ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
Данная заявка испрашивает приоритет на основе Заявки США № 61/261145, поданной 13 ноября 2009 года, которая включена здесь посредством ссылки в ее полном объеме.
ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Обеспечены способы, композиции и наборы для vWF-содержащих препаратов, в том числе способы, наборы и применение таких препаратов для получения Фактора VIII (FVIII). Предоставляются также полипептиды vWF и молекулы нуклеиновых кислот, кодирующие их.
УРОВЕНЬ ТЕХНИКИ
FVIII, экспрессируемый клетками млекопитающих, является часто специфически или неспецифически абсорбируемым на клеточных поверхностях взаимодействием с компонентами поверхности (например, протеогликанами) или рецептор-опосредованными событиями (например, взаимодействием с LRP-рецептором). Возможно также, что экспрессируемый FVIII ферментативно расщепляется и/или деградируется в средах культивируемых клеток. На протяжении культивирования концентрация экспрессированного FVIII уменьшается в среде, если этот секретируемый материал не удаляется быстро после экспрессии (например, перфузионными способами).
При обычных обстоятельствах, комплекс FVIII-vWF может быть удален из среды общепринятыми хроматографическими способами, включающими в себя абсорбцию на заряженных матриксах, или псевдоаффинной хроматографией. Затем FVIII очищают из комплекса FVIII:vWF селективными стадиями промывания с получением обогащенной молекулами FVIII популяции, минимально загрязненной vWF.
vWF образуется в васкулярных эндотелиальных клетках, которые являются основным источником этого белка плазмы, конститутивным или стимулируемым высвобождением, но он также синтезируется в меньших количествах мегакариоцитами. Считается, что первичный продукт трансляции состоит из 2813 аминокислот. После отщепления сигнального пептида (22 аминокислот) имеет место димеризация. Дальнейший процессинг выполняется в аппарате Гольджи, где эти димеры полимеризуются после отщепления и удаления пропептида (741 аминокислот). Этот пропептид играет важную роль в последующем связывании этих димеров, где он катализирует образование дисульфидных мостиков на амино-концевой стороне. Таким образом, могут образовываться олигомеры разного размера в диапазоне размеров от димера 500000 дальтон до больших мультимеров до 20 миллионов дальтон. Кроме протеолитических процедур, vWF подвергается другим посттрансляционным модификациям, в том числе гликозилированию и сульфатированию.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
В одном аспекте, данное изобретение обеспечивает полипептид, содержащий первую аминокислотную последовательность, присутствующую в полипептиде vWF, и другую аминокислотную последовательность, гетерологичную относительно первой, где этот полипептид способен связывать FVIII.
В другом аспекте, данное изобретение обеспечивает композицию, содержащую полипептид, содержащий эту первую и вторую аминокислотную последовательность.
В некоторых аспектах, данное изобретение обеспечивает белковый комплекс, содержащий этот полипептид и FVIII.
В других аспектах, данное изобретение обеспечивает композицию, содержащую этот белковый комплекс.
В других дополнительных аспектах, данное изобретение обеспечивает нуклеотидную последовательность, кодирующую полипептид, содержащий первую и вторую аминокислотную последовательность.
В одном аспекте, данное изобретение обеспечивает экспрессирующий вектор, содержащий эту нуклеотидную последовательность.
В другом аспекте, данное изобретение обеспечивает клетку, экспрессирующую полипептид, содержащий эту первую и вторую аминокислотную последовательность.
В некоторых аспектах, данное изобретение обеспечивает клетку, экспрессирующую белковый комплекс, содержащий этот полипептид и FVIII.
В других аспектах, данное изобретение обеспечивает способ получения этого белкового комплекса, предусматривающий контактирование этого полипептида с FVIII.
В одном аспекте, данное изобретение обеспечивает способ получения FVIII, предусматривающий: контактирование FVIII с полипептидом, содержащим первую и вторую аминокислотную последовательность, с образованием белкового комплекса, содержащего этот полипептид и FVIII.
В другом аспекте, данное изобретение обеспечивает способ усиления фармакокинетического свойства FVIII в плазме, предусматривающий введение субъекту композиции, содержащей этот белковый комплекс, включающий в себя этот полипептид и FVIII.
В некоторых аспектах, данное изобретение обеспечивает композицию, содержащую этот белковый комплекс и фармацевтически приемлемый носитель.
В других аспектах, данное изобретение обеспечивает способ лечения связанного с кровью состояния, предусматривающий введение композиции, содержащей белковый комплекс, содержащий этот полипептид и FVIII, где этот полипептид содержит первую и вторую аминокислотную последовательность.
В дополнительных аспектах обеспечен набор.
КРАТКОЕ ОПИСАНИЕ ФИГУР
Фигура 1 показывает схематическое представление структуры, процессинга и созревания белка фактора фон Виллебранда. (A) Структура доменов первичного полипептида фактора фон Виллебранда: SS = сигнальный пептид; D1 и D2 = последовательность пропептида; D'-D3 = включает в себя номинальный район связывания фактора FVIII; A1 и A3 = домены связывания коллагена (и других взаимодействий); (B) Во время секреции и процессинга сигнальный пептид удаляется и пропептид позднее отщепляется от полипептида vWF фурин-подобной стадией процессинга с образованием зрелого полипептида vWF, который начинается обычно в месте присоединения D'-домена; (C) Пропептид, ассоциированный со зрелым полипептидом vWF, стимулирует увеличенное связывание и мультимеризацию FVIII; и (D) Остатки цистеина, обеспечиваемые комплексом пропептид-зрелый vWF, обеспечивают ковалентные мостики, которые позволяют интра- и интермолекулярное образование мультимера.
Фигура 2 показывает схематическое представление укорочений доменов фактора фон Виллебранда с ковалентным слиянием с Fc IgG1. (А) Структура доменов первичного полипептида фактора фон Виллебранда: SS = сигнальный пептид; D1 и D2 = последовательность пропептида; D'-D3 = включает в себя номинальный район связывания фактора FVIII; A1 и A3 = домены связывания коллагена (и других взаимодействий); (B) Эти первичные укороченные полипептиды vWF с Fc представлены в виде диаграммы, показывающей ожидаемую структуру доменов отщепления сигнального пептида. Последовательности включают в себя пропептид, за которым следует либо D'-D3-, либо D'-A1-, либо D'-A3-домен, каждый из которых, в свою очередь, слит ковалентно в шарнирной области константного района IgG1; и (C) Пропептид, ассоциированный со слияниями укороченный полипептид vWF-Fc (т.е., D'-D3-Fc, D'-A1-Fc или D'A3-Fc), стимулирует увеличенное связывание FVIII и мультимеризацию, точно так, как это происходит в процессинге зрелого vWF.
Фигура 3 показывает схематическое представление процессинга, созревания и мультимеризации слияния укороченный vWF-домен-Fc первичного продукта трансляции. (A) Представление пропептида и слияния укороченный vWF-домен-Fc после отщепления сигнального пептида. В этом представлении, эти полипептиды получены из единственного первичного продукта трансляции, изображенного, например, на фигуре 1; (B) Представление, показывающее пропептид и мономер слияния vWF-домен-Fc после процессинга первичного продукта трансляции и ассоциации пропептида с полипептидом укороченный vWF-домен-Fc для стимуляции правильной укладки (фолдинга), и (C) Остатки цистеина, содержащиеся в шарнирной области Fc IgG1, обеспечивают внутримолекулярное образование мостика для содействия созданию функционального димера vWF-Fc, который также связывает FVIII, способом, сходным со способом, обнаруживаемым в полученным из плазмы vWF; пропептиды vWF, в свою очередь, стимулируют мультимеризацию этих слитых полипептидов.
Фигура 4 показывает схематическое представление процессинга, созревания и мультимеризации vWF-Fc двух первичных продуктов трансляции из независимых кодирующих районов. (А) Представление отдельных и независимых первичных продуктов трансляции пропептида и слитых полипептидов укороченный vWF-домен-Fc после отщепления сигнального пептида; не требуется фурин-подобный процессинг, как в нормальном процессинге vWF. В этом представлении, полипептиды получают из двух независимых промоторных кассет, транскрибируемых либо из одного экспрессирующего вектора, либо из двух экспрессирующих векторов, коэкспрессируемых в одной и той же клетке, первый способ экспрессии изображен, например, на фигуре 6; (B) Представление, показывающее слитый мономер vWF-домен-Fc после процессинга первичного продукта трансляции и ассоциации коэкспрессируемого пропептида с полипептидом укороченный vWF-домен-Fc для стимуляции правильной укладки; и (C) Остатки цистеина, содержащиеся в шарнирной области Fc IgG1, обеспечивают внутримолекулярное образование мостика для содействия созданию функционального димера vWF-Fc, который также связывает FVIII, способом, сходным со способом, обнаруживаемым в полученном из плазмы vWF; пропептиды vWF, в свою очередь, стимулируют мультимеризацию этих слитых полипептидов.
Фигура 5 является схематической иллюстрацией, в некоторых вариантах осуществления, трансфекции плазмидных экспрессирующих векторов в клетки млекопитающих, которые экспрессируют полипептиды зрелого vWF или слитые полипептиды укороченный vWF-домен-Fc. (A) Экспрессирующая плазмида с кодирующей последовательностью для vWF или слитых белков укороченный vWF-домен-Fc, каждый из которых содержит домены последовательностей сигнального пептида и пропептида в виде части первичного продукта трансляции. (B) Представление плазмид, трансфицированных и поглощенных в клетки млекопитающих при отборе для создания стабильно экспрессирующей клеточной линии; (C) Экспрессирующая плазмида с кодирующей кассетой для Фактора VIII, использующая другой селектируемый маркер (неомицин), чем в (А); и (D) Представление плазмид FVIII (C) и vWF или vWF-Fc (D), ко-трансфицированных и поглощенных в клетки млекопитающих при отборе для создания стабильно экспрессирующей клеточной линии, которая экспрессирует FVIII и vWF-Fc (или vWF).
Фигура 6 является схематической иллюстрацией, в других вариантах осуществления, трансфекции плазмидных экспрессирующих векторов в клетки млекопитающих, которые экспрессируют полипептиды зрелого vWF или слитых полипептидов укороченный vWF-домен-Fc от независимых промоторов либо на одних и тех же, либо на различных плазмидных векторах. (A) Экспрессирующая плазмида с кодирующими последовательностями для vWF или слитых белков укороченный vWF-домен-Fc, которые не экспрессируют домен пропептида в виде части их первичного продукта трансляции, и экспрессирующая плазмида для домена пропептида vWF (каждая с соответствующими последовательностями сигнального пептида) от независимых промоторов; (B) Представление плазмид полипептидов пропептид-минус vWF или слитых полипептидов vWF-Fc и плазмид полипептида пропептида, трансфицированных и поглощенных в клетки млекопитающих при отборе для создания стабильной клеточной линии, которая коэкспрессирует как vWF, так и белки vWF-Fc, а также полипептид пропептида; (С) Экспрессирующая плазмида с кодирующей кассетой для Фактора VIII, использующая другой селектируемый маркер (неомицин), чем в (А); и (D) Представление плазмид FVIII (C) и vWF или vWF-Fc (D), ко-трансфицированных и поглощенных в клетки млекопитающих при отборе для создания стабильно экспрессирующей клеточной линии, которая экспрессирует FVIII, и либо vWF-Fc, либо vWF, и, независимо, пропептид vWF.
Фигура 7 показывает окрашенные Кумасси белки экспрессированных белков, подвергнутых электрофорезу на 4-12% Бис-Трис-ПААГ гелях при восстанавливающих и денатурирующих условиях, либо непосредственно из культуральных супернатантов (А), либо из Белок G-иммунопреципитаций (В) этих белков, присутствующих в культуральных супернатантах из (А). Клетки PER.C6, трансфицированные плазмидами, содержащими конструкт укороченный vWF-Fc, отбирали на стабильные культуры в виде пула с использованием отбора с применением антибиотиков. Для непосредственных проб супернатантов, 20 микролитров наносили на гель. Для иммунопреципитации, 20 микролитров гранул Белка G добавляли либо к 1 мл среды с D'-D3-Fc, либо к 0,2 мл среды с D'-Al-Fc или D'-A3-Fc. Дорожки на обоих гелях представляют: Дорожка 1: Стандарт молекулярных масс; Дорожка 2: D'-D3-Fc; Дорожка 3: pro-D'-D3-Fc; Дорожка 4: D'-A1-Fc; Дорожка 5: pro-D'-Al-Fc; Дорожка 6: D'-A3-Fc; Дорожка 7: pro-D'-A3-Fc; и Дорожка 8: кондиционированная среда нетрансфицированных PER.C6 в качестве контроля. Полосы более высоких молекулярных масс в дорожках 3, 5 и 8 представляют непроцессированный пропептид, все еще присоединенный к соответствующим доменам vWF.
Фигура 8 является диаграммой в виде столбцов, показывающей извлеченную активность FVIII с использованием слитых белков vWF-Fc или полученный из плазмы белок vWF. Заштрихованный столбец показывает увеличенное извлечение выше контроля (т.e., BDD-FVIII без добавленных белков vWF). Супернатанты клеток, содержащие экспрессируемые, слитые белки укороченный vWF-Fc или полноразмерный vWF смешивали с клетками BDD-078, экспрессирующими рекомбинантный FVIII. После двух дней, пробы анализировали на экспрессию FVIII с использованием хромогенного анализа FVIII.
На Фигуре 9 показан гель, показывающий мультимеризацию слитых белков pro-vWF-Fc в сравнении с нормальными полученными из плазмы мультимерами vWF. Полученный из плазмы фактор VIII (Koate-DVI®) подвергался электрофорезу в качестве стандарта для мультимеризации на денатурирующих, а не на восстанавливающих, 1,6% (дорожка 1) и 2% HGT(P) (дорожка 2) агарозных гелях, тогда как пробы белков pro-D'-D3-Fc (дорожка 3), pro-D'-Al-Fc (дорожка 4) и pro-D'-A3-Fc (дорожка 5) подвергались электрофорезу на 1,6% гелях для визуализации различий в размерах “лэддера”. Скобки идентифицируют положение триплетов vWF-димеров в дорожке 1 и соответствующее положение в дорожке 2. Как и ожидалось, увеличивающийся размер полипептидной цепи pro-vWF-Fc приводит к созданию мультимерных полос увеличивающейся молекулярной массы, в порядке: pro-D'-D3-Fc < pro-D'-A1-Fc < pro-D'-A3-Fc.
Фигура 10 показывает очистку FVIII из супернатантов, содержащих комплексы pro-D'A3-Fc/FVIII: (A) Хроматографическое отслеживание пиков, элюированных во время различных буферных условий; и (B) Кумасси-окрашенный 7,5% гель электрофореза в ПААГ с пробами, взятыми из колоночного отслеживания, представленного в (А), показывает специфическую элюцию FVIII из комплексов pro-D'A3-Fc/FVIII в дорожке 4. Дорожки 1-9 представляют, соответственно, (1) исходный материал, (2) проходящий поток, (3) 0,1 M CaCl2-элюат, (4) 0,3M CaCl2-элюат, (5) pH 5,5 цитрат-элюат, (6) концентрированный препарат BDD-FVIII, (7) маркеры молекулярных масс (размеры на правой стороне), (8) клеточный супернатант из экспрессирующих pro-D'-A3-Fc клеток PER.C6, (9) коммерческий BDD-FVIII (Xyntha®), причем звездочки показывают три полосы приблизительно 170, 90 и 80 кД, соответствующие полноразмерному BDD-FVIII, тяжелой цепи и легкой цепи, соответственно.
Фигура 11 показывает очистку FVIII из супернатантов, содержащих коммерческий рекомбинантный FVIII с делетированным В-доменом (Xyntha), с супернатантом, содержащим белок pro-D'D3-Fc. После элюции из гелей различными буферными композициями эти фракции анализировали электрофорезом на 7%-ных восстанавливающих/денатурирующих полиакриламидных гелях (NuPAGE) и окрашивали Кумасси бриллиантовым синим. Дорожка 7 показывает по существу чистый FVIII, извлеченный после элюции из комплексов pro-D'-D3-Fc/FVIII, связанный с Белком А. Дорожки 1-9 представляют, соответственно, (1) маркеры молекулярных масс полипептидов с молекулярными массами, перечисленными слева, (2) клеточный супернатант pro-D'-D3-Fc, (3) коммерческий FVIII с делетированным B-доменом (Xyntha®), (4) Xyntha®, смешанный с супернатантом pro-D'-D3, (5) супернатант (pro-D'D3-Fc + Xyntha®), загруженный на колонку белок А, (6) проходящий через колонку промывочный поток с 20 мМ Tris-HCl, pH 7,0, (7) 0,3 M CaCl2-элюат и (8) промывку глицином с pH 3,9, которая разделяет эту колонку на дополнительные Белок А-связывающие белки. Три звездочки в дорожке 3 соответствуют полосам белков приблизительно 170, 90 и 80 кД, соответствующим полноразмерному BDD-FVIII, тяжелой цепи и легкой цепи, соответственно.
ПОДРОБНОЕ ОПИСАНИЕ
В одном аспекте данное изобретение обеспечивает полипептид, содержащий первую аминокислотную последовательность, присутствующую в полипептиде vWF, и вторую аминокислотную последовательность, гетерологичную относительно первой, где этот полипептид способен к связыванию FVIII.
В данном контексте термин "способный к связыванию" подразумевает варианты, в которых эта способность полипептида связываться с FVIII осуществляется сборкой белка высокого порядка и/или одной или несколькими посттрансляционными модификациями, такими как, например, отщепление сигнального пептида, отщеплением пропептида, ассоциацией пропептида, фосфорилированием, гликозилированием и т.п. Например, В некоторых вариантах осуществления, этот полипептид является "способным к связыванию" с FVIII в виде димерного, тримерного, четырехмерного или более высокого порядка мультимерного комплекса, который образуется после мультимеризации этого полипептида. Или, например, в других вариантах осуществления, этот полипептид "способен к связыванию" FVIII после мультимеризации этого полипептида после ассоциации пропептида с этим полипептидом. "Мультимеризация" и "олигомеризация" используются здесь взаимозаменяемо и относятся к ассоциации двух или более молекул белков, опосредуемой ковалентными (например, межмолекулярными дисульфидными связями) и/или нековалентными взаимодействиями. Таким образом, термины "мультимер (мультимеры)" и "олигомер (олигомеры)" также используются здесь взаимозаменяемо.
Данное изобретение рассматривает полипептиды vWF и FVIII человека и не человека (происходящие, например, из приматов, собак, кошек, лошадей, свиней, мышей, крыс, морских свинок, кроликов, коров, других позвоночных), включающие в себя природные, синтетические и рекомбинантные белки. В объеме данного изобретения находятся также полипептиды vWF и FVIII, соответствующие белкам дикого типа, или их мутанты, варианты и/или их укороченные (процессированные) формы. Например, в некоторых вариантах осуществления, первая аминокислотная последовательность соответствует фрагменту полипептида vWF из человека, причем гетерологичная вторая аминокислотная последовательность содержит последовательность или состоит из последовательности, не присутствующей в каком-либо белке vWF, человека, или наоборот. FVIII и/или vWF включают в себя природные белки, а также их производные, например, белки, мутированные делецией, заменой или инсерцией, или химическое производное или его фрагмент.
I. Первая аминокислотная последовательность
В одном варианте осуществления, первая аминокислотная последовательность определяет структуру или домен, которые взаимодействуют с моноклональным анти-vWF-антителом, способным специфически связываться с районом полипептида VWF, содержащим домен связывания FVIII. В одном варианте осуществления, этим моноклональным антителом является моноклональное антитело С3, описанное, например, в статьях Foster et al, JBC, 262:8443 (1987) и Fulcher et al, J. Clin. Invest, 76:117 (1985), каждая из которых включена здесь посредством ссылки на их описания моноклонального антитела С3 и способа получения моноклональных антител, в частности, моноклонального антитела С3.
Не ограничивающие примеры аминокислотных последовательностей vWF и последовательностей нуклеиновых кислот, кодирующих vWF или его часть, описаны например, под номерами доступа GenBank: NP_000543, NM_000552, AAE20723, AAB59512, P04275, EAW88815, ACP57027, EAW88816 и AAD04919; U.S Patent No. 5,260,274; Titani et al, Biochemistry, 25:3171-84 (1986); и Sadler et al, PNAS, 82:6391-6398 (1985), каждые из которых включены здесь посредством ссылки на их описания аминокислотных последовательностей и последовательностей нуклеиновых кислот, соответствующих vWF.
Специалистам с обычной квалификацией в данной области известно, что прототипическим preprop-vWF является полипептид из 2813 аминокислот с сигнальным пептидом из 22 аминокислот и повторяющимися функциональными доменами, A, B, C, D и CK, которые распределены от амино-конца в порядке "D1", "D2", "D'", "D3", "A1", "A2", "A3", "D4", "B1", "B2", "B3” (последние три вместе считаются "B"), "C1", "C2" и "CK". Субъединица "зрелого" vWF состоит, от N-конца к C-концу в этом порядке, из доменов: D'-D3-A1-A2-A3-D4-B1-B2-B3-C1-C2-CK.
Аминокислотная последовательность примерного полноразмерного vWF человека показана посредством SEQ ID NO:29, которая кодируется нуклеотидами 251-8689 SEQ ID NO:30. Со ссылкой на SEQ ID NO:29, часть "сигнального пептида" простирается от положения 1 аминокислот по Cys-22, включительно, часть "пропептида" (D1-D2) простирается от положения 23 аминокислот по Arg-763, включительно, и "зрелый" белок vWF простирается от положения 764 по 2813, включительно. Индивидуальные домены были также приближенно картированы, как D': 764-865; D3: 866-1242; A1: 1260-1479; A2: 1480-1672; A3: 1673-1874; D4: 1947-2298; B: 2296-2399; C1: 2400-2516; C2: 2544-2663 и CK: 2720-2813. Одна альтернативная система картирования и обозначения доменов vWF была использована конвенцией Базы данных белков EXPASY (worldwideweb.uniprot.org/uniprot/P04275) в виде D1: 34-240; D2: 387-598; D': 776-827; D3: 866-1074; A1: 1277-1453; A2: 1498-1665; A3: 1691-1871; D4: 1949-2153; B: 2255-2328 (который называемый C1 в EXPASY); C1: 2429-2495 (называемый C2 в EXPASY); C2: 2580-2645 (называемый C3 в EXPASY); и CK: 2724-2812.
Не ограничивающие примеры аминокислотной последовательности и последовательности нуклеиновых кислот FVIII описаны, например, номерами доступа GenBank 1012296A AAA52420.1, CAA25619.1, AAA52484.1, 1012298A, EAW72647.1, EAW72646.1, XP_001498954.1, ACK44290.1, AC095359.1, NP_001138980.1, ABZ10503.1, NP_032003.2, Патент США с номером 6,307,032, и Wood et al, Nature, 312:330-7 (1984), каждые из которых включены здесь посредством ссылки на их описания последовательностей FVIII. Варианты, производные, модификации и комплексы FVIII также известны в данной области и охватываются данным изобретением. Например, варианты Фактора FVIII, описанные в Патенте США № 5668108, описывают варианты FVIII, посредством которых аспарагиновая кислота в положении 1241 заменена глутаминовой кислотой с сопутствующими изменениями также нуклеиновых кислот; Патент США № 5149637 описывает варианты FVIII, содержащие С-концевую часть, либо гликозилированную, либо негликозилированную; и Патент США 5661008 описывает вариант FVIII, содержащий аминокислоты 1-740, связанные с аминокислотами 1649-2332 по меньшей мере 3 аминокислотными остатками, которые включены здесь посредством ссылки на описания каждой из этих вариантных последовательностей FVIII.
В одном варианте осуществления, FVIII является происходящим из плазмы или сыворотки FVIII. В другом варианте осуществления, этот FVIII является рекомбинантным FVIII, например, активным FVIII человека, экспрессируемым в культивируемых клетках млекопитающих из рекомбинантных ДНК-клонов. Экспрессионные системы для получения Фактора FVIII известны в данной области и включают в себя прокариотические и эукариотические клетки, как описано Патентами США с номерами 5633150, 5804420 и 5422250, каждый из которых включен здесь посредством ссылки на описание получения FVIII.
Специалисту с обычной квалификацией в данной области известно, что способность полипептида связывать FVIII может быть определена различными способами. В частности, полипептид данного изобретения может анализироваться на его способность связывать FVIII с использованием способов, описанных здесь, и/или адаптацией способов, известных в данной области. Например, для анализа/определения связывания могут быть использованы иммуноанализы, включающие в себя, но не ограничивающиеся ими, конкурентные и неконкурентные системы анализа, использующие такие способы, как вестерн-блоты, радиоиммуноанализы, ELISA (твердофазный иммуноферментный анализ), "сэндвич"-иммуноанализы, анализы иммунопреципитации, преципитиновые реакции, гель-диффузионные преципитиновые реакции, иммунодиффузионные анализы, анализы агглютинации, анализы фиксации комплемента, иммунорадиометрические анализы, флуоресцентные иммуноанализы и т.д. (см., например, руководство Ausubel et al, eds, 1994, Current Protocols in Molecular Biology, Vol. 1, John Wiley & Sons, Inc., New York, которое включено здесь посредством ссылки в его полном объеме).
Например, полипептид, содержащий первую и вторую аминокислотные последовательности, может быть контактирован с FVIII в подходящем буфере, таком как TBS, в присутствии моноклонального антитела, связанного с Сефарозой. Это антитело может быть направлено против района этого полипептида, так что связывание этого антитела с полипептидом не мешает связыванию этого полипептида с FVIII (например, это антитело может быть направлено против второй аминокислотной последовательности или против повторяющегося района "A1" или "A2" или "A3" vWF, где такой район также присутствует на этом полипептиде. После контакта, FVIII, связанный с этим полипептидом/антителом, и несвязанный FVIII могут быть разделены, например, центрифугированием, и FVIII может быть измерен, например, с использованием анализа с хромогенным субстратом (Factor VIII Coatest; Chromogenix, Molndal, Sweden).
В предпочтительных вариантах осуществления, первой аминокислотной последовательностью является укороченный полипептид vWF. Например, укороченные формы vWF, в некоторых вариантах осуществления, включают в себя (i) укороченные полипептиды vWF, которые лишены последовательности "пропептида"; и (ii) укороченные полипептиды vWF, которые лишены "A1", "A2", "A3", "D4", "B" (также известные как "B1", "B2" и "B3"), "C1", "C2" и/или "CK" домена зрелой последовательности. Встречаются также и другие укороченные или иным образом модифицированные формы vWF.
В одном варианте осуществления, первая аминокислотная последовательность является такой как последовательность, представленная в SEQ ID NO:1, SEQ ID NO:2, SEQ ID NO:3, SEQ ID NO:4, SEQ ID NO:5, SEQ ID NO:6, SEQ ID NO:7, SEQ ID NO:8, SEQ ID NO:9, SEQ ID NO:10, SEQ ID NO:11, SEQ ID NO:12, SEQ ID NO:13, SEQ ID NO:14, SEQ ID NO:15, SEQ ID NO:33, SEQ ID NO:34 или SEQ ID NO:35.
II. Вторая аминокислотная последовательность
В других вариантах осуществления, вторая аминокислотная последовательность этого полипептида обеспечивает структуру или домен, имеющие аффинность в отношении партнера связывания.
Эта вторая аминокислотная последовательность является гетерологичной относительно первой. В одном варианте осуществления, эта гетерологичная вторая аминокислотная последовательность содержит последовательность или состоит из последовательности, которая не присутствует ни в каком белке vWF. В одном варианте осуществления, по меньшей мере часть (например, смежная часть) гетерологичной второй аминокислотной последовательности соответствует последовательности, не присутствующей ни в каком полипептиде vWF.
Предпочтительно, в некоторых вариантах осуществления, эта вторая аминокислотная последовательность соответствует Fc-полипептиду антитела, такому как, например, Fc-район IgG1 человека. Например, вторая аминокислотная последовательность может соответствовать аминокислотным остаткам, которые простираются от N-концевой шарнирной области до природного С-конца, т.е. является по существу Fc-районом полноразмерного антитела. Могут быть также использованы фрагменты Fc-районов, например, Fc-районов, которые были усечены на С-концевой стороне. В некоторых вариантах осуществления, эти фрагменты предпочтительно содержат один или несколько остатков цистеина (по меньшей мере остатки цистеина в шарнирной области) для получения возможности образования межцепочечных дисульфидных связей между частями Fc-полипептидов двух отдельных полипептидов данного изобретения, с образованием димеров.
Другие Fc-районы антитела могут заменять Fc-район IgG1 человека. Например, другими подходящими Fc-районами являются Fc-районы, которые могут связываться с аффинностью с белком А или белком G или другими подобными Fc-связывающими матриксами и включают в себя Fc-районы мышиных IgG, IgA, IgE, IgD, IgM или фрагменты Fc-района IgG, IgA, IgE, IgD, IgM человека, например, фрагменты, содержащие по меньшей мере шарнирную область, так что будут образовываться межцепочечные дисульфидные связи.
Fc-район IgG1 описан, например, номером доступа Х70421 GenBank, который включен здесь посредством ссылки в его полном объеме.
В одном варианте осуществления эта вторая аминокислотная последовательность содержит последовательность, представленную в SEQ ID NO:16.
В некоторых вариантах осуществления вторая аминокислотная последовательность предпочтительно является С-концом для первой аминокислотной последовательности. Получение слитых полипептидов, содержащих гетерологичную аминокислотную последовательность, слитую с различными частями другой аминокислотной последовательности, описано, например, в статьях Ashkenazi et al, PNAS, 88:10535 (1991) и Byrn et al, Nature 344:677 (1990), каждая из которых включена здесь посредством ссылки в ее полном объеме. Например, слияние генов, кодирующее полипептид, содержащий первую и вторую аминокислотные последовательности, может быть инсертировано в подходящий экспрессирующий вектор. Экспрессированные слитые белки могут быть собраны, посредством чего межцепочечные дисульфидные связи могут образовываться между этими полипептидами с образованием димеров. В других вариантах осуществления, эти слитые полимеры данного изобретения могут быть экспрессированы со спейсерными связывающими аминокислоты группами или без спейсерных связывающих аминокислоты групп. Например, в некоторых вариантах осуществления, полипептиды данного изобретения могут дополнительно содержать линкер между первой и второй аминокислотными последовательностями, причем этот линкер содержит один или несколько аминокислотных остатков, разделяющих первую и вторую последовательности.
В другом варианте осуществления полипептид данного изобретения содержит аминокислотную последовательность, представленную в SEQ ID NO:17, SEQ ID NO:18, SEQ ID NO:19, SEQ ID NO:20, SEQ ID NO:21, SEQ ID NO:22, SEQ ID NO:36, SEQ ID NO:38 или SEQ ID NO:39.
В одном варианте осуществления этот полипептид кодируется молекулой нуклеиновой кислоты, имеющей нуклеотидную последовательность, представленную в SEQ ID NO:23, SEQ ID NO:24, SEQ ID NO:25, SEQ ID NO:26, SEQ ID NO:27, SEQ ID NO:28, SEQ ID NO:37, SEQ ID NO:42 или SEQ ID NO:43.
Варианты описанных здесь последовательностей также находятся в объеме данного изобретения. Вариантом полипептида может называться аминокислотная последовательность, которая изменена одной или несколькими аминокислотами. Этот вариант может иметь "консервативные" изменения, в которых замененная аминокислота имеет сходные структурные или химические свойства, например, замену лейцина изолейцином. Альтернативно, вариант может иметь "не-консервативные" изменения, например, замену глицина триптофаном. Аналогичная минорная вариация может также включать в себя делецию или инсерцию аминокислоты или и то, и другое. Конкретной формой "вариантного" полипептида является "функционально эквивалентный" полипептид, т.е. полипептид, который проявляет по существу сходную активность in vivo или in vitro и/или сходное связывание, что и полипептид этого изобретения. Руководство в определении, какие аминокислотные остатки могут быть заменены, инсертированы или делетированы без элиминации биологической или иммунологической активности, может быть найдено с использованием компьютерных программ, хорошо известных в данной области, например, в программе DNASTAR (DNASTAR, Inc., Madison, WI). Дополнительно, конкретное руководство обеспечено ниже, в том числе руководство, обеспеченное в цитируемых ссылках, которые включены здесь посредством ссылки.
В других вариантах осуществления конкретные положения названных остатков могут несколько варьироваться, все еще при присутствии их в полипептиде в структурно и функционально аналогичных положениях (см. Chang, Y., et al, Biochemistry 57:3258-3271 (1998).
Кроме того, конкретные варианты этого изобретения могут характеризоваться функционально в отношении полипептида vWF, или его фрагмента, по FVIII-связывающей способности. В некоторых вариантах осуществления, полипептид этого изобретения проявляет способность связывания в отношении FVIII, которая является меньшей, приблизительно равной или большей, чем способность связывания ссылочного белка vWF (например, эндогенного vWF дикого типа) или его фрагмента, который способен связываться с белком FVIII (например, эндогенным FVIII дикого типа).
Таким образом, это изобретение включает в себя такие вариации полипептидов, описанных здесь. Такие варианты включают в себя делеции, инсерции, инверсии, повторы и замены. Дополнительное руководство в отношении того, какие аминокислотные изменения являются, вероятно, молчащими, может быть найдено в Bowie, J. U., et al, "Deciphering the Message in Protein Sequences: Tolerance to Amino Acid Substitutions", Science 247:1306-1310 (1990).
Таким образом, фрагменты, производные или аналоги полипептидов данного изобретения включают в себя фрагменты, производные или аналоги, имеющие последовательности, которые имеют, в сравнении с полипептидами данного изобретения, (i) один или несколько из аминокислотных остатков (например, 1, 3, 5, 8, 10, 15 или 20 остатков), замененных консервативным или неконсервативным аминокислотным остатком (предпочтительно консервативным аминокислотным остатком). Такие замененные аминокислотные остатки могут быть кодированы или могут не быть кодированы генетическим кодом; или (ii) один или несколько из аминокислотных остатков (например, 1, 3, 5, 8, 10, 15 или 20 остатков) включают в себя группу-заместитель. В других вариантах осуществления фрагменты, производные или аналоги полипептида данного изобретения включают в себя полипептид данного изобретения, который связан с другим соединением, таким как соединение для увеличения времени полужизни этого полипептида (например, полиэтиленгликоль), или таким соединением, в котором дополнительные аминокислоты слиты с этим полипептидом. Предполагается, что такие фрагменты, производные и аналоги находятся в рамках квалификации специалистов в данной области на основе описанного здесь.
Как указано, изменения предпочтительно являются незначительными, например, включают в себя консервативные аминокислотные замены, которые не влияют значимо на укладку или способность связывания FVIII. Конечно, количество аминокислотных замен, которое может производить специалист в данной области, зависит от многих факторов, в том числе описанных выше факторов. В некоторых вариантах осуществления количество замен для любого конкретного полипептида будет не более чем 50, 40, 30, 25, 20, 15, 10, 5, 3, 2 или 1.
Аминокислотные остатки (полипептидов данного изобретения), которые являются существенными для связывания с FVIII, могут быть идентифицированы способами, известными в данной области, такими как сайт-направленный мутагенез или аланин-сканирующий мутагенез (Cunningham and Wells, Science 244:1081-1085 (1989)). Последняя процедура вводит одиночные аланиновые мутации при каждом остатке в молекуле. Затем полученные мутантные молекулы тестируют на связывание с FVIII, например, как описано здесь. Сайты, которые являются критическими для связывания с FVIII, могут быть также определены структурным анализом, таким как кристаллизация, ядерный магнитный резонанс или фотоаффинное мечение (Smith, et al., J. Mol. Biol. 224:399-904 (1992) и de Vos, et al. Science 255:306-312 (1992)).
В одном варианте осуществления этот рекомбинантный полипептид имеет аминокислотные последовательности, которые являются по меньшей мере на 70%, 80%, 90%, 95%, 98% или более идентичными любой из аминокислотных последовательностей, представленных здесь.
В другом варианте осуществления первая аминокислотная последовательность присутствует в полипептиде vWF, содержащем аминокислотную последовательность, представленную в SEQ ID NO:29, или ее вариант или фрагмент.
В других вариантах осуществления первая аминокислотная последовательность присутствует в полипептиде vWF, кодируемом последовательностью нуклеиновой кислоты, представленной в SEQ ID NO:30 или ее варианте или фрагменте.
В других аспектах данное изобретение обеспечивает рекомбинантный слитый белок vWF-Fc, в котором vWF-часть этого слитого белка является укороченным vWF, который лишен по меньшей мере одного домена зрелого полноразмерного полипептида vWF, причем этот слитый белок способен образовывать мультимеры, которые способны связывать белок FVIII. В одном варианте осуществления, этот укороченный vWF имеет домены D' и D3, при условии, что этот укороченный vWF лишен домена A1, A2, A3, D4, B1, B2, B3, C1, C2, CK или их комбинации. В другом варианте осуществления, этот укороченный vWF имеет домены D', D3 и A1, при условии, что этот укороченный vWF лишен доменов A2, A3, D4, B1, B2, B3, C1, C2 и CK. В некоторых вариантах осуществления, этот укороченный vWF имеет домены D', D3 и A1 и A2, при условии, что этот укороченный vWF лишен доменов A3, D4, B1, B2, B3, C1, C2 и CK. В других вариантах осуществления, этот укороченный vWF имеет домены D', D3, A1, A2, и A3, при условии, что этот укороченный vWF лишен доменов D4, B1, B2, B3, C1, C2 и CK. В других дополнительных вариантах осуществления, этот укороченный vWF лишен доменов D4, B1, B2, B3, C1, C2 и CK.
III. Нуклеиновые кислоты, векторы и системы экспрессии
В других аспектах данное изобретение обеспечивает рекомбинантные экспрессирующие векторы для экспрессии полипептида, содержащего первую и вторую аминокислотные последовательности, и клетки-хозяева, трансформированные этими экспрессирующими векторами. Может быть использована любая подходящая система экспрессии. Эти векторы содержат первую и вторую ДНК-последовательность, кодирующие первую и вторую аминокислотные последовательности, соответственно, функционально связанные с подходящими транскрипционными или трансляционными регуляторными нуклеотидными последовательностями, такими как последовательности, полученные из гена млекопитающего, микробного, вирусного гена или гена насекомого. Примеры регуляторных последовательностей включают в себя транскрипционные промоторы, операторы или энхансеры, сайт связывания рибосом мРНК и подходящие последовательности, которые контролируют инициацию и терминацию транскрипции и трансляции. Нуклеотидные последовательности функционально связаны, когда эта регуляторная последовательность функционально связывается с кодирующей ДНК-последовательностью. Таким образом, промоторная нуклеотидная последовательность функционально связана с кодирующей ДНК-последовательностью, если эта промоторная нуклеотидная последовательность регулирует транскрипцию этой кодирующей ДНК-последовательности. Способность реплицироваться в желаемых клетках-хозяевах, обычно придаваемая сайтом инициации репликации, и ген отбора, при помощи которого идентифицируют трансформанты, могут дополнительно быть включены в этот экспрессирующий вектор.
В других дополнительных вариантах осуществления ДНК-последовательности, кодирующие подходящие сигнальные пептиды, которые могут быть или могут не быть природными относительно первой аминокислотной последовательности, могут быть включены в экспрессирующие векторы. Например, ДНК-последовательность для сигнального пептида (секреторного лидера) может быть обеспечена в рамке считывания с первой последовательностью, так что экспрессируемый полипептид первоначально транслируется в виде слитого белка, содержащего сигнальный пептид. Сигнальный пептид, который является функциональным в предполагаемых клетках-хозяевах, усиливает внеклеточную секрецию полипептида, содержащего первую и вторую аминокислотные последовательности. В некоторых вариантах осуществления, сигнальный пептид отщепляется от этого полипептида после секреции полипептида из клетки. В других вариантах осуществления, подходящие сигнальные пептиды, которые не являются природными для первой аминокислотной последовательности, могут быть обеспечены в качестве альтернативы природной сигнальной последовательности или в дополнение к природной сигнальной последовательности.
В некоторых вариантах осуществления сигнальный пептид имеет аминокислотную последовательность, показанную в виде SEQ ID NO:40.
Подходящие клетки-хозяева для экспрессии полипептида данного изобретения включают в себя клетки прокариотов, дрожжей, мицелиальных грибов или высшие эукариотические клетки. Подходящие клонирующие и экспрессирующие векторы для применения с бактериальными, грибными, дрожжевыми клетками-хозяевами или клетками-хозяевами млекопитающих описаны, например, в Pouwels et al. Cloning Vectors: A Laboratory Manual, Elsevier, New York, (1985). Бесклеточные системы трансляции могут быть также использованы для получения полипептидов данного изобретения с использованием РНК, полученных из ДНК-конструктов.
Прокариоты включают в себя грамотрицательные или грамположительные организмы, например, E. coli. Подходящие прокариотические клетки-хозяева для трансформации включают в себя, например, E. coli, Bacillus subtilis, Salmonella typhimurium, и различные другие виды в родах Pseudomonas, Streptomyces и Staphylococcus.
Экспрессирующие векторы для применения в прокариотических клетках-хозяевах содержат один или несколько фенотипических генов селектируемых маркеров. Геном фенотипического селектируемого маркера является, например, ген, кодирующий белок, который придает устойчивость к антибиотику или который обеспечивает автотрофную потребность. Векторы, предпочтительные для применения в бактериях, включают в себя, например, pET24b или pET22b, доступные из Novagen, Madison, WI (pET-24b(+) и pET-22b(+) = pET Expression System 24b (Cat. No. 69750) и 22b (Cat. No. 70765), соответственно, EMD Biosciences, Inc., Novagen Brand, Madison, WI; см. http://worldwideweb.emdbiosciences.com раздел информации о продукте, рассматривающий pET-24b и pET-22b, в отношении деталей для этого вектора), pQE70, pQE60 и pQE-9, доступные из Qiagen Inc., Valencia, CA; векторы pBS, векторы PHAGES CRIPT, векторы BLUESCRIPT, pNH8A, pNH16a, pNH18A, pNH46A, доступные из Stratagene, LaJolla, CA; и ptrc99a, pKK223-3, pKK233-3, pDR540, pRIT5, доступные из Pharmacia (теперь Pfizer, Inc., New York, NY). Среди предпочтительных эукариотических векторов находятся pWLNEO, pSV2CAT, pOG44, pXT1 и pSG, доступные из Stratagene; и pSVK3, pBPV, pMSG и pSVL, доступные из Pharmacia. Другие подходящие векторы будут очевидными для квалифицированного специалиста.
Бактериальные промоторы, подходящие для применения в данном изобретении включают в себя промоторы lacI и lacZ E. coli, промоторы T3 и T7, промотор gpt, промоторы лямбда PR и PL и промотор trp. Подходящие эукариотические промоторы включают в себя немедленно ранний промотор CMV, промотор тимидинкиназы HSV, ранний и поздний промоторы SV40, промоторы ретровирусных LTR, такие как промоторы вируса саркомы Рауса (RSV), и промоторы металлотионеина, такие как промотор металлотионеина-I мыши. Например, последовательности промоторов, используемых для экспрессирующих векторов рекомбинантных прокариотических клеток-хозяев, включают в себя, но не ограничиваются ими, промотор β-лактамазы (пенициллиназы), промотор лактозы, систему промоторов триптофана (trp) и промотор tac (Maniatis, Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, 1982). Особенно применимая система экспрессии прокариотических клеток-хозяев использует промотор фага λΡL и термолабильную репрессорную последовательность cI857ts. Плазмидные векторы, доступные из Американской Коллекции Типовых Культур, которые включают в себя производные промотора λPL, включают в себя плазмиду pHUB2 (обитателя в штамме E. coli JMB9 (ATCC 37092)) и pPLc28 (обитателя в E. coli RR1 (ATCC 53082)).
Полипептиды данного изобретения могут быть также экспрессированы в дрожжевых клетках-хозяевах, предпочтительно из рода Saccharomyces (например, S. cerevisiae). Могут быть также использованы другие роды дрожжей, такие как Pichia или Kluyveromyces. Дрожжевые векторы часто содержат сайт инициации репликации из плазмиды 2μ дрожжей, автономно реплицирующуюся последовательность (ARS), район промотора, последовательности для полиаденилирования, последовательности для терминации транскрипции и ген селектируемого маркера. Подходящие последовательности промоторов для векторов дрожжей включают в себя, среди других, промоторы для металлотионеина, 3-фосфоглицераткиназы или других гликолитических ферментов, таких как энолаза, глицеральдегид-3-фосфатдегидрогеназа, гексокиназа, пируватдекарбоксилаза, фосфофруктокиназа, глюкозо-6-фосфатизомераза, 3-фосфоглицератмутаза, пируваткиназа, триозофосфатизомераза, фосфоглюкозоизомераза и глюкокиназа. Другие подходящие векторы и промоторы для применения в экспрессии дрожжей, включают в себя глюкозо-репрессируемый промотор ADH2. Могут быть сконструированы челночные векторы, реплицируемые как в дрожжах, так и в E. coli, инсертированием ДНК-последовательностей из pBR322 для отбора и репликации в E. coli (ген Ampr и сайт инициации репликации) в эти дрожжевые векторы.
В некоторых вариантах осуществления может быть использована лидерная последовательность а-фактора дрожжей для управления секрецией этого полипептида. Эта лидерная последовательность а-фактора может быть инсертирована между промоторной последовательностью и последовательностью структурного гена. Другие лидерные последовательности, подходящие для облегчения секреции рекомбинантных полипептидов из дрожжей-хозяев, известны квалифицированным в данной области специалистам. Лидерная последовательность может быть модифицирована вблизи ее 3'-конца для содержания одного или нескольких сайтов рестрикции. Это будет облегчать слияние лидерной последовательности со структурным геном.
Протоколы трансформации дрожжей известны квалифицированным в данной области специалистам. Один такой протокол описан Hinnen et al, Proc. Natl. Acad. Sci. USA 75:1929, 1978. Этот протокол Hinnen et al. отбирает на Trp+ трансформанты в селективной среде, где эта селективная среда состоит из 0,67% азотистого основания дрожжей, 0,5% казаминокислот, 2% глюкозы, 10 мкг/мл аденина и 20 мкг/мл урацила. Клетки-хозяева дрожжей, трансформированные векторами, содержащими последовательность промотора ADH2, могут выращиваться для индукции экспрессии в богатой среде. Примером богатой среды является среда, состоящая из 1% дрожжевого экстракта, 2% пептона и 1% глюкозы, дополненных 80 мкг/мл аденина и 80 мкг/мл урацила. Дерепрессия промотора ADH2 происходит, когда глюкоза в среде истощается.
Системы культивирования клеток-хозяев млекопитающих или насекомых также могут быть использованы для экспрессии рекомбинантных полипептидов. Бакуловирусные системы для получения гетерологичных белков в клетках насекомых обсуждаются Luckow et al., Bio/Technology 6:47 (1988). Установленные клеточные линии, происходящие из животных, могут быть также использованы. Примеры подходящих линий клеток-хозяев млекопитающих включают в себя линию COS-7 клеток почки обезьяны (ATCC CRL 1651), L-клетки, C127-клетки, 3T3-клетки (ATCC CCL 163), клетки яичника китайского хомячка (CHO), HeLa-клетки и линии клеток BHK (ATCC CRL 10) и линия клеток CV-1/EBNA-1, полученная из клеточной линии почки Африканской зеленой мартышки CVI (ATCC CCL 70), как описано McMahan et al, EMBO J. 10:2821 (1991).
Другие подходящие клеточные линии включают в себя, но не ограничиваются ими, HeLa-клетки, клетки почки baby-хомячка (BHK), клетки почки обезьяны (COS-1), клетки гепатоцеллюлярной карциномы человека (например, HepG2), трансформированные аденовирусом клетки 293 человека, L-929-клетки мыши, клеточные линии HaK хомячка, мышиные клетки 3T3, полученные из Swiss, Balb-c или NIH мышей и ряд других клеточных линий. Другой подходящей линией клеток млекопитающих является клеточная линия CV-1. Подходящими также являются нормальные диплоидные клетки, клеточные штаммы, происходящие из in vitro культуры первичной ткани, а также первичных эксплантатов. Кандидатные клетки могут быть генотипически недостаточными в отношении гена селективного маркера или могут содержать доминантно действующий ген отбора.
В некоторых вариантах осуществления введение векторной конструкции в культивируемую клетку-хозяина может осуществляться трансфекцией с использованием фосфата кальция, ДЭАЭ-декстран-опосредуемой трансфекцией, опосредуемой катионным липидом трансфекцией, электропорацией, трансдукцией, инфекцией или другими способами. Такие способы описаны во многих стандартных лабораторных справочниках, таких как Davis et al, Basic Methods In Molecular Biology, 2nd Edition (1995).
Например, эти клетки-хозяева могут быть трансформированы одним или несколькими векторами, несущими ДНК, содержащую нуклеотидную последовательность, кодирующую полипептид данного изобретения, например, способами, известными в данной области, и затем могут культивироваться при подходящих условиях, если желательно, с амплификацией одного или обоих введенных генов. Затем экспрессированный полипептид может быть извлечен и очищен из культуральной среды (или из клетки, например, при внутриклеточной экспрессии) способами, известными квалифицированному в данной области специалисту. В некоторых вариантах осуществления, экспрессированный полипептид может быть получен в виде белкового комплекса, например, в виде гомодимера благодаря одной или нескольким межцепочечным дисульфидным связям между двумя разделенными полипептидами с FVIII или без FVIII.
Регуляторные последовательности транскрипции и трансляции для экспрессирующих векторов клеток-хозяев млекопитающих могут происходить из вирусных геномов. Обычно используемые последовательности промоторов и последовательности энхансеров получают из полиомавируса, Аденовируса 2, Вируса 40 обезьяны (SV40) и цитомегаловируса (CMV). ДНК-последовательности, полученные из вирусного генома SV40, например, сайта инициации репликации SV40, раннего и позднего промотора, энхансера, сайтов сплайсинга и полиаденилирования, могут быть использованы для обеспечения других генетических элементов для экспрессии последовательности структурного гена в клетке-хозяине млекопитающего. Особенно применимы вирусные ранние и поздние промоторы, так как и те, и другие легко могут быть получены из вирусного генома в виде фрагмента, который может также содержать вирусный сайт инициации репликации.
Векторы, подходящие для репликации в клетках млекопитающих, могут включать в себя вирусные репликоны или последовательности, которые обеспечивают интеграцию последовательности, кодирующей этот полипептид, в геном хозяина. Подходящие векторы могут включать в себя, например, векторы, которые произведены из обезьяньего вируса SV40, ретровирусов, вируса папилломы коров, вируса коровьей оспы и аденовируса. Компоненты этих векторов, например, репликоны, гены отбора, энхансеры, промоторы и т.п., могут быть получены из природных источников или синтезированы известными процедурами.
Например, подходящим вектором может быть вектор, полученный из вирусов коровьей оспы. В этом случае, гетерологичнную ДНК инсертируют в геном вируса коровьей оспы. Способы инсертирования чужеродной ДНК в геном вируса коровьей оспы известны в данной области и используют, например, гомологичную рекомбинацию. Инсертирование гетерологичной ДНК производят обычно в ген, который не является незаменимым по природе, например, ген тимидинкиназы (tk), который также обеспечивает селектируемый маркер.
Таким образом, экспрессирующие векторы млекопитающих могут содержать одну или несколько эукариотических единиц транскрипции, которые способны экспрессировать клетки млекопитающих. Например, единица транскрипции может содержать по меньшей мере один промоторный элемент для опосредования транскрипции чужеродных ДНК-последовательностей. В некоторых вариантах осуществления, промоторы для клеток млекопитающих включают в себя вирусные промоторы, такие как промоторы из SV40, CMV, вируса саркомы Рауса (RSV), аденовируса (ADV) и вируса папилломы коров (BPV).
Единица транскрипции может также содержать последовательность терминации и последовательности присоединения поли(А), функционально связанные с последовательностью, кодирующей этот полипептид. Единица транскрипции может также содержать энхансерную последовательность для усиления экспрессии.
Необязательно, могут быть также включены последовательности, позволяющие амплификацию гена, как и последовательности, кодирующие селектируемые маркеры. Селектируемые маркеры для клеток млекопитающих известны в данной области и включают в себя, например, гены тимидинкиназы, дигидрофолатредуктазы (вместе с метотрексатом в качестве усилителя DHFR), аминогликозидфосфотрансферазы, гигромицин В-фосфотрансферазы, аспарагинсинтетазы, аденозиндезаминазы, металлотионеина и устойчивые к антибиотикам гены, такие как ген неомицина. Или, например, векторная ДНК может содержать весь геном или часть генома вируса папилломы коров или переноситься в клеточных линиях, таких как клетки мыши С127, в виде стабильного эписомного элемента.
Могут быть сконструированы неограничивающие примеры экспрессирующих векторов и систем применения клеток-хозяев млекопитающих, например, как описано в статьях Okayama et al, Mol. Cell. Biol. 3:280 (1983), Cosman et al, Mol. Immunol. 23:935 (1986) (система для стабильной экспрессии высокого уровня ДНК в мышиных эпителиальных клетках С127 молочной железы), Cosman et al, Nature 312:768 (1984) (экспрессирующий вектор PMLSV N1/N4; ATCC 39890), EP-A-0367566, и Патенте США № 5,350,683, все из которых включены здесь посредством ссылки на описания экспрессирующих векторов и/или систем экспрессии. Векторы могут быть произведены из ретровирусов. В некоторых вариантах осуществления, вместо природной сигнальной последовательности, может быть включена гетерологичная сигнальная последовательность, такая как сигнальная последовательность для интерлейкина-7 (IL-7), описанная, например, в Патенте США № 4965195; сигнальная последовательность для рецептора интерлейкина-2, описанная, например, Cosman et al, Nature 312:768 (1984); сигнальная последовательность для интерлейкина-4, описанная, например, в EP 367566; сигнальный пептид рецептора интерлейкина типа I, описанный, например, в Патенте США № 4968607; и сигнальный пептид рецептора интерлейкина-1 типа II, описанный, например, в EP 460846, каждый из которых включен здесь посредством ссылки на описание сигнальных последовательностей.
В одном варианте осуществления рекомбинантный полипептид может быть получен при помощи технологии PER.C6® (Crucell, Holland, The Netherlands). Экспрессия рекомбинантных белков описана, например, в Патенте США 6855544, который включен здесь посредством ссылки на описание способов и композиций для получения рекомбинантных белков в линии клеток человека.
Обсуждается также, что полипептиды этого изобретения могут быть получены твердофазными синтетическими способами. См. Houghten, R. A., Proc. Natl. Acad. Set USA 52:5131-5135 (1985); и Патент США № 4631211, выданный Houghten et al. (1986).
В других вариантах осуществления данное изобретение включает в себя также рекомбинантные полипептиды, содержащие первую и вторую аминокислотные последовательности, где эти полипептиды дифференциально модифицированы во время трансляции или после трансляции, например, гликозилированием, ацетилированием, фосфорилированием, амидированием, дериватизацией известными защитными/блокирующими группами, протеолитическим расщеплением, образованием связи с молекулой антитела или другим клеточным лигандом и т.д. Любая из многочисленных химических модификаций может проводиться известными способами, в том числе, но не только, специфическим химическим расщеплением цианогенбромидом, трипсином, химотрипсином, папаином, протеазой S. aureus V8, NaBEL4; ацетилированием, деамидированием, формилированием, метилированием, окислением, восстановлением; метаболическим синтезом в присутствии туникамицина; и т.д. Дополнительные посттрансляционные модификации, включенные в это изобретение, включают в себя, например, N-связанные или O-связанные углеводные цепи, процессинг N-концевой или C-концевой сторон, присоединение химических цепей к аминокислотному скелету, химические модификации N-связанных или О-связанных углеводных цепей и добавление N-концевого остатка метионина, как результат векторов и конструкций, приспособленных для экспрессии рекомбинантных полипептидов, например, для экспрессии в прокариотических культивируемых клетках-хозяевах.
В некоторых вариантах осуществления, в которых нерастворимые полипептиды выделяют из клетки-хозяина (например, прокариотической клетки), эта клетка может быть подвергнута действию буфера подходящей ионной силы для солюбилизации большинства белков хозяев, в которых агрегированные представляющие интерес полипептиды могут быть по существу нерастворимыми, и разрушением этих клеток для высвобождения телец включения и приданием им доступности для высвобождения, например, центрифугированием. Этот способ известен квалифицированному в данной области специалисту, и его вариация описана, например, в Патенте США № 4511503, который включен здесь посредством ссылки на описание способа солюбилизации гетерологичного белка, продуцируемого в нерастворимой преломляющей свет форме в культуре рекомбинантных клеток. Не ограничивая себя какой-либо конкретной теорией, авторы изобретения считают, что экспрессия рекомбинантного белка, например, в E. coli, может приводить к внутриклеточному отложению этого рекомбинантного белка в нерастворимых агрегатах, называемых тельцами включения. Отложение рекомбинантных белков в тельцах включения может быть выгодным как вследствие того, что эти тельца включения накапливают высокоочищенный рекомбинантный белок, так и вследствие того, что белок, заключенный в тельцах включения, защищен от действия бактериальных протеаз.
Обычно клетки-хозяева (например, клетки E. coli) собирают после подходящего периода роста и суспендируют в подходящем буфере перед разрушением посредством лизиса с использованием таких способов, как, например, механические способы (например, ультразвуковой осциллятор) или химические или ферментативные способы. Примеры химических или ферментативных способов разрушения клеток включают в себя образование сферопластов, которое предусматривает использование лизосомы для лизиса бактериальной стенки, и осмотический шок, который предусматривает обработку жизнеспособных клеток раствором высокой тоничности и холодной водой низкой тоничности для высвобождения полипептидов.
После разрушения клеток-хозяев суспензию обычно центрифугируют для осаждения телец включения. Полученный осадок содержит по существу все полипептиды из фракции нерастворимых полипептидов, но, если процесс разрушения клеток является неполным, эта фракция может содержать также интактные клетки или фрагменты разрушенных клеток. Полнота разрушения клеток может быть анализирована ресуспендированием этого осадка в небольшом количестве того же самого буферного раствора и испытанием этой суспензии с использованием фазово-контрастного микроскопа. Присутствие фрагментов разрушенных клеток или целых клеток указывает на то, что необходимо дополнительное разрушение для удаления фрагментов или клеток и ассоциированных не преломляющих свет полипептидов. После такого разрушения, если необходимо, эта суспензия может быть снова центрифугирована, и осадок может быть извлечен, ресуспендирован и анализирован. Этот процесс может повторяться, пока визуальное обследование не выявит отсутствие фрагментов разрушенных клеток в осажденном материале или пока дальнейшая обработка не перестанет уменьшать размер (количество) полученного осадка. После получения из солюбилизированных телец включения или на более поздней стадии очистки, этот полипептид может быть подходящим образом повторно уложен в подходящем буфере для рефолдинга, таким как буферы, известные в данной области. Степень любого отсутствия укладки может быть определена хроматографией, в том числе обращенно-фазовой высокоразрешающей жидкостной хроматографией (ОФ-ВЖХ (RP-HPLC).
Если эти рекомбинантно экспрессируемые полипептиды данного изобретения не находятся уже в растворимой форме, перед тем, как они подвергаются рефолдингу, они могут быть солюбилизированы инкубированием в буфере для солюбилизации, содержащем хаотропный агент (например, мочевину, гуанидин) и восстанавливающий агент (например, глутатион, дитиотреитол (ДТТ), цистеин) в количествах, необходимых для солюбилизации по существу этих полипептидов. Эта инкубация происходит при условиях концентрирования полипептидов, времени инкубации и температуре инкубации, которые будут позволять солюбилизацию этого полипептида. Измерение степени солюбилизации может проводиться по определению мутности, анализом фракционирования полипептидов между супернатантом и осадком после центрифугирования на восстанавливающих ДСН-гелях, анализом белка (например, с использованием набора Bio-Rad для анализа белков) или высокоэффективной жидкостной хроматографией (ВЖХ).
pH буфера для солюбилизации может быть щелочным, предпочтительно приблизительно pH 7,5, причем предпочтительным является диапазон от приблизительно pH 7,5 до приблизительно pH 11. Концентрация полипептида данного изобретения в буферном растворе для солюбилизации должна быть такой, чтобы полипептид был по существу солюбилизированным и частично или полностью восстановленным и денатурированным. Альтернативно, рекомбинантный полипептид может быть первоначально нерастворимым. Точное количество для использования будет зависеть, например, от концентраций и типов других ингредиентов в этом буферном растворе, особенно от типа и количества восстанавливающего агента, типа и количества хаотропного агента и рН этого буфера. Например, концентрация рекомбинантного полипептида может быть увеличена, если одновременно увеличивается концентрация восстанавливающего агента, например, глутатиона.
В дополнительных вариантах осуществления данное изобретение обеспечивает гомогенные или по существу гомогенные полипептиды, содержащие первую и вторую аминокислотные последовательности. В одном варианте осуществления, данное изобретение обеспечивает выделенный полипептид, содержащий первую аминокислотную последовательность, присутствующую в полипептиде vWF, и вторую аминокислотную последовательность, гетерологичную относительно первой, причем этот полипептид способен связывать FVIII. В других вариантах осуществления, этот полипептид очищают по существу до гомогенности, как показано единственной белковой полосой после анализа при помощи электрофореза на ДСН-полиакриламидном геле (электрофореза на ДСН-ПААГ).
Как будет понятно квалифицированному в данной области специалисту, процедуры для очистки рекомбинантных белков будут варьироваться в соответствии с такими факторами, как тип используемых клеток-хозяев, и от того, секретируются или не секретируются эти белки в культуральную среду. Например, при использовании систем экспрессии, которые секретируют рекомбинантный белок, эта культуральная среда сначала может быть сконцентрирована с использованием коммерчески доступного фильтра для концентрирования белка, например, ультрафильтрационного устройства Amicon или Millipore Pellicon. После стадии концентрирования, этот концентрат может быть нанесен на матрикс очистки, такой как гель-фильтрационная среда. Альтернативно, может быть использована анионообменная смола, например, матрикс или субстрат, имеющие боковые диэтиламиноэтильные группы (DEAE). Этими матриксами могут быть акриламид, агароза, декстран, целлюлоза или другие типы, обычно используемые в очистке белка. Альтернативно, может быть использована катионообменная стадия. Подходящие катионообменники включают в себя матриксы, содержащие сульфопропильные или карбоксиметильные группы. Предпочтительными являются сульфопропильные группы. Наконец, могут быть использованы одна или несколько стадий обращенно-фазовой высокоэффективной жидкостной хроматографии (ОФ-ВЖХ), использующие гидрофобные ОФ-ВЖХ-среды (например, силикагель, имеющий боковые метильные или другие алифатические группы), для дополнительной очистки рекомбинантно экспрессируемого полипептида. Некоторые или все из предыдущих стадий очистки, в различных комбинациях, могут быть использованы для обеспечения по существу гомогенного рекомбинантного белка.
В некоторых вариантах осуществления используют аффинную колонку, содержащую партнер связывания структуры или домена, определяемого второй аминокислотной последовательностью, для аффинной очистки экспрессируемых рекомбинантных полипептидов или содержащих их белковых комплексов. Например, когда вторая аминокислотная последовательность соответствует полипептиду Fc антитела, может быть использована аффинная колонка, содержащая белок А или белок G, для аффинной очистки этого полипептида или содержащих его белковых комплексов. В некоторых вариантах осуществления, связанные полипептиды и/или комплексы могут быть удалены из аффинной колонки в высокосолевом буфере для элюции и затем диализованы в низкосолевой буфер для применения. В способе другого примера, эта аффинная колонка может содержать антитело, которое связывает полипептиды или содержащие их белковые комплексы, например, антитело против структуры или домена, определяемого первой или второй аминокислотной последовательностью.
В других аспектах обеспечена нуклеотидная последовательность, кодирующая полипептид данного изобретения, где этот полипептид содержит первую аминокислотную последовательность, присутствующую в полипептиде vWF, и вторую аминокислотную последовательность, гетерологичную относительно первой, где этот полипептид способен связывать FVIII. В одном варианте осуществления, данное изобретение обеспечивает выделенную молекулу нуклеиновой кислоты, содержащую нуклеотидную последовательность, представленную в SEQ ID NO:23, SEQ ID NO:24, SEQ ID NO:25, SEQ ID NO:26, SEQ ID NO:27, SEQ ID NO:28, 37, 42 и 43.
Полинуклеотиды этого изобретения могут включать в себя варианты, которые имеют замены, делеции и/или добавления, которые могут включать в себя один или несколько нуклеотидов. Эти варианты могут быть изменены в кодирующих районах, некодирующих районах или в обоих. Изменения в кодирующих районах могут продуцировать консервативные или неконсервативные аминокислотные замены, делеции и добавления. Особенно предпочтительными среди них являются молчащие замены, добавления и делеции, которые не изменяют свойств и способности связывания FVIII полипептида данного изобретения.
Дополнительные варианты осуществления включают в себя молекулы нуклеиновых кислот, содержащие полинуклеотид, имеющий нуклеотидную последовательность, по меньшей мере на 90% идентичную и более предпочтительно по меньшей мере на 95%, 96%, 97%, 98% или 99% идентичную (a) нуклеотидной последовательности, кодирующей полипептид, имеющий аминокислотные последовательности, представленные здесь; и (b) нуклеотидную последовательность, комплементарную любой из нуклеотидных последовательностей в (a) выше.
Под полинуклеотидом, имеющим нуклеотидную последовательность, по меньшей мере, например, на 95% "идентичную" ссылочной нуклеотидной последовательности, кодирующей полипептид, имеется в виду, что нуклеотидная последовательность этого полинуклеотида является идентичной ссылочной последовательности, за исключением того, что эта полинуклеотидная последовательность может включать в себя до пяти точковых мутаций на каждые 100 нуклеотидов ссылочной нуклеотидной последовательности, кодирующей этот полипептид. Другими словами, для получения полинуклеотида, имеющего нуклеотидную последовательность, по меньшей мере на 95% идентичную ссылочной нуклеотидной последовательности, до 5% нуклеотидов в этой ссылочной последовательности могут быть делетированы или заменены другим нуклеотидом, или число нуклеотидов до 5% общих нуклеотидов в ссылочной последовательности могут быть инсертированы в эту ссылочную последовательность. Эти мутации могут встречаться при 5'- или 3'-концевых положениях этой ссылочной последовательности в любом месте между этими концевыми положениями, рассеянными либо индивидуально среди нуклеотидов в ссылочной последовательности, либо в одной или нескольких смежных группах в этой ссылочной последовательности.
Две или более полинуклеотидных последовательностей могут сравниваться определением их процентной идентичности. Две или более аминокислотных последовательностей подобным образом могут сравниваться определением их процентной идентичности. Процентная идентичность двух последовательностей, последовательностей нуклеиновых кислот или пептидных последовательностей, обычно описывается как количество точных совпадений между двумя сопоставляемыми последовательностями, деленное на длину более короткой последовательности и умноженное на 100. Приближенное сопоставление для последовательностей нуклеиновых кислот обеспечивается алгоритмом локальной гомологии Smith and Waterman, Advances in Applied Mathematics 2:482-489 (1981). Этот алгоритм может быть распространен для применения с пептидными последовательностями с использованием матрицы замен, разработанной Dayhoff, Atlas of Protein Sequences and Structure, M. O. Dayhoff ed., 5 suppl. 3:353-358, National Biomedical Research Foundation, Washington, D.C., USA, и нормализованной Gribskov, Nucl. Acids Res. 14(6):6Ί 5-6763 (1986). Одно воплощение этого алгоритма для последовательностей нуклеиновых кислот и пептидных последовательностей обеспечено Genetics Computer Group (Madison, Wis.) в их BESTFIT utility application. Параметры по умолчанию для этого способа описаны в Wisconsin Sequence Analysis Package Program Manual, Version 8 (1995) (доступном из Genetics Computer Group, Madison, Wis.).
Например, вследствие вырожденности генетического кода, квалифицированному в данной области специалисту будет понятно, что ряд молекул нуклеиновых кислот, имеющих последовательность, по меньшей мере на 90%, 95%, 96%), 97%, 98% или 99% идентичную любой из описанных здесь последовательностей нуклеиновых кислот, могут кодировать этот полипептид.
Действительно, так как вырожденные варианты этих нуклеотидных последовательностей, все, кодируют один и тот же полипептид, это будет ясно квалифицированному в данной области специалисту даже без выполнения каких-либо функциональных анализов или измерений, описанных здесь. Кроме того, будет также известно в данной области, что, для таких молекул нуклеиновых кислот, которые не являются вырожденными вариантами, некоторое разумное количество будет также кодировать полипептид, имеющий способность связывания FVIII. Это объясняется тем, что квалифицированному в данной области специалисту полностью известны аминокислотные замены, которые либо имеют меньшую вероятность значимого влияния на связывание белка, либо не имеют вероятности значимо влиять на связывание белка (например, замена одной алифатической аминокислоты второй алифатической аминокислотой).
Недавно, успехи в синтетическом получении более длинных полинуклеотидных последовательностей позволили синтетическое получение нуклеиновых кислот, кодирующих более длинные полипептиды без применения традиционных способов клонирования. Коммерческие поставщики таких служб включают в себя Blue Heron, Inc., Bothell, WA (http://worldwideweb.blueheronbio.com). Технология, используемая Blue Heron, Inc., описана в Патентах США с номерами 6664112; 6623928; 6613508; 6444422; 6312893; 4652639; Опубликованных Заявках на патент США с номерами 20020119456A1; 20020077471A1; и Опубликованных заявках на международный патент (с номерами) WO 03054232A3; WO 0194366A1; WO 9727331A2 и WO 9905322A1, все из которых включены здесь посредством ссылки.
Конечно, общие способы молекулярной биологии, микробиологии и рекомбинантных нуклеиновых кислот могут также быть использованы для получения полинуклеотидов этого изобретения. Эти способы хорошо известны и объяснены, например, в Current Protocols in Molecular Biology, F. M. Ausebel, ed., Vols. I, II and III (1997); Sambrook et at, Molecular Cloning: A Laboratory Manual, 2nd Edition, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. (1989); DNA Cloning: A Practical Approach, D. N. Glover, ed., Vols. I and II (1985); Oligonucleotide Synthesis, M. L. Gait, ed. (1984); Nucleic Acid Hybridization, Hames and Higgins, eds. (1985); Transcription and Translation, Hames and Higgins, eds. (1984); Animal Cell Culture, R. I. Freshney, ed. (1986); Immobilized Cells and Enzymes, IRL Press (1986); Perbal, "A Practical Guide to Molecular Cloning"; the series, Methods in Enzymology, Academic Press, Inc. (1984); Gene Transfer Vectors for Mammalian Cells, J. H. Miller and M. P. Calos, eds., Cold Spring Harbor Laboratory (1987); and Methods in Enzymology, Wu and Grossman and Wu, eds., соответственно, Vols. 154 and 155, все, включенные здесь посредством ссылки.
Обеспечен также, в других аспектах, экспрессирующий вектор, содержащий молекулы нуклеиновых кислот, кодирующие полипептиды данного изобретения. Обеспечены также клетки-хозяева, которые экспрессируют полипептиды данного изобретения. В одном варианте осуществления, данное изобретение обеспечивает клетку, экспрессирующую полипептид, содержащий первую аминокислотную последовательность, присутствующую в полипептиде vWF, и вторую аминокислотную последовательность, гетерологичную относительно первой, где этот полипептид способен связывать FVIII, причем эта клетка дополнительно экспрессирует FVIII. В другом варианте осуществления, этим FVIII является рекомбинантный FVIII.
IV. Белковый комплекс
В другом аспекте данное изобретение обеспечивает белковый комплекс, содержащий полипептид и FVIII, где этот полипептид содержит первую аминокислотную последовательность, присутствующую в полипептиде vWF, и вторую аминокислотную последовательность, гетерологичную относительно первой, где этот полипептид способен связывать FVIII. В одном варианте осуществления, этот комплекс содержит две отдельные цепи этого полипептида в форме димера в комплексе с FVIII.
В другом варианте осуществления данное изобретение обеспечивает гомодимерный белковый комплекс, содержащий две полипептидные цепи данного изобретения, где между этими цепями образуются одна или несколько дисульфидных связей. В одном варианте осуществления, одна или несколько дисульфидных связей образуются между первыми аминокислотными последовательностями двух отдельных цепей с образованием посредством этого димера. В других вариантах осуществления, одна или несколько дисульфидных связей образуются между Fc-районами двух отдельных цепей с образованием посредством этого димера. В некоторых вариантах осуществления, этот гомодимерный комплекс состоит из или по существу состоит из двух полипептидных цепей данного изобретения. В других дополнительных вариантах осуществления, в объеме данного изобретения находятся гетеродимеры.
В другом варианте осуществления данное изобретение обеспечивает олигомеры, например, дополнительным связыванием димеров. В некоторых вариантах осуществления, обеспечены олигомеры разного размера, предпочтительно с образованием дисульфидных мостиков при амино-концевых сторонах полипептида данного изобретения. Таким образом, в других вариантах осуществления, обеспечены олигомеры с разными размерами в диапазоне от димера по меньшей мере приблизительно: 100000, 250000, 500000 дальтон или более, в том числе мультимеры приблизительно: 5, 10, 20, 30, 40 или 50 миллионов дальтон или более.
В других дополнительных вариантах осуществления эти олигомеры являются гомо- или гетероолигомерами. В другом варианте осуществления, этот димер является гетеродимером.
В других вариантах осуществления этот белковый комплекс получают из системы экспрессии культуры клеток или ткани, которая экспрессирует этот полипептид и FVIII. В одном варианте осуществления, этот полипептид и FVIII коэкспрессируются в одной и той же клетке.
В одном варианте осуществления данное изобретение обеспечивает растворимый слитый белок, содержащий первую аминокислотную последовательность, слитую с N-концом полипептида Fc, где эта первая аминокислотная последовательность присутствует в полипептиде vWF, причем этот полипептид способен связывать FVIII. В некоторых вариантах осуществления, этот полипептид способен связывать FVIII в виде димера, содержащего два растворимых слитых белка, соединенных дисульфидными связями.
В другом варианте осуществления данное изобретение обеспечивает димер, содержащий два растворимых слитых белка, соединенных дисульфидными связями, где каждый белок содержит первую аминокислотную последовательность, слитую с N-концом полипептида Fc, где эта первая аминокислотная последовательность присутствует в полипептиде vWF, причем этот димер способен связывать FVIII.
В других аспектах данное изобретение обеспечивает белковый комплекс, содержащий дисульфид-связанные мультимеры, содержащие два или более полипептида, имеющих первую и вторую аминокислотную последовательность.
В одном варианте осуществления эти дисульфид-связанные мультимеры получают контактированием полипептидов с пропептидным фрагментом vWF, посредством чего этот пропептидный фрагмент vWF действует "in trans" для управления сборкой дисульфид-связанных мультимеров.
В некоторых вариантах осуществления пропептидный фрагмент vWF содержит аминокислотную последовательность, представленную в SEQ ID NO:31, или ее вариант. В другом варианте осуществления пропептидный фрагмент vWF кодируется нуклеотидной последовательностью, представленной в SEQ ID NO:32, или ее вариантом.
В другом варианте осуществления это контактирование включает в себя коэкспрессию этого полипептида с пропептидным фрагментом vWF.
Например, в одном варианте осуществления, данное изобретение обеспечивает белковый комплекс, содержащий дисульфид-связанные мультимеры, содержащие два или более из полипептидов, имеющих первую и вторую аминокислотную последовательность, где эта первая аминокислотная последовательность представлена в SEQ ID NO:1, SEQ ID NO:4 или SEQ ID NO:7. В других вариантах осуществления, эта первая аминокислотная последовательность представлена в SEQ ID NO:3, SEQ ID NO:6 или SEQ ID NO:9. В некоторых вариантах осуществления, эта первая аминокислотная последовательность представлена в SEQ ID NO:17, SEQ ID NO:18 или SEQ ID NO:19. В другом варианте осуществления, этот белковый комплекс получают коэкспрессией, с использованием рекомбинантной системы экспрессии, полипептида с пропептидным фрагментом, содержащим аминокислотную последовательность, представленную в SEQ ID NO:31, посредством чего этот фрагмент действует "in trans" для управления сборкой дисульфид-связанных мультимеров, содержащих эти полипептиды.
V. Способы
В других дополнительных аспектах данное изобретение обеспечивает способ получения белкового комплекса данного изобретения. В некоторых вариантах осуществления, используют аффинную колонку, содержащую партнер связывания структуры или домена, определяемого второй аминокислотной последовательностью. Например, если вторая аминокислотная последовательность соответствует полипептиду Fc антитела, для аффинной очистки этого комплекса может быть использована аффинная колонка, содержащая белок А или белок G. В некоторых вариантах осуществления, партнер связывания может быть иммобилизован. Альтернативно, эта аффинная колонка может содержать антитело, которое связывает Fc-часть этого полипептида или которая связывает структуру или домен, определяемый первой аминокислотной последовательностью этого полипептида. В некоторых вариантах осуществления, получаемый комплекс содержит димер, содержащий два растворимых слитых белка, соединенные дисульфидными связями, где каждый белок содержит первую аминокислотную последовательность, слитую с N-концом Fc-полипептида, где эта первая аминокислотная последовательность присутствует в полипептиде vWF, причем этот димер способен связывать FVIII.
В другом варианте осуществления этот комплекс дополнительно содержит FVIII, связанный с этим димером. FVIII может быть удаляемым/диссоциируемым из этого комплекса и, необязательно, подвергаемым одной или нескольким дополнительным стадиям очистки для получения частично очищенного, по существу чистого или чистого FVIII.
В одном аспекте данное изобретение обеспечивает способ получения FVIII, предусматривающий: контактирование FVIII с полипептидом с образованием белкового комплекса, содержащего FVIII и этот полипептид, где этот полипептид содержит первую аминокислотную последовательность, присутствующую в полипептиде vWF, и вторую аминокислотную последовательность, гетерологичную относительно первой, причем этот полипептид способен связывать FVIII с образованием белкового комплекса.
В одном варианте осуществления этот способ дополнительно предусматривает селективное прикрепление этого комплекса к среде разделения, содержащей партнер связывания, имеющий аффинность в отношении района или домена, определяемого второй аминокислотной последовательностью. В другом варианте осуществления, эта вторая аминокислотная последовательность соответствует Fc-району иммуноглобулина. В других вариантах осуществления, этим партнером связывания является белок А или белок G. В одном варианте осуществления, этим партнером связывания является антитело. В некоторых вариантах осуществления, этим партнером связывания является антитело против Fc-района иммуноглобулина. Еще в одном варианте осуществления, этот комплекс содержит две цепи этого полипептида в форме димера, где этот димер аффинно связан с FVIII.
Для иммобилизации партнера связывания может быть использовано любое число различных твердых носителей. Например, материал твердого носителя может состоять из полисахаридов, таких как целлюлоза, крахмал, декстран, агар или агароза, или гидрофильных синтетических полимеров, таких как замещенные или незамещенные полиакриламиды, полиметакриламиды, полиакрилаты, полиметакрилаты, поливиниловые гидрофильные полимеры, полистирол, полисульфон или т.п. Другие подходящие материалы для применения в качестве материала твердого носителя, включают в себя пористые минеральные материалы, такие как диоксид кремния, оксид алюминия, оксид титана, оксид циркония и другие церамические структуры. Альтернативно, в качестве материала твердого носителя могут быть использованы композиционные материалы. Такие композиционные материалы могут быть образованы сополимеризацией или с использованием взаимопроникающей сети двух или нескольких из вышеупомянутых молекулярных частиц. Примеры подходящих композиционных материалов включают в себя структуры полисахарид-синтетические полимеры и/или структуры полисахарид-минерал и/или структуры синтетический полимер-минерал, такие как описанные в Патентах США с номерами 5268097, 5234991 и 5075371, каждый из которых включен здесь посредством ссылки на описание композиционных материалов.
Материал для твердого носителя данного изобретения может быть в форме гранул или неровных частиц в диапазоне от приблизительно 0,1 мм до 1000 мм в диаметре, волокон (полых или иных) любого размера, мембран, плоских поверхностей в диапазоне по толщине от приблизительно 0,1 мм до 1 мм, и пористых материалов с порами от мкм до нескольких мм в диаметре.
Предпочтительно, партнеры связывания являются химически иммобилизованными на материале твердого носителя через ковалентную связь, образованную между, например, меркаптогруппой партнера связывания и реакционноспособной группой, присутствующей на этом твердом носителе. Реакционноспособные группы, способные реагировать с меркаптогруппой данного лиганда, включают в себя эпоксигруппы, тозилаты, трезилаты, галогениды и винильные группы. Поскольку многие вышеупомянутые материалы твердого носителя не включают в себя ни одной из вышеуказанных реакционноспособных групп, могут быть использованы бифункциональные активирующие агенты, способные как реагировать с материалами твердого носителя, так и обеспечивать необходимые реакционноспособные группы. Примеры подходящих активирующих агентов включают в себя эпихлоргидрин, эпибромгидрин, дибром- и дихлорпропанол, дибромбутан, простой диглицидиловый эфир этиленгликоля, простой диглицидиловый эфир бутандиола, дивинилсульфон и т.п.
Типичными примерами подходящих носителей являются Сефароза™, агароза, смола активированная СН-Sepharose™ 4B (N-гидроксисукцинимидсодержащая агароза) из Pharmacia (Sweden), смола NHS-активированная Sepharose™ 4 Fast Flow (активированная 6-аминогексановой кислотой для образования активных N-гидроксисукцинимидных эфиров; Amersham Biosciences), смола CNBr-активированная Sepahrose™ Fast Flow (активированная цианогенбромидом; Amersham Biosciences) смола PROTEIN PAK™ эпокси-активированная аффинная смола (Waters, USA), смола EUPERGIT™ C30 N (Rohm & Haas, Germany), UltraLink Biosupport Medium (Pierce), Trisacryl GF-2000 (Pierce) или AFFI-GEL™ из BioRad (USA). Предпочтительно, этот носитель для аффинной хроматографии предварительно активируют эпоксидными группами для прямого связывания с пептидами и белками.
Смолы для аффинной хроматографии, применимые для применения на практике способов этого изобретения, включают в себя, но не ограничиваются ими, любую комбинацию лиганда или вышеописанного соединения, с любым из носителей, описанных выше. Неограничивающими примерами конкретных смол для аффинной хроматографии являются Белок A-Sepharose™, Белок A-агароза, Белок A-агароза CL-4B, Белок G-Sepharose™, Белок G-агароза, Белок G-агароза CL-4B, Белок A/G агароза (различные версии всех из них доступны от различных производителей, например, Sigma-Aldrich, Amersham, и Pierce), Белок A Ultraflow™ (Sterogene), Белок A Cellthru™ 300 (Sterogene), QuickMab (Sterogene), QuickProtein A™ (Sterogene), Thruput™ и Thruput Plus (Sterogene), PROSEP-A или PROSEP-G (Millipore) и любые вариации вышеописанных смол.
Способы, используемые для аффинной хроматографии, могут зависеть, по меньшей мере частично, от конкретного используемого и обычно поставляемого производителем или известного в данной области реагента. Например, реагент для аффинной хроматографии может быть упакован в хроматографической колонке, уравновешен буфером, способным стимулировать взаимодействие между белковым комплексом и партнером связывания и затем контактирован с композицией, содержащей этот комплекс. Затем эта колонка может быть промыта по меньшей мере одним раствором, способным элюировать примеси без нарушения взаимодействия между комплексом и аффинным лигандом. Целый комплекс, содержащий этот полипептид и FVIII, может быть затем элюирован с использованием подходящего элюента для получения этого комплекса. Альтернативно, FVIII может быть диссоциирован из этого комплекса, посредством чего этот комплекс остается связанным с партнером связывания.
Например, в способе получения белкового комплекса, содержащего этот полипептид (например, димер этого полипептида) со связанным с ним FVIII или без связанного FVIII, композиция, содержащая этот комплекс, может быть контактирована по меньшей мере с одной смолой для аффинной хроматографии, которая имеет Белок А в качестве лиганда, при условиях, которые позволяют связывание этого комплекса с этой смолой. Желательно, этот Белок А является природно-встречающейся или рекомбинантной формой Белка А. Затем эта хроматографическая смола может быть промыта рядом промывочных буферов, имеющих увеличивающуюся кислотность (например, pH 7,0, 6,5, 6,0, 5,8, 5,5, 5,2, 5,0, 4,8, 4,6, 4,5, 4,4 или 4,0), так что это промывание вызывает диссоциацию материала, не находящегося в комплексе из смолы, но по существу не диссоциирует этот комплекс. Эта смола может быть промыта по меньшей мере один раз, предпочтительно по меньшей мере два раза и наиболее предпочтительно по меньшей мере три раза промывочными буферами, где первый буфер имеет pH приблизительно 5,0-6,0, предпочтительно приблизительно 5,2, и каждый последующий промывочный буфер имеет pH, который является более кислотным, чем предыдущий промывочный буфер. В предпочтительных вариантах осуществления, промывочные буферы не будут диссоциировать более 80%, 70%, 60%, 50%, 40%, 30%, 20%, 10% или 5% этого комплекса из смолы.
В некоторых вариантах осуществления после промывания смолы этот комплекс может быть элюирован из хроматографической смолы с использованием элюента, имеющего кислотный pH приблизительно 2,5-3,5 (например, pH 2,5, 3,0, 3,5) и являющегося более кислотным, чем любой из промывочных буферов. Этот элюат содержит очищенный, частично очищенный или по существу очищенный комплекс с предпочтительной чистотой по меньшей мере 50%, 60%, 70%, 80%, 85%, 90% или 95% или большей.
В других вариантах осуществления, когда белковый комплекс, который должен быть получен, содержит FVIII, связанный с димерной формой этого полипептида, связывание со смолой и промывание могут выполняться при таких условиях, что FVIII остается аффинно связанным с димером, который, в свою очередь остается связанным со смолой. Затем FVIII может быть отделен от связанного со смолой димера с получением композиции, содержащей FVIII, где эта композиция является частично, по существу или полностью свободной от димера.
Таким образом, в некоторых вариантах осуществления, полипептид, содержащий первую и вторую аминокислотные последовательности, может быть использован для получения FVIII, благодаря его способности связывать FVIII, для образования белкового комплекса, содержащего этот полипептид и FVIII. В некоторых вариантах осуществления, такой белковый комплекс может быть подвергнут аффинной хроматографии для получения этого комплекса и/или ассоциированного с ним FVIII.
В других аспектах данное изобретение обеспечивает композицию, содержащую FVIII, где этот FVIII получают в соответствии с описанными здесь способами. Обеспечены также композиции, содержащие белковый комплекс, содержащий полипептид, имеющий первую и вторую аминокислотную последовательность, с FVIII, связанным с полипептидом этого комплекса, или без FVIII, связанного с полипептидом этого комплекса. В одном варианте осуществления, эта композиция содержит белковый комплекс, содержащий два этих полипептида в форме димера (например, гомо- или гетеродимера), связанного с FVIII.
Таким образом, в некоторых аспектах, данное изобретение обеспечивает композиции (например, слитый белок укороченного рекомбинантного vWF) и способы усиления экспрессии рекомбинантного FVIII клетками млекопитающих защитой его от удаления и/или деградации, а также для создания возможности быстрой хроматографической очистки полученного комплекса FVIII:рекомбинантный vWF с использованием регулятора слитого белка (например, второй аминокислотной последовательности), включенной в рекомбинантный полипептид vWF данного изобретения.
Таким образом, в различных вариантах осуществления, обеспечен надежный и простой способ для использования укороченной версии vWF, связанной с Fc-частью иммуноглобулина (например, иммуноглобулина G1), для образования комплекса с FVIII и защиты FVIII во время экспрессии и получения, аналогично полной молекуле vWF. Описанный здесь уникальный подход является более эффективным (1) по меньшей мере вследствие меньшего размера и более легкой экспрессии слитых полипептидов vWF данного изобретения, и/или (2) возможности быстрой очистки слитого комплекса FVIII:vWF посредством Fc-района на слитом белке, следовательно, с селективным обогащением желаемой молекулой FVIII с меньшим загрязнением от других белков, а также от vWF. Кроме того, высокоаффинное связывание Fc-района обеспечивает то, что слитый комплекс FVIII:vWF может быть также удален in situ в подключенном картридже с белком А, например, для увеличения выхода и чистоты.
В одном варианте осуществления данное изобретение обеспечивает способ подключенной очистки во время процесса перфузии для непрерывного очищения слитого комплекса FVIII:рекомбинантный vWF посредством селективного аффинного присоединения vWF-слитого Fc-района к матриксу Белка A (или другому сходному матриксу).
Данное изобретение в некоторых вариантах осуществления использует преимущество по меньшей мере двух наблюдений для создания превосходного способа очистки FVIII: (а) использование рекомбинантных полипептидов vWF, в частности, рекомбинантных укороченных полипептидов vWF, которые сохраняют связывание с FVIII, но являются значимо меньшими, чем полноразмерная молекула, и (b) создание слитого белка с Fc-районом иммуноглобулина (например, IgG1), который связывается с высокой аффинностью с лигандом, Белком А. Эта комбинация обеспечивает сильную экспрессию в системах клеток млекопитающих, с возможностью создания быстрого способа сбора комплекса FVIII:recvWF независимо от среды или от других компонентов в супернатанте экспрессии, а также от проблем, связанных с ионной силой и т.д. Кроме того, она позволяет использовать различные реагенты/растворы для удаления FVIII с сохранением полной активности и с превосходным выходом продукта. Высокоаффинное связывание Fc-части с Белок А-колонкой (или высокоаффинным антителом) не препятствует взаимодействию FVIII:vWF, так как она является отдельной и независимой молекулярной частицей.
В дополнительных аспектах данное изобретение обеспечивает способ рекомбинантного получения полипептидов данного изобретения. В некоторых вариантах осуществления, этот способ предусматривает: (a) генерирование линии клеток млекопитающих трансформацией этой линии клеток экспрессирующим вектором, кодирующим полипептид данного изобретения; (b) выращивание этой линии клеток при условиях, достаточных для экспрессии этого полипептида; и (c) очистку экспрессированного белка из стадии (b) для получения слитого белка vWF-Fc, где vWF-часть этого слитого белка является укороченным vWF, который лишен по меньшей мере одного домена зрелого полноразмерного полипептида vWF, причем этот слитый белок способен образовывать мультимеры, которые способны связывать белок FVIII. В одном варианте осуществления, этот укороченный vWF имеет домены D' и D3, при условии, что этот укороченный vWF лишен домена A1, A2, A3, D4, B, C1, C2, CK или их комбинации. В другом варианте осуществления, этот укороченный vWF имеет домены D', D3 и A1, при условии, что этот укороченный vWF лишен доменов A2, A3, D4, B, C1, C2 и CK. В некоторых вариантах осуществления, этот укороченный vWF имеет домены D', D3, A1 и A2, при условии, что этот укороченный vWF лишен доменов A3, D4, B, C1, C2 и CK. В других вариантах осуществления, этот укороченный vWF имеет домены D', D3, A1, A2 и A3, при условии, что этот укороченный vWF лишен доменов D4, B, C1, C2 и CK. В других дополнительных вариантах осуществления, этот укороченный vWF лишен доменов D4, B, C1, C2 и CK. В других вариантах осуществления, этот способ дополнительно предусматривает коэкспрессию рекомбинантного пропептида, где этот слитый белок экспрессируется в виде рекомбинантного слитого белка, который дополнительно лишен пропептидной последовательности, причем этот рекомбинантный пропептид и рекомбинантный слитый белок vWF-Fc ассоциируются с образованием комплекса пропептид/vWF-Fc после рекомбинантной экспрессии.
VI. Другие композиции и способы
В других аспектах данное изобретение обеспечивает способы и композиции для FVIII, имеющего пролонгированный период полувыведения из плазмы. Например, в некоторых вариантах осуществления, эти слитые полипептиды рекомбинантный vWF-Fc данного изобретения могут быть использованы в качестве добавки к рекомбинантному или полученному из плазмы FVIII, которая может стимулировать пролонгированный период полувыведения для FVIII и/или комбинированного комплекса. В соответствии с данным изобретением, в некоторых вариантах осуществления, Fc-район сливают с укороченным фрагментом vWF, так что обеспечивается связывание FVIII и этот Fc-район дополнительно обеспечивает дополнительное время полужизни этому комплексу. Например, в одном варианте осуществления, слитый комплекс рекомбинантный FVIII:рекомбинантный vWF-Fc может быть очищен непосредственно из культуральной среды для экспрессии, затем инъецирован (например, внутривенно) в пациентов в виде фармацевтической композиции, например, с избытком слитого белка рекомбинантный vWF-Fc (т.е., полипептида данного изобретения), или без него. В некоторых вариантах осуществления, дополнение этого комплекса избытком слитого белка vWF-Fc (например, 2-, 3-, 4-, 5-, 6-, 7-, 8-, 9-, 10-кратного в расчете на моль) обеспечено для увеличения концентрации этого слитого полипептида в плазме и обеспечения повторного связывания FVIII со слитым белком vWF-Fc, если он диссоциируется в плазме.
В других дополнительных вариантах осуществления слитые полипептиды данного изобретения могут быть приготовлены в более высокой концентрации с FVIII, введены человеку или субъекту не человеку внутривенно или другими способами, позволяя этому FVIII быть ассоциированным со слитым белком vWF во время хода его циркуляции и до расщепления тромбином, высвобождаемым в участке повреждения.
Таким образом, в некоторых вариантах осуществления, полипептиды данного изобретения и/или белковый комплекс, содержащий их, могут быть приготовлены для лечения. Например, полипептиды и/или белковые комплексы данного изобретения могут быть приготовлены в соответствии с известными способами приготовления фармацевтически приемлемых композиций, предпочтительно объединенных в смеси с фармацевтически приемлемым носителем. Подходящие носители и их приготовление, в том числе другие белки человека, например, сывороточный альбумин человека, описаны, например, в руководстве Remington's Pharmaceutical Sciences by E. W. Hartin, которое включено посредством ссылки. В одном варианте осуществления, такие композиции содержат эффективное количество белка FVIII в комплексе с рекомбинантным полипептидом vWF, вместе с подходящим количеством носителя для приготовления фармацевтически приемлемых композиций, подходящих для эффективного введения субъекту, например, парентерально вводимых субъекту, страдающему, например, от гемофилии А.
Средняя находящаяся в обращении доза для лечения пациентов с гемофилией варьируется в зависимости от тяжести приступа кровотечения. Например, средние дозы, вводимые внутривенно, могут находиться в диапазоне: приблизительно 40 единиц на килограмм FVIII для предоперационных показаний, приблизительно 15 - приблизительно 20 единиц на килограмм для минорного кровотечения и приблизительно 20 - приблизительно 40 единиц на килограмм, вводимые на протяжении приблизительно 8-часового периода времени, для поддерживающей дозы. Другие дозы и схемы введения могут быть легко определены специалистом с обычной квалификацией в области лечения пациентов с гемофилией.
VII. Набор
В дополнительных аспектах обеспечены также наборы, содержащие полипептиды, последовательности нуклеиновых кислот, белковые комплексы и/или композиции данного изобретения. Эти наборы могут иметь единый контейнер, или они могут иметь отдельный контейнер для каждого желаемого компонента. Рассматриваются также наборы, содержащие реагенты, необходимые для приготовления рекомбинантных полипептидов и/или белковых комплексов, полученных из них, например, таких как, но не только, экспрессирующие векторы, рекомбинантные клетки-хозяева, содержащие эти экспрессирующие векторы, и реагенты для очистки. Кроме того, когда компоненты этого набора обеспечены в виде одного или нескольких жидких растворов, этот жидкий раствор является водным раствором, причем особенно предпочтительным является стерильный водный раствор. Однако, компоненты этого набора могут быть обеспечены в виде высушенного порошка (высушенных порошков). Когда реагенты или компоненты обеспечены в виде сухого порошка, этот порошок может быть воспроизведен добавлением подходящего растворителя. Предполагается, что этот растворитель также может быть обеспечен.
Следующие примеры приводятся только для иллюстрации данного способа и не должны ограничивать это изобретение. Специалисту, квалифицированному в данной области, будет понятно, что приведенные примеры только иллюстрируют то, является заявленным, и что данное изобретение ограничено только объемом прилагаемой формулы изобретения.
ПРИМЕРЫ
Пример 1
Конструирование экспрессионных плазмид для
слитых полипептидов укороченный vWF-Fc
Шесть молекул ДНК, каждая из которых имеет нуклеотидную последовательность, которая кодирует слитый полипептид укороченный vWF-Fc, синтезировали коммерчески (GENEART AG, Regensburg, Germany). Идентификаторы последовательностей для нуклеотидных последовательностей этих ДНК-молекул и для соответствующих кодируемых аминокислотных последовательностей представлены в таблице 1.
Нуклеотидные и аминокислотные последовательности для полипептидов укороченный vWF-Fc
Для каждой молекулы ДНК часть нуклеотидной последовательности молекулы, кодирующей укороченный район vWF, кодон-оптимизировали с использованием алгоритмов, которые ответственны за использование кодонов, вторичную структуру, ингибирующие последовательности и т.п. ДНК-район Fc каждой молекулы, который соответствует аминокислотной последовательности, показанной в SEQ ID NO:16 и полученной из IgG1, не подвергали оптимизации в отношении последовательности. Кроме того, каждая молекула ДНК содержит нуклеотидную последовательность, которая кодирует последовательность сигнального пептида, показанную в виде SEQ ID NO:40. SEQ ID NO:20-22 дополнительно включают в себя аминокислотную последовательность пропептида, показанную в SEQ ID NO:41.
Сайты рестрикции, включенные в эти синтетические гены, расщепляли и повторно клонировали в соответствующие сайты рестрикции в плазмидном экспрессирующем векторе pcDNA3002Neo (Crucell, Netherlands). Рекомбинантные ДНК получали из клонов ДСН-щелочным способом лизиса в соответствии с обычными известными протоколами, например, как описано Sambrook et al, (1989) Molecular Cloning: A Laboratory Manual, CSH; Jones (1995) Gel Electrophoresis: Nucleic Acids Essential Techniques, Wiley. Расщепленные рестрикционными ферментами ДНК фракционировали на 1% агарозных гелях для определения размера и идентичности. Подтверждение последовательностей плазмидных клонов выполняли автоматизированным секвенированием ДНК с использованием флуоресцентных дезоксирибонуклеотидных праймеров (Applied Biosystems, Carlsbad, CA). Полученные ДНК плазмидных экспрессирующих векторов, кодирующие различные конструкции укороченный vWF-Fc, в каждом случае стерилизовали промывкой 70% этанолом после осаждения и ресуспендировали в стерильной воде до конечной концентрации приблизительно 0,2 - приблизительно 1,0 микрограммов на микролитр.
Пример 2
Электропорация экспрессирующих плазмид в клетках PER.C6 млекопитающих
Клетки PER.C6 и клетки клональной линии 078 (клетки BDD-078), которые являются клетками PER.C6, экспрессирующими B-домен-делетированный FVIII (BDD-FVIII), поддерживали в каждом случае в непрерывной культуре рутинным (4 дня) пассированием в среде PER-MAb media (Hyclone, Logan, UT). За сорок восемь часов перед электропорацией, клетки освежали посевом клеток при 1 x 106 клеток/мл в свежей среде PER-MAb. В день электропорации, 5 x 106 клеток ресуспендировали в 100 микролитров раствора Amaxa Nucleofector® Kit V (Lonza Walkersville Inc., Walkersville, MD).
Для каждой электропорации, клеточную суспензию смешивали приблизительно с 2 - приблизительно 5 микрограммами ДНК экспрессионной плазмиды для экспрессии полипептидов, описанных в таблице 1. Затем эту смесь клеток переносили в кювету для электропорации Amaxa Nucleofector® Device (Lonza Walkersville Inc., Walkersville, MD); и использовали предварительно установленную программу X-001 для электропорации этой ДНК в клетки. После электропорации, эту суспензию клеток переносили немедленно в предварительно нагретую (37°С) среду Mab (SAFC Biosciences, Lenexa, Kansas) в лунке 6-луночного пластикового микропланшета. Клетки инкубировали без встряхивания в увлажненной камере термостата, установленной при 37°С, с 5% СО2 и 95% влажностью.
Спустя приблизительно 48 часов культивирования, каждую реакционную смесь электропорации извлекали из 6-луночного микропланшета в культуральную колбу на 125 мл с 20 мл свежей среды MAb (SAFC Bioscience, Lenexa, Kansas), содержащей подходящий антибиотик (подходящие антибиотики) для отбора: Генетицин/G418 или Зеоцин использовали в конечных концентрациях 125 или 100 микрограмм/мл, соответственно; Гигромицин использовали в конечной концентрации 50 микрограмм/мл). Затем эти клетки культивировали при 37°С, при 5% СО2 и 95% влажности без встряхивания. Затем, спустя 7-10 дней, клетки адаптировали к росту в среде PER-MAb со встряхиванием и рутинным пассированием для получения поликлонального пула трансфицированных клеток. Альтернативно, отобранные клетки получали клонированием с использованием лимитирующих разведений, как описано, например, Harlow, E and Lane, D. (1988). Antibodies: A Laboratory Manual, стр. 116-117 и стр. 222-223. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY. Клонированные клетки проверяли регулярно на рост и со временем перемещали во встряхиваемые колбы для размножения и роста в виде суспензионных клеток в среде PER-MAb (Hyclone, Logan, UT).
Пример 3
Анализы
FVIII: Коммерческий белок BDD-FVIII (a/k/a Xyntha®; Antihemophilic Factor (Recombinant)) (Wyeth (теперь дочерняя компания Pfizer Inc., New York, NY)) использовали в качестве стандарта для разработки надежного линейного анализа с использованием твердофазного иммуноферментного анализа (ELISA). Иммобилизованным антителом, связанным с микропланшетом, было коммерческое мышиное моноклональное антитело, а именно, клон GMA-012 (R8B12) (Green Mountain Antibodies, Inc., Burlington, VT), направленное против домена A2 FVIII человека; и детектирующим антителом было коммерческое биотинилированное овечье поликлональное антитело, а именно, SAF8C-APBIO, (Affinity Biologicals, Ontario, Canada), которое узнает FVIII человека. Промывание микролунок между добавлениями антитела выполняли в присутствии забуференного трисом солевого раствора (TBS), pH 7,5. Колориметрическое детектирование при 405 нм выполняли с использованием опосредуемого стрептавидин-щелочной фосфатазой расщепления субстрата pNpp. Альтернативно, колориметрическое определение в единицах активности FVIII выполняли с использованием Beckman ACL Coagulation Analyzer в соответствии с указаниями изготовителя и с использованием внутреннего контроля калибратора плазмы или при помощи набора Chromogenix Coatest SP4 FVIII (Diapharma Group, Columbus, OH) с использованием BDD-FVIII-стандарта.
vWF: Полученный из плазмы vWF категории, подходящей для исследований, (Haemtech, Essex Junction, VT) использовали в качестве стандарта-антигена для измерений ELISA содержания белка. Используемым иммобилизованным антителом является коммерческое мышиное моноклональное антитело против белка vWF человека; и для детектирования использовали HRP-конъюгированное козье антитело против vWF человека. Колориметрическое детектирование при 450 нм выполняют инкубированием с субстратом фермента, тетраметилбензидином.
Для слитых белков рекомбинантный vWF-Fc, использовали сходные форматы ELISA, такие как захват Fc-района белком А или антителом против Fc человека, иммобилизованным на микропланшете с последующим детектированием связывания мышиных поликлональных антител против vWF человека с доменами D'-D3, D'-A1 или D'-A3 vWF, в зависимости от экспрессионного конструкта.
Пример 3
Характеристика полипептидов укороченный vWF-Ас
Рекомбинантные полипептиды, соответствующие белкам укороченный vWF-Fc (т.е., D'-D3-Fc, Pro-D'-D3-Fc, D'-A1-Fc, Pro-D'-A1-Fc, D'-A3-Fc, Pro-D'-A3-Fc; таблица 1), которые экспрессировали в клетках PER.C6, иммунопреципитировали инкубированием 0,2-1 мл супернатанта клеток с 20 микролитрами гранул белка G. Гранулы концентрировали центрифугированием, затем промывали Трис-буфером. Центрифугированные гранулы суспендировали в 25 микролитрах буфера Лэммли и нагревали до 95°C в течение 10 минут перед нанесением на 4-12% градиентный Bis-Tris-гель для электрофореза в ПААГ. Визуализацию полос выполняли окрашиванием/удалением красителя Кумасси бриллиантового синего.
Для экспериментов, включающих в себя либо коэкспрессию BDD-FVIII и слитого pro-D'-A3-Fc, либо смешивание коммерческого BDD-FVIII, Xyntha® (Wyeth) с супернатантами клеток PER.C6, экспрессирующих pro-D'-D3-Fc, пробы, элюированные из аффинных колонок, электрофоретически разделяли на восстанавливающих и денатурирующих 7% NuPAGE Трис-ацетат полиакриламидных гелях, окрашивали красителем Кумасси синим, удаляли краситель и фотографировали для сравнения с известными маркерами молекулярных масс, подвергаемыми электрофорезу на том же самом геле.
Анализ последовательности
Рекомбинантные полипептиды, разделенные при помощи электрофореза в полиакриламидном геле (PAGE), подвергали блоттингу на PVDF-мембраны; полосы белков, визуализированные быстрым окрашиванием Ponceau S и удалением красителя, вырезали из этих мембран в подготовке для расщепления по Эдману (AIBioTech, Richmond VA) для получения информации в отношении N-концевой последовательности. Эти полученные амино-концевые последовательности сопоставляли с известными последовательностями либо из N-конца зрелого vWF, либо из N-концевой пропептидной последовательности, которая появлялась в полосах, предположительно представляющих неполностью процессированные продукты vWF.
In vitro ассоциация BDD-FVIII и белков укороченный vWF-Fc
Как только клетки, экспрессирующие белки D'-D3-Fc, Pro-D'-D3-Fc, D'-A1-Fc, Pro-D'-A1-Fc, D'-A3-Fc и Pro-D'-A3-Fc, были отобраны и размножены, пробы клеточного супернатанта осветляли и добавляли к активно растущим клеткам BDD-078 (клону клеток млекопитающих, экспрессирующему B-домен-делетированный FVIII). Клетки BDD-078 высевали при 12,5×106 клеток/мл с супернатантами клеток PER.C6 от клеток, трансфицированных плазмидными конструктами, для экспрессии слитых белков укороченный vWF-Fc (т.е., D'-D3-Fc, Pro-D'-D3-Fc, D'-A1-Fc, Pro-D'-A1-Fc, D'-A3-Fc, Pro-D'-A3-Fc). Для "BDD-контроля" кондиционированную среду заменяли. Клетки выращивали в течение 2 дней со встряхиванием при 37°С, как описано. В этой временной точке, брали аликвоты, центрифугировали и супернатанты испытывали как на активность FVIII, так и на антиген FVIII и vWF. Только активность FVIII показана на фигуре 8. Заштрихованный столбец показывает увеличение выше контроля BDD-FVIII без добавленных белков vWF-Fc.
При нормальных условиях, BDD-FVIII, экспрессируемый из клеток BDD-078, является, по-видимому, катаболизированным или секвестрированным (отторгнутым) этими клетками (Kalind et al., J. Biotechnology, 147:198-204 (2010)). Однако, добавление либо полученного из плазмы белка vWF, либо супернатантов клеток, экспрессирующих один из слитых белков укороченный vWF-Fc, приводило к более высокому извлечению активности FVIII, что позволяет предполагать защитное действие против нежелательного поглощения или катаболизма FVIII растущими клетками. Этот результат демонстрирует, что слитые белки укороченный vWF-Fc могут функционировать, увеличивая выход FVIII в культуре. После 2 дней в культуре при 37°С с экспрессируемым FVIII, брали аликвоты и оценивали на накапливание активности FVIII. Наблюдали существенное увеличение извлечения активности FVIII (фигура 8).
Анализ мультимеров
Полученный из плазмы vWF (pd-vWF) и рекомбинантные полипептиды оценивали на их способность образовывать высокомолекулярные комплексы с использованием электрофоретического анализа на невосстанавливающих 1,6% или 2% плавящихся при высокой температуре (HGT-P) агарозных гелях (модифицированного из Raines et al, Thrombosis Res., 60:201-212 (1990). Полученный из плазмы FVIII (Koate-DVI®) использовали в качестве стандарта для электрофореза для оценивания мультимеров vWF-Fc. Белки переносили способами полусухого блоттинга на нитроцеллюлозную бумагу на устройстве iBlot (Invitrogen Corp., Rockville, MD) и блокировали с использованием раствора SuperBlock (Pierce, Rockford, IL). Пробы инкубировали с кроличьим поликлональным антителом против vWF человека (Abcam cat# ab6994) с последующим добавлением конъюгированного со щелочной фосфатазой козьего антитела против фрагмента F(ab')2 IgG кролика (cat# A3937; Sigma); детектирование полос выполняли инкубированием с раствором Western Blue (Invitrogen Corp., Carlsbad, CA). Промывание в дистиллированной воде использовали для гашения реакции; и полосы визуализировали на BioRad Molecular Imager ChemiDoc XRS+ Imaging System (Bio-Rad Laboratories Hercules, CA). Эти результаты показаны на фигуре 9.
Очистка белков и хроматография на Белке А
Осветленные супернатанты из культур клеток, экспрессирующих BDD-FVIII и белок pro-D'-A3-Fc, получали трехкратным центрифугированием и разделением при 2500 x g в течение 7 минут, с последующим центрифугированием при 2,500 x g в течение 11 минут. Затем супернатанты фильтровали через глубинный фильтр Sartobran 150 (0,45 микрометров), затем через фильтр из ацетата целлюлозы (0,2 микрометра) (#5231307-H4-00) (Sartorius Stedim Stedim Biotech S.A., Aubagne, France). Фильтры предварительно увлажняли с использованием насоса 100-200 мл 20 mM Tris, pH 7,0, затем клеточным супернатантом и, наконец, промывали приблизительно 25 мл 20 мМ Трис, рН 7,0. Конечный фильтрат был разбавленным в два раза клеточным супернатантом и служил материалом, используемым для колоночной хроматографии. Отфильтрованные супернатанты наносили на колонку Protein A-HiTrap размером 5 или 10 мл (часть 17-1403-01) (GE Healthcare, Piscataway, NJ) на системе AKTA Explorer Chromatography System (GE Healthcare, Piscataway, NJ). Систему трубок предварительно промывали 200 мМ Трис, рН 7,0 и колонку Protein A промывали пятью колоночными объемами 20 мМ Трис, рН 7,0 для гарантии стабильного фона перед нанесением пробы на эту колонку. Отфильтрованные пробы проходили через колонку при 5 миллилитрах в минуту и элюат собирали в виде "протекающего через колонку потока". После нанесения всего материала, колонку Protein A-HiTrap дополнительно промывали пятью колоночными объемами 20 мМ Трис, рН 7,0, пока не достигалась стабильная фоновая линия. В этой временной точке колонку промывали четырьмя колоночными объемами 20 мМ Трис, рН 7,0, содержащим 0,1 M CaCl2, и элюированный материал собирали. Затем Фактор VIII, связанный со слитыми белками укороченный vWF-Fc, элюировали 20 мМ Трис, рН 7,0, содержащим 0,3 M CaCl2, пока эта фракция не элюировалась (приблизительно тремя колоночными объемами). Затем эту колонку промывали пятью дополнительными колоночными объемами 20 мМ Трис, рН 7,0. Белки укороченный vWF-Fc, остающиеся связанными с белком А, снимали из этой колонки добавлением 250 мМ Глицина, 150 мМ NaCl, рН 3,9. Альтернативные способы элюции были описаны, например, Arakawa et al, Prot. Expr. Purif, 63: 158-163 (2009). Все собранные пробы из колонки Protein A-HiTrap сохраняли, испытывали на активность Фактора VIII с использованием хромогенного анализа и/или анализа свертывания (описанных выше) и готовили аликвоты для электрофореза в ДСН-ПААГ. В некоторых случаях, например, когда был детектирован пик активности FVIII, этот белок наносили на колонки PD10 (GE Healthcare 45000148), которые были предварительно промыты 25 мл желаемого буфера для хранения FVIII. Затем два с половиной миллилитра элюированного BDD-FVIII наносили на каждую колонку и обессоленный BDD-FVIII элюировали 3,5 мл буфера для хранения FVIII. Затем белки помещали в условия -80°C для долгосрочного хранения; и, в некоторых случаях, добавляли сывороточный альбумин до 10 мг/мл.
Очистка FVIII из pro-D'-A3-Fc/FVIII связывающего комплекса
Для определения, способны ли полипептиды укороченный vWF-Fc связывать FVIII, клетки, коэкспрессирующие BDD-FVIII и pro-D'-A3-Fc, выращивали в течение 5 дней в среде PER-MAb с 125 микрограммами на мл каждого из неомицина и зеоцина. Готовили супернатанты и хроматографировали по существу, как описано выше. В различных временных точках во время элюции пробы сохраняли и подвергали электрофорезу (фигура 10). На фигуре 10А показано хроматографическое отслеживание пиков, элюированных с различными буферными условиями, после нанесения 10 мл супернатанта клеток PER.C6, содержащих BDD-FVIII/pro-D'A3-Fc, на колонку HiTrap Protein A 5 мл (стадия 5 является промывочной стадией, стадия 6 является элюатом после 0,1 M CaCl2, стадия 7 является элюатом FVIII после промывания 0,3 MM CaCl2, стадия 8 является элюатом после промывания 1 M CaCl2, стадия 9 является низкосолевой промывкой и стадия 10 является элюатом после очистки колонки цитратом, pH 5,5). Фигура 10B показывает электрофорез в полиакриламидном геле проб элюции. Дорожки 1-9 представляют, соответственно, (1) исходный материал, (2) проходящий через колонку поток, (3) элюат 0,1 M CaCl2, (4) элюат 0,3 M CaCl2, (5) элюат цитратом pH 5,5 (6) препарат концентрированного BDD-FVIII, (7) маркеры молекулярных масс (размеры на правой стороне), (8) клеточный супернатант от экспрессирующих pro-D'-A3-Fc клеток PER.C6, (9) коммерческий BDD-FVIII (Xyntha®), звездочки, показывающие три полосы 170, 90 и 80 кД, соответствующие полноразмерному BDD-FVIII, тяжелой цепи и легкой цепи (цепям), соответственно.
Точки рядом с полосами в дорожках 5 и 8 показывают миграцию ожидаемых размеров для содержащих пропептид белков proD'-A3-Fc и зрелого proD'-A3-Fc (высокой молекулярной массы и низкой молекулярной массы, соответственно).
По существу чистую фракцию BDD-FVIII, связанную с pro-D'A3-Fc, иммобилизовали на матриксе белка А. Этот комплекс был устойчивым к условиям промывания перед его отделением и элюированием из матрикса vWF-Fc только при стадии элюции 0,3 M CaCl2 (фигура 10А, стадия 7; и фигура 10B, дорожка 4). Сам белок pro-D'-A3-Fc визуализировался только очисткой матрикса при более жестких промывочных условиях, которые включали в себя кислотный рН 3,9 (фигура 10А, стадия 10; и фигура 10В, дорожка 5). Белки, элюированные промывкой рН 3,9, совместно мигрируют с набором белков, которые предварительно готовили и идентифицировали как аутентичные молекулы укороченный pro-D'-A3-Fc (фигура 10В, дорожка 8).
Очистка FVIII из D'-D3-Fc связывающего комплекса
10 мл раствора супернатанта клеток PER.C6, содержащего экспрессированный белок pro-D'-D3-Fc (фигура 11, дорожка 2), добавляли к 250 единицам коммерческого BDD-FVIII (Xyntha®, дорожка 3) и инкубировали при 37°C в течение 4 часов (дорожки 4 и 5). Эту смесь разбавляли в два раза 20 мМ Трис-HCl, pH 7,0 и наносили на колонку 1 мл Белок A-HiTrap по существу, как описано для очистки FVIII из комплексов pro-D'-A3-Fc выше и показано на фигуре 10. Несвязанные белки вымывали из колонки 20 мМ Трис-HCl, pH 7,0 (дорожка 6). Имеется явное удерживание FVIII, находящегося в комплексе с белком pro-D'-D3, как видно по отсутствию FVIII и/или полипептидов vWF-Fc на фигуре 11, дорожке 6 (в сравнении с дорожкой 7), с последующей сильной элюцией FVIII (дорожка 7) с использованием 0,3 M CaCl2; это вполне сравнимо с FVIII, элюированным из комплексов FVIII/pro-D'-A3-Fc (фиг. 10B, дорожка 4). Удержанные полипептиды, представляющие укороченный vWF-Fc, элюируются буфером с низким рН, 0,1 М глицином, 0,15 M NaCl, pH 3,9 (фигура 11, дорожка 8). Три звездочки в дорожке 3 совпадают с полосами белков приблизительно 170, 90 и 80 кД, соответствующих полноразмерному BDD-FVIII, тяжелой цепи и легкой цепи (цепей), соответственно. Эти результаты из очистки pro-D'D3-Fc/FVIII демонстрируют, что добавление pro-D'D3-Fc с FVIII приводит к захвату, удерживанию и последующей специфической элюции FVIII, с результатами, фактически идентичными результатам, наблюдаемым на фигуре 10.
Фармакокинетика
Очищенные слитые белки рекомбинантный vWF-Fc, связанные с FVIII, или в виде природного белка, инъецируют внутривенно в хвостовую вену мышей при приблизительно 5 микрограммах на мышь в забуференном фосфатом солевом растворе со стабилизирующим агентом, таким как альбумин. В качестве контрольных вариантов, FVIII с/без vWF инъецируют сходным образом и оценивают на клиренс в животных. Пять - восемь животных (либо дикого типа, либо имеющих заболевание с кровотечением вследствие генетической недостаточности либо FVIII, либо vWF, либо обоих) используют для испытания потери крови. В различных временных точках (например, 0 минут, 3 минуты, 15 минут, 30 минут, 1 ч, 2 ч, 4 ч, 8 ч, 16 ч, 24 ч) после инъекции, животных умерщвляют, кровь выпускают через нижнюю полую вену и получают плазму и замораживают. Затем пробы плазмы оценивают на антиген и/или функциональную активность при помощи ELISA и/или хромогенного анализа или анализа свертывания, соответственно, в зависимости от генетической среды животного. Клиренс плазмы определяют фармакокинетическим анализом антигена и/или активности в зависимости от времени (например, как описано в статьях Mordenti et al , Toxicol. Appl. Pharmacol., 137:75-78 (1996) и Lenting et al, J. Biol. Chem., 279:12102-12109 (2004), каждая из которых включена здесь для представления способа для фармакокинетического анализа), и посредством сравнения с контрольным вариантом.
| название | год | авторы | номер документа |
|---|---|---|---|
| МОДИФИЦИРОВАННЫЙ ФАКТОР ВИЛЛЕБРАНДА С УДЛИНЕННЫМ ПОЛУПЕРИОДОМ СУЩЕСТВОВАНИЯ IN VIVO, ЕГО ПРИМЕНЕНИЯ И СПОСОБЫ ПОЛУЧЕНИЯ | 2009 |
|
RU2528855C2 |
| ПРИМЕНЕНИЕ ЛЕНТИВИРУСНЫХ ВЕКТОРОВ, ЭКСПРЕССИРУЮЩИХ ФАКТОР VIII | 2019 |
|
RU2803163C2 |
| ПОЛИПЕПТИДЫ, МОДУЛИРУЮЩИЕ SIGLEC-ЗАВИСИМЫЕ ИММУННЫЕ ОТВЕТЫ | 2017 |
|
RU2776807C2 |
| ГЛИКОЗИЛИРОВАННЫЕ СЛИТЫЕ БЕЛКИ VWF С УЛУЧШЕННОЙ ФАРМАКОКИНЕТИКОЙ | 2017 |
|
RU2782212C2 |
| ВЫДЕЛЕННЫЙ ПОЛИПЕПТИД IB АЛЬФА ГЛИКОПРОТЕИНА ТРОМБОЦИТОВ ЧЕЛОВЕКА, СЛИТЫЙ БЕЛОК, МОЛЕКУЛА ДНК (ВАРИАНТЫ), ЭКСПРЕССИРУЮЩИЙ ВЕКТОР (ВАРИАНТЫ), КЛЕТКА (ВАРИАНТЫ), СПОСОБ ЭКСПРЕССИИ ПОЛИПЕПТИДА, СПОСОБ ЭКСПРЕССИИ СЛИТОГО БЕЛКА, ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ (ВАРИАНТЫ), СПОСОБ ИНГИБИРОВАНИЯ ПРИКРЕПЛЕНИЯ КЛЕТКИ КРОВИ К БИОЛОГИЧЕСКОЙ ТКАНИ В БИОЛОГИЧЕСКОЙ СИСТЕМЕ, СПОСОБ ИНГИБИРОВАНИЯ ПРИКРЕПЛЕНИЯ БЕЛКА К БИОЛОГИЧЕСКОЙ ТКАНИ В БИОЛОГИЧЕСКОЙ СИСТЕМЕ И СПОСОБ ЛЕЧЕНИЯ НАРУШЕНИЯ | 2004 |
|
RU2403262C2 |
| Выделенная нуклеиновая кислота, которая кодирует слитый белок на основе FVIII-BDD и гетерологичного сигнального пептида, и ее применение | 2022 |
|
RU2818229C2 |
| МОЛЕКУЛЫ НУКЛЕИНОВОЙ КИСЛОТЫ И ПУТИ ИХ ПРИМЕНЕНИЯ | 2018 |
|
RU2819144C2 |
| КОНЪЮГИРОВАННЫЕ МОЛЕКУЛЫ ФАКТОРА VIII | 2009 |
|
RU2573587C2 |
| ВЫСОКОГЛИКОЗИЛИРОВАННЫЙ СЛИТЫЙ БЕЛОК НА ОСНОВЕ ФАКТОРА СВЕРТЫВАНИЯ КРОВИ ЧЕЛОВЕКА VIII, СПОСОБ ЕГО ПОЛУЧЕНИЯ И ЕГО ПРИМЕНЕНИЕ | 2016 |
|
RU2722374C1 |
| МОЛЕКУЛЫ НУКЛЕИНОВОЙ КИСЛОТЫ И ПУТИ ИХ ПРИМЕНЕНИЯ ДЛЯ НЕВИРУСНОЙ ГЕННОЙ ТЕРАПИИ | 2019 |
|
RU2834625C2 |
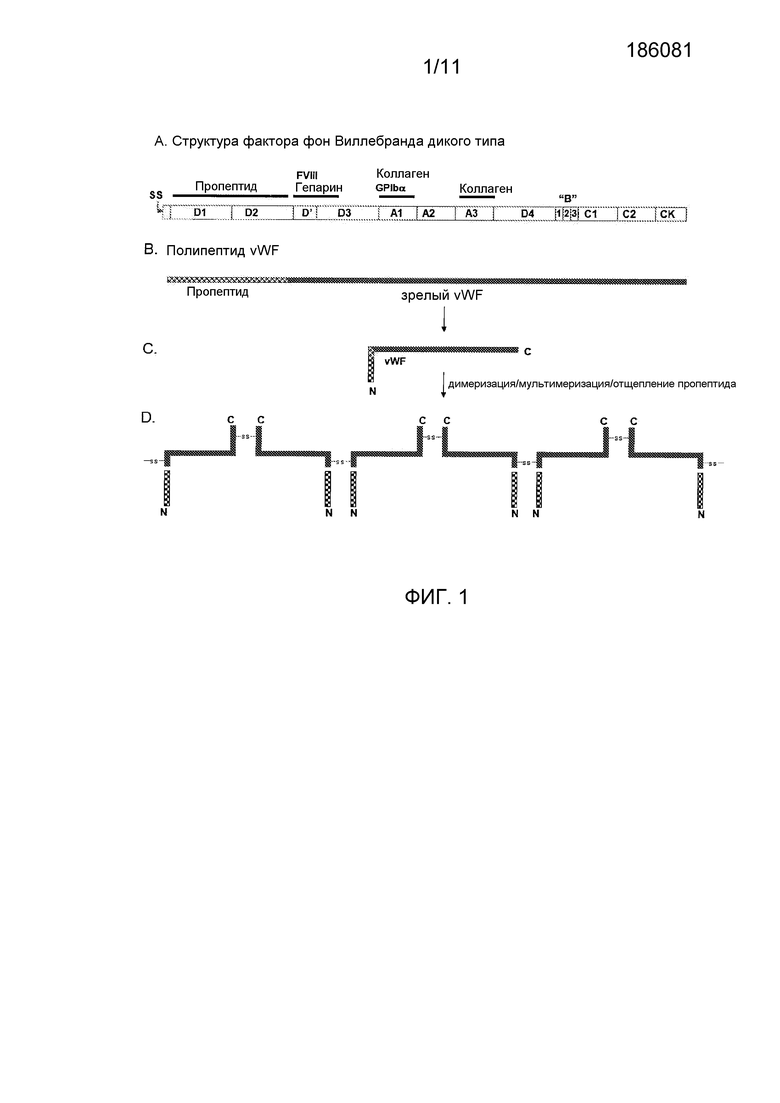

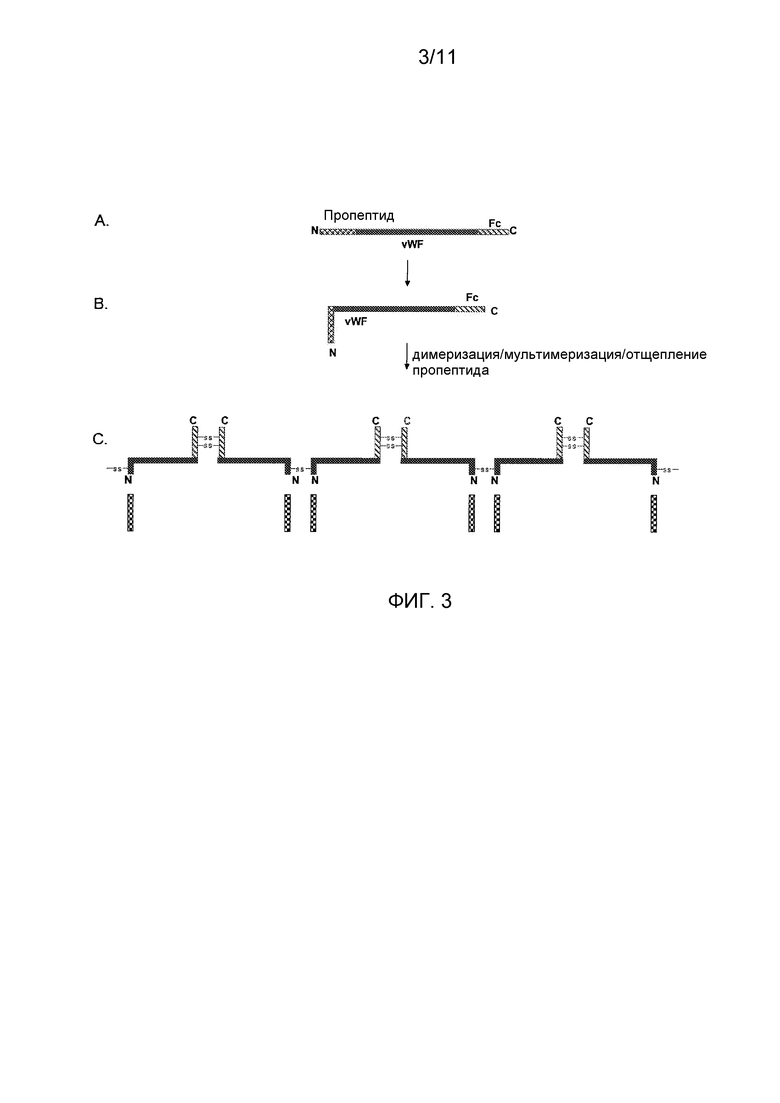
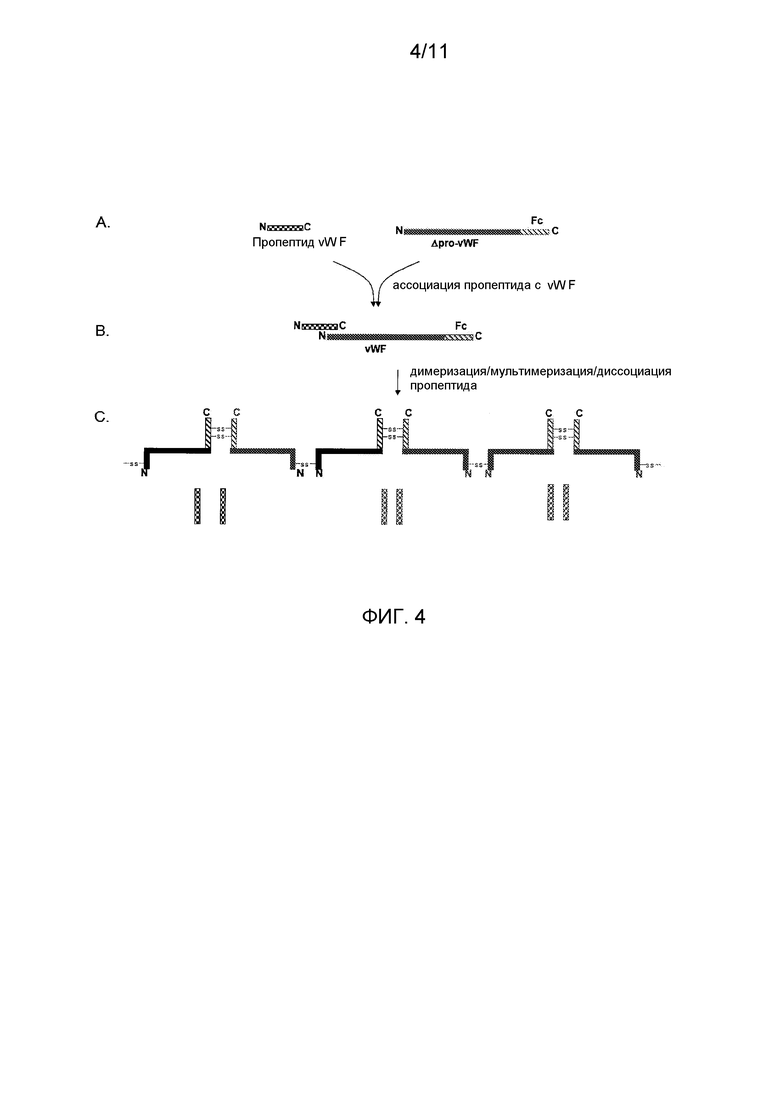
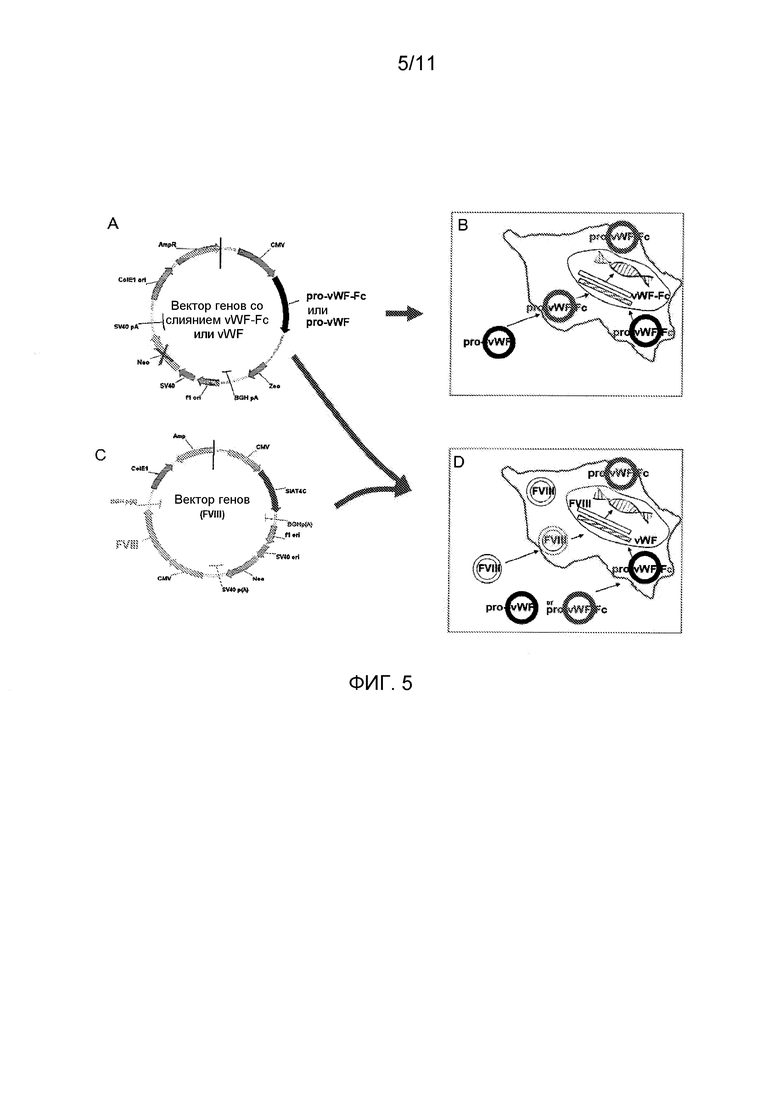
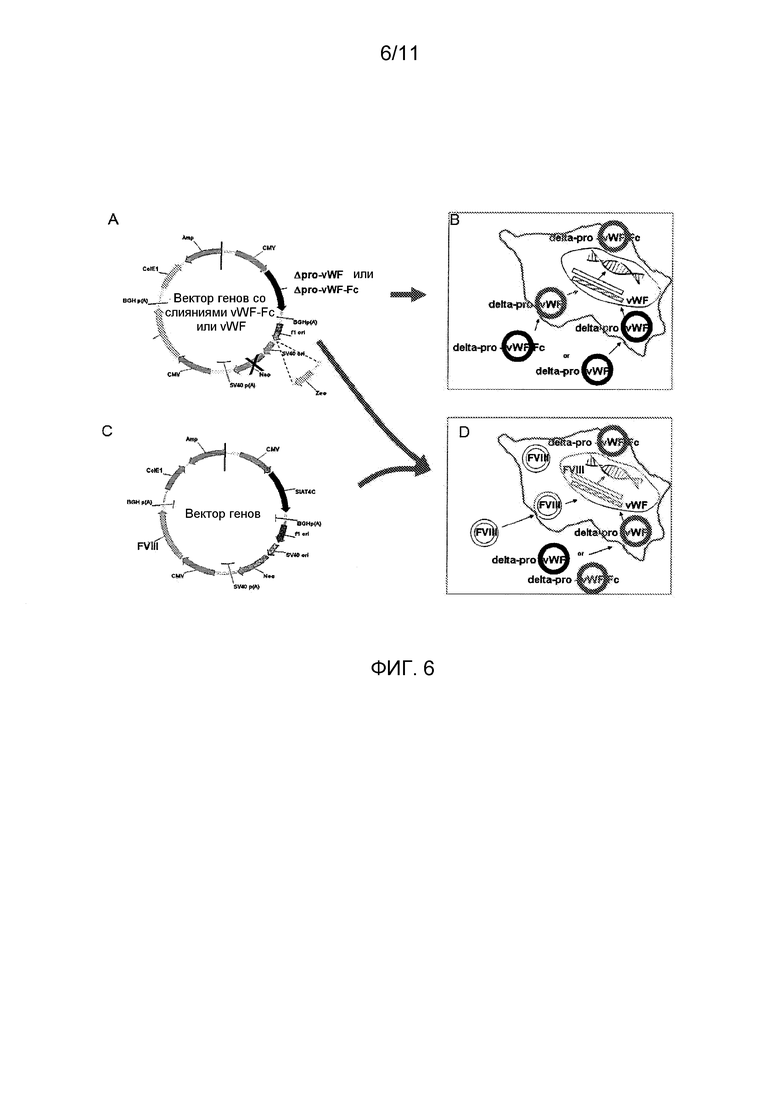
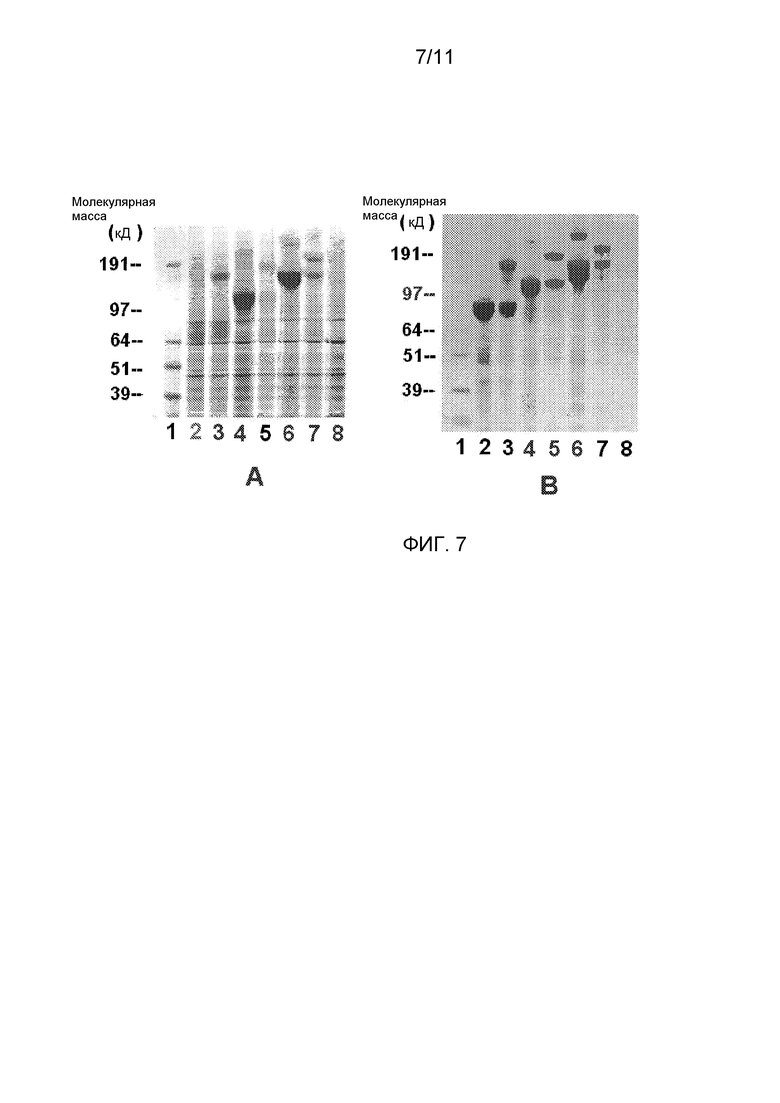
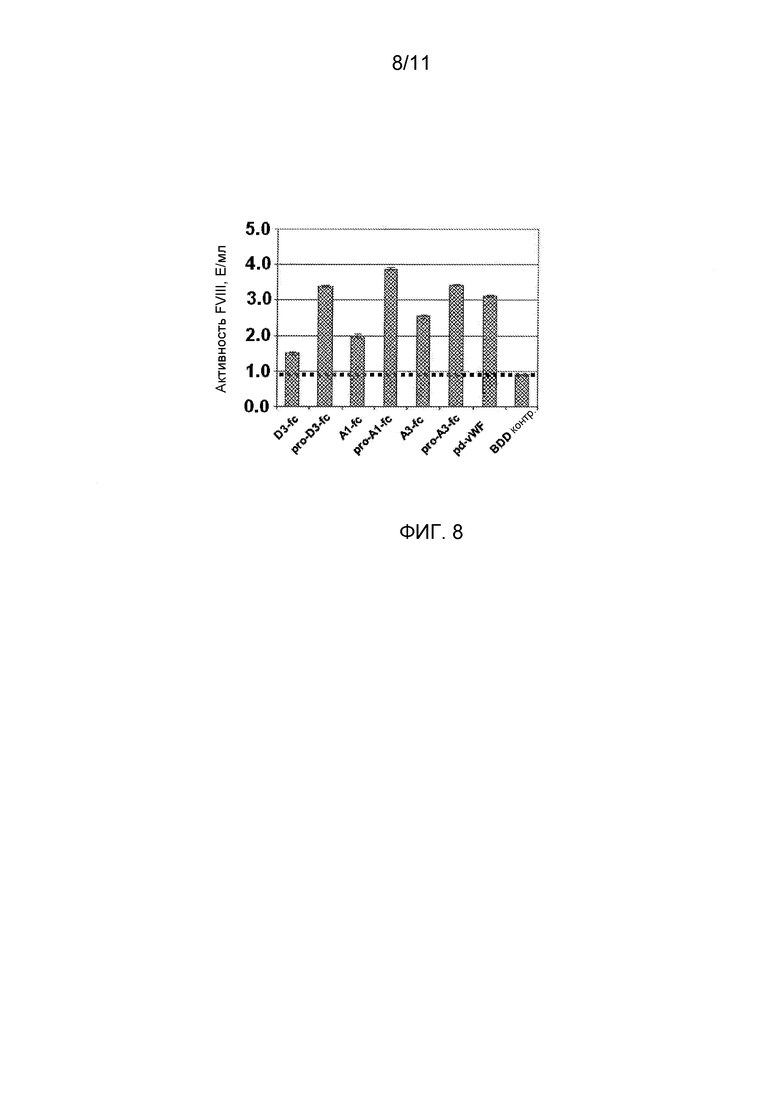

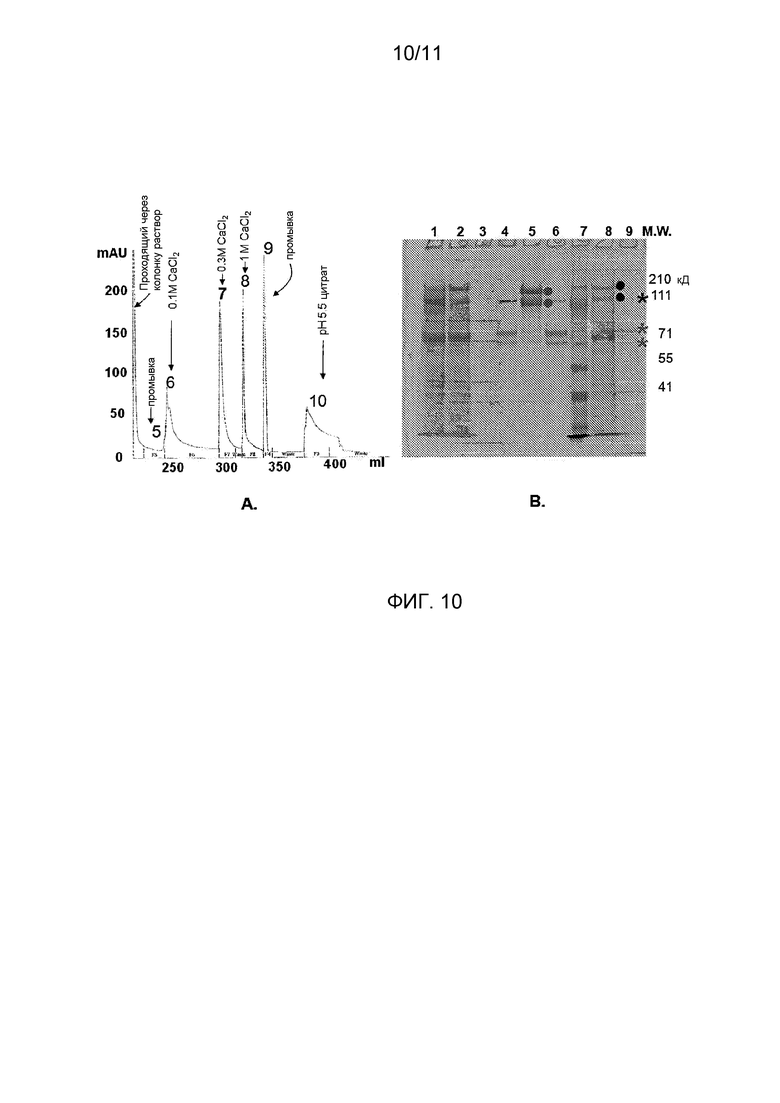
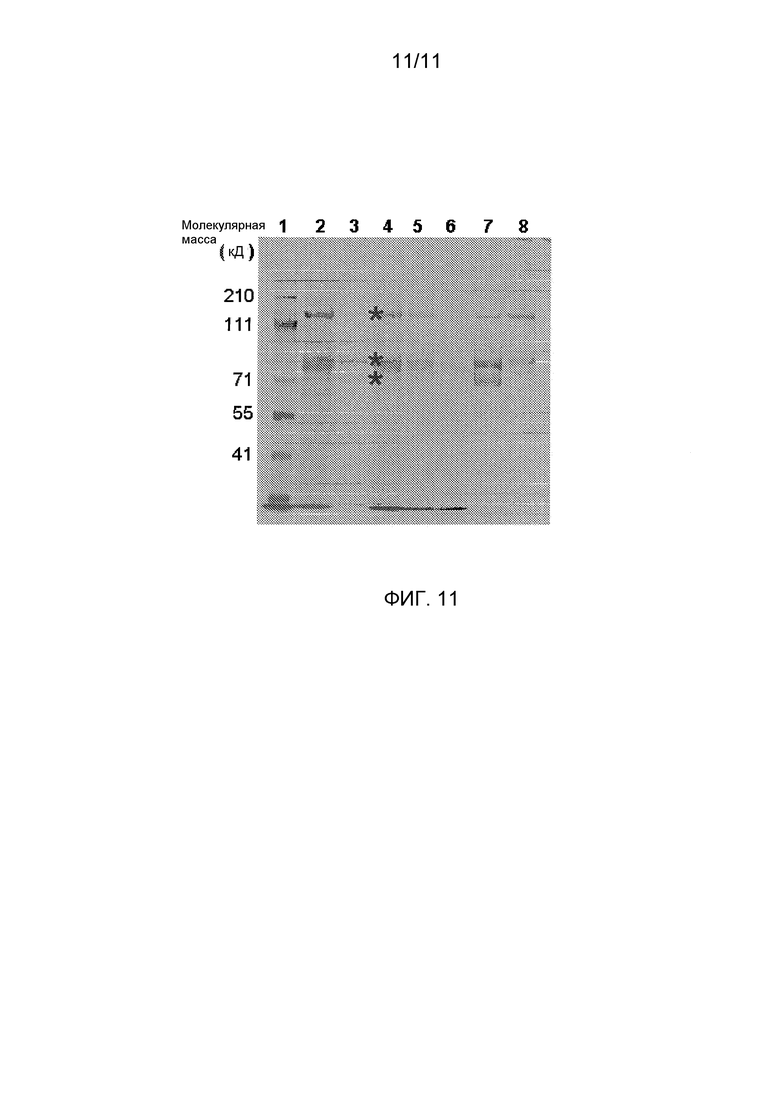









































































































































































Изобретение относится к области биотехнологии, конкретно к рекомбинантному получению модифицированного vWF, и может быть использовано в медицине для лечения гемофилии. Получают укороченные варианты vWF, связанные с Fc-частью иммуноглобулина, способные связывать FVIII. Изобретение позволяет облегчить получение и очистку FVIII, а также способствует пролонгированию полужизни FVIII в крови. 10 н. и 2 з.п. ф-лы, 11 ил., 1 табл., 3 пр.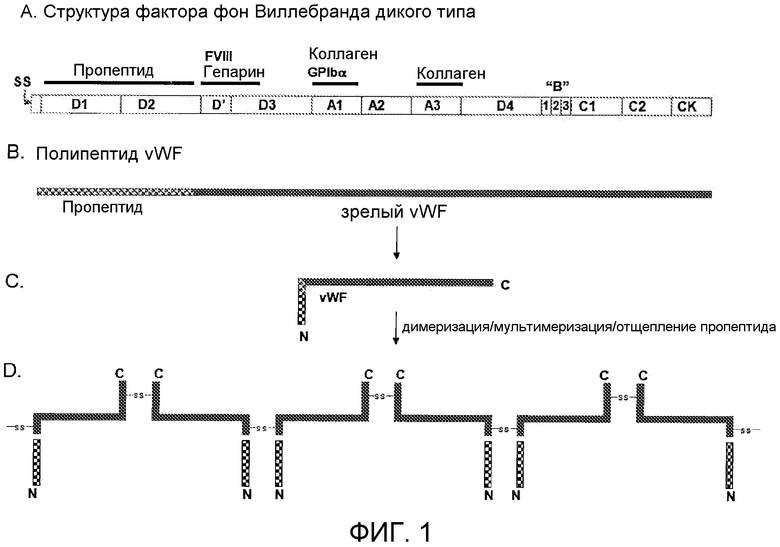
1. Полипептид, связывающий FVIII, где этот полипептид содержит последовательность, представленную в SEQ ID NO: 20, SEQ ID NO:21, SEQ ID NO: 22, SEQ ID NO: 36, SEQ ID NO: 38 или SEQ ID NO: 39.
2. Полипептид по п. 1, где этот полипептид способен образовывать димер.
3. Композиция, связывающая FVIII, содержащая полипептид по п. 1 в эффективном количестве.
4. Белковый комплекс для лечения гемофилии, содержащий полипептид по п. 1 и связанный с FVIII.
5. Фармацевтическая композиция для лечения гемофилии, содержащая белковый комплекс по п. 4 в эффективном количестве и фармацевтически приемлемый носитель.
6. Молекула нуклеиновой кислоты, кодирующая полипептид по п. 1.
7. Молекула нуклеиновой кислоты по п. 6, где эта молекула нуклеиновой кислоты имеет нуклеотидную последовательность представленную в SEQ ID NO: 23, SEQ ID NO: 24, SEQ ID NO: 28, SEQ ID NO: 37, SEQ ID NO: 42 или SEQ ID NO: 43.
8. Экспрессирующий вектор, содержащий молекулу нуклеиновой кислоты по п. 6.
9. Клетка-хозяин, экспрессирующая полипептид по п. 1, содержащая вектор по п. 8.
10. Способ получения белкового комплекса по п. 4, предусматривающий рекомбинантную экспрессию полипептида по п. 1 в клетке, которая экспрессирует FVIII, с их дальнейшим контактированием с образованием комплекса.
11. Способ выделения FVIII, предусматривающий контактирование FVIII с полипептидом по п. 1 для образования белкового комплекса, содержащего этот полипептид и FVIII.
12. Способ лечения гемофилии, предусматривающий введение композиции по п. 5 субъекту, страдающему гемофилией.

