ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
[0001] В данной заявке заявлен приоритет по предварительной заявке на патент США №62/247,469, поданной 28 октября, 2015 года; предварительной заявке на патент США №62/307,897, поданной 14 марта, 2016 года; предварительной заявке на патент США №62/326,229, поданной 22 апреля, 2016 года; предварительной заявке на патент США №62/315,438, поданной 30 марта, 2016 года; и предварительной заявке на патент США №62/355,106, поданной 27 июня 2016 года, раскрытие которых включено в данный документ посредством ссылки в полном объеме.
ОБЛАСТЬ ТЕХНИКИ
[0002] Данное изобретение относится к области генной терапии, в частности целенаправленной доставки конструкций, кодирующих трансген в печень для экспрессии полезных (терапевтических) белков. В частности, изобретение относится к лечению гемофилии, таких как гемофилия А.
УРОВЕНЬ ТЕХНИКИ
[0003] Генная терапия может применяться для генетической инженерии клетки с целью иметь один или более инактивированных генов и/или вызывать в этой клетке экспрессию продукта, который ранее не производился в этой клетке (например, посредством введения трансгена и/или путем коррекции эндогенной последовательности). Примеры применения вставки трансгена включают введение одного или более генов, кодирующих один или более новых терапевтических белков, введение кодирующей последовательности, кодирующей белок, который отсутствует в клетке или в индивидууме, введение гена дикого типа в клетку, которая содержит последовательность мутированного гена и/или введение последовательности, которая кодирует структурную нуклеиновую кислоту, такую как микроРНК или киРНК (siRNA - короткая интерферирующая РНК). Примеры полезных применений "коррекции" эндогенной последовательности гена включают в себя изменения мутаций, связанных с заболеванием, изменения в последовательностях, кодирующих сайты сплайсинга, изменения в регуляторных последовательностях и/или целевые изменения последовательностей, кодирующих структурные характеристики белка.
[0004] Перенос генов в печень обеспечивает эффективное средство доставки трансгенов субъекту для лечения и/или профилактики различных расстройств, включая гемофилии и лизосомальные болезни накопления. См., например, патент США №9,150,847 и публикации США №20130177983 и 20140017212. Также были описаны векторы, специфичные для генной терапии печени. См., например, WO 2014064277; WO 2009130208; ЕР 2451474 В1, Chuah и соавт., (2014 год) Molecular Therapy, 22, 1605-1613; и Nair и соавт. (2014 год) Blood 123:3195-3199. Эти векторы могут включать интронную последовательность мелкого мышиного вируса (MVM) дикого типа. См., например, Haut and Pintel (1998 год) J. Virol. 72:1834-1843; Haut and Pintel (1998 год) Virol. 258:84-94.
[0005] У всех млекопитающих существуют сложные механизмы, которые могут регулировать либо активацию, либо супрессию клеточных элементов иммунной системы. Например, дендритные клетки (ДК) были признаны как центральные игроки в балансе между иммунной активацией и иммунной толерантностью. Они являются наиболее мощными антиген-представляющими клетками в иммунной системе и специфически захватывают, и представляют антигены наивным Т-клеткам. Незрелые ДК взаимодействуют с потенциальными антигенами через специфические рецепторы, такие как Toll-подобные рецепторы, где антиген доставляется в клетку с помощью микропиноцитоза. Затем антиген разрушается на более мелкие пептиды, которые представлены Т-клеткам с помощью основных комплексов гистосовместимости. Кроме того, зрелые ДК секретируют медиаторы воспаления, такие как ИЛ-1β, ИЛ-12, ИЛ-6 и ФНО, которые также служат для активации Т-клеток. С другой стороны, ДК также играют роль в вызывании толерантности организма к некоторым антигенам для поддержания центральной и периферической толерантности. Толерогенные ДК (толДК) имеют низкие количества костимулирующих сигналов на поверхностях клеток и имеют уменьшенную экспрессию медиаторов воспаления, описанных выше. Однако эти толДК экспрессируют большое количество противовоспалительных цитокинов, таких как ИЛ-10, и когда эти клетки взаимодействуют с наивными Т-клетками, Т-клетки становятся анергическими/регуляторными Т-клетками (CD8+ Трег). Фактически, было показано, что этот процесс усиливается при повторной стимуляции Т-клеток этими незрелыми/толерогенными ДК. Было также определено несколько факторов, которые работают совместно с толДК, чтобы индуцировать различные типы Трег. Например, наивные Т-клетки, совместно представленные с толДК и ФРГ (HGF - hepatocyte growth factor, фактор роста гепатоцитов), пептидом VIP, ТСЛП (TSLP - thymic stromal lymphopoietin, тимусный стромальный лимфопоэтин) или витамином D3, приводят к индукции CD4+CD25+Foxp3+Трег, совместное представление с ТРФ-β (TGF - transforming growth factor, трансформирующий ростовой фактор) или ИЛ-10 приводит к Tr1 Трег и совместное представление с кортикостероидами, рапамицином, ретиноевой кислотой может приводить к CD4+/CD8+Трег (Raker и соавт (2015 год) Front Immunol 6:art 569 и Osorio и соавт (2015 год) Front Immunol 6:art 535).
[0006] Гемофилии, такие как гемофилия А и гемофилия В, представляют собой генетические нарушения системы свертывания крови, характеризующиеся кровотечением в суставы и мягкие ткани, а также чрезмерным кровотечением в любое место, где произошла травма или происходит операция. Гемофилия А клинически неотличима от гемофилии В, но фактор VIII (FVIII или F8) недостаточен или отсутствует при гемофилии А, а фактор IX (FIX или F.IX) недостаточен или отсутствует у пациентов с гемофилией В. Ген F8 кодирует гликопротеин плазмы, который циркулирует в сочетании с фактором фон Виллебранда в его неактивной форме. При повреждении поверхности инициируется каскад внутреннего свертывания, и FVIII высвобождается из комплекса и активируется. Активированная форма взаимодействует с фактором IX с целью активировать фактор X, чтобы фактор X стал активированным Ха, что в конечном итоге приводит к изменению фибриногена на фибрин и индукции сгустка. См, Levinson и соавт.(1990 год) Genomics 7(1): 1-11. 40-50% пациентов с гемофилией А имеют хромосомную инверсию, включающую интрон 22 F8 (также известный как IVS22). Инверсия вызвана событием внутрихромосомной рекомбинации между последовательностью 9,6 т.п.н. внутри интрона 22 гена F8 и одной из двух близкородственных последовательностей с обратной ориентацией, расположенных на расстоянии около 300 т.п.н. дистально к гену F8, что приводит к инверсии экзонов 1-22 по отношению к экзонам 23-26. См, Textbook of Hemophilia. Lee и соавт. (изд.) 2005 год, Blackwell Publishing. Другие пациенты с гемофилией А имеют дефекты в F8, включая мутации активного сайта, а также нонсенс-мутация и миссенс-мутации.
[0007] Клинически пациенты с гемофилией А оцениваются и стратифицируются в зависимости от того, насколько часто у пациента происходит эпизод кровотечения и как долго длятся эти эпизоды. Обе эти характеристики напрямую зависят от количества белка FVIII в крови пациента. Пациенты с тяжелой формой гемофилии обычно имеют менее 1% от нормального уровня FVIII в крови, испытывают кровотечение после травмы и часто имеют спонтанное кровотечение в суставах. Пациенты с умеренной формой имеют 1-5% от нормального уровня FVIII, в то время как пациенты с легкой формой имеют 6% или более нормального уровня FVIII и имеют эпизоды кровотечения только после серьезных повреждений, травм или хирургии (Kulkarni и соавт (2009 год) Haemophilia 15:1281-90). Пациенты с гемоАилией А лечатся заменой белка FVIII, полученного либо из плазмы человека, либо продуцируемого рекомбинантно, причем частота лечения основывается на характере кровотечений и тяжести гемофилии. Пациенты с тяжелой гемофилией А регулярно получают профилактическое лечение, чтобы предотвратить кровотечения, в то время как пациенты с менее тяжелой формой могут получать лечение только по мере необходимости после повреждения.
[0008] Описана генная терапия для пациентов с гемофилией А или В, включающая введение плазмиды и других векторов (например, AAV), кодирующих функциональные белки FVIII или F.IX. (См., например, патенты США №6,936,243; 7,238,346 и 6,200,560; Shi и соавт. (2007 год) J Thromb Haemost. (2):352-61; Lee и соавт. (2004 год) Pharm. Res. 7:1229-1232; Graham и соавт. (2008 год) Genet Vaccines Ther. 3:6-9; Manno и соавт. (2003 год) Blood 101(8):2963-72; Manno и соавт. (2006 год) Nature Medicine 12(3):342-7; Nathwani и соавт. (2011 год) Mol Ther 19(5):876-85; Nathwani и соавт. (2011 год); N Engl J Med. 365(25):2357-65 и Mcintosh и соавт. (2013 год) Blood 121(17):3335-44). Однако в этих протоколах образование ингибирующих антител к фактору VIII или IX (антитело к FVIII или к F.IX) и антител к средству доставки является основным осложнением лечения гемофилии на основе замещения FVIII и F.IX. См., например, Scott & Lozier (2012 год) Br J Haematol. 156(3):295-302.
[0009] Однако остается потребность в печень-специфичных полинуклеотидах (экспрессионные конструкции и модули транскрипции), которые приводят к экспрессии одного или более трансгенов (включая трансгены, кодирующие один или более белков, отсутствующих при гемофилии) в клетках печени на высоких уровнях.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
[0010] Данное изобретение описывает композиции и способы экспрессии трансгена в клетке печени. Трансген может быть экспрессирован внехромосомно (эписомально) или может быть интегрирован в геном клетки печени (например, посредством опосредованной нуклеазой целенаправленной интеграции, например, в локус альбумина). В некоторых вариантах реализации изобретения трансген кодирует белок, вовлеченный в каскад свертывания крови. В предпочтительных вариантах реализации изобретения трансген кодирует полипептид FVIII. Композиции и способы, описанные в данном документе, приводят к высоким уровням продуцирования белка как in vitro, так и in vivo, в том числе на уровнях, достаточных для проявления клинически значимых (терапевтических) эффектов in vivo.
[0011] В одном аспекте, описанном в данном документе, представлена полинуклеотидная экспрессионная конструкция, содержащая по меньшей мере одну спейсерную последовательность, содержащую последовательность инсулятора, печень-специфичную последовательность энхансера (например, последовательность энхансера Серпин 1 дикого типа или мутантную), последовательность промотора (например, промотор транстиретина (TTR) дикого типа или мутантный) и трансген (например, нуклеаза и/или терапевтический белок, такой как белок, который отсутствует и/или уровень которого недостаточен при гемофилии или лизосомальной болезни накопления). В некоторых вариантах реализации изобретения экспрессионная полинуклеотидная конструкция дополнительно содержит последовательность интрона (например, последовательность интрона мелкого мышиного вируса (MVM) дикого типа или мутантную). В некоторых вариантах реализации изобретения полинуклеотидная экспрессионная кассета содержит две спейсерные последовательности, фланкирующие печень-специфическую последовательность энхансера, последовательность промотора, последовательность интрона и трансген и, необязательно, сигнал полиаденилирования. Может применяться любая последовательность инсулятора(ов), включая, но без ограничений, любую последовательность инсулятора дикого типа или мутантную (например, одну или более из SEQ ID NO: 28, 29, 30 и/или 38 в любой комбинации). В некоторых вариантах реализации изобретения полинуклеотидная экспрессионная конструкция содержит полинуклеотид, обозначенный CRMSBS1 (SEQ ID NO: 37) или CRMSBS2 (SEQ ID NO: 34).
[0012] В другом аспекте, описанном в данном документе, представлена полинуклеотидная экспрессионная конструкция, содержащая печень-специфичную последовательность энхансера (например, последовательность энхансера Серпин 1 дикого типа или мутантную), последовательность промотора (например, промотор транстиретина (TTR) дикого типа или мутантный), последовательность интрона, как показано в любой из SEQ ID NO: 15, 16 или 17 и трансген (например, нуклеаза и/или терапевтический белок, такой как белок, который отсутствует и/или уровень которого недостаточен при гемофилии или лизосомальной болезни накопления). В некоторых вариантах реализации изобретения полинуклеотидная экспрессионная кассета дополнительно содержит по меньшей мере одну спейсерную последовательность, содержащую последовательность инсулятора и/или сигнал полиаденилирования.
[0013] В еще одном аспекте, описанном в данном документе, представлена полинуклеотидная экспрессионная конструкция, содержащая печень-специфическую последовательность энхансера, имеющую мутации в положениях 1, 5, 14, 32 и/или 39 любой из SEQ ID NO: 1-13 (например, как показано в SEQ ID NO: 35 или 36), последовательность промотора (например, промотор транстиретина (TTR) дикого типа или мутантный) и трансген (например, нуклеаза и/или терапевтический белок, такой как белок, который отсутствует и/или уровень которого недостаточен при гемофилии или лизосомальной болезни накопления). В некоторых вариантах реализации изобретения экспрессионные полинуклеотидные конструкции дополнительно содержат последовательность интрона (например, последовательность интрона мелкого мышиного вируса (MVM) дикого типа или мутантную) и/или по меньшей мере одну спейсерную последовательность, содержащую последовательность инсулятора и/или сигнал полиаденилирования.
[0014] В других аспектах представлен вектор AAV, содержащий любую полинуклеотидную экспрессионную конструкцию, описанную в данном документе (например, в котором полинуклеотидная экспрессионная конструкция находится между 5' и 3' инвертированными концевыми повторами (ИКП) вектора AAV).
[0015] В других аспектах, в данном документе представлены фармацевтические композиции, содержащие один или более векторов AAV и/или одну или более полинуклеотидных экспрессионных конструкций, как описано в данном документе.
[0016] Также представлены способы обеспечения белком субъекта, нуждающегося в этом, при этом способ включает введение в печень субъекта полинуклеотидной экспрессионной конструкции, вектора AAV или фармацевтической композиции, как описано в данном документе, субъекту, при этом трансген кодирует белок и белок продуцируется у субъекта. В некоторых вариантах реализации изобретения трансген интегрируется в геном клетки печени субъекта. Необязательно, способы дополнительно включают введение одной или более нуклеаз субъекту, при этом нуклеаза расщепляет эндогенный ген альбумина и трансген интегрируется в эндогенный ген альбумина. Также предлагаются способы генетической модификации клетки с включением трансгена (например, который продуцирует белок), включая способы введения трансгена в клетку (эписомально или интегрированного с помощью опосредованной нуклеазой направленной интеграции). Также предложены способы индуцирования толерантности у млекопитающего к терапевтическому белку, причем способ включает генетическую модификацию клетки (например, клетки, которая была модифицирована для получения белка, как описано в данном документе) субъекта, как описано в данном документе, и применение одного или более стероидов и/или ингибиторов В-клеток, так что млекопитающее становится толерантным к терапевтическому белку.
[0017] В одном аспекте, описанном в данном документе, представлена полинуклеотидная экспрессионная конструкция, содержащая последовательность энхансера (например, последовательность энхансера Серпин 1 дикого типа или мутантную), последовательность промотора (например, минимальный промотор гена транстиретина (TTRm)) и трансген и, необязательно, последовательность полиаденилирования (например, синтетическую последовательность полиаденилирования (SPA) и/или сигнальный пептид (СП). В некоторых вариантах реализации изобретения экспрессионная конструкция дополнительно содержит последовательность интрона (например, последовательность MVM дикого типа или мутантную последовательность MVM и/или химерный интрон). В некоторых вариантах реализации изобретения экспрессионные конструкции содержат в направлении от 5' к 3', последовательность энхансера, последовательность промотора, последовательность интрона, трансген (необязательно содержащий сигнальный пептид) и сигнал полиаденилирования.
[0018] Экспрессионная кассета может быть включена в любой вирусный или невирусный вектор, включая, но без ограничений, плазмидные векторы, аденовирусный вектор, ретровирусные векторы и вектор на основе аденоассоциированного вируса (AAV). В предпочтительном варианте реализации изобретения экспрессионная конструкция переносится на конструкцию AAV и дополнительно содержит 5' и 3' ИКП, фланкирующие экспрессионные конструкции, как описано в данном документе. Необязательно, молекулы инсулятора (спейсера) также включены между одним или более компонентами экспрессионной конструкции, например, между 5'-ИКП и энхансером и/или между сигналом полиаденилирования и 3'-ИКП. В некоторых вариантах реализации изобретения области инсулятора (спейсера) содержат гомологические плечи для облегчения направленной интеграции. В некоторых вариантах реализации изобретения конструкция представляет собой конструкцию, как изображено на любой из Фиг. 1, 2, 4, 6, 7, 10, 19, 20 или 25. Две типовые конструкции показаны в SEQ ID NO: 34 и в SEQ ID NO: 37. Очевидно, что отдельные компоненты (промотор, изолятор(ы), энхансер, трансген) могут быть объединены в любой комбинации с другими компонентами, как описано в данном документе.
[0019] В любом из описанных в данном документе полинуклеотидов энхансер может быть получен из энхансера Серпин-1. В некоторых вариантах реализации изобретения энхансер представляет собой последовательность дикого типа. В других вариантах реализации изобретения энхансер содержит одну или более модификаций по сравнению с диким типом, например, энхансер Серпин1 содержит одну или более нуклеотидных модификаций, как изображено на Фиг. 5, модификации нуклеотидов в одном или более остатках 1, 5, 14, 32 и/или 39 последовательности, показанной в любой из SEQ ID NO: 1-13. В некоторых вариантах реализации изобретения энхансер Серпин1 содержит модификации в положениях 1,5, 14 и 32 любой из SEQ ID NO: 1-13, в то время как в других вариантах реализации изобретения энхансер Серпин1 содержит модификацию в положениях 1, 14, 32 и 39 любой из SEQ ID NO: 1-13. Типовые последовательности энхансера показаны в SEQ ID NO: 35 и 36. В некоторых вариантах реализации изобретения полинуклеотиды, описанные в данном документе, содержат 1, 2, 3, 4, 5 или более энхансерных элементов. В некоторых вариантах реализации изобретения 1, 2, 3, 4, 5 или более энхансерных элементов идентичны, причем в других вариантах реализации изобретения применяется более одного типа энхансера.
[0020] Любой из описанных в данном документе полинуклеотидов может дополнительно необязательно содержать интронную последовательность. В некоторых вариантах реализации изобретения экспрессионная конструкция включает химерную последовательность интрона, например, как изображено на нижних панелях Фиг. 6 и 7. Т-химерный интрон представляет собой усеченную версию химерного интрона в pCI-neo (GenBank U47120). Химерный интрон в pCI-neo представляет собой 5' сайт донора сплайсинга из гена β-глобина человека, а также точку разветвления и 3' акцепторный сайт вариабельной области тяжелой цепи гена иммуноглобулина. Т-химерный интрон содержит делецию 45 п.н. между 5' донором сплайсинга и точкой разветвления. В других вариантах реализации изобретения экспрессионные конструкции включают мутантную последовательность интрона MVM (например, одну или более мутаций, изображенных на Фиг. 12, 13 и 14 (SEQ ID NO: 15-17). Альтернативно, экспрессионные конструкции, как описано в данном документе, могут не иметь интронной последовательности, например, как показано в конструкциях, изображенных на средних панелях Фиг. 6, 7, 19 и 20.
[0021] Экспрессионные конструкции по изобретению также могут включать оптимизированные последовательности инсулятора между ИКП AAV и экспрессионной кассетой. В определенных вариантах реализации изобретения экспрессионная конструкция содержит инсуляторные (спейсерные) области (например, Инс1 и Инс3, SEQ ID NO: 15 или 28 и SEQ ID NO: 17 или 30, (соответственно), а также Инс2 (SEQ ID NO: 16 или 29). В определенных вариантах реализации изобретения экспрессионная конструкция содержит Инс4 (SEQ ID NO: 38). Любая из последовательностей инсулятора может применяться как в 5' так и в 3' положении, и любая комбинация последовательностей инсулятора может применяться в экспрессионной конструкции. Особенно предпочтительными являются комбинации Инс1 в положении 5' и Инс3 в положении 3'.
[0022] Экспрессионные конструкции, как описано в данном документе, также содержат и экспрессируют один или более трансгенов. Любой трансген(ы) может быть экспрессирован с применением описанных в данном документе полинуклеотидов, включая, но без ограничений, трансгены, кодирующие функциональные варианты белков, которые отсутствуют или присутствуют в недостаточных количествах при любом генетическом заболевании, включая, но без ограничений, лизосомальные болезни накопления (например, болезнь Гоше, Фабри, Хантера, Гурлера, Ниманна-Пика, Фенилкетонурию (ФКУ) и тому подобные), болезни нарушения обмена веществ и/или заболевания крови, такие как гемофилии и гемоглобинопатии и тому подобное. См., например, Публикация США №20140017212 и 20140093913; патенты США №9,255,250 и 9,175,280. Неограничивающие примеры белков, которые могут быть экспрессированы, как описано в данном документе, включают фибриноген, протромбин, тканевой фактор, фактор V, фактор VII, фактор VIII, фактор IX, фактор X, фактор XI, фактор XII (фактор Хагемана), фактор XIII (фибринстабилизирующий фактор), фактор Виллебранда, прекалликреин, высокомолекулярный кининоген (фактор Фицджеральда), фибронектин, антитромбин III, кофактор II гепарина, белок С, белок S, белок Z, белок Z-подобного ингибитора протеазы, плазминоген, альфа 2-антиплазмин, тканевой активатор плазминогена, урокиназу, ингибитор активатора плазминогена-1, ингибитор активатора плазминогена-2, глюкоцереброзидазу (GBA), α-галактозидазу А (ГЛА), идуронатсульфатазу (НДС), идуронидазу (IDUA), кислую сфингомиелиназу (SMPD1), ММАА, ММАВ, ММАСНС, MMADHC (C2orf25), MTRR, LMBRD1, MTR, пропионил-СоА-карбоксилазу (РСС) (субъединицы РССА и/или РССВ), белок транспортера глюкозо-6-фосфатна (G6PT) или глюкозо-6-фосфатазу (Г6Фазу), рецептор ЛНП (РЛНП), АроВ, LDLRAP-1, PCSK9, митохондриальный белок, такой как NAGS (N-ацетилглутамат синтетазу), CPS1 (карбамоилфосфат синтетазу I) и ОТС (орнитинтранскарбамилазу), ASS (аргининосукцинат-синтетазу), ASL (аргининосукцинатлиазу) и/или ARG1 (аргиназу), и/или белок семейства транспортеров растворенных веществ типа 25 (SLC25A13, переносчик аспартата/глутамата), полипептид А1 глюкуронилтрансферазы UGT1A1 или UDP, фумарилацетоацетатгидролиазу (FAH), белок аланин-глиоксилат-аминотрансферазы (AGXT), белок глиоксилатредуктазы/гидроксипируватредуктазы (GRHPR), белок гена транстиретина (TTR), белок АТР7 В, белок фенилаланингидроксилазы (ФАГ), белок липопротеинлиазы (ЛПЛ), сконструированную нуклеазу, сконструированный транскрипционный фактор и/или сконструированный одноцепочечный вариабельный фрагмент антитела (диатело, верблюжье антитело и тому подобное). В одном предпочтительном варианте реализации изобретения трансген кодирует полипептид FVIII. В некоторых вариантах реализации изобретения полипептид FVIII содержит делецию домена В.
[0023] В некоторых вариантах реализации изобретения один или более трансгенов включают последовательности, кодирующие сконструированные нуклеазы (например, ZFN (нуклеазы с цинковыми пальцами), TALEN (эффекторные нуклеазы, подобные активаторам транскрипции), TtAgo и системы CRISPR/Cas). В других вариантах реализации изобретения трансгены включают последовательности, кодирующие сконструированные факторы транскрипции (например, ZFP-TF (факторы транскрипции белков "цинковые пальцы"), TALE-TFs, системы CRISPR/Cas-TF). Трансгены могут также включать последовательности, кодирующие одноцепочечное антитело, специфичное к мишени интереса. Кроме того, трансген может включать последовательности, кодирующие структурную РНК (например, РНКи, shPHК (малые шпилечные РНК), миРНК (микроРНК)).
[0024] В определенных аспектах полинуклеотиды, как описано в данном документе, вводят в клетку таким образом, что они поддерживаются эписомально во время обеспечения экспрессии трансгена. В других аспектах экспрессионная конструкции случайным образом интегрируются в геном клетки, в которую они вводятся. В дополнительных аспектах экспрессионные конструкции, обеспечивающие экспрессию трансгена, интегрируются в геном посредством опосредованной нуклеазой целенаправленной интеграции.
[0025] В дополнительных аспектах, в данном документе описаны способы экспрессии одного или более трансгенов в клетке печени, причем способы включают введение одной или более экспрессионных конструкций, как описано в данном документе, в клетку, так что трансген экспрессируется в клетке. В некоторых вариантах реализации изобретения экспрессионная конструкция переносится вирусным или невирусным вектором, предпочтительно вектором AAV (например, AAV2 или AAV2/6).
[0026] В другом аспекте, в данном документе предложен способ экспрессии одного или более трансгенов в живом животном, причем способы включают введение одной или более экспрессионных кассет, как описано в данном документе, живому животному. В некоторых вариантах реализации изобретения экспрессионные кассеты вводят в печень животного. В некоторых вариантах реализации изобретения экспрессионная конструкция переносится вирусным или невирусным вектором, предпочтительно вектором AAV (например, AAV2, AAV2/6 или AAV2/8).
[0027] В некоторых вариантах реализации изобретения, в данном документе предложены способы и композиции для экспрессии терапевтически значимых уровней одного или более терапевтических белков одного или более трансгенов. В некоторых вариантах реализации изобретения экспрессия трансгенной конструкции, кодирующей замещающий белок, приводит к 1% от нормальных уровней продуцируемого белка, тогда как в других к 2%, 3%, 4%, 5%, 10%, 15%, 20%, 30%, 50%, 80%, 100%, 150%, 200% или более от нормальных уровней продуцируемого белка. В некоторых предпочтительных вариантах реализации изобретения трансген кодирует белок FVIII и продуцируется терапевтически значимое количество белка. В некоторых вариантах реализации изобретения в результате применения способов и композиций по изобретению, у пациента-человека в крови увеличивается количество терапевтического белка, что приводит к уменьшению клинических симптомов. В некоторых аспектах продуцирование терапевтического белка способами и композициями, описанными в данном документе у пациента-человека, приводит к уменьшению времени свертывания крови после повреждения по сравнению с пациентом, который не получал лечения или по сравнению с пациентом до лечения. В некоторых аспектах пациент-человек, лечившийся способами и композициями по изобретению, требует уменьшенного количества заместительной терапии, чем пациент, который не лечился или по сравнению с пациентом до лечения.
[0028] В некоторых вариантах реализации изобретения способ и композиции по изобретению, как описано в данном документе, могут быть применены для индуцирования толерантности у млекопитающего к терапевтическому белку, таким образом, что уровни терапевтического белка, кодируемого трансгеном, остаются на терапевтически значимых уровнях после кратковременного повышения количества антител к терапевтическим белкам. Таким образом, в данном документе представлен способ индуцирования толерантности к терапевтическому белку у субъекта, причем способ включает генетическую модификацию клетки у субъекта с использованием способа, описанного в данном документе (например, таким образом что клетка продуцирует терапевтический белок), необязательно лечением субъекта дополнительными композициями (например, стероидами и/или ингибиторами В-клеток), таким образом, что животное становится толерантным к терапевтическому белку. В некоторых вариантах реализации изобретения введение (интеграция) терапевтического белка в клетки-реципиенты проводят одновременно с лечением иммуноингибирующим стероидом или ингибитором В-клеток, тогда как в других случаях иммуномодулирующие вещества не вводят животному. В некоторых случаях иммуномодулирующий агент вводят только в том случае, если вырабатываются антитела к терапевтическому белку. В дополнительных случаях введение иммуномодулирующего агента прекращают через некоторое время.
[0029] В другом аспекте предлагаются фармацевтические композиции, содержащие одну или более клеток, экспрессионные конструкции и/или, необязательно, нуклеазы, описанные в данном документе.
[0030] В некоторых аспектах, в данном документе описаны способы и системы для направленной интеграции печень-специфичной экспрессионной кассеты. Способы и системы включают введение одной или более экспрессионных кассет, как описано в данном документе, и введение одной или более нуклеаз, специфичных к целевому гену, в клетку. После опосредованного нуклеазой расщепления целевого гена экспрессионная кассета интегрируется в ген с помощью механизмов, зависящих или не зависящих от гомологической рекомбинации. В некоторых вариантах реализации изобретения целевой ген представляет собой эндогенный ген альбумина.
[0031] Для опосредованной нуклеазой направленной интеграции экспрессионных конструкций по данному изобретению может быть применена любая нуклеаза, включая, но без ограничений, одну или более нуклеаз с цинковыми пальцами (ZFN), TALEN, нуклеаз CRISPR/Cas и/или нуклеаз TtAgo, так что экспрессионная конструкция интегрируется в область (ген), расщепленную нуклеазой(ами). В определенных вариантах реализации изобретения используют одну или более пар нуклеаз. Нуклеазы могут быть введены в форме мРНК или могут быть введены в клетку с использованием невирусных или вирусных векторов. В некоторых аспектах полинуклеотиды нуклеаз могут быть доставлены с помощью лентивируса или неинтегрирующего лентивируса. В других экспрессионная кассета может быть доставлена с помощью AAV и/или олигонуклеотидами ДНК.
[0032] В другом аспекте, в данном документе представлены способы для обеспечения одного или более функциональных белков, которые отсутствуют или присутствуют в недостаточных количествах у млекопитающего, или у примата, такого как человекоподобный примат, такого как пациента-человека с заболеванием (например, метаболическое заболевание, лизосомальная болезнь накопления (LSD), гемоглобинопатия и/или гемофилия), например, для лечения заболевания путем обеспечения белком(ами), которые отсутствуют или присутствуют в недостаточных количествах у субъекта. В другом аспекте, в данном документе представлены способы обеспечения функционального белка для лечения расстройства, при котором белок отсутствует, присутствуют в недостаточных количествах или аберрантно экспрессируется. В дополнительных вариантах реализации изобретения способы включают введение экспрессионной кассеты, кодирующей терапевтический белок, полезный для профилактики или лечения расстройства. В дополнительном аспекте в данном документе описаны способы для обеспечения терапевтического белка с целью лечения расстройства, в которых терапевтический белок представляет собой одноцепочечное антитело. В некоторых вариантах реализации изобретения способы включают введение экспрессионной кассеты (например, вектор AAV), как описано в данном документе, в печень субъекта, нуждающегося в этом. В других вариантах реализации изобретения способ включает введение модифицированной клетки (экспрессирующей функциональную версию белка, которая аберрантно экспрессируется у субъекта, из экспрессионной кассеты, как описано) субъекту. Таким образом, выделенная клетка может быть введена субъекту (клеточная терапия ex vivo) или клетка может быть модифицирована, в случае если она является частью субъекта (in vivo).
[0033] В любой из описанных композиций и способов экспрессионные кассеты и/или нуклеазы могут переноситься вектором AAV, включая, но без ограничений, AAV1, AAV3, AAV4, AAV5, AAV6, AAV8, AAV9 и AAVrh10 или псевдотипированным AAV, таким как AAV2/8, AAV8. 2, AAV2/5 и AAV2/6 и тому подобное. В определенных вариантах реализации изобретения полинуклеотиды (экспрессионные конструкции и/или нуклеазы) доставляются с применением тех же типов векторов AAV. В других вариантах реализации изобретения полинуклеотиды доставляются с применением различных типов векторов AAV. Полинуклеотиды могут быть доставлены с применением одного или более векторов. В некоторых вариантах реализации изобретения полинуклеотиды доставляются путем внутривенного (например, через портальную вену) введения в печень интактного животного. В других вариантах реализации изобретения полинуклеотиды доставляются путем внутривенного введения в периферическую вену.
[0034] В любой из описанных в данном документе композиций и способов белок, кодируемый трансгеном, может содержать белок F8, например, фактор VIII с удаленным доменом В (BDD-F8). В других вариантах реализации изобретения белок, кодируемый трансгеном, содержит белок фактора IX. В других вариантах реализации изобретения белок, кодируемый трансгеном, содержит белок фактора VII. В других вариантах реализации изобретения белок, кодируемый трансгеном, содержит белок фактора X. В некоторых вариантах реализации изобретения белок, кодируемый трансгеном, содержит глюкоцереброзидазу. В других вариантах реализации изобретения белок, кодируемый трансгеном, содержит α-галактозидазу. В дополнительных вариантах реализации изобретения белок, кодируемый трансгеном, содержит идуронат-2-сульфатазу. В некоторых вариантах реализации изобретения белок, кодируемый трансгеном, содержит альфа-L-идуронидазу. В дополнительных вариантах реализации изобретения белок, кодируемый трансгеном, содержит сфингомиелинфосфодиэстеразу. В некоторых вариантах реализации изобретения трансген кодирует одноцепочечное антитело. В других вариантах реализации изобретения трансген кодирует структурную РНК. В любой из описанных в данном документе композиций или способов трансген также содержит регулятор транскрипции, а в других - не содержит, а транскрипция регулируется эндогенным регулятором. В другом аспекте способы по изобретению включают композицию для терапевтического лечения субъекта, нуждающегося в этом. В некоторых вариантах реализации изобретения композиция содержит сконструированные стволовые клетки, содержащие специфическую нуклеазу, безопасную для своих генов, и кодируемый трансгеном фактор VII, F8, F.IX, фактор X, GBA, GLA, IDS, IDUA, одноцепочечное антитело и/или белок SMPD1, или функциональный фрагмент и/или их усечение. В других вариантах реализации изобретения композиция содержит сконструированные стволовые клетки, которые были модифицированы и экспрессируют трансген, кодирующий фактор VII, F8, F.IX, фактор X, GBA, GLA, IDS, IDUA, одноцепочечное антитело и/или белок SMPD1, или функциональный фрагмент и/или их усечение.
[0035] Способы, описанные в данном документе, могут быть осуществлены на практике in vitro, ex vivo или in vivo. В определенных вариантах реализации изобретения композиции вводят в живое, интактное млекопитающее. Млекопитающее может находиться на любой стадии развития во время доставки, например, зачаточной, эмбриональной, неонатальной, ранней, ювенильной или взрослой. Дополнительно, целевые клетки могут быть здоровыми или больными. В определенных вариантах реализации изобретения одна или более композиций доставляются внутривенно (например, в печень через портальную вену, например инъекцией в хвостовую вену), внутриартериально, внутрибрюшинно, внутримышечно, в паренхиму печени (например, инъекцией), в печеночную артерию (например, инъекцией) и/или через желчные протоки (например, инъекцией).
[0036] Для нацеливания композиций на конкретный тип клеток, например, тромбоциты, фибробласты, гепатоциты и тому подобное, одна или более введенных композиций могут быть связаны с агентом "хоуминга", который специфически связывается с поверхностным рецептором клетки. Например, вектор может быть конъюгирован с лигандом (например, галактозой) к которому определенные клетки системы печени имеют рецепторы. Конъюгация может быть ковалентной, например, сшивающий агент, такой как глутаральдегид, или нековалентной, например, связывание авидинированного лиганда с биотинилированным вектором. Другая форма ковалентной конъюгации обеспечивается путем конструирования плазмиды-помощника AAV, применяемой для получения основного количества вектора, так что один или более кодированных белков оболочки представляют собой гибрид нативного белка оболочки AAV и пептидного или белкового лиганда, так что лиганд выставлен на поверхности вирусной частицы.
[0037] Также предлагается набор, содержащий одну или более экспрессирующих конструкций, векторов AAV, клеточных и/или фармацевтических композиций, описанных в данном документе. Набор может дополнительно содержать нуклеиновые кислоты, кодирующие нуклеазы (например, молекулы РНК, кодирующие ZFN, TALEN или Cas и модифицированные белки Cas, и гидовые РНК), или аликвоты нуклеазных белков, клетки, инструкции для осуществления способов по изобретению и тому подобное.
[0038] Эти и другие аспекты будут очевидны для квалифицированного специалиста в контексте изложенной в данном документе информации в целом.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
[0039] На Фиг. 1 изображена схема, показывающая элементы кассеты трансгена, а также шаги, предпринятые для идентификации улучшенных энхансеров и интронов.
[0040] На Фиг. 2 изображена схема, показывающая исходный промотор TTRm и конструкции HLP с инсуляторами 1-3 ("Инс1-3"). СерпЭ относится к энхансеру Серпина из гена SERPINA1, который является регуляторным элементом Серпина, специфичным для печени. TTRm относится к минимальному промотору гена транстиретина. HLP относится к гибридному печень-специфичному промотору (Mcintosh и соавт., из того же источника). чFVIII относится к трансгену фактора VIII человека с удаленным В-доменом. СП относится к сигнальному пептиду. ИКП относится к инвертированному концевому повтору. SPA относится к синтетической последовательности полиаденилирования.
[0041] На Фиг. 3 изображен график, показывающий выход различных конструкций AAV, содержащих различные последовательности инсуляторов или промоторные области. На фигуре изображены улучшенные выходы от Инс1-Инс3 по сравнению с Инс1-Инс2 в контексте исходного промотора TTRm или конструкций HLP. Вирус продуцировался в клетках HEK293, выход из двух клеточных фабрик (2 КФ). СерпЭ относится к энхансеру Серпина из гена SERPINA1, который является регуляторным элементом Серпина, специфичным для печени. TTRm относится к минимальному промотору гена транстиретина. HLP относится к гибридному печень-специфичному промотору (Mcintosh и соавт., из того же источника). чFVIII относится к трансгену фактора VIII человека с удаленным В-доменом. СП относится к сигнальному пептиду. ИКП относится к инвертированному концевому повтору. SPA относится к синтетической последовательности полиаденилирования.
[0042] На Фиг. 4 схематически изображена исходная конструкция, и конструкции с новыми энхансерами, обозначенными CRMSBS1 и CRMSBS2, каждый из которых несет трансген фактора VIII человека с удаленным (Фактор VIII-BDD) В-доменом. "СерпЭ" относится к энхансеру Серпина из гена SERPINA1, который является регуляторным элементом Серпина, специфичным для печени. "TTRm" относится к минимальному промотору гена транстиретина. "SBS" относится к внутренней числовой ссылке Sangamo Biosciences. "чFVIII" относится к трансгену человеческого фактора VIII BDD. "СП" относится к сигнальному пептиду. ИКП относится к инвертированному концевому повтору. "SPA" относится к синтетической последовательности полиаденилирования. "Инс1" и "Инс3" являются такими, как изображено выше на Фиг. 2.
[0043] На Фиг. 5 (SEQ ID NO: 1-13) изображено выравнивание гена SERPINA1 от нескольких видов, с использованием выравнивания ENCODE, нескольких видов гена SERPINA1. Идентичные остатки не показаны (обозначаются знаком "."). Области в рамках, обозначенные 1-5, представляют собой сайты модификации последовательности. Также области чувствительности к ДНКазе I в клетках HepG2 показаны черными полосками в средней панели. CRMSBS1 включает изменения в положениях 1, 2, 3 и 4 в рамке, а CRMSBS2 включает изменения в положениях 1, 3, 4 и 5 в рамке. "CRM" относится к цис-регулирующему модулю. "SBS1/2" относится к внутренним числовым ссылкам Sangamo Biosciences для конструкций 1 и 2. Светло-серая область в рамке представляет собой последовательности гена SERPINA 1, которые были пропущены.
[0044] На Фиг. 6 схематически изображены CRMSBS1 и 2 конструкции, полученные из CRMSBS1 без интрона MVM (CRMSBS1 Без Интрона), или усеченный химерный интрон (CRMSBS1 Т-Химерный Интрон), каждая конструкция включает трансген фактора VIII BDD. "CRM" относится к цис-регулирующему элементу. "SBS" относится к внутренней числовой ссылке Sangamo Biosciences. "чFVIII" относится к человеческому фактору VIII-BDD. "СП" относится к сигнальному пептиду. ИКП относится к инвертированному концевому повтору. "SPA" относится к синтетической последовательности полиаденилирования. "Инс1" и "Инс3" являются такими, как изображено выше на Фиг. 2.
[0045] На Фиг. 7 схематически изображены CRMSBS2 и 2 конструкции, полученные из CRMSBS2 без интрона MVM (CRMSBS2 Без Интрона), или усеченный химерный интрон (CRMSBS2 Т-Химерный Интрон), каждая конструкция включает трансген фактора VIII BDD. "CRM" относится к цис-регулирующему элементу. "SBS" относится к внутренней числовой ссылке Sangamo Biosciences. "чFVIII" относится к фактору VIII BDD человека. "СП" относится к сигнальному пептиду. ИКП относится к инвертированному концевому повтору. "SPA" относится к синтетической последовательности полиаденилирования. "Инс1" и "Инс3" являются такими, как изображено выше на Фиг. 2.
[0046] На Фиг. 8А и 8В изображены графики, показывающие in vivo продуцирование секретируемого фактора VIII BDD человека после введения указанных конструкций мышам. Мышей C57BL/6 трансдуцировали 6Е+12 гв/кг (геномов вируса на кг) конструкций AAV2/6 чFVIII-BDD. На Фиг. 8А изображены уровни секретируемого чFVIII-BDD на 7-й день после введения, а на Фиг. 8В изображены уровни секретируемого 4FVIII-BDD в течение 28-дневного исследования. Левая ось "у" каждого графика показывает нг/мл, а правая ось "у" показывает процент от нормы, где 1 Ед = 100% от нормы = 200 нг/мл. *** обозначает р >. 001, * обозначает р >. 05. Каждая индивидуальная форма в пределах группирований представляет результаты от одного животного.
[0047] На Фиг. 9А и 9В изображена печень-специфичная экспрессия кДНК F8-BDD, после введения векторов, как описано на Фиг. 7. На Фиг. 9А изображены уровни мРНК фактора VIII-BDD (чFVIII) человека, которые анализируются из указанных тканей (головного мозга, сердца, почек, легких, печени, селезенки, яичек). Как показано, мРНК чFVIII-BDD была обнаружена только в печени и не обнаружена в других тканях как из исходной кДНК конструкции F8-BDD (незаштрихованные обозначения) так из CRMSBS2 без интрона (заштрихованные обозначения). На Фиг. 9В изображено биораспределение генома вектора (ГВ) из тех же тканей, изображенных на Фиг. 9А. Серотипом AAV2/6 преимущественно трансдуцировали печень.
[0048] На Фиг. 10 схематично изображена конструкция CRMSBS2, и дополнительные конструкции (обозначенные CRMSBS2 SBR Интроны 1, 2 или 3). Все конструкции включают в себя трансген фактора VIII BDD человека. "CRM" относится к цис-регулирующему элементу. "SBS" относится к внутренней числовой ссылке Sangamo Biosciences. "чFVIII" относится к трансгену человеческого фактора VIII-BDD. "СП" относится к сигнальному пептиду. ИКП относится к инвертированному концевому повтору. "SPA" относится к синтетической последовательности полиаденилирования. "MVM" относится к последовательности интрона MVM.
[0049] На Фиг. 11 схематично изображен интрон мелкого мышиного вируса (MVM) дикого типа ("ДТ MVM"), включая частичную последовательность (SEQ ID NO: 14) и расположение доноров и акцепторов." Д1" и" Д2" относятся к донорам 1 и 2 соответственно. "А1" и "А2" относятся к акцепторам 1 и 2 соответственно. "IES" относится к последовательности интронного энхансера. См, также, Haut and Pintel (1998 год) J. Virology 72:1834-1843 и Haut and Pintel (1998 год) Virology J., 258:84-94.
[0050] На Фиг. 12 (SEQ ID NO: 15) изображен обзор изменений в интроне MVM дикого типа в SBR Интрон 1, который применяется в конструкциях, описанных в данном документе. Изменения отмечены серым цветом и включают: незначительную мутацию для потенциального акцептора точки разветвления 1 (РВРА1); мутированный акцепторный сайт 1 полипиримидинового тракта, А1РРТ(-); мутированный акцепторный сайт 1, А1(-); улучшенный потенциальный акцепторный сайт точки разветвления 2, РВРА2(+); усиленный акцепторный сайт 2 полипиримидинового тракта, А2РРТ (+, 8Т) путем добавления большего количества тимидинов (Т); Улучшенный акцепторный сайт 2, А2(+).
[0051] На Фиг. 13 (SEQ ID NO: 16) изображен обзор изменений в интроне MVM дикого типа в SBR Интрон 2, который применяется в конструкциях, как описано в данном документе. Изменения отмечены серым цветом и включают: незначительную мутацию для потенциального акцептора точки разветвления 1 (РВРА1); мутированный акцепторный сайт 1 полипиримидинового тракта, А1РРТ(-); мутированный акцепторный сайт 1, А1(-); усиленный акцепторный сайт 2 полипиримидинового тракта, А2РРТ (+, 8Т) путем добавления большего количества тимидинов (Т); Улучшенный акцепторный сайт 2, А2(+).
[0052] На Фиг. 14 (SEQ ID NO: 17) изображен обзор изменений в интроне MVM дикого типа в SBR Интрон 3, который применяется в конструкциях, как описано в данном документе. Изменения отмечены серым цветом и включают: мутацию богатой на G последовательности интронного энхансера, нокаут IES (IES ko); незначительную мутацию для потенциального акцептора точки разветвления 1 (РВРА1); мутированный акцепторный сайт 1 полипиримидинового тракта, А1РРТ(-); мутированный акцепторный сайт 1, А1(-); усиленный акцепторный сайт 2 полипиримидинового тракта, А2РРТ (+, 9Т) путем добавления большего количества тимидинов (Т); Улучшенный акцепторный сайт 2, А2(+).
[0053] На Фиг. 15А и 15В изображено in vivo продуцирование секретируемого фактора VIII человека с удаленным В-доменом у мышей с новыми последовательностями интрона. На Фиг. 15 изображены графики, показывающие уровни секретируемого фактора VIII человека с удаленным В-доменом у мышей, представленные как либо нг/мл (левая ось "у"), либо процент от нормы (правая ось "у"), 1 Ед = 100% нормальности = 200 нг/мл. Мышей C57BL/6 трансдуцировали 6Е+12 гв/кг конструкций AAV2/6 чFVIII, как обозначено на оси "х". На Фиг. 15А изображены уровни секретируемого чFVIII на 7-й день. На Фиг. 15В изображены пиковые уровни секретируемого чFVIII в течение 28-дневного исследования. Числа обозначают средний процент от нормальных уровней чFVIII для группы.
[0054] На Фиг. 16А и 16В изображено, что экспрессия кДНК FVIII-BDD специфична для печени. На Фиг. 16А изображен график, показывающий значения мРНК, кодирующей человеческий фактор VIII (чFVIII), которые анализировали из тканей (головного мозга, сердца, почек, легких, печени, селезенки, яичек) в исследовании, представленном на Фиг. 15. мРНК чFVIII была обнаружена только в печени и не обнаружена в других тканях, как из исходной кДНК конструкции F8 (незаштрихованные обозначения), так из CRMSBS2 SBR Интрон 3 (заштрихованные обозначения). На Фиг. 16В изображено биораспределение генома вектора (ГВ) из тех же тканей, проанализированных на Фиг. 16А. Серотипом AAV2/6 преимущественно трансдуцировали печень, как было опубликовано ранее.
[0055] На Фиг. 17А и 17В изображены супрафизиологические уровни ферментативно активного фактора VIII-BDD человека в плазме мышей, страдающих гемофилей А. На Фиг. 17 изображены графики, показывающие уровни ферментативно активного секретируемого человеческого фактора VIII (чFVIII) с удаленным В-доменом у мышей R593C, страдающих гемофилией А, представленные в процентах от нормы, 1 Ед = 100%. Нокаутные мыши R593C, страдающие гемофилией А, были трансдуцированы ~ 7Е+12 гв/кг конструкции AAV2/6 чFVIII-BDD, как показано на оси "х". На Фиг. 17А изображены уровни активности секретируемого чFVIII на 14-й день. На Фиг. 17В изображены уровни секретируемого чFVIII на 42-й день. Числа обозначают средний процент от нормальных уровней чFVIII для группы.
[0056] На фигуре 18 изображен график, показывающий время кровотечения у мышей R593C, страдающих гемофилией А, получавших кДНК F8 CRMSBS2 SBR Интрон 3. Результаты демонстрируют значительное сокращение времени для достижения гемостаза (р < 0,0001) у мышей, страдающих гемофилией, после рассечения хвостовой вены.
[0057] На Фиг. 19 схематически изображены CRMSBS1 и новые типовые конструкции без интрона MVM или химерного интрона вместе с трансгеном фактора VIII с удаленным доменом В и Инс1-Инс3. CRM относится к цис-регулирующему элементу. SBS относится к Sangamo Biosciences. чFVIII относится к фактору VIII человека. СП относится к сигнальному пептиду. ИКП относится к инвертированному концевому повтору. SPA относится к синтетической последовательности полиаденилирования. "Инс1" и "Инс3" являются такими, как изображено выше на Фиг. 2.
[0058] На Фиг. 20 схематически изображены CRMSBS2 и новые типовые конструкции без интрона MVM, химерного интрона или SBR Интрон 3 вместе с трансгеном фактора VIII с удаленным доменом В и Инс1-Инс3. CRM относится к цис-регулирующему элементу. SBS относится к Sangamo Biosciences. чFVIII относится к фактору VIII человека. СП относится к сигнальному пептиду. ИКП относится к инвертированному концевому повтору. SPA относится к синтетической последовательности полиаденилирования. "Инс1" и "Инс3" являются такими, как изображено выше на Фиг. 2.
[0059] На Фиг. 21 изображен график, показывающий оценку уровней чFVIII и активности путем экспрессии кДНК AAV F8 in vitro в клетках HepG2. кДНК AAV2/6 F8 (CRMSBS2 SBR Интрон 3) добавляли к клеткам HepG2 в дозах вирусных геномов 4,8Е+06, 2.4Е+06, 1,2Е+- 6 и 0,6Е+06 на 1Е+05 клеток в 24-луночной чаше (обозначается как время t0 дней). Супернатанты анализировали на уровни секретируемого чFVIII с помощью ИФА (крайние левые столбцы) и проводили анализ АЧТВ (активированное частичное тромбопластиновое время) коагулирующей активности (крайние правые столбцы) и хромогенный анализ активности (средние столбцы) в течение семи дней (t7) после добавления вируса AAV2/6. Результаты показали хорошую корреляцию между уровнями секретируемого чFVIII и активностью (сообщалось как Ед/мл на левой оси и Процент от Нормы на правой оси "у", 1 Ед/мл = 100 Процент от Нормы). Показанные данные представляют собой n=6 биологических повторов. "Усы" погрешностей представляют собой стандартную ошибку среднего.
[0060] На Фиг. 22 изображен график, показывающий улучшенные выходы продукции в контексте интронных серий CRMSBS1 или CRMSBS2. Вирус продуцировался в клетках HEK293, выход из двух клеточных фабрик (2 КФ). CRM относится к цис-регулирующему элементу. SBS относится к Sangamo Biosciences. чFVIII относится к фактору VIII человека. СП относится к сигнальному пептиду. ИКП относится к инвертированному концевому повтору. SPA относится к синтетической последовательности полиаденилирования. Инс1 и Инс3 относятся к инсулятору 1 и инсулятору 3, соответственно.
[0061] На Фиг. 23 изображен график, показывающий уровни продуцирования чFVIII-BD у мышей дикого типа с использованием нескольких серотипов AAV, как показано. Самцам мышей C57BL/6 внутривенно вводили 6Е+10 гв/мышь (~ 2Е+12 гв/кг) для конструкций AAV2/5 и AAV2/9 или 6Е+10 гв/мышь (~ 2Е+12 гв/кг) и 1,8Е+11 гв/мышь (~ 7Е+12 гв/кг) для конструкций AAV2/6 и AAV2/8. На фигуре изображен супрафизиологический чFVIII-BDD, обнаруженный в плазме всех образцов, за исключением некоторых из более низких доз AAV2/6 и AAV 2/5.
[0062] На Фиг. 24 изображен график, показывающий относительную экспрессию трансгена чFVIII-BDD у мышей дикого типа, в случае если трансген был доставлен через AAV2/6, очищенным либо продуцированием HEK293 (HEK), либо продуцированием бакуловируса в клетках Sf9 (Sf9). Трансген, используемый с исходной кДНК F8 (Фиг. 4). Самцам мышей C57BL/6 вводили исходную кДНК F8 путем внутривенной инъекции (в хвостовую вену) 1,8Е+11 гв/мышь (~ 7Е+12 гв/кг) исходной кДНК AAV2/6 F8, продуцируемой либо клетками HEK293, либо Sf9/rBV (рекомбинантный бакуловирус). Обработка исходной кДНК F8 из клеток HEK293 достигала среднего пикового значения 142,1% ± 8,3% СОС (n=8) (измерено с помощью ИФА чFVIII) от нормальных уровней человеческого FVIII в плазме. Аналогичный уровень 132,8% ± 18,6% СОС (n=5) (измерено с помощью ИФА чFVIII) был достигнут после введения мышам исходной кДНК F8 (Sf9/rBV).
[0063] На Фиг. 25А и 25В изображен уровень экспрессии у мышей кДНК донора трансгена CRBSBS2 SBR Интрон 3 (Инс1-Инс3). На Фиг. 25А схематически изображен донор, который также изображен на Фиг. 10. На Фиг. 25В изображено количество чFVIII-BDD, обнаруживаемое в плазме, выраженное как в виде нг/мл, так и в процентах от нормы (в нормальной человеческой плазме). Самцам мышей C57BL/6 вводили внутривенно 1,8Е + 11 гв/мышь (~ 7Е+12 гв/кг) кДНК AAV2/6 CRMSBS2 SBR Интрон 3 (n=8). Показаны средние пиковые уровни чFVIII-BDD в плазме мышей C57BL/6, измеренные с помощью ИФА чFVIII.
[0064] На Фиг. 26А и 26В изображена схема дозирования для FVIII-BDD человека в различных исследованиях приматов, отличных от человека (NHP - non-human primate), включая исследования с удалением всей иммуносупрессии на 103-й день. На Фиг. 26А изображен обзор дозирования Ритуксана и Солу-Медрола. Дозировка Ритуксана (10 мг/кг, в/в) была введением дотестового препарата, в то время как ежедневная дозировка метилпреднизолона (Солу-Медрол) (10 мг/кг, в/м) производилась до дня 103. На рисунке 26В изображены схемы дозирования для экспериментов с добавлением дотестового и послетестового препарата, а также изображено время дозирования Ритуксана и Солу-Медрола. Группы 1-5 следовали схеме иммуносупрессии при инъекции дотестового препарата, тогда как Группы 6-8 следовали схеме иммуносупрессии при инъекции послетестового препарата. Продолжительность исследования составила 56 дней.
[0065] На Фиг. 27 изображен график, показывающий корреляцию между активностью чFVIII и уровнями в NHP, получавших кДНК AAV2/6 4F8. Яванским макакам вводили рецептурный буфер или 6Е+12 гв/кг кДНК AAV2/6 4F8 (исходный вектор). На Фиг. 27 изображены отдельные животные для контрольной группы состава (Группа 1, идентификаторы животных 1101-1102) и дозовая группа 6Е+12 гв/кг (Группа 3, идентификаторы животных 3101-3103). На 14-й день после добавления тестового препарата уровни и активность циркулирующего чFVIII анализировали с помощью ИФА или активности свертывания АЧТВ. Нормальные уровни FVIII яванских макак составляют ~ 1 Ед/мл (~ 100% от нормы), отраженные в данных активности свертывания для контрольной группы состава, поскольку анализ активности свертывания не является специфическим для человеческого FVIII. ИФА специфичен для человеческого FVIII, а не для FVIII яванских макак, таким образом, как ожидается, не будет уровней чFVIII, измеренных с помощью ИФА в контрольной группе состава. У животных Группы 3 наблюдаются супрафизиологические уровни и активность циркулирующего чFVIII, выше 8 Ед/мл (800% от нормы).
[0066] На Фиг. 28А и 28В изображены графики, показывающие профили ферментов печени в NHP, получавших кДНК AAV 4F8. Яванским макакам вводили рецептурный буфер или 6Е+12 гв/кг кДНК AAV2/6 4F8 (исходный вектор). Показаны репрезентативные профили ферментов печени у животных для контрольной группы состава (Группа 1, идентификатор животных 1101) и дозовая группа 6Е+12 гв/кг (Группа 3, идентификатор животных 3102) в качестве индикатора состояния печени. Показаны аланинаминотрансфераза (АЛТ) и аспартатаминотрансфераза (ACT). Допустимые референтные значения верхнего предела, которые все еще находятся в нормальном диапазоне для яванских макак для АЛТ составляют 126 Ед/л и 120 Ед/л для ACT. Как для контрольной группы состава, так и для группы кДНК AAV 4F8 6А+12 гв/кг (исходный вектор) уровни АЛТ/АСТ были увеличены после биопсии печени (биопсия печени осуществлялась на 41-й день), обозначены звездочкой. С другой стороны, кДНК AAV 4F8 хорошо переносилась на протяжении всего исследования (247 дней).
[0067] На Фиг. с 29А по 29D изображены графики, показывающие сводку уровней человеческого FVIII в плазме крови для исследования NHP с использованием доноров AAV, несущих протеины фактора VIII (F8) с удаленным доменом В (FVIII-BDD). На Фиг. 29А изображены результаты для животных Группы 2 (AAV2/6, 2Е+12 гв/кг); На Фиг. 29В изображены результаты для животных Группы 3 (AAV2/6, 6Е+12 гв/кг); На Фиг. 29С изображены результаты для животных Группы 4 (AAV2/8, 6Е+12 гв/кг); и на Фиг. 29D изображены результаты для животных Группы 5 (AAV2/8, 6Е+12 гв/кг). 1 Ед/мл фактора VIII человека считается физиологически нормальным и, таким образом, равен 100% нормального физиологического циркулирующего фактора VIII человека.
[0068] На Фиг. 30 изображен график, показывающий пиковые уровни антигена FVIII человека в течение исследования после лечения у приматов, отличных от человека (NHP) с указанными конструкциями (в векторах AAV6 или AAV8). При уровнях дозы 2Е+12 гв/кг (n=3) достигнуты пиковые значения 111,0%, 493,9% и 840,0% (общее среднее значение 481,6% при измерении с помощью ИФА чFVIII) от нормальных уровней чFVIII человека в плазме крови. При более высокой дозе, составляющей 6Е+12 гв/кг (n=3), достигнуты пиковые значения 450,0%, 625,6% и 886,7% [общее среднее значение 654,1%] уровней чFVIII в плазме крови. Общее среднее значение для AAV8 составило 147,1%.
[0069] На Фиг. с 31А по 31С изображены графики, показывающие результаты от отдельных яванских макак (n=3, животные 2101, 2102 и 2103), получавших низкую дозу (2Е+12 гв/кг, Группа 2) кДНК AAV2/6-FVIII-BDD, в течение 168 дней после дозирования. На всех трех графиках концентрации FVIII-BDD в плазме крови, измеренные с помощью ИФА, показаны черным цветом. Кроме того, концентрации нейтрализующего антитела к FVIII (показаны как единицы Бетесды) в плазме крови показаны серым цветом. Пунктирная горизонтальная линия представляет предельную точку единицы Бетесды, ниже которой значения не будут считаться положительными для нейтрализующих антител к FVIII. Прием Солу-Медрола был остановлен на 103-й день, как показано пунктирной вертикальной линией. Каждый график показывает результаты для одной обезьяны: на Фиг. 31А изображено животное 2101; на Фиг. 31В изображено животное 2102; и на Фиг. 31С изображено животное 2103.
[0070] На Фиг. с 32А по 32С изображены графики, показывающие результаты от отдельных яванских макак (n=3, животные 3101, 3102 и 3103), получавших высокую дозу (6Е+12 гв/кг, Группа 3) кДНК AAV2/6-FVIII-BDD, в течение 168 дней после дозирования. На всех трех графиках концентрации FVIII-BDD в плазме крови, измеренные с помощью ИФА, показаны черным цветом. Кроме того, концентрации нейтрализующего антитела к FVIII (показаны как единицы Бетесды) в плазме крови показаны серым цветом. Пунктирная горизонтальная линия представляет предельную точку единицы Бетесды, ниже которой значения не будут считаться положительными для нейтрализующих антител к FVIII. Прием Солу-Медрола был остановлен на 103-й день, как показано пунктирной вертикальной линией. Каждый график показывает результаты для одной обезьяны: на Фиг. 32А изображено животное 3101; на Фиг. 32В изображено животное 3102; и на Фиг. 32С изображено животное 3103.
[0071] На Фиг. с 33А по 33D изображены графики, показывающие результаты от отдельных яванских макак (n=3, животные 4101, 4102 и 4103), получавших высокую дозу (6Е+12 гв/кг, Группа 4) кДНК AAV2/8-FVIII-BDD, в течение 168 дней после дозирования. На графиках 33А-33С концентрации FVIII-BDD в плазме крови, измеренные с помощью ИФА, показаны черным цветом. Кроме того, концентрации нейтрализующего антитела к FVIII (показаны как единицы Бетесды) в плазме крови показаны серым цветом. На Фиг. 33D изображено "раздутие" нижних значений на графике для животного 4103 (следует отметить, что ось "у" на Фиг. 33А-33С идет от 0-5 Ед/мл антигена FVIII, тогда как на Фиг. 33D идет от 0-1 Ед/мл антигена FVIII). Пунктирная горизонтальная линия представляет предельную точку единицы Бетесды, ниже которой значения не будут считаться положительными для нейтрализующих антител к FVIII. Прием Солу-Медрола был остановлен на 103-й день, как показано пунктирной вертикальной линией. Каждый график показывает результаты для одной обезьяны: на Фиг. 33А изображено животное 4101; на Фиг. 33В изображено животное 4102; на Фиг. 33С и 33D изображено животное 4103.
[0072] На Фиг. с 34А по 34Е изображены графики, показывающие результаты от отдельных яванских макак (n=3), получавших высокую дозу (6Е+12 гв/кг, Группа 5) кДНК AAV2/8-FVIII-BDD, в течение 168 дней после дозирования. На Фиг. 34А-34С концентрации FVIII-BDD в плазме крови, измеренные с помощью ИФА, показаны черным цветом. Кроме того, концентрации нейтрализующего антитела к FVIII (показаны как единицы Бетесды) в плазме крови показаны серым цветом. На Фиг. 34D и 34Е изображены "раздутия" нижних значений на графике (следует отметить, что ось "у" на Фиг. 34А-С идет от 0-5 Ед/мл антигена FVIII, тогда как ось на Фиг. 34D и 34Е идет от 0-1 Ед/мл антигена FVIII). Пунктирная горизонтальная линия представляет предельную точку единицы Бетесды, ниже которой значения не будут считаться положительными для нейтрализующих антител к FVIII. Прием Солу-Медрола был остановлен на 103-й день, как показано пунктирной вертикальной линией. Каждый график показывает результаты для одной обезьяны (животное 5101: Фиг. 34А, 5102: Фиг. 34В и 5103: Фиг. 34С).
[0073] На Фиг. 35А и 35F изображены графики, показывающие введение терапевтического препарата чFVIII животным, экспрессирующим трансгенный чFVIII. Яванским макакам из Групп 1, 3, 4 и 5 (идентификаторы животных 1101, 3101, 3103, 4103, 5101, 5102) был введен биологический препарат чFVIII Xyntha®, введения состояли из четырех недельных инфузий 25 Ед/кг Xyntha®, как изображено на Фиг. 35A-35F в виде перевернутых треугольников. Введения Xyntha® соответствовали дням 198, 205, 212, 219 (после добавления кДНК тестового препарата AAV 4F8 в день 0). Из-за короткого периода полувыведения чFVIII и недельного сбора плазмы не наблюдалось увеличения уровней чFVIII от биологического препарата Xyntha® чFVIII. Однако наблюдалось увеличение ингибирующих антител после введения Xyntha® (измерение по единицам Бетесда, БЕ) в контрольной группе 1 состава (см., идентификатор животного 1101, которое не получало кДНК AAV 4F8, Фиг. 35А). Как показано на Фиг. 35А, БЕ достигла ~ 0,9 БЕ/мл после введения Xyntha®, обозначено серой областью графика. Животное с уникальным идентификатором 3103 имело высокий уровень БЕ, выше БЕ/мл в течение многих недель, а добавленное испытание биологическим препаратом чFVIII (обозначено пунктирными вертикальными линиями) не вызывало дополнительных детектируемых ингибирующих антител (БЕ) (Фиг. 35В). Во всех группах, которые получали кДНК AAV 4F8, и имели устойчивые уровни чFVIII, не наблюдалось увеличения ингибирующих антител, БЕ (Фиг. с 35С по F, обозначено серыми стрелками). На Фиг. 35С изображено, что уровни чFVIII для животного с идентификатором 3101, были устойчивыми в течение 19-ти недель при ~ 150% от нормального уровня чFVIII, а на Фиг. 35D изображено, что уровни чFVIII для животного с идентификатором 4103 были устойчивыми в течение 19-ти недель при ~ 10% от нормального уровня чFVIII. На Фиг. 35Е изображено, что уровни чFVIII для животного с идентификатором 5101, были устойчивыми в течение 19-ти недель при ~ 15% от нормального уровня чFVIII, в то время как на Фиг. 35F уровни чFVIII для животного с идентификатором 5102 были устойчивыми в течение 19-ти недель при ~ 35% от нормального уровня чFVIII. Пунктирная горизонтальная линия представляет точку сочленения единицы Бетесды, ниже которой значения не будут считаться положительными для нейтрализующих антител к FVIII. Вертикальная пунктирная линия представляет собой отмену Солу-Медрола на 103-й день введения послетестового препарата на Фиг. 35В, 35Е и 35F.
[0074] На Фиг. 36А и 36В изображена in vitro экспрессия ч VIII-BDD из эндогенного локуса альбумина в клетках HepG2. На Фиг. 36А изображен график, показывающий чFVIII-BDD, обнаруженный в клеточном супернатанте HepG2 с течением времени (t = дни) после введения ZFN, нацеленных на альбумин (SBS#47171/47898, AAV2/6-ZFN) и кДНК чFVIII-BDD AAV2/6-FVIII-BDD (CRMSBS2 Без Интрона) в клетки HepG2 AAV2/6-FVIII-BDD, введенный при 3,0Е+04, 6,0Е+04 и 1,2Е+05 вместе с AAV2/6-ZFN 3.0Е+05. На Фиг. 36В изображен график, показывающий отдельно кДНК чFVIII (без ZFN). Показанные данные представляют собой n=3 биологических повторов. "Усы" погрешностей представляют собой стандартную ошибку среднего повторов. Пунктирная линия представляет собой предел выявления чFVIII с помощью ИФА.
[0075] На Фиг. 37 изображен график, показывающий in vitro обнаружение направленной интеграции трансгена чFVIII-BDD в эндогенный локус альбумина. Направленная интеграция была обнаружена с помощью количественной ПЦР, содержащей сайт вставки альбумина. 5'-праймер ПЦР был расположен в эндогенном локусе альбумина человека, зонд ПЦР расположен в пределах ИКП кассеты чFVIII-BDD, а 3'-праймер расположен в пределах трансгена чFVIII-BDD. Отображаемые числа нормализуются к гену "домашнего хозяйства" GAPDH.
[0076] На Фиг. 38 изображен график, показывающий пиковые уровни антигена человека FVIII после лечения у приматов, отличных от человека (NHP) в соответствии с графиком, изображенным на Фиг. 26В. Группы 1-5 следовали схеме иммуносупрессии (ИС) при инъекции дотестового препарата. При уровнях дозы 6Е+11 гв/кг (n=3, Группа 3) общее среднее значение пиковых значений составляло 5,7% (измеренное с помощью ИФА чFVIII) от нормальных уровней чFVIII в плазме крови у людей. При более высокой дозе, составляющей 2Е+12 гв/кг (n=3, Группа 4), общее среднее значений пиковых значений составляло 56,4%, а при 6Е+12 гв/кг (n=3, Группа 5) общее среднее значений пиковых значений составляло 229,0% (измеренное с помощью ИФА чFVIII) от уровней чFVIII в плазме крови. Заштрихованный серый цвет - это Группы 7 и 8, которые следовали режиму ИС послетестового препарата. На уровне дозы 2Е+12 гв/кг (n=3, Группа 7) общее среднее значение пиковых значений составляло 87,9%, а для 6Е+12 гв/кг (n=3, Группа 8) - 101,7%. Группы 1 и 6 представляли собой контрольные группы состава, обозначенные как Состав.
[0077] На Фиг. 39 изображен график, показывающий результаты после повторного введения субъектам в когорте 2Е+11 гв/кг, изображенной на Фиг. 38, с применением того же серотипа, что приводит к обнаружению уровней циркулирующего антигена FVIII человека. Группе 2, представляющей исходную дозу 2Е+11 гв/кг (n=3), была повторно назначена доза 9Е+11 гв/кг (n=3) на 56-й день исследования (Фиг. 26В). Показаны уровни циркулирующего антигена FVIII человека через семь дней после повторного введения, обозначены полуоткрытым кружком (7-й день при новой дозе 9Е+11 гв/кг). Все остальные данные те же, что и на Фиг. 38 (пиковые уровни такие, как на 56-й день исследования).
[0078] На Фиг. 40А и 40В изображены графики, показывающие in vitro получение и обнаружение чVIII из эндогенного локуса альбумина. На Фиг. 40А изображены суммарные уровни чFVIII, обнаруженные в клеточном супернатанте HepG2, после введения ZFN, нацеленных на альбумин (SBS#42906/43043) и кДНК чFVIII (CRMSBS2 SBR Интрон 3) в клетки HepG2 (анализировали на 19-й день). В этом примере кДНК чFVIII применяли в возрастающих дозах от 3.0Е+04, 6,0Е+04 и 1,2Е+05 вместе с ZFN 3,0Е+05. Также показана кДНК чFVIII с ЗФБ (без ZFN) для контроля общего вируса, добавленного к клеткам, и одна кДНК чFVIII (без ZFN), причем обе из них демонстрируют едва обнаруживаемый секретируемый чFVIII. Это связано с тем, что эписомальный чFVIII деградирует до накопления достаточного количества обнаруживаемого секретируемого чFVIII. Показанные данные представляют собой n=2 биологических повторов. "Усы" погрешностей представляют собой стандартную ошибку среднего технических и биологических повторов. Пунктирная линия представляет собой предел обнаружения чFVIII с помощью ИФА. На Фиг. 40В изображены уровни целевой интеграции эндогенного альбумина-чFVIII с помощью NHEJ (негомологичного соединения концов) с применением количественной ПЦР. 5'-праймер расположен в эндогенном локусе альбумина человека, зонд расположен в пределах ИКП кассеты чFVIII, а 3'-праймер расположен в пределах чFVIII.
ПОДРОБНОЕ ОПИСАНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
[0079] В данном документе описаны экспрессионные кассеты для экспрессии трансгена, в частности в клетках печени. Конструкции могут быть использованы для доставки любого трансгена(ов) в клетки печени, in vivo или in vitro и могут применяться для лечения и/или профилактики любого заболевания или расстройства, которое может быть облегчено путем предоставления одного или более трансгенов. В отличие от применяемых в данное время нацеленных на печень конструкций, описанные в данном документе конструкции включают модифицированные энхансерные и/или интронные последовательности и, кроме того, экспрессируют трансген на высоких уровнях даже без использования интрона MVM. Эти конструкции также малы, что делает возможным их успешное применение с трансгенами, которые доставляются с помощью небольших векторных систем, таких как AAV.
[0080] Описанные в данном документе конструкции могут быть применены для экспрессии чFVIII BDD в печени приматов, отличных от человека. В зависимости от начальной дозы кДНК экспрессионной кассеты AAV чF8 уровни циркулирующего в плазме чFVIII были выше 800% от нормального циркулирующего чFVIII. После этих начальных высоких доз многие животные экспрессировали 10-150% от нормальных уровней в течение более восьми недель. Кроме того, некоторые животные не демонстрировали ответ антитела к чFVIII при введении инъецированного белка чFVIII (Xyntha®), что наводит на мысль о развитии толерантности к белку чFVIIII у этих животных в течение эксперимента.
Общие положения
[0081] Для реализации способов на практике, а также при приготовлении и применении описанных в данном документе композиций используют, если не указано иное, обычные методы молекулярной биологии, биохимии, структуры и анализа хроматина, вычислительной химии, культуры клеток, рекомбинантной ДНК и методы смежных областей, которые находятся в пределах компетентности специалиста данной области техники. Эти методы в полной мере описаны в литературе. См., например, Sambrook и соавт. MOLECULAR CLONING: A LABORATORY MANUAL, Второе издание, Cold Spring Harbor Laboratory Press, 1989 год и Третье издание, 2001 год; Ausubel и соавт., CURRENT PROTOCOLS IN MOLECULAR BIOLOGY, John Wiley & Sons, Нью-Йорк, 1987 год и периодические обновления; серии METHODS IN ENZYMOLOGY, Academic Press, Сан Диего; Wolffe, CHROMATIN STRUCTURE AND FUNCTION, Третье издание, Academic Press, Сан Диего, 1998 год; METHODS IN ENZYMOLOGY, Том. 304, "Chromatin" (P.M. Wassarman and A.P. Wolffe, ред.), Academic Press, Сан Диего, 1999 год; и METHODS IN MOLECULAR BIOLOGY, Том. 119, "Chromatin Protocols" (P.B. Becker, ред.) Humana Press, Тотова, 1999 год.
Определения
[0082] Термины "нуклеиновая кислота", "полинуклеотид" и "олигонуклеотид" применяются взаимозаменяемо для обозначения дезоксирибонуклеотидного или рибонуклеотидного полимера, линейной или кольцевой конформации, а также находящегося либо в одно-, либо в двух-цепочечной форме. Для целей данного изобретения эти термины не следует рассматривать как ограничивающие длину полимера. Термины могут включать известные аналоги природных нуклеотидов, а также нуклеотиды, которые модифицированы в основной, сахарной и/или фосфатных частях (например, фосфортиоатные остовы). В общем, аналог конкретного нуклеотида имеет одинаковую специфичность спаривания оснований; то есть, аналог А будет парой оснований с Т.
[0083] Термины "полипептид", "пептид" и "белок" применяются взаимозаменяемо для обозначения полимера аминокислотных остатков. Этот термин также относится к аминокислотным полимерам, в которых одна или большее количество аминокислот являются химическими аналогами или модифицированными производными соответствующих встречающихся в природе аминокислот.
[0084] "Рекомбинация" относится к процессу обмена генетической информацией между двумя полинуклеотидами, включая, но не ограничиваясь этим, захват с помощью негомологичного соединения концов (NHEJ) и гомологичной рекомбинации. Для целей данного изобретения термин "гомологичная рекомбинация (ГР)" относится к специализированной форме такого обмена, которая имеет место, например, при репарации двухцепочечных разрывов в клетках с помощью гомологичных механизмов репарации.
[0085] В некоторых способах описания одна или более целевых нуклеаз, как описано в данном документе, создают двухцепочечный разрыв (ДЦР) в целевой последовательности (например, клеточный хроматин) на предопределенном участке (например, ген альбумина). ДЦР обеспечивает интеграцию конструкции, как описано в данном документе. Необязательно, конструкция имеет гомологию с нуклеотидной последовательностью в области разрыва. Экспрессионная конструкция может быть физически интегрирована или, в альтернативном варианте, экспрессионная кассета применяется в качестве матрицы для репарации разрыва с помощью гомологичной рекомбинации, что приводит к введению всей нуклеотидной последовательности или ее части в виде экспрессионной кассеты в клеточный хроматин. Таким образом, первая последовательность в клеточном хроматине может быть изменена и в определенных вариантах реализации изобретения может быть преобразована в последовательность, присутствующую в экспрессионной кассете. Таким образом, использование терминов "заменить" или "замена" можно понимать, как замену одной нуклеотидной последовательности другой (то есть, замена последовательности в информационном смысле) и не обязательно требует физической или химической замены одного полинуклеотида другим.
[0086] В любом из способов, описанных в данном документе, экзогенная нуклеотидная последовательность ("экспрессионная конструкция" или "экспрессионная кассета" или "вектор") может содержать последовательности, которые являются гомологичными, но не идентичными, геномным последовательностям в области интереса, тем самым стимулируя гомологичную рекомбинацию для вставки неидентичной последовательности в область интереса. Таким образом, в определенных вариантах реализации изобретения части последовательности экспрессионной кассеты, которые представляют собой гомологичные последовательности в области интереса, демонстрируют от около 80 до 99% (или любое целое число в этом диапазоне) идентичности последовательности с геномной последовательностью, которая заменена. В других вариантах реализации изобретения гомология между экспрессирующей кассетой и геномной последовательностью составляет более чем 99%, например, если только один нуклеотид отличается между гомологическими областями экспрессионной кассеты и геномными последовательностями на более чем 100 смежных пар оснований. В некоторых случаях негомологичная часть экспрессионной кассеты может содержать последовательности, не присутствующие в области интереса, так что новые последовательности вводятся в область интереса. В этих случаях негомологичная последовательность обычно фланкируется последовательностями из 50-1000 пар оснований (или любым целым значением в этом диапазоне) или любым числом пар оснований большим чем 1000, которые являются гомологичными или идентичными последовательностям в области интереса.
[0087] Термин "последовательность" относится к нуклеотидной последовательности любой длины, которая может быть ДНК или РНК; может быть линейной, кольцевой или разветвленной, а также может быть одноцепочечной или двухцепочечной. Термин "трансген" относится к нуклеотидной последовательности, которую вводят в геном. Трансген может быть любой длины, например, длиной от 2 до 100000000 нуклеотидов (или любое целое число в этом диапазоне или больше), предпочтительно от около 100 и до 100000 нуклеотидов в длину (или любое целое число в этом диапазоне), более предпочтительно от около 2000 и до 20000 нуклеотидов в длину (или любое значение в этом диапазоне) и еще более предпочтительно, от около 5 и до 15 т.п. н (или любое значение в этом диапазоне).
[0088] "Хромосома" представляет собой хроматиновый комплекс, содержащий весь геном или часть генома клетки. Геном клетки часто характеризуется своим кариотипом, который представляет собой совокупность всех хромосом, которые содержат геном клетки. Геном клетки может содержать одну или более хромосом.
[0089] "Эписома" представляет собой реплицирующуюся нуклеиновую кислоту, нуклеопротеиновый комплекс или другую структуру, включающую нуклеиновую кислоту, которая не является частью хромосомного кариотипа клетки. Примеры эписом включают плазмиды и некоторые вирусные геномы. Конструкции, специфичные для печени, описанные в данном документе, могут поддерживаться эписомально или, в альтернативном варианте, могут быть стабильно интегрированы в клетку.
[0090] "Экзогенная" молекула представляет собой молекулу, которая обычно не присутствует в клетке, но может быть введена в клетку с помощью одного или большего количества генетических, биохимических или других способов. "Нормальное присутствие в клетке" определяется в отношении конкретного этапа развития и условий окружающей среды клетки. Так, например, молекула, которая присутствует только во время эмбрионального развития мышц, является экзогенной молекулой по отношению к зрелой мышечной клетке. Аналогично, молекула, индуцированная тепловым шоком, представляет собой экзогенную молекулу по отношению к клетке, на которую не воздействовали тепловым шоком. Экзогенная молекула может содержать, например, функционирующий вариант неправильно функционирующей эндогенной молекулы или неправильно функционирующий вариант нормально функционирующей эндогенной молекулы.
[0091] Экзогенная молекула может быть, среди прочего, небольшой молекулой, например, сгенерированной с помощью процесса комбинаторной химии, или макромолекулой, такой как белок, нуклеиновая кислота, углевод, липид, гликопротеин, липопротеин, полисахарид, любое модифицированное производное вышеупомянутых молекул, или любым комплексом, содержащим одну или более вышеупомянутых молекул. Нуклеиновые кислоты включают ДНК и РНК, могут быть одно- или двухцепочечными; могут быть линейными, разветвленными или кольцевими; и могут быть любой длины. К нуклеиновым кислотам относятся те, которые способны образовывать дуплексы, а также триплекс-образующие нуклеиновые кислоты. Смотри, например, патенты США №5,176,996 и 5,422,251. Белки включают, но не ограничиваются ими, ДНК-связывающие белки, факторы транскрипции, факторы ремоделирования хроматина, метилированные ДНК-связывающие белки, полимеразы, метилазы, деметилазы, ацетилазы, деацетилазы, киназы, фосфатазы, деубиквитиназы, интегразы, рекомбиназы, лигазы, топоизомеразы, гиразы и геликазы.
[0092] Экзогенная молекула может быть тем же типом молекулы, что и эндогенная молекула, например, экзогенным белком или нуклеиновой кислотой. Например, экзогенная нуклеиновая кислота может содержать инфицирующий вирусный геном, плазмиду или эписому, вводимые в клетку, или хромосому, которая обычно не присутствует в клетке. Способы введения экзогенных молекул в клетки известны специалистам в данной области техники и включают, но не ограничиваются ими, липид-опосредованный перенос (то есть, с помощью липосом, включая нейтральные и катионные липиды), электропорацию, непосредственное инъекционное введение, слияние клеток, бомбардировку частицами, совместное осаждение фосфатом кальция, перенос, опосредуемый DEAE-декстраном, и перенос, опосредуемый вирусным вектором. Экзогенная молекула также может быть молекулой того же типа, что и эндогенная молекула, но получена из другого вида, чем клетка. Например, последовательность нуклеиновой кислоты человека может быть введена в линию клеток, первоначально полученную от мыши или хомяка. Способы введения экзогенных молекул в клетки растений известны специалистам в данной области техники и включают, но не ограничиваются ими, трансформацию протопласта, карбид кремния (например, WHISKERS™), трансформацию, опосредованную Agrobacterium, липид-опосредованный перенос (то есть, с помощью липосом, включая нейтральные и катионные липиды), электропорацию, непосредственное инъекционное введение, слияние клеток, бомбардировку частицами (например, с применением, "генной пушки") совместное осаждение фосфатом кальция, перенос, опосредуемый DEAE-декстраном, и перенос, опосредуемый вирусным вектором.
[0093] Напротив, "эндогенная" молекула представляет собой ту, которая обычно присутствует в конкретной клетке на определенном этапе развития в конкретных условиях окружающей среды. Например, эндогенная нуклеиновая кислота может содержать хромосому, геном митохондрии, хлоропласта или другой органеллы, или встречающуюся в природе эписомальную нуклеиновую кислоту. Дополнительные эндогенные молекулы могут включать белки, например, транскрипционные факторы и ферменты.
[0094] Используемый в данном документе термин "продукт экзогенной нуклеиновой кислоты" включает как полинуклеотидные, так и полипептидные продукты, например, продукты транскрипции (полинуклеотиды, такие как РНК) и продукты трансляции (полипептиды).
[0095] "Слитая" молекула представляет собой молекулу, в которой две или большее количество субъединичных молекул являются связанными, предпочтительно ковалентно. Субъединичные молекулы могут быть молекулами одного и того же химического типа или могут быть молекулами разных химических типов. Примеры слитых молекул включают, но не ограничиваются ими, слитые белки (например, слияние между белковым ДНК-связывающим доменом и доменом расщепления), слияния между полинуклеотидным ДНК-связывающим доменом (например, sgPHК), функционально ассоциированным с доменом расщепления, и слитые нуклеиновые кислоты (например, нуклеиновые кислоты, кодирующие слитый белок).
[0096] Экспрессия слитого белка в клетке может быть результатом доставки слитого белка в клетку или доставки полинуклеотида, кодирующего слитый белок, в клетку, при этом полинуклеотид транскрибируется, а транскрипт транслируется с целью генерирования слитого белка. Транс-сплайсинг, расщепление полипептида и лигирование полипептидов также могут быть вовлечены в экспрессию белка в клетке. Способы доставки полинуклеотидов и полипептидов в клетки представлены в другом месте этого описания.
[0097] "Ген" для целей данного описания включает область ДНК, кодирующую генный продукт (см. ниже), а также все области ДНК, которые регулируют продукцию гена, независимо от того, являются ли такие регуляторные последовательности смежными с кодирующими и/или транскрибированными последовательностями. Соответственно, ген включает, но не обязательно ограничивается этим, промоторные последовательности, терминаторы, трансляционные регуляторные последовательности, такие как участки связывания рибосом и участки внутренней посадки рибосомы, энхансеры, сайленсеры, инсуляторы, граничные элементы, точки начала репликации, участки прикрепления матрикса и области контроля локусов.
[0098] Термин "экспрессия гена" относится к преобразованию информации, содержащейся в гене, в продукт гена. Продукт гена может быть прямым транскрипционным продуктом гена (например, мРНК, тРНК, рРНК, антисмысловая РНК, рибозим, структурная РНК или любой другой тип РНК) или белком, продуцируемым в результате трансляции мРНК. Продукты генов также включают РНК, которые модифицируются такими способами, как "кэппинг", полиаденилирование, метилирование и редактирование, а также белки, модифицированные, например, метилированием, ацетилированием, фосфорилированием, убиквитинированием, АДФ-рибозилированием, миристилированием и гликозилированием.
[0099] Термин "модуляция" экспрессии генов относится к изменению активности гена. Модуляция экспрессии может включать, но не ограничивается этим, активацию гена и репрессию гена. Редактирование генома (например, расщепление, изменение, инактивация, случайная мутация) можно применять для модуляции экспрессии. Термин "инактивация гена" относится к любому уменьшению экспрессии гена по сравнению с клеткой, которая не содержит ZFP, TALE или систему CRISPR/Cas, как описано в данном документе. Таким образом, инактивация гена может быть частичной или полной.
[0100] "Область интереса" представляет собой любую область клеточного хроматина, такую как, например, ген или некодирующая последовательность в пределах или рядом с геном, в которой является желательным связывание экзогенной молекулы. Связывание может быть предназначено для целенаправленного расщепления ДНК и/или целенаправленной рекомбинации. Область интереса, может присутствовать в хромосоме, эписоме, геноме органелл (например, митохондрия, хлоропласт) или, например, инфицирующем вирусном геноме. Область интереса, может находиться в кодирующей области гена, в пределах транскрибируемых некодирующих областей, таких как, например, лидерные последовательности, трейлерные последовательности или интроны, или в пределах нетранскрибируемых областей либо против, либо по ходу транскрипции от кодирующей области. Область, представляющая интерес, может быть такой же небольшой по размеру, что и одиночная нуклеотидная пара или до 2000 пар нуклеотидов в длину, или любое целое число пар нуклеотидов.
[0101] Термин "эукариотические" клетки включает в себя, но не ограничиваются ими, грибковые клетки (такие как дрожжи), растительные клетки, клетки животных, клетки млекопитающих и клетки человека (например, Т-клетки), включая стволовые клетки (плюрипотентные и мультипотентные).
[0102] Термины "функциональная связь" и "функционально связанный" (или "функционально ассоциированный") применяются взаимозаменяемо со ссылкой на сопоставление двух или более компонентов (таких как элементы последовательности), в которых компоненты расположены так, что оба компонента функционируют нормально, что дает возможность по меньшей мере одному из компонентов опосредовать функцию, которая воздействует по меньшей мере на один из других компонентов. В качестве иллюстрации регуляторная последовательность транскрипции, такая как промотор, является функционально связанной с кодирующей последовательностью, если регуляторная последовательность транскрипции контролирует уровень транскрипции кодирующей последовательности в ответ на наличие или отсутствие одного, или более регуляторных факторов транскрипции. Регуляторная последовательность транскрипции, как правило, является функционально связанной в цис-конфигурации с кодирующей последовательностью, но не обязательно является непосредственно примыкающей к ней. Например, энхансер представляет собой регуляторную последовательность транскрипции, которая функционально связана с кодирующей последовательностью, хотя они не являются смежными.
[0103] "Функциональный фрагмент" белка, полипептида или нуклеиновой кислоты представляет собой белок, полипептид или нуклеиновую кислоту, последовательность которых не идентична полноразмерному белку, полипептиду или нуклеиновой кислоте, но которые при этом сохраняют ту же функцию, которой обладает полноразмерный белок, полипептид или нуклеиновая кислота. Функциональный фрагмент может обладать большим, меньшим или таким же количеством остатков, что и соответствующая нативная молекула, и/или может содержать одну или большее количество аминокислот или нуклеотидных замен. Способы определения функции нуклеиновой кислоты (например, кодирующей функции, способности к гибридизации с другой нуклеиновой кислоты) хорошо известны в данной области техники. Аналогично, способы определения функции белка также хорошо известны. Например, человеческий фактор VIII с удаленным В-доменом, является функциональным фрагментом полноразмерного белка фактора VIII.
[0104] Полинуклеотидный "вектор" или "конструкция" способен переносить последовательности генов в целевые клетки. Как правило, "векторная конструкция", "экспрессионный вектор", "экспрессионная конструкция", "экспрессионная кассета" и "вектор переноса гена" означают любую конструкцию нуклеиновой кислоты, способную направлять экспрессию представляющего интерес гена, и которая может переносить последовательности генов в целевые клетки. Таким образом, этот термин включает клонирование и экспрессионные носители, а также интегрирующие векторы.
[0105] Термины "субъект" и "пациент" применяются взаимозаменяемо и относятся к млекопитающим, таким как пациенты-люди и приматы, отличные от людей, а также к экспериментальным животным, таким как кролики, собаки, кошки, крысы, мыши и другие животные. Соответственно, термин "субъект" или "пациент", применяемый в данном документе, означает любого млекопитающего пациента или субъекта, которому могут быть введены экспрессионные кассеты по данному изобретению. Субъекты по данному изобретению включают пациентов с расстройством.
Печень-специфичные экспрессионные конструкции
[0106] В данном документе описаны экспрессионные кассеты (конструкции) для применения в направлении экспрессии трансгена в клетке печени, включая in vivo после введения экспрессионной кассеты(сет) субъекту (например, доставку в печень). Экспрессионная конструкция может поддерживаться эписомально и направлять экспрессию трансгена внехромосомно или, в альтернативном варианте, экспрессионную конструкцию можно интегрировать в геном клетки печени, например, посредством опосредованной нуклеазой целенаправленной интеграции.
[0107] Полинуклеотидная экспрессионная конструкция содержит последовательность энхансера, последовательность промотора и один или более трансгенов. Необязательно включены одно или более из следующего: последовательность интрона, последовательность полиаденилирования и/или сигнальный пептид. Любая последовательность энхансера может быть применена в экспрессионных конструкциях, описанных в данном документе. В определенных вариантах реализации изобретения энхансер представляет собой энхансер дикого типа или модифицированный Серпин1 (Chuah и соавт., (2014 год) Molecular Therapy, 22, 1605-1613; Nair и соавт., (2014 год) Blood, 123, 3195-3199).
[0108] В предпочтительных вариантах реализации изобретения энхансер Серпин1 содержит одну или более мутаций (например, точечные мутации) по сравнению с диким типом, например, энхансер Серпин1 содержит одну или более нуклеотидных модификаций, как изображено на Фиг. 5, а именно модификации нуклеотидов в одном или более остатках 1, 5, 14, 32 и/или 39 последовательности, показанной в любой из SEQ ID NO: 1-13. В определенных вариантах реализации изобретения энхансер Серпин1 содержит модификации в положениях 1, 5, 14 и 32 (конструкции, обозначенные CRMSBS1) любой из SEQ ID NO: 1-13, в то время как в других вариантах реализации изобретения энхансер Серпин1 содержит модификацию в положениях 1, 14, 32 и 39 (конструкции, обозначенные CRMSBS2) или любой из SEQ ID NO: 1-13. Типовые последовательности энхансера показаны ниже:
CRMSBS1 (SEQ ID NO: 35):
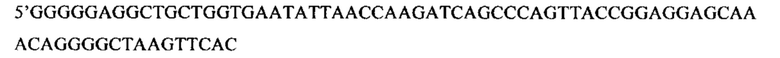
CRMSBS2 (SEQ ID NO: 36): 5'
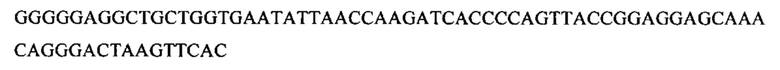
[0109] Подобным образом, любая последовательность промотора может быть применена в экспрессионных кассетах по данному изобретению. В определенных вариантах реализации изобретения промотор представляет собой конститутивный промотор. В других вариантах реализации изобретения промотор представляет собой индуцибельный или тканеспецифичный промотор. В некоторых вариантах реализации изобретения промотор представляет собой печень-специфичный промотор. В определенных вариантах реализации изобретения промотор представляет собой минимальный промотор гена транстиретина (TTRm). В других вариантах реализации изобретения промотор представляет собой промотор гена альфа-1-антитрипсина (чААТ).
[0110] Любой из описанных в данном документе полинуклеотидов может дополнительно необязательно содержать интронную последовательность. В определенных вариантах реализации изобретения экспрессионная конструкция включает усеченную химерную последовательность интрона (Т-химерный интрон), например, как изображено на нижних панелях Фиг. 6, 7, 19 и 20. Т-химерный интрон представляет собой усеченную версию химерного интрона в pCI-neo (GenBank U47120). Химерный интрон в pCI-neo представляет собой 5' сайт донора сплайсинга из гена β-глобина человека, а также точку разветвления и 3' акцепторный сайт вариабельной области тяжелой цепи гена иммуноглобулина. Т-химерный интрон содержит делецию 45 п.н. между 5' донором сплайсинга и точкой разветвления. В других вариантах реализации изобретения экспрессионные конструкции включают мутантную последовательность интрона MVM (например, одну или более мутаций, изображенных на Фиг. 12, 13 и 14 (SEQ ID NO: 15-17).
[0111] В альтернативном варианте, экспрессионные конструкции, как описано в данном документе, могут не иметь интронной последовательности, например, как показано в конструкциях, изображенных на средних панелях Фиг. 6, 7, 19 и 20. Типовая конструкция с аннотациями, включающими трансген фактора VIII, показана ниже, и будет очевидно, что трансген F8 может быть заменен любым трансгеном и что последовательности промотора и инсулятора могут быть дополнительно модифицированы, как описано в данном документе: кДНК чF8, без интрона (полная последовательность, включающая модуль промотора, поли А и Инс, CRMSBS2 Без интрона) (SEQ ID NO: 34):




5'
[0112] Другая типовая конструкция с аннотациями, включающими трансген фактора VIII, показана ниже, и также будет очевидно, что трансген F8 может быть заменен любым трансгеном и что последовательности промотора и инсулятора могут быть дополнительно модифицированы, как описано в данном документе:
[0113] кДНК 4F8, включая интрон (полная последовательность, включающая модуль промотора, поли А и Инс, CRMSBS2 SBR Интрон 3) (SEQ ID NO: 37):





[0114] В предпочтительных вариантах реализации изобретения конструкции, описанные в данном документе содержит последовательности инсулятора (спейсера), как изображено на Фиг. 1, 2, 4, 6, 7, 10, 19, 20 и 25. Типовые последовательностей инсуляторов включают Инс1: 5' GGAACCATTGCCACCTTCA (SEQ ID NO: 28); Инс2: 5' CTATCCATTGCACTATGCT (SEQ ID NO: 29); Инс3: 5' TTTCCTGTAACGATCGGG (SEQ ID NO: 30) и Инс4: 5' TTGAATTCATAACTATCCCAA (SEQ ID NO: 38).
[0115] Очевидно, что любой трансген может быть применен в конструкциях, описанных в данном документе. Кроме того, отдельные компоненты (промотор, энхансер, инсулятор, трансген и тому подобное) описанных в данном документе конструкций могут быть смешаны и согласованы в любой комбинации.
[0116] Конструкции, описанные в данном документе, могут содержаться в любом вирусном или невирусном векторе. Конструкции могут поддерживаться эписомально или могут быть интегрированы в геном клетки (например, посредством опосредованной нуклеазой целенаправленной интеграции).
[0117] Невирусные векторы включают ДНК или РНК-плазмиды, МС ДНК, оголенную нуклеиновую кислоту и нуклеиновую кислоту, входящую в состав носителя для доставки, такого как липосома, наночастица или полоксамер. Вирусные векторы, которые могут быть применены для переноса экспрессионных кассет, описанных в данном документе, включают, но не ограничиваются ими, ретровирусные, лентивирусные, аденовирусные, аденоассоциированные вирусные векторы, векторы на основе вируса коровьей оспы и простого герпеса. Интеграция в геном хозяина возможна с помощью способов переноса генов с помощью векторов ретровирусов, лентивирусов и аденоассоциированных вирусов, и, как описано в данном документе, может быть облегчена посредством опосредованной нуклеазой интеграции.
[0118] В определенных предпочтительных вариантах реализации изобретения конструкции включены в вектор на основе аденоассоциированного вируса ("AAV") или в векторную систему, которая может поддерживаться эписомально или интегрироваться в геном клетки печени (например, посредством опосредованной нуклеазой целенаправленной интеграции). Конструкции рекомбинантных векторов AAV представлены в ряде публикаций, включая патент США №5,173,414; Tratschin и соавт., Mol. Cell. Biol. 5:3251-3260 (1985 год); Tratschin, и соавт., Mol. Cell. Biol. 4:2072-2081 (1984 год); Hermonat (Muzyczka, PNAS 81:6466-6470 (1984 год); и Samulski и соавт., J. Virol. 63:03822-3828 (1989 год).
[0119] Таким образом, в определенных вариантах реализации изобретения экспрессионная конструкция переносится на конструкции AAV и дополнительно содержит 5' и 3' ИКП, фланкирующие элементы экспрессионной конструкции (например, энхансер, промотор, факультативный интрон, трансген и тому подобное), как описано в данном документе. Необязательно, молекулы спейсера также включены между одним или более компонентами экспрессионной конструкции, например, между 5' ИКП и энхансером и/или между сигналом полиаденилирования и 3' ИКП. Спейсеры могут функционировать как гомологические плечи для облегчения рекомбинации в "безопасный" локус (например, альбумина). В определенных вариантах реализации изобретения конструкция представляет собой конструкцию, как изображено на любой из Фиг. 1, 2, 4, 6, 7, 10, 18, 19, 20 или 25.
[0120] В определенных вариантах реализации изобретения векторы AAV, как описано в данном документе, могут быть получены из любого AAV. В определенных вариантах реализации изобретения вектор AAV получают из дефектного и непатогенного парвовируса аденоассоциированного вируса типа 2. Все такие векторы являются производными от плазмиды, которая сохраняет только инвертированные концевые повторы AAV 145 п.о., фланкирующие экспрессионную кассету трансгена. Ключевыми особенностями этой векторной системы являются эффективный перенос гена и стабильная доставка трансгена, благодаря интеграции в геномы трансдуцированной клетки. (Wagner и соавт., Lancet 351:9117 1702-3 (1998 год), Kearns и соавт., Gene Ther. 9:748-55 (1996 год)). Другие серотипы AAV, включая AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8, AAV9 и AAVrh. 10 и любой новый серотип AAV также можно применять в соответствии с данным изобретением. В некоторых вариантах реализации изобретения применяется химерный AAV, где вирусные происхождения последовательностей ДКП (длинного концевого повтора) вирусной нуклеиновой кислоты являются гетерологичными по отношению к вирусному происхождению последовательностей капсида. Неограничивающие примеры включают химерный вирус с ДКП, полученным из AAV2, и капсиды, полученные из AAV5, AAV6, AAV8 или AAV9 (то есть ААУ2/5, AAV2/6, AAV2/8 и AAV2/9, соответственно).
[0121] Ретровирусные векторы включают те, которые основаны на вирусе лейкемии мышей (MuLV), вирусе лейкоза гиббонов (GaLV), вирусе иммунодефицита обезьян (ВИО), вирусе иммунодефицита человека (ВИЧ) и их комбинации (см, например, Buchscher и соавт., J. Virol. 66:2731-2739 (1992 год); Joharm и соавт., J. Virol. 66:1635-1640 (1992 год); Sommerfelt и соавт., Virol. 176:58-59 (1990 год); Wilson и соавт., J. Virol. 63:2374-2378 (1989 год); Miller и соавт., J. Virol. 65:2220-2224 (1991 год); PCT/US94/05700).
[0122] Конструкции, описанные в данном документе, также могут быть включены в систему аденовирусных векторов. Аденовирусные векторы способны к очень высокоэффективной трансдукции во многих типах клеток и не требуют деления клеток. С помощью таких векторов получают высокий титр и высокий уровень экспрессии. Этот вектор можно получить в больших количествах в относительно простой системе.
[0123] pLASN и MFG-S являются примерами ретровирусных векторов, которые применялись в клинических исследованиях (Dunbar и соавт., Blood 85:3048-305 (1995 год); Kohn и соавт., Nat. Med. 1:1017-102 (1995 год); Malech и соавт., PNAS 94:22 12133-12138 (1997 год)). PA317/pLASN был первым терапевтическим вектором, который применялся в исследовании генной терапии. (Blaese и соавт., Science 270:475-480 (1995 год)). Эффективность трансдукции, составляющая 50% или более, наблюдалась при применении MFG-S-упакованных векторов. (Ellem и соавт., Immunol Immunother. 44(1):10-20 (1997 год); Dranoff и соавт., Hum. Gene Ther. 1:111-2 (1997 год).
[0124] Дефектные по репликации рекомбинантные аденовирусные векторы (Ad) также могут быть применены с описанными в данном документе полинуклеотидами. Большинство аденовирусных векторов сконструированы таким образом, что трансген заменяет гены Ad E1a, E1b и/или Е3; впоследствии дефектный по репликации вектор распространяется в клетках человека 293, которые обеспечивают функционирование удаленного гена в транс-положении. Векторы Ad могут трансформировать несколько типов тканей in vivo, включая неделящиеся, дифференцированные клетки, такие как обнаруженные в печени, почках и мышцах. Обычные векторы Ad характеризуются возможностью переносить большой объем. Пример применения вектора Ad в клиническом исследовании включал полинуклеотидную терапию с целью противоопухолевой иммунизации с помощью внутримышечного введения (Sterman и соавт., Hum. Gene Ther. 7:1083-9 (1998 год)). Дополнительные примеры применения аденовирусных векторов для переноса генов в клинических исследованиях включают Rosenecker и соавт., Infection 24:1 5-10 (1996 год); Sterman и соавт., Hum. Gene Ther. 9:7 1083-1089 (1998 год); Welsh и соавт., Hum. Gene Ther. 2:205-18 (1995 год); Alvarez и соавт., Hum. Gene Ther. 5:597-613 (1997 год); Topf и соавт., Gene Ther. 5:507-513 (1998 год); Sterman и соавт., Hum. Gene Ther. 7:1083-1089 (1998 год).
[0125] Упаковочные клетки применяются для образования вирусных частиц, которые способны инфицировать клетку-хозяина. Такие клетки включают клетки HEK293 и Sf9, которые могут применяться для упаковки AAV и аденовируса, и клетки ψ2 или клетки РА317, которые упаковывают ретровирус. Как правило, вирусные векторы, применяемые в генной терапии, генерируются линией клеток-продуцентов, которая упаковывает вектор нуклеиновой кислоты в вирусную частицу. Векторы обычно содержат минимальные вирусные последовательности, необходимые для упаковки и последующей интеграции в хозяина (если применимо), причем другие вирусные последовательности заменяются экспрессионной кассетой, кодирующей белок, который должен быть экспрессирован. Отсутствующие вирусные функции обеспечиваются в транс-положении линией упаковочных клеток. Например, векторы AAV, применяемые в генной терапии, как правило, имеют только последовательности инвертированного концевого повтора (ИКП) из генома AAV, которые необходимы для упаковки и интеграции в геном хозяина. Вирусную ДНК упаковывают в линию клеток, которая содержит плазмиду-помощник, кодирующую другие AAV гены, а именно rep и cap, но не содержит последовательности ИКП. Линию клеток также инфицируют аденовирусом в качестве помощника. Вирус-помощник способствует репликации вектора AAV и экспрессии генов AAV из плазмиды-помощника. Плазмида-помощник не упаковывается в значительных количествах из-за отсутствия последовательности ИКП. Уровень контаминации аденовирусом можно снизить, например, при термической обработке, к которой аденовирус более чувствителен, чем AAV. В некоторых вариантах реализации изобретения AAV получают с применением системы экспрессии бакуловируса.
[0126] При применении многих методов генной терапии является желательным, чтобы вектор генной терапии был доставлен к определенному типу ткани с высокой степенью специфичности. Соответственно, вирусный вектор можно модифицировать, чтобы получить специфичность для данного типа клеток, путем экспрессии лиганда в виде слитого белка с белком вирусной оболочки на внешней поверхности вируса. Лиганд выбирают на основе его аффинности к рецептору, о котором известно, что он присутствует на типе клеток, представляющем интерес. Например, в публикации Han и соавт., Proc. Natl. Acad. Sci. USA 92:9747-9751 (1995 год), сообщается, что вирус мышиного лейкоза Молони можно модифицировать для экспрессии герегулина человека, слитого с gp70, при этом рекомбинантный вирус инфицирует определенные клетки рака молочной железы человека, экспрессирующие рецептор к эпидермальному фактору роста человека. Этот принцип можно распространить на другие пары "вирус - целевая клетка", в которых целевая клетка экспрессирует рецептор, а вирус экспрессирует слитый белок, содержащий лиганд для рецептора клеточной поверхности. Например, можно сконструировать нитчатый фаг с целью выставления фрагментов антител (например, FAB или Fv), обладающих специфической аффинностью связывания для практически любого выбранного клеточного рецептора. Хотя вышеприведенное описание относится в первую очередь к вирусным векторам, те же самые принципы могут быть применены к невирусным векторам. Такие векторы могут быть сконструированы так, чтобы содержать специфические последовательности поглощения, которые способствуют поглощению вектора определенными целевыми клетками.
[0127] Описанные в данном документе полинуклеотиды могут включать в себя одно или более оснований и/или остовов неестественного происхождения. В частности, экспрессионная кассета, как описано в данном документе, может включать в себя метилированные цитозины для достижения состояния покоя транскрипции в области, представляющей интерес.
[0128] Кроме того, экспрессионные конструкции, как описано в данном документе, могут также включать в себя дополнительные транскрипционные или трансляционные регуляторные, или другие последовательности, например, последовательности Козак, дополнительные промоторы, энхансеры, инсуляторы, участки внутренней посадки рибосомы, последовательности, кодирующие 2А-пептиды, сайты расщепления фурином и/или сигналы полиаденилирования. Кроме того, контрольные элементы генов, представляющих интерес, могут быть функционально связаны с репортерными генами для создания химерных генов (например, репортерные экспрессионные кассеты).
Трансгены
[0129] Описанные в данном документе конструкции могут быть применены для доставки любого трансгена в печень. Типовые трансгены (также называемые генами интереса и/или экзогенными последовательностями) включают, но без ограничений, любую кодирующую полипептид последовательность (например, кДНК), последовательности промотора, последовательности энхансера, метки эпитопа, маркерные гены, сайты распознавания ферментами расщепления и различные типы экспрессионных конструкций. Маркерные гены включают, но без ограничений, последовательности, кодирующие белки, которые опосредуют устойчивость к антибиотикам (например, устойчивость к ампициллину, устойчивость к неомицину, устойчивость к G418, устойчивость к пуромицину), последовательности, кодирующие окрашенные или флуоресцентные, или люминесцирующие белки (например, зеленый флуоресцентный белок, усиленный зеленый флуоресцентный белок, красный флуоресцентный белок, люцифераза) и белки, которые опосредуют усиленный рост клеток и/или амплификацию генов (например, дигидрофолатредуктаза). Теги эпитопа включают, например, одну или более копий FLAG, His, myc, Тар, НА или любую обнаруживаемую аминокислотную последовательность.
[0130] В предпочтительном варианте реализации изобретения трансген содержит полинуклеотид, кодирующий любой полипептид, экспрессия которого желательна в клетке, включая, но без ограничений, антитела, антигены, ферменты, рецепторы (клеточной поверхности или ядерные), гормоны, лимфокины, цитокины, репортерные полипептиды, факторы роста и функциональные фрагменты любого из вышеперечисленных. Кодирующими последовательностями могут быть, например, кДНК.
[0131] В определенных вариантах реализации изобретения трансген(ы) кодирует(ют) функциональные варианты белков, которые отсутствуют или присутствуют в недостаточных количествах при любом генетическом заболевании, включая, но без ограничений, лизосомальные болезни накопления (например, болезнь Гоше, Фабри, Хантера, Гурлера, Ниманна-Пика и тому подобные), болезни нарушения обмена веществ и/или заболевания крови, такие как гемофилии и гемоглобинопатии и тому подобное. См., например, патент США №20140017212 и 20140093913; патенты США №9,255,250 и 9,175,280.
[0132] Например, трансген может содержать последовательность, кодирующую полипептид, который отсутствует или нефункционален у субъекта, имеющего генетическое заболевание, включая, но без ограничений, любое из следующих генетических заболеваний: ахондроплазию, ахроматопсию, дефицит кислой мальтазы, дефицит аденозиндезаминазы (OMIM №102700), адренолейкодистрофию, синдром Экарди, дефицит антитрипсина альфа-1, альфа-талассемию, синдром нечувствительности к андрогенам, синдром Аперта, аритмогенную дисплазию правого желудочка, телеангиоэктатическую атаксию, синдром Барта, бета-талассемию, синдром голубого пузырчатого невуса, болезнь Канавана, хронические гранулематозные болезни (ХГБ), синдром кошачьего крика, муковисцидоз, болезнь Деркума, эктодермальную дисплазию, анемию Фанкони, фибродисплазию оссифицирующую прогрессирующую, синдром ломкой Х-хромосомы, галактоземию, болезнь Гоше, генерализированные ганглиозидозы (например, GM1), гемохроматоз, мутацию гемоглобина С в 6-м кодоне бета-глобина (HbC), гемофилию, болезнь Хантингтона, синдром Гурлера, гипофосфатазию, синдром Клайнфельтера, болезнь Краббе, синдром Лангера-Гиедиона, дефицит адгезии лейкоцитов (LAD, OMIM №116920), лейкодистрофию, синдром удлинения интервала QT, синдром Марфана, синдром Мебиуса, мукополисахаридоз (МПС), синдром ногтей-надколенника, нефрогенный несахарный диабет, нейрофиброматоз, болезнь Ниманна-Пика, несовершенный остеогенез, порфирию, синдром Прадера-Вилли, прогерию, синдром Протея, ретинобластому, синдром Ретта, синдром Рубинштейна-Тэйби, синдром Санфилиппо, тяжелый комбинированный иммунодефицит (ТКИД), синдром Швачмана, серповидно-клеточное заболевание (серповидно-клеточную анемию), синдром Смита-Магениса, синдром Стиклера, болезнь Тея-Сакса, синдром тромбоцитопении аплазии лучевой кости (TAR-синдром), синдром Тричера-Коллинза, трисомию, туберозный склероз, синдром Тернера, нарушение цикла мочевины, болезнь Гиппеля-Линдау, синдром Ваарденбурга, синдром Вильямса, болезнь Вильсона, синдром Вискотта-Олдрича, X-сцепленную лимфопролиферативную болезнь (XLP, OMIM №308240), иммунные приобретенные дефициты, лизосомные болезни накопления (например, болезнь Гоше, GM1, болезнь Фабри и болезнь Тея-Сакса), мукополисахаридозы (напримерболезнь Хантера, болезнь Гурлера), гемоглобинопатии (например, серповидно-клеточные заболевания, HbC, α-талассемию, β-талассемию) и гемофилии.
[0133] Неограничивающие примеры белков (включая их функциональные фрагменты, такие как усеченные версии), которые могут быть экспрессированы, как описано в данном документе, включают фибриноген, протромбин, тканевой фактор, фактор V, фактор VII, фактор VIII, фактор IX, фактор X, фактор XI, фактор XII (фактор Хагемана), фактор XIII (фибринстабилизирующий фактор), фактор Виллебранда, прекалликреин, высокомолекулярный кининоген (фактор Фицджеральда), фибронектин, антитромбин III, кофактор II гепарина, белок С, белок S, белок Z, белок Z-подобного ингибитора протеазы, плазминоген, альфа 2-антиплазмин, тканевой активатор плазминогена, урокиназу, ингибитор активатора плазминогена-1, ингибитор активатора плазминогена-2, глюкоцереброзидазу (GBA), α-галактозидазу А (ГЛА), идуронатсульфатазу (НДС), идуронидазу (IDUA), кислую сфингомиелиназу (SMPD1), ММАА, ММАВ, ММАСНС, MMADHC (C2orf25), MTRR, LMBRD1, MTR, пропионил-СоА-карбоксилазу (РСС) (субъединицы РССА и/или РССВ), белок транспортера глюкозо-6-фосфата (G6PT) или глюкозо-6-фосфатазу (Г6Фазу), рецептор ЛНП (РЛНП), АроВ, LDLRAP-1, PCSK9, митохондриальный белок, такой как NAGS (N-ацетилглутамат синтетазу), CPS1 (карбамоилфосфат синтетазу I) и ОТС (орнитинтранскарбамилазу), ASS (аргининосукцинат-синтетазу), ASL (аргининосукцинатлиазу) и/или ARG1 (аргиназу), и/или белок семейства транспортеров растворенных веществ типа 25 (SLC25A13, переносчик аспартата/глутамата), полипептид А1 глюкуронилтрансферазы UGT1A1 или UDP, фумарилацетоацетатгидролиазу (FAH), белок аланин-глиоксилат-аминотрансферазы (AGXT), белок глиоксилатредуктазы/гидроксипируватредуктазы (GRHPR), белок гена транстиретина (TTR), белок АТР7В, белок фенилаланингидроксилазы (ФАГ), белок липопротеинлиазы (ЛПЛ), сконструированную нуклеазу, сконструированный транскрипционный фактор и/или терапевтическое одноцепочечное антитело.
[0134] В определенных вариантах реализации изобретения трансген может содержать маркерный ген (описанный выше), позволяющий отбирать клетки, которые подверглись целенаправленной интеграции, и связанную последовательность, кодирующую дополнительные функциональные возможности. Неограничивающие примеры маркерных генов включают ЗФБ, маркер(ы) отбора по чувствительности к лекарственному препарату и тому подобное.
[0135] Конструкции, описанные в данном документе, могут также применяться для доставки некодирующих трансгенов. Последовательные кодирующие антисмысловые РНК, РНКи, shPHK и микроРНК (миРНК) также могут быть применены для целенаправленных инсерций.
[0136] В определенных вариантах реализации изобретения трансген включает последовательности (например, кодирующие последовательности, также называемые трансгенами) длиной более 1 т.п.н., например, от 2 до 200 т.п.н., от 2 до 10 т.п.н. (или любое значение в этом диапазоне). Трансген может также включать один или более целевых сайтов нуклеазы.
[0137] При интеграции (например, посредством опосредованной нуклеазой интеграции), трансген может быть вставлен в эндогенный ген таким образом, что все, некоторые или ни один эндогенный ген не экспрессируются.
Нуклеазы
[0138] Как отмечено выше, экспрессионные кассеты могут поддерживаться эписомально или могут быть интегрированы в геном клетки. Интеграция может быть случайной. Предпочтительно, чтобы интеграция конструкции (конструкций) трансгена нацеливалась расщеплением целевого гена одной или более нуклеазами (например, нуклеазами с цинковыми пальцами ("ZFN"), TALEN, TtAgo, системами нуклеаз CRISPR/Cas и хоуминг-эндонуклеазами) а конструкция была интегрирована либо с помощью репарации, направляемой гомологией (HDR), либо путем захвата конца во время процессов, запускающих негомологичное соединение концов (NHEJ). См., например, патенты США №9,255,250; 9,200,266; 9,045,763; 9,005,973; 9,150,847; 8,956,828; 8,945,868; 8,703,489; 8,586,526; 6,534,261; 6,599,692; 6,503,717; 6,689,558; 7,067,317; 7,262,054; 7,888,121; 7,972,854; 7,914,796; 7,951,925; 8,110,379; 8,409,861; публикации патентов США 20030232410; 20050208489; 20050026157; 20050064474; 20060063231; 20080159996; 201000218264; 20120017290; 20110265198; 20130137104; 20130122591; 20130177983; 20130196373 и 20150056705, которые включены в данное описание посредством ссылки в полном объеме для всех целей.
[0139] Любая нуклеаза может быть применена для целенаправленной интеграции экспрессионной конструкции трансгена.
[0140] В определенных вариантах реализации изобретения нуклеаза содержит нуклеазу с цинковыми пальцами (ZFN), которая содержит ДНК-связывающий домен цинкового пальца и домен расщепления (нуклеазы). См., например, патенты США №9,255,250; 9,200,266; 9,045,763; 9,005,973; 9,150,847; 8,956,828; 8,945,868; 8,703,489; 8,586,526; 6,534,261; 6,599,692; 6,503,717; 6,689,558; 7,067,317; 7,262,054; 7,888,121; 7,972,854; 7,914,796; 7,951,925; 8,110,379; 8,409,861.
[0141] В других вариантах реализации изобретения нуклеаза содержит TALEN, который включает в себя ДНК-связывающий домен эффектора TAL и домен расщепления (нуклеазы). См., например, патент США №8,586,526 и публикацию США №20130196373.
[0142] В дополнительных вариантах реализации изобретения нуклеаза содержит систему нуклеазы CRISPR/Cas, которая включает в себя одну гидовую РНК для распознавания целевого сайта и один или более доменов расщепления. См., например, публикация патента США №20150056705. В некоторых вариантах реализации изобретения применяется система CRISPR-Cpf1 (см. Fagerlund и соавт., (2015 год) Genom Bio 16:251). Понятно, что термин "система CRISPR/Cas" относится к системам CRISPR/Cas и CRISPR/Cfp1.
[0143] Нуклеазные домены расщепления могут быть дикого типа или мутированными, включая не встречающиеся в природе (сконструированные) домены расщепления, которые образуют облигатные гетеродимеры. См., например, патенты США №8,623,618; 7,888,121; 7,914,796; и 8,034,598 и патент США №20110201055.
[0144] Нуклеаза(ы) может сделать один или более двухцепочечных и/или одноцепочечных разрезов в целевом участке. В определенных вариантах реализации изобретения нуклеаза содержит каталитически неактивный домен расщепления (например, белок FokI и/или Cas). См., например, патенты США №9,200,266; 8,703,489 и Guillinger и соавт.(2014 год) Nature Biotech. 32(6):577-582. Каталитически неактивный домен расщепления может в сочетании с каталитически активным доменом действовать как никаза, чтобы сделать одноцепочечный разрез. Поэтому две никазы могут применяться в комбинации для создания двухцепочечного разреза в определенном регионе. В данной области техники известны дополнительные никазы, например, McCaffery и соавт. (2016 год) Nucleic Acids Res. 44(2):e11.doi:10.1093/nar/gkv878. Epub 19-го октября 2015 года.
[0145] В определенных вариантах реализации изобретения нуклеаза расщепляет "безопасный" ген (например, CCR5, Rosa, альбумин, AAVS1, и тому подобное. См., например, патенты США №7,888,121; 7,972,854; 7,914,796; 7,951,925; 8,110,379; 8,409,861; 8,586,526; Публикации патентов США 20030232410; 20050208489; 20050026157; 20060063231; 20080159996; 201000218264; 20120017290; 20110265198; 20130137104; 20130122591; 20130177983 и 20130177960. В предпочтительных вариантах реализации изобретения нуклеаза расщепляет эндогенный ген альбумина, так что экспрессионная кассета интегрируется в эндогенный локус альбумина клетки печени. Специфичные к альбумину нуклеазы описаны, например, в патенте США №9,150,847; и публикациях США №20130177983 и 20150056705.
Доставка
[0146] Конструкции, описанные в данном документе (и/или нуклеазы), могут быть доставлены in vivo или ex vivo любым подходящим способом в любой тип клеток, предпочтительно в печень (доставка в печень). Аналогичным образом, при применении в сочетании с нуклеазами для целенаправленной интеграции, нуклеазы могут быть доставлены в форме полинуклеотида и/или белка, например, с применением невирусного вектора(ов), вирусного вектора(ов) и/или в форме РНК, например, в виде мРНК.
[0147] Способы невирусной доставки нуклеиновых кислот включают электропорацию, липофекцию, микроинъекцию, биолистику, виросомы, липосомы, иммунолипосомы, другие наночастицы, конъюгаты "поликатион или липид : нуклеиновая кислота", оголенную ДНК, искусственные вирионы и усиленное агентом поглощение ДНК. Для доставки нуклеиновых кислот также может применяться порация ультразвуком, например, система Sonitron 2000 (Rich-Mar). Дополнительные типовые системы доставки нуклеиновой кислоты включают системы, предоставляемые AmaxaBiosystems (Кельн, Германия), Maxcyte, Inc. (Роквилл, штат Мэриленд), системы молекулярной доставки ВТХ (Холлистон, штат Массачусетс) и Copernicus Therapeutics Inc., (см., например, US 6008336).
[0148] В предпочтительных вариантах реализации изобретения экспрессионная конструкции представляют собой векторы AAV. Необязательные нуклеазы могут вводиться в форме мРНК или с применением одного или более вирусных векторов (AAV, Ad и тому подобного). Введение может осуществляться любым способом, в котором полинуклеотиды доставляются в желаемые целевые клетки. Предполагаются как in vivo, так и ex vivo способы. Внутривенная инъекция в портальную вену представляет собой предпочтительный метод введения. Другие in vivo режимы введения включают, например, прямую инъекцию в доли печени или желчную протоку и внутривенную инъекцию, дистально по отношению к печени, в том числе через печеночную артерию, прямую инъекцию в паренхиму печени, инъекцию через печеночную артерию и/или ретроградную инъекцию через панкреатическое дерево. Ex vivo режимы введения включают трансдукцию in vitro резецированных гепатоцитов или других клеток печени с последующей инфузией трансдуцированных резецированных гепатоцитов обратно в портальную васкулатуру, паренхиму печени или билиарное дерево пациента-человека, см., например, Grossman и соавт., (1994 год) Nature Genetics, 6:335-341.
[0149] В системах, включающих доставку более одного полинуклеотида (например, конструкция, как описано в данном документе, и нуклеаза в полинуклеотидной форме), два или более полинуклеотида(ов) доставляются с применением одного или более одинаковых и/или разных векторов. Например, нуклеаза в полинуклеотидной форме может быть доставлена в форме мРНК, а печень-специфичные конструкции, как описано в данном документе, могут быть доставлены с помощью других средств, таких как вирусные векторы (например, AAV), миникольцевая ДНК, плазмидная ДНК, линейная ДНК, липосомы, наночастицы и тому подобное.
[0150] Фармацевтически приемлемые носители частично определяются конкретной вводимой композицией, а также конкретным способом, применяемым для введения указанной композиции. Соответственно, существует широкий спектр пригодных составов доступных фармацевтических композиций, как описано ниже (см., например, Remington's Pharmaceutical Sciences, 17e изд., 1989 год).
[0151] Эффективное количество экспрессионной кассеты (и необязательной нуклеазы(нуклеаз) и/или модифицированных клеток), подлежащей введению, будет варьироваться от пациента к пациенту. Соответственно, эффективные количества лучше всего определит лечащий врач, назначающий указанные композиции (например, клетки), а соответствующие дозы сможет легко определить специалист в данной области техники. Анализ уровня терапевтического полипептида в сыворотке, плазме или других тканях, а также сравнение с начальным уровнем до введения, может определить, является ли вводимое количество слишком низким, находится в правильном диапазоне, или является слишком высоким. Пригодные схемы для начального и последующего введений также являются вариабельными, но типичны для начального введения, после которого при необходимости осуществляют последующие введения. Последующие введения могут осуществляться с переменными интервалами: от ежедневного введения до ежегодного введения, а также введения один раз в нескольких лет. Специалисту в данной области техники будет понятно, что подходящие иммуносупрессивные методы могут быть рекомендованы для предотвращения ингибирования или блокирования трансдукции иммуносупрессией векторов доставки, см., например, Vilquin и соавт., (1995 год) Human Gene Ther., 6:1391-1401.
[0152] Составы для введения как ex vivo, так и in vivo включают суспензии (например, генетически модифицированных клеток, липосом или наночастиц) в жидких или эмульгированных жидкостях. Активные ингредиенты часто смешивают с наполнителями, которые являются фармацевтически приемлемыми и совместимыми с активным ингредиентом. Подходящие наполнители включают, например, воду, физиологический раствор, декстрозу, глицерин, этанол или тому подобное и их комбинации. Кроме того, композиция может содержать незначительные количества вспомогательных веществ, таких как смачивающие или эмульгирующие агенты, рН-буферные средства, стабилизирующие агенты или другие реагенты, которые повышают эффективность фармацевтической композиции.
Сферы применения
[0153] Способы и композиции, раскрытые в данном документе, предназначены для обеспечения терапии любого заболевания путем предоставления трансгена, который экспрессирует продукт, который отсутствует или присутствует в недостаточных количествах при заболевании, или иным образом лечит или предотвращает болезнь. Клетка может быть модифицирована in vivo или может быть модифицирована ex vivo и затем введена субъекту. Таким образом, способы и композиции обеспечивают лечение и/или профилактику таких генетических заболеваний.
[0154] Следующие примеры включают типовые варианты реализации данного изобретения, в которых необязательно используемая нуклеаза содержит нуклеазу с цинковыми пальцами (ZFN). Следует понимать, что примеры приведены только в целях иллюстрации и что можно применять другие нуклеазы, например, TALEN, системы CRISPR/Cas, хоуминг-эндонуклеазы (мегануклеазы), имеющие сконструированные ДНК-связывающие домены и/или слияния ДНК-связывающих доменов хоуминг-эндонуклеаз (мегануклеаз) естественного происхождения и гетерологичных доменов расщепления и/или слияния мегануклеаз и белков TALE. Кроме того, будет понятно, что экспрессионные конструкции, как описано в данном документе, могут переноситься на другие векторы (отличные от AAV) для получения тех же результатов при лечении и/или профилактике нарушений, вызванных недостаточным продуцированием белка.
ПРИМЕРЫ Пример 1: Способы Конструкции ДНК
[0155] Общая структура и подход, применяемый для разработки иллюстративных конструкций, содержащих кДНК фактора VIII человека, изображены на Фиг. 1. Конструкции включают печень-специфичный энхансер, печень-специфичный промотор, факультативный интрон, последовательности инсулятора и трансген (например, кодон-оптимизированный трансген фактора VIII человека с удаленным В-доменом), синтетическая последовательность полиаденилирования, спейсер и 5'/3' инвентированные концевые повторы. Все конструкции были собраны с использованием стандартных способов клонирования молекулярной биологии.
[0156] Как показано на Фиг. 1, во время разработки вектора выполнялось несколько этапов. Сначала мы исследовали влияние компонентов конструкции на выход вируса, полученный в процессе производства. Влияние последовательности областей инсулятора в конструкциях изучалось с использованием трех потенциальных последовательностей инсулятора, как показано: Инс1: 5' GGAACCATTGCCACCTTCA (SEQ ID NO: 28), Инс2: 5' CTATCCATTGCACTATGCT (SEQ ID NO: 29), и Инс3: 5' TTTCCTGTAACGATCGGG (SEQ ID NO: 30). Затем области инсулятора были вставлены в экспрессионную кассету трансгена, где инсулятор, который использовался между 5' ИКП последовательностью и началом последовательности энхансера/промотора, всегда был Инс1. Однако последовательности Инс2 либо Инс3 были связаны на 3'-конце конструкции с синтетической поли А последовательностью SPA51 (5' AATAAAATATCTTTATTTTCATTACATCTGTGTGTTGGTTTTTTGTGTGTT, SEQ ID NO: 31) следующим образом: Hhc2-SPA51: 5' ATAAAATATCTTTATTTTCATTACATCTGTGTGTTGGTTTTTTGTGTGTTCTATCCATT GCACTATGCT, SEQ ID NO: 32, и Hhc3-SPA51: 5' AATAAAATATCTTTATTTTCATTACATCTGTGTGTTGGTTTTTTGTGTGTTTTCCTGTA ACGATCGGG, SEQ ID NO: 33. В некоторых вариантах реализации изобретения Инс4 связан с SPA51.
[0157] Варианты последовательности Инс также использовались в экспрессионных конструкциях, где область промотора SerpE-TTRm-MVM была заменена гибридным промотором печени (HLP - hybrid liver promoter, McIntosh и соавт., там же). Эти конструкции изображены на Фиг. 2. Эти конструкции затем упаковывали в частицы AAV2/6 с использованием клеток HEK293, где выходы были получены из двух клеточных фабрик (Yuan и соавт., (2011) Hum Gene Ther., 22, 613-624, 2011 год). Было обнаружено, что выход (векторные геномы) для конструкций, содержащих модифицированные последовательности инсулятора, почти на три log выше для конструкции, содержащей исходную область промотора, где экспрессионная кассета была фланкирована областями инсулятора Инс1 и Инс3 на 5' и 3' концах, соответственно (Фиг. 3). Кроме того, эти последовательности инсулятора также были вставлены в модифицированные конструкции CRMSBS1 и CRMSBS2, описанные ниже на этапе 2 (Фиг. 19 и 20), и были протестированы на вирусный выход (Фиг. 22). Данные показали 8-10-кратное улучшение вирусного выхода для новой конструкции по сравнению с исходной конструкцией.
[0158] На этапе 2 точечные мутации вводили в энхансер Серпин 1 для создания первых двух производных векторов CRMSBS1 и CRMSBS2 (Фиг. 4). Эти векторы различаются по своим энхансерным последовательностям, где были введены точечные мутации, как показано на Фиг. 5. Конкретные изменения заключаются в следующем: для CRMSBS1, обозначены как 1, 2, 3 и 4, как показано на Фиг. 5, были сделаны следующие замены, соответственно: 1=G на А, 2=С на G, 3=Т на С и 4=G на А. Аналогично, для CRMSBS2 позиции 1, 3, 4 и 5 были изменены следующим образом: 1=G на А, 3=Т на С, 4=G на А и 5=С на Т.
[0159] На этапе 3 были сделаны конструкции для исследования последовательности интрона в векторе. Как изображено на Фиг. 6 и 7, последовательности интрона в векторах CRMSBS2 и CRMSBS2 были модифицированы путем удаления интрона MVM ("CRMSBS1 Без интрона", Фиг. 6, и "CRMSBS2 Без интрона", Фиг. 7). В другом варианте была протестирована Т-химерная последовательность интрона ("Т-химерный интрон CRMSBS1", Фиг. 6, и "Т-химерный интрон CRMSBS2", Фиг. 7).
[0160] Для этапа 4 были внесены изменения в интрон MVM путем введения мутаций, нацеленные на сайты донора и акцептора сплайсинга. Карты этих векторов изображены на Фиг. 10, где измененные интроны называются SBR Интрон 1-3. Интрон MVM имеет два потенциальных донорных сайта и два потенциальных акцепторных сайта этих доноров (для иллюстрации см. Фиг. 11). На Фиг. 11 два донорных сайта обозначены D1 и D2, а их соответствующие акцепторные сайты изображены как А1 и А2. Границы сплайсинга, образованные в результате сплайсинга D1-A1 и D2-A2, изображены над геном в виде пунктирных линий. Частичная последовательность интрона MVM изображена под геном на Фиг. 11 (SEQ ID NO: 14) и также указано местоположение (подчеркнуто) последовательности интронного энхансера (IES - intronic enhancer sequence) и сайтов акцепторов А1 и А2 (Haut и Pintel, (1998) J Virol, 72:1834-1843; Haut и Pintel, (1998) Virol J, 258:84-94). Три интрона были сконструированы для тестирования, и они изображены на Фиг. 10 и обозначены как SBR Интрон 1-3 (последовательности интрона указаны в SEQ ID NO: 15-17). Активность экспрессированного чFVIII, доставленного с помощью кДНК AAV F8, также оценивали in vitro. кДНК AAV2/6 F8 (CRMSBS2 SBR Интрон 3) добавляли к клеткам HepG2 в дозах 4,8Е+06, 2,4Е+06, 1,2Е+06 и 0,6Е+06 векторных геномов/мл на 1Е+05 клеток в 24-луночной чаше (обозначается как время, t0 дней). Супернатанты анализировали на уровни секретируемого чFVIII с помощью ИФА и проводили анализ АЧТВ (активированное частичное тромбопластиновое время) коагулирующей активности и хромогенный анализ активности в течение семи дней (t7) после добавления вируса AAV2/6 (способы см. ниже). Результаты (Фиг. 21) продемонстрировали хорошую корреляцию между уровнями и активностью секретируемого чFVIII. Показанные данные представляют собой n=6 биологических повторов. "Усы" погрешностей представляют собой стандартную ошибку среднего повторов.
[0161] Затем эти конструкции были протестированы in vivo, как описано ниже.
Количественная ПЦР
[0162] кОТ-ПЦР (для уровней мРНК фактора VIII человека): Ткань мыши лизировали с использованием FastPrep и Lysing Matrix D (MP Biomedicals, Санта-Ана, штат Калифорния) в соответствии с инструкциями производителя. РНК/ДНК выделяли из ткани мыши с использованием набора ДНК/РНК AllPrep согласно инструкциям производителя (Qiagen, Карлсбад, штат Калифорния). Затем экстрагированную РНК использовали для создания кДНК с использованием набора для синтеза кДНК Quantitect (Qiagen, Карлсбад, штат Калифорния). Затем количественную ПЦР проводили с использованием универсального супермикса SsoAdvanced Univesal Probes Supermix (Biorad, Геркулес, штат Калифорния) на Biorad CFX 96, используя анализы с меченым праймером/зондом от IDT (Коралвиль, штат Айова). Анализ GAPDH мыши являлся Mm.PT.39a.1. Для специфического обнаружения мРНК фактора VIII человека обычным был анализ с использованием праймера/зонда; Прямой праймер (GGAGATGAAGAAGGAGGACTTTG) (SEQ ID NO: 18), зонд (ACATCTACGACGAGGACGAGAACCA) (SEQ ID NO: 19) и Обратный праймер (TCCACAGCAGCAATGAAGTAG) (SEQ ID NO: 20). Количественная кОТ-ПЦР (не абсолютная) использовалась с нормализацией к GAPDH для каждого образца, а окончательный анализ данных приводился как относительный к одному образцу мыши, который был установлен в 1.0. Контроль без матрицы и контроли без обратной транскриптазы были выполнены со всеми образцами и не показывали детектируемого сигнала.
[0163] кПЦР (для анализов генома вектора, ГВ): Ткань мыши лизировали с использованием FastPrep и Lysing Matrix D (MP Biomedicals, Санта-Ана, штат Калифорния) в соответствии с инструкциями производителя. РНК/ДНК выделяли из ткани мыши с использованием набора ДНК/РНК AllPrep согласно инструкциям производителя (Qiagen, Карлсбад, штат Калифорния). Экстрагированную ДНК использовали для количественной ПЦР с помощью мастер-микса TaqMan Fast Universal PCR Master Mix, без AmpErase UNG (Applied Biosystems, Фотер Сити, штат Калифорния) в системе ПЦР в реальном времени АВ 7300 (Applied Biosystems, Фотер Сити, штат Калифорния). Для специфического обнаружения фактора VIII человека обычным был анализ с использованием праймера/зонда; прямой праймер (CCTGGGCCAGTTCCTGCT) (SEQ ID NO: 21), зонд (TTCTGCCACATCAGCAGCCACCA) (SEQ ID NO: 22) и обратный праймер (GGCCTCCATGCCATCATG) (SEQ ID NO: 23). Контроли без матрицы были выполнены со всеми образцами и не показывали детектируемого сигнала. Стандартная кривая ДНК кПЦР генерировалась из семи последовательных 4-кратных разведений известного количества очищенной, линеаризованной плазмиды фактора VIII человека.
[0164] кПЦР (для целевой интеграции посредством NHEJ с использованием клеток HepG2) - ДНК была выделена из человеческих клеток HepG2 с использованием QIAamp DNA micro в соответствии с инструкциями производителя (Qiagen, Карлсбад, штат Калифорния). Затем количественную ПЦР проводили с применением универсального супермикса SsoAdvanced Univesal Probes Supermix (Biorad, Геркулес, штат Калифорния) на Biorad CFX 96, используя анализы с меченым праймером/зондом от IDT (Коралвиль, штат Айова). Анализ GAPDH мыши представлял собой Hs.PT.39a.22214836. Для специфического обнаружения целевой интеграции фактора VIII человека посредством NHEJ в эндогенном локусе альбумина человека обычным был анализ с использованием праймера/зонда; Прямой праймер (AGTGCAAAGTAACTTAGAGTGACT) (SEQ ID NO: 24), зонд (CCATCACTAGGGGTTCCTGCGGCCT) (SEQ ID NO: 25) и Обратный праймер (CCTGAAGGTGGCAATGGT) (SEQ ID NO: 26). Для целей данного исследования мы использовали количественную ПЦР (не абсолютную) с нормализацией к GAPDH для каждого образца, а окончательный анализ данных приводился как относительный к одному образцу, который был установлен в 1.0. Контроль без матрицы, контроль без транскриптазы и контроли без ZFN были выполнены и не показывали детектируемого сигнала.
[0165] кПЦР (для целевой интеграции в ткани мыши посредством NHEJ и HDR) - Ткань мыши лизировали с использованием FastPrep и Lysing Matrix D (MP Biomedicals, Санта-Ана, штат Калифорния) в соответствии с инструкциями производителя. РНК/ДНК выделяли из ткани мыши с использованием набора ДНК/РНК AllPrep согласно инструкциям производителя (Qiagen, Карлсбад, штат Калифорния). Затем количественную ПЦР проводили с использованием универсального супермикса SsoAdvanced Univesal Probes Supermix (Biorad, Геркулес, штат Калифорния) на Biorad CFX 96, используя анализы с меченым праймером/зондом от IDT (Коралвиль, штат Айова). Анализ GAPDH мыши являлся Mm.PT.39a.1. Для специфического обнаружения целевой интеграции кДНК фактора 8 человека, произведенной посредством NHEJ, в эндогенном локусе альбумина мыши обычным был анализ с использованием праймера/зонда; Прямой праймер (GTGTAGCAGAGAGGAACCATT, SEQ ID NO: 39), зонд (CCATCACTAGGGGTTCCTGCGGCCT, SEQ ID NO: 40) и Обратный праймер (GTTAATATTCACCAGCAGCCT, SEQ ID NO: 41). Для специфического обнаружения целевой интеграции ZFN посредством NHEJ в эндогенном локусе альбумина мыши обычным был анализ с использованием праймера/зонда; Прямой праймер (AGTGTAGCAGAGAGGAACCA, SEQ ID NO: 42), зонд (CCATCACTAGGGGTTCCTGCGGCCT, SEQ ID NO: 43) и Обратный праймер (CAGGGTGAGCCCAGAAAC, SEQ ID NO: 44). Для специфического обнаружения целевой интеграции фактора 8 человека посредством HDR в эндогенном локусе альбумина мыши обычным был анализ с использованием праймера/зонда; Прямой праймер (AACTTTGAGTGTAGCAGAGAGG, SEQ ID NO: 45), зонд (TACCGGAGGAGCAAACAGGGACTA, SEQ ID NO: 46) и Обратный праймер (CTCTACGAAATGTGCAGACAGA, SEQ ID NO: 47). Для целей данного исследования мы использовали количественную кПЦР (не абсолютную) с окончательным анализом данных, приведенным как относительный к одному образцу, который был установлен в 1.0. Контроль без матрицы, контроль без транскриптазы и контроли без ZFN были выполнены и не показывали детектируемого сигнала.
[0166] Индел (инсерции и делеции) - Ткань мыши лизировали с использованием FastPrep и Lysing Matrix D (MP Biomedicals, Санта-Ана, штат Калифорния) в соответствии с инструкциями производителя. РНК/ДНК выделяли из ткани мыши с использованием набора ДНК/РНК AllPrep согласно инструкциям производителя (Qiagen, Карлсбад, штат Калифорния). Экстрагированную ДНК использовали для ПЦР и глубокого секвенирования для измерения инделов в локусе альбумина мыши.
[0167] Плазма - Кровь собирали у всех мышей на День 7, 14, 21 (без летального исхода) и на День 28 или дольше, как указано (с летальным исходом) для исследований на C57BL/6. Кровь собирали у всех мышей на День 8, 14, 21, 28, 35 и 42 (без летального исхода) для исследований на мышах с гемофилией А. Вся кровь была собрана в пробирки, содержащие цитрат натрия, и перерабатывалась в плазму. Заборы крови с не летальным исходом собирали через подчелюстную вену или ретро-орбитальную пазуху. Кровь во время умерщвление подопытного животного была собрана с помощью сердечной пункции или из полой вены. Плазму отделяли и хранили при температуре от -60 до -80°С до применения в анализе ИФА или анализе активности Chromogenix Coamatic, описанном ниже.
[0168] Печень - Для исследований на C57BL/6, мышей умерщвляли на 28-й день, а печень, селезенку, яички, мозг, сердце, легкие и почки собирали и взвешивали. Левую боковую долю печени отделяли и делили на 3 части и быстро замораживали в жидком азоте отдельно от остальной части печени. Оставшиеся доли печени и другие ткани (целые) быстро замораживали в жидком азоте. Замороженные образцы хранили при температуре от -60 до -80°С до обработки для экстракции РНК/ДНК.
[0169] Исследования In vitro, HepG2 AAV F8 кДНК/ZFN. Человеческие клетки печени HepG2 поддерживались в соответствии с рекомендациями производителя (АТСС, Манассас, штат Виргиния). В день эксперимента клетки HepG2 промывали, трипсинизировали и подсчитывали. Применяемые ZFN были специфичными к локусу интрона человеческого альбумина, слева SBS47171 и справа SBS47898. ZFN вместе с кДНК 4F8 (CRMSBS2 Без Интрона) были доставлены в виде AAV2/6, и обозначены как нулевой момент времени. AAV2/6 ZFN были доставлены в количестве 3,0Е+05, a AAV2/6 кДНК чFVIII CRMSBS2 Без интрона в количестве 3,0Е+04, 6,0Е+04 и 1,2Е+05 для 1Е+05 клеток на лунку 24-луночного планшета. На следующий день среды меняли. Супернатанты анализировали на секретируемый чFVIII с использованием ИФА чFVIII, описано ниже, в точки времени t3, t5 и t7 дней после добавления вируса AAV2/6.
[0170] Исследования in vitro, HepG2 кДНК AAV F8. Человеческие клетки печени HepG2 поддерживались в соответствии с рекомендациями производителя (АТСС, Манассас, штат Виргиния). В день эксперимента клетки HepG2 промывали, трипсинизировали и подсчитывали. кДНК AAV2/6 чF8 (CRMSBS2 SBR Интрон 3) была доставлена в дозах 6,0Е+06, 1,2Е+06, 2,4Е+06 и 4,8Е+06 на 1Е+05 клеток в лунке 24-луночного планшета. Обмен носителями осуществлялся в момент времени t3 (t=день). Супернатанты анализировали в точки времени t5 и t7 после добавления AAV 2/6 вируса на уровни секретируемого чFVIII с использованием ИФА 4FVIII от Affinity Biologies, и на активность чFVIII с помощью хромогенного анализа и анализа коагулирующей активности, описанных ниже.
[0171] ИФА фактора VIII человека. ИФА чFVIII Affinity Biologies (Мышь и клетки HepG2). Уровни секретируемого человеческого фактора VIII определяли с использованием набора ИФА Affinity Biologicals (Канада) (FVIII-AG) в соответствии с протоколом производителя, за исключением стандарта фактора VIII человека. Стандарт человеческого фактора VIII, применяемый в анализе ИФА, представляет собой рекомбинантный очищенный человеческий фактор VIII (#F0016-06) от US Biologicals (Сейлем, штат Массачусетс). Коротко, плазму мыши добавляли в планшет, инкубировали покачивая при комнатной температуре в течение полутора часов с последующим промыванием три раза промывочным буфером, предусмотренным в наборе. Антитело для обнаружения, предоставляемое с набором, добавляли и инкубировали в течение сорока пяти минут при комнатной температуре с последующим промыванием три раза промывочным буфером, предусмотренным в наборе. Добавляли субстрат ТМБ, предусмотренный набором, и оставляли на 10 минут. Реакцию останавливали останавливающим раствором и считывали поглощение при 450 нМ с помощью сканирующего спектрофотометра для прочтения планшетов.
[0172] Штатная ИФА чFVIII (яванский, макак). Уровни чFVIII, секретируемые в плазме NHP, определяли с использованием стандартной ИФА. Полистирольные микропланшеты (Corning, 96-луночные половинчатые с высокой степенью связывания) покрывали в течение ночи при 4°С моноклональным антителом к чFVIII мыши (Green Mountain, Берлингтон, штат Вермонт) в 0,2 М карбонатного бикарбонатного буфера при рН 9,4 (Thermo Fisher Scientific, Уолтем, штат Массачусетс). На следующий день планшеты промывали четыре раза с использованием 1-кратного TBST (Thermo Fisher Scientific, Уолтем, штат Массачусетс). 96-луночные планшеты затем блокировали два часа при комнатной температуре, используя 3% блокирующего буфера БСА/TBS, а затем промывали четыре раза 1-кратным TBST. Плазму добавляли в планшет и инкубировали, покачивая при комнатной температуре в течение двух часов с последующим промыванием четыре раза 1-кратным TBST. Добавляли детекторное антитело, биотинилированное моноклональное антитело к FVIII мыши (Green Mountain, Берлингтон, штат Вермонт), и инкубировали в течение одного часа при комнатной температуре, а затем промывали четыре раза 1-кратным TBST. Стрептавидин HRP (пероксидаза хрена) (Jackson ImmunoResearch, Вест Гров, штат Пенсильвания) добавляли и инкубировали в течение одного часа при комнатной температуре, а затем промывали четыре раза 1-кратным TBST. Добавляли ТМВ Ultra (Thermo Fisher Scientific, Уолтем, штат Массачусетс) и выдерживали в течение десяти минут, реакцию останавливали останавливающим раствором и измеряли оптическую плотность при 450 нМ с помощью сканирующего спектрофотометра для прочтения планшетов. Показания фоновой абсорбции были незначительными (обычно 0).
[0173] Хромогенный анализ активности фактора VIII человека. Активность секретируемого человеческого фактора VIII в плазме определяли с использованием анализа Diapharma Chromogenic Coamatic Factor VIII (Уэст-Честер, штат Огайо) в соответствии с протоколом производства, за исключением стандарта фактора VIII человека. Стандарт человеческого фактора VIII, применяемый в анализе ИФА, представляет собой рекомбинантный очищенный человеческий фактор VIII (#F0016-06) от US Biologicals (Сейлем, штат Массачусетс).
[0174] Анализ коагулирующей активности Активность секретируемого человеческого фактора VIII в супернатанте HepG2 (или плазме яванского макака) определяли, используя временной анализ активированного частичного тромбопластина (аЧТВ) с помощью Diagnostica Stago (Бостон, штат Массачусетс) в соответствии с протоколом производства, за исключением стандарта фактора VIII человека и плазмы с дефицитом фактора VIII человека. Стандарт фактора VIII человека был таким же, как применяемый в анализе ИФА (рекомбинантный очищенный человеческий фактор VIII, #F0016-06 от US Biologicals, Сейлем, штат Массачусетс). Реактив с дефицитным FVIII, используемый в анализ коагулирующей активности, представлял собой FVIII-CD<1% активности FVIII (замороженный, дефицитный FVIII) от Haematologic Technologies, Inc. (Эссекс Джанкшен, штат Вермонт). Коротко, стандарт или образец добавляли в кюветы, содержащие стальной шарик. Добавляли FVIII-CD<1% активности FVIII и РТТ Automate, предоставляемый с набором, инкубировали при 37°С в течение ста восьмидесяти секунд. STA CaCl2, предоставляемый с набором, добавляли в реакцию в то время, как стальной шарик приходил в движение в кювете. Время коагуляции измеряли в течение от добавления STA CaCl2 до тех пор, пока движение стального шарика не прекращается. Инкубацию и запись времени для этого анализа проводили с использованием анализатора гемостаза Start Stago.
Пример 2: Исследования in vivo
[0175] Мыши дикого типа: В исследованиях in vivo использовали мышей C57BL/6 возрастом от восьми до десяти недель. Исследование проводилось в соответствии с законом о благополучии животных для гуманного ухода и использования животных. Тестовые препараты (конструкции, содержащие вирусы AAV) оттаивали при комнатной температуре до дозирования, и все животные получали одну внутривенную инъекцию (в/в) 200 мкл. В Таблице 1 ниже показана схема исследования для исследования конструкций in vivo на Фиг. 8 (изображены на Фиг. 4, 6 и 7). В Таблице 2 показана схема исследования для исследования конструкций, изображенных на Фиг. 15. Дозы составляли 1,8Е+11 гв на мышь, что составляло примерно 7Е+12 гв/кг. Все животные получали последующую внутрибрюшинную инъекцию 200 мкл циклофосфамида на День 0 и 14. Заборы крови с летальным и не летальным исходами были выполнены, как указано в Таблицах 1 и 2.


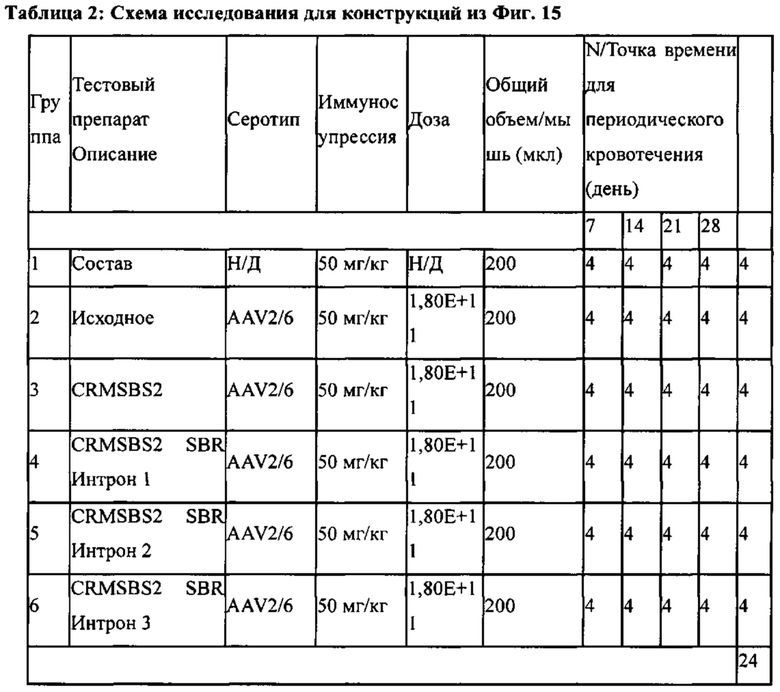
[0176] Кровь собирали у всех мышей на День 7, 14, 21 (без летального исхода) и на День 28 (с летальным исходом). Вся кровь была собрана в пробирки, содержащие цитрат натрия, и перерабатывалась в плазму. Заборы крови с не летальным исходом собирали через подчелюстную вену или ретро-орбитальную пазуху. Кровь во время умерщвления подопытного животного была собрана с помощью сердечной пункции или из полой вены. Плазму отделяли и хранили при температуре от -60 до -80°С до применения в анализе ИФА, описанном ниже.
[0177] Всех животных умерщвляли на 28-й день, а печень, селезенку, яички, мозг, сердце, легкие и почки собирали и взвешивали. Левую боковую долю печени отделяли и делили на 3 части и быстро замораживали в жидком азоте отдельно от остальной части печени. Оставшиеся доли печени и другие ткани (целые) быстро замораживали в жидком азоте. Замороженные образцы хранили при температуре от -60 до -80°С до обработки для экстракции РНК/ДНК.
[0178] Как изображено на Фиг. 8 и 15, после введения все тестируемые конструкции приводили к in vivo продуцированию трансгена (секретируемого фактора VIII человека с удаленным В-доменом) in vivo на 7-й день (Фиг. 8А и Фиг. 15А) и 28-й день (Фиг. 8В и Фиг. 15В). Кроме того, как изображено на Фиг. 9 и 16, трансген экспрессировался только в печени, и не экспрессировался в других тканях (Фиг. 9А и 16А) и вектор AAV трансдуцирует в основном клетки печени (анализ биораспределения генома вектора изображен на Фиг. 9В и 16В). В этих наборах данных мРНК человеческого фактора VIII-BDD (чFVIII-BDD) анализировали из тканей (головного мозга, сердца, почек, легких, печени, селезенки, яичек) в исследовании, представленном в Таблице 1, с помощью кОТ-ПЦР, как описано выше в Примере 1. Кроме того, распределение генома вектора анализировалось в этих же тканях с помощью кПЦР (см. Пример 1), а результаты изображены на Фиг. 9В/16В. Эти данные свидетельствуют о том, что вектор AAV2/6 в основном трансдуцирует печень.
[0179] Конструкции, сконструированные для изучения эффективности модификаций интронов, были протестированы in vivo, а также в исследовании, приведенном в Таблице 2, а данные за 7 дней и на пиковых уровнях изображены на Фиг. 15. мРНК чFVIII-BDD также была проанализирована (Фиг. 16А), как и распределение генома вектора (Фиг. 16 В), и показала, что экспрессия мРНК чF8 специфична для печени, a AAV2/6 преимущественно трансдуцирует печень.
[0180] Таким образом, описанные в данном документе конструкции обеспечивают надежную экспрессию трансгена после доставки в печень.
[0181] Было проведено дополнительное исследование, чтобы проверить, были ли наблюдаемые уровни чFVIII-BDD in vivo чувствительны к серотипу применяемого AAV. Самцам мышей C57BL/6 внутривенно вводили 6Е+10 гв/мышь (~2Е+12 гв/кг) или 1,8Е+11 гв/мышь (~7Е+12 гв/кг) исходной кДНК F8 AAV2/5, AAV2/6, AAV2/8 или AAV2/9. Было достигнуто трансдуцирование мыши с серотипом AAV2/6, которые, как известно, неэффективны при трансдукции печени мыши, при 6Е+10 гв/мышь (~2Е+12 гв/кг) со средним значением 91,9% +/- 15,5 СОС (n=6) от нормального уровня чFVIII в плазме у людей. При более высокой дозе, составляющей ~7Е+12 гв/кг было достигнуто среднее пиковое значение 4FVIII плазмы после шести независимых исследований мыши in vivo 169,2% +/- 10,1 СОС (n=36). Средние пиковые уровни для AAV2/8 при 2Е+12 гв/кг составляли 320% и 323,6% при 6Е+12 гв/кг. Для AAV2/9 при 2Е+12 гв/кг средний пиковый уровень составлял 389,6% (см. Фиг. 23, также изображающую результаты с применением AAV2/5 и AAV2/6).
[0182] Были также проведены исследования для определения того, имел ли способ получения, применяемый для создания AAV, воздействие на полученную экспрессию чFVIII-BDD in vivo. Самцам мышей C57BL/6 вводили исходную кДНК F8 путем внутривенной инъекции (в хвостовую вену) 1,8Е+11 гв/мышь (~7Е+12 гв/кг) исходной кДНК AAV2/6 F8, продуцируемой либо клетками HEK293, либо Sf9/rBV (рекомбинантный бакуловирус). Обработка исходной кДНК F8 из клеток HEK293 достигала среднего пикового значения 142,1%±8,3% СОС (n=8) (измерено с помощью ИФА чFVIII) от нормальных уровней человеческого FVIII в плазме. Аналогичный уровень 132,8%±18,6% СОС (n=5) (измерено с помощью ИФА чFVIII) был достигнут после введения мышам исходной кДНК F8 (Sf9/rBV) (Фиг. 24).
[0183] Исследования были повторены для сравнения с кДНК F8 CRMSBS2 SBR Интрон 3 (см. Фиг. 10). Самцам мышей C57BL/6 вводили внутривенно 1,8Е+11 гв/мышь (~6Е+12 гв/кг) кДНК AAV2/6 CRMSBS2 SBR Интрон 3 (n=8). Показаны средние пиковые уровни 4FVIII в плазме мышей C57BL/6, измеренные с помощью ИФА чFVIII (Фиг. 25).
[0184] Мыши, страдающие гемофилией А: Конструкции также тестировали на мышиной модели R593A гемофилии A (Bril и соавт.(2006 год) Thromb Haemost 95(2):341-7; Chavez и соавт., (2012 год) Hum Gen Ther., 23(4):390). Эти мыши имели инактивацию эндогенного гена F8 мыши. Кроме того, они были носителями мутантного трансгена FVIII-R593C человека под контролем промотора мышиного альбумина, так что они экспрессировали необнаруживаемые количества мутантного человеческого белка, но считались толерантными к экспрессии FVIII человека из-за незначительного количества произведенного мутантного белка FVIII R593A. Первоначально линия была FVB, но была обратно скрещена с мышами C57BL/6 в течение по меньшей мере 5 поколений в Jackson Laboratory. Исследование проводилось в соответствии с законом о благополучии животных для гуманного ухода и использования животных. Тестовые препараты оттаивали при комнатной температуре до дозирования, и все животные получали одну внутривенную инъекцию (в/в) 200 мкл. Дозы составляли 1,8Е+11 гв на мышь, что составляло примерно 7Е+12 гв/кг. Схема исследования показана ниже в Таблице 3:
Таблица 3: CRMSBS2 SBR Интрон 3 в мутантных мышах, страдающих гемофилией А

[0185] Исследование проводилось в течение трех месяцев, показаны репрезентативные результаты трехмесячного исследования, результаты как 14-го дня, так и 42-го дня (Фиг. 17) показывают, что трансген FVIII-BDD экспрессировался на уровнях >300% от уровней белка FVIII, обнаруживаемого в нормальной плазме человека в оба дня 14 (Фиг. 17А) и 42 (Фиг. 17 В).
[0186] Для тестирования функциональности и терапевтической эффективности чFVIII в мышиной модели гемофилии А применялась модель рассечения хвостовой вены (TVT). Кратко, мышей первоначально обезболивали изофлураном, а анестезию поддерживали с помощью наркозной маски на протяжении всего исследования. Сразу после начала анестезии мышей помещали на нагревательную подушку (устанавливали на 37°С) с датчиком температуры между желудком и нагревательной подушкой, обеспечивая правильное расположение головки мыши в наркозной маске. Используя маркерный блок для измерения, хвост был отмечен маркерной ручкой на "12 часов", что точно соответствовало 2,5 мм в диаметре, после чего хвост был погружен в собирательную трубку с физиологическим раствором (37°С физиологического раствора). Хвост погружали в физиологический раствор с заданной температурой в течение 10 минут до начала кровотечения. Затем хвост помещали в режущий блок с отметкой от "13", указывающей на "15 часов" примерно за десять секунд до разреза (чтобы облегчить перерезание левой боковой хвостовой вены). Точно при t=0 минут производили разрез хвоста глубиной 1 мм сбоку на левой стороне хвоста мыши, тем самым перерезая боковую хвостовую вену. Сразу после этого хвост погружали в предварительно нагретую собирательную трубку с физиологическим раствором. Первый эпизод кровотечения регистрировали в течение трех минут. Если первое кровотечение превышало три минуты, животное подвергалось эвтаназии и замене. Через три минуты после повреждения собирательная трубка была заменена на новую предварительно нагретую собирательную трубку с физиологическим раствором. Все эпизоды второго кровотечения регистрировали дополнительные 57 минут. Если кровотечение прекращалось через 15, 30 или 45 минут, рану осторожно дважды вытирали намоченной в физиологическом растворе марлевой салфеткой. Сразу после испытания хвост снова погружали в физиологический раствор. Через t=60 минут мышей подвергали эвтаназии, в то время как они все еще находились под полным наркозом. Время кровотечения сообщалось как сумма первого и второго кровотечений. Первые и вторые кровотечения лизировали и хранили для последующих измерений гемоглобина, связанных с потерей крови. (Johansen и соавт., Haemophilia, (2016 год) doi:10.1111/hae.12907).
[0187] Результаты демонстрируют значительное сокращение времени для достижения гемостаза (р<0,0001) у мышей R593C, страдающих гемофилией А, после рассечения хвостовой вены (Фиг. 18), демонстрируя терапевтическую эффективность у этих мышей.
[0188] Приматы, отличные от человека: Эксперименты проводили на NHP (приматах, отличных от человека) с применением конструкций кДНК чFVIII-BDD, где оценивали серотипы AAV2/6 и AAV2/8. В Таблице 4 ниже и на Фиг. 26В показаны отличительные черты групп дозирования. Отличия между экспрессионной кассетой трансгена F8, обозначений "кДНК1" (Групп 2-4 Таблицы 4 ниже) и "кДНК2" (Группы 5 Таблицы 4 ниже), заключается в том, что донор "кДНК2" имел слегка отличающийся модуль промотора (гибридный промотор печени, см. Mcintosh и соавт (2013 год) Blood 121(17):3335), но остальная часть экспрессионной кассеты трансгена F8-BDD (включая кодирующую область) была одинаковой. Для этих экспериментов использовали режим дозирования, изображенный на Фиг. 26А (где ритуксимаб был введенным дотестовым препаратом, а стероид вводили одновременно с тестовым препаратом и ежедневно после этого).
Таблица 4: Группы NHP с кассетой трансгена FVIII
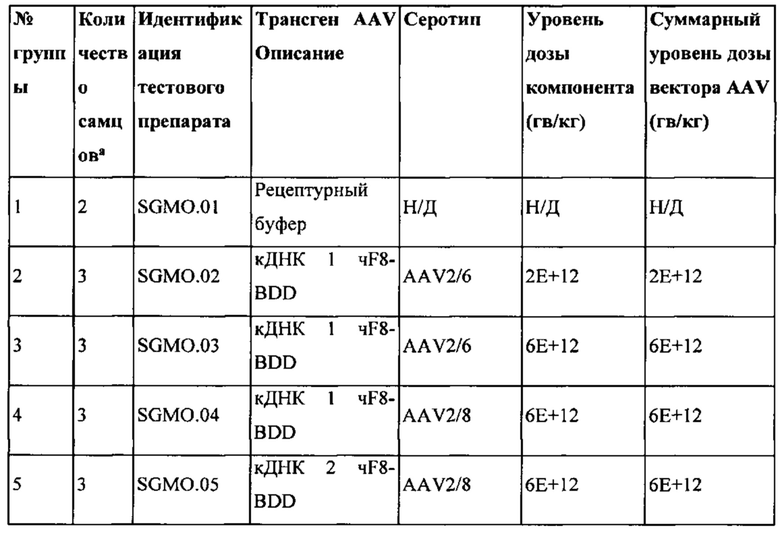
[0189] Яванским макакам вводили рецептурный буфер или 6Е+12 гв/кг кДНК
AAV2/6 чF8 (исходный). На 14-й день после добавления тестового препарата уровни и активность циркулирующего чFVIII анализировали с помощью ИФА или активности свертывания (см. Фиг. 27). Нормальные уровни FVIII яванских макак составляют ~1Ед/мл, отраженные в данных активности свертывания для контрольной группы состава, поскольку анализ активности свертывания не является специфическим для человеческого FVIII. ИФА специфичен для человеческого FVIII, а не для FVIII яванских макак, таким образом, как ожидается, не будет уровней чFVIII, измеренных с помощью ИФА в контрольной группе состава. Показаны отдельные животные для контрольной группы состава (Группа 1, идентификаторы животных 1101-1102) и дозовая группа 6Е+12 гв/кг (Группа 3, идентификаторы животных 3101-3103). У животных Группы 3 наблюдаются супрафизиологические уровни и активность циркулирующего чFVIII, выше 8 Ед/мл.
[0190] В качестве индикатора состояния печени после лечения измеряли ферменты печени для контрольной группы состава (Группа 1, идентификатор животного 1101) и дозовой группы 6Е+12 гв/кг (Группа 3, идентификатор животного 3102). Измеряли аланинаминотрансферазу (АЛТ) и аспартатаминотрансферазу (ACT). Допустимые референтные значения верхнего предела для яванских макак для АЛТ составляют 126 Ед/л и 120 Ед/л для ACT. Как для контрольной группы состава, так и для группы кДНК AAV mF8 6Е+12 гв/кг (исходный), уровни АЛТ/АСТ были увеличены после биопсии печени (биопсия печени осуществлялась на 41-й день), что обозначено звездочкой. В остальном, кДНК AAV 4F8 хорошо переносилась на протяжении всего исследования (247 дней) (см. Фиг. 28А и 28В).
[0191] Данные изображены на Фиг. 29, при этом данные, установленные для каждой обезьяны, изображены на графиках. Данные показывают, что более высокие дозы (сравнение Фиг. 29А с Фиг. 29В) тестового препарата на фоне серотипа AAV6 дали экспрессию FVIII-BDD почти на 10-кратном уровне, по сравнению с уровнем в нормальной человеческой плазме. Данные для тестового препарата в серотипе AAV2/8 показали увеличение активности FVIII, но не в той же степени, что и для AAV2/6.
[0192] После начального 14-дневного периода, описанного выше, эксперимент продолжался до 247 дней после однократной дозы AAV-FVIII-BDD. Совместное дозирование стероида прекратилось на 103-й день. Уровни чFVIII-BDD в плазме обезьян определяли с использованием стандартной ИФА следующим образом. 96-луночные половинчатые НВ (high binding) - с высокой степенью связывания) полистирольные микропланшеты (Corning) покрывали в течение ночи при 4°С моноклональным антителом к чFVIII мыши (Green Mountain, Берлингтон, штат Вермонт) в 0,2 М карбонатного бикарбонатного буфера при рН 9,4 (Thermo Fisher Scientific, Уолтем, штат Массачусетс). На следующий день планшеты промывали четыре раза с использованием 1-кратного TBST (Thermo Fisher Scientific, Уолтем, штат Массачусетс). 96-луночные планшеты затем блокировали два часа при комнатной температуре, используя 3% блокирующего буфера БСА/TBS, а затем промывали четыре раза 1-кратным TBST. Плазму добавляли в планшет и инкубировали, покачивая при комнатной температуре в течение двух часов с последующим промыванием четыре раза 1-кратным TBST. Добавляли детекторное антитело, биотинилированное моноклональное антитело к FVIII мыши (Green Mountain, Берлингтон, штат Вермонт), и инкубировали в течение одного часа при комнатной температуре, а затем промывали четыре раза 1-кратным TBST. Стрептавидин HRP (пероксидаза хрена) (Jackson ImmunoResearch, Вест Гров, штат Пенсильвания) добавляли и инкубировали в течение одного часа при комнатной температуре, а затем промывали четыре раза 1-кратным TBST. Добавляли ТМВ Ultra (Thermo Fisher Scientific, Уолтем, штат Массачусетс) и выдерживали в течение десяти минут, реакцию останавливали останавливающим раствором и измеряли оптическую плотность при 450 нМ с помощью сканирующего спектрофотометра для прочтения планшетов. Показания фоновой абсорбции были незначительными (обычно 0). Присутствие ингибирующих антител к FVIII определяли с использованием анализа Бетесда (например, см. Rasper и соавт (1975 год) Thromb Diath Haemorrh 34:869-72). Анализ ИФА применяли для оценки пиковых уровней антигена FVIII человека в ходе исследования. При уровнях дозы 2Е+12 гв/кг (n=3) достигнуты пиковые значения 111,0%, 493,9% и 840,0% (общее среднее значение 481,6% при измерении с помощью ИФА чFVIII) от нормальных уровней чFVIII человека в плазме крови. При более высокой дозе, составляющей 6Е+12 гв/кг (n=3), достигнуты пиковые значения 450,0%, 625,6% и 886,7% [общее среднее значение 654,1%] уровней чFVIII в плазме крови (Фиг. 30).
[0193] Было проведено исследование, описанное в Таблице 4, и были измерены уровни FVIII-BDD и любых ингибирующих антител к FVIII. Для животных с низкой дозой (n=3), содержащих кДНК FVIII-BDD в AAV2/6, получавших дозу 2Е+12 гв/кг, после обнаружения устойчивых уровней антигена (Аг) чFVIII, уровни чFVIII-BDD снижались при одновременном увеличении единиц Бетесда (БЕ). БЕ уменьшались с течением времени, а Аг чFVIII увеличился (Фиг. 31). Результаты показали, что после отмены иммуносупрессивной терапии уровни (как измерено с помощью ИФА) антигена FVIII человека снизились.
[0194] Для животных с высокой дозой (n=3), содержащих кДНК FVIII-BDD в AAV2/6, получавших дозу 6Е+12 гв/кг, наблюдалась аналогичная картина (см. Фиг. 32). Однако у одного животного, 3101, после удаления Солу-Медрола, не были обнаружены антитела к FVIII, несмотря на выявляемый и устойчивый уровень антигена FVIII (что составляет 200% от нормальных уровней чFVIII), что может свидетельствовать о толерантности животного к человеческому антигену FVIII.
[0195] Когда серотип AAV2/8 применялся для доставки высокой дозы, наблюдались аналогичные результаты, за исключением того, что количество антигена чFVIII, обнаруживаемого в плазме, было меньше, чем при применении вектора AAV2/6 (Фиг. 33). Аналогичным образом, когда была протестирована кДНК FVIII-BDD другого промоторного модуля (Группа 5, описанная выше) в векторе AAV2/8, уровни чFVIII-BDD в плазме крови были такими же, как и в Группе 4 (Фиг. 34). Однако, как указано выше, было две особи, которые поддерживали обнаруживаемый уровень экспрессии FVIII-BDD (5101 и 5102) и особь в Группе 4 (4103, Фиг. 33D) без заметного ответа антитела после удаления Солу-Медрола, что снова наводит на мысль о толеризации антигена после устойчивых уровней ответа, наблюдавшихся в первые дни эксперимента.
[0196] Как показано на Фиг. 34D, животное №5101, по-видимому, приобрело толерантность к чFVIII-BDD, так как после отмены метилпреднизолона уровни чFVIII-BDD оставались стабильными в течение 8 недель при приблизительно 0,1 Ед/мл (что составляет 10% от нормальных уровней nFVIII). Также, как показано на Фиг. 34Е, животное №5102, по-видимому, приобрело толерантность к чFVIII-BDD, так как после отмены метилпреднизолона уровни чFVIII-BDD оставались стабильными в течение 8 недель при приблизительно 0,6 Ед/мл (что составляет 60% от нормальных уровней чFVIII). Стоит отметить, что нормальные уровни 4FVIII в плазме человека составляют приблизительно 1 Ед/мл или 200 нг/мл, и что экспрессия даже 1% - 5% от нормального уровня (>0,001 Ед/мл) может иметь терапевтическую эффективность (Llung, RC (1999 год) Thromb Haemost 82(2):525-530).
[0197] Также проводили эксперименты с NHP, как описано выше, за исключением того, что были исследованы различные режимы иммуносупрессии. Для этих экспериментов применяли режим дозирования, изображенный на Фиг. 26В, при этом некоторые группы (1-5) получали иммуносупрессивное лечение, введенное до введения тестового препарата, тогда как другие группы (6-8) получали иммуносупрессию после введения тестового препарата.
[0198] Как изображено на Фиг. 38, при уровнях дозы 6Е+11 гв/кг (n=3, Группа 3) общее среднее значение пиковых значений составляло 5,7% (измеренное с помощью ИФА чFVIII) от нормальных уровней чFVIII в плазме крови у людей. При более высокой дозе, составляющей 2Е+12 гв/кг (n=3, Группа 4), общее среднее значений пиковых значений составляло 56,5%, а при 6Е+12 гв/кг (n=3, Группа 5) общее среднее значений пиковых значений составляло 229,0% (измеренное с помощью ИФА чFVIII) от уровней чFVIII в плазме крови. На уровне дозы 2Е+12 гв/кг (n=3, Группа 7) общее среднее значение пиковых значений составляло 87,9%, а для 6Е+12 гв/кг (n=3, Группа 8) - 101,7%. Группы 1 и 6 представляли собой контрольные группы состава, обозначенные как Состав. Кроме того, как показано на Фиг. 39, повторное введение дозы с применением того же серотипа приводило к выявляемым уровням циркулирующего антигена FVIII человека. Эти результаты показывают, что предварительное иммуносупрессивное лечение животных может быть полезно для максимальной экспрессии терапевтического белка. Кроме того, предварительная лечение позволяет повторное введение дозы, когда после первой дозы не было выработано нейтрализующих антител. Фиг. 39 также демонстрирует эффект дозы экспрессии чFVIII у животных получавших лечение в ответ на различные диапазоны доз кДНК FVIII-BDD. Поскольку, как полагается, наличие даже 1-5% белка FVIII в плазме обладает значительной терапевтической эффективностью у людей, клинические дозы в диапазоне Е11 (приводящие к 5,7-12,0 процентам от нормальных уровней в этом эксперименте), вероятно, будут оказывать значительную терапевтическую пользу. Таким образом, данные показывают, что описанные в данном документе конструкции приводят к терапевтическим уровням продуцирования трансгена in vivo.
Пример 3: Опосредованная нуклеазой целенаправленная интеграция
[0199] Конструкции, описанные в Примерах 1 и 2, также оценивали на экспрессию
при применении в сочетании с альбуминспецифическими нуклеазами для целенаправленной интеграции экспрессионной конструкции в локус гена альбумина. В частности, конструкции вводили клеткам HepG2 с альбуминспецифическими нуклеазами с цинковыми пальцами. См., например, патент США №9,150,847; 9,255,250 и публикации патентов США №20130177983; 20150159172; 20150056705 и 20150166618.
[0200] Человеческие клетки печени HepG2 поддерживались в соответствии с рекомендациями производителя (АТСС, Манассас, штат Виргиния). В день эксперимента клетки HepG2 промывали, трипсинизировали и подсчитывали. Применяемые ZFN были специфичными к локусу интрона человеческого альбумина вместе с кДНК человеческого фактора VIII-BDD (чFVIII) CRMSBS2 Без интрона, были доставлены в виде AAV2/6, и обозначены как нулевой момент времени. AAV2/6 ZFN были доставлены в количестве 3,0Е+05, a AAV2/6 кДНК чFVIII CRMSBS2 Без интрона в количестве 3,0Е+04, 6,0Е+04 и 1,2Е+05 для 1Е+05 клеток на лунку 24-луночного планшета. Таким образом, экспрессионные конструкции вводили в возрастающих дозах от 3,0Е+04,6,0Е+04 и 1,2Е+05 вместе с ZFN 3,0Е+05. Контрольные образцы включали введение только трансгена CRMSGS2 SBR Интрон 3 или с ЗФБ, только ZFN или только ЗФБ. На следующий день среды меняли. Супернатанты анализировали на секретируемый чFVIII с использованием ИФА чFVIII, описано ниже, через 19 дней после добавления вируса AAV2/6.
[0201] Секретируемые уровни человеческого фактора VIII-BDD определяли с использованием набора ИФА Affinity Biologicals (Канада) (FVIII-AG) в соответствии с протоколом производителя, за исключением стандарта фактора VIII человека. Стандарт человеческого фактора VIII, применяемый в анализе ИФА, представляет собой рекомбинантный очищенный человеческий фактор VIII (#F0016-06) из US Biologicals (Сейлем, штат Массачусетс). Коротко, супернатант HepG2 добавляли в планшет, инкубировали покачивая при комнатной температуре в течение полутора часов с последующим промыванием три раза промывочным буфером, предусмотренным в наборе. Антитело для обнаружения, предоставляемое с набором, добавляли и инкубировали в течение сорока пяти минут при комнатной температуре с последующим промыванием три раза промывочным буфером, предусмотренным в наборе. Добавляли субстрат ТМБ, предусмотренный набором, и оставляли на 10 минут. Реакцию останавливали останавливающим раствором и считывали поглощение при 450 нМ с помощью сканирующего спектрофотометра для прочтения планшетов.
[0202] Были проанализированы общий уровень белка чFVIII-BDD, обнаруженный в клеточном супернатанте HepG2 в течение времени (t=дни) после введения нацеленных на альбумин ZFN (SBS#47171/47898, см. публикацию патентной заявки РСТ WO 2015/089077) и указанная конструкция (Фиг. 36). В присутствии как экспрессионной конструкции FVIII BDD, так и ZFN, секретируемый BDD FVIII был обнаружен в супернатанте через три дня после лечения с самой высокой дозой трансгена BDD FVIII, а затем обнаруживался при более низкой дозе на пятый день и обнаруживался на всех уровнях на седьмой день. В отсутствие ZFN, FVIII BDD в супернатанте практически не обнаруживался выше фоновог уровня анализа на седьмой день. Показанные данные представляют собой n=3 биологических повторов. "Усы" погрешностей представляют собой стандартную ошибку среднего технических повторов. Пунктирная линия представляет собой предел обнаружения чFVIII с помощью ИФА. Секретируемый чFVIII-BDD слабо выявлялся в отсутствие ZFN, так как эписомальный чFVIII-BDD, вероятно, был разведен или деградирован до накопления достаточного количества детектируемого секретируемого чFVIII-BDD из этой эписомы.
[0203] Кроме того, как определено количественной ПЦР (с применением 5'-праймера, находящегося внутри эндогенного локуса альбумина человека, зонд находится в пределах ИКП кассеты чFVIII-BDD, а 3'-праймер в пределах чFVIII-BDD), целенаправленная интеграция (с помощью NHEJ) указанных конструкций была достигнута только при наличии альбуминспецифических ZFN (Фиг. 37). Таким образом, эти данные демонстрируют успешную интеграцию кассеты чFVIII-BDD и экспрессии кодированного белка F8.
[0204] Эти эксперименты были повторены, испытывая другой набор
альбуминспецифических ZFN (SBS#42906/43043, см. публикацию патентной заявки WO 2015/089046). Результаты (Фиг. 40А и 40В) показывают экспрессию белка FVIII в супернатанте этих клеток и целенаправленную интеграцию трансгена.
[0205] Альбуминспецифические ZFN и конструкции затем доставляют in vivo животным, как, по существу, описано в Примерах 1 и 2. Конструкции интегрированы в эндогенный локус альбумина и экспрессируют трансген.
[0206] Экспериментальная схема исследований in vivo приведена ниже в Таблице 5.
Кратко, поколение F1 детенышей самцов мыши C57BL/6 использовали для исследования in vivo (тестовый препарат обозначен в Таблице 5). В исследовании применяли ZFN SBS#48641 и SBS#31523 (см. публикацию патента США 2014-0017212). Исследование проводилось в соответствии с законом о благополучии животных для гуманного ухода и использования животных. Тестовые препараты оттаивали при комнатной температуре до дозирования, и все животные получали одну подкожную инъекцию 100 мкл, как описано ниже. Доза ZFN составляла 1,5 Е+11 гв на мышь, а доза кДНК F8 составляла 1,5 Е+11 гв на мышь. Кровь собирали после эвтаназии из двух или трех детенышей на группу в каждый из дней 7, 14, 21 и 28 и перерабатывали до плазмы для анализа уровней циркулирующего 4FVIII в плазме с использованием ИФА чFVIII для мыши. Ткани собирали у всех мышей для анализа уровней модификации генов с помощью глубокого секвенирования (инделы, вставки и делеции), анализа мРНК с помощью ОТ-ПЦР, анализа векторного генома с использованием кПЦР и анализа целенаправленной интеграции с использованием кПЦР.
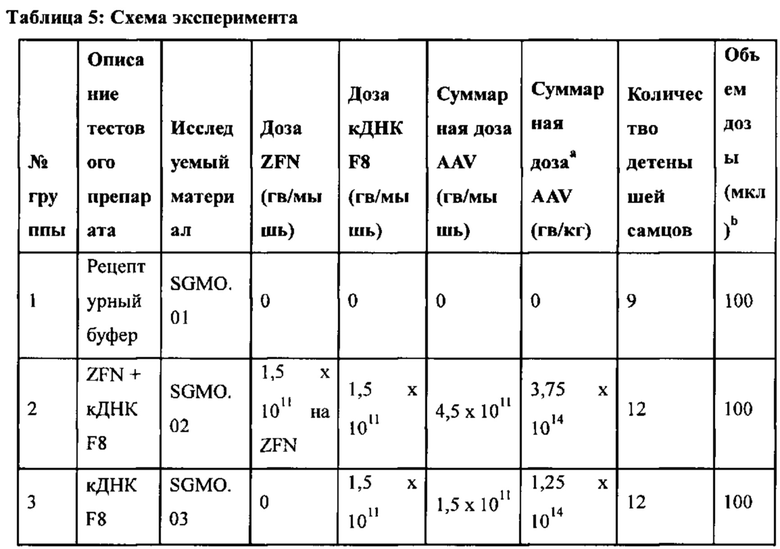
а: Суммарная доза AAV, рассчитанная с использованием массы тела 1,2 г
b: Вводится один раз в одном объеме 100 мкл.
[0207] Уровни F8, обнаруженные в плазме, представлены ниже в Таблице 6, как и количество целенаправленной интеграции трансгена FVIII-BDD.
Таблица 6: Экспрессия F8 и интеграция трансгена
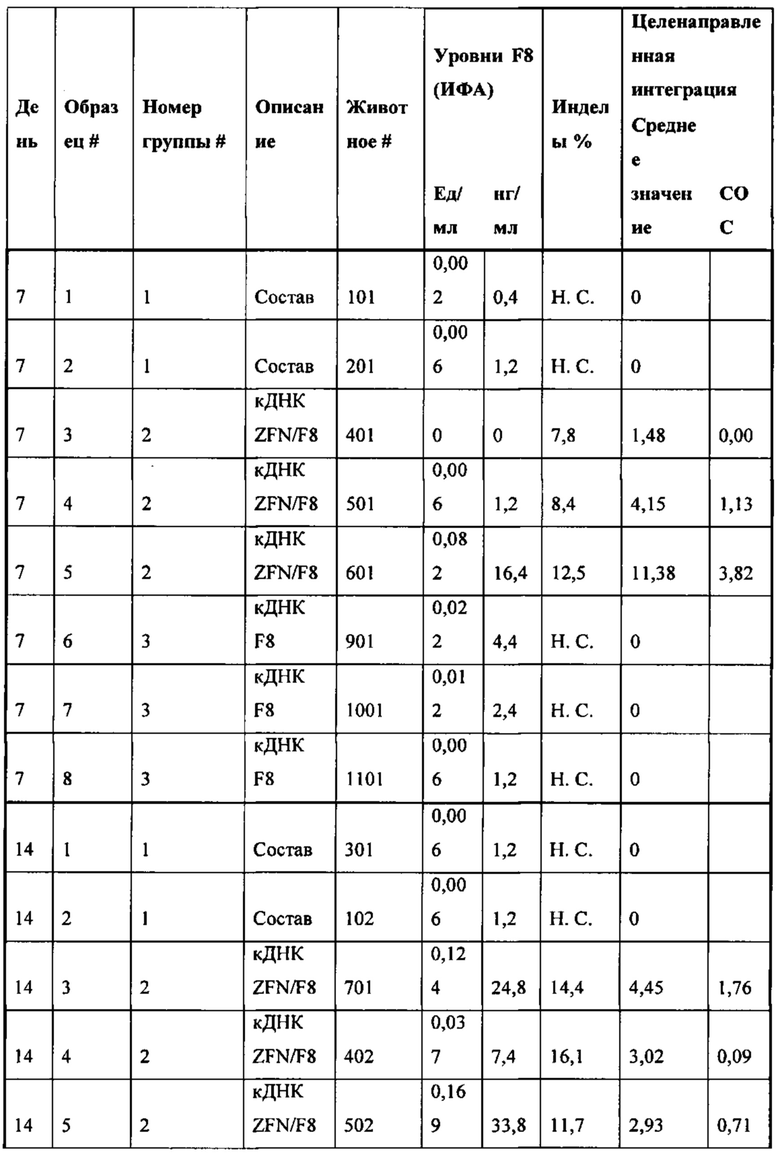
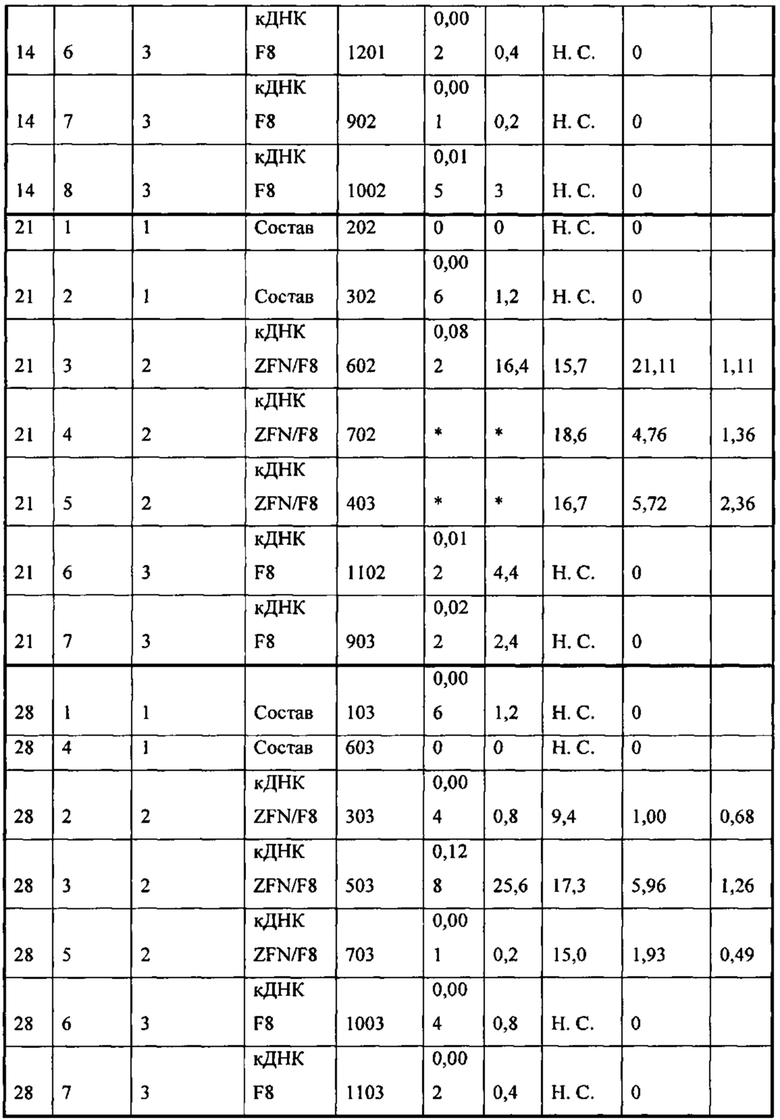
*: данные не приведены
НС: не существенно выше фона
[0208] Таким образом, FVIII выявлялся у детенышей мыши после введения альбуминспецифических нуклеаз и трансгена FVIII-BDD. Вставка трансгена также выявлялась в большем количестве у мышей, получавших как ZFN, так и трансген.
[0209] Таким образом, опосредованная нуклеазой целенаправленная интеграция описанных в данном документе конструкций обеспечивает экспрессию трансгена в клетках печени.
[0210] Все публикации, патенты и патентные заявки, цитируемые в данном документе, включены в данное описание посредством ссылки во всей своей полноте.
[0211] Несмотря на то, что настоящее изобретение было подробно описано в качестве иллюстрации и примера в целях ясности понимания, специалистам в данной области техники будет очевидно, что различные изменения и модификации могут быть реализованы на практике без отхода от сущности или объема изобретения. Соответственно, приведенные выше описания и примеры не должны рассматриваться как ограничивающие.
Позиции, указанные на графических материалах:
ФИГ. 5
1ʺ Шкала
2ʺ хр14
3ʺ Пробелы
4ʺ Человек
5ʺ Шимпанзе
6ʺ Горилла
7ʺ Орангутанг
8ʺ Гиббон
9ʺ Резус
10ʺ Макак_крабоед
11ʺ Бабуин
12ʺ Зеленая мартышка
13ʺ Галаго
14ʺ Мышь
15ʺ HepG2 Сиг
16ʺ Последовательность Вlat
17ʺ HepG2 Исходная
18ʺ Выравнивание Multiz 100 Позвоночных
19ʺ HepG2 ДНКаза I DGF Горячие точки из ENCODE/UW
20ʺ HepG2 ДНКаза I DGF Пики из ENCODE/UW
21ʺ HepG2 ДНКаза I DGF Сигнал на Каждое Основание из ENCODE/UW
22ʺ Ваша Последовательность из Поиска Blat
23ʺ (RefSeq, GenBank, CCDS, Rfam, тРНК и Сравнительная Геномика)
24ʺ HepG2 ДНКаза I DGF Необработанный Сигнал из ENCODE/U
--->
SEQUENCE LISTING
<110> SANGAMO BIOSCIENCES, INC.
<120> LIVER-SPECIFIC CONSTRUCTS, FACTOR VIII EXPRESSION CASSETTES AND
METHODS OF USE THEREOF
<130> 8325-0150.40
<140> PCT/US2016/042099
<141> 2016-07-13
<150> 62/355,106
<151> 2016-06-27
<150> 62/326,229
<151> 2016-04-22
<150> 62/315,438
<151> 2016-03-30
<150> 62/307,897
<151> 2016-03-14
<150> 62/247,469
<151> 2015-10-28
<160> 47
<170> PatentIn version 3.5
<210> 1
<211> 40
<212> DNA
<213> Homo sapiens
<400> 1
gtcaccccag ttatcggagg agcaaacagg ggctaagtcc 40
<210> 2
<211> 40
<212> DNA
<213> Pan troglodytes
<400> 2
gtcaccccag ttatcggagg agcaaacagg ggctaagtcc 40
<210> 3
<211> 40
<212> DNA
<213> Gorilla sp.
<400> 3
gtcaccccag ttatcggagg agcaaacagg ggctaagtcc 40
<210> 4
<211> 40
<212> DNA
<213> Pongo sp.
<400> 4
gtcaccccag ttatcggagg agcaaacagg ggctaagtcc 40
<210> 5
<211> 40
<212> DNA
<213> Unknown
<220>
<223> Description of Unknown: Oligonucleotide from
Gibbon species
<400> 5
gtcaccccag ttatcggagg agcaaacagg ggctaagtcc 40
<210> 6
<211> 40
<212> DNA
<213> Macaca mulatta
<400> 6
atcaccccag ttaccggagg agcaaacagg gactaagttc 40
<210> 7
<211> 40
<212> DNA
<213> Macaca fascicularis
<400> 7
atcaccccag ttaccggagg agcaaacagg gactaagttc 40
<210> 8
<211> 40
<212> DNA
<213> Papio sp.
<400> 8
atcaccccag ttaccggagg agcaaacagg gactaagttc 40
<210> 9
<211> 40
<212> DNA
<213> Chlorocebus sabaeus
<400> 9
atcaccccag ttaccggagg agcaaacagg gactaagttc 40
<210> 10
<211> 40
<212> DNA
<213> Unknown
<220>
<223> Description of Unknown: Oligonucleotide from
Callitrichidae family
<400> 10
gtcagcccag ttatcggagg agcaaacagg ggctaagtcc 40
<210> 11
<211> 39
<212> DNA
<213> Saimiri sp.
<400> 11
gtcagcccag ttaccggagg agcaaacagg gctaagtcc 39
<210> 12
<211> 38
<212> DNA
<213> Unknown
<220>
<223> Description of Unknown: Oligonucleotide from
Bush baby species
<400> 12
gtcacccagt tatcagggag caaacaggag ctaagtcc 38
<210> 13
<211> 40
<212> DNA
<213> Mus sp.
<400> 13
gtcaccacag ttattggtag agcaaacagg ggctatgtcc 40
<210> 14
<211> 88
<212> DNA
<213> Minute virus of mice
<400> 14
aagaggtaag ggtttaaggg atggttggtt ggtggggtat taatgtttaa ttacctgttt 60
tacaggcctg aaatcacttg gttttagg 88
<210> 15
<211> 92
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 15
aagaggtaag ggtttaaggg atggttggtt ggtggggcat taatgtttaa ttacctgaac 60
gacgcgccac taatcacttt ttttcaggtt gg 92
<210> 16
<211> 92
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 16
aagaggtaag ggtttaaggg atggttggtt ggtggggcat taatgtttaa ttacgacaac 60
gacgcgcctg aaatcacttt ttttcaggtt gg 92
<210> 17
<211> 94
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 17
aagaggtaag ggtttaagtt atcgttagtt cgtgcaccat taatgtttaa ttacctggag 60
cacctgcctg aaatcacttt ttttttcagg ttgg 94
<210> 18
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
primer
<400> 18
ggagatgaag aaggaggact ttg 23
<210> 19
<211> 25
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
probe
<400> 19
acatctacga cgaggacgag aacca 25
<210> 20
<211> 21
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
primer
<400> 20
tccacagcag caatgaagta g 21
<210> 21
<211> 18
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
primer
<400> 21
cctgggccag ttcctgct 18
<210> 22
<211> 23
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
probe
<400> 22
ttctgccaca tcagcagcca cca 23
<210> 23
<211> 18
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
primer
<400> 23
ggcctccatg ccatcatg 18
<210> 24
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
primer
<400> 24
agtgcaaagt aacttagagt gact 24
<210> 25
<211> 25
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
probe
<400> 25
ccatcactag gggttcctgc ggcct 25
<210> 26
<211> 18
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
primer
<400> 26
cctgaaggtg gcaatggt 18
<210> 27
<400> 27
000
<210> 28
<211> 19
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 28
ggaaccattg ccaccttca 19
<210> 29
<211> 19
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 29
ctatccattg cactatgct 19
<210> 30
<211> 18
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 30
tttcctgtaa cgatcggg 18
<210> 31
<211> 51
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 31
aataaaatat ctttattttc attacatctg tgtgttggtt ttttgtgtgt t 51
<210> 32
<211> 69
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 32
ataaaatatc tttattttca ttacatctgt gtgttggttt tttgtgtgtt ctatccattg 60
cactatgct 69
<210> 33
<211> 68
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 33
aataaaatat ctttattttc attacatctg tgtgttggtt ttttgtgtgt tttcctgtaa 60
cgatcggg 68
<210> 34
<211> 4800
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 34
gcggcctaag cttggaacca ttgccacctt cagggggagg ctgctggtga atattaacca 60
agatcacccc agttaccgga ggagcaaaca gggactaagt tcacacgcgt ggtaccgtct 120
gtctgcacat ttcgtagagc gagtgttccg atactctaat ctccctaggc aaggttcata 180
tttgtgtagg ttacttattc tccttttgtt gactaagtca ataatcagaa tcagcaggtt 240
tggagtcagc ttggcaggga tcagcagcct gggttggaag gagggggtat aaaagcccct 300
tcaccaggag aagccgtcac acagatccac aagctcctgc tagtatgcag atcgagctct 360
ccacctgctt ctttctgtgc ctgttgagat tctgcttcag cgccaccagg agatactacc 420
tgggggctgt ggagctgagc tgggactaca tgcagtctga cctgggggag ctgcctgtgg 480
atgccaggtt cccccccaga gtgcccaaga gcttcccctt caacacctct gtggtgtaca 540
agaagaccct gtttgtggag ttcactgacc acctgttcaa cattgccaag cccaggcccc 600
cctggatggg cctgctgggc cccaccatcc aggctgaggt gtatgacact gtggtgatca 660
ccctgaagaa catggccagc caccctgtga gcctgcatgc tgtgggggtg agctactgga 720
aggcctctga gggggctgag tatgatgacc agaccagcca gagggagaag gaggatgaca 780
aggtgttccc tgggggcagc cacacctatg tgtggcaggt gctgaaggag aatggcccca 840
tggcctctga ccccctgtgc ctgacctaca gctacctgag ccatgtggac ctggtgaagg 900
acctgaactc tggcctgatt ggggccctgc tggtgtgcag ggagggcagc ctggccaagg 960
agaagaccca gaccctgcac aagttcatcc tgctgtttgc tgtgtttgat gagggcaaga 1020
gctggcactc tgaaaccaag aacagcctga tgcaggacag ggatgctgcc tctgccaggg 1080
cctggcccaa gatgcacact gtgaatggct atgtgaacag gagcctgcct ggcctgattg 1140
gctgccacag gaagtctgtg tactggcatg tgattggcat gggcaccacc cctgaggtgc 1200
acagcatctt cctggagggc cacaccttcc tggtcaggaa ccacaggcag gccagcctgg 1260
agatcagccc catcaccttc ctgactgccc agaccctgct gatggacctg ggccagttcc 1320
tgctgttctg ccacatcagc agccaccagc atgatggcat ggaggcctat gtgaaggtgg 1380
acagctgccc tgaggagccc cagctgagga tgaagaacaa tgaggaggct gaggactatg 1440
atgatgacct gactgactct gagatggatg tggtgaggtt tgatgatgac aacagcccca 1500
gcttcatcca gatcaggtct gtggccaaga agcaccccaa gacctgggtg cactacattg 1560
ctgctgagga ggaggactgg gactatgccc ccctggtgct ggcccctgat gacaggagct 1620
acaagagcca gtacctgaac aatggccccc agaggattgg caggaagtac aagaaggtca 1680
ggttcatggc ctacactgat gaaaccttca agaccaggga ggccatccag catgagtctg 1740
gcatcctggg ccccctgctg tatggggagg tgggggacac cctgctgatc atcttcaaga 1800
accaggccag caggccctac aacatctacc cccatggcat cactgatgtg aggcccctgt 1860
acagcaggag gctgcccaag ggggtgaagc acctgaagga cttccccatc ctgcctgggg 1920
agatcttcaa gtacaagtgg actgtgactg tggaggatgg ccccaccaag tctgacccca 1980
ggtgcctgac cagatactac agcagctttg tgaacatgga gagggacctg gcctctggcc 2040
tgattggccc cctgctgatc tgctacaagg agtctgtgga ccagaggggc aaccagatca 2100
tgtctgacaa gaggaatgtg atcctgttct ctgtgtttga tgagaacagg agctggtacc 2160
tgactgagaa catccagagg ttcctgccca accctgctgg ggtgcagctg gaggaccctg 2220
agttccaggc cagcaacatc atgcacagca tcaatggcta tgtgtttgac agcctgcagc 2280
tgtctgtgtg cctgcatgag gtggcctact ggtacatcct gagcattggg gcccagactg 2340
acttcctgtc tgtgttcttc tctggctaca ccttcaagca caagatggtg tatgaggaca 2400
ccctgaccct gttccccttc tctggggaga ctgtgttcat gagcatggag aaccctggcc 2460
tgtggattct gggctgccac aactctgact tcaggaacag gggcatgact gccctgctga 2520
aagtctccag ctgtgacaag aacactgggg actactatga ggacagctat gaggacatct 2580
ctgcctacct gctgagcaag aacaatgcca ttgagcccag gagcttcagc cagaatccac 2640
ccgtccttaa gcgccatcag cgcgagatca ccaggaccac cctgcagtct gaccaggagg 2700
agattgacta tgatgacacc atctctgtgg agatgaagaa ggaggacttt gacatctacg 2760
acgaggacga gaaccagagc cccaggagct tccagaagaa gaccaggcac tacttcattg 2820
ctgctgtgga gaggctgtgg gactatggca tgagcagcag cccccatgtg ctgaggaaca 2880
gggcccagtc tggctctgtg ccccagttca agaaggtggt gttccaggag ttcactgatg 2940
gcagcttcac ccagcccctg tacagagggg agctgaatga gcacctgggc ctgctgggcc 3000
cctacatcag ggctgaggtg gaggacaaca tcatggtgac cttcaggaac caggccagca 3060
ggccctacag cttctacagc agcctgatca gctatgagga ggaccagagg cagggggctg 3120
agcccaggaa gaactttgtg aagcccaatg aaaccaagac ctacttctgg aaggtgcagc 3180
accacatggc ccccaccaag gatgagtttg actgcaaggc ctgggcctac ttctctgatg 3240
tggacctgga gaaggatgtg cactctggcc tgattggccc cctgctggtg tgccacacca 3300
acaccctgaa ccctgcccat ggcaggcagg tgactgtgca ggagtttgcc ctgttcttca 3360
ccatctttga tgaaaccaag agctggtact tcactgagaa catggagagg aactgcaggg 3420
ccccctgcaa catccagatg gaggacccca ccttcaagga gaactacagg ttccatgcca 3480
tcaatggcta catcatggac accctgcctg gcctggtgat ggcccaggac cagaggatca 3540
ggtggtacct gctgagcatg ggcagcaatg agaacatcca cagcatccac ttctctggcc 3600
atgtgttcac tgtgaggaag aaggaggagt acaagatggc cctgtacaac ctgtaccctg 3660
gggtgtttga gactgtggag atgctgccca gcaaggctgg catctggagg gtggagtgcc 3720
tgattgggga gcacctgcat gctggcatga gcaccctgtt cctggtgtac agcaacaagt 3780
gccagacccc cctgggcatg gcctctggcc acatcaggga cttccagatc actgcctctg 3840
gccagtatgg ccagtgggcc cccaagctgg ccaggctgca ctactctggc agcatcaatg 3900
cctggagcac caaggagccc ttcagctgga tcaaggtgga cctgctggcc cccatgatca 3960
tccatggcat caagacccag ggggccaggc agaagttcag cagcctgtac atcagccagt 4020
tcatcatcat gtacagcctg gatggcaaga agtggcagac ctacaggggc aacagcactg 4080
gcaccctgat ggtgttcttt ggcaatgtgg acagctctgg catcaagcac aacatcttca 4140
acccccccat cattgccaga tacatcaggc tgcaccccac ccactacagc atcaggagca 4200
ccctgaggat ggagctgatg ggctgtgacc tgaacagctg cagcatgccc ctgggcatgg 4260
agagcaaggc catctctgat gcccagatca ctgccagcag ctacttcacc aacatgtttg 4320
ccacctggag ccccagcaag gccaggctgc atctgcaggg caggagcaat gcctggaggc 4380
cccaggtcaa caaccccaag gagtggctgc aggtggactt ccagaagacc atgaaggtga 4440
ctggggtgac cacccagggg gtgaagagcc tgctgaccag catgtatgtg aaggagttcc 4500
tgatcagcag cagccaggat ggccaccagt ggaccctgtt cttccagaat ggcaaggtga 4560
aggtgttcca gggcaaccag gacagcttca cccctgtggt gaacagcctg gacccccccc 4620
tgctgaccag atacctgagg attcaccccc agagctgggt gcaccagatt gccctgagga 4680
tggaggtgct gggctgtgag gcccaggacc tgtactgagg atccaataaa atatctttat 4740
tttcattaca tctgtgtgtt ggttttttgt gtgttttcct gtaacgatcg ggctcgagcg 4800
<210> 35
<211> 72
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 35
gggggaggct gctggtgaat attaaccaag atcagcccag ttaccggagg agcaaacagg 60
ggctaagttc ac 72
<210> 36
<211> 72
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 36
gggggaggct gctggtgaat attaaccaag atcaccccag ttaccggagg agcaaacagg 60
gactaagttc ac 72
<210> 37
<211> 4894
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
polynucleotide
<400> 37
gcggcctaag cttggaacca ttgccacctt cagggggagg ctgctggtga atattaacca 60
agatcacccc agttaccgga ggagcaaaca gggactaagt tcacacgcgt ggtaccgtct 120
gtctgcacat ttcgtagagc gagtgttccg atactctaat ctccctaggc aaggttcata 180
tttgtgtagg ttacttattc tccttttgtt gactaagtca ataatcagaa tcagcaggtt 240
tggagtcagc ttggcaggga tcagcagcct gggttggaag gagggggtat aaaagcccct 300
tcaccaggag aagccgtcac acagatccac aagctcctga agaggtaagg gtttaagtta 360
tcgttagttc gtgcaccatt aatgtttaat tacctggagc acctgcctga aatcattttt 420
ttttcaggtt ggctagtatg cagatcgagc tctccacctg cttctttctg tgcctgttga 480
gattctgctt cagcgccacc aggagatact acctgggggc tgtggagctg agctgggact 540
acatgcagtc tgacctgggg gagctgcctg tggatgccag gttccccccc agagtgccca 600
agagcttccc cttcaacacc tctgtggtgt acaagaagac cctgtttgtg gagttcactg 660
accacctgtt caacattgcc aagcccaggc ccccctggat gggcctgctg ggccccacca 720
tccaggctga ggtgtatgac actgtggtga tcaccctgaa gaacatggcc agccaccctg 780
tgagcctgca tgctgtgggg gtgagctact ggaaggcctc tgagggggct gagtatgatg 840
accagaccag ccagagggag aaggaggatg acaaggtgtt ccctgggggc agccacacct 900
atgtgtggca ggtgctgaag gagaatggcc ccatggcctc tgaccccctg tgcctgacct 960
acagctacct gagccatgtg gacctggtga aggacctgaa ctctggcctg attggggccc 1020
tgctggtgtg cagggagggc agcctggcca aggagaagac ccagaccctg cacaagttca 1080
tcctgctgtt tgctgtgttt gatgagggca agagctggca ctctgaaacc aagaacagcc 1140
tgatgcagga cagggatgct gcctctgcca gggcctggcc caagatgcac actgtgaatg 1200
gctatgtgaa caggagcctg cctggcctga ttggctgcca caggaagtct gtgtactggc 1260
atgtgattgg catgggcacc acccctgagg tgcacagcat cttcctggag ggccacacct 1320
tcctggtcag gaaccacagg caggccagcc tggagatcag ccccatcacc ttcctgactg 1380
cccagaccct gctgatggac ctgggccagt tcctgctgtt ctgccacatc agcagccacc 1440
agcatgatgg catggaggcc tatgtgaagg tggacagctg ccctgaggag ccccagctga 1500
ggatgaagaa caatgaggag gctgaggact atgatgatga cctgactgac tctgagatgg 1560
atgtggtgag gtttgatgat gacaacagcc ccagcttcat ccagatcagg tctgtggcca 1620
agaagcaccc caagacctgg gtgcactaca ttgctgctga ggaggaggac tgggactatg 1680
cccccctggt gctggcccct gatgacagga gctacaagag ccagtacctg aacaatggcc 1740
cccagaggat tggcaggaag tacaagaagg tcaggttcat ggcctacact gatgaaacct 1800
tcaagaccag ggaggccatc cagcatgagt ctggcatcct gggccccctg ctgtatgggg 1860
aggtggggga caccctgctg atcatcttca agaaccaggc cagcaggccc tacaacatct 1920
acccccatgg catcactgat gtgaggcccc tgtacagcag gaggctgccc aagggggtga 1980
agcacctgaa ggacttcccc atcctgcctg gggagatctt caagtacaag tggactgtga 2040
ctgtggagga tggccccacc aagtctgacc ccaggtgcct gaccagatac tacagcagct 2100
ttgtgaacat ggagagggac ctggcctctg gcctgattgg ccccctgctg atctgctaca 2160
aggagtctgt ggaccagagg ggcaaccaga tcatgtctga caagaggaat gtgatcctgt 2220
tctctgtgtt tgatgagaac aggagctggt acctgactga gaacatccag aggttcctgc 2280
ccaaccctgc tggggtgcag ctggaggacc ctgagttcca ggccagcaac atcatgcaca 2340
gcatcaatgg ctatgtgttt gacagcctgc agctgtctgt gtgcctgcat gaggtggcct 2400
actggtacat cctgagcatt ggggcccaga ctgacttcct gtctgtgttc ttctctggct 2460
acaccttcaa gcacaagatg gtgtatgagg acaccctgac cctgttcccc ttctctgggg 2520
agactgtgtt catgagcatg gagaaccctg gcctgtggat tctgggctgc cacaactctg 2580
acttcaggaa caggggcatg actgccctgc tgaaagtctc cagctgtgac aagaacactg 2640
gggactacta tgaggacagc tatgaggaca tctctgccta cctgctgagc aagaacaatg 2700
ccattgagcc caggagcttc agccagaatc cacccgtcct taagcgccat cagcgcgaga 2760
tcaccaggac caccctgcag tctgaccagg aggagattga ctatgatgac accatctctg 2820
tggagatgaa gaaggaggac tttgacatct acgacgagga cgagaaccag agccccagga 2880
gcttccagaa gaagaccagg cactacttca ttgctgctgt ggagaggctg tgggactatg 2940
gcatgagcag cagcccccat gtgctgagga acagggccca gtctggctct gtgccccagt 3000
tcaagaaggt ggtgttccag gagttcactg atggcagctt cacccagccc ctgtacagag 3060
gggagctgaa tgagcacctg ggcctgctgg gcccctacat cagggctgag gtggaggaca 3120
acatcatggt gaccttcagg aaccaggcca gcaggcccta cagcttctac agcagcctga 3180
tcagctatga ggaggaccag aggcaggggg ctgagcccag gaagaacttt gtgaagccca 3240
atgaaaccaa gacctacttc tggaaggtgc agcaccacat ggcccccacc aaggatgagt 3300
ttgactgcaa ggcctgggcc tacttctctg atgtggacct ggagaaggat gtgcactctg 3360
gcctgattgg ccccctgctg gtgtgccaca ccaacaccct gaaccctgcc catggcaggc 3420
aggtgactgt gcaggagttt gccctgttct tcaccatctt tgatgaaacc aagagctggt 3480
acttcactga gaacatggag aggaactgca gggccccctg caacatccag atggaggacc 3540
ccaccttcaa ggagaactac aggttccatg ccatcaatgg ctacatcatg gacaccctgc 3600
ctggcctggt gatggcccag gaccagagga tcaggtggta cctgctgagc atgggcagca 3660
atgagaacat ccacagcatc cacttctctg gccatgtgtt cactgtgagg aagaaggagg 3720
agtacaagat ggccctgtac aacctgtacc ctggggtgtt tgagactgtg gagatgctgc 3780
ccagcaaggc tggcatctgg agggtggagt gcctgattgg ggagcacctg catgctggca 3840
tgagcaccct gttcctggtg tacagcaaca agtgccagac ccccctgggc atggcctctg 3900
gccacatcag ggacttccag atcactgcct ctggccagta tggccagtgg gcccccaagc 3960
tggccaggct gcactactct ggcagcatca atgcctggag caccaaggag cccttcagct 4020
ggatcaaggt ggacctgctg gcccccatga tcatccatgg catcaagacc cagggggcca 4080
ggcagaagtt cagcagcctg tacatcagcc agttcatcat catgtacagc ctggatggca 4140
agaagtggca gacctacagg ggcaacagca ctggcaccct gatggtgttc tttggcaatg 4200
tggacagctc tggcatcaag cacaacatct tcaacccccc catcattgcc agatacatca 4260
ggctgcaccc cacccactac agcatcagga gcaccctgag gatggagctg atgggctgtg 4320
acctgaacag ctgcagcatg cccctgggca tggagagcaa ggccatctct gatgcccaga 4380
tcactgccag cagctacttc accaacatgt ttgccacctg gagccccagc aaggccaggc 4440
tgcatctgca gggcaggagc aatgcctgga ggccccaggt caacaacccc aaggagtggc 4500
tgcaggtgga cttccagaag accatgaagg tgactggggt gaccacccag ggggtgaaga 4560
gcctgctgac cagcatgtat gtgaaggagt tcctgatcag cagcagccag gatggccacc 4620
agtggaccct gttcttccag aatggcaagg tgaaggtgtt ccagggcaac caggacagct 4680
tcacccctgt ggtgaacagc ctggaccccc ccctgctgac cagatacctg aggattcacc 4740
cccagagctg ggtgcaccag attgccctga ggatggaggt gctgggctgt gaggcccagg 4800
acctgtactg aggatccaat aaaatatctt tattttcatt acatctgtgt gttggttttt 4860
tgtgtgtttt cctgtaacga tcgggctcga gcgc 4894
<210> 38
<211> 21
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
oligonucleotide
<400> 38
ttgaattcat aactatccca a 21
<210> 39
<211> 21
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
primer
<400> 39
gtgtagcaga gaggaaccat t 21
<210> 40
<211> 25
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
probe
<400> 40
ccatcactag gggttcctgc ggcct 25
<210> 41
<211> 21
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
primer
<400> 41
gttaatattc accagcagcc t 21
<210> 42
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
primer
<400> 42
agtgtagcag agaggaacca 20
<210> 43
<211> 25
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
probe
<400> 43
ccatcactag gggttcctgc ggcct 25
<210> 44
<211> 18
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
primer
<400> 44
cagggtgagc ccagaaac 18
<210> 45
<211> 22
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
primer
<400> 45
aactttgagt gtagcagaga gg 22
<210> 46
<211> 24
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
probe
<400> 46
taccggagga gcaaacaggg acta 24
<210> 47
<211> 22
<212> DNA
<213> Artificial Sequence
<220>
<223> Description of Artificial Sequence: Synthetic
primer
<400> 47
ctctacgaaa tgtgcagaca ga 22
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| УЛУЧШЕНИЕ КЛИНИЧЕСКИХ ПАРАМЕТРОВ ПОСРЕДСТВОМ ЭКСПРЕССИИ ФАКТОРА VIII | 2019 |
|
RU2799048C2 |
| ГЕННАЯ ТЕРАПИЯ ДЛЯ ЛЕЧЕНИЯ ГЕМОФИЛИИ A | 2017 |
|
RU2762257C2 |
| КОМПОЗИЦИИ ВЕКТОРА AAV И СПОСОБЫ ПЕРЕНОСА ГЕНОВ В КЛЕТКИ, ОРГАНЫ И ТКАНИ | 2013 |
|
RU2653444C2 |
| Кодон-оптимизированная нуклеиновая кислота, которая кодирует белок фактора свёртывания крови VIII c делетированным B доменом, и ее применение | 2022 |
|
RU2808564C2 |
| КОДИРУЮЩИЕ ФАКТОР VIII ВАРИАНТЫ С ПОНИЖЕННЫМ СОДЕРЖАНИЕМ CPG, КОМПОЗИЦИИ И СПОСОБЫ И ПРИМЕНЕНИЕ В ЛЕЧЕНИИ НАРУШЕНИЙ ГЕМОСТАЗА | 2016 |
|
RU2745506C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ЛЕЧЕНИЯ БОЛЕЗНИ ФАБРИ | 2020 |
|
RU2822369C2 |
| МОДИФИКАЦИЯ B-КЛЕТОК | 2018 |
|
RU2783116C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ЛЕЧЕНИЯ БОЛЕЗНИ ФАБРИ | 2017 |
|
RU2788133C2 |
| Выделенная нуклеиновая кислота, которая кодирует слитый белок на основе FVIII-BDD и гетерологичного сигнального пептида, и ее применение | 2022 |
|
RU2818229C2 |
| СПОСОБЫ ГЕНОТЕРАПИИ С ИСПОЛЬЗОВАНИЕМ ГЕНА ФАКТОРА VIII (FVIII) | 2018 |
|
RU2808274C2 |
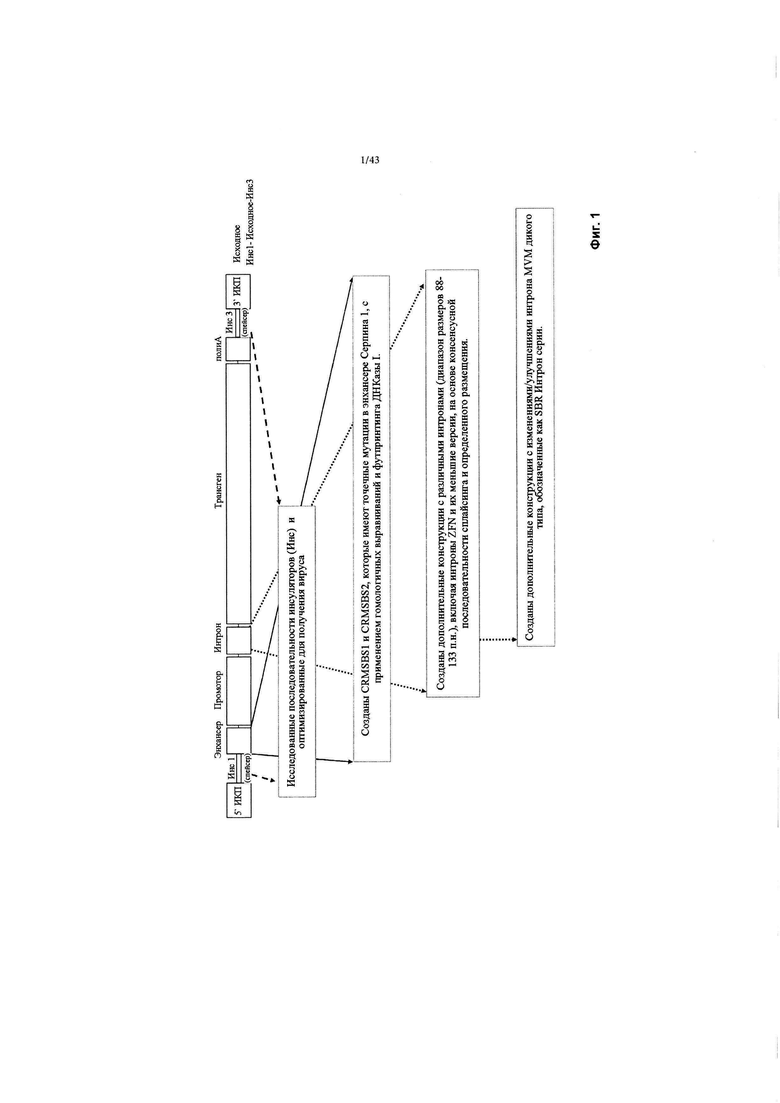
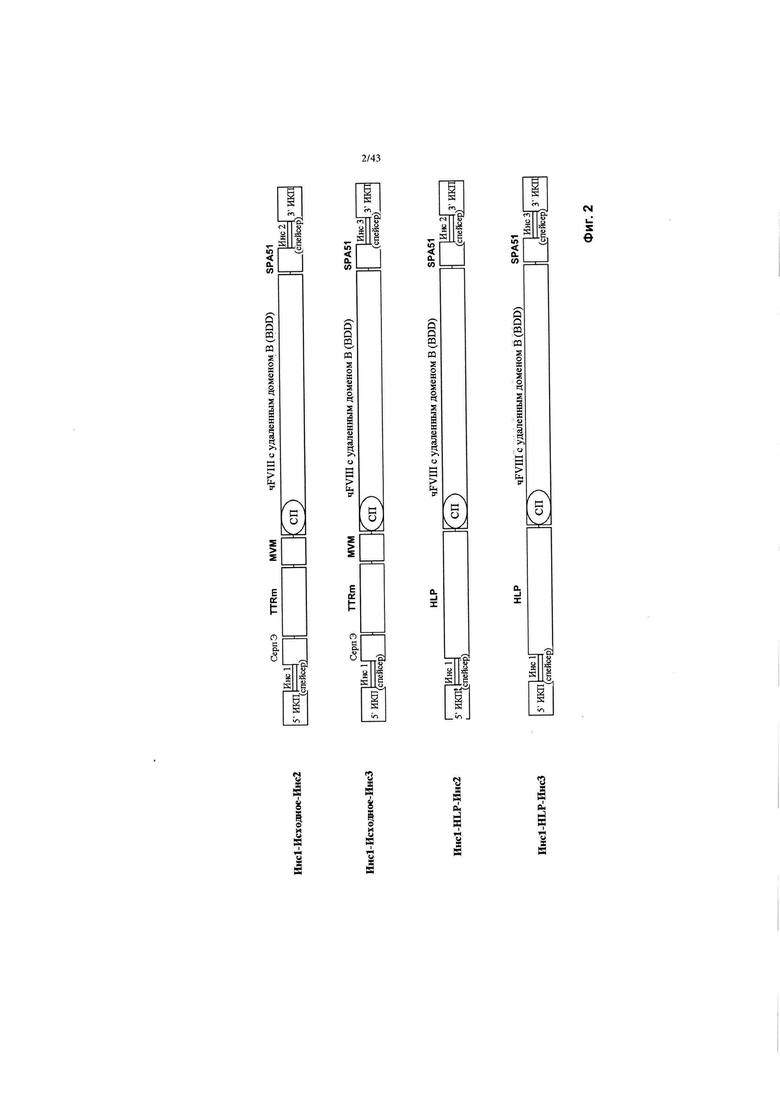
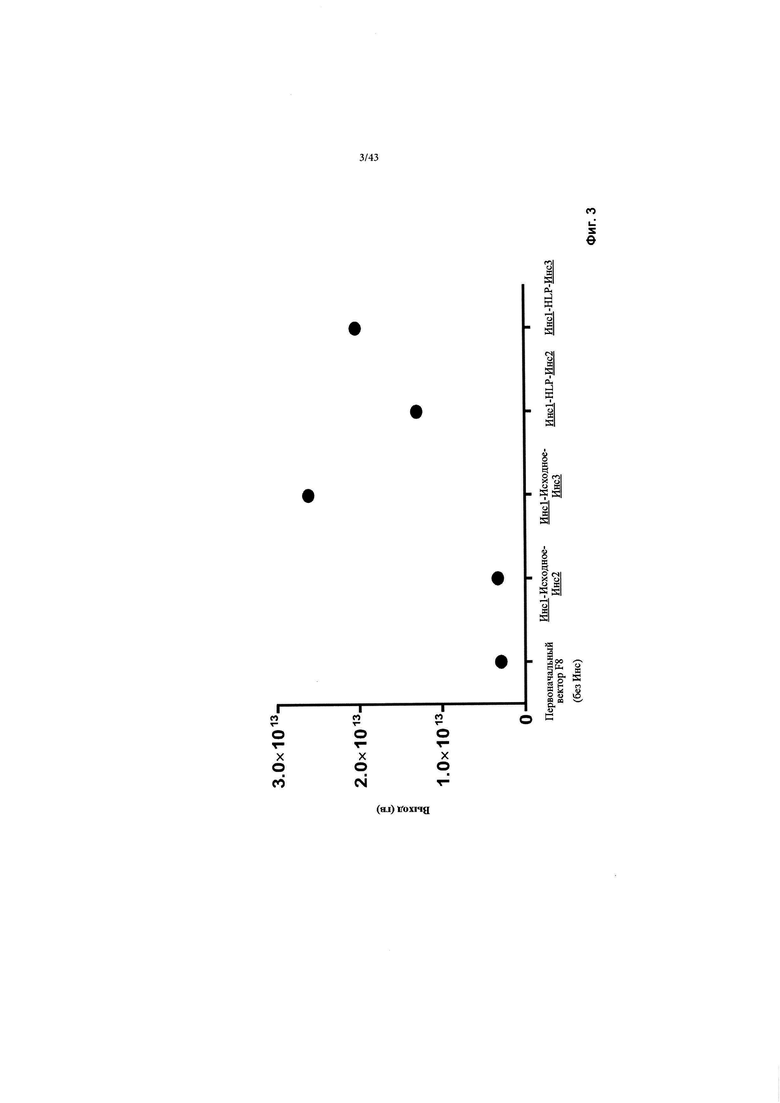
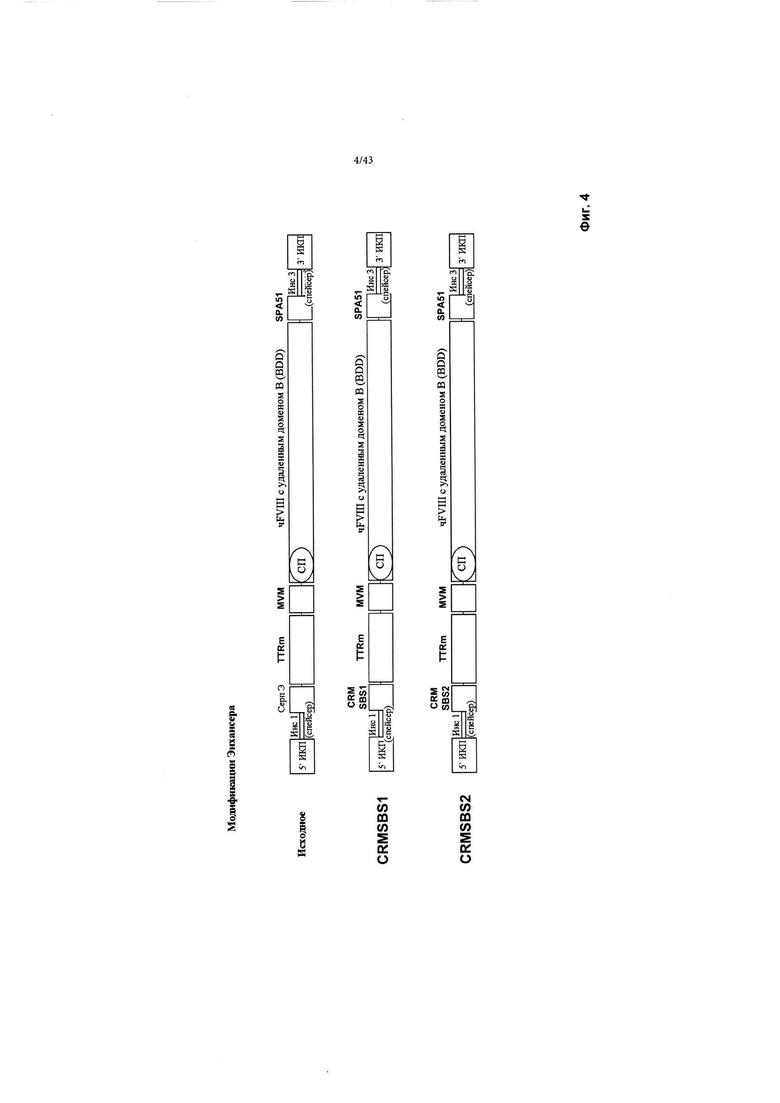
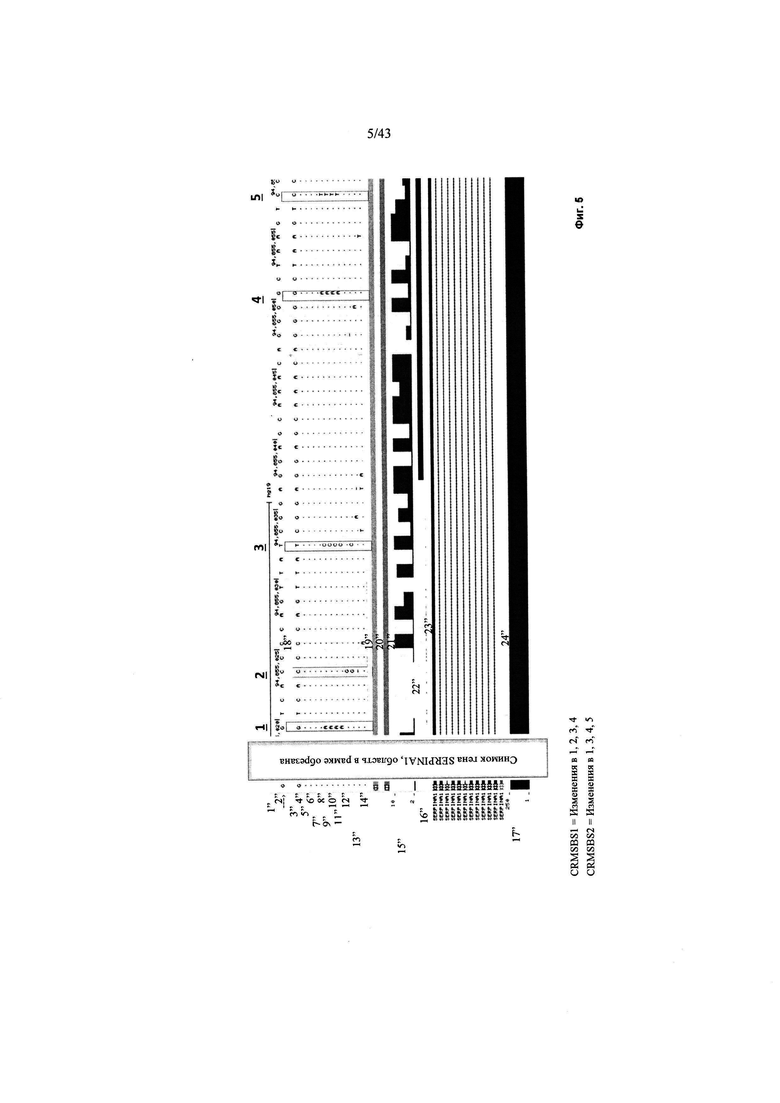
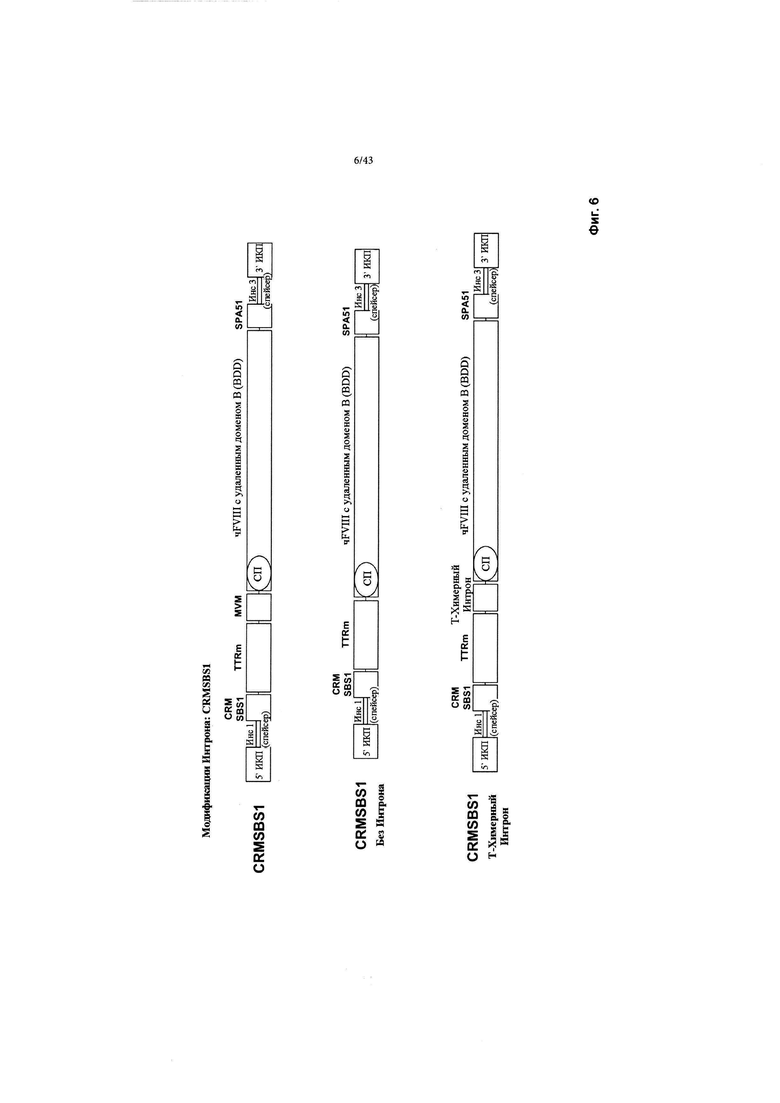
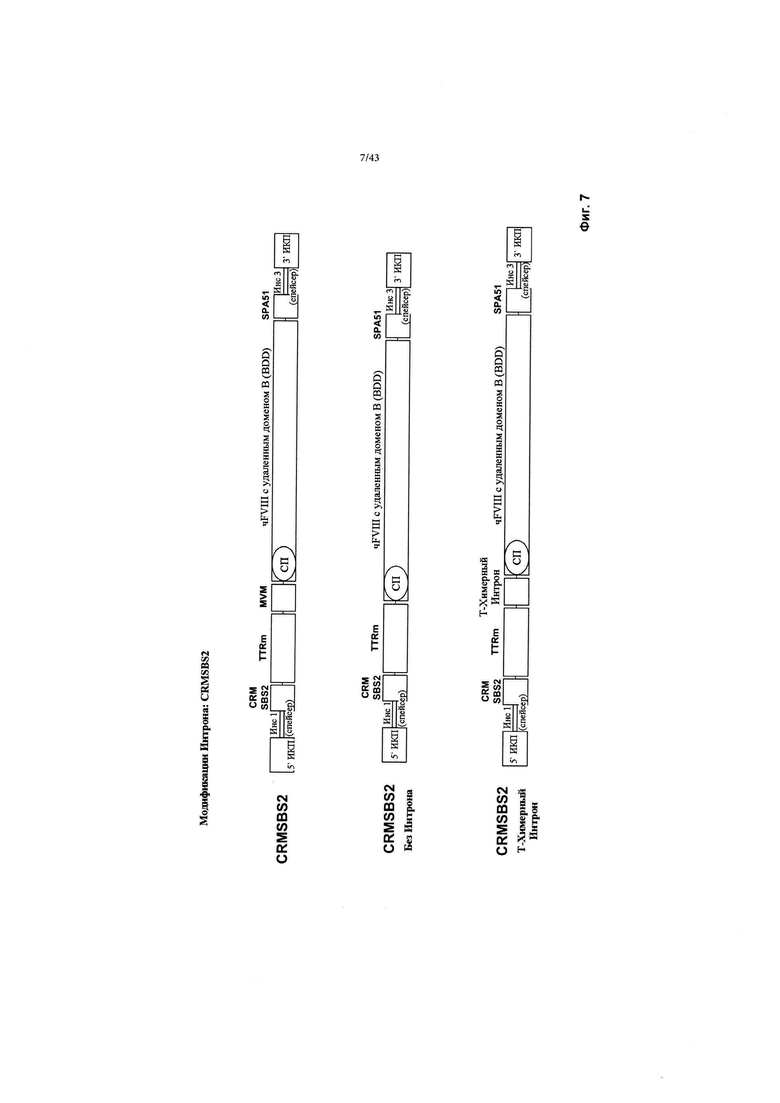
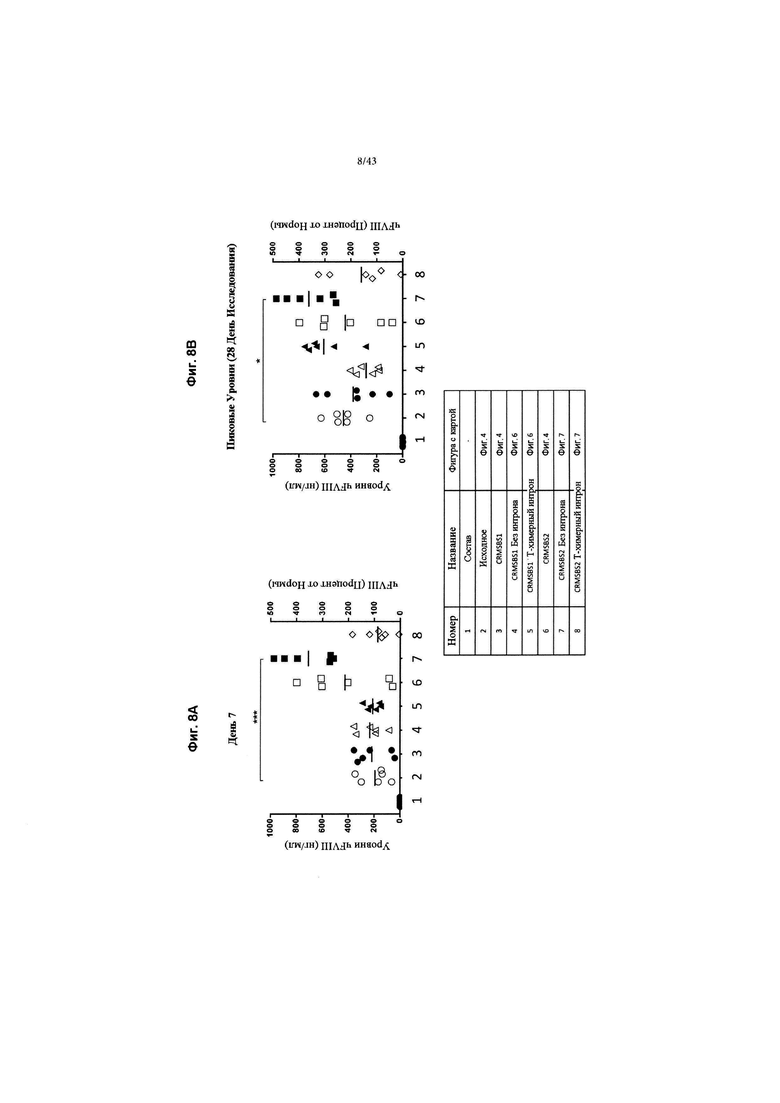

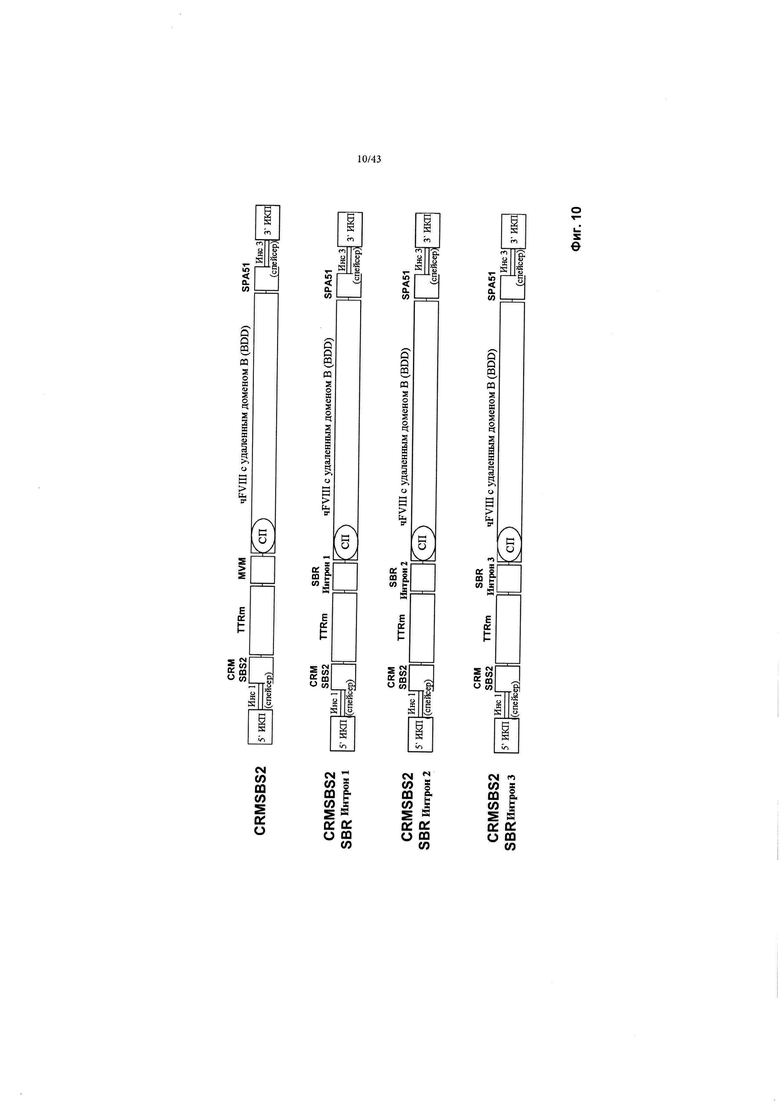
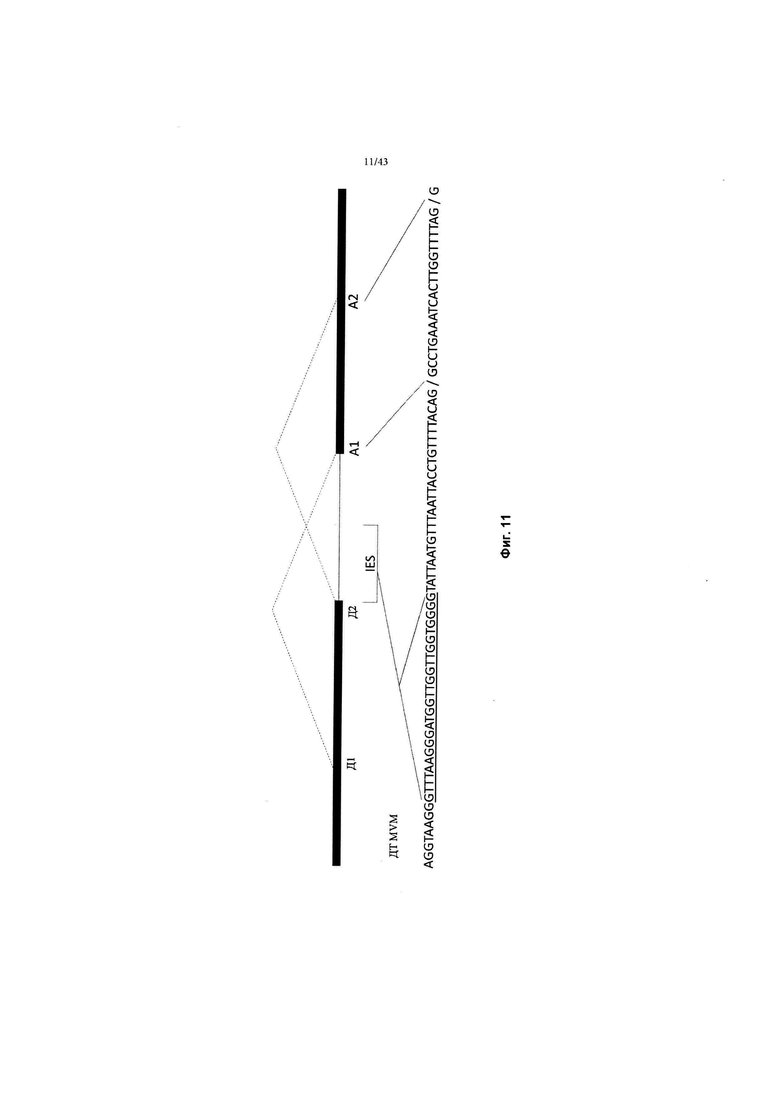
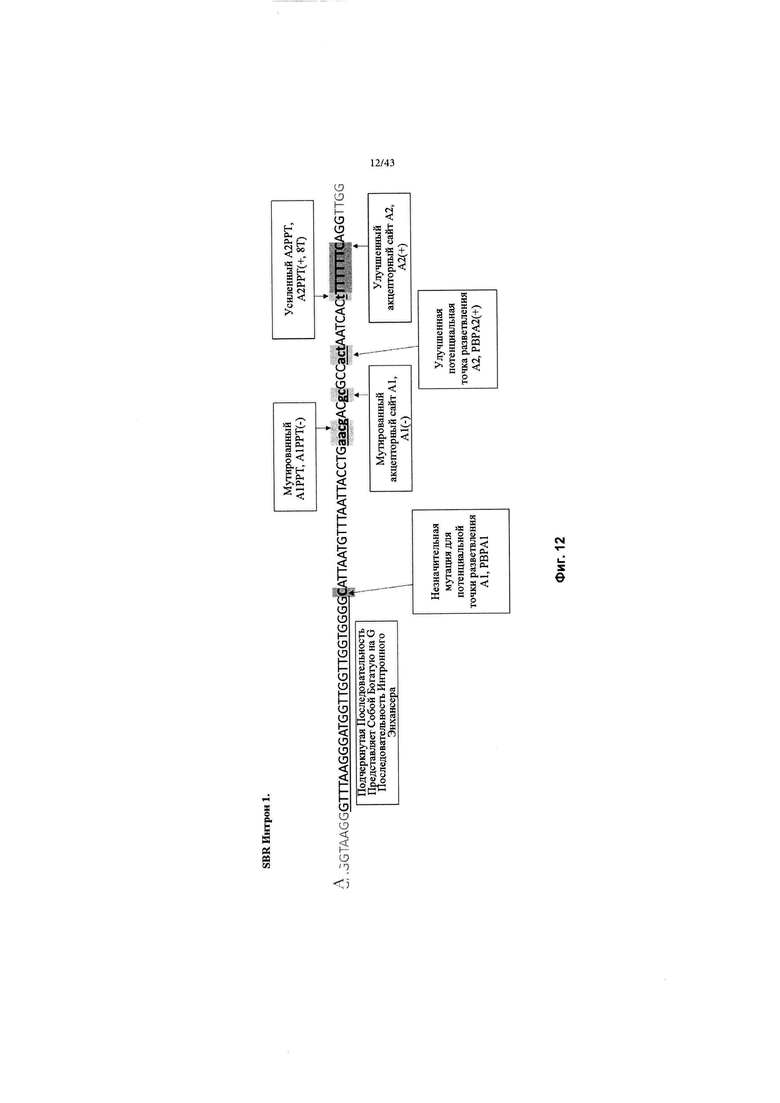
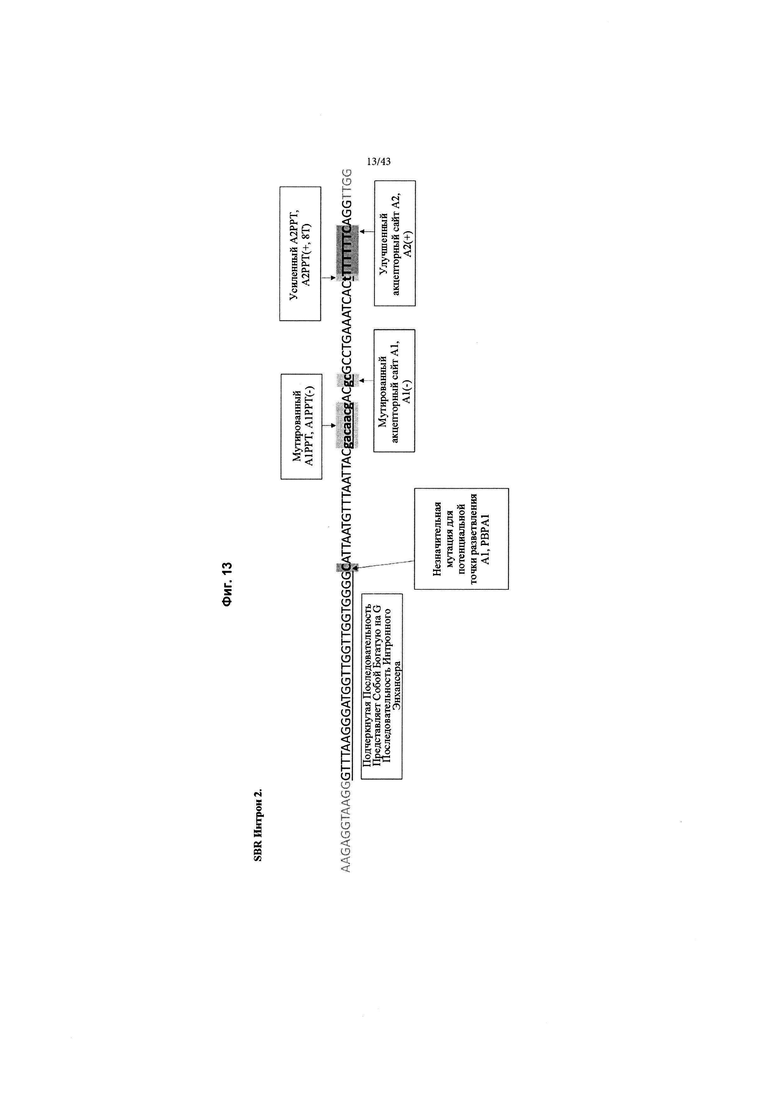
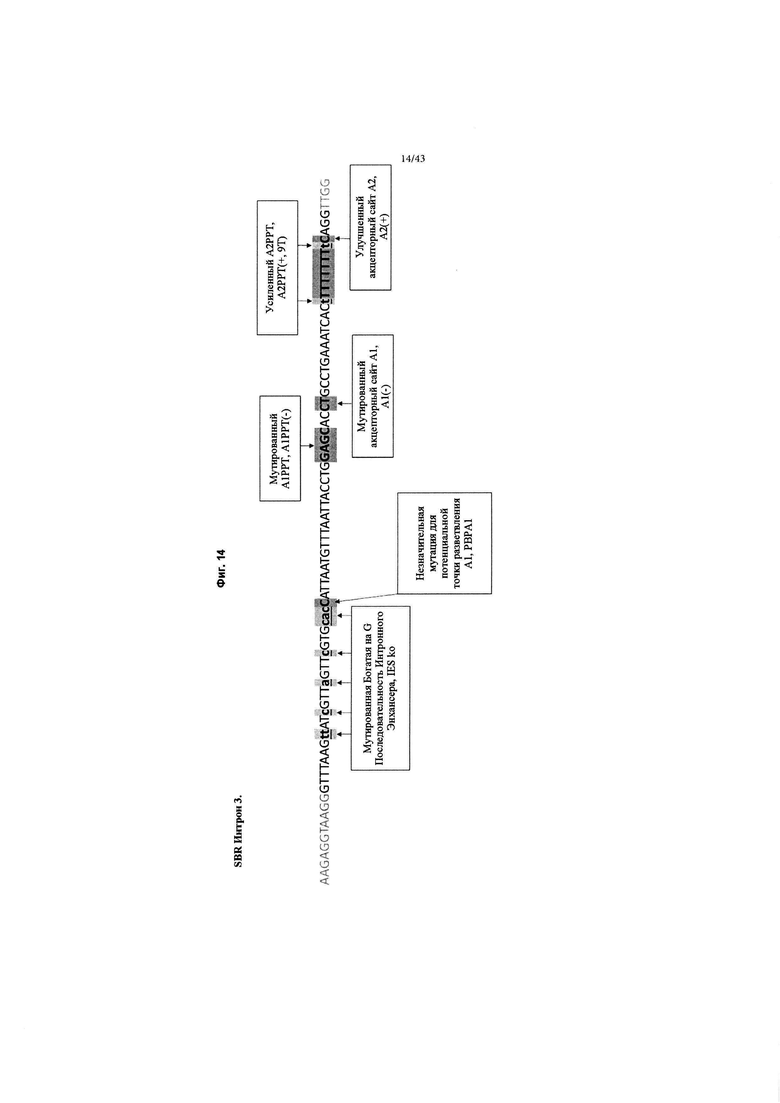
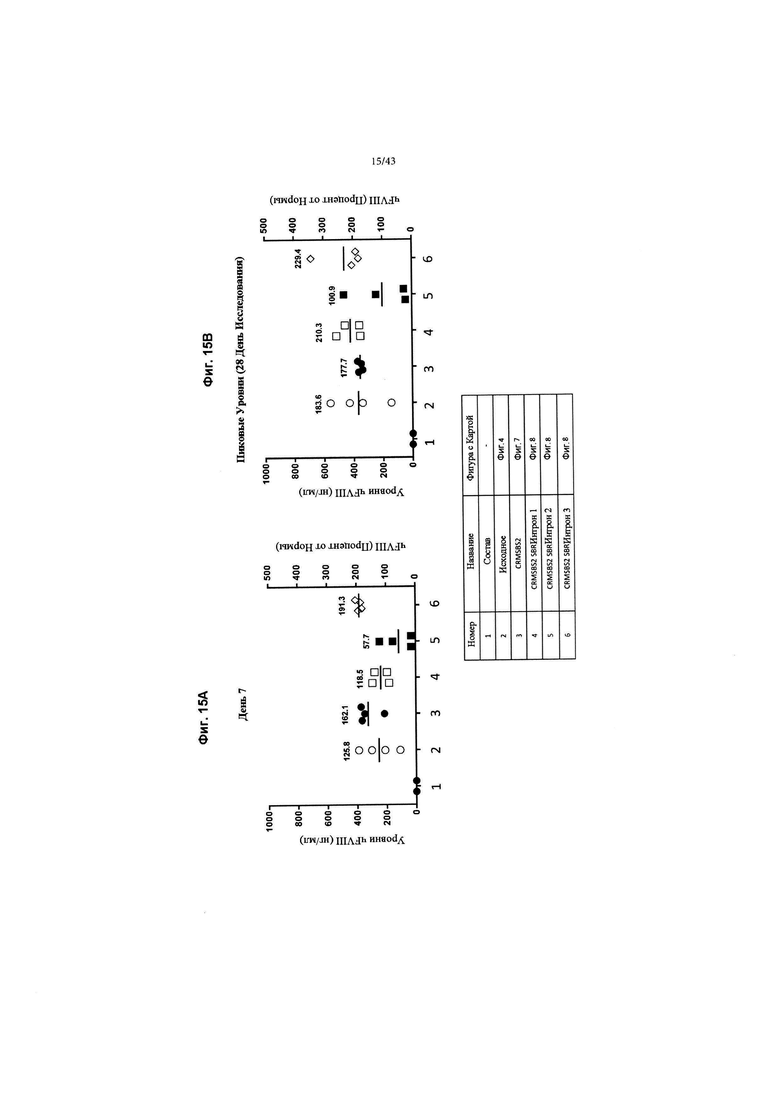


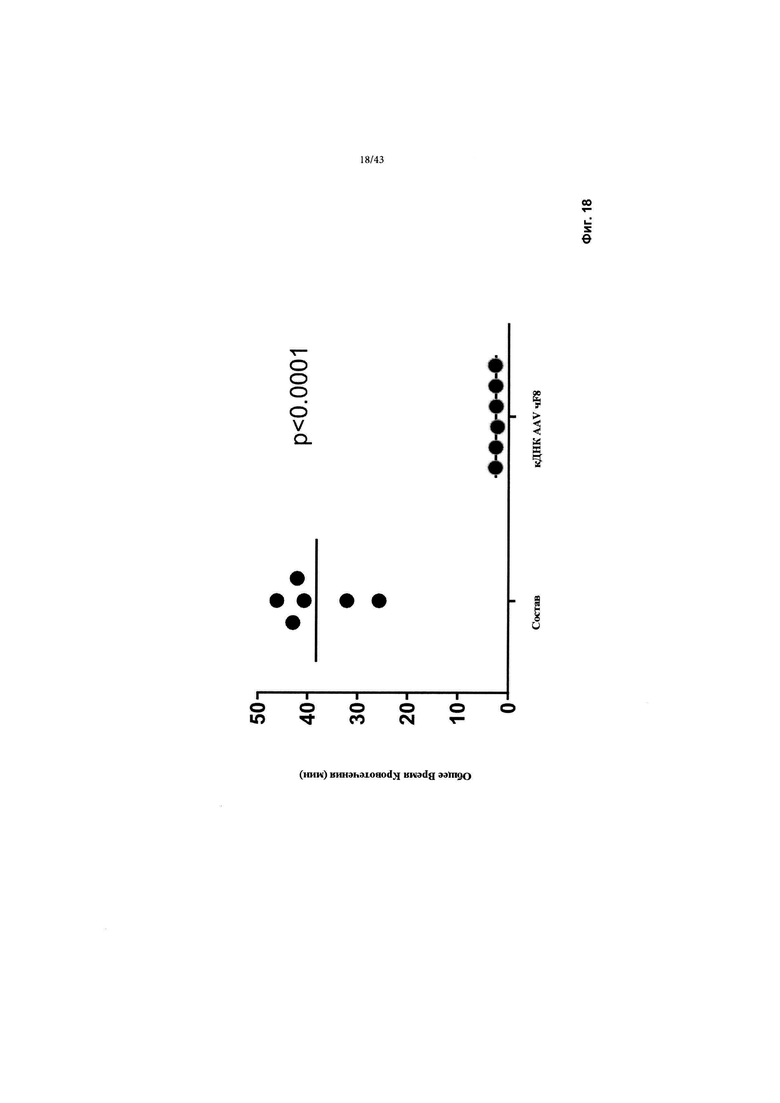
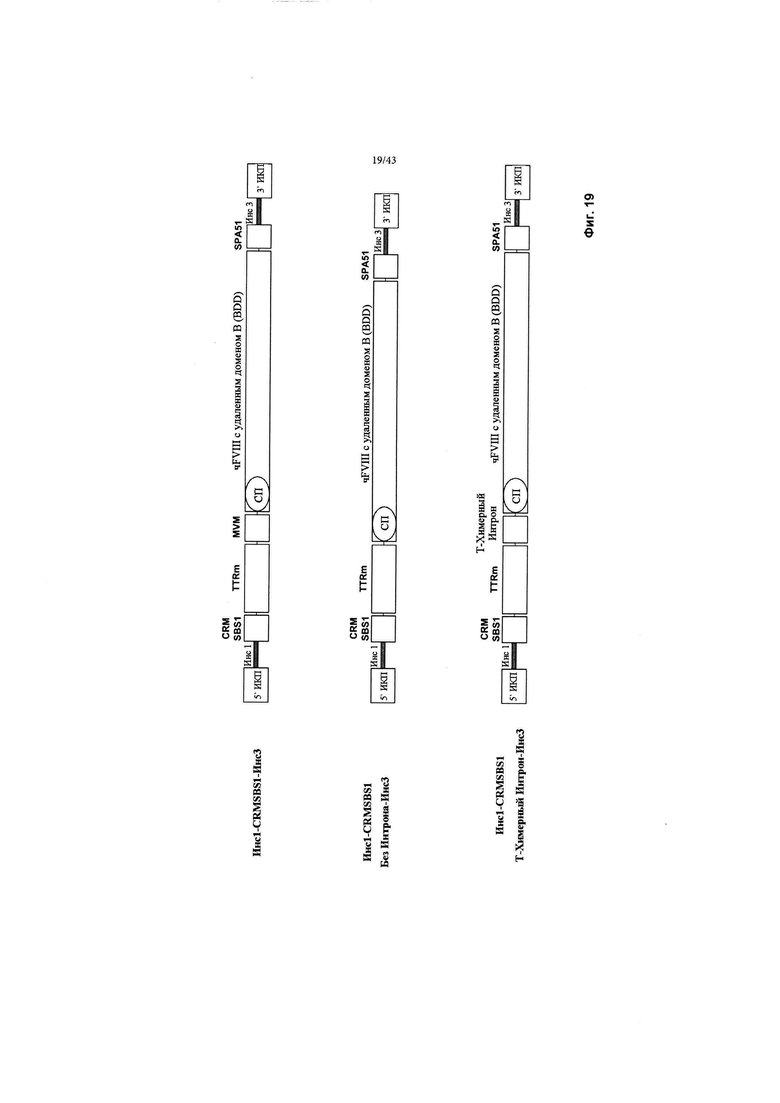
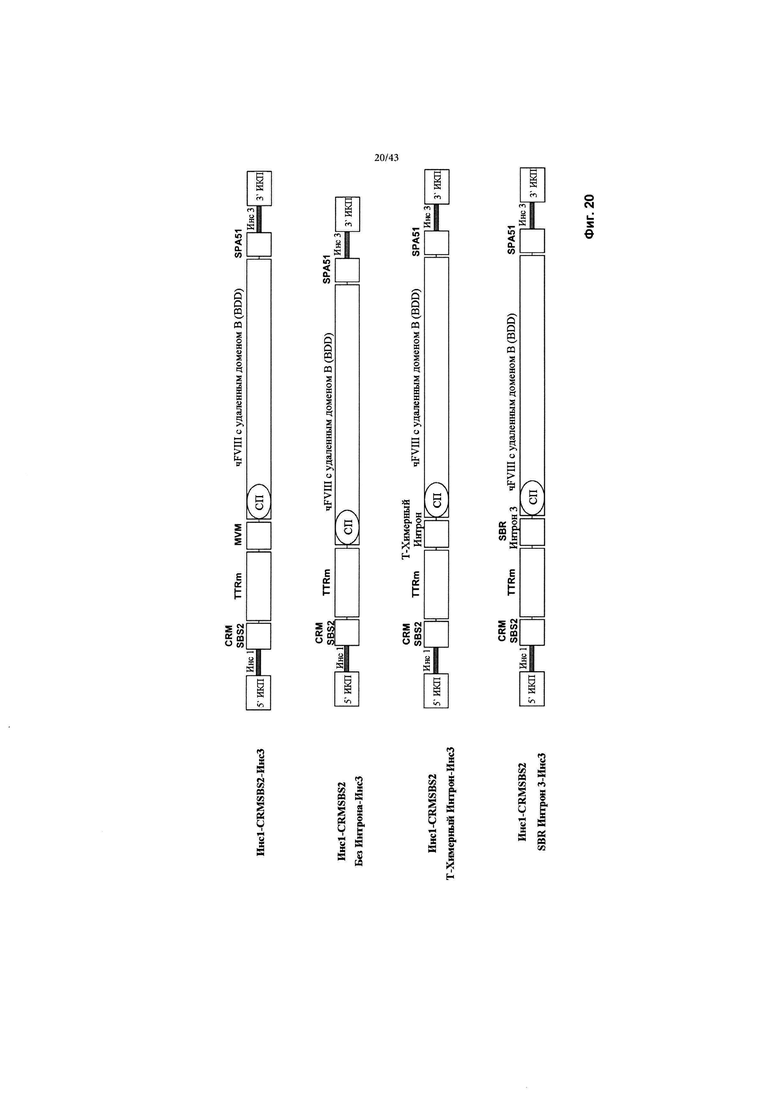
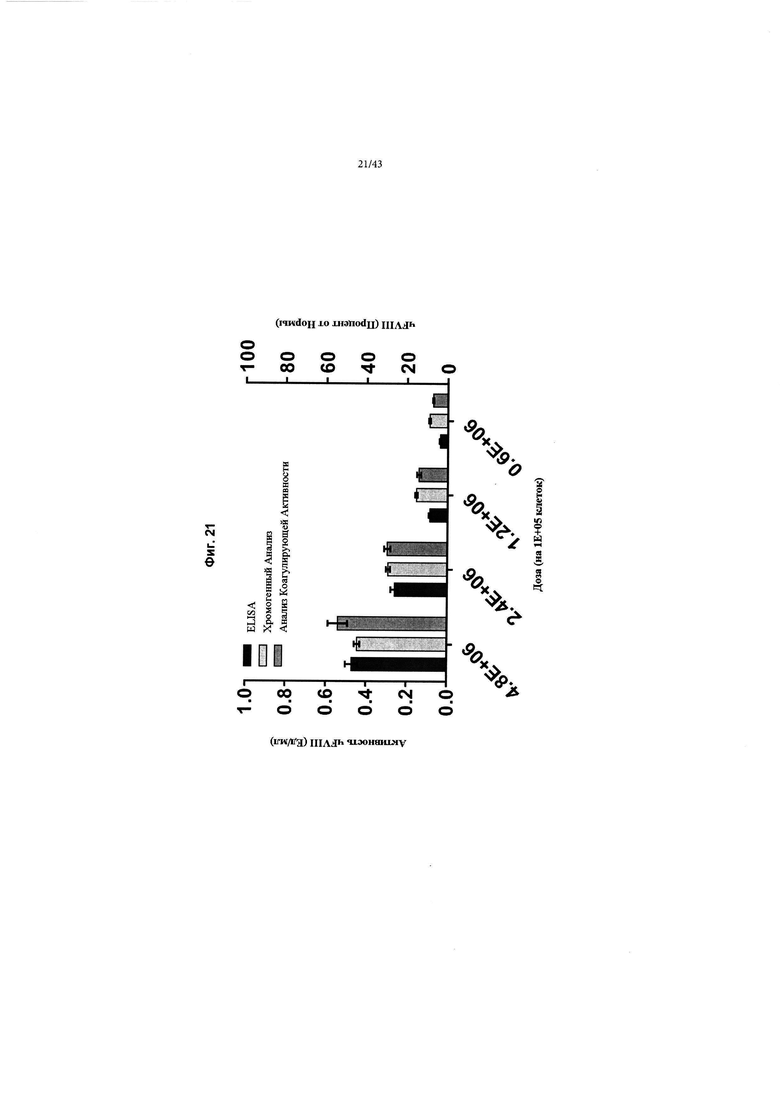
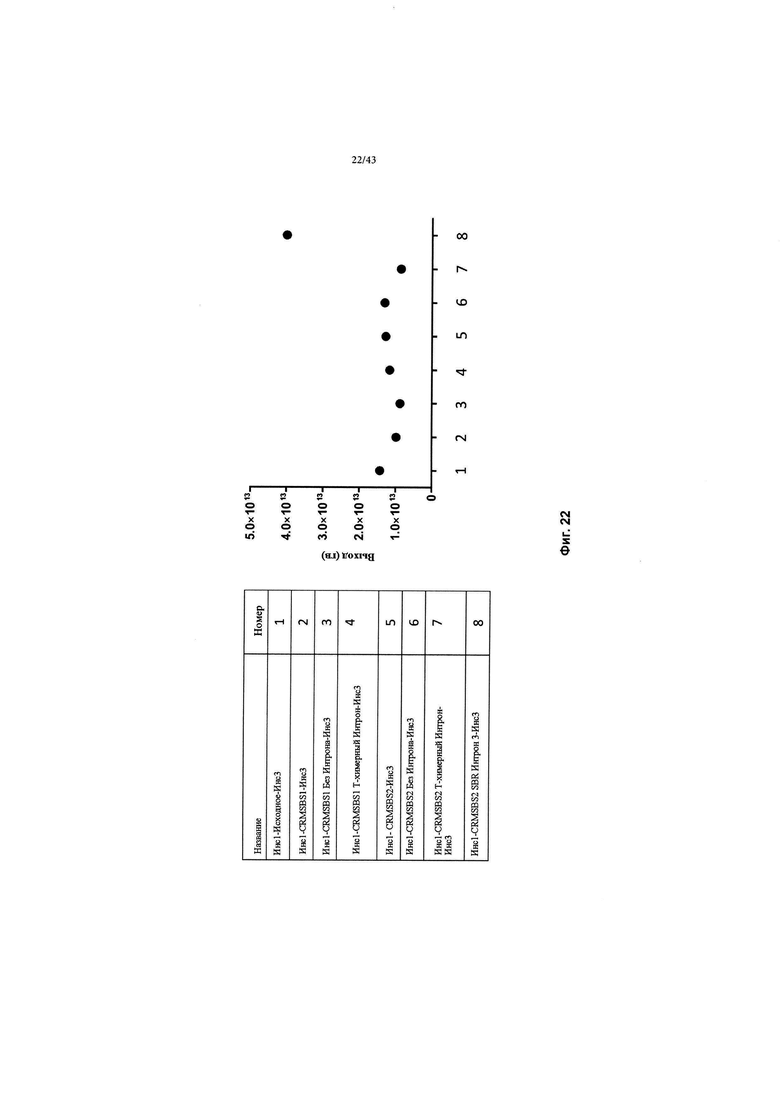
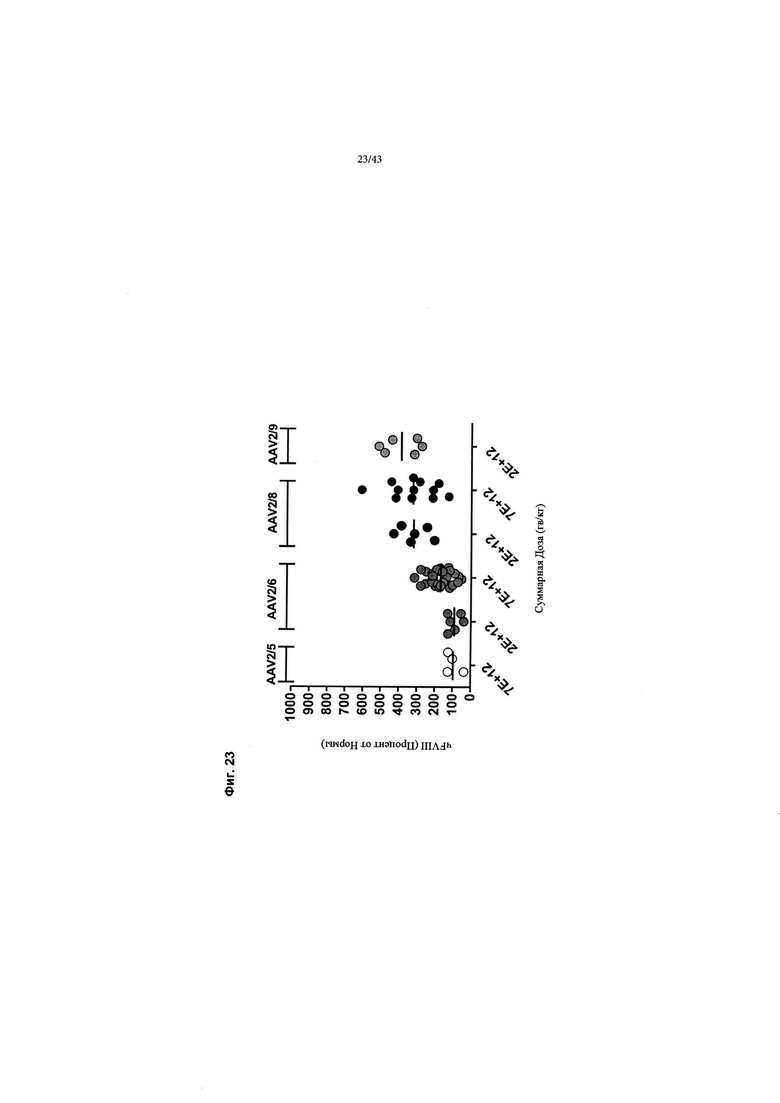
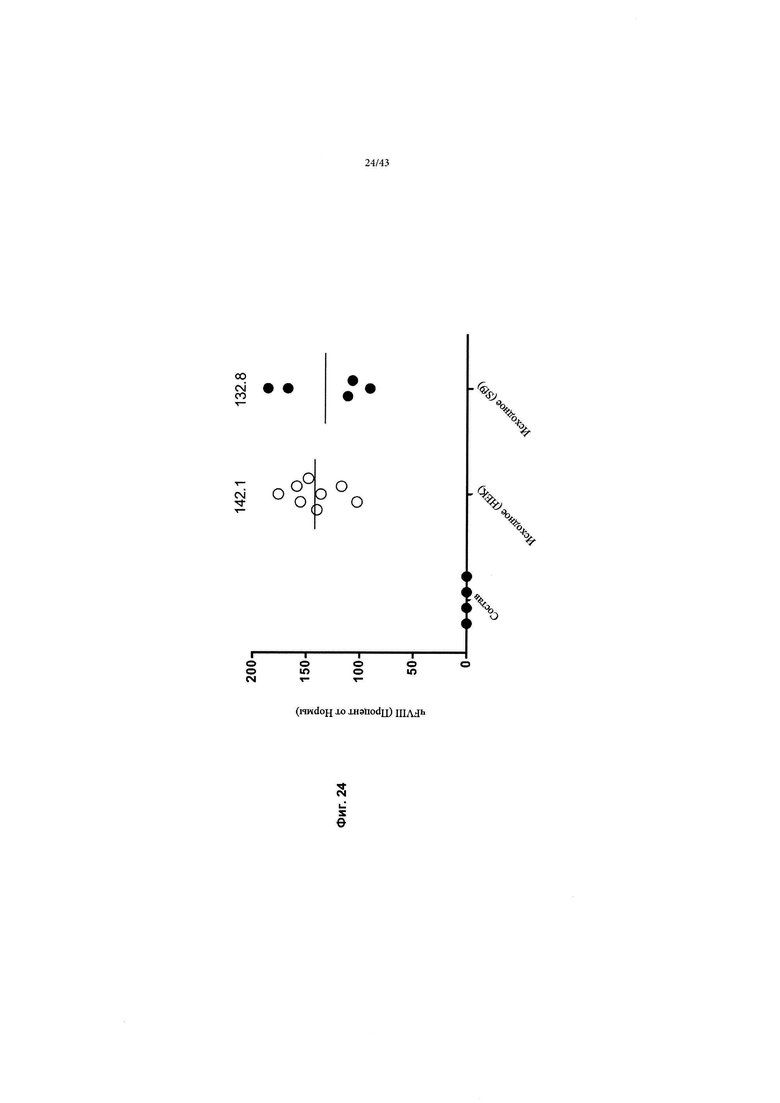
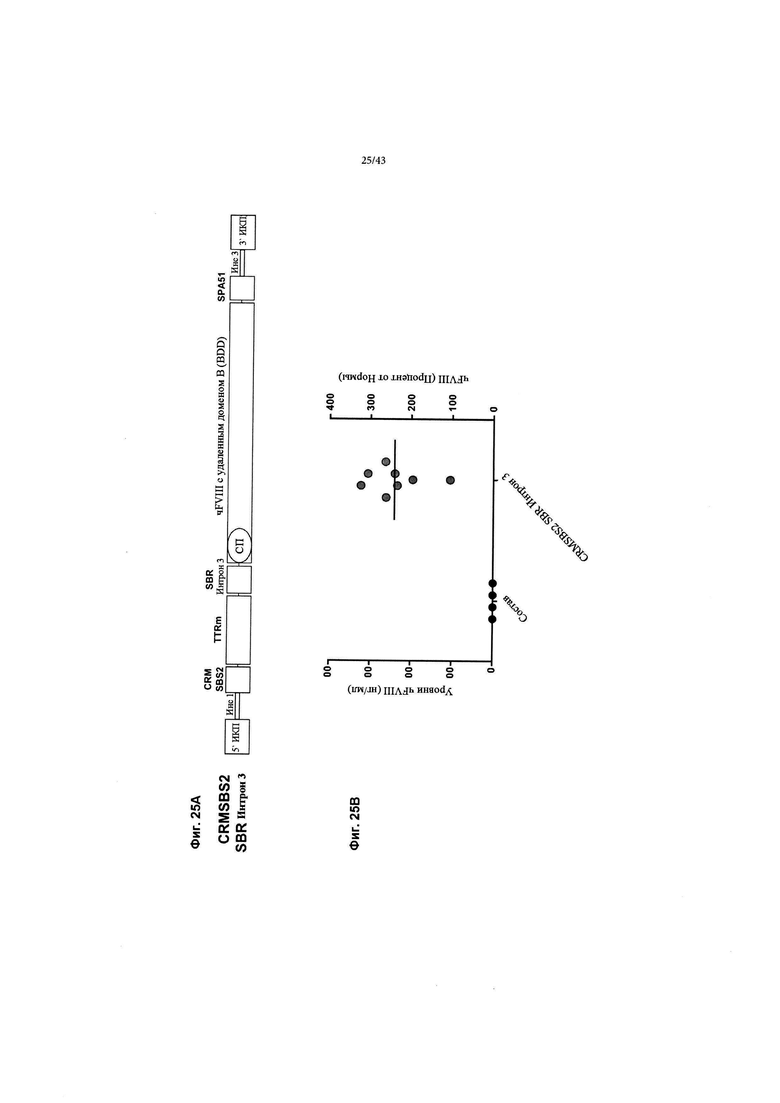

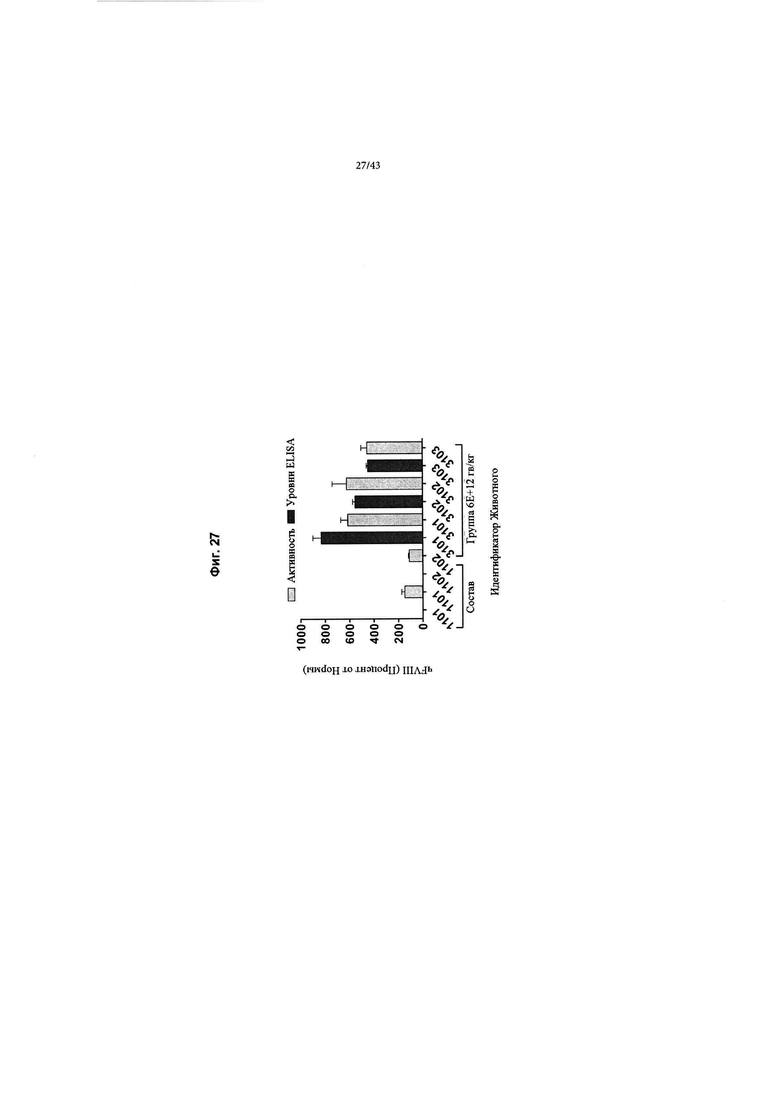
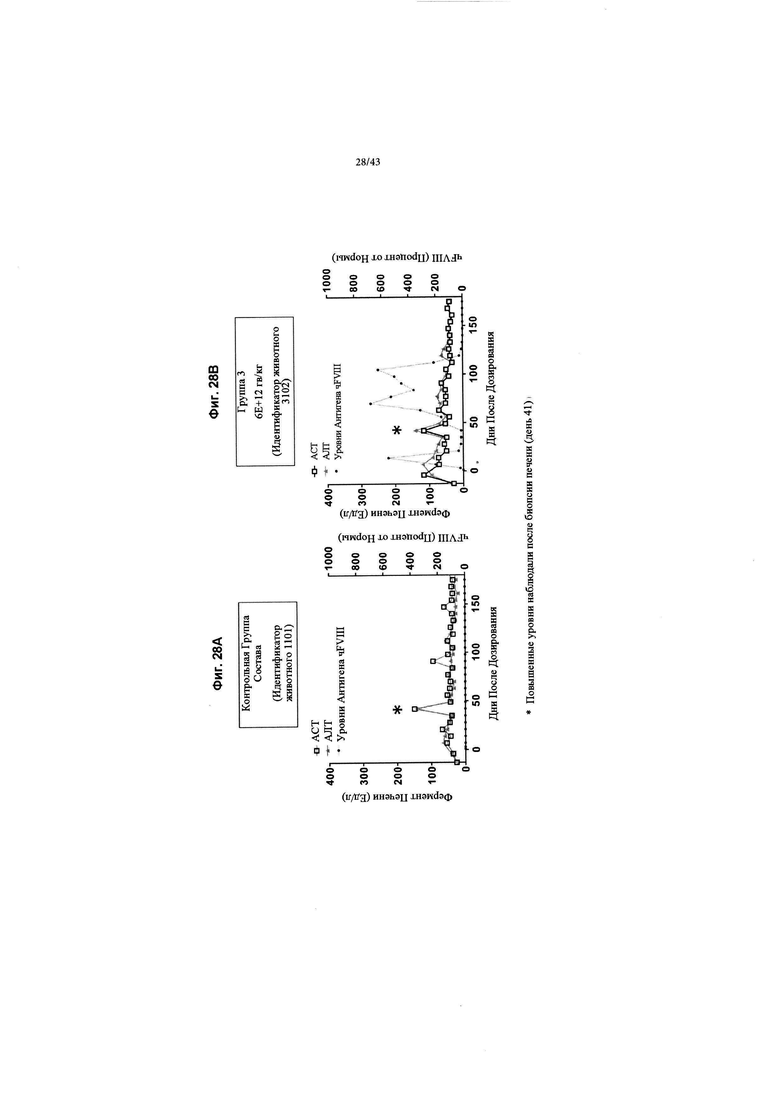
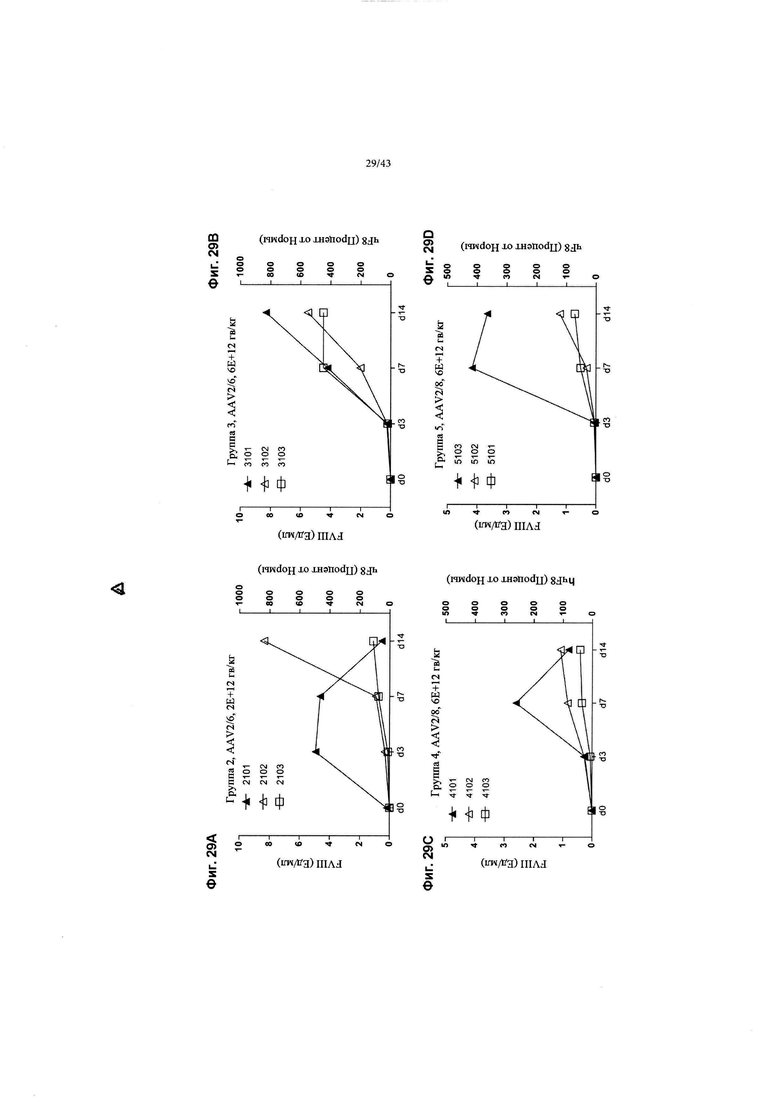
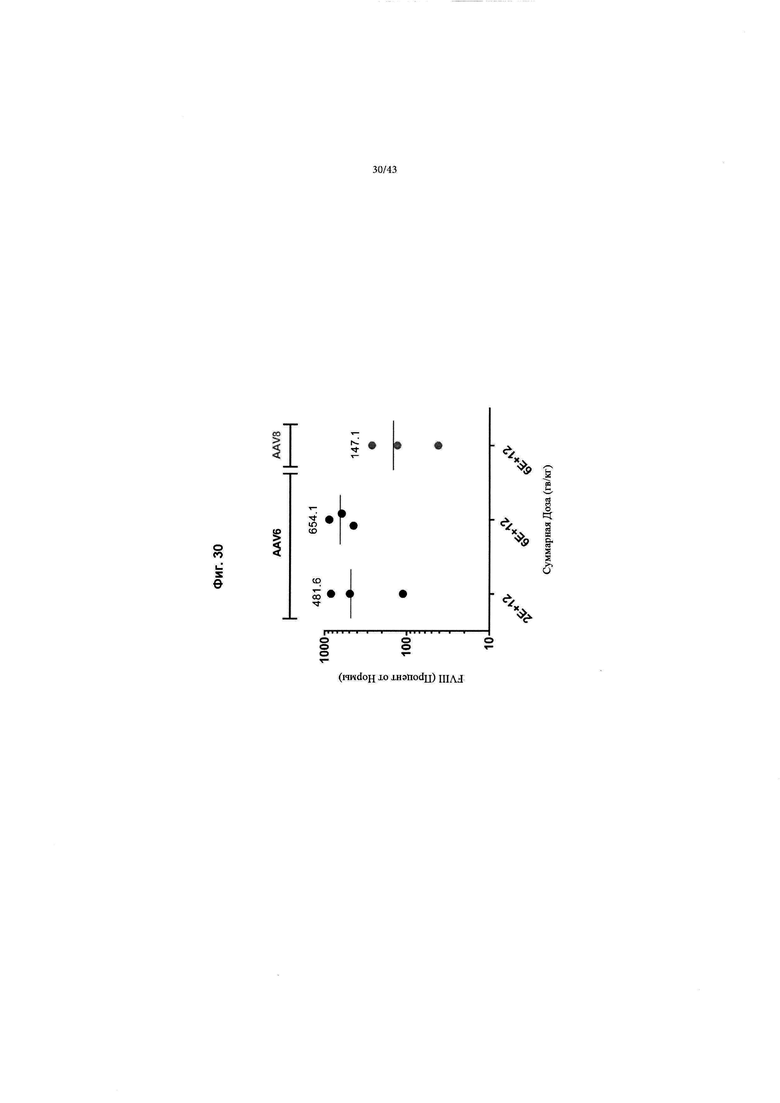
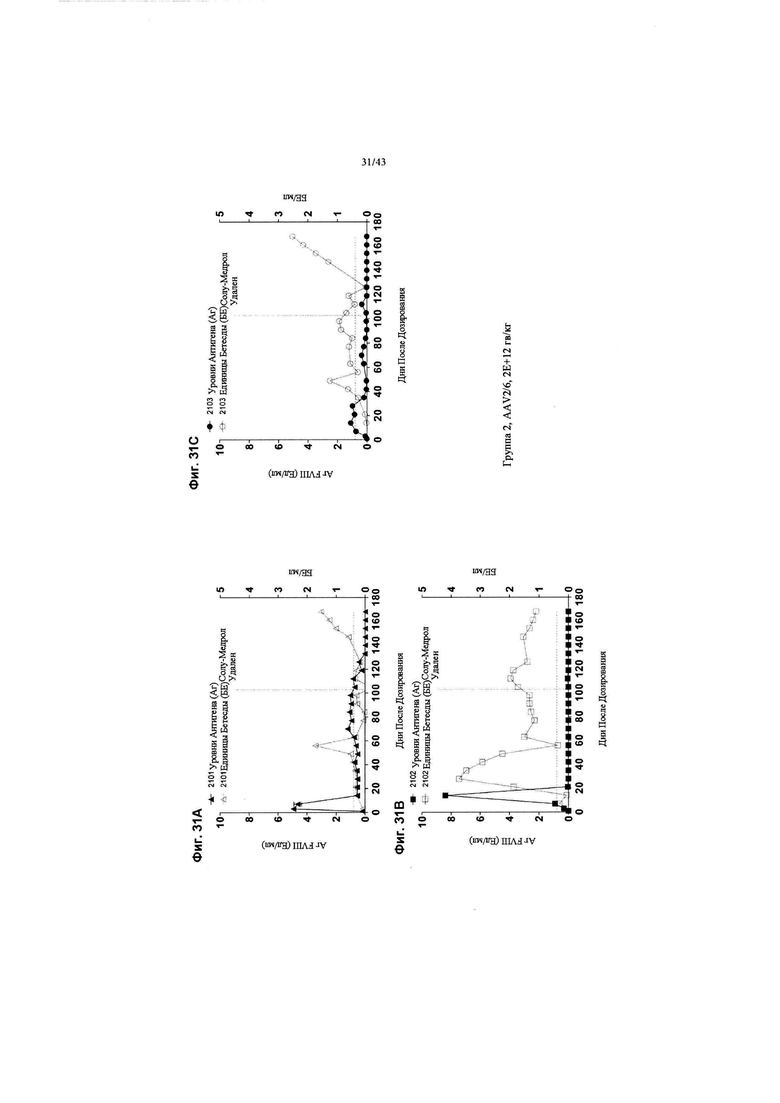
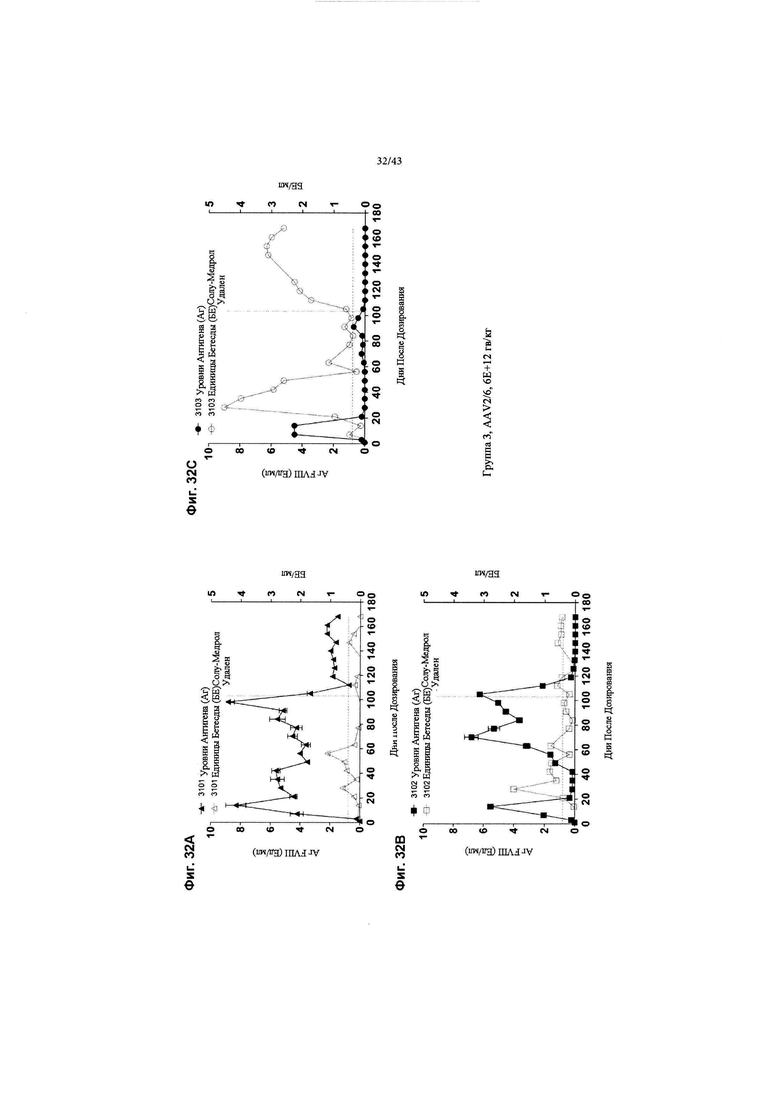

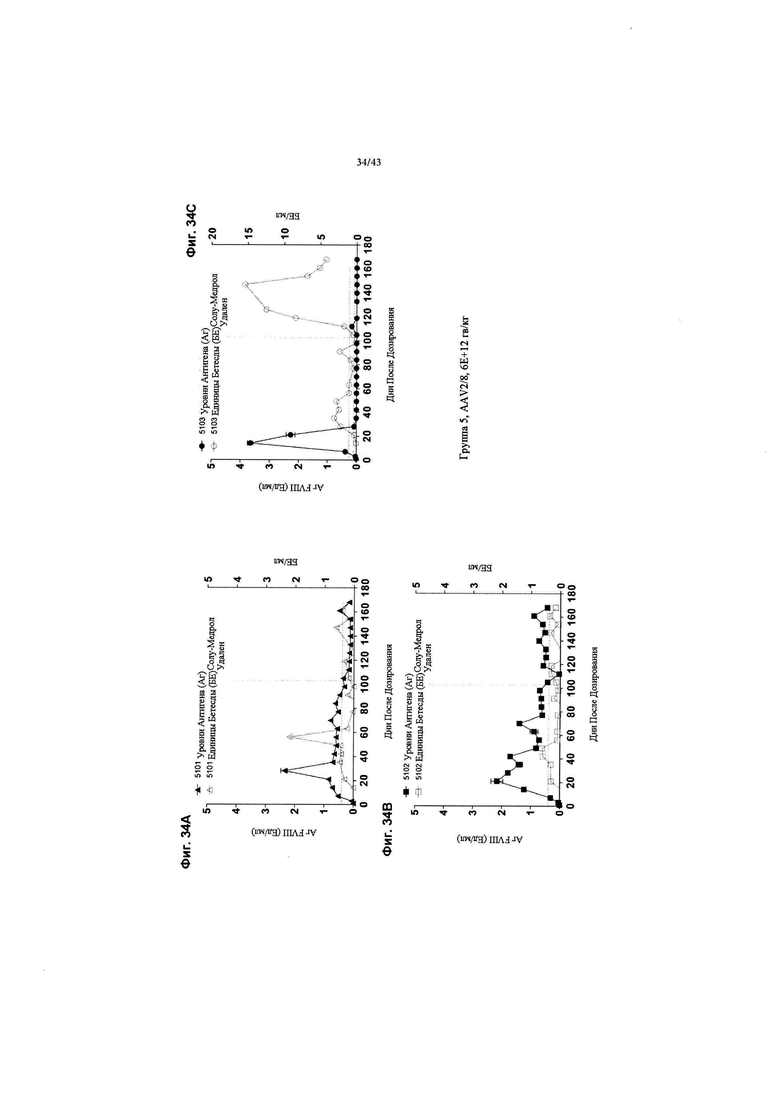
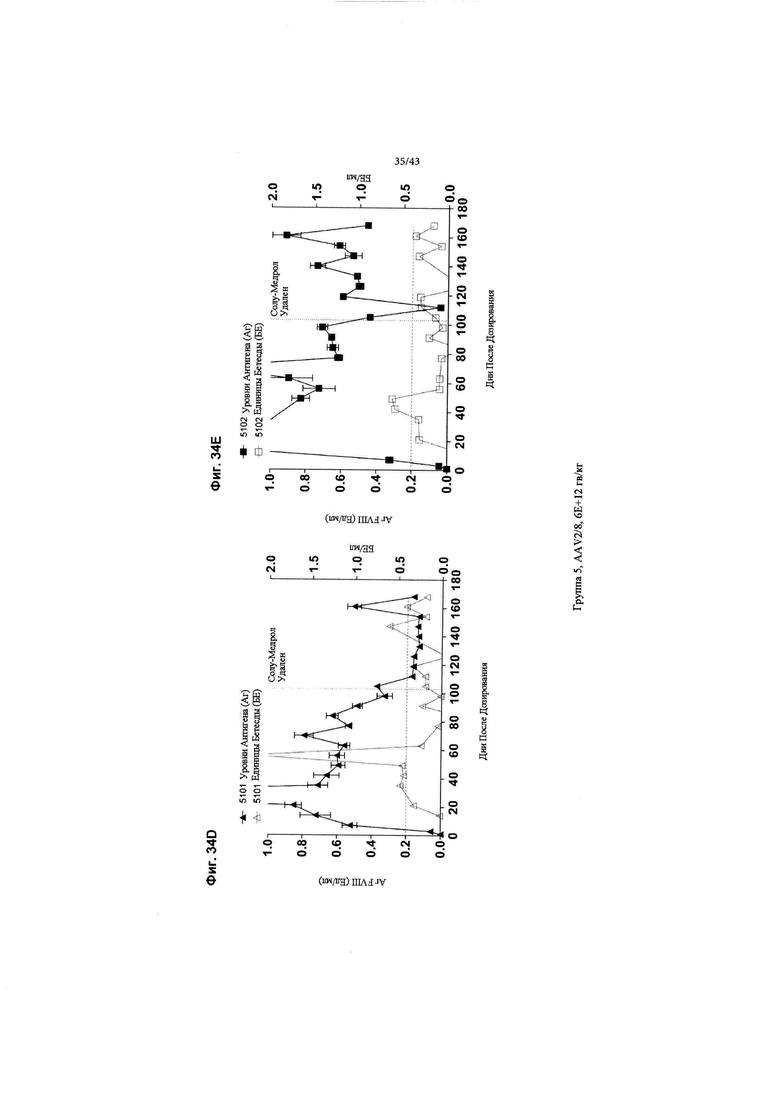

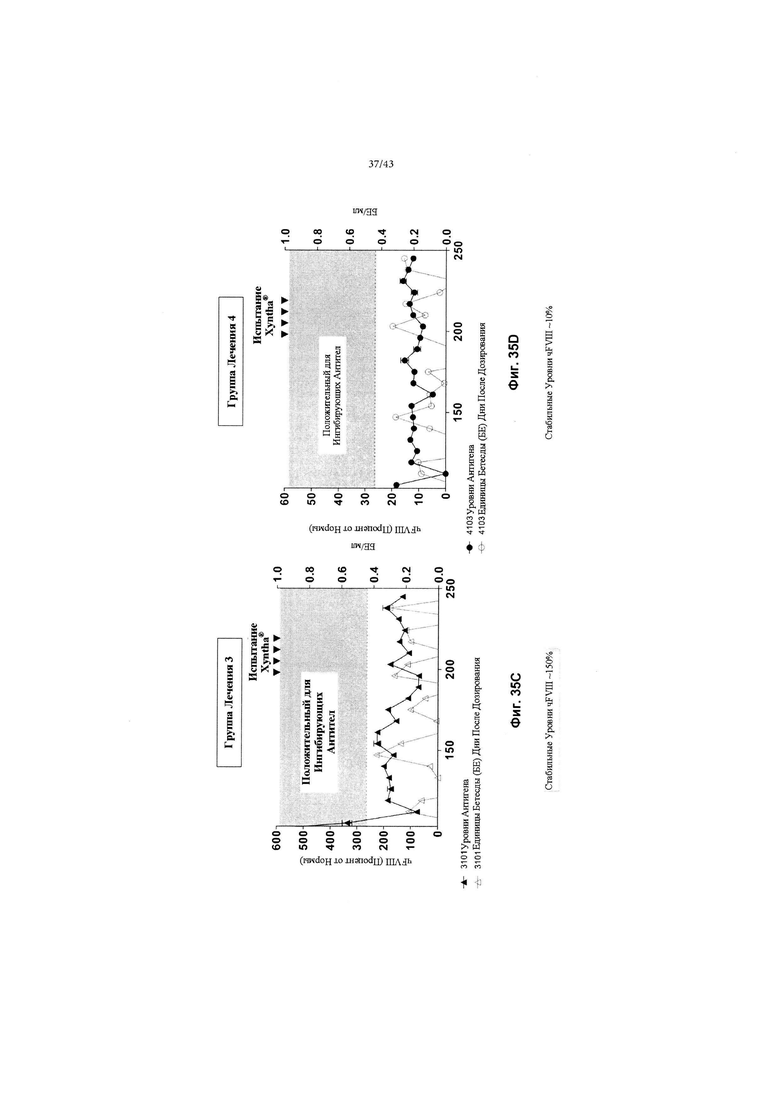
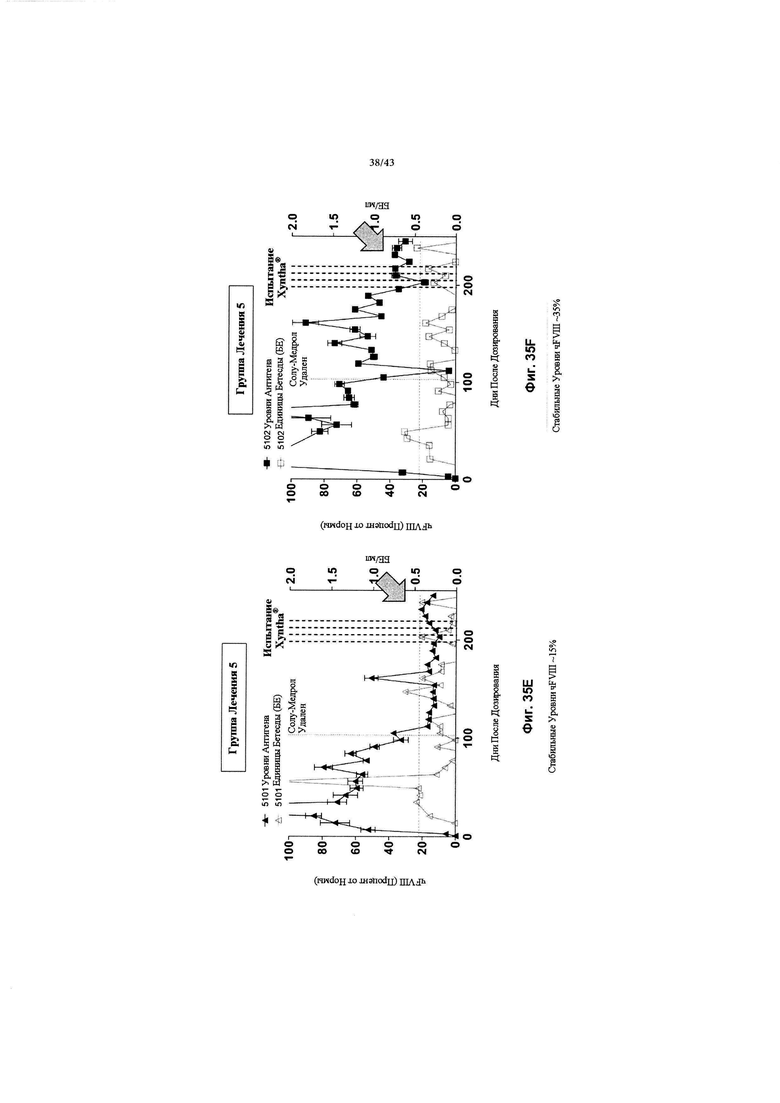
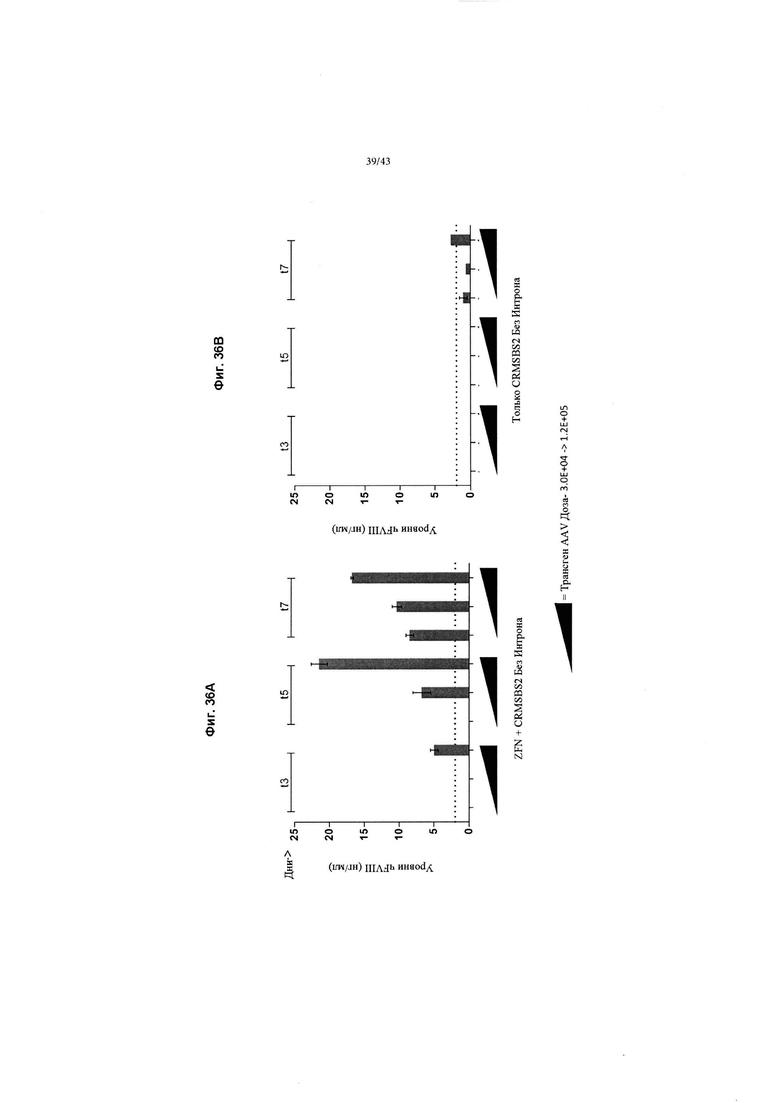
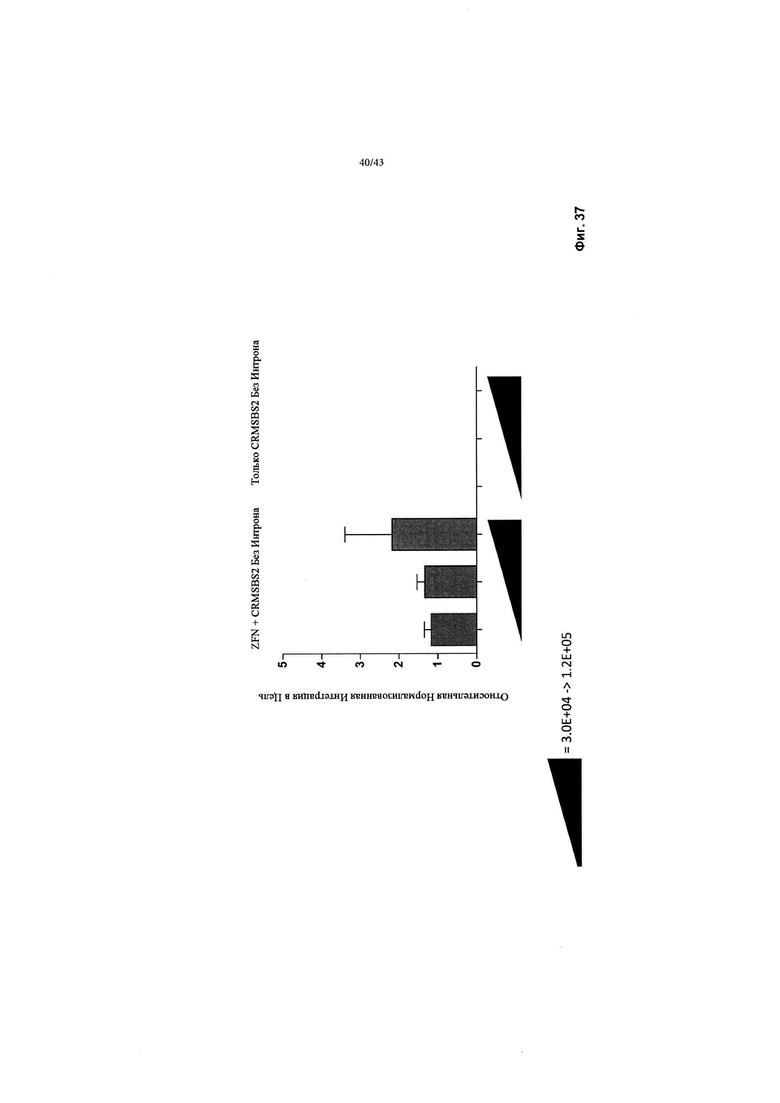
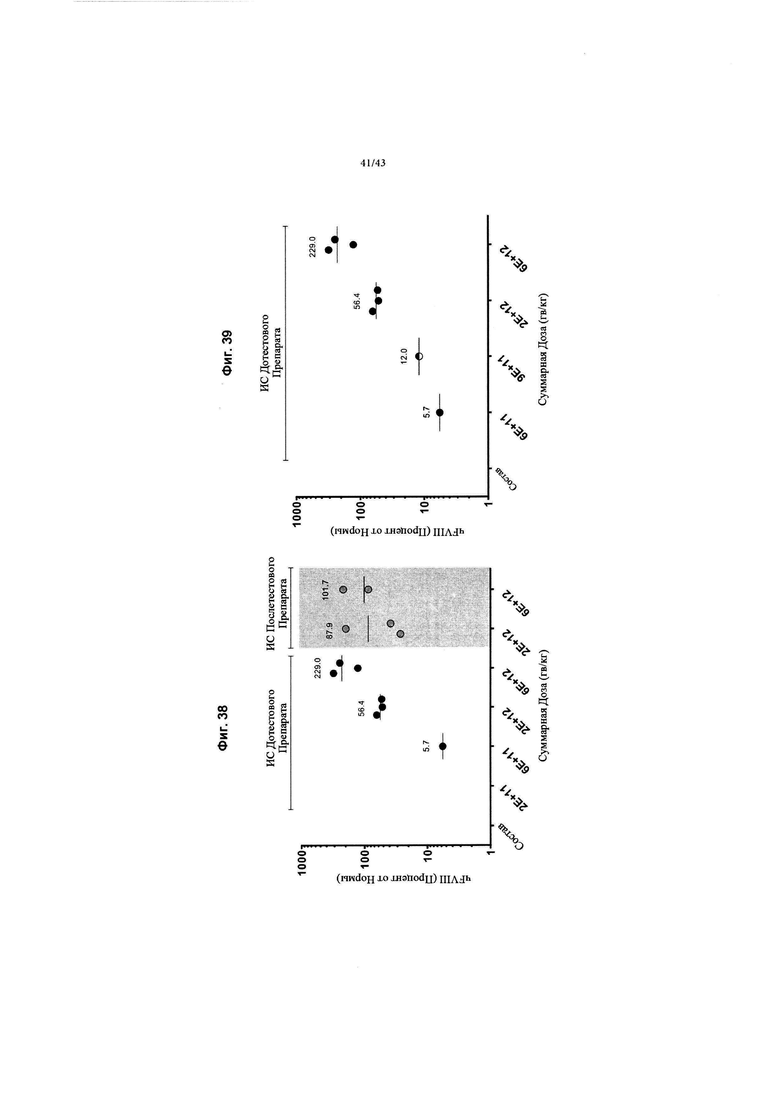

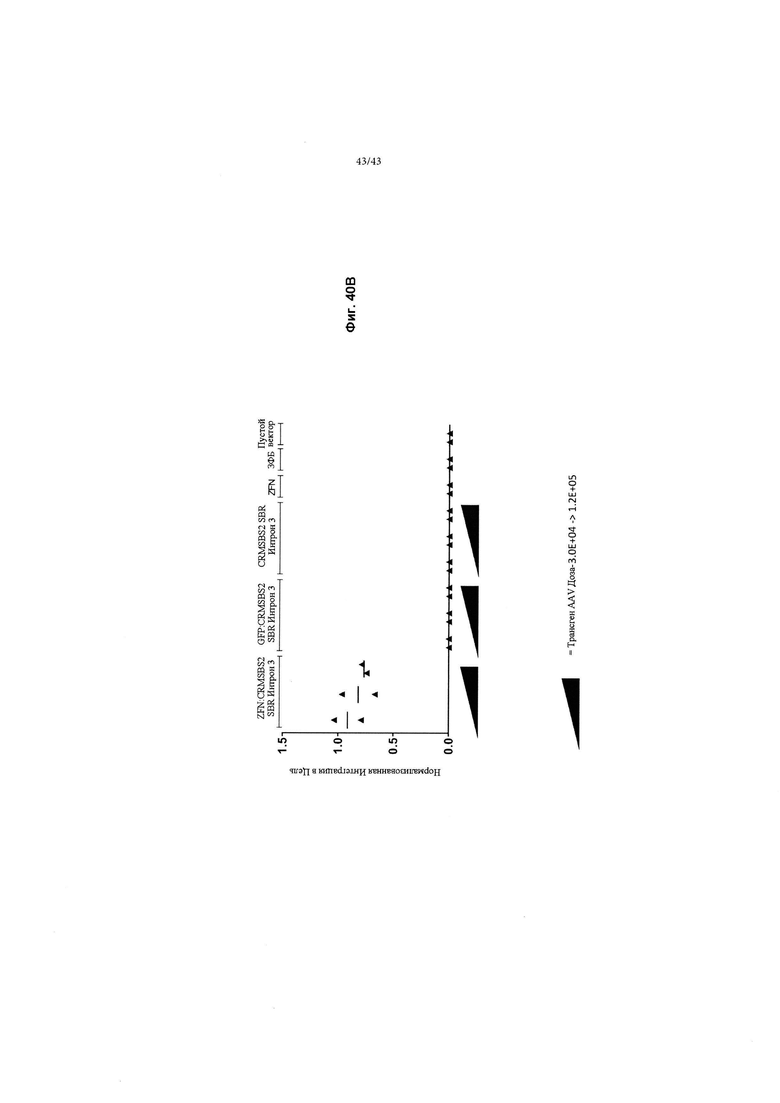
Группа изобретений относится к генной терапии, в частности к целенаправленной доставке трансген-кодирующих конструкций в печень для экспрессии терапевтических белков. Предложены полинуклеотидные экспрессионные конструкции, вектор AAV, содержащий полинуклеотидную экспрессионную конструкцию. Предложены также содержащие указанный вектор AAV фармацевтическая композиция для лечения гемофилии у субъекта, нуждающегося в этом, и фармацевтическая композиция для обеспечения трансгеном, который кодирует человеческий Фактор VIII, субъекта, нуждающегося в этом. Также предложены способ обеспечения человеческим Фактором VIII субъекта, нуждающегося в этом, и способ индуцирования толерантности у субъекта к человеческому Фактору VIII. Группа изобретений обеспечивает экспрессию трансгенов на высоких уровнях в клетках печени при использовании указанных экспрессионных конструкций. 9 н. и 6 з.п. ф-лы, 40 ил., 9 табл., 3 пр.
1. Полинуклеотидная экспрессионная конструкция, содержащая, от 5’ к 3’:
первую последовательность инсулятора, содержащую SEQ ID NO: 28,
последовательность энхансера Серпина 1, содержащую SEQ ID NO: 36,
последовательность минимального промотора TTR,
последовательность интрона, содержащую нуклеотиды 340-432 последовательности SEQ ID NO: 37,
трансген, кодирующий человеческий Фактор VIII,
последовательность сигнала полиаденилирования и
вторую последовательность инсулятора, содержащую SEQ ID NO: 30.
2. Полинуклеотидная экспрессионная конструкция, содержащая SEQ ID NO: 37.
3. Полинуклеотидная экспрессионная конструкция, содержащая SEQ ID NO: 34.
4. Вектор AAV, содержащий полинуклеотидную экспрессионную конструкцию по любому из пп. 1-3, отличающийся тем, что полинуклеотидная экспрессионная конструкция находится между 5' и 3' инвертированными концевыми повторами (ИКП) вектора AAV, причем указанный вектор AAV обеспечивает возможность экспрессии трансгена в клетке, трансдуцированной указанным вектором.
5. Вектор AAV по п. 4, отличающийся тем, что ИКП представляют собой ИКП AAV2.
6. Вектор AAV по п. 4 или 5, отличающийся тем, что вектор содержит капсид AAV6.
7. Фармацевтическая композиция для лечения гемофилии у субъекта, нуждающегося в этом, содержащая вектор AAV по пп. 4-6.
8. Фармацевтическая композиция для обеспечения трансгеном, который кодирует человеческий Фактор VIII, субъекта, нуждающегося в этом, причем указанная композиция содержит вектор AAV по любому из пп. 4-6.
9. Способ обеспечения человеческим Фактором VIII субъекта, нуждающегося в этом, при этом способ включает введение в печень субъекта вектора AAV по любому из пп. 4-6 или фармацевтической композиции по п. 7 или 8, при этом человеческий Фактор VIII продуцируется в субъекте, причем у указанного субъекта недостаточен уровень Фактора VIII, и необязательно указанный субъект страдает гемофилией.
10. Способ по п. 9, согласно которому трансген интегрируется в геном клетки печени.
11. Способ по п. 9 или 10, дополнительно включающий введение одной или более нуклеаз субъекту, при этом нуклеаза расщепляет эндогенный ген альбумина и трансген интегрируется в эндогенный ген альбумина.
12. Способ индуцирования толерантности у субъекта к человеческому Фактору VIII, причем способ включает модификацию клетки у указанного субъекта с целью получения человеческого Фактора VIII в соответствии со способом по любому из пп. 9-11, и
лечение указанного субъекта одним или более стероидами и/или ингибиторами В-клеток, так что субъект становится толерантным к человеческому Фактору VIII.
13. Способ обеспечения человеческим Фактором VIII субъекта, нуждающегося в этом, причем способ включает введение в печень указанного субъекта вектора AAV, содержащего полинуклеотидную экспрессионную конструкцию, содержащую SEQ ID NO: 37, при этом человеческий Фактор VIII продуцируется в субъекте, причем у указанного субъекта недостаточен уровень Фактора VIII.
14. Способ по п. 13, отличающийся тем, что субъект страдает гемофилией A.
15. Способ по любому из пп. 9-14, отличающийся тем, что субъект представляет собой человека.
| WO 2014064277 A1, 01.05.2014 | |||
| MCINTOSH J | |||
| ET AL | |||
| Therapeutic levels of FVIII following a single peripheral vein administration of rAAV vector encoding a novel human factor VIII variant | |||
| Blood | |||
| Многоступенчатая активно-реактивная турбина | 1924 |
|
SU2013A1 |
| NATHWANI A.C | |||
| ET AL | |||
| Adenovirus-Associated Virus Vector-Mediated Gene Transfer in Hemophilia B | |||
| Способ приготовления лака | 1924 |
|
SU2011A1 |

