Уровень техники
Настоящее изобретение относится к способам диагностики и лечения злокачественной опухоли, ингибиторам пролиферации клеток и средствам против злокачественной опухоли.
Предпосылки изобретения
Член семейства эпидермальных факторов роста (в дальнейшем в настоящем описании обозначаемого как EGF), очищенный Toyoda et al., был назван "эпирегулином (EREG)". Известно, что EREG функционирует в качестве ингибитора роста злокачественной опухоли, который индуцирует морфологические изменения клеток HeLa (непатентный документ 1). Аминокислотная последовательность полученного из мыши EREG (зрелый белок), очищенного Toyoda et al., состоит из 46 аминокислотных остатков и обладает степенью идентичности последовательности с другими членами семейства EGF, составляющей приблизительно 24-50%. EREG мыши демонстрировал низкую аффинность в отношении рецептора EGF на клетках A431 (клеточная линия эпителиальной карциномы человека). Анализ с клонированием и экспрессией гена EREG человека, проведенный Toyoda et al., показал, что, несмотря на то, что другие члены семейства EGF экспрессируются повсеместно в тканях человека, экспрессия EREG детектируется в макрофагах, плаценте и различных типах злокачественных клеток (непатентный документ 2). Кроме того, было показано, что растворимая форма EREG оказывает эффект подавления пролиферации на несколько типов злокачественных клеток (WO 94/29340).
Takahashi et al. показали, что активация Erk (MPK3) и p38 (MAPK14) в дифференцированных гладкомышечных клетках (в дальнейшем в настоящем описании обозначаемых как VSMC) от крыс индуцирует дифференцировку клеток. Более того, было показано, что EREG, секретируемый VSMC, действует в качестве аутокринного и/или паракринного фактора дифференцировки. Гомодимер ненасыщенной лизофосфатидной кислоты и PDGFB, который может действовать в качестве факторов дифференцировки VSMC, быстро повышал экспрессию мРНК EREG зависимым от MAPK Erk и p38 образом. Анализ посредством полимеразной цепной реакции с обратной транскриптазой (в дальнейшем в настоящем описании обозначаемой как ОТ-ПЦР) и иммуногистохимический анализ или иммуногистохимия (в дальнейшем в настоящем описании обозначаемая как IHC) выявил локализованную экспрессию EREG в атеросклеротических артериях и поврежденных баллоном артериях крысы. Исходя из этих результатов, Takahashi и другие сделали предположение, что EREG может быть вовлечен в прогрессирование ремоделирования сосудов, такого как атеросклероз (непатентный документ 3).
Minn et al. идентифицировали несколько генных кластеров, связанных с метастазами в легкое рака молочной железы, с помощью селекции in vivo, транскриптомного анализа, функционального анализа и клинического исследования, и показали, что EREG является одним из указанных генов (непатентный документ 4).
Более того, Shirasawa et al. показали, что EREG экспрессируется не только в кератиноцитах, но также в тканевых макрофагах, и что у мышей, нокаутированных EREG, развивается хронический дерматит. Осмотр при анализе таких мышей показал, что EREG играет важную роль в иммунных и воспалительных ответах кератиноцитов и макрофагов на границе с внешней средой (непатентный документ 5).
Как описано выше, была показана связь между EREG и дерматитом, метастазами злокачественной опухоли и атеросклерозом. Однако в настоящее время все еще отсутствует конкретное описание эффекта связывающих EREG антител, которые обладают нейтрализующей активностью и цитотоксической активностью на экспрессирующие EREG злокачественные клетки.
[Непатентный документ 1] J. Biol. Chem. 270: 7495-7500, 1995.
[Непатентный документ 2] Biochem. J. 326: 69-75, 1997.
[Непатентный документ 3] Circulation 108: 2524-2529, 2003.
[Непатентный документ 4] Nature 436: 518-524, 2005.
[Непатентный документ 5] Proc. Nat. Acad. Sci. 101: 13921-13926, 2004.
Описание изобретения
[Проблемы, решаемые посредством изобретения]
Задачей настоящего изобретения является предоставление антител против EREG и их применение. Более конкретно, задачей настоящего изобретения является предоставление новых способов диагностики и лечения злокачественной опухоли с использованием антител против EREG, новых ингибиторов пролиферации клеток и средств против злокачественной опухоли, содержащих антитела против EREG, и новых антител против EREG.
[Способы решения проблем]
Авторы настоящего изобретения открыли, что EREG высоко экспрессируется в злокачественных клетках, таких как клетки рака толстого кишечника. Более того, когда определяли активность комплемент-зависимой цитотоксичности (CDC) и антитело-зависимой клеточно-опосредуемой цитотоксичности (ADCC) антител против EREG, было обнаружено, что антитела против EREG обладают активностью CDC и активностью ADCC в экспрессирующих EREG клетках. Авторы настоящего изобретения также показали, что антитела против EREG оказывают эффект подавления пролиферации на злокачественные клеточные линии посредством нейтрализации. Более того, исходя из указанных выше открытий, авторы настоящего изобретения обнаружили, что антитела против EREG были эффективны в отношении диагностики, профилактики и лечения различных типов первичных и метастазирующих злокачественных опухолей, и, таким образом, дополнили настоящее изобретение. Более конкретно, авторы настоящего изобретения дополнили настоящее изобретение открытием того, EREG пригоден в качестве инструмента для лечения или диагностики злокачественных опухолей, при которых усиливается экспрессия EREG, включая рак толстого кишечника, аденокарциному легкого, рак поджелудочной железы, рак желудка и рак почки.
Настоящее изобретение относится к фармацевтическим композициям, содержащим в качестве активного ингредиента антитело, которое связывается с белком EREG. Также настоящее изобретение относится к ингибиторам пролиферации клеток, содержащим в качестве активного ингредиента антитело, которое связывается с белком EREG. Кроме того, настоящее изобретение относится к средствам против злокачественной опухоли, содержащим в качестве активного ингредиента антитело, которое связывается с белком EREG. Предпочтительно антитело, которое связывается с белком EREG, обладает цитотоксической активностью. Более предпочтительно антитело также обладает нейтрализующей активностью. В предпочтительном варианте осуществления настоящего изобретения злокачественные опухоли, на которые можно нацеливать лечение, представляют собой рак толстого кишечника, аденокарциному легкого, рак поджелудочной железы, рак желудка и рак почки. Средства против злокачественной опухоли, содержащие антитело против EREG согласно изобретению, пригодны для лечения таких злокачественных опухолей, которые, главным образом, представляют собой первичные или метастазирующие злокачественные опухоли и обладают повышенной экспрессией EREG. Особенно предпочтительными мишенями для лечения согласно изобретению являются первичный рак толстого кишечника, метастазирующий рак толстого кишечника и рак поджелудочной железы.
Более того, настоящее изобретение относится к фармацевтическим композициям, содержащим антитело, которое связывается с белком EREG, и фармацевтически приемлемый носитель. Фармацевтические композиции согласно изобретению пригодны для лечения и/или профилактики злокачественных опухолей, которые обладают повышенной экспрессией EREG. Таким образом, настоящее изобретение относится к применению антитела, которое связывается с белком EREG, для изготовления фармацевтических композиций для лечения и/или профилактики злокачественной опухоли.
В другом варианте осуществления настоящее изобретение относится к способам индукции клеточного повреждения в клетках, которые экспрессируют белок EREG, посредством контактирования экспрессирующих EREG клеток с антителом, которое связывается с белком EREG. Также настоящее изобретение относится к способам подавления пролиферации клеток, которые экспрессируют белок EREG, посредством контактирования экспрессирующих белок EREG клеток с антителом, которое связывается с белком EREG. Антитело, которое связывается с белком EREG, предпочтительно обладает цитотоксической активностью. Клетки, которые экспрессируют белок EREG, предпочтительно представляют собой злокачественные клетки.
Более того, в другом варианте осуществления настоящее изобретение относится к антителам, которые связываются с белком EREG и обладают цитотоксической активностью в клетках, которые экспрессируют белок EREG. Предпочтительно цитотоксическая активность представляет собой активность ADCC. Предпочтительно цитотоксическая активность представляет собой активность CDC. Также настоящее изобретение относится к антителам, с которыми связано цитотоксическое вещество. В настоящем изобретении цитотоксические вещества, которые могут быть связаны с антителом, включают химиотерапевтические средства, радиоактивные изотопы и токсические пептиды. Предпочтительно в настоящем изобретении антитело само по себе обладает цитотоксической активностью.
Кроме того, настоящее изобретение относится к антителам, которые связываются с белком EREG и которые обладают цитотоксической активностью и нейтрализующей активностью против экспрессирующих белок EREG клеток.
В другом варианте осуществления настоящее изобретение относится к применению белка EREG в качестве диагностического маркера злокачественной опухоли.
Более того, в другом варианте осуществления настоящее изобретение относится к способам диагностики злокачественной опухоли, которые включают детектирование белка EREG с использованием антитела, которое связывается с белком EREG. В способах согласно изобретению предпочтительно проводят детектирование внеклеточного участка белка EREG. Предпочтительно способы согласно изобретению осуществляют с использованием антитела, которое распознает белок EREG. Предпочтительно в способах согласно изобретению проводят детектирование белка EREG в крови, сыворотке или плазме или белка EREG, выделенного из клеток.
В другом варианте осуществления настоящее изобретение относится к способам диагностики злокачественной опухоли, которые включает следующие стадии:
(a) получение образца от субъекта и
(b) детектирование белка EREG, присутствующего в полученном образце, с использованием антитела, которое связывается с белком EREG.
В настоящем изобретении в качестве указанного выше образца можно использовать любой образец при условии, что его можно получить от субъекта. В одном варианте осуществления используют образец крови, полученный от субъекта. Предпочтительным образцом крови в настоящем изобретении является сыворотка. В другом варианте осуществления можно использовать образцы, полученные от субъекта хирургически или посредством биопсии. Способы диагностики можно использовать для любой злокачественной опухоли, при условии, что она представляет собой злокачественную опухоль, при которой злокачественные клетки-мишени экспрессируют белок EREG. Злокачественные опухоли, которые являются предпочтительными для настоящего изобретения, представляют собой рак толстого кишечника, аденокарциному легкого, рак поджелудочной железы, рак желудка и рак почки. С помощью настоящего изобретения можно диагностировать как первичные, так и метастатические очаги таких злокачественных опухолей. Злокачественные опухоли, которые являются особенно предпочтительными, представляют собой первичный рак толстого кишечника, метастазирующий рак толстого кишечника и рак поджелудочной железы.
В настоящем изобретении стадию получения образца от субъекта также можно выразить как стадию предоставления образца, полученного от субъекта.
Более того, в другом варианте осуществления настоящее изобретение относится к способам диагностики злокачественной опухоли, которые включают в себя стадии: (1) введения субъекту меченного радиоактивным изотопом антитела, которое связывается с белком EREG; и (2) детектирования накопления радиоактивного изотопа. В определенном варианте осуществления радиоактивный изотоп представляет собой позитронно-активный нуклид. Предпочтительный позитронно-активный нуклид согласно изобретению может быть выбран, например, из группы, состоящей из 11C, 13N, 15O, 18F, 45Ti, 55Co, 64Cu, 66Ga, 68Ga, 76Br, 89Zr и 124I.
Более того, в другом варианте осуществления настоящее изобретение относится к способам диагностики злокачественной опухоли, где выявляют экспрессию гена, кодирующего белок EREG.
Более того, в другом варианте осуществления настоящее изобретение относится к диагностическим средствам и наборам, предназначенным для применения в способах диагностики согласно изобретению.
Кроме того, настоящее изобретение относится к способам скрининга соединений-кандидатов в качестве лекарственных средств против злокачественной опухоли. В настоящем изобретении соединения могут быть выбраны в качестве лекарственного средства-кандидата против злокачественной опухоли с использованием в качестве показателя, например, уровня экспрессии EREG. Альтернативно, соединение может быть выбрано в качестве лекарственного средства-кандидата против злокачественной опухоли с использованием в качестве показателя нейтрализующего эффекта против активности EREG в отношении стимуляции клеточной пролиферации.
Более конкретно, настоящее изобретение относится к следующему:
[1] фармацевтической композиции, содержащей в качестве активного ингредиента антитело, которое связывается с белком EREG;
[2] ингибитору клеточной пролиферации, содержащему в качестве активного ингредиента антитело, которое связывается с белком EREG;
[3] средству против злокачественной опухоли содержащему в качестве активного ингредиента антитело, которое связывается с белком EREG;
[4] средству против злокачественной опухоли по [3], где антитело, которое связывается с белком EREG, представляет собой антитело, которое обладает ингибирующей активностью в отношении клеточной пролиферации;
[5] средству против злокачественной опухоли по [3] или [4], где антитело, которое связывается с белком EREG, представляет собой антитело согласно любому из (1)-(57), ниже:
(1) антитело, содержащее H-цепь, имеющую аминокислотную последовательность SEQ ID NO: 2 в качестве CDR1, аминокислотную последовательность SEQ ID NO: 4 в качестве CDR2 и аминокислотную последовательность SEQ ID NO: 6 в качестве CDR3;
(2) антитело, содержащее H-цепь согласно (1), где H-цепь имеет аминокислотную последовательность из положений 117-452 в аминокислотной последовательности SEQ ID NO: 8 в качестве CH;
(3) антитело, содержащее H-цепь согласно (1), где H-цепь имеет аминокислотную последовательность из положений 117-446 в аминокислотной последовательности SEQ ID NO: 10 в качестве CH;
(4) антитело, содержащее L-цепь, имеющую аминокислотную последовательность SEQ ID NO: 12 в качестве CDR1, аминокислотную последовательность SEQ ID NO: 14 в качестве CDR2 и аминокислотную последовательность SEQ ID NO: 16 в качестве CDR3;
(5) антитело, содержащее L-цепь согласно (4), где L-цепь имеет аминокислотную последовательность из положений 107-213 в аминокислотной последовательности SEQ ID NO: 18 в качестве CL;
(6) антитело, содержащее L-цепь согласно (4), где L-цепь имеет аминокислотную последовательность из положений 107-213 в аминокислотной последовательности SEQ ID NO: 20 в качестве CL;
(7) антитело, содержащее H-цепь согласно (1) и L-цепь согласно (4);
(8) антитело, содержащее H-цепь согласно (2) и L-цепь согласно (5);
(9) антитело, содержащее H-цепь согласно (3) и L-цепь согласно (6);
(10) антитело, содержащее H-цепь, имеющую аминокислотную последовательность SEQ ID NO: 49 в качестве CDR1, аминокислотную последовательность SEQ ID NO: 51 в качестве CDR2 и аминокислотную последовательность SEQ ID NO: 53 в качестве CDR3;
(11) антитело, содержащее H-цепь согласно (10), где H-цепь имеет CH, полученную из мышиного IgG1;
(12) антитело, содержащее H-цепь согласно (10), где H-цепь имеет аминокислотную последовательность из положений 117-446 в аминокислотной последовательности SEQ ID NO: 10 в качестве CH;
(13) антитело, содержащее L-цепь, имеющую аминокислотную последовательность SEQ ID NO: 55 в качестве CDR1, аминокислотную последовательность SEQ ID NO: 57 в качестве CDR2 и аминокислотную последовательность SEQ ID NO: 59 в качестве CDR3;
(14) антитело, содержащее L-цепь согласно (13), где L-цепь имеет CL, полученную из мышиной κ-цепи;
(15) антитело, содержащее L-цепь согласно (13), где L-цепь имеет аминокислотную последовательность из положений 107-213 в аминокислотной последовательности SEQ ID NO: 20 в качестве CL;
(16) антитело, содержащее H-цепь согласно (10) и L-цепь согласно (13);
(17) антитело, содержащее H-цепь согласно (11) и L-цепь согласно (14);
(18) антитело, содержащее H-цепь согласно (12) и L-цепь согласно (15);
(19) антитело, содержащее H-цепь, имеющую аминокислотную последовательность SEQ ID NO: 61 в качестве CDR1, аминокислотную последовательность SEQ ID NO: 63 в качестве CDR2 и аминокислотную последовательность SEQ ID NO: 65 в качестве CDR3;
(20) антитело, содержащее H-цепь согласно (19), где H-цепь имеет CH, полученную из мышиного IgG1;
(21) антитело, содержащее H-цепь согласно (19), где H-цепь имеет аминокислотную последовательность из положений 117-446 в аминокислотной последовательности SEQ ID NO: 10 в качестве CH;
(22) антитело, содержащее L-цепь, имеющую аминокислотную последовательность SEQ ID NO: 67 в качестве CDR1, аминокислотную последовательность SEQ ID NO: 69 в качестве CDR2 и аминокислотную последовательность SEQ ID NO: 71 в качестве CDR3;
(23) антитело, содержащее L-цепь согласно (22), где L-цепь имеет CL, полученную из мышиной κ-цепи;
(24) антитело, содержащее L-цепь согласно (22), где L-цепь имеет аминокислотную последовательность из положений 107-213 в аминокислотной последовательности SEQ ID NO: 20 в качестве CL;
(25) антитело, содержащее H-цепь согласно (19) и L-цепь согласно (22);
(26) антитело, содержащее H-цепь согласно (20) и L-цепь согласно (23);
(27) антитело, содержащее H-цепь согласно (21) и L-цепь согласно (24);
(28) антитело, содержащее H-цепь, имеющую аминокислотную последовательность SEQ ID NO: 73 в качестве CDR1, аминокислотную последовательность SEQ ID NO: 75 в качестве CDR2 и аминокислотную последовательность SEQ ID NO: 77 в качестве CDR3;
(29) антитело, содержащее H-цепь согласно (28), где H-цепь имеет CH, полученную из мышиного IgG1;
(30) антитело, содержащее H-цепь согласно (28), где H-цепь имеет аминокислотную последовательность из положений 117-446 в аминокислотной последовательности SEQ ID NO: 10 в качестве CH;
(31) антитело, содержащее L-цепь, имеющую аминокислотную последовательность SEQ ID NO: 79 в качестве CDR1, аминокислотную последовательность SEQ ID NO: 81 в качестве CDR2 и аминокислотную последовательность SEQ ID NO: 83 в качестве CDR3;
(32) антитело, содержащее L-цепь согласно (31), где L-цепь имеет CL, полученную из мышиной κ-цепи;
(33) антитело, содержащее L-цепь согласно (31), где L-цепь имеет аминокислотную последовательность из положений 107-213 в аминокислотной последовательности SEQ ID NO: 20 в качестве CL;
(34) антитело, содержащее H-цепь согласно (28) и L-цепь согласно (31);
(35) антитело, содержащее H-цепь согласно (29) и L-цепь согласно (32);
(36) антитело, содержащее H-цепь согласно (30) и L-цепь согласно (33);
(37) антитело, содержащее H-цепь, имеющую аминокислотную последовательность SEQ ID NO: 85 в качестве CDR1, аминокислотную последовательность SEQ ID NO: 87 в качестве CDR2 и аминокислотную последовательность SEQ ID NO: 89 в качестве CDR3;
(38) антитело, содержащее H-цепь согласно (37), где H-цепь имеет CH, полученную из мышиного IgG1;
(39) антитело, содержащее H-цепь согласно (37), где H-цепь имеет аминокислотную последовательность из положений 117-446 в аминокислотной последовательности SEQ ID NO: 10 в качестве CH;
(40) антитело, содержащее L-цепь, имеющую аминокислотную последовательность SEQ ID NO: 91 в качестве CDR1, аминокислотную последовательность SEQ ID NO: 93 в качестве CDR2 и аминокислотную последовательность SEQ ID NO: 95 в качестве CDR3;
(41) антитело, содержащее L-цепь согласно (40), где L-цепь имеет CL, полученную из мышиной κ-цепи;
(42) антитело, содержащее L-цепь согласно (40), где L-цепь имеет аминокислотную последовательность из положений 107-213 в аминокислотной последовательности SEQ ID NO: 20 в качестве CL;
(43) антитело, содержащее H-цепь согласно (37) и L-цепь согласно (40);
(44) антитело, содержащее H-цепь согласно (38) и L-цепь согласно (41);
(45) антитело, содержащее H-цепь согласно (39) и L-цепь согласно (42);
(46) антитело, содержащее H-цепь, имеющую аминокислотную последовательность SEQ ID NO: 97 в качестве CDR1, аминокислотную последовательность SEQ ID NO: 99 в качестве CDR2 и аминокислотную последовательность SEQ ID NO: 101 в качестве CDR3;
(47) антитело, содержащее H-цепь согласно (46), где H-цепь имеет CH, полученную из мышиного IgG1;
(48) антитело, содержащее H-цепь согласно (46), где H-цепь имеет аминокислотную последовательность из положений 117-446 в аминокислотной последовательности SEQ ID NO: 10 в качестве CH;
(49) антитело, содержащее L-цепь, имеющую аминокислотную последовательность SEQ ID NO: 103 в качестве CDR1, аминокислотную последовательность SEQ ID NO: 105 в качестве CDR2 и аминокислотную последовательность SEQ ID NO: 107 в качестве CDR3;
(50) антитело, содержащее L-цепь согласно (49), где L-цепь имеет CL, полученную из мышиной κ-цепи;
(51) антитело, содержащее L-цепь согласно (49), где L-цепь имеет аминокислотную последовательность из положений 107-213 в аминокислотной последовательности SEQ ID NO: 20 в качестве CL;
(52) антитело, содержащее H-цепь согласно (46) и L-цепь согласно (49);
(53) антитело, содержащее H-цепь согласно (47) и L-цепь согласно (50);
(54) антитело, содержащее H-цепь согласно (48) и L-цепь согласно (51);
(55) антитело, содержащее одну или несколько аминокислотных замен, делеций, добавлений и/или инсерций в антителе согласно любому из (1)-(54), которое обладает активностью, эквивалентной активности антитела согласно любому из (1)-(54);
(56) антитело, которое связывается с тем же эпитопом белка EREG, что и антитело согласно любому из (1)-(55); и
(57) мини-антитело антитела согласно любому из (1)-(56);
[6] средству против злокачественной опухоли по любому из [3]-[5], где антитело, которое связывается с белком EREG, распознает участок с Ala в положении 29 по Ser в положении 69, или с Val в положении 63 по Leu в положении 108, в аминокислотной последовательности SEQ ID NO: 21;
[7] средству против злокачественной опухоли по любому из [3]-[6], где злокачественная опухоль представляет собой любую злокачественную опухоль, выбранную из группы, состоящей из рака толстого кишечника, аденокарциномы легкого, рака поджелудочной железы, рака желудка и рака почки;
[8] средству против злокачественной опухоли по [7], где злокачественная опухоль представляет собой первичную злокачественную опухоль;
[9] средству против злокачественной опухоли по [7], где злокачественная опухоль представляет собой метастазирующую злокачественную опухоль;
[10] антителу, которое связывается с белком EREG и которое обладает активностью ингибирования клеточной пролиферации против клеток, экспрессирующих белок EREG;
[11] антителу по [10], где активность ингибирования клеточной пролиферации представляет собой цитотоксическую активность;
[12] антителу по [11], где цитотоксическая активность представляет собой активность ADCC;
[13] антителу по [11], где цитотоксическая активность представляет собой активность CDC;
[14] антителу по любому из [10]-[13], которое дополнительно обладает нейтрализующей активностью против белка EREG;
[15] антителу по [10], где активность ингибирования клеточной пролиферации представляет собой нейтрализующую активность;
[16] мини-антителу, полученному из антитела по [10];
[17] антителу по любому из [10]-[16], где с антителом связано химиотерапевтическое средство или токсический пептид;
[18] антителу, которое связывается с белком EREG, где с антителом связано любое цитотоксическое вещество, выбранное из группы, состоящей из химиотерапевтического средства, токсического пептида и радиоактивного изотопа;
[19] антителу по любому из [10]-[18], которое представляет собой антитело согласно любому из (1)-(57), ниже:
(1) антитело, содержащее H-цепь, имеющую аминокислотную последовательность SEQ ID NO: 2 в качестве CDR1, аминокислотную последовательность SEQ ID NO: 4 в качестве CDR2 и аминокислотную последовательность SEQ ID NO: 6 в качестве CDR3;
(2) антитело, содержащее H-цепь согласно (1), где H-цепь имеет аминокислотную последовательность из положений 117-452 в аминокислотной последовательности SEQ ID NO: 8 в качестве CH;
(3) антитело, содержащее H-цепь согласно (1), где H-цепь имеет аминокислотную последовательность из положений 117-446 в аминокислотной последовательности SEQ ID NO: 10 в качестве CH;
(4) антитело, содержащее L-цепь, имеющую аминокислотную последовательность SEQ ID NO: 12 в качестве CDR1, аминокислотную последовательность SEQ ID NO: 14 в качестве CDR2 и аминокислотную последовательность SEQ ID NO: 16 в качестве CDR3;
(5) антитело, содержащее L-цепь согласно (4), где L-цепь имеет аминокислотную последовательность из положений 107-213 в аминокислотной последовательности SEQ ID NO: 18 в качестве CL;
(6) антитело, содержащее L-цепь согласно (4), где L-цепь имеет аминокислотную последовательность из положений 107-213 в аминокислотной последовательности SEQ ID NO: 20 в качестве CL;
(7) антитело, содержащее H-цепь согласно (1) и L-цепь согласно (4);
(8) антитело, содержащее H-цепь согласно (2) и L-цепь согласно (5);
(9) антитело, содержащее H-цепь согласно (3) и L-цепь согласно (6);
(10) антитело, содержащее H-цепь, имеющую аминокислотную последовательность SEQ ID NO: 49 в качестве CDR1, аминокислотную последовательность SEQ ID NO: 51 в качестве CDR2 и аминокислотную последовательность SEQ ID NO: 53 в качестве CDR3;
(11) антитело, содержащее H-цепь согласно (10), где H-цепь имеет CH, полученную из мышиного IgG1;
(12) антитело, содержащее H-цепь согласно (10), где H-цепь имеет аминокислотную последовательность из положений 117-446 в аминокислотной последовательности SEQ ID NO: 10 в качестве CH;
(13) антитело, содержащее L-цепь, имеющую аминокислотную последовательность SEQ ID NO: 55 в качестве CDR1, аминокислотную последовательность SEQ ID NO: 57 в качестве CDR2 и аминокислотную последовательность SEQ ID NO: 59 в качестве CDR3;
(14) антитело, содержащее L-цепь согласно (13), где L-цепь имеет CL, полученную из мышиной κ-цепи;
(15) антитело, содержащее L-цепь согласно (13), где L-цепь имеет аминокислотную последовательность из положений 107-213 в аминокислотной последовательности SEQ ID NO: 20 в качестве CL;
(16) антитело, содержащее H-цепь согласно (10) и L-цепь согласно (13);
(17) антитело, содержащее H-цепь согласно (11) и L-цепь согласно (14);
(18) антитело, содержащее H-цепь согласно (12) и L-цепь согласно (15);
(19) антитело, содержащее H-цепь, имеющую аминокислотную последовательность SEQ ID NO: 61 в качестве CDR1, аминокислотную последовательность SEQ ID NO: 63 в качестве CDR2 и аминокислотную последовательность SEQ ID NO: 65 в качестве CDR3;
(20) антитело, содержащее H-цепь согласно (19), где H-цепь имеет CH, полученную из мышиного IgG1;
(21) антитело, содержащее H-цепь согласно (19), где H-цепь имеет аминокислотную последовательность из положений 117-446 в аминокислотной последовательности SEQ ID NO: 10 в качестве CH;
(22) антитело, содержащее L-цепь, имеющую аминокислотную последовательность SEQ ID NO: 67 в качестве CDR1, аминокислотную последовательность SEQ ID NO: 69 в качестве CDR2 и аминокислотную последовательность SEQ ID NO: 71 в качестве CDR3;
(23) антитело, содержащее L-цепь согласно (22), где L-цепь имеет CL, полученную из мышиной κ-цепи;
(24) антитело, содержащее L-цепь согласно (22), где L-цепь имеет аминокислотную последовательность из положений 107-213 в аминокислотной последовательности SEQ ID NO: 20 в качестве CL;
(25) антитело, содержащее H-цепь согласно (19) и L-цепь согласно (22);
(26) антитело, содержащее H-цепь согласно (20) и L-цепь согласно (23);
(27) антитело, содержащее H-цепь согласно (21) и L-цепь согласно (24);
(28) антитело, содержащее H-цепь, имеющую аминокислотную последовательность SEQ ID NO: 73 в качестве CDR1, аминокислотную последовательность SEQ ID NO: 75 в качестве CDR2 и аминокислотную последовательность SEQ ID NO: 77 в качестве CDR3;
(29) антитело, содержащее H-цепь согласно (28), где H-цепь имеет CH, полученную из мышиного IgG1;
(30) антитело, содержащее H-цепь согласно (28), где H-цепь имеет аминокислотную последовательность из положений 117-446 в аминокислотной последовательности SEQ ID NO: 10 в качестве CH;
(31) антитело, содержащее L-цепь, имеющую аминокислотную последовательность SEQ ID NO: 79 в качестве CDR1, аминокислотную последовательность SEQ ID NO: 81 в качестве CDR2 и аминокислотную последовательность SEQ ID NO: 83 в качестве CDR3;
(32) антитело, содержащее L-цепь согласно (31), где L-цепь имеет CL, полученную из мышиной κ-цепи;
(33) антитело, содержащее L-цепь согласно (31), где L-цепь имеет аминокислотную последовательность из положений 107-213 в аминокислотной последовательности SEQ ID NO: 20 в качестве CL;
(34) антитело, содержащее H-цепь согласно (28) и L-цепь согласно (31);
(35) антитело, содержащее H-цепь согласно (29) и L-цепь согласно (32);
(36) антитело, содержащее H-цепь согласно (30) и L-цепь согласно (33);
(37) антитело, содержащее H-цепь, имеющую аминокислотную последовательность SEQ ID NO: 85 в качестве CDR1, аминокислотную последовательность SEQ ID NO: 87 в качестве CDR2 и аминокислотную последовательность SEQ ID NO: 89 в качестве CDR3;
(38) антитело, содержащее H-цепь согласно (37), где H-цепь имеет CH, полученную из мышиного IgG1;
(39) антитело, содержащее H-цепь согласно (37), где H-цепь имеет аминокислотную последовательность из положений 117-446 в аминокислотной последовательности SEQ ID NO: 10 в качестве CH;
(40) антитело, содержащее L-цепь, имеющую аминокислотную последовательность SEQ ID NO: 91 в качестве CDR1, аминокислотную последовательность SEQ ID NO: 93 в качестве CDR2 и аминокислотную последовательность SEQ ID NO: 95 в качестве CDR3;
(41) антитело, содержащее L-цепь согласно (40), где L-цепь имеет CL, полученную из мышиной κ-цепи;
(42) антитело, содержащее L-цепь согласно (40), где L-цепь имеет аминокислотную последовательность из положений 107-213 в аминокислотной последовательности SEQ ID NO: 20 в качестве CL;
(43) антитело, содержащее H-цепь согласно (37) и L-цепь согласно (40);
(44) антитело, содержащее H-цепь согласно (38) и L-цепь согласно (41);
(45) антитело, содержащее H-цепь согласно (39) и L-цепь согласно (42);
(46) антитело, содержащее H-цепь, имеющую аминокислотную последовательность SEQ ID NO: 97 в качестве CDR1, аминокислотную последовательность SEQ ID NO: 99 в качестве CDR2 и аминокислотную последовательность SEQ ID NO: 101 в качестве CDR3;
(47) антитело, содержащее H-цепь согласно (46), где H-цепь имеет CH, полученную из мышиного IgG1;
(48) антитело, содержащее H-цепь согласно (46), где H-цепь имеет аминокислотную последовательность из положений 117-446 в аминокислотной последовательности SEQ ID NO: 10 в качестве CH;
(49) антитело, содержащее L-цепь, имеющую аминокислотную последовательность SEQ ID NO: 103 в качестве CDR1, аминокислотную последовательность SEQ ID NO: 105 в качестве CDR2 и аминокислотную последовательность SEQ ID NO: 107 в качестве CDR3;
(50) антитело, содержащее L-цепь согласно (49), где L-цепь имеет CL, полученную из мышиной κ-цепи;
(51) антитело, содержащее L-цепь согласно (49), где L-цепь имеет аминокислотную последовательность из положений 107-213 в аминокислотной последовательности SEQ ID NO: 20 в качестве CL;
(52) антитело, содержащее H-цепь согласно (46) и L-цепь согласно (49);
(53) антитело, содержащее H-цепь согласно (47) и L-цепь согласно (50);
(54) антитело, содержащее H-цепь согласно (48) и L-цепь согласно (51);
(55) антитело, содержащее одну или несколько аминокислотных замен, делеций, добавлений и/или инсерций в антителе согласно любому из (1)-(54), которое обладает активностью, эквивалентной активности антитела согласно любому из (1)-(54);
(56) антитело, которое связывается с тем же эпитопом белка EREG, что и антитело согласно любому из (1)-(55); и
(57) мини-антитело антитела согласно любому из (1)-(56);
[20] антителу по любому из [10]-[19], где антитело распознает участок с Ala в положении 29 по Ser в положении 69, или с Val в положении 63 по Leu в положении 108 в аминокислотной последовательности SEQ ID NO: 21;
[21] применению белка EREG, или мРНК EREG, или их обоих в качестве диагностического маркера злокачественной опухоли;
[22] способу диагностики злокачественной опухоли, включающему комбинирование белка EREG с антителом, которое связывается с белком EREG;
[23] способу диагностики злокачественной опухоли, включающему стадии
(a) получения образца от субъекта и
(b) детектирования белка EREG, содержащегося в собранном образце, с использованием антитела, которое связывается с белком EREG;
[24] способу диагностики злокачественной опухоли, включающему стадии (1) введения субъекту меченного радиоактивным изотопом антитела, которое связывается с белком EREG; и (2) детектирования накопления указанного радиоактивного изотопа;
[25] способу диагностики по [22], где радиоактивный изотоп представляет собой позитронно-активный нуклид;
[26] способу диагностики согласно [25], где позитронно-активный нуклид представляет собой любой нуклид, выбранный из группы, состоящей из 11C, 13N, 15O, 18F, 45Ti, 55Co, 64Cu, 66Ga, 68Ga, 76Br, 89Zr и 124I;
[27] способу диагностики злокачественной опухоли, включающему детектирование экспрессии гена, кодирующего белок EREG;
[28] способу диагностики по любому из [22]-[27], где злокачественная опухоль представляет собой любую злокачественную опухоль, выбранную из группы, состоящей из рака толстого кишечника, аденокарциномы легкого, рака поджелудочной железы, рака желудка и рака почки;
[29] способу диагностики по [28], где злокачественная опухоль представляет собой первичную злокачественную опухоль;
[30] способу диагностики по [28], где злокачественная опухоль представляет собой метастазирующую злокачественную опухоль;
[31] диагностическому средству для применения в способе диагностики злокачественной опухоли, которое содержит антитело, которое связывается с белком EREG;
[32] набору для применения в способе диагностики злокачественной опухоли, который содержит антитело, которое связывается с белком EREG, и биологический образец, содержащий белок EREG;
[33] способу скрининга соединения-кандидата в качестве лекарственного средства против злокачественной опухоли, который включает стадии:
(1) контактирования экспрессирующих EREG клеток с тестируемым соединением;
(2) измерения уровня экспрессии EREG в экспрессирующих EREG клетках и
(3) отбора соединения, которое снижает уровень экспрессии EREG по сравнению с контролем, в качестве лекарственного средства-кандидата против злокачественной опухоли;
[34] способу по [33], где уровень экспрессии EREG оценивают по уровню белка EREG, секретируемого в культуральный супернатант;
[35] способу скрининга соединения-кандидата в качестве лекарственного средства против злокачественной опухоли, который включает стадии:
(1) культивирования клеток, которые пролиферируют зависимым от EREG образом, в присутствии EREG и тестируемого соединения;
(2) измерения уровня клеточной пролиферации и
(3) отбора тестируемого соединения, которое подавляет клеточную пролиферацию по сравнению с контролем, в качестве лекарственного средства-кандидата против злокачественной опухоли.
[Эффекты изобретения]
С помощью анализа экспрессии генов в различных злокачественных тканях и злокачественных клеточных линиях было подтверждено, что экспрессия гена EREG значительно повышена в злокачественных клетках. С другой стороны, экспрессия EREG является очень низкой в нормальных клетках. Таким образом, EREG пригоден в качестве специфичного маркера для детектирования злокачественной опухоли.
Цитотоксический эффект антител против EREG в отношении экспрессирующих EREG клеток был подтвержден с помощью настоящего изобретения. Несмотря на то, что экспрессия EREG является очень низкой в нормальных клетках, она повышена в злокачественных клетках. Это подтверждает возможность того, что можно достигнуть специфичного цитотоксического эффекта на злокачественные клетки in vivo, например, посредством введения антитела против EREG.
Более того, в настоящем изобретении было подтверждено, что зависимая от EREG клеточная пролиферация нейтрализуется посредством антител против EREG. Таким образом, в предпочтительном варианте осуществления, в дополнение к цитотоксическому эффекту, антитела против EREG ингибируют пролиферацию злокачественных клеток посредством нейтрализации эффекта EREG на клеточную пролиферацию.
Краткое описание чертежей
На фиг.1 показаны уровни мРНК-транскрипта EREG в нормальных тканях и злокачественных тканях. На вертикальной оси указана интенсивность сигнала зонда c ID: 205767_at HG-U133A. Интенсивность представлена относительно среднего значения показателей экспрессии для всех генов на GeneChip U133A, которому было присвоено значение 100.
На фиг.2 показаны уровни мРНК-транскрипта EREG в злокачественных клеточных линиях. На вертикальной оси указана интенсивность сигнала зонда с ID: 205767_at HG-U133A. Интенсивность представлена относительно среднего значения показателей экспрессии для всех генов на GeneChip U133A, которому было присвоено значение 100.
На фиг.3 представлено связывание моноклональных антител против EREG с клетками EREG-DG посредством проточной цитометрии. На вертикальной оси указано количество клеток (флуоресцентные импульсы), и на горизонтальной оси указана величина интенсивности флуоресценции.
На фиг.4 показано связывание моноклональных антител против EREG с клеточной линией рака толстого кишечника DLD1 посредством проточной цитометрии. На вертикальной оси указано количество клеток (флуоресцентные импульсы), и на горизонтальной оси указана величина интенсивности флуоресценции.
На фиг.5 показана активность CDC моноклональных антител против EREG в отношении клеточной линии рака толстого кишечника DLD1. На вертикальной оси указан уровень высвобождения хрома (%), и на горизонтальной оси указано название антитела.
На фиг.6 представлена активность ADCC моноклональных антител против EREG в отношении клеточной линии рака толстого кишечника DLD1. На вертикальной оси указан уровень высвобождения хрома (%), и на горизонтальной оси указано название антитела.
На фиг.7 показана зависимость пролиферации клеток EGFR-BAF от EREG человека. На вертикальной оси указано поглощение при 450 нм (длина волны сравнения 655 нм), и на горизонтальной оси указана концентрация EREG (нг/мл).
На фиг.8 показана зависимость пролиферации клеточной линии рака поджелудочной железы AsPC1 от EREG человека. На вертикальной оси указано поглощение при 450 нм (длина волны сравнения 655 нм), и на горизонтальной оси указана концентрация EREG (нг/мл).
На фиг.9 показана зависимость пролиферации клеточной линии рака толстого кишечника DLD1 от EREG человека. На вертикальной оси указано поглощение при 450 нм (длина волны сравнения 655 нм), и на горизонтальной оси указана концентрация EREG (нг/мл).
На фиг.10 показана нейтрализующая активность моноклональных антител против EREG человека в клетках EGFR-BAF. На вертикальной оси указана скорость клеточной пролиферации (поглощение при каждой концентрации антитела/поглощение контроля×100%), и на горизонтальной оси указана концентрация антитела (мкг/мл).
На фиг.11 показана нейтрализующая активность моноклональных антител против EREG человека в клеточной линии рака поджелудочной железы AsPC1. На вертикальной оси указана скорость клеточной пролиферации (поглощение при каждой концентрации антитела/поглощение контроля×100%), и на горизонтальной оси указана концентрация антитела (мкг/мл).
На фиг.12 показано связывание химерных антител к EREG, которые были реконструированы посредством идентификации последовательности вариабельной области.
На фиг.13 показана активность химерных антител в отношении индукции ADCC против злокачественных клеточных линий.
На фиг.14 показано связывание химерных антител с ортологами EREG.
На фиг.15 показан результат сравнения активации рецептора EGFR рекомбинантными растворимыми EREG и EGF.
На фиг.16 показана активация рецептора EGF посредством кокультивирования с клетками, искусственно экспрессирующими EREG. Такая активация подавляется антителами против EREG.
На фиг.17 показана активация рецептора EGF посредством кокультивирования с клетками, искусственно экспрессирующими EREG, и подавление такой активации антителами против EREG.
На фиг.18 показана активация рецептора EGFR посредством кокультивирования с клетками DLD-1 и подавление такой активации посредством добавления антител против EREG.
На фиг.19 показано детектирование растворимого белка EREG посредством сэндвич-ELISA с использованием антител против EREG.
На фиг.20 показан результат иммунного окрашивания моноклональным антителом мыши против EREG (EP27).
На фиг.21-1 представлены результаты сэндвич-ELISA для антигенных белков с использованием антител мыши против EREG и химерных антител человека против EREG.
На фиг.21-2 представлено продолжение фиг.22-1.
На фиг.21-3 представлено продолжение фиг.22-2.
На фиг.22 показан результат, который был получен из данных сэндвич-ELISA, посредством кластерного анализа фингерпринтов связывания антител.
Способ осуществления изобретения
EREG представляет собой мембраносвязанный белок эпидермального фактора роста. Его аминокислотная последовательность и кодирующая последовательность его гена описаны в GenBank под регистрационными номерами NP_001423 (SEQ ID NO: 22) и NM_001432 (SEQ ID NO: 21) соответственно. В настоящем изобретении белок EREG относится как к полноразмерному белку, так и к его фрагментам. Термин "фрагменты" относится к полипептидам, содержащим любой участок белка EREG, и он может не обладать функцией природного белка EREG. Не ограничиваясь этим, примером фрагментов является фрагмент, содержащий внеклеточный участок белка EREG. Положения 29-122 в аминокислотной последовательности SEQ ID NO: 22 соответствуют внеклеточному участку белка EREG. Положения 123-140 в аминокислотной последовательности SEQ ID NO: 22 соответствуют трансмембранному участку.
В настоящем изобретении посредством анализов клинических образцов и злокачественных клеточных линий было показано, что ген EREG в высокой степени и часто экспрессируется в тканях первичного рака толстого кишечника, метастазирующего рака толстого кишечника, аденокарциномы легкого, рака поджелудочной железы, рака желудка и рака почки. Более того, также было показано, что белок EREG в высокой степени экспрессируется в злокачественных клеточных линиях. Таким образом, белок EREG пригоден в качестве диагностического маркера для злокачественной опухоли.
Детектирование экспрессии гена EREG
В настоящем изобретении способы диагностики злокачественной опухоли включают в себя стадию детектирования экспрессии гена EREG. В одном варианте осуществления способов согласно изобретению выявляют экспрессию белка EREG.
В настоящем изобретении детектирование включает количественное и качественное детектирование. Примеры качественного детектирования включают следующее определение:
определение только присутствия или отсутствия белка EREG;
определение с целью выявить, присутствует или нет белок EREG в количестве, превышающем определенное количество; и
определение для сравнения количества белка EREG с количеством белка в других образцах (например, в контрольном образце).
С другой стороны, примеры количественного детектирования включают измерение концентрации белка EREG и измерение количества белка EREG.
Тестируемые образцы согласно изобретению, конкретно, не ограничиваются, при условии, что они представляют собой образцы, которые могут содержать белок EREG. Конкретно, предпочтительными являются образцы, полученные из организмов, таких как млекопитающие. Более предпочтительными являются образцы, полученные от человека. Конкретные примеры тестируемых образцов включают кровь, интерстициальную жидкость, плазму, внесосудистую жидкость, цереброспинальную жидкость, синовиальную жидкость, плевральную жидкость, сыворотку, лимфу, слюну и мочу. Предпочтительными образцами являются образцы крови. Образцы крови включают сыворотку, плазму и цельную кровь. Среди указанных образцов крови, сыворотка является предпочтительной. Тестируемые образцы, полученные из образцов, таких как иммобилизованные образцы ткани или клетки, полученные из организма, и раствор с культурой клеток также относятся к тестируемым образцам согласно изобретению.
Злокачественные опухоли, которые можно диагностировать посредством настоящего изобретения, особенно не ограничены и могут представлять собой любую злокачественную опухоль. Конкретные примеры включают рак толстого кишечника, метастазирующий рак толстого кишечника, аденокарциному легкого, рак поджелудочной железы, рак желудка и рак почки. В настоящем изобретении можно диагностировать как первичные, так и метастатические очаги указанных злокачественных опухолей. В частности, предпочтительными для настоящего изобретения являются первичный рак толстого кишечника, метастазирующий рак толстого кишечника и рак поджелудочной железы.
В настоящем изобретении злокачественную опухоль выявляют при детектировании в тестируемом образце белка EREG. Более конкретно, если количество белка EREG, детектированного в тестируемом образце, превышает количество белка EREG в отрицательном контроле или у здорового индивидуума, определяют, что субъект обладает злокачественной опухолью или имеет риск развития злокачественной опухоли в будущем. Таким образом, настоящее изобретение относится к способам детектирования злокачественной опухоли, которые включают в себя стадии
(1) детектирования экспрессии EREG в биологическом образце, полученном от субъекта; и
(2) определения того, что субъект имеет злокачественную опухоль, если экспрессия EREG, выявленная на стадии (1), превышает экспрессию контроля.
В настоящем изобретении контроль включает отрицательный контроль и биологический образец от здорового индивидуума. Отрицательный контроль можно получить забором биологических образцов от здоровых индивидуумов и смешиванием биологических образцов при необходимости. Детектирование экспрессии EREG в контроле можно проводить совместно с детекцией экспрессии EREG в биологическом образце субъекта. Альтернативно, детектирование экспрессии EREG в биологических образцах многих здоровых индивидуумов можно проводить заблаговременно и можно статистически определять стандартный уровень экспрессии у здоровых индивидуумов. Более конкретно, например, в качестве стандартной величины можно использовать средние значение±2× стандартное отклонение (S.D.) или среднее значение±3× стандартное отклонение (S.D.). Статистически значение для 80% здоровых индивидуумов находится в пределах ±2× стандартного отклонения (S.D.) от среднего значения, и значение для 90% здоровых индивидуумов находится в пределах ±3× стандартного отклонения (S.D.) от среднего значения.
Альтернативно, уровень экспрессии EREG в контроле можно определять с использованием ROC-кривой. Кривая зависимости чувствительности от вероятности ложноположительных результатов (ROC) представляет собой график с чувствительностью детектирования по вертикальной оси и частотой ложноположительных результатов (т.е. "1-специфичность") по горизонтальной оси. В настоящем изобретении ROC-кривую можно получить нанесением на график изменений чувствительности к частоте ложноположительных результатов в результате постоянно меняющихся стандартных значений при определении уровня экспрессии EREG в биологическом образце.
"Стандартное значение", используемое для получения кривой ROC, представляет собой числовое значение, временно используемое для статистического анализа. Как правило, "стандартные значения" для получения ROC-кривой постоянно варьируют в диапазоне, который охватывает все выбираемые стандартные значения. Например, стандартные значения могут варьировать между минимальным и максимальным значениями EREG, измеренными в анализируемой популяции.
Исходя из полученной ROC-кривой, можно выбрать стандартные значения, которые, как ожидают, приведут к требуемой чувствительности и точности детектирования. Стандартные значения, которые статистически определяют с помощью ROC-кривой или подобной ей кривой, также называют "значениями нижнего предела". На стадии (2) описанных выше способов детектирования на основе значений нижнего предела, экспрессию EREG, выявленную на стадии (1) сравнивают со значением нижнего предела. Детектирование более высокой экспрессии EREG на стадии (1), чем значение нижнего предела, означает детектирование злокачественной опухоли у субъекта.
В настоящем изобретении уровень экспрессии EREG можно определить любым способом. Более конкретно, уровень экспрессии EREG можно определить посредством оценки количества мРНК EREG, количества белка EREG и биологической активности белка EREG. Количество мРНК EREG и белка EREG можно определить способами, описанными в настоящем описании. Альтернативно, в качестве биологической активности EREG можно определять активность индукции клеточной пролиферации, с использованием клеток, пролиферация которых индуцируется зависимым от EREG образом.
Субъектами в настоящем изобретении могут быть любые виды животных, у которых экспрессируются белок EREG. Например, известно, что белок EREG экспрессируется у многих не относящихся к человеку млекопитающих, таких как обезьяны, крупный рогатый скот, овцы, мыши, собаки, кошки и хомяки. Таким образом, приведенные животные относятся к субъектам согласно изобретению. Субъектами, которые являются особенно предпочтительными, являются люди. Безусловно, когда в качестве животного используют не относящееся к человеку животное, выявляют белок EREG этого вида животного.
Предпочтительные варианты осуществления диагностических способов согласно изобретению включают способы диагностики злокачественной опухоли, которые включают в себя стадию детектирования белка EREG на срезе иммобилизованной ткани или в клетках, полученных от пациента, страдающего указанной выше злокачественной опухолью. Более того, другие варианты осуществления настоящего изобретения включают в себя диагностические способы, включающие в себя стадию детектирования высвобожденного клетками белка EREG в крови. В частности, настоящее изобретение предпочтительно представляет собой способ диагностики злокачественной опухоли, который включает в себя стадию детектирования фрагмента, содержащего внеклеточный домен белка EREG, присутствующего в крови.
Способы детектирования белка EREG, содержащегося в тестируемого образце, особенно не ограничены. Предпочтительным является иммунологический способ, где для детектирования используют антитело против EREG. Иммунологические способы, которые можно использовать, включают:
радиоиммунный анализ (RIA);
ферментный иммунологический анализ (EIA);
флуоресцентный иммунологический анализ (FIA);
люминесцентный иммунологический анализ (LIA);
иммунопреципитацию (IP);
турбодиметрический иммунологический анализ (TIA);
Вестерн-блоттинг (WB);
иммуногистохимическое окрашивание (IHC) и
простую радиальную иммунодиффузию (SRID).
Среди указанных выше способов предпочтительным иммунологическим анализом является ферментный иммунологический анализ. Более конкретно, предпочтительным ферментным иммунологическим анализом является твердофазный иммуноферментный анализ (ELISA). Вариантом осуществления ELISA, например, является сэндвич-ELISA. Указанные выше иммунологические способы, такие как ELISA, являются способами, известными специалистам в данной области.
Следующий способ представляет собой, например, обычный способ детектирования, где используют антитело против EREG. После иммобилизации антитела против EREG на подложке подложку блокируют для избежания неспецифического связывания белков с подложкой. Например, для блокирования используют бычий сывороточный альбумин (BSA), желатин или альбумин. Известны различные способы связывания антител с подложкой. Например, синтетические смолы, такие как полистирольные смолы, физически адсорбируют антитела. Альтернативно, антитела могут быть химически связаны с подложкой, в которую внесены функциональные группы. Для химического связывания антител можно использовать бифункциональные линкеры.
Затем к подложке добавляют тестируемый образец и проводят инкубацию. В ходе инкубации антитела против EREG, связанные с подложкой, связываются с белком EREG в тестируемом образце. После этого проводят детектирование EREG, связанного с подложкой через антитело против EREG. Перед детекцией EREG, связанного с подложкой, подложка может быть промыта. Детектирование EREG, связанного с подложкой, можно проводить, например, с использованием второго антитела, которое распознает EREG. Второе антитело может быть мечено материалом для мечения. Альтернативно, второе антитело можно метить косвенно с использованием третьего антитела (вторичное антитело), которое распознает второе антитело. Как описано выше, детектирование белка EREG в тестируемом образце можно проводить посредством качественного или количественного детектирования белка EREG, связанного с антителом против EREG на подложке. Далее описано несколько конкретных примеров.
В настоящем изобретении в качестве подложки для иммобилизации антитела против EREG можно использовать следующие материалы:
нерастворимые полисахариды: агарозу, целлюлозу и подобные;
синтетические смолы: кремнийорганическую смолу, полистирольную смолу, полиакриламидную смолу, нейлоновую смолу, поликарбонатную смолу и подобные; и
нерастворимую подложку, такую как стекло.
Такую подложку используют в форме гранул или планшетов. В случае гранул можно использовать колонку, заполненную гранулами, или подобные. В случае планшета можно использовать многолуночный планшет (96-луночный планшет или подобные) или биосенсорный чип. В случае связывания антитела против EREG с подложкой антитело против EREG можно связывать с подложкой широко используемыми способами, такими как химическое присоединение или физическая адсорбция. Пригодным является применение коммерчески доступных подложек.
Связывание антитела против EREG с белком EREG, как правило, проводят в буфере. Например, в качестве буфера используют фосфатный буфер, Tris-буфер, буфер на основе лимонной кислоты, боратный буфер, карбонатный буфер или подобные. Более того, пригодным является проведение инкубации с использованием широко используемых условий, таких как инкубация при температуре от 4°C до комнатной температуры в течение одного-24 часов. Если связывание между белком EREG и антителом против EREG не прерывают, после инкубации можно использовать любое средство для промывания, и, например, пригодным является применение буфера, содержащего поверхностно-активное вещество, такое как Tween 20 или подобные.
В способе детектирования белка EREG согласно изобретению, в дополнение к тестируемому образцу, в котором будут проводить детектирование содержания белка EREG, пригодным является получение контрольного образца. Контрольный образец включает, например, отрицательный контрольный образец, не содержащий белка EREG, и положительный контрольный образец, содержащий белок EREG. В этом случае, сравнивая результаты, полученные от отрицательного контрольного образца, не содержащего белка EREG, с результатами, полученными от положительного контрольного образца, содержащего белок EREG, можно подтвердить присутствие или отсутствие белка EREG в тестируемом образце. Более того, после получения серии контрольных образцов с постепенно изменяющейся концентрацией и получения результатов детектирования для каждого контрольного образца в качестве числовой величины можно строить стандартную кривую, исходя из значений концентрации белка EREG и их соответствующих измеренных значений. Белок EREG, содержащийся в тестируемом образце, можно количественно определить, исходя из стандартной кривой и результата количественного определения белка EREG, содержащегося в тестируемом образце.
В предпочтительном варианте осуществления детектирования белка EREG, связанного с подложкой посредством антитела против EREG, применяют способ, где используется антитело против EREG, меченное материалом для мечения. Например, детектирование белка EREG можно проводить посредством контактирования тестируемого образца с антителом против EREG, иммобилизованным на подложке, промывания его при необходимости и последующего добавления меченого антитела, которое специфично распознает белок EREG.
Антитела против EREG можно метить широко известными способами. В качестве материала для мечения можно использовать материал для мечения известный специалистам в данной области, такой как флуоресцентные красители, ферменты, коферменты, хемилюминесцентные вещества и радиоактивные вещества. Конкретно, для мечения антитела можно использовать следующие материалы для мечения:
радиоактивные изотопы: 32P, 14C, 125I, 3H, 131I и т.п.;
флуоресцентные красители: флуоресцеин, родамин, дансилхлорид, умбеллиферон и т.п.;
ферменты: люциферазу, пероксидазу, щелочную фосфатазу, β-галактозидазу, β-глюкозидазу, пероксидазу хрена, глюкоамилазу, лизоцим, сахаридоксидазу, микропероксидазу и т.п.; и
вещества с аффинным связыванием: биотин и т.п.
При применении биотина в качестве материала для мечения детектирование меченных биотином антител можно проводить с использованием авидина, связанного с ферментом, таким как щелочная фосфатаза. Для связывания материала для мечения с антителом против EREG можно использовать известные способы, такие как способ с глутаральдегидом, способ с малеинимидом, способ с пиридилдисульфидом или способ с перйодистой кислотой.
Способы детектирования материала для мечения также известны. Например, при детектировании антитела против EREG, меченного радиоактивным веществом, детектирование радиоактивности можно проводить с помощью жидкостного сцинтилляционного счетчика. Антитело против EREG, меченное ферментом, можно выявлять добавлением субстрата к меченому антителу против EREG и последующей детекцией ферментативного изменения субстрата. Комбинирование фермента, который катализирует цветную реакцию или люминесцентную реакцию с субстратом, известно. Конкретные примеры субстрата для детектирования посредством пероксидазы включают диаммониевую соль 2,2-азинобис(3-этилбензотиазолин-6-сульфоновой кислоты) (ABTS), 1,2-фенилендиамин (орто-фенилендиамин) и 3,3',5,5'-тетраметилбензидин (TMB). Когда используют такие окрашивающие вещества, реакцию можно отслеживать с помощью абсорбционного фотометра. Когда вещество представляет собой испускающее флуоресцентное излучение вещество, детектирование ферментативного изменения субстрата можно проводить с помощью спектрофлуориметра.
В настоящем изобретении особенно предпочтительным вариантом осуществления способов детектирования белка EREG является, например, способ, где используют меченное биотином антитело против EREG и авидин. Конкретно, для детектирования меченного биотином антитела против EREG можно использовать авидин или стрептавидин, связанный ферментом. Авидин и стрептавидин, связанный с ферментом, таким как щелочная фосфатаза или пероксидаза, известны.
В настоящем изобретении другой вариант осуществления способов детектирования белка EREG включает способ, где используют один или несколько типов первичных антител, которые специфично распознают белок EREG, и один или несколько типов вторичных антител, которые специфично распознают первичные антитела.
При применении вторичного антитела в качестве антитела жидкой фазы используют антитело против EREG, полученное из видов, отличающихся от вида, являющегося источником антитела, связанного с подложкой (антитело твердой фазы). Антитело жидкой фазы связывается с белком EREG, захваченным антителом твердой фазы, и с антителом жидкой фазы связывается вторичное антитело. Вторичное антитело может связываться с антителом жидкой фазы, но не может связываться с антителом твердой фазы. Таким образом, количество вторичного антитела, остающегося на подложке, зависит от количества антитела жидкой фазы, связанного с подложкой через белок EREG. В результате указанного выше действия детектирование белка EREG в тестируемом образце можно проводить посредством качественного или количественного детектирования связанных вторичных антител. В этом случае вторичное антитело может быть пригодным образом мечено материалом для мечения.
В настоящем изобретении другой вариант осуществления способов детектирования белка EREG представляет собой, например, способ детектирования, где используется реакция агрегации. В данном способе детектирование белка EREG можно проводить с использованием носителя, на котором адсорбировано антитело против EREG. Для адсорбции антитела можно использовать любой носитель при условии, что он является нерастворимым и стабильным, и не вызывает неспецифичных реакций. Например, можно использовать латексные частицы, бентонит, коллодий, каолин или иммобилизованные эритроциты овцы. Латексные частицы обладают превосходной однородностью и стабильностью и, таким образом, являются предпочтительными носителями. Латексные частицы, которые можно использовать, представляют собой, например, латексные частицы из полистирола, латексные частицы из сополимера стирола и бутадиена или латексные частицы из поливинилтолуола. Альтернативно, также известны латексные частицы, на которые внесены функциональные группы, такие как карбоксильные группы. Например, предпочтительными латексными частицами являются латексные частицы из полистирола.
При смешивании с антителом латексные частицы, которые обладают гидрофобной поверхностью, физически адсорбируют антитело. Альтернативно, если латексные частицы имеют функциональные группы, антитело может связываться химически. Частицы со связанным антителом смешивают с образцом и перемешивают в течение определенного периода времени. Поскольку степень агрегации частиц повышается при повышении концентрации белка EREG в образце, детектирование белка EREG можно проводить посредством оценки степени агрегации невооруженным глазом. Более того, также детектирование белка EREG можно проводить посредством измерения повышения помутнения или рассеяния света, вызываемых агрегацией с использованием спектрофотометра или подобного.
В настоящем изобретении другой вариант осуществления способов детектирования белка EREG включает, например, способ, где используют биосенсор с использованием явления поверхностного плазмонного резонанса. Применение биосенсора с использованием явления поверхностного плазмонного резонанса обеспечивает наблюдение в режиме реального времени белок-белковых взаимодействий в качестве сигналов поверхностного плазмонного резонанса без необходимости мечения белка. Например, с использованием биосенсора, такого как BIAcore (Biacore), можно проводить детектирование связывания между белком EREG и антителом против EREG. Конкретно, тестируемый образец подвергают контактированию с сенсорным чипом, на котором иммобилизовано антитело против EREG, и детектирование белка EREG, который связывается с антителом против EREG, можно проводить в качестве изменения резонансных сигналов.
EREG представляет собой мембранный белок, экспрессия которого специфично повышается в злокачественных клетках. Таким образом, злокачественные клетки или злокачественные ткани можно выявлять с помощью антител против EREG. Например, злокачественные клетки, содержащиеся в клетках или тканях, полученных из живого организма, выявляют с помощью иммуногистохимического анализа с использованием антитела против EREG. Альтернативно, злокачественные ткани в живом организме можно выявлять с помощью антител против EREG. Более конкретно, настоящее изобретение относится к способам выявления злокачественной опухоли, которые включают в себя стадии: (1) введения субъекту меченного радиоактивным изотопом антитела, которое связывается с белком EREG; и (2) детектирования накопления радиоактивного изотопа. В целях отслеживания антитела, введенного в живой организм, антитело можно метить для детектирования. Например, можно отслеживать in vivo поведение антител, меченных флуоресцентным веществом, люминесцентным веществом или радиоактивным изотопом. Антитела, меченные флуоресцентным веществом или люминесцентным веществом, можно наблюдать с использованием эндоскопа или лапароскопа. При использовании радиоактивного изотопа локализацию антитела можно визуализировать, отслеживая радиоактивность радиоактивного изотопа. В настоящем изобретении локализация антитела против EREG in vivo демонстрирует присутствие злокачественных клеток.
Для детектирования злокачественной опухоли в живом организме в качестве радиоактивного изотопа для мечения антитела можно использовать позитронно-активный нуклид. Например, антитела можно метить позитронно-активными нуклидами, такими как 18F, 55Co, 64Cu, 66Ga, 68Ga, 76Br, 89Zr и 124I. Антитела против EREG можно метить указанными позитронно-активными нуклидами известными способами (Acta Oncol. 32, 825-830, 1993).
После введения антитела против EREG, меченного позитронно-активным нуклидом, человеку или животным измеряют излучение, испускаемое радиоактивным нуклеидом извне организма с использованием PET (сканнер позитронной эмиссионной томографии), и затем превращают в изображение способами компьютерной томографии. PET представляет собой инструмент для неинвазивного получения данных о поведении in vivo лекарственных средств и т.п. Интенсивность излучения может быть количественно преобразована в изображение в качестве интенсивности сигнала с использованием PET. С использованием PET, как описано выше, антигенные молекулы, которые в высокой степени экспрессируются при конкретной злокачественной опухоли, можно выявлять без забора образцов от пациентов. В дополнение к указанным выше нуклидам антитела против EREG можно подвергать радиоактивному мечению коротко живущими нуклидами с использованием позитронно-активных нуклидов, таких как 11C, 13N, 15O, 18F и 45Ti.
В настоящее время исследуют и разрабатывают применение медицинского циклотрона для получения коротко живущих нуклидов с использованием указанных выше нуклидов, способы получения коротко живущих радиоактивно меченных соединений и т.п. С использованием таких способов антитела против EREG можно метить различными радиоактивными изотопами. Антитела против EREG, вводимые пациентам, накапливаются в первичных очагах и метастатических очагах благодаря специфичности антител против EREG к любому участку патологической ткани. Если антитела против EREG метят позитронно-активными нуклидами, присутствие первичных очагов и метастатических очагов можно выявлять, исходя из локализации их радиоактивности посредством детектирования радиоактивности. Для применения в таких диагностических целях пригодным образом можно использовать значения активности испускания гамма-частиц или позитронов 25-4000 кэВ. Более того, можно ожидать терапевтических эффектов путем выбора пригодного нуклида и введения его в больших количествах. Для достижения эффекта против злокачественной опухоли посредством облучения можно использовать нуклиды, которые обеспечивают величины испускания гамма-частиц или позитронов 70-700 кэВ.
В другом варианте осуществления способов согласно изобретению проводят детектирование экспрессии мРНК EREG. В настоящем изобретении детектирование включает количественное и качественное детектирование. Примеры качественного детектирования включают следующие способы определения:
простое определение присутствия или отсутствия мРНК EREG;
определение с целью выявить, присутствует или нет мРНК EREG в количестве, превышающем определенное количество; и
определение, в котором сравнивают количество мРНК EREG с количеством с мРНК EREG в других образцах (например, контрольном образце).
С другой стороны, количественная детектирование включает, например, измерение концентрации мРНК EREG и измерение количества мРНК EREG.
В качестве тестируемого образца согласно изобретению можно использовать любой образец, который может содержать мРНК EREG. Предпочтительными являются образцы, полученные из такого организма, как млекопитающее, и более предпочтительными являются образцы, полученные от человека. Конкретные примеры тестируемых образцов включают кровь, интерстициальную жидкость, плазму, внесосудистую жидкость, цереброспинальную жидкость, синовиальную жидкость, плевральную жидкость, сыворотку, лимфу, слюну и мочу. Предпочтительными образцами являются образцы крови. Образцы крови включают сыворотку, плазму и цельную кровь. Также тестируемые образцы согласно изобретению включают образцы, полученные из тестируемых образцов, таких как растворы с клеточными культурами и образцы иммобилизованных тканей или клеток, полученные из организма.
Злокачественная опухоль для диагностики особенно не ограничена. Конкретные примеры включают рак толстого кишечника, аденокарциному легкого, рак поджелудочной железы, рак желудка и рак почки. В настоящем изобретении можно диагностировать как первичные, так и метастатические очаги указанных злокачественных опухолей. Особенно предпочтительными являются первичный рак толстого кишечника, метастазирующий рак толстого кишечника и рак поджелудочной железы.
Субъектами для настоящего изобретения могут являться любые виды животных, у которых экспрессируется белок EREG. Например, известно, что EREG экспрессируются у многих не относящихся к человеку млекопитающих, таких как обезьяны, крупный рогатый скот, овцы, мыши, собаки, кошки и хомяки. В частности, пригодными субъектами являются люди. Когда в качестве субъекта используют не относящийся к человеку вид животных, проводят детектирование мРНК EREG этого вида животных.
Конкретные варианты осуществления способа детектирования описаны ниже. Сначала получают образец от субъекта. Затем проводят детектирование мРНК EREG, находящейся в образце. В настоящем изобретении также можно проводить детектирование кДНК, синтезированных с мРНК. В настоящем изобретении детектирование мРНК EREG или кодирующей EREG кДНК в тестируемом образце означает детектирование злокачественной опухоли у пациента. Например, если в тестируемом образце выявляют большее количество мРНК EREG или кодирующей EREG кДНК, чем в отрицательном контроле или у здорового индивидуума, может быть определено, что субъект обладает злокачественной опухолью или имеет высокий риск возникновения злокачественной опухоли в будущем.
Способы детектирования мРНК известны. Конкретно, например, для настоящего изобретения можно использовать способ Нозерн-блоттинга, способ ОТ-ПЦР, способ матриц ДНК и т.п.
Способы детектирования согласно изобретению, описанные выше, могут быть автоматизированы с использованием различных автоматических тестирующих устройств. С помощью автоматизации можно исследовать большие количества образцов одновременно.
Следующей задачей настоящего изобретения является предоставление диагностических средств или наборов для диагностики злокачественной опухоли, которые содержат реагенты для детектирования белка EREG в тестируемом образце. Диагностические средства согласно изобретению включают в себя по меньшей мере одно антитело против EREG. Когда диагностические средства или наборы согласно изобретению основаны на способе EIA, таком как ELISA, может быть включен носитель для иммобилизации антитела. Альтернативно, может быть предоставлено связанное с носителем антитело. Если диагностические средства или наборы согласно изобретению основаны на способе агрегации, где используется носитель, такой как латекс, они могут включать носитель с адсорбированным антителом.
Наборы для диагностики злокачественной опухоли можно получить, комбинируя средства для диагностики злокачественной опухоли согласно изобретению с другим элементом, используемым для детектирования EREG. Более конкретно, настоящее изобретение относится к наборам для диагностики злокачественной опухоли, которые содержат EREG-связывающее антитело и контрольный образец, содержащий биологический образец, содержащий EREG. Наборы согласно изобретению, кроме того, могут содержать реагент для иммунологического анализа EREG. Например, в качестве реагентов для ELISA можно комбинировать хромогенные субстраты для детектирования ферментных меток или промывочные растворы для промывания твердой фазы. Кроме того, к наборам могут прилагаться инструкции, в которых описан процесс определения.
Более того, настоящее изобретение относится к диагностическим средствам или наборам для диагностики злокачественных опухолей, которые содержат реагент для детектирования мРНК EREG или кодирующей EREG кДНК в тестируемом образце. Диагностические средства согласно изобретению содержат по меньшей мере одну кодирующую EREG ДНК (SEQ ID NO: 21; ДНК, состоящая из нуклеотидной последовательности NM_001423) или один олигонуклеотид, содержащий по меньшей мере 15 нуклеотидов, комплементарных комплементарной цепи ДНК.
В настоящем описании термин "комплементарная цепь" относится к цепи, противоположной одной из цепей двухцепочечной нуклеиновой кислоты, состоящей из пар оснований A:T (U в случае РНК) и G:C. Кроме того, "комплементарный" относится не только к случаям полностью комплементарных последовательностей в пределах участка по меньшей мере 15 последовательно расположенных нуклеотидов, но также к случаем по меньшей мере 70%, предпочтительно по меньшей мере 80%, более предпочтительно 90% и еще более предпочтительно 95% гомологии или более в нуклеотидной последовательности. Гомологию можно определить с использованием алгоритма, описанного в настоящем описании.
Такие олигонуклеотиды можно использовать в качестве зондов или праймеров для детектирования или амплификации кодирующей EREG ДНК и зондов или праймеров для детектирования экспрессии таких ДНК. Более того, зонды можно использовать в форме субстрата для матрицы ДНК.
Когда такие олигонуклеотиды используют в качестве праймеров, их длины в норме составляют от 15 п.о. до 100 п.о. Предпочтительные длины праймеров составляют 17 п.о.-30 п.о. В качестве праймера можно использовать любой праймер, который может амплифицировать по меньшей мере часть кодирующей EREG ДНК или комплементарной ему цепи. Более того, когда они используются в качестве праймеров, их 3'-концевые участки могут быть сделаны таким образом, чтобы быть комплементарными, и на 5'-концы можно добавлять последовательности распознавания ферментами рестрикции или метки.
Более того, в качестве зонда можно использовать любой олигонуклеотид, который специфично гибридизуется по меньшей мере с частью кодирующей EREG ДНК или комплементарной ему цепи. Нуклеотидную последовательность зонда для детектирования мРНК выбирают из нуклеотидных последовательностей, комплементарных смысловой цепи EREG. Длина цепи зонда, как правило, составляет по меньшей мере 15 п.о. или более.
В настоящем изобретении олигонуклеотиды можно превращать в зонды мечением пригодным образом. Способы мечения олигонуклеотидов известны. Например, олигонуклеотиды можно метить фосфорилированием 5'-концов олигонуклеотидов, с использованием T4-полинуклеотидкиназы с меченным 32P АТФ в качестве субстрата. Альтернативно, примеры способов включают способы, которые встраивают меченый субстратный нуклеотид с использованием ДНК-полимеразы и случайный гексамерный олигонуклеотид или подобных в качестве праймера (способы со случайными праймерами и т.д.). В способах со случайными праймерами в качестве ДНК-полимеразы используют фрагмент Кленова или подобные. Более того, нуклеотиды можно метить радиоактивным изотопом, таким как 32P, флуоресцентный краситель, биотин, дигоксин или подобные.
Олигонуклеотиды согласно изобретению можно получать с использованием, например, коммерчески доступного устройства для синтеза олигонуклеотидов. Зонды можно получать в виде двухцепочечных фрагментов ДНК, полученных путем обработки ферментом рестрикции или сходных способов.
В диагностические средства или наборы, указанные выше, в дополнение к олигонуклеотидам и антителам, которые представляют собой активные ингредиенты, при необходимости можно включать стерилизованную воду, физиологический раствор, растительное масло, поверхностно-активные вещества, липиды, солюбилизаторы, буферы, белковые стабилизаторы (BSA, желатин или подобные), консерванты, блокирующие растворы, реакционный раствор, раствор для остановки реакции, реагенты для обработки образцов и т.п.
Получение антител против EREG
Антитела против EREG, используемые в настоящем изобретении, могут быть получены из любого источника, и они могут представлять собой антитела любого типа и любой формы, при условии, что они специфично связываются с белком EREG. Конкретно, можно использовать известные антитела, такие как антитела не относящегося к человеку животного (например, антитела мыши, антитела крысы и антитела верблюда), антитела человека, химерные антитела и гуманизированные антитела. В настоящем изобретении используемые антитела могут представлять собой моноклональные антитела или поликлональные антитела. Моноклональные антитела являются предпочтительными.
Антитела против EREG для применения в настоящем изобретении можно получать в виде поликлональных или моноклональных антител с использованием известных способов. В частности, моноклональные антитела, полученные от млекопитающего, являются предпочтительными в качестве антител против EREG для применения в настоящем изобретении. Моноклональные антитела, полученные от млекопитающего, включают антитела, продуцируемые гибридомой, и антитела, продуцируемые хозяином, трансформированным экспрессирующим вектором, содержащим ген антитела, способами генной инженерии.
Продуцирующую моноклональное антитело гибридому можно получать, в основном, с использованием следующего способа. Сначала проводят иммунизацию с использованием белка EREG в качестве сенсибилизирующего антигена общим способом иммунизации. Затем иммуноциты, которые получают из иммунизированных животных, подвергают слиянию с известными родительскими клетками посредством основного способа слияния клеток с получением гибридом. Более того, гибридомы, которые продуцируют антитело против EREG, можно отбирать скринингом клеток, которые продуцируют представляющие интерес антитела с использованием основных способов скрининга.
Конкретно, моноклональные антитела получают следующим образом. Сначала можно экспрессировать ген EREG с получением белка EREG для применения в качестве сенсибилизирующего антигена для продукции антитела. Нуклеотидная последовательность гена EREG описана в GenBank под регистрационным номером No. NM_001432 (SEQ ID NO: 21) и т.п. Конкретно, кодирующую ген EREG последовательность встраивают в известный экспрессирующий вектор и его используют для трансформации соответствующей клетки-хозяина. Затем представляющий интерес белок EREG человека можно выделить известным способом из клетки-хозяина или ее культурального супернатанта. Альтернативно, можно сходным образом использовать очищенный природный белок EREG. Более того, как используют в настоящем изобретении, слитый белок, получаемый слиянием требуемого неполного пептида белка EREG с другим полипептидом, можно использовать в качестве иммуногена. Например, для получения слитого белка для применения в качестве иммуногена можно использовать Fc-фрагменты антител, пептидные метки или подобные. Вектор, который экспрессирует слитый белок, можно получать слиянием требуемых генов, кодирующих два или несколько типов полипептидных фрагментов в рамке считывания, и встраиванием слитых генов в экспрессирующий вектор, как описано выше. Способы получения слитых белков описаны в Molecular Cloning 2nd ed. (Sambrook, J. et al., Molecular Cloning 2nd ed., 9.47-9.58, Cold Spring Harbor Lab. press, 1989).
Белок EREG, очищенный таким способом, можно использовать в качестве сенсибилизирующего антигена для иммунизации млекопитающих. Также в качестве сенсибилизирующего антигена можно использовать неполный пептид EREG. Например, в качестве сенсибилизирующего антигена можно использовать следующие пептиды:
пептиды, полученные из аминокислотной последовательности EREG человека химическим синтезом;
пептиды, полученные встраиванием части гена EREG в экспрессирующий вектор и его экспрессией; и
пептиды, полученные деградацией белка EREG протеазой.
Не существует ограничений в отношении участка или размера используемых неполных пептидов EREG. Предпочтительный участок может быть выбран из аминокислотной последовательности, составляющей внеклеточный домен EREG (положения 29-122 в аминокислотной последовательности SEQ ID NO: 22). Количество аминокислот, составляющих пептид, подлежащий применению в качестве сенсибилизирующего антигена, предпочтительно составляет по меньшей мере три или более, например пять или более или шесть или более. Более конкретно, в качестве сенсибилизирующего антигена можно использовать пептид, содержащий 8-50 остатков или предпочтительно 10-30 остатков.
Млекопитающее, подлежащее иммунизации сенсибилизирующим антигеном, особенно не ограничено. Для получения моноклональных антител способом слияния клеток животное, подлежащее иммунизации, предпочтительно выбирают с учетом совместимости с родительскими клетками, подлежащими применению для клеточного слияния. Как правило, в качестве животного, подлежащего иммунизации, предпочтительными являются грызуны. Конкретно, в качестве животного, подлежащего иммунизации, можно использовать мышей, крыс, хомяков или кроликов. Кроме того, в качестве животного, подлежащего иммунизации, можно использовать обезьян и подобных.
Описанных выше животных можно иммунизировать сенсибилизирующим антигеном известным способом. Например, в качестве основного способа иммунизацию можно проводить посредством внутрибрюшинной или подкожной инъекции млекопитающему сенсибилизирующего антигена. Конкретно, сенсибилизирующий антиген вводят млекопитающим несколько раз каждые четвертые-21 сутки. Сенсибилизирующий антиген разводят в соответствующем разведении PBS (забуференный фосфатом солевой раствор), физиологическим раствором или подобным и затем используют для иммунизации. Более того, сенсибилизирующий антиген можно вводить вместе с адъювантом. Например, сенсибилизирующий антиген можно изготавливать смешиванием с полным адъювантом Фрейнда, с последующим эмульгированием полученной смеси. Более того, для иммунизации с использованием сенсибилизирующего антигена можно использовать пригодный носитель. В частности, когда в качестве сенсибилизирующего антигена используют неполный пептид с низкой молекулярной массой, сенсибилизирующий антигенный пептид желательно связывают с белком-переносчиком, таким как альбумин или гемоцианин лимфы улитки, и затем используют для иммунизации.
Млекопитающих иммунизируют, как описано выше, и после подтверждения повышения количества требуемого антитела в сыворотке иммуноциты млекопитающего собирают и подвергают слиянию клеток. Особенно предпочтительным иммуноцитом является спленоцит.
В качестве клетки, подлежащей слиянию с указанным выше иммуноцитом, используют клетку миеломы млекопитающих. Клетки миеломы предпочтительно содержат пригодный селективный маркер для скрининга. Селективный маркер придает клеткам признаки для их выживания (или не выживания) в конкретных условиях культивирования. В качестве селективных маркеров известны дефицит гипоксантин-гуанинфосфорибозилтрансферазы (в дальнейшем в настоящем описании сокращаемый как дефицит HGPRT) и дефицит тимидинкиназы (в дальнейшем в настоящем описании сокращаемый как дефицит TK). Клетки, имеющие дефицит HGPRT или TK, обладают чувствительностью к гипоксантин-аминоптерин-тимидину (в дальнейшем в настоящем описании сокращаемую как чувствительность к HAT). Чувствительные к HAT клетки не могут осуществлять синтез ДНК в селективной среде HAT и, таким образом, погибают. Однако когда клетки слиты с нормальными клетками, они могут продолжать синтезировать ДНК с использованием каскада спасения нормальных клеток и, таким образом, могут расти в селективной среде HAT.
Селекцию клеток с дефицитом HGPRT и TK можно проводить в среде, содержащей 6-тиогуанин или 8-азагуанин (в дальнейшем в настоящем описании сокращаемый как 8AG) и 5'-бромдезоксиуридин соответственно. Нормальные клети погибают, поскольку они встраивают аналоги пиримидина в свою ДНК. Однако клетки с дефицитом указанных ферментов могут выживать в селективной среде, поскольку они не могут встраивать аналоги пиримидина. Альтернативно, селективный маркер, обозначаемый как устойчивость к G418, обеспечивает устойчивость к антибиотикам типа 2-дезоксистрептамина (аналоги гентамицина) с помощью гена устойчивости к неомицину. Известны различные типы клеток миеломы, которые пригодны для слияния клеток. Например, для получения моноклональных антител согласно изобретению можно использовать клетки миеломы, включающие следующие клетки:
P3 (P3x63Ag8.653) (J. Immunol. (1979) 123, 1548-1550);
P3x63Ag8U.1 (Current Topics in Microbiology and Immunology (1978) 81, 1-7);
NS-1 (Kohler. G. and Milstein, C. Eur. J. Immunol. (1976) 6, 511-519);
MPC-11 (Margulies. D.H. et al., Cell (1976) 8, 405-415);
SP2/0 (Shulman, M. et al., Nature (1978) 276, 269-270);
FO (de St. Groth, S.F. et al., J. Immunol. Способы (1980) 35, 1-21);
S194 (Trowbridge, I.S. J. Exp. Med. (1978) 148, 313-323) и
R210 (Galfre, G. et al., Nature (1979) 277, 131-133).
Клеточное слияние указанных выше иммуноцитов с клетками миеломы, как правило, проводят в соответствии с известным способом, например, способом Kohler and Milstein et al. (Kohler. G. and Milstein, C., Methods Enzymol. (1981) 73, 3-46).
Более конкретно, указанное выше слияние клеток можно проводить в стандартной питательной культуральной среде в присутствии, например, агента, усиливающего слияние клеток. Агент, усиливающий слияние клеток, может представлять собой, например, полиэтиленгликоль (PEG), вирус Сендай (HVJ) или подобные. Если желательно, для дополнительного усиления эффективности слияния можно добавлять вспомогательное вещество, такое как диметилсульфоксид.
Используемое соотношение иммуноцитов и клеток миеломы может быть установлено в соответствии с мнением специалиста. Например, предпочтительно количество иммуноцитов установлено как один к десяти относительно количества клеток миеломы. В качестве среды, подлежащей применению для указанного выше слияния клеток, можно использовать, например, среду RPMI1640 и среду MEM, которые пригодны для роста указанной выше миеломной клеточной линии, или другие стандартные среды, которые используют для данного типа клеточной культуры. Более того, к средам можно добавлять вспомогательный раствор сыворотки, такой как фетальная телячья сыворотка (FCS).
Слияние клеток проводят, осторожно перемешивая заданные количества указанных выше иммуноцитов и клеток миеломы в указанной выше среде, добавляя и перемешивая раствор PEG, предварительно нагретый приблизительно до 37°C, таким образом, чтобы образовывались требуемые слитые клетки (гибридомы). В способе слияния клеток, например, PEG со средней молекулярной массой приблизительно 1000-6000 можно, главным образом, добавлять в концентрации от 30-60% (масс./об.). После этого агент для слияния клеток или подобные, который является неблагоприятным для роста гибридом, может быть удален последовательным добавлением пригодной среды, такой как среды, приведенные выше, удалением супернатанта после центрифугирования и повторением указанных стадий.
Селекцию гибридом, полученных таким способом, можно проводить с использованием селективной среды, пригодной для селективных маркеров, которыми обладают миеломы, используемые для слияния клеток. Например, селекцию клеток с дефицитом HGPRT и TK можно проводить посредством культивирования их в среде HAT (среде, содержащей гипоксантин, аминоптерин и тимидин). Более конкретно, когда для слияния используют чувствительные к HAT клетки миеломы, клетки, успешно слитые с нормальными клетками, можно селективно выращивать в среде HAT. Культивирование с использованием указанной выше среды HAT продолжают в течение периода времени, достаточного для уничтожения клеток, отличных от представляющей интерес гибридомы (неслитых клеток). Более конкретно, как правило, селекцию представляющей интерес гибридомы можно проводить культивированием в течение от нескольких суток до нескольких недель. После этого гибридомы, которые продуцируют представляющее интерес антитело, можно подвергнуть скринингу и моноклональному клонированию путем проведения стандартного способа лимитирующих разведений. Альтернативно, распознающее EREG антитело можно получить с использованием способа, описанного в международной патентной публикации No. WO 03/104453.
Представляющее интерес антитело можно пригодным образом подвергнуть скринингу и отдельно клонировать с помощью способа скрининга на основе реакции известный антиген-антитело. Например, антиген связывают с носителем, таким как гранулы из полистирола или подобные, или коммерчески доступным 96-луночным микропланшетом для титрования с последующей реакцией с культуральным супернатантом гибридом. Затем, после промывания носителя, его подвергают реакции с меченным ферментом вторичным антителом или подобным. Если представляющее интерес антитело, которое реагирует с сенсибилизирующим антигеном, содержится в культуральном супернатанте, вторичное антитело будет связываться с носителем через антитело. В конечном итоге, присутствие представляющего интерес антитела в культуральном супернатанте можно определить посредством детектирования вторичных антител, связанных с носителем. Гибридомы, продуцирующие требуемые антитела, которые могут связываться с антигеном, можно клонировать способом лимитирующих разведений или подобным. В этом случае можно пригодным образом использовать антигены, используемые для иммунизации, а также функционально эквивалентный белок EREG. Например, в качестве антигена можно использовать внеклеточный домен EREG или олигопептида, содержащего неполную аминокислотную последовательность, составляющую данный участок.
В дополнение к указанному выше способу, где гибридомы получают посредством иммунизации антигеном не относящихся к человеку животных, представляющее интерес антитело можно получить посредством сенсибилизации антигеном лимфоцитов человека. Более конкретно, сначала лимфоциты человека сенсибилизируют белком EREG in vitro. Затем иммунносенсибилизированные лимфоциты подвергают слиянию с пригодным партнером по слиянию. Например, в качестве партнера по слиянию можно использовать клетки миеломы человека, которые обладают бесконечным потенциалом к делению (см. публикацию патента Японии Kokoku No. (JP-B) H01-59878 (рассмотренная, одобренная патентная заявка Японии, опубликованная после экспертизы)). Антитела против EREG, полученные таким способом, представляют собой антитела человека, которые обладают активностью связывания с белком EREG.
Альтернативно, антитела человека против EREG также можно получить посредством введения белка EREG, который служит в качестве антигена, трансгенному животному, имеющему полный набор генов антител человека. Антитело продуцирующие клетки иммунизированного животного можно иммортализовать клеточным слиянием с пригодным партнером по слиянию или посредством обработки, такой как инфицирование вирусом Эпштейна-Барр. Антитела человека против белка EREG можно выделить из иммортализованных клеток, полученных таким способом (см. международные патентные публикации No. WO 94/25585, WO 93/12227, WO 92/03918 и WO 94/02602). Более того, клетки, которые продуцируют антитело, обладающее представляющей интерес специфичностью реакции, можно клонировать посредством клонирования иммортализованных клеток. Когда в качестве животного, подлежащего иммунизации, используют трансгенное животное, иммунная система данного животного распознает EREG человека как чужеродный материал. Таким образом, можно легко получить антитела человека против EREG человека.
Продуцирующие моноклональное антитело гибридомы, полученные таким способом, можно пассировать и культивировать в стандартной среде. Альтернативно, гибридомы можно хранить в течение длительного периода времени в жидком азоте.
Гибридомы можно культивировать стандартным способом, и из культуральных супернатантов можно получать представляющее интерес моноклональное антитело. Альтернативно, гибридомы можно выращивать посредством введения их совместимому млекопитающему, и моноклональные антитела можно получать из их асцитных жидкостей. Первый из таких способов пригоден для получения в высокой степени очищенных антител.
В настоящем изобретении можно использовать антитело, кодируемое геном антитела, клонированным из антитело продуцирующих клеток. Клонированный ген антитела можно встраивать в пригодный вектор и затем инкубировать в хозяине для экспрессии антитела. Способы выделения гена антитела, встраивания гена в вектор и трансформации клеток были разработаны (см., например, Vandamme, A.M. et al., Eur. J. Biochem. (1990) 192, 767-775).
Например, кДНК, кодирующую вариабельную область (V-область) антитела против EREG можно получить из гибридомных клеток, продуцирующих антитело против EREG. Как правило, для этого сначала из гибридомы экстрагируют тотальную РНК. Например, в качестве способов экстракции тотальной мРНК из клеток можно использовать следующие способы:
способ ультрацентрифугирования с гуанидином (Chirgwin, J.M. et al., Biochemistry (1979) 18, 5294-5299) и
способ AGPC (Chomczynski, P. et al., Anal. Biochem. (1987) 162, 156-159).
Экстрагированную мРНК можно очистить с использованием набора для очистки мРНК (GE Healthcare Bio-Sciences) или подобных. Альтернативно, коммерчески доступны также наборы для прямой экстракции тотальной мРНК из клеток, такие как набор для очистки мРНК QuickPrep (GE Healthcare Bio-Sciences). Тотальную РНК можно получить из гибридомы с использованием таких наборов. С полученной мРНК можно синтезировать кДНК, кодирующую V-область антитела, с использованием обратной транскриптазы. кДНК можно синтезировать с использованием набора для синтеза первой цепи кДНК с помощью обратной транскриптазы AMV (SEIKAGAKU CORPORATION) или подобного. Для синтеза и амплификации кДНК можно использовать набор 5'-Ampli FINDER RACE (Clontech) и способ с использованием ПЦР с 5'-RACE (Frohman, M.A. et al., Proc. Natl. Acad. Sci. USA (1988) 85, 8998-9002; Belyavsky, A. et al., Nucleic Acids Res. (1989) 17, 2919-2932). Более того, в процессе такого синтеза кДНК на оба конца кДНК могут быть встроены соответствующие участки для ферментов рестрикции, которые описаны ниже.
Представляющий интерес фрагмент кДНК очищают из полученного продукта ПЦР и затем лигируют в ДНК вектор. Таким образом, получают рекомбинантный вектор и вводят в Escherichia coli или подобные, и после селекции колоний из E. coli, которые образовали колонии, можно получить требуемый рекомбинантный вектор. Имеет или нет рекомбинантный вектор представляющую интерес нуклеотидную последовательность кДНК, можно подтвердить известным способом, таким как способ терминации цепи дидезоксинуклеотидами.
Для получения генов, кодирующих вариабельные области, можно использовать способ ПЦР, где используются праймеры для амплификации генов вариабельных областей. Сначала получают библиотеку кДНК посредством синтеза кДНК с использованием экстрагированных мРНК в качестве матриц. Для синтеза библиотеки кДНК удобно использовать коммерчески доступный набор. На практике, поскольку количество мРНК, получаемое из только небольшого количества клеток, является крайне малым, выход мРНК после такой прямой очистки является низким. Таким образом, очистку, как правило, проводят после добавления РНК-носителя, которая не содержит ген антитела. Альтернативно, когда можно экстрагировать определенное количество РНК, эффективную экстракцию можно проводить с использованием только РНК антитело продуцирующих клеток. Например, добавление РНК-носителя может не требоваться, когда РНК экстрагируют из десяти или более, 30 или более, или предпочтительно 50 или более антитело продуцирующих клеток.
Гены антител амплифицируют способом ПЦР с использованием полученной библиотеки кДНК в качестве матрицы. Праймеры, используемые для амплификации генов антител способом ПЦР, известны. Например, праймеры для амплификации гена антитела человека можно конструировать, исходя из описания в статье (J. Mol. Biol. (1991) 222, 581-597) и т.п. Нуклеотидные последовательности таких праймеров варьируют в зависимости от подкласса иммуноглобулина. Таким образом, когда в качестве матрицы используют библиотеку кДНК неизвестного подкласса, при проведении способа ПЦР необходимо рассматривать различные возможности.
Более конкретно, когда задачей является получение генов, кодирующих IgG человека, можно использовать праймеры, способные амплифицировать гены, кодирующие γ1-γ5 для тяжелой цепи, и гены, кодирующие κ-цепь и λ-цепь легкой цепи. Для амплификации генов вариабельной области IgG, как правило, в качестве 3'-концевого праймера используют праймер, который гибридизуется с частью соответствующей шарнирной области. Между тем, в качестве 5'-концевого праймера используют праймер, соответствующий каждому подклассу.
Из продуктов ПЦР, полученных посредством праймеров для амплификации генов для подклассов тяжелой цепи и легкой цепи, создают независимые библиотеки. С использованием библиотек, синтезированных таким способом, можно восстанавливать иммуноглобулины, содержащие комбинацию легких и тяжелых цепей. Представляющие интерес антитела можно подвергнуть скринингу с использованием в качестве показателя активности восстановленных иммуноглобулинов в отношении связывания EREG.
Например, когда задачей является получение антител против EREG, более предпочтительным является, чтобы связывание антител с EREG являлось специфическим. Например, можно проводить скрининг антител, которые связываются с EREG, как описано ниже:
(1) контактированием EREG с антителом, содержащим V-область, кодируемую кДНК, полученной из гибридомы;
(2) детекцией связывания между EREG и антителом и
(3) селекцией антитела, которое связывается с EREG.
Способы детектирования связывания между антителом и EREG известны. Конкретно, тестируемое антитело подвергают реакции с EREG, иммобилизованным на подложке, и затем его подвергают реакции с меченым антителом, которое распознает тестируемое антитело. Если меченое антитело на подложке выявляется после промывания, это показывает, что тестируемое антитело связывается с EREG. Для мечения можно использовать ферментативно активные белки, такие как пероксидаза или β-галактозидаза, или флуоресцентные вещества, такие как FITC. Для оценки активности связывания антитела можно использовать образцы иммобилизованных экспрессирующих EREG клеток.
Альтернативно, для способа скрининга антитела, где в качестве показателя используется активность связывания, можно использовать способ пэннинга на основе фагового вектора. Когда гены антитела получают в виде библиотек субклассов тяжелых цепей и легких цепей, как описано выше, преимущественными являются способы скрининга на основе фагового вектора. Гены, кодирующие вариабельные области тяжелой и легкой цепей, могут быть преобразованы в ген одноцепочечного Fv (scFv) посредством связывания генов через пригодные линкерные последовательности. Фаги, экспрессирующие на своей поверхности scFv, можно получить встраиванием гена, кодирующего scFv, в фаговый вектор. ДНК, кодирующую scFv, обладающий представляющей интерес активностью связывания, можно получить контактированием фага с представляющим интерес антигеном, с последующим сбором фага со связанным с ним антигеном. scFv, обладающий представляющей интерес активностью связывания, можно концентрировать повторением указанной процедуры при необходимости.
Кодирующий антитело полинуклеотид согласно изобретению может кодировать полноразмерное антитело или часть антитела. "Часть антитела" относится к любой части молекулы антитела. В дальнейшем в настоящем описании термин "фрагмент антитела" можно использовать для обозначения части антитела. Предпочтительный фрагмент антитела согласно изобретению содержит определяющую комплементарность область (CDR) антитела. Более предпочтительно фрагмент антитела согласно изобретению содержит все из трех CDR, которые составляют вариабельную область.
После получения кДНК, кодирующей V-область представляющего интерес антитела против EREG, кДНК расщепляют ферментами рестрикции, которые распознают участки для фермента рестрикции, встроенные на оба конца кДНК. Предпочтительный фермент рестрикции распознает и расщепляет нуклеотидную последовательность, которая с меньшей вероятностью встретится в нуклеотидной последовательности, составляющей ген антитела. Более того, для встраивания отдельной копии расщепленного фрагмента в вектор в правильном направлении, предпочтительным является фермент рестрикции, обеспечивающий "липкие" концы. кДНК, кодирующую V-область антитела против EREG, расщепленную, как описано выше, встраивают в пригодный экспрессирующий вектор с получением экспрессирующего вектора антитела. На этой стадии можно получать химерное антитело посредством слияния в рамке считывания гена, кодирующего константную область антитела (C-область), с указанным выше геном, кодирующим V-область. В настоящем описании "химерное антитело" относится к антителу, константные и вариабельные области которого имеют различные источники. Таким образом, в дополнение к межвидовым химерным антителам, таким как химерные антитела мыши-человека, межвидовые химерные антитела человека-человека, также включены в химерные антитела согласно изобретению. Экспрессирующий химерное антитело вектор также можно конструировать посредством встраивания указанного выше гена V-области в экспрессирующий вектор, в который встроен ген константной области.
Более конкретно, например, на 5'-конец экспрессирующего вектора, обладающего ДНК, кодирующей константную область (C-область) требуемого антитела, можно поместить последовательность для распознавания ферментом рестрикции для фермента рестрикции, который расщепляет указанный выше ген V-области. Экспрессирующий химерное антитело вектор конструируют расщеплением двух генов с использованием одинакового сочетания ферментов рестрикции и слияния их в рамке считывания.
Для получения антитела против EREG для применения в настоящем изобретении ген антитела может быть встроен в экспрессирующий вектор, таким образом, чтобы он экспрессировался под регуляцией контролирующего экспрессию участка. Регулирующий экспрессию участок для экспрессии антитела включает, например, энхансер или промотор. Затем посредством трансформации пригодных клеток-хозяев данным экспрессирующим вектором можно получить рекомбинантные клетки, которые экспрессируют ДНК, кодирующую антитело против EREG.
Для экспрессии гена антитела, ДНК, кодирующую тяжелую цепь (H-цепь) антитела, и ДНК, кодирующую легкую цепь (L-цепь) антитела, можно встраивать по отдельности в экспрессирующие векторы. Молекулу антитела, содержащую H-цепь и L-цепь, можно экспрессировать посредством одновременной трансфекции (котрансфекции) объединенных векторов для H-цепи и L-цепи в одну и ту же клетку-хозяина. Альтернативно, ДНК, кодирующие H-цепь и L-цепь, можно встраивать в один экспрессирующий вектор для трансформации клетки-хозяина данным вектором (см. международную публикацию патента No. WO 94/11523).
Известно множество комбинаций хозяев и экспрессирующих векторов для выделения гена антитела с последующим введением гена в соответствующего хозяина, с получением антитела. Для настоящего изобретения можно использовать любые из таких экспрессирующих систем. При использовании эукариотических клеток в качестве хозяина можно использовать клетки животных, клетки растений и клетки грибов. Более конкретно, клетки животных, которые можно использовать в настоящем изобретении, представляют собой, например, следующие клетки:
(1) клетки млекопитающих, такие как клетки CHO, COS, миеломы, почки детенышей хомяка (BHK), HeLa и Vero;
(2) клетки амфибий, такие как овоциты Xenopus и
(3) клетки насекомых, такие как sf9, sf21, Tn5.
Кроме того, в качестве растительной клеточной системы, известна система экспрессии генов антител с использованием клеток, полученных из рода Nicotiana, таких как Nicotiana tabacum. Для трансформации клеток растений можно использовать каллусные культивируемые клетки.
Более того, в качестве клеток грибов можно использовать следующие клетки; дрожжи: рода Saccharomyces, например, Saccharomyces cerevisiae, и рода Pichia, например, Pichia pastoris; и нитчатые грибы: рода Aspergillus, например, Aspergillus niger.
Системы экспрессии генов антител, которые используются прокариотическими клетками, также известны. Например, при использовании бактериальных клеток в настоящем изобретении можно использовать клетки E. coli, клетки Bacillus subtilis и т.п.
Экспрессирующие векторы, содержащие представляющие интерес гены антител, вводят в данные клетки посредством трансформации. Посредством культивирования трансформированных клеток in vitro из культуры трансформированных клеток можно получить требуемые антитела.
В дополнение к указанным выше клеткам-хозяевам для продукции рекомбинантного антитела также можно использовать трансгенных животных. Таким образом, антитело можно получить из животного, у которого встроен ген, кодирующий представляющее интерес антитело. Например, ген антитела можно встраивать в рамке считывания в ген, который кодирует белок, продуцируемый природным образом в молоке, для конструирования слитого гена. Например, в качестве белка, секретируемого в молоке, можно использовать β-казеин козы или подобные. Фрагмент ДНК, содержащий ген для слияния, в который встроен ген антитела, инъецируют в эмбрион козы и затем этот эмбрион вводят самке козы. Требуемые антитела можно получать в качестве белка, слитого с белком молока, из молока, продуцируемого трансгенной козой (или ее потомком), рожденной козой, которой ввели эмбрион. Для повышения объема молока, содержащего требуемое антитело, продуцируемое трансгенной козой, при необходимости трансгенной козе можно вводить гормоны (Ebert, K.M. et al., Bio/Technology (1994) 12, 699-702).
C-области антитела из животного можно использовать для C-областей рекомбинантного антитела согласно изобретению. Например, Cγ1, Cγ2a, Cγ2b, Cγ3, Cμ, Cδ, Cα1, Cα2 и Cε можно использовать для C-области H-цепи антитела мыши и Cκ и Cλ можно использовать для C-области L-цепи. В дополнение к антителам мыши, в качестве антител животных можно использовать антитела животных, таких как крысы, кролики, козы, овцы, верблюды и обезьяны. Их последовательности известны. Более того, для повышения стабильности антител или их продукции C-область можно модифицировать.
В настоящем изобретении при введении антител людям можно использовать генетически рекомбинантные антитела, которые искусственно модифицированы для снижения ксеноантигенности против человека, или подобные. Примеры генетически рекомбинантных антител включают химерные антитела и гуманизированные антитела. Такие модифицированные антитела можно получить с использованием известных способов.
Химерное антитело представляет собой антитело, вариабельные области и константные области которого имеют различное происхождение. Например, антитело, содержащее вариабельные области тяжелой цепи и легкой цепи антитела мыши и константные области тяжелой цепи и легкой цепи антитела человека, представляет собой межвидовое химерное антитело мыши-человека. Рекомбинантный вектор, экспрессирующий химерное антитело, можно получать лигированием ДНК, кодирующей вариабельную область антитела мыши, с ДНК, кодирующей константную область антитела человека, с последующим встраиванием его в экспрессирующий вектор. Рекомбинантные клетки, которые трансформированы вектором, культивируют и включенную ДНК экспрессируют с получением химерного антитела, продуцируемого в культуре. C-области человека используют для C-областей химерных антител и гуманизированных антител.
Например, Cγ1, Cγ2, Cγ3, Cγ4, Cμ, Cδ, Cα1, Cα2 и Cε можно использовать в качестве C-области H-цепи. Cκ и Cλ можно использовать в качестве C-области L-цепи. Аминокислотные последовательности таких C-областей и нуклеотидные последовательности, кодирующие их, известны. Более того, для повышения стабильности антитела и его продукции C-область антитела человека можно модифицировать.
Как правило, химерное антитело состоит из V-области антитела из не относящегося к человеку животного, и C-области из антитела человека. С другой стороны, гуманизированное антитело состоит из определяющей комплементарность области (CDR) антитела из не относящегося к человеку животного и каркасной области (FR) и C-области из антитела человека. Поскольку антигенность гуманизированного антитела в организме человека снижается, гуманизированное антитело пригодно в качестве активного ингредиента для лекарственных средств согласно изобретению.
Например, химерные антитела мыши человека, которые можно получить связыванием аминокислотных последовательностей, составляющих константные области человека и вариабельные области моноклонального антитела против EREG (B3#18), полученного на основе настоящего изобретения, являются предпочтительными моноклональными антителами согласно изобретению. Более конкретно, настоящее изобретение относится к химерным моноклональным антителам мыши-человека, содержащим H и L-цепи, имеющие следующие аминокислотные последовательности.
H-цепь: аминокислотная последовательность из положений 1-446 в аминокислотной последовательности SEQ ID NO: 10 (сигнальная последовательность из положений (-19)-(-1) удалена);
L-цепь: аминокислотная последовательность из положений 1-213 в аминокислотной последовательности SEQ ID NO: 20 (сигнальная последовательность из положений (-20)-(-1) удалена).
Более того, другие предпочтительные примеры химерных антител согласно изобретению включают следующие антитела.
Антитело, содержащее, H-цепь, имеющую последовательность, в которой сигнальная последовательность (-19)-(-1) удалена из аминокислотной последовательности SEQ ID NO: 129, и L-цепь, имеющую последовательность, в которой сигнальная последовательность (-19)-(-1) удалена из аминокислотной последовательности SEQ ID NO: 131.
Антитело, содержащее H-цепь, имеющую последовательность, в которой сигнальная последовательность (-19)-(-1) удалена из аминокислотной последовательности SEQ ID NO: 133, и L-цепь, имеющую последовательность, в которой сигнальная последовательность (-19)-(-1) удалена из аминокислотной последовательности SEQ ID NO: 135.
Антитело, содержащее H-цепь, имеющую последовательность, в которой сигнальная последовательность (-19)-(-1) удалена из аминокислотной последовательности SEQ ID NO: 137, и L-цепь, имеющую последовательность, в которой сигнальная последовательность (-19)-(-1) удалена из аминокислотной последовательности SEQ ID NO: 139.
Антитело, содержащее H-цепь, имеющую последовательность, в которой сигнальная последовательность (-19)-(-1) удалена из аминокислотной последовательности SEQ ID NO: 141, и L-цепь, имеющую последовательность, в которой сигнальная последовательность (-19)-(-1) удалена из аминокислотной последовательности SEQ ID NO: 143.
Антитело, содержащее H-цепь, имеющую последовательность, в которой сигнальная последовательность (-19)-(-1) удалена из аминокислотной последовательности SEQ ID NO: 145, и L-цепь, имеющую последовательность, в которой сигнальная последовательность (-19)-(-1) удалена из аминокислотной последовательности SEQ ID NO: 147.
Вариабельная область антитела, как правило, содержит три определяющих комплементарность участка (CDR), разделенных четырьмя каркасными областями (FR). CDR представляет собой участок, который по существу определяет специфичность связывания антитела. Аминокислотные последовательности CDR весьма разнообразны. С другой стороны, составляющие FR аминокислотные последовательности часто являются высоко гомологичными, даже среди антител с различной специфичностью связывания. Таким образом, как правило, специфичность связывания определенного антитела можно переносить в другое антитело посредством пересадки CDR.
Гуманизированное антитело также называют переформированным антителом человека. Конкретно, гуманизированные антитела, полученные прививкой CDR из антитела не относящегося к человеку животного в антитело человека и подобные, известны. Также известны общие способы генной инженерии для получения гуманизированных антител.
Конкретно, например, в качестве способа прививки CDR антитела мыши в FR человека известна ПЦР с перекрывающимся удлинением. В ПЦР с перекрывающимся удлинением, нуклеотидную последовательность, кодирующую CDR антитела мыши, подлежащую прививке, добавляют к праймерам для синтеза FR антитела человека. Праймеры получают для каждого из четырех FR. Как правило, считается, что при прививке CDR мыши в FR человека выбор FR человека, который высоко гомологичен FR мыши, является преимущественным для сохранения функции CDR. Таким образом, как правило, предпочтительно использовать FR человека, содержащий аминокислотную последовательность, высоко гомологичную аминокислотной последовательности FR, соседней с CDR мыши, подлежащей прививке.
Нуклеотидные последовательности, подлежащие лигированию, конструируют таким образом, чтобы они были соединены друг с другом в рамке считывания. FR человека по отдельности синтезируют с использованием соответствующих праймеров. В результате получают продукты, в которых ДНК, кодирующая CDR мыши, связана с ДНК, кодирующей отдельные FR. Нуклеотидные последовательности, кодирующие CDR мыши каждого продукта, конструируют таким образом, чтобы они перекрывались друг с другом. Затем проводят отжиг перекрывающихся участков CDR продуктов, синтезированных с использованием гена антитела человека в качестве матрицы, для реакции синтеза комплементарной цепи. Посредством такой реакции FR человека лигируют через последовательности CDR мыши.
Полноразмерный ген V-области, в котором в конечном итоге лигированы три CDR и четыре FR, амплифицируют с использованием праймеров, которые гибридизуются на его 5'- и 3'-концах и которые обладают пригодными последовательностями для распознавания ферментом рестрикции. Вектор для экспрессии антитела человека можно получить встраиванием ДНК, полученной, как описано выше, и ДНК, которая кодирует C-область антитела человека, в экспрессирующий вектор, таким образом, он будет лигироваться в рамке считывания. После встраивания единого вектора в хозяина для получения рекомбинантных клеток, рекомбинантные клетки культивируют и ДНК, кодирующую гуманизированное антитело, экспрессируют с получением гуманизированного антитела в клеточной культуре (см. публикацию Европейского патента No. EP 239400 и международную публикацию патента No. WO 96/02576).
Посредством качественного или количественного измерения и оценки антигенсвязывающей активности гуманизированного антитела, полученного, как описано выше, можно пригодным образом выбрать FR антитела человека, которые дают возможность CDR образовывать подходящий антигенсвязывающий участок при лигировании через CDR. При необходимости, аминокислотные остатки в FR могут быть замещены таким образом, чтобы CDR переформированного антитела человека образовывало соответствующий антигенсвязывающий участок. Например, в FR можно вносить мутации аминокислотной последовательности применением способа ПЦР, используемого для слияния CDR мыши с FR человека. Более конкретно, в праймеры, которые гибридизуются с последовательностью FR, можно вносить частичные мутации нуклеотидной последовательности. Мутации нуклеотидной последовательности вносят в FR, синтезированные с использованием таких праймеров. Мутантные последовательности FR, имеющие требуемые характеристики, можно отбирать путем определения и оценки активности мутантного антитела с замещенной аминокислотой в отношении связывания антигена с помощью указанного выше способа (Sato, K. et al., Cancer Res. 1993, 53, 851-856).
Способы получения антител человека также известны. Например, лимфоциты человека сенсибилизируют in vitro требуемым антигеном или клетками, экспрессирующими требуемый антиген. Затем посредством слияния сенсибилизированных лимфоцитов с клетками миеломы человека можно получить требуемые антитела человека, обладающие антигенсвязывающей активностью (см. JP-B H01-59878). В качестве партнера для слияния можно использовать клетки миеломы человека U266 или подобные.
Альтернативно, требуемое антитело человека можно получить с использованием требуемого антигена для иммунизации трансгенного животного, которое содержит полный набор генов антител человека (см. международные патентные публикации No. WO 93/12227, WO 92/03918, WO 94/02602, WO 94/25585, WO 96/34096 и WO 96/33735). Более того, также известны способы получения антител человека посредством пэннинга библиотеки антител человека. Например, V-область антитела человека экспрессируют в качестве одноцепочечного антитела (scFv) на поверхности фага с использованием способа фагового дисплея, и можно проводить селекцию фагов, которые связываются с антигеном. Посредством анализа генов выбранных фагов можно определять последовательности ДНК, кодирующие V-области антител человека, которые связываются с антигеном. После определения последовательностей ДНК scFv, которые связываются с антигеном, последовательность V-области подвергают слиянию в рамке считывания с требуемой последовательностью C-области антитела человека и ее встраивают в пригодный экспрессирующий вектор с получением экспрессирующего вектора. Экспрессирующий вектор можно встраивать в пригодные экспрессирующие клетки, такие как клетки, описанные выше, и ген, кодирующий антитело человека, можно экспрессировать с получением антител человека. Такие способы хорошо известны (международные патентные публикации No. WO 92/01047, WO 92/20791, WO 93/06213, WO 93/11236, WO 93/19172, WO 95/01438 и WO 95/15388).
Антитело, используемое в настоящем изобретении, не ограничивается двухвалентными антителами в виде IgG, но также они включают одновалентные антитела и поливалентные антитела в виде IgM, при условии, что они связываются с белком EREG. Поливалентное антитело согласно изобретению включает поливалентное антитело, которое обладает одинаковыми антигенсвязывающими участками, и поливалентное антитело, которое обладает частично или полностью отличающимися антигенсвязывающими участками. Антитело, используемое в настоящем изобретении, не ограничивается целой молекулой антитела, но также включает мини-антитела и их модифицированные продукты, при условии, что они связываются с белком EREG.
Мини-антитело содержит фрагмент антитела, лишенный части целого антитела (например, целого IgG). При условии наличия способности связывать антиген EREG, допустимы частичные делеции молекулы антитела. Фрагменты антител согласно изобретению предпочтительно содержат вариабельную область тяжелой цепи (VH) и/или вариабельную область легкой цепи (VL). Аминокислотная последовательность VH или VL может обладать заменами, делециями, добавлениями и/или инсерциями. Более того, при условии, что антитело обладает способностью связывать антиген EREG, VH и/или VL, могут быть частично удалены. Вариабельная область может быть химеризованной или гуманизированной. Конкретные примеры фрагментов антител включают Fab, Fab', F(ab')2 и Fv. Конкретные примеры мини-антител включают Fab, Fab', F(ab')2, Fv, scFv (одноцепочечный Fv), димерное антитело и sc(Fv)2 (одноцепочечный (Fv)2). Мультимеры таких антител (например, димеры, тримеры, тетрамеры и полимеры) также включены в мини-антитела согласно изобретению.
Фрагменты антител можно получить обработкой антитела ферментом, с получением фрагментов антител. Известные ферменты, которые образуют фрагменты антител, представляют собой, например, папаин, пепсин и плазмин. Альтернативно, можно конструировать гены, кодирующие указанные фрагменты антител, встраивать их в экспрессирующие векторы и затем экспрессировать в подходящих клетках-хозяевах (см., например, Co, M.S. et al., J. Immunol. (1994) 152, 2968-2976; Better, M. and Horwitz, A.H., Methods in Enzymology (1989) 178, 476-496; Plueckthun, A. and Skerra, A., Methods in Enzymology (1989) 178, 476-496; Lamoyi, E., Methods in Enzymology (1989) 121, 652-663; Rousseaux, J. et al., Methods in Enzymology (1989) 121, 663-669; и Bird, R.E. et al., TIBTECH (1991) 9, 132-137).
Расщепляющие ферменты расщепляют конкретные участки фрагмента антитела и приводят к фрагментам антител с представленными ниже конкретными структурами. С использованием способов генной инженерии для таких ферментативно полученных фрагментов антител можно удалить любую часть антитела.
Расщепление папаином: F(ab)2 или Fab
Расщепление пепсином: F(ab')2 или Fab'
Расщепление плазмином: Facb
Таким образом, мини-антитела согласно изобретению могут представлять собой фрагменты антител, лишенные какого-либо участка, при условии, что они обладают аффинностью связывания в отношении EREG. Более того, в соответствии с настоящим изобретением желательно, чтобы антитела сохраняли их эффекторную активность, в частности, при лечении клеточно-пролиферативных заболеваний, таких как злокачественная опухоль. Более конкретно, предпочтительные мини-антитела согласно изобретению обладают как аффинностью связывания в отношении EREG, так и эффекторной функцией. Эффекторная функция антитела включает активность ADCC и активность CDC. В частности, предпочтительно, чтобы терапевтические антитела согласно изобретению обладали в качестве эффекторной функции активностью ADCC и/или активностью CDC.
Димерное антитело относится к двухвалентному фрагменту антитела, сконструированному слиянием генов (Hollinger P. et al., Proc. Natl. Acad. Sci. USA 90: 6444-6448 (1993); EP 404,097; WO 93/11161 и т.п.). Димерное антитело представляет собой димер, состоящий из двух полипептидных цепей. Как правило, в каждой полипептидной цепи, составляющей димер, VL и VH связаны линкером в пределах одной цепи. Линкер в димерном антителе, как правило, является достаточно коротким, чтобы препятствовать связыванию между VL и VH. Конкретно, аминокислотные остатки, составляющие линкер, представляют собой, например, пять остатков или около этого. Таким образом, VL и VH, которые кодируются одной пептидной цепью, не могут образовывать одноцепочечный фрагмент вариабельной области и образуют димер с другим одноцепочечным фрагментом вариабельной области. В результате, димерные антитела обладают двумя антигенсвязывающими участками.
scFv можно получить лигированием V-области H-цепи и V-области L-цепи антитела. В scFv, V-область H-цепи и V-область L-цепи лигированы через линкер, предпочтительно пептидный линкер (Huston, J.S. et al., Proc. Natl. Acad. Sci. U.S.A., 1988, 85, 5879-5883). Источником V-области H-цепи и V-области L-цепи в scFv могут быть любые антитела, описанные в настоящем описании. Пептидный линкер для лигирования V-областей особенно не ограничен. Например, в качестве линкера можно использовать любой одноцепочечный пептид, состоящий из 3-25 остатков или около того. Более конкретно, например, можно использовать пептидные линкеры, описанные ниже, или подобные.
Для лигирования V-областей можно использовать способы ПЦР, такие как способы, описанные выше. Для лигирования V-областей способами ПЦР сначала в качестве матрицы можно использовать целую ДНК или ДНК, кодирующую требуемую неполную аминокислотную последовательность, выбранную из следующих ДНК:
последовательности ДНК, кодирующей H-цепь или V-область H-цепи указанного выше антитела; и
последовательности ДНК, кодирующей L-цепь или V-область L-цепи указанного выше антитела.
ДНК, кодирующие V-области H-цепи и L-цепи, по отдельности амплифицируют способами ПЦР с использованием пары праймеров, которые обладают последовательностями, соответствующими последовательностям обоих концов ДНК, подлежащей амплификации. Затем получают ДНК, кодирующую часть, представляющую собой пептидный линкер. ДНК, кодирующую пептидный линкер, также можно синтезировать с использованием ПЦР. На 5'-конец используемых праймеров добавляют нуклеотидные последовательности, которые можно лигировать с каждым из синтезированных по отдельности продуктов амплификации V-областей. Затем проводят реакцию ПЦР с использованием "ДНК V-области H-цепи", "ДНК пептидного линкера" и "ДНК V-области L-цепи" и праймеров для ПЦР со сборкой.
Праймеры для ПЦР со сборкой состоят из комбинации праймера, который гибридизуется с 5'-концом "ДНК V-области H-цепи", и праймера, который гибридизуется с 3'-концом "ДНК V-области L-цепи". Таким образом, праймеры для ПЦР со сборкой представляют собой набор праймеров, которые могут амплифицировать ДНК, кодирующую полноразмерную последовательность scFv, подлежащего синтезу. С другой стороны, нуклеотидные последовательности, которые можно лигировать с каждой ДНК V-области, добавляют к "ДНК пептидного линкера". Таким образом, ДНК лигируются, и в конечном итоге получают полноразмерный scFv в качестве продукта амплификации с использованием праймеров для ПЦР со сборкой. После конструирования кодирующей scFv ДНК, экспрессирующие векторы, содержащие ДНК, и рекомбинантные клетки, трансформированные этими экспрессирующими векторами, можно получить в соответствии с общепринятыми способами. Более того, scFv можно получить культивированием полученных рекомбинантных клеток и экспрессией кодирующей scFv ДНК.
sc(Fv)2 представляет собой мини-антитело, полученное лигированием двух VH и двух VL с линкерами или подобными с образованием одной цепи (Hudson et al., J. Immunol. Methods 1999; 231: 177-189). sc(Fv)2 можно получить, например, связыванием scFv с помощью линкера.
Более того, предпочтительными являются антитела, в которых два VH и два VL расположены в порядке VH, VL, VH и VL ([VH]-линкер-[VL]-линкер-[VH]-линкер-[VL]), начиная с N-концевой стороны одноцепочечного полипептида.
Порядок двух VH и двух VL особенно не ограничивается указанным выше расположением, и они могут быть расположены в любом порядке. Примеры включают следующее расположение:
[VL]-линкер-[VH]-линкер-[VH]-линкер-[VL]
[VH]-линкер-[VL]-линкер-[VL]-линкер-[VH]
[VH]-линкер-[VH]-линкер-[VL]-линкер-[VL]
[VL]-линкер-[VL]-линкер-[VH]-линкер-[VH]
[VL]-линкер-[VH]-линкер-[VL]-линкер-[VH]
Любой произвольный пептидный линкер можно встраивать способами генной инженерии, и в качестве линкеров для связывания вариабельных областей антитела можно использовать синтетические линкеры (см., например, линкеры, описанные в Protein Engineering, 9(3), 299-305, 1996) или подобные. В настоящем изобретении пептидные линкеры являются предпочтительными. Длина пептидных линкеров особенно не ограничена, и ее могут пригодным образом выбрать специалисты в данной области в зависимости от цели. Длина аминокислотных остатков, составляющих пептидный линкер, как правило, составляет 1-100 аминокислот, предпочтительно 3-50 аминокислот, более предпочтительно 5-30 аминокислот и особенно предпочтительно 12-18 аминокислот (например, 15 аминокислот).
Можно использовать любые аминокислотные последовательности, составляющие пептидные линкеры, при условии, что они не ингибируют активность связывания scFv. Примеры аминокислотных последовательностей, используемых в пептидных линкерах, включают:
Ser
Gly-Ser
Gly-Gly-Ser
Ser-Gly-Gly
Gly-Gly-Gly-Ser (SEQ ID NO: 23)
Ser-Gly-Gly-Gly (SEQ ID NO: 24)
Gly-Gly-Gly-Gly-Ser (SEQ ID NO: 25)
Ser-Gly-Gly-Gly-Gly (SEQ ID NO: 26)
Gly-Gly-Gly-Gly-Gly-Ser (SEQ ID NO: 27)
Ser-Gly-Gly-Gly-Gly-Gly (SEQ ID NO: 28)
Gly-Gly-Gly-Gly-Gly-Gly-Ser (SEQ ID NO: 29)
Ser-Gly-Gly-Gly-Gly-Gly-Gly (SEQ ID NO: 30)
(Gly-Gly-Gly-Gly-Ser (SEQ ID NO: 31))n
(Ser-Gly-Gly-Gly-Gly (SEQ ID NO: 32))n
где n является целым числом, равным 1 или более. Аминокислотные последовательности пептидных линкеров могут пригодным образом выбрать специалисты в данной области в зависимости от цели. Например, n, которое определяет длину пептидного линкера, как правило, равен 1-5, предпочтительно 1-3, более предпочтительно 1 или 2.
Таким образом, особенно предпочтительный вариант осуществления sc(Fv)2 в настоящем изобретении представляет собой, например, следующий sc(Fv)2:
[VH]-пептид линкер (15 аминокислот)-[VL]-пептид линкер (15 аминокислот)-[VH]-пептид линкер (15 аминокислот)-[VL].
Альтернативно, для связывания V-областей можно использовать синтетические химические линкеры (химические поперечно-сшивающие агенты). В настоящем изобретении можно использовать поперечно-сшивающие агенты, обычно используемые для поперечного сшивания пептидных соединений и подобных. Например, следующие химические поперечно-сшивающие агенты известны. Такие поперечно-сшивающие агенты коммерчески доступны:
N-гидроксисукцинимид (NHS);
дисукцинимидилсуберат (DSS);
бис(сульфосукцинимидил)суберат (BS3);
дитиобис(сукцинимидилпропионат) (DSP);
дитиобис(сульфосукцинимидилпропионат) (DTSSP);
бис(сукцинимидилсукцинат) этиленгликоля (EGS);
бис(сульфосукцинимидилсукцинат) этиленгликоля (сульфо-EGS);
дисукцинимидилтартрат (DST);
дисульфосукцинимидилтартрат (сульфо-DST);
бис[2-(сукцинимидоксикарбонилокси)этил]сульфон (BSOCOES) и
бис[2-(сульфосукцинимидоксикарбонилокси)этил]сульфон (сульфо-BSOCOES).
Обычно для связывания четырех вариабельных областей антитела требуются три линкера. Несколько линкеров, для применения, могут быть все одного типа или различных типов. В настоящем изобретении предпочтительным мини-антителом является димерное антитело или sc(Fv)2. Такое мини-антитело можно получить обработкой антитела ферментом, таким как папаин или пепсин, с получением фрагментов антител или конструированием ДНК, которые кодируют фрагменты антител, встраиванием их в экспрессирующие векторы с последующей экспрессией их в подходящих клетках-хозяевах (см., например, Co, M.S. et al., J. Immunol. (1994) 152, 2968-2976; Better, M. and Horwitz, A.H., Methods Enzymol. (1989) 178, 476-496; Pluckthun, A. and Skerra, A., Methods Enzymol. (1989) 178, 497-515; Lamoyi, E., Methods Enzymol. (1986) 121, 652-663; Rousseaux, J. et al., Methods Enzymol. (1986) 121, 663-669; и Bird, R.E. and Walker, B.W., Trends Biotechnol. (1991) 9, 132-137).
В качестве антитела согласно изобретению можно использовать любое распознающее EREG антитело. Предпочтительные антитела включают антитела согласно (1)-(57), представленные ниже. Эти антитела могут представлять собой, например, полноразмерные антитела, мини-антитела, антитела животных, химерные антитела, гуманизированные антитела или антитела человека:
(1) антитело, содержащее H-цепь, имеющую аминокислотную последовательность SEQ ID NO: 2 (последовательность CDR1 H-цепи антитела B3#18) в качестве CDR1, аминокислотную последовательность SEQ ID NO: 4 (последовательность CDR2 H-цепи антитела B3#18) в качестве CDR2 и аминокислотную последовательность SEQ ID NO: 6 (последовательность CDR3 H-цепи антитела B3#18) в качестве CDR3;
(2) антитело, содержащее H-цепь согласно (1), где H-цепь имеет аминокислотную последовательность из положений 117-452 в аминокислотной последовательности SEQ ID NO: 8 (последовательность CH антитела B3#18) в качестве CH (константной области H-цепи);
(3) антитело, содержащее H-цепь согласно (1), где H-цепь имеет аминокислотную последовательность из положений 117-446 в аминокислотной последовательности SEQ ID NO: 10 (последовательность CH химерного антитела мыши-человека DF151) в качестве CH;
(4) антитело, содержащее L-цепь, имеющую аминокислотную последовательность SEQ ID NO: 12 (последовательность CDR1 L-цепи антитела B3#18) в качестве CDR1, аминокислотную последовательность SEQ ID NO: 14 (последовательность CDR2 L-цепи антитела B3#18) в качестве CDR2 и аминокислотную последовательность SEQ ID NO: 16 (последовательность CDR3 L-цепи антитела B3#18) в качестве CDR3;
(5) антитело, содержащее L-цепь согласно (4), где L-цепь имеет аминокислотную последовательность из положений 107-213 в аминокислотной последовательности SEQ ID NO: 18 (последовательность CL антитела B3#18) в качестве CL (константной области L-цепи);
(6) антитело, содержащее L-цепь согласно (4), где L-цепь имеет аминокислотную последовательность из положений 107-213 в аминокислотной последовательности SEQ ID NO: 20 (последовательность CL химерного антитела мыши-человека B3#18) в качестве CL;
(7) антитело, содержащее H-цепь согласно (1) и L-цепь согласно (4);
(8) антитело, содержащее H-цепь согласно (2) и L-цепь согласно (5);
(9) антитело, содержащее H-цепь согласно (3) и L-цепь согласно (6);
(10) антитело, содержащее H-цепь, имеющую аминокислотную последовательность SEQ ID NO: 49 в качестве CDR1, аминокислотную последовательность SEQ ID NO: 51 в качестве CDR2 и аминокислотную последовательность SEQ ID NO: 53 в качестве CDR3;
(11) антитело, содержащее H-цепь согласно (10), где H-цепь имеет CH, полученную из мышиного IgG1;
(12) антитело, содержащее H-цепь согласно (10), где H-цепь имеет аминокислотную последовательность из положений 117-446 в аминокислотной последовательности SEQ ID NO: 10 в качестве CH;
(13) антитело, содержащее L-цепь, имеющую аминокислотную последовательность SEQ ID NO: 55 в качестве CDR1, аминокислотную последовательность SEQ ID NO: 57 в качестве CDR2 и аминокислотную последовательность SEQ ID NO: 59 в качестве CDR3;
(14) антитело, содержащее L-цепь согласно (13), где L-цепь имеет CL, полученную из мышиной κ-цепи;
(15) антитело, содержащее L-цепь согласно (13), где L-цепь имеет аминокислотную последовательность из положений 107-213 в аминокислотной последовательности SEQ ID NO: 20 в качестве CL;
(16) антитело, содержащее H-цепь согласно (10) и L-цепь согласно (13);
(17) антитело, содержащее H-цепь согласно (11) и L-цепь согласно (14);
(18) антитело, содержащее H-цепь согласно (12) и L-цепь согласно (15);
(19) антитело, содержащее H-цепь, имеющую аминокислотную последовательность SEQ ID NO: 61 в качестве CDR1, аминокислотную последовательность SEQ ID NO: 63 в качестве CDR2, и аминокислотную последовательность SEQ ID NO: 65 в качестве CDR3;
(20) антитело, содержащее H-цепь согласно (19), где H-цепь имеет CH, полученную из мышиного IgG1;
(21) антитело, содержащее H-цепь согласно (19), где H-цепь имеет аминокислотную последовательность из положений 117-446 в аминокислотной последовательности SEQ ID NO: 10 в качестве CH;
(22) антитело, содержащее L-цепь, имеющую аминокислотную последовательность SEQ ID NO: 67 в качестве CDR1, аминокислотную последовательность SEQ ID NO: 69 в качестве CDR2 и аминокислотную последовательность SEQ ID NO: 71 в качестве CDR3;
(23) антитело, содержащее L-цепь согласно (22), где L-цепь имеет CL, полученную из мышиной κ-цепи;
(24) антитело, содержащее L-цепь согласно (22), где L-цепь имеет аминокислотную последовательность из положений 107-213 в аминокислотной последовательности SEQ ID NO: 20 в качестве CL;
(25) антитело, содержащее H-цепь согласно (19) и L-цепь согласно (22);
(26) антитело, содержащее H-цепь согласно (20) и L-цепь согласно (23);
(27) антитело, содержащее H-цепь согласно (21) и L-цепь согласно (24);
(28) антитело, содержащее H-цепь, имеющую аминокислотную последовательность SEQ ID NO: 73 в качестве CDR1, аминокислотную последовательность SEQ ID NO: 75 в качестве CDR2 и аминокислотную последовательность SEQ ID NO: 77 в качестве CDR3;
(29) антитело, содержащее H-цепь согласно (28), где H-цепь имеет CH, полученную из мышиного IgG1;
(30) антитело, содержащее H-цепь согласно (28), где H-цепь имеет аминокислотную последовательность из положений 117-446 в аминокислотной последовательности SEQ ID NO: 10 в качестве CH;
(31) антитело, содержащее L-цепь, имеющую аминокислотную последовательность SEQ ID NO: 79 в качестве CDR1, аминокислотную последовательность SEQ ID NO: 81 в качестве CDR2 и аминокислотную последовательность SEQ ID NO: 83 в качестве CDR3;
(32) антитело, содержащее L-цепь согласно (31), где L-цепь имеет CL, полученную из мышиной κ-цепи;
(33) антитело, содержащее L-цепь согласно (31), где L-цепь имеет аминокислотную последовательность из положений 107-213 в аминокислотной последовательности SEQ ID NO: 20 в качестве CL;
(34) антитело, содержащее H-цепь согласно (28) и L-цепь согласно (31);
(35) антитело, содержащее H-цепь согласно (29) и L-цепь согласно (32);
(36) антитело, содержащее H-цепь согласно (30) и L-цепь согласно (33);
(37) антитело, содержащее H-цепь, имеющую аминокислотную последовательность SEQ ID NO: 85 в качестве CDR1, аминокислотную последовательность SEQ ID NO: 87 в качестве CDR2 и аминокислотную последовательность SEQ ID NO: 89 в качестве CDR3;
(38) антитело, содержащее H-цепь согласно (37), где H-цепь имеет CH, полученную из мышиного IgG1;
(39) антитело, содержащее H-цепь согласно (37), где H-цепь имеет аминокислотную последовательность из положений 117-446 в аминокислотной последовательности SEQ ID NO: 10 в качестве CH;
(40) антитело, содержащее L-цепь, имеющую аминокислотную последовательность SEQ ID NO: 91 в качестве CDR1, аминокислотную последовательность SEQ ID NO: 93 в качестве CDR2 и аминокислотную последовательность SEQ ID NO: 95 в качестве CDR3;
(41) антитело, содержащее L-цепь согласно (40), где L-цепь имеет CL, полученную из мышиной κ-цепи;
(42) антитело, содержащее L-цепь согласно (40), где L-цепь имеет аминокислотную последовательность из положений 107-213 в аминокислотной последовательности SEQ ID NO: 20 в качестве CL;
(43) антитело, содержащее H-цепь согласно (37) и L-цепь согласно (40);
(44) антитело, содержащее H-цепь согласно (38) и L-цепь согласно (41);
(45) антитело, содержащее H-цепь согласно (39) и L-цепь согласно (42);
(46) антитело, содержащее H-цепь, имеющую аминокислотную последовательность SEQ ID NO: 97 в качестве CDR1, аминокислотную последовательность SEQ ID NO: 99 в качестве CDR2 и аминокислотную последовательность SEQ ID NO: 101 в качестве CDR3;
(47) антитело, содержащее H-цепь согласно (46), где H-цепь имеет CH, полученную из мышиного IgG1;
(48) антитело, содержащее H-цепь согласно (46), где H-цепь имеет аминокислотную последовательность из положений 117-446 в аминокислотной последовательности SEQ ID NO: 10 в качестве CH;
(49) антитело, содержащее L-цепь, имеющую аминокислотную последовательность SEQ ID NO: 103 в качестве CDR1, аминокислотную последовательность SEQ ID NO: 105 в качестве CDR2 и аминокислотную последовательность SEQ ID NO: 107 в качестве CDR3;
(50) антитело, содержащее L-цепь согласно (49), где L-цепь имеет CL, полученную из мышиной κ-цепи;
(51) антитело, содержащее L-цепь согласно (49), где L-цепь имеет аминокислотную последовательность из положений 107-213 в аминокислотной последовательности SEQ ID NO: 20 в качестве CL;
(52) антитело, содержащее H-цепь согласно (46) и L-цепь согласно (49);
(53) антитело, содержащее H-цепь согласно (47) и L-цепь согласно (50);
(54) антитело, содержащее H-цепь согласно (48) и L-цепь согласно (51);
(55) антитело, содержащее одну или несколько аминокислотных замен, делеций, добавлений и/или инсерций в антителе согласно любому из (1)-(54), которое обладает активностью, эквивалентной активности антитела согласно любому из (1)-(54);
(56) антитело, которое связывается с тем же эпитопом белка EREG, что и антитело согласно любому из (1)-(55); и
(57) мини-антитело антитела согласно любому из (1)-(56).
Пример VH в указанной выше H-цепи согласно (1), которая имеет приведенные ниже CDR, включает VH, имеющую аминокислотную последовательность SEQ ID NO: 44 (последовательность VH антитела B3#18):
CDR1: аминокислотная последовательность SEQ ID NO: 2 (последовательность CDR1 H-цепи антитела B3#18 (EP27));
CDR2: аминокислотная последовательность SEQ ID NO: 4 (последовательность CDR2 H-цепи антитела B3#18 (EP27)) и
CDR3: аминокислотная последовательность SEQ ID NO: 6 (последовательность CDR3 H-цепи антитела B3#18 (EP27)).
Пример VL в указанной выше L-цепи согласно (4), которая имеет приведенные ниже CDR, включает VL, имеющую аминокислотную последовательность SEQ ID NO: 46 (последовательность VL антитела B3#18):
CDR1: аминокислотная последовательность SEQ ID NO: 12 (последовательность CDR1 L-цепи антитела B3#18 (EP27));
CDR2: аминокислотная последовательность SEQ ID NO: 14 (последовательность CDR2 L-цепи антитела B3#18 (EP27)) и
CDR3: аминокислотная последовательность SEQ ID NO: 16 (последовательность CDR3 L-цепи антитела B3#18 (EP27)).
Пример VH в указанной выше H-цепи согласно (10), которая имеет приведенные ниже CDR, включает VH, имеющую аминокислотную последовательность SEQ ID NO: 109 (последовательность VH C7 (EP03)):
CDR1: аминокислотная последовательность SEQ ID NO: 49 (последовательность CDR1 H-цепи антитела C7 (EP03));
CDR2: аминокислотная последовательность SEQ ID NO: 51 (последовательность CDR2 H-цепи антитела C7 (EP03)) и
CDR3: аминокислотная последовательность SEQ ID NO: 53 (последовательность CDR3 H-цепи антитела C7 (EP03)).
Пример VL в указанной выше L-цепи согласно (13), которая имеет приведенные ниже CDR, включает VL, имеющую аминокислотную последовательность SEQ ID NO: 111 (последовательность VL антитела C7 (EP03)):
CDR1: аминокислотная последовательность SEQ ID NO: 55 (последовательность CDR1 L-цепи антитела C7 (EP03));
CDR2: аминокислотная последовательность SEQ ID NO: 57 (последовательность CDR2 L-цепи антитела C7 (EP03)) и
CDR3: аминокислотная последовательность SEQ ID NO: 59 (последовательность CDR3 L-цепи антитела C7 (EP03)).
Пример VH в указанной выше H-цепи согласно (19), которая имеет приведенные ниже CDR, включает VH, имеющую аминокислотную последовательность SEQ ID NO: 113 (последовательность VH #15 (EP08)):
CDR1: аминокислотная последовательность SEQ ID NO: 61 (последовательность CDR1 H-цепи антитела #15 (EP08));
CDR2: аминокислотная последовательность SEQ ID NO: 63 (последовательность CDR2 H-цепи антитела #15 (EP08)) и
CDR3: аминокислотная последовательность SEQ ID NO: 65 (последовательность CDR3 H-цепи антитела #15 (EP08)).
Пример VL в указанной выше L-цепи согласно (22), которая имеет приведенные ниже CDR, включает VL, имеющую аминокислотную последовательность SEQ ID NO: 115 (последовательность VL антитела #15 (EP08)):
CDR1: аминокислотная последовательность SEQ ID NO: 67 (последовательность CDR1 L-цепи антитела #15 (EP08));
CDR2: аминокислотная последовательность SEQ ID NO: 69 (последовательность CDR2 L-цепи антитела #15 (EP08)) и
CDR3: аминокислотная последовательность SEQ ID NO: 71 (последовательность CDR3 L-цепи антитела #15 (EP08)).
Пример VH в указанной выше H-цепи согласно (28), которая имеет приведенные ниже CDR, включает VH, имеющую аминокислотную последовательность SEQ ID NO: 117 (последовательность VH B2#30 (EP20)):
CDR1: аминокислотная последовательность SEQ ID NO: 73 (последовательность CDR1 H-цепи антитела B2#30 (EP20));
CDR2: аминокислотная последовательность SEQ ID NO: 75 (последовательность CDR2 H-цепи антитела B2#30 (EP20)) и
CDR3: аминокислотная последовательность SEQ ID NO: 77 (последовательность CDR3 H-цепи антитела B2#30 (EP20)).
Пример VL в указанной выше L-цепи согласно (31), которая имеет приведенные ниже CDR, включает VL, имеющую аминокислотную последовательность SEQ ID NO: 119 (последовательность VL антитела B2#30 (EP20)):
CDR1: аминокислотная последовательность SEQ ID NO: 79 (последовательность CDR1 L-цепи антитела B2#30 (EP20));
CDR2: аминокислотная последовательность SEQ ID NO: 81 (последовательность CDR2 L-цепи антитела B2#30 (EP20)) и
CDR3: аминокислотная последовательность SEQ ID NO: 83 (последовательность CDR3 L-цепи антитела B2#30 (EP20)).
Пример VH в указанной выше H-цепи согласно (37), которая имеет приведенные ниже CDR, включает VH, имеющую аминокислотную последовательность SEQ ID NO: 121 (последовательность VH B3#8 (EP24)):
CDR1: аминокислотная последовательность SEQ ID NO: 85 (последовательность CDR1 H-цепи антитела B3#8 (EP24));
CDR2: аминокислотная последовательность SEQ ID NO: 87 (последовательность CDR2 H-цепи антитела B3#8 (EP24)) и
CDR3: аминокислотная последовательность SEQ ID NO: 89 (последовательность CDR3 H-цепи антитела B3#8 (EP24)).
Пример VL в указанной выше L-цепи согласно (40), которая имеет приведенные ниже CDR, включает VL, имеющую аминокислотную последовательность SEQ ID NO: 123 (последовательность VL антитела B3#8 (EP24)):
CDR1: аминокислотная последовательность SEQ ID NO: 91 (последовательность CDR1 L-цепи антитела B3#8 (EP24));
CDR2: аминокислотная последовательность SEQ ID NO: 93 (последовательность CDR2 L-цепи антитела B3#8 (EP24)); и
CDR3: аминокислотная последовательность SEQ ID NO: 95 (последовательность CDR3 L-цепи антитела B3#8 (EP24)).
Пример VH в указанной выше H-цепи согласно (46), которая имеет приведенные ниже CDR, включает VH, имеющую аминокислотную последовательность SEQ ID NO: 125 (последовательность VH B3#41 (EP29)):
CDR1: аминокислотная последовательность SEQ ID NO: 97 (последовательность CDR1 H-цепи антитела B3#41 (EP29));
CDR2: аминокислотная последовательность SEQ ID NO: 99 (последовательность CDR2 H-цепи антитела B3#41 (EP29)) и
CDR3: аминокислотная последовательность SEQ ID NO: 101 (последовательность CDR3 H-цепи антитела B3#41 (EP29)).
Пример VL в указанной выше L-цепи согласно (49), которая имеет приведенные ниже CDR, включает VL, имеющую аминокислотную последовательность SEQ ID NO: 127 (последовательность VL B3#41 антитела (EP29)):
CDR1: аминокислотная последовательность SEQ ID NO: 103 (последовательность CDR1 L-цепи антитела B3#41 (EP29));
CDR2: аминокислотная последовательность SEQ ID NO: 105 (последовательность CDR2 L-цепи антитела B3#41 (EP29)) и
CDR3: аминокислотная последовательность SEQ ID NO: 107 (последовательность CDR3 L-цепи антитела B3#41 (EP29)).
Указанные выше антитела согласно (1)-(57) включают не только одновалентные антитела, но также поливалентные антитела с двумя или более валентностями. Поливалентные антитела согласно изобретению включают поливалентные антитела, все антигенсвязывающие участки которых являются одинаковыми, и поливалентные антитела, антигенсвязывающие участки которых частично или полностью отличаются.
Предпочтительным вариантом осуществления указанного выше антитела (55) является антитело, в котором CDR не модифицирован. Например, среди указанных выше антител (55) предпочтительным вариантом осуществления "антитела, имеющего одну или несколько аминокислотных замен, делеций, добавлений и/или инсерций в антителе согласно (1) и обладающего активностью, эквивалентной активности антитела согласно (1)" является "антитело, обладающее активностью, эквивалентной активности антитела согласно (1), и имеющее одну или несколько аминокислотных замен, делеций, добавлений и/или инсерций в антителе согласно (1), а также содержащее H-цепь, имеющую аминокислотную последовательность SEQ ID NO: 2 в качестве CDR1, аминокислотную последовательность SEQ ID NO: 4 в качестве CDR2 и аминокислотную последовательность SEQ ID NO: 6 в качестве CDR3". Предпочтительные варианты осуществления других антител, включенных в указанное выше антитело согласно (55), могут быть определены сходным образом.
Способ внесения мутаций в полипептиды представляет собой один из хорошо известных специалистам в данной области способов получения полипептидов, которые функционально эквивалентны определенному полипептиду. Например, специалисты в данной области могут получить антитело, функционально эквивалентное антителу согласно изобретению, посредством внесения соответствующих мутаций в антитело с использованием сайт-направленного мутагенеза (Hashimoto-Gotoh, T. et al. (1995) Gene 152, 271-275; Zoller, MJ, and Smith, M. (1983) Methods Enzymol. 100, 468-500; Kramer, W. et al. (1984) Nucleic Acids Res. 12, 9441-9456; Kramer W, and Fritz HJ (1987) Methods Enzymol. 154, 350-367; Kunkel, TA (1985) Proc. Natl. Acad. Sci. USA. 82, 488-492; Kunkel (1988) Methods Enzymol. 85, 2763-2766) и подобных. Также могут встречаться природные аминокислотные мутации. Таким образом, антитела согласно изобретению также включают антитела, содержащие аминокислотные последовательности с одной или несколькими аминокислотными мутациями в аминокислотных последовательностях антител согласно изобретению и функционально эквивалентные антителам согласно изобретению.
Считают, что число аминокислот, которые являются мутантными в таких мутантных формах, главным образом, составляет 50 аминокислот или менее, предпочтительно 30 аминокислот или менее и более предпочтительно 10 аминокислот или менее (например, 5 аминокислот или менее).
Желательно, чтобы аминокислотные остатки были подвергнуты мутации в аминокислоты, в которых сохранены свойства боковых цепей аминокислот. Например, в зависимости от свойств боковых цепей аминокислот были установлены следующие категории:
гидрофобные аминокислоты (A, I, L, M, F, P, W, Y и V);
гидрофильные аминокислоты (R, D, N, C, E, Q, G, H, K, S и T);
аминокислоты, имеющие алифатические боковые цепи (G, A, V, L, I и P);
аминокислоты, имеющие гидроксилсодержащие боковые цепи (S, T и Y);
аминокислоты, имеющие содержащие серу боковые цепи (C и M);
аминокислоты, имеющие содержащие карбоновую кислоту и амид боковые цепи (D, N, E и Q);
аминокислоты, имеющие основные боковые цепи (R, K и H), и
аминокислоты, имеющие содержащие ароматическое кольцо боковые группы (H, F, Y и W)
(в скобках представлены аминокислоты посредством однобуквенных кодов).
Известно, что полипептиды, обладающие модифицированной аминокислотной последовательностью, в которой один или несколько аминокислотных остатков в определенной аминокислотной последовательности удалены, добавлены и/или заменены другими аминокислотами, сохраняют их исходные виды биологической активности (Mark, D.F. et al., Proc. Natl. Acad. Sci. USA (1984) 81, 5662-5666; Zoller, M.J. & Smith, M. Nucleic Acids Research (1982) 10, 6487-6500; Wang, A. et al., Science 224, 1431-1433; Dalbadie-McFarland, G. et al., Proc. Natl. Acad. Sci. USA (1982) 79, 6409-6413). Таким образом, как правило, в аминокислотной последовательности, составляющей определенный полипептид, активность полипептида с высокой вероятностью сохраняется при взаимной замене аминокислот, классифицированных в одну группу. В настоящем изобретении указанную выше замену между аминокислотами в одной группе аминокислот называют консервативной заменой.
Настоящее изобретение относится к антителам, которые связываются с тем же эпитопом, что и антитела против EREG, описанные в настоящем описании. Более конкретно, настоящее изобретение относится к антителам, которые распознают тот же эпитоп, что и антитело согласно любому из (1)-(57), указанное выше, и к их применению. Такие антитела можно получить, например, следующим способом.
Наличие у антитела общего эпитопа с определенным антителом может быть подтверждено проверкой того, конкурируют ли эти два антитела за один эпитоп. Конкуренцию между антителами можно выявить анализом поперечного сшивания и подобным. Например, конкурентный ELISA является предпочтительным перекрестно-блокирующим анализом.
Конкретно, в анализе перекрестного блокирования на лунки микропланшета для титрования наносят белок EREG, который затем предварительно инкубируют с конкурирующим антителом-кандидатом или без него, и затем добавляют антитело против EREG согласно изобретению. Количество антитела против EREG согласно изобретению, связанного с белком EREG в лунках, косвенно коррелирует со способностью к связыванию конкурирующего антитела-кандидата (тестируемого антитела), которое конкурирует за связывание того же эпитопа. Более конкретно, чем большей аффинностью тестируемое антитело обладает в отношении того же эпитопа, тем более низкой является активность антитела против EREG согласно изобретению в отношении связывания с лунками, на которые нанесен белок EREG. Иными словами, чем более высокой является аффинность тестируемого антитела в отношении того же эпитопа, тем более высокой является активность тестируемого антитела в отношении связывания с лунками, на которые нанесен белок EREG.
Количество антитела, связанного с лунками, можно легко измерить посредством мечения антител заранее. Например, меченное биотином антитело можно определить с использованием конъюгата авидин-пероксидаза и подходящего субстрата. В частности, анализы перекрестного блокирования с использованием ферментных меток, таких как пероксидаза, называют конкурентными анализами ELISA. Антитела можно метить другими поддающимися детектированию или определению материалами для мечения. Более конкретно, известны радиоактивные метки и флуоресцентные метки.
Более того, когда тестируемое антитело содержит константную область из вида, отличного от вида, который является источником антитела против EREG согласно изобретению, определение можно проводить для любого из антител, связанных с лунками с использованием меченого антитела, которое распознает их константную область. Если источником антител является один вид, но они принадлежат различным классам, можно определить антитела, связанные с лунками, с использованием антител, которые отчетливо распознают отдельные классы.
Если конкурирующее антитело-кандидат может блокировать связывание антитела с EREG по меньшей мере на 20%, предпочтительно по меньшей мере на 20-50% и еще более предпочтительно по меньшей мере на 50%, по сравнению с активностью связывания, полученной в контрольном эксперименте, проведенном в отсутствие конкурирующего антитела-кандидата, конкурирующее антитело-кандидат представляет собой либо антитело, которое связывается по существу с тем же эпитопом, либо антитело, которое конкурирует за связывание с тем же эпитопом, что и антитело против EREG согласно изобретению. Антитела, которые связывают тот же эпитоп, что и антитела против EREG, включают, например, указанное выше антитело согласно (57), но не ограничиваются им.
Как описано выше, указанные выше антитела согласно (1)-(57) включают не только одновалентные антитела, но также поливалентные антитела. Поливалентные антитела согласно изобретению включают поливалентные антитела, антигенсвязывающие участки которых являются одинаковыми, и поливалентные антитела, антигенсвязывающие участки которых частично или полностью отличаются.
В качестве модифицированных антител также можно использовать антитела, связанные с молекулами различного типа, такими как полиэтиленгликоль (PEG). Более того, с антителами могут быть связаны химиотерапевтические средства, токсические пептиды или радиоактивные химические вещества. Такие модифицированные антитела (в дальнейшем в настоящем описании обозначаемые как конъюгаты антитела) можно получить, подвергая полученные антитела химической модификации. Способы модификации антител уже разработаны в данной области. Более того, как описано ниже, такие антитела также можно получить в молекулярной форме биспецифичного антитела, сконструированного с использованием способов генной инженерии для распознавания не только белков EREG, но также химиотерапевтических средств, токсических пептидов, радиоактивных химических соединений или подобных. Такие антитела включены в термин "антитела" согласно изобретению.
Химиотерапевтические средства, которые связывают с антителами против EREG для направления цитотоксической активности, включают следующие средства:
азарибин, анастрозол, азацитидин, блеомицин, бортезомиб, бриостатин-1, бисульфан, камптотецин, 10-гидроксикамптотецин, кармустин, целебрекс, хлорамбуцил, цисплатин, иринотекан, карбоплатин, кладрибин, циклофосфамид, цитарабин, декарбазин, доцетаксел, дактиномицин, дауномицина глюкуронид, даунорубицин, дексаметазон, диэтилстилбестрол, доксорубицин, доксорубицина глюкуронид, эпирубицин, этинилэстрадиол, эстрамустин, этопозид, этопозида глюкуронид, флоксуридин, флударабин, флутамид, фторурацил, флуоксиместерон, гемцитабин, гидроксипрогестерона капроат, гидроксимочевина, идарубицин, ифосфамид, лейковорин, ломустин, мехлорэтамин, медроксипрогестерона ацетат, магестрола ацетат, мелфалан, меркаптопурин, метотрексат, митоксантрон, митрамицин, митомицин, митотан, фенилбутират, преднизон, прокарбазин, паклитаксел, пентостатин, семустин стрептозоцин, тамоксифен, таксаны, таксол, тестостерона пропионат, талидомид, тиогуанин, тиотепа, тенипозид, топотекан, иприт урацила, винбластин, винорелбин и винкристин.
В настоящем изобретении предпочтительными химиотерапевтическими средствами являются низкомолекулярные химиотерапевтические средства. Низкомолекулярные химиотерапевтические средства с малой вероятностью препятствуют функции антител, даже после связывания с антителами. В настоящем изобретении низкомолекулярные химиотерапевтические средства, как правило, имеют молекулярную массу от 100 до 2000, предпочтительно 200-1000. Все примеры химиотерапевтических средств, представленных в настоящем описании, являются низкомолекулярными химиотерапевтическими средствами. Химиотерапевтические средства согласно изобретению включают пролекарства, которые превращаются в активные химиотерапевтические средства in vivo. Пролекарство может активировать ферментативное превращение или неферментативное превращение.
Более того, антитела можно модифицировать с использованием токсических пептидов, таких как рицин, абрин, рибонуклеаза, онконаза, ДНКаза I, стафилококковый энтеротоксин-A, противовирусный белок лаконоса, гелонин, дифтерийный токсин, экзотоксин Pseudomonas, эндотоксин Pseudomonas, L-аспарагиназа и PEG-L-аспарагиназа. В другом варианте осуществления один, или два, или более низкомолекулярных химиотерапевтических средств и токсических пептидов можно комбинировать и использовать для модификации антитела. Связывание между антителом против EREG и указанным выше низкомолекулярным химиотерапевтическим средством может представлять собой ковалентное связывание или нековалентное связывание. Способы получения антител, связанных с такими химиотерапевтическими средствами, известны.
Более того, белковые фармацевтические средства или токсины можно связывать с антителами способами рекомбинации генов. Конкретно, например, можно конструировать рекомбинантный вектор посредством слияния ДНК, кодирующей указанный выше токсический пептид, с ДНК, кодирующей антитело против EREG, в рамке считывания и встраивания его в экспрессирующий вектор. Этот вектор вводят в пригодную клетку-хозяина, полученные трансформированные клетки культивируют и встроенную ДНК экспрессируют. Таким образом, антитело против EREG, связанное с токсическим пептидом, можно получить в качестве слитого белка. При получении антитела в качестве слитого белка белковое фармацевтическое средство или токсин, как правило, помещают на C-конце антитела. Между антителом и белковым фармацевтическим средством или токсином может находиться пептидный линкер.
Более того, антитело, используемое в настоящем изобретении, может представлять собой биспецифичное антитело. Биспецифичное антитело относится к антителу, которое обладает вариабельными областями, которые распознают различные эпитопы в одной молекуле антитела. Биспецифичное антитело может иметь антигенсвязывающие участки, которые распознают различные эпитопы на молекуле EREG. Две молекулы такого биспецифичного антитела могут связываться с одной молекулой EREG. В результате можно ожидать более сильного цитотоксического действия.
Альтернативно, биспецифичное антитело может представлять собой антитело, в котором один антигенсвязывающий участок распознает EREG и другой антигенсвязывающий участок распознает цитотоксическое вещество. Конкретно, цитотоксические вещества включают химиотерапевтические средства, токсические пептиды и радиоактивные химические вещества. Такое биспецифичное антитело связывается с экспрессирующими EREG клетками и в то же время захватывает цитотоксические вещества. Это обеспечивает действие цитотоксических веществ непосредственно на экспрессирующие EREG клетки. Таким образом, биспецифичные антитела, которые распознают цитотоксические вещества, специфично повреждают опухолевые клетки и подавляют пролиферацию опухолевых клеток.
Более того, в настоящем изобретении можно комбинировать биспецифичные антитела, которые распознают антигены, отличные от EREG. Например, можно комбинировать биспецифичные антитела, которые распознают не являющиеся EREG антигены, которые специфично экспрессируются на поверхности злокачественных клеток-мишеней, таких как EREG.
Способы получения биспецифичных антител известны. Например, два типа антител, распознающих различные антигены, можно связывать с получением биспецифичного антитела. Антитела, подлежащие связыванию, могут представлять собой половины молекул, каждая из которых имеет H-цепь или L-цепь, или они могут представлять собой четверти молекул, состоящие только из H-цепи. Альтернативно, можно получить продуцирующие биспецифичное антитело слитые клетки слиянием гибридом, продуцирующих различные моноклональные антитела. Биспецифичные антитела также можно получать способами генной инженерии.
Гены антител, сконструированные, как описано выше, можно получить посредством экспрессии известными способами. В случае клеток млекопитающих гены антител можно экспрессировать функциональным связыванием гена антитела, подлежащего экспрессии, с эффективным обычно используемым промотором и сигнальной последовательностью поли-A с 3'-стороны от гена антитела. Примером промотора/энхансера является предранний промотор/энхансер цитомегаловируса.
Другие промоторы/энхансеры, которые можно использовать для экспрессии антитела, включают вирусные промоторы/энхансеры или промоторы/энхансеры клеток млекопитающих, такие как промотор фактора элонгации 1α человека (HEF1α). Конкретные примеры вирусов, чьи промоторы/энхансеры можно использовать, включают ретровирус, вирус полиомы, аденовирус и вирус обезьян 40 (SV40).
Когда используют промотор/энхансер SV40, можно использовать способ Mulligan et al. (Nature (1979) 277, 108). Промотор/энхансер HEF1α можно использовать для экспрессии представляющего интерес гена способом Mizushima et al. (Nucleic Acids Res. (1990) 18, 5322).
В случае E. coli для экспрессии гена ген антитела, подлежащий экспрессии, функционально связывают с эффективным общепринятым промотором и сигнальной последовательностью для секреции антитела. Примеры промотора включают промотор lacZ и промотор araB. Когда используют промотор lacZ, можно использовать способ Ward et al., (Nature (1989) 341, 544-546; FASEB J. (1992) 6, 2422-2427). Альтернативно, промотор araB можно использовать для экспрессии представляющего интерес гена способом Better et al. (Science (1988) 240, 1041-1043).
В отношении сигнальной последовательности для секреции антитела сигнальную последовательность pelB (Lei, S.P. et al., J. Bacteriol. (1987) 169, 4379) можно использовать для продукции в периплазме E. coli. После выделения антитела, продуцированного в периплазме, структуру антитела подвергают повторному сворачиванию с использованием денатурирующего агента для белка, такого как гидрохлорид гуанидина или мочевина, чтобы антитело обладало требуемой активностью связывания.
Ориджин репликации, встроенный в экспрессирующий вектор, включает, например, ориджин репликации из SV40, вируса полиомы, аденовируса или вируса папилломы крупного рогатого скота (BPV). Для амплификации количества копий гена в системе клетки-хозяина, в экспрессирующий вектор можно встраивать селективный маркер. Конкретно можно использовать следующие селективные маркеры:
ген аминогликозидтрансферазы (APH);
ген тимидинкиназы (TK);
ген ксантин-гуанин-фосфорибозилтрансферазы E. coli (Ecogpt);
ген дигидрофолатредуктазы (dhfr) и т.д.
Для продукции антител, используемых в настоящем изобретении, можно использовать любую экспрессирующую систему, например систему эукариотических клеток или систему прокариотических клеток. Примеры эукариотических клеток включают клетки животных, такие как общепризнанная система клеток млекопитающих, система клеток насекомых и клеток нитчатых грибов и дрожжевые клетки. Примеры прокариотических клеток включают бактериальные клетки, такие как клетки E. coli. Антитела, используемые в настоящем изобретении, предпочтительно экспрессируются в клетках млекопитающих. Например, можно использовать клетки млекопитающих, такие как клетки CHO, COS, миеломы, BHK, Vero или HeLa.
Затем трансформированную клетку-хозяина культивируют in vitro или in vivo для индукции продукции представляющего интерес антитела. Клетки-хозяева культивируют известными способами. Например, в качестве культуральной среды можно использовать DMEM, MEM, RPMI 1640 или IMDM. Также вместе со средой можно использовать вспомогательный раствор сыворотки, такой как фетальная телячья сыворотка (FCS).
Антитела, экспрессированные и продуцированные, как описано выше, можно очистить с использованием одного из известных способов или пригодной комбинации известных способов, обычно используемых для очистки белков. Антитела можно выделить и очистить, например, правильным выбором и комбинированием аффинных колонок, таких как колонка с белком A, хроматографическая колонка, фильтрации, ультрафильтрации, осаждения солью, диализа и т.п. (Antibodies A Laboratory Manual. Ed Harlow, David Lane, Cold Spring Harbor Laboratory, 1988).
Для определения антигенсвязывающей активности антител можно использовать известные способы (Antibodies A Laboratory Manual. Ed Harlow, David Lane, Cold Spring Harbor Laboratory, 1988). Например, можно использовать твердофазный иммуноферментный анализ (ELISA), ферментный иммунологический анализ (EIA), радиоиммунный анализ (RIA) или флуороиммунный анализ.
Антитела, используемые в настоящем изобретении, могут представлять собой антитела с модифицированной цепью сахаров. Известно, что цитотоксическую активность антитела можно повышать модификацией его цепи сахаров. Известные антитела, имеющие модифицированные цепи сахаров, включают следующие антитела:
антитела с модифицированным гликозилированием (например, WO 99/54342);
антитела с дефицитом фукозы, присоединенной к цепям сахаров (например, WO 00/61739 и WO 02/31140);
антитела, имеющую цепь сахаров с биссекторным GlcNAc (например, WO 02/79255) и т.д.
Антитела, используемые в настоящем изобретении, предпочтительно представляют собой антитела, обладающие цитотоксической активностью.
В настоящем изобретении цитотоксическая активность включает, например, активность антителозависимой клеточно-опосредуемой цитотоксичности (ADCC) и активность комплементзависимой цитотоксичности (CDC). В настоящем изобретении активность CDC относится к цитотоксической активности, опосредуемой системой комплемента. Активность ADCC относится к активности повреждения клетки-мишени, когда специфичное антитело связывается с ее антигеном клеточной поверхности. Клетка, обладающая Fcγ-рецептором (иммунная клетка или подобные) связывается Fc-областью антигена через Fcγ-рецептор, и клетка-мишень повреждается.
Антитело против EREG можно тестировать, чтобы видеть, обладает ли оно активностью ADCC или активностью CDC, с использованием известных способов (например, Current Protocols in Immunology, Chapter 7. Immunologic studies in humans, Editor, John E. Coligan et al., John Wiley & Sons, Inc., (1993) и т.п.).
Сначала обычно получают эффекторные клетки, раствор комплемента и клетки-мишени.
(1) Получение эффекторных клеток
Селезенку удаляют из мыши CBA/N или подобной и выделяют клетки селезенки в среде RPMI1640 (изготовленной Invitrogen). После промывания той же средой, содержащей 10% фетальную телячью сыворотку (FBS, изготовленную HyClone), концентрацию клеток доводят до 5×106 /мл с получением эффекторных клеток.
(2) Получение раствора комплемента
Комплемент детенышей кролика (изготовленный CEDARLANE) разводят в 10 раз в культуральной среде (изготовленной Invitrogen), содержащей 10% FBS, с получением раствора комплемента.
(3) Получение клеток-мишеней
Клетки-мишени можно подвергнуть радиоактивному мечению посредством инкубации клеток, экспрессирующих белок EREG, с 0,2 мКи хромата-51Cr натрия (изготовленного GE Healthcare Bio-Sciences) в среде DMEM, содержащей 10% FBS, в течение одного часа при 37°C. Для экспрессирующих белок EREG клеток можно использовать клетки, трансформированные геном, кодирующим белок EREG, клетки первичного рака толстого кишечника, метастазирующего рака толстого кишечника, аденокарциномы легкого, рака поджелудочной железы, рака желудка, клетки рака почки, клетки рака толстого кишечника, клетки рака пищевода, клетки рака желудка, клетки рака поджелудочной железы или подобные. После мечения радиоактивной меткой клетки промывают три раза в среде RPMI1640, содержащей 10% FBS, и клетки-мишени можно получить доведением концентрации клеток до 2×105 /мл.
Активность ADCC или активность CDC можно определить способом, описанным ниже. В случае определения активности ADCC, клетку-мишень и антитело против EREG (50 мкл каждого) добавляют в 96-луночный планшет с U-образным дном (изготовленный Becton Dickinson) и дают возможность реагировать в течение 15 минут на льду. После этого добавляют 100 мкл эффекторных клеток и инкубируют в инкубаторе с диоксидом углерода в течение четырех часов. Конечную концентрацию антитела доводят до 0 или 10 мкг/мл. После культивирования 100 мкл супернатанта собирают и с помощью гамма-счетчика (COBRAII AUTO-GAMMA, MODEL D5005, изготовленный Packard Instrument Company) измеряют радиоактивность. Цитотоксическую активность (%) можно вычислить с использованием полученных значений в соответствии с уравнением: (A-C)/(B-C)×100, где A представляет собой радиоактивность (cpm) в каждом образце, B представляет собой радиоактивность (cpm) в образце, в который добавлен 1% NP-40 (изготовленный Nacalai Tesque), и C представляет собой радиоактивность (cpm) в образце, содержащем только клетки-мишени.
Между тем, в случае определения активности CDC 50 мкл клеток-мишеней и 50 мкл антитела против EREG добавляют в 96-луночный планшет с плоским дном (изготовленный Becton Dickinson) и дают возможность реагировать в течение 15 на льду. После этого добавляют 100 мкл раствора комплемента и инкубируют в инкубаторе с диоксидом углерода в течение четырех часов. Конечную концентрацию антитела доводят до 0 или 3 мкг/мл. После инкубации 100 мкл супернатанта собирают и с помощью гамма-счетчика измеряют радиоактивность. Цитотоксическую активность можно вычислить аналогичным способом, как и в случае определения активности ADCC.
С другой стороны, в случае определения цитотоксической активности конъюгата антитела 50 мкл клеток-мишеней и 50 мкл конъюгата антитела против EREG добавляют в 96-луночный планшет с плоским дном (изготовленный Becton Dickinson) и дают возможность реагировать в течение 15 минут на льду. Затем проводят инкубацию в инкубаторе с диоксидом углерода в течение от одного-четырех часов. Конечную концентрацию антитела доводят до 0 или 3 мкг/мл. После культивирования 100 мкл супернатанта собирают и с помощью гамма-счетчика определяют радиоактивность. Цитотоксическую активность можно вычислить аналогичным способом, что и в случае определения активности ADCC.
Антитело согласно изобретению, обладающее цитотоксической активностью, более предпочтительно представляет собой антитело, которое обладает нейтрализующей активностью. Как правило, "нейтрализующая активность" относится к активности ингибирования биологической активности лиганда, такого как вирусы или токсины, в отношении клеток. Более конкретно, "вещества, обладающие нейтрализующей активностью" относятся к веществам, которые связываются с лигандом или с рецептором, связывающимся с лигандом, и ингибируют связывание между лигандом и рецептором. Когда связывание лигандом рецептора блокируется посредством нейтрализующей активности, опосредуемая рецептором биологическая активность может не проявляться. Антитела, которые обладают такой нейтрализующей активностью, как правило, называют нейтрализующими антителами. Нейтрализующую активность можно определить сравнением видов биологической активности в присутствии и в отсутствие тестируемого вещества, нейтрализующая активность которого подлежит оценке, в присутствии представляющего интерес лиганда.
В настоящем изобретении связывание лигандом рецептора EGF, который считают основным рецептором EREG, приводит к димеризации рецептора и активации внутриклеточного тирозинкиназного домена рецептора. Активированная тирозинкиназа образует содержащие фосфорилированный тирозин пептиды посредством аутофосфорилирования, и пептиды связываются с различными вспомогательными молекулами передачи сигнала. Они, главным образом, представляют собой PLCγ (фосфолипаза Cγ), Shc, Grb2 и т.п. Среди таких вспомогательных молекул первые две далее фосфорилируются тирозинкиназой рецептора EGF. Основной каскад передачи сигнала с рецептора EGF представляет собой каскад, в котором фосфорилирование передается в следующем порядке: Shc, Grb2, Sos, Ras, Raf/MAPK-киназа/MAP-киназа. Более того, считают, что существует альтернативный каскад от PLCγ-PKC.
Поскольку такие внутриклеточные каскады передачи сигнала варьируют в зависимости от типа клеток, пригодные молекулы могут быть нацелены на представляющую интерес клетку-мишень, и молекулы-мишени не ограничены указанными выше факторами. Для определения активации передачи сигнала in vivo можно пригодным образом использовать коммерчески доступные наборы (например, систему для анализа активности протеинкиназы C (GE Healthcare Bio-Sciences)).
Более того, активацию передачи сигнала in vivo можно выявить с использованием в качестве показателя эффекта индукции транскрипции гена-мишени, расположенного далее в каскаде передачи сигнала in vivo. Изменения активности транскрипции можно выявлять на основе принципа репортерного анализа. Более конкретно, репортерный ген, такой как ген зеленого флуоресцентного белка (GFP) или люциферазы, помещают ниже фактора транскрипции или промоторной области гена-мишени и определяют активность репортерного гена. Изменение активности транскрипции можно проводить на основе репортерной активности.
Более того, поскольку рецептор EGF обычно функционирует, обеспечивая клеточную пролиферацию, активацию передачи сигнала in vivo можно оценивать посредством определения активности пролиферации клеток-мишеней. В настоящем изобретении нейтрализующую активность нейтрализующего антитела согласно изобретению оценивают посредством оценки активности клеточной пролиферации. Однако настоящее изобретение не ограничивается этим способом, и нейтрализующую активность можно оценивать, подходящим образом применяя указанные выше способы в отношении выбранных клеток-мишеней.
Конкретно, например, посредством измерения указанной ниже активности клеточной пролиферации можно оценивать или измерять нейтрализующую активность антитела против EREG. Например, используют способ, где измеряют включение [3H]-меченного тимидина, добавленного в среду, живыми клетками в качестве показателя способности к репликации ДНК.
В качестве более удобного способа используют способ вытеснения красителя, где под микроскопом определяют способность клеток высвобождать краситель, такой как трипановый синий, во внешнюю среду клетки, или способ MTT. В последнем используется способность живых клеток превращать бромид 3-(4,5-диметилтиазол-2-ил)-2,5-дифенилтетразолия (MTT), который представляет собой соль тетразолия, в синий продукт формазана. Более конкретно, тестируемое антитело добавляют в культуральный раствор с тестируемой клеткой и после определенного периода времени в культуральный раствор добавляют раствор MTT и оставляют стоять в течение определенного периода времени для включения MTT в клетку. В результате MTT, который представляет собой желтое соединение, в митохондриях клетки под действием сукцинатдегидрогеназы превращается в синее соединение. После растворения синего продукта для окрашивания измеряют поглощение и его используют в качестве показателя количества жизнеспособных клеток.
Помимо MTT, коммерчески доступны реагенты, такие как MTS, XTT, WST-1 и WST-8 (Nacalai Tesque и подобные), и их можно пригодным образом использовать. Более того, известны способы, где в качестве показателя оценивают активность клеточной пролиферации с использованием клеточного АТФ или сопротивления клеточной культуры. Для определения активности связывающее антитело, которое имеет то же изотип, что и антитело против EREG, но не обладает нейтрализующей активностью, можно использовать в качестве контрольного антитела таким же образом, как и антитело против EREG, и наличие активности можно определить, если антитело против EREG обладает более сильной нейтрализующей активностью, чем контрольное антитело.
Клетки, пролиферацию которых подавляет антитело против EREG, особенно не ограничиваются, при условии, что они экспрессируют белок EREG. Предпочтительными экспрессирующими EREG клетками являются, например, злокачественные клетки. Конкретно, клетки, источником которых является рак толстого кишечника, аденокарцинома легкого, рак поджелудочной железы, рак желудка или рак почки, пригодны в качестве экспрессирующих EREG клеток согласно изобретению. В соответствии с настоящим изобретением эффекта подавления эффективной клеточной пролиферации можно достигнуть как для первичных, так и для метастатических очагов указанных злокачественных опухолей. Более предпочтительными злокачественными клетками являются клетки первичного рака толстого кишечника, метастазирующего рака толстого кишечника, аденокарциномы легкого, рака поджелудочной железы, рака желудка и рака почки. Таким образом, антитело против EREG можно использовать для лечения или профилактики индуцируемых клеточной пролиферацией заболеваний, таких как рак толстого кишечника, аденокарцинома легкого, рак поджелудочной железы, рак желудка и рак почки. Такие злокачественные опухоли могут быть мишенями для лечения или профилактики, независимо от того, являются ли они первичными очагами или метастатическими очагами. Более предпочтительно антитело против EREG можно использовать для лечения и/или профилактики первичного рака толстого кишечника, метастазирующего рака толстого кишечника или рака поджелудочной железы. Более того, среди таких злокачественных опухолей злокачественные опухоли, которые пролиферируют зависимым от EREG образом, являются предпочтительными мишенями для лечения и/или профилактики согласно изобретению.
Настоящее изобретение также относится к полинуклеотидам, кодирующим антитела согласно изобретению, и полинуклеотидам, которые гибридизуются в строгих условиях с данными полинуклеотидами и кодируют антитела, обладающие активностью, эквивалентной активности антител согласно изобретению. Настоящее изобретение также относится к векторам, содержащим указанные полинуклеотиды, и трансформантам (включая трансформированные клетки), содержащим такие векторы.
Полинуклеотиды согласно изобретению представляют собой полимеры, содержащие множество нуклеотидов или пар оснований дезоксирибонуклеиновых кислот (ДНК) или рибонуклеиновых кислот (РНК), и они особенно не ограничиваются при условии, что они кодируют антитела согласно изобретению. Полинуклеотиды согласно изобретению также могут содержать неприродные нуклеотиды.
Полинуклеотиды согласно изобретению можно использовать для экспрессии антител с использованием способов генной инженерии. Более того, их можно использовать в качестве зондов при скрининге антител, которые функционально эквивалентны антителам согласно изобретению. Конкретно, ДНК, которая гибридизуется в строгих условиях с полинуклеотидом, кодирующим антитело согласно изобретению, и кодирует антитело, обладающее активностью, эквивалентной активности антитела согласно изобретению, можно получить способами, такими как гибридизация и способы амплификации генов (например, ПЦР), с использованием полинуклеотида, кодирующего антитело согласно изобретению, или его части в качестве зонда. Такие ДНК включены в полинуклеотиды согласно изобретению. Способы гибридизации хорошо известны специалистам в данной области (Sambrook, J. et al., Molecular Cloning 2nd ed., 9,47-9,58, Cold Spring Harbor Lab. press, 1989).
Условия гибридизации включают, например, условия низкой строгости. Примеры условий низкой строгости включают промывание после гибридизации в условиях 0,1× SSC и 0,1% SDS при 42°C и предпочтительно в условиях 0,1× SSC и 0,1% SDS при 50°C. Более предпочтительные условия гибридизации включают условия высокой строгости. Условия высокой строгости включают, например, условия 5× SSC и 0,1% SDS при 65°C. В этих условиях можно ожидать, что более высокая температура приведет к эффективному получению полинуклеотидов с высокой гомологией. Однако на строгость гибридизации могут влиять несколько факторов, таких как температура и концентрация соли, и специалисты в данной области могут подходящим образом выбрать указанные факторы для достижения условий сходной строгости.
Функционально эквивалентное антитело, кодируемое полинуклеотидом, полученным указанными способами гибридизации и амплификации генов, в настоящем изобретении, как правило, имеет высокую гомологию с аминокислотными последовательностями антител согласно изобретению. Антитела согласно изобретению также включают антитела, которые функционально эквивалентны антителам согласно изобретению и имеют высокую гомологию аминокислотной последовательности с ними. Термин "высокая гомология", главным образом, относится к идентичности аминокислот по меньшей мере 50% или более, предпочтительно 75% или более, более предпочтительно 85% или более, более предпочтительно 95% или более. Гомологию полипептида можно определить с помощью алгоритма, описанного в литературе (Wilbur, W.J. и Lipman, D.J. Proc. Natl. Acad. Sci. USA (1983) 80, 726-730).
Фармацевтические композиции
В другом аспекте настоящее изобретение относится фармацевтическим композициям, содержащим в качестве активного ингредиента антитело, которое связывается с белком EREG. Кроме того, настоящее изобретение относится к ингибитору клеточной пролиферации, в частности к средству против злокачественной опухоли, содержащему в качестве активного ингредиента антитело, которое связывается с белком EREG. Ингибиторы пролиферации клеток и средства против злокачественной опухоли согласно изобретению предпочтительно вводят субъекту, страдающему злокачественной опухолью, или субъекту, который имеет вероятность развития злокачественной опухоли.
В настоящем изобретении ингибитор клеточной пролиферации, содержащий в качестве активного ингредиента антитело, которое связывается с белком EREG, также может быть описан в качестве способа подавления клеточной пролиферации, который включает стадию введения субъекту антитела, которое связывается с белком EREG, или в качестве применения антитела, которое связывается с белком EREG, для получения ингибитора клеточной пролиферации.
Более того, в настоящем изобретении средство против злокачественной опухоли, содержащее в качестве активного ингредиента антитело, которое связывается с белком EREG, также может быть описано в качестве способа профилактики или лечения злокачественной опухоли, который включает стадию введения антитела, которое связывается с белком EREG, субъекту, или в качестве применения антитела, которое связывается с белком EREG, для получения средства против злокачественной опухоли.
В настоящем изобретении выражение "содержащий в качестве активного ингредиента антитело, которое связывается с EREG" означает "содержащий антитело против EREG в качестве основного активного вещества" и не ограничивает процентное содержание антитела против EREG.
Антитело, включенное в фармацевтическую композицию согласно изобретению (например, ингибитор клеточной пролиферации и средство против злокачественной опухоли; то же в дальнейшем в настоящем описании), особенно не ограничено при условии, что оно связывается с белком EREG, и его примеры включают антитела, описанные в настоящем описании.
Фармацевтические композиции согласно изобретению можно вводить пациенту перорально или парентерально. Предпочтительно введение представляет собой парентеральное введение. Конкретно, способ введения представляет собой, например, введение посредством инъекции, трансназальное введение, транспульмонарное введение или трансдермальное введение. Примеры введения посредством инъекции включат системное и местное введение фармацевтической композиции согласно изобретению посредством внутривенной инъекции, внутримышечной инъекции, внутрибрюшинной инъекции, подкожной инъекции или подобными. Подходящий способ введения можно выбрать в зависимости от возраста пациента и симптомов. Дозировку можно выбрать, например, в диапазоне от 0,0001 мг до 1000 мг на кг массы тела при каждом введении. Альтернативно, например, дозировку для каждого пациента можно выбрать в диапазоне от 0,001 до 100000 мг/масса тела. Однако фармацевтическая композиция согласно изобретению не ограничивается указанными дозами.
Фармацевтические композиции согласно изобретению можно изготавливать общепринятыми способами (например, Remington's Pharmaceutical Science, последнее издание, Mark Publishing Company, Easton, U.S.A), а также они могут содержать фармацевтически приемлемые носители и добавки. Их примеры включают, но не ограничиваются ими, поверхностно-активные вещества, эксципиенты, красители, ароматизаторы, консерванты, стабилизаторы, буферы, суспендирующие вещества, изотонические вещества, связующие вещества, дезинтегрирующие вещества, лубриканты, обеспечивающие текучесть вещества и вкусовые добавки; и можно подходящим образом использовать другие широко используемые носители. Конкретные примеры носителей включают легкую безводную кремниевую кислоту, лактозу, кристаллическую целлюлозу, маннит, крахмал, кармеллозу кальция, кармеллозу натрия, гидроксипропилцеллюлозу, гидроксипропилметилцеллюлозу, поливинилацетальдиэтиламиноацетат, поливинилпирролидон, желатин, триглицерид жирных кислот со средней длиной цепи, упрочненное полиоксиэтиленом касторовое масло 60, сахарозу, карбоксиметилцеллюлозу, кукурузный крахмал, неорганическую соль и т.п.
Также настоящее изобретение относится к способам индукции повреждений в экспрессирующих EREG клетках и к способам подавления клеточной пролиферации посредством контактирования экспрессирующих EREG клеток с антителами, которые связываются с белком EREG.
Более конкретно, настоящее изобретение включает следующие варианты осуществления.
Способ индукции повреждения клеток в экспрессирующих EREG клетках, который включает стадию контактирования экспрессирующих белок EREG клеток со связывающим белок EREG антителом.
Способ подавления пролиферации экспрессирующих EREG клеток, который включает стадию контактирования экспрессирующих белок EREG клеток со связывающим белок EREG антителом.
Указанный выше способ, где связывающее белок EREG антитело обладает цитотоксической активностью.
Указанный выше способ, где связывающее белок EREG антитело обладает нейтрализующей активностью.
Указанный выше способ, где связывающее белок EREG антитело представляет собой антитело согласно любому из (1)-(57), представленных ниже:
(1) антитело, содержащее H-цепь, имеющую аминокислотную последовательность SEQ ID NO: 2 в качестве CDR1, аминокислотную последовательность SEQ ID NO: 4 в качестве CDR2 и аминокислотную последовательность SEQ ID NO: 6 в качестве CDR3;
(2) антитело, содержащее H-цепь согласно (1), где H-цепь имеет аминокислотную последовательность из положений 117-452 в аминокислотной последовательности SEQ ID NO: 8 в качестве CH;
(3) антитело, содержащее H-цепь согласно (1), где H-цепь имеет аминокислотную последовательность из положений 117-446 в аминокислотной последовательности SEQ ID NO: 10 в качестве CH;
(4) антитело, содержащее L-цепь, имеющую аминокислотную последовательность SEQ ID NO: 12 в качестве CDR1, аминокислотную последовательность SEQ ID NO: 14 в качестве CDR2 и аминокислотную последовательность SEQ ID NO: 16 в качестве CDR3;
(5) антитело, содержащее L-цепь согласно (4), где L-цепь имеет аминокислотную последовательность из положений 107-213 в аминокислотной последовательности SEQ ID NO: 18 в качестве CL;
(6) антитело, содержащее L-цепь согласно (4), где L-цепь имеет аминокислотную последовательность из положений 107-213 в аминокислотной последовательности SEQ ID NO: 20 в качестве CL;
(7) антитело, содержащее H-цепь согласно (1) и L-цепь согласно (4);
(8) антитело, содержащее H-цепь согласно (2) и L-цепь согласно (5);
(9) антитело, содержащее H-цепь согласно (3) и L-цепь согласно (6);
(10) антитело, содержащее H-цепь, имеющую аминокислотную последовательность SEQ ID NO: 49 в качестве CDR1, аминокислотную последовательность SEQ ID NO: 51 в качестве CDR2 и аминокислотную последовательность SEQ ID NO: 53 в качестве CDR3;
(11) антитело, содержащее H-цепь согласно (10), где H-цепь имеет CH, полученную из мышиного IgG1;
(12) антитело, содержащее H-цепь согласно (10), где H-цепь имеет аминокислотную последовательность из положений 117-446 в аминокислотной последовательности SEQ ID NO: 10 в качестве CH;
(13) антитело, содержащее L-цепь, имеющую аминокислотную последовательность SEQ ID NO: 55 в качестве CDR1, аминокислотную последовательность SEQ ID NO: 57 в качестве CDR2 и аминокислотную последовательность SEQ ID NO: 59 в качестве CDR3;
(14) антитело, содержащее L-цепь согласно (13), где L-цепь имеет CL, полученную из мышиной κ-цепи;
(15) антитело, содержащее L-цепь согласно (13), где L-цепь имеет аминокислотную последовательность из положений 107-213 в аминокислотной последовательности SEQ ID NO: 20 в качестве CL;
(16) антитело, содержащее H-цепь согласно (10) и L-цепь согласно (13);
(17) антитело, содержащее H-цепь согласно (11) и L-цепь согласно (14);
(18) антитело, содержащее H-цепь согласно (12) и L-цепь согласно (15);
(19) антитело, содержащее H-цепь, имеющую аминокислотную последовательность SEQ ID NO: 61 в качестве CDR1, аминокислотную последовательность SEQ ID NO: 63 в качестве CDR2 и аминокислотную последовательность SEQ ID NO: 65 в качестве CDR3;
(20) антитело, содержащее H-цепь согласно (19), где H-цепь имеет CH, полученную из мышиного IgG1;
(21) антитело, содержащее H-цепь согласно (19), где H-цепь имеет аминокислотную последовательность из положений 117-446 в аминокислотной последовательности SEQ ID NO: 10 в качестве CH;
(22) антитело, содержащее L-цепь, имеющую аминокислотную последовательность SEQ ID NO: 67 в качестве CDR1, аминокислотную последовательность SEQ ID NO: 69 в качестве CDR2 и аминокислотную последовательность SEQ ID NO: 71 в качестве CDR3;
(23) антитело, содержащее L-цепь согласно (22), где L-цепь имеет CL, полученную из мышиной κ-цепи;
(24) антитело, содержащее L-цепь согласно (22), где L-цепь имеет аминокислотную последовательность из положений 107-213 в аминокислотной последовательности SEQ ID NO: 20 в качестве CL;
(25) антитело, содержащее H-цепь согласно (19) и L-цепь согласно (22);
(26) антитело, содержащее H-цепь согласно (20) и L-цепь согласно (23);
(27) антитело, содержащее H-цепь согласно (21) и L-цепь согласно (24);
(28) антитело, содержащее H-цепь, имеющую аминокислотную последовательность SEQ ID NO: 73 в качестве CDR1, аминокислотную последовательность SEQ ID NO: 75 в качестве CDR2 и аминокислотную последовательность SEQ ID NO: 77 в качестве CDR3;
(29) антитело, содержащее H-цепь согласно (28), где H-цепь имеет CH, полученную из мышиного IgG1;
(30) антитело, содержащее H-цепь согласно (28), где H-цепь имеет аминокислотную последовательность из положений 117-446 в аминокислотной последовательности SEQ ID NO: 10 в качестве CH;
(31) антитело, содержащее L-цепь, имеющую аминокислотную последовательность SEQ ID NO: 79 в качестве CDR1, аминокислотную последовательность SEQ ID NO: 81 в качестве CDR2 и аминокислотную последовательность SEQ ID NO: 83 в качестве CDR3;
(32) антитело, содержащее L-цепь согласно (31), где L-цепь имеет CL, полученную из мышиной κ-цепи;
(33) антитело, содержащее L-цепь согласно (31), где L-цепь имеет аминокислотную последовательность из положений 107-213 в аминокислотной последовательности SEQ ID NO: 20 в качестве CL;
(34) антитело, содержащее H-цепь согласно (28) и L-цепь согласно (31);
(35) антитело, содержащее H-цепь согласно (29) и L-цепь согласно (32);
(36) антитело, содержащее H-цепь согласно (30) и L-цепь согласно (33);
(37) антитело, содержащее H-цепь, имеющую аминокислотную последовательность SEQ ID NO: 85 в качестве CDR1, аминокислотную последовательность SEQ ID NO: 87 в качестве CDR2 и аминокислотную последовательность SEQ ID NO: 89 в качестве CDR3;
(38) антитело, содержащее H-цепь согласно (37), где H-цепь имеет CH, полученную из мышиного IgG1;
(39) антитело, содержащее H-цепь согласно (37), где H-цепь имеет аминокислотную последовательность из положений 117-446 в аминокислотной последовательности SEQ ID NO: 10 в качестве CH;
(40) антитело, содержащее L-цепь, имеющую аминокислотную последовательность SEQ ID NO: 91 в качестве CDR1, аминокислотную последовательность SEQ ID NO: 93 в качестве CDR2 и аминокислотную последовательность SEQ ID NO: 95 в качестве CDR3;
(41) антитело, содержащее L-цепь согласно (40), где L-цепь имеет CL, полученную из мышиной κ-цепи;
(42) антитело, содержащее L-цепь согласно (40), где L-цепь имеет аминокислотную последовательность из положений 107-213 в аминокислотной последовательности SEQ ID NO: 20 в качестве CL;
(43) антитело, содержащее H-цепь согласно (37) и L-цепь согласно (40);
(44) антитело, содержащее H-цепь согласно (38) и L-цепь согласно (41);
(45) антитело, содержащее H-цепь согласно (39) и L-цепь согласно (42);
(46) антитело, содержащее H-цепь, имеющую аминокислотную последовательность SEQ ID NO: 97 в качестве CDR1, аминокислотную последовательность SEQ ID NO: 99 в качестве CDR2 и аминокислотную последовательность SEQ ID NO: 101 в качестве CDR3;
(47) антитело, содержащее H-цепь согласно (46), где H-цепь имеет CH, полученную из мышиного IgG1;
(48) антитело, содержащее H-цепь согласно (46), где H-цепь имеет аминокислотную последовательность из положений 117-446 в аминокислотной последовательности SEQ ID NO: 10 в качестве CH;
(49) антитело, содержащее L-цепь, имеющую аминокислотную последовательность SEQ ID NO: 103 в качестве CDR1, аминокислотную последовательность SEQ ID NO: 105 в качестве CDR2 и аминокислотную последовательность SEQ ID NO: 107 в качестве CDR3;
(50) антитело, содержащее L-цепь согласно (49), где L-цепь имеет CL, полученную из мышиной κ-цепи;
(51) антитело, содержащее L-цепь согласно (49), где L-цепь имеет аминокислотную последовательность из положений 107-213 в аминокислотной последовательности SEQ ID NO: 20 в качестве CL;
(52) антитело, содержащее H-цепь согласно (46) и L-цепь согласно (49);
(53) антитело, содержащее H-цепь согласно (47) и L-цепь согласно (50);
(54) антитело, содержащее H-цепь согласно (48) и L-цепь согласно (51);
(55) антитело, содержащее одну или несколько аминокислотных замен, делеций, добавлений и/или инсерций в антителе согласно любому из (1)-(54), которое обладает активностью, эквивалентной активности антитела согласно любому из (1)-(54);
(56) антитело, которое связывается с тем же эпитопом белка EREG, что и антитело согласно любому из (1)-(55); и
(57) мини-антитело антитела согласно любому из (1)-(56).
Указанный выше способ, где экспрессирующие белок EREG клетки представляют собой злокачественные клетки.
Как описано выше, указанные выше антитела согласно (1)-(57) включают не только одновалентные антитела, но также поливалентные антитела с двумя или более валентностями. Поливалентные антитела согласно изобретению включают поливалентные антитела, все антигенсвязывающие участки которых являются одинаковыми, и поливалентные антитела, антигенсвязывающие участки которых частично или полностью отличаются.
Связывающие белок EREG антитела описаны выше в качестве связывающих белок EREG антител, включенных в ингибиторы пролиферации клеток согласно изобретению. Клетки, которые связываются антителами против EREG, особенно не ограничены, при условии, что клетки представляют собой экспрессирующие EREG клетки. Предпочтительными экспрессирующими EREG клетками согласно изобретению являются злокачественные клетки. Более предпочтительно клетки представляют собой клетки рака толстого кишечника, аденокарциномы легкого, рака поджелудочной железы, рака желудка и рака почки. Способы согласно изобретению можно применять как к первичным, так и к метастатическим очагам данных злокачественных опухолей. Более предпочтительными злокачественными клетками являются клетки первичного рака толстого кишечника, метастазирующего рака толстого кишечника и рака поджелудочной железы.
В настоящем изобретении "контактирование" проводят, например, добавлением антитела в культуральный раствор экспрессирующих EREG клеток, культивируемых в тестируемой пробирке. В этом случае антитело можно добавлять в форме, например, раствора или твердого вещества, полученного сушкой вымораживанием или подобным образом. При добавлении антитела в виде водного раствора используемый водный раствор может содержать только антитело или раствор может включать, например, указанные выше поверхностно-активные вещества, эксципиенты, красители, ароматизаторы, консерванты, стабилизаторы, буферы, суспендирующие вещества, изотонические вещества, связующие вещества, дезинтегрирующие вещества, лубриканты, обеспечивающие текучесть вещества или вкусовые добавки. Концентрация для добавления особенно не ограничена, однако конечная концентрация в культуре, которую можно пригодным образом использовать, предпочтительно находится в диапазоне от 1 пг/мл до 1 г/мл, более предпочтительно от 1 нг/мл до 1 мг/мл и более предпочтительно от 1 мкг/мл до 1 мг/мл.
Более того, в другом варианте осуществления "контактирование" в настоящем изобретении проводят посредством введения не относящемуся к человеку животному, в организм которого трансплантировали экспрессирующую EREG клетку, или животному, обладающему злокачественными клетками, эндогенно экспрессирующими EREG. Способ введения может представлять собой пероральное или парентеральное введение. Способ введения, в частности, предпочтительно представляет собой парентеральное введение, и, конкретно, способ введения представляет собой, например, введение посредством инъекции, трансназальное введение, транспульмонарное введение или трансдермальное введение. Примеры введения посредством инъекции включают системное и местное введение фармацевтических композиций, ингибиторов пролиферации клеток и средств против злокачественной опухоли согласно изобретению посредством внутривенной инъекции, внутримышечной инъекции, внутрибрюшинной инъекции, подкожной инъекции или подобных. Пригодный способ введения может быть выбран в зависимости от возраста тестируемого животного и симптомов. При введении в виде водного раствора используемый водный раствор может содержать только антитело, или раствор может включать, например, указанные выше поверхностно-активные вещества, эксципиенты, красители, ароматизаторы, консерванты, стабилизаторы, буферы, суспендирующие вещества, изотонические вещества, связующие вещества, дезинтегрирующие вещества, лубриканты, обеспечивающие текучесть вещества или вкусовые добавки. Дозировку можно выбрать, например, в диапазоне от 0,0001 мг до 1000 мг на кг массы тела при каждом введении. Альтернативно, например, дозировку для каждого пациента можно выбрать в диапазоне от 0,001 до 100000 мг/масса тела. Однако доза антитела согласно изобретению не ограничивается указанными дозами.
Следующий способ подходящим образом используют в качестве способа оценки или определения повреждения клеток, индуцируемого контактированием экспрессирующих EREG клеток с антителом против EREG. Примеры способа оценки или определения цитотоксической активности в тестируемой пробирке включают способы измерения указанной выше активности антителозависимой клеточно-опосредуемой цитотоксичности (ADCC), активности комплементзависимой цитотоксичности (CDC) и т.п. Наличие или отсутствие у антитела против EREG активности ADCC или активности CDC можно определить известными способами (например, Current protocols in Immunology, Chapter 7. Immunologic studies in humans, Editor, John E. Coligan et al., John Wiley & Sons, Inc., (1993) и т.п.). Для определения активности, в качестве контрольного антитела можно использовать связывающее антитело, обладающее тем же изотипом, что и антитело против EREG, не обладающее какой-либо активностью повреждения клеток, таким же образом, как и антитело против EREG, и наличие активности можно определить, если антитело против EREG демонстрирует более сильную цитотоксическую активность, чем контрольное антитело.
Изотип антитела определяют по последовательности его константной области H-цепи в аминокислотной последовательности антитела. Изотип антитела в конечном итоге определяется in vivo переключением классов, которое происходит при генетических рекомбинациях в хромосомах, происходящих в ходе созревания антителопродуцирующих B-клеток. Различие в изотипе отражается в различии в физиологических и патологических функциях антител. Конкретно, например, известно, что в дополнение к уровню экспрессии антигена на силу цитотоксической активности влияет изотип антитела. Таким образом, при измерении описанной выше активности повреждения клеток в качестве контроля предпочтительно используют антитело того же изотипа, что и тестируемое антитело.
Для оценки или определения активности повреждения клеток in vivo, например, экспрессирующие EREG злокачественные клетки внутрикожно или подкожно трансплантируют не относящемуся к человеку тестируемому животному и затем внутривенно или внутрибрюшинно вводят тестируемое антитело каждые сутки или с интервалом несколько суток, начиная со дня трансплантации или на следующий день. Цитотоксичность можно определить посредством каждодневного измерения размера опухоли. Аналогично оценке в тестируемой пробирке цитотоксичность можно определить посредством введения контрольного антитела, имеющего тот же изотип, и наблюдением того, что размер опухоли в группе введения антитела против EREG значительно меньше, чем размер опухоли в группе введения контрольного антитела. При использовании мыши в качестве не относящегося к человеку тестируемого животного пригодным является использование мышей nude (nu/nu), тимус которых был сделан генетически дефектным, так что их функция T-лимфоцитов утрачена. Применение такой мыши может устранить участие T-лимфоцитов в тестах на животных при оценке или определении цитотоксичности вводимого антитела.
Способ аналогично используемому для определения нейтрализующей активности, указанной выше, можно подходящим образом использовать в качестве способа оценки или определения опосредуемого контактом ингибиторного эффекта антитела против EREG на пролиферацию экспрессирующих EREG клеток.
В качестве способа оценки или определения активности ингибирования клеточной пролиферации in vivo можно подходящим образом использовать способ аналогично описанному выше для оценки или определения цитотоксичности in vivo.
Способы скрининга
Авторы настоящего изобретения показали, что EREG, секретируемый клетками, индуцирует пролиферацию клеток, растущих зависимым от EREG образом. Более того, также было подтверждено, что экспрессия EREG специфично усиливается в злокачественных клетках. Таким образом, лечение злокачественной опухоли можно проводить, подавляя экспрессию EREG в экспрессирующих EREG клетках. Более конкретно, в настоящем изобретении показано, что соединения, которые подавляют экспрессию EREG в злокачественных клетках, пригодны в качестве соединений-кандидатов в качестве лекарственных средств против злокачественной опухоли. Исходя из этих открытий, настоящее изобретение относится к способам скрининга соединений-кандидатов в качестве лекарственных средств, которые включают в себя стадии:
(1) контактирования экспрессирующих EREG клеток с тестируемым соединением;
(2) измерения уровня экспрессии EREG в экспрессирующих EREG клетках и
(3) селекции соединения, которое снижает экспрессию EREG, по сравнению с контролем в качестве лекарственного средства-кандидата против злокачественной опухоли.
В настоящем изобретении в качестве экспрессирующих EREG клеток, например, можно использовать экспрессирующие EREG злокачественные клетки. Конкретно, например, для способов скрининга согласно изобретению можно использовать любую злокачественную клеточную линию, демонстрирующую усиленную экспрессию EREG, представленную на фиг.2. Более конкретно, клетки, которые можно использовать для способов скрининга согласно изобретению, включают представленные ниже злокачественные клеточные линии. Все примеры клеточных линий, приведенные ниже, могут быть получены из банка клеток. Условия культивирования для таких клеточных линий уже установлены. Более конкретно, клетки можно культивировать, например, в условиях, представленных в таблице 2.
MKN45
2M
2MD3
DLD1
hCT116
LOVO
Paca2
PK-1
Caki2
H157
H1648
H2009
H2347
H522
Приведенные клеточные линии культивируют в присутствии тестируемого соединения и определяют уровень экспрессии EREG в клеточных линиях. Уровень экспрессии EREG можно оценивать посредством определения количества внутриклеточной мРНК или количества белка EREG, секретируемого внутри клетки, на клеточную поверхность или во внешнюю среду клетки. В качестве способа определения мРНК EREG или белка EREG можно использовать любой способ, описанный в настоящем описании. Авторы настоящего изобретения подтвердили эффект индукции клеточной пролиферации секретируемого EREG в клетках, которые пролиферируют зависимым от EREG образом. Таким образом, предпочтительным вариантом осуществления настоящего изобретения является способ скрининга соединений-кандидатов в качестве средств против злокачественной опухоли, в которых в качестве показателя используется количество внеклеточно секретируемого EREG.
В настоящем изобретении в качестве представляющего интерес соединения-кандидата выбирают тестируемое соединение, которое снижает экспрессию EREG по сравнению с контролем. В качестве контроля в настоящем изобретении, например, можно использовать ту же клеточную линию, культивированную в отсутствие тестируемого соединения. Альтернативно, для контроля можно использовать клеточную линию, культивируемую в присутствии соединения, эффект которого на экспрессию EREG уже известен. С использованием такого контроля можно выбрать соединение, обладающее большим эффектом, чем определенное соединение.
В настоящем изобретении можно проводить скрининг лекарственных средств против злокачественной опухоли с использованием нейтрализующего эффекта тестируемого соединения против EREG в качестве показателя. Более конкретно, настоящее изобретение относится к способам скрининга соединений-кандидатов в качестве лекарственных средств против злокачественной опухоли, которые включают в себя стадии:
(1) культивирования клеток, которые пролиферируют зависимым от EREG образом, в присутствии EREG и тестируемого соединения;
(2) измерения уровня клеточной пролиферации и
(3) выбора тестируемого соединения, которое подавляет клеточную пролиферацию по сравнению с контролем, в качестве лекарственного средства-кандидата против злокачественной опухоли.
В настоящем изобретении "клетки, которые пролиферируют зависимым от EREG образам", относятся к клеткам, демонстрирующим дозозависимую клеточную пролиферацию, опосредуемую EREG. Например, в разделе "Примеры", приведенном ниже, клеточная линия рака поджелудочной железы AsPC1 демонстрирует дозозависимую клеточную пролиферацию, опосредуемую EREG. Более того, в соответствии с открытиями авторов настоящего изобретения зависимость от EREG можно обеспечить посредством экспрессии химерного рецептора рецептор EGF/рецептор G-CSF. Более конкретно, клетки мыши, трансформированные ДНК, кодирующей химерный рецептор, содержащий внеклеточный домен рецептора EGF и внутриклеточный домен рецептора G-CSF, проявляют зависимую от дозы EREG клеточную пролиферацию. Таким образом, клетки, в которых зависимость от EGFR искусственно индуцирована таким образом, можно использовать для способов скрининга согласно изобретению.
Затем измеряют уровень пролиферации клеток, пролиферирующих зависимым от EREG образом, культивируемых в присутствии EREG и тестируемого соединения. Клеточную пролиферацию можно определить любым способом. Например, количество жизнеспособных клеток можно сравнить с использованием коммерчески доступного реагента для подсчета жизнеспособных клеток, как описано в разделе "Примеры". Альтернативно, также коммерчески доступны системы для эффективного крупномасштабного скрининга. Например, на практике применяют систему, которая может оценивать количество жизнеспособных клеток в каждой лунке с использованием многолуночного планшета. На основе результата сравнения количества жизнеспособных клеток тестируемые соединения, которые подавляют повышение количества клеток по сравнению с контролем, можно выбрать в качестве соединений-кандидатов для лекарственных средств против злокачественной опухоли.
В способах скрининга согласно изобретению, например, в качестве контроля можно использовать ту же клеточную линию, культивированную в отсутствие тестируемого соединения. Альтернативно, в качестве контроля можно использовать клеточную линию, культивированную в присутствии соединения, которое обладает очевидным эффектом на действие индукции клеточной пролиферации, зависящее от секреции EREG. С использованием такого контроля можно выбрать соединение, которое обладает большим эффектом, чем определенное соединение.
Соединения-кандидаты, выбранные способами скрининга согласно изобретению, пригодны в качестве соединений-кандидатов для противоопухолевого лекарственного средства, которое действует, подавляя функцию EREG. В настоящем изобретении терапевтический эффект антител против EREG был подтвержден. Таким образом, можно ожидать, что соединения, выбранные способами скрининга согласно изобретению, обладают эффектами, сходными с антителами. Если необходимо, соединения-кандидаты, выбранные способами скрининга согласно изобретению, далее оценивают в отношении их эффекта на другие злокачественные клеточные линии или клетки первичной культуры, а также в отношении такого их эффекта, как токсичность против нормальных клеток. Посредством такой оценки можно выбрать соединения, которые пригодны в качестве лекарственных средств против злокачественной опухоли. Уже разработан ряд способов оценки лекарственных средств против злокачественной опухоли.
В способах скрининга согласно изобретению в качестве тестируемых соединений можно использовать различные встречающиеся в природе и искусственно синтезированные соединения. Например, в качестве библиотек тестируемых соединений согласно изобретению предпочтительными являются библиотеки белков и библиотеки антител. Альтернативно, в качестве тестируемых соединений можно использовать фаговые библиотеки, презентирующие белки или антитела. Более того, в качестве тестируемых соединений можно использовать библиотеки искусственно синтезированных соединений, такие как комбинаторные библиотеки.
Все ссылки предшествующего уровня техники, цитированные в настоящем описании, включены в настоящее описании посредством ссылок.
Примеры
Ниже настоящее изобретение описано конкретно с помощью примеров, однако его не следует истолковывать, как ограниченное ими.
[Пример 1]
Анализ экспрессии гена EREG человека в различных типах злокачественных опухолей
1-1. Анализ экспрессии гена EREG человека с помощью GeneChip
Для поиска гена, экспрессия которого специфично усиливается в злокачественных тканях, таких как ткани рака толстого кишечника и рака поджелудочной железы, был проведен обширный анализ экспрессии генов в нормальных тканях, злокачественных тканях и злокачественных клеточных линиях с использованием GeneChip U-133A (изготовленного Affymetrix).
Тотальную РНК из нормальных тканей, злокачественных тканей и злокачественных клеточных линий, представленных в таблицах 1 и 2, получали общепринятыми способами с использованием ISOGEN (изготовленного Nippon Gene). Анализ экспрессии генов проводили, используя 10 мкг каждой из указанных тотальных РНК и подвергая их анализу транскриптов на GeneChip U-133A (изготовленном Affymetrix). Данный способ осуществляли в соответствии с техническим описанием анализа экспрессии (изготовленным Affymetrix). Для поиска генов, экспрессия которых усиливается в злокачественных тканях или злокачественных клетках, среднее значение показателей экспрессии для всех генов нормализовали к 100.
Было обнаружено, что экспрессия мРНК EREG человека (ID образца: 205767_at HG-U133A) усилена в тканях первичного рака толстого кишечника, метастазирующего рака толстого кишечника, аденокарциномы легкого и рака поджелудочной железы, и клеточных линиях рака толстого кишечника (CACO2, DLD1, HCT116 и LOVO), клеточных линиях рака желудка (2M и 2MD3), клеточной линии рака печени (Alexander), клеточных линиях рака поджелудочной железы (Capan1 и Paca2), клеточных линиях рака почки (Caki1 и Caki2) и клеточных линиях первичного рака толстого кишечника, метастазирующего рака толстого кишечника, аденокарциномы легкого, рака поджелудочной железы, рака желудка и рака почки (H157, H1648, H2009, H23 и H2347), в то время как в исследованных нормальных тканях не было обнаружено заметной экспрессии мРНК EREG человека (фиг.1 и 2).
Из указанного выше становится очевидно, что, несмотря на то, что экспрессия гена EREG человека (ID образца: 205767_at HG-U133A) является очень низкой в нормальных тканях, экспрессия гена EREG человека усиливается в широком множестве злокачественных опухолей, включая первичный рак толстого кишечника, метастазирующий рак толстого кишечника, аденокарциному легкого, рак поджелудочной железы, рак желудка, рак почки, рак толстого кишечника, рак поджелудочной железы, рак желудка и рак почки.
Ткани, используемые для анализа экспрессии гена EREG
Клеточные линии и условия культивирования, используемые для анализа экспрессии гена EREG
[Пример 2]
Получение моноклонального антитела против EREG
2-1. Получение клеток, экспрессирующих полноразмерный EREG человека
Полноразмерную кДНК EREG человека (NM_001432), содержащую последовательность, представленную в SEQ ID NO: 21, выделяли общепринятыми способами. Фрагмент гена клонировали в экспрессирующий вектор (pMCN) для клеток млекопитающих и полученный вектор назвали hEREG/pMCN. pMCN обеспечивает индуцируемую экспрессию чужеродного гена под контролем промотора CMV мыши (регистрационный No. U68299) и представляет собой вектор, в который встроен ген устойчивости к неомицину. Клеточную линию CHO, которая стабильно экспрессирует полноразмерный EREG человека, получали введением hEREG/pMCN в клеточную линию CHO DG44 (Invitrogen) электропорацией и селекцией клеток в присутствии 500 мкг/мл генетицина. Полученную клеточную линию назвали EREG_DG.
2-2. Получение растворимого слитого белка EREG человека/Fc IgG2a мыши (sEREG_mFc)
Растворимый слитый белок EREG человека/Fc IgG2a мыши (в дальнейшем в настоящем описании sEREG_mFc) получали в качестве иммунизирующего антигена для продукции антитела против EREG.
sEREG_mFc конструировали слиянием внеклеточного домена EREG человека (аминокислоты 7-122) с константной областью IgG2a мыши через последовательность распознавания CpoI в шарнирной области и клонированием в вектор pMCDN, который получали встраиванием гена DHFR в экспрессирующий вектор pMCN. Сконструированный вектор назвали sEREG-mFc/pMCDN. Последовательность, представленная в SEQ ID NO: 33, демонстрирует нуклеотидную последовательность гена sEREG-mFc, и последовательность, представленная в SEQ ID NO: 34, демонстрирует аминокислотную последовательность sEREG-mFc. Экспрессирующую sEREG-mFc клеточную линию CHO получали введением sEREG-mFc/pMCDN в клеточную линию CHO DG44 (Invitrogen) электропорацией и селекцией клеток в присутствии 500 мкг/мл генетицина.
Затем из культурального супернатанта очищали sEREG-mFc. Культуральный супернатант наносили на колонку HiTrap Protein G HP (Amersham, CAT# 17-0404-01), не связавшиеся материалы смывали буфером для связывания (20 мМ фосфат натрия (pH 7,0)) и представляющий интерес sEREG-mFc элюировали с помощью буфера для элюирования (0,1 M глицин-HCl (pH 2,7)). Элюат сразу нейтрализовали сбором в пробирку, содержащую нейтрализующий буфер (1M Tris-HCl (pH 9,0)). Затем элюированный sEREG-mFc разделяли гель-фильтрацией с использованием Superdex 200 HR 10/30 (Amersham) для того, чтобы растворитель данного раствора заменить PBS. Очищенный белок количественно определяли с использованием анализа белка DC (BIO-RAD), в котором концентрацию белка пересчитывали с использованием включенного бычьего IgG в качестве стандарта.
2-3. Получение моноклонального антитела против EREG
В качестве животных для иммунизации использовали мышей Balb/c и мышей MRL/MpJ-Tnfrsf6<lpr>/Crlj (Charles River Japan). Иммунизацию начинали на с 5 по 8 неделю. Для первой иммунизации sEREG-mFc эмульгировали с использованием полного адъюванта Фрейнда (FCA, Beckton Dickinson) и вводили подкожно (100 мкг/особь). Через две недели sEREG-mFc эмульгировали с использованием неполного адъюванта Фрейнда (FIA, Beckton Dickinson) и вводили подкожно (50 мкг/особь). После этого два или три раза проводили повторные иммунизации с интервалами, составляющими одну неделю, и для конечной иммунизации sEREG-mFc разводили PBS и затем вводили в хвостовую вену (50 мкг/особь).
Через четверо суток после последней иммунизации клетки селезенки извлекали и смешивали с клетками миеломы мыши P3-X63Ag8U1 (P3U1, приобретенными от ATCC) в соотношении 2:1 и посредством пошагового добавления PEG 1500 (Roche Diagnostics) проводили слияние клеток. Для разведения PEG 1500 осторожно добавляли среду RPMI1640 (GIBCO BRL) и затем PEG 1500 удаляли центрифугированием. Затем клетки ресуспендировали в RPMI1640, содержащей 10% FBS, 1× добавку для среды HAT (SIGMA) и 0,5× добавку для клонирования гибридом BM-Condimed H1 (Roche Diagnostics) (в дальнейшем в настоящем описании обозначаемую как среда HAT) и высевали в 96-луночный культуральный планшет в количестве 200 мкл/лунка. Скрининг проводили посредством оценки активности связывания с EREG_DG человека с использованием проточного цитометра. Клоны, демонстрирующие положительную активность связывания, полученные посредством данного анализа, подвергали моноклональному клонированию способом лимитирующих разведений.
Более того, моноклональные антитела получали иммунизацией мышей способом инокуляции ДНК с использованием системы генной пушки Helios (Bio-Rad). На трубку кассеты наносили частицы из золота-ДНК (экспрессирующий вектор полноразмерного EREG человека) в соответствии с руководством по эксплуатации генной пушки Helios. 50 мг 1,0-мкм частиц золота взвешивали, суспендировали и перемешивали в 0,1 мл 0,05 M раствора спермидина. К ним добавляли 0,1 мл раствора 1 мг/мл плазмиды и затем встряхивали. После этого добавляли 0,1 мл 1 M CaCl2 и его оставляли стоять в течение десяти минут. После кратковременного центрифугирования супернатант удаляли. Осадок суспендировали в этаноле и затем центрифугировали. После повторения стадии дегидратации этанолом осадок в конечном итоге суспендировали в 6 мл раствора 0,05 мг/мл поливинилпирролидона-этанол. Раствор помещали в трубку для нанесения и трубку покрывали, сушили и разрезали на сегменты длиной 0,5 дюймов с использованием устройства для разрезания трубок.
Иммунизацию посредством ДНК проводили на мышах в возрасте от четырех-пяти недель MRL/MpJ-Tnfrsf6<lpr>/Crlj (давление гелия приблизительно 200 фунт/кв. дюйм) с интервалом от одного до трех раз в неделю и в ходе этого периода периодически проводили мониторинг титра антитела против EREG в сыворотке. Клеточную линию, искусственно экспрессирующую EREG (5×106 клеток/особь), вводили внутрибрюшинно или в хвостовую вену особей, у которых было подтверждено наличие повышенного титра антитела в сыворотке. После выдерживания в течение от двух до трех суток селезенку извлекали и выделяли мононуклеарные клетки, содержащие антителопродуцирующие клетки. Слияние клеток и клонирование проводили аналогичным способом, как в случае иммунизации посредством sEREG-mFc.
Антитела очищали из культурального супернатанта гибридом, культивированных в среде HAT, дополненной сывороткой. Сыворотка, добавляемая к среде, представляла собой FBS (с крайне низким содержанием IgG) (GIBCO BRL). Антитела очищали из культурального супернатанта с использованием HiTrap Protein G HP аналогичным способом, как при очистке sEREG_mFc, описанной в примере 2-2. Растворитель фракций элюата HiTrap Protein G HP заменяли PBS с использованием колонки PD-10 (Amersham) и затем хранили при 4°C. Очищенные антитела количественно определяли с использованием анализа белка DC (BIO-RAD), в котором концентрацию белка вычисляли с использованием включенного бычьего IgG в качестве стандарта.
В результате были успешно выделены клоны B2#30 (IgG1, каппа), B3#18 (IgG2b, каппа), B3#41 (IgG1, каппа) и т.п., которые представлены в таблице 3.
Подтипы антитела и перекрестная видовая реактивность выделенных моноклональных антител мыши против EREG и эпитопные участки, распознаваемые ими
NA - не анализировали
2-4. Оценка активности связывания посредством проточной цитометрии
Проточную цитометрию использовали для оценки связывания антител, полученных указанными выше способами, с EREG_DG. Антитела против EREG, разведенные до пригодной концентрации, добавляли к EREG_DG, суспендированным в буфере FACS (1% FBS/PBS) с плотностью 1×105 клеток/мл, и затем смеси давали возможность реагировать на льду в течение 60 минут. После промывания клеток один раз буфером FACS добавляли меченное FITC антитело против IgG мыши и смеси давали возможность реагировать на льду в течение 30 минут. После завершения реакции супернатант удаляли центрифугированием. Затем клетки ресуспендировали в 150 мкл буфера FACS и затем подвергали анализу посредством проточной цитометрии.
В качестве проточного цитометра использовали FACS Calibur (Becton Dickinson). Популяцию живых клеток выбирали посредством получения гистограммы прямого рассеяния и бокового рассеяния. В дополнение к активности связывания EREG_DG также оценивали активность связывания с клеточной линией рака толстого кишечника человека DLD1. Как подтверждается результатами, представленными на фиг.2, экспрессия гена EREG человека сходным образом усиливалась в клеточной линии рака толстого кишечника DLD1, как в клетках EREG_DG.
Как представлено на фиг.3, моноклональные антитела против EREG (B2#30, B3#18 и B3#41), продуцированные гибридомами, связывались сильно с EREG_DG, но не с родительскими клетками DG44. Эти результаты показали, что эти моноклональные антитела специфично связываются с EREG, присутствующим на клеточной мембране.
Более того, было подтверждено, что описанные выше моноклональные антитела против EREG согласно изобретению специфично связываются с DLD1, которые обладают усиленной экспрессией гена EREG человека (фиг.4), а также с EREG_DG.
Описана усиленная экспрессия гена EREG человека при раке мочевого пузыря (Thogersen, V. et al., Cancer Res. 61:6227-6233, 2001), раке поджелудочной железы (Zhu. Z., et al., Biochem. Biophys. Res. Commun. 273: 1019-1024, 2000), раке предстательной железы (Torring, N., et al., Anticancer Res. 20: 91-95, 2000), раке толстого кишечника (Baba, I., et al., Cancer Res. 60: 6886-6889, 2000) и т.п. Однако впервые было показано, что усиленная экспрессия гена EREG человека коррелирует с усиленной экспрессией белка EREG человека.
[Пример 3]
Определение активности комплементзависимой цитотоксичности (CDC) и активности антителозависимой клеточной цитотоксичности (ADCC), индуцируемой моноклональными антителами против EREG
3-1. Определение активности CDC, индуцируемой моноклональными антителами против EREG
В качестве клеток-мишеней использовали клеточную линю рака толстого кишечника человека DLD1. Для поддержания клеток DLD1 использовали среду RPMI1640 (GIBCO BRL), содержащую 10% FBS. 5×105 DLD1 клеток собирали центрифугированием (1000 об/мин, пять минут, 4°C). Клеточный осадок суспендировали приблизительно в 200 мкл среды, содержащей 3,7 MBq хрома-51 (код No. CJS4, Amersham Pharmacia Biotech), и затем культивировали в инкубаторе с 5% диоксидом углерода в течение одного часа при 37°C. Клетки промывали три раза средой, затем доводили до плотности 1×104 клеток/мл в среде и затем распределяли в 96-луночный планшет с плоским дном в количестве 100 мкл/лунка.
Затем добавляли моноклональное антитело против (B3#18_IgG2b) и контрольное антитело IgG2a мыши (каталожный No. 553453, BD Biosciences Pharmingen), разведенные в среде, в количестве 50 мл на лунку. Конечную концентрацию антитела доводили до 10 мкг/мл. Затем добавляли комплемент детенышей кролика (каталожный No. CL3441, Cederlane), разведенный в 5 раз в среде в количестве 50 мл/лунка, и затем планшет оставляли инкубаторе с 5% диоксидом углерода на 1,5 часа при 37°C. После этого планшет центрифугировали (1000 об/мин в течение пяти минут при 4°C), 100 мл супернатанта собирали из каждой лунки планшета и измеряли радиоактивность собранного супернатанта с использованием гамма-счетчика (1480 WIZARD 3", Wallac). Удельный уровень высвобождения определяли на основе следующего уравнения:
Удельный уровень высвобождения хрома (%)=(A-C)×100/(B-C),
где
A представляет собой радиоактивность (cpm) в каждой лунке;
B представляет собой среднее значение радиоактивности (cpm) в лунках, где к 100 мкл клеток добавляли 100 мкл раствора 2% NP-40 (Nonidet P-40, код No. 252-23, Nacalai Tesque); и
C представляет собой среднее значение радиоактивности (cpm) в лунках, где к 100 мкл клеток добавляли 100 мкл среды. Измерения проводили в двух повторах и среднее значение и стандартное отклонение вычисляли для удельного уровня высвобождения хрома для супернатанта, полученного из каждой лунки.
Было подтверждено, что моноклональные антитела против EREG, используемые в эксперименте, обладают активностью CDC, как представлено на фиг.5. С другой стороны, контрольное антитело IgG2a мыши не демонстрировало активности CDC в той же концентрации.
3-2. Определение активности ADCC, индуцируемой моноклональными антителами против EREG
Как и в случае определения активности CDC, для определения активности ADCC использовали клеточную линию рака толстого кишечника человека DLD1. Конкретно, клетки культивировали в 96-луночном планшете с плоским дном и затем инкубировали с хромом-51. Каждую лунку промывали средой RPMI1640 (Invitrogen), содержащей 10% FBS, и затем добавляли 100 мкл среды. Затем в лунки планшета добавляли 50 мкл каждого моноклонального антитела против EREG (B2#30, B3#18 и B3#41), разведенного в среде. Антитело добавляли до конечной концентрации 1 мкг/мл. Затем в каждую лунку добавляли 50 мкл раствора с эффекторными клетками (1×107 клеток/мл), затем планшет оставляли в инкубаторе с 5% диоксидом углерода на четыре часа при 37°C и затем определяли удельный уровень высвобождения хрома. Эффекторные клетки получали культивированием клеток селезенки мыши Balb/c (Charles River Japan) в среде, содержащей 50 нг/мл рекомбинантного интерлейкина-2 человека (каталожный No. 200-02, Peprotech) в течение пяти суток, или культивированием клеток костного мозга из той же мыши в среде, содержащей 50 нг/мл рекомбинантного интерлейкина-2 человека и 10 нг/мл рекомбинантного GM-CSF мыши (каталожный No. 315-03, Peprotech) в течение шести суток.
Результаты показали, что все из моноклональных антител против EREG, использованных в эксперименте, индуцировали активность ADCC против DLD1 (фиг.6).
[Пример 4]
Определение нейтрализующей активности моноклональных антител против EREG в отношении клеточной пролиферации, усиленной посредством EREG человека
4-1. Получение клеточной линии Ba/F3, которая экспрессирует химерный рецептор EGFR-mG-CSF (EGFR_BAF)
С использованием стандартных способов выделяли рецептор EGF человека, содержащий последовательность, представленную в SEQ ID NO: 35 (GenBank Acc. No. NM_005228) (в дальнейшем в настоящем описании обозначаемый как "EGFR"), и затем получали вектор, который экспрессирует химерный рецептор рецептора EGF человека, слитый с mG-CSFR. В кратком изложении, вектор (pCV_hEGFR/mG-CSFR) экспрессирует химерный рецептор (hEGFR/mG-CSFR), содержащий внеклеточный домен (Met7-Ser645) EGFR человека и внутриклеточный домен G-CSFR мыши (NM_007782). Нуклеотидная последовательность и аминокислотная последовательность химерного рецептора hEGFR/mG-CSFR представлены в SEQ ID NO: 39 и 40 соответственно.
Пятнадцатью мкг линеаризованного экспрессирующего вектора (pCV_hEGFR/mG-CSFR) химерного рецептора (hEGFR/mG-CSFR), полученного расщеплением посредством PvuI, трансфицировали клетки Ba/F3 посредством электропорации (Gene Pulser; BioRad) в условиях 0,33 кВ и 950 мкФ. Селекцию трансфицированных клеток проводили в среде RPMI1640, содержащей 10% FBS (GIBCO), 200 мкг/мл генетицина и рекомбинантный EREG человека (R&D Systems, каталожный: 1195-EP/CF, 200 нг/мл), и выделяли клеточную линию EGFR_BAF.
4-2. Оценка зависимости EGFR_BAF от EREG
Эксперимент проводили для измерения зависимой от концентрации EREG клеточной пролиферации EGFR_BAF, выделенных способом, описанным в 4-1. Для удаления EREG из культуры клетки центрифугировали и ресуспендировали в среде RPMI1640+10% FBS и затем клетки высевали в каждую лунку 96-луночного планшета с плотностью 2×104 клеток/100 мкл. К клеткам добавляли EREG человека (R&D Systems), разведенный в среде в различных концентрациях (от 7,8 до 250 нг/мл), и затем клетки инкубировали с 5% CO2 при 37°C. После инкубации в течение четырех суток добавляли реагент для подсчета клеток Reagent SF (Nacalai Tesque), проводили окрашивание в течение двух часов в соответствии с инструкциями и определяли поглощение (450/655 нм) реакционного раствора с использованием Benchmark Plus (Bio-Rad).
Аналогично исследовали зависимую от EREG пролиферацию клеточной линии рака поджелудочной железы AsPC1 и клеточной линии рака толстого кишечника DLD1 аналогичным способом, как описано выше. В качестве среды использовали CHO-S-SFMII (GIBCO).
Результат показал, что EGFR_BAF (фиг.7) и AsPC1 (фиг.8) пролиферировали зависимым от концентрации EREG образом. Однако было обнаружено, что DLD1 не обладают зависимостью от EREG человека (фиг.9).
4-3. Определение нейтрализующей активности моноклональных антител против EREG на зависимую от EREG человека клеточную пролиферацию
На пролиферации EGFR_BAF, зависимой от EREG человека, исследовали нейтрализующую активность моноклональных антител против EREG. EGFR_BAF суспендировали в среде RPMI1640+10% FBS, содержащей EREG человека (конечная концентрация 125 нг/мл), и клетки высевали в 96-луночный планшет с плотностью 2×104 клеток на лунку. К клеткам добавляли моноклональное антитело против EREG, разведенное в среде в различных концентрациях (от 0,008 до 25 мкг/мл), и затем клетки культивировали в инкубаторе с 5% CO2 в течение трех суток при 37°C. После культивирования добавляли реагент для подсчета клеток Reagent SF (Nacalai Tesque) и в течение двух часов проводили окрашивание. Затем определяли поглощение реакционного раствора при 450 нм с использованием Benchmark Plus (Bio-Rad) посредством установки длины волны для сравнения на 655 нм.
Скорость клеточной пролиферации вычисляли с использованием измеренного значения при концентрации антитела 0 мкг/мл в качестве сравнения (значение OD для каждой концентрации антитела/значение OD сравнения×100 (%)). В качестве положительного контроля использовали поликлональное антитело козы против EREG человека, о котором известно, что оно обладает ингибирующей активностью (R&D Systems, каталожный: AF1195). В прилагаемой к данному продукту антитела инструкции указана его нейтрализующая активность против зависимой от EREG пролиферации клеточной линии фибробластов Balb/3T3. Однако в прилагаемой инструкции не описана какая-либо информация, касающаяся эффекта подавления пролиферации в злокачественных клеточных линиях.
Более того, определяли нейтрализующую активность моноклональных антител против EREG в AsPC1 в качестве клеток-мишеней аналогичным способом, как описано выше, и измерение проводили в условиях следующих трех концентраций антитела: 25, 2,5 и 0,25 мкг/мл.
Результаты представлены на фиг.10 и 11 соответственно.
Как в EGFR_BAF, так и в AsPC1, B2#30 и B3#18 демонстрировали нейтрализующую активность, и B3#41 демонстрировало очень слабую нейтрализующую активность. Более того, было обнаружено, что нейтрализующая активность известного поликлонального антитела козы против hEREG является очень слабой по сравнению с B2#30 и B3#18.
Таким образом, было обнаружено, что B2#30 и B3#18 обладают нейтрализующей активностью в отношении EREG и подавляют пролиферацию клеточной линии рака поджелудочной железы AsPC1.
Данное исследование показывает, что моноклональные антитела против EREG (B2#30 и B3#18), выделенные в данном исследовании, обладают эффектом уничтожения клеток, опосредуемым активностью ADCC и активностью CDC, а также эффектом подавления зависимой от EREG человека клеточной пролиферации (нейтрализующей активностью), и что антитела можно использовать в качестве фармацевтических средств на основе антитела с новым механизмом против злокачественных клеток, которые обладают усиленной экспрессией гена EREG человека.
[Пример 5]
Определение нуклеотидной последовательности вариабельных областей моноклонального антитела против EREG
Нуклеотидные последовательности вариабельных областей антитела определяли для гибридомного антитела B3#18 (EP27), которое показало нейтрализующую активность в отношении EREG, а также активность ADCC и активность CDC против клеточной линии DLD1. Последовательности вариабельных областей антитела амплифицировали посредством ОТ-ПЦР с тотальной РНК гибридом B3#18, продуцирующих антитело против EREG. Тотальную РНК экстрагировали из 1×107 гибридомных клеток посредством набора RNeasy Plant Mini Kit (QIAGEN).
Библиотеку RACE конструировали из 1 мкг тотальной РНК с использованием набора для амплификации кДНК SMART RACE (CLONTECH). Реакцию обратной транскрипции проводили в течение одного часа и тридцати минут при 42°C. 5'-концевую последовательность кДНК амплифицировали с помощью синтетического олигонуклеотида MHC-IgG2b (SEQ ID NO: 41), который комплементарен последовательности константной области IgG2b мыши, или синтетического олигонуклеотида каппа (SEQ ID NO: 42), который комплементарен нуклеотидной последовательности константной области κ-цепи мыши. Компоненты раствора для ПЦР (в 50 мкл) являются следующими:
5 мкл 10× буфера для ПЦР Advantage 2;
5 мкл 10× смеси праймеров Universal Primer A;
0,2 мМ дНТФ (дАТФ, дГТФ, дЦТФ и дТТФ);
1 мкл смеси полимеразы Advantage 2 (все указанные выше реактивы были изготовлены CLONTECH);
2,5 мкл продукта реакции с обратной трансриптазой; и
10 пмоль синтетического олигонуклеотида MHC-IgG2b или каппа.
Условия ПЦР являются следующими.
Исходная температура, составляющая 94°C в течение 30 секунд, затем пять циклов, состоящих из 94°C в течение пяти секунд и 72°C в течение трех минут;
пять циклов из 94°C в течение пяти секунд, 70°C в течение десяти секунд и 72°C в течение трех минут; и
25 циклов из 94°C в течение пяти секунд, 68°C в течение десяти секунд и 72°C в течение трех минут.
В конце проводили элонгацию продукта реакции при 72°C в течение семи минут. Каждый продукт ПЦР, разделенный в агарозном геле, очищали с использованием набора для экстракции из геля QIAquick (изготовленного QIAGEN) и затем клонировали в вектор pGEM-T Easy (изготовленный Promega). Определяли нуклеотидную последовательность клона. Для B3#18 нуклеотидная последовательность и аминокислотная последовательность вариабельной области H-цепи представлены в SEQ ID NO: 43 и SEQ ID NO: 44 соответственно, и нуклеотидная последовательность и аминокислотная последовательность вариабельной области L-цепи представлены в SEQ ID NO: 45 и SEQ ID NO: 46 соответственно.
Последовательности вариабельной области C7 (EP03), #15 (EP08), B2#30 (EP20), B3#8 (EP24) и B3#41 (EP29) определяли аналогичными способами. Синтетический олигонуклеотид MHC-IgG1 (SEQ ID NO: 47, 5'-ccatggagttagtttgggcagcagatcc-3'), который полностью комплементарен последовательности константной области IgG1 мыши, использовали для амплификации вариабельных областей тяжелой цепи. Определенные нуклеотидные последовательности вариабельной области представлены в SEQ ID NO: 108 (EP03, тяжелая цепь), 110 (EP03, легкая цепь), 112 (EP08, тяжелая цепь), 114 (EP08, легкая цепь), 116 (EP20, тяжелая цепь), 118 (EP20, легкая цепь), 120 (EP24, тяжелая цепь), 122 (EP24, легкая цепь), 124 (EP29, тяжелая цепь) и 126 (EP29, легкая цепь). Транслированные последовательности вариабельных областей представлены в SEQ ID NO: 109 (EP03, тяжелая цепь), 111 (EP03, легкая цепь), 113 (EP08, тяжелая цепь), 115 (EP08, легкая цепь), 117 (EP20, тяжелая цепь), 119 (EP20, легкая цепь), 121 (EP24, тяжелая цепь), 123 (EP24, легкая цепь), 125 (EP29, тяжелая цепь) и 127 (EP29, легкая цепь). Номера ID для последовательностей вариабельных областей и аминокислотные последовательности CDR1, CDR2 и CDR3 каждого антитела обобщенно представлены в таблице 4.
Последовательности CDR моноклонального антитела против EREG и их номера ID последовательности
(SEQ ID NO: 49)
(SEQ ID NO: 51)
(SEQ ID NO: 53)
(SEQ ID NO: 55)
(SEQ ID NO: 57)
(SEQ ID NO: 59)
(SEQ ID NO: 61)
(SEQ ID NO: 63)
(SEQ ID NO: 65)
(SEQ ID NO: 67)
(SEQ ID NO: 69)
(SEQ ID NO: 71)
(SEQ ID NO: 73)
(SEQ ID NO: 75)
(SEQ ID NO: 77)
(SEQ ID NO: 79)
(SEQ ID NO: 81)
(SEQ ID NO: 83)
(SEQ ID NO: 85)
(SEQ ID NO: 87)
(SEQ ID NO: 89)
(SEQ ID NO: 91)
(SEQ ID NO: 93)
(SEQ ID NO: 95)
(SEQ ID NO: 2)
(SEQ ID NO: 4)
(SEQ ID NO: 6)
(SEQ ID NO: 12)
(SEQ ID NO: 14)
(SEQ ID NO: 16)
(SEQ ID NO: 97)
(SEQ ID NO: 99)
(SEQ ID NO: 101)
(SEQ ID NO: 103)
(SEQ ID NO: 105)
(SEQ ID NO: 107)
[Пример 6]
Подтверждение активности связывания EREG рекомбинантных химерных антител
Было подтверждено, что рекомбинантные химерные антитела, содержащие выделенные последовательности вариабельной области антитела и константной области IgG1/IgK человека связываются с EREG, следующим образом. Последовательность каждой вариабельной области тяжелой цепи амплифицировали посредством ПЦР и затем встраивали в участок для клонирования тяжелой цепи в экспрессирующем векторе химерного антитела человека. Затем последовательность вариабельной области легкой цепи амплифицировали посредством ПЦР и затем встраивали в участок для клонирования легкой цепи в указанном выше экспрессирующем векторе химерного антитела человека, в который встроена последовательность тяжелой цепи. В сконструированных экспрессирующих векторах клеток млекопитающих гены как тяжелой, так и легкой цепи сконструированы, чтобы они транскрибировались под контролем промоторов CMV мыши. Номера ID последовательностей для нуклеотидных последовательностей и транслированных последовательностей каждого химерного антитела человека обобщенно представлены в таблице 5.
Номера ID последовательности химерных антител человека
Клетки DG44 трансформировали экспрессирующим вектором химерного антитела человека способом электропорации. Селекцию рекомбинантных клеточных клонов проводили посредством устойчивости к генетицину, которая обеспечивалась селективным маркером в экспрессирующем векторе химерного антитела человека. Антитело в культуральном супернатанте рекомбинантных клонов количественно определяли посредством сэндвич-ELISA с использованием антител против антител человека и отбирали рекомбинантные клетки, экспрессирующие антитело. Химерные антитела человека очищали из культурального супернатанта выбранных рекомбинантных клеток с использованием колонки HiTrap Protein A (Amersham Bioscience) в соответствии с прилагаемым руководством.
Для анализа связывания очищенных химерных антител с EREG использовали планшеты Nunc-Immuno, на которые наносили sEREG-mFc и затем блокировали раствором, содержащим BSA. После добавления химерных антител для связывания планшеты инкубировали в течение одного часа и затем промывали. После этого добавляли меченное щелочной фосфатазой антитело против IgG человека (BIOSOURCE, AHI0305) и давали возможность реагировать. После промывания связанное химерное антитело количественно определяли добавлением хромогенного реагента Sigma 104. Как представлено на фиг.12, химерные антитела связывались с EREG дозозависимым образом. На основе указанных выше результатов было подтверждено, что последовательности вариабельной области выделенного антитела в действительности являются последовательностями антител против EREG.
[Пример 7]
Активность ADCC in vitro, индуцируемая химерными антителами против EREG
На фиг.13 представлена активность ADCC in vitro, индуцируемая химерными антителами против EREG и PBMC человека. С использованием инъецированного шприцом 200 мкл раствора 1000 единиц/мл гепарина (гепарин для инъекций Novo, 5000 единиц, Novo Nordisk) проводили забор 50 мл периферической крови от здорового индивида. Периферическую кровь разводили в два раза посредством PBS(-) и разделяли на четыре аликвоты. Для выделения лимфоцитов их добавляли в пробирки Leucosep (каталожный No. 227290, Greiner Bio-One), в которых заранее было добавлено 15 мл Ficoll-Paque PLUS (GE Healthcare), и подвергали центрифугированию. После центрифугирования при 2150 об/мин при комнатной температуре в течение десяти минут, собирали фракцию моноцитов. Клетки промывали один раз модифицированной способом Дульбекко средой Игла (изготовленной SIGMA), содержащей 10% FBS (в дальнейшем в настоящем описании обозначаемой как 10% FBS/D-MEM), и затем клеточную плотность доводили до 5×106/мл в 10% FBS/D-MEM, с получением раствора с PBMC человека.
Клетки DLD-1 и MIA-PaCa2 метили хромом-51 способом аналогично способу примера 3. К клеткам-мишеням добавляли раствор антитела, доведенный до каждой концентрации, и давали возможность связываться при комнатной температуре в течение 15 минут. Затем добавляли раствор PBMC человека (5×105 клеток/лунка) и клетки инкубировали в инкубаторе с 5% диоксидом углерода в течение приблизительно четырех часов. Радиоактивность в супернатанте после инкубации измеряли с использованием гамма-счетчика и скорость высвобождения хрома вычисляли способом, описанным в примере 3. Активность ADCC индуцировалась всеми из химерных антител, а химерные антитела EP20, EP27 и EP08 обладали особенно сильной активностью. Была продемонстрирована сходная сила индукции ADCC каждым из антител в клеточной линии рака поджелудочной железы MIA-PaCa2 и клеточной линии рака толстого кишечника DLD-1.
[Пример 8]
Связывание с ортологами EREG и анализ эпитопов антител против EREG
Анализировали связывание выделенных антител с EREG мыши и EREG обезьяны. Полноразмерную кДНК EREG мыши (NM_007950) и кДНК EREG обезьяны (XM_001102069) амплифицировали посредством ПЦР из библиотек кДНК и клонировали. Выделенные последовательности представлены в SEQ ID NO: 148 и 149 соответственно. Растворимый слитый белок EREG мыши/Fc IgG2a мыши (в дальнейшем в настоящем описании EREG мыши-mFc) и растворимый слитый белок EREG обезьяны/Fc IgG2a мыши (в дальнейшем в настоящем описании EREG обезьяны-mFc) получали способом, описанным в примере 2. Последовательности EREG мыши-mFc и EREG обезьяны-mFc представлены в SEQ ID NO: 150 и 151 соответственно. На планшеты Nunc-Immuno наносили EREG человека-mFc, EREG мыши-mFc и EREG обезьяны-mFc соответственно и затем блокировали раствором, содержащим BSA. Затем анализировали связывание очищенных химерных антител. После добавления химерных антител планшеты инкубировали в течение одного часа и затем промывали. После этого добавляли меченное щелочной фосфатазой антитело против IgG человека (BIOSOURCE, AHI0305) и давали возможность реагировать. После промывания связанное химерное антитело количественно определяли добавлением хромогенного реагента Sigma 104 (фиг.14). Различия в степени насыщения при связывании антитела с EREG при одинаковой концентрации антител демонстрировали нанесением реактивности в отношении различных типов EREG с одинаковыми концентрациями антител в качестве "EREG человека-mFc" на горизонтальной оси против "EREG мыши-mFc" на вертикальной оси или "EREG мыши-mFc" на горизонтальной оси против "EREG обезьяны-mFc" на вертикальной оси. График вдоль горизонтальной оси указывает на селективность связывания в отношении EREG человека, и это демонстрирует, что EP08 связывается с EREG человека, но не связывается с EREG мыши. Параболический график для EP27 указывает на то, что EP27 связывается с молекулой мыши, но для насыщения связывания требуется более высокая концентрация антитела. Иными словами, EP27 обладает более низкой аффинностью связывания в отношении EREG мыши, чем в отношении EREG человека. Антитело, которое образует диагональный график, означает, что антитело обладает активностью в отношении EREG не относящегося к человеку вида, эквивалентной EREG человека. Результат, полученный посредством указанного выше анализа продуцированных гибридомой антител мыши, обобщенно представлен в таблице 3. Результат показан посредством четырех уровней реактивности относительно молекулы человека: нет реактивности (-); очень низкая реактивность (+); немного более низкая реактивность (++); и эквивалентная реактивность (+++).
Для определения эпитопа антитела анализировали связывание с неполными фрагментами внеклеточного домена EREG человека. N-концевую неполную последовательность фрагмента от 29Ala до 69Ser EREG человека SEQ ID NO: 21 экспрессировали в качестве слитого белка GST и очищали до гомогенности. Последовательность полученного рекомбинантного белка представлена в SEQ ID NO: 152. Растворимый фрагмент EREG человека (домен EGF; соответствующий 63Val-108Leu SEQ ID NO: 21), содержащий зрелую структуру, приобретали от R&D Systems (каталожный no. 1195-EP/CF). Фрагменты белка EREG иммобилизовали на иммунологическом планшете и посредством ELISA исследовали связывание антител. Результат обобщенно представлен в таблице 3. Большинство из антител связывались с растворимым фрагментом EREG, который содержит домен EGF (указанным как "ereg" в таблице 3). EP25 и EP32 связывались с N-концевым фрагментом ("Nterm"). Все из выделенных антител связывались с каким-либо одним из пептидных фрагментов, и ни одно из антител не распознавало последовательность после остатка 109.
[Пример 9]
Активация передачи сигнала рецептора EGF экспрессирующими EREG клетками
Растворимый EREG был описан как низкоаффинный лиганд для рецепторов EGF (ссылочные документы: Shelly et al. (1998) J. Biol. Chem. 273, 10496-505; Jones et al. (1999) FEBS Lett 447, 227-31). Посредством сравнения эффектов на EGFR_BaF было подтверждено, что EREG обладает низкой активностью (аффинностью) в отношении рецептора EGF, которая составляет одну сотую аффинности EGF или менее (фиг.15). EREG не является обильно представленным белком в крови в физиологических условиях. В то время как была описана повышенная экспрессия гена EREG в ходе злокачественной трансформации, не существует описаний, демонстрирующих, что белок EREG был выявлен в высокой концентрации в крови пациентов со злокачественной опухолью. Авторы настоящего изобретения предположили, что EREG мембранного типа, экспрессированный на клеточной мембране, отличный от растворимого EREG, стимулирует ближайший рецептор EGF, и исследовали гипотезу о том, что может присутствовать локальный механизм, следующим образом. Было исследовано, стимулируется ли пролиферация EGFR_BaF не только растворимым EREG, но и кокультивированием EGFR_BaF с клетками, искусственно экспрессирующими EREG. Экспрессирующие EREG клетки EREG_DG и клетки DG44 (отрицательный контроль) высевали в 96-луночный планшет в количестве 5×103 клеток/лунка и инкубировали в течение ночи со средой, содержащей FCS. На следующие сутки добавляли 1×104 клеток EGFR_BaF или клеток Ba/F3 (отрицательный контроль) и затем клетки инкубировали в течение трех суток. Как представлено на фиг.16, кокультивирование с клетками EREG_DG, которые обладали искусственной экспрессией EREG, значительно усиливало пролиферацию EGFR_BaF. Однако кокультивирование с DG44, которые не экспрессируют EREG, совсем не стимулировало пролиферацию EGFR_BaF. Стимулированную пролиферацию EREG_DG подавляли добавлением антитела EP20 против EREG (10 мкг/мл). Клеточную пролиферацию EGFR_BaF количественно определяли добавлением реагента WST-8 (фиг.17). Было показано, что кокультивирование с клетками EREG_DG не стимулировало пролиферацию клеток Ba/F3 и что клеточную пролиферацию, запускаемую кокультивированием EGFR_BaF с EREG_DG, можно полностью подавлять добавлением антитела против EREG.
Анализ посредством GeneChip показал, что в дополнение к EREG, экспрессия мРНК амфирегулина (члена семейства лигандов EGF) также усиливалась в клетках рака толстого кишечника DLD-1. При передачи сигнала рецептора EGF существует избыточность молекул семейства лигандов, которые активируют рецептор, и, таким образом, теоретически ингибирование одного из лигандов не эквивалентно ингибированию передачи сигнала рецептора. Было показано, что посредством кокультивирования с DLD-1 пролиферация EGFR_BaF была значительно усилена по сравнению с пролиферацией Ba/F3 и что такую усиленную пролиферацию подавляло добавление нейтрализующего антитела против EREG EP20 или EP27 (фиг.18).
Растворимый белок EREG в культуральных супернатантах клеток EREG_DG и DLD-1 количественно определяли посредством сэндвич-ELISA. На иммунологический планшет наносили EP30 мыши и подвергали блокированию. Затем для исследования добавляли растворимый рекомбинантный EREG человека (R&D Systems), используемый в качестве стандарта для вычисления концентрации, и супернатант культуры клеток после трех суток культивирования. После инкубации проводили детектирование белка EREG путем добавления по порядку химеризованного антитела EP20 и меченного щелочной фосфатазой антитела против IgG человека. Детектирование растворимых белков EREG с использованием стандарта представлена на фиг.19. Концентрация EREG в культуральном супернатанте была ниже предела детектирования (1 нг/мл). Выявленные значения A405/655 составляют 0,157 для EREG_DG и 0,184 для DLD-1. Указанный выше результат демонстрирует, что между соседними клетками происходит опосредуемая EREG активация передачи сигнала рецептором EGF и что EREG мембранного типа с высокой вероятностью вовлечен в активацию.
Данный результат, что опосредуемая EREG активация рецептора EGF эффективно происходит посредством межклеточного взаимодействия, был впервые открыт с помощью настоящего изобретения. Открытие того, что активацию можно подавлять антителом против EREG, также было сделано впервые.
[Пример 10]
Детектирование белка EREG в опухолях толстого кишечника иммуногистохимическим окрашиванием
Экспрессию белка EREG в опухолях и локализацию белка на мембранах злокачественных клеток демонстрировали иммуногистохимическим окрашиванием. Иммуногистохимическое окрашивание проводили с использованием ряда тканей, полученных из клинических злокачественных тканей, и образцы тканей получали следующим образом. Ткани, удаленные при хирургической операции, фиксировали в течение ночи в 4% параформальдегиде, затем промывали в течение двух часов 0,01 M забуференным фосфатом солевым раствором и затем погружали в парафин способом AMeX (ссылочный документ: Sato et al. (1986) Am. J. Pathol. 125, 431-5). Злокачественную ткань диаметром 1,5-мм вырезали из данного блока, переносили в новый блок размером приблизительно 2,5×3,0 см и различные ткани располагали в ряд для получения ряда тканей. Ряд тканей разрезали и подвергали депарафинизации и срезы иммуногистохимически окрашивали с использованием системы Ventana HX Discovery System (Ventana Medical Systems, Inc.). К срезам тканей добавляли пятнадцать мкг/мл моноклонального антитела мыши против EREG (EP27) и затем давали возможность реагировать. В качестве вторичного антитела использовали меченное пероксидазой антитело против IgG мыши и затем проводили реакцию цветного проявления с использованием DAB. Выявили экспрессию белка EREG и его локализацию на клеточной мембране в первичном раке толстого кишечника (фиг.20). Между тем, белок EREG совсем не был выявлен в незлокачественных тканях в тех же условиях окрашивания. Детектирование белка EREG и локализация белка на мембране в злокачественных тканях были показаны впервые в настоящем изобретении.
[Пример 11]
Классификация моноклональных антител против EREG по распознаванию эпитопа в сэндвич-ELISA
В примере 8 было обнаружено, что распознаваемые антителом эпитопы, главным образом, были расположены в зрелом домене EGF EREG. Более конкретно, эпитопы антитела классифицировали посредством идентификации сочетаний антител, которые могут неконкурентно связываться с одной молекулой антигена в сэндвич-ELISA. Антитело мыши против EREG разводили до 1 мкг/мл и иммобилизовали на планшетах Nunc-Immuno. После блокирования добавляли 50 мкг/мл растворимого фрагмента EREG человека, содержащего зрелый домен EGF (63Val-108Leu SEQ ID NO: 21) или 100 мг/мл EREG человека-mFc (SEQ ID NO: 34), в количестве 100 мкл/лунка, и, таким образом, молекула антигена улавливалась антителом мыши. После промывания добавляли химерное антитело против EREG (концентрация антитела: 1, 0,33, 0,11 или 0,037 мкг/мл) и количественно определяли связывание химерного антитела с уловленным антигеном посредством добавления меченного щелочной фосфатазой антитела против IgG человека и хромогенного реагента Sigma 104. Полученные данные (фиг.21-1)-(фиг.21-3) подвергали кластерному анализу с использованием R-программного обеспечения для статистического анализа (URL http://www.R-project.org.), и дендрограмма представлена на фиг.22. На фиг.22 чем больше расстояние до точки ветвления на горизонтальной оси, тем более отличающимися являются паттерны реакций в системе сэндвич-ELISA и тем более отличающимся являются типы распознавания антигена. EP20 может связываться с EREG одновременно со многими антителами, включая EP27, и оказалось, что характерным является эпитоп EP20. Антитела, относящиеся к кластеру EP29-EP25 на фиг.22, почти не давали возможность одновременного связывания EREG с другими антителами, и, таким образом, полагают, что молекула антигена прочно связывалась данными антителами.
Промышленная применимость
Специфичные к белку EREG антитела согласно изобретению можно использовать в качестве диагностических средств при раке толстого кишечника, аденокарциноме легкого, раке поджелудочной железы, раке желудка, раке почки или подобными. Диагностические средства согласно изобретению пригодны для диагностики первичных или метастатических злокачественных опухолей. Более конкретно, злокачественные опухоли можно выявлять посредством детектирования белка EREG или кодирующей EREG мРНК, содержащихся в биологическом образце, полученном от пациента. Альтернативно, наличие первичного рака толстого кишечника, метастазирующего рака толстого кишечника, аденокарциномы легкого, рака поджелудочной железы, рака желудка или рака почки можно выявлять in vivo посредством детектирования локализации экспрессирующих EREG клеток in vivo.
Более того, антитела против EREG, обладающие цитотоксической активностью в настоящем изобретении, пригодны для лечения или профилактики злокачественных опухолей, которые экспрессируют белок EREG. Более конкретно, посредством настоящего изобретения предоставляются цитотоксические средства или ингибиторы пролиферации клеток для различных типов злокачественных клеток, таких как клетки рака толстого кишечника, аденокарциномы легкого, рака поджелудочной железы, рака желудка или рака почки. Цитотоксические средства или ингибиторы пролиферации клеток для злокачественных клеток в настоящем изобретении можно применять как к первичным, так и к метастатическим злокачественным опухолям.
Более того, в соответствии с настоящим изобретением антитела против EREG, обладающие цитотоксической активностью, можно использовать в качестве лекарственных средств против различных типов злокачественных опухолей, таких как рак толстого кишечника, аденокарцинома легкого, рак поджелудочной железы, рак желудка или рак почки. В настоящем изобретении антитела против EREG пригодны в качестве лекарственных средств как для первичных, так и метастатических злокачественных опухолей.
Кроме того, гены, кодирующие антитела согласно изобретению, и рекомбинантные клетки, трансформированные этими генами, можно использовать для получения рекомбинантных антител, которые проявляют указанные выше эффекты или более предпочтительные эффекты.
Более того, на основе открытий согласно изобретению можно проводить скрининг соединений-кандидатов с целью выявления полезных лекарственных средств против злокачественной опухоли. Посредством настоящего изобретения была показана зависимая от EREG пролиферация злокачественной опухоли. Таким образом, соединения, выбранные способами скрининга согласно изобретению, можно считать соединениями, которые подавляют зависимую от EREG пролиферацию злокачественной опухоли. Соединения, выбранные посредством настоящего изобретения, пригодны в качестве соединений-кандидатов для оценки пригодности средств против злокачественной опухоли посредством оценки безопасности и специфичности к злокачественной опухоли.
| название | год | авторы | номер документа |
|---|---|---|---|
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ И ПРОФИЛАКТИКИ ЗЛОКАЧЕСТВЕННОЙ ОПУХОЛИ | 2012 |
|
RU2641260C2 |
| ЛЕКАРСТВЕННЫЙ ПРЕПАРАТ ДЛЯ ЛЕЧЕНИЯ И/ИЛИ ПРОФИЛАКТИКИ РАКА | 2011 |
|
RU2624029C2 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ И ПРОФИЛАКТИКИ ЗЛОКАЧЕСТВЕННЫХ ОПУХОЛЕЙ | 2009 |
|
RU2498819C2 |
| АНТИТЕЛО ПРОТИВ ЗЛОКАЧЕСТВЕННОЙ КЛЕТКИ, ПРОТИВОРАКОВОЕ ЛЕКАРСТВЕННОЕ СРЕДСТВО И СПОСОБ ТЕСТИРОВАНИЯ ЗЛОКАЧЕСТВЕННОГО НОВООБРАЗОВАНИЯ | 2015 |
|
RU2753512C2 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ И/ИЛИ ПРОФИЛАКТИКИ ЗЛОКАЧЕСТВЕННОЙ ОПУХОЛИ | 2012 |
|
RU2611197C2 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ ИЛИ ПРОФИЛАКТИКИ РАКА | 2013 |
|
RU2631804C2 |
| СПОСОБЫ РАЗРУШЕНИЯ КЛЕТОК С ИСПОЛЬЗОВАНИЕМ ЭФФЕКТОРНЫХ ФУНКЦИЙ АНТИ-EphA4 АНТИТЕЛ | 2007 |
|
RU2429246C2 |
| ИНДУЦИРУЮЩЕЕ ПОВРЕЖДЕНИЕ КЛЕТОК ТЕРАПЕВТИЧЕСКОЕ ЛЕКАРСТВЕННОЕ СРЕДСТВО, ПРЕДНАЗНАЧЕННОЕ ДЛЯ ПРОТИВОРАКОВОЙ ТЕРАПИИ | 2017 |
|
RU2746754C2 |
| Молекулы, связывающиеся с 4-1ВВ | 2011 |
|
RU2710717C2 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ И/ИЛИ ПРОФИЛАКТИКИ ЗЛОКАЧЕСТВЕННОЙ ОПУХОЛИ | 2012 |
|
RU2610428C2 |
























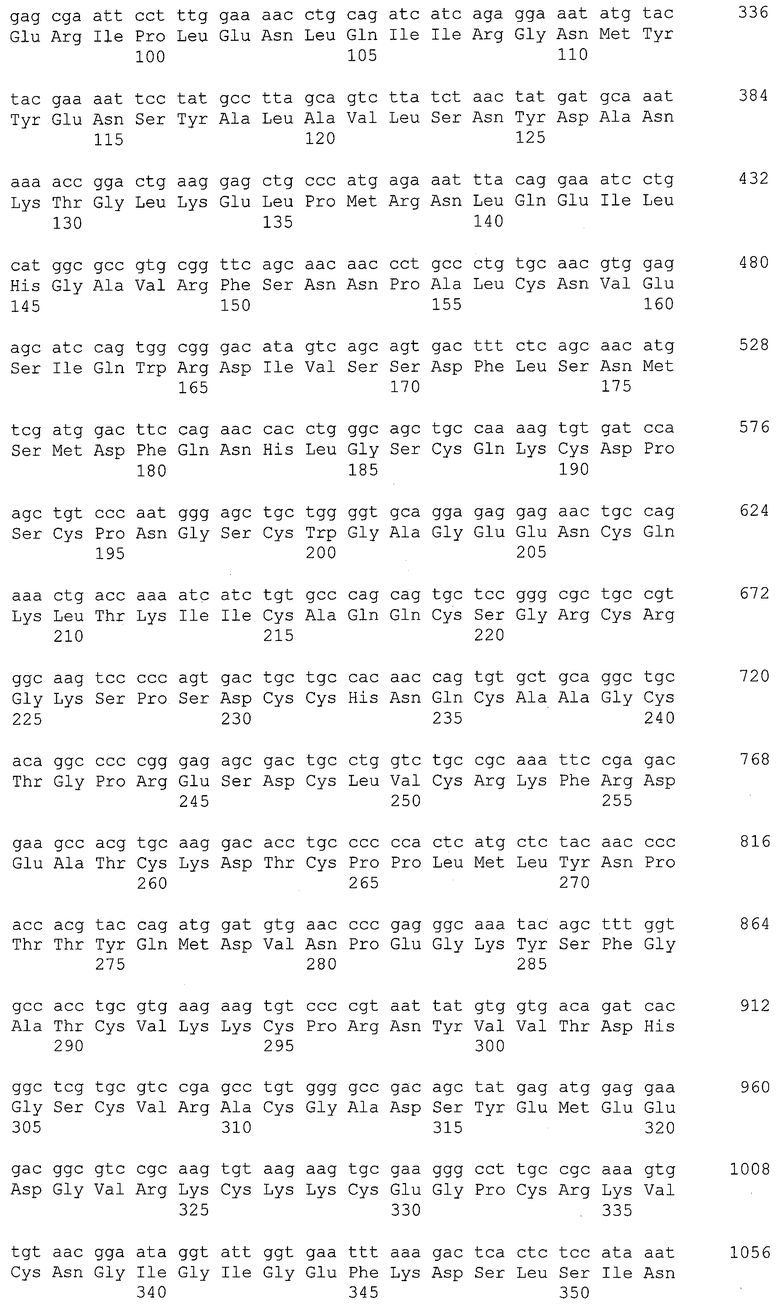


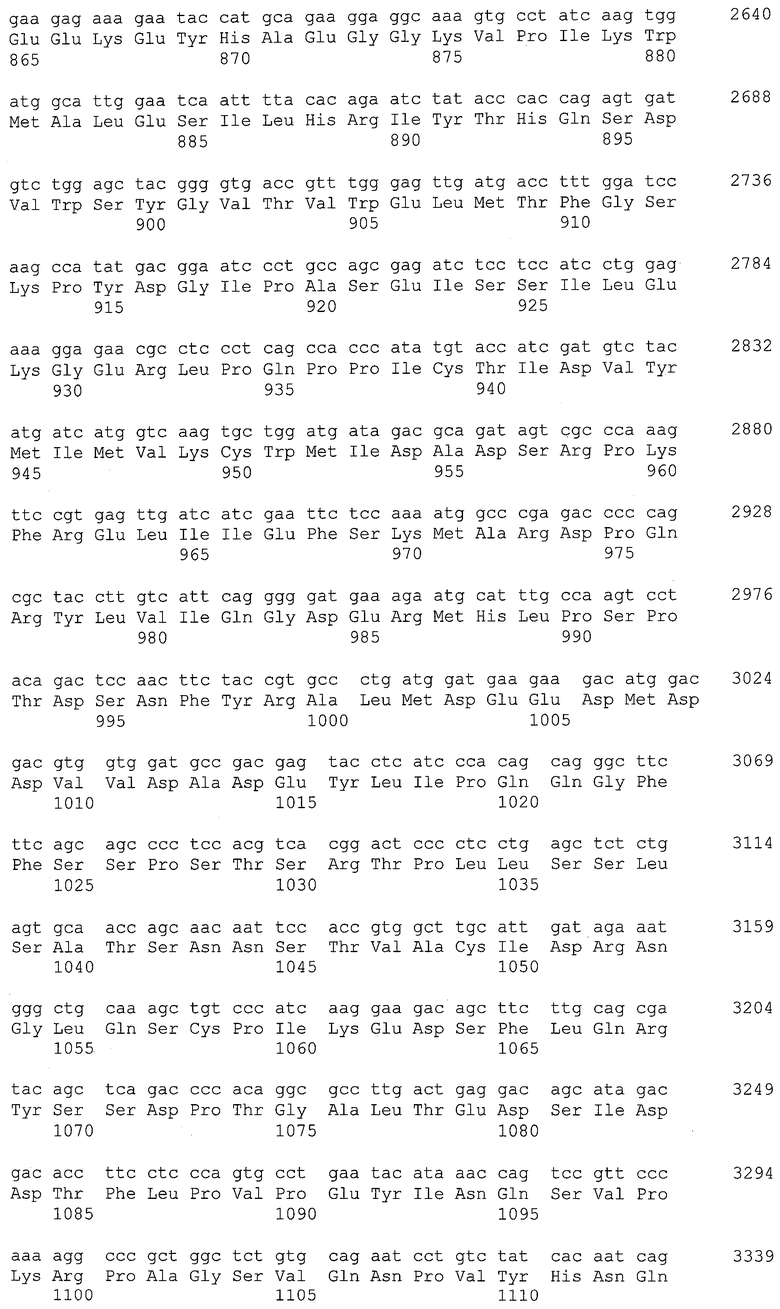








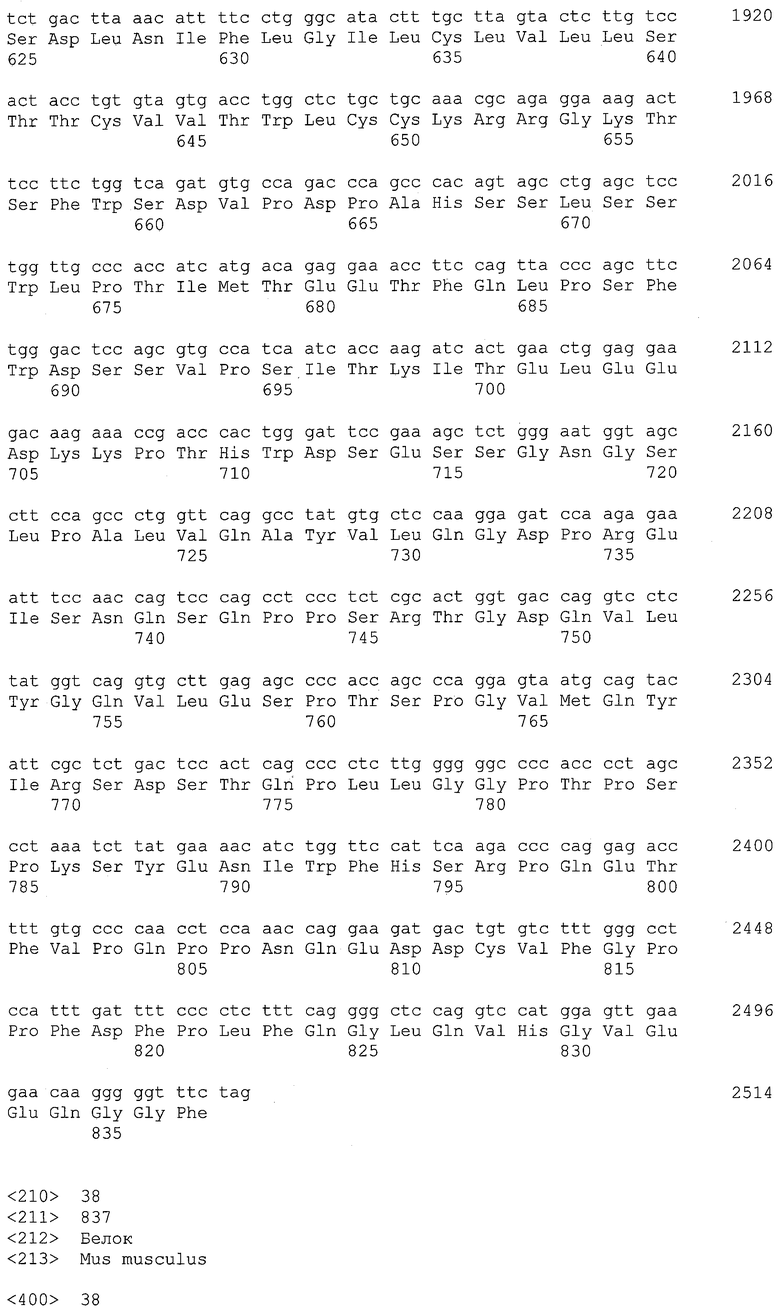
















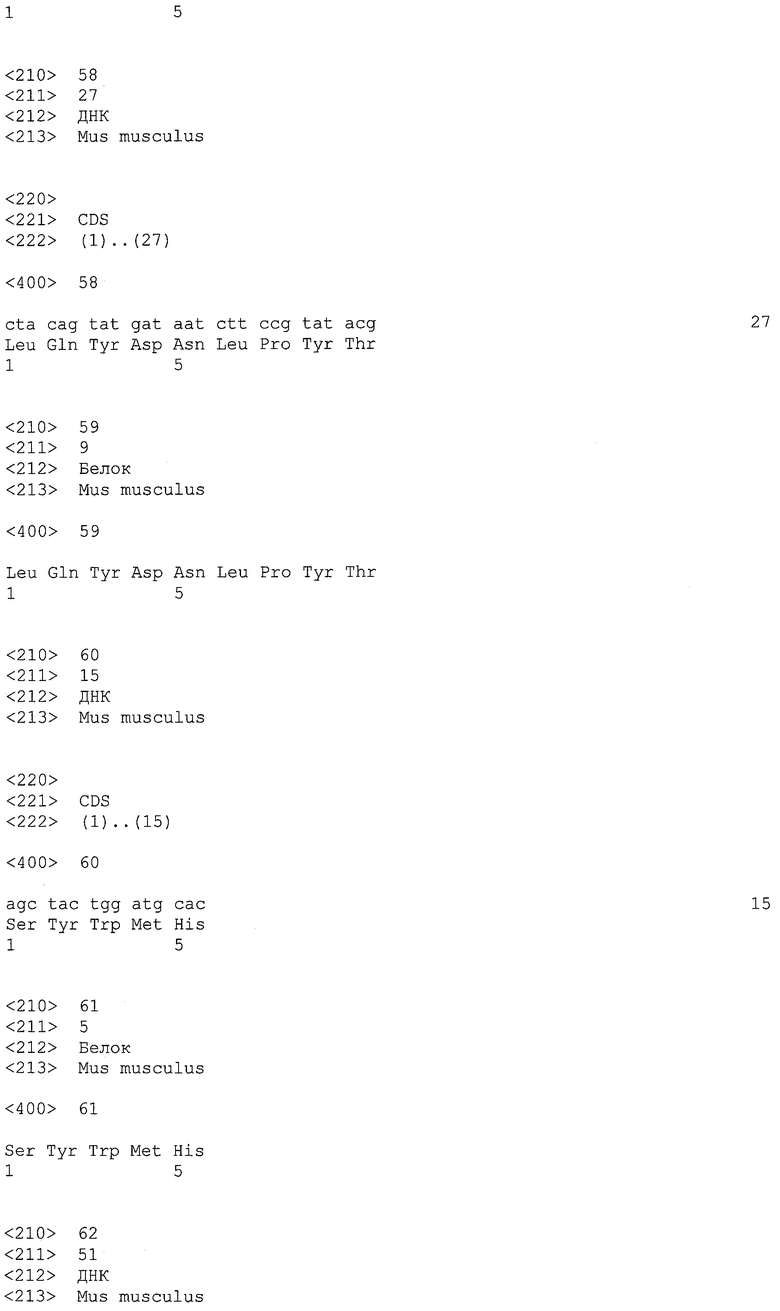








































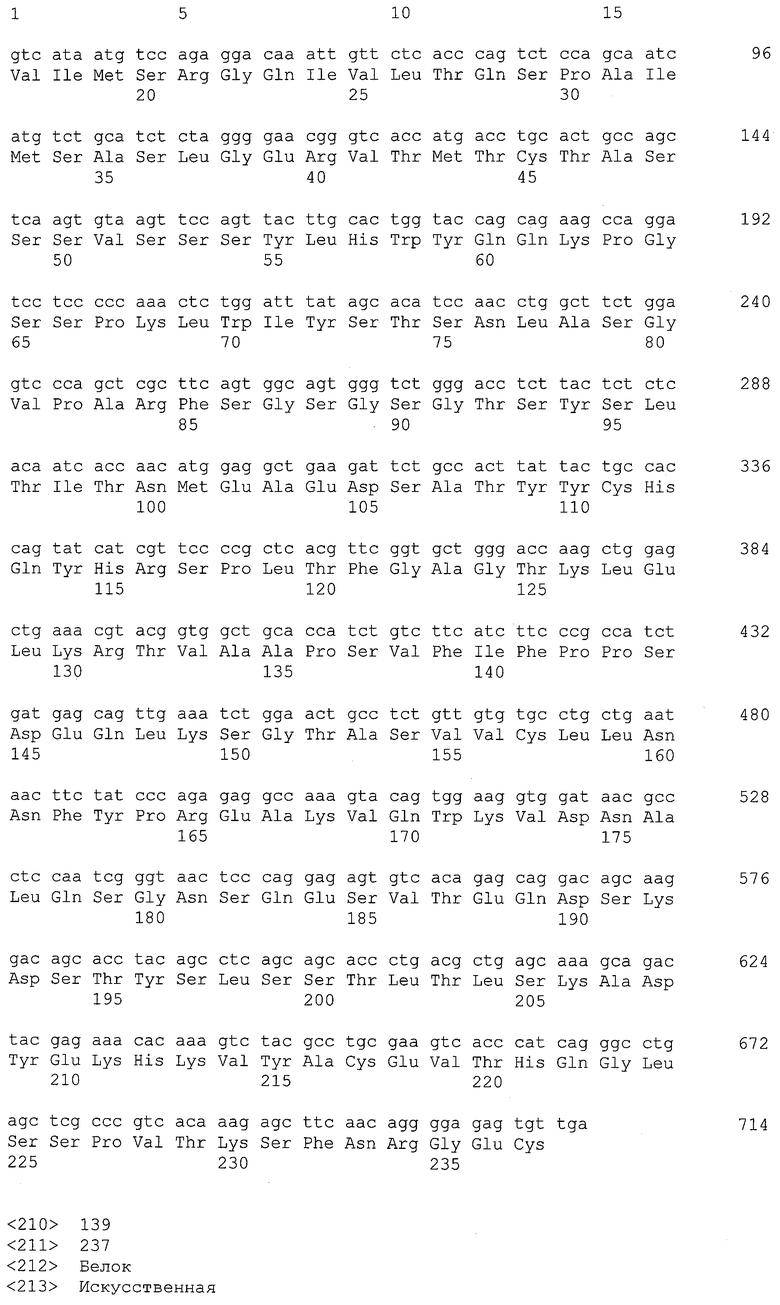




















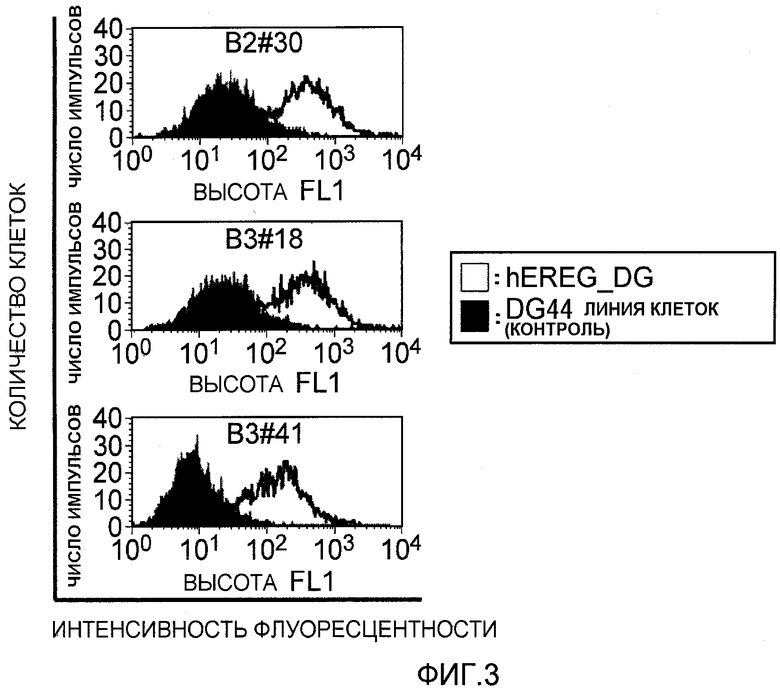
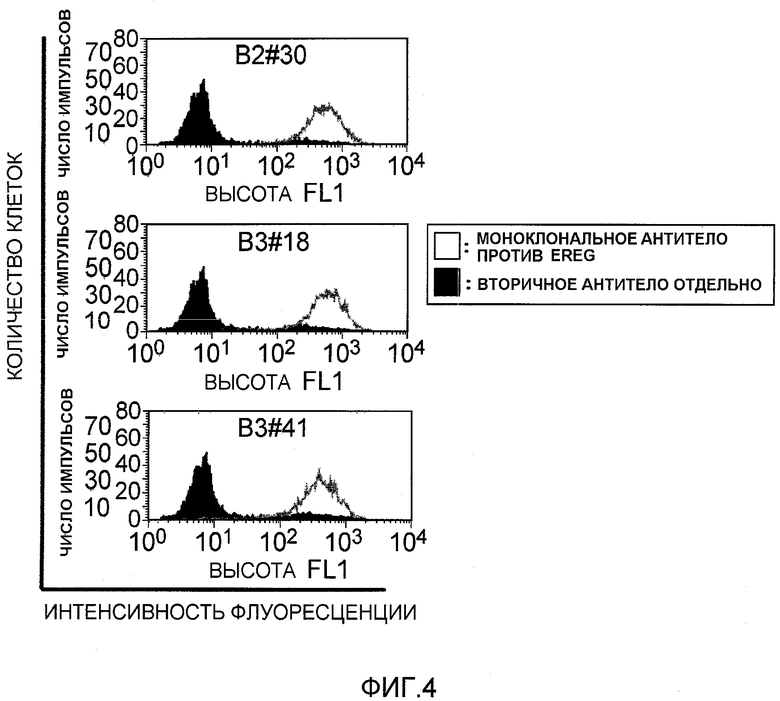
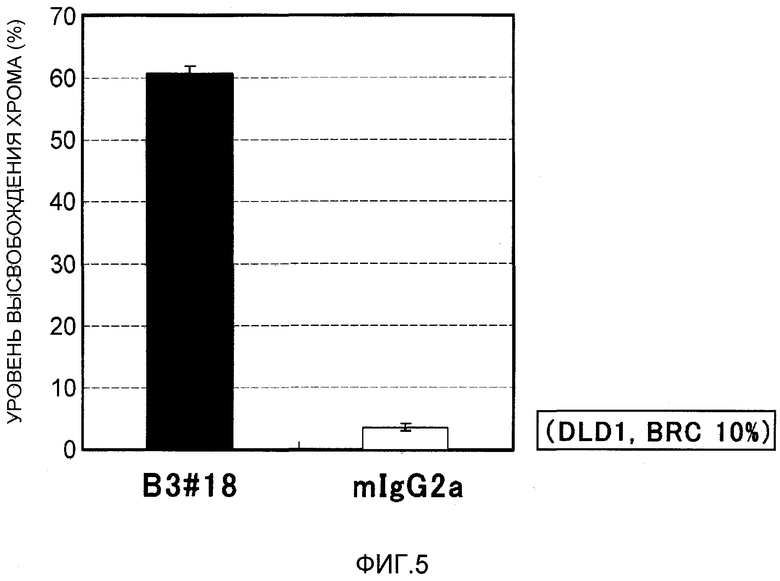
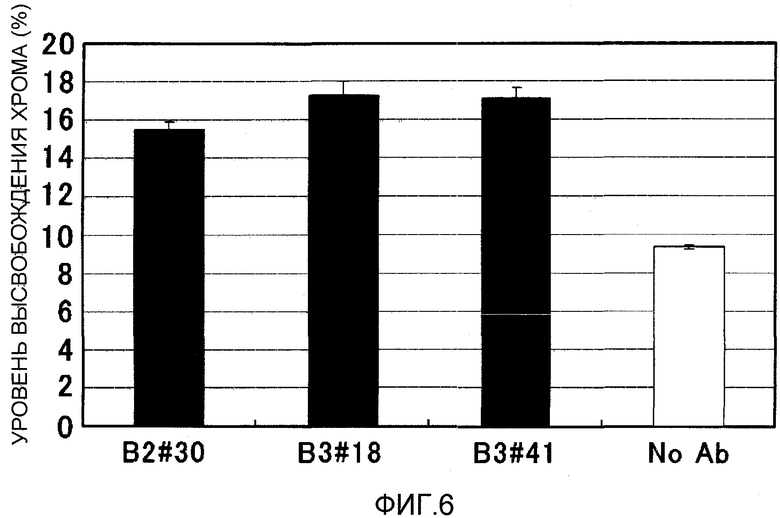
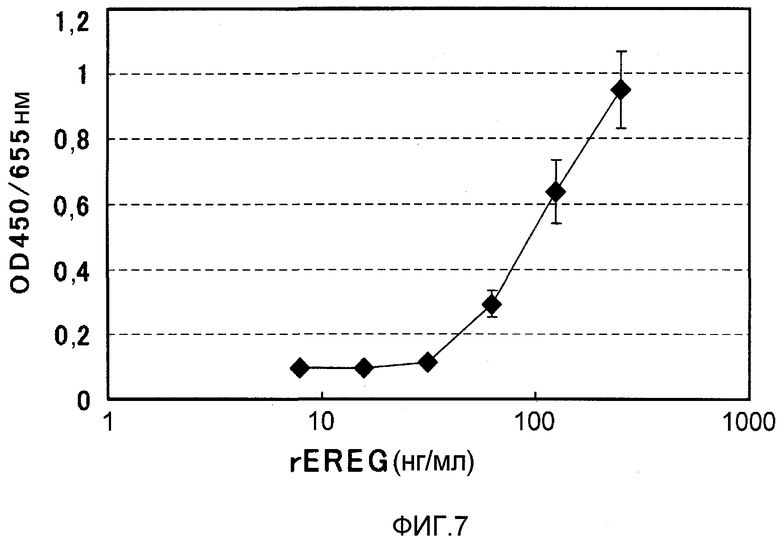
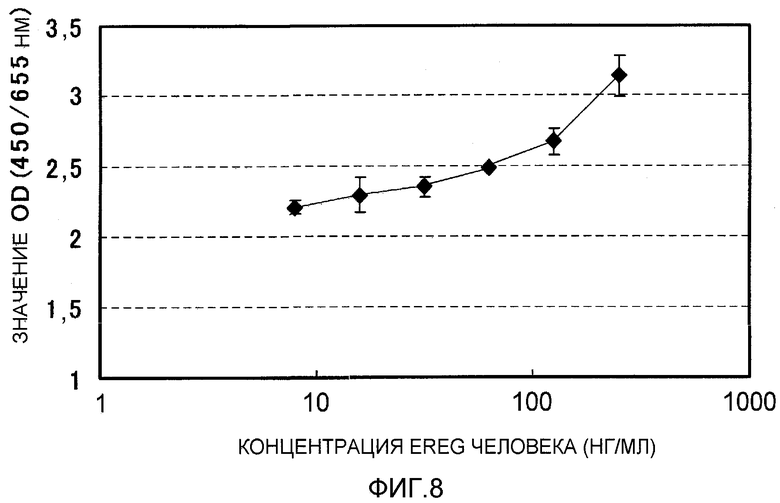
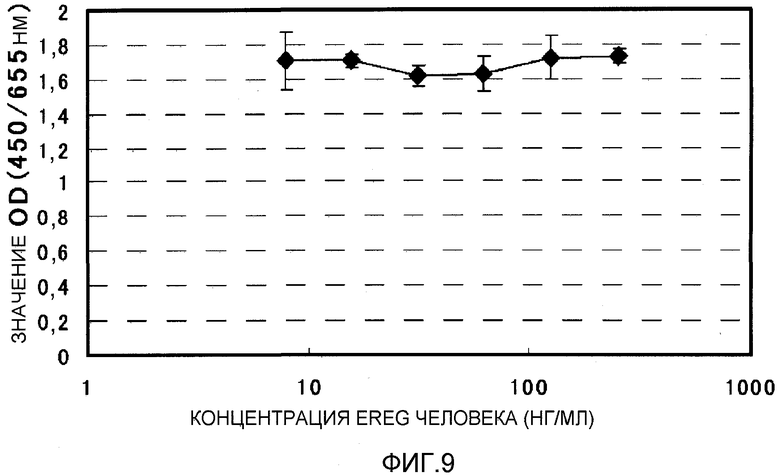
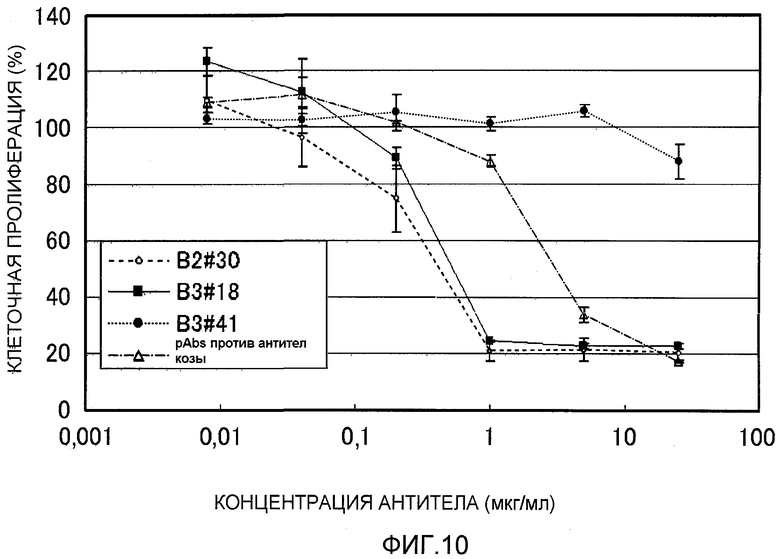
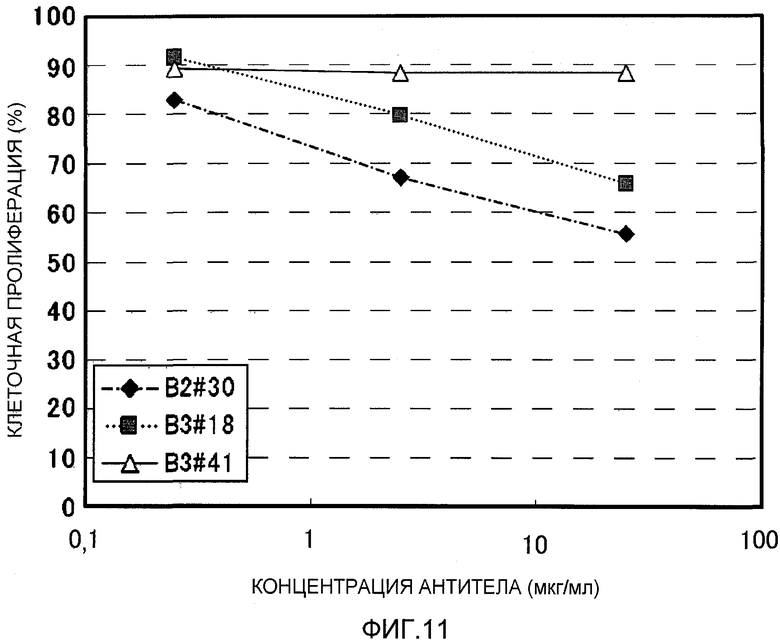

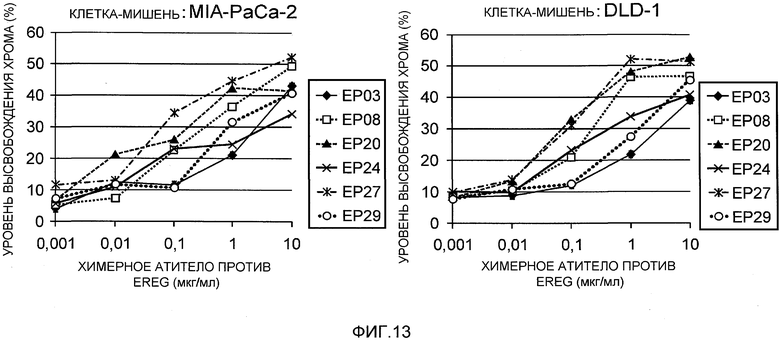
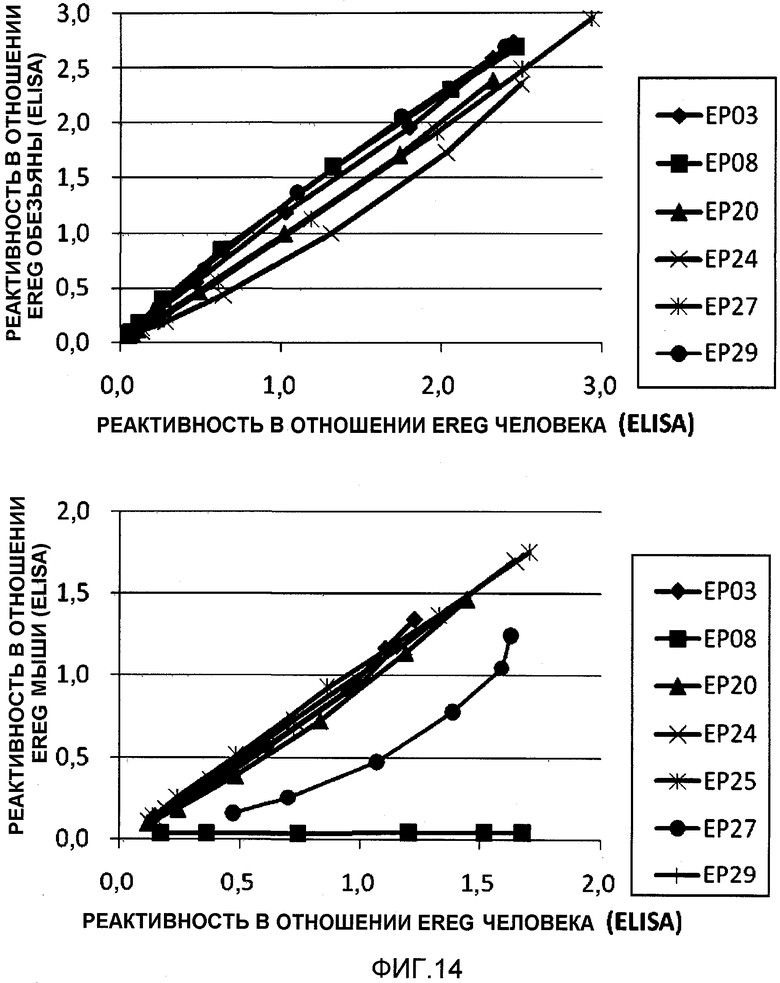
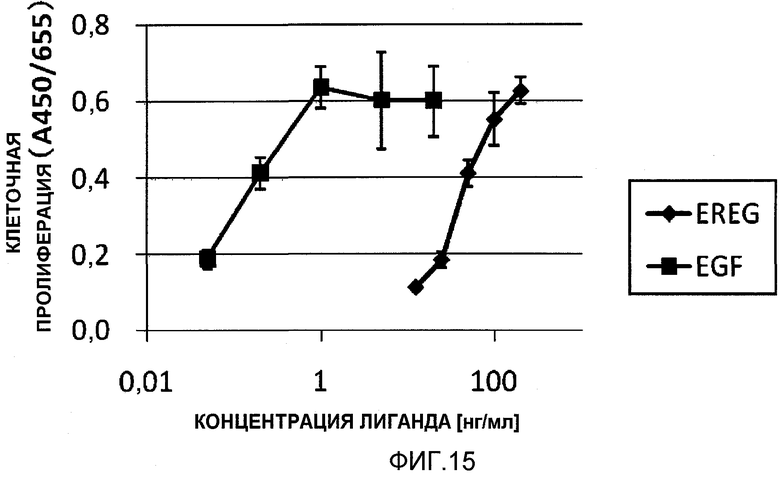

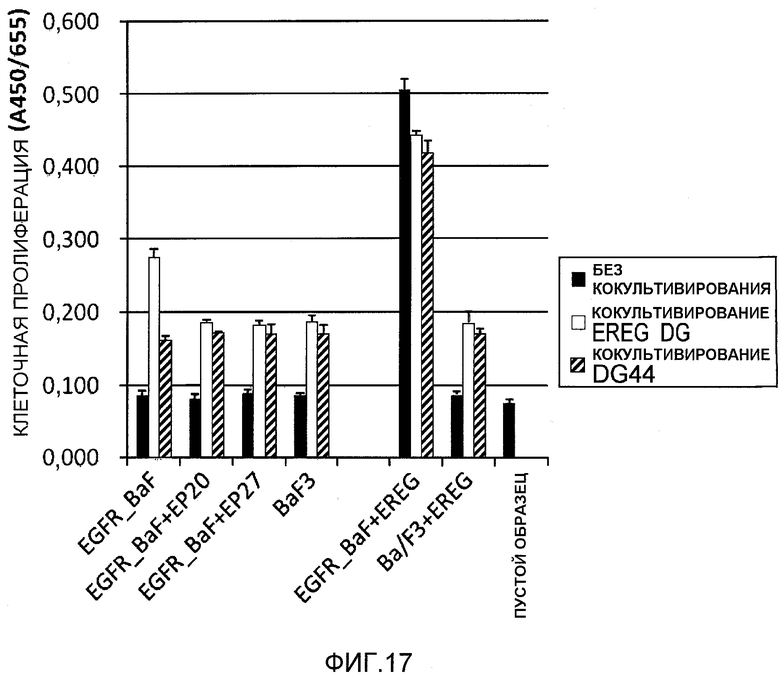
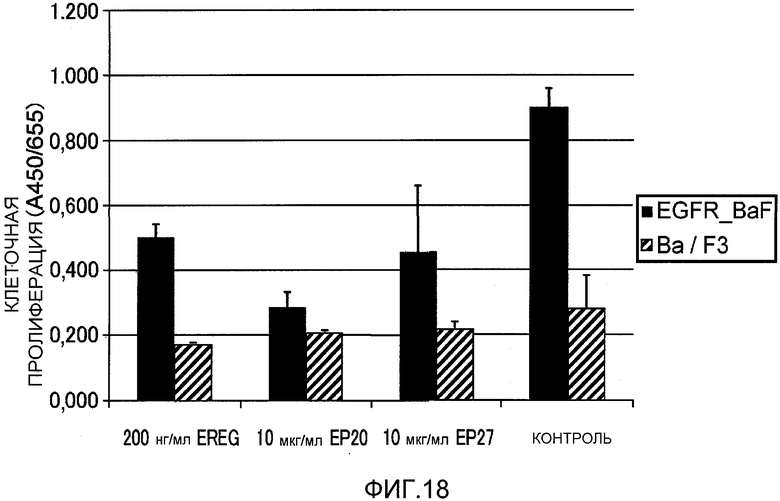
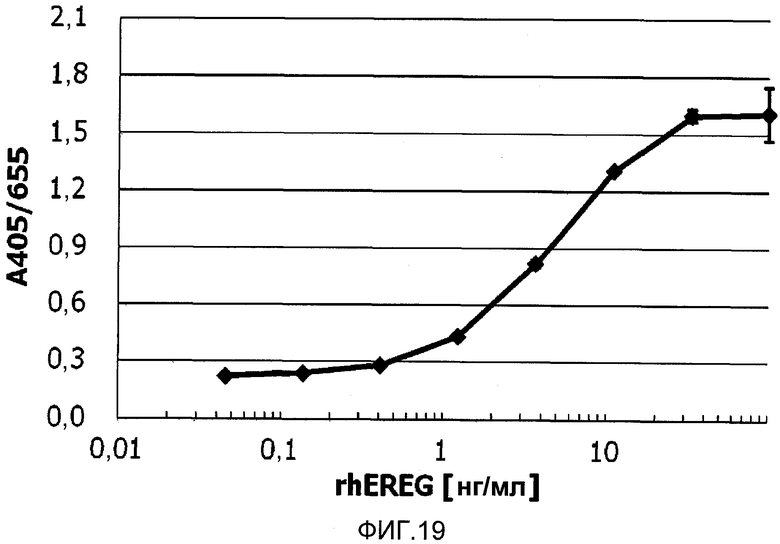


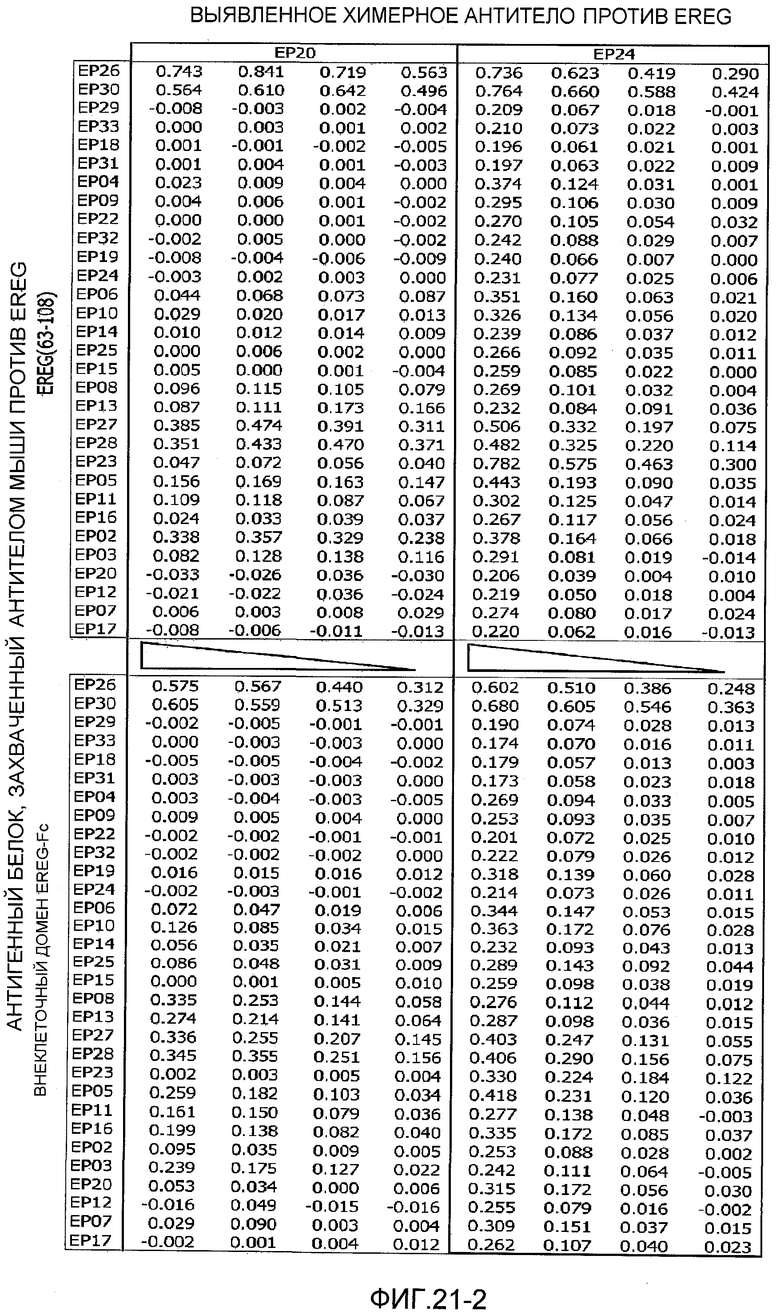

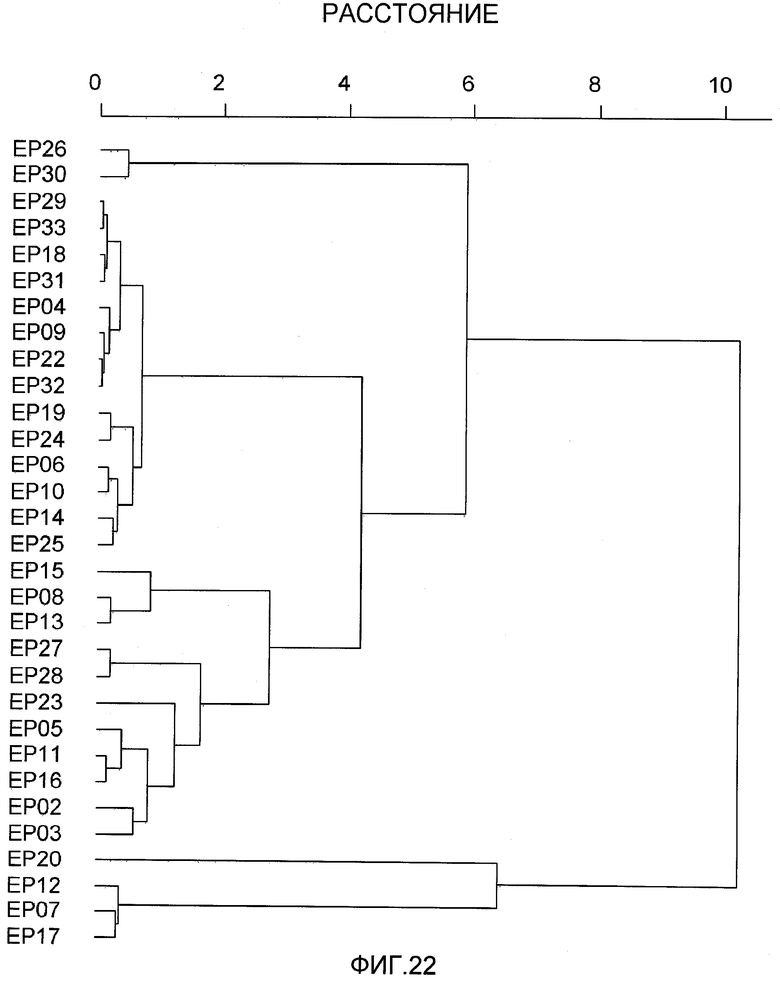
Группа изобретений основана на выявлении повышенной экспрессии гена и белка EREG при раке толстого кишечника, аденокарциноме легкого, раке поджелудочной железы, раке желудка и раке почки. Группа изобретений раскрывает антитела, которые распознают белок EREG, используют для лечения злокачественной опухоли. Группа изобретений относится к средствам и фармацевтическим композициям против злокачественной опухоли, содержащим в качестве активного ингредиента связывающее EREG антитело. Группа изобретений эффективна в лечении вышеуказанных опухолей. 2 н. и 10 з.п. ф-лы, 22 ил., 5 табл., 11 пр.
1. Средство против злокачественной опухоли, содержащее в качестве активного ингредиента антитело, которое связывается с белком EREG, где указанное антитело обладает ADCC или CDC активностью, где антитело, которое связывается с белком EREG, представляет собой антитело согласно любому из подпунктов (1)-(42), представленных ниже:
(1) антитело, содержащее Н-цепь, имеющую аминокислотную последовательность SEQ ID NO:2 в качестве CDR1, аминокислотную последовательность SEQ ID NO:4 в качестве CDR2 и аминокислотную последовательность SEQ ID NO:6 в качестве CDR3, и содержащее L-цепь, имеющую аминокислотную последовательность SEQ ID NO:12 в качестве CDR1, аминокислотную последовательность SEQ ID NO:14 в качестве CDR2 и аминокислотную последовательность SEQ ID NO:16 в качестве CDR3;
(2) антитело согласно (1), где Н-цепь имеет аминокислотную последовательность из положений 117-452 в аминокислотной последовательности SEQ ID NO:8 в качестве СН;
(3) антитело согласно (1), где Н-цепь имеет аминокислотную последовательность из положений 117-446 в аминокислотной последовательности SEQ ID NO:10 в качестве СН;
(4) антитело согласно (1), где L-цепь имеет аминокислотную последовательность из положений 107-213 в аминокислотной последовательности SEQ ID NO:18 в качестве CL;
(5) антитело согласно (1), где L-цепь имеет аминокислотную последовательность из положений 107-213 в аминокислотной последовательности SEQ ID NO:20 в качестве CL;
(6) антитело, содержащее Н-цепь согласно (2) и L-цепь согласно (4);
(7) антитело, содержащее Н-цепь согласно (3) и L-цепь согласно (5);
(8) антитело, содержащее Н-цепь, имеющую аминокислотную последовательность SEQ ID NO:49 в качестве CDR1, аминокислотную последовательность SEQ ID NO:51 в качестве CDR2 и аминокислотную последовательность SEQ ID NO:53 в качестве CDR3, и содержащее L-цепь, имеющую аминокислотную последовательность SEQ ID NO:55 в качестве CDR1, аминокислотную последовательность SEQ ID NO:57 в качестве CDR2 и аминокислотную последовательность SEQ ID NO:59 в качестве CDR3;
(9) антитело согласно (8), где Н-цепь имеет СН, полученную из мышиного IgG1;
(10) антитело согласно (8), где Н-цепь имеет аминокислотную последовательность из положений 117-446 в аминокислотной последовательности SEQ ID NO:10 в качестве СН;
(11) антитело согласно (8), где L-цепь имеет CL, полученную из мышиной κ-цепи;
(12) антитело согласно (8), где L-цепь имеет аминокислотную последовательность из положений 107-213 в аминокислотной последовательности SEQ ID NO:20 в качестве CL;
(13) антитело, содержащее Н-цепь согласно (9) и L-цепь согласно (11);
(14) антитело, содержащее Н-цепь согласно (10) и L-цепь согласно (12);
(15) антитело, содержащее Н-цепь, имеющую аминокислотную последовательность SEQ ID NO:61 в качестве CDR1, аминокислотную последовательность SEQ ID NO:63 в качестве CDR2 и аминокислотную последовательность SEQ ID NO:65 в качестве CDR3, и содержащее L-цепь, имеющую аминокислотную последовательность SEQ ID NO:67 в качестве CDR1, аминокислотную последовательность SEQ ID NO:69 в качестве CDR2 и аминокислотную последовательность SEQ ID NO:71 в качестве CDR3;
(16) антитело согласно (15), где Н-цепь имеет СН, полученную из мышиного IgG1;
(17) антитело согласно (15), где Н-цепь имеет аминокислотную последовательность из положений 117-446 в аминокислотной последовательности SEQ ID NO:10 в качестве СН;
(18) антитело согласно (15), где L-цепь имеет CL, полученную из мышиной κ-цепи;
(19) антитело согласно (15), где L-цепь имеет аминокислотную последовательность из положений 107-213 в аминокислотной последовательности SEQ ID NO:20 в качестве CL;
(20) антитело, содержащее Н-цепь согласно (16) и L-цепь согласно (18);
(21) антитело, содержащее Н-цепь согласно (17) и L-цепь согласно (19);
(22) антитело, содержащее Н-цепь, имеющую аминокислотную последовательность SEQ ID NO:73 в качестве CDR1, аминокислотную последовательность SEQ ID NO:75 в качестве CDR2 и аминокислотную последовательность SEQ ID NO:77 в качестве CDR3, и содержащее L-цепь, имеющую аминокислотную последовательность SEQ ID NO:79 в качестве CDR1, аминокислотную последовательность SEQ ID NO:81 в качестве CDR2, и аминокислотную последовательность SEQ ID NO:83 в качестве CDR3;
(23) антитело согласно (22), где Н-цепь имеет СН, полученную из мышиного IgG1;
(24) антитело согласно (22), где Н-цепь имеет аминокислотную последовательность из положений 117-446 в аминокислотной последовательности SEQ ID NO:10 в качестве СН;
(25) антитело согласно (22), где L-цепь имеет CL, полученную из мышиной κ-цепи;
(26) антитело согласно (22), где L-цепь имеет аминокислотную последовательность из положений 107-213 в аминокислотной последовательности SEQ ID NO:20 в качестве CL;
(27) антитело, содержащее Н-цепь согласно (23) и L-цепь согласно (25);
(28) антитело, содержащее Н-цепь согласно (24) и L-цепь согласно (26);
(29) антитело, содержащее Н-цепь, имеющую аминокислотную последовательность SEQ ID NO:85 в качестве CDR1, аминокислотную последовательность SEQ ID NO:87 в качестве CDR2 и аминокислотную последовательность SEQ ID NO:89 в качестве CDR3, и содержащее L-цепь, имеющую аминокислотную последовательность SEQ ID NO:91 в качестве CDR1, аминокислотную последовательность SEQ ID NO:93 в качестве CDR2 и аминокислотную последовательность SEQ ID NO:95 в качестве CDR3;
(30) антитело согласно (29), где Н-цепь имеет СН, полученную из мышиного IgG1;
(31) антитело согласно (29), где Н-цепь имеет аминокислотную последовательность из положений 117-446 в аминокислотной последовательности SEQ ID NO:10 в качестве СН;
(32) антитело согласно (29), где L-цепь имеет CL, полученную из мышиной κ-цепи;
(33) антитело согласно (29), где L-цепь имеет аминокислотную последовательность из положений 107-213 в аминокислотной последовательности SEQ ID NO:20 в качестве CL;
(34) антитело, содержащее Н-цепь согласно (30) и L-цепь согласно (32);
(35) антитело, содержащее Н-цепь согласно (31) и L-цепь согласно (33);
(36) антитело, содержащее Н-цепь, имеющую аминокислотную последовательность SEQ ID NO:97 в качестве CDR1, аминокислотную последовательность SEQ ID NO:99 в качестве CDR2 и аминокислотную последовательность SEQ ID NO:101 в качестве CDR3, и содержащее L-цепь, имеющую аминокислотную последовательность SEQ ID NO:103 в качестве CDR1, аминокислотную последовательность SEQ ID NO:105 в качестве CDR2 и аминокислотную последовательность SEQ ID NO:107 в качестве CDR3;
(37) антитело согласно (36), где Н-цепь имеет СН, полученную из мышиного IgG1;
(38) антитело согласно (36), где Н-цепь имеет аминокислотную последовательность из положений 117-446 в аминокислотной последовательности SEQ ID NO:10 в качестве СН;
(39) антитело согласно (36), где L-цепь имеет CL, полученную из мышиной κ-цепи;
(40) антитело согласно (36), где L-цепь имеет аминокислотную последовательность из положений 107-213 в аминокислотной последовательности SEQ ID NO:20 в качестве CL;
(41) антитело, содержащее Н-цепь согласно (37) и L-цепь согласно (39); и
(42) антитело, содержащее Н-цепь согласно (38) и L-цепь согласно (40).
2. Средство по п.1, которое дополнительно обладает нейтрализующей активностью против белка EREG.
3. Средство против злокачественной опухоли по п.1 или 2, где антитело, которое связывается с белком EREG, распознает участок от Ala в положении 29 до Ser в положении 69, или от Val в положении 63 до Leu в положении 108 в аминокислотной последовательности SEQ ID NO:21.
4. Средство против злокачественной опухоли по п.1 или 2, где злокачественная опухоль представляет собой любую злокачественную опухоль, выбранную из группы, состоящей из рака толстого кишечника, аденокарциномы легкого, рака поджелудочной железы, рака желудка и рака почки.
5. Средство против злокачественной опухоли по п.4, где злокачественная опухоль представляет собой первичную злокачественную опухоль.
6. Средство против злокачественной опухоли по п.4, где злокачественная опухоль представляет собой метастазирующую злокачественную опухоль.
7. Фармацевтическая композиция для лечения рака, содержащая в качестве активного агента антитело, которое связывается с белком EREG, где указанное антитело обладает ADCC или CDC активностью в отношении клеток, экспрессирующих белок EREG, для лечения рака,
где антитело представляет собой антитело согласно любому из подпунктов (1)-(42), представленных ниже:
(1) антитело, содержащее Н-цепь, имеющую аминокислотную последовательность SEQ ID NO:2 в качестве CDR1, аминокислотную последовательность SEQ ID NO:4 в качестве CDR2 и аминокислотную последовательность SEQ ID NO:6 в качестве CDR3, и содержащее L-цепь, имеющую аминокислотную последовательность SEQ ID NO:12 в качестве CDR1, аминокислотную последовательность SEQ ID NO:14 в качестве CDR2 и аминокислотную последовательность SEQ ID NO:16 в качестве CDR3;
(2) антитело согласно (1), где Н-цепь имеет аминокислотную последовательность из положений 117-452 в аминокислотной последовательности SEQ ID NO:8 в качестве СН;
(3) антитело согласно (1), где Н-цепь имеет аминокислотную последовательность из положений 117-446 в аминокислотной последовательности SEQ ID NO:10 в качестве СН;
(4) антитело согласно (1), где L-цепь имеет аминокислотную последовательность из положений 107-213 в аминокислотной последовательности SEQ ID NO:18 в качестве CL;
(5) антитело согласно (1), где L-цепь имеет аминокислотную последовательность из положений 107-213 в аминокислотной последовательности SEQ ID NO:20 в качестве CL;
(6) антитело, содержащее Н-цепь согласно (2) и L-цепь согласно (4);
(7) антитело, содержащее Н-цепь согласно (3) и L-цепь согласно (5);
(8) антитело, содержащее Н-цепь, имеющую аминокислотную последовательность SEQ ID NO:49 в качестве CDR1, аминокислотную последовательность SEQ ID NO:51 в качестве CDR2 и аминокислотную последовательность SEQ ID NO:53 в качестве CDR3, и содержащее L-цепь, имеющую аминокислотную последовательность SEQ ID NO:55 в качестве CDR1, аминокислотную последовательность SEQ ID NO:57 в качестве CDR2 и аминокислотную последовательность SEQ ID NO:59 в качестве CDR3;
(9) антитело согласно (8), где Н-цепь имеет СН, полученную из мышиного IgG1;
(10) антитело согласно (8), где Н-цепь имеет аминокислотную последовательность из положений 117-446 в аминокислотной последовательности SEQ ID NO:10 в качестве СН;
(11) антитело согласно (8), где L-цепь имеет CL, полученную из мышиной κ-цепи;
(12) антитело согласно (8), где L-цепь имеет аминокислотную последовательность из положений 107-213 в аминокислотной последовательности SEQ ID NO:20 в качестве CL;
(13) антитело, содержащее Н-цепь согласно (9) и L-цепь согласно (11);
(14) антитело, содержащее Н-цепь согласно (10) и L-цепь согласно (12);
(15) антитело, содержащее Н-цепь, имеющую аминокислотную последовательность SEQ ID NO:61 в качестве CDR1, аминокислотную последовательность SEQ ID NO:63 в качестве CDR2 и аминокислотную последовательность SEQ ID NO:65 в качестве CDR3, и содержащее L-цепь, имеющую аминокислотную последовательность SEQ ID NO:67 в качестве CDR1, аминокислотную последовательность SEQ ID NO:69 в качестве CDR2 и аминокислотную последовательность SEQ ID NO:71 в качестве CDR3;
(16) антитело согласно (15), где Н-цепь имеет СН, полученную из мышиного IgG1;
(17) антитело согласно (15), где Н-цепь имеет аминокислотную последовательность из положений 117-446 в аминокислотной последовательности SEQ ID NO:10 в качестве СН;
(18) антитело согласно (15), где L-цепь имеет CL, полученную из мышиной κ-цепи;
(19) антитело согласно (15), где L-цепь имеет аминокислотную последовательность из положений 107-213 в аминокислотной последовательности SEQ ID NO:20 в качестве CL;
(20) антитело, содержащее Н-цепь согласно (16) и L-цепь согласно (18);
(21) антитело, содержащее Н-цепь согласно (17) и L-цепь согласно (19);
(22) антитело, содержащее Н-цепь, имеющую аминокислотную последовательность SEQ ID NO:73 в качестве CDR1, аминокислотную последовательность SEQ ID NO:75 в качестве CDR2 и аминокислотную последовательность SEQ ID NO:77 в качестве CDR3, и содержащее L-цепь, имеющую аминокислотную последовательность SEQ ID NO:79 в качестве CDR1, аминокислотную последовательность SEQ ID NO:81 в качестве CDR2, и аминокислотную последовательность SEQ ID NO:83 в качестве CDR3;
(23) антитело согласно (22), где Н-цепь имеет СН, полученную из мышиного IgG1;
(24) антитело согласно (22), где Н-цепь имеет аминокислотную последовательность из положений 117-446 в аминокислотной последовательности SEQ ID NO:10 в качестве СН;
(25) антитело согласно (22), где L-цепь имеет CL, полученную из мышиной κ-цепи;
(26) антитело согласно (22), где L-цепь имеет аминокислотную последовательность из положений 107-213 в аминокислотной последовательности SEQ ID NO:20 в качестве CL;
(27) антитело, содержащее Н-цепь согласно (23) и L-цепь согласно (25);
(28) антитело, содержащее Н-цепь согласно (24) и L-цепь согласно (26);
(29) антитело, содержащее Н-цепь, имеющую аминокислотную последовательность SEQ ID NO:85 в качестве CDR1, аминокислотную последовательность SEQ ID NO:87 в качестве CDR2 и аминокислотную последовательность SEQ ID NO:89 в качестве CDR3, и содержащее L-цепь, имеющую аминокислотную последовательность SEQ ID NO:91 в качестве CDR1, аминокислотную последовательность SEQ ID NO:93 в качестве CDR2 и аминокислотную последовательность SEQ ID NO:95 в качестве CDR3;
(30) антитело согласно (29), где Н-цепь имеет СН, полученную из мышиного IgG1;
(31) антитело согласно (29), где Н-цепь имеет аминокислотную последовательность из положений 117-446 в аминокислотной последовательности SEQ ID NO:10 в качестве СН;
(32) антитело согласно (29), где L-цепь имеет CL, полученную из мышиной κ-цепи;
(33) антитело согласно (29), где L-цепь имеет аминокислотную последовательность из положений 107-213 в аминокислотной последовательности SEQ ID NO:20 в качестве CL;
(34) антитело, содержащее Н-цепь согласно (30) и L-цепь согласно (32);
(35) антитело, содержащее Н-цепь согласно (31) и L-цепь согласно (33);
(36) антитело, содержащее Н-цепь, имеющую аминокислотную последовательность SEQ ID NO:97 в качестве CDR1, аминокислотную последовательность SEQ ID NO:99 в качестве CDR2 и аминокислотную последовательность SEQ ID NO:101 в качестве CDR3, и содержащее L-цепь, имеющую аминокислотную последовательность SEQ ID NO:103 в качестве CDR1, аминокислотную последовательность SEQ ID NO:105 в качестве CDR2 и аминокислотную последовательность SEQ ID NO:107 в качестве CDR3;
(37) антитело согласно (36), где Н-цепь имеет СН, полученную из мышиного IgG1;
(38) антитело согласно (36), где Н-цепь имеет аминокислотную последовательность из положений 117-446 в аминокислотной последовательности SEQ ID NO:10 в качестве СН;
(39) антитело согласно (36), где L-цепь имеет CL, полученную из мышиной κ-цепи;
(40) антитело согласно (36), где L-цепь имеет аминокислотную последовательность из положений 107-213 в аминокислотной последовательности SEQ ID NO:20 в качестве CL;
(41) антитело, содержащее Н-цепь согласно (37) и L-цепь согласно (39); и
(42) антитело, содержащее Н-цепь согласно (38) и L-цепь согласно (40).
8. Фармацевтическая композиция по п.7, которая дополнительно обладает нейтрализующей активностью против белка EREG.
9. Фармацевтическая композиция по п.7 или 8, где антитело распознает участок от Ala в положении 29 до Ser в положении 69, или от Val в положении 63 до Leu в положении 108 в аминокислотной последовательности SEQ ID NO:21.
10. Фармацевтическая композиция по п.7 или 8, где злокачественная опухоль представляет собой любую злокачественную опухоль, выбранную из группы, состоящей из рака толстого кишечника, аденокарциномы легкого, рака поджелудочной железы, рака желудка и рака почки.
11. Фармацевтическая композиция по п.10, где злокачественная опухоль представляет собой первичную злокачественную опухоль.
12. Фармацевтическая композиция по п.10, где злокачественная опухоль представляет собой метастазирующую злокачественную опухоль.
| EP13505211 A1, 08.10.2003 |

