Настоящее изобретение относится к полиспецифичным, особенно биспецифичным антителам, включающим антитела полной длины и одноцепочечные фрагменты Fab, к способам их получения, фармацевтическим композициям, содержащим указанные антитела, и к их применению.
Предпосылки создания изобретения
В недавнем прошлом разработан широкий спектр форматов полиспецифичных рекомбинантных антител, например, четырехвалентных биспецифичных антител, полученных путем слияния, например, антител формата IgG и одноцепочечных доменов (см., например, M.J.Coloma и др., Nature Biotech, 15, 1997, cc.159-163, WO 2001/077342 и S.L.Morrison, Nature Biotech, 25,2007, cc.1233-1234).
Также разработано несколько других новых форматов, в которых уже не сохраняется структура ядра антитела (IgA, IgD, IgE, IgG или IgM), например, ди-, три- или тетратела, минитела, несколько одноцепочечных форматов (scFv, Bis-scFv), которые способны к связыванию с двумя или несколькими антигенами (Р.Holliger и др., Nature Biotech, 23, 2005, cc. 1126-1136, N.Fischer, O. Léger, Pathobiology, 74, 2007, cc.3-14, J.Shen и др., Journal of Immunological Methods, 318, 2007, cc.65-74, С.Wu и др., Nature Biotech., 25, 2007, cc.1290-1297).
Во всех таких форматах используют линкеры, либо для слияния ядра антитела (IgA, IgD, IgE, IgG и IgM) с дополнительным связывающим белком (например, ScFv), либо для слияния, например, двух Фрагментов Fab или scFv (N.Fischer, О.Léger, Pathobiology, 74, 2007, cc.3-14). Следует учитывать, что возможно сохранение эффекторных функций, например, комплементзависимой цитотоксичности (complement dependent cytotoxicity - CDC) или антителозависимой клеточной цитотоксичности (antibody dependent cellular cytotoxicity - ADCC), которые опосредованы через связывание с рецептором Fc, путем поддержания высокой степени сходства с природными антителами.
В WO 2007/024715 описывают иммуноглобулины с двойственными вариабельными доменами, сконструированными в виде поливалентных и полиспецифичных связывающих белках. Способ получения димеров антител биологического действия описан в US 6897044. Поливалентная конструкция фрагмента антитела Fy, имеющая по меньшей мере четыре вариабельных домена, которые связаны друг с другом посредством пептидных линкеров, описана в US 7129330. Димерные и полимерные антигенсвязывающие структуры описаны в US 2005/0079170. В US 6511663 сообщают о трех- или четырехвалентном моноспецифичном антигенсвязывающем белке, включающем три или четыре фрагмента Fab, ковалентно связанных друг с другом через связывающую структуру, при этом белок не является природным иммуноглобулином. В WO 2006/020258 сообщают о четырехвалентных биспецифичных антителах, которые могут эффективно экспрессироваться в прокариотических и эукариотических клетках и которые полезны в терапевтических и диагностических методах. Метод разделения или предпочтительно синтеза димеров, которые связаны с помощью по меньшей мере одной межцепочечной дисульфидной связи, от димеров, которые не связаны с помощью по меньшей мере одной межцепочечной дисульфидной связи, в смеси, включающей два типа полипептидных димеров, сообщают в US 2005/0163782. В US 5959083 сообщают о биспецифичных четырехвалентных рецепторах. В WO 2001/077342 C сообщают о создании антител с тремя и более функциональными антигенсвязывающими участками в WO 2001/077342.
Полиспецифичные и поливалентные антигенсвязывающие полипептиды описаны в WO 1997/001580. В WO 1992/004053 сообщают о гомоконъюгатах, обычно получаемых из моноклональных антител класса IgG, которые связываются с одной и той же антигенной детерминантой и которые ковалентно связаны посредством синтетических перекрестных сшивок. В заявке WO 1991/06305 сообщают об олигомерных моноклональных антителах с высокой авидностью к антигену, причем секретируются олигомеры, обычно класса IgG, с двумя или более мономерами иммуноглобулина, связаны вместе для формирования четырехвалентных или шестивалентных молекул IgG. В US 6350860 сообщают об антителах, производных от овец, и о сконструированных конструкциях антител, которые можно применять для лечения заболеваний, в которых активность интерферона гамма является патогенной. В US 2005/0100543 сообщают о конструкциях, нацеливаемых на мишень, которые являются поливалентными носителями биспецифичных антител, т.е. каждая молекула конструкции, нацеливаемой на мишень, может служить в качестве носителя двух или нескольких биспецифичных антител. В WO 1995/009917 сообщают о генно-инженерных биспецифичных четырехвалентных антителах. В WO 2007/109254 сообщают о стабилизированных связывающих молекулах, которые состоят из стабилизированного фрагмента scFv или включают его.
В кн.: D.Müller и др. «Handbook of Therapeutic antibodies», 2008, ч. III, гл. 2, cc.345-378, сообщают о биспецифичных антителах, например, об антителе полной длины, к которому посредством пептидного линкера на С-конце тяжелой цепи присоединены два фрагмента scFv (см. также WO 1995/009917). В работе М.Hust и др., ВМС Biotechnology, 2007, с.7, сообщают об одноцепочечных фрагментах Fab (scFab).
Поскольку существуют различные проблемы и задачи, связанные с полиспецифичными антителами (например, фармакокинетические и биологические свойства, стабильность, агрегация, выход экспрессии), имеется необходимость в дальнейшей разработке других форматов полиспецифичных антител. Например, генно-инженерные биспецифичные четырехвалентные антитела, описанные в WO 1995/009917 и в кн.: D. Müller и др. «Handbook of Therapeutic antibodies», 2008, ч.III, гл.2, cc.345-378, показали слишком низкие выходы экспрессии.
Краткое описание изобретения
Первым объектом настоящего изобретения является полиспецифичное антитело, включающее
а) антитело полной длины, специфически связывающееся с первым антигеном и состоящее из двух тяжелых цепей антитела и двух легких цепей антитела, и
б) один или несколько одноцепочечных фрагментов Fab, специфически связывающихся с одним или до четырех дополнительных антигенов (предпочтительно специфически связывающихся с одним дополнительным антигеном),
в котором указанные одноцепочечные фрагменты Fab по пункту б) гибридизированы с указанным антителом полной длины по пункту а) посредством пептидного коннектора с С- или N-конца тяжелой или легкой цепи указанного антитела полной длины.
Предпочтительным объектом настоящего изобретения является полиспецифичное антитело, включающее
а) антитело полной длины, специфически связывающееся с первым антигеном и состоящее из двух тяжелых цепей антитела и двух легких цепей антитела, и
б) от одного до четырех одноцепочечных фрагментов Fab, специфически связывающихся с одним или до четырех дополнительных антигенов (предпочтительно специфически связывающихся с одним дополнительным антигеном),
в котором указанные одноцепочечные фрагменты Fab по пункту б) гибридизированы с указанным антителом полной длины по пункту а) посредством пептидного коннектора с С- или N-конца тяжелой или легкой цепи указанного антитела полной длины.
Предпочтительно указанное полиспецифичное антитело включает один или два одноцепочечных фрагмента Fab, связывающихся со вторым антигеном (биспецифичное антитело).
Предпочтительно указанное полиспецифичное антитело включает два одноцепочечных фрагмента Fab, связывающихся со вторым антигеном (биспецифичное антитело).
Предпочтительно указанное полиспецифичное антитело включает два одноцепочечных фрагмента Fab, связывающихся со вторым антигеном и третьим антигеном (трехспецифичное антитело).
Другим объектом настоящего изобретения является молекула нуклеиновой кислоты, кодирующая указанное полиспецифичное антитело, в котором одноцепочечный фрагмент Fab гибридизирован с С- или N-концом тяжелой или легкой цепи указанного антитела полной длины.
Кроме того, дополнительным объектом настоящего изобретения является фармацевтическая композиция, включающая указанное полиспецифичное антитело.
Полиспецифичные антитела по настоящему изобретению показали ценные свойства, например, высокую стабильность, низкую тенденцию к агрегации (см., например, пример 2), например, по сравнению с антителом полной длины, к которому присоединены два фрагмента scFv посредством пептидного линкера с С-конца тяжелой цепи (см. WO 1995/009917 или кн.: D.Müller и др. «Handbook of Therapeutic antibodies», 2008, ч.III, гл.2, c. 345-378).
С одной стороны полиспецифичные антитела по настоящему изобретению показывают новые свойства из-за способности связываться с другими антигенами, а с другой стороны они применимы для получения и технологии приготовления лекарственного средства, благодаря их хорошей стабильности, низкой агрегации и ценным фармакокинетическим и биологическим свойствам. Благодаря ядру Ig и способности вырабатываться в экспрессирующих системах млекопитающих, они все же сохраняют свойства природных антител, например ADCC и CDC.
Подробное описание изобретения
Первым объектом настоящего изобретения является полиспецифичное антитело, включающее
а) антитело полной длины, специфически связывающееся с первым антигеном и состоящее из двух тяжелых цепей антитела и двух легких цепей антитела, и
б) один или несколько одноцепочечных фрагментов Fab, специфически связывающихся с одним или до четырех дополнительных антигенов (предпочтительно специфически связывающихся с одним дополнительным антигеном),
в котором указанные одноцепочечные фрагменты Fab по пункту б) соединены с указанным антителом полной длины по пункту а) посредством пептидного коннектора с С- или N-конца тяжелой или легкой цепи указанного антитела полной длины.
Предпочтительным объектом настоящего изобретения является полиспецифичное антитело, включающее
а) антитело полной длины, специфически связывающееся с первым антигеном и состоящее из двух тяжелых цепей антитела и двух легких цепей антитела, и
б) от одного до четырех одноцепочечных фрагментов Fab, специфически связывающихся с одним или до четырех дополнительных антигенов (предпочтительно специфически связывающихся с одним дополнительным антигеном),
в котором указанные одноцепочечные фрагменты Fab по пункту б) гибридизированы с указанным антителом полной длины по пункту а) посредством пептидного коннектора с С- или N-конца тяжелой или легкой цепи указанного антитела полной длины.
В одном варианте осуществления настоящего изобретения один или два идентичных одноцепочечных фрагмента Fab, связывающихся со вторым антигеном, соединены с указанным антителом полной длины посредством пептидного коннектора с С-конца тяжелых или легких цепей указанного антитела полной длины.
В одном варианте осуществления настоящего изобретения один или два идентичных одноцепочечных фрагмента Fab, связывающихся со вторым антигеном, соединены с указанным антителом полной длины посредством пептидного коннектора с С-конца тяжелых цепей указанного антитела полной длины.
В одном варианте осуществления настоящего изобретения один или два идентичных одноцепочечных фрагмента Fab, связывающихся со вторым антигеном, соединены с указанным антителом полной длины посредством пептидного коннектора с С-конца легких цепей указанного антитела полной длины.
В одном варианте осуществления настоящего изобретения два идентичных одноцепочечных фрагмента Fab, связывающихся со вторым антигеном, соединены с указанным антителом полной длины посредством пептидного коннектора с С-конца каждой тяжелой или легкой цепи указанного антитела полной длины.
В одном варианте осуществления настоящего изобретения два идентичных одноцепочечных фрагмента Fab, связывающихся со вторым антигеном, гибридизированы с указанным антителом полной длины посредством пептидного коннектора с С-конца каждой тяжелой цепи указанного антитела полной длины.
В одном варианте осуществления настоящего изобретения два идентичных одноцепочечных фрагмента Fab, связывающихся со вторым антигеном, гибридизированы с указанным антителом полной длины посредством пептидного коннектора с С-конца каждой легкой цепи указанного антитела полной длины.
Понятие «антитело полной длины» означает антитело, состоящее из двух «тяжелых цепей антитела полной длины» и двух «легких цепей антитела полной длины» (см. фиг.1). «Тяжелая цепь антитела полной длины» представляет собой полипептид, состоящий в направлении от N-конца к С-концу из вариабельного домена тяжелой цепи антитела (VH), константного домена 1 тяжелой цепи антитела (СН1), шарнирной области антитела (HR), константного домена 2 тяжелой цепи антитела (СН2) и константного домена 3 тяжелой цепи антитела (СН3), сокращенно обозначаемый VH-CH1-HR-CH2-CH3, и необязательно константного домена 4 тяжелой цепи антитела (СН4) в случае антитела подкласса IgE. Предпочтительно «тяжелая цепь антитела полной длины» является полипептидом, состоящим в направлении от N-конца к С-концу из VH, СН1, HR, CH2 и СН3. «Легкая цепь антитела полной длины» представляет собой полипептид, состоящий в направлении от N-конца к С-концу из вариабельного домена легкой цепи антитела (VL) и константного домена легкой цепи антитела (CL), сокращенно обозначаемый VL-CL. Константный домен легкой цепи антитела (CL) может быть доменом к (каппа) или λ (лямбда). Две цепи антитела полной длины связаны вместе посредством межполипептидных дисульфидных связей между CL доменом и СН1 доменом, и между шарнирными областями тяжелых цепей антитела полной длины. Примерами типичных антител полной длины являются природные антитела, например IgG (например, IgG 1 или IgG2), IgM, IgA, IgD и IgE. Антитела полной длины по настоящему изобретению могут происходить от одного вида, например человека, или они могут являться химерными или гуманизированными антителами. Антитела полной длины по настоящему изобретению включают два сайта связывания антигена, каждый из которых образован парой VH и VL, причем оба специфически связываются с одним и тем же антигеном. С-конец тяжелой или легкой цепи указанного антитела полной длины означает последнюю аминокислоту на С-конце указанной тяжелой или легкой цепи. N-конец тяжелой или легкой цепи указанного антитела полной длины означает последнюю аминокислоту на N-конце указанной тяжелой или легкой цепи.
«Одноцепочечный фрагмент Fab» (см. фиг.2) представляет полипептид, состоящий из вариабельного домена тяжелой цепи антитела (VH), константного домена 1 антитела (СН1), вариабельного домена легкой цепи антитела (VL), константного домена легкой цепи антитела (CL) и линкера, в котором указанные домены антитела и указанный линкер имеют одну из следующих последовательностей в направлении от N-конца к С-концу:
a) VH-CH1-линкер-VL-CL, б) VL-CL-линкер-VH-CH1, в) VH-CL-линкер-VL-CH1 или г) VL-CH1-линкер-VH-CL, и в котором указанный линкер представляет полипептид, состоящий по меньшей мере из 30 аминокислот, предпочтительно из 32-50 аминокислот. Указанные одноцепочечные фрагменты Fab a) VH-CH1-линкер-VL-CL, б) VL-CL-линкер-VH-CH1, в) VH-CL-линкер-VL-СН1 и г) VL-CH1-линкер-VH-CL стабилизированы посредством природной дисульфидной связи между доменом CL и доменом СН1. Понятие «N-конец» означает последнюю аминокислоту N-конца. Понятие «С-конец» означает последнюю аминокислоту С-конца.
В предпочтительном варианте осуществления настоящего изобретения указанные домены антитела и указанный линкер в указанном одноцепочечном фрагменте Fab имеют одну из следующих последовательностей в направлении от N-конца к С-концу: a) VH-CH1-линкер-VL-CL или б) VL-CL-линкер-VH-СН1, более предпочтительно VL-CL-линкер-VH-CH1.
В другом предпочтительном варианте осуществления настоящего изобретения указанные домены антитела и указанный линкер в указанном одноцепочечном фрагменте Fab имеют одну из следующих последовательностей в направлении от N-конца к С-концу: а) VH-CL-линкер-VL-СН1 или б) VL-CH1-линкер-VH-CL.
Необязательно в указанном одноцепочечном фрагменте Fab, помимо природной дисульфидной связи между доменом CL и доменом СН1, также вариабельный домен тяжелой цепи антитела (VH) и вариабельный домен легкой цепи антитела (VL) стабилизированы дисульфидом посредством введения дисульфидной связи между следующими положениями:
i) положением 44 вариабельного домена тяжелой цепи и положением 100 вариабельного домена легкой цепи,
ii) положением 105 вариабельного домена тяжелой цепи и положением 43 вариабельного домена легкой цепи или
iii) положением 101 вариабельного домена тяжелой цепи и положением 100 вариабельного домена легкой цепи, (нумерация всегда соответствует индексу ЕС по К-abat).
Такая дополнительная дисульфидная стабилизация одноцепочечных фрагментов Fab достигается посредством введения дисульфидной связи между вариабельными доменами VH и VL одноцепочечных фрагментов Fab. Методы введения неприродных дисульфидных мостиков с целью стабилизации одноцепочечного фрагмента Fv описаны, например, в WO 94/029350, V.Rajagopal и др., Prot. Engin., 10, 1997, c.1453-1459, H.Kobayashi и др., Nuclear Medicine & Biology, 25, 1998, c.387-393 или М.Schmidt и др., Oncogene, 18, 1999, cc.1711-1721. В одном из вариантов осуществления настоящего изобретения необязательная дисульфидная связь между вариабельными доменами одноцепочечных фрагментов Fab, включенных в антитело по настоящему изобретению, находится между положением 44 вариабельного домена тяжелой цепи и положением 100 вариабельного домена легкой цепи. В одном из вариантов осуществления настоящего изобретения необязательная дисульфидная связь между вариабельными доменами одноцепочечных фрагментов Fab, включенных в антитело по настоящему изобретению, находится между положением 105 вариабельного домена тяжелой цепи и положением 43 вариабельного домена легкой цепи (нумерация всегда соответствует индексу ЕС по Kabat).
В одном из вариантов осуществления настоящего изобретения предпочтительны одноцепочечные фрагменты Fab без указанной необязательной дисульфидной стабилизацией между вариабельными доменами VH и VL одноцепочечных фрагментов Fab.
Понятие «пептидный коннектор» в контексте настоящего изобретения означает пептид с аминокислотной последовательностью, предпочтительно синтетического происхождения. Эти пептидные коннекторы по настоящему изобретению используют для присоединения одноцепочечных фрагментов Fab к С- или N-концу антитела полной длины для формирования полиспецифичного антитела по настоящему изобретению. Предпочтительно указанные пептидные коннекторы по пункту б) являются пептидами с аминокислотной последовательностью, длина которой состоит по меньшей мере из 5 аминокислот, предпочтительно длиной по меньшей мере из 5-100, более предпочтительно из 10-50 аминокислот. В одном варианте осуществления настоящего изобретения указанным пептидным коннектором является (GxS)n или (GxS)nGm, где G = глицин, S = серин и (х=3, n=3, 4, 5 или 6, и m=0, 1, 2 или 3) или (х=4, n=2, 3, 4 или 5, и m=0, 1, 2 или 3), предпочтительно х=4 и n=2 или 3, более предпочтительно х=4, n=2. В одном варианте осуществления настоящего изобретения указанным пептидным коннектором является (G4S)2.
Понятие «линкер» в контексте настоящего изобретения означает пептид с аминокислотными последовательностями, предпочтительно синтетического происхождения. Эти пептиды по настоящему изобретению используют для связывания a) VH-CH1 с VL-CL, б) VL-CL с VH-CH1, в) VH-CL с VL-CH1 или г) VL-CH1 с VH-CL для образования следующих одноцепочечных фрагментов Fab по настоящему изобретению а) VH-CH1-линкер-VL-CL, б) VL-CL-линкер-VH-СН1, в) VH-CL-линкер-VL-СН1 или г) VL-СН1-линкер-VH-CL. Указанный линкер в одноцепочечных фрагментах Fab является пептидом с аминокислотной последовательностью, длина которой состоит по меньшей мере из 30 аминокислот, предпочтительно длиной из 32-50 аминокислот. В одном варианте осуществления настоящего изобретения указанным линкером является (GxS)n, где G = глицин, S = серин, (х=3, n=8, 9 или 10 и m=0, 1, 2 или 3) или (х=4 и n=6, 7 или 8 и m=0, 1, 2 или 3), предпочтительно х=4, n=6 или 7 и m=0, 1, 2 или 3, более предпочтительно х=4, n=7 и m=2. В одном варианте осуществления настоящего изобретения указанным линкером является (G4S)6G2.
К каждому С- или N-концу тяжелой или легкой цепи указанного антитела полной длины одновременно можно присоединить только один из одноцепочечных фрагментов Fab по пункту б). Таким образом, с указанным антителом полной длины можно гибридизировать до восьми одноцепочечных фрагментов Fab. Предпочтительно полиспецифичное антитело по настоящему изобретению включает от одного до четырех одноцепочечных фрагментов Fab. Более предпочтительно полиспецифичное антитело по настоящему изобретению включает два идентичных одноцепочечных фрагмента Fab (предпочтительно VL-CL-линкер-VH-СН1), оба из которых присоединены к двум С-концам двух тяжелых цепей или к двум С-концам двух легких цепей указанного антитела полной длины по пункту а). Такая гибридизация приводит к двум идентичным гибридным пептидам [либо i) тяжелой цепи и одноцепочечного фрагмента Fab, либо ii) легкой цепи и одноцепочечного Фрагмента Fab], которые совместно экспрессируются либо с i) легкой цепи, либо с тяжелой цепи антитела полной длины с образованием полиспецифичного антитела по настоящему изобретению (см. фиг.3, 4 и 5).
В другом предпочтительном варианте осуществления настоящего изобретения полиспецифичное антитело по настоящему изобретению включает два идентичных одноцепочечных фрагмента Fab (предпочтительно VH-CH1-линкер-VL-CL), оба из которых присоединены к двум N-концам двух тяжелых цепей или к двум N-концам двух легких цепей указанного антитела полной длины по пункту а). Такая гибридизация приводит к двум идентичным гибридным пептидам (либо i) тяжелой цепи и одноцепочечного фрагмента Fab, либо ii) легкой цепи и одноцепочечного фрагмента Fab), которые совместно экспрессируются либо с i) легкой цепи, либо с тяжелой цепи антитела полной длины с образованием полиспецифичного антитела по настоящему изобретению.
Обе части полиспецифичного антитела по настоящему изобретению включают сайты связывания антигена (антитело полной длины по настоящему изобретению включает два, и каждый одноцепочечный Fab-фрагмент включает один сайт связывания антигена). Понятия «сайт связывания» или «сайт связывания антигена» в контексте настоящего изобретения означают область (области) указанного полиспецифичного антитела по настоящему изобретению, с которыми действительно специфически связывается соответствующий антиген. Каждый из сайтов связывания антигена, или в антителе полной длины, или в одноцепочечном фрагменте Fab, сформирован парой, состоящей из вариабельного домена легкой цепи антитела (VL) и вариабельного домена тяжелой цепи антитела (VH).
Сайты связывания антигена, которые специфически связываются с требуемым антигеном (например, EGFR), могут быть получены а) из известных антител к антигену (например, анти-EGFR антител), или б) из новых антител или фрагментов антител, вновь полученных посредством методов иммунизации, используя среди прочего или антигенный белок, или нуклеиновую кислоту, или их фрагменты, или посредством фагового дисплея.
Сайт связывания антигена у антитела по настоящему изобретению содержит шесть комплементарность-детерминирующих областей (complementary determining region - CDR), которые влияют на варьирующую степень сродства сайта связывания с антигеном. Имеется три области CDR вариабельного домена тяжелой цепи (CDRH1, CDRH2 и CDRH3) и три области CDR вариабельного домена легкой цепи (CDRL1, CDRL2 и CDRL3). Протяженность CDR и каркасных участков (framework regions - FR) определяют путем сравнения с аминокислотными последовательностями сводной базы данных, в которых эти участки установлены по вариабельности среди последовательностей.
Специфичность антитела относится к селективному распознаванию антителом конкретного эпитопа антигена. Природные антитела, например, являются моноспецифичными. Понятие «полиспецифичное» антитело в контексте настоящего изобретения означает антитело, которое имеет два или несколько сайтов связывания антигена, из которых по меньшей мере два связываются с различными антигенами или различными эпитопами одного и того же антигена. «Биспецифичными антителами» по настоящему изобретению являются антитела, которые обладают двумя различными антигенсвязывающими специфичностями. Антитела настоящего изобретения являются, например, полиспецифичными по отношению, по меньшей мере, к двум различным антигенам, т.е. EGFR в качестве первого антигена и IGF-1R в качестве второго антигена. В одном варианте осуществления настоящего изобретения полиспецифичное антитело по настоящему изобретению является биспецифичным. В другом варианте осуществления настоящего изобретения полиспецифичное антитело по настоящему изобретению является трехспецифичным.
Понятие «моноспецифичное» антитело в контексте настоящего изобретения означает антитело, которое имеет один или несколько сайтов связывания, каждый из которых связывается с одним и тем же эпитопом одного и того же антигена.
Понятие «валентный» в контексте настоящей заявки означает присутствие конкретного числа сайтов связывания в молекуле антитела. Например, природное антитело или антитело полной длины по настоящему изобретению имеет два сайта связывания и является двухвалентным. Таким образом, понятия «трехвалентное», «четырехвалентное», «пятивалентное» и «шестивалентное» относятся к присутствию двух сайтов связывания, трех сайтов связывания, четырех сайтов связывания, пяти сайтов связывания и шести сайтов связывания, соответственно, в молекуле антитела. Полиспецифичные антитела по настоящему изобретению являются по меньшей мере «трехвалентными». Предпочтительно они являются «трехвалентными», «четырехвалентными», «пятивалентными» или «шестивалентными», более предпочтительно они являются «трехвалентными» или «четырехвалентными».
Антитела по настоящему изобретению имеют три или более сайтов связывания и являются полиспецифичными, предпочтительно биспецифичными или трехспецифичными. Поскольку полиспецифичные антитела по настоящему изобретению могут быть биспецифичными даже в тех случаях, когда присутствует более трех сайтов связывания (т.е. когда антитело является четырехвалентным, пятивалентным, шестивалентным или поливалентным). У антител более чем с двумя сайтами связывания антигена, некоторые сайты связывания могут быть идентичными, при этом белок имеет сайты связывания для двух разных антигенов.
Другим вариантом осуществления настоящего изобретения является полиспецифичное антитело, включающее
а) антитело полной длины, специфически связывающееся с первым антигеном и состоящее из:
аа) двух идентичных тяжелых цепей антитела, состоящих в направлении от N-конца к С-концу из вариабельного домена тяжелой цепи антитела (VH), константного домена тяжелой цепи антитела 1 (СН1), шарнирного участка антитела (HR), константного домена тяжелой цепи антитела 2 (СН2) и константного домена тяжелой цепи антитела 3 (CH3), и
аб) двух идентичных легких цепей антитела, состоящих в направлении от N-конца к С-концу из вариабельного домена легкой цепи антитела (VL) и константного домена легкой цепи антитела (CL) (VL-CL), и
б) от одного до четырех одноцепочечных фрагментов Fab специфически связывающихся с одним или до четырех дополнительных антигенов (предпочтительно специфически связывающихся с одним дополнительным антигеном), в котором одноцепочечные фрагменты Fab состоят из вариабельного домена тяжелой цепи антитела (VH), константного домена антитела 1 (СН1), вариабельного домена легкой цепи антитела (VL), константного домена легкой цепи антитела (CL) и линкера, и в котором указанные домены антитела и указанный линкер имеют одну из следующих последовательностей в направлении от N-конца к С-концу:
ба) VH-CH1-линкер-VL-CL, бб) VL-CL-линкер-VH-CH1, бв) VH-CL-линкер-VL-CH1 или бг) VL-CH1-линкер-VH-CL,
в котором указанный линкер является пептидом по меньшей мере из 30 аминокислот, предпочтительно из 32-50 аминокислот,
и в котором указанные одноцепочечные фрагменты Fab по пункту б) гибридизированы с указанным антителом полной длины по пункту а) посредством пептидного коннектора с С- или N-конца тяжелой или легкой цепи указанного антитела полной длины,
в котором указанный пептидный коннектор является пептидом по меньшей мере из 5 аминокислот, предпочтительно из 10-50 аминокислот.
В рамках указанного осуществления настоящего изобретения предпочтительно один или два, более предпочтительно два, одноцепочечных фрагмента Fab ба) VH-СН1-линкер-VL-CL или бб) VL-CL-линкер-VH-СН1, предпочтительно бб) VL-CL-линкер-VH-СН1, специфически связывающихся со вторым антигеном, гибридизированы с указанным антителом полной длины посредством пептидного коннектора с С-конца тяжелой цепи указанного антитела полной длины, и одноцепочечные фрагменты Fab не стабилизированы дисульфидами.
Один из вариантов осуществления настоящего изобретения представляет полиспецифичное антитело по настоящему изобретению, в котором один или два одноцепочечных фрагмента Fab, связывающихся со вторым антигеном, гибридизированы с указанным антителом полной длины посредством пептидного коннектора с С-конца тяжелых цепей указанного антитела полной длины (биспецифичное антитело).
Предпочтительно полиспецифичное антитело по настоящему изобретению включает два идентичных одноцепочечных фрагмента Fab, связывающихся со вторым антигеном, которые оба либо соединены с тяжелой цепью, либо оба из которых соединены с С- или N-концами легкой цепи (биспецифичное антитело).
Один вариант осуществления настоящего изобретения представляет полиспецифичное антитело по настоящему изобретению, в котором два идентичных одноцепочечных фрагмента Fab VL-CL-линкер-VH-СН1 или VH-СH1-линкер-VL-CL, предпочтительно VL-CL-линкер-VH-CH1, связывающихся со вторым антигеном, гибридизированы своими N-концами с указанным антителом полной длины посредством пептидного коннектора с двух С-концов двух тяжелых цепей или с двух С-концов двух легких цепей указанного антитела полной длины (четырехвалентное, биспецифичное антитело). В предпочтительном варианте осуществления настоящего изобретения указанное полиспецифичное антитело (предпочтительно указанное четырехвалентное, биспецифичное антитело) по настоящему изобретению содержит IgG полной длины и два идентичных одноцепочечных фрагмента Fab по настоящему изобретению, согласно вышеуказанному, и специфически связывается с IGF-1R человека, а также с EGFR человека. Эти молекулы предпочтительно основаны на сайтах связывания антигена антител человека против IGF-1R антител <IGF-1R>HUMAB клон 18 (DSM АСС 2587, WO 2005/005635, обозначение в виде аббревиатуры <IGF-1R> клон 18 или <IGF-1R>AK18) и гуманизированном <EGFR>ICR62 (в WO 2006/082515 обозначение в виде аббревиатуры <EGFR>ICR62). Эти молекулы одновременно нацелены и интерферируют с действием двух рецепторных тирозинкиназ на опухолевых клетках. Такое двойное действие приводит к явному улучшению противоопухолевого действия по сравнению с антителами, которые интерферируют только с одним из этих рецепторов. Схема, состав, получение и описание этих молекул показаны в примерах 1-6.
Таким образом, в одном из вариантов осуществления настоящего изобретения такое полиспецифичное антитело по настоящему изобретению отличается тем, что
i) указанное антитело полной длины специфически связывается с IGF1R и включает в вариабельном домене тяжелой цепи участок CDR3 последовательности SEQ ID NO:1, участок CDR2 последовательности SEQ ID NO:2 и участок CDR1 последовательности SEQ ID NO:3, а в вариабельном домене легкой цепи участок CDR3 последовательности SEQ ID NO:4, участок CDR2 последовательности SEQ ID NO:5 и участок CDR1 последовательности SEQ ID NO:6, и
ii) указанный одноцепочечный фрагмент Fab специфически связывается с EGFR и включает в вариабельном домене тяжелой цепи участок CDR3 последовательности SEQ ID NO:9, участок CDR2 последовательности SEQ ID NO:10 и участок CDR1 последовательности SEQ ID NO:11, а в вариабельном домене легкой цепи участок CDR3 последовательности SEQ ID NO:12, участок CDR2 последовательности SEQ ID NO:13 и участок CDR1 последовательности SEQ ID NO:14.
В одном варианте осуществления настоящего изобретения такое полиспецифичное антитело по настоящему изобретению отличается тем, что
i) указанное антитело полной длины специфически связывается с IGF-1R и включает в качестве вариабельного домена тяжелой цепи последовательность SEQ ID NO:7 и в качестве вариабельного домена легкой цепи последовательность SEQ ID NO:8, и
ii) указанный одноцепочечный фрагмент Fab специфически связывается с EGFR и включает в качестве вариабельного домена тяжелой цепи последовательность SEQ ID NO:15 и в качестве вариабельного домена легкой цепи последовательность SEQ ID NO:16.
В одном из вариантов осуществления настоящего изобретения такое полиспецифичное антитело по настоящему изобретению отличается тем, что
i) указанное антитело полной длины специфически связывается с EGFR и включает в вариабельном домене тяжелой цепи участок CDR3 последовательности SEQ ID NO:9, участок CDR2 последовательности SEQ ID NO:10 и участок CDR1 последовательности SEQ ID NO:11, а в вариабельном домене легкой цепи участок CDR3 последовательности SEQ ID NO:12, участок CDR2 последовательности SEQ ID NO:13 и участок CDR1 последовательности SEQ ID NO:14, и
ii) указанный одноцепочечный фрагмент Fab специфически связывается с IGF-1R и включает в вариабельном домене тяжелой цепи участок CDR3 последовательности SEQ ID NO:1, участок CDR2 последовательности SEQ ID NO:2 и участок CDR1 последовательности SEQ ID N0:3, а в вариабельном домене легкой цепи участок CDR3 последовательности SEQ ID NO:4, участок CDR2 последовательности SEQ ID N0:5 и участок CDR1 последовательности SEQ ID N0:6.
В одном варианте осуществления настоящего изобретения такое полиспецифичное антитело по настоящему изобретению отличается тем, что
i) указанное антитело полной длины специфически связывается с EGFR и включает в качестве вариабельного домена тяжелой цепи SEQ ID NO:15 и в качестве вариабельного домена легкой цепи SEQ ID NO:16, и
ii) указанный одноцепочечный фрагмент Fab специфически связывается с IGF1R и включает в качестве вариабельного домена тяжелой цепи SEQ ID NO:7 и в качестве вариабельного домена легкой цепи SEQ ID NO:8.
Одним из вариантов осуществления настоящего изобретения является полиспецифичное антитело по настоящему изобретению, в котором два идентичных одноцепочечных фрагмента Fab VL-CL-линкер-VH-CH1 или VH-CH1-линкер-VL-CL, предпочтительно VL-CL-линкер-VH-CH1, связывающихся со вторым антигеном, присоединены своими С-концами к указанному антителу полной длины посредством пептидного коннектора с двух N-концов двух тяжелых цепей или с двух N-концов двух легких цепей указанного антитела полной длины.
Одним из вариантов осуществления настоящего изобретения является полиспецифичное антитело по настоящему изобретению, в котором один одноцепочечный фрагмент Fab, связывающийся со вторым антигеном, соединен с указанным антителом полной длины посредством пептидного коннектора с С-или N-конца одной тяжелой цепи или одной легкой цепи указанного антитела полной длины. Одним из вариантов осуществления настоящего изобретения является полиспецифичное антитело по настоящему изобретению, в котором один одноцепочечный фрагмент Fab, связывающийся со вторым антигеном, соединен с указанным антителом полной длины посредством пептидного коннектора с N-конца одной тяжелой цепи или одной легкой цепи указанного антитела полной длины. Одним вариантом осуществления настоящего изобретения является полиспецифичное антитело по настоящему изобретению, в котором один одноцепочечный фрагмент Fab, связывающийся со вторым антигеном, соединен с указанным антителом полной длины посредством пептидного коннектора на С-конце одной тяжелой цепи или одной легкой цепи указанного антитела полной длины (см., например, фиг.6).
Предпочтительно полиспецифичное антитело по настоящему изобретению включает два одноцепочечных фрагмента Fab, связывающихся со вторым антигеном и третьим антигеном (трехспецифичное антитело) (см., например, фиг.7).
В другом объекте настоящего изобретения полиспецифичное антитело по настоящему изобретению включает
а) антитело полной длины, связывающееся с одним антигеном, состоящее из двух идентичных тяжелых цепей антитела VH-CH1-HR-CH2-CH3 и двух идентичных легких цепей антитела VL-CL, и
б) от одного до четырех одноцепочечных фрагментов Fab ба) VH-CH1-линкер-VL-CL или бб) VL-CL-линкер-VH-CH1, связывающихся с одним или до четырех дополнительных антигенов, причем указанные одноцепочечные фрагменты Fab связаны с указанным антителом полной длины посредством пептидного коннектора с С- или N-конца тяжелой и легкой цепи указанного антитела полной длины.
Антитела полной длины настоящего изобретения включают константные участки иммуноглобулина одного или нескольких классов иммуноглобулинов. Классы иммуноглобулинов включают изотипы IgG, IgM, IgA, IgD и IgE и, в случае IgG и IgA, их подтипы. В предпочтительном варианте осуществления настоящего изобретения антитело полной длины настоящего изобретения имеет структуру константного домена антитела типа IgG.
Понятия «моноклональное антитело» или «композиция моноклонального антитела» в контексте настоящего изобретения относится к препарату молекул антител единого аминокислотного состава.
Понятие «химерное антитело» относится к антителу, включающему вариабельную область, т.е. связывающую область, из одного источника или от одного вида и по меньшей мере часть константной области, полученной из другого источника или от другого вида, обычно получаемому методами рекомбинации ДНК. Предпочтительны химерные антитела, включающие вариабельную область мыши и константную область человека. Другими предпочтительными формами «химерных антител», которые охватывает настоящее изобретение, являются химерные антитела, константная область которых модифицирована или изменена по сравнению с исходным антителом с тем, чтобы получить свойства в соответствии с настоящим изобретением, особенно в отношении связывания с C1q и/или связывания с Fc-рецептором (FcR). Такие химерные антитела также относятся к так называемым «антителам переключенного класса». Химерные антитела являются продуктом экспрессии генов иммуноглобулина, включающих сегменты ДНК, кодирующие вариабельные участки иммуноглобулина, и сегменты ДНК, кодирующие константные области иммуноглобулина. Методы получения химерных антител включают традиционные методы рекомбинации ДНК и методы генной трансфекции, известные в данной области. См., например, S.L.Morrison и др., Proc. Natl. Acad. Sci. USA, 81, 1984, cc.6851-6855, US 5202238 и US 5204244.
Понятие «гуманизированное антитело» относится к антителам, у которых каркасные участки или «комплементарность-детерминирующие области» (CDR) модифицированы, чтобы включить CDR иммуноглобулина иной специфичности по сравнению с таковой исходного иммуноглобулина. В предпочтительном варианте осуществления настоящего изобретения CDR мыши трансплантируют в каркасный участок антитела человека, чтобы получить «гуманизированное антитело». См., например, L.Riechmann и др., Nature, 332, 1988, c.323-327 и M.S.Neuberger и др., Nature, 314, 1985, c.268-270. Особо предпочтительны области CDR, соответствующие тем репрезентативным последовательностям, которые распознают антигены, указанные выше для химерных антител. Другие формы «гуманизированных антител», которые охватывает настоящее изобретение, являются гуманизированными антителами, у которых константная область дополнительно модифицирована или изменена по сравнению с исходным антителом для того, чтобы получить свойства в соответствии с настоящим изобретением, особенно в отношении связывания с C1q и/или связывания с рецептором Fc (FcR).
Понятие «антитело человека» в контексте настоящего изобретения включает антитела, имеющие вариабельные и константные области, полученные из последовательностей иммуноглобулина зародышевой линии человека. Антитела человека известны в данной области (van M.A.Dijk и van de J.G.Winkel, Curr. Opin. Chem. Biol., 5, 2001, cc.368-374). Человеческие антитела также можно получать в трансгенных животных (например, в мышах), которые способны после иммунизации к выработке полного набора или выбранного элемента человеческих антител при отсутствии продукции эндогенных иммуноглобулинов. Перенос набора генов иммуноглобулинов зародышевой линии человека в такую зародышевую линию мутантных мышей приводит к получению антител человека в ответ на антигенную стимуляцию (см., например, A.Jakobovits и др., Proc. Natl. Acad. Sci. USA, 90, 1993, c.2551-2555, А.Jakobovits и др., Nature, 362, 1993, c.255-258, M.Brueggemann и др., Year Immunol., 7, 1993, c.33-40). Антитела человека также можно получить в библиотеках фагового дисплея (H.R.Hoogenboom и G.J.Winter, Mol. BioL, 227, 1992, cc.381-388, J.D.Marks и др., J.Mol. Biol., 222, 1991, c.581-597). Для получения моноклональных антител человека также доступны методы Cole и др. и Boerner и др. (S.P.C. Cole и др., Monoclonal Antibodies and Cancer Therapy, Alan R. Liss, 1985, с.77 и Р.Boerner и др., J. Immunol., 147, 1991, c.86-95). Согласно указанному выше для химерных и гуманизированных антител по настоящему изобретению, понятие «антитело человека» в контексте настоящего изобретения также включает такие антитела, которые модифицированы в константной области для получения свойств по настоящему изобретению, особенно в отношении связывания с C1q и/или связывания с FcR, например, путем «переключения класса», т.е. изменения или мутации частей Fc (например, от IgG1 до IgG4 и/или IgGl/IgG4 мутации).
В случае, когда полиспецифичное антитело по настоящему изобретению включает один или три одноцепочечных фрагмента Fab (или в случае двух неидентичных одноцепочечных фрагментов, которые оба присоединены к С-или N-концам либо тяжелой цепи, либо легкой цепи), в результате чего образуются гетеродимерные гибридные пептиды, СН3-домены указанного антитела полной длины по настоящему изобретению можно изменить методом «выступ-во-впадины», который подробно описан в нескольких примерах, например, в WO 96/027011, J.B.Ridgway и др., Protein Eng, 9, 1996, c.617-621 и A.M.Merchant и др., Nat Biotechnol., 16, 1998, c.677-681. В этом методе поверхности взаимодействия двух СН3 доменов изменены так, чтобы увеличить гетеродимеризацию обеих тяжелых цепей, содержащих эти два СН3-домена. Каждый из двух СН3-доменов (двух тяжелых цепей) может представлять «выступ», в то время как другой - «впадину». Введение дисульфидного мостика стабилизирует гетеродимеры (A.M.Merchant и др., Nature Biotech., 16, 1998, c.677-681, S.Atwell и др., J.Mol. Biol., 270, 1997, c.26-35) и повышает выход.
Таким образом, в одном из объектов настоящего изобретения указанное полиспецифичное антитело по настоящему изобретению включает только один одноцепочечный фрагмент Fab и дополнительно отличается тем, что каждый из доменов, и домен CH3 одной тяжелой цепи, и домен CH3 другой тяжелой цепи, присутствует на поверхности раздела, которая представляет исходную поверхность раздела между доменами CH3 антитела;
причем указанная поверхность раздела изменена таким образом, чтобы индуцировать формирование двухвалентного, биспецифичного антитела, причем изменение отличается тем, что:
а) домен CH3 одной тяжелой цепи изменен так,
что в исходной поверхности раздела домена CH3 одной тяжелой цепи, которая противостоит исходной поверхности раздела домена CH3 другой тяжелой цепи в двухвалентном биспецифичном антителе,
аминокислотный остаток замещен аминокислотным остатком, имеющим более длинную боковую цепь, таким образом, образуя выпуклость внутри поверхности раздела домена CH3 одной тяжелой цепи, которая помещается в полости внутри поверхности раздела домена CH3 другой тяжелой цепи, и
б) домен CH3 другой тяжелой цепи изменен так,
что внутри исходной поверхности раздела второго домена CH3, которая противостоит исходной поверхности раздела первого домена CH3 в трехвалентном биспецифичном антителе,
аминокислотный остаток замещен аминокислотным остатком, имеющим меньший объем боковой цепи, таким образом образуя полость в исходной поверхности раздела второго домена CH3, в которой размещается выпуклость в поверхности раздела первого домена CH3.
Предпочтительно указанный аминокислотный остаток, имеющий более длинный объем боковой цепи, выбран из группы, состоящей из аргинина (R), фенилаланина (F), тирозина (Y), триптофана (W).
Предпочтительно указанный аминокислотный остаток, имеющий меньший объем боковой цепи, выбран из группы, состоящей из ананина (А), серина (S), треонина (Т), валина (V).
В одном объекте настоящего изобретения оба домена CH3 дополнительно изменены путем введения цистеина (С) в качестве аминокислоты в соответствующих положениях каждого домена CH3 так, чтобы между обоими доменами CH3 мог образоваться дисульфидный мостик.
В предпочтительном варианте осуществления настоящего изобретения указанное полиспецифичное антитело включает только один одноцепочечный фрагмент Fab и является трехвалентным биспецифичным антителом. Указанное трехвалентное биспецифичное антитело включает мутацию T366W в домене CH3 «цепи с выступом» и мутации T366S, L368A, Y407V в домене CH3 «цепи со впадиной». Также можно использовать дополнительный дисульфидный мостик между цепочками доменов CH3 (A.M.Merchant и др., Nature Biotech, 16, 1998, c.677-681), например, путем введения мутации Y349C в домен CH3 «цепи с выступом» и мутации Е356С или мутации S354C в домен CH3 «цепи со впадиной». Таким образом, в другом предпочтительном варианте осуществления настоящего изобретения указанное трехвалентное биспецифичное антитело включает мутации Y349C, T366W в одном из двух доменов CH3, и мутации Е356С, T366S, L368A, Y407V в другом из двух доменов CH3, или указанное трехвалентное биспецифичное антитело включает мутации Y349C, T366W в одном из двух доменов CH3 и мутации S354C, T366S, L368A, Y407V в другом из двух доменов CH3 (дополнительную мутацию Y349C в одном домене CH3 и дополнительную мутацию Е356С или S354C в другом домене CH3 с образованием между цепями дисульфидного мостика) (нумерация всегда соответствует индексу ЕС по Kabat). Кроме того, в другом варианте или дополнительно можно использовать другие методы «выступ-во-впадины», описанные в ЕР 1870459А1. Предпочтительным примером для указанного трехвалентного биспецифичного антитела являются мутации R409D, К370Е в домене CH3 «цепи с выступами» и мутации D399K, Е357К в домене CH3 «цепи со впадиной» (нумерация всегда соответствует индексу ЕС по Kabat).
В другом предпочтительном варианте осуществления настоящего изобретения указанное трехвалентное биспецифичное антитело (полиспецифичное антитело, включающее только один одноцепочечный фрагмент Fab) включает мутацию T366W в домене CH3 «цепи с выступами» и мутации T366S, L368A, Y407V в домене CH3 «цепи со впадиной» и дополнительно мутации R409D, К370Е в домене CH3 «цепи с выступами» и мутации D399K, Е357К в СН3-домене «цепи со впадиной».
В другом предпочтительном варианте осуществления настоящего изобретения указанное трехвалентное биспецифичное антитело (полиспецифичное антитело, включающее только один одноцепочечный фрагмент Fab) включает мутации Y349C, T366W в одном из двух доменов CH3 и мутации S354C, T366S, L368A, Y407V в другом из двух доменов CH3, или указанное трехвалентное биспецифичное антитело включает мутации Y349C, T366W в одном из двух доменов CH3 и мутации S354C, T366S, L368A, Y407V в другом из двух доменов CH3 и дополнительно мутации R409D, К370Е в домене CH3 «цепи с выступами»и мутации D399K, Е357К в домене CH3 «цепи со впадиной».
Таким образом, одним из вариантов осуществления настоящего изобретения является полиспецифичное антитело по настоящему изобретению, в котором один одноцепочечный фрагмент Fab, связывающийся со вторым антигеном, гибридизирован с указанным антителом полной длины посредством пептидного коннектора с С- или N-конца одной тяжелой цепи или одной легкой цепи указанного антитела полной длины (предпочтительно с С-конце одной тяжелой цепи), причем антитело полной длины включает мутацию T366W в одном из двух доменов CH3 и мутации T366S, L368A, Y407V в другом из двух доменов CH3. Другим вариантом осуществления настоящего изобретения является полиспецифичное антитело по настоящему изобретению, в котором один одноцепочечный фрагмент Fab, связывающийся со вторым антигеном, гибридизирован с указанным антителом полной длины посредством пептидного коннектора с С- или N-конца одной тяжелой цепи или одной легкой цепи указанного антитела полной длины (предпочтительно с С-конца одной тяжелой цепи), причем антитело полной длины включает мутации Y349C, T366W в одном из двух доменов CH3 и мутации S354C, T366S, L368A, Y407V в другом из двух доменов CH3 (см., например, фиг.6).
Понятие «рекомбинантное антитело человека» в контексте настоящего изобретения относится ко всем антителам человека, которые получают, экспрессируют, создают или выделяют способами рекомбинации, например, к антителам, выделенным из клетки-хозяина, например клетки NSO или СНО, или из животного (например мыши), которое является трансгенным для генов иммуноглобулина человека, или к антителам, экспрессируемым, используя рекомбинантный вектор экспрессии, трансфицированный в клетку-хозяина. Такие рекомбинантные антитела человека содержат вариабельные и константные области в перестроенной форме. Рекомбинантные антитела человека по настоящему изобретению подвергаются in vivo соматической сверхмутации. Таким образом, аминокислотные последовательности областей VH и VL рекомбинантных антител являются последовательностями, которые хотя и производны от последовательностей VH и VL зародышевой линии человека или связаны с ними, могут естественным образом отсутствовать в зародышевой линии человека in vivo.
«Вариабельный домен» (вариабельный домен легкой цепи (VL), вариабельный домен тяжелой цепи (VH)) в контексте настоящего изобретения означает каждую из пары легкой и тяжелой цепей, которая вовлечена непосредственно в связывание антитела с антигеном. Вариабельные домены легких и тяжелых цепей человека имеют одну и ту же общую структуру, и каждый домен включает четыре каркасных участка (FR), последовательности которых являются в значительной степени консервативными соединенными тремя «гипервариабельными областями» (или комплементарность-детерминирующими областями (CDR)). Каркасные участки принимают β-складчатую конформацию, и CDR могут образовывать петли, объединяющие β-складчатую структуру. Области CDR в каждой цепи поддерживаются в виде трехмерной структуры с помощью каркасных участков и формируют вместе с областями CDR другой цепи сайт связывания антигена. Области CDR тяжелой и легкой цепи антитела играют очень важную роль в специфичности/аффинности связывания антител по настоящему изобретению, и вследствие этого представляют еще один объект изобретения.
Понятия «гипервариабельная область» или «антигенсвязывающий участок антитела» в контексте настоящего описания относятся к аминокислотным остаткам антитела, которые ответственны за связывание с антигеном. Гипервариабельная область включает аминокислотные остатки из «комплементарность-детерминирующих областей (complementary determining region - CDR)». В контексте настоящего описания «каркасные участки (framework - FR)» представляют собой участки вариабельных доменов, остатки которых отличны от остатков гипервариабельной области. Таким образом, легкие и тяжелые цепи антитела включают в направлении от N- к С-концу домены FR1, CDR1, FR2, CDR2, FR3, CDR3 и FR4. Области CDR на каждой цепи разделены такими каркасными аминокислотами. Прежде всего, CDR3 тяжелой цепи представляет область, которая главным образом обеспечивает связывание с антигеном. Области CDR и каркасные участки FR определяют в соответствии со стандартным принципом, описанным в кн.: Kabat и др., «Sequences of Proteins of Immunological Interest», Public Health Service, National Institutes of Health, Бетесда, Мэриленд, 1991, 5-е изд.
В контексте настоящего описания понятие «связывание» или «специфичное связывание» относится к связыванию антитела с эпитопом антигена в оценке in vitro, предпочтительно методом плазменного резонанса (анализ BIAcore, фирма GE-Healthcare, Упсала, Швеция) с очищенным антигеном дикого типа. Сродство связывания характеризуют с помощью параметров ka (константа скорости реакции ассоциации антитела из комплекса антитело/антиген), kD (константа скорости диссоциации) и kD (kD/ka). Связывание или специфичное связывание означает связывающее сродство (КD) 10-8 моль/л или менее, предпочтительно от 10-9 до 10-13 молей/л. Таким образом, полиспецифичное антитело по настоящему изобретению специфически связывается с каждым из антигенов, для которых оно является специфичным, со связывающим сродством (КD) 10-8 молей/л или менее, предпочтительно от 10-9 до 10-13 молей/л.
Связывание антитела с рецептором FcγRIII можно исследовать с помощью анализа BIAcore (фирма GE-Healthcare, Упсала, Швеция). Сродство связывания характеризуют с помощью параметров ka (константа скорости реакции ассоциации антитела из комплекса антитело/антиген), kD (константа диссоциации) и kD (kD/ka).
Понятие «эпитоп» включает какой-либо полипептидный детерминант, обладающий способностью специфически связываться с антителом. В определенных вариантах осуществления настоящего изобретения детерминант эпитопа включает химически активные поверхностные группы молекул, например, аминокислоты, боковые цепи сахаров, фосфорильную или сульфонильную группу и, в определенных вариантах осуществления настоящего изобретения может иметь характеристики специфичной трехмерной структуры или специфичные характеристики зарядов. Эпитоп представляет собой участок антигена, который связывается антителом.
В определенных вариантах осуществления настоящего изобретения антитело антитело рассматривают в качестве специфически связывающегося с антигеном, если оно предпочтительно распознается его антигеном-мишенью в сложной смеси белков и/или макромолекул.
Понятие «константная область» в контексте настоящего изобретения означает совокупность доменов антитела, отличных от вариабельной области.
Константный участок непосредственно не участвует в связывании антигена, но проявляет различные эффекторные функции. В зависимости от аминокислотной последовательности константных областей их тяжелых цепей антитела подразделяются на классы: IgA, IgD, IgE, IgG и IgM, и некоторые из них могут дополнительно подразделяться на подклассы, например, IgG1, IgG2, IgG3, и IgG4, IgAl и IgA2. Константные области тяжелых цепей, соответствующие разным классам антител, называются α, δ, ε, γ и µ, соответственно. Константные области легких цепей (CL), которые можно обнаружить во всех пяти классах антител, называются к (каппа) и λ (лямбда).
Понятие «константная область, происходящая от человека» в контексте настоящей заявки означает константный участок тяжелой цепи антитела человека подкласса IgG1, IgG2, IgG3 или IgG4 и/или константная область участок каппа или лямбда легкой цепи. Такие константные области хорошо известны в данной области и, например, описаны Е.А.Kabat (см., например, G.Johnson и Т.Т.Wu, Nucleic Acids Res., 28, 2000, c.214-218, Е.А.Kabat и др., Proc. Natl. Acad. Sci. USA, 72, 1975, c.2785-2788).
Хотя антитела подкласса IgG4 показывают пониженное связывание с Fc-рецептором (FcyRIIIa), антитела других подклассов IgG показывают сильное связывание. Однако Pro238, Asp265, Asp270, Asn297 (потеря углевода фрагмента Fc), Pro329, Leu234, Leu235, Gly236, Gly237, Ile253, Ser254, Lys288, Thr307, Gln311, Asn434 и His435 являются остатками, изменение которых также приводит к снижению связывания с рецептором Fc (R.L.Shields и др., J.Biol. Chem., 276, 2001, c.6591-6604, J.Lund и др., FASEB J., 9, 1995, cc.115-119, А.Morgan и др., Immunology, 86, 1995, c.319-324, ЕР 0307434).
В одном из вариантов осуществления настоящего изобретения антитело по настоящему изобретению обладает пониженным связыванием с FcR по сравнению с антителом IgG1 и исходным антителом полной длины в отношении связывания с FcR подкласса IgG4, или IgG1, или IgG2 подкласса с мутацией по положению S228, L234, L235 и/или D265, и/или содержит мутацию PVA236. В одном из вариантов осуществления настоящего изобретения мутации в исходном антителе полной длины расположены в местах S228P, L234A, L235A, L235E и/или PVA236. В другом варианте осуществления настоящего изобретения мутации в исходном антителе полной длины расположены в местах S228P в IgG4 и L234A и L235A в IgG1. Константные области тяжелых цепей показаны в виде последовательностей SEQ ID NO:17 и 18. В одном варианте осуществления настоящего изобретения константный участок тяжелой цепи исходного антитела полной длины имеет последовательность SEQ ID NO:17 с мутациями L234A и L235A. В другом варианте осуществления настоящего изобретения константная область тяжелой цепи исходного антитела полной длины имеет последовательность SEQ ID NO:18 с мутацией S228P. В другом варианте осуществления настоящего изобретения константная область легкой цепи исходного антитела полной длины является участок легкой цепи каппа последовательности SEQ ID NO:19 или участок легкой цепи лямбда. Предпочтительно константная область тяжелой цепи исходного антитела полной длины представляет последовательность SEQ ID NO:17 или SEQ ID NO:18 с мутацией S228P.
Константная область антитела непосредственно участвует в антитело-обусловленной клеточнозависимой цитотоксичности (antibody dependent cellular cytotoxicity - ADCC) и в комплементзависимой цитотоксичности (complement dependent cytotoxicity - CDC). Активация комплемента (CDC) инициируется связыванием фактора комплемента C1q с константной областью большинства подклассов IgG антител. Связывание C1q с антителом вызывается определенными межбелковыми взаимодействиями в так называемом сайте связывания. Такие сайты связывания константной области известны в данной области и описаны, например, T.J.Lukas и др., J. Immunol., 127, 1981, cc.2555-2560, R.Brunhouse и J.J.Cebra, Mol. Immunol., 16, 1979, cc.907-917, D.R.Burton и др., Nature, 288, 1980, cc.338-344, J.E.Thommesen и др., Mol. Immunol., 37, 2000, c.995-1004, E.E.Idusogie и др., J. Immunol., 164, 2000, c.4178-4184, М.Hezareh и др., J.Virol., 75, 2001, cc.12161-12168, A.Morgan и др., Immunology, 86, 1995, c.319-324 и ЕР 0307434. Такие сайты связывания константных областей отличаются, например, аминокислотами L234, L235, D270, N297, Е318, К320, К322, Р331 и Р329 (нумерация соответствует индексу ЕС по Kabat).
Понятие «антитело-обусловленной клеточнозависимой цитотоксичности (antibody dependent cellular cytotoxicity - ADCC)» относится к лизису клеток-мишеней человека антителом по настоящему изобретению в присутствии эффекторных клеток. ADCC оценивают предпочтительно путем обработки препарата антиген-экспрессирующих клеток антителом по настоящему изобретению в присутствии эффекторных клеток, например, свежевыделенных мононуклеарных клеток периферической крови (МКПК) или очищенных эффекторных клеток из светлого слоя кровяных сгустков (лейкоцитной пленки), например, моноцитов или естественных клеток-киллеров (NK), или линии постоянно растущих клеток NK.
Понятие «комплементзависимой цитотоксичности (complement dependent cytotoxicity - CDC» означает процесс, инициируемый связыванием фактора комплемента C1q с частью Fc большинства антител подклассов IgG. Связывание C1q с антителом вызывается определенными межбелковыми взаимодействиями на так называемом сайте связывания. Такие сайты связывания части Fc известны в данной области (см. выше). Такие сайты связывания части Fc отличаются, например, аминокислотами L234, L235, D270, N297, Е318, К320, К322, Р331 и Р329 (нумерация соответствует индексу ЕС по Kabat). Антитела подклассов IgG1, IgG2 и IgG3 обычно показывают активацию комплемента, включающую связывание C1q и С3, в то же время IgG4 не активирует систему комплемента и не связывает C1q и/или С3.
В еще одном варианте осуществления настоящего изобретения полиспецифичное антитело по настоящему изобретению отличается тем, что указанное антитело полной длины принадлежит подклассу IgG1 человека или подклассу IgG1 человека с мутациями L234A и L235A.
В еще одном варианте осуществления настоящего изобретения полиспецифичное антитело по настоящему изобретению отличается тем, что указанное антитело полной длины принадлежит подклассу IgG2 человека.
В еще одном варианте осуществления настоящего изобретения полиспецифичное антитело по настоящему изобретению отличается тем, что указанное антитело полной длины принадлежит подклассу IgG3 человека.
В еще одном варианте осуществления настоящего изобретения полиспецифичное антитело по настоящему изобретению отличается тем, что указанное антитело полной длины принадлежит подклассу IgG4 человека или подклассу IgG4 человека с дополнительной мутацией S228P.
Предпочтительно полиспецифичное антитело по настоящему изобретению отличается тем, что указанное антитело полной длины принадлежит подклассу IgG1 человека, подклассу IgG4 человека с дополнительной мутацией S228P.
В еще одном варианте осуществления настоящего изобретения полиспецифичное антитело по настоящему изобретению отличается тем, что указанное антитело полной длины модифицировано (либо путем мутаций в участках Fc, либо путем гликоинженерии) с целью повышения сродства к Fc-гамма рецептору IIIa человека, чтобы увеличить их способность к опосредованию ADCC. Способы усиления ADCC антител путем уменьшения количества фукозы описаны, например в WO 2005/018572, WO 2006/116260, WO 2006/114700, WO 2004/065540, WO 2005/011735, WO 2005/027966, WO 1997/028267, US 2006/0134709, US 2005/0054048, US 2005/0152894, WO 2003/035835, WO 2000/061739, R.Niwa и др., J. Immunol. Methods, 306, 2005, cc.151-160, Т.Shinkawa и др., J. Biol. Chem., 278, 2003, 3466-3473, WO 03/055993 или US 2005/0249722. Поэтому в одном варианте осуществления настоящего изобретения полиспецифичное антитело по настоящему изобретению отличается тем, что указанное антитело полной длины представляет афукозилированный изотип IgG1 или IgG3, в котором количество фукозы составляет 60% или менее от общего количества олигосахаридов (сахаров) при Asn297 (это означает, что по меньшей мере 40% или более олигосахаридов области Fc в положении Asn297 являются афукозилированными).
Антитело по настоящему изобретению получают способами рекомбинации. Поэтому одним из объектом настоящего изобретения является нуклеиновая кислота, кодирующая антитело по настоящему изобретению, и другим объектом является клетка, включающая указанную нуклеиновую кислоту, кодирующую антитело по настоящему изобретению. Способы рекомбинации широко известны в данной области и включают экспрессию белка в прокариотических или эукариотических клетках с последующим выделением антитела и обычно очисткой до фармацевтически приемлемой чистоты. Для экспрессии вышеуказанных антител в клетке-хозяине нуклеиновую кислоту, кодирующую соответствующие модифицированные легкие и тяжелые цепи, вставляют в векторы экспрессии посредством стандартных методов. Экспрессию проводят в подходящих прокариотических или эукариотических клетках-хозяевах, например, клетках СНО, клетках NSO, клетках SP2/0, клетках НЕК293, клетках COS, клетках PER.C6, клетках дрожжей или клетках Е. coli, и из клеток (супернатанта или клеток после лизиса) выделяют антитело. Общие способы рекомбинации для получения антител хорошо известны в данной области и описаны, например, в обзорных статьях S.C.Makrides, Protein Expr. Purif., 17, 1999, cc.183-202, S.Geisse и др., Protein Expr. Purif., 8, 1996, c.271-282, R.J.Kaufman, Mol. BiotechnoL, 16, 2000, c.151-160, R.G.Werner, Drug Res., 48, 1998, cc.870-880.
Полиспецифичные антитела по настоящему изобретению соответствующим образом выделяют из культуральной среды путем традиционных методов очистки иммуноглобулина, например, методами хроматографии на белок А-сефарозе, гидроксилаппатите, гель-электрофорезом, диализом или аффинной хроматографией. ДНК и РНК, кодирующие моноклональные антитела, легко выделяют и секвенируют, используя традиционные методы. В качестве источника таких РНК и ДНК могут служить гибридомные клетки. После изоляции ДНК можно вставить в векторы экспрессии, которые затем трансфицируют в клетки-хозяева, например, клетки НЕК 293, клетки СНО или миеломные клетки, которые иным способом не продуцируют иммуноглобулиновый белок, чтобы получить синтез рекомбинантных моноклональных антител в клетках-хозяевах.
Варианты последовательностей аминокислот (или мутантов) полиспецифичного антитела получают путем введения соответствующих нуклеотидных замен в ДНК антитела или путем синтеза нуклеотидов. Такие модификации, однако, можно осуществлять только в очень ограниченном диапазоне, например, описанном выше. Например, модификации не должны изменять вышеуказанные характеристики антитела, например, изотип IgG и связывание антигена, но могут повышать выход рекомбинантной продукции, стабильность белка или облегчать очистку.
Понятие «клетка-хозяин» в контексте настоящей заявки означает какую-либо клеточную систему, которую можно создать для получения антител по настоящему изобретению. В одном варианте осуществления настоящего изобретения в качестве клеток-хозяев используют клетки НЕК293 и клетки СНО.
В контексте настоящего изобретения экспрессирующие «клетки», «клеточные линии» и «клеточные культуры» используются взаимозаменяемо, и все эти обозначения включают потомство клеток. Таким образом, слова «трансформанты» и «трансформированные клетки» включают первичные клетки субъекта и культуры клеток, полученные из них, независимо от количества пассажей. Также следует понимать, что все потомство может не являться точно идентичным по содержанию ДНК, из-за неслучайных или случайных мутаций. К этим понятиям также относятся варианты потомства, которые обладают тем же функциональным или биологическим действием, что и первоначальная трансформированная клетка. Если подразумеваются другие значения, это будет ясно из контекста.
Экспрессия в клетках NS0 описана, например, L.M.Barnes и др., Cytotechnology, 32, 2000, c.109-123, L.M.Barnes и др., Biotech. Bioeng., 73 2001, c.261-270. Временная экспрессия описана, например, Y.Durocher и др., Nucl. Acids. Res., 30, 2002, с. Е9. Клонирование вариабельных доменов описано R.Orlandi и др., Proc. Natl. Acad. Sci. USA, 86, 1989, c.3833-3837, P.Carter и др., Proc. Natl. Acad. Sci. USA, 89, 1992, c.4285-4289 и L.Norderhaug и др., J. Immunol. Methods, 204, 1997, c.77-87. Предпочтительная система временной экспрессии (НЕК 293) описана E.-J.Schlaeger и К.Christensen, Cytotechnology, 30, 1999, c.71-83 и E.-J.Schlaeger, J. Immunol. Methods, 194, 1996, c.191-199.
Контрольные последовательности, пригодные для прокариот, включают, например, промотор, необязательно последовательность оператора и сайт связывания рибосомы. Известно, что для эукариотических клеток используют промоторы, энхансеры и сигналы полиаденилирования.
Нуклеиновая кислота является «оперативно связанной», если она находится в функциональной связи с другой нуклеотидной последовательностью. Например, ДНК предшествующей последовательности или лидерной секреторной последовательности оперативно связана с ДНК полипептида, если при ее экспрессии образуется белок-предшественник, который участвует в секреции полипептида; промотор или энхансер оперативно связаны с кодирующей последовательностью, если они оказывают воздействие на транскрипцию последовательности; или сайт связывания рибосомы оперативно связан с кодирующей последовательностью, если он расположен так, что облегчает трансляцию. Обычно, понятие «оперативно связанный» означает, что последовательности ДНК, будучи связаны, являются смежными, а в случае лидера секреции, смежными и находятся в рамке считывания. Однако энхансеры не должны быть смежными. Связывание осуществляют путем лигирования в соответствующих сайтах рестрикции. Если такие сайты отсутствуют, то в соответствии с общепринятой практикой применяют синтетические олигонуклеотидные адаптеры или линкеры.
Чтобы элиминировать другие клеточные компоненты или другие примеси, например другие клеточные нуклеиновые кислоты или белки, проводят очистку антител, используя стандартные методы, включая обработку щелочью/додецилсульфатом натрия, центрифугирование в градиенте CsCI, колоночную хроматографию, электрофорез в агарозном геле и другие методы, хорошо известные в данной участки. См. кн.: «Current Protocols in Molecular Biology», под ред. F.Ausubel и др., изд-во Greene Publishing and Wiley Interscience, Нью-Йорк, 1987. Для очистки белка хорошо известны и широко используются различные методы, например аффинную хроматографию с микробными белками (например, аффинная хроматография с белком А или белком G), ионообменную хроматографию (например, катионный обмен (карбоксиметильные смолы), анионный обмен (аминоэтильные смолы) и обмен смешанного типа), тиофильнаую абсорбцию (например с использованием бета-меркаптоэтанола и других SH-содержащих лигандов), гидрофобное взаимодействие или абсорбционную хроматографию с ароматическими соединениями (например, с использованием фенилсефарозы, аза-аренофильных смол или т-аминофенилборной кислоты), металло-хелатную аффинную хроматографию (например, с использованием Ni(II)- и Cu(II)-аффинного материала), эксклюзионную хроматографию и методы электрофореза (например гель-электрофореза, капиллярного электрофореза) (М.А.Vijayalakshmi, Appl. Biochem. Biotech., 75, 1998, cc.93-102).
Понятие «трансформация» в контексте настоящего изобретения относится к процессу переноса векторов/нуклеиновой кислоты в клетку-хозяина. Если в качестве клеток-хозяев используются клетки без труднопреодолимых барьеров клеточной стенки, трансфекцию проводят, например, методом преципитации кальцием фосфатом, согласно описанию F.L.Graham и A.J. van der Eb, Virology, 52, 1973, c.456-467. Однако для введения ДНК в клетки можно использовать и другие методы, например ядерную инъекцию или слияние протопластов. Если используются прокариотические клетки или клетки, содержащие прочные конструкции клеточных стенок, одним из методов трансфекции является, например, обработка кальцием с использованием хлорида кальция, согласно описанию S.N.Cohen и др., PNAS., 69, 1972, cc.2110-2114.
В контексте настоящего изобретения «экспрессия» относится к процессу, при котором нуклеиновая кислота транскрибируется в мРНК, и/или к процессу, при котором транскрибируемая РНК (также известная в качестве транскрипта) впоследствии транслируется в пептиды, полипептиды или белки. Транскрипты и кодируемые полипептиды в совокупности называются генным продуктом. Если полинуклеотид производят на основе геномной ДНК, экспрессия в эукариотической клетке может включать сплайсинг иРНК.
Понятие «вектор» относится к молекуле нуклеиновой кислоты, в частности самореплицирующейся, которая переносит инсертированную молекулу нуклеиновой кислоты в клетки-хозяева и/или между ними. Понятие включает векторы, которые главным образом функционируют для инсерции ДНК или РНК в клетку (например, хромосомная интеграция), репликативные векторы, которые главным образом функционируют для репликации ДНК или РНК, и векторы экспрессии, которые главным образом функционируют для транскрипции и/или трансляции ДНК или РНК. Также к понятию относятся векторы, которые обеспечивают более одной из указанных функций.
Понятие «вектор экспрессии» относится к полинуклеотиду, который при введении в подходящую клетку-хозяина может транскрибироваться и транслироваться в полипептид. Понятие «экспрессирующей системы» обычно относится к соответствующей клетке-хозяину, содержащей вектор экспрессии, который может функционировать с образованием требуемого продукта экспрессии.
Теперь установлено, что полиспецифичные антитела по настоящему изобретению имеют улучшенные характеристики, например, биологическое или фармакологическое действие, фармакокинетические свойства или токсичность. Они могут применяться, например, для лечения заболеваний, например, рака.
Одним из объектов настоящего изобретения является фармацевтическая композиция, включающая антитело по настоящему изобретению. Другим объектом настоящего изобретения является применение антитела по настоящему изобретению для получения фармацевтической композиции. Еще одним объектом настоящего изобретения является способ получения фармацевтической композиции, включающей антитело по настоящему изобретению. В другом объекте настоящее изобретение предусматривает композицию, например фармацевтическую композицию, содержащую антитело по настоящему изобретению вместе с фармацевтическим носителем.
В одном варианте осуществления настоящего изобретения предусматривают полиспецифичное, предпочтительно биспецифичное, антитело по настоящему изобретению для лечения рака.
Другим объектом настоящего изобретения является указанная фармацевтическая композиция для лечения рака.
Другим объектом настоящего изобретения является применение антитела по настоящему изобретению для получения лекарственного средства для лечения рака.
Другим объектом настоящего изобретения является способ лечения пациента, болеющего раком, посредством введения пациенту, нуждающемуся в таком лечении, антитела по настоящему изобретению.
В контексте настоящего описания понятие «фармацевтический носитель» включает какой-либо или все растворители, дисперсионные среды, покрытия, антибактериальные и противогрибковые агенты, придающие изотоничность и замедляющие абсорбцию агенты и т.п., которые являются физиологически совместимыми. Предпочтительно носитель можно применять для внутривенного, внутримышечного, подкожного, парентерального, спинального или эпидермального введения (например, путем инъекции или инфузии).
Композицию, предлагаемую в настоящем изобретении, можно вводить различными методами, известными в данной области. Для специалиста в данной области очевидно, что путь и/или способ введения должен варьироваться в зависимости от требуемых результатов. Для введения соединения, предлагаемого в изобретении, с помощью конкретных путей введения может оказаться необходимым нанесение на соединение покрытия из агента, препятствующего его инактивации, или осуществление совместного введения соединения с таким агентом. Например, соединение можно вводить субъекту в соответствующем носителе, например в липосомах или в разбавителе. Фармацевтически приемлемые разбавители включают физиологические и водные буферные растворы. Фармацевтически приемлемые носители включают стерильные водные растворы или дисперсии и стерильные порошки для приготовления и немедленного применения стерильных инъецируемых растворов или дисперсий. Применение таких сред и агентов для фармацевтически действующих субстанций известно в данной области.
Понятия «парентеральное введение» и «введенный парентерально» в контексте настоящего описания означают способы введения, отличные от энтерального и местного введения, обычно, путем инъекции, и включают, но ими не ограничиваются, внутривенную, внутримышечную, внутриартериальную, подоболочечную, внутрикапсулярную, внутриглазную, внутрисердечную, внутрикожную, внутрибрюшинную, транстрахеальную, подкожную, подкутикулярную, внутрисуставную, субкапсулярную, субарахноидальную, интраспинальную, эпидуральную и интрастернальную инъекцию и инфузию.
Понятие рак в контексте настоящего описания относится к пролиферативным заболеваниям, например, к лимфоме, лимфолейкозу, раку легкого, немелкоклеточному раку легкого, бронхиолоальвеолярному раку легкого, раку костей, раку поджелудочной железы, раку кожи, раку головы и шеи, кожной или внутриглазной меланоме, раку матки, раку яичников, раку прямой кишки, раку анальной области, раку желудка, раку толстой кишки, раку груди, раку матки, карциноме фаллопиевых труб, карциноме эндометрия, раку шейки матки, раку влагалища, карциноме вульвы, Болезни Ходжкина, раку пищевода, раку тонкой кишки, раку эндокринной системы, раку щитовидной железы, раку паращитовидных желез, раку надпочечников, саркоме мягких тканей, раку уретры, раку полового члена, раку предстательной железы, раку мочевого пузыря, раку почки или мочеточников, гипернефроме, раку почечной лоханки, мезотелиоме, гепатоцеллюлярному раку, раку желчных путей, опухолям центральной нервной системы (ЦНС), опухолям спинного мозга, глиоме стволовой части мозга, мультиформной глиобластоме, астроцитоме, невриномам, эпендимомам, медуллобластоме, менингиоме, плоскоклеточной карциноме, аденоме гипофиза и саркоме Юинга, включая устойчивые формы любого из вышеуказанного вида рака или сочетания одного или нескольких вышеуказанных видов рака.
Указанные композиции могут также содержать вспомогательные средства, например консерванты, увлажняющие агенты, эмульгаторы и диспергирующие агенты. Предупреждение присутствия микроорганизмов можно гарантировать применением процессов стерилизации, а также путем введения различных антибактериальных и противогрибковых агентов, например парабена, хлорбутанола, фенола, сорбиновой кислоты и т.п. Может также потребоваться введение в композицию агентов изотоничности, например сахаров, хлорида натрия и т.п. Кроме того, для пролонгированной абсорбции инъецируемой фармацевтической формы можно вводить агенты, которые замедляют абсорбцию, например моностеарат алюминия и желатин.
Независимо от выбранного способа введения соединения по настоящему изобретению можно применять в виде пригодной гидратированной формы и/или фармацевтические композиции по настоящему изобретению готовят в виде фармацевтически приемлемых лекарственных форм с помощью общепринятых методов, известных специалисту в данной области.
Фактические уровни доз действующих веществ в фармацевтических композициях по настоящему изобретению могут меняться так, чтобы они содержали такое количество действующего ингредиента, которое является эффективным для получения требуемого терапевтического ответа у конкретного пациента в зависимости от состава и пути введения, не являясь при этом токсичными для пациента. Выбранный уровень доз должен зависеть от разнообразных фармакокинетических факторов, включая активность конкретных применяемых композиций по настоящему изобретению, пути введения, времени введения, скорости выведения конкретного применяемого соединения, продолжительности лечения, других лекарственных средств, соединений и/или агентов, применяемых в сочетании с конкретными применяемыми композициями, возраста, пола, веса, состояния, общего состояния здоровья и предыдущей истории болезни пациента, подлежащего лечению, и подобных факторов, которые хорошо известны в медицине.
Композиция должна быть стерильной и жидкой до такой степени, чтобы композицию можно было вводить с помощью шприца. Помимо воды носитель предпочтительно может представлять собой изотонический забуференный физиологический раствор.
Требуемую текучесть можно поддерживать, например, нанося покрытие, например из лецитина, поддерживая требуемый размер частиц в случае дисперсии и применяя поверхностно-активные агенты. Во многих случаях предпочтительным является добавление в композицию придающих изотоничность агентов, например Сахаров, многоатомных спиртов, например маннита или сорбита, и хлорида натрия.
Описание аминокислотных последовательностей
SEQ ID NO:1 CDR3 тяжелой цепи, <IGF-1R> HUMAB- клон 18
SEQ ID NO:2 CDR2 тяжелой цепи, <IGF-1R> HUMAB- клон 18
SEQ ID NO:3 CDR1 тяжелой цепи, <IGF-1R> HUMAB- клон 18
SEQ ID NO:4 CDR3 легкой цепи, <IGF-1R> HUMAB- клон 18
SEQ ID NO:5 CDR2 легкой цепи, <IGF-1R> HUMAB- клон 18
SEQ ID NO:6 CDR1 легкой цепи, <IGF-1R> HUMAB- клон 18
SEQ ID NO:7 вариабельный домен тяжелой цепи, <IGF-1R> HUMAB-клон 18
SEQ ID NO:8 вариабельный домен легкой цепи, <IGF-1R> HUMAB-клон 18
SEQ ID NO:9 CDR3 тяжелой цепи, гуманизированное <EGFR> ICR62
SEQ ID NO:10 CDR2 тяжелой цепи, гуманизированное <EGFR> ICR62
SEQ ID NO:11 CDR1 тяжелой цепи, гуманизированное <EGFR> ICR62
SEQ ID NO:12 CDR3 легкой цепи, гуманизированное <EGFR> ICR62
SEQ ID NO:13 CDR2 легкой цепи, гуманизированное <EGFR> ICR62
SEQ ID NO:14 CDR1 легкой цепи, гуманизированное <EGFR> ICR62
SEQ ID NO:15 вариабельный домен тяжелой цепи, гуманизированное <EGFR> ICR62-I-HHD
SEQ ID NO:16 вариабельный домен легкой цепи, гуманизированное <EGFR>ICR62-I-KC
SEQ ID NO:17 константная область тяжелой цепи человека, производная от IgG1
SEQ ID NO:18 константная область тяжелой цепи человека, производная от IgG4
SEQ ID NO:19 константный участок легкой цепи каппа
Описание фигур
Фиг.1. Схематическая структура антитела полной длины без домена СН4, специфически связывающегося с первым антигеном 1, с двумя парами тяжелых и легких цепей, которые включают вариабельный и константный домены в определенной последовательности.
Фиг.2. Схематическая структура четырех возможных одноцепочечных фрагментов Fab, специфически связывающихся, например, со вторым антигеном 2.
Фиг.3. Схематическая структура полиспецифичных антител по настоящему изобретению, включающих антитело полной длины, специфически связывающееся с первым антигеном 1, и два одноцепочечных фрагмента Fab, специфически связывающихся со вторым антигеном 2 (пример биспецифичного четырехвалентного антитела).
Фиг.4. Биспецифичные антитела по настоящему изобретению, включающие антитело полной длины, специфически связывающееся с IGF-1R, и два идентичных одноцепочечных фрагмента Fab, специфически связывающихся с EGFR-ScFab-XGFR1 молекулами А, Б, В и Г, и уровни экспрессии после очистки.
A: scFab (VH-CH1-линкер-VL-CL), гибридизированный с С-концом тяжелой цепи,
Б: scFab (VH-CH1-линкер-VL-CL, гибридизированный с дополнительным VH44-VL100 дисульфидным мостиком) с С-концом тяжелой цепи,
В: scFab (VH-CH1-линкер-VL-CL), гибридизированный с С-концом легкой цепи,
Г: scFab (VH-CH1-линкер-VL-CL, гибридизированный с дополнительным VH44-VL100 дисульфидным мостиком) с С-концом легкой цепи.
Фиг.5. Биспецифичные антитела по настоящему изобретению, включающие антитело полной длины, специфически связывающееся с EGFR, и два идентичных одноцепочечных фрагмента Fab, специфически связывающихся с IGF-1R-ScFab-XGFR2 молекулами А, Б, В и Г.
A: scFab (VH-CH1-линкер-VL-CL), гибридизированный с С-концом тяжелой цепи,
Б: scFab (VH-CH1-линкер-VL-CL, гибридизированный с дополнительным VH44-VL100 дисульфидным мостиком) с С-концом тяжелой цепи,
В: scFab (VH-CH1-линкер-VL-CL), гибридизированный с С-концом легкой цепи,
Г: scFab (VH-CH1-линкер-VL-CL, гибридизированный с дополнительным VH44-VL100 дисульфидным мостиком) с С-концом легкой цепи.
Фиг.6. Схематическая структура полиспецифичных антител по настоящему изобретению, включающих антитело полной длины, специфически связывающееся с первым антигеном 1 и один одноцепочечный фрагмент Fab, специфически связывающийся со вторым антигеном 2 (пример биспецифичных трехвалентных антител с выступами и впадинами).
Фиг.7. Схематическая структура полиспецифичных антител по настоящему изобретению, включающих антитело полной длины, специфически связывающееся с первым антигеном 1, один одноцепочечный фрагмент Fab, специфически связывающийся со вторым антигеном 2, и один одноцепочечный фрагмент Fab, специфически связывающийся с третьим антигеном 3 (пример трехспецифичных четырехвалентных антител с выступами и впадинами).
Фиг.8. Данные анализа методом электрофореза белков в полиакриламидом геле в присутствии додецилсульфата натрия (sodium dodecyi sulfate polyacrylamide gel electrophoresis - SDS-PAGE) производных биспецифичных антител, содержащих фрагменты Fab, scFab-XGFR1
1: scFab-XGFR1_4720 (невосстановленное антитело),
2: scFab-XGFR1_4721 (невосстановленное антитело),
3: scFab-XGFR1_4720 (восстановленное антитело),
4: scFab-XGFR1_4721 (восстановленное антитело).
Фиг.9. Данные анализа методом высокоэффективной эксклюзионной хроматографии (High Performance Size Exclusion Chromatography - HP-SEC) производных биспецифичных антител scFab-XGFR1, содержащих фрагменты scFab.
Фиг.9а. scFab-XGFR1-4720; 7,7%, агрегатов (отмечены рамкой).
Фиг.9б. scFab-XGFR1-4721; 3,5% агрегатов (отмечены рамкой).
Фиг.10. Связывание scFab-XGFR1 и scFab-XGFR2 с EGFR и IGF1R.
Фиг.10а. Диаграмма Biacore. Связывание scFab-XGFR1_2720 с EGFR, KD=2нМ.
Фиг.10б. Диаграмма Biacore. Связывание scFab-XGFR1_2720 с IGF-1R, KD=2 нМ.
Фиг.10в. Диаграмма Biacore. Связывание scFab-XGFR2_2720 с EGFR, KD=0,5 нМ.
Фиг.10г. Диаграмма Biacore. Связывание scFab-XGFR2_2720 с IGF-1R, KD=11 нМ.
Фиг.11. Схема связывания scFab-XGFR с клетками, исследованного конкурентным методом сортировки клеток по флуоресценции (fluorescence-activated cell sorting - FACS) в следующем общем порядке осуществления действий:
- параллельно добавляют моноклональное антитело <IGF1R> с меткой Аlеха647 (1 мкг/мл), + антитело без метки scFab-XGFR (100-0,001 мкг/мл),
- инкубируют на льду в течение 45 мин, отмывают и удаляют несвязанные антитела,
- фиксируют 1% НСНО, затем проводят метод FACS.
Фиг.12. Связывание scFab-XGFR_2721 и исходного <IGF1R> клон 18 с клетками на основе конкурентного анализа FACS.
Фиг.12а. Сравнение значений IC50 <IGF-1R> клон 18 (0,18 мкг/мл) и scFab-XGFR_2721 (0,15 мкг/мл).
Фиг.12б. Кривая связывания <IGF-1R> клон 18 (точка поворота 0,11 мкг/мл); ось y - относительные световые единицы (ОСЕ), ось х - концентрация антител (мкг/мл).
Фиг.12в. Кривая связывания scFab-XGFR_2721 (точка поворота 0,10 мкг/мл); ось y - относительные световые единицы (ОСЕ), ось х - концентрация антител (мкг/мл).
Фиг.13. Регуляция по типу отрицательной (обратной) связи IGF1-R на Н322М-клетках после инкубации в течение 24 ч с различными вариантами scFab-XGFR (100 нМ).
Фиг.14. Регуляция по типу отрицательной (обратной) связи EGFR на Н322М-клетках после инкубации в течение 24 ч с различными вариантами scFab-XGFR (100 нМ).
Фиг.15. Ингибирование пролиферации Н322М-клеток различными вариантами scFab-XGFR (100 нМ).
Экспериментальная часть
Примеры
Материалы и основные методы
Общая информация, касающаяся нуклеотидных последовательностей легких и тяжелых цепей иммуноглобулинов человека, представлена Е.А.Kabat и др. в кн.: «Sequences of Proteins of Immunological Interest», Public Health Service, National Institutes of Health, Бетесда, Мэрилэнд,1991, 5-е изд. Нумерация и названия аминокислот цепей антител соответствуют нумерации ЕС (G.M.Edelman и др., Proc. Natl. Acad. Sci. USA, 63, 1969, сс.78-85, Е.А.Kabat и др. в кн.: «Sequences of Proteins of Immunological Interest», Public Health Service, National Institutes of Health, Бетесда, Мэрилэнд, 1991, 5-е изд.
Методы рекомбинации ДНК
Для манипуляций с ДНК используют стандартные методы, описанные J. Sambrook и др. в кн.: «Molecular cloning: A laboratory manual». Cold Spring Harbor Laboratory Press, Cold Spring Harbor, Нью-Йорк, 1989. Реагенты для работы в области молекулярной биологии применяют согласно инструкциям производителей.
Синтез генов
Требуемые сегменты генов получают из олигонуклеотидов, полученных химическим синтезом. Генные сегменты длиной 600-1800 п.о., которые фланкированы особыми сайтами расщепления эндонуклеазой рестрикции, собирают путем отжига и лигирования олигонуклеотидов, включая амплификацию методом ПЦР, и впоследствии клонируют по указанным сайтам рестрикции, например BamHI/BstEII, BamHI/BsiWI, BstEII/NotI или BsiWI/NotI, в pcDNA 3.1/Zeo(+) (фирма Invitrogen) на основе клонирующего вектора pUC. Последовательности ДНК субклонированных генных фрагментов контролируют путем секвенирования ДНК. Фрагменты синтеза генов упорядочивают в соответствии с заданными условиями в компании Geneart (Регенсбург, Германия).
Определение последовательности ДНК
Последовательности ДНК определяют путем двунитевого секвенирования, выполненного в компании Sequiserve GmbH (Ватерштеттен, Германия).
Секвенирование ДНК и белков и использование данных по секвенированию
Для создания последовательностей, картирования, анализа, комментирования и иллюстрирования используют пакет программ GCG (фирма Genetics Computer Group, Мэдисон, Висконсин), версия 10.2, и пакет программ Vector NT1 Advance фирмы Invitrogen, версия 9.1.
Методы клеточных культур
Используют стандартные методы клеточных культур, описанные в кн.: «Current Protocols in Cell Biology», 2000, под ред. J.S.Bonifacino, M., Dasso, J.B.Harford, J.Lippincott-Schwartz и K.M.Yamada, John Wiley & Sons, Inc.
Временная экспрессия вариантов иммуноглобулинов в клетках HEK293F
Полиспецифичные антитела экспрессируют путем временной трансфекции клеток первичной почки 293-F, используя экспрессирующую систему FreeStyle™ 293 Expression System в соответствии с инструкцией производителя (фирма Invitrogen, США). Вкратце, суспензию клеток FreeStyle™ 293-F культивируют в среде FreeStyle™ 293 Expression Medium при 37°С в атмосфере 8% СО2 и в день трансфекции клетки пересевают в свежую среду при плотности 1-2×106 жизнеспособных клеток/мл. Комплексы ДНК-293 fectin™ готовят в среде Opti-MEM® I (фирма Invitrogen, США), используя 333 мкл 293fectin™ (фирма Invitrogen, Германия) и 250 мкг плазмидной ДНК легкой и тяжелой цепей в мольном соотношении 1:1 для тансфекции в финальном объеме 250 мл. Спустя 7 суток после трансфекции супернатанты клеточной культуры, содержащие биспецифичные антитела, осветляют путем центрифугирования при 14000 g в течение 30 мин и фильтрации через стерильный фильтр (0,22 мкм). До очистки супернатанты хранят при -20°С.
Определение белка
Концентрацию белка очищенных антител и производных устанавливают на основе определения оптической плотности (ОН) при длине волны 280 нм, с ОП в качестве поправки на фон при длине волны 320 нм, используя коэффициент молярной экстинкции, вычисленный по аминокислотной последовательности, в соответствии с C.N.Расе и др., Protein Science, 4, 1995, cc.2411-2423.
Определение концентрации антител в супернатантах
Концентрацию антител и производных в супернатантах клеточных культур оценивают с помощью аффинной ВЭЖХ хроматографии. Вкратце, супернатанты клеточных культур, содержащие антитела и производные, которые связываются с белком А, наносят на колонку Applied Biosystems Poros A/20 в растворе 200 мМ КН2РO4, 100 мМ цитрата натрия, рН 7,4, и элюируют с матрицы раствором 200 мМ NaCl, 100 мМ лимонной кислоты, рН 2,5, на системе ВЭЖХ UltiMate 3000 (фирма Dionex). Количество элюированного белка устанавливают с помощью УФ поглощения и интеграции площадей пиков. Стандартом является очищенное стандартное антитело IgG1.
Очистка белка
Выделенные антитела подвергают двухступенчатой очистке от супернатанта с помощью аффинной хроматографии, используя Protein А-Sepharose™ (фирма GE Healthcare, Швеция), и экслюзионной хроматографии на колонке Superdex200. Вкратце, осветленные культуральные супернатанты, содержащие биспецифичные и триспецифичные антитела, наносят на колонку HiTrap ProteinA HP (5 мл), уравновешенную ФСБ буфером (10 мМ Na2HPO4, 1 мМ КН2РO4, 137 мМ NaCl и 2,7 мМ КСl, рН 7,4). Несвязанные белки отмывают уравновешивающим буфером. Биспецифичные антитела элюируют 0,1 М цитратным буфером, рН 2,8, и фракции, содержащие белок, нейтрализуют, используя 0,1 мл 1 М триса, рН 8,5. Затем, элюированные фракции белка объединяют, концентрируют, используя фильтрующую центрифугу Amicon Ultra (MWCO: 30 К, фирма Millipore), до объема 3 мл и наносят на гельфильтрующую колонку Superdex200 HiLoad 120 мл 16/60 (фирма GE Healthcare, Швеция), уравновешенную 20 мМ гистидина, 140 мМ NaCl, рН 6,0. Фракции мономерных антител объединяют, мгновенно замораживают и хранят при -80°С. Части образцов используют для последующего анализа и описания белков.
Анализ очищенных белков
Концентрацию белка в образцах очищенных белков определяют посредством измерения оптической плотности (ОП) при длине волны 280 нм, используя коэффициент мольной экстинкции, рассчитанный на основе аминокислотной последовательности. Чистоту биспецифичных антител анализируют методом электрофореза белков в полиакриламидом геле в присутствии додецилсульфата натрия (sodium dodecyl sulfate polyacrylamide gel electrophoresis - SDS-PAGE) в присутствии или в отсутствие восстанавливающего агента (5 мМ 1,4-дитиотреитола) и окрашивают, используя кумасси бриллиантовый голубой. Гелевую систему NuPAGE® Pre-Cast (фирма Invitrogen, США) применяют согласно инструкции производителя (4-20% гели трис-глицин). Содержание совокупности образцов биспецифических антител анализируют методом высокоэффективной эксклюзионной хроматографии (High Performance Size Exclusion Chromatography - HP-SEC) в системе ВЭЖХ UltiMate 3000 (фирма Dionex), используя аналитическую эксклюзионную колонку Superdex 200 (фирма GE Healthcare, Швеция) в подвижном буфере, 200 мМ КН2РO4, 250 мМ КСl, рН 7,0, при температуре 25°С. 25 мкг белка наносят на колонку при скорости потока 0,5 мл/мин и элюируют кратным объемом через 50 мин. Для анализа стабильности готовят концентрации 0,1 мг/мл, 1 мг/мл и 3 мг/мл очищенных белков и инкубируют при 4°С, 37°С в течение 7 суток и затем оценивают методом высокоэффективной эксклюзионной хроматографии (HP-SEC). Целостность аминокислотного каркаса легких и тяжелых цепей восстановленных биспецифических антител подтверждают квадруполь-времяпролетной тандемной масс-спектрометрией с наноэлектроспрейным источником ионов (NanoElectrospray Q-TOF mass spectrometry - TOF MS) после удаления N-гликанов путем ферментативной обработки пептид-ТМ-гликозидазой F (фирма Roche Molecular Biochemicals).
Пример 1. Конструирование молекул полиспецифичных антител по настоящему изобретению, которые распознают IGF1-рецептор человека и EGF-рецептор человека
Ниже приводят в качестве примера одного из вариантов осуществления настоящего изобретения четырехвалентные биспецифичные антитела, включающие антитело полной длины, связывающееся с первым антигеном (IGF-1R или EGFR), и два одноцепочечных фрагмента Fab, связывающихся со вторым отличным от первого антигеном (отличным от IGF-1R или EGFR), связанным с антителом полной длины посредством пептидного коннектора (либо оба одноцепочечных фрагмента Fab с двух С-концов тяжелой цепи, либо с двух С-концов легкой цепи). Домены антитела и линкер в указанном одноцепочечном фрагменте Fab расположены в следующем порядке в направлении от N-конца к С-концу: VL-CL-линкер-VH-CH1.
В качестве вариабельного домена тяжелой цепи VH для <IGF-1R> сайта связывания антигена используют последовательность SEQ ID NO:15. В качестве вариабельного домена легкой цепи VL для <IGF-1R> сайта связывания антигена используют последовательность SEQ ID NO:16.
В качестве вариабельного домена тяжелой цепи VH для <EGFR> сайта связывания антигена используют последовательность SEQ ID NO:7. В качестве вариабельного домена легкой цепи VL для <EGFR> сайта связывания антигена используют последовательность SEQ ID NO:8.
Путем генного синтеза и методов рекомбинации в молекулярной биологии VL-CL и VH-CH1, включающие VH и VL соответствующего сайта связывания антигена связываются посредством глицин-серинового (G4S)n одноцепочечного линкера, чтобы получить одноцепочечный фрагмент Fab VL-CL-линкер-VH-СН1, который прикрепляется к С-концу тяжелой или легкой цепи антитела с помощью (G4S)n линкера.
Необязательно в домены VH (включая положение 44 по Kabat) и VL (включая положение 100 по Kabat) одноцепочечного фрагмента Fab включают остатки цистеина, следуя технологии, описанной ранее (например, WO 94/029350, Y.Reiter и др., Nature biotechnology, 14, 1996, cc.1239-1245, N.M.Young и др., FEBS Letters, 377, 1995, cc.135-139 или V.Rajagopal и др., Protein Engineering, 10, 1997, cc.1453-1459).
Все указанные молекулы получают с помощью рекомбинации, очищают и описывают, а также проводят оценку экспрессии, стабильности и биологического действия белка.
Сущность конструкции полиспецифичных антител, которые используют для получения тетравалентных, биспецифичных <EGFR-IGF-1R>, <IGF-1R-EGFR> антител, представлена в табл.1. Для этого исследования для описания различных моделей четырехвалентных белков в настоящем изобретении используют обозначение «scFab-Ab». Описание сконструированных форматов показано на фиг.4 и 5 и перечислено в табл.1.
Пример 2. Экспрессия и очистка молекул scFabXGFR1 биспецифичного <EGFR-IGF1R> антитела
Легкие и тяжелые цепи соответствующих биспецифичных антител встраивают в векторы экспрессии, несущие про- или эукариотические селективные маркеры. Эти плазмиды амплифицируют в Е. coli, очищают и затем трансфецируют для временной экспрессии в клетки HEK293F (используя коммерчески доступную систему фирмы Invitrogen). Спустя 7 суток супернатанты клеток НЕК 293 собирают и подвергают очистке с помощью белка А и эксклюзионной хроматографии. Гомогенность всех конструкций биспецифичных антител подтверждают с помощью SDS-PAGE в невосстанавливающих и восстанавливающих условиях. В восстанавливающих условиях (фиг.8), цепи полипептида, несущие С- и N-концевые гибриды scFab, показали в результате электрофореза SDS-PAGE точно установленные молекулярные размеры, аналогичные рассчитанным молекулярным массам. Уровни экспрессии всех конструкций анализируют с помощью белок А-ВЭЖХ, и они являются близкими к выходам экспрессии «стандартных» IgG или в некоторых случаях немного ниже. Средние выходы белка составляют 1,5-10 мг белка на литр супернатанта клеточной культуры в таких экспериментах без оптимизации временной экспрессии (фиг.4 и 5).
Анализы очищенных белков с помощью высокоэффективной эксклюзионной хроматографии показывают некоторую тенденцию к агрегации рекомбинантных молекул. В отношении проблем с агрегацией биспецифичных антител используют дисульфидную стабилизации. Между VH и VL дополнительных связывающих частей молекул. Для этого вводят одиночные замены цистеина внутри VH и VL фрагмента scFab в определенных положениях (положения VH44/VL100 в соответствии со схемой нумерации по Kabat). Эти мутации допускают возможность образования стабильных дисульфидных связей между цепочками VH и VL, которые в свою очередь стабилизируют полученный в результате дисульфид-стабилизированный scFab модуль. Введение VH44/VL100 дисульфидов в scFab незначительно влияет на уровни экспрессии белка, а в некоторых случаях даже повышает выход экспрессии (см. фиг.4 и 5).
Биспецифичные антитела экспрессируют посредством временной трансфекции клеток эмбриональной почки человека 293-F, используя экспрессирующую систему FreeStyle™ 293 Expression System в соответствии с инструкцией производителя (фирма Invitrogen, США). Вкратце, суспензию клеток FreeStyle™ 293-F культивируют в среде для экспрессии FreeStyle™ 293 Expression Medium при 37°С в атмосфере 8% СО2 и в день трансфекции клетки пересевают в свежую среду при плотности 1-2×106 жизнеспособных клеток/мл.
Комплексы ДНК-293fectin™ готовят в среде Opti-MEM® I (фирма Invitrogen, США), используя 333 мкл 293fectin™ (фирма Invitrogen, Германия) и 250 мкг плазмидной ДНК легкой и тяжелой цепей в мольном соотношении 1:1 для тансфекции в конечном объеме 250 мл. Спустя 7 суток после трансфекции супернатанты клеточных культур, содержащие производные рекомбинантных антител, осветляют центрифугированием в режиме 14000 g в течение 30 мин и фильтрацией через стерильный фильтр (0,22 мкм). До очистки супернатанты хранят при -20°С.
Секретированные производные антител подвергают двухступенчатой очистке от супернатанта с помощью аффинной хроматографии, используя Protein A-Sepharose™ (фирма GE Healthcare, Швеция), и экслюзионной хроматографии на колонке Superdex 200. Вкратце, осветленные культуральные супернатанты, содержащие биспецифичные и трехспецифичные антитела, наносят на колонку HiTrap ProteinA HP (5 мл), уравновешенную ФСБ буфером (10 мМ Na2НРO4, 1 мМ КН2РO4, 137 мМ NaCl и 2,7 мМ КСl, рН 7,4). Несвязанные белки отмывают уравновешивающим буфером. Производные антител элюируют 0,1 М цитратным буфером, рН 2,8, и фракции, содержащие белок, нейтрализуют, используя 0,1 мл 1 М триса, рН 8,5. Затем, элюированные фракции белка объединяют, концентрируют, используя фильтрующую центрифугу Amicon Ultra (MWCO: 30 К, фирма Millipore) до объема 3 мл и наносят на гельфильтрационную колонку Superdex200 HiLoad 120 мл 16/60 (фирма GE Healthcare, Швеция), уравновешенную раствором 20 мМ гистидина, 140 мМ NaCl, рН 6,0. Фракции мономерных антител объединяют, мгновенно замораживают и хранят при -80°С. Часть образцов используют для последующих анализов и описания белков. В качестве иллюстрации результаты анализа методом SDS-PAGE очищенных белков и профили, полученные на основе высокоэффективной эксклюзионной хроматографии (ВЭЭХ) производных биспецифичных антител, показаны на фиг.8 и 9.
Фиг.5 показывает выходы экспрессии, которые наблюдались в системах временной экспрессии: все сконструированные варианты производных антител можно экспрессировать и подвергать очистке в достаточных для дальнейших анализов количествах.
Для сравнения получают четырехвалентное биспецифичное антитело, основанное на антителе полной длины, к которому через пептидный линкер с С-конца тяжелой цепи присоединены два scFv-фрагмента, согласно описанию в WO 1995/009917 и в кн.: D.Müller и др. «Handbook of Therapeutic antibodies», 2008, ч.III, гл.2, cc.345-378. В качестве вариабельного домена тяжелой цепи VH для <IGF-1R> сайта связывания антигена используют последовательность SEQ ID NO:15. В качестве вариабельного домена легкой цепи VL для <IGF-1R> сайта связывания антигена используют последовательность SEQ ID NO:16. В качестве вариабельного домена тяжелой цепи VH для <EGFR> сайта связывания антигена используют последовательность SEQ ID NO:7. В качестве вариабельного домена легкой цепи VL для <EGFR> сайта связывания антигена используют последовательность SEQ ID NO:8. Эту сравнительную молекулу называют «XGFR1_2320» (она также описана в РСТ РСТ/ЕР 2009/006782).
Биспецифичная одноцепочечная молекула Fv XGFR1-2320 имеет конечный выход после очистки 0,27 мг, а конечный выход соответствующей одноцепочечной молекулы Fab XGFR1-2720 составляет 6,8 мг (см. фиг.4, соединение А), что представляет повышение выхода более чем в 200 раз.
Пример 3. Стабильность и тенденция к агрегации молекул scFab-XGFR биспецифичного антитела <EGFR-IGF1R>
Для определения количества агрегатов, присутствующих в препарате производных рекомбинантных антител, проводят анализ с помощью высокоэффективной эксклюзионной хроматографии (HP-SEC). Для этого образцы биспецифичных антител анализируют с помощью HP-SEC в системе ВЭЖХ UltiMate 3000 (фирма Dionex), используя аналитическую эксклюзионную колонку Superdex 200 (фирма GE Healthcare, Швеция). Фиг.9 показывает пример этих анализов. Агрегаты проявляются в качестве отдельного пика или плеча перед фракциями, которые содержат производное мономерного антитела. Для этой работы выявляют искомые «мономерные» молекулы, объединяемые в два гетеродимера тяжелой и легкой цепей и фрагментов scFab, связанных с любым из них. Целостность аминокислотного каркаса легкой и тяжелой цепей восстановленного биспецифичного антитела и гибридных белков подтверждают квадруполь-времяпролетной тандемной масс-спектрометрией с наноэлектроспрейным источником ионов (NanoElectrospray Q-TOF mass spectrometry - TOF MS) после удаления N-гликанов путем ферментативной обработки пептид-N-гликозидазой F (фирма Roche Molecular Biochemicals).
Высокоэффективная эксклюзионная хроматография (HP-SEC) очищенных белков при различных условиях (варьирующие концентрации и время) показала слабую тенденцию к агрегации у молекул, содержащих scFab-фрагменты, по сравнению с нормальными IgG. Эту слабую тенденцию к агрегации, которая наблюдалась для некоторых молекул, можно уменьшить путем введения межцепьевой дисульфидной связи VH44/VL100 в scFab-модулях.
Пример 4. Связывание scFab-молекул биспецифичного <EGFR-IGF1R> антитела с рецептором тирозинкиназы (receptor tyrosine kinase - RTK), EGFR и IGF1R
Связывание scFab-модулей и сайтов связывания антигена, сохраненных в полноразмерном IgG-модуле различных форматов биспецифичных антител scFab-XGFR, сравнивают со связыванием IgG «дикого типа», из которых получены связывающие модули и биспецифичные антитела. Эти анализы проводят, используя поверхностный плазменный резонанс (анализ Biacore), a также клеточный иммуноферментный анализ ELISA.
Связывающие свойства биспецифичных <IGF-1R-EGFR> антител анализируют методом поверхностного плазменного резонанса (ППР), используя прибор Biacore T100 (фирма GE Healthcare Bio-Sciences AB, Упсала). Эта система хорошо приспособлена для исследований взаимодействий молекул. Она позволяет в условиях реального времени проводить длительный мониторинг связываний лиганд/анализируемое вещество и таким образом определять константы скорости ассоциации (ka), константы скорости диссоциации (kd) и константы равновесия (KD) в различных условиях эксперимента. Метод ППР основан на измерении показателя преломления близкого к поверхности покрытого золотом биосенсорного чипа. Изменения в показателе преломления указывают на изменения массы на поверхности, вызываемые взаимодействием иммобилизованного лиганда с анализируемым веществом, который вводят в раствор инъекцией. Если молекулы связываются с иммобилизованным на поверхности лигандом, масса увеличивается, а в случае диссоциации масса снижается.
Захватывающее антитело против IgG человека иммобилизуют на поверхности биосенсорного чипа С1, используя химию аминного сочетания. Проточные кюветы активируют с помощью смеси 1:1 0,1 М N-гидроксисукцинимида и 0,1 М 3-(N,N-диметиламино)пропил-N-этилкарбодиимида при скорости потока 5 мкл/мин. Антитело против IgG человека вносят инъекцией в ацетат натрия, рН 5,0, в количестве 5 мкг/мл, в результате чего создается поверхностная плотность приблизительно 200 RU. Контрольную проточную кювету обрабатывают тем же способом, но только буферами-разбавителями вместо захватывающего антитела. Поверхности блокируют путем инъекции 1 М этаноламина/HCl, рН 8,5. Биспецифичные антитела разводят в буфере HBS-P и вносят инъекцией при скорости потока 5 мкл/мин. Время контакта (фаза ассоциации) составляет 1 мин для антител при концентрации 1-5 нМ. EGFR-ECD вводят инъекцией при возрастающих концентрациях 1,2, 3,7, 11,1, 33,3, 100 и 300 нМ, IGF-1R при концентрациях 0,37, 1,11, 3,33, 10, 30 и 90 нМ. Время контакта (фаза ассоциации) составляет 3 мин, время диссоциации (отмывка подвижным буфером) 5 мин для обеих молекул при скорости потока 30 мкл/мин. Все взаимодействия осуществляют при 25°С (стандартная температура). После каждого цикла связывания для удаления любых нековалентно связанных белков вносят инъекцией регенирующие растворы 0,85% фосфорной кислоты и 5 мМ гидроксида натрия, каждый в течение 60 сек при скорости потока 5 мкл/мин. Сигналы определяют при скорости один сигнал в сек. Образцы вносят инъекцией при увеличивающихся концентрациях.
Пример одновременного связывания биспецифичных <IGF-1R-EGFR> антител с EGFR и IGF1R показан на фиг.10а-г.
Для оценки способности к связыванию производных биспецифичных антител с RTK, расположенной на поверхности клеток, можно также использовать анализ связывания и конкурентный анализ на основе FACS на культивируемых клетках. Фиг.11 показывает экспериментальную установку, которую используют для тестирования способности к связыванию содержащих scFab биспецифичных производных XGFR на раковых клетках А549. Для этих клеточных конкурентных анализов отделяют и подсчитывают клетки А549, которые экспрессируют антигены EGFR и IGF1R. 1,5×10 клеток на лунку высевают в 96-луночный планшет с коническими лунками. Клетки осаждают центрифугированием (1500 об/мин, 4°С, 5 мин) и инкубируют в течение 45 мин на льду в 50 мкл серийных разведении соответствующего биспецифичного антитела в ФСБ с 2% ФСТ (фетальной сывороткой теленка), содержащих 1 мкг/мл Аlеха647-меченного IGFIR-специфичного антитела. Клетки снова осаждают центрифугированием и дважды промывают 200 мкл ФСБ, содержащим 2% ФСТ. В итоге клетки ресуспендируют в растворе BD CellFix (фирма BD Biosciences) и инкубируют по меньшее мере в течение 10 мин на льду. Среднюю интенсивность флуоресценции клеток определяют с помощью проточной цитометрии (прибор FACS Canto). Среднюю интенсивность флуоресценции определяют по меньшей мере в двух повторностях двух независимых окрашиваний. Спектры, полученные на основе проточной цитометрии, в дальнейшем обрабатывают, используя программное обеспечение FlowJo software (фирма TreeStar). Половину максимального связывания (IC50) определяют, используя программу XLFit 4.0 (фирма IDBS) и дозозависимую одностороннюю модель 205.
Результаты этих анализов, представленные на фиг.12а-в, показывают функциональность связывания биспецифичных производных scFab-содержащих антител на поверхности опухолевых клеток. Например, показатель IC50 в экспериментах по конкурентному связыванию биспецифичного производного антитела scFab-XGFR1_2721 составляет 0,11 мкг/мл, а значение IC50 моноспецифичного антитела превышает 50% (0,18 мкг/мл). Такое повышенное действие в конкурентных анализах биспецифичного производного scFab-XGFR_2721 по сравнению с исходным антителом позволяет предположить, что биспецифичная молекула лучше связывается с поверхностью клеток, чем моноспецифичное антитело.
Пример 5. Регуляция по типу отрицательной (обратной) связи EGFR- и IGF-1R- биспецифичными scFab-XGFR молекулами <EGFR-IGF-1R> антител
Антитело человека против IGF-1R <IGF-1R> HUMAB клон 18 (DSM АСС 2587) ингибирует передачу сигнала через IGFR1 и гуманизированное антитело крысы против EGFR <EGFR> ICR62 ингибирует передачу сигнала через EGFR. Для оценки потенциального ингибирующего действия различных вариантов scFab-XGFR1 анализируют степень регуляции по типу отрицательной (обратной) связи_обоих рецепторов.
Чтобы выявить воздействие антитела по настоящему изобретению на количество рецептора IGF-I (IGF-1R) в опухолевых клетках проводят эксперименты зависимости от времени и последующий анализ ELISA со специфичными антителами к IGF-1R и EGFR.
6-луночный планшет инокулируют в количестве 1 мл на лунку опухолевыми клетками (Н322М, 5×105 клеток/мл) в среде RPMI 1640 с добавлением 10% ФСТ (фирма РАА, номер в каталоге Е15-039) и 1% пенициллина/стрептомицина. В каждую лунку добавляют 3 мл среды, клетки культивируют в течение 24 ч при 37°С и 5% СО2.
Среду осторожно удаляют и замещают 2 мл 100 нМ XGFR антителами, разведенными средой RPMI-VM. В контрольных лунках среду замещают либо средой, либо буфером без антитела и средой с контрольными антителами (<IGF-1R> HUMAB клон 18 и <EGFR> ICR62 с конечной концентрацией 100 нМ). Клетки инкубируют при 37°С и 5% СО2 и отдельные планшеты берут для дальнейшего процессинга в течение 24 ч.
Среду осторожно удаляют отсасыванием, и клетки промывают 1 мл ФСБ. Добавляют 300 мкл/лунку холодного лизирующего буфера MES (MES, 10 мМ Na3VO4 и протеазный ингибитор Complete). Через один час клетки отделяют на льду, используя клеточный скрепер (фирма Corning, кат.№3010), и содержимое лунок переносят в пробирки Эппендорф для проведения реакций. Фрагменты клеток удаляют центрифугированием в течение 10 мин при 13000 об/мин и 4°С.
Для выявления EGFR
96-луночные планшеты для микротитрования (МТП) подготавливают в соответствии с протоколом (DuoSet ELISA for Human EGFR, фирма RnD systems, номер в каталоге DY231). Козье антитело к EGFR человека в количестве 144 мкг/мл в ФСБ разводят в соотношении 1:180 в ФСБ и добавляют 100 мкл/лунку в МТП. МТП инкубируют в течение ночи при комнатной температуре в условиях встряхивания. Планшеты промывают три раза ФСБ, содержащим 0,1% Tween® 20, и блокируют добавлением 300 мкл/лунку раствора ФСБ, содержащего 3% БСА (бычий сывороточный альбумин) и 0,1% Tween® 20, в течение 1 ч при комнатной температуре в условиях встряхивания. Планшеты промывают три раза ФСБ, содержащим 0,1% Tween® 20.
В лизатах клеток определяют количество белка, используя набор ВСА Protein Assay kit (фирма Pierce), затем лизаты клеток доводят до концентрации белка 0,04 мг/мл литическим буфером MES с добавлением 100 мМ Na3VO4, 1:100, и протеазного ингибитора Complete®, 1:20, и 100 мкл/лунку лизата добавляют к подготовленному МТП. Для фонового измерения в лунки МТП добавляют по 100 мкл литического буфера.
Вторую концентрацию клеточного лизата используют в количестве 0,025 мг/мл, лизат разбавляют 1:2 и 100 мкл на лунку добавляют в подготовленный МТП. МТП инкубируют дополнительно в течение 2 ч при комнатной температуре в условиях встряхивания и затем 3 раза промывают раствором ФСБ, содержащим 0,1% Tween® 20.
Детекторным антителом для EGFR является биотинилированное антитело козы к EGFR человека в концентрации 36 мкг/мл, разведенное 1:180 в ФСБ, содержащем 3% БСА и 0,2% Tween® 20. Добавляют 100 мкл на лунку и инкубируют при комнатной температуре в течение 2 ч в условиях встряхивания. Затем МТП трижды промывают раствором ФСБ, содержащим 0,1% Tween® 20, 200 мкл/лунку. После этого добавляют по 100 мкг/лунку Streptavidin-HRP 1:200 в ФСБ, содержащем 3% БСА и 0,2% Tween® 20, и инкубируют при встряхивании в течение 20 мин при КТ. Затем планшет шесть раз промывают раствором ФСБ, содержащим 0,1% Tween® 20. Добавляют 100 мкл на лунку 3,3′-5,5′-тетраметилбензидина (фирма Roche, BM-Blue ID-No.: 11484581) и инкубируют в течение 20 мин при комнатной температуре при встряхивании. Цветную реакцию останавливают, добавляя 25 мкл на лунку 1М Н2SO4 и инкубируя в течение еще 5 мин при комнатной температуре. Оптическую плотность измеряют при длине волны 450 нм.
Для выявления IGF-1R
МТП со стрептавидином (фирма Roche, ID. No.: 11965891001) готовят, добавляя 100 мкг на лунку антитела AK1a-Biotinylated (фирма Genmab, Дания), которое разводят 1:200 в ФСБ, содержащем 3% БСА и 0,2% Tween® 20. МТП со стрептавидином инкубируют в течение 1 ч при комнатной температуре в условиях встряхивания и затем промывают трижды, добавляя по 200 мкл в лунку раствора ФСБ, содержащего 0,1% Tween® 20.
В лизатах клеток определяют количество белка, используя набор ВСА Protein Assay kit (фирма Pierce), затем лизаты клеток доводят до концентрации белка 0,3 мг/мл, используя 50 мМ трис, рН 7,4, 100 мМ Na3VO4 1:100 и протеазный ингибитор Complete® 1:20, и к подготовленным МТП со стрептавидином добавляют по 100 мкл/лунку лизата.
Используют вторую концентрацию клеточного лизата 0,15 мг/мл, для чего лизат разбавляют и по 100 мкл на лунку добавляют в подготовленный МТП со стрептавидином. Для фонового измерения в лунку МТП со стрептавидином добавляют 100 мкл лизирующего буфера.
МТП инкубируют дополнительно в течение 1 ч при комнатной температуре в условиях встряхивания и затем 3 раза промывают раствором ФСБ, содержащим 0,l%Tween® 20.
Детекторным антителом к IGF-1R является антитело кролика к IGF-IRβ человека (фирма Santa Cruz Biotechnology, номер в каталоге sc-713), разведенное 1:750 в ФСБ, содержащем 3% БСА и 0,2% Tween® 20. Добавляют по 100 мкл на лунку и инкубируют при комнатной температуре в течение 1 ч в условиях встряхивания. Затем МТП три раза промывают раствором ФСБ, содержащим 0,1% Tween® 20, в количестве 200 мкл/лунку. Затем добавляют вторичное антитело IgG-POD кролика (фирма Cell signaling, номер в каталоге 7074) 1:4000 в ФСБ, содержащем 3% БСА и 0,2% Tween® 20, 100 мкл на лунку, и инкубируют при встряхивании в течение 1 ч при комнатной температуре. Затем планшет шесть раз промывают раствором ФСБ, содержащим 0,1% Tween® 20. Добавляют 100 мкл на лунку 3,3′-5,5′-тетраметилбензидина (фирма Roche, BM-Blue ID-No.: 11484581) и инкубируют в течение 20 мин при комнатной температуре при встряхивании. Цветную реакцию останавливают, добавляя 25 мкл на лунку 1М H2SO4 и инкубируя в течение дополнительных 5 мин при комнатной температуре. Оптическую плотность измеряют при длине волны 450 нм.
Результаты выявления негативной регуляции рецепторов биспецифичными scFab-содержащими молекулами XGFR по сравнению с исходными моноспецифичными антителами <EGFR> ICR62 и <IGF-1R> HUMAB- клон 18 в клетках А549 показаны на фиг.13 и 14. Биспецифичные антитела scFab-XGFR подавляют и EGFR, и IGF1R. Это показывает, что полная функциональность (биологическая функциональность) и фенотипическое опосредование связывающих молекул сохраняется. Фиг.14 также показывает, что неожиданно биспецифичные антитела scFab-XGFR_2720 показали повышенное подавление EGFR по сравнению с одним исходным антителом <EGFR> ICR62.
Тот факт, что scFab-содержащие варианты XGFR при применении в таких же анализах при идентичной молярности показывают такую же активность или выше, что и антитела дикого типа, указывает на то, что молекулы scFab-XGFR 1 способны интерферировать с обоими сигнальными путями.
Пример 6. scFab-XGFR1 и scFab-ХGFR2-опосредованное ингибирование роста опухолевых клеточных линий in vitro
Антитело человека против IGF-1R <IGF-1R> HUMAB клон 18 (DSM АСС 2587) ингибирует рост опухолевых клеточных линий, которые экспрессируют IGF1R (WO 2005/005635). Сходным образом, показано, что гуманизированное антитело крысы против EGFR <EGFR> ICR62 ингибирует рост опухолевых клеточных линий, которые экспрессируют EGFR (WO 2006/082515). Для оценки потенциальной ингибиторной активности различных вариантов scFab-XGFR1 в исследованиях роста опухолевых клеточных линий степень ингибирования анализируют в клетках Н322М, которые экспресируют и EGFR, и IGF1R.
Клетки Н322М (5000 клеток/лунку) культивируют в среде RPMI 1640, содержащей 10% ФСТ, на покрытых поли-ГЭМА (поли(2-гидроксиэтилметакрилатом)) чашках, чтобы предотвратить адгезию на пластиковой поверхности. При этих условиях клетки Н322М образуют плотные сфероиды, которые растут в трех направлениях (свойство, которое называется якорной независимостью). Эти сфероиды имеют близкое сходство с трехмерной гистоархитектурой и организацией плотных опухолей in-situ. Сфероидные культуры инкубируют в течение 7 суток в присутствии 100 нМ антител. Для измерения ингибирования роста используют люминесцентный анализ Celltiter Glow. При обработке сфероидных культур Н322М антителом <IGF-1R> HUMAB-клон 18 наблюдают ингибирование роста.
Фиг.15 показывает, что применение 100 нМ <IGF-1R> HUMAB- клон 18 редуцирует клеточный рост на 72%, и что в том же анализе применение 100 нМ <EGFR> ICR62 редуцирует клеточный рост на 77%. Одновременное применение обоих антител (оба в одной и той же концентрации 100 нМ) приводит к полному подавлению жизнеспособности клеток (100% ингибирование). Это указывает на то, что одновременная интерференция с обоими метаболическими путями RTK имеет более сильное действие на опухолевые клеточные линии, чем интерференция только с одним путем. Применение различных scFab-XGFR1-вариантов в молярной концентрации 100 нМ приводит к более высокому-ингибированию роста, которое более ярко выражено по сравнению с эффектом, наблюдаемым в случае применения только одиночных молекул. В действительности, при концентрации антител 100 нМ, различные scFab-XGFR1-варианты показывают полное (100%) ингибирование клеточного роста, а применение одиночных модулей вызывает только частичное ингибирование.
В настоящем изобретении сделан вывод о том, что молекулы scFab-XGFR1 имеют крайне высокую активность ингибирования роста по сравнению с иммуноглобулинами IgG, которые исключительно интерферируют либо с передачей сигнала через EGFR, либо с передачей сигнала через IGF1R.
| название | год | авторы | номер документа |
|---|---|---|---|
| ТРИ- ИЛИ ТЕТРАСПЕЦИФИЧЕСКИЕ АНТИТЕЛА | 2010 |
|
RU2570633C2 |
| БИСПЕЦИФИЧЕСКИЕ АНТИ-VEGF/АНТИ-ANG-2 АНТИТЕЛА | 2009 |
|
RU2542382C2 |
| БИСПЕЦИФИЧЕСКИЕ АНТИ-VEGF/АНТИ-ANG-2 АНТИТЕЛА | 2014 |
|
RU2640253C2 |
| ГУМАНИЗИРОВАННЫЕ АНТИТЕЛА К CD19 ЧЕЛОВЕКА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2731156C1 |
| КОНСТРУКЦИИ МУЛЬТИСПЕЦИФИЧНЫХ АНТИТЕЛ | 2015 |
|
RU2710730C2 |
| АНТИТЕЛО-ПОДОБНЫЕ СВЯЗЫВАЮЩИЕ БЕЛКИ С ДВОЙНЫМИ ВАРИАБЕЛЬНЫМИ ОБЛАСТЯМИ, ИМЕЮЩИЕ ОРИЕНТАЦИЮ СВЯЗЫВАЮЩИХ ОБЛАСТЕЙ КРЕСТ-НАКРЕСТ | 2012 |
|
RU2695880C2 |
| АНТИ-LRP6 АНТИТЕЛА | 2011 |
|
RU2587625C2 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С GPRC5D | 2019 |
|
RU2797268C2 |
| ОДНОВАЛЕНТНЫЕ МОДУЛИ-ПЕРЕНОСЧИКИ ЧЕРЕЗ ГЕМАТОЭНЦЕФАЛИТИЧЕСКИЙ БАРЬЕР | 2014 |
|
RU2799436C1 |
| ОДНОВАЛЕНТНЫЕ МОДУЛИ-ПЕРЕНОСЧИКИ ЧЕРЕЗ ГЕМАТОЭНЦЕФАЛИЧЕСКИЙ БАРЬЕР | 2014 |
|
RU2694659C2 |












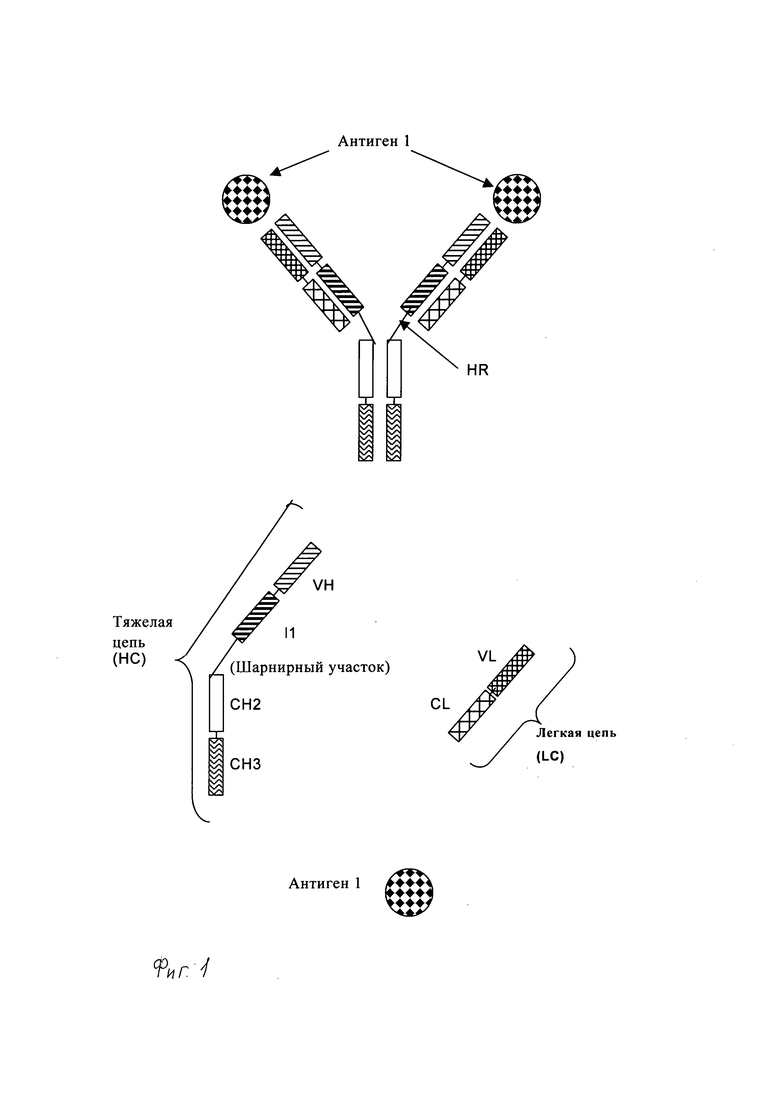
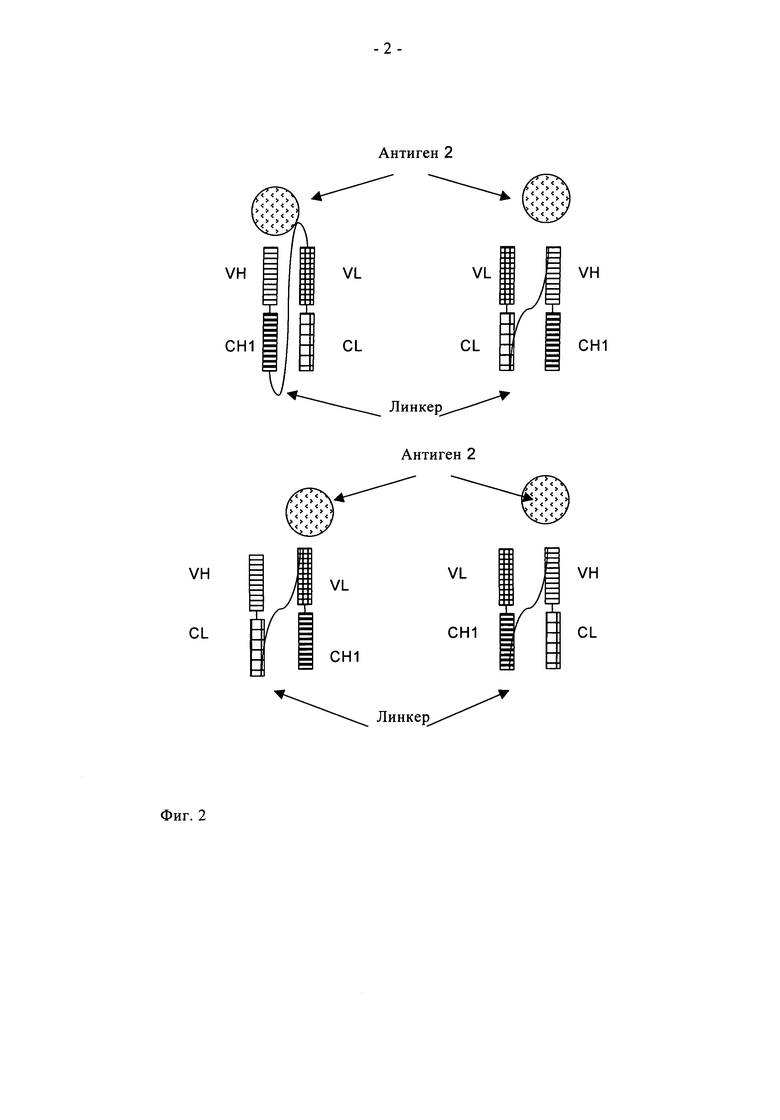
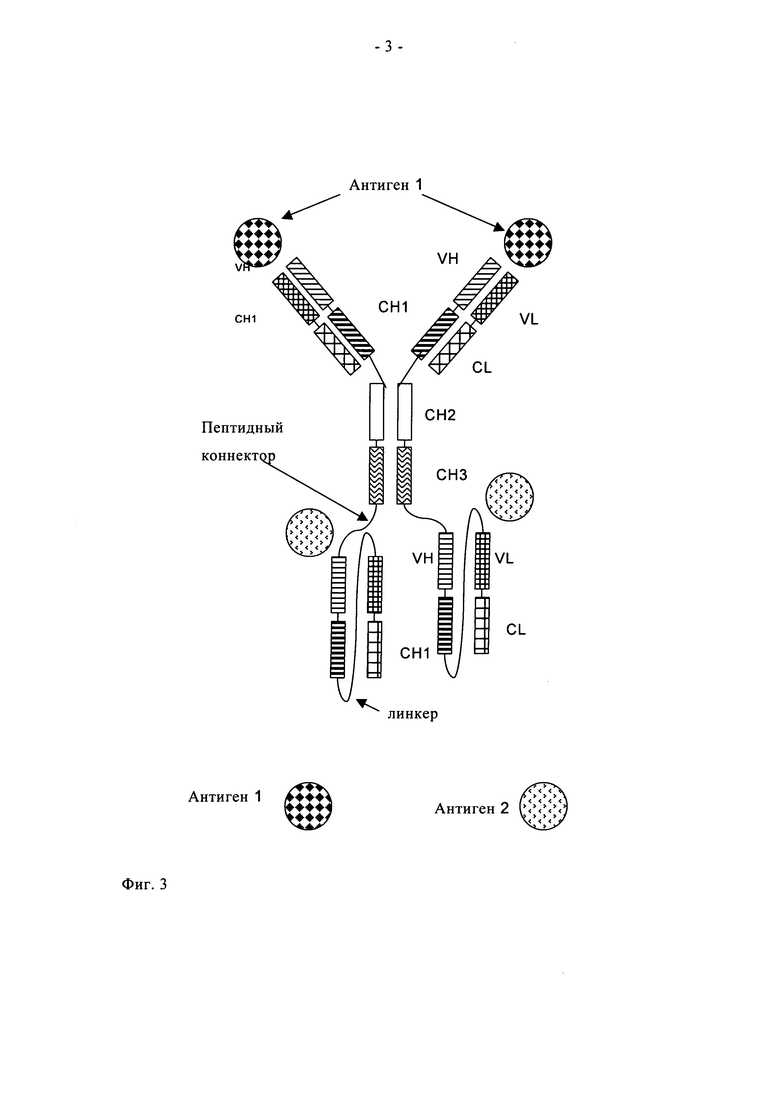
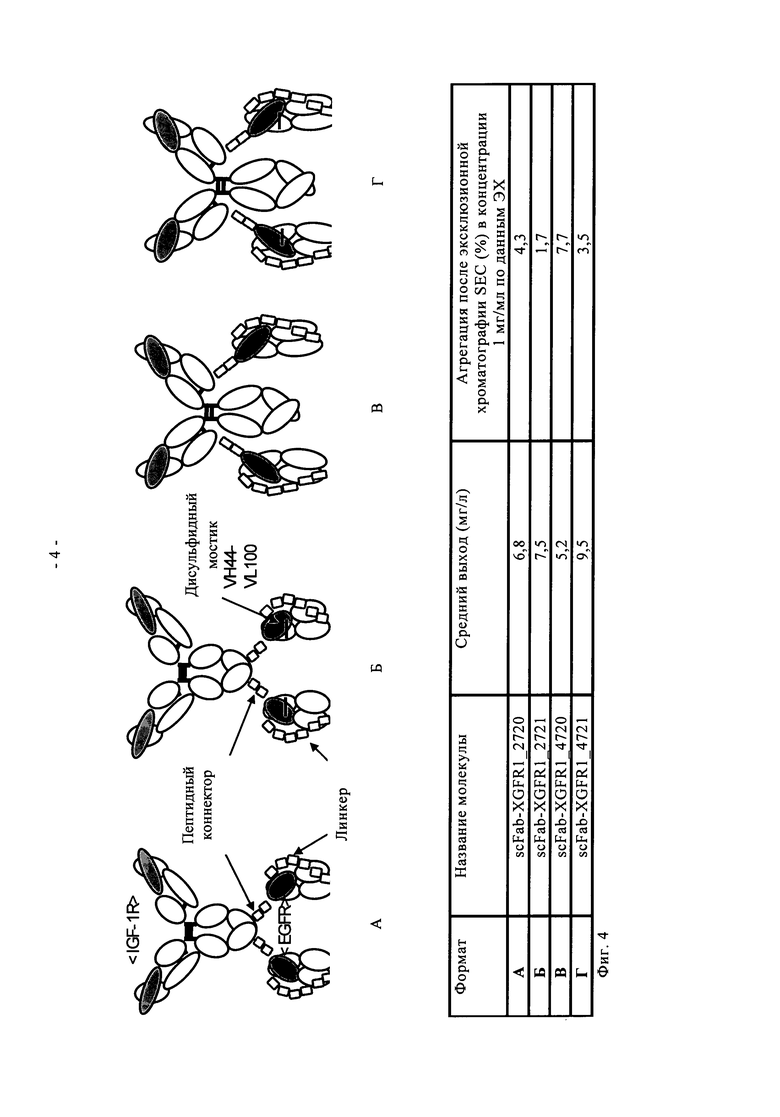
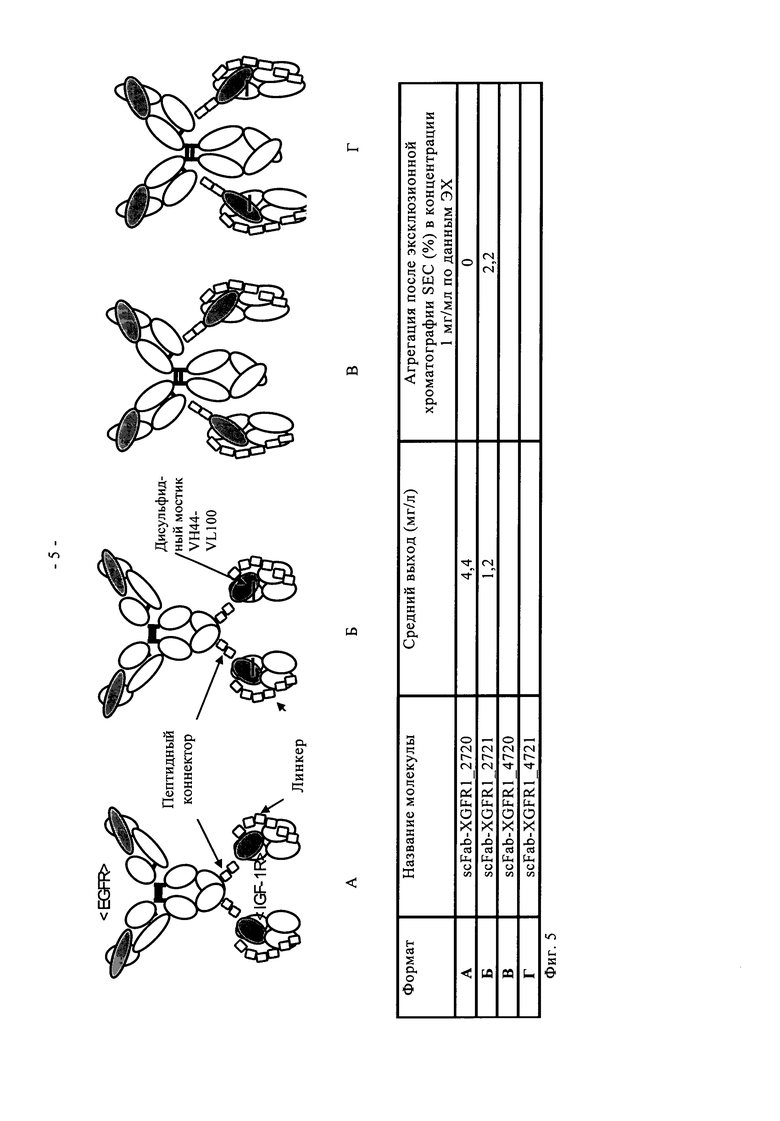
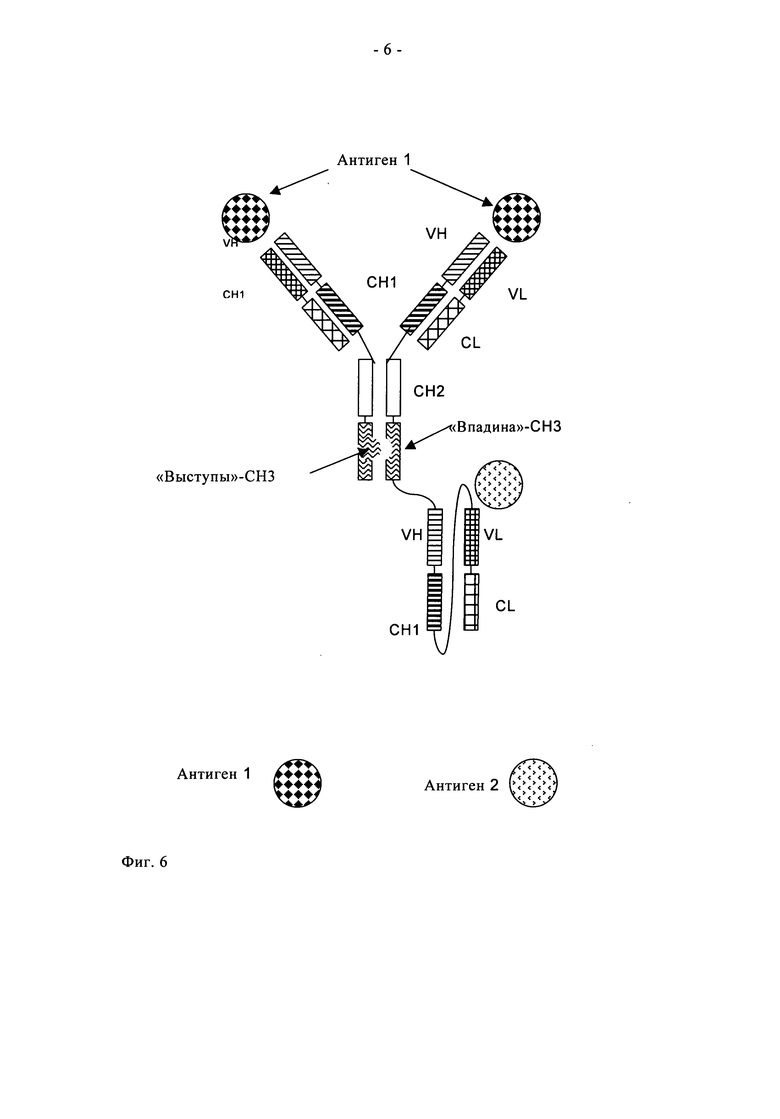
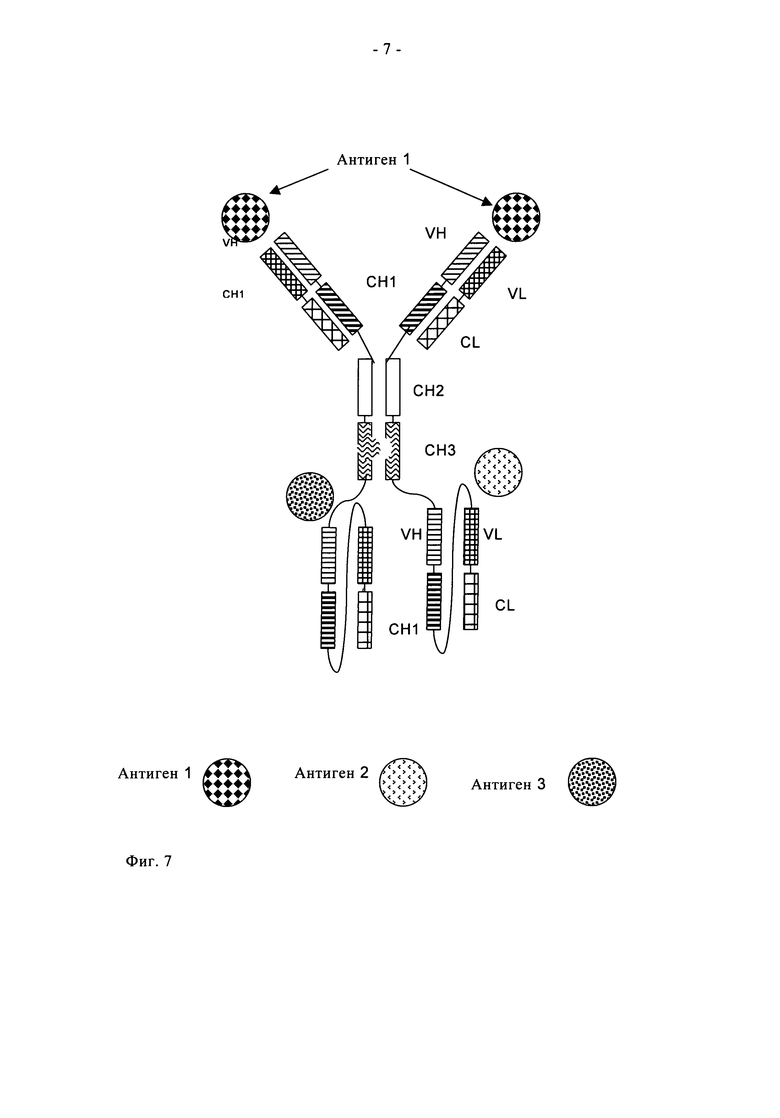

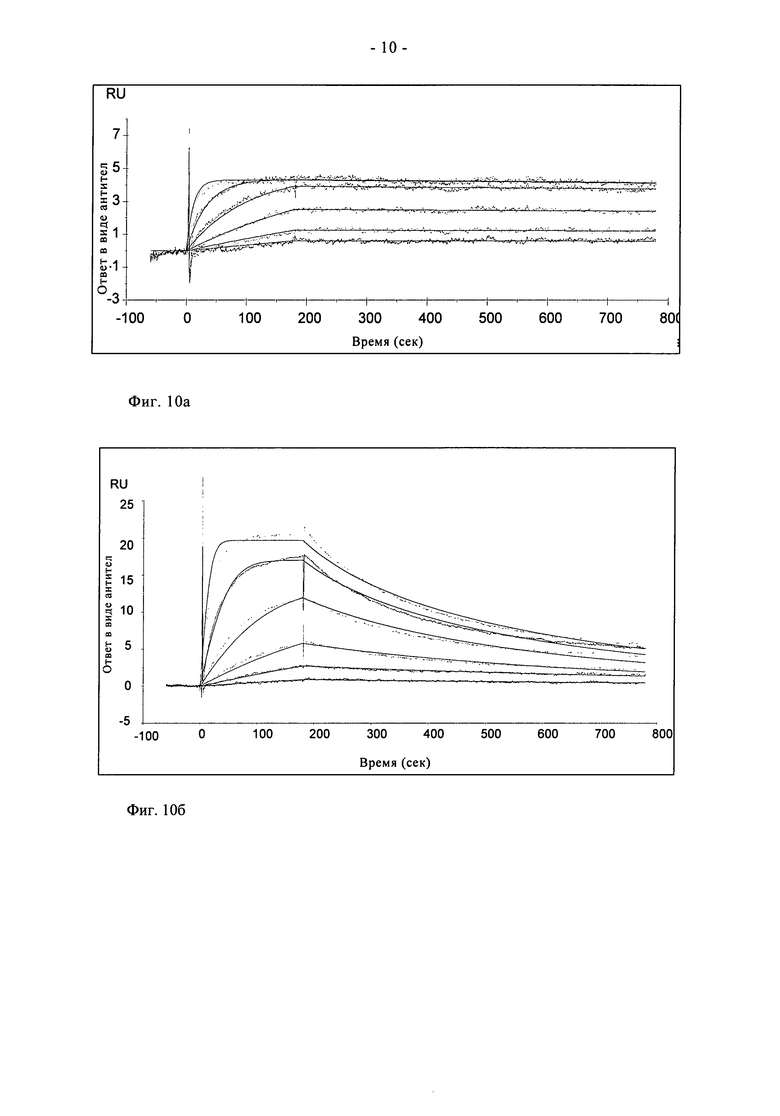
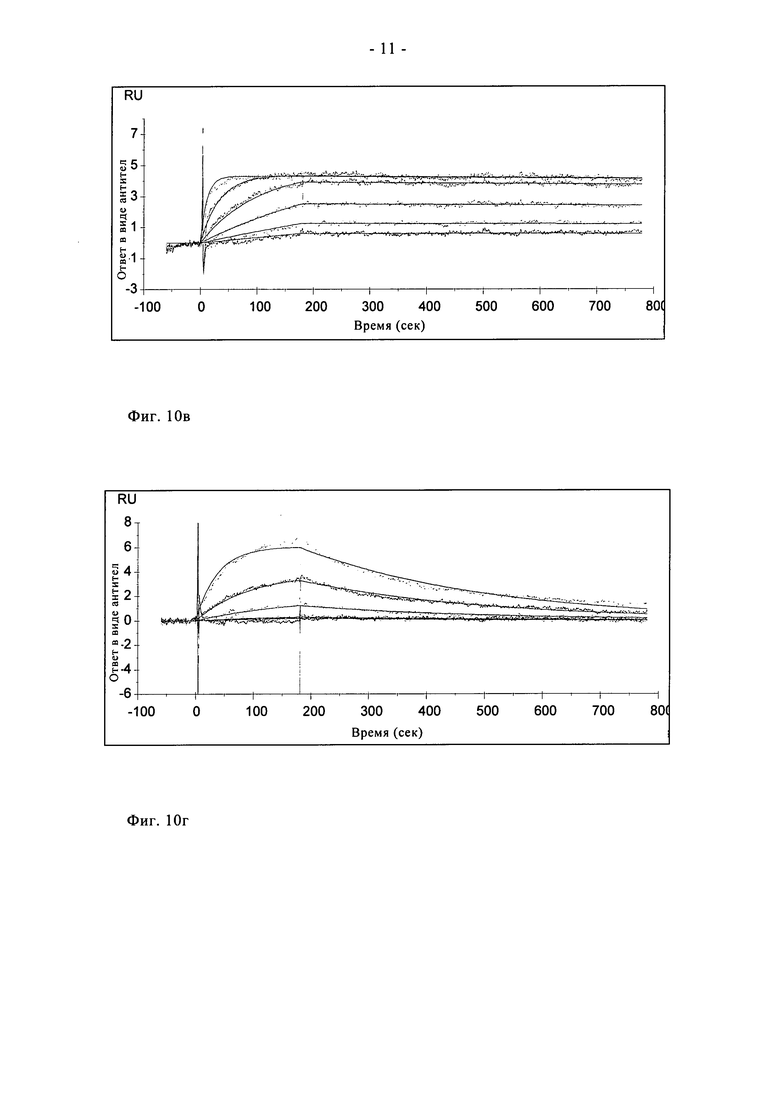
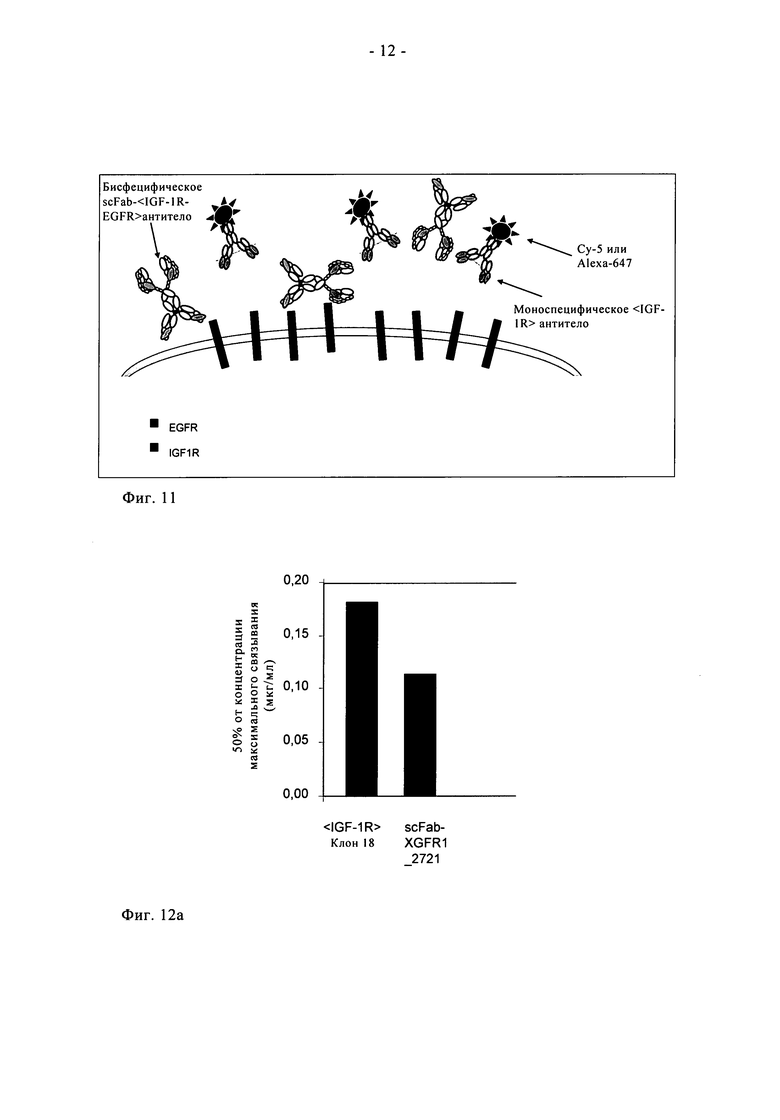
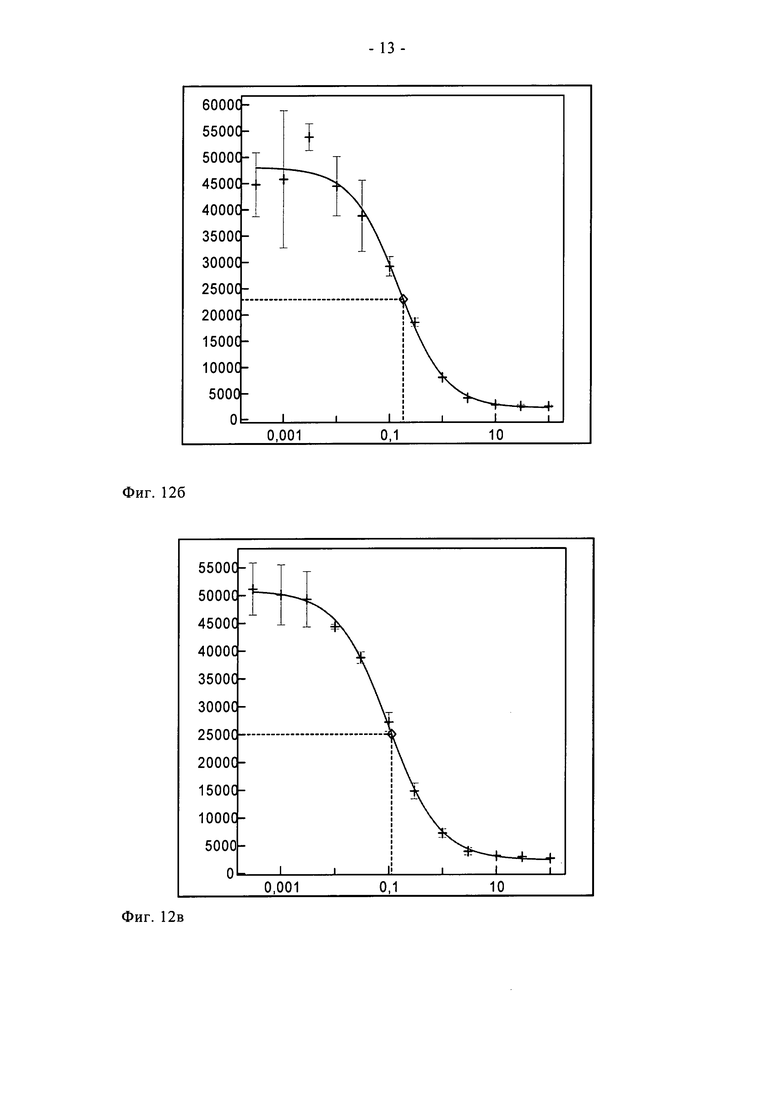
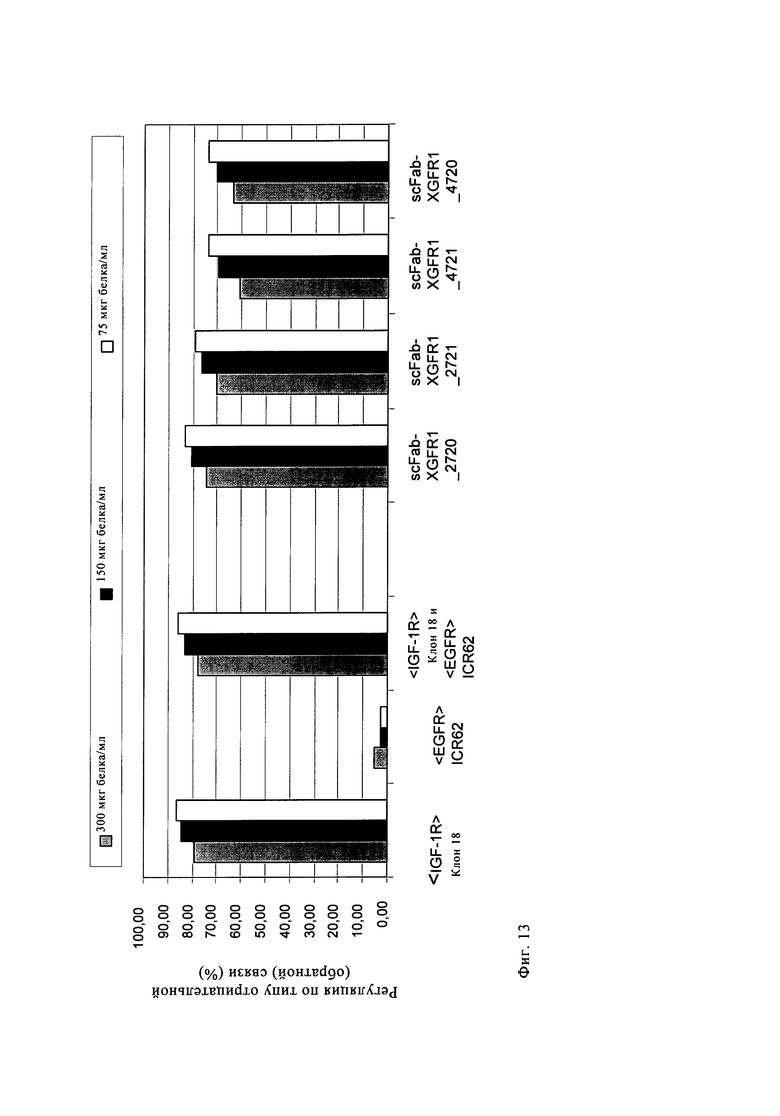
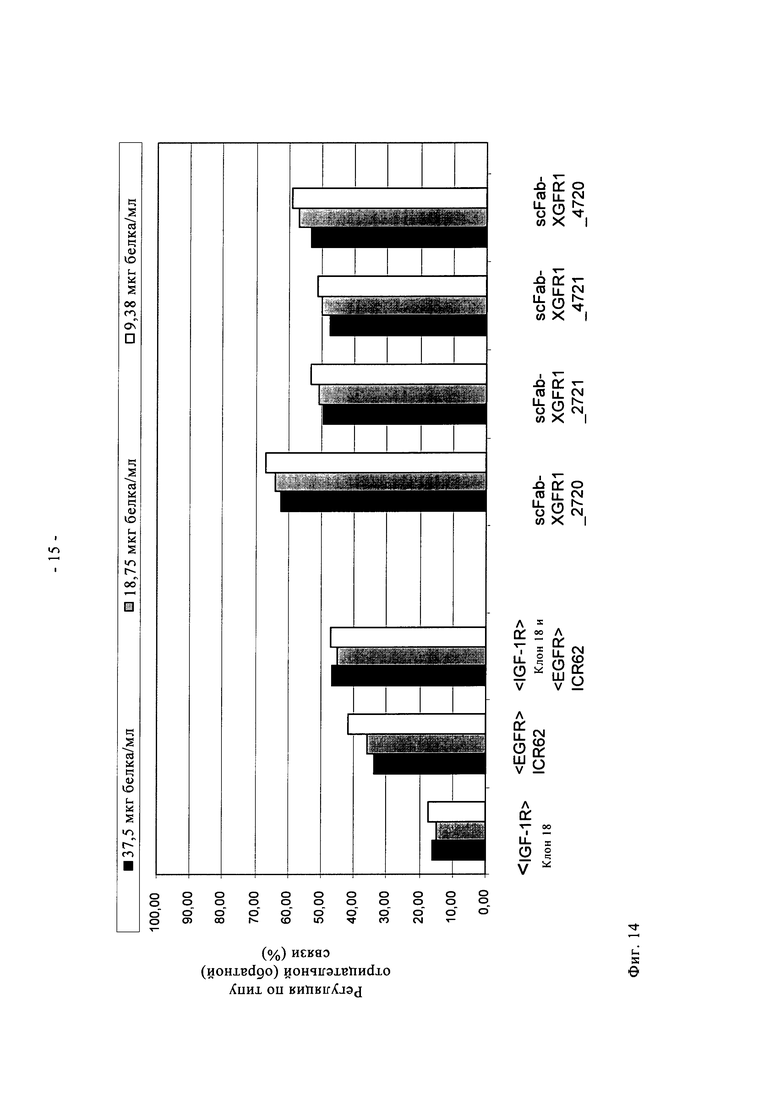

Настоящее изобретение относится к области биотехнологии и иммунологии. Предложен способ получения полиспецифичного антитела, включающего полноразмерное антитело, специфически связывающееся с первым антигеном и состоящее из двух тяжелых цепей антитела и двух легких цепей антитела, и один или несколько одноцепочечных фрагментов Fab, связывающихся с одним или несколькими дополнительными антигенами, в котором указанные одноцепочечные Fab-фрагменты соединены с указанным полноразмерным антителом через пептидный коннектор с С-конца тяжелой или легкой цепи указанного полноразмерного антитела, причем пептидный коннектор имеет структуру (GxS)n или (GxS)Gm, где G = глицин, S = серин, и (х = 3, n = 3, 4, 5 или 6 и m = 0, 1, 2 или 3) или (х = 4, n = 2, 3, 4 или 5 и m = 0, 1, 2 или 3). Полиспецифические антитела, полученные рассмотренным способом, обладают хорошей стабильностью и низкой склонностью к агрегации. В этой связи способ по настоящему изобретению может найти применение в приготовлении лекарственных средств. 4 з.п. ф-лы, 15 ил., 3 табл., 6 пр.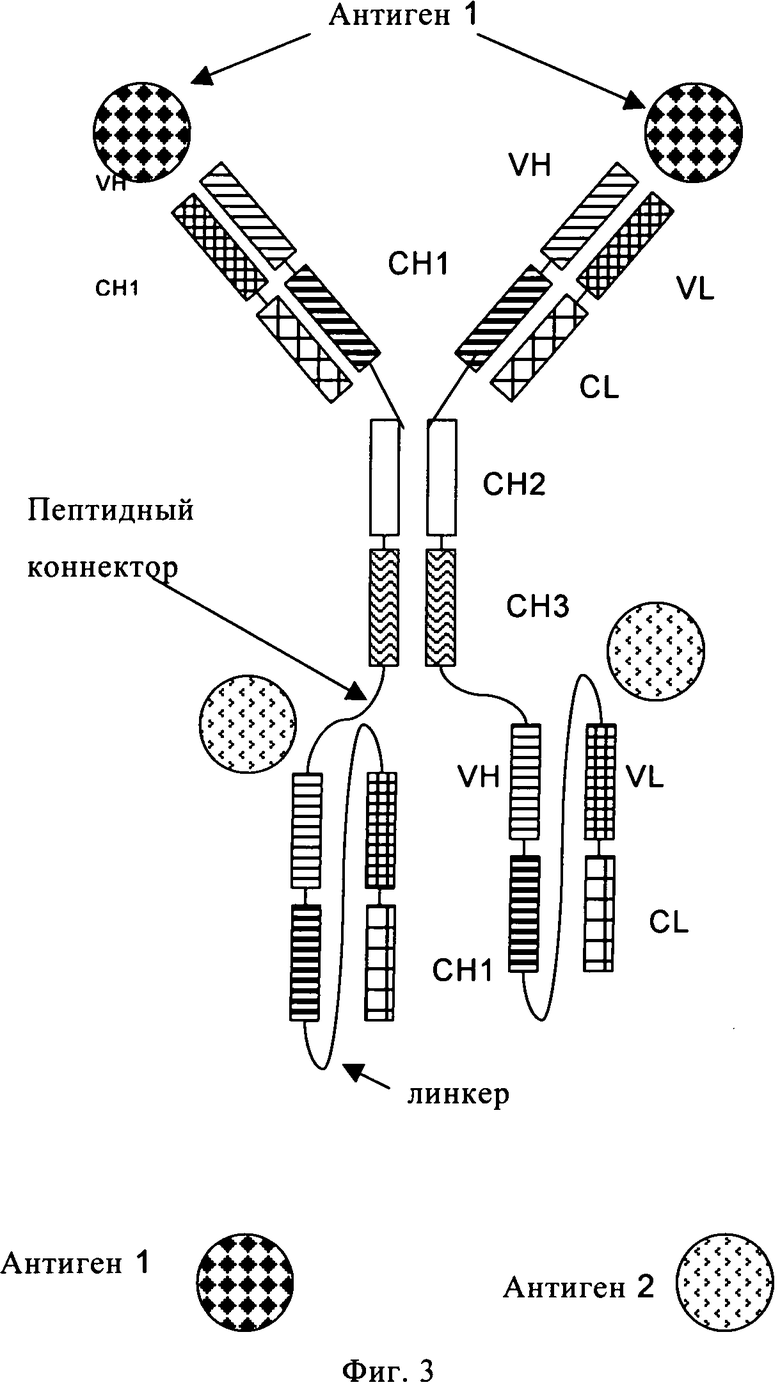
1. Способ получения полиспецифичного антитела, включающего
а) полноразмерное антитело, специфически связывающееся с первым антигеном и состоящее из двух тяжелых цепей антитела и двух легких цепей антитела, и
б) один или несколько одноцепочечных фрагментов Fab, связывающихся с одним или несколькими дополнительными антигенами,
в котором указанные одноцепочечные Fab-фрагменты по пункту б) соединены с указанным полноразмерным антителом по пункту а) через пептидный коннектор с С-конца тяжелой или легкой цепи указанного полноразмерного антитела, причем пептидный коннектор имеет структуру (GxS)n или (GxS)Gm, где G = глицин, S = серин, и (х = 3, n = 3, 4, 5 или 6 и m = 0, 1, 2 или 3) или (х = 4, n = 2, 3, 4 или 5 и m = 0, 1, 2 или 3),
где способ включает:
синтез нуклеиновых кислот, кодирующих соответствующие полипептиды полиспецифичного антитела,
встраивание этих нуклеиновых кислот в векторы экспрессии,
трансформацию полученными векторами клетки-хозяина,
культивирование клетки-хозяина,
с последующим выделением и очисткой из культуральной среды полиспецифичного антитела.
2. Способ по п.1,
где из культуральной среды выделяют и очищают
полиспецифичное антитело, включающее
а) полноразмерное антитело, специфически связывающееся с первым антигеном и состоящее из двух тяжелых цепей антитела и двух легких цепей антитела, и
б) от одного до четырех одноцепочечных фрагментов Fab, которые связывают от одного до четырех дополнительных антигенов,
в котором указанные одноцепочечные фрагменты Fab по пункту б) соединены с указанным полноразмерным антителом по пункту а) через пептидный коннектор с С-конца тяжелой или легкой цепи указанного полноразмерного антитела.
3. Способ по п.1 или 2,
где из культуральной среды выделяют и очищают
полиспецифичное антитело, в котором один или два одноцепочечных фрагмента Fab, связывающихся со вторым антигеном, соединены с указанным полноразмерным антителом через пептидный коннектор с С-конца тяжелых цепей указанного полноразмерного антитела.
4. Способ по п.1 или 2,
где из культуральной среды выделяют и очищают
полиспецифичное антитело, в котором один одноцепочечный фрагмент Fab, связывающийся со вторым антигеном, соединен с указанным полноразмерным антителом через пептидный коннектор с С-конца одной тяжелой цепи или одной легкой цепи указанного полноразмерного антитела.
5. Способ по п.1 или 2,
где из культуральной среды выделяют и очищают
полиспецифичное антитело, в котором два идентичных одноцепочечных фрагмента Fab VL-CL-линкер-VH-СН1 или VH-СН1-линкер-VL-CL, связывающихся со вторым антигеном, соединены своими N-концами с указанным полноразмерным антителом через пептидный коннектор с двух С-концов двух тяжелых цепей или с двух С-концов двух легких цепей указанного полноразмерного антитела.
| Пресс для выдавливания из деревянных дисков заготовок для ниточных катушек | 1923 |
|
SU2007A1 |
| LU D | |||
| et al | |||
| "Simultaneous blockade of both the epidermal growth factor receptor and the insulin-like growth factor receptor signaling pathways in cancer cells with a fully human recombinant bispecific antibody", Journal of Biological Chemistry, 2004, 279(4): 2856-2865 | |||
| Прибор для очистки паром от сажи дымогарных трубок в паровозных котлах | 1913 |
|
SU95A1 |
| MILLER K | |||
| et al., | |||

