Область техники, к которой относится изобретение
Настоящее изобретение относится к слитым белкам, подавляющим индуцирующие ангиогенез факторы, и их применению. В частности, настоящее изобретение относится к слитым белкам, которые ингибируют множество ангиогенных факторов, и их применению. Более конкретно, настоящее изобретение относится к слитым белкам рецепторов VEGF и FGF и их применению в лечении заболеваний, связанных с нарушением ангиогенеза.
Уровень техники
Ангиогенез является одним из основных факторов, приводящих к росту и метастазированию злокачественной опухоли [1]. Процесс ангиогенеза регулируется многими факторами: некоторые из них стимулируют ангиогенез, в то время как другие факторы ингибируют ангиогенез; таким образом, регуляция ангиогенеза является очень сложным динамическим равновесным процессом [2]. Основной целью антиангиогенной терапии является остановка роста опухоли за счет блокирования стимулирующих ангиогенез факторов или предотвращения ангиогенеза опухоли при помощи использования ингибиторов ангиогенеза. В настоящее время известно большое количество стимулирующих ангиогенез факторов таких как, пример, фактор роста сосудистого эндотелия (VEGF), фактор роста фибробластов (FGF), фактор роста гепатоцитов (HGF), DDR1, EphA1, EphA2, EphAS, EphB1, EphB4, EGFR, HER-2, ErbB3, MET, RON, CSF1R, KIT, PDGFR-A, PDGFR-B, ТЕК, Tie-1 и т.п., которые способны стимулировать деление и дифференцировку клеток сосудистого эндотелия и морфогенез кровеносных сосудов. Известно, что среди этих факторов самым специфическим ангиогенным фактором и самым эффективным фактором роста является VEGF [3, 4].
В гипоксической среде внутри опухолевой ткани клетками этой ткани секретируется большое количество VEGF, которые индуцируют деление и миграцию сосудистых эндотелиоцитов, приводя к образованию сосудистой сети опухоли. В ходе большого количества экспериментов с использованием животных было показано, что ингибирование VEGF может предотвращать ангиогенез, а также подавлять рост опухоли. По этой причине VEGF и его рецепторы стали важной мишенью для антиангиогенных лекарственных средств. К настоящему времени в клинических испытаниях было показано, что антиангиогенные лекарственные средства обладают необычайной эффективностью, включая бевацизумаб (известный как Авастин), который способен блокировать непосредственно VEGF и ингибировать ангиогенез опухоли. Продажа бевацизумаба была разрешена Управлением по надзору за качеством пищевых продуктов и лекарственных средств США в 2004 в качестве лекарственного средства первой линии для лечения рака прямой кишки, это был первый разрешенный для коммерческого использования препарат, предупреждающий канцерогенез за счет ингибирования ангиогенеза. Авастин представляет собой гуманизированное анти-VEGF моноклональное антитело, который производится известной американской биотехнологической компанией Genentech. В III фазе крупномасштабных клинических испытаний было показано, что комбинированное с химиотерапией лечение Авастином может существенно удлинять время жизни пациентов с различными видами рака, том числе раком прямой кишки, легких, молочной железы почек и т.д. [5, 6] Клинический успех авастина является исторически значимым, являясь демонстрацией того, что антиангиогенная терапия, мишенью которой является сосудистая система опухоли, является клинически эффективным мероприятием и обеспечивает новый подход к лечению опухолей. В западных странах авастин уже нашел широкое применение в терапии опухолей и является одним из лекарственных средств, удерживающих лидирующее положение на мировом рынке.
На последней фазе клинических испытаний с участием людей, помимо авастина, находится еще несколько лекарственных средств, подавляющих передачу сигналов VEGF, клиническое применение которых ожидается в течение следующих нескольких лет. В настоящее время на III фазе крупномасштабных клинических испытаний, среди прочих препаратов, находится афлиберцепт (также известный как VEGF-Trap), разработанный американской биотехнологической компанией Regeneron совместно с компанией Sanofi-Aventis [7]. На III фазе клинических испытаний также находится лекарственное средство IMC-1121B (имклон), представляющее собой моноклональное антитело к VEGF рецептору II (VEGFR2) [8]. Эти новые лекарственные средства несомненно обеспечат новые возможности для терапии рака, а также надежду для пациентов.
Использование анти-VEGF препарата позволило достичь большого прогресса в лечении рака, однако в клинических испытаниях было показано, что анти-VEGF терапия также имеет существенные ограничения. С точки зрения эффективности лечения опухоли авастин способен увеличить время выживания у 50% пациентов с раком прямой кишки примерно на 3-4 месяца [9, 10], и на 7-8 месяцев у 5% пациентов с раком груди [11], и следовательно, авастин не способен эффективно подавлять рост кровеносных сосудов в опухолях в течение длительного времени. Таким образом, проблема, требующая решения заключается в том, как улучшить эффективность антиангиогенной терапии, при этом не менее важной является задача изучения и разработки антиангиогенных препаратов следующего поколения.
Основные причины, приводящие к неудачам в лечении анти-VEGF препаратами или появлению резистентности, могут быть связаны с регуляцией ангиогенеза опухоли, опосредованной множеством факторов. Хотя VEGF играет важную роль в ангиогенезе, он не является единственным фактором, стимулирующим ангиогенез. Тем не менее, в силу гетерогенности опухолевых клеток, сложности микроокружения опухоли и ответных компенсаторных механизмов тела при длительном подавлении активности VEGF могут экспрессироваться другие стимулирующие ангиогенез факторы [12], и, таким образом, рост кровеносной системы опухоли перестает зависеть от пути передачи VEGF сигналов. Группа Hanahan изучала изменение факторов ангиогенеза, экспресируемых опухолью, во время лечения опухоли поджелудочной железы анти-VEGFR2 препаратами, показав, что во время лечения анти-VEGF препаратами наблюдается изменение уровня экспрессия нескольких генов, причем экспрессия FGF-2 значительно усиливается. Было показано, что в резистентной к лечению анти-VEGF препаратами опухоли уровень экспрессии FGF, в частности FGF-2, увеличивается настолько, что процесс ангиогенеза вновь активируется, но после блокирования пути передачи сигналов FGF происходит подавление репопуляции опухоли [13]. Отсюда следует, что сверхэкспрессия FGF-2 тесно связана со способностью опухоли избегать лечения анти-VEGF препаратами. Следовательно, пролиферативные сосудистые заболевания, такие как опухоли, можно лечить более эффективно путем одновременного ингибирования как VEGF, так и FGF-2.
В настоящее время, в области изучения антагонизма, направленного на ингибирование двух или множества мишеней, достигаемого благодаря применению лекарственного средства на основе низкомолекулярного активного соединения достигнут некоторый прогресс, и продемонстрировано, что противоопухолевый эффект, достигаемый в результате одновременного подавления действия как VEGF, так и FGF-2, превосходит эффект, получаемый в результате противоопухолевой терапии, нацеленной на единственную мишень [14]. Однако отсутствие специфичности в действии низкомолекулярных многоцелевых антагонистов может приводить к появлению непредвиденных побочных эффектов, которые порой могут проявляться только на поздней стадии клинических исследований, и, таким образом, их применение является очень рискованным. В то же время, лекарственные средства на основе макромолекулярных белков, в частности Fc-слитого белка и моноклонального антитела, имеют преимущества, не свойственные лекарственным средствам, созданным на основе низкомолекулярных активных соединений, такие как высокая специфичность и длительный in vivo период полувыведения и т.д., что делает их важным объектом изучения и разработки лекарственных средств.
Фактор роста фибробластов (FGF) относится к семейству гепарин-связывающих факторов роста, которое у млекопитающих состоит из 22 членов (FGF 1-14, 16-23). FGF выполняет множество важных биологических функций, таких как размножение, дифференцировка и миграция клеток, ангиогенез и онкогенез. Многие биологические функции FGF осуществляются через связывание и активацию FGF рецепторов клеточной поверхности (FGFR). (См, например, Eswarakumar et al. Cytokine Growth Factor Rev.16:139-149, 2005). Рецепторы фактора роста фибробластов (FGFR) представляют собой рецепторы, которые связываются с членами семейства факторов роста фибробластов. Часть рецепторов факторов роста фибробластов участвует в развитии патологического процесса. У млекопитающих к ним относятся 4 гена FGFR: fgfRl-fgfR4. Рецепторы факторов роста фибробластов состоят из внеклеточного домена, трансмембранного домена и внутриклеточного домена. Члены семейства FGFR отличаются друг от друга лиганд-связывающими свойствами и киназными доменами. Однако имеют одинаковые внеклеточные домены. В составе их внеклеточных доменов различают три иммуноглобулин-подобных (Ig-подобных) домена: первый Ig-подобный домен, второй Ig-подобный домен и третий Ig-подобный домен, при этом между первым и вторым Ig-подобными доменами имеется последовательность. Эта последовательность, расположенная между первым и вторым Ig-подобными доменами, в настоящем описании названа участком промежуточной функциональной последовательности Ig-подобного домена FGFR. Этот участок промежуточной функциональной последовательности содержит область кислых аминокислот, названную кислым боксом (АВ).
Учитывая важное влияние VEGF и FGF (особенно FGF-2) на ангиогенез опухоли, авторами изобретения было выдвинуто предположение, что лекарственные средства на основе макромолекулярных белков, ингибирующие ангиогенную активность как VEGF, так и FGF (особенно FGF-2), смогут оказывать более сильное ингибирующее действие на ангиогенез опухоли и позволят достигнуть улучшенного терапевтического эффекта в медицинской практике. Однако на сегодняшний день нет ни одной публикации, описывающей успешно сконструированный макромолекулярный слитый белок, подавляющий активность как VEGF, так и FGF. Следовательно, настоящее изобретение направлено на изучение вопроса создания Fc-слитого белка, который способен одновременно подавлять ангиогенную активность как VEGF, так и FGF (особенно FGF-2), при этом указанный слитый белок способен одновременно блокировать путь передачи сигналов как VEGF, так и FGF благодаря специфичности и высокой аффинности к этим рецепторам. Слитый белок, одновременно подавляющий ангиогенную активность как VEGF, так и FGF, полностью состоит из последовательности человеческого белка и обладает превосходными фармакокинетическими свойствами Fc-слитого белка, демонстрируя при этом благоприятные фармацевтические параметры и хорошие возможности для применения в клинике. В то же время, аффинность слитого белка, направленного на подавление ангиогенной активности двух мишеней, зависит от правильной конфигурации белка, при этом рецепторный сегмент в таком слитом белке обнаруживает тенденцию к потере лиганд-связывающей способности. Следовательно, разработка лекарственного средства, содержащего высокоэффективный слитый белок, который мог бы специфично подавлять как VEGF, так и FGF (например, FGF-2), сопряжена с трудностями. На первой стадии этого исследования было сконструировано более 20 слитых белков, содержащих разные VEGFR сегменты, сегменты FGFR и Fc IgG1, и был получен слитый белок, обладающий высокой аффинностью как к VEGF, так и к FGF (в частности, к FGF-2).
Несмотря на наличие публикаций, описывающих многочисленное количество ингибирующих ангиогенез слитых белков, например, FGFR-слитый белок (WO/2008/065543), Notch3-слитый белок (WO/2010/021729). VEGFR-слитый белок (WO/2010/105573), LK8-слитый белок (WO/2008/075833) и т.д., все эти слитые белки направлены на одну мишень, при этом ингибирование ангиогенеза реализуется за счет слияния части единственного ингибитора ангиогенеза с Fc-сегментом иммуноглобулина. Отсутствуют публикации уровня техники, описывающие слитый белок, эффективность ингибирования ангиогенеза которого достигалась бы за счет ингибирования двух мишеней благодаря успешному слиянию двух структурных единиц ингибирующих образование кровеносных сосудов. Автору изобретения впервые удалось достичь слияния по меньшей мере двух ингибирующих ангиогенез структурных единиц, выделенных из по меньшей мере двух ингибиторов ангиогенеза, для получения слитого белка, ингибирующего две мишени, а также ингибирующего ангиогенез, и неожиданно получить отличный эффект. Следовательно, настоящее изобретение обеспечивает новое направление в разработке лекарственных средств, ингибирующих ангиогенез.
Сущность изобретения
Один из аспектов настоящего изобретения относится к слитому белку, содержащему по меньшей мере две ингибирующие ангиогенез структурные единицы, полученные по меньшей мере из двух ингибиторов ангиогенеза. В одном из вариантов осуществления изобретения указанный слитый белок ингибирует ангиогенез. В другом варианте осуществления указанный слитый белок связывается с FGF и VEGF in vivo и/или in vitro.
В одном из вариантов осуществления изобретения заявленный слитый белок содержит по меньшей мере две ингибирующие ангиогенез структурные единицы, полученные из по меньшей мере двух ингибиторов ангиогенеза, где указанные по меньшей мере два ингибитора ангиогенеза выбирают из группы состоящей из: VEGFR, например, VEGFR1, VEGFR2, VEGFR3, и FGFR, например, FGFR1, FGFR2, FGFR4. Слитый белок по изобретению предпочтительно содержит по меньшей мере две ингибирующие ангиогенез структурные единицы, полученные из VEGFR1, VEGFR2 и FGFR1. Более предпочтительно, слитый белок по изобретению содержит две ингибирующие ангиогенез структурные единицы, полученные из VEGFR1, VEGFR2 и FGFR1, где одна из ингибирующих ангиогенез структурных единиц получена из VEGFR1 и VEGFR2, в то время как другая получена из FGFR1.
В другом варианте осуществления изобретения слитый белок по изобретению содержит по меньшей мере две ингибирующие ангиогенез структурные единицы, полученные из по меньшей двух ингибиторов ангиогенеза, где по меньшей два ингибитора ангиогенеза представляют собой растворимый сегмент ингибирующего ангиогенез рецептора, причем указанный растворимый сегмент рецептора, например, может быть выбран из группы, состоящей из: DDR1, EphA1, EphA2, EphAS, EphB1, EphB4, EGFR, HER-2, ЕrbВ3, FGFR1, FGFR2, FGFR4, MET, RON, CSF1R, KIT, PDGFR-A, PDGFR-B, ТЕК, Tie-1, HGF, VEGFR1, VEGFR2, VEGFR3 и их аллельных вариантов. В одном из конкретных вариантов осуществления указанный ингибитор ангиогенеза представляет собой ингибирующий ангиогенез рецептор VEGFR И FGFR.
В некоторых вариантах осуществления слитый белок по изобретению содержит по меньшей мере две ингибирующие ангиогенез структурные единицы, полученные из по меньшей мере двух (предпочтительно, двух или трех) ингибиторов ангиогенеза, где по меньшей мере две ингибирующие ангиогенез структурные единицы содержат по меньшей мере одну ингибирующую ангиогенез структурную единицу, полученную из внеклеточного домена VEGFR, и по меньшей мере одну ингибирующую ангиогенез структурную единицу, полученную из внеклеточного домена FGFR, при этом, внеклеточный домен VEGFR предпочтительно представляет собой внеклеточный домен VEGFR1, VEGFR2 и/или VEGFR3, а внеклеточный домен FGFR предпочтительно представляет собой внеклеточный домен FGFR1 и/или FGFR2.
В некоторых вариантах осуществления слитый белок по изобретению содержит по меньшей мере две ингибирующие ангиогенез структурные единицы, полученные из по меньшей мере двух (предпочтительно, двух или трех) ингибиторов ангиогенеза, где по меньшей мере две ингибирующие ангиогенез структурные единицы содержат по меньшей мере одну ингибирующую ангиогенез структурную единицу, полученную из внеклеточного домена VEGFR1 и/или VEGFR2, и одну ингибирующую ангиогенез структурную единицу, полученную из внеклеточного домена FGFR1.
В некоторых вариантах осуществления часть, полученная из внеклеточного домена VEGFR (такого как VEGFR1 и/или VEGFR2), содержит один или более элементов, выбранных из группы, состоящей из, или состоит из: первого Ig-подобного домена VEGFR (такого как VEGFR1 или VEGFR2) или его фрагмента, второго Ig-подобного домена VEGFR (такого как VEGFR1 или VEGFR2) или его фрагмента, третьего Ig-подобного домена VEGFR (такого как VEGFR1 или VEGFR2) или его фрагмента, четвертого Ig-подобного домена VEGFR (такого как VEGFR1 или VEGFR2) или его фрагмента, пятого Ig-подобного домена VEGFR (такого как VEGFR1 или VEGFR2) или его фрагмента, шестого Ig-подобного домена VEGFR (такого как VEGFR1 или VEGFR2) или его фрагмента и седьмого Ig-подобного домена VEGFR (такого как VEGFR1 или VEGFR2) или его фрагмента.
В некоторых вариантах осуществления часть, полученная из внеклеточного домена FGFR (такого как FGFR1), содержит один или более элементов, выбранных из группы, состоящей из, или состоит из: первого Ig-подобного домена FGFR (такого как FGFR1) или его фрагмента, где указанная часть получена из участка промежуточной функциональной последовательности Ig-подобного домена FGFR (такого как FGFR1), второго Ig-подобного домена FGFR (такого как FGFR1) или его фрагмента и третьего Ig-подобного домена FGFR (такого как FGFR1) или его фрагмента. В частности, домены и/или сегменты, содержащиеся в слитом белке по изобретению, могут быть связаны друг с другом непосредственно и/или через линкеры. В одном из конкретных вариантов осуществления слитый белок по изобретению содержит часть, полученную из участка промежуточной функциональной последовательности Ig-подобного домена FGFR (такого как FGFR1); предпочтительно, указанная часть, полученная из участка промежуточной функциональной последовательности Ig-подобного домена FGFR (такого как FGFR1), не содержит кислый бокс (АВ). В одном из конкретных вариантов осуществления часть, полученная из участка промежуточной функциональной последовательности имеет последовательность, выбранную из группы, состоящей из: аминокислотной последовательности, соответствующей положениям 134-162, положениям 145-162 или положениям 151-162 SEQ ID NO:1.
В одном из вариантов осуществления часть, полученная из внеклеточного домена VEGFR, содержит: второй Ig-подобный домен VEGFR1 или VEGFR2 и третий Ig-подобный домен VEGFR1 или VEGFR2; часть, полученная из внеклеточного домена FGFR (такого как FGFR1), содержит: часть, полученную из участка промежуточной функциональной последовательности Ig-подобного домена FGFR, второй Ig-подобный домен FGFR и третий Ig-подобный домен FGFR. Часть, полученная из участка промежуточной функциональной последовательности Ig-подобного домена FGFR, предпочтительно не содержит кислый бокс. В частности, указанный FGFR, пример, представляет собой FGFR1 или FGFR2.
В другом варианте осуществления часть, полученная из внеклеточного домена VEGFR, содержит: второй Ig-подобный домен VEGFR1 и третий Ig-подобный домен VEGFR2; часть, полученная из внеклеточного домена FGFR, содержит: часть, полученную из участка промежуточной функциональной последовательности Ig-подобного домена FGFR, второй Ig-подобный домен FGFR третий Ig-подобный домен FGFR. Часть, полученная из участка промежуточной функциональной последовательности Ig-подобного домена FGFR, предпочтительно не содержит кислый бокс. В частности, указанный FGFR, например, представляет собой FGFR1 или FGFR2.
В другом варианте осуществления часть, полученная из внеклеточного домена VEGFR, содержит последовательно в направлении от N-конца к С-концу второй Ig-подобный домен VEGFR1 и третий Ig-подобный домен VEGFR2; часть, полученная из внеклеточного домена FGFR, содержит последовательно в направлении от N-конца к С-концу, часть, полученную из участка промежуточной функциональной последовательности Ig-подобного домена FGFR, второй Ig-подобный домен FGFR и третий Ig-подобный домен FGFR. Часть, полученная из участка промежуточной функциональной последовательности Ig-подобного домена FGFR, предпочтительно не содержит кислый бокс. В частности, указанный FGFR, например, представляет собой FGFR1 или FGFR2.
В некоторых конкретных вариантах осуществления часть, полученная из внеклеточного домена VEGFR, также содержит первый Ig-подобный домен VEGFR1. Например, за первым Ig-подобным доменом VEGFR1 следует второй Ig-подобный домен VEGFR1. Первый Ig-подобный домен VEGFR1 предпочтительно имеет аминокислотную последовательность, соответствующую положениям 32-123 SEQ ID NO:2, или аминокислотную последовательность, имеющую по меньшей мере 70% идентичность, предпочтительно по меньшей мере 80%, 90%, 93%, 95%, 97%, 98% или 99% идентичность, с аминокислотной последовательностью, соответствующей положениям 32-123 SEQ ID NO:2.
В некоторых конкретных вариантах осуществления часть, полученная из внеклеточного домена FGFR (такого как FGFR1) дополнительно содержит первый Ig-подобный домен FGFR или его фрагмент. Первый Ig-подобный домен FGFR или его фрагмент предпочтительно имеет аминокислотную последовательность, соответствующую положениям 40-118 SEQ ID NO:1, или аминокислотную последовательность, имеющую по меньшей мере 70% идентичность, предпочтительно по меньшей мере 80%, 90%, 93%, 95%, 97%, 98% или 99% идентичность, с последовательностью, соответствующей положениям 40-118 SEQ ID NO:1; или
аминокислотную последовательность, соответствующую положениям 77-118 SEQ ID NO:1, или аминокислотную последовательность, имеющую по меньшей мере 70% идентичность, предпочтительно по меньшей мере 80%, 90%, 93%, 95%, 97%, 98% или 99% идентичность, с аминокислотной последовательностью, соответствующей положениям 77-118 SEQ ID NO:1.
В некоторых предпочтительных вариантах осуществления изобретения второй Ig-подобный домен VEGFR1 имеет аминокислотную последовательность, соответствующую положениям 151-214 SEQ ID NO:2, или аминокислотную последовательность, имеющую по меньшей мере 70% идентичность, предпочтительно по меньшей мере 80%, 90%, 93%, 95%, 97%, 98% или 99% идентичность, с аминокислотной последовательностью, соответствующей положениям 151-214 SEQ ID NO:2.
В некоторых других предпочтительных вариантах осуществления изобретения третий Ig-подобный домен VEGFR2 имеет аминокислотную последовательность, соответствующую положениям 224-320 SEQ ID NO:3, или аминокислотную последовательность, имеющую по меньшей мере 70% идентичность, предпочтительно по меньшей мере 80%, 90%, 93%, 95%, 97%, 98% или 99% идентичность, с аминокислотной последовательностью, соответствующей положениям 224-320 SEQ ID NO:3.
В некоторых других предпочтительных вариантах осуществления изобретения второй Ig-подобный домен FGFR1 имеет аминокислотную последовательность, соответствующую положениям 163-247 SEQ ID NO:1, или аминокислотную последовательность, имеющую по меньшей мере 70% идентичность, предпочтительно по меньшей мере 80%, 90%, 93%, 95%, 97%, 98% или 99% идентичность, с аминокислотной последовательностью, соответствующей положениям 163-247 SEQ ID NO:1.
В других предпочтительных вариантах осуществления изобретения третий Ig-подобный домен FGFR1 имеет аминокислотную последовательность, соответствующую положениям 270-359 SEQ ID NO:1, или аминокислотную последовательность, имеющую по меньшей мере 70% идентичность, предпочтительно по меньшей мере 80%, 90%, 93%, 95%, 97%, 98% или 99% идентичность, с аминокислотной последовательностью, соответствующей положениям 270-359 SEQ ID NO:1.
Предпочтительно, слитый белок по изобретению содержит дополнительную структурную единицу, например, Fc-область иммуноглобулина, предпочтительно, Fc-область человеческого IgG, более предпочтительно, Fc-область человеческого IgGI, более предпочтительно, слитый белок содержит:
аминокислотную последовательность, соответствующую SEQ NO:7. или аминокислотную последовательность, имеющую по меньшей мере 70% идентичность, предпочтительно по меньшей мере 80%, 90%, 93%, 95%, 97%, 98% или 99% идентичность, с аминокислотной последовательностью SEQ ID NO:7; или
аминокислотную последовательность, кодируемую нуклеотидной последовательностью, соответствующей SEQ ID NO:8, или аминокислотную последовательность, кодируемую нуклеотидной последовательностью, имеющей по меньшей мере 70% идентичность, предпочтительно по меньшей мере 80%, 90%, 93%, 95%, 97%, 98% или 99% идентичность, с нуклеотидной последовательностью SEQ ID NO:8.
В некоторых других вариантах осуществления слитый белок по изобретению дополнительно содержит область секреторного сигнального пептида, например, область сигнального пептида VEGFR1, и предпочтительно указанная область секреторного сигнального пептида имеет аминокислотную последовательность положений 1-26 SEQ ID NO:2 или аминокислотную последовательность, кодируемую нуклеотидной последовательностью SEQ ID NO:25.
В другом аспекте настоящее изобретение предоставляет Fc-слитый белок, содержащий:
(1) аминокислотную последовательность, имеющую по меньшей мере 70% идентичность, предпочтительно по меньшей мере 80%, 90%, 93%, 95%, 97%, 98% или 99% идентичность, с любой аминокислотной последовательностью из SEQ ID NO:9-24;
(2) аминокислотную последовательность, кодируемую нуклеотидной последовательностью, имеющей о меньшей мере 70% идентичность, предпочтительно по меньшей мере 80%, 90%, 93%, 95%, 97%, 98% или 99% идентичность, с любой нуклеотидной последовательностью из SEQ ID NO:26-41; или
(3) любую аминокислотную последовательность из SEQ ID NO:9-24, или аминокислотную последовательность, кодируемую любой нуклеотидной последовательностью из SEQ ID NO:26-41.
В некоторых вариантах осуществления настоящего изобретения части и/или домены, содержащиеся в VEGFR-FGFR-Fc слитом белке, могут быть расположены в любом порядке в направлении от N-конца к С-концу, например, как показано на Фиг. 1.
В некоторых вариантах осуществления VEGFR-FGFR-Fc слитый белок по настоящему изобретению дополнительно содержит сигнальный пептид (SP), предпочтительно секреторный сигнальный пептид, например, сигнальный пептид VEGFR1, который имеет аминокислотную последовательность, соответствующей положениям 1-26 SEQ ID NO:2 или аминокислотную последовательность, кодируемую нуклеотидной последовательностью SEQ ID NO:25. Сигнальный пептид предпочтительно локализован на N-конце слитого белка.
Предпочтительно, слитый белок по изобретению содержит последовательно в направлении от N-конца к С-концу часть, полученную из внеклеточного домена VEGFR, и часть, полученную из внеклеточного домена FGFR.
В слитом белке по настоящему изобретению Fc-область иммуноглобулина предпочтительно представляет собой Fc-область человеческого IgG, например, Fc-область человеческого IgG1, более предпочтительно, слитый белок содержит:
аминокислотную последовательность, соответствующую SEQ NO:1, или аминокислотную последовательность, имеющую по меньшей мере 70% идентичность, предпочтительно по меньшей мере 80%, 90%, 93%, 95%, 97%, 98% или 99% идентичность, с аминокислотной последовательностью SEQ ID NO:7; или
аминокислотную последовательность, кодируемую нуклеотидной последовательностью, соответствующей SEQ ID NO:8, или аминокислотную последовательность, кодируемую нуклеотидной последовательностью, имеющей по меньшей мере 70% идентичность, предпочтительно по меньшей мере 80%, 90%, 93%, 95%, 97%, 98% или 99% идентичность, с нуклеотидной последовательностью SEQ ID NO:8.
В одном из вариантов настоящего изобретения Fc-область иммуноглобулина локализована на С-конце слитого белка.
В некоторых вариантах настоящего изобретения области и/или домены, содержащиеся в VEGFR-FGFR-Fc слитом белке, могут находиться в любом порядке в направлении от N-конца до С-конца слитого белка. В некоторых других вариантах осуществления указанный порядок может быть таким, как показано на Фиг. 1. В некоторых конкретных вариантах осуществления часть VEGFR и часть FGFR могут быть расположены в прямом, в направлении от N-конца к С-концу, или противоположном порядке по отношению друг к другу, соответственно. Более того, Fc-область может находиться выше или ниже ингибирующей ангиогенез структурной единицы; например, Fc-область может находиться на С-конце слитого белка.
В некоторых вариантах VEGFR-FGFR-Fc слитый белок по настоящему изобретению дополнительно содержит одну или более межцепочечных дисульфидных связей, предпочтительно содержит одну или более межцепочечных дисульфидных связей в Ig-подобном домене.
В одном из аспектов настоящего изобретения VEGFR-FGFR-Fc слитый белок может быть получен в результате экспрессии слитого белка по изобретению в прокариотической или эукариотической клетке, например, клеточными линиями бактерий, грибов (таких как дрожжи) и млекопитающих. В частности, клеточная линия млекопитающих может представлять собой клеточную линию СНО. Клетки СНО представляют собой клеточную линию, часто используемую для экспрессии рекомбинантного белка. Помимо этого, исходную клетку СНО модифицируют, руководствуясь различными требованиями для крупномасштабной экспрессии, и, таким образом, для специализированной экспрессии была получена серия производных клеточных линий СНО, используемых для продуцирования рекомбинантных белков, например, для культуры, не содержащей серу, и т.п. Все они известны в области экспрессии рекомбинантных белков.
В другом аспекте домены и/или области, включенные в слитый белок по изобретению, связаны непосредственно друг с другом и/или через линкер, и, например, часть, полученная из внеклеточного домена VEGFR, часть, полученная из внеклеточного домена FGFR, и часть, полученная из Fc-области иммуноглобулина, могут быть связаны через линкер или непосредственно друг с другом. В одном из вариантов осуществления часть, полученная из внеклеточного домена VEGFR, часть, полученная из внеклеточного домена FGFR, и Fc-область иммуноглобулина непосредственно связаны друг с другом. В другом варианте осуществления часть, полученная из внеклеточного домена VEGFR, часть, полученная из внеклеточного домена FGFR, и Fc-область иммуноглобулина связаны посредством линкера, например, связаны посредством (С4S)3 линкера.
В другом аспекте настоящее изобретение относится к изолированным молекулам нуклеиновой кислоты, кодирующей слитый белок. Молекула нуклеиновой кислоты предпочтительно содержит любую нуклеотидную последовательность из SEQ ID NO:26-41.
В другом аспекте настоящего изобретения предоставляется вектор, содержащий молекулу нуклеиновой кислоты. Также предоставляются клетки, предпочтительно клетки СНО, трансформированные/трансфицированные вектором.
В другом аспекте настоящего изобретения предоставляется фармацевтическая композиция, содержащая слитый белок по изобретению, молекулу нуклеиновой кислоты, вектор или клетку, и фармацевтически приемлемый носитель.
Настоящее изобретение также относится к способу продуцирования слитого белка для ингибирования ангиогенеза, который осуществляется путем экспрессии слитого белка по изобретению в прокариотических клетках или эукариотических клетках, например, клеточных линиях бактерий, грибов (таких как дрожжи) или млекопитающих. Предпочтительно, клеточная линия млекопитающих может представлять собой клеточную линию СНО.
В другом аспекте настоящего изобретения предоставляется способ ингибирования ангиогенеза, где способ содержит введение эффективного для ингибирования ангиогенеза количества слитого белка по изобретению, молекулы нуклеиновой кислоты, вектора, клетки или фармацевтической композиции, нуждающемуся в этом субъекту. Предпочтительно, способ осуществляется у млекопитающих.
В другом аспекте настоящего изобретения предоставляется способ лечения или профилактики опухолей у млекопитающих, где способ содержит ведение терапевтически или превентивно эффективного количества слитого белка по изобретению, молекулы нуклеиновой кислоты, вектора, клетки или фармацевтической композиции нуждающемуся в этом субъекту, предпочтительно, опухоль представляет собой солидную опухоль.
В другом аспекте настоящего изобретения предоставляется способ лечения или профилактики у млекопитающих заболеваний, связанных с офтальмологическим ангиогенезом, где способ содержит введение терапевтически или превентивно эффективного количества слитого белка по изобретению, молекулы нуклеиновой кислоты, вектора, клетки или фармацевтической композиции нуждающемуся в этом субъекту, при этом, заболевания, связанные с офтальмологическим ангиогенезом предпочтительно выбирают из группы, состоящей из возрастной дегенерации желтого пятна, диабетической ретинопатии т.п.
В другом аспекте настоящего изобретения предоставляется способ связывания FGF и/или VEGF in vitro или in vivo, содержащий обеспечение контакта FGF и VEGF со слитым белком по изобретению.
Настоящее изобретение также относится к применению слитого белка, молекулы нуклеиновой кислоты, кодирующей указанный слитый белок, вектора, содержащего указанную молекулу нуклеиновой кислоты, клетки, трансформированной/трансфицированной указанным вектором, или фармацевтической композиции, содержащей его, согласно настоящему изобретению для производства лекарственного средства для ингибирования ангиогенеза. Более того, настоящее изобретение также относится к применению слитого белка по настоящему изобретению, молекулы нуклеиновой кислоты, кодирующей указанный слиты белок, вектора, содержащего указанную молекулу нуклеиновой кислоты, клетки, трансформированной/трансфицированной указанным вектором, или фармацевтической композиции, содержащей его, для производства лекарственного средства для лечения или профилактики связанных с ангиогенезом заболеваний, причем связанные с ангиогенезом заболевания предпочтительно представляют собой опухоль или заболевание, связанное с офтальмологическим ангиогенезом.
Учитывая разные требования в разных странах к объекту, защищаемому в патентных системах, указанное раскрытие также предусматривает фармацевтическое применение, соответствующее указанным выше способам, и лекарственные средства для медицинского применения. Фармацевтическое применение и лекарственные средства также входят в объем защиты настоящего изобретения, как если бы они уже были конкретно описаны в настоящем раскрытии.
В настоящем описании приведены примеры только для нескольких конкретных вариантов осуществления изобретения, в отношении которого испрашивается защита, при этом технические признаки, описанные в одном или более технических предложениях, могут быть объединены с любым одним или более признаками одного и/или более технических предложений, и эти технические предложения, полученные путем комбинации, также входят в объем защиты заявки, как если бы эти технические предложения, полученные путем комбинации, были уже конкретно раскрыты в настоящем описании.
Ниже приводится более подробное описание настоящего изобретения со ссылкой на прилагаемые чертежи и описание в виде примеров, приведенных исключительно с целью иллюстрации. Следует иметь в виду, что приведенное ниже описание проиллюстрировано путем предоставления примеров технических предложений, заявленных в рамках настоящего изобретения, причем указанные примеры не следует рассматривать в качестве ограничения этих технических предложений. Объем защиты настоящего изобретения, определяется исключительно прилагаемой формулой изобретения.
Краткое описание чертежей
На Фиг. 1 представлена структура VEGFR-FGFR-Fc слитого белка, на которой отдельно указаны VEGFR1, VEGFR2 и FGFR1 белки. Fc-слитый белок показан жирной линией, а удаленная аминокислота показана пунктирной линией; антителоподобный домен представлен в виде окружности; разные антителоподобные домены обозначены буквой + номером, где домен VEGFR1 представлен как а1-а7, домен VEGFR2 представлен как b1-b7, домен FGFR1 представлен как с1-с3; дисульфидная связь обозначена s s; человеческий Fc IgG1 представлен в виде серого прямоугольника; сигнальный пептид обозначен SP; (G4S)3-связывающая последовательность представлена в виде трех ромбов; последовательность кислого бокса представлена в виде прямоугольника, обозначенного АВ.
Фиг. 2 представляет собой сравнительную диаграмму, демонстрирующую связывание VEGF и FGF-2 соответствующими Fc-слитыми белками, на которой связывание между соответствующим Fc-слитым белком (20 нг/мл) и VEGF165 и/или FGF-2 (содержащими 100 нг/мл гепарина), иммобилизованными на планшете, определено методом ELISA.
На Фиг. 3 показано изображение SDS-PAGE геля для слитого белка.
На Фиг. 4 показано связывание VEGF (А) и FGF-2 (В) в зависимости от градиента концентрации слитых белков.
На Фиг. 5 показана аффинность слитого белка к VEGF (А) и FGF-2 (В).
На Фиг. 6 показано влияние слитого белка на деление клеток HUVEC, индуцированное VEGF или FGF-2, и относительные уровни ингибирования. На Фиг. 6А показано влияние слитого белка на пролиферацию клеток HUVEC, индуцированную VEGF, а на Фиг. 6В показаны относительные уровни ингибирования слитым белком пролиферации клеток HUVEC, индуцированной VEGF, на Фиг. 6С показано влияние слитого белка на пролиферацию клеток HUVEC, индуцированную FGF2, и на Фиг. 6D показаны относительные уровни ингибирования слитым белком пролиферации клеток HUVEC, индуцированной FGF2.
Описание предпочтительных вариантов осуществления
Определения
Если не указано иное, все используемые в этом описании научные термины имеют общепринятое значение, известное специалисту в данной области техники. Для прояснения определений и терминов, общепринятых в данной области техники, специалист может обратиться к имеющимся на сегодняшний день в молекулярной биологии протоколам (Ausubel). Стандартное трех и/или однобуквенное обозначение, используемое для указания одной из 20 обычных L-амино кислот, является общепринятым в данной области сокращенным названием аминокислотных остатков.
Хотя в настоящем изобретении приведены широкие числовые диапазоны, а приблизительные значения параметров заданы в широких пределах, все числа, встречающиеся в конкретных примерах, определены с максимальной точностью. Тем не менее, по существу в любом числовом значении имеется некоторая погрешность, которая может быть результатом стандартного отклонения, возникающего при измерении любого параметра. К тому же, следует иметь в виду, что все приведенные здесь диапазоны значений охватывают все возможные содержащиеся в нем поддиапазоны. Например, следует иметь в виду, что диапазон "от 1 до 10", как указано в этом описании, охватывает все возможные поддиапазоны, начиная с минимального значения 1 вплоть до максимального значения 10 (включая конечные точки); т.е., все поддиапазоны, начиная с минимального значения 1 и более, например, от 1 до 6,1, при этом, все поддиапазоны ограничены максимальным значением 10 или менее, например, от 5,5 до 10. Кроме того, также следует иметь в виду, что любая ссылка, обозначенная как "включенный в данное описание", рассматривается включенной в описание во всей ее полноте.
Также следует иметь в виду, что, если четко и однозначно не указано иное, форма единственного числа также включает множественное число, согласно тому, как это используется в данном описании. Термин "или" и термин "и/или" являются взаимозаменяемыми, если иное не следует из контекста очевидным образом.
Используемые здесь термины "Fc", "Fc-область", "Fc-фрагмент" или " Fc-область иммуноглобулина" относятся к кристаллизуемому фрагменту иммуноглобулина, и в настоящем изобретении эта Fc-область предпочтительно представляет собой человеческую Fc-область IgG1.
Термин "Fc-слитый белок" относится к антителоподобной молекуле, которая одновременно обладает специфичностью связывания гетерологичного белка и эффекторной функцией константной области иммуноглобулина. С точки зрения молекулярной структуры, Fc-слитый белок содержит аминокислотную последовательность, имеющую требуемую специфичность связывания, и последовательность константной области иммуноглобулина. Как правило, молекула Fc-слитого белка содержит сайт связывания рецептора или лиганда. Последовательность константной области иммуноглобулина может быть получена из любого иммуноглобулина, например, подтипа IgG-1, IgG-2, IgG-3 или IgG-4, IgA (включая IgA-1 и IgA-2), IgE, IgD или IgM.
Термин "растворимый" белок, согласно тому как используется в настоящем описании, относится к белку, который может быть растворен в водном растворе при температуре, значении рН и осмотическом давлении, которые характерны для биологических сред. Выражение "растворимый слитый белок", согласно тому как используется в настоящем описании, означает, что слитый белок не содержит трансмембранной области или внутриклеточной области.
Используемый здесь термин "изолированный" относится к следующему веществу и/или объекту: (1) изолированному от по меньшей мере некоторых компонентов, которые присутствуют изначально при его получении (в природной окружающей среде и/или в экспериментальном устройстве) и связаны с ним, и/или (2) продуцированному, полученному и/или произведенному искусственно. Изолированное вещество и/или объект могут быть изолированы от других исходно связанных с ним компонентов на примерно 10%, примерно 20%, примерно 30%, примерно 40%, примерно 50%, примерно 60%, примерно 70%, примерно 80%, примерно 90%, примерно 95%, примерно 98%, примерно 99%, по существу 100% или 100%.
Термины "часть" и "сегмент" взаимозаменяемо относятся к части полипептида, нуклеиновой кислоты или другим молекулярным структурам.
Используемый здесь термин "VEGFR" относится к рецептору фактора роста эндотелия сосудов, который может представлять собой VEGFR1, VEGFR2 и/или VEGFR3. Предпочтительно, VEGFR в настоящем изобретении представляет собой VEGFR1 и/или VEGFR2, и предпочтительно является человеческим VEGFR.
Используемый здесь термин "FGFR" относится к рецептору фактора роста фибробластов, который может представлять собой FGFR1, FGFR2, FGFR3 и/или FGFR4. В настоящем изобретении FGFR предпочтительно представляет собой FGFR1, и более предпочтительно является человеческим FGFR1.
Используемый здесь термин "Ig-подобный домен" относится к иммуноглобулин-подобному домену, который встречается во множестве семейств белков и задействован во многих биологических функциях, включая распознавание клетка-клетка, рецептор клеточной поверхности, иммунную функцию т.д.
Используемый здесь термин "первый Ig-подобный домен VEGFR" относится к первому от N-конца Ig-подобному домену белка VEGFR, и предпочтительно, первому от N-конца Ig-подобному домену белка VEGFR1 (называемому здесь первым Ig-подобным доменом VEGFR1) или первому от N-конца Ig-подобному домену белка VEGFR2 (называемому здесь первым Ig-подобным доменом VEGFR2), в котором, например, аминокислотная последовательность соответствует положениям 32-123 SEQ ID NO:2 или положениям 46-110 SEQ ID NO:3. Аналогично, второй Ig-подобный домен VEGFR имеет, например, аминокислотную последовательность, соответствующую положениям 151-214 SEQ ID NO:2 или положениям 141-207 SEQ ID NO:3, третий Ig-подобный домен VEGFR имеет, например, аминокислотную последовательность, соответствующую положениям 230-327 SEQ ID NO:2 или положениям 224-320 SEQ ID NO:3, четвертый Ig-подобный домен VEGFR имеет, например, аминокислотную последовательность, соответствующую положениям 335-421 SEQ ID NO:2 или положениям 328-414 SEQ ID NO:3, пятый Ig-подобный домен VEGFR имеет, например, аминокислотную последовательность, соответствующую положениям 428-553 SEQ ID NO:2 или положениям 421-548 SEQ ID NO:3, шестой Ig-подобный домен VEGFR имеет, например, аминокислотную последовательность, соответствующую положениям 556-654 SEQ ID NO:2 или положениям 551-660 SEQ ID NO:3, и седьмой Ig-подобный домен VEGFR имеет, например, аминокислотную последовательность, соответствующую положениям 661-747 SEQ ID NO:2 или положениям 667-753 SEQ ID NO:3. Предпочтительно, VEGFR может представлять собой VEGFR1 или VEGFR2.
Используемый здесь термин "первый Ig-подобный домен FGFR" или "первый Ig-подобный домен FGFR1" относится к первому от N-конца Ig-подобному домену белка FGFR или FGFR1, который имеет, например, аминокислотную последовательность, соответствующую положениям 40-118 SEQ ID NO:1. Аналогично, термин "второй Ig-подобный домен FGFR" или "второй Ig-подобный домен" относится ко второму от N-конца Ig-подобному домену белка FGFR, который имеет, например, аминокислотную последовательность, соответствующую положениям 163-247 SEQ ID NO:1; термин "третий Ig-подобный домен FGFR" или "третий Ig-подобный домен" относится к третьему от N-конца Ig-подобному домену белка FGFR, который имеет, например, аминокислотную последовательность, соответствующую положениям 270-359 SEQ ID NO:1. Предпочтительно, FGFR представляет собой FGFR1, а первый Ig-подобный домен FGFR представляет собой первый Ig-подобный домен FGFR1, второй Ig-подобный домен FGFR представляет собой второй Ig-подобный домен FGFR1, и третий Ig-подобный домен FGFR представляет собой третий Ig-подобный домен FGFR1.
Используемый здесь термин "участок промежуточной функциональной последовательности Ig-подобного домена FGFR" или "участок промежуточной функциональной последовательности" относится к последовательности, расположенной между первым Ig-подобным доменом и вторым Ig-подобным доменом белка FGFR, при этом, последовательность IFS предпочтительно имеет аминокислотную последовательность, соответствующую положениям 118-162 SEQ ID NO:1. Авторами настоящего изобретения неожиданно было обнаружено, что участок промежуточной функциональной последовательности оказывает влияние на функцию Ig-подобного домена. Белок FGFR предпочтительно представляет собой FGFR1 (SEQ ID NO:1), в частности человеческий белок FGFR1. Аминокислотная последовательность человеческого белка FGFR1 представлена в SEQ ID NO:1, а последовательность его кДНК представлена в SEQ ID NO:4.
Ниже приведена часть последовательности hFGFR1, в которой каждый из Ig-подобных доменов показан областью, выделенной жирным шрифтом, в порядке их следования, см. http: // www.ncbi.nlm.nih.gov/protein/AAH15035.1

Аминокислотная последовательность FGFR1 представлена в SEQ ID NO:1, а кодирующая ее нуклеотидная последовательность представлена в SEQ ID NO:4.
Ниже приведена часть последовательности hVEGFR1, в которой каждый из Ig-подобных доменов показан областью, выделенной жирным шрифтом, в порядке их следования, а природные линкерные последовательности находятся между отдельными доменами, см. http: //www.uniprot.org/uniprot/P17948:

Аминокислотная последовательность VEGFR1 представлена в SEQ ID NO:2, а кодирующая ее нуклеотидная последовательность представлена в SEQ ID NO:5.
Ниже приведена часть последовательности hVEGFR2, в которой каждый из Ig-подобных доменов показан областью, выделенной жирным шрифтом, в порядке их следования, между которыми находятся природные связывающие последовательности, см. http://www.uniprot.org/uniprot/P35968

Аминокислотная последовательность VEGFR2 представлена в SEQ ID NO:3, а кодирующая ее нуклеотидная последовательность представлена в SEQ ID NO:6.
Используемый в настоящем описании термин "ингибирующая ангиогенез структурная единица" относится к полинуклеотидной секции, сегменту, мотиву или домену, который имеет функцию ингибирования ангиогенеза. Ингибирующая ангиогенез структурная единица может относиться к любому сегменту аминокислотной последовательности слитого белка по изобретению при условии, что она обладает способностью ингибировать ангиогенез. Например, ингибирующая ангиогенез единица по настоящему изобретению может содержать часть, полученную из внеклеточного домена VEGFR, и часть, полученную из внеклеточного домена FGFR.
Используемый в настоящем описании термин "вырожденный вариант" означает, что вырожденный вариант содержит вырожденное изменение в третьем положении аминокислотного кодона, при котором вырожденные варианты кодируют одну и ту же аминокислоту, например, положение неоднозначного соответствия триплетного кода содержит один или более измененных вариантов (также называемых измененным вариантом).
Используемый в настоящем описании термин "аллельный вариант" относится к двум или более генам, находящимся в определенном положении хромосомы.
Используемый в настоящем описании термин "субъект" относится к млекопитающему, такому как человек. Однако могут быть и другие животные, например домашние животные (такие как собаки, кошки и т.д.), сельскохозяйственные животные (такие как, крупный рогатый скот, овцы, свиньи, лошади и т.д.) или экспериментальные животные (такие как обезьяны, крысы, мыши, кролики, морские свинки и т.д.).
Используемый в настоящем описании термин "процент идентичности", "гомология" или "идентичность" относится к идентичности двух аминокислотных последовательностей или последовательностей нуклеиновых кислот. Процент идентичности определяют путем выравнивания двух последовательностей, причем процент идентичности относится к количеству одинаковых остатков (т.е. аминокислотных или нуклеотидных) в одинаковых положениях выравненных последовательностей. Выравнивание последовательностей и их сравнение можно осуществлять, используя стандартные в данной области алгоритмы (см., например. Smith and Waterman, 1981, Adv. Appl. Math. 2: 482; Needleman and Wunsch, 1970, J. Mol. Biol. 48: 443; Pearson and Lipman, 1988, Proc. Natl. Acad. Sci., USA, 85: 2444) или компьютерные версии этих алгоритмов (Wisconsin Genetics Software Package Release 7.0, Genetics Computer Group, 575 Science Drive, Madison, WI). К доступным версиям относятся программы BLAST и FASTA. Помимо этого, для выравнивания последовательностей можно использовать программу ENTREZ, предоставляемую национальным институтом здравоохранения (Bethesda MD). Если используются программы BLAST и GAP-BLAST, для каждой из них (например, BLASTN можно найти на сайте национального центра биотехнологической информации (National Center for Biotechnology Information) можно использовать параметры по умолчанию. В одном из вариантов осуществления относительную идентичность между двумя последовательностями можно определять при помощи программного пакета GCG, используя вес участка разрыва, равный 1, с тем, чтобы заданный вес для каждого аминокислотного разрыва соответствовал несовпадению между двумя последовательностями по одной аминокислоте. В качестве альтернативы можно использовать программный пакет для выравнивания последовательностей ALIGN (версия 2.0), включенный в пакет программ GCG (Accelrys, San Diego, CA).
Используемый в настоящем описании термин "гибридизация" относится к процессу, согласно которому стабильный двухнитевой полинуклеотид образуется через нековалентное связывание двух однонитевых полинуклеотидов. Термин "гибридизация" также может относиться к трехнитевой гибридизации. Полученный двухнитевой полинуклеотид (обычно) представляет собой "гибрид" или "дуплекс". "Условие гибридизации" обычно включает концентрацию соли ниже чем примерно 1 М, более предпочтительно, ниже чем примерно 500 мМ, и ниже чем примерно 200 мМ. Температура гибридизации может быть ниже 5°С, но обычно выше чем примерно 22°С, и лучше, если температура выше чем примерно 30°С, более предпочтительно выше чем примерно 37°С. Гибридизацию обычно выполняют в жестких условиях (т.е., условиях, при которых зонд гибридизуется со своей целевой последовательностью). Жесткие условия гибридизации зависят от последовательности и в разных случаях будут разными. Для гибридизации более длинных сегментов может потребоваться более высокая температура гибридизации. Поскольку на силу гибридизации могут влиять другие факторы (включая состав оснований и длину комплементарной нити, наличие органического растворителя и степень несоответствия оснований), комбинация параметров является более важной, чем абсолютное значение любого одного параметра. Обычно, выбирают жесткие условия, при которых температура на 5°С ниже чем Тm последовательности при определенной ионной силе и рН. Жесткие условия, например, включают рН 7,0-8,3, концентрацию ионов натрия (или других солей) по меньшей мере от 0,01 М до более чем 1 М, температуру по меньшей мере 25°С. Информацию о жестких условиях можно найти, например, у Sambrook, Fritsche и Maniatis. "Molecular Cloning A laboratory Manual", 2nd edition. Cold Spring Harbor Press (1989) and Anderson "Nucleic Acid Hybridization", 1st edition, BIOS Scientific Publishers Limited (1999), этот документ включен в настоящее описание во всей своей полноте в виде ссылки.
Используемые в настоящем описании термины "линкер", "пептидный линкер", "связывающая последовательность" или "линкерная последовательность" относятся к короткой аминокислотной последовательности, которая связывает вместе отдельные домены и/или области, составляющие слитый белок, и длина такой короткой аминокислотной последовательности обычно составляет 0-20 аминокислот, предпочтительно, 2-10 аминокислоты.
Используемый здесь термин "аминокислотная последовательность, соответствующая SEQ ID NO:N" в слитом белке или части или домене означает, что указанный слитый белок или часть или домен имеет аминокислотную последовательность по существу такую как указано в SEQ ID NO:N, и предпочтительно, содержащую не более чем 1, 2, 3, 4, 5, 10 или 20 замен, добавлений и делеций аминокислот, и более предпочтительно, указанный слитый белок или часть или домен имеет 80%, 90%, 93%, 95%, 97%, 98% или 99% идентичность с аминокислотной последовательностью SEQ ID NO:N, и более предпочтительно, указанный слитый белок или часть или домен имеет аминокислотную последовательность, приведенную в SEQ ID NO:N.
Используемый здесь термин "слитый белок VEGFR-FGFR-Fc" относится к слитому белку, который содержит часть, полученную из внеклеточного домена VEGFR, часть, полученную из внеклеточного домена FGFR и Fc-область. В некоторых вариантах осуществления часть, полученная из внеклеточного домена VEGFR, и часть, полученная из внеклеточного домена FGFR, может: (1) иметь любую аминокислотную последовательность из: 1-443 SEQ ID NO:9, 1-364 SEQ ID NO:10, 1-379 SEQ ID NO:11, 1-531 SEQ ID NO:12, 1-611 SEQ ID NO:13, 1-531 SEQ ID NO:14, 1-312 SEQ ID NO:15, 1-611 SEQ ID NO:16, 1-207 SEQ ID NO:17, 1-665 SEQ ID NO:18, 1-610 SEQ ID NO:19, 1-611 SEQ ID NO:20, 1-580 SEQ ID NO:21, 1-540 SEQ ID NO:22, 1-542 SEQ ID NO:23 и 1-435 SEQ ID NO:24, или последовательность, кодируемую любой нуклеотидной последовательностью из: 1-1326 SEQ ID NO:26, 1-1092 SEQ ID NO:27, 1-1137 SEQ ID NO:28, 1-1593 SEQ ID NO:29, 1-1833 SEQ ID NO:30, 1-1593 SEQ ID NO:31, 1-936 SEQ ID NO:32, 1-1833 SEQ ID NO:33, 1-621 SEQ ID NO:34, 1-1995 SEQ ID NO:35, 1-1830 SEQ ID NO:36, 1-1833 SEQ ID NO:37, 1-1740 SEQ ID NO:38, 1-1620 SEQ ID NO:39, 1-1626 SEQ ID NO:40 и 1-1305 SEQ ID NO:41; (2) содержать или состоять из аминокислотной последовательности, которая имеет по меньшей мере 70% идентичность, предпочтительно по меньшей мере 80%, 90%, 93%, 95%, 97%, 98% или 99% идентичность, с любой аминокислотной последовательностью из: 1-443 SEQ ID NO: 9, 1-364 SEQ ID NO: 10, 1-379 SEQ ID NO: 11, 1-531 SEQ ID NO: 12, 1-611 SEQ ID NO: 13, 1-531 SEQ ID NO: 14, 1-312 SEQ ID NO: 15, 1-611 SEQ ID NO: 16, 1-207 SEQ ID NO: 17, 1-665 SEQ ID NO: 18, 1-610 SEQ ID NO: 19, 1-611 SEQ ID NO: 20, 1-580 SEQ ID NO: 21, 1-540 SEQ ID NO: 22, 1-542 SEQ ID NO: 23 и 1-435 SEQ ID NO: 24; или (3) содержать или состоять из аминокислотной последовательности, кодируемой нуклеотидной последовательностью, которая имеет по меньшей мере 70% идентичность, предпочтительно 80%, 90%, 93%, 95%, 97%, 98% или 99% идентичность с любой нуклеотидной последовательностью из: 1-1326 SEQ ID NO: 26, 1-1092 SEQ ID NO: 27, 1-1137 SEQ ID NO: 28, 1-1593 SEQ ID NO: 29, 1-1833 SEQ ID NO: 30, 1-1593 SEQ ID NO:31, 1-936 SEQ ID NO: 32, 1-1833 SEQ ID NO: 33, 1-621 SEQ ID NO:34, 1-1995 SEQ ID NO: 35, 1-1830 SEQ ID NO: 36, 1-1833 SEQ ID NO: 37, 1-1740 SEQ ID NO: 38, 1-1620 SEQ ID NO: 39. 1-1626 SEQ ID NO: 40 и 1-1305 SEQ ID NO:41.
В некоторых предпочтительных вариантах осуществления изобретения слитый белок VEGFR-FGFR-Fc может: (1) иметь любую аминокислотную последовательность из SEQ ID NO: 9-24 или последовательность, кодируемую любой нуклеотидной последовательностью из SEQ ID NO: 26-41; (2) содержать или состоять из аминокислотной последовательности, которая имеет по меньшей мере 70% идентичность, предпочтительно по меньшей мере 80%, 90%, 93%, 95%, 97%, 98% или 99% идентичность, с любой аминокислотной последовательностью из SEQ ID NO: 9-24; или (3) содержать или состоять из аминокислотной последовательности, кодируемой нуклеотидной последовательностью, которая имеет по меньшей мере 70% идентичность, предпочтительно по меньшей мере 80%, 90%, 93%, 95%, 97%, 98% или 99% идентичность, с любой нуклеотидной последовательностью из SEQ ID NO: 26-41.
В некоторых предпочтительных вариантах осуществления слитый белок VEGFR-FGFR-Fc может кодироваться нуклеиновой кислотой, содержащей последовательность, комплементарная последовательность которой гибридизуется с любой нуклеотидной последовательностью из SEQ ID NO: 26-41 в жестких условиях, или содержащей вырожденный вариант любой нуклеотидной последовательности из SEQ ID NO: 26-41. В некоторых предпочтительных вариантах осуществления часть, полученная из Fc-области иммуноглобулина слитого белка VEGFR-FGFR-Fc может кодироваться нуклеиновой кислотой, содержащей последовательность, комплементарная последовательность которой гибридизуется с нуклеотидной последовательностью SEQ ID NO: 8 в жестких условиях, или содержащей вырожденный вариант нуклеотидной последовательности SEQ ID NO: 8.
В других предпочтительных вариантах осуществления слитый белок VEGFR-FGFR-Fc содержит вариант слитого белка VEGFR-FGFR-Fc, где указанный вариант содержит не более 2, 3, 4, 5, 10, 20, 30 или 50 замен, добавлений или делеций аминокислот в любой аминокислотной последовательности из SEQ ID NO: 26-41, и предпочтительно, указанный вариант сохраняет способность ингибировать ангиогенез. В одном из вариантов осуществления замена, добавление или делеция находится в части, полученной из внеклеточного домена VEGFR. В другом варианте осуществления замена, добавление или делеция находится в части, полученной из внеклеточного домена FGFR. В другом варианте осуществления замена, добавление или делеция находится в части, полученной из Fc-области иммуноглобулина. В другом варианте осуществления замена, добавление или делеция находится в линкере или связывающей части.
Дополнительно к встречающимся в природе модификациям в части, полученной из внеклеточного домена FGFR, и части, полученной из Fc-области иммуноглобулина, слитый белок VEGFR-FGFR-Fc также может содержать другие пост-трансляционные модификации. Такие модификации включают, без ограничений, ацетилирование, карбоксилирование, гликозилирование, фосфорилирование, эстерификацию и ацилирование. В результате, модифицированный слитый белок VEGFR-FGFR-Fc может содержать неаминокислотный компонент, например, полиэтиленгликоль, липид, полисахарид или моносахарид и фосфорную кислоту. Влияние таких неаминокислотных компонентов на функцию слитого белка VEGFR-FGFR-Fc может быть протестировано согласно тому, как описано здесь для других вариантов слитого белка VEGFR-FGFR-Fc. Если слитый белок VEGFR-FGFR-Fc продуцируется в клетке, посттрансляционный процессинг также может оказаться важным для корректировки укладки и/или функции белка. В разных клетках (например, СНО, HeLa, MDCK, 293, WI38, NIH-3T3 или НЕК293) существуют специальные клеточные машины и уникальные механизмы для такого пост-трансляционного процессинга, и для гарантированного осуществления модификации и процессинга слитого белка VEGFR-FGFR-Fc могут быть выбраны разные клетки.
Используемый здесь слитый белок может быть получен любым методом, известным в данной области техники. Например, он может быть получен методом химического синтеза или в результате экспрессии нуклеиновой кислоты. Используемые в настоящем описании пептиды могут быть легко получены стандартным методом жидкофазного или предпочтительно твердофазного пептидного синтеза, известным в данной области (см., например, J. М. Stewart and J. D. Young, Solid Phase Peptide Synthesis, 2nd edition. Pierce Chemical Company, Rockford, Illinois (1984), in М. Bodanzsky, and A. Bodanzsky, The Practice of Peptide Synthesis, Springer Verlag, New York (1984)). Слитый белок может быть продуцирован способами, известными в данной области, при этом между цистеиновыми остатками, находящимися в полипептидной последовательности, которая предположительно содержится в белке, может быть образована одна или более межмолекулярных поперечных связей (см., например патент США №5478925). Кроме того, в описанном здесь белке могут быть выполнены обычные модификации путем добавления цистеина или биотина к С-концу или N-концу этого белка.
Используемые здесь термины "терапевтически эффективное количество" или "эффективное количество" относится к дозам, которые являются достаточными для демонстрации положительного эффекта у субъекта, которому введено такое количество. Фактически вводимая доза, скорость и продолжительность курса введения зависят от состояния пациента и тяжести заболевания. Наконец, врач несет ответственность за назначение лечения (например, решение относительно дозировки т.д.) и принятие решения о лечении, как правило исходя из предназначенного для лечения заболевания, состояния пациента, места доставки, способа введения и других известных врачу факторов.
В одном из аспектов настоящего изобретения предоставляется слитый белок, способный ингибировать ангиогенез, который содержит ингибирующие ангиогенез структурные единицы, полученные из по меньшей мере двух ингибиторов ангиогенеза. Предпочтительно, по меньшей мере два ингибитора ангиогенеза представляют собой ингибирующий ангиогенез рецептор, при этом указанный рецептор может быть выбран, например, из группы, состоящей из: DDR1, EphA1, EphA2, EphA8, EphB1, EphB4, EGFR, HER-2, ЕrbВ3, FGFR1, FGFR2, FGFR4, MET, RON, CSF1R, KIT, PDGFR-A, PDGFR-B, ТЕК, Tie-1, HGF. VEGFR1, VEGFR2, VEGFR3, Notch рецептора, LK8, ангиостатина, эндостатина, плазминогена, коллагена XVIII и их аллельных вариантов. В одном из конкретных вариантов указанный ингибитор ангиогенеза представляет собой ингибирующие ангиогенез рецепторы VEGFR и FGFR.
В частности, авторами изобретения была сконструирована серия слитых белков VEGFR-FGFR-Fc, которые могут связываться с VEGF и FGF с высокой аффинностью и эффективно подавлять клеточное деление, индуцируемое VEGF и FGF.
В некоторых вариантах осуществления авторы изобретения с неожиданностью обнаружили, что слитый белок, который содержит последовательно от N-конца к С-концу часть, полученную из внеклеточного домена VEGFR, и часть, полученную из внеклеточного домена FGFR, обладает превосходной способностью связываться с VEGF и FGF, при этом указанная часть, полученная из внеклеточного домена VEGFR, предпочтительно содержит первый Ig-подобный домен VEGFR1, второй Ig-подобный домен VEGFR1 и третий Ig-подобный домен VEGFR2; более предпочтительно, указанная часть, полученная из внеклеточного домена FGFR, содержит часть, полученную из участка промежуточной функциональной последовательности Ig-подобного домена FGFR1, второй Ig-подобный домен FGFR1 и третий Ig-подобный домен FGFR1. Более предпочтительно, часть, полученная из участка промежуточной функциональной последовательности Ig-подобного домена FGFR1, не содержит кислый бокс, и предпочтительно она имеет аминокислотную последовательность, соответствующую положениям 134-162, 145-162 или 151-162 SEQ ID NO:1.
В некоторых вариантах осуществления настоящее изобретение включает применение (i) слитого белка VEGFR-FGFR-Fc, или (ii) полинуклеотида, кодирующего такой слитый белок, при изготовлении композиций или лекарственных средств для лечения заболеваний, опосредованных или связанных с ангиогенезом. Например, в одном из вариантов осуществления настоящее изобретение включает применение (i) слитого белка VEGFR-FGFR-Fc, или (ii) полинуклеотида, кодирующего такой слитый белок, при изготовлении лекарственных средств в качестве ингибитора ангиогенеза.
В некоторых вариантах осуществления слитый белок VEGFR-FGFR-Fc по настоящему изобретению может быть продуцирован в клеточной линии млекопитающих путем экспрессии любой нуклеиновой кислоты из SEQ ID NOs: 26-41. В частности, клеточная линия млекопитающих может представлять собой клеточную линию СНО.
Кроме того, в настоящем изобретении также предоставляется описанный ниже слитый белок VEGFR-FGFR-Fc, в котором часть, полученная из внеклеточного домена VEGFR, часть, полученная из внеклеточного домена FGFR, и часть, полученная из Fc-области иммуноглобулина, связаны или не связаны линкером.
В некоторых вариантах осуществления настоящее изобретение включает изолированные молекулы нуклеиновых кислот, кодирующие слитый белок VEGFR-FGFR-Fc. Настоящее изобретение также включает применение этих молекул в производстве лекарственных средств. Нуклеиновая кислота может быть рекомбинантной, синтетической или может быть получена любыми способами, известными в уровне техники, а способ включает клонирование стандартными методами.
В некоторых вариантах осуществления настоящее изобретение включает вектор, содержащий изолированную молекулу нуклеиновой кислоты по настоящему изобретению. Такой вектор может представлять собой экспрессирующий вектор, в котором нуклеиновая кислота эффективно связана с регуляторной последовательностью, способной экспрессировать нуклеиновую кислоту в клетке хозяине. Может быть использовано множество векторов. Например, подходящие векторы могут включать вирус (например, поксвирус, аденовирус, бакуловирус т.п.); дрожжевой вектор, бактериофаг, хромосому, искусственную хромосому, плазмиду, космид.
В некоторых вариантах осуществления настоящее изобретение также включает клетки, трансфицированные этими векторами для экспрессии слитого белка VEGFR-FGFR-Fc. Клетка хозяин, подходящая для настоящего изобретения, может быть прокариотической клеткой или эукариотической клеткой. Эти клетки включают бактерии, например, E.coli, дрожжи, клетку насекомых или клетку млекопитающих. Клеточные линии млекопитающих, которые могут быть использованы, включают, без ограничения, овариальные клетки китайского хомячка (СНО), клетки почки новорожденного хомячка, клетки мышиной миеломы NS0, клеточные линии обезьян и человека, и полученные от них клеточные линии т.п.
В другом аспекте настоящего изобретения предоставляется способ ингибирования ангиогенеза, содержащий введение слитого белка VEGFR-FGFR-Fc по настоящему изобретению нуждающемуся в этом субъекту. Предпочтительно, способ применяется в отношении млекопитающих.
В другом аспекте настоящего изобретения предоставляется способ лечения и профилактики опухолей у млекопитающих, содержащий введение слитого белка VEGFR-FGFR-Fc по настоящему изобретению нуждающемуся в этом субъекту, и предпочтительно, опухоль представляет собой солидную опухоль.
В другом аспекте настоящего изобретения предоставляется способ лечения и профилактики заболеваний, связанных с офтальмологическим ангиогенезом, у млекопитающих, содержащий введение слитого белка VEGFR-FGFR-Fc по настоящему изобретению нуждающемуся в этом субъекту, и предпочтительно, заболевания, связанные с офтальмологическим ангиогенезом, представляют собой возрастную дегенерацию желтого пятна и диабетическую ретинопатию.
Настоящее изобретение также относится к применению слитого белка VEGFR-FGFR-Fc в производстве лекарственных средств для ингибирования ангиогенеза. Кроме того, настоящее изобретение также относится к применению слитого белка VEGFR-FGFR-Fc в производстве лекарственных средств для лечения или профилактики связанных с ангиогенезом заболеваний, при этом связанные с ангиогенезом заболевания предпочтительно представляют собой опухоли или заболевания, связанные с офтальмологическим ангиогенезом.
Описанные в настоящем изобретении заболевания, связанные с ангиогенезом, включают, без ограничения, зависимые от ангиогенеза виды рака, включающие, например, твердую опухоль, гематогенную опухоль (например, лейкемию) и метастаз опухоли; доброкачественную опухоль, например, ангиому, нейрому слухового нерва, нейрофиброму, трахому и пиогенную гранулему; ревматоидный артрит; псориаз; рубеоз радужной оболочки; синдром Рандю-Ослера; миокардиальный ангиогенез; неоваскуляризацию бляшки; телеангиэктазию; гемофилический гемартроз и ангиофиброму.
В некоторых вариантах осуществления описанных способов один или более слитых белков VEGFR-FGFR-Fc могут вводиться совместно (одновременно) или в разное время (последовательно). Кроме того, слитый белок может вводиться совместно с дополнительным лекарственным средством, используемым для лечения рака или ингибирования ангиогенеза.
В некоторых вариантах осуществления может использоваться только способ, описанный в настоящем изобретении. В качестве альтернативы, способ по изобретению может использоваться в комбинации с другими традиционными видами противоопухолевой терапии для лечения или профилактики пролиферативных заболеваний (например, опухоли). Например, эти способы могут применяться для лечения рака, профилактики рецидива рака и послеоперационного метастаза, и могут быть использованы в качестве дополнительного средства для терапии других видов рака. Согласно тому как описано в настоящем изобретении, эффективность и традиционные методы терапии рака (например, химиотерапия, радиотерапия, фототерапия, иммунотерапия операция) могут быть улучшены благодаря использованию целевых полипептидных терапевтических агентов.
В офтальмологии ангиогенез связан, например, с диабетической ретинопатией, ретинопатией недоношенных, возрастной дегенерацией желтого пятна, отторжением роговичного трансплантата, неоваскулярной глаукомой и RLF (ретролентальной фиброплазией). Раскрытый здесь слитый белок VEGFR-FGFR-Fc можно вводить непосредственно внутрь глаза или другими способами. Другие офтальмологические заболевания, связанные с ангиогенезом, включают, но не ограничены, эпидемический кератоконъюктивит, дефицит витамина А, чрезмерное ношение контактных линз, атопический кератит, кератоконъюктивит, птеригиум синдром сухого глаза, болезнь Шегрена, красные угри, фликтена, сифилис, микобактериальную инфекцию, липидную дегенерацию, химический ожог, бактериальную язву, грибковую язву, инфекцию, вызванную вирусом простого герпеса (Негрез simplex), инфекцию, вызванную вирусом опоясывающего герпеса (Негрез zoster), протозойную инфекцию, саркому Калоши, разъедающую язву роговицы, краевую дегенерацию Террьена, краевой кератит, ревматоидный артрит, системную волчанку, полиартрит, травму, саркоидоз Вегенера, склерит, болезнь Стивенса-Джонса, пемфигоидную радиальную кератотомию и отторжение роговичного трансплантата, серповидно-клеточную анемию, саркоид, эластическую псевдоксантому, болезнь Педжета, венозную окклюзию, артериальную окклюзию, каротидное обструктивное заболевание, хронический увеит/витрит, микобактериальную инфекцию, болезнь Лайма, системную красную волчанку, ретролетальную фиброплазию, болезнь Илза, болезнь Бехчета, вызываемые ретинитом или хлороидитом инфекции, предполагаемый глазной гистоплазмоз, болезнь Беста, миопию, врожденные ямки в диске зрительного нерва, болезнь Старгардта, парспланит, хроническое отслоение сетчатки, синдром повышенной вязкости, токсоплазмоз, травматические или постлазерные осложнения. Другие заболевания включают, без ограничения, заболевания, связанные с рубеозом радужки (неоваскуляризацию угла передней камеры), и заболевания, индуцируемые аномальной гиперплазией фиброзного кровеносного сосуда или фиброзной ткани, в том числе все виды пролиферативной витреоретинопатии.
Введение
Слитый белок по настоящему изобретению можно вводить отдельно, но предпочтительно, в виде фармацевтической композиции, которая обычно содержит подходящий фармацевтический наполнитель, разбавитель или носитель, выбираемый в зависимости от используемого способа введения. Слитый белок можно вводить нуждающемуся в этом пациенту любым подходящим способом. Точная дозировка зависит от многих факторов, в том числе и от четко определенных свойств слитого белка.
Некоторые подходящие способы введения включают (без ограничения) пероральное, ректальное, назальное, топическое (включая буккальное и сублингвальное), подкожное, вагинальное или парентеральное (включая подкожное, внутримышечное, внутривенное, внутрикожное, интратекальное и экстрадуральное) введение.
Для внутривенной инъекции и инъекции в место повреждения активные ингредиенты находятся в форме парентерально доступного водного раствора, который не содержит пирогена и имеет приемлемое значение рН, изотоничность и стабильность.
Специалистом в данной области слитый белок может быть составлен в виде лекарственной формы вместе с подходящим растворителем, или, например, изотоническими наполнителями, такими как раствор хлорида натрия для инъекций, раствор Рингера для инъекций, раствор Рингера лактата для инъекций. При необходимости может быть добавлен консервант, стабилизатор, буферный агент, антиоксидант и/или некоторые другие добавки.
Вводимая перорально фармацевтическая композиция может быть в форме таблетки, капсулы, порошка или перорально вводимой жидкости и т.п. Таблетка может содержать твердый носитель, такой как желатин или адъювант. Жидкая фармацевтическая композиция обычно содержит жидкий носитель, такой как вода, вазелин, животное или растительное масло, минеральное масло или синтетическое масло. Композиция также может содержать обычный физраствор, раствор глюкозы или других Сахаров или гликолей, таких как этиленгликоль, пропиленгликоль или полиэтиленгликоль.
Примеры описанных выше методов и схем и другие методы и схемы, используемые согласно настоящему изобретению, можно найти в справочнике Ремингтона "Фармацевтические науки" (Pharmaceutical Sciences), 16-oe издание, Oslo, А. (ред.), 1980.
Клонирование слитого белка и конструкции экспрессионной плаэмиды
Фрагмент рецептора VEGF и рецептора FGF получают амплификацией кДНК-матрицы соответствующего рецептора методом ПЦР, а Fc-фрагмент IgG1 получают амплификацией кДНК человеческого IgG1 методом ПЦР.При разработке праймеров для ПЦР связывающие последовательности вводят между разными фрагментами таким образом, чтобы эти разные фрагменты были в результате связаны методом перекрывающегося ПЦР для образования рамок считывания разных слитых белков, и в оба конца кДНК вводят дополнительные сайты рестрикции для эндонуклеаз BspE I и Pst I. кДНК для разных слитых белков могут быть клонированы в экспрессионной плазмиде после расщепления эндонуклеазами BspE I и Pst I. После клонирования эту плазмиду можно определить расщеплением эндонуклеазами, электрофорезом и наконец секвенированием ДНК.
В настоящем изобретении конструируют рекомбинантные экспрессионные плазмиды 2#, 4#, 7#, 9#, 10#, 11#, 12#, 14#, 15#, 16#, 20#, 21#, 24#, 25#, 27# и 28#.
Слитые белки 2#, 4# и 7# содержат часть внеклеточного домена VEGFR1, часть внеклеточного домена FGFR1 и Fc IgGi;
слитые белки 9#, 10#, 16#, 20#, 21#, 24#, 25# и 28# содержат часть внеклеточного домена VEGFR1, часть внеклеточного домена VEGFR2, часть внеклеточного домена FGFR1 и IgG1 Fc, причем для указанных выше 11 рекомбинантных экспрессионных плазмид используется один и тот же обратный праймер внеклеточного домена FGFR1 (2#-FGFR1 обратный праймер: GTTTTGTCCTCCAGGTACAGGGGCGAGGTC), прямой праймер для Fc IgG1 (CTGTACCTGGAGGACAAAACTCACACATGC) и обратный праймер для Fc IgG1 (GATATCTGCAGTCATTTACCCGGAGACAGG).
Ниже перечислены праймеры для остальных слитых белков.
2# слитый белок:
2# - VEGFR1 прямой праймер:
ATAGTTCCGGAGGTAGACCATTCGTAGAGATG,
2# - VEGFR1 обратный праймер:
CCTGTGATGCGGGTGCGATTTTTTTCATCAGGGTAACTCC,
2# - FGFR1 прямой праймер: CTGATGAAAAAAATCGCACCCGCATCACAG.
4# слитый белок:
4# VEGFR1 прямой праймер является таким же как для 2#: 2# - VEGFR1 прямой праймер,
4# - VEGFR1 обратный праймер: TTTTTCATCAGGGTAACTCCAGGTCATTTG,
4# FGFR1 прямой праймер:
GGAGTTACCCTGATGAAAAACCAGAAAAGATGGAAAAGAAAT,
4# FGFR1 обратный праймер является таким же как для 2#: 2# - FGFR1 обратный праймер.
7# слитый белок:
7# VEGFR1 прямой праймер является таким же как для 2#: 2# - VEGFR1 прямой праймер,
7# VEGFR1 обратный праймер:
ACCGCCAGAGCCACCTCCGCCTGAACCGCCACCACCTTTTTCATCAGGGTAACTCCAG,
7# FGFR1 прямой праймер:
AGGCGGAGGTGGCTCTGGCGGTGGCGGATCCCCAGAAAAGATGGAAAAGAAATTG,
7# FGFR1 обратный праймер является таким же как для 2#: 2# - FGFR1 обратный праймер.
9# слитый белок: 9# слитый белок содержит 5 сегментов;
VEGFR1D1, VEGFR1D2, VEGFR2D3, FGFR и fc.
VEGFR1D1 амплифицируют, используя pBLAST45-hFLT1s7cDNA и следующие праимеры:
9# - VEGFR1D1 прямой праймер:
TAGTTCCGGAAGCAAATTAAAAGATCCTGAACTGAG,
9# - VEGFR1D1 обратный праймер:
ATCTCTACGAAAGGTCTACCTGTATCACTAATAAATATATAG.
VEGFR1D2 амплифицируют, используя pBLAST45-hFLT1s7cDNA и
следующие праимеры:
9# - VEGFR1D2R2D3 прямой праймер: GGTAGACCTTTCGTAGAGATGT,
9# - VEGFR1D2 обратный праймер:
CATGAGACGGACTCAGAACCACATCTATGATTGTATTGGTTTG.
VEGFR2D3 амплифицируют, используя в качестве матрицы pBLAST45-hFLK1s7 и следующие праймеры:
9# - VEGFR2D3 прямой праймер:
CAAACCAATACAATCATAGATGTGGTTCTGAGTCCGTCTCATG,
9# - VEGFRlD2R2D3 обратный праймер: AGGTTTTTCATGGACCCTGAC,
9# FGFR1 прямой праймер:
TCAGGGTCCATGAAAAACCTCCAGAAAAGATGGAAAAGAAATTGC.
10# слитый белок:
праймеры для ПЦР для последовательностей доменов фрагмента 10#VEGFR1 являются такими же как для 9#,
10# - FGFR1 прямой праймер:
TCAGGGTCCATGAAAAACCTAAAAATCGCACCCGCATCACAGG.
16# слитый белок:
16# VEGFR1D1 прямой праймер является таким же как для 9# - VEGFR1D1 прямой праймер,
16# VR1D2R2D3R обратный праймер является таким же как для 9# - VRlD2R2D3R обратный праймер,
16# FGFR1 прямой праймер:
GTCAGGGTCCATGAAAAACCTAGGCCGTCCCCGACCTTGCCTG.
20# слитый белок:
20# - VEGFR1D1 прямой праймер является таким же как для 9# - VEGFR1D1 прямой праймер,
20# - VR1D2R2D3R обратный праймер является таким же как для 9# - VR1D2R2D3R обратный праймер,
20# - FGFR1 прямой праймер:
CCTGTGATGCGGGTGCGATTAGGTTTTTCATGGACCCTGAC.
21# слитый белок:
10# используется в качестве матрицы для 21#, при этом для замены одного основания в 10#FGFR1D1h таким образом, чтобы Суз был заменен Ser, используются следующие праймеры:
21# - mutF: CTCCGGCCTCTATGCTTCCGTAACCAGCAGCCCCTC,
21# - mutR: GAGGGGCTGCTGGTTACGGAAGCATAGAGGCCGGAG.
24# слитый белок:
24# VEGFR1D1 прямой праймер является таким же как для 9# - VEGFR1D1 прямой праймер,
24# VR1D2R2D3R обратный праймер является таким же как для 9# - VR1D2R2D3R обратный праймер,
24# FGFR1 прямой праймер:
TCAGGGTCCATGAAAAACCTTCGGGCAGTGACACCACCTAC.
25# слитый белок:
25# VEGFR1D1 прямой праймер является таким же как для 9# - EGFR1D1 прямой праймер,
25# VR1D2R2D3R обратный праймер является таким же как для 9# - VR1D2R2D3R обратный праймер,
25# FGFR1 прямой праймер:
TCAGGGTCCATGAAAAACCTAACCCCGTAGCTCCATATTGG.
28# слитый белок:
ПЦР для 28# выполняют, используя в качестве матрицы 25#, при этом прямой праймер является таким же как для 2#-VEGFR1, a обратный праймер является таким же как для Fc IgG1.
Слитые белки 11#, 14# и 27# содержат часть внеклеточного домена FGFR1, часть внеклеточного домена VEGFR1, часть внеклеточного домена VEGFR2 и Fc IgG1. Для указанных выше рекомбинантных экспрессионных плазмид используется прямой праймер, который является таким же как для Fc IgG1, а обратный праймер является таким же как для Fc IgG1.
Ниже приведены остальные праймеры:
11# слитый белок:
11#-FGFR1 прямой праймер:
CTAGCTCCGGACCAGAAAAGATGGAAAAGAAATTGC,
11#-FGFR1 обратный праймер:
TCAGGATCTTTTAATTTTGACTCCAGGTACAGGGGCGAGGTC,
11#-VEGFR1D1 прямой праймер: TCAAAATTAAAAGATCCTGAACTG,
11#-VEGFR1D1 обратный праймер является таким же как для 9#-VEGFR1D1 обратный праймер,
11#VEGFR1D2R2D3 прямой праймер является таким же как для 9#-VR1D2R2D3 прямой праймер,
11#VEGFR1D2D3 обратный праймер:
TGGGCATGTGTGAGTTTTGTCAGGTTTTTCATGGACCCTGAC.
14# слитый белок:
14#-FGFR1 прямой праймер: TAGTTCCGGAAAAAATCGCACCCGCATCACAG,
14# FGFR1 обратный праймер является таким же как для 11#-VEGFR1D1 обратный праймер,
14# VEGFR1D1 прямой праймер является таким же как для 11#-VEGFR1D1 прямой праймер,
14# VEGFR1D1 обратный праймер является таким же как для 9#-VEGFR1D1 обратный праймер,
14#VEGFR1D2R2D3 прямой праймер является таким же как для 9#-VEGFR1D2R2D3 прямой праймер,
14# VEGFR1D2R2D3 обратный праймер является таким же как для 11#-VEGFR1D2R2D3 обратный праймер.
27# слитый белок:
ПЦР для 27# выполняют, используя в качестве матрицы 14#, при этом прямой праймер является таким же как для 27#-FGFR1 (TAGTTCCGGAAAACCTAACCCCGTAGCTCCAT), а обратный праймер является таким же как для Fc IgG1.
12# слитый белок содержит часть внеклеточного домена VEGFR1, часть внеклеточного домена VEGFR2 и Fc IgG1. Используются следующие праймеры:
прямой праймер для 12# является таким же как для 9#-FGFR1 прямой праймер,
12# VEGFR2 обратный праймер является таким же как для 11#-VEGFR1D2D3 обратный праймер.
Прямой праймер 1 для Fc: FcFor1: GACAAAACTCACACATGCCCACC.
Обратный праймер для Fc является таким как указанный выше обратный праймер для Fc IgG1.
Слитый белок 15# содержит часть внеклеточного домена VEGFR1 и Fc IgG1. Используются следующие праймеры:
прямой праймер для 15# является таким же как для 2#-VEGFR1 прямой праймер: ATAGTTCCGGAGGTAGACCATTCGTAGAGATG,
обратный праймер 15#VEGFR2 является таким же как 11#VEGFR1D2D3 обратный праймер.
Прямой праймер 1 для Fc 1: FcFor1: GACAAAACTCACACATGCCCACC
Обратный праймер для Fc является таким же как указанный выше обратный праймер для Fc IgG1.
Экспрессия и очистка слитого белка
Слитый белок по изобретению может быть экспрессирован и очищен методами, обычно используемыми в данной области. ДНК соответствующей плазмиды слитого белка очищают, используя набор для очистки плазмид (МАХ), предоставляемый Qiagen, a концентрацию плазмидной ДНК определяют методом УФ-спектрофотомерии, клетки СНО трансфицируют плазмидой, используя липосому FUGENE 6 (Roche). Конкретный способ трансфекции осуществляют согласно спецификации продукта. Исходя из требуемого количества экспрессируемого белка, в настоящем описании использовали два способа экспрессии белка: (1) транзиторную экспрессию, в которой слитый белок, содержащийся в супернатанте культуры, обычно собирают через 48-72 часов после трансфекции, а затем определяют относительное содержание слитого белка ELISA анализом (человеческий IgG) для быстрого и эффективного получения слитого белка; (2) метод, включающий определение стабильной клеточной линии и создание обычной экспрессионной системы в DHFR-дефектных клетках СНО для экспрессии рекомбинантного белка для получения лекарственного средства, где основной способ содержит трансфекцию клетки, выбор стабильно трансфицированной клетки, скрининг клона, амплификацию в стрессовых условиях, культуральную среду и процесс оптимизации т.д., и наконец получение массовой суспензионной культуры созданного штамма клеточной линии СНО в культуральной среде, не содержащей сыворотку. Продукт культуры собирают, а слитый белок очищают методом аффинной хроматографии в колонке с белком А. Очищенный белок анализируют методом электрофореза в додецилсульфат полиакриламидном геле (SDS-PAGE), затем все элюаты, содержащие требуемый продукт экспрессии, объединяют, фильтруют, используя 0,22 мкм фильтр, затем определяют количество белка любым из многочисленных способов, например, анализом белка методом Лоури. В настоящем изобретении объем клеточной культуры СНО соответствовал объему 10 л биореактора, что позволяет получить после очистки объем слитого белка, требуемый для проведения экспериментов на животных, а также служит в качестве основы для будущего увеличения масштабов производства.
Оценка нейтрализации VEGF и FGF на уровне белка
После получения слитого белка, экспрессированного клетками СНО, в настоящем изобретении проводится оценка способности слитого белка связываться с VEGF и FGF на уровне белка. Для оценки связывающей способности и аффинности проводили эксперименты, в которых оценку связывающей способности выполняли следующим образом: после покрытия 96-луночного ELISA планшета VEGF и FGF-2, покрытую лунку блокировали BSA, после чего добавляли каждый из слитых белков с одинаковыми концентрациями; после промывки к человеческому Fc-HRP IgG добавляли вторичное антитело, образцы проявляли, реакцию останавливали, с ELISA планшета считывали данные при 450 нм, и на завершающем этапе по силе сигнала отбирали слитые белки, который обладали способностью связываться с VEGF и FGF-2. Эксперимент по оценке аффинности проводили для определения аффинности слитого белка к VEGF или FGF-2 в системе раствора, который заключался в следующем: сначала 96-луночный ELISA планшет покрывали VEGF или FGF-2 для захвата антител, затем покрытую лунку блокировали BSA, и, пользуясь методом последовательных разведении, добавляли заранее приготовленную и инкубированную смесь слитого белка и VEGF или FGF-2, и после инкубации для детектирования добавляли HRP-меченное антитело (использовали антитело 2, которое используется для специфического детектирования свободного VEGF или FGF-2), после чего образцы проявляли, реакцию останавливали, с ELISA планшета считывали данные при 450 нм, и на завершающем этапе определяли относительную концентрацию свободного VEGF или FGF-2 в смеси слитого белка и VEGF или FGF-2. В результате описанного выше эксперимента был отобран слитый белок, обладающий эффектом двойного блокирования: VEGF и FGF-2.
Оценка нейтрализации VEGF и FGF слитым белком на клеточном уровне
После оценки способности слитого белка связываться с VEGF и FGF-2, проведенной на уровне белка, в настоящем изобретении проводили проверку наличия эффекта ингибирования ангиогенеза на клеточном уровне. Ингибирующую способность слитого белка в отношении деления и миграции сосудистых эндотелиоцитов оценивали в тестах на деление и миграцию клеток, используя человеческие эндотелиальные клетки пупочной вены (HUVEC). Ингибирующая способность слитого белка в отношении деления клеток HUVEC может быть оценена при помощи теста на деление клеток HUVEC, который содержит следующий этапы: 3000 HUVEC клеток на лунку инкубировали в 96-луночном планшете и культивировали при 37°С в 5% СO2 инкубаторе, затем добавляли VEGF или FGF-2, а также смесь слитого белка с разной концентрацией с VEGF или FGF-2, соответственно, и после культивирования в течение 3-4 дней добавляли 10% ССК-8 и культивировали еще 2 часа перед считыванием данных с ELISA планшета при 450 нм. Способность слитого белка ингибировать деление сосудистых эндотелиоцитов, индуцированное VEGF или FGF-2, оценивали по разнице в уровнях поглощения, определяли среднюю эффективную концентрацию слитого белка для ингибирования VEGF или FGF-2. Способность слитого белка ингибировать миграцию клеток HUVEC оценивали при помощи теста на миграцию клеток HUVEC, который содержит следующие этапы: 50000 HUVEC клеток на лунку, а также слитый белок с разными концентрациями сначала инкубировали в верхней камере, при этом в нижнюю камеру добавляли культуральную жидкость, содержащую 600 мкл VEGF или FGF-2, затем образец культивировали при 37°С в 5% СO2 инкубаторе в течение 20-24 часов, после чего удаляли клетки, находящиеся на лицевой стороне мембраны верхней камеры, затем клетки, расположенные на задней стороне мембраны фиксировали, окрашивали и промывали PBS перед осмотром и подсчетом клеток под инвертированным микроскопом. Миграцию клеток HUVEC, индуцированную стимуляцией VEGF или FGF-2, определяли подсчетом клеток HUVEC, находящихся на задней стороне мембраны, а способность слитого белка ингибировать миграцию сосудистых эндотелиоцитов тестировали путем добавления в культуральную жидкость слитого белка с разными концентрациями. Описанные выше эксперименты подтвердили способность нового слитого белка, разработанного в настоящем изобретении, ингибировать деление и миграцию сосудистых эндотелиоцитов, индуцированные VEGF и FGF-2, что также послужило основанием для проведения экспериментов на животных.
Оценка способности слитого белка ингибировать рост опухоли с использованием модели опухоли
После того, как в проведенных в настоящем изобретении экспериментах на уровне белка и клетки было показано, что новый слитый белок обладает способностью блокировать сигналы VEGF и FGF-2, была протестирована противоопухолевая активность заявленного слитого белка на моделях опухолей животных. В настоящем изобретении антиангиогенная активность и противоопухолевая активность слитого белка проверялась на моделях, которые обычно используются для подбора лекарственного средства для лечения ангиогенеза и опухоли, например, мышиная модель рака легких LLC, глиоцитома U87, меланома В16 т.д. В экспериментах на животных дополнительно к традиционным контрольным группам, также были включены контрольные лекарственные средства, такие как VEGF-Trap и FP-1039, для получения сравнительных данных в отношении противоопухолевой активности. В ходе эксперимента мышам С57 на одной стороне в области спины вводили подкожно 100 мкл жидкости, содержащей клетки опухоли в подходящем количестве, и дважды в неделю штангенциркулем с нониусом измеряли объем опухоли. Когда объем опухоли достиг примерно 200 мм3, подкожно инъецировали слитый белок с разными концентрациями, через 2-3 недели мышей умерщвляли. Затем, при помощи штангенциркуля с нониусом измеряли объем опухоли, противоопухолевую активность слитого белка оценивали по размеру опухоли. Помимо этого, для изучения механизма регуляции ангиогенеза отдельно анализировали опухолевую ткань такими методами как иммуногистохимия.
Примеры
Пример 1: Конструкция рекомбинантной экспрессионной плазмиды для двунаправленного слитого белка
Коммерчески доступную кДНК (готовую для синтеза методом ПЦР первую цепь кДНК, полученную из пораженной раком ткани прямой кишки взрослого человека, BioChain) использовали в качестве матрицы для фрагмента FGFR1. Коммерчески доступную плазмиду pBLAST45-hFLT1s7cDNA (InvivoGen) использовали в качестве матрицы для фрагмента VEGFR1, и коммерчески доступную плазмиду pBLAST45-hFLKls7 (InvivoGen) использовали в качестве матрицы для фрагмента VEGFR2. Общую РНК экстрагировали из крови здорового человека, используя набор для экстракции РНК из крови человека (QIAGEN). Согласно инструкции производителя для набора по проведению обратной транскрипции (Promega) выполняли ОТ-ПЦР с обратной транскриптазой M-MLV (Promega) для осуществления обратной транскрипции РНК в кДНК, которую затем использовали в качестве матрицы для Fc-фрагмента IgG1. ОТ-ПЦР выполняли согласно инструкции производителя для набора для проведения обратной транскрипции, которая состояла в следующем: Oligo dT, dNTP, общую РНК и DEPC НгО смешивали до получения гомогенной смеси, и проводили реакцию при 70°С в течение 10 мин, затем смесь помещали на лед на 5 мин, осле чего добавляли ингибитор RNase, обратную транскриптазу M-MLV и реакционный буфер. Температуру смеси доводили до 42°С в течение 1 часа, а затем до 70°С за 15 мин. Полученную кДНК использовали в качестве матрицы.
Отдельно методом ПЦР амплифицировали различные фрагменты FGFR1, используя в качестве матрицы кДНК, полученную из пораженной раком ткани прямой кишки взрослого человека (праймеры приведены в Таблице 1), а различные фрагменты VEGFR1 и VEGFR2 амплифицировали, используя в качестве матрицы коммерчески доступную плазмиду (праймеры приведены в Таблице 1), фрагмент Fc IgG1 амплифицировали методом ПЦР, используя в качестве матрицы кДНК, полученную из крови человека (праймеры приведены в Таблицах 1 и 2). При конструировании праймеров для ПЦР последовательности с 20 или более комплементарными основаниями вводили в качестве последовательностей, связывающих фрагменты VEGFR1, VEGFR2, FGFR1 и Fc IgG1, для получения фрагментов, связанных последовательно методом перекрывающегося ПЦР, с образованием рамок считывания для различных слитых белков, при этом в оба конца продуктов ПЦР вводили сайты рестрикции для эндонуклеаз BspE I и Pst I.
Затем методом перекрывающегося ПЦР амплифицировали каждый фрагмент слитого белка FGFR1-Fc. Условия реакции ПЦР были следующими: 5 минутная предварительная денатурация при 98°С, 30 циклов 30 секундной денатурации при 98°С, 45 секундный отжиг при 56°С и 2 минутное удлинение при 72°С и, наконец, еще одно 10 минутное удлинение. При конструировании праймеров для ПЦР между разными фрагментами вводили связывающую последовательность для связывания этих разных фрагментов методом перекрывающегося ПЦР. Процесс проведения реакции методом перекрывающегося ПЦР можно разделить на два этапа, при этом на первом этапе вводят фрагмент, необходимый для связывания и не содержащий праймер, условия проведения реакции являются следующими: 5 минутная предварительная денатурация при 98°С, 6 циклов 30 секундной денатурации при 98°С, 45 секундный отжиг при 56°С и 5 минутное удлинение при 72°С и, наконец, еще одно 10 минутное удлинение при 72°С; после первого этапа выполняют второй этап ПЦР, в котором на оба конца добавляют праймеры, условия проведения реакции являются следующими: 5 минутная предварительная денатурация при 98°С, 30 циклов 30 секундной денатурации при 98°С, 45 секундный отжиг при 56°С и 2 минутное удлинение при 72°С и, наконец, еще одно 10 минутное удлинение при 72°С; получая таким образом сплайсированные рамки считывания для разных слитых белков и одновременно вводя на оба конца кДНК сайты рестрикции для эндонуклеаз ВзрЕ I и Pst I.
После амплификации амплифицированные методом ПЦР фрагменты очищали, используя набор для очистки QIAquick PCR (QIAGEN). кДНК различных слитых белков и эукариотическую экспрессируемую плазмиду pSV2-dhfr (ATCC) расщепляли ВзрЕ I и Pst I, соответственно. Затем расщепленные образцы анализировали методом электрофореза в 1% агарозном геле, используя напряжение 90 В. Целевые фрагменты выделяли при помощи набора для гелевой экстракции QIAquick (QIAGEN) затем, используя лигазу (NEB), лигировали в течение 1 часа при 16°С. Смесь, используемую для реакции лигирования, трансформировали в соответствующую Тор10 Е.coli в условиях 90 сек реакции при 42°С, а затем в течении 3 мин выдерживали на льду. После добавления стерильного LB культурального бульона (без антител) смесь встряхивали при 250 об/мин на шейкере при 37°С в течение 1 часа перед нанесением на LB планшет, в который был добавлен ампициллин. Планшет культивировали в течение ночи в термостатируемом инкубаторе при 37°С, а затем отбирали единичные колонии и переносили в содержащий ампициллин LB культуральный бульон. Инокулированный культуральный бульон встряхивали при 250 об/мин на шейкере при 37°С в течение ночи, после чего выделяли плазмиду методом щелочного лизиса. Затем перед оценкой образца электрофорезом в 1% агарозном геле при 90 В его расщепляли рестрикционной эндонуклеазой. Рекомбинантную плазмиду, правильно расщепленную эндонуклеазами, подтверждали методом секвенирования ДНК. В результате осуществления описанных выше этапов были сконструированы рекомбинантные экспрессионные плазмиды 2#, 4#, 7#, 9#, 10#, 11#, 12#, 14#, 15#, 16#, 20#, 21#, 24#, 25#, 27# и 28#.
Среди других полученных белков, слитые белки 2#, 4# и 7# содержали часть внеклеточного домена VEGFR1, часть внеклеточного домена FGFR1 и Fc IgG1, где указанную часть внеклеточного домена VEGFR1 амплифицировали методом ПЦР, используя в качестве матрицы плазмиду pBLAST45-hFLT1s7cDNA, указанную часть внеклеточного домена FGFR1 амплифицировали методом ПЦР, используя в качестве матрицы кДНК, выделенную из пораженной раком ткани прямой кишки, а Fc-область человеческого IgG1 амплифицировали, используя в качестве матрицы кДНК, выделенную из крови человека. Затем методом перекрывающегося ПЦР получали продукт ПЦР с тремя связанными фрагментами, который вводили в экспрессирующий вектор после расщепления и лигирования. Использованные праймеры приведены в таблице 1.
9#, 10#, 16#, 20#, 21#, 24#, 25# и 28# содержат часть внеклеточного домена VEGFR1, часть внеклеточного домена VEGFR2, часть внеклеточного домена FGFR1 и Fc IgG1. Сначала была получена конструкция 9#, в которой фрагмент VEGFR1D1 и фрагмент VEGFR1D2 амплифицировали методом ПЦР, используя в качестве матрицы pBLAST45-hFLT1s7cDNA, фрагмент VEGFR2D3 амплифицировали методом ПЦР, используя в качестве матрицы pBLAST45-hFLK1s7, указанную часть внеклеточного домена FGFR1 амплифицировали методом ПЦР, используя в качестве матрицы кДНК пораженной раком ткани прямой кишки человека, и Fc-область человеческого IgG1 амплифицировали методом ПЦР, используя в качестве матрицы кДНК, полученную из крови человека. Затем методом перекрывающегося ПЦР получали продукт ПЦР с пятью связанными фрагментами, который вводили в экспрессирующий вектор после расщепления и лигирования. После амплификации фрагмента VEGFR методом ПЦР, в котором в качестве матрицы была использована конструкцию 9#, указанную часть внеклеточного домена FGFR1 амплифицировали методом ПЦР, используя в качестве матрицы кДНК из пораженной раком ткани прямой кишки, а Fc-область человеческого IgG1 амплифицировали методом ПЦР, используя в качестве матрицы кДНК, полученной из крови человека. Затем методом перекрывающегося ПЦР получали продукт ПЦР с тремя связанными фрагментами, который вводили в экспрессирующий вектор после расщепления и лигирования для получения конструкций 10#, 16#, 20#, 24# и 25#. После этого методом ПЦР, используя в качестве матрицы конструкцию 10# и примеры, приведенные в таблице 1, осуществляли мутацию одного основания в 10#FGFR1D1h. В частности, получали конструкцию 21# путем замены Cys на Ser, следуя инструкции производителя для набора сайт-направленного мутагенеза (QuikChangeTM Site-Directed Mutagenesis Kit, STRATAGEM), следующим образом: реакцию ПЦР проводили путем добавления в пробирку для ПЦР деионизованной воды, плазмиды 10#, dNTP, приведенных в таблице 1 праймеров для мутации, раствор реакционного буфера и фермент Pfu при следующих условиях: 2 минутная предварительная денатурация при 95°С, 18 циклов 50 секундной денатурации при 95°С, 50 секундный отжиг при 60°С и 8 минутное удлинение при 68°С и, наконец, еще одно 10 минутное удлинение. После реакции ПЦР добавляли 1 мкл Dpn I (NEB) в течение 1 часа при 37°С.2 мкл реакционного продукта отбирали для трансформации соответствующей Тор10 Е.coli в условиях 90 секундной реакция при 42°С, затем выдерживание на льду в течение 3 мин; добавляли стерильный LB культуральный буфер (без антител), затем культивировали в шейкере при 250 об/мин в течение 1 часа при 37°С перед нанесением на LB планшет с добавленным в него ампициллином; планшет культивировали в течение ночи в термостатируемом инкубаторе при 37°С, затем отбирали единичные колонии и переносили в содержащий ампициллин LB культуральный бульон; инокулированный культуральный бульон культивировали при 250 об/мин в шейкере при 37°С в течение ночи, затем выделяли плазмиду методом щелочного лизиса; после чего образец оценивали методом секвенирования ДНК. Конструкцию 28# получали методом ПЦР, используя в качестве матрицы конструкцию 25#, а также 2#-VEGFR1 прямой праймер и Fc IgG1 обратный праймер. Использованные праймеры перечислены в таблице 1.
Конструкции 11#, 14# и 27# содержат часть внеклеточного домена FGFR1, часть внеклеточного домена VEGFR1, часть внеклеточного домена VEGFR2 и Fc IgG1. Указанную часть внеклеточного домена FGFR1 амплифицировали методом ПЦР, используя в качестве матрицы кДНК, полученную из пораженной раком ткани прямой кишки, фрагмент VEGFR амплифицировали методом ПЦР, используя в качестве матрицы конструкцию 9#, и Fc-область человеческого IgG1 амплифицировали методом ПЦР, используя в качестве матрицы кДНК, полученную из крови человека. Затем методом перекрывающегося ПЦР получали продукт ПЦР с тремя слитыми фрагментами, который вводили в экспрессирующий вектор после расщепления и лигирования, получая таким образом конструкции 11# и 14#. Затем получали конструкцию 27# методом ПЦР, используя в качестве матрицы конструкцию 14#. Использованные праймеры приведены в таблице 1.
Слитый белок 12# содержит часть внеклеточного домена VEGFR1, часть внеклеточного домена VEGFR2 и Fc IgG1. Используя в качестве матрицы конструкцию 9#, Fc-область человеческого IgG1 амплифицировали с 9#-VEGFR1D1 прямым праймером и 11#VEGFR1D2D3 обратным праймером, где используемую в качестве матрицы кДНК получали из крови человека. После этого методом перекрывающегося ПЦР получали продукт ПЦР с двумя связанными фрагментами, который вводили в экспрессирующий вектор после расщепления и лигирования, получая таким образом конструкцию 12#.
Слитый белок 15# содержит часть внеклеточного домена VEGFR1, часть внеклеточного домена VEGFR2 и Fc IgG1. Используя в качестве матрицы конструкцию 9#, Fc-область человеческого IgG1 амплифицировали с 2#-VEGFR1 прямым праймером и 11#VEGFR1D2D3 обратным праймером, где используемую в качестве матрицы кДНК получали из крови человека. После этого методом перекрывающегося ПЦР получали продукт ПЦР с двумя связанными фрагментами, который вводили в экспрессирующий вектор после расщепления и лигирования, получая таким образом конструкцию 15#.
Слитый белок 26# содержит часть внеклеточного домена FGFR1 и Fc IgG1. Указанную часть внеклеточного домена FGFR1 амплифицировали методом ПЦР, используя в качестве матрицы кДНК, полученную из пораженной раком ткани прямой кишки, и Fc-область человеческого IgG1 амплифицировали с прямым праймером с последовательностью SEQ ID NO: 73 и обратным праймером с последовательностью SEQ ID NO: 74, используя в качестве матрицы кДНК, полученную из крови человека. После этого методом перекрывающегося ПЦР получали продукт ПЦР с двумя связанными фрагментами, который вводили в экспрессирующий вектор после расщепления и лигирования, получая таким образом конструкцию 26#.
Последовательность белка VEGFR-FGFR-Fc для каждого слитого белка и кодирующие нуклеотидные последовательности приведены в таблице 2. Структурная схема слитого белка показана на Фиг. 1.
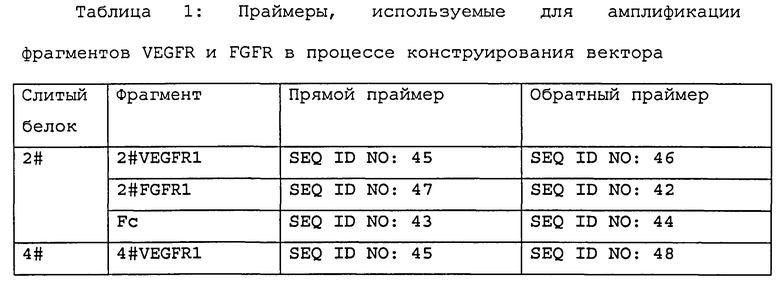
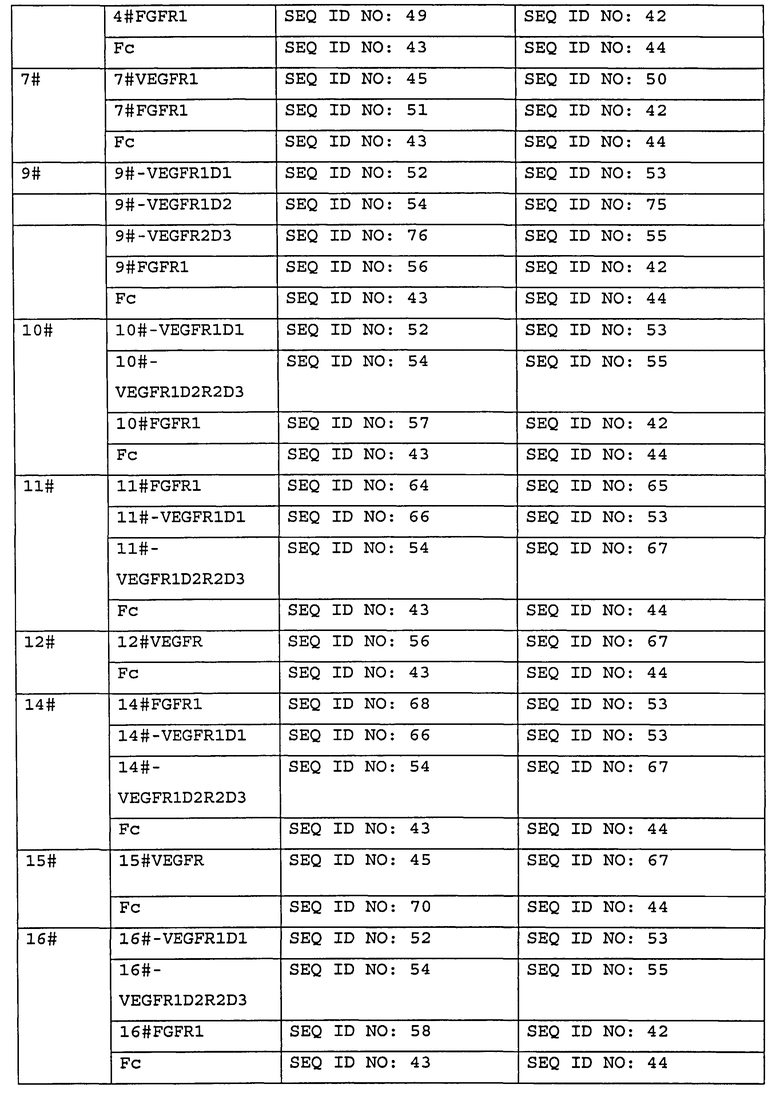

Пример 2: Транзитная экспрессия и количественное определение слитых белков
ДНК каждой плазмиды, используемой для получения слитого белка, очищали при помощи набора МАХ для очистки плазмид (Qiagen). Концентрацию ДНК плазмиды определяли УФ-спектрометрией. 1 мкг рекомбинантной плазмиды и 6 мкл липосом (FuGENE 6 Transfection Reagent, Roche) перемешивали до гомогенного состояния в 100 мкл свежеприготовленного IMDM культурального бульона (GIBCO); смесь отстаивали в течение 15 мин, затем эту смесь добавляли к клеткам СНО (АТСС), которые культивировали в течение ночи после посева с плотностью клеток 3×105/мл в 6-луночном планшете; смесь культивировали при 37°С в 5% СO2 инкубаторе в течение 48 часов в подходящем для этих клеток культуральном бульоне, содержащем 88% IMDM, 10% FBS, 1% НТ и 1% глутамин (приобретенные у компании GIBCO); затем собирали супернатант, и, используя набор IgG ELISA для количественного определения белков (BETHYL), определяли относительное количество слитого белка. Относительное количество слитого белка, экспрессированного и секретированного клетками СНО, определяли следующим образом: 100 мкл аффинно очищенного античеловеческого IgG-Fc белка (10 мкг/мл) наносили на 96-луночный планшет ELISA (IMMULON), который затем промывали 5 раз 300 мкл раствора PBST для промывки; каждую покрытую лунку блокировали 200 мкл свежеприготовленного рабочего блокирующего раствора (маточный блокирующий раствор:PBS=1:19) и инкубировали при 37°С в течение 1 часа; после 5-ти разовой промывки 300 мкл раствора PBST для промывки в каждую лунку добавляли 100 мкл раствора IgG, используя градиентное разбавление (концентрация исходного раствора - 200 нг/мл, и разбавленный PBS с кратностью разбавления 1:2), используемого в качестве стандарта, и 100 мкл культурального супернатанта каждого слитого белка, используя градиентное разбавление (начиная с концентрации каждого культурального супернатанта и разбавляя PBS с кратностью разбавления 1:5), и инкубировали при 37°С в течение 2 ч; после 5-ти разовой промывки 300 мкл раствора для промывки добавляли 100 мкл античеловеческих IgG Fc-HRP вторичных антител, разбавленных PBS в отношении 1:10000, и инкубировали при 37°С течение 1 часа; после 5-ти разовой промывки 300 мкл раствора для промывки планшет проявляли путем добавления в лунку 100 мкл раствора для проявки (KPL); и наконец, реакцию останавливали путем добавления 100 мкл останавливающего раствора (KPL), с планшеты ELISA считывали показатели поглощения при длине волны 450 нм, используя ELISA ридер. Используя стандартную кривую, определяли концентрации различных слитых белков.
Пример 3: Связывание слитых белков
Способность сконструированных слитых белков связываться с VEGF и FGF-2 определяли методом ELISA. Сначала 96-луночный ELISA планшет (IMMULON Company) покрывали 100 мкл раствора (100 нг/мл гепарина (Sigma)), содержащего 20 нг/мл VEGF (R&D Systems) и 50 нг/мл FGF-2 (R&D Systems). Затем планшет 5 раз промывали 300 мкл раствора PBST для промывки, после чего каждую лунку блокировали добавлением 200 мкл свежеприготовленного рабочего блокирующего раствора (KPL Company) (маточный блокирующий pacтвоp: PBS=1:19) и инкубировали при 37°С в течение 1 часа. После 5-ти разовой промывки 300 мкл раствора PBST для промывки добавляли по 100 мкл растворов, содержащих различные слитые белки (растворенные в PBS, рН=7,2, 20 нг/мл), и инкубировали при 37°С в течение 2 часов. После 5-ти разовой промывки 300 мкл раствора PBST для промывки добавляли 100 мкл античеловеческих IgG Fc-HRP вторичных антител (BETHYL Company), разбавленных PBS в отношении 1:10000, и инкубировали при 37°С в течение 1 часа. После 5-ти разовой промывки 300 мкл раствора PBST для промывки планшет проявляли путем добавления в лунку 100 мкл раствора для проявки (KPL Company), и, наконец, реакцию останавливали добавлением 100 мкл останавливающего раствора (KPL Company), после чего с ELISA планшета считывали поглощение при длине волны 450 нм на ELISA ридере. Более высокая способность слитого белка связываться с VEGF или FGF2 соответствует более высокому уровню поглощения и, соответственно, более сильному сигналу. Исходя из интенсивности сигнала было выявлено, что слитые белки 25# и 28# обладают более высокой связывающей способностью как по отношению к VEGF, так и по отношению к FGF-2. Результаты сравнения способности слитых белков связываться с VEGF и FGF-2 показаны на Фиг. 2.
Пример 4: Стабильная экспрессия и очистка слитых белков
DHFR-дефектные клетки СНО (АТСС) трансфицировали 25# слитым белком (высокая связывающая способность) при помощи рекомбинантной экспрессионной плазмиды, а также слитыми белками 12# и 15# (контроль), используя липосомы (Roche). В частности, 5 мкг рекомбинантной плазмиды и 30 мкл липосом (FuGENE 6 Transfection Reagent, Roche) смешивали до получения гомогенной смеси в 100 мкл свежеприготовленного культурального бульона IMDM (GIBCO); затем выдерживали 15 мин, после чего эту смесь добавляли к DHFR-дефектным клеткам СНО (АТСС), которые культивировали в течение ночи после посева клеток на 10 см культуральные чашки (Corning) с плотностью посева клеток 3×105/мл; смесь культивировали при 37°С в 5% СO2 инкубаторе в течение 2-3 дней с соответствующим для этих клеток культуральным бульоном, содержащим 10% FBS, 1% НТ и 1% глутамин в культуральной среде IMDM (все компоненты были приобретены у компании GIBCO); затем клетки обрабатывали трипсином (GIBCO), засевали с плотностью посева клеток 3×105/мл в колбу, содержащую 30 мл бессывороточной культуральной среды 302 (SAFC), и селективно культивировали при 37°С в 5% СО2 инкубаторе при 100 об/мин при плотности клеток 106/мл. Затем 3000 клеток засевали на 10 см культуральные чашки (Corning) (с культуральным бульоном, содержащим в культуральной среде IMDM 10% FBS и 1% глутамин) и культивировали при 37°С в 5% СO2 инкубаторе для формирования единичных клонов. Полученные единичные клоны отбирали и культивировали в 96-луночном планшете (Corning). Относительное количество слитого белка, экспрессированного и секретированного каждым отдельным клоном, определяли при помощи набора ELISA (человеческий IgG) для количественного определения белка (BETHYL) в тех же самых условиях и таким же образом, как описано в Примере 2 для определения относительного количества слитого белка. Отбирали клон с самым высоким уровнем экспрессии и переносили в 6-луночный планшет для культивирования со степенью смыкания монослоя примерно 70%. Клетки обрабатывали трипсином и переносили в 10 см культуральную чашку. Затем проводили амплификацию в условиях стресса, силу которого увеличивали путем добавления метатрексата (МТХ, Sigma) с разной концентрацией (10 нМ, 20 нМ, 50 нМ, 100 нМ, 200 нМ и 500 нМ). После амплификации в условиях стресса клетки обрабатывали трипсином и засевали в колбу с плотностью посева 3×105/мл. Определяли величину экспрессии одной клетки с тем, чтобы получить линии клеток СНО, разработанные методом генной инженерии, экспрессирующие конкретный белок. Наконец, массовую суспензионную культуру (объем 10 л) линии клеток СНО, созданную методом генной инженерии, получали культивированием в бессывороточной культуральной среде 302 (рН 7,0, SAFC) при 37°С, в 5% СO2, 40% растворенном кислороде и при перемешивании со скоростью 80 об/мин. Культивированный продукт собирали центрифугированием. Затем супернатант фильтровали через 0,45 мкм мембранный фильтр (Millipore), аффинную хроматографию проводили согласно инструкции в колонке с белком A (GE) следующим образом: сначала колонку для аффинной хроматографии с белком А уравновешивали буфером PBS (рН 7,0); затем в колонку загружали супернатант и снова промывали буфером PBS; наконец, колонку элюировали цитратным буфером (рН 3,0), элюент собирали и фильтровали через 0,45 мкм мембранный фильтр. После инактивации вируса путем добавления S/D (0,3% трибутилфосфат/1% Твин 80) при 24°С в течение 6 часов, проводили дополнительную очистку целевого белка хроматографией на молекулярных ситах следующим образом: сначала в диализном мешке проводили диализ элюента, полученного после аффинной хроматографии с белком А, против буфера PBS; затем образец концентрировали в 10 KD ультрафильтрационной ячейке (Millipore); после чего концентрированный в ультрафильтрационной ячейке образец загружали в уравновешенную буфером PBS колонну с Superdex 200 (GE) для хроматографии с молекулярными ситами, затем колонну элюировали буфером PBS, и собирали фракцию с пиком элюции. Очищенный белок анализировали при помощи SDS-PAGE (Фиг.3); затем элюаты, содержащие требуемый продукт экспрессии, объединяли и фильтровали через 0,22 мкм мембранный фильтр (Millipore) с последующим определением содержания белка, осуществляемым множеством методов, например, определением белка методом Лоури.
Пример 5: Изучение связывающей способности слитого белка в зависимости от его концентрации
Способность сконструированных описанным выше образом слитых белков связываться с VEGF и FGF-2 определяли методом ELISA, аналогично тому, как описано в Примере 3. Сначала 96-луночный ELISA планшет покрывали 100 мкл раствора, содержащего 20 нг/мл VEGF и 50 нг/мл FGF-2 (R&D Systems). Затем планшет 5 раз промывали 300 мкл буфера PBST, после чего каждую покрытую лунку блокировали 200 мкл свежеприготовленного рабочего блокирующего раствора (KPL) (маточный блокирующий раствор:PBS=1:19) и инкубировали при 37°C в течение 1 ч. После 5-ти кратной, промывки 300 мкл раствора PBST для промывки добавляли по 100 мкл растворов, содержащих различные слитые белки с разными концентрациями (исходное содержание белка составляло 1000000 пМ, которое разбавляли в отношении 1:5), инкубировали при 37°С в течение 2 часов. После 5-ти кратной промывки 300 мкл раствора PBST для промывки добавляли 100 мкл античеловеческих IgG Fc-HRP вторичных антител (BETHYL), разбавленных буфером PBS в отношении 1:10000, и инкубировали при 37°С в течение 2 часов. После 5-ти кратной промывки 300 мкл раствора PBST для промывки планшет проявляли путем добавления в лунку 100 мкл раствора для проявки (KPL), и, наконец, реакцию останавливали путем добавления 100 мкл останавливающего раствора (KPL), после чего с ELISA планшета считывали показатели поглощения при длине волны 450 нм на ELISA ридере. Исходя из интенсивности сигнала, определяли способность слитых белков связываться с VEGF и FGF-2 в зависимости от их концентрации. Результаты сравнения способности слитых белков 12#, 15# и 25# связываться с VEGF и FGF-2 в зависимости от их концентрации показаны на Фиг. 4А. Более того, конструкцию 25# также сравнивали со слитым белком FGFR-Fc (обозначенным 26#), см. Фиг. 4В. Наряду с прочим, FGFR-Fc слитый белок 26# содержит второй Ig-подобный домен FGFR, третий Ig-подобный домен FGFR и Fc-область, его аминокислотная последовательность приведена в SEQ ID NO: 71, а кодирующая его нуклеотидная последовательность приведена в SEQ ID NO: 72. Слитый белок 26# получали следующим образом: также как в примере 1, фрагмент FGFR1 амплифицировали методом ПЦР, используя в качестве матрицы кДНК пораженную раком ткань прямой кишки (праймеры: SEQ ID NO: 73 и SEQ ID NO: 74), a Fc-фрагмент IgGI амплифицировали методом ПЦР, используя в качестве матрицы кДНК, полученную из крови человека (праймеры: SEQ ID NO: 43 и SEQ ID NO: 44), затем эти фрагменты были клонированы в экспрессирующий вектор лигированием при помощи метода перекрывающегося ПЦР.В этом примере было показано, что способность связываться с VEGF и FGF-2 увеличивается с увеличением молярной концентрации VEGFR-FGFR-Fc слитого белка по изобретению, что определяется более сильным сигналом при длине волны 450 нм; при этом способность связываться с VEGF и FGF-2 уменьшается по мере разбавления молярной концентрации слитого белка по изобретению.
Пример 6: Изучение аффинности слитых белков
Аффинность слитого белка к VEGF или FGF-2 в системе растворов определяли в эксперименте по изучению аффинности. Сначала 96-луночный ELISA планшет покрывали 100 мкл раствора, содержащего 1,0 мкг/мл VEGF, или 100 мкл раствора, содержащего 2,0 мкг/мл антитела для захвата FGF-2 (R&D Systems). Затем планшет 5 раз промывали 300 мкл раствора PBST для промывки, каждую покрытую лунку блокировали рабочим блокирующим раствором (KPL) (см. пример 3) и инкубировали при 37°C в течение 1 часа. После 5-ти кратной промывки 300 мкл раствора PBST для промывки добавляли заранее приготовленную и инкубированную смесь слитых белков и FGF-2, а также стандарт, приготовленный методом серийных разведении, при этом смесь готовили следующим образом: исходную концентрацию Fc-слитых белков 12#, 15# и 25#, которая составляла 800000 пМ (растворенных в PBS), разбавляли в 10 раз, растворы слитых белков смешивали в отношении 1:1 с 20 пМ раствором VEGF, другими словами, итоговая исходная концентрация каждого слитого белка составляла 400000 пМ, а итоговая концентрация VEGF составляла 10 пМ; исходную концентрацию Fc-слитых белков 25# и 26#, которая составляла 200 пМ (растворенных в PBS), разбавляли в 2 раза, и растворы слитых белков смешивали в отношении 1:1 с 20 пМ раствором FGF-2, другими словами, итоговая исходная концентрация каждого слитого белка составляла 200 пМ, итоговая концентрация FGF-2 составляла 10 пМ. Планшет инкубировали при 37°C в течение 2 часов и 5 раз промывали 300 мкл раствора PBST для промывки, затем добавляли 100 мкл раствора антител для детектирования VEGF (100 нг/мл) или 100 мкл раствора антител для детектирования FGF-2 (250 нг/мл) (R&D systems, которая может специфически детектировать свободные антитела к VEGF или FGF-2). Планшет инкубировали при 37°С в течение 2 часов и 5 раз промывали 300 мкл раствора PBST для промывки, после чего добавляли HRP-меченный стрептавидин (R&D systems) (разбавленный PBS в отношении 1:200). Планшет инкубировали при 37°С в течение 2 часов и 5 раз промывали в 300 мкл раствора PBST для промывки, затем планшет проявляли в течение соответствующего времени (примерно 15-30 мин) путем добавления в лунку 100 мкл раствора для проявки (KPL). Наконец, реакцию останавливали путем добавления 100 мкл останавливающего раствора (KPL), с ELISA планшета считывали показатели поглощения при длине волны 450 нм на ELISA ридере. В смеси слитого белка и VEGF или FGF-2 определяли относительную концентрацию VEGF или FGF-2. Сравнивали аффинности слитых белков 12#, 15# и 25# к VEGF или FGF-2 в системе растворов, данные сравнения приведены на Фиг. 5А и 5В. Более того, слитый белок 25# сравнивали с FGFR-Fc слитым белком (обозначенным 26#), см. Фиг. 5С и 5D. Среди прочего, FGFR-Fc слитый белок 26# содержит второй Ig-подобный домен FGFR, третий Ig-подобный домен FGFR и Fc-белок. В этом примере было показано, что VEGFR-FGFR-Fc слитые белки, сконструированные в настоящем изобретении (например, 12#, 15# и 25#), имеют в системе растворов высокую аффинность как по отношению к VEGF, так и по отношению к FGF-2. Аффинность увеличивается с увеличением концентрации, на что указывает уменьшение количества VEGF и FGF-2 с увеличением концентрации слитого белка. Показатели аффинности VEGFR-FGFR-Fc слитых белков 12#, 15# и 25# по изобретению к VEGF или FGF-2 в системе растворов показаны на Фиг. 5. В этом примере было продемонстрировано, что в системе растворов сконструированные в настоящем изобретении VEGFR-FGFR слитые белки имеют аффинность как по отношению к VEGF, так и по отношению к FGF-2. Аффинность увеличивается с увеличением концентрации, на что указывает уменьшенное количество свободных VEGF и FGF-2.
Пример 7: Тест на ингибирование деления человеческих эндотелиальных клеток пупочной вены
Способность слитых белков подавлять деление клеток сосудистого эндотелия оценивали при помощи теста на деление клеток человеческие эндотелиальные клетки пупочной вены (HUVEC). Клетки HUVEC (AllCells) культивировали на фазе экспоненциального роста в полной среде HUVEC (AllCells) при 37°C в 5% СO2 инкубаторе. После обработки трипсином клетки HUVEC подсчитывали. В каждую лунку засевали 3000 клеток HUVEC в минимальной среде HUVEC, содержащей 1% FBS (AllCells) в 96-луночном планшете. Планшет культивировали в течение ночи при 37°C в 5% СО2 инкубаторе. Добавляли 100 мкл раствора VEGF (R&D Systems) (конечная концентрация 20 нг/мл) или раствора FGF-2 (R&D Systems) (конечная концентрация 5 нг/мл), разбавленного минимальным буфером HUVEC, содержащим 1% FBS, а также 100 мкл смеси с разным количеством слитых белков и FGF-2 (в которой конечная концентрация слитого белка составляла 40 пМ, разбавленного минимальной средой HUVEC, содержащей 1% FBS, в отношении 1:10, а конечная концентрация FGF-2 составляла 5 нг/мл) и культивировали в течение еще 3-4 дней. Затем эту культуральную среду удаляли и добавляли культуральную среду, содержащую 10% ССК-8 (DOJINDO), культивировали в течение 2 часов, а затем с ELISA планшета считывали показатели поглощения при длине волны 450 нм на ELISA ридере. Исходя из разницы в показателях поглощения, и определяли способность слитых белков ингибировать деление сосудистых эндотелиальных клеток, индуцированных VEGF или FGF-2. Влияние слитых белков на деление клеток HUVEC, индуцированное VEGF или FGF-2, относительные уровни ингибирования показаны на Фиг. 6. В этом примере было показано, что VEGFR-FGFR-Fc слитые белки, сконструированные в настоящем изобретении (например, 12#, 15# и 25#), на клеточном уровне проявляют биологическую активность и способны ингибировать индуцированное VEGF или FGF-2 деление клеток HUVEC, а также обладают способностью связываться с VEGF и FGF-2. Такая связывающая способность увеличивается с увеличением молярной концентрации, на что указывает ингибирование деления клеток HUVEC, индуцированного VEGF или FGF-2.
Настоящее изобретение было проиллюстрировано конкретными примерами. Однако для специалиста в данной области очевидно, что настоящее изобретение не ограничено этими конкретными вариантами осуществления. В пределах настоящего изобретения специалист в данной области может осуществлять различные изменения и модификации, и любые технические признаки, упомянутые в описании, могут быть объединены без изменения сущности изобретения и не выходя за его объем. Такие изменения и модификации входят в объем настоящего изобретения.
Список литературы
[1] Hanahan D, Weinberg RA. The hallmarks of cancer. Cell, 2000, 100(1): 57-70.
[2] Hanahan D, Folkman J. Patterns and emerging mechanisms of the angiogenic switch during tumorigenesis. Cell. 1996, 86:353-64.
[3] Ferrara N, Gerber HP, LeCouter J. The biology of VEGF and its receptors. Nat Med. 2003, 9: 669-76.
[4] Ferrara N. Vascular endothelial growth factor as a target for anticancer therapy. Oncologist. 2004, 1: 2-10.
[5] Jenab-Wolcott J, Giantonio BJ. Bevacizumab: current indications and future development for management of solid tumors. Expert Opin Biol Ther. 2009, 9(4): 507-17.
[6] Summers J, Cohen MH, Keegan P, Pazdur R. PDA drug approval summary: bevacizumab plus interferon for advanced renal cell carcinoma. Oncologist. 2010, 15(1): 104-11.
[7] Hsu JY, Wakelee НА. Monoclonal antibodies targeting vascular endothelial growth factor: current status and future challenges in cancer therapy. BioDrugs. 2009, 23(5): 289-304.
[8] Krupitskaya Y, Wakelee НА. Ramucirumab, a fully human mAb to the transmembrane signaling tyrosine kinase VEGFR-2 for the potential treatment of cancer. Curr Opin Investig Drugs. 2009, 10(6): 597-605.
[9] Hurwitz H, Fehrenbacher L, Novotny W, Cartwright T, Hainsworth J, Heim W, Berlin J, Baron A, Griffing S, Holmgren E, Ferrara N, Fyfe G, Rogers B, Ross R, Kabbinavar F. Bevacizumab plus irinotecan, fluorouracil, and leucovorin for metastatic colorectal cancer. N Engl J Med. 2004, 350(23): 2335-42.
[10] Sandler A, Gray R, Perry MC, Brahmer J, Schiller JH, Dowlati A, Lilenbaum R, Johnson DH. Paclitaxel-carboplatin alone or with bevacizumab for non-small-cell lung cancer. N Engi J Med. 2006, 355(24): 2542-50.
[11] Jenab-Wolcott J, Giantonio BJ. Bevacizumab: current indications and future development for management of solid tumors. Expert Opin Biol Ther, 2009, 9(4): 507-17.
[12] Dorrell MI, Aguilar E, Scheppke L, Barnett FH, Friedlander M. Combination angiostatic therapy completely inhibits ocular and tumor angiogenesis. Proc Natl Acad Sci U S A. 2007, 104(3): 967-72.
[13] Casanovas 0, Hicklin DJ, Bergers G, Hanahan D. Drug resistance by evasion of antiangiogenic targeting of VEGF signaling in late-stage pancreatic islet tumors. Cancer Cell. 2005, 8(4): 299-309.
[14] Welti JC, Gourlaouen M, Powles T, Kudahetti SC, Wilson P, Berney DM, Reynolds AR. Fibroblast growth factor 2 regulates endothelial cell sensitivity to sunitinib. Oncogene. 2010, doi:10.1038/onc.2010.503.
| название | год | авторы | номер документа |
|---|---|---|---|
| FGF-R-Fc СЛИТЫЙ БЕЛОК И ЕГО ИСПОЛЬЗОВАНИЕ | 2012 |
|
RU2560573C2 |
| СЛИТЫЕ БЕЛКИ, СОДЕРЖАЩИЕ СВЯЗЫВАЮЩИЕ PDGF И VEGF ЧАСТИ, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2014 |
|
RU2692652C2 |
| ВНЕКЛЕТОЧНЫЙ ДОМЕН ТИРОЗИНКИНАЗНОГО РЕЦЕПТОРА, СВЯЗЫВАЮЩИЙ АЛЛОСТЕРИЧЕСКИЙ ИНГИБИТОР | 2010 |
|
RU2604805C2 |
| БИФУНКЦИОНАЛЬНЫЙ ИНГИБИТОР АНГИОГЕНЕЗА И ЕГО ПРИМЕНЕНИЕ | 2019 |
|
RU2760738C1 |
| ОСНОВАННЫЕ НА Notch3 ЧЕЛОВЕКА ГИБРИДНЫЕ БЕЛКИ В КАЧЕСТВЕ ЛОВУШЕК-ИНГИБИТОРОВ ПЕРЕДАЧИ СИГНАЛА Notch3 | 2009 |
|
RU2567662C2 |
| БИСПЕЦИФИЧНЫЙ СЛИТЫЙ БЕЛОК И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2801528C2 |
| ХИМЕРНЫЕ БЕЛКИ, ИНГИБИРУЮЩИЕ АНГИОГЕНЕЗ, И ИХ ПРИМЕНЕНИЕ | 2005 |
|
RU2355414C2 |
| КОМПОЗИЦИИ ГУМАНИЗИРОВАННЫХ СЛИТЫХ БЕЛКОВ NOTCH И СПОСОБЫ ЛЕЧЕНИЯ | 2008 |
|
RU2532830C2 |
| АНТАГОНИСТЫ ALK1 И ИХ ПРИМЕНЕНИЕ В ЛЕЧЕНИИ ПОЧЕЧНО-КЛЕТОЧНОЙ КАРЦИНОМЫ | 2013 |
|
RU2633638C2 |
| ИНГИБИТОРЫ РЕЦЕПТОРОВ ФАКТОРА РОСТА ЭНДОТЕЛИЯ СОСУДОВ ТИПА 2 | 2004 |
|
RU2402567C2 |
































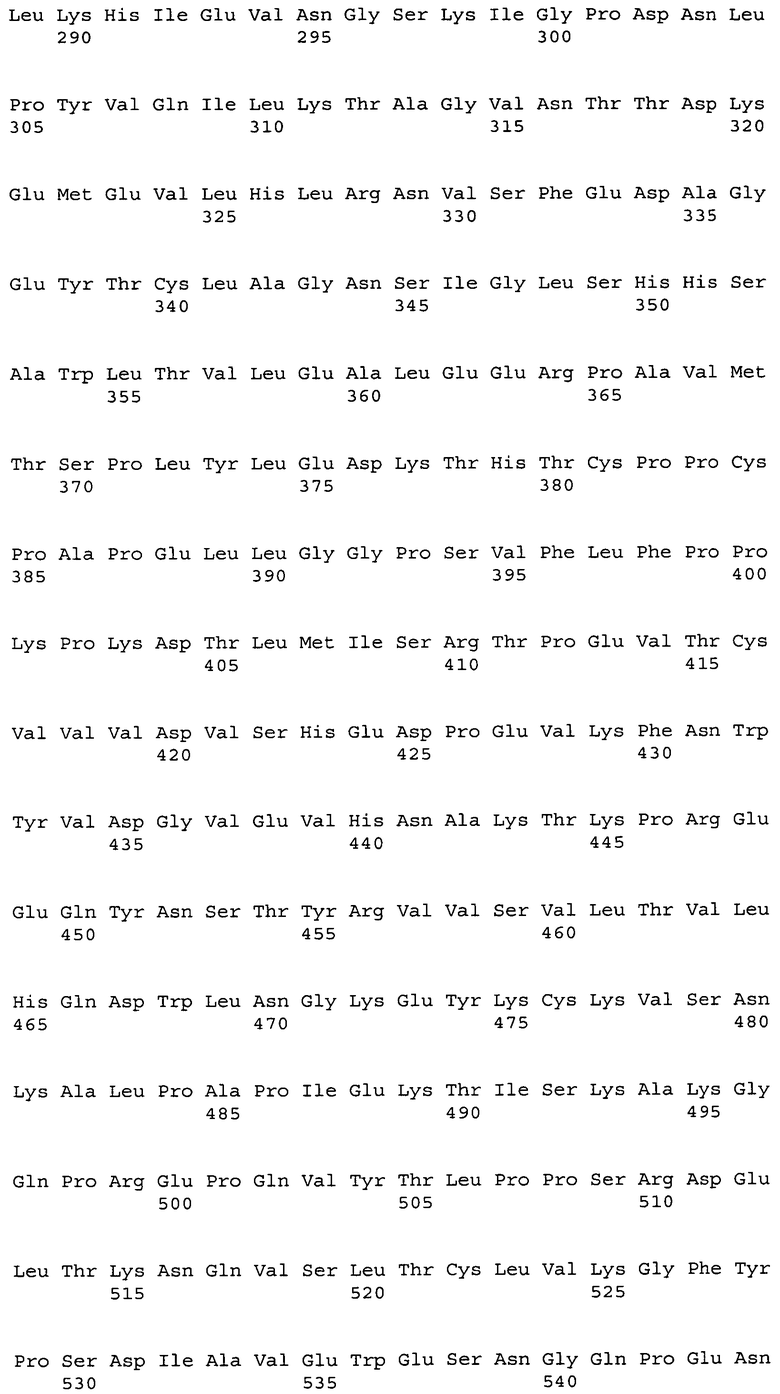







































































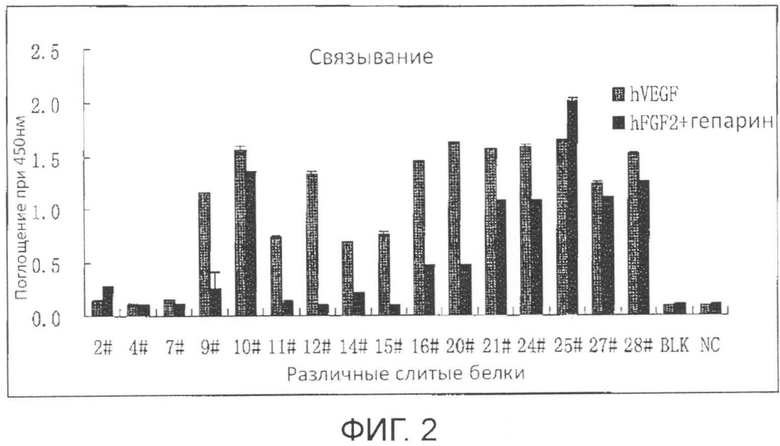
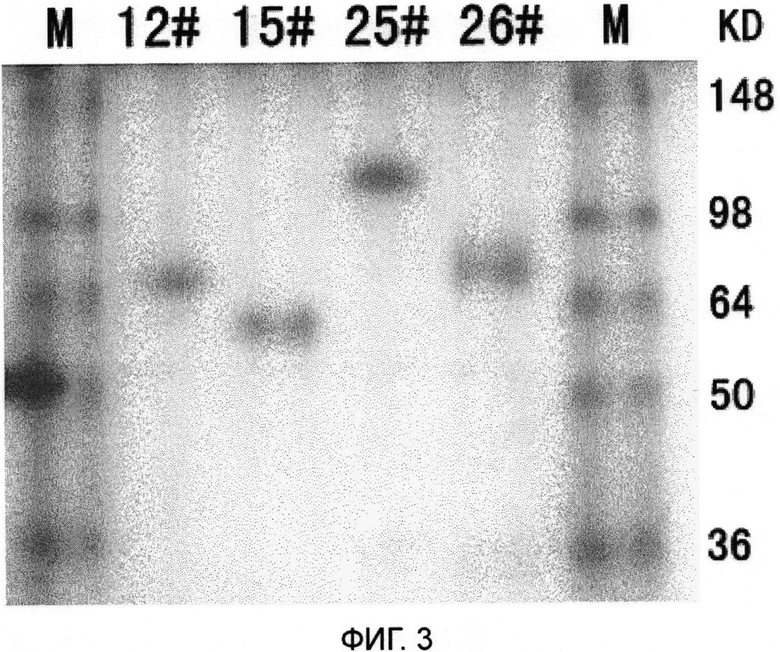
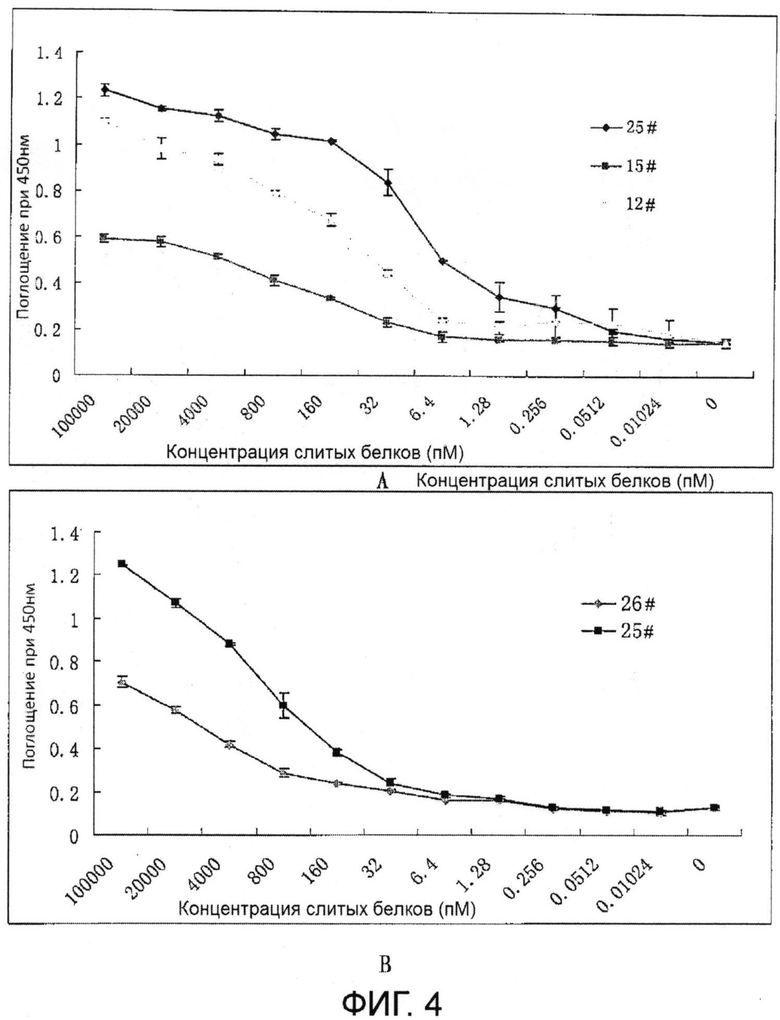
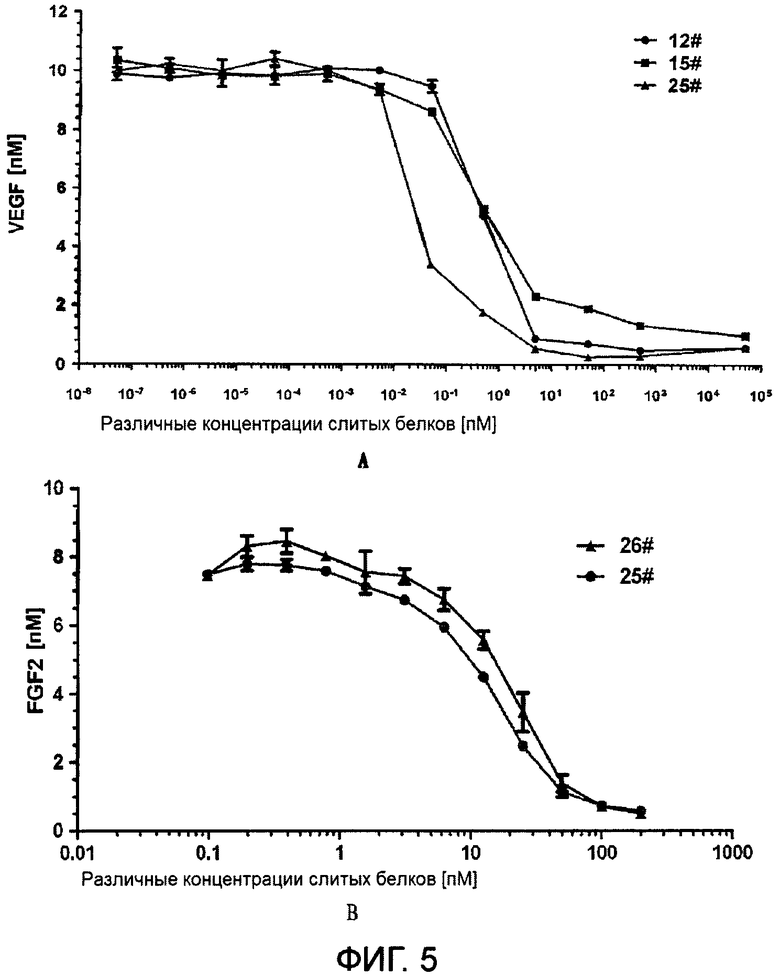
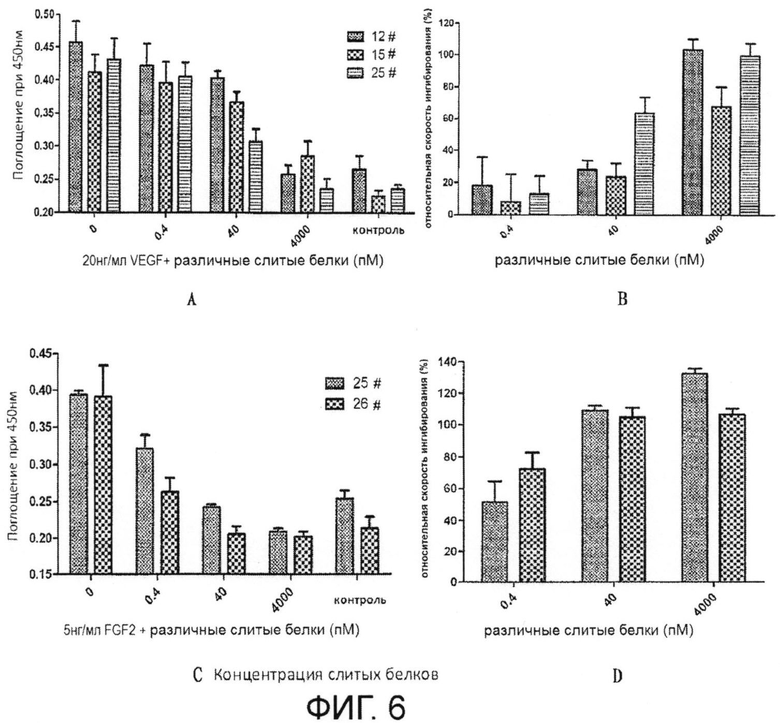
Настоящее изобретение относится к области биотехнологии, конкретно к ингибиторам ангиогенеза, и может быть использовано в медицине для ингибирования патологического ангиогенеза. Получают слитый белок, состоящий из (A) части, полученной из внеклеточного домена VEGFR, (B) части, полученной из внеклеточного домена FGFR, и (C) Fc-области иммуноглобулина. Часть (A), полученная из внеклеточного домена VEGFR, состоит из второго и третьего Ig-подобных доменов VEGFR2 или из первого, второго и третьего Ig-подобных доменов VEGFR2. Часть (В), полученная из внеклеточного домена FGFR, состоит из (i) части, полученной из участка промежуточной функциональной последовательности Ig-подобного домена FGFR, которая представляет собой последовательность между первым и вторым Ig-подобным доменом в белке FGFR, (ii) из второго Ig-подобного домена FGFR и (iii) третьего Ig-подобного домена FGFR. Изобретение позволяет эффективно использовать полученные слитые белки для лечения заболеваний, связанных с нарушением регуляции ангиогенеза. 10 н. и 11 з.п. ф-лы, 6 ил., 2 табл., 7 пр.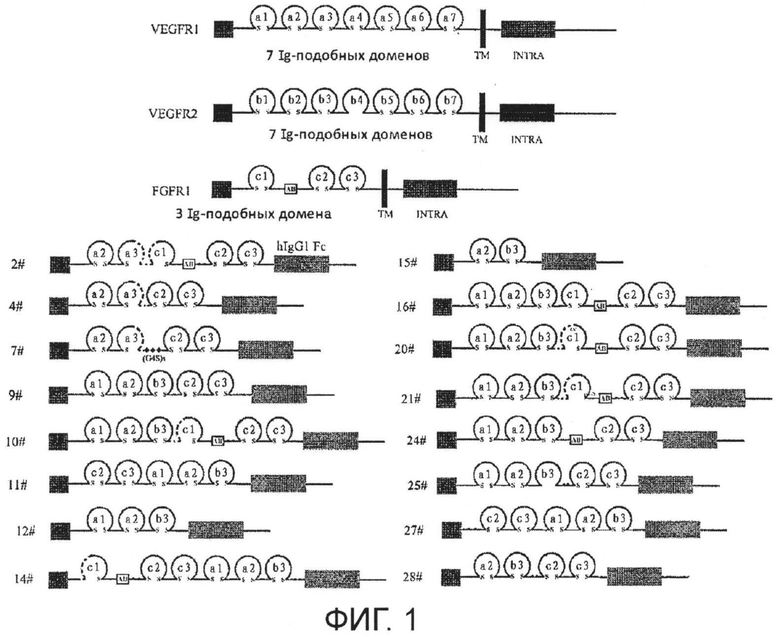
1. Ингибирующий ангиогенез слитый белок, состоящий из по меньшей мере одной части, полученной из внеклеточного домена VEGFR, по меньшей мере одной части, полученной из внеклеточного домена FGFR, и Fc-области иммуноглобулина,
где указанная часть, полученная из внеклеточного домена VEGFR, состоит из второго Ig-подобного домена VEGFR1 и третьего Ig-подобного домена VEGFR2 или из первого Ig-подобного домена VEGFR1, второго Ig-подобного домена VEGFR1 и третьего Ig-подобного домена VEGFR2; указанная часть, полученная из внеклеточного домена FGFR, состоит из части, полученной из участка промежуточной функциональной последовательности Ig-подобного домена FGFR, второго Ig-подобного домена FGFR и третьего Ig-подобного домена FGFR,
где указанный участок промежуточной функциональной последовательности Ig-подобного домена FGFR представляет собой последовательность между первым Ig-подобным доменом и вторым Ig-подобным доменом в белке FGFR.
2. Слитый белок по п. 1, где указанный слитый белок состоит последовательно от конца N к концу С из части, полученной из внеклеточного домена VEGFR, части, полученной из внеклеточного домена FGFR, и Fc-области иммуноглобулина.
3. Слитый белок по п. 2, где часть, полученная из внеклеточного домена VEGFR, состоит последовательно от конца N к концу С из второго Ig-подобный домена VEGFR1 и третьего Ig-подобного домена VEGFR2 или из первого Ig-подобного домена VEGFR1, второго Ig-подобного домена VEGFR1 и третьего Ig-подобного домена VEGFR2, а
часть, полученная из внеклеточного домена FGFR, состоит последовательно от конца N к концу С из части, полученной из участка промежуточной функциональной последовательности Ig-подобного домена FGFR, второго Ig-подобного домена FGFR и третьего Ig-подобного домена FGFR.
4. Слитый белок по п. 2 или 3, где часть, полученная из внеклеточного домена VEGFR, состоит последовательно от конца N к концу С из первого Ig-подобного домена VEGFR1, второго Ig-подобного домена VEGFR1 и третьего Ig-подобного домена VEGFR2; часть, полученная из внеклеточного домена FGFR, состоит последовательно от конца N к концу С из части, полученной из участка промежуточной функциональной последовательности Ig-подобного домена FGFR, второго Ig-подобного домена FGFR и третьего Ig-подобного домена FGFR.
5. Слитый белок по п. 2 или 3, где:
второй Ig-подобный домен VEGFR1 имеет аминокислотную последовательность, соответствующую положениям 151-214 SEQ ID NO: 2,
третий Ig-подобный домен VEGFR2 имеет аминокислотную последовательность, соответствующую положениям 224-320 SEQ ID NO: 3,
второй Ig-подобный домен FGFR1 имеет аминокислотную последовательность, соответствующую положениям 163-247 SEQ ID NO: 1, и
третий Ig-подобный домен FGFR1 имеет аминокислотную последовательность, соответствующую положениям 270-359 SEQ ID NO: 1.
6. Слитый белок по п. 4, где первый Ig-подобный домен VEGFR1 имеет аминокислотную последовательность, соответствующую положениям 32-123 SEQ ID NO: 2.
7. Слитый белок по п. 1, где часть, полученная из участка промежуточной функциональной последовательности Ig-подобного домена FGFR, не содержит кислый бокс.
8. Слитый белок по п. 1, где часть, полученная из участка промежуточной функциональной последовательности Ig-подобного домена FGFR, обладает аминокислотной последовательностью, соответствующей аминокислотной последовательности, которая начинается в положении от 118 до 151 и заканчивается в положении 162 в SEQ ID NO: 1.
9. Слитый белок по п. 8, где часть, полученная из участка промежуточной функциональной последовательности Ig-подобного домена FGFR, обладает аминокислотной последовательностью, соответствующей аминокислотной последовательности, которая начинается в положении 134 и заканчивается в положении 162 в SEQ ID NO: 1, аминокислотной последовательности, которая начинается в положении 145 и заканчивается в положении 162 в SEQ ID NO: 1, или аминокислотной последовательности, которая начинается в положении 151 и заканчивается в положении 162 в SEQ ID NO: 1.
10. Слитый белок по п. 1, где Fc-область иммуноглобулина представляет собой Fc-область человеческого IgG, например, Fc-область человеческого IgGl, более предпочтительно обладает аминокислотной последовательностью, соответствующей SEQ ID NO: 7, или аминокислотной последовательностью, кодируемой нуклеотидной последовательностью, соответствующей SEQ ID NO: 8.
11. Ингибирующий ангиогенез слитый белок, состоящий из аминокислотной последовательности, указанной в любой из SEQ ID NO: 9-24, или аминокислотной последовательности, кодируемой нуклеотидной последовательностью, указанной в любой из SEQ ID NO: 26-41.
12. Ингибирующий ангиогенез слитый белок, состоящий из слитого белка по любому из предшествующих пунктов и участка секреторного участка сигнального пептида, например, участка сигнального пептида VEGFR1, где, предпочтительно, указанный секреторный участок сигнального пептида имеет аминокислотную последовательность, соответствующую положениям 1-26 SEQ ID NO: 2 или аминокислотную последовательность, кодируемую нуклеотидной последовательностью SEQ ID NO: 25.
13. Изолированная молекула нуклеиновой кислоты, кодирующая слитый белок по любому из предшествующих пунктов.
14. Молекула нуклеиновой кислоты по п. 13, состоящая из нуклеотидной последовательности, указанной в любой из SEQ ID NO: 26-41.
15. Экспрессионный вектор, содержащий молекулу нуклеиновой кислоты по п. 13 или 14.
16. Клетка, способная экспрессировать слитый белок по любому из пп. 1-12, предпочтительно клетка СНО, трансфицированная вектором по п. 15, не являющаяся эмбриональной клеткой человека.
17. Фармацевтическая композиция для ингибирования ангиогенеза, содержащая эффективное количество слитого белка по любому из пп. 1-12, молекулы нуклеиновой кислоты по п. 13 или 14, вектора по п. 15 или клетки по п. 16, а также фармацевтически приемлемый носитель.
18. Способ получения ингибирующего ангиогенез слитого белка, осуществляемый экспрессией слитого белка по любому из пп. 1-12 в прокариотических или эукариотических клетках.
19. Способ по п. 18, осуществляемый экспрессией слитого белка по любому из пп. 1-12 в клеточной линии СНО.
20. Применение слитого белка по любому из пп. 1-12, молекулы нуклеиновой кислоты по п. 13 или 14, вектора по п. 15, клетки по п. 16 или фармацевтической композиции по п. 17 в приготовлении лекарственного средства для ингибирования патологического ангиогенеза у млекопитающих.
21. Применение слитого белка по любому из пп. 1-12, молекулы нуклеиновой кислоты по п. 13 или 14, вектора по п. 15, клетки по п. 16 или фармацевтической композиции по п. 17 в приготовлении лекарственного средства для лечения или предотвращения опухолей и/или заболеваний, связанных с офтальмологическим ангиогенезом, предпочтительно, где указанная опухоль представляет собой солидную опухоль, а указанные заболевания, связанные с офтальмологическим ангиогенезом, выбраны из группы, состоящей из возрастной дегенерации желтого пятна и диабетической ретинопатии.
| WO 2006113277 A2, 26.10.2006 | |||
| ГИДРОМЕТЕОРОЛОГИЧЕСКИЙ РЕАКТИВНЫЙ СНАРЯД | 1995 |
|
RU2083081C1 |
| US 20100055093 A1, 04.03.2010 | |||
| CN 101679988 А, 24.03.2010 | |||
| АНТИАНГИОГЕННЫЕ СОЕДИНЕНИЯ | 2007 |
|
RU2418003C2 |
| EA 200970515 A1, 30.12.2009 | |||

