВАНИЯ КРОВИ, ЕГО ПОЛУЧЕНИЕ И ПРИМЕНЕНИЕ | 2001 |
|
RU2326126C2 |
| ПРИМЕНЕНИЕ ЧИГЛИТАЗАРА И РОДСТВЕННЫХ ЕМУ СОЕДИНЕНИЙ | 2019 |
|
RU2769446C1 |
| ХИМЕРНЫЕ МОЛЕКУЛЫ ФАКТОРА VII | 2010 |
|
RU2563231C2 |
| ПОЛИПЕПТИДЫ НА ОСНОВЕ МОДИФИЦИРОВАННОГО ФАКТОРА VII И ИХ ПРИМЕНЕНИЕ | 2008 |
|
RU2571931C2 |
| МОДИФИЦИРОВАННЫЙ КОАГУЛИРУЮЩИЙ ФАКТОР VIIa С ПРОДЛЕННЫМ ВРЕМЕНЕМ ПОЛУЖИЗНИ | 2007 |
|
RU2466141C2 |
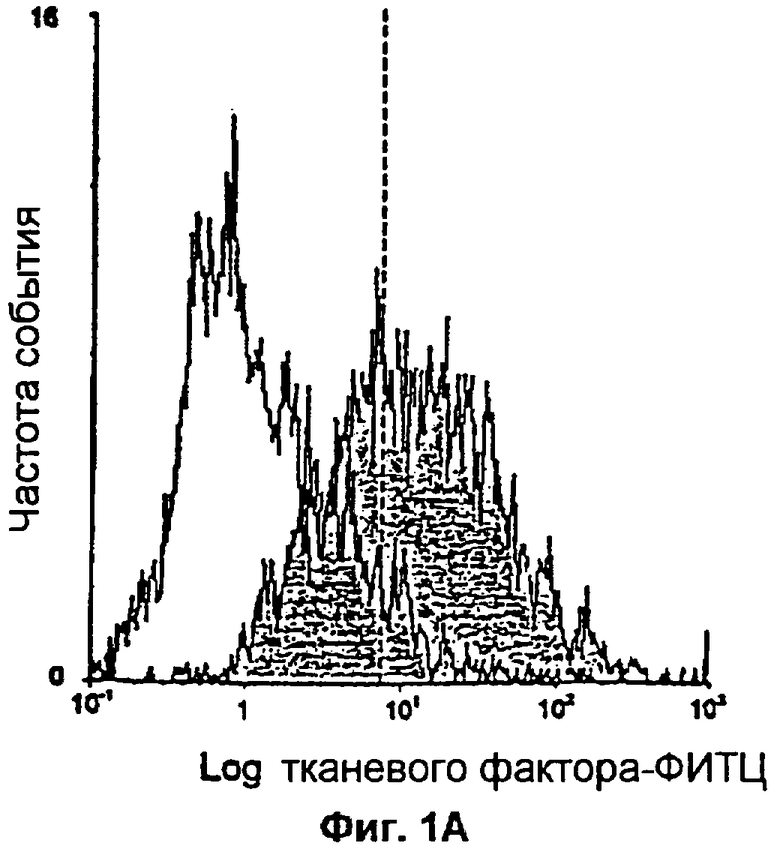

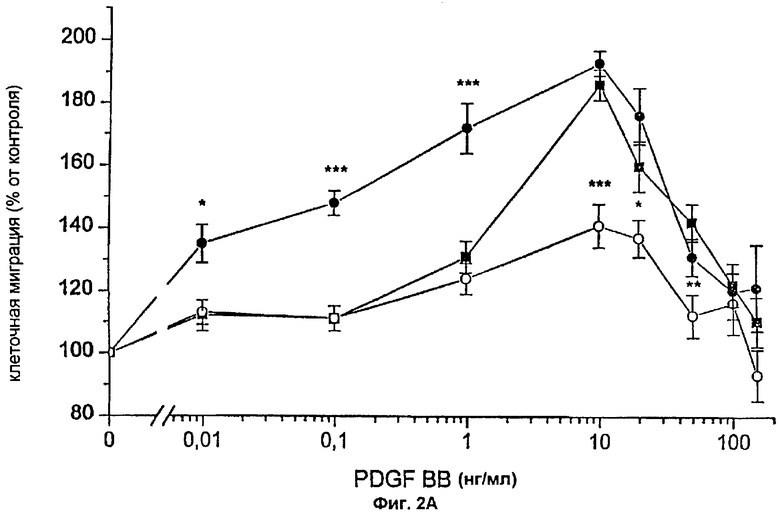

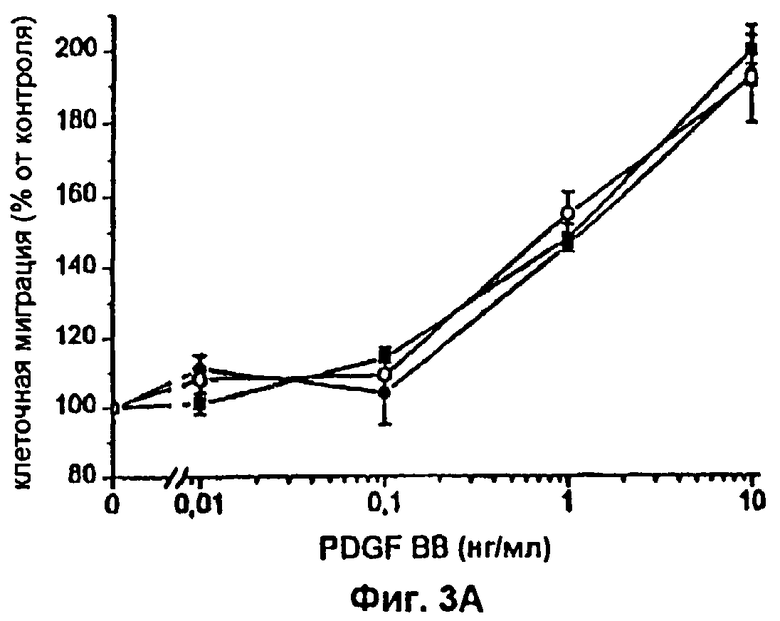
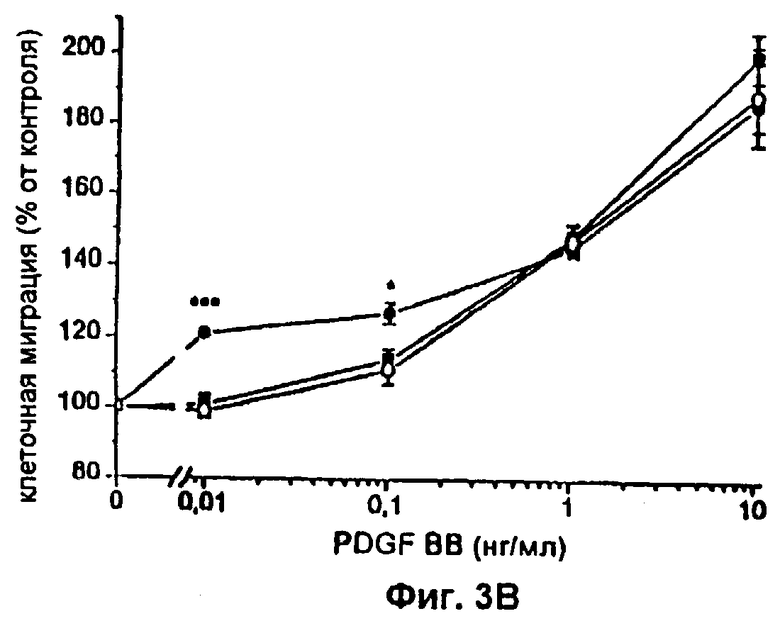

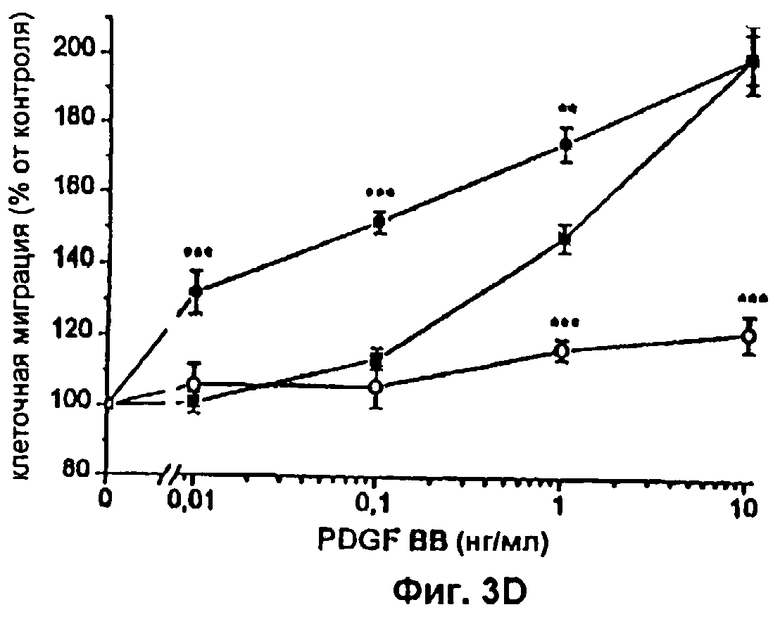
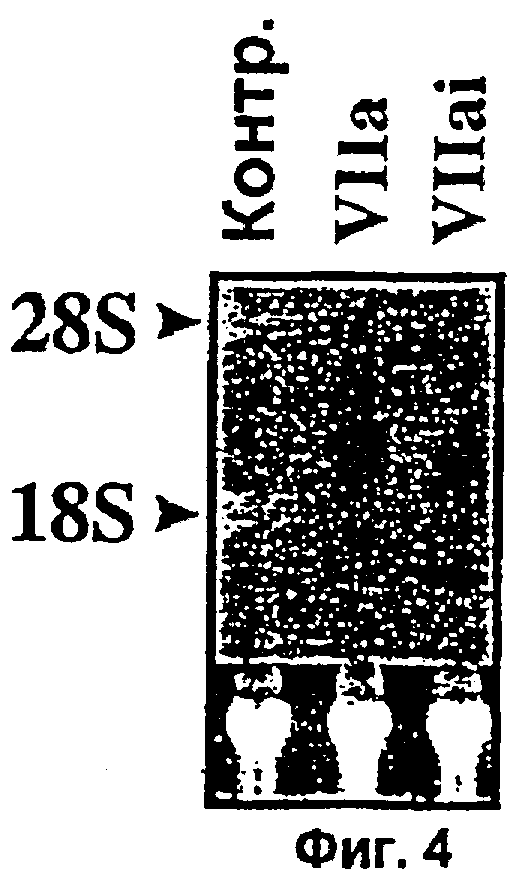
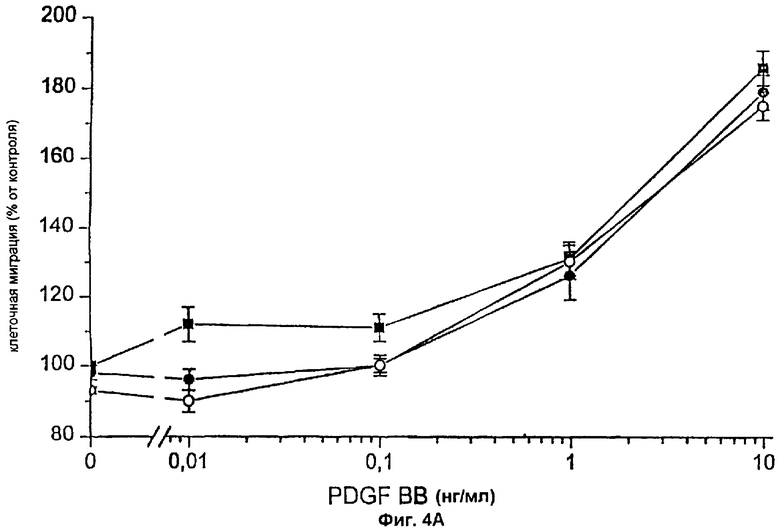

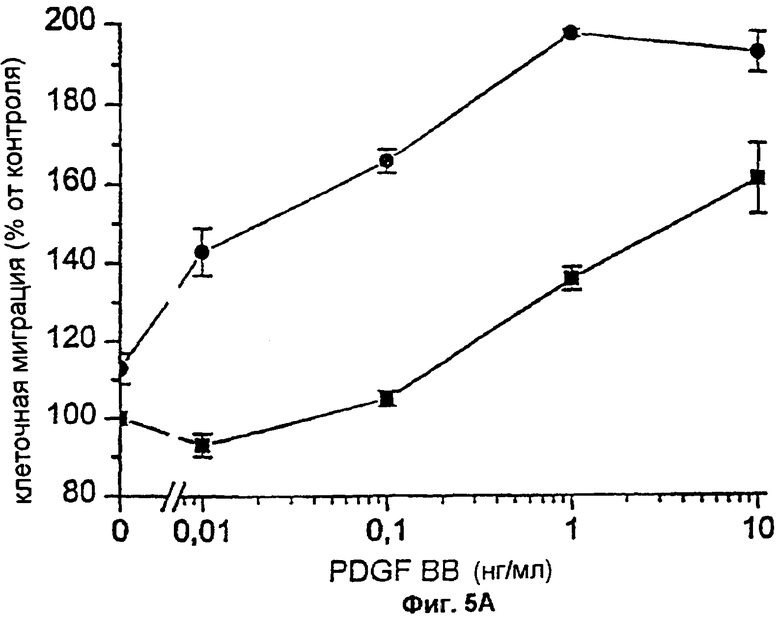
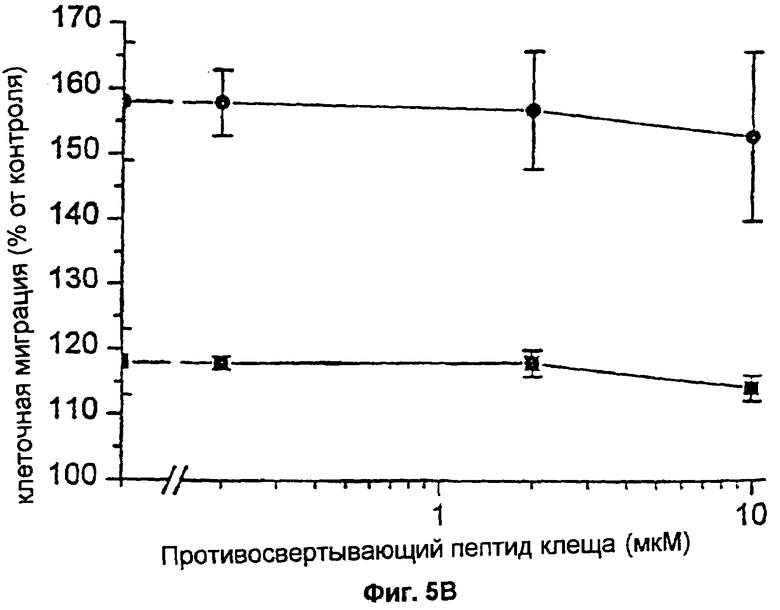
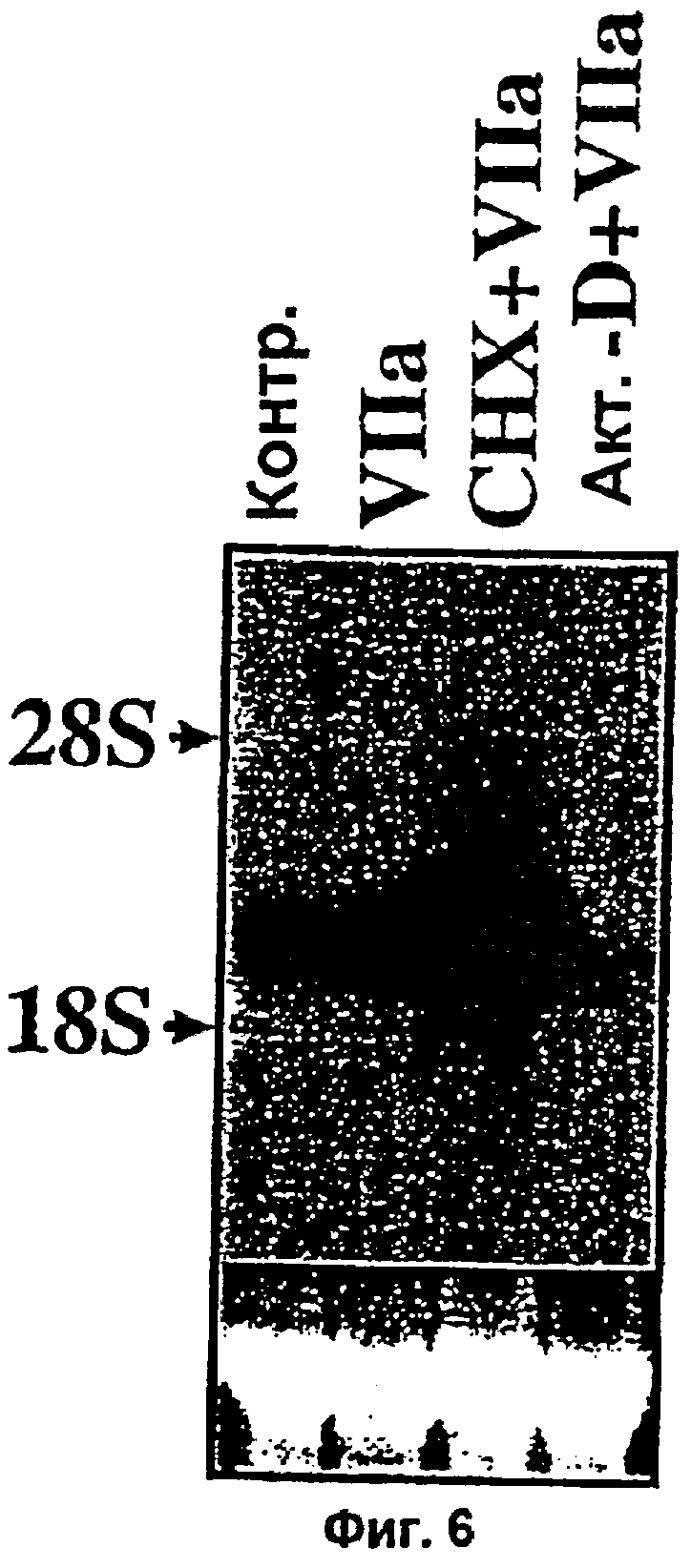
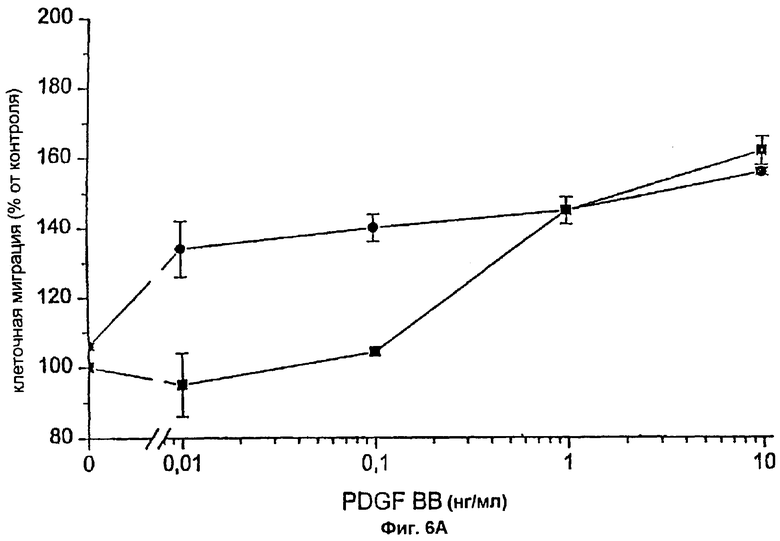
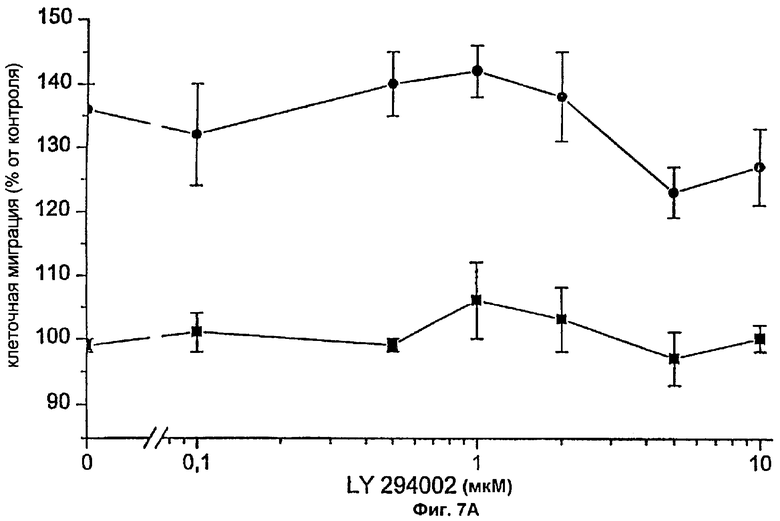

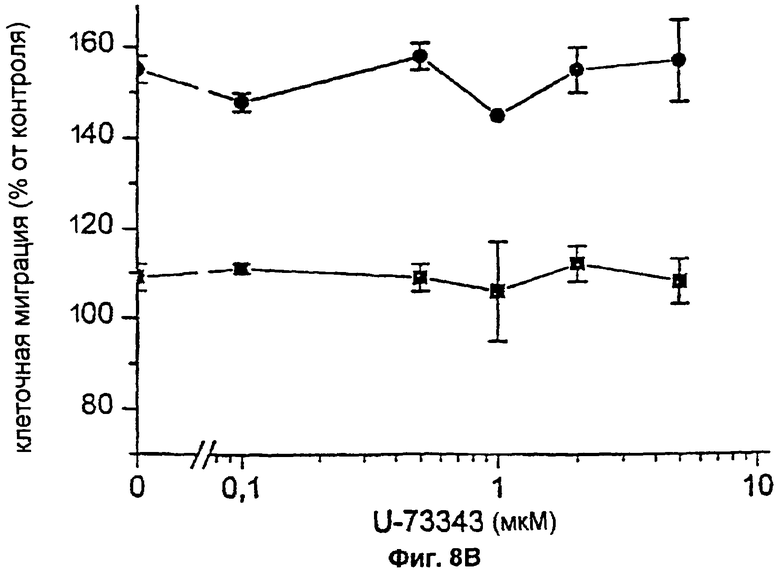
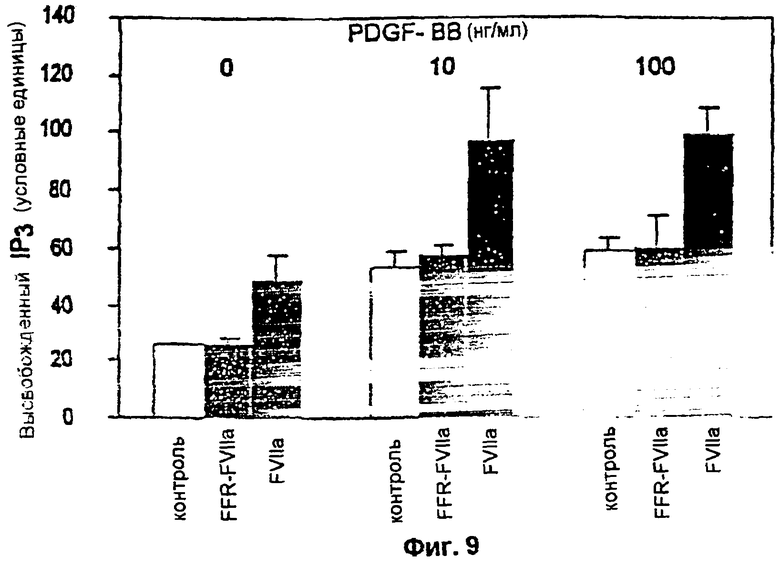

Изобретение относится к медицине, биологии и касается применения агониста тканевого фактора, которым является FVII или FVIIa для индукции или увеличения клеточной миграции, а также к применению антагониста тканевого фактора-модифицированного FVII для снижения или прекращения клеточной миграции при лечении патологических состояний, связанных с специфической регуляцией клеточной миграцией или хемотаксиса. Изобретение обеспечивает повышение эффективности лечения. 7 н. и 7 з.п. ф-лы, 10 ил.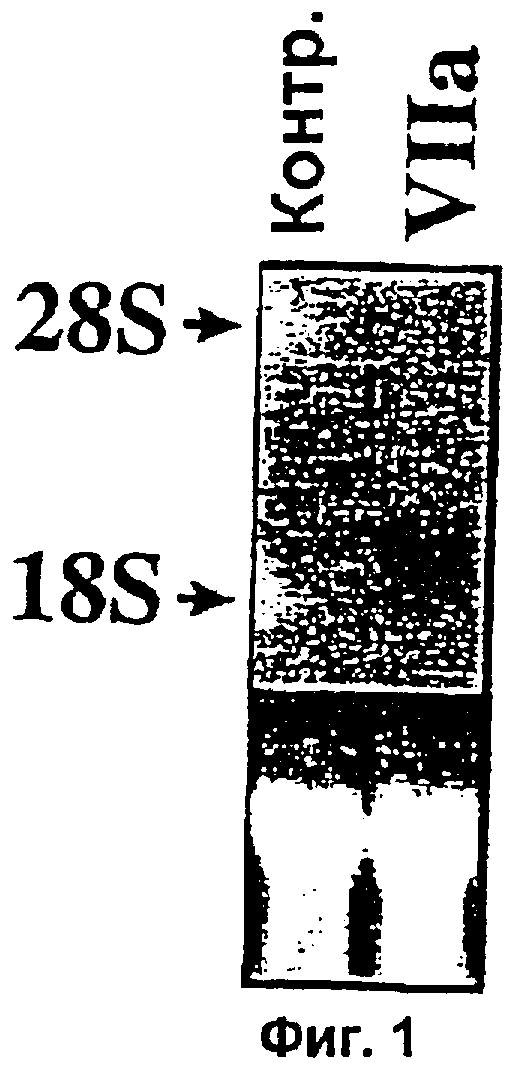
Приоритет по пунктам:
| WO 9733995 А, 18.09.1997 | |||
| WO 9903498 А, 28.01.1999 | |||
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ НА ОСНОВЕ МЕДИАТОРА МЕЖКЛЕТОЧНОЙ АДГЕЗИИ (ВАРИАНТЫ), СПОСОБ ИНГИБИРОВАНИЯ МЕЖКЛЕТОЧНОЙ АДГЕЗИИ, СПОСОБ ЛЕЧЕНИЯ ВОСПАЛИТЕЛЬНОГО ЗАБОЛЕВАНИЯ, СПОСОБ АНАЛИЗА ТЕСТИРУЕМОГО СОЕДИНЕНИЯ НА СПОСОБНОСТЬ ИНГИБИРОВАТЬ КЛЕТОЧНУЮ АДГЕЗИЮ, СПОСОБ ПОЛУЧЕНИЯ СОЕДИНЕНИЯ | 1991 |
|
RU2123338C1 |

