Настоящее изобретение относится к способу получения аминокислот и кетокислот с использованием микроорганизмов, в которых промотор, индуцируемый пропионатом, дает возможность регулировать экспрессию определенных генов.
Уровень техники
Аминокислоты и кетокислоты используют в медицине человека, в фармацевтической промышленности, в косметике, в пищевой промышленности и в кормлении животных.
Многие из этих соединений получают путем ферментации штаммов коринеформных бактерий, в частности, Corynebacterium glutamicum. Из-за большой важности постоянно проводится работа по усовершенствованию способа получения. Усовершенствования способа может включать в себя воздействия на технологию ферментации, такую как, например, перемешивание и подача кислорода, или воздействия на состав питательных сред, такой как, например, изменение концентрации сахара во время ферментации, или воздействие на форму продукта с помощью, например, ионообменной хроматографии или внутренних характеристик продуктивности самого микроорганизма.
Для усовершенствования характеристик продуктивности этих микроорганизмов используют способы мутагенеза, селекции и отбора мутантов. Таким образом, получают штаммы, устойчивые к антиметаболитам, таким как, например, аналог валина 2-тиазолаланин или аналог лейцина 4-азалейцин, или 5,5,5-трифторлейцин, и которые производят химические соединения, например, L-аминокислоты L-валин или L-лейцин.
В течение нескольких лет также использовались способы с методиками рекомбинантной ДНК для усовершенствования производящих L-аминокислоты штаммов Corynebacterium glutamicum, например, путем усиления или ослабления генов биосинтеза отдельных аминокислот, например, также с временной регуляцией экспрессии генов в ходе производства, и исследования влияния на получение химического соединения.
Обобщающие описания биологии, генетики и биотехнологии Corynebacterium glutamicum можно обнаружить в “Handbook of Corynebacterium glutamicum” (Ред.: L. Eggeling and M. Bott, CRC Press, Taylor & Francis, 2005), в специальном издании Journal of Biotechnology (главный редактор: A.  ) под названием “A New Era in Corynebacterium glutamicum Biotechnology” (Journal of Biotechnology 104/1-3, (2003)) и в книге T. Scheper (Ответственный редактор) “Microbial Production of L-Amino Acids” (Advances in Biochemical Engineering/Biotechnology 79, Springer Verlag, Berlin, Germany 2003).
) под названием “A New Era in Corynebacterium glutamicum Biotechnology” (Journal of Biotechnology 104/1-3, (2003)) и в книге T. Scheper (Ответственный редактор) “Microbial Production of L-Amino Acids” (Advances in Biochemical Engineering/Biotechnology 79, Springer Verlag, Berlin, Germany 2003).
Нуклеотидная последовательность генома Corynebacterium glutamicum описана у Ikeda and Nakagawa (Applied Microbiology and Biotechnology 62, 99-109 (2003)), в EP 1108790 и в Kalinowski et al. (Journal of Biotechnology 104/1-3, (2003)).
Нуклеотидные последовательности генома Corynebacterium glutamicum также доступны в базе данных Национального Центра Биотехнологической информации (NCBI) Национальной библиотеки медицины (Бетесда, Мэриленд, США), в Банке данных ДНК Японии (DDBJ, Мишима, Япония) или в базе данных нуклеотидных последовательностей Европейских лабораторий молекулярной биологии (EMBL, Хейдельберг, Германия и Кембридж, Соединенное Королевство).
По сути, существует две возможности для экспрессии генов. При непрерывной экспрессии ген непрерывно экспрессируют посредством конститутивного промотора и соответствующий белок накапливается в клетке.
С другой стороны, индуцибельный промотор используют при индуцированной экспрессии. Экспрессию гена-мишени индуцируют, то есть запускают, посредством индуктора. Этот способ используют, если (сверх)экспрессия имеет отрицательное воздействие на производящий организм. Причинами этого может быть высокая загрузка метаболических ресурсов во время фазы роста.Результатом является более медленный рост и, таким образом, удлиненный рабочий цикл биореактора и связанное с этим увеличение затрат в случае промышленного получения. Индуцированная экспрессия также является преимущественной в случае цитотоксических продуктов. При этом аутоинтоксикация и гибель клетки происходит после индукции экспрессии. В связи с экономикой способа получения существует, следовательно, попытка подразделить способ на фазу роста и фазу образования продукта. В фазе роста получают как можно большее количество биомассы и затем в фазе образования продукта получают целевой белок путем индукции промотора. Таким образом можно получить максимальный выход продукта, посредством чего процесс становится заметно более экономичным.
Для регулируемых промоторов из Escherichia coli, lac, lambda PL и trp, уже было показано, что их можно использовать в коринеформных бактериях для регулируемой экспрессии различных генов (Tsuchiya and Morinaga, Bio/Technology 6 (1988) 428-431).
Идеальным случаем для индуцибельного промотора является коринеформный промотор, который регулируется легкодоступным, недорогим веществом.
В ЕР 0530765 В1 (Kyowa Hakko) описано применение индуцибельного промотора из коринебактерий, в данном случае из гена изоцитратлиазы, для получения ферментов, таких как [бета]-галактозидаза, хлорамфениколацетилтрансфераза и ICL, а также физиологически активных белков, таких как инсулин или [альфа]-, [бета]- или [гамма]-интерферон. Этот промотор приводит к экспрессии генов так долго, пока источник углерода (источник С), отличный от сахара, находится в среде; в присутствии сахара он репрессирован. Поскольку, однако, сахара используют в качестве источника С в обычных ферментационных средах, было бы полезно получить регулируемый промотор, который даже в присутствии сахаров приводит к экспрессии гена с использованием недорогого индуктора.
В DE 4440118 C1 (таком как US 5965391, F.Z.  ) заявлено применение индуцибельного промотора из коринебактерий, в данном случае из гена aceB малатсинтазы, для производства белков; индукторами при этом являются источники углерода лактат, пируват или ацетат.
) заявлено применение индуцибельного промотора из коринебактерий, в данном случае из гена aceB малатсинтазы, для производства белков; индукторами при этом являются источники углерода лактат, пируват или ацетат.
Промотор оперона prpDBC2 из Corynebacterium glutamicum, гены которого необходимы в качестве основного источника С для роста на пропионате и кодируют ферменты 2-метилцитратдегидратазу (PrpD2), 2-метилизоцитратлиазу (PrpB2) и 2-метилцитратсинтазу (PrpC2), описан Plassmeier et al. (Journal of Biotechnology 159/1-2 (2012)). Анализ промоторных конструкций тестируемых векторов привел к идентификации операторного участка длиной 121 пара оснований выше оперона prpDBC2, который необходим для индуцируемой пропионатом транскрипции с помощью активатора PrpR. Исследования EMSA показали, что 2-метилцитрат вероятно функционирует в качестве коактиватора PrpR.
Цель настоящего изобретения
Настоящее изобретение основано на намерении сделать доступным новый способ получения L-аминокислот L-лейцина, L-валина или L-изолейцина, предпочтительно L-валина, или α-кетокислот α-кетоизовалерата, α-кетометилвалерата или α-кетоизокапроата с предпочтительно улучшенным выходом и/или более высокой конечной концентрацией продукта внутриклеточно и/или в среде. При этом предпочтительно должно присутствовать улучшение специфического выхода (т.е. выхода желаемого продукта относительно использованного источника углерода).
Новый способ предпочтительно должен делать возможной регуляцию получения L-аминокислот и/или α-кетокислот независимо от главного источника углерода в среде.
В новом способе образование нежелательных побочных продуктов, в частности, образование нежелательного побочного продукта аланина, должно, кроме того, предпочтительно быть подавлено, если это возможно, поскольку отделение побочного продукта является очень трудоемким и дорогостоящим и, кроме того, имеет отрицательное воздействие на чистоту продуктов бульона и выход углерода.
Используемый способ должен, кроме того, предпочтительно приводить к увеличению генетической стабильности штамма, используемого для получения, и тем самым обеспечивать большое количество поколений в ферментационном процессе (стадий культивирования и основной ферментер) без снижения параметров производительности.
Описание настоящего изобретения
Цели согласно настоящему изобретению достигают путем применения оператора, с которым связывается активатор PrpR для регуляции экспрессии гена ilvBN.
ilvBN (EC No. 4.1.3.18) представляет собой ген, кодирующий субъединицы ацетолактатсинтазы.
Объектом настоящего изобретения, таким образом, является способ получения L-аминокислоты, выбранной из L-лейцина, L-валина и L-изолейцина, предпочтительно L-валина, или α-кетокислоты, выбранной из кетоизовалерата, кетометилвалерата и кетоизокапроата путем ферментации микроорганизмов из рода Corynebacterium, содержащих в реплицируемой форме полинуклеотид с операторной активностью, последовательность которого по меньшей мере на 85%, предпочтительно, по меньшей мере на 90, 92, 94 или 96%, в частности, по меньшей мере на 97, 98 или 99%, особенно предпочтительно на 100% идентична последовательности от положения с 1 по 121 согласно SEQ ID NO: 1, 2 или 3, с которым связывается активатор PrpR и функционально ниже которого на 3'-конце находится второй полинуклеотид, имеющий промоторную активность, а также гены ilvB и ilvN, кодирующие субъединицы ацетолактатсинтазы, и который регулирует транскрипцию генов ilvBN в зависимости от добавления активатора PrpR, который активируют посредством коактиватора 2-метилцитрата в среде, к которой после первой фазы без индуктора добавляют пропионат или 2- метилцитрат в последующей второй фазе в качестве индуктора, после чего экспрессируют гены ilvBN и таким образом синтезируют желаемую L-аминокислоту или α-кетокислоту в условиях, при которых увеличивается содержание желаемой L-аминокислоты или α-кетокислоты в среде или клетках.
Полинуклеотид, характеризующийся операторной активностью, предпочтительно представляет собой оператор, который естественным образом регулирует экспрессию 2-метилцитратдегидратазы у коринеформных бактерий, или полинуклеотид, полученный из такого оператора.
Полинуклеотид с операторной активностью предпочтительно содержит полинуклеотид, последовательность которого по меньшей мере на 90%, предпочтительно, по меньшей мере на 92, 94 или 96%, в частности, по меньшей мере на 97, 98 или 99%, особенно предпочтительно на 100% идентична последовательности согласно SEQ ID NO: 11 или последовательности от положения 22 до положения 49 согласно SEQ ID NO: 1, 2 или 3 (в дальнейшем также названная “IR 1”), а также полинуклеотид, последовательность которого по меньшей мере на 90%, предпочтительно, по меньшей мере на 92, 94 или 96%, в частности, по меньшей мере на 97, 98 или 99%, особенно предпочтительно на 100% идентична последовательности согласно SEQ ID NO: 12 или последовательности от положения 77 до положения 105 согласно SEQ ID NO: 1, 2 или 3 (в дальнейшем также названная “IR 2”). Последовательность “IR 1” при этом расположена в положении с 22 по 49 в предпочтительном варианте осуществления в полинуклеотиде с операторной активностью, в то время как последовательность “IR 2” расположена в положении с 77 по 105 в предпочтительном варианте осуществления.
В предпочтительном варианте осуществления согласно настоящему изобретению полинуклеотид с операторной активностью является частью более длинного полинуклеотида, предпочтительно полинуклеотида, имеющего идентичность последовательности по меньшей мере 90%, предпочтительно, по меньшей мере 92, 94 или 96%, в частности, по меньшей мере 97, 98 или 99%, особенно предпочтительно 100% с последовательностью от положения 1 до 177 согласно SEQ ID NO: 1, 2 или 3.
Ген ilvB предпочтительно представляет собой полинуклеотид, кодирующий полипептид, имеющий аминокислотную последовательность, которая имеет идентичность по меньшей мере 90%, предпочтительно, по меньшей мере 92, 94 или 96%, в частности, по меньшей мере 97, 98 или 99%, особенно предпочтительно 100% с аминокислотной последовательностью согласно SEQ ID NO: 9.
Особенно предпочтительным при этом является полинуклеотид, последовательность которого по меньшей мере на 90%, предпочтительно, по меньшей мере на 92, 94 или 96%, в частности, по меньшей мере на 97, 98 или 99%, особенно предпочтительно на 100% идентична последовательности от положения 499 до 2379 согласно SEQ ID NO: 8.
Ген ilvN предпочтительно представляет собой полинуклеотид, кодирующий полипептид, имеющий аминокислотную последовательность, которая имеет идентичность по меньшей мере 90%, предпочтительно, по меньшей мере 92, 94 или 96%, в частности, по меньшей мере 97, 98 или 99%, особенно предпочтительно 100% с аминокислотной последовательностью согласно SEQ ID NO: 10.
Особенно предпочтительным при этом является полинуклеотид, последовательность которого по меньшей мере на 90%, предпочтительно, по меньшей мере на 92, 94 или 96%, в частности, по меньшей мере на 97, 98 или 99%, особенно предпочтительно на 100% идентична последовательности от положения 2393 до 2911 согласно SEQ ID NO: 8.
Полипептиды, кодируемые генами ilvB и ilvN, агломерируют друг с другом с получением функциональной ацетолактатсинтазы.
Пропионат или пропионовую кислоту предпочтительно используют для индукции экспрессии. В этом отношении имеет место "индукция пропионатом". Пропионат превращают in vitro в фактический коактиватор 2-метилцитрат. В качестве альтернативы, 2-метилцитрат также можно использовать непосредственно для индукции, но его применяют с меньшей предпочтительностью из-за более низкой доступности. Согласно настоящему изобретению выражение "индукция пропионатом" также включает индукцию 2-метилцитратом.
После завершения получения желаемую L-аминокислоту или α-кетокислоту предпочтительно выделяют, другие компоненты ферментационного бульона и/или биомассы необязательно остаются во всей их полноте или частично (> 0 до 100%) в выделенном продукте или полностью удаляются.
В способе согласно настоящему изобретению сначала имеет место фаза культивирования без индукции (фаза роста, первая фаза) для обеспечения биомассы. В этой фазе предпочтительно не образуют или почти не образуют никакую аминокислоту или кетокислоту (< 5 г/л). В последующей фазе получения (фаза индукции, вторая фаза) получение индуцируют путем индукции генов биосинтеза ilvBN с помощью пропионата или 2-метилцитрата. Фаза культивирования включает в себя все этапы культивирования, начиная с сохраненного образца, через предпочтительно используемые стадии встряхиваемого сосуда до предпочтительно используемого культурального ферментера. Фаза культивирования может также еще включать в себя первую фазу основной ферментации. Предпочтительно, фазу культивирования завершают не позднее, чем после первых 3-15 часов, предпочтительно 5-10 часов основной ферментации.
Фаза индукции предпочтительно включает в себя время от 0 до 20 часов после инокуляции основного ферментера вплоть до конца основной ферментации. Индукция в более ранний момент времени, то есть в культуральном ферментере, может быть преимущественной и представляет собой специфический вариант осуществления способа согласно настоящему изобретению.
Точная регуляция/замер концентрации пропионовой кислоты в фазе получения продукта необходим для того, чтобы, с одной стороны, поддерживать индукцию синтеза аминокислот или кетокислот, а с другой стороны, чтобы предотвратить токсическое действие пропионата или одного из промежуточных продуктов его разложения (пропионил-КоА, 2-метилцитрат). Предпочтительная концентрация пропионовой кислоты в фазе индукции лежит в диапазоне от 0,1 - 10 г/л. Пропионовую кислоту можно добавлять непрерывно в виде подкормки с пропионовой кислотой или периодически в виде одного или нескольких импульсов пропионовой кислоты в различные моменты времени в течение фазы индукции. Однократная доза пропионовой кислоты в качестве компонента среды в основной ферментационной среде также является возможной и представляет собой специфический вариант осуществления способа согласно настоящему изобретению.
Большой проблемой в производстве аминокислот и кетокислот является, как правило, образование побочных продуктов. Образованные побочные продукты снижают выход углерода и должны быть, кроме того, необязательно отделены, что является очень трудоемким и дорогостоящим.
Примером образования побочного продукта является образование побочного продукта аланина при биосинтезе валина. Образование аланина увеличивается в общепринятых способах получения, при которых обычно имеет место конститутивная экспрессия, в зависимости от числа клеточных поколений.
От архивных образцов до сбора урожая из ферментера для получения продукта в способе, состоящем из трех стадий (встряхиваемый сосуд, культуральный ферментер, основной ферментер), проходят приблизительно 20-25 поколений, способе, состоящем из четырех стадий (встряхиваемый сосуд, предпосевный ферментер, культуральный ферментер, основной ферментер), проходят даже более чем 30 поколений. При прохождении через такое большое количество поколений образование побочного продукта и образование биомассы, как правило, соответственно значительно увеличено.
Согласно настоящему изобретению, однако, обнаружено очень сильное снижение образования побочного продукта и образования биомассы. Более того, было обнаружено, что образование побочного продукта и образование биомассы в способах получения согласно настоящему изобретению не зависит от числа пройденных поколений. Это предполагает, что управление процессом согласно настоящему изобретению ведет к повышению генетической стабильности используемого штамма. Фазу культивирования можно поэтому, в принципе, расширить до любой продолжительности, что особенно выгодно для управления процессом.
Особенно предпочтительный способ согласно настоящему изобретению, таким образом, отличается тем, что он включает в себя по меньшей мере четыре стадии, а именно, по меньшей мере три стадии культивирования и стадию получения продукта. Культивирование при этом предпочтительно включает в себя культивирование во встряхиваемом сосуде, в предпосевном ферментере, а также в посевном ферментере. Производство предпочтительно имеет место в ферментере для получения продукта.
Дополнительный особенно предпочтительный способ согласно настоящему изобретению, таким образом, отличается тем, что бактерии, используемые в фазе культивирования, проходят по меньшей мере 16, предпочтительно, по меньшей мере 24 поколения и/или в течение всей ферментации (включая культивирование и получение) проходят по меньшей мере 25, предпочтительно, по меньшей мере 30 поколений.
Настоящее изобретение также относится к применению полинуклеотида, характеризующегося операторной активностью, с которым связывается активатор PrpR, при этом полинуклеотид обладает последовательностью, которая по меньшей мере на 85%, 90%, 92%, 94%, 96%, 97%, 98%, 99% или 100%, предпочтительно, по меньшей мере на 97%, особенно предпочтительно, по меньшей мере на 98%, наиболее предпочтительно, по меньшей мере на 99% и чрезвычайно предпочтительно, на 100% идентична последовательности от положения 1 до 121 согласно SEQ ID NO: 1, 2 или 3 для регуляции экспрессии генов ilvBN, предпочтительно в комбинации с промотором, расположенным выше данных генов.
Настоящее изобретение также относится к кассете экспрессии, содержащей полинуклеотид, характеризующийся операторной активностью, последовательность которого по меньшей мере на 85%, 90%, 92%, 94%, 96%, 97%, 98%, 99% или 100%, предпочтительно, по меньшей мере на 97%, особенно предпочтительно, по меньшей мере на 98%, наиболее предпочтительно, по меньшей мере на 99% и чрезвычайно предпочтительно, на 100% идентична последовательности от положения 1 до 121 согласно SEQ ID NO:1, 2 или 3, расположенный ниже промотор и гены ilvBN, кодирующие ацетолактатсинтазу.
Полинуклеотид, характеризующийся операторной активностью, при этом предпочтительно всегда имеет характеристики, выделенные ранее как предпочтительные, в частности, консервативные области “IR 1” и “IR 2”.
Промотор, расположенный выше генов ilvBN, или промотор, расположенный ниже оператора, может быть любым желательным промотором.
Примеры промоторов согласно настоящему изобретению, предпочтительно используемых в Corynebacterium glutamicum, описаны, например, на Фиг. 1 обзорной статьи Patek et al. (Journal of Biotechnology 104(1-3), 311-323 (2003)). Таким же образом можно использовать варианты промотора dapA, например, промотор A25, описанный Vasicova et al. (Journal of Bacteriology 181, 6188-6191 (1999)). Кроме того, можно использовать промотор gap из Corynebacterium glutamicum (EP 06007373). Наконец, можно использовать хорошо известные промоторы T3, T7, SP6, M13, lac, tac и trc, описанные Amann et al. (Genes 69(2), 301-315 (1988)) и Amann and Brosius (Genes 40(2-3), 183-190 (1985)).
В предпочтительном варианте осуществления промотор, используемый согласно настоящему изобретению, представляет собой полинуклеотид, имеющий промоторную активность, последовательность которого по меньшей мере на 90%, 92%, 94%, 96%, 97%, 98%, 99% или 100%, предпочтительно, по меньшей мере на 97%, особенно предпочтительно, по меньшей мере на 98%, наиболее предпочтительно, по меньшей мере на 99% и, чрезвычайно предпочтительно, на 100% идентична последовательности от положения 122 до 206 согласно SEQ ID NO: 4.
Кассета экспрессии согласно настоящему изобретению представляет собой предпочтительно кассету экспрессии, имеющую последовательность согласно SEQ ID NO: 13.
Дополнительным объектом настоящего изобретения является вектор, который содержит кассету экспрессии согласно настоящему изобретению.
Kirchner and Tauch (Journal of Biotechnology 104:287-299 (2003)) описывают выбор векторов, которые предпочтительно используют в Corynebacterium glutamicum.
Гомологичная рекомбинация с использованием векторов согласно настоящему изобретению позволяет заменить участки ДНК в хромосоме на кассеты экспрессии согласно настоящему изобретению, которые транспортируют в клетку с помощью вектора. Для эффективной рекомбинации между кольцевой молекулой ДНК вектора и ДНК-мишенью в хромосоме область ДНК, подлежащую замене, которая содержит кассету экспрессии согласно настоящему изобретению, обеспечивают с нуклеотидными последовательностями на концах, гомологичными целевому сайту, в результате чего точно определяют сайт интеграции вектора или замены ДНК.
Включение кассеты экспрессии согласно настоящему изобретению может иметь место при этом в сайте нативного гена в генах ilvBN, предпочтительно, нативных генов ilvBN и необязательно также нативного промотора генов ilvBN, заменяемого при этом кассетой экспрессии согласно настоящему изобретению.
Альтернативно, кассету экспрессии согласно настоящему изобретению можно также интегрировать в межгенную область в хромосоме, которая не имеет кодирующей функции, или в сайт другого гена, при этом сайт другого гена предпочтительно представляет собой нуклеотидную последовательность в хромосоме, которая не является необходимой для роста клеток и получения аминокислоты или кетокислоты.
Кассету экспрессии согласно настоящему изобретению можно также включить согласно настоящему изобретению в хромосому в предпочтительном варианте осуществления в нескольких копиях, а также необязательно в сайтах различных генов.
Вместо включения в хромосому кассеты экспрессии согласно настоящему изобретению, альтернативно можно также включить в хромосому оператор, используемый согласно настоящему изобретению, в комбинации с промотором по сайту нативного гена в генах ilvBN, где предпочтительно нативный промотор генов ilvBN заменяют конструкцией оператора и промотора.
Вместо включения кассеты экспрессии согласно настоящему изобретению в хромосому, согласно настоящему изобретению альтернативно также можно конечно использовать внехромосомно реплицирующийся вектор, который содержит кассету экспрессии согласно настоящему изобретению.
Настоящее изобретение также дополнительно относится к микроорганизму, предпочтительно к Corynebacterium, особенно Corynebacterium, которая производит L-аминокислоту, выбранную из L-лейцина, L-валина и L-изолейцина, или α-кетокислоту, выбранную из кетоизовалерата, кетометилвалерата и кетоизокапроата, которая содержит кассету экспрессии согласно настоящему изобретению и/или вектор согласно настоящему изобретению.
Детали биохимии и химической структуры полинуклеотидов, таких какие встречаются в живых организмах, таких как, например, микроорганизмы, можно найти, помимо прочего, в учебнике “Biochemie” [Biochemistry] Berg et al. (Spektrum Akademischer Verlag Heidelberg Berlin, 2003; ISBN 3-8274-1303-6).
Если полинуклеотид состоит из дезоксирибонуклеотидных мономеров, содержащих нуклеиновые основания или основания аденин (A), гуанин(G), цитозин(C) и тимин (T), говорят о дезоксирибополинуклеотидах или дезоксирибонуклеиновой кислоте (ДНК). Если полинуклеотид состоит из рибонуклеотидных мономеров, содержащих нуклеиновые основания или основания аденин (A), гуанин(G), цитозин(C) и урацил (U), говорят о рибополинуклеотидах или рибонуклеиновой кислоте (РНК). В упомянутых полинуклеотидах мономеры соединены ковалентно друг с другом при помощи сложной 3'→ 5'-фосфодиэфирной связи.
Выражение "полинуклеотид с операторной активностью" или "оператор" следует понимать как полинуклеотид, предпочтительно дезоксирибополинуклеотид, или нуклеиновую кислоту, предпочтительно дезоксирибонуклеиновую кислоту (ДНК), которая функционально связана с помощью полинуклеотида с промоторной активностью с полинуклеотидом, подлежащим транскрибированию, включает или выключает транскрипцию этого полинуклеотида посредством взаимодействия с различными регуляторными белками (активаторами или репрессорами, которые в свою очередь взаимодействуют с лигандами или эффекторными молекулами).
Выражение "полинуклеотид с промоторной активностью" или "промотор" следует понимать как полинуклеотид, предпочтительно дезоксирибополинуклеотид, или нуклеиновую кислоту, предпочтительно дезоксирибонуклеиновую кислоту (ДНК), которая функционально связана с полинуклеотидом, подлежащим транскрибированию, определяющий точку инициации и частоту инициации транскрипции этого полинуклеотида, посредством чего можно влиять на уровень экспрессии контролируемого полинуклеотида.
Вследствие двухцепочечной структуры ДНК настоящее изобретение также относится к комплементарной цепи цепи последовательности, перечисленной в SEQ ID NO: 1, 2 или 3.
Выражение "транскрипция" следует понимать как способ, посредством которого, начиная с матрицы ДНК, получают комплементарную молекулу РНК. Белки, такие как РНК-полимеразы, "сигма-факторы" и белки-регуляторы транскрипции участвуют в этом способе. Затем синтезированная РНК (информационная РНК, м-РНК) служит в качестве матрицы в процессе трансляции, который затем ведет к полипептиду или белку.
Ген, рассматриваемый с химической точки зрения, является полинуклеотидом. Полинуклеотид, который кодирует белок/полипептид, используют при этом синонимично выражению "ген". Соответственно, эти два выражения "ген" и "кодирующая область" используют синонимично, как и два выражения "белок" и "полипептид".
Выражение "ниже расположенное функциональное соединение или связь" следует понимать в этой связи как означающее последовательное расположение полинуклеотида, характеризующегося операторной активностью согласно настоящему изобретению, со вторым полинуклеотидом, имеющим промоторную активность, и с дополнительным олиго- или полинуклеотидом, которое ведет к транскрипции этого дополнительного полинуклеотида.
Если дополнительный полинуклеотид представляет собой полинуклеотид, который кодирует полипептид/белок, состоящий из кодирующей области для полипептида, начиная с инициирующего кодона, в том числе стоп-кодон и необязательно в том числе терминатор транскрипции, выражение "ниже расположенное функциональное соединение или связь" означает последовательное расположение, которое ведет к транскрипции дополнительного полинуклеотида и трансляции синтезированной РНК.
Дополнительый полинуклеотид кодирует один или более полипептид(полипептиды). Полинуклеотид, кодирующий белок/полипептид, в целом состоит из инициирующего кодона, выбранного из группы, состоящей из ATG, GTG и TTG, предпочтительно ATG или GTG, особенно предпочтительно ATG, из кодирующей белок последовательности и одного или более стоп-кодона(кодонов), выбранного из группы, состоящей из TAA, TAG и TGA.
Если дополнительный полинуклеотид кодирует ряд белков/полипептидов, сайт связывания рибосомы может быть расположен перед каждым геном. После последнего гена необязательно расположен терминатор.
Дополнительный полинуклеотид состоит согласно настоящему изобретению из генов ilvB и ilvN, которые кодируют субъединицы ацетолактатсинтазы (IlvBN, EC No.: 4.1.3.18).
Настоящее изобретение, кроме того, относится к применению кассеты экспрессии согласно настоящему изобретению или вектора согласно настоящему изобретению для экспрессии генов ilvBN в микроорганизмах. Кассета экспрессии согласно настоящему изобретению гарантирует транскрипцию и трансляцию синтезированной РНК, предпочтительно мРНК, в полипептиды, а именно, в две субъединицы ацетолактатсинтазы.
С использованием кассеты экспрессии согласно настоящему изобретению гены ilvBN в микроорганизмах можно экспрессировать или сверхэкспрессировать в желаемый момент времени.
Сверхэкспрессию обычно следует понимать как увеличение внутриклеточной концентрации или активности рибонуклеиновой кислоты, белка (полипептида) или фермента по сравнению с исходным штаммом (родительским штаммом) или штаммом дикого типа, если он является исходным штаммом. Исходный штамм (родительский штамм) следует понимать как штамм, на который производят воздействие, ведущее к сверхэкспрессии.
В случае сверхэкспрессии предпочтительными являются способы рекомбинантной сверхэкспрессии. Среди них сгруппированы все способы, в которых микроорганизм получают с использованием молекулы ДНК, полученной in vitro. Такие молекулы ДНК включают в себя, например, промоторы, кассеты экспрессии, гены, аллели, кодирующие области и т.д. Их превращают способами трансформации, конъюгации, трансдукции или аналогичными способами в желаемый микроорганизм.
С помощью воздействия сверхэкспрессии с использованием оператора, подлежащего использованию согласно настоящему изобретению, и/или с использованием кассеты экспрессии согласно настоящему изобретению активность или концентрацию ацетолактатсинтазы в целом предпочтительно повышают по меньшей мере на 10%, 25%, 50%, 75%, 100%, 150%, 200%, 300%, 400% или 500%, предпочтительно, максимально до 1000%, 2000%, 4000%, 10000% или 20000% на основании активности или концентрации полипептида в штамме до воздействия, ведущего к сверхэкспрессии.
Степень экспрессии или сверхэкспрессии можно определить путем измерения количества мРНК, транскрибированной с гена, путем определения количества полипептида и путем определения активности фермента.
Для определения количества мРНК, помимо прочего, используют способ "нозерн-блоттинга" и количественную RT-PCR (ПЦР с обратной транскрипцией). При количественной RT-PCR полимеразной цепной реакции предшествует обратная транскрипция. Для этой цели можно использовать LightCyclerTM System от компании Roche Diagnostics (Boehringer Mannheim GmbH, Roche Molecular Biochemicals, Маннхейм, Германия), как описано, например, в Jungwirth et al. (FEMS Microbiology Letters 281, 190-197 (2008)). Концентрацию белка в геле можно определить с помощью 1- и 2-мерного гель-разделения белков и последующей оптической идентификации концентрации белка с использованием соответствующего аналитического программного обеспечения. Общепринятым способом для получения белковых гелей в случае коринеформных бактерий и для идентификации белков является процедура, описанная Hermann et al. (Electrophoresis, 22:1712-23 (2001)). Концентрацию белка также можно определить с помощью вестерн-блот-гибридизации с использованием антитела, специфичного для белка, подлежащего обнаружению (Sambrook et al., Molecular cloning: a laboratory manual. 2nd Ed. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., 1989) и последующей оптической оценки с использованием соответствующего программного обеспечения для определения концентрации (Lohaus and Meyer (1998) Biospektrum 5:32-39; Lottspeich, Angewandte Chemie 321: 2630-2647 (1999)). Статистическую значимость собранных данных определяют с использованием Т-теста (Gosset, Biometrika 6(1): 1-25 (1908)).
Воздействие для сверхэкспрессии генов ilvBN с использованием оператора, подлежащего использованию согласно настоящему изобретению, можно комбинировать подходящим образом с дополнительными воздействиями для сверхэкспрессии.
В уровне техники доступно множество способов для достижения сверхэкспрессии. В дополнение к модификации нуклеотидных последовательностей, которые регулируют или контролируют экспрессию гена, они также включают увеличение числа копий.
Увеличение числа копий может иметь место посредством плазмид, которые реплицируются в цитоплазме микроорганизма. Для этой цели в уровне техники описано множество плазмид для различных групп микроорганизмов, с помощью которых можно регулировать желаемое увеличение числа копий гена. Подходящие плазмиды для рода Corynebacterium описаны, например, в Tauch et al. (Journal of Biotechnology 104 (1-3), 27-40, (2003)) или в Stansen et al. (Applied and Environmental Microbiology 71, 5920-5928 (2005)).
Увеличение числа копий на по меньшей мере одну (1) копию может, кроме того, иметь место посредством вставки дополнительных копий в хромосому микроорганизма. Подходящие способы для рода Corynebacterium описаны, например, в описаниях изобретения к патентам WO 03/014330, WO 03/040373 и WO 04/069996.
Увеличение экспрессии гена может, кроме того, иметь место тогда, когда ряд промоторов располагают перед желаемым геном или функционально связывают с геном, подлежащим экспрессии, и таким образом достигают повышенной экспрессии. Примеры этого описаны в описании изобретения к патенту WO 2006/069711.
Степень продления зависит от использования кодона; при применении кодонов для т(транспортных) РНК, часто встречающихся в исходных штаммах, трансляцию можно увеличить. Кроме того, замена инициирующего кодона на кодон ATG, наиболее часто возникающая во многих микроорганизмах (77% в Escherichia coli), может значительно улучшить трансляцию, поскольку на уровне РНК кодон AUG является от двух до трех раз более эффективным, чем, например, кодоны GUG и UUG (Khudyakov et al., FEBS Letters 232(2):369-71 (1988); Reddy et al., Proceedings of the National Academy of Sciences of the USA 82(17):5656-60 (1985)). Окружение последовательности инициирующего кодона также можно оптимизировать, поскольку описаны эффекты взаимодействия между инициирующим кодоном и фланкирующими областями (Stenstrom et al., Gene 273(2):259-65 (2001); Hui et al., EMBO Journal 3(3):623-9 (1984)).
Инструкции для обработки ДНК, расщепления и лигирования ДНК, трансформации и селекции трансформантов находятся, помимо прочего, в известном Справочнике Sambrook et al. Molecular Cloning: A Laboratory Manual, Second Edition (Cold Spring Harbor Laboratory Press, 1989).
В предпочтительном варианте осуществления согласно настоящему изобретению используют микроорганизмы, в которых дополнительные гены биосинтетического пути желаемой L-аминокислоты или α-кетокислоты дополнительно присутствуют в амплифицированной форме, в частности, сверхэкспрессированы.
В связи с получением L-валина, L-изолейцина, α-кетоизовалериановой кислоты, α-кето-β-метилвалериановой кислоты или α-кетоизокапроновой кислоты, предпочтительно один или более генов или полинуклеотидов, которые кодируют ферменты биосинтеза L-валина, L-изолейцина, α-кетоизовалериановой кислоты, α-кето-β-метилвалериановой кислоты или α-кетоизокапроновой кислоты, выбранные из группы:
a) полинуклеотид (ген ilvC), который кодирует изомероредуктазу (IlvC, EC No.: 1.1.1.86),
b) полинуклеотид (ген ilvD), который кодирует дигидроксиациддегидратазу (IlvD, EC No.: 4.2.1.9),
c) полинуклеотид (ген ilvE), который кодирует трансаминазу (IlvE, EC No.: 2.6.1.42),
d) полинуклеотид (ген ilvA), который кодирует треониндегидратазу (IlvA, EC No.: 4.3.1.19),
e) полинуклеотид (ген hom), который кодирует гомосериндегидрогеназу (Hom, EC No.: 1.2.1.11),
f) полинуклеотид (ген thrB), который кодирует гомосеринкиназу (ThrB, EC No.: 2.7.1.39),
g) полинуклеотид (ген thrC), который кодирует треонинсинтазу (ThrC, EC No.: 4.2.3.1),
h) полинуклеотид (ген leuA), который кодирует изопропилмалатсинтазу (LeuA, EC No.: 2.3.3.13),
i) полинуклеотид (ген leuB), который кодирует изопропилмалатдегидрогеназу (LeuB, EC No.: 1.1.1.85),
j) полинуклеотид (ген leuC), который кодирует большую субъединицу изопропилмалатизомеразы (LeuC, EC No.: 4.2.1.33),
k) полинуклеотид (ген leuD), который кодирует малую субъединицу изопропилмалатизомеразы (LeuD, EC No.: 4.2.1.33),
можно дополнительно сверхэкспрессировать, при этом гены hom, ilvA, ilvC, ilvD и ilvE являются особенно предпочтительными для изолейцина и валина, гены ilvC и ilvD являются особенно предпочтительными для α-кетоизовалериановой кислоты (KIV) и α-кето-β-метилвалериановой кислоты (KMV) и гены ilvC, ilvD, leuA, leuB, leuC и leuD являются особенно предпочтительными для α-кетоизокапроновой кислоты (KIC).
В предпочтительном варианте осуществления используют микроорганизмы, в которых метаболические пути, которые снижают образование желаемой L-аминокислоты или α-кетокислоты, по меньшей мере частично ослаблены.
Если предпочтительным является получение валина или кетоизовалерата, тогда метаболические пути для изолейцина (для образования предшественника - альфа-кетомасляной кислоты: ilvA, thrB, thrC, hom) и/или для лейцина (leuA, leuB, leuCD) могут быть ослаблены. Если предпочтительным является получение изолейцина или кетометилвалерата, то биосинтетический путь лейцина может быть ослаблен. Если предпочтительным является производство лейцина или кетоизокапроата, тогда биосинтетический путь изолейцина (или предшественника - альфа-кетомасляной кислоты) может быть ослаблен.
Выражение "ослабление" в этой связи описывает снижение или выключение внутриклеточной активности одного или нескольких ферментов (белков) в бактерии, которые кодируются соответствующей ДНК, посредством, например, использования слабого промотора или с использованием гена или аллеля, который кодирует соответствующий фермент, имеющий низкую активность, или инактивирует соответствующий ген или фермент (белок) и необязательно сочетает в себе эти воздействия. Полного или частичного ослабления индивидуальных генов-мишеней можно достичь, например, посредством полной или частичной делеции генов или посредством вставки точечных мутаций в структурный ген или в промоторный участок или в сайт связывания рибосомы. Дополнительным способом для специфического снижения экспрессии гена является антисмысловая техника, когда короткие олигодезоксирибонуклеотиды или векторы для синтеза более длинной антисмысловой РНК вводят в клетки-мишени. Антисмысловая РНК может связываться там с комплементарными участками специфических мРНК и снижать их стабильность или блокировать способность к трансляции. Специалист в данной области найдет пример этого в Srivastava et al. (Applied Environmental Microbiology 2000 Oct; 66 (10): 4366 – 4371). Только что описанную антисмысловую технику можно также выполнять путем применения операторов/промотора согласно настоящему изобретению, в котором их клонируют в "антисмысловой" ориентации после гена-мишени. После добавления индуктора - пропионата индуцируют образование мРНК комплементарной цепи гена-мишени, подлежащего ослаблению. Путем добавления этой антисмысловой мРНК к мРНК гена-мишени уменьшают экспрессию гена-мишени. Регулируемую экспрессию или сверхэкспрессию генов ilvBN или получение L-аминокислот или α-кетокислот предпочтительно проводят в микроорганизмах из рода Corynebacterium. В роде Corynebacterium предпочтительными штаммами являются те, которые основаны на следующих видах: Corynebacterium efficiens, типовой штамм, размещенный как DSM44549, Corynebacterium glutamicum, типовой штамм, размещенный как ATCC13032, и Corynebacterium ammoniagenes, типовой штамм, размещенный как ATCC6871. Вид Corynebacterium glutamicum является наиболее предпочтительным.
Некоторые представители вида Corynebacterium glutamicum также известны из уровня техники под другими названиями. Они включают в себя, например: штамм ATCC13870, который обозначили как Corynebacterium acetoacidophilum, штамм DSM20137, который обозначили как Corynebacterium lilium, штамм ATCC17965, который обозначили как Corynebacterium melassecola, штамм ATCC14067, который обозначили как Brevibacterium flavum, штамм ATCC13869, который обозначили как Brevibacterium lactofermentum, и штамм ATCC14020, который обозначили как Brevibacterium divaricatum.
Также для Corynebacterium glutamicum было распространено выражение "Micrococcus glutamicus". Некоторые представители вида Corynebacterium efficiens были также обозначены в уровне техники как Corynebacterium thermoaminogenes, такие, например, как штамм FERM BP-1539.
Микроорганизмы или штаммы (исходные штаммы), используемые для воздействий согласно настоящему изобретению, предпочтительно уже имеют способность секретировать желаемую L-аминокислоту или α-кетокислоту в питательную среду, окружающую их, и накапливать ее там. В дальнейшем для этого также используется выражение "производить". В частности, штаммы, используемые согласно настоящему изобретению, предпочтительно имеют способность после индукции увеличивать содержание или накапливать ≥ (по меньшей мере) 0,5 г/л*ч, предпочтительно, по меньшей мере 1,0 или 2,0 г/л*ч, желаемой аминокислоты или кетокислоты в клетке или в питательной среде. Исходные штаммы предпочтительно представляют собой штаммы, которые были получены путем мутагенеза и селекции, с помощью методов рекомбинантной ДНК или путем комбинации обоих способов.
Специалисту в данной области понятно, что также можно получить микроорганизм, пригодный для воздействий согласно настоящему изобретению, путем первоначального использования в диком штамме, таком как, например, типовой штамм Corynebacterium glutamicum ATCC 13032 или штамм АТСС 14067, оператора, подлежащего использованию согласно настоящему изобретению, для регулируемой экспрессии желаемых генов, а затем путем индукции микроорганизма для получения аминокислоты или кетокислоты путем дополнительных генетических воздействий, описанных в уровне техники.
Известными представителями штаммов коринеформных бактерий, производящих или секретирующих L-валин, являются, например: Brevibacterium lactofermentum FERM BP-1763 (описан в US 5188948); Brevibacterium lactofermentum FERM BP-3007 (описан в US 5521074); Corynebacterium glutamicum FERM BP-3006 (описан в US 5521074) и Corynebacterium glutamicum FERM BP-1764 (описан в US 5188948).
Микроорганизмы, производящие L-валин, как правило, имеют устойчивую к торможению по механизму обратной связи или десенсибилизированную ацетолактатсинтазу (AHAS, EC 4.1.3.18). Она представляет собой первый фермент параллельных метаболических путей для синтеза изолейцина, валина и лейцина (Umbarger, H. E. 1987, Biosynthesis of the branched-chain amino acids, pp. 352-367, в F. C. Neidhardt, J. L. Ingraham, K. B. Low, B. Magasanik, M. Schaechter and H.E. Umbarger (ed.), Escherichia coli and Salmonella typhimurium: Cellular and Molecular Biology, American Society for Microbiology, Washington, D.C.). Холофермент всегда состоит из 2 больших субъединиц и 2 малых субъединиц. Большая субъединица AHAS образует каталитический домен и кодируется ilvB; малая субъединица, которая функционирует как регуляторный домен, кодируется ilvN. Под устойчивыми к торможению по механизму обратной связи ацетолактатсинтазами подразумевают ацетолактатсинтазы, которые по сравнению с дикой формой (диким типом) имеют более низкую чувствительность к ингибированию аминокислотами с разветвленной цепью валином, изолейцином и лейцином или их смесями. В случае ацетолактатсинтаз из видов Corynebacterium glutamicum, штаммы ATCC13032, ATCC14067 (также известного как Brevibacterium flavum) или ATCC13869 (также известного как Brevibacterium lactofermentum) представляют собой подходящий дикий тип.
Гены ilvBN в Corynebacterium glutamicum, кодирующие ацетолактатсинтазу, описаны, например, Keilhauer et al. (Journal of Bacteriology 175(17):5595-603 (1993)) или в EP1108790. По номеру доступа L09232 (GenBank, NCBI) показана последовательность данных генов.
Варианты фермента AHAS, которые больше не подвержены ингибированию по механизму обратной связи аминокислотами с разветвленной цепью (лейцин, валин, изолейцин), описаны, например, в Mendel et al. (Journal of Molecular Biology 325, 275-284 (2003)), Elisakova et al. (Applied and Environmental Microbiology 71, 207-213 (2005)), Wada et al. (Bioscience Biotechnology & Biochemistry, 72 (11), 2959-2965, (2008)) и в EP 1491634. Предпочтительными являются такие варианты устойчивой к торможению по механизму обратной связи ацетолактатсинтазы, которые несут одну или более из следующих аминокислотных замен в малой субъединице, кодируемой ilvN, выбранных из группы: в положении 20 аминокислотной последовательности L-аспарагиновая кислота вместо глицина, в положении 21 аминокислотной последовательности L-аспарагиновая кислота вместо L-изолейцина, в положении 22 аминокислотной последовательности L-фенилаланин вместо L-изолейцина, в положении 42 любая протеиногенная аминокислота за исключением L-аланина, предпочтительно L-валин, L-изолейцин и L-лейцин, особенно предпочтительно L-валин, и необязательно в положении 47 L-лейцин вместо L-гистидина (описано в DE 102011118019 А1).
Известными представителями L-изолейцин-производящих или секретирующих штаммов коринеформных бактерий являются, например, Brevibacterium flavum FERM BP-760 (описан в US 4656135); Brevibacterium flavum FERM BP-2215 (описан в US 5294547) и Corynebacterium glutamicum FERM BP-758 (описан в US 4656135).
Секретирующие или производящие α-кетокислоту штаммы основаны, например, на: Corynebacterium glutamicum, штамм ATCC13032; Brevibacterium flavum, штамм ATCC 14067 и Brevibacterium lactofermentum, штамм ATCC 13869.
Настоящее изобретение обеспечивает микроорганизм, который производит L-аминокислоту, выбранную из L-лейцина, L-валина и L-изолейцина, или α-кетокислоту, выбранную из α-кетоизовалерата, α-кетометилвалерата и α-кетоизокапроата, при этом микроорганизм делает возможным или имеет при применении оператора, подлежащего использованию согласно настоящему изобретению, регулируемую экспрессию генов ilvBN, кодирующих ацетолактатсинтазу.
Кроме того, настоящее изобретение делает доступным способ ферментативного получения L-аминокислоты, выбранной из L-лейцина, L-валина и L-изолейцина, или α-кетокислоты, выбранной из α-кетоизовалерата, α-кетометилвалерата и α-кетоизокапроата, включающий следующие этапы:
a) культивирование микроорганизма согласно настоящему изобретению в подходящей среде с получением ферментационного бульона и
b) увеличение содержания L-аминокислоты или α-кетокислоты в данном ферментационном бульоне a) и/или в клетках данного микроорганизма.
При этом предпочтительно, что L-аминокислоту или α-кетокислоту или жидкий или твердый продукт, который содержит L-аминокислоту или α-кетокислоту, получают из ферментационного бульона, содержащего L-аминокислоту или α-кетокислоту.
Полученные микроорганизмы можно культивировать непрерывно - как описано, например, в WO 05/021772 - или по партиям в периодическом способе (периодическое культивирование или периодический способ) или в периодическом с подпиткой (способ с подкормкой) или периодическом способе с повторной подпиткой (способ с повторяющейся подкормкой) с целью получения желаемого органического химического соединения. Общий обзор известных способов культивирования доступен в учебнике Chmiel (Bioprozesstechnik 1,  in die Bioverfahrenstechnik [Bioprocess Technology 1, Introduction to Bioprocess Technology] (Gustav Fischer Verlag, Штуттгарт, 1991)) или в учебнике Storhas (Bioreaktoren and periphere Einrichtungen [Bioreactors and Peripheral Devices] (Vieweg Verlag, Брауншвейг/Висбаден, 1994)).
in die Bioverfahrenstechnik [Bioprocess Technology 1, Introduction to Bioprocess Technology] (Gustav Fischer Verlag, Штуттгарт, 1991)) или в учебнике Storhas (Bioreaktoren and periphere Einrichtungen [Bioreactors and Peripheral Devices] (Vieweg Verlag, Брауншвейг/Висбаден, 1994)).
Культуральная среда или ферментационная среда, которая будет использоваться, должна удовлетворять требованиям соответствующих штаммов подходящим образом. Описания культуральных сред для различных микроорганизмов содержатся в справочнике “Manual of Methods for General Bacteriology” от Американского общества по бактериологии (Вашингтон, округ Колумбия, США, 1981). Выражения культуральная среда или ферментационная среда являются взаимозаменяемыми.
Используемым источником углерода могут быть сахара и углеводы, такие как, например, глюкоза, сахароза, лактоза, фруктоза, мальтоза, меласса, содержащие сахарозу растворы от переработки сахарной свеклы или сахарного тростника, крахмал, гидролизат крахмала и целлюлоза, масла и жиры, такие как, например, соевое масло, подсолнечное масло, арахисовое масло и кокосовое масло, жирные кислоты, такие как, например, пальмитиновая кислота, стеариновая кислота и линолевая кислота, спирты, такие как, например, глицерин, метанол и этанол, и органические кислоты, такие как, например, уксусная кислота или молочная кислота.
Используемым источником азота могут быть органические азот-содержащие соединения, такие как пептоны, дрожжевой экстракт, мясной экстракт, солодовый экстракт, жидкий кукурузный экстракт, соевая мука и мочевина, или неорганические соединения, такие как сульфат аммония, хлорид аммония, фосфат аммония, карбонат аммония и нитрат аммония. Источники азота можно использовать по отдельности или в виде смеси.
Используемым источником фосфора может быть фосфорная кислота, фосфат аммония, дигидрофосфат калия или гидроортофосфат калия или соответствующие натрий-содержащие соли.
Культуральная среда должна, кроме того, содержать соли, например, в виде хлоридов или сульфатов металлов, таких как, например, натрий, калий, магний, кальций и железо, такие как, например, сульфат магния или сульфат железа, которые необходимы для роста. Наконец, незаменимые ростовые вещества, такие как аминокислоты, например гомосерин, и витамины, например тиамин, биотин или пантотеновую кислоту, можно использовать дополнительно к вышеупомянутым веществам.
Пропионат предпочтительно добавляют в среду в виде соли, но можно также добавить в виде пропионовой кислоты. Подходящими солями пропионовой кислоты являются пропионат магния, пропионат натрия, пропионат кальция, пропионат аммония и пропионат калия. Пропионат присутствует в среде растворенным в виде свободной кислоты или в виде аниона пропионата.
Упомянутое сырье можно добавить к культуре в виде одной партии или подходящим образом во время культивирования.
Для контроля рН культуры подходящими для использования являются основные соединения, такие как гидроксид натрия, гидроксид калия, аммиак, гидроксид аммония или аммиачную воду, или кислотные соединения, такие как фосфорная кислота или серная кислота. pH обычно доводят до значения от 6,0 до 8,5, предпочтительно от 6,5 до 8. Для контроля образования пены можно использовать противовспенивающие средства, такие как, например, полигликолевые эфиры жирных кислот. Для поддержания стабильности плазмид можно добавить в среду пригодные избирательно действующие вещества, такие как, например, антибиотики. Ферментацию предпочтительно проводят в аэробных условиях. Для поддержания этого в культуру добавляют кислород или кислород-содержащие смеси газов, такие как, например, воздух. Также возможно применение жидкостей, которые обогащены перекисью водорода. Ферментация в условиях ограниченного содержания кислорода представляет собой дополнительный специфический вариант осуществления согласно настоящему изобретению. Необязательно ферментацию проводят при избыточном давлении, например, при избыточном давлении от 0,03 до 0,2 МПа. Температура культуры обычно составляет от 20°C до 45°C и предпочтительно от 25°C до 40°C, особенно предпочтительно, от 30° до 37°C. В периодическом или периодическом с подпиткой способах культивирование предпочтительно продолжается до тех пор, пока не получено количество, достаточное для измерения извлечения желаемого органического химического соединения. Эта цель обычно достигается в пределах от 10 часов до 160 часов. При непрерывных способах возможны более длительные сроки культивирования. Благодаря активности микроорганизмов происходит увеличение содержания (накопление) органического химического соединения в ферментационной среде и/или в клетках микроорганизмов.
Примеры подходящих ферментационных сред находятся, помимо прочего, в патентных описаниях US 5770409, US 5990350, US 5275940, WO 2007/012078, US 5827698, WO 2009/043803, US 5756345, US 7138266.
Анализ L-аминокислот для определения концентрации в один или более момент(моменты) времени в ходе ферментации можно выполнить путем разделения L-аминокислот с помощью ионообменной хроматографии, предпочтительно катионообменной хроматографии, с последующей пост-колоночным получением производных с использованием нингидрина, как описано в Spackman et al. (Analytical Chemistry 30: 1190-1206 (1958)). Вместо нингидрина для пост-колоночного получения производных также можно использовать ортофталевый диальдегид. Обзорная статья по ионообменной хроматографии находится в Pickering (LC⋅GC (Magazine of Chromatographic Science) 7(6), 484-487 (1989)).
Также можно выполнить пред-колоночное получение производных, например, с помощью ортофталевого диальдегида или фенилизотиоцианата, и отделить полученные аминокислотные производные с помощью обращенно-фазовой хроматографии (RPC), предпочтительно в форме высокоэффективной жидкостной хроматографии (HPLC). Такой способ описан, например, в Lindroth et al. (Analytical Chemistry 51: 1167-1174 (1979)).
Детекцию выполняют фотометрически (поглощение, флуоресценция).
Всеобъемлющее представление об аминокислотном анализе можно получить, помимо прочего, из учебника “Bioanalytik” от Lottspeich and Zorbas (Spektrum Akademischer Verlag, Heidelberg, Germany 1998).
Анализ α-кетокислот для определения концентрации в один или более момент(моменты) времени в ходе ферментации можно выполнить путем разделения кетокислот и других продуктов экскреции с помощью ионообменной хроматографии, предпочтительно катионообменной хроматографии, на сульфонированном стирол/дивинилбензольном полимере в H+-форме, например, с использованием 0,025 н серной кислоты, с последующей УФ-детекцией при 215 нм (альтернативно, также при 230 или 275 нм). Предпочтительно, можно использовать колонку REZEX RFQ - Fast Fruit H+ (Phenomenex); возможны другие поставщики для разделительной фазы (например, Aminex от BioRad). Аналогичные разделения описаны в соответствующих примерах применения от поставщика.
Анализ пропионовой кислоты для определения концентрации в один или более момент(моменты) времени в ходе ферментации можно осуществить путем разделения органических кислот с помощью HPLC. В качестве колонки использовали VARIAN MetaCarb H+ 300 x 7,8 мм A5215 (длина 300 мм, диаметр 7,8 мм). Смесь серной кислоты и ацетонитрила (215 мл 0,5 М серной кислоты, 50 мл ацетонитрила, 5 л дистиллированной воды) служила в качестве элюента. 0,005 М серная кислота служила в качестве растворителя для подвижных образцов. При этом бесклеточные подвижные образцы развели 1:20.
Параметры разделения были следующими: скорость потока 0,4 мл/мин; объем инъекции образца 20 мкл; температура 35°C. Детекция происходит фотометрически при 215 нм с помощью УФ. Время удержания пропионовой кислоты составило 30,258 мин. Измеряемый диапазон находился между 0,09 и 7,514 г/л.
Продуктивность способов или ферментационных способов согласно настоящему изобретению по отношению к одному или нескольким параметрам, выбранным из группы, состоящей из концентрации (соединение, образованное на единицу объема), выхода (соединение, образованное на единицу потребляемого источника углерода), образования (соединение, образованное на единицу объема и времени), специфического образования (соединение, образованное на единицу сухой массы клеток или единицу биологической сухой массы и времени, или соединение, образованное на единицу белка клеток и времени), или также другим параметрам способа и их комбинациям, предпочтительно повышена согласно настоящему изобретению по меньшей мере на 0,5%, по меньшей мере на 1%, по меньшей мере на 1,5% или по меньшей мере на 2% по сравнению со способами или ферментационными способами с использованием микроорганизмов, в которых не присутствует кассета экспрессии согласно настоящему изобретению. Это следует рассматривать как очень значимое в контексте крупномасштабного способа.
С помощью данных воздействий на ферментацию получают ферментационный бульон, который содержит желаемую аминокислоту или кетокислоту.
Впоследствии происходит запасание или получение или извлечение продукта, содержащего аминокислоту или кетокислоту, в жидком или твердом виде.
Ферментационный бульон следует понимать как ферментационную среду или питательную среду, в которой микроорганизм культивировали в течение определенного времени и при определенной температуре. Ферментационная среда или среды, используемые в ходе ферментации, содержат/содержит все вещества или компоненты, которые обеспечивают получение желаемого соединения и, как правило, репликацию и жизнеспособность.
По завершении ферментации полученный ферментационный бульон, соответственно, содержит
a) биомассу (клеточную массу) микроорганизма, образованную в результате размножения клеток микроорганизма,
b) желаемую аминокислоту или кетокислоту, образованную в ходе ферментации,
c) органические побочные продукты, необязательно образованные в ходе ферментации, и
d) компоненты использованной ферментационной среды или сырья, такие как, например, витамины, такие как биотин, или соли, такие как сульфат магния, не поглощенные при ферментации.
Органические побочные продукты включают вещества, которые произведены использованными микроорганизмами при ферментации в дополнение к соответствующему желаемому соединению и необязательно выделены.
Ферментационный бульон удаляют из культурального сосуда или ферментационного контейнера, необязательно собирают и используют для подготовки продукта, содержащего аминокислоту или кетокислоту в жидком или твердом виде. Для этого также используют выражение «получение содержащего чистое химическое вещество продукта». В простейшем случае содержащий чистое химическое вещество ферментационный бульон, удаленный из ферментационного контейнера, сам по себе является полученным продуктом.
С помощью одного или нескольких воздействий, выбранных из группы, состоящей из
a) частичного (от > 0% до < 80%) до полного (100%) или практически полного (≥ 80%, ≥ 90%, ≥ 95%, ≥ 96%, ≥ 97%, ≥ 98%, ≥ 99%) удаления воды,
b) частичного (от > 0% до < 80%) до полного (100%) или практически полного (≥ 80%, ≥ 90%, ≥ 95%, ≥ 96%, ≥ 97%, ≥ 98%, ≥ 99%) удаления биомассы, которую необязательно инактивируют перед удалением,
c) частичного (от > 0% до < 80%) до полного (100%) или практически полного (≥ 80%, ≥ 90%, ≥ 95%, ≥ 96%, ≥ 97%, ≥ 98%, ≥ 99%, ≥ 99,3%, ≥ 99,7%) удаления органических побочных продуктов, образованных в ходе ферментации, и
d) частичного (> 0%) до полного (100%) или практически полного (≥ 80%, ≥ 90%, ≥ 95%, ≥ 96%, ≥ 97%, ≥ 98%, ≥ 99%, ≥ 99,3%, ≥ 99,7%) удаления компонентов использованной ферментационной среды или сырья - тех компонентов, которые не были поглощены при ферментации,
проводят концентрирование или очистку желаемого органического химического соединения от ферментационного бульона. Таким образом выделяют продукты, которые имеют желаемое содержание соединения.
Частичное (от > 0% до < 80%) до полного (100%) или практически полное(≥ от 80% до < 100%) удаление воды (воздействие a) также обозначено как высушивание.
В одном варианте настоящего способа чистые (≥ 80% по весу, ≥ 90% по весу) или высокочистые (≥ 95% по весу, ≥ 97% по весу, ≥ 99% по весу) формы продукта желаемого органического химического соединения, предпочтительно L-аминокислоты, получают посредством полного или практически полного удаления воды, биомассы, органических побочных продуктов и непоглощенных компонентов использованной ферментационной среды. Для воздействий согласно a), b), c) или d) в уровне техники доступно множество технических инструкций.
В случае способов получения L-аминокислот L-лейцина, L-валина или L-изолейцина или α-кетокислот кетоизовалерата, кетометилвалерата или кетоизокапроата с использованием бактерий из рода Corynebacterium предпочтительными являются такие способы, в которых получают продукты, не содержащие компоненты ферментационного бульона. Их используют, в частности, в медицине человека, в фармацевтической промышленности и в пищевой промышленности.
Способ согласно настоящему изобретению служит для ферментативного получения L-аминокислот L-лейцина, L-валина или L-изолейцина или α-кетокислот кетоизовалерата, кетометилвалерата или кетоизокапроата.
Настоящее изобретение в конечном итоге относится к применению микроорганизма согласно настоящему изобретению для ферментативного получения L-аминокислот L-лейцина, L-валина или L-изолейцина или α-кетокислот кетоизовалерата, кетометилвалерата или кетоизокапроата.
Настоящее изобретение объяснено более подробно ниже с помощью иллюстративных вариантов осуществления.
Пример 1. Клонирование замещающей конструкции pK18mobsacB_PprpD2-ilvB
Исходя из геномной последовательности Corynebacterium glutamicum ATCC14067 в компании Life Technologies GmbH (Дармштадт, Германия) синтезировали фрагмент ДНК размером 1390 п.о. (SEQ ID NO: 5), который состоит из следующих компонентов:
• гомологичного участка ДНК выше ilvB
• промотора PprpD2 из C. glutamicum ATCC14067
• сайта связывания рибосомы гена gap из C. glutamicum ATCC14067
• гомологичного участка гена ilvB, который вместо инициирующего кодона GTG несет инициирующий кодон ATG.
Фрагмент клонировали при помощи терминально введенных сайтов расщепления EcoRI и HindIII посредством соответствующего расщепления с помощью двух упомянутых ферментов рестрикции и последующего лигирования в вектор pK18mobsacB, аналогично расщепленный с помощью EcoRI и HindIII. Плазмида имеет обозначение pK18mobsacB_PprpD2-ilvB. Она позволяет получить мутанта, в котором нативный промотор гена ilvB удален и заменен индуцибельным промотором PprpD2. В нем нативный инициирующий кодон (GTG), кроме того, заменен инициирующим кодоном ATG, предпочтительным для рибосомы.
Пример 2. Конструирование замещающей конструкции pK18mobsacB_ilvN(M13)
Исходя из геномной последовательности Corynebacterium glutamicum ATCC14067 в компании Life Technologies GmbH (Дармштадт, Германия) синтезировали фрагмент ДНК размером 1421 п.о. (SEQ ID NO: 14), который содержит часть гена ilvB, межгенную область между геном ilvB и геном ilvN, а также часть гена ilvN. В нем нативную последовательность “GGAATCATT” в гене ilvN (от +58 до +66 п.о. ниже начала гена ilvN, при этом начало гена определяется как +1) заменили на „GATGACTTT“. Таким образом, аминокислотную последовательность белка IlvN в положениях 20, 21 и 22 меняют с Gly (20), Ile(21), Ile(22) на Asp(20), Asp(21), Phe(22).
Фрагмент лигировали при помощи терминально введенных сайтов расщепления EcoRI и HindIII посредством соответствующего расщепления с помощью двух упомянутых ферментов рестрикции и последующего лигирования в вектор pK18mobsacB, аналогично расщепленный с помощью EcoRI и HindIII. Плазмида имеет обозначение pK18mobsacB_ilvN(M13). Она позволяет получить мутанта, в котором нативную последовательность гена “GGAATCATT” (от +58 до +66 п.о. ниже начала гена ilvN, при этом начало гена определяется как +1) заменили на “GATGACTTT”.
Пример 3. Конструирование мутантов C. glutamicum ATCC14067_PprpD2-ilvBN, C. glutamicum ATCC14067_ilvN(M13)_PprpD2-ilvBN и C. glutamicum VPS_PprpD2-ilvBN.
Вектор pK18mobsacB_PprpD2-ilvB, упомянутый в примере 1, перенесли с помощью электропорации согласно протоколу Liebl et al. (FEMS Microbiology Letters 65, 299-304 (1989)) в штамм Corynebacterium glutamicum ATCC14067 и в производящие валин штаммы Corynebacterium glutamicum ATCC14067_ilvN(M13) (см. пример 4) и штамм Corynebacterium glutamicum, производящий валин, C. glutamicum VPS. Вектор pK18mobsacB или pK18mobsacB_PprpD2-ilvB не способен независимо реплицироваться в C. glutamicum ATCC14067, C. glutamicum ATCC14067_ilvN(M13) и C. glutamicum VPS, и сохраняется в клетках, только если был интегрирован в хромосому в результате акта рекомбинации. Селекцию клонов, содержащих интегрированный pK18mobsacB_PprpD2_ilvB, выполняют путем осаждения конъюгационной серии на агар LB (Sambrook et al., Molecular Cloning: A Laboratory Manual, 2nd Ed., Cold Spring Habor, Нью-Йорк, 1989), который был дополнен 15 мг/л канамицина и 50 мг/мл налидиксовой кислоты. Наросшие клоны наносят штрихом на чашки с агаром LB, содержащим 25 мг/л канамицина, и инкубируют при 33°C в течение 16 часов. Для селекции мутантов, в которых в результате второго акта рекомбинации имело место вырезание плазмиды, клоны культивируют неселективно в жидкой среде LB в течение 20 часов, затем наносят штрихом на агар LB, содержащий 10% сахарозу, и инкубируют в течение 24 часов.
Плазмида pK18mobsacB_PprpD2-ilvB, а также исходная плазмида pK18mobsacB, содержит в дополнение к гену устойчивости к канамицину копию гена sacB, кодирующего левансахаразу из Bacillus subtilis. Экспрессия, индуцированная сахарозой, ведет к образованию левансахаразы, которая катализирует синтез продукта левана, токсичного для C. glutamicum. На LB-агаре, содержащем сахарозу, следовательно, растут только те клоны, в которых интегрированная pK18mobsacB_PprpD2-ilvB в свою очередь была вырезана. В случае вырезания вместе с плазмидой может быть вырезана либо полная копия гена ilvB дикого типа, включая промоторный участок дикого типа, либо рекомбинантная копия гена ilvB, содержащая промотор PprpD2.
Приблизительно от 40 до 50 колоний протестировали на фенотип "рост в присутствии сахарозы" и "отсутствие роста в присутствии канамицина". Для того, чтобы доказать, что рекомбинантный аллель PprpD2-ilvB остался в хромосоме, приблизительно 20 колоний, которые содержали фенотип "рост в присутствии сахарозы" и "отсутствие роста в присутствии канамицина", исследовали с помощью полимеразной цепной реакции согласно стандартному способу PCR по Innis et al. (PCR Protocols. A Guide to Methods and Applications, 1990, Academic Press). В связи с этим фрагмент ДНК, который несет модифицированные участки рекомбинантного аллеля PprpD2-ilvB, амплифицировали из хромосомной ДНК колоний. Следующие праймеры-олигонуклеотиды выбирали для проверочной PCR.
Контрольный праймер 1 (SEQ ID NO: 6)
5'-AAA GCC TGC ATC GCG GAG AC-3'
Контрольный праймер 2 (SEQ ID NO: 7)
5'-TGG TGA TGC CGC GGA TAT CG-3'
Праймеры делают возможной амплификацию фрагмента ДНК размером приблизительно 880 п.о. в клонах, содержащих рекомбинантный локус PprpD2-ilvBN. В клонах, содержащих локус PilvBN-ilvBN дикого типа, амплифицируются фрагменты ДНК, имеющие размер приблизительно 1136 п.о.
Амплифицированные фрагменты ДНК идентифицировали с помощью электрофореза в агарозном геле с концентрацией 0,8%. Таким образом можно было показать, что штаммы несут в хромосоме модифицированный рекомбинантный аллель PprpD2-ilvBN. Штаммы обозначили как Corynebacterium glutamicum ATCC14067_PprpD2-ilvBN, ATCC14067_ilvN(M13)_PprpD2-ilvBN и VPS_PprpD2-ilvBN.
Пример 4. Конструирование штамма C. glutamicum ATCC14067_ilvN(M13)
Вектор pK18mobsacB_ilvN(M13), упомянутый в примере 2, перенесли в штамм Corynebacterium glutamicum ATCC14067 с помощью электропорации аналогично способу, описанному в примере 3. Селекцию клонов выполняли с помощью методов культивирования, упомянутых в примере 3. Детекцию позитивных клонов провели на основе хромосомной ДНК, которая была выделена из 20 клонов, путем амплификации продукта длиной 947 п.о. с помощью полимеразной цепной реакции с использованием контрольных праймеров 3 и 4
Контрольный праймер 3 (SEQ ID NO: 15)
5'- CCC AGT AGT CAT CGA CTT C -3'
Контрольный праймер 4 (SEQ ID NO: 16)
5'- CAG CGT CAG CAT CAT AAA GC -3'
и последующего секвенирования продукта PCR.
Пример 5. Тест на продуктивность Corynebacterium glutamicum ATCC14067_PprpD2-ilvBN применительно к получению L-валина
Для исследования способности производить L-валин пять клонов штамма Corynebacterium glutamicum ATCC14067_PprpD2-ilvBN и в качестве эталона штамм Corynebacterium glutamicum ATCC14067 предварительно культивировали в 10 мл контрольной среды в каждом случае в течение 16 ч при 33°С. Для теста на продуктивность каждые 10 мл контрольной среды инокулировали полученной предварительной культурой, так что начальная OD 600 (оптическая плотность при 600 нм) составляла 0,1. Каждый клон тестировали в трех встряхиваемых сосудах, так что иллюстративный штамм представлен в общей сложности пятнадцатью встряхиваемыми сосудами.
Контрольная среда была идентична среде CgXII, описанной в Keilhauer et al. (Journal of Bacteriology (1993) 175: 5593-5603), но дополнительно содержала 7,5 г/л дрожжевого экстракта (Difco (Becton Dickinson GmbH), Гейдельберг). Состав контрольной среды обобщен в нижеследующей таблице 1. Контрольная среда для индуцирования синтеза валина дополнительно содержала пропионат в концентрации 0,6 г/л (в пересчете на свободную кислоту).
Таблица 1
Культивирование выполняли при 33°C и 200 об./мин. во встряхиваемых сосудах на 100 мл. Смещение шейкера составило 5 см. Через 24 и 48 часов образцы извлекали из культур, определяли оптическую плотность содержания декстрозы и содержания L-валина и клетки короткое время центрифугировали (центрифуга настольного типа 5415D (Eppendorf), 13000 об./мин., 10 мин при комнатной температуре).
Оптическую плотность определяли при длине волны 660 нм с использованием фотометра для микротитровальных планшетов GENios (Tecan, Рединг, Соединенное Королевство). Образцы перед измерением разводили 1:100 деминерализованной водой.
Декстрозу определяли с помощью теста с сопряженными ферментами (гексокиназа/глюкозо-6-фосфатдегидрогеназа) посредством образования NADH.
Концентрации внеклеточных аминокислот определяли количественно в культуральном супернатанте с использованием HPLC с обращенной фазой (Lindroth et al., Analytical chemistry (1979) 51: 1167-1174). Использовали прибор HPLC серии HP1100 (Hewlett-Packard, Вальдброн, Германия) с присоединенным детектором флуоресценции (G1321A); управление системой и анализ данных выполняли с помощью HP Chem-Station (Hewlett-Packard). 1 мкл раствора аминокислоты, подлежащей анализу, смешивали в автоматическом предколоночном получении производных с 20 мкл готового реагента - ортофталевого альдегида/2-меркаптоэтанола (Pierce Europe BV, Ауд-Бейерланд, Нидерланды). Флуоресцирующие тиозамещенные изоиндолы, полученные при этом (Jones et al., Journal of Chromatography (1983) 266: 471-482), разделяли с использованием комбинированной предколонки (40x4 мм Hypersil ODS 5) и основной колонки (Hypersil ODS 5, обе колонки от компании CS-Chromatographie Service GmbH, Лангервее, Германия) с помощью градиентной программы с увеличением неполярной фазы (метанол). Полярным элюентом был ацетат натрия (0,1 M; pH 7,2); скорость потока составила 0,8 мл в минуту. Детекцию флуоресценции производных аминокислот проводили при длине волны возбуждения 230 нм и длине волны излучения 450 нм. Концентрации валина рассчитывали путем сравнения с внешним стандартом.
Для расчета выхода количество образованного L-валина делили на количество поглощенной декстрозы.
Результаты представлены в таблице 2 и показывают, что иллюстративный штамм Corynebacterium glutamicum ATCC14067_PprpD2-ilvBN значительно выделяет валин в среду в присутствии пропионата, в то время как без пропионата он не отличается от контрольного штамма Corynebacterium glutamicum ATCC14067, который незначительно производит валин при любых условиях.
Таблица 2. Образование L-валина после инкубирования в течение 24 часов без пропионовой кислоты в среде (таблица 2A) или с 0,6 г/л пропионовой кислоты в среде (таблица 2B). Сокращения: *: ATCC 14067_PprpD2-ilvBN, станд. откл.: стандартное отклонение.
Таблица 2A. Результаты без пропионовой кислоты
г/л
(± станд. откл.)
г/г
(± станд. откл.)
(± станд. откл.)
Таблица 2B. Результаты с пропионовой кислотой
г/л
(± станд. откл.)
г/г
(± станд. откл.)
(± станд. откл.)
Пример 6. Тест на продуктивность с использованием производящих валин штаммов Corynebacterium glutamicum
Аналогично примеру 5, штаммы Corynebacterium glutamicum ATCC14067_ilvN(M13)_PprpD2-ilvBN и Corynebacterium glutamicum VPS_PprpD2-ilvBN исследовали в системе встряхиваемых сосудов.
Для исследования способности производить L-валин штамм Corynebacterium glutamicum ATCC14067_ilvN(M13)_PprpD2-ilvBN и в качестве эталона штамм Corynebacterium glutamicum ATCC14067_ilvN(M13), или штамм Corynebacterium glutamicum VPS_PprpD2-ilvBN и в качестве эталона штамм Corynebacterium glutamicum VPS, в каждом случае 10 мл контрольной среды (таблица 3), предварительно культивировали при 33°С в течение 16 ч. Для теста на продуктивность каждые 10 мл контрольной среды инокулировали полученной предварительной культурой, так что начальная OD600 (оптическая плотность при 600 нм) составляла 0,1. Каждый клон тестировали в трех встряхиваемых сосудах, так что иллюстративный штамм представлен в общей сложности 15 встряхиваемыми сосудами.
Таблица 3. SK1039 использовали в качестве контрольной среды
Контрольная среда для индуцирования синтеза валина дополнительно содержала пропионат в концентрации 0,6 г/л (в пересчете на свободную кислоту).
Условия культивирования и определение концентрации биомассы, декстрозы и валина выполняли аналогично тому, как описано в примере 5.
Результаты представлены в таблице 4 и показывают, что производящие валин штаммы Corynebacterium glutamicum ATCC14067_ilvN(M13)_PprpD2-ilvBN и VPS_PprpD2-ilvBN при присутствии пропионата в среде имеют более высокий специфический выход валина относительно использованного источника углерода, чем в каждом из случаев немодифицированных исходных штаммов Corynebacterium glutamicum ATCC14067_ilvN(M13) и Corynebacterium glutamicum VPS.
Таблица 4. Выход L-валина после инкубирования в течение 24 часов без пропионовой кислоты в среде (таблица 4A) или с 0,6 г/л пропионовой кислоты в среде (таблица 4B); сокращения: станд. откл. = стандартное отклонение.
Таблица 4A. Результаты без пропионовой кислоты
г/г
(± станд. откл.)
Таблица 4B. Результаты с пропионовой кислотой
г/г
(± станд. откл.)
Пример 7. Тест на стабильность валина для штаммов VPS_PprpD2-ilvBN и VPS
Тест на стабильность проводили в 10 мл жидких культур (как в примере 6).
Предварительное культивирование штаммов VPS и VPS_PprpD2-ilvBN
В 10 мл жидкой культуры во встряхиваемом сосуде на 100 мл (с перегородками) инокулировали по 50 мкл каждой непрерывной глицериновой культуры и инкубировали в течение 22 ч (33°C, 200 об./мин., амплитуда 5 см).
Оптическую плотность культуры измеряли и 1,5 мл культуры обрабатывали глицерином (10% глицерин в конечной концентрации) и замораживали в качестве криокультуры при -80°C в сосуде с завинчивающейся крышкой.
В следующие 10 мл жидкой культуры инокулировали по 50 мкл культур в каждую и инкубировали снова при 33°C в течение 22 ч со встряхиванием.
Эту процедуру повторяли дополнительно дважды, так что каждую глицериновую культуру культивировали в общей сложности в четырех последовательных жидких культурах. Каждое культивирование соответствует приблизительно 8 клеточным поколениям. Четыре пассажа в жидких культурах, таким образом, соответствовали в общей сложности более чем 30 (приблизительно 32) клеточным поколениям.
Основное культивирование штаммов VPS и VPS_PprpD2-ilvBN
В культуры во встряхиваемых сосудах каждой непрерывной культуры или криокультуры инокулировали по 10 мл жидкой культуры с начальной OD 0,1. Затем культуры инкубировали в течение 24 ч. Каждое культивирование выполняли в двойном определении. В конце инкубирования (через 24 ч) образцы брали для анализа оптической плотности, титра валина и остаточной концентрации сахаров. Анализы выполняли, как описано в примере 5.
оптическая плотность (OD)
валин/декстроза
Y (P/S) [г/г]
Результат теста на продуктивность (таб. 5) показал, что с каждым пассажем титр валина (указанный в относительном изменении процента), отношение валин/биомасса (указанное в валин/OD) и выход (г образованного продукта/г поглощенного субстрата) для штамма VPS становился слабее или ниже. Это является свидетельством того факта, что в популяции закрепляются мутации, которые являются негативными для образования продукта и позитивными для образования биомассы. После двух стадий культивирования (15,6 поколений) штамм VPS уже показывает спад в данных продуктивности в контрольном культивировании (результаты основного культивирования). Таким образом, в 4-стадийном процессе (3 стадии культивирования + 1 стадия получения продукта) можно ожидать выраженное снижение данных продуктивности (титр, выход, специфичное для биомассы образование продукта). Однако для штамма VPS_PprpD2-ilvBN согласно настоящему изобретению данный негативный эффект не наблюдался даже при пересеве в четырех стадиях культивирования с более чем 30 поколениями. В отличие от этого специфичное для биомассы образование валина (валин/OD) даже несколько увеличивается. Таким образом, по меньшей мере 4-стадийный способ получения, состоящий из 3 стадий культивирования и основного культивирования, воспроизведен или необходимое условие даже превзойдено.
Таблица 5. Данные продуктивности в тесте на стабильность (выход L-валина, специфичное для биомассы образование продукта (валин/OD) и относительные изменения образования валина в зависимости от дополнительных клеточных поколений относительно контроля (=0) после инкубирования в течение 24 часов (средние значения))
Использованные сокращения и обозначения имеют следующие значения.
oriV: ColE1-подобная точка начала репликации pMB1
sacB: ген sacB, кодирующий белок левансахаразы
RP4mob: сайт мобилизации RP4
Kan: ген устойчивости к канамицину
PprpD2: индуцируемый пропионатом промотор
'ilvB: 5'-область гена ilvB
HindIII: сайт расщепления фермента рестрикции HindIII
EcoRI: сайт расщепления фермента рестрикции EcoRI

















Группа изобретений относится к области биотехнологии. Предложен способ получения L-валина путем ферментации микроорганизмов из рода Corynebacterium, содержащих в реплицируемой форме полинуклеотид с операторной активностью, последовательность которого по меньшей мере на 85% идентична последовательности от положения 1 до 121 согласно SEQ ID NO: 1, 2 или 3, с которым связывается активатор PrpR, и функционально ниже которого на 3’-конце находится второй полинуклеотид, характеризующийся промоторной активностью, а также гены ilvB и ilvN, и который регулирует транскрипцию генов ilvBN в зависимости от добавления активатора PrpR в среду, к которой после первой фазы (фазы роста), проходящей без индуктора, во второй фазе в качестве индуктора добавляют пропионат или 2-метилцитрат. Предложено применение указанного полинуклеотида с операторной активностью для регуляции экспрессии генов ilvBN. Предложена кассета экспрессии для использования в указанном способе, содержащая указанный полинуклеотид с операторной активностью, расположенный ниже промотор и гены ilvBN. Предложен вектор экспрессии, содержащий указанную кассету экспрессии. Предложено применение указанной кассеты экспрессии и вектора экспрессии для экспрессии генов ilvBN в микроорганизмах рода Corynebacterium. Предложен микроорганизм рода Corynebacterium для получения L-валина указанным способом, содержащий в реплицируемой форме указанную кассету экспрессии или указанный вектор экспрессии. Группа изобретений позволяет увеличить выход L-валина. 7 н. и 10 з.п. ф-лы, 1 ил., 7 табл., 7 пр. 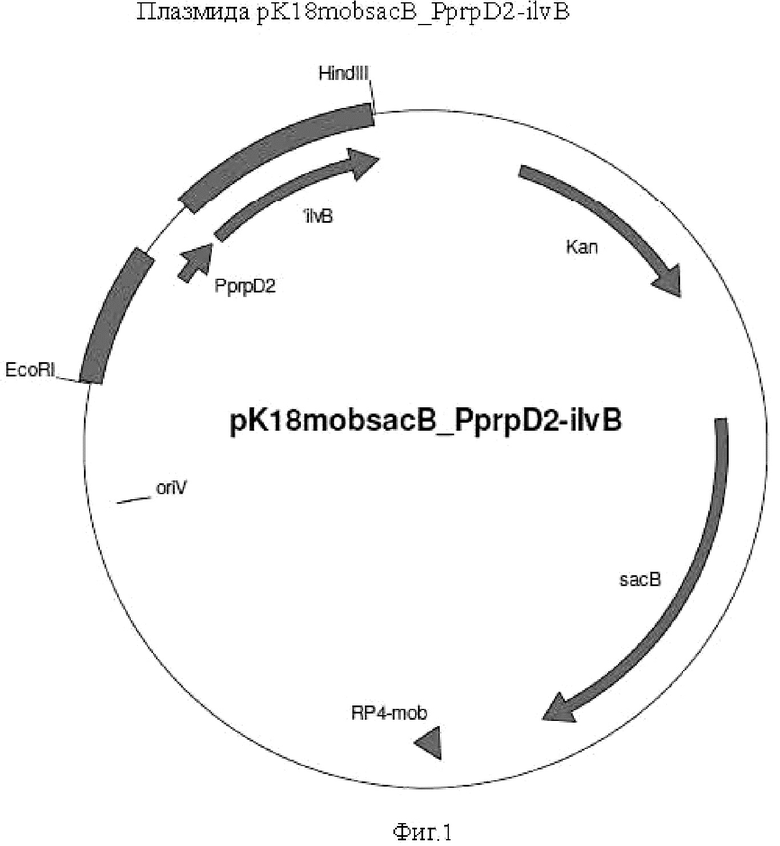
1. Способ получения L-валина путем ферментации микроорганизмов из рода Corynebacterium, содержащих в реплицируемой форме полинуклеотид с операторной активностью, последовательность которого по меньшей мере на 85% идентична последовательности от положения 1 до 121 согласно SEQ ID NO: 1, 2 или 3, с которым связывается активатор PrpR, и функционально ниже которого на 3'-конце находится второй полинуклеотид, характеризующийся промоторной активностью, а также гены ilvB и ilvN, кодирующие субъединицы ацетолактатсинтазы, и который регулирует транскрипцию генов ilvBN в зависимости от добавления активатора PrpR в среду, к которой после первой фазы (фазы роста), проходящей без индуктора, во второй фазе в качестве индуктора добавляют пропионат или 2-метилцитрат, после чего синтезируют L-валин в условиях, при которых увеличивается содержание L-валина в среде и/или в клетках.
2. Способ по п.1, отличающийся тем, что полинуклеотид с операторной активностью предусматривает полинуклеотид ("IR 1"), последовательность которого по меньшей мере на 90%, предпочтительно по меньшей мере на 92, 94 или 96%, особенно предпочтительно на 100% идентична последовательности от положения 22 до положения 49 согласно SEQ ID NO: 1, 2 или 3, а также полинуклеотид ("IR 2"), последовательность которого по меньшей мере на 90%, предпочтительно по меньшей мере на 92, 94 или 96%, особенно предпочтительно на 100% идентична последовательности от положения 77 до положения 105 согласно SEQ ID NO: 1, 2 или 3, при этом полинуклеотид с операторной активностью предпочтительно представляет собой полинуклеотид с последовательностью, которая по меньшей мере на 90, 92, 94 или 96%, особенно предпочтительно на 100% идентична последовательности от положения 1 до 121 согласно SEQ ID NO: 1, 2 или 3.
3. Способ по п.1 или 2, отличающийся тем, что полинуклеотид с промоторной активностью представляет собой полинуклеотид, характеризующийся последовательностью, которая по меньшей мере на 90, 92, 94 или 96%, в частности, по меньшей мере на 97, 98 или 99%, особенно предпочтительно на 100% идентична последовательности от положения 122 до 206 согласно SEQ ID NO: 4.
4. Способ по п.1, отличающийся тем, что ген ilvB является полинуклеотидом, который кодирует полипептид, последовательность которого по меньшей мере на 90, 92, 94 или 96%, в частности, по меньшей мере на 97, 98 или 99%, особенно предпочтительно на 100% идентична последовательности согласно SEQ ID NO: 9, и ген ilvN является полинуклеотидом, который кодирует полипептид, последовательность которого по меньшей мере на 90, 92, 94 или 96%, в частности, по меньшей мере на 97, 98 или 99%, особенно предпочтительно на 100% идентична последовательности согласно SEQ ID NO: 10.
5. Способ по п.1, отличающийся тем, что используют микроорганизмы, в которых присутствующие дополнительные ферменты биосинтетического пути L-валина дополнительно амплифицированы, в частности сверхэкспрессированы, и/или в которых по меньшей мере частично ослаблены метаболические пути, которые снижают образование L-валина.
6. Способ по п.1, отличающийся тем, что коринебактерии, используемые в фазе культивирования, проходят по меньшей мере 16, предпочтительно по меньшей мере 24 поколения, и ферментация предпочтительно включает по меньшей мере четыре стадии, предпочтительно предусматривающие встряхиваемый сосуд, предпосевной ферментер, посевной ферментер и ферментер для получения продукта.
7. Применение полинуклеотида, характеризующегося операторной активностью, последовательность которого по меньшей мере на 85% идентична последовательности от положения 1 до 121 согласно SEQ ID NO: 1, 2 или 3, для регуляции экспрессии генов ilvBN, кодирующих ацетолактатсинтазу.
8. Применение по п.7, где указанный полинуклеотид применяют в комбинации с промотором, расположенным выше генов.
9. Кассета экспрессии для использования в способе по любому из пп.1-6, содержащая полинуклеотид, характеризующийся операторной активностью, последовательность которого по меньшей мере на 85% идентична последовательности от положения 1 до 121 согласно SEQ ID NO: 1, 2 или 3, расположенный ниже промотор, а также гены ilvBN, кодирующие ацетолактатсинтазу.
10. Кассета экспрессии по п.9, где указанный промотор характеризуется последовательностью, по меньшей мере на 90% идентичной последовательности от положения 122 до 206 согласно SEQ ID NO: 4.
11. Вектор экспрессии, содержащий кассету экспрессии по п.9 или 10.
12. Применение кассеты экспрессии по п.9 или 10 для экспрессии генов ilvBN в микроорганизмах из рода Corynebacterium.
13. Применение вектора по п.11 для экспрессии генов ilvBN в микроорганизмах из рода Corynebacterium.
14. Микроорганизм из рода Corynebacterium для получения L-валина для осуществления способа по любому из пп.1-6, содержащий в реплицируемой форме кассету экспрессии по п.9 или 10 или вектор экспрессии по п.11.
15. Микроорганизм по п.14, отличающийся тем, что является микроорганизмом из вида Corynebacterium glutamicum.
16. Микроорганизм по п.14 или 15, отличающийся тем, что кассета экспрессии была интегрирована в хромосому микроорганизма.
17. Микроорганизм по любому из пп.14-16, отличающийся тем, что он производит L-валин, и в котором предпочтительно в повышенном содержании дополнительно присутствуют дополнительные ферменты биосинтетического пути L-валина, в частности сверхэкспрессированые, и/или в котором метаболические пути, которые снижают образование L-валина, по меньшей мере частично ослаблены.
| PLASSMEIER J.K | |||
| et al., A propionate-inducible expression system based on the Corynebacterium glutamicum prpD2 promoter and PrpR activator and its application for the redirection of amino acid biosynthesis pathways, Journal of Biotechnology Vol | |||
| Деревянное стыковое устройство | 1920 |
|
SU163A1 |
| Синхронизирующее устройство для аппарата, служащего для передачи изображений на расстояние | 1920 |
|
SU225A1 |
| DE 102011118019 A1, 03.01.2013 | |||
| Устройство для зачистки деталей | 1983 |
|
SU1096010A1 |
| PLASSMEIER J | |||
| et al., Molecular characterization of PrpR, the transcriptional activator of propionate catabolism in Corynebacterium glutamicum, Journal of Biotechnology Vol | |||
| Катодное реле | 1918 |
|
SU159A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| DATABASE EMBL, accession no | |||
| AF434799, Corynebacterium glutamicum prpDBC2 gene cluster, complete sequence, 15.04.2005; Найдено в Интернет 30.03.2018 по адресу: www.ebi.ac.uk/ena/data/view/AF434799 | |||
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ | 2003 |
|
RU2307165C2 |
| СПОСОБ ФЕРМЕНТАТИВНОГО ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ С ИСПОЛЬЗОВАНИЕМ КОРИНЕФОРМНЫХ БАКТЕРИЙ | 2000 |
|
RU2247778C2 |

